Embed Size (px)
Citation preview

European Journal of Pharmacology 626 (2010) 159–165
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Molecular and Cellular Pharmacology
Estrogen-like activities of saikosaponin-d in vitro: A pilot study
Peng Wang a, Jianlin Ren b, Jian Tang a, Dali Zhang a, Bai Li c, Yong Li a,⁎a Department of Gastroenterology, Shanghai Municipal Hospital of Traditional Chinese Medicine, Shanghai University of Traditional Chinese Medicine, Shanghai 200071, Chinab Department of Medical Intelligence, Shanghai Research Institute of Medical Technique Intelligence, Shanghai 200031, Chinac Department of Chinese Traditional Medicine, Changhai Hospital, Second Military Medical University, Shanghai 200433, China
⁎ Corresponding author. Tel.: +86 21 56639828 2303E-mail address: [email protected] (Y. Li).
0014-2999/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.ejphar.2009.09.047
a b s t r a c t
a r t i c l e i n f oArticle history:Received 8 March 2009Received in revised form 6 September 2009Accepted 28 September 2009Available online 9 October 2009
Keywords:Saikosaponin-dPhytoestrogenEstrogen receptorEstrogen response element
Saikosaponin-d (SSd), a saponin derivativewith a similar structure to estradiol, was extracted from Bupleurumfalcatum L. (Umbelliferae). It was found that SSd stimulated the proliferation of MCF-7 cells by usingMCF-7 cellproliferation assay. Cell-cycle analysis revealed that the proliferation-stimulating effect was associated with amarked increase in the number of MCF-7 cells in S phase. These actions of SSd were dose-dependent at dosesranging from 10 nM to 10 µM and could be significantly inhibited by the specific estrogen receptor (ER)antagonist ICI-182780. Co-incubation of MCF-7 cells with 1 μM of ER antagonist ICI-182780 abolished theinductive effects of SSd on estrogen response element (ERE)-luciferase activity, suggesting that the estro-genic effects of SSd were mediated through the estrogen receptors. To evaluate the relative involvement ofestrogen receptor alpha (ERα) and estrogen receptor beta (ERβ) in mediating the actions of SSd, ER-negativehuman cervical carcinoma (HeLa) cells were cotransfected with the ERE-luciferase reporter construct andeither ERα or ERβ construct. The results showed that SSd could activate ERE-luciferase activity via the ERα-mediated pathway in a dose-dependent manner (10 nM to 10 µM); whereas, the activation of ERβ-mediatedERE-luciferase activity by SSd only occurred at a high concentration (10 µM). Furthermore, the ERα proteinand mRNA levels were increased by treatment with SSd within 24 h. These data support our hypothesis thatSSd acts as a weak phytoestrogen. Presumably, the estrogenic effect of SSd is mediated by the estrogenreceptor.
; fax: +86 21 56639310.
l rights reserved.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Radix Bupleuri (Chinese name: Chaihu) has been used for over2000 years for the treatment of common cold with fever, hepatitis,kidney syndrome, inflammatory diseases, menoxenia, etc. (Yang et al.,2007; Zhao et al., 1996). It is believed that saikosaponins areresponsible for part of the pharmaceutical properties of Radix Bupleuri(Zhu et al., 2006). In particular, saikosaponin-d (SSd) is especiallyknown for its pharmacological activities such as anticancer, anti-inflammation, and liver protection, as well as corticosterone-likeactivity (Aoyagi et al., 2001; Hsu et al., 2004; Leung et al., 2005;Morinaga et al., 2006). Furthermore, Xiao-Chai-Hu-Tang (Japanesename: Sho-Saiko-To), a Bupleurum soup for oral administration, themain active constituent of which is SSd, has achieved great success inthe treatment of menoxenia, menopausal symptoms and hepaticfibrosis (Ikegami et al., 2003). Various studies have indicated thatBupleurum has estrogenic activity, although the detailed mechanismof action, the effective dosages as well as its efficacy in differentclinical conditions are not clear. We assumed that, as the main
constituent of Bupleurum soup, saikosaponin-d might be a new typeof phytoestrogen.
Many flavone glycosides, including isoflavones such as daidzeinand genistein, are structurally similar to estrogens and are believed topossess the estrogenic activities through ER-dependent pathway andER-independent pathway (Singhal et al., 2009). The estrogenicactivities of phytoestrogens are closely associated with their struc-tures. The number and position of hydroxyl groups in the molecularstructure are other important factors which contribute to strongbinding to estrogen receptors and the resulting estrogenic activity(Ding et al., 2005). Interestingly, it seems that SSd has similarstructure with estrogen mimetics (Fig. 1).
There are two major types of estrogen receptors, ERα and ERβ,which differ in their ligand specificities and physiological functions.Certain natural hormones or phytochemicals are being shown topossess differential affinity for and transactivation of ERα versus ERβ.For example, the natural hormone estradiol binds with equal affinityto ERα and ERβ, while the well-known phytoestrogens genistein andcoumestrol, preferentially interact with and activate ERβ to mediateestrogenic effects in vitro assays (Brigitte and Johannes, 2001). Thus,one of our aims in the present study is to observe whether SSdactivates ER isoforms α and β differentially.
In this study, by using the estrogen-dependent human breastcancer cell line MCF-7 and the ER-negative human cervical carcinoma

Fig.1. Chemical structures of related compounds.
160 P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
cell line HeLa as model systems, we investigated the estrogenic-likeactivity of SSd as well as the associated mechanisms.
2. Materials and methods
2.1. Materials
SSd was purchased from Sigma-Aldrich (St. Louis. MO, USA) with apurity greater than 95%. The extract is quite stable at roomtemperature and retains its activity after exposure with organicsolvents such as dimethyl sulphoxide (DMSO). ICI-182780, DMSO,estradiol, 3-(4, 5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazoliumbromile (MTT), phenol red-free Dulbecco's modified Eagle's medium(phenol red-free DMEM), Triton-x 100, ribonuclease A, propidiumiodide and LipofectamineTM 2000 reagent were purchased fromSigma-Aldrich (St. Louis. MO, USA). The mouse monoclonal antibodyagainst β-actin and the rabbit polyclonal antihuman estrogen receptorα antibody were purchased from Upstate (Charlottesville, VA, USA).Goat antirabbit-HRP and goat antimouse-HRP conjugates were pur-chased from Bio-Rad Laboratories (Hercules, CA). Fetal bovine serumand charcoal-stripped fetal bovine serum were obtained from GibcoBRL Life Technologies.
