Embed Size (px)
Citation preview

CENTRO UNIVERSITÁRIO FAVENI
Genética
GUARULHOS – SP

SUMÁRIO
1 HISTÓRIA E FILOSOFIA DA GENÉTICA ............................................................. 3
2 EX OVO OMNI ...................................................................................................... 5
3 MENDELISMO: OS PRINCÍPIOS BÁSICOS DA HERANÇA ................................ 7
3.1 Monoibridismo ................................................................................................ 10
3.2 Diibridismo ...................................................................................................... 12
4 HEREDOGRAMAS ............................................................................................. 15
5 DOMINÂNCIA INCOMPLETA, CO-DOMINÂNCIA E PLEIOTROPIA ................. 22
5.1 Alelos Múltiplos ............................................................................................... 23
5.2 Variação descontínua ..................................................................................... 24
5.3 Norma de reação dos Genes .......................................................................... 25
5.4 Herança dos Grupos Sanguíneos ................................................................... 25
5.5 Determinação do tipo sanguíneo .................................................................... 27
5.6 Genética dos grupos sanguíneos ................................................................... 27
6 GRUPOS SANGUÍNEOS .................................................................................... 29
6.1 Determinação genética do grupo RH .............................................................. 29
6.2 O fator RH e a Eritroblastose Fetal ................................................................. 30
6.3 Probabilidade em genética ............................................................................. 31
7 CRUZAMENTOS DIÍBRIDOS ............................................................................. 32
8 INTERAÇÃO GÊNICA ........................................................................................ 34
9 HERANÇA POLIGÊNICA .................................................................................... 36
9.1 Genética da Cor dos Olhos ............................................................................. 38
9.2 Epistasia ......................................................................................................... 40
9.3 Ligação Gênica ............................................................................................... 43
9.4 Arranjos “cis” e “trans” .................................................................................... 44
9.5 Mapeamento Cromossômico .......................................................................... 45

9.6 Sistemas de Determinação Sexual ................................................................. 49
9.7 Herança Sexual .............................................................................................. 52
9.8 Composição do Material Genético .................................................................. 55
10 BIBLIOGRAFIA BÁSICA ..................................................................................... 58

3
1 HISTÓRIA E FILOSOFIA DA GENÉTICA
A disciplina de Genética é uma área da Biologia que tem como objetivo estudar
a herança biológica, também conhecida como hereditariedade, ou seja, a transmissão
das características dos pais para os filhos, seus descendentes, ao longo das
gerações. O termo Genética deriva do grego genno (que significa fazer nascer) e foi
utilizado pela primeira vez pelo cientista Willian Bateson em uma carta dirigida a
Sedgewick, datada de 18 de abril de 1905.Apesar de a hereditariedade despertar a
curiosidade das pessoas desde a pré-história, quando já faziam a seleção e
domesticação de animais e plantas de acordo com características que mais lhes
convinham, somente no século XX essa área da ciência se desenvolveu de maneira
mais expressiva.
Fonte: blog.ipog.edu.br
Uma viagem rápida à Grécia Antiga nos dá uma clara visão de como as
pessoas tentavam explicar as semelhanças entre pais e filhos, por exemplo, por volta
de 500 a.C., um dos discípulos de Pitágoras de Samos, chamado Alcmeon de
Crotona, acreditava que os homens e as mulheres tinham sêmen, e este se formava

4
no cérebro. Ainda segundo ele, o gênero da criança seria determinado de acordo com
o predomínio do sêmen de um dos pais e, caso tivessem a mesma proporção,
ocorreria o que chamamos de hermafroditismo. Um pouco mais adiante na história da
Grécia podemos encontrar Empédocles de Acragas (492-432 a.C.), que dizia que se
o útero materno fosse quente nasceria um menino; e, se fosse frio, nasceria uma
menina. Já Anaxágoras de Clazomene (500-428 a.C.) postulava que somente no
homem ocorria o sêmen e que este sêmen continha o protótipo de cada órgão que
formaria o futuro ser.
Além disso, Anaxágoras também postu-lou que os meninos seriam gerados do
lado direito do corpo, enquanto as meninas seriam geradas do lado esquerdo do
corpo, sendo que essa teoria ficou conhecida como “Teoria direita e esquerda”. A partir
do Renascimento, as ideias dos filósofos gregos Hipócrates e Aristóteles passaram a
exercer uma forte influência no pensamento ocidental. Hipócrates de Cos (460-370
a.C.) desenvolveu a hipótese da pangênese, segundo a qual cada parte do corpo de
um organismo vivo produziria gêmulas, as quais seriam partículas hereditárias que
migrariam para o sêmen tanto do macho quanto da fêmea e seriam passadas aos
descendentes no momento da concepção, explicando o motivo pelo qual os
descendentes apresentam semelhanças com seus genitores. Entretanto, Aristóteles
(384-322 a.C.), um século depois das ideias de Hipócrates, escreveu um tratado que
trazia novas ideias sobre a hereditariedade e o desenvolvimento dos animais. No seu
livro, De Generatione Animalium (Geração de Animais), Aristóteles distinguiu 4 tipos
de geração: 1) reprodução sexuada com cópula; 2) reprodução sexuada sem cópula;
3) brotamento e 4) abiogênese. No que se entendeu por reprodução sexuada,
Aristóteles acreditava que o indivíduo era formado por uma contribuição diferencial
dos gêneros, sendo que a fêmea forneceria a “matéria” básica que constituiria e
nutriria o novo ser que se desenvolve, ao passo que o macho forneceria a “essência”,
transmitindo-lhe a alma, a qual seria a fonte da forma e do movimento. Mas não para
por ai. Segundo o mesmo tratado, caso o novo indivíduo nascesse normal, isso
significaria que a forma paterna havia prevalecido e que o novo ser seria igual ao pai.

5
Fonte: ilmattino.it.com.br
Entretanto, se houvesse uma falha no desenvolvimento, o indivíduo seria
parecido com a mãe. Caso ocorressem mais falhas, o indivíduo estaria mais
relacionado com os avós e, sucessivamente, de ancestrais mais distantes até que,
por fim, estaria relacionado a um ser inumano. Desde a época de Aristóteles até o
final do século XIX houve pouco avanço sobre as ideias da hereditariedade. Somente
na segunda metade do século XIX ocorreu um aumento considerável no
conhecimento dos princípios básicos da reprodução dos seres vivos, o que foi
essencial para o posterior desenvolvimento do conhecimento da herança biológica.
2 EX OVO OMNI
A frase acima pode ser traduzida da seguinte maneira: todo animal se origina
de um ovo. É com essa frase, de autoria do médico inglês William Harvey (1578-1657),
que vamos começar a nossa aula a respeito das bases da hereditariedade, como
proposto ao final da aula anterior. Durante a época de Harvey, as ideias mais
difundidas e aceitas sobre a hereditariedade partiam do pressuposto da geração
espontânea. A teoria de Harvey é de fundamental importância, pois se opunha
totalmente a esta. Harvey propunha, em sua teoria, que o ovo produzido pela fêmea
deveria ser fertilizado pelo sêmen, que provinha do macho, para poder gerar um novo

6
indivíduo. Após sua fertilização, Harvey acreditava em duas possibilidades para o
desenvolvimento do ovo:
Que o material necessário para produzir um novo ser já estaria presente
dentro do ovo
Que o material necessário para constituir o novo ser seria produzido
conforme o desenvolvimento e o tempo que esse organismo seria moldado.
Um dos postos-chaves para o desenvolvimento da Genética foi a descoberta
de que um novo ser se origina da fusão de duas células, os gametas (do grego gamos,
união, casamento). Somente após essa descoberta as leis que controlam a herança
biológica foram compreendidas. O espermatozoide (do grego spermatos, semente;
zoon, animal; oide, que se origina de) foi observado pela primeira vez em 1667 por
Antonie van Leeuwnhoek. Embora tenha ocorrido um grande avanço na compreensão
dos mecanismos reprodutivos durante esses últimos séculos, ainda ficava uma dúvida
no ar: se os gametas são as estruturas físicas que unem as gerações, então eles
devem conter toda a informação hereditária para originar um novo ser vivo. De que
forma eles continham essas informações?
Essa dúvida levou os cientistas da época a voltarem seus estudos para as
células gaméticas. Para responder essa questão, os cientistas contaram com o
desenvolvimento da Teoria Celular por Henri Dutrochet (1776-1847), François Raspail
(1794-1878), Mathias Jakob Schneider (1804-1881), Theodor Schwann (1810-1882)
e Rudolph Virchow (1821-1902), dentre outros. De acordo com essa teoria, a célula é
a unidade fundamental de qualquer ser vivo. O próprio Rudolph Virchow, em 1855,
resumiu toda a teoria em uma única frase: “omnis cellula ex cellula”. Uma das
primeiras descrições dos eventos que ocorrem durante a mitose foi feita em 1873 por
Friedrich Anton Schneider (1831-1890). Em 1888, os filamentos observados na mitose
receberam o nome de cromossomos (do grego krôma, cor, e soma, corpo) pelo fato
da sua alta afinidade por determinados corantes. O responsável por esse feito foi o
biólogo alemão Wilhelm Gottfried Waldeyer (1836-1921). A partir desse ponto, a
genética experimentou um grande desenvolvimento. Ele imaginou que essas células
estavam envolvidas com o processo de reprodução e que dentro de cada célula existia
um ser em miniatura já pré-formado. Entretanto, muitos cientistas da época
contestaram a ideia de que essas células estariam envolvidas em tal processo,