2.2. Cell culture
MCF-7 cells (human breast cancer) and HeLa cells (human cervicalcarcinoma), (from Cell Biological Research Institution of ChineseAcademy of Sciences, Shanghai, China) were routinely cultured inDulbecco's modified Eagle's medium (DMEM) supplemented with 5%fetal bovine serum (FBS), maintained in 5% CO2 atmosphere at 37 °C.Cells grown to 85% confluence were transferred to phenol red-freeDMEM supplemented with 5% charcoal-stripped fetal bovine serum(sFBS) for 2 days, in order to minimize the estrogenic activity ofserum. Then cells were treated with SSd or estradiol at differentconcentrations. SSd, estradiol and ICI-182780 were dissolved inDMSO. All were diluted in the medium immediately before use
(final concentration of MSO <0.1%). DMSO (<0.1%) alone did nothave any effect on the parameters measured.
2.3. MTT growth assay
MCF-7 cells were washed twice with phosphate-buffered salinesolution (PBS), counted and seeded into 96-well plates at a density of0.5×104 cells/well. After 24 h, the cells were completely attached tothe well bottom. Cell proliferation was assessed after 24 h. In theassessment method, cells were incubated with 100 µl of 0.5 mg/mlMTT solution for 4 h. Following this, the medium was discarded, and150 µl DMSO was added. Absorbance at 492 nmwas determined withan ELx800 universal microplate reader (Bio-Rad, USA). By MTTmethod, cell numbers were obtained as absorbance values. Theresults are expressed as proliferation compared with that induced bytreatment with estradiol.
2.4. Interaction with ICI-182780
Some phytoestrogens, such as genistein and daidzein, act both asagonists and antagonists at estrogen receptors. In the present study,we examined the effect of SSd alone as well as its interaction withICI-182780, a pure estrogen receptor antagonist. The effects of dif-ferent concentrations of SSdwith orwithout 1 µM ICI-182780 on cellproliferation were assessed with the MTT method after 24 h, asdescribed above.
2.5. Cell-cycle analysis
Exponentially growing cells were plated in 6-well plates in phenolred-free DMEM containing 5% sFBS for 24 h and treated with testcompounds. After 24 h, the cells were harvested, and 1×106 cells wereplaced into a polypropylene tube and centrifuged. The supernatant wasremoved and 1 ml 4 °C 95% ethanol was added dropwise to the cellpellet during vortexing. Cells were kept at 4 °C until DNA staining. Fixedcells were treated with 200 µg/ml ribonuclease A in PBS, followed by
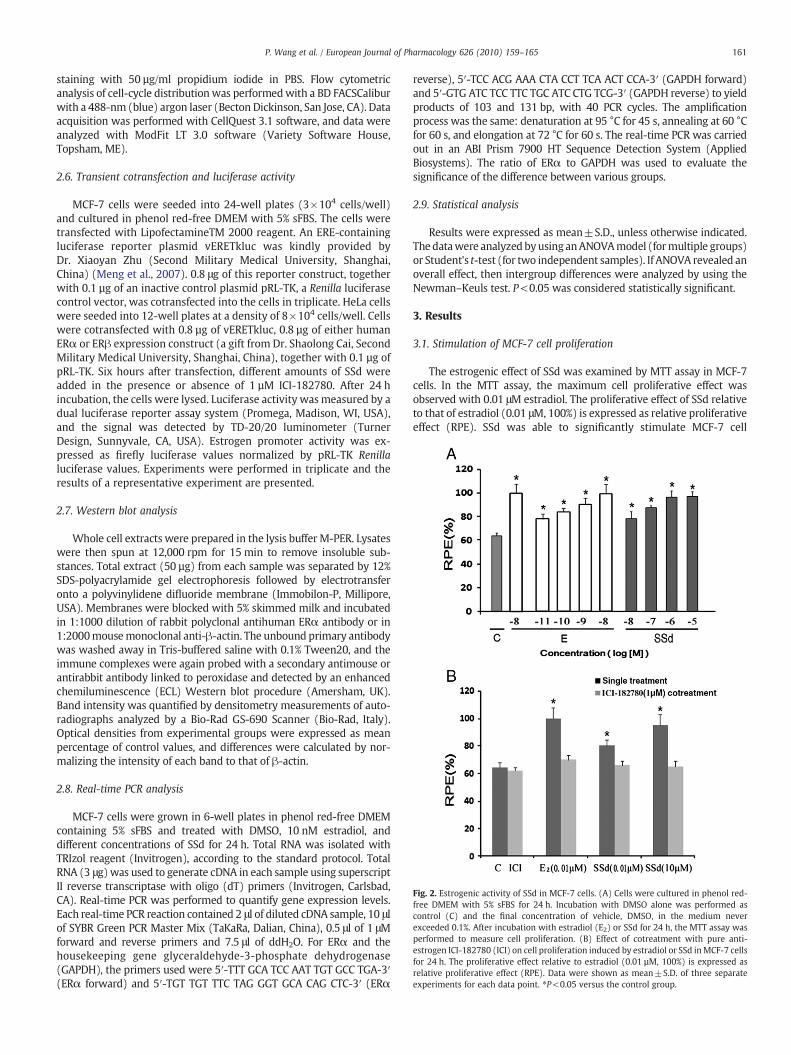
Fig. 2. Estrogenic activity of SSd in MCF-7 cells. (A) Cells were cultured in phenol red-free DMEM with 5% sFBS for 24 h. Incubation with DMSO alone was performed ascontrol (C) and the final concentration of vehicle, DMSO, in the medium neverexceeded 0.1%. After incubation with estradiol (E2) or SSd for 24 h, the MTT assay wasperformed to measure cell proliferation. (B) Effect of cotreatment with pure anti-estrogen ICI-182780 (ICI) on cell proliferation induced by estradiol or SSd inMCF-7 cellsfor 24 h. The proliferative effect relative to estradiol (0.01 µM, 100%) is expressed asrelative proliferative effect (RPE). Data were shown as mean±S.D. of three separateexperiments for each data point. ⁎P<0.05 versus the control group.
161P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
staining with 50 µg/ml propidium iodide in PBS. Flow cytometricanalysis of cell-cycle distributionwas performedwith a BD FACSCaliburwith a 488-nm (blue) argon laser (Becton Dickinson, San Jose, CA). Dataacquisition was performed with CellQuest 3.1 software, and data wereanalyzed with ModFit LT 3.0 software (Variety Software House,Topsham, ME).
2.6. Transient cotransfection and luciferase activity
MCF-7 cells were seeded into 24-well plates (3×104 cells/well)and cultured in phenol red-free DMEM with 5% sFBS. The cells weretransfected with LipofectamineTM 2000 reagent. An ERE-containingluciferase reporter plasmid vERETkluc was kindly provided byDr. Xiaoyan Zhu (Second Military Medical University, Shanghai,China) (Meng et al., 2007). 0.8 μg of this reporter construct, togetherwith 0.1 µg of an inactive control plasmid pRL-TK, a Renilla luciferasecontrol vector, was cotransfected into the cells in triplicate. HeLa cellswere seeded into 12-well plates at a density of 8×104 cells/well. Cellswere cotransfected with 0.8 µg of vERETkluc, 0.8 µg of either humanERα or ERβ expression construct (a gift from Dr. Shaolong Cai, SecondMilitary Medical University, Shanghai, China), together with 0.1 µg ofpRL-TK. Six hours after transfection, different amounts of SSd wereadded in the presence or absence of 1 µM ICI-182780. After 24 hincubation, the cells were lysed. Luciferase activity wasmeasured by adual luciferase reporter assay system (Promega, Madison, WI, USA),and the signal was detected by TD-20/20 luminometer (TurnerDesign, Sunnyvale, CA, USA). Estrogen promoter activity was ex-pressed as firefly luciferase values normalized by pRL-TK Renillaluciferase values. Experiments were performed in triplicate and theresults of a representative experiment are presented.
2.7. Western blot analysis
Whole cell extracts were prepared in the lysis buffer M-PER. Lysateswere then spun at 12,000 rpm for 15 min to remove insoluble sub-stances. Total extract (50 µg) from each sample was separated by 12%SDS-polyacrylamide gel electrophoresis followed by electrotransferonto a polyvinylidene difluoride membrane (Immobilon-P, Millipore,USA). Membranes were blocked with 5% skimmed milk and incubatedin 1:1000 dilution of rabbit polyclonal antihuman ERα antibody or in1:2000mousemonoclonal anti-β-actin. The unbound primary antibodywas washed away in Tris-buffered saline with 0.1% Tween20, and theimmune complexes were again probed with a secondary antimouse orantirabbit antibody linked to peroxidase and detected by an enhancedchemiluminescence (ECL) Western blot procedure (Amersham, UK).Band intensity was quantified by densitometry measurements of auto-radiographs analyzed by a Bio-Rad GS-690 Scanner (Bio-Rad, Italy).Optical densities from experimental groups were expressed as meanpercentage of control values, and differences were calculated by nor-malizing the intensity of each band to that of β-actin.
2.8. Real-time PCR analysis
MCF-7 cells were grown in 6-well plates in phenol red-free DMEMcontaining 5% sFBS and treated with DMSO, 10 nM estradiol, anddifferent concentrations of SSd for 24 h. Total RNA was isolated withTRIzol reagent (Invitrogen), according to the standard protocol. TotalRNA (3 μg) was used to generate cDNA in each sample using superscriptII reverse transcriptase with oligo (dT) primers (Invitrogen, Carlsbad,CA). Real-time PCR was performed to quantify gene expression levels.Each real-time PCR reaction contained 2 μl of diluted cDNA sample, 10 μlof SYBR Green PCR Master Mix (TaKaRa, Dalian, China), 0.5 μl of 1 μMforward and reverse primers and 7.5 μl of ddH2O. For ERα and thehousekeeping gene glyceraldehyde-3-phosphate dehydrogenase(GAPDH), the primers used were 5′-TTT GCA TCC AAT TGT GCC TGA-3′(ERα forward) and 5′-TGT TGT TTC TAG GGT GCA CAG CTC-3′ (ERα
reverse), 5′-TCC ACG AAA CTA CCT TCA ACT CCA-3′ (GAPDH forward)and 5′-GTG ATC TCC TTC TGC ATC CTG TCG-3′ (GAPDH reverse) to yieldproducts of 103 and 131 bp, with 40 PCR cycles. The amplificationprocess was the same: denaturation at 95 °C for 45 s, annealing at 60 °Cfor 60 s, and elongation at 72 °C for 60 s. The real-time PCR was carriedout in an ABI Prism 7900 HT Sequence Detection System (AppliedBiosystems). The ratio of ERα to GAPDH was used to evaluate thesignificance of the difference between various groups.
2.9. Statistical analysis
Results were expressed as mean±S.D., unless otherwise indicated.The datawere analyzed byusing anANOVAmodel (formultiple groups)or Student's t-test (for two independent samples). If ANOVA revealed anoverall effect, then intergroup differences were analyzed by using theNewman–Keuls test. P<0.05 was considered statistically significant.