7
acreditando que elas eram micróbios parasitas do sistema genital masculino. Somente
em 1841, o anatomista e fisiologista Rudolf Albert Von Kölliker (1817-1905) estudando
a estrutura microscópica dos testículos, provou que os espermatozoides não eram
parasitas do sistema genital masculino. A descoberta dos óvulos dos animais
vivíparos ocorreu na segunda metade do século XVII, pelo médico holandês Regnier
de Graaf (1641-1673), quando relacionou os folículos ovarianos (inchaços
encontrados no ovário) com elementos reprodutivos. O óvulo em si só foi descoberto
em 1828 pelo alemão Karl Ernst Von Baer (1792-1876), quando ele estudava o interior
dos folículos ovarianos. Somente no ano de 1861, o anatomista alemão Karl
Gegenbaur (1826-1903) conseguiu demonstrar, definitivamente, que o óvulo dos
animais vertebrados é uma única célula. Apesar de a descoberta dos gametas ter
ocorrido no século XVII, a consolidação da ideia da formação de um novo ser pela
fusão de duas células diferentes se deu somente na segunda metade do século XIX,
processo esse que passou a ser denominado fecundação (do latim fecundus,
produtivo, fértil) ou fertilização (do latim fertilis, produtivo, fértil).
3 MENDELISMO: OS PRINCÍPIOS BÁSICOS DA HERANÇA
A Genética em si começou a existir de maneira formal somente em 1900,
quando três biólogos, o austríaco Erich von Tschermark-Seysenegg (1871-1962), o
alemão Carl Erich Correns (1864-1933) e o holandês Hugo de Vries (1848-1935),
estudando de forma independente, chegaram juntos às mesmas explicações para a
hereditariedade. Entretanto, quando esses cientistas pesquisaram trabalhos de seus
antecessores, descobriram que suas ideias a respeito da hereditariedade não eram
originais, uma vez que o monge austríaco Gregor Johann Mendel (1822-1884) já havia
chegado aos mesmos resultados 35 anos antes. Gregor Johann Mendel nasceu em
1822 no nordeste da Morávia, em um vilarejo chamado Heinzendorf, região que na
época pertencia à Áustria e hoje pertence à República Tcheca. Em 1847, ordenou-se
padre no mosteiro agostiniano de São Tomás, na cidade de Brünn. Durante o período
do seu noviciado, Mendel teve formação básica, no qual aprendeu técnicas de
polinização artificial e Ciências Agrárias.

8
Ao terminar essa etapa, Mendel tornou-se professor substituto de uma escola,
passando a lecionar Matemática e Grego. Almejando o cargo de professor titular,
Mendel submeteu-se a exames de competência em Viena, sendo reprovado duas
vezes. Embora o curso que Mendel escolheu formalmente tenha sido física, ele
frequentou cursos como Fisiologia Vegetal, Paleontologia, Zoologia, Botânica,
Química e Matemática. Apesar de não ter sido aprovado para o cargo de professor,
os seus estudos o colocaram na presença de professores renomados, os quais
tiveram grande influência em sua vida e obra. Durante o período que passou em
Viena, Mendel se deparou com grandes questões a serem respondidas pela Biologia,
dentre elas a hereditariedade. Como sua formação básica incluía a polinização
artificial, Mendel concluiu que o caminho para a compreensão da hereditariedade seria
por meio do cruzamento entre variedades que diferissem quanto a suas
características hereditárias. Dessa maneira, Gregor Mendel deu início a suas
pesquisas, escolhendo como material para estudo a ervilha-de-cheiro Pisum sativum.
A escolha dessa espécie não foi aleatória.
Mendel se baseou em alguns pontos que o levaram a optar por essa espécie:
facilidade de cultivo; ciclo de vida curto (o que permite obter várias gerações em um
espaço curto de tempo); existência de variedades facilmente identificáveis por
características dis-tintas; facilidade de realização de polinização; obtenção de
descendência fértil no cruzamento de variedades diferentes. Para iniciar seus
experimentos, Mendel utilizou 34 variedades diferentes de ervilhas, dentre as quais
selecionou as que mais lhe convinham para seus estudos. Nesse caso, Mendel
desejava trabalhar com variedades cujas características não sofressem alterações de
uma geração para outra, o que lhe garantiria estar trabalhando com características
hereditárias e não decorrentes das variações do meio ambiente. Mendel também só
avaliava uma característica de cada vez, ou seja, se ele estava verificando a cor da
semente, as características secundárias, como tamanho e forma, eram deixadas de
lado. Além disso, era necessária a utilização somente de plantas puras em seus
cruzamentos. Para Mendel, plantas puras eram plantas que, por autofecundação,
geravam somente descendentes iguais entre si. Por exemplo, se cruzarmos plantas
puras que possuam vagem verde, todos os descendentes também terão vagens
verdes e, se cruzados entre si, continuarão a aparecer somente vagens verdes.

9
Para uma melhor identificação, Mendel denominou as plantas puras como
geração parental, ou geração P, em sua forma abreviada. Os descendentes diretos
da geração P são chamados primeira geração híbrida, ou geração F1. Essa geração
F1, quando autofecundada, dá origem a segunda geração híbrida, ou geração F2.
Gregor Mendel observou que quando cruzadas plantas puras que possuíam sementes
de cor amarela com plantas, também puras, que possuíam sementes verdes, todos
os descendentes em F1 possuíam sementes de cor amarela. De acordo com a
observação desse fato, Gregor Mendel chamou de caráter recessivo o fator que não
se manifestava em F1, nesse caso a cor verde, e de caráter dominante o caráter que
se manifestava nesse caso a cor amarela. Embora descontente com os resultados
desse primeiro cruzamento, uma vez que se esperava obter metade das plantas com
sementes de cor verde e a outra metade com sementes de cor amarela, Mendel
decidiu autofecundar F1. Para sua surpresa, em F2, as sementes de cor verde que
haviam desaparecido em F1 voltaram a se manifestar. Observe a imagem a seguir:
Como exposto anteriormente, três outros biólogos haviam chegado às mesmas
conclusões 35 anos depois de Mendel. A diferença entre os trabalhos deles e o de
Gregor Mendel foi a relação matemática estabelecida por Mendel e que fugiu aos
outros biólogos.
Na verdade, Mendel cruzou milhares de plantas. Utilizando o exemplo da cor
das sementes, Mendel cruzou plantas puras verdes com plantas puras amarelas e
obteve em F2 um total de 8023 sementes, das quais 6022 eram amarelas e 2001 eram
verdes. Ao se dividir o número de sementes amarelas pelo número de sementes

10
verdes obtém-se a proporção 3:1. Como Mendel não analisou apenas uma
característica, podemos demonstrar os resultados obtidos por ele ao considerar
sementes puras lisas e sementes puras rugosas, onde de um total de 7324 sementes,
5474 sementes eram lisas e 1850 eram rugosas. Novamente dividindo-se o valor das
sementes lisas pelo das sementes rugosas encontramos a proporção 2,96:1 ou seja,
aproximadamente 3:1.
3.1 Monoibridismo
Quando realizou autofecundação de F1 voltaram a aparecer, em F2, as plantas
com sementes verdes, na proporção 3:1 (para cada 3 sementes amarelas existe 1
verde). Nesse momento vamos avançar no sentido de como os caracteres realmente
se transmite aos descendentes, uma vez que já sabemos a respeito dos gametas.
Como ficou claro, um indivíduo é formado pela união de 2 células (os gametas) que
contém todas as instruções para a formação desse indivíduo, ou seja, tanto o macho
quanto a fêmea têm contribuição igual na formação desse ser. Como são dois os
envolvidos na formação do novo ser, devemos trabalhar com dois conjuntos de
instruções, um conjunto paterno e outro conjunto materno. A partir de agora vamos
usar letras que passarão a representar uma dada característica dos indivíduos, sendo
que cada letra representa a herança adquirida de um dos genitores. A letra maiúscula
representa o caráter que se manifesta durante as gerações no indivíduo, o mesmo
que Mendel chamou de dominante e a letra minúscula representa o caráter recessivo,
aquele que não se manifesta no indivíduo. Tomando como exemplo a letra a para uma
dada característica podemos ter as seguintes combinações nas células de um
indivíduo;
AA, Aa, aa
Como podemos observar existem 3 possíveis combinações para qualquer tipo
de característica (consideramos que Aa e aA são a mesma coisa, uma vez que a
ordem dos fatores não altera o produto final). O indivíduo AA apresentará a
característica pertencente ao gene A; o indivíduo Aa também apresentará a

11
característica pertencente ao gene A, mesmo tendo o gene a uma vez que este só irá
se manifestar se estiver em dose dupla, que ocorre com o indivíduo aa
Voltando mais uma vez ao exemplo das ervilhas, a primeira decisão a ser
tomada é a escolha da letra a ser utilizada em nosso problema. A letra escolhida, por
convenção, é a letra da característica recessiva. Dessa maneira, para resolver o
problema das ervilhas de Mendel, iremos utilizar a letra v, (pois o caráter recessivo
é verde). A letra a ser utilizada na resolução é a letra que representa a característica
recessiva. Portanto, a resolução do problema inicial de Mendel seria a seguinte: como
F1 eram linhagens puras, uma verde (vv) e outra amarela (VV), ao se cruzar os genes
temos que 100% das novas plantas terão sementes amarelas (Vv), observe:
Analisando a tabela acima, podemos ver que o único resultado possível de F1
é ser 100% Aa, uma vez que a planta 1 só pode doar o gene A e a planta 2 somente
o gene a, portanto, só podem existir plantas Aa. Quando essas plantas são
autofecundadas elas dão origem a uma prole que é 25% AA, 50% Aa e 25% aa. Isso
é feito da seguinte maneira: Em um lado (linha) colocamos os genes Aa em colunas
diferentes (em azul) e na coluna colocamos os outros genes Aa em linhas diferentes
(em vermelho). Ai virou um jogo de batalha naval! É só misturar o gene que se
encontra em uma linha com sua respectiva coluna! Fácil.