3. Results
3.1. Stimulation of MCF-7 cell proliferation
The estrogenic effect of SSd was examined by MTT assay in MCF-7cells. In the MTT assay, the maximum cell proliferative effect wasobserved with 0.01 μM estradiol. The proliferative effect of SSd relativeto that of estradiol (0.01 μM, 100%) is expressed as relative proliferativeeffect (RPE). SSd was able to significantly stimulate MCF-7 cell
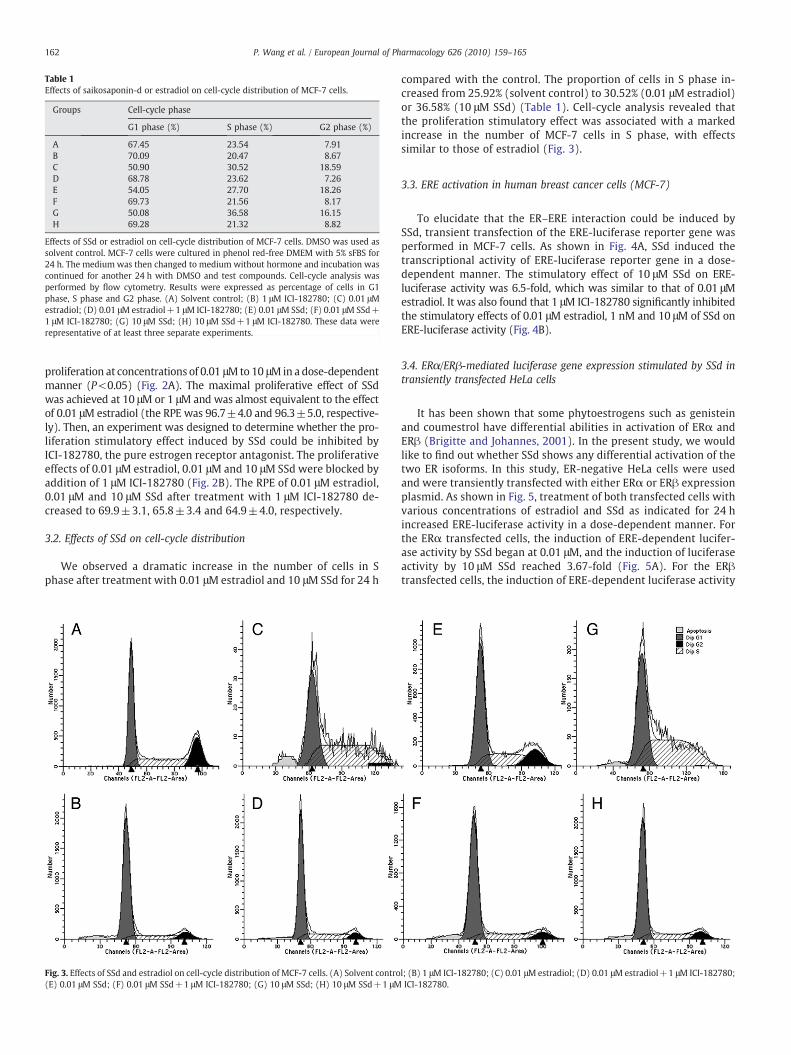
Table 1Effects of saikosaponin-d or estradiol on cell-cycle distribution of MCF-7 cells.
Groups Cell-cycle phase
G1 phase (%) S phase (%) G2 phase (%)
A 67.45 23.54 7.91B 70.09 20.47 8.67C 50.90 30.52 18.59D 68.78 23.62 7.26E 54.05 27.70 18.26F 69.73 21.56 8.17G 50.08 36.58 16.15H 69.28 21.32 8.82
Effects of SSd or estradiol on cell-cycle distribution of MCF-7 cells. DMSO was used assolvent control. MCF-7 cells were cultured in phenol red-free DMEM with 5% sFBS for24 h. The medium was then changed to medium without hormone and incubation wascontinued for another 24 h with DMSO and test compounds. Cell-cycle analysis wasperformed by flow cytometry. Results were expressed as percentage of cells in G1phase, S phase and G2 phase. (A) Solvent control; (B) 1 μM ICI-182780; (C) 0.01 μMestradiol; (D) 0.01 μM estradiol+1 μM ICI-182780; (E) 0.01 μM SSd; (F) 0.01 μM SSd+1 μM ICI-182780; (G) 10 μM SSd; (H) 10 μM SSd+1 μM ICI-182780. These data wererepresentative of at least three separate experiments.
162 P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
proliferation at concentrations of 0.01 μMto 10 μMin a dose-dependentmanner (P<0.05) (Fig. 2A). The maximal proliferative effect of SSdwas achieved at 10 μM or 1 μM and was almost equivalent to the effectof 0.01 μM estradiol (the RPE was 96.7±4.0 and 96.3±5.0, respective-ly). Then, an experiment was designed to determine whether the pro-liferation stimulatory effect induced by SSd could be inhibited byICI-182780, the pure estrogen receptor antagonist. The proliferativeeffects of 0.01 μM estradiol, 0.01 μM and 10 μM SSd were blocked byaddition of 1 μM ICI-182780 (Fig. 2B). The RPE of 0.01 μM estradiol,0.01 μM and 10 μM SSd after treatment with 1 μM ICI-182780 de-creased to 69.9±3.1, 65.8±3.4 and 64.9±4.0, respectively.
3.2. Effects of SSd on cell-cycle distribution
We observed a dramatic increase in the number of cells in Sphase after treatment with 0.01 μM estradiol and 10 μM SSd for 24 h
Fig. 3. Effects of SSd and estradiol on cell-cycle distribution of MCF-7 cells. (A) Solvent contro(E) 0.01 μM SSd; (F) 0.01 μM SSd+1 μM ICI-182780; (G) 10 μM SSd; (H) 10 μM SSd+1 μM
compared with the control. The proportion of cells in S phase in-creased from 25.92% (solvent control) to 30.52% (0.01 μM estradiol)or 36.58% (10 μM SSd) (Table 1). Cell-cycle analysis revealed thatthe proliferation stimulatory effect was associated with a markedincrease in the number of MCF-7 cells in S phase, with effectssimilar to those of estradiol (Fig. 3).
3.3. ERE activation in human breast cancer cells (MCF-7)
To elucidate that the ER–ERE interaction could be induced bySSd, transient transfection of the ERE-luciferase reporter gene wasperformed in MCF-7 cells. As shown in Fig. 4A, SSd induced thetranscriptional activity of ERE-luciferase reporter gene in a dose-dependent manner. The stimulatory effect of 10 μM SSd on ERE-luciferase activity was 6.5-fold, which was similar to that of 0.01 μMestradiol. It was also found that 1 μM ICI-182780 significantly inhibitedthe stimulatory effects of 0.01 μM estradiol, 1 nM and 10 μM of SSd onERE-luciferase activity (Fig. 4B).
3.4. ERα/ERβ-mediated luciferase gene expression stimulated by SSd intransiently transfected HeLa cells
It has been shown that some phytoestrogens such as genisteinand coumestrol have differential abilities in activation of ERα andERβ (Brigitte and Johannes, 2001). In the present study, we wouldlike to find out whether SSd shows any differential activation of thetwo ER isoforms. In this study, ER-negative HeLa cells were usedand were transiently transfected with either ERα or ERβ expressionplasmid. As shown in Fig. 5, treatment of both transfected cells withvarious concentrations of estradiol and SSd as indicated for 24 hincreased ERE-luciferase activity in a dose-dependent manner. Forthe ERα transfected cells, the induction of ERE-dependent lucifer-ase activity by SSd began at 0.01 μM, and the induction of luciferaseactivity by 10 μM SSd reached 3.67-fold (Fig. 5A). For the ERβtransfected cells, the induction of ERE-dependent luciferase activity
l; (B) 1 μM ICI-182780; (C) 0.01 μM estradiol; (D) 0.01 μM estradiol+1 μM ICI-182780;ICI-182780.
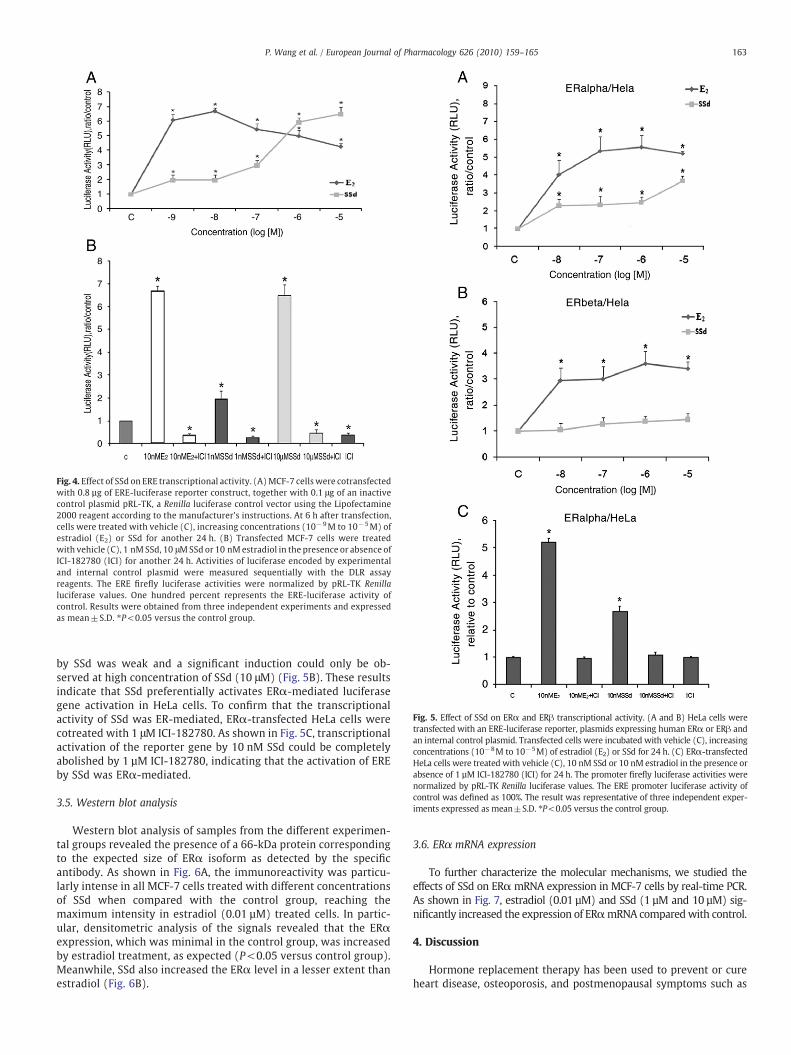
Fig. 4. Effect of SSd on ERE transcriptional activity. (A)MCF-7 cells were cotransfectedwith 0.8 µg of ERE-luciferase reporter construct, together with 0.1 μg of an inactivecontrol plasmid pRL-TK, a Renilla luciferase control vector using the Lipofectamine2000 reagent according to the manufacturer's instructions. At 6 h after transfection,cells were treated with vehicle (C), increasing concentrations (10−9M to 10−5M) ofestradiol (E2) or SSd for another 24 h. (B) Transfected MCF-7 cells were treatedwith vehicle (C), 1 nM SSd, 10 μMSSd or 10 nM estradiol in the presence or absence ofICI-182780 (ICI) for another 24 h. Activities of luciferase encoded by experimentaland internal control plasmid were measured sequentially with the DLR assayreagents. The ERE firefly luciferase activities were normalized by pRL-TK Renillaluciferase values. One hundred percent represents the ERE-luciferase activity ofcontrol. Results were obtained from three independent experiments and expressedas mean±S.D. ⁎P<0.05 versus the control group.
Fig. 5. Effect of SSd on ERα and ERβ transcriptional activity. (A and B) HeLa cells weretransfected with an ERE-luciferase reporter, plasmids expressing human ERα or ERβ andan internal control plasmid. Transfected cells were incubated with vehicle (C), increasingconcentrations (10−8M to 10−5M) of estradiol (E2) or SSd for 24 h. (C) ERα-transfectedHeLa cells were treated with vehicle (C), 10 nM SSd or 10 nM estradiol in the presence orabsence of 1 µM ICI-182780 (ICI) for 24 h. The promoter firefly luciferase activities werenormalized by pRL-TK Renilla luciferase values. The ERE promoter luciferase activity ofcontrol was defined as 100%. The result was representative of three independent exper-iments expressed as mean±S.D. ⁎P<0.05 versus the control group.