12
Em relação à proporção, temos ¼ AA que nos dá 0,25. Quando multiplicamos
esse resultado por 100 temos um valor expresso em porcentagem (a famosa regra de
3). Em relação a Aa, temos 2/4, que é a mesma coisa que ½, ou seja, meio, ou 50%.
Em relação a aa temos os mesmos ¼ e, portanto, 25%. Agora ficou claro como Mendel
explicou os resultados obtidos durante os anos de cruzamentos. Todas as plantas que
apareceram em F1 eram Aa. Assim, como o gene a não se manifesta e A sim, todas
as plantas tinham sementes amarelas. Como existe uma possibilidade de 25% de
aparecer sementes verdes em F2, elas voltam a aparecer numa proporção de 3:1.
Com isso já temos uma base para as próximas aulas. O importante é que, a partir de
agora, todo o restante depende da compreensão dessa aula. Assim, se você ficou
com alguma dúvida estude mais, busque outros livros ou peça ajuda para nossa
tutoria. Essa aula perdida significa perder também quase todo o restante do conteúdo
que envolve cruzamentos.
3.2 Diibridismo
Após um período de letrinhas e porcentagens vamos voltar rapidamente aos
conceitos introduzidos na genética durante o seu desenvolvimento. Wilhelm L.
Johannsen (1857-1927), cientista dinamarquês, introduziu os conceitos de genótipo
(do grego genos, originar, e typos, característica) e fenótipo (do grego pheno,
evidente, e typos, característica).O termo genótipo refere-se aos genes dos
indivíduos, entidades invisíveis que determinam as características, ou seja, os tipos
de alelos que esse indivíduo possui. Já o termo fenótipo refere-se à expressão desses
genes, ou seja, a maneira como eles se manifestam. Assim, o genótipo de uma pessoa
albina é aa (nós vamos discutir isso mais detalhadamente nas próximas aulas) e o

13
seu fenótipo é ter a pele, cabelos e pelos brancos, ou seja, as características visíveis
da ação do gene. Tanto genótipo quanto fenótipo são conceitos que devem estar muito
bem digeridos a partir de agora. Por exemplo: o genótipo ou o fenótipo resultam da
sua interação com o meio ambiente? Vamos considerar, inicialmente, duas pessoas,
uma loira e outra de cabelos castanhos. Elas podem tranquilamente alterar a cor do
cabelo (basta pintar; pode ser verde, laranja, azul, como está na moda nos dias de
hoje!). Quando essa pessoa pinta o cabelo ela alterou o seu fenótipo. Imagine, duas
pessoas, ambas de cabelos castanhos, pintam os cabelos de laranja. Nesse período
resolvem ter um filho e ele nasce com cabelos laranja! Impossível, você não acha?
Assim, quando se altera o fenótipo não se alteram os genes, o genótipo. O mesmo
vale para o uso de lentes de contato, aumento da massa muscular por meio de
exercícios físicos e bronzeamento da pele.
A pessoa pode ter olhos castanhos escuros (geno-tipicamente falando) e usar
uma lente de contato azul (o fenótipo), ou ter pela branca e passar um longo período
tomando sol na praia e escurecer a cor da pele pela deposição de melanina. Outro
exemplo muito bem estudado é a coloração dos pelos em coelhos da raça himalaia.
Esses coelhos têm como característica apresentar as orelhas, os focinhos e as patas
com pelos pretos e, no restante do corpo, pelos brancos. Isso ocorre somente se
esses coelhos estiverem em ambiente cuja temperatura varie entre 15°C e 24°C.
Como as extremidades do corpo tendem a ser mais frias (perdem mais calor para o
meio), se possuírem pelagem escura nessas áreas, ela absorverá mais calor (da
mesma forma quando se usa roupa escura). Se você pegar um coelho desse e criá-
lo em uma região cuja temperatura seja menor que 2°C, esse coelho passará a
apresentar pelagem totalmente escura.
O inverso também é verdadeiro. Se você criá-lo em uma região na qual a
temperatura seja maior que 29°C, a pelagem passa a ser totalmente branca. Agora
quero mais ainda sua atenção: vamos imaginar um experimento (um biólogo que não
faz experimentos não é um biólogo). Vamos pegar um desses coelhos e criá-los em
um local onde a temperatura seja, digamos, 20°C. Fazendo isso a sua pelagem será
escura nas extremidades (focinho, orelhas e patas) e a do restante do corpo será
branca. Agora vamos raspar totalmente o pelo do dorso desse animal e amarrar uma
bolsa com gelo sobre o local raspado. Será que você consegue imaginar o que
aconteceria? Se sua resposta foi: - nesse local crescerão pelos pretos, você está

14
totalmente correto. Assim, o animal terá, além das extremidades, pelos escuros no
meio do corpo, bem no local onde nós raspamos anteriormente. Agora quero lhe
propor outro problema. Suponha que você receba, para realizar experimentos,
algumas sementes amarelas (iguais às de Mendel). Entretanto, o fornecedor não lhe
informou se essas sementes eram homozigotas (plantas puras) ou heterozigotas
(híbridas). Como você pode perceber, surgiram no período anterior, dois termos que
ainda não discutimos: homozigotos e heterozigotos. O termo homozigoto refere-se ao
indivíduo que apresenta o mesmo tipo de gene em alelos diferentes. Assim, são
homozigotos os indivíduos AA, aa, BB, bb, etc. Já o termo heterozigoto faz referência
aos indivíduos que possuem genes diferentes em seus alelos, exemplificando, como
no caso anterior, como Aa e Bb (lembre-se: homo significa igual e hetero significa
diferente). Enfim, para descobrir se um indivíduo é heterozigoto ou homozigoto basta
realizar o chamado cruzamento-teste. Esse experimento consiste em cruzar o
indivíduo em questão com um indivíduo totalmente recessivo e analisar a sua
descendência. No exemplo clássico de Mendel, vamos cruzar as plantas com
sementes amarelas (as quais em F1 nós não sabíamos o genótipo) com plantas
verdes, ou seja, plantas recessivas. Ao se analisar os resultados, podemos chegar as
seguintes conclusões: entre os descendentes não apareceram indivíduos verdes ou,
entre os descendentes apareceram indivíduos verdes. Vamos analisar o primeiro
caso. Se não apareceram indivíduos verdes em momento algum, podemos concluir
que o genótipo da semente amarela só pode ser VV, observe o cruzamento abaixo:
VV x vv
100% Vv (amarelas)
Analisando o segundo caso, no qual aparecem sementes verdes, o genótipo
da semente amarela só pode ser Vv, observe o cruzamento abaixo:
Vv x vv
50% Vv (amarelas) e 50% vv (verdes)
Observe que tanto no primeiro caso quanto no segundo, as proporções
genotípicas são os genes (ex.: 50% Vv) e as proporções fenotípicas estão entre