163P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
by SSd was weak and a significant induction could only be ob-served at high concentration of SSd (10 μM) (Fig. 5B). These resultsindicate that SSd preferentially activates ERα-mediated luciferasegene activation in HeLa cells. To confirm that the transcriptionalactivity of SSd was ER-mediated, ERα-transfected HeLa cells werecotreated with 1 μM ICI-182780. As shown in Fig. 5C, transcriptionalactivation of the reporter gene by 10 nM SSd could be completelyabolished by 1 µM ICI-182780, indicating that the activation of EREby SSd was ERα-mediated.
3.5. Western blot analysis
Western blot analysis of samples from the different experimen-tal groups revealed the presence of a 66-kDa protein correspondingto the expected size of ERα isoform as detected by the specificantibody. As shown in Fig. 6A, the immunoreactivity was particu-larly intense in all MCF-7 cells treated with different concentrationsof SSd when compared with the control group, reaching themaximum intensity in estradiol (0.01 µM) treated cells. In partic-ular, densitometric analysis of the signals revealed that the ERαexpression, which was minimal in the control group, was increasedby estradiol treatment, as expected (P<0.05 versus control group).Meanwhile, SSd also increased the ERα level in a lesser extent thanestradiol (Fig. 6B).
3.6. ERα mRNA expression
To further characterize the molecular mechanisms, we studied theeffects of SSd on ERαmRNA expression in MCF-7 cells by real-time PCR.As shown in Fig. 7, estradiol (0.01 µM) and SSd (1 µM and 10 µM) sig-nificantly increased the expression of ERαmRNA comparedwith control.
4. Discussion
Hormone replacement therapy has been used to prevent or cureheart disease, osteoporosis, and postmenopausal symptoms such as
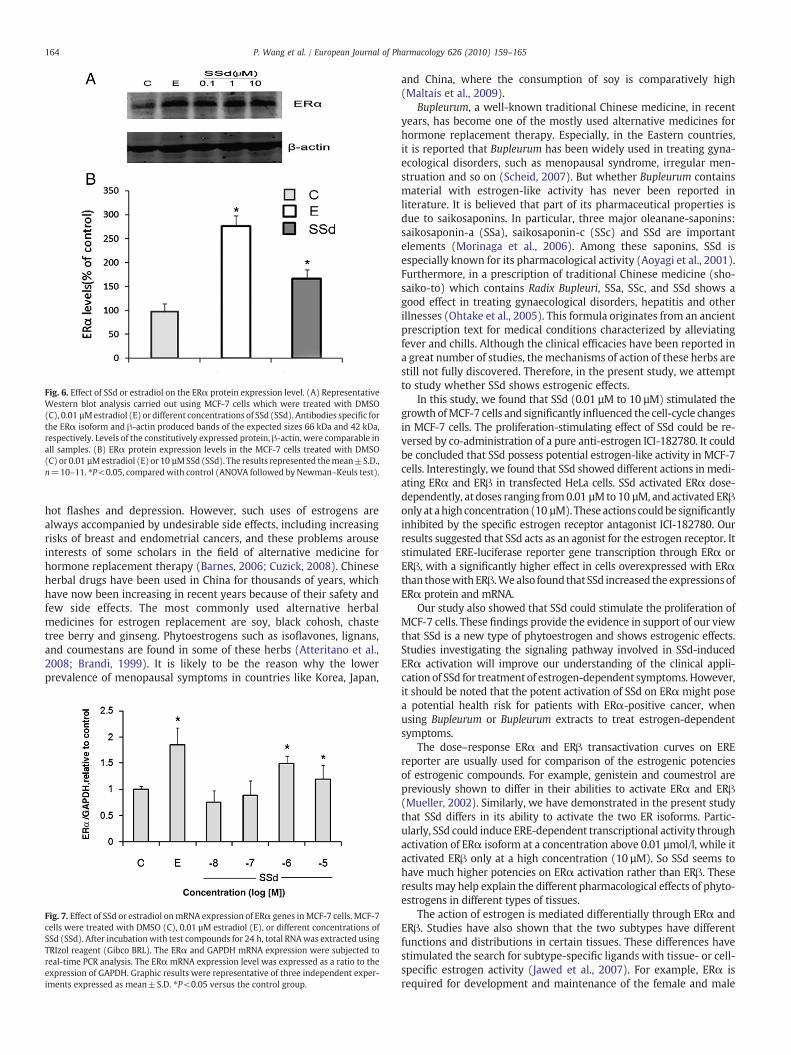
Fig. 6. Effect of SSd or estradiol on the ERα protein expression level. (A) RepresentativeWestern blot analysis carried out using MCF-7 cells which were treated with DMSO(C), 0.01 µMestradiol (E) or different concentrations of SSd (SSd). Antibodies specific forthe ERα isoform and β-actin produced bands of the expected sizes 66 kDa and 42 kDa,respectively. Levels of the constitutively expressed protein, β-actin, were comparable inall samples. (B) ERα protein expression levels in the MCF-7 cells treated with DMSO(C) or 0.01 µMestradiol (E) or 10 µMSSd (SSd). The results represented themean±S.D.,n=10–11. ⁎P<0.05, comparedwith control (ANOVA followed by Newman–Keuls test).