15
parênteses, ou seja, as cores das sementes. Uma observação importante a ser feita
sobre o cruzamento-teste é que ele é válido para cruzamentos que deixam muitos
descendentes. Cruzamento que deixam poucos descendentes tem de ser repetidos
várias vezes, uma vez que, como são poucos os descendentes, a chance de se
equivocar com o resultado é grande. Se um homem que apresenta uma pigmentação
normal de pele (albino) casa-se com uma mulher albina e tem um filho normal, não
significa que esse homem seja homozigoto (AA), pois mesmo sendo heterozigoto (Aa)
possui 50% de chance de ter um filho normal. Para confirmar isso, faça você mesmo
os cruzamentos e veja os resultados. Bom, a nossa aula de hoje acaba por aqui. Na
próxima vamos aprender como se constroem árvores genealógicas.
4 HEREDOGRAMAS
Um dos problemas enfrentados pela genética é a não possibilidade, por motivos
óbvios, de se realizarem cruzamentos dirigidos entre seres humanos. Dessa forma,
conhecer as características dos familiares envolvidos em uma dada problemática
genética é de fundamental importância para a determinação dos genótipos dos
envolvidos. Assim, surgiu o que conhecemos popularmente como árvores
genealógicas, mas que em âmbito científico chamamos de heredogramas (do grego
heredium, herança). Um heredograma nada mais é do que a representação gráfica do
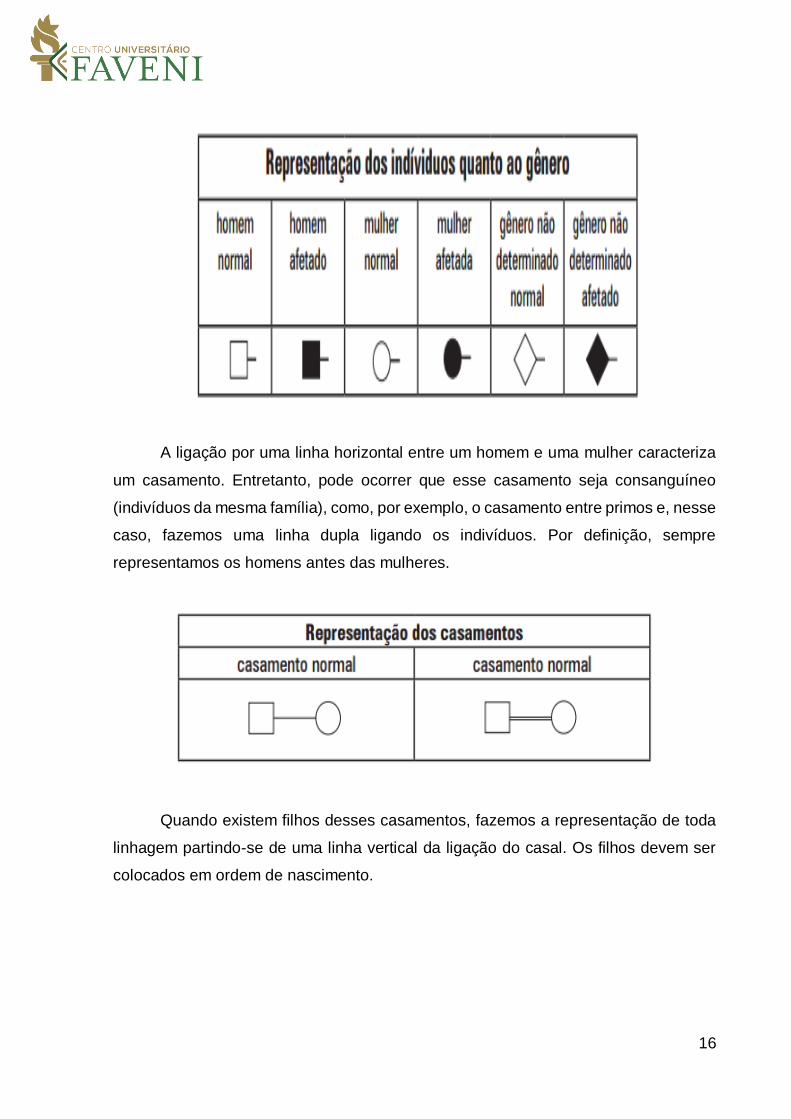
histórico de uma ou mais famílias para um dado traço genético. Os indivíduos
masculinos são representados com quadrados e os indivíduos femininos com círculos.
Veja a ilustração abaixo:
16
A ligação por uma linha horizontal entre um homem e uma mulher caracteriza
um casamento. Entretanto, pode ocorrer que esse casamento seja consanguíneo
(indivíduos da mesma família), como, por exemplo, o casamento entre primos e, nesse
caso, fazemos uma linha dupla ligando os indivíduos. Por definição, sempre
representamos os homens antes das mulheres.
Quando existem filhos desses casamentos, fazemos a representação de toda
linhagem partindo-se de uma linha vertical da ligação do casal. Os filhos devem ser
colocados em ordem de nascimento.
17
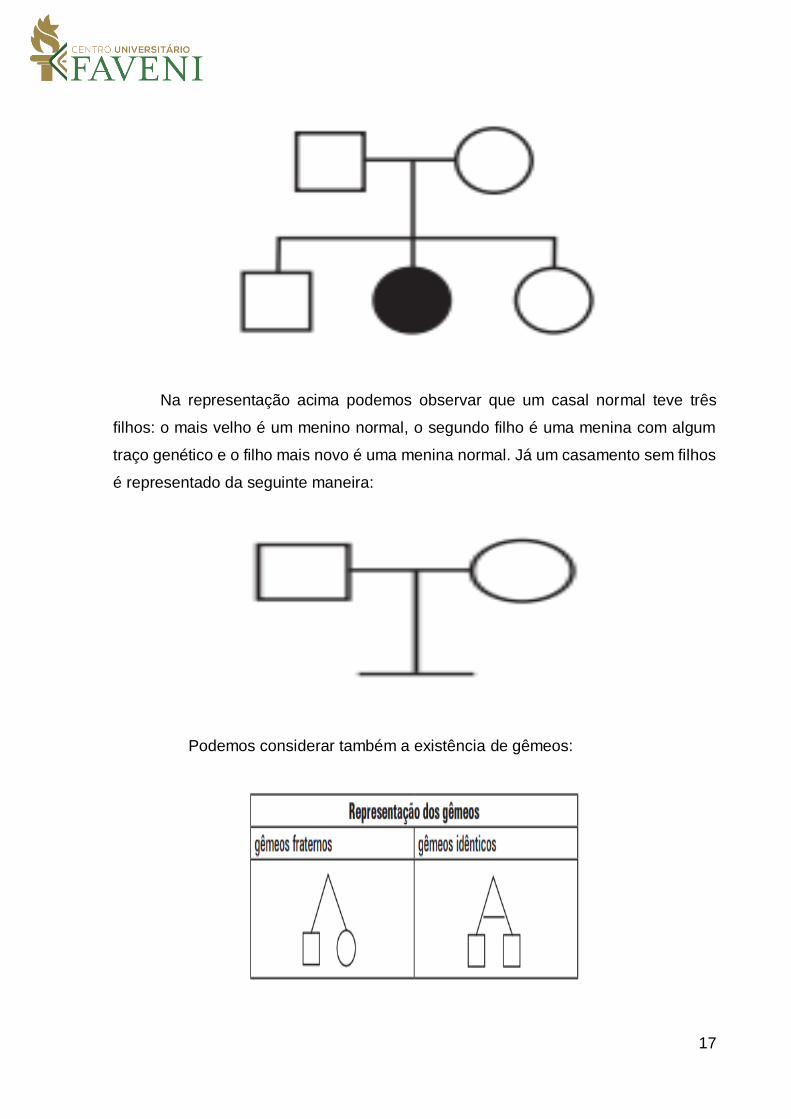
Na representação acima podemos observar que um casal normal teve três
filhos: o mais velho é um menino normal, o segundo filho é uma menina com algum
traço genético e o filho mais novo é uma menina normal. Já um casamento sem filhos
é representado da seguinte maneira:
Podemos considerar também a existência de gêmeos:

18
Quando falamos em casamento temos de levar em consideração que a mulher
pode estar grávida. Nesse caso, usaremos as representações abaixo:
Para representar um indivíduo já falecido, usamos o seguinte símbolo:
Se uma mulher for portadora de um caráter ligado ao cromossomo X, iremos
representá-la da seguinte maneira:
Para a localização dos indivíduos em um heredograma, foi adotada por
convenção, a utilização dos números romanos para indicar gerações e números
arábicos para indicar os indivíduos em uma geração. Veja:
19
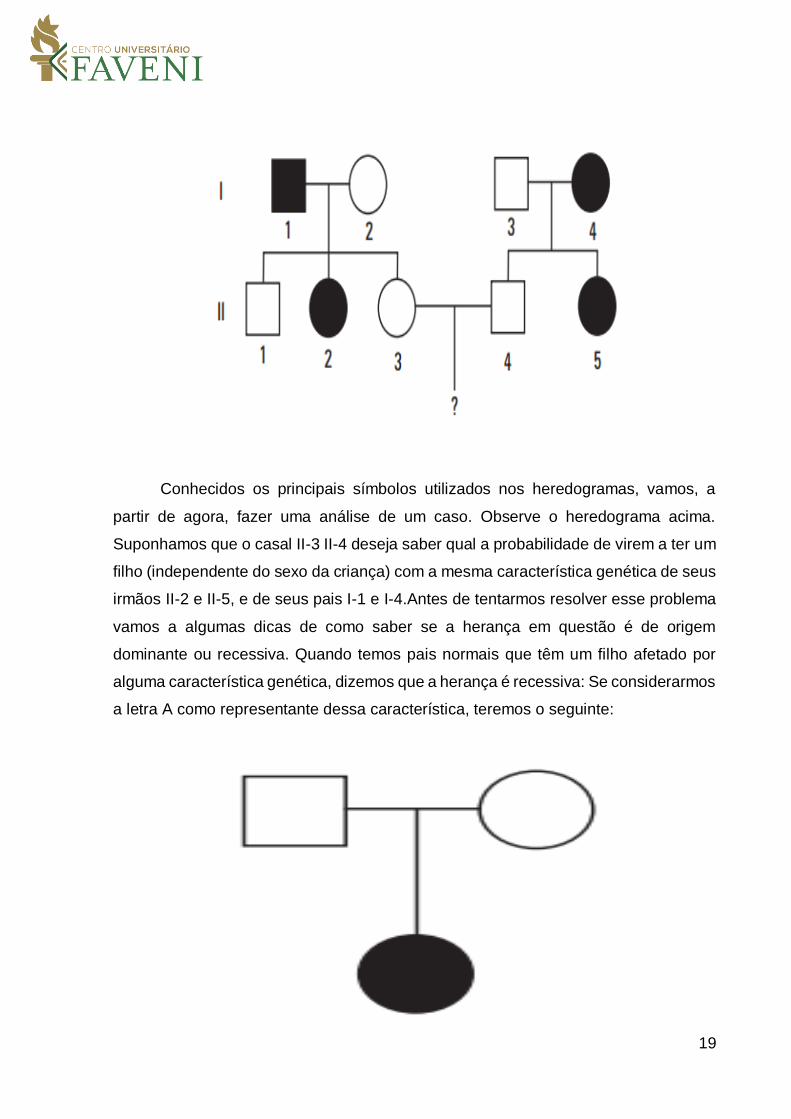
Conhecidos os principais símbolos utilizados nos heredogramas, vamos, a
partir de agora, fazer uma análise de um caso. Observe o heredograma acima.
Suponhamos que o casal II-3 II-4 deseja saber qual a probabilidade de virem a ter um
filho (independente do sexo da criança) com a mesma característica genética de seus
irmãos II-2 e II-5, e de seus pais I-1 e I-4.Antes de tentarmos resolver esse problema
vamos a algumas dicas de como saber se a herança em questão é de origem
dominante ou recessiva. Quando temos pais normais que têm um filho afetado por
alguma característica genética, dizemos que a herança é recessiva: Se considerarmos
a letra A como representante dessa característica, teremos o seguinte:
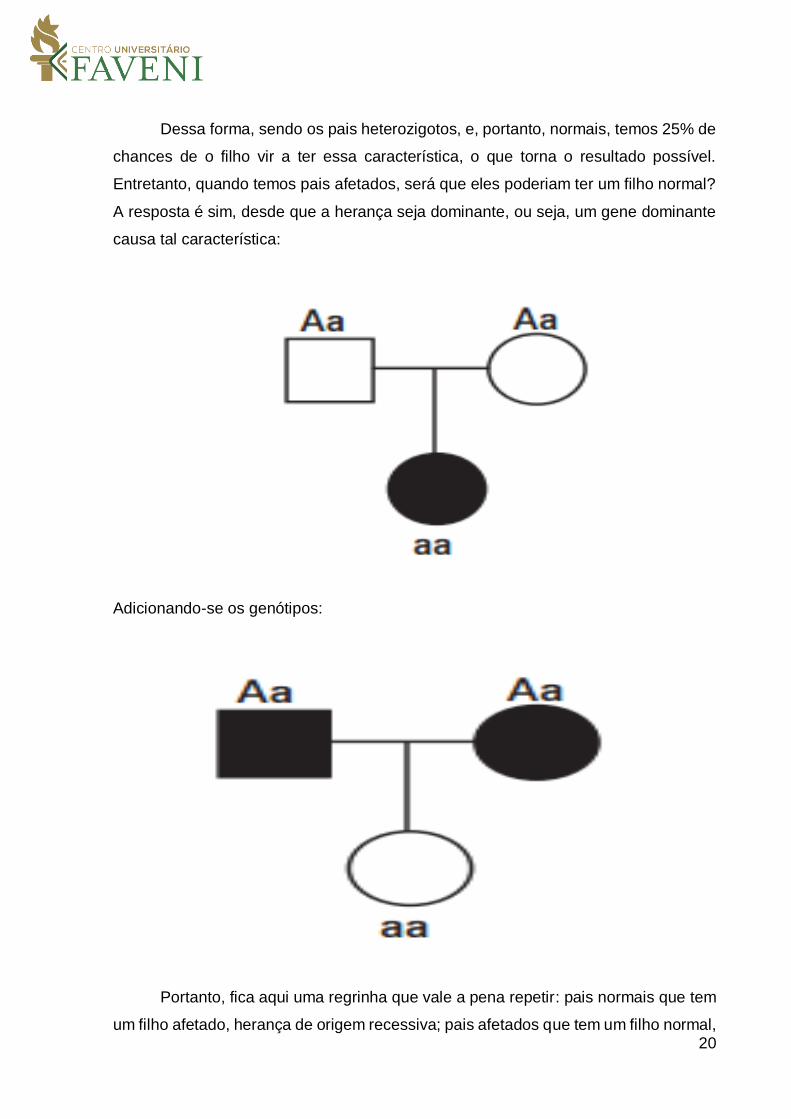
20
Dessa forma, sendo os pais heterozigotos, e, portanto, normais, temos 25% de
chances de o filho vir a ter essa característica, o que torna o resultado possível.
Entretanto, quando temos pais afetados, será que eles poderiam ter um filho normal?
A resposta é sim, desde que a herança seja dominante, ou seja, um gene dominante
causa tal característica:
Adicionando-se os genótipos:
Portanto, fica aqui uma regrinha que vale a pena repetir: pais normais que tem
um filho afetado, herança de origem recessiva; pais afetados que tem um filho normal,

21
herança de origem dominante. Retornando ao problema proposto inicialmente,
podemos verificar que a herança tem caráter recessivo. Assim, podemos marcar todos
os indivíduos afetados com genes recessivos (aqui usaremos a letra t, só para sair da
rotina um pouco!).Como os genes tt nos indivíduos II-2 e II-5 têm de ter vindos um da
mãe e outro do pai pode concluir que I-2 tem um gene t e, sendo normal, deve ter um
gene T. Usando o mesmo raciocínio vale para o restante do problema e chegamos ao
resultado final:
Infelizmente não existe milagre nesse caso. Embora possa ter parecido um
pouco confuso, a resolução é simples, mas somente se tornará simples quando você
tentar resolver por si só. Embora o exemplo acima tenha possibilitado a determinação
de todos os genótipos nem sempre isso vai acontecer. Algumas vezes você não terá
dados suficientes para saber se um indivíduo é TT ou Tt. Nesse caso iremos deixar
indicado como T_. Na próxima aula estudaremos os conceitos de dominância
incompleta, codominância e pleiotropia. Espero que você já esteja craque nesse tipo
de cruzamento que acabamos de fazer.

22
5 DOMINÂNCIA INCOMPLETA, CO-DOMINÂNCIA E PLEIOTROPIA
Na aula de hoje estudaremos os conceitos de Dominância Incompleta,
Codominância e Pleiotropia. As formas de cruzamento seguem o mesmo padrão das
últimas aulas, só apresentando algumas diferenças na análise dos seus resultados.
Ao se cruzar plantas boca-de-leão, uma vermelha e outra branca, esperar-se-ia que
os resultados fossem parecidos com os obtidos por Mendel: a característica
dominante se manifestando em F1 e a característica recessiva somente voltando a se
manifestar em F2. Entretanto, ao se cruzar essas duas plantas, observamos a
presença de um fenótipo intermediário: plantas de cor rosa. O cruzamento pode ser
visto abaixo:
P: VV x BB
F1: 100% VB (rosa)
F2: 25% VV (vermelha), 50% VB (rosa) e 25% BB (branca)
A esse fenômeno de característica intermediária damos o nome de Dominância
incompleta. Esse fenômeno não ocorre somente em plantas. Por exemplo, quando
cruzamos galinhas pretas puras com galinhas brancas puras da raça Andaluza
obtemos indivíduos de coloração intermediária (cinza). O cruzamento nesse caso é o
seguinte
P: PBPB (brancas) x PPPP (pretas)
F1: 100% PBPP (cinzas)
F2: 25% PBPB (brancas), 50% PBPP (cinzas) e 25% PPPP (pretas)
A questão agora é a seguinte: como explicar logicamente esse tipo de ação
gênica? Estudos atuais têm demonstrado que na dominância incompleta o fenótipo
depende da quantidade de proteínas expressas por aquele gene, ou seja, pela
quantidade do produto do gene. A codominância é um fenômeno parecido, mas não
igual, à dominância incompleta. Nesse caso não há uma mistura que resulta em um
produto intermediário, mas há a expressão simultânea dos genes. Um caso claro é o
sistema MN do sangue humano (iremos estudar a genética dos grupos sanguíneos
nas próximas aulas). Resumidamente, os indivíduos podem ser do grupo M (AgMAgM),
do grupo N (AgNAgN) ou do grupo MN (AgMAgN). O grupo MN é a expressão dos dois

23
produtos gênicos ao mesmo tempo. Outro exemplo de codominância pode ser
encontrado em pessoas portadoras da Síndrome de Marfan. As pessoas afetadas por
essa síndrome apresentam um alelo responsável pela aracnodactilia, que se
caracteriza por dedos anormalmente longos, curvos e finos.
Esse termo (aracnodactilia) é assim usado, pois os dedos da pessoa afetada
se parecem com patas de aranhas. Essa característica vem acompanhada de outras,
como defeitos nos olhos, coração e pulmões e anomalias ósseas. Na verdade, um
gene não atua sozinho determinando uma característica. Hoje sabemos que mesmo
as características mais simples são determinadas pela atuação de vários genes.
Existe o fenômeno, portanto, de que um gene pode influenciar, ou condicionar, mais
de uma característica. A esse fenômeno chamamos Pleiotropia. Ainda em Mendel
podemos encontrar um bom exemplo de pleiotropia: o mesmo gene que condiciona a
cor da semente também condiciona a cor da flor e da presença de uma mancha roxa
nas folhas. Assim, dizemos que o gene da cor da semente da ervilha é pleiotrópico.
Outro exemplo de pleiotropia são os alelos letais. Se apenas um alelo for suficiente
para causar a morte falamos em letal dominante; se forem necessários dois alelos
para que ocorra a morte do indivíduo falamos em letal recessivo. A acondroplasia é
uma forma de nanismo humano que é condicionada por um alelo D, o qual prejudica
o crescimento dos ossos. As pessoas normais são dd, enquanto as pessoas afetadas
são Dd. Como nunca se encontraram pessoas DD, os cientistas concluíram que
quando o gene D está em dose dupla (homozigose) ele tem um efeito muito severo e
o portador é levado à morte. Portanto, na acondroplasia, o gene D é letal dominante.
Com isso concluímos a aula de hoje. Na próxima aula vamos começar a estudar os
Alelos Múltiplos, os quais estão envolvidos na determinação da cor dos olhos, da pele,
do tamanho e peso de certos organismos.
5.1 Alelos Múltiplos
Podemos então definir como indivíduos diploides (2n) aqueles que têm esse
conjunto duplo, e indivíduos haploides (n) aqueles que têm apenas um conjunto de
cromossomos. O mesmo vale para classificação das células: uma célula haploide, por
exemplo, seria um gameta humano e uma diploide seria, por exemplo, um neurônio.
Embora tenhamos visto até o momento apenas um alelo em cada cromossomo,

24
existem algumas características no qual se encontram três ou mais alelos diferentes
na população, como por exemplo, os alelos do grupo sanguíneo, onde podemos
encontrar os fenótipos A, B, AB e O. Nesses casos, portanto, estamos falando
em Alelos Múltiplos. Vamos começar com um exemplo muito conhecido: a pelagem
dos coelhos. Em coelhos, o gene que determina a cor da pelagem apresenta-se sobre
4 formas alélicas diferentes: C (determina a pelagem castanho-acinzentada, também
conhecida como selvagem ou aguti), cch (determina a pelagem cinzenta-prateada,
também conhecida como chinchila), ch (determina uma pelagem branca com
extremidades escuras, também conhecida como himalaia) e c (determina a pelagem
totalmente branca, também conhecida como albina). Existe uma relação de
dominância entre esses genes, sendo que o gene C domina todos os outros, o gene
cch domina o ch e c, o gene ch domina apenas o c e o c não domina ninguém, apenas
se expressando em homozigose.
C > cch > ch > c
Vamos a um exemplo prático: um coelho selvagem, heterozigoto para a
himalaia, é cruzado com uma coelha chinchila, heterozigota para albino.
Quais são os possíveis descendentes desse cruzamento?
Genótipo do macho: Cch
Genótipo da fêmea: cchc
P: Cch x cchc
F1: Ccch (selvagem), Cc (selvagem), cchch (chinchila), chc (himalaia)
Proporção fenotípica: 50% selvagem, 25% chinchila, 25% himalaia
5.2 Variação descontínua
Vimos até agora que os diferentes genótipos são bem distintos, produzindo
proporções genotípicas e fenotípicas estritamente como o esperado pela lei de Mendel
(lei da segregação). Em alguns casos vamos perceber que diversos genótipos podem
corresponder a fenótipos alternativos, um bem diferente do outro. Quando isso
ocorrer, falaremos então de variação descontínua.