164 P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
hot flashes and depression. However, such uses of estrogens arealways accompanied by undesirable side effects, including increasingrisks of breast and endometrial cancers, and these problems arouseinterests of some scholars in the field of alternative medicine forhormone replacement therapy (Barnes, 2006; Cuzick, 2008). Chineseherbal drugs have been used in China for thousands of years, whichhave now been increasing in recent years because of their safety andfew side effects. The most commonly used alternative herbalmedicines for estrogen replacement are soy, black cohosh, chastetree berry and ginseng. Phytoestrogens such as isoflavones, lignans,and coumestans are found in some of these herbs (Atteritano et al.,2008; Brandi, 1999). It is likely to be the reason why the lowerprevalence of menopausal symptoms in countries like Korea, Japan,
Fig. 7. Effect of SSd or estradiol onmRNA expression of ERα genes inMCF-7 cells. MCF-7cells were treated with DMSO (C), 0.01 µM estradiol (E), or different concentrations ofSSd (SSd). After incubation with test compounds for 24 h, total RNAwas extracted usingTRIzol reagent (Gibco BRL). The ERα and GAPDH mRNA expression were subjected toreal-time PCR analysis. The ERα mRNA expression level was expressed as a ratio to theexpression of GAPDH. Graphic results were representative of three independent exper-iments expressed as mean±S.D. ⁎P<0.05 versus the control group.
and China, where the consumption of soy is comparatively high(Maltais et al., 2009).
Bupleurum, a well-known traditional Chinese medicine, in recentyears, has become one of the mostly used alternative medicines forhormone replacement therapy. Especially, in the Eastern countries,it is reported that Bupleurum has been widely used in treating gyna-ecological disorders, such as menopausal syndrome, irregular men-struation and so on (Scheid, 2007). But whether Bupleurum containsmaterial with estrogen-like activity has never been reported inliterature. It is believed that part of its pharmaceutical properties isdue to saikosaponins. In particular, three major oleanane-saponins:saikosaponin-a (SSa), saikosaponin-c (SSc) and SSd are importantelements (Morinaga et al., 2006). Among these saponins, SSd isespecially known for its pharmacological activity (Aoyagi et al., 2001).Furthermore, in a prescription of traditional Chinese medicine (sho-saiko-to) which contains Radix Bupleuri, SSa, SSc, and SSd shows agood effect in treating gynaecological disorders, hepatitis and otherillnesses (Ohtake et al., 2005). This formula originates from an ancientprescription text for medical conditions characterized by alleviatingfever and chills. Although the clinical efficacies have been reported ina great number of studies, themechanisms of action of these herbs arestill not fully discovered. Therefore, in the present study, we attemptto study whether SSd shows estrogenic effects.
In this study, we found that SSd (0.01 µM to 10 µM) stimulated thegrowth ofMCF-7 cells and significantly influenced the cell-cycle changesin MCF-7 cells. The proliferation-stimulating effect of SSd could be re-versed by co-administration of a pure anti-estrogen ICI-182780. It couldbe concluded that SSd possess potential estrogen-like activity in MCF-7cells. Interestingly, we found that SSd showed different actions in medi-ating ERα and ERβ in transfected HeLa cells. SSd activated ERα dose-dependently, at doses ranging from0.01 µM to 10 µM, andactivated ERβonly at ahighconcentration (10 µM). Theseactionscouldbe significantlyinhibited by the specific estrogen receptor antagonist ICI-182780. Ourresults suggested that SSd acts as an agonist for the estrogen receptor. Itstimulated ERE-luciferase reporter gene transcription through ERα orERβ, with a significantly higher effect in cells overexpressed with ERαthan thosewith ERβ.Wealso found that SSd increased the expressionsofERα protein and mRNA.
Our study also showed that SSd could stimulate the proliferation ofMCF-7 cells. These findings provide the evidence in support of our viewthat SSd is a new type of phytoestrogen and shows estrogenic effects.Studies investigating the signaling pathway involved in SSd-inducedERα activation will improve our understanding of the clinical appli-cation of SSd for treatment of estrogen-dependent symptoms. However,it should be noted that the potent activation of SSd on ERα might posea potential health risk for patients with ERα-positive cancer, whenusing Bupleurum or Bupleurum extracts to treat estrogen-dependentsymptoms.
The dose–response ERα and ERβ transactivation curves on EREreporter are usually used for comparison of the estrogenic potenciesof estrogenic compounds. For example, genistein and coumestrol arepreviously shown to differ in their abilities to activate ERα and ERβ(Mueller, 2002). Similarly, we have demonstrated in the present studythat SSd differs in its ability to activate the two ER isoforms. Partic-ularly, SSd could induce ERE-dependent transcriptional activity throughactivation of ERα isoform at a concentration above 0.01 µmol/l, while itactivated ERβ only at a high concentration (10 µM). So SSd seems tohave much higher potencies on ERα activation rather than ERβ. Theseresults may help explain the different pharmacological effects of phyto-estrogens in different types of tissues.
The action of estrogen is mediated differentially through ERα andERβ. Studies have also shown that the two subtypes have differentfunctions and distributions in certain tissues. These differences havestimulated the search for subtype-specific ligands with tissue- or cell-specific estrogen activity (Jawed et al., 2007). For example, ERα isrequired for development and maintenance of the female and male

165P. Wang et al. / European Journal of Pharmacology 626 (2010) 159–165
reproductive systems, bone maintenance and regulation of the car-diovascular system, and it is also significant for certain brain functions(Beck and Wade, 2008). ERα is dominant in the breast and uterus,suggesting that ERβ-selective ligands may be used as hormonereplacement therapy without increasing the risk of breast or uterinecancer (Fox et al., 2008). Our study demonstrates that SSd shows apreferential activation of ERα in mediating estrogenic effects, sug-gesting that SSd may play an important role in other ERα-dominanttissues. As far as we know, the activation of ERα pathway is only wellcharacterized in the reproductive systems, while the information inother cell types, such as bone and brain, is limited. Thus, SSd might bea good tool for understanding the role of such pathway in non-reproductive systems. In addition, it is intriguing to determine theeffects as well as molecular actions of SSd in bone and brain tissues soas to delineate the potential health benefits of SSd for treatment ofmenopausal symptoms.
In summary, our study indicates that SSd represents a new type ofphytoestrogen as it exerts estrogen-like actions via the activation ofERα-mediated pathway. This discovery provides new views forunderstanding the complex actions of Bupleurum in treatment ofmenopausal and postmenopausal disorders.