25
5.3 Norma de reação dos Genes
Devido ao fato da variação descontínua ser pouco frequente na natureza, o
mais comum é um mesmo genótipo produzir uma gama variada de fenótipos. A essa
variação chamamos de norma de reação. Aqui podemos usar como exemplo o feijão
carioca, que pode ser obtido em qualquer supermercado. Esse tipo de feijão possui
variegações em sua casca, com listras escuras e que não seguem um padrão e,
portanto, são irregulares. Essa característica é determinada pelo alelo dominante L.
O seu alelo recessivo l determina um feijão totalmente pigmentado, não apresentando
variegações, passando a ser chamado de feijão mulatinho. Se você possuir em sua
casa um pacote de feijão carioca você observará que a maioria dos grãos é variegada,
sendo que aproximadamente 5% dos grãos são totalmente pigmentados, parecendo-
se com o feijão mulatinho. Como o feijão carioca é homozigoto (LL), esses 5% não
expressaram o fenótipo correspondente. A explicação para esse fato é que o alelo L
tem uma norma de reação tão grande que alguns indivíduos LL não exibem esse
fenótipo. O caso do feijão-carioca nos dá a ideia de penetrância gênica, ou seja,
mesmo sendo LL, 5% dos feijões não expressaram o fenótipo apropriado, em outras
palavras, o gene L tem uma penetrância de 95%, que é calculado subtraindo-se esses
5% dos 100% esperados inicialmente. Vamos exemplificar a penetrância gênica com
um caso em humanos. A presença de 1 dedo extra na mão ou no pé, ao lado do quinto
dedo, é conhecida como polidactilia postaxial (do grego polys, muitos; daktylos,
dedos). A existência desse dedo extra é determinada pela presença de um alelo
dominante com dominância incompleta. Na África foi realizado um estudo que estimou
que a penetrância desse alelo é de 64,9%, ou seja, 35,1% (100– 64,9) da população
que tenha esse alelo não apresentará o dedo extra.
5.4 Herança dos Grupos Sanguíneos
Karl Landsteiner (1868-1943), médico austríaco que se naturalizou norte-
americano, identificou, no início do século XX, uma incompatibilidade sanguínea entre
determinadas pessoas. Ele observou o fato de que quando o sangue de duas pessoas
era misturado, vez ou outra, as células conhecidas como hemácias (eritrócitos)
formavam aglomerados, que ele chamou de aglutinações. A classificação dos tipos

26
sanguíneos como conhecemos hoje, data de 1902, quando Landsteiner e seus
colaboradores conseguiram classificar o sangue. Embora inicialmente eles tenham
classificado os tipos sanguíneos em A, B, AB e 0 (zero), durante o decorrer dos anos
o fenótipo 0 (zero) passou a ser chamado de O (ó), uma clara referência a se
classificar os tipos sanguíneos apenas com letras. A descoberta da incompatibilidade
entre os grupos sanguíneos foi de grande importância para a medicina, pois
possibilitou a transfusão de sangue somente entre pessoas compatíveis, evitando
assim a aglutinação do sangue que poderia levar ao entupimento de um vaso
sanguíneo. Por seus trabalhos sobre grupos sanguíneos da espécie humana,
Landsteiner recebeu o Prêmio Nobel de Medicina e Fisiologia no ano de 1930.A
incompatibilidade entre os grupos sanguíneos se dá pela reação de anticorpos
(aglutininas) presentes no plasma sanguíneo com substâncias presentes na
membrana plasmática das hemácias (aglutinogênios). Assim, pessoas do tipo
sanguíneo A tem aglutininas anti-B; pessoas do grupo sanguíneo B tem aglutininas
anti-A; pessoas do grupo sanguíneo AB não tem nenhuma aglutinina, pois se as
tivesse aglutinaria o seu próprio sangue, uma vez que possui os dois aglutinogênios;
pessoas do grupo sanguíneo tipo O possuem os dois tipos de aglutininas (anti-A e
anti-B). Veja a tabela abaixo:
Assim, classificamos o tipo sanguíneo AB como receptor universal, pois não
tem nenhum anticorpo contra outros grupos sanguíneos e como doador universal o
tipo sanguíneo O, uma vez que as suas hemácias não possuem nenhum
aglutinogênio e, assim, não são reconhecidas pelos anticorpos do receptor.
Entretanto, fica uma dúvida: se uma pessoa recebe sangue do tipo O, este contém

27
além das hemácias, o seu plasma, no qual se encontram os anticorpos anti-A e anti-
B. Esses anticorpos não farão o sangue do receptor se aglutinar do mesmo jeito? A
resposta é não, e se explica da seguinte forma: a quantidade de plasma que o paciente
recebe é muito pequena quando comparada com a quantidade total de sangue que
possui. Assim, as aglutininas do doador se diluem no plasma do receptor causando
pouca ou nenhuma aglutinação das suas células.
5.5 Determinação do tipo sanguíneo
A determinação dos grupos sanguíneos, procedimento rotineiro em qualquer
posto de saúde, é tecnicamente simples de ser realizado. Para realizar esse
procedimento você irá precisar de apenas uma lâmina de vidro (igual àquelas usadas
em microscopia), aglutinina anti-A e anti-B e duas gotinhas de sangue. Na lâmina
pingam-se as duas gotas de sangue, uma separada da outra e sobre a primeira
coloca-se uma gota de aglutinina anti-A e sobre a outra gota de sangue coloca-se uma
gota de aglutinina anti-B. A definição do grupo sanguíneo da pessoa é feita pela
análise macroscópica dos resultados: se somente a gota de sangue na qual foi jogada
aglutinina anti-A aglutinar, o sangue é do tipo A. Se somente a gota de sangue na qual
foi jogado aglutinina anti-B aglutinar, o sangue é do tipo B. Se ambas as gotas
aglutinares, o sangue é do tipo AB e, se nenhuma das gotas de sangue aglutinarem,
o sangue é do tipo O.
5.6 Genética dos grupos sanguíneos
Como comentamos na aula passada, os grupos sanguíneos são determinados
por alelos múltiplos. Na espécie humana existem 3 genes que determinam os 4 grupos
sanguíneos descritos acima: os alelos IA, IB e i. O alelo IA determina a presença do
aglutinogênio A nas hemácias. O alelo IB determina a presença do aglutinogênio B e
o alelo recessivo i não determina a presença de nenhum aglutinogênio. Observe a
tabela abaixo para cada tipo de genótipo com seu respectivo fenótipo:

28
Atualmente sabemos que a determinação dos grupos sanguíneos por esses 3
alelos ocorre da seguinte maneira. O alelo IA sintetiza uma enzima que transforma o
precursor H no aglutinogênio A; o alelo IB sintetiza uma outra enzima que transforma
o precursor H no aglutinogênio B. Já o alelo i é inativo, não sintetizando nenhuma
enzima e, portanto, não determinando a transformação de nenhum precursor. Como
vínhamos comentando, um gene não atua sozinho na determinação de um dado traço
e com os grupos sanguíneos não poderia ser diferente. A própria síntese do precursor
H depende de outro gene, o qual possui um alelo recessivo que não é funcional.
Assim, indivíduos hh, mesmo possuindo o gene IA, IB ou ambos, não irá produzir o
precursor e, portanto, apresentará o fenótipo do grupo O. Esses indivíduos
classificados erroneamente como pertencentes ao grupo O foram inicialmente
descritos em Bombain, na Índia e o fenômeno passou a ser chamado de fenótipo de
Bombain. Alguns anos antes de receber o Prêmio Nobel, Landsteiner e seus
colaboradores passaram a testar a interação entre o sangue humano e o de coelhos.
Eles injetaram sangue humano nos coelhos para testar a capacidade dos anticorpos
dos coelhos aglutinar as hemácias humanas. Com esse experimento eles
conseguiram identificar outro sistema de grupo sanguíneo humano, o sistema MN.
Como já foi estudado anteriormente, embora de forma muito generalista, verificamos
a existência de indivíduos com genótipo AgMAgM (grupo M), AgNAgN (grupo N) e
AgMAgN (grupo MN). Uma diferença fundamental existe entre o sistema MN e o
sistema ABO: no sistema MN não existe a presença de aglutininas contra os antígenos

29
M e N e, portanto, não há incompatibilidade entre indivíduos pertencentes a grupos
diferentes.
6 GRUPOS SANGUÍNEOS
Landsteiner e seus colaboradores, na década de 1940, injetaram hemácias de
macacos da espécie Macaca Rhesus, hoje conhecidos como macaca mullata, em
coelhos. Nesse experimento eles descobriram um anticorpo contra as hemácias, os
quais foram chamados de fator anti-Rh (abreviatura de anti-rhesus). Ao verificar a
ação desse anticorpo em humanos, eles descobriram que em 85% das pessoas as
hemácias aglutinavam. As pessoas em que as hemácias aglutinavam foram
chamadas de Rh positivas e as pessoas em que as hemácias não aglutinavam foram
chamadas Rh negativas, o que indica a ausência desse fator na membrana das
hemácias. Para saber se uma pessoa possui fator Rh positivo ou negativo utilizamos
o mesmo método da tipagem sanguínea, entretanto, trocamos o anticorpo anti-A e
anti-B por um anticorpo anti-Rh. Se as hemácias aglutinarem, o Rh é positivo, se não
aglutinarem, o Rh é negativo.
6.1 Determinação genética do grupo RH
O fator Rh é determinado por um alelo dominante R e seu alelo recessivo r não
determina sua expressão. Assim, indivíduos RR ou Rr (para facilitar vamos passar a
usar um traço: R_) são Rh positivos e somente os indivíduos rr são Rh negativos.

30
6.2 O fator RH e a Eritroblastose Fetal
Assim como no sistema MN, o fator Rh não possui anticorpos naturalmente
produzidos pelo corpo, sendo somente produzidos pela sensibilização da pessoa.
Essa sensibilização se dá pelo contato com os antígenos, os quais estimulam o
sistema imune a produzir anticorpos contra eles. Na eritroblastose fetal, também
conhecida como Doença Hemolítica do Recém-Nascido, ou simplesmente DHRN, há
a destruição das hemácias do bebê por anticorpos produzidos pela mãe e passados
ao recém-nascido no momento do parto. O termo correto para “destruição das
hemácias” é hemólise (do grego haimos, sangue, lise, quebra, destruição).
Fonte: worldpdfdatabase.us.com.br
Essa doença só existirá no caso em que a mãe for Rh- e o bebê Rh+. Na
primeira gestação, durante o momento do parto, as hemácias Rh+ do bebê entram em
contato com o sangue materno, misturando-se com ele. Essas hemácias Rh+ irão
estimular a produção de anticorpos anti-Rh, deixando, assim, a mãe sensibilizada
contra esses antígenos. Se, em um eventual segundo filho, esse também for Rh+, os
anticorpos produzidos pela mãe durante a primeira gestação passarão para o filho no
momento do parto, causando a destruição das hemácias do bebê, o que causa anemia
profunda e icterícia.

31
A icterícia (amarelamento da pele) é causada pelo acúmulo de bilirrubina no
sangue, sendo que a bilirrubina é produzida a partir da hemoglobina no fígado do
recém-nascido. Dessa forma, toda hemoglobina liberada pela destruição das
hemácias é convertida em bilirrubina, tornando a pele do bebê amarelada. Para
compensar a falta de hemácias no sangue devido à sua destruição, a medula óssea
então passa a liberar hemácias imaturas conhecidas como eritoblastos. Um quadro
clínico de icterícia, anemia e eritroblastos circulantes é um forte indicativo de Doença
Hemolítica do Recém-nascido. Mas, enfim, todo bebê Rh+, filho de mãe Rh- terá
DHRN? A resposta é não. Hoje em dia, a mulher Rh- que vai dar à luz a um bebê
Rh+ recebe, no momento do parto, uma injeção de anticorpos anti-Rh que destrói as
hemácias Rh+ que se misturaram com o seu sangue, evitando, assim, a formação de
anticorpos anti-Rh.
6.3 Probabilidade em genética
Vamos neste momento lembrar um pouquinho das regras de probabilidade já
discutidas durante o Ensino Médio. Suponhamos que se jogue uma moeda para cima.
Qual seria a probabilidade de cair cara? Obviamente a resposta é que a chance é de
50% (ou irá cair cara ou coroa). Jogando-se um dado, qual seria a probabilidade de
cair o número 3? Nesse caso, a chance é de 1 em 6 (lembre-se que o dado tem 6
faces), ou seja, 1/6. Então, qual seria a chance de, jogando-se o dado, se obter um
número par? Bom, os números pares encontrados no dado são 2, 4 e 6, ou seja,
existem 3 chances em 6, o que, simplificando, dá um valor igual a 2½ ou 50%.Em
genética, há a necessidade de se saber qual a chance de se obter dois ou mais
eventos simultaneamente, como por exemplo, qual a chance de uma criança ser loira
e de olhos azuis. Nesse caso vamos conhecer duas regrinhas básicas em
probabilidade: a regra do e e a do ou. Quando queremos saber a probabilidade de
dois eventos ocorrerem simultaneamente, como por exemplo, a chance de uma
criança ter olhos azuis e ser loira, usamos a multiplicação da probabilidade de ocorrer
cada evento. Assim, se a chance de uma criança ter olhos azuis é de ¼ e a chance
de ter cabelos loiros é de ½, temos que:½. ¼ = 1/8 assim, dividindo-se 1 por 8 temos
0,125. Multiplicando-se o resultado por 100 obtemos 12,5%.

32
Quando queremos saber a probabilidade no qual aconteça um evento ou outro
iremos somar as probabilidades. Qual seria então, a chance de a mesma criança ter
olhos azuis ou cabelos loiros? Somando-se as probabilidades, temos:½ + ¼ =
¾assim, dividindo-se 3 por 4 temos 0,75. Multiplicando-se esse resultado por 100
chegamos ao valor de 75%.
7 CRUZAMENTOS DIÍBRIDOS
Quais são as proporções genotípicas e fenotípicas obtidas do cruzamento de
dois duplo-heterozigotos CcRr, onde o gene C caracteriza a pessoa destra e o
gene c a canhota, e o gene R ter Rh+ e o gene r Rh-. Resolvendo

33
Como você percebeu após a geração P nós colocamos a letra G (gametas),
cujo resultado foi obtido cruzando-se os genes C e c com R e r, ou seja, aplicando-se
uma primeira lei de Mendel em P. Em seguida colocamos os resultados em uma
tabela, como se fosse um jogo de batalha naval, e cruzamos os resultados das linhas
com as colunas, como demonstrado acima.
Podemos definir que sempre que tivermos o cruzamento entre dois duplo-
heterozigotos (também chamados diíbridos) teremos a proporção 9:3:3:1. Em outros
cruzamentos diferentes, o número dos tipos de gametas que podem ser produzidos é
menor, o que reduz também o tamanho do quadro mostrado acima, dando menos
trabalho de ser feito. Existem casos em que o quadro pode ser resumido em apenas
uma linha, como por exemplo, o cruzamento entre um indivíduo AaBb com outro aabb.
Observe

34
Como vimos, estes tipos de cruzamentos parecem, à primeira vista, de difícil
resolução, mas com a prática isso se torna mais simples e lógico. Na próxima aula
estudaremos a respeito da Interação Gênica, mas, para tanto, precisamos ter os
conceitos apresentados nessa aula muito bem definidos em nossas cabeças.
8 INTERAÇÃO GÊNICA
Relembrando só um pouquinho o que vimos sobre as leis de Mendel, quando
temos um único par de genes que determina o caráter em questão, nós tínhamos a
proporção 3:1, no qual o número 3 indica a possibilidade de existirem indivíduos com
o gene dominante e o número 1 indica a possibilidade de termos indivíduos com o
gene recessivo. Quando analisamos duas características ao mesmo tempo,
chegamos à proporção 9:3:3:1, como ocorreu na aula passada. Já havíamos
comentado também que os genes agem em conjunto na determinação de várias
características. Assim, passamos a denominar Interação Gênica essa interação entre
diferentes genes que resultarão em diferentes fenótipos.
Muitas características encontradas nos seres vivos são decorrentes desses
tipos de interações. Por exemplo, a pigmentação dos olhos da mosca-da-fruta
(Drosophila melanogater). Nessas moscas, mais de 100 genes atuam em conjunto
para determinar a coloração dos olhos. Vamos agora a um exemplo quase que
cotidiano para muitas pessoas: a coloração da plumagem dos periquitos. Esses
periquitos, também chamados de periquitos australianos, possuem uma grande
diversidade de cores, as quais são determinadas por dezenas de genes. As cores
básicas desses periquitos verde, amarelo, azul e branco, entretanto, são
determinadas pela ação conjunta de apenas dois alelos, que chamaremos de A/a e
B/b, os quais se segregam de forma independente. O alelo A condiciona a produção
de uma proteína chamada melanina, um pigmento escuro que, quando presentes nas
penas, caracteriza a cor azul. Essa coloração se dá graças a um fenômeno óptico
chamado de “dispersão Rayleigh”. Assim, a luz, ao sofrer a dispersão nas camadas
superficiais da pena contra um fundo escuro composto de melanina no centro da pena,
aparece como azul. O alelo recessivo do gene A o gene a, não produz melanina, ou
seja, é uma forma alterada do gene. Do outro lado está o gene B. Esse gene, por sua
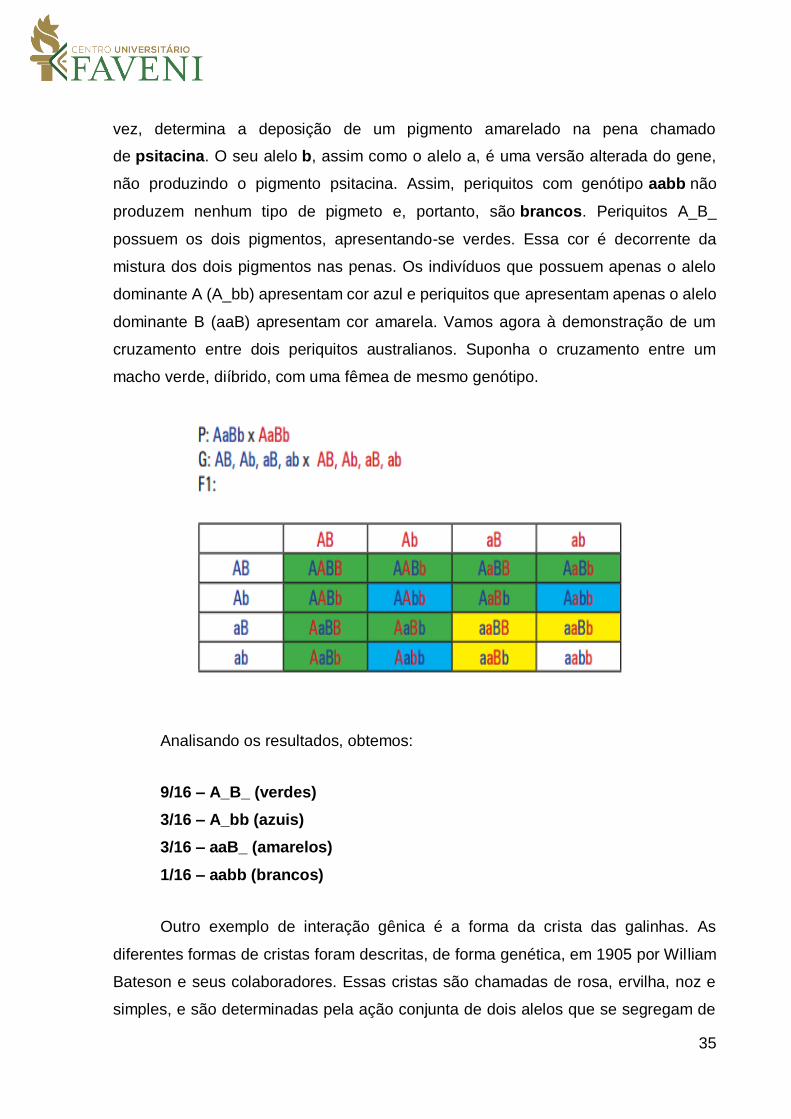
35
vez, determina a deposição de um pigmento amarelado na pena chamado
de psitacina. O seu alelo b, assim como o alelo a, é uma versão alterada do gene,
não produzindo o pigmento psitacina. Assim, periquitos com genótipo aabb não
produzem nenhum tipo de pigmeto e, portanto, são brancos. Periquitos A_B_
possuem os dois pigmentos, apresentando-se verdes. Essa cor é decorrente da
mistura dos dois pigmentos nas penas. Os indivíduos que possuem apenas o alelo
dominante A (A_bb) apresentam cor azul e periquitos que apresentam apenas o alelo
dominante B (aaB) apresentam cor amarela. Vamos agora à demonstração de um
cruzamento entre dois periquitos australianos. Suponha o cruzamento entre um
macho verde, diíbrido, com uma fêmea de mesmo genótipo.
Analisando os resultados, obtemos:
9/16 – A_B_ (verdes)
3/16 – A_bb (azuis)
3/16 – aaB_ (amarelos)
1/16 – aabb (brancos)
Outro exemplo de interação gênica é a forma da crista das galinhas. As
diferentes formas de cristas foram descritas, de forma genética, em 1905 por William
Bateson e seus colaboradores. Essas cristas são chamadas de rosa, ervilha, noz e
simples, e são determinadas pela ação conjunta de dois alelos que se segregam de