Acknowledgments
Thisworkwas supported by grants from theNational Natural ScienceFoundation of China (30472271), Shanghai Natural Science Foundation(09ZR1429700), Innovation Program of Shanghai Municipal EducationCommission (09YZ134), Shanghai Municipal Education Commission(06cz013) and Shanghai Municipal Public Health Bureau (054030). Wethank Dr. Changquan Ling, Qinghui Zhou,Yani Zhang and Binbin Chenfor their critical reading of this manuscript.
References
Atteritano,M., Pernice, F.,Mazzaferro, S.,Mantuano, S., Frisina, A., D'Anna, R., Cannata,M.L.,Bitto, A., Squadrito, F., Frisina, N., Buemi, M., 2008. Effects of phytoestrogen genisteinon cytogenetic biomarkers in postmenopausal women: 1 year randomized, placebo-controlled study. Eur. J. Pharmacol. 589, 22–26.
Aoyagi, H., Kobayashi, Y., Yamada, K., Yokoyama,M., Kusakari, K., Tanaka,H., 2001. Efficientproduction of saikosaponins in Bupleurum falcatum root fragments combined withsignal transducers. Appl. Microbiol. Biotechnol. 57, 482–488.
Barnes, P.J., 2006. Corticosteroids: the drugs to beat. Eur. J. Pharmacol. 533, 2–14.Beck, L.A.,Wade, J., 2008. Steroid receptor expression in the developing copulatory system
of the green anole lizard (Anolis carolinensis). Gen. Comp. Endocrinol. 157, 70–74.Brandi,M.L., 1999. Phytoestrogensandmenopause. Environ. Toxicol. Pharmacol. 7, 213–216.Brigitte,G., Johannes,W., 2001. Comparisonof an arrayof in vitro assays for theassessment
of the estrogenic potential of natural and synthetic estrogens, phytoestrogens andxenoestrogens. Toxicology 166, 79–89.
Cuzick, J., 2008. Hormone replacement therapy and the risk of breast cancer. Eur. J. Cancer44, 2344–2349.
Ding, W.X., Qi, X.R., Li, P., Maitani, Y., Nagai, T., 2005. Cholesteryl hemisuccinate as amembrane stabilizer in dipalmitoylphosphatidylcholine liposomes containingsaikosaponin-d. Int. J. Pharm. 300, 38–47.
Fox, E.M., Davisb, R.J., Shupnik, M.A., 2008. ERβ in breast cancer—onlooker, passive player,or active protector? Steroids 73, 1039–1051.
Hsu, Y.L., Kuo, P.L., Lin, C.C., 2004. The proliferative inhibition and apoptotic mechanism ofsaikosaponinD inhumannon-small cell lungcancerA549cells. Life Sci. 75, 1231–1242.
Ikegami, F., Fujii, Y., Ishihara, K., Satoh, T., 2003. Toxicological aspects of Kampomedicines in clinical use. Chem.-Biol. Interact. 145, 235–250.
Jawed, S., Kim, B., Ottenhof, T., Brown, G.M., Werstiuk, E.S., Niles, L.P., 2007. HumanmelatoninMT1 receptor induction by valproic acid and its effects in combinationwithmelatonin on MCF-7 breast cancer cell proliferation. Eur. J. Pharmacol. 560, 17–22.
Leung, C.Y., Liu, L., Wong, R.N.S., Zeng, Y.Y., Li, M., Zhou, H., 2005. Saikosaponin-d inhibitsT cell activation through the modulation of PKCθ, JNK, and NF-κB transcription factor.Biochem. Biophys. Res. Commun. 338, 1920–1927.
Maltais, A., Remondetto, G.E., Subirade,M., 2009. Soy protein cold-set hydrogels as controlleddelivery devices for nutraceutical compounds. Food Hydrocoll. 23, 1647–1653.
Meng, Q.S., Zhu, X.Y., Tang, X.L., Ma, B., Ni, X., 2007. Effect of isoflavones in regulatingthe transcription of target genes through estrogen receptors [in Chinese]. J. Chin.Integr. Med. 5, 577–580.
Morinaga, O., Zhu, S.H., Tanaka, H., Shoyama, Y., 2006. Visual detection of saikosaponinsby on-membrane immunoassay and estimation of traditional Chinese medicinescontaining Bupleuri radix. Biochem. Biophys. Res. Commun. 346, 687–692.
Mueller, S.O., 2002. Overview of in vitro tools to assess the estrogenic and antiestrogenicactivity of phytoestrogens. J. Chromatogr. B 777, 155–165.
Ohtake,N., Yamamoto,M., Takeda, S., Aburada,M., Ishige, A.,Watanabe, K., Inoue,M., 2005.Theherbalmedicine Sho-Saiko-To selectively inhibits CD8+T-cell proliferation. Eur. J.Pharmacol. 507, 301–310.
Scheid, V., 2007. Traditional Chinese medicine—what are we investigating? The case ofmenopause. Complement. Ther. Med. 15, 54–68.
Singhal, R., Badger, T.M., Ronis, M.J., 2009. Estrogenic status modulates the effect of soyon hepatic responses to 7, 12-dimethylbenz (a) anthracene (DMBA). Toxicol. Appl.Pharmcol. 234, 89–97.
Yang, Z.Y., Chao, Z., Huo, K.K., Xie, H., Tian, Z.P., Pan, S.L., 2007. ITS sequence analysis used formolecular identification of the Bupleurum species from northwestern China. Phytome-dicine 14, 416–423.
Zhao, Y.Y., Luo, H.S., Wang, B., Ma, L.B., Tu, G.Z., Zhang, R.Y., 1996. Triterpenoid saponinsfrom Bupleurum smithii var. parvifolium. Phytochemistry 42, 1673–1675.
Zhu, S., Shimokawa, S., Shoyama, Y., Tanaka, H., 2006. A novel analytical ELISA-basedmethodology for pharmacologically active saikosaponins. Fitoterapia 77, 100–108.