36
forma independente, R/r e E/e. A interação entre os genes E e R resulta em crista noz;
entre o alelo R e e resulta em indivíduos com crista rosa; entre E e r resultam
indivíduos com crista ervilha e, entre o gene r e e, resultam indivíduos com crista
simples.
9 HERANÇA POLIGÊNICA
Embora as características sejam determinadas pelo número de genes
dominantes, esse tipo de herança sofre grande influência do meio ambiente. Por
exemplo, uma pessoa possui um conjunto de genes dominantes que determinaria sua
altura como, digamos, 1,90 m. Entretanto, se essa pessoa, determinada
geneticamente a possuir 1,90 m de altura, não possuir um ambiente favorável a esse
fenótipo, como por exemplo, a disponibilidade de alimento, ela não alcançará 1,90 m.
Em outras palavras, de que vale possuir os genes se não se possui o alimento? Uma
rápida analogia pode ser feita com uma construção: de que adianta ter uma planta de
uma casa de três andares se não possui tijolos suficientes para esse tamanho?
Quando analisamos os fatos por esse lado podemos concluir que Determinação
Genética não é o termo apropriado para a definição dos problemas naturais. Nesse
caso, o termo mais correto seria Pré-disposição Genética. Para exemplificar, vamos
falar sobre a herança da cor da pele na espécie humana. O primeiro problema que
enfrentamos é determinar o número de genes envolvidos em tal evento. Para
solucionar esse problema aplicamos uma equação de primeiro grau bem simples:
2n + 1 = x
onde n é o número de genes envolvidos e x é o número de fenótipos existentes
nesse problema. No caso da cor da pele humana, encontramos indivíduos brancos,
mulatos claros, mulatos médios, mulatos escuros e negros, ou seja, 5 fenótipos
diferentes. Aplicando-se a equação acima temos:

37
2n + 1 = 5
2n = 5 – 1
2n = 4 n = 4/2
n = 2
assim, nesse caso estão envolvidos 2 genes, os quais chamaremos de A/a e B/b.
Como nesse tipo de herança o que importa é a quantidade de genes dominantes,
indivíduos aabb serão brancos (sem nenhum gene dominante),
Aabb ou aaBb serão mulatos claros (lembre-se que a herança é quantitativa e
não qualitativa, ou seja, o indivíduo Aabb e aaBb terão o mesmo fenótipo).
AAbb, aaBB e AaBb serão mulatos médios, AABb ou AaBB serão mulatos
escuros e AABB serão negros. Observe a tabela abaixo:
Outros modelos para a cor da pele humana admitem a existência de 3 pares
de genes envolvidos no processo, nesse caso adicionaríamos o alelo C/c. Isso
demonstraria a existência de pessoas de pele muito clara (aabbcc) e pessoas de pele
muito escura (AABBCC). Vale lembrar que a cor da pele é determinada pela
quantidade de melanina presente, a qual é codificada nos alelos discutidos acima.
Lembrando um pouquinho da disciplina de histologia, a melanina deposita-se na forma
de um capuz em cima do núcleo, filtrando os raios UV provenientes do sol. Ou seja,
quanto mais escura for a pele maior será a quantidade de melanina protegendo o
núcleo, e menores serão as mutações causadas por esse tipo de radiação. Para
treinar, vamos a um exemplo prático que foi cobrado no vestibular da UNESP: a altura
de uma certa espécie de planta é determinada por dois pares de genes A e B e seus
respectivos alelos a e b. Os alelos A e B apresentam efeito aditivo e, quando

38
presentes, cada alelo acrescenta à planta 0,15m. Verificou-se que as plantas dessa
espécie variam de 1,00m a 1,60m de altura. Cruzando-se plantas AaBB com aabb
pode-se prever que, entre os descendentes,
a) 100% terão 1,30m de altura,
b) 75% terão 1,30m e 25% terão 1,45m de altura,
c) 25% terão 1,00m e 75% terão 1,60m de altura,
d) 50% terão 1,15m e 50% terão 1,30m de altura,
e) 25% terão 1,15m, 25% 1,30m, 25% 1,45m e 25% 1,60m de altura.
Resolvendo o problema: cruzando-se os indivíduos AaBB com aabb teremos:
P: AaBB x aabb
G: AB, aB x ab
F1:
Analisando-se os resultados encontramos 50% das plantas AaBb (1,30m) e
50% das plantas aaBb (1,15m), sendo, portanto, a resposta d correta.
9.1 Genética da Cor dos Olhos
A primeira tentativa de explicar a determinação genética da cor dos olhos foi
proposta em 1907. Essa proposta admitia apenas 1 par de genes nessa característica.
Assim, indivíduos A_ teriam olhos pretos ou castanhos e indivíduos aa teriam
olhos azuis. Obviamente que essa explicação é insatisfatória, uma vez que não
consegue explicar a existência de cor de olhos como o verde, as variações do
castanho (claro, médio e escuro) etc. A cor da íris no olho humano varia do preto ao

39
azul-claro e cinza. Entretanto, não existem pigmentos azuis ou verdes na íris, sendo
que as diversas cores são produzidas pelas diferentes quantidades de melanina e por
efeitos ópticos. Assim, o olho escuro é resultante do acúmulo de células com muita
melanina (chamadas melanócitos) encontradas na camada de tecido anterior à íris. A
melanina presente nessas células absorve a maior parte da luz incidente, refletindo a
luz marrom-amarelada. Já em casos de pessoas de olhos claros, a quantidade de
melanócitos encontrados na camada anterior da íris é muito reduzida e assim, apenas
pouca luz marrom-amarelada é refletida pela melanina. Dessa forma, a maior parte
da luz incidente atravessa a camada sem pigmento da íris no qual os comprimentos
de onda mais curtos (azul) são refletidos seletivamente, sendo esse fenômeno de
reflexão seletiva conhecido como “dispersão Rayleigh”, como já discutido brevemente
em aula anterior. É esse o efeito que determina a existência da cor azul dos olhos. Já
a cor verde é determinada no caso de a camada anterior da íris conter uma quantidade
intermediária do pigmento melanina, onde a cor marrom-amarelada se combina com
a luz azul que foi produzida pela dispersão Rayleigh. Assim, quanto menor for a
quantidade de melanina na camada anterior da íris produzirá uma variação de
tonalidades da cor castanha dos olhos. A ausência quase que total da melanina
produz olhos que variam e tonalidade do verde ao cinza. Os genes que atuam na
determinação da cor dos olhos em humanos já foram identificados pelos cientistas: o
gene EYCL1, mais conhecido como GEY, localizado no cromossomo número 19 e o
gene EYCL3, localizado no cromossomo 15 e mais conhecido com BEY.O gene GEY
possui dois alelos bem caracterizados, embora exista a possibilidade da existência de
mais alelos. O alelo dominante é chamado de Gv, o qual condiciona a cor verde à íris
e seu alelo recessivo GA, o qual condiciona a cor azul. O termo GEY deriva do termo
inglês green eye color gene. O gene BEY também possui dois alelos, o dominante BM,
o qual condiciona a cor castanha e seu alelo recessivo BA, que condiciona a cor azul.
A denominação do termo BEY deriva do inglês brown eye color gene. O modo de
interação entre esses dois genes será discutido na próxima aula, cujo tema é
epistasia.

40
9.2 Epistasia
Existem alguns casos no qual a ação de um gene impede ou inibe a ação de
outro gene, estando ele localizado no mesmo cromossomo ou não. O gene que
impede a ação do outro é chamado de epistático e o gene que é inibido é chamado
de hipostático. A epistasia, assim como todas as outras formas de herança estudadas
até aqui, pode ser dominante ou recessiva. Um exemplo da ação desse tipo de gene
ocorre na determinação da cor da pelagem de alguns camundongos. Esses
camundongos podem ter pelagem aguti, preta ou albina. Ao se cruzar camundongos
pretos com camundongos albinos, o resultado de F1 é 100% aguti. Fazendo-se o
cruzamento entre os indivíduos de F2, obtemos 9/16 aguti, 3/16 de pretos e 4/16 de
albinos. Observe o exemplo abaixo:
P: AApp x aaPP
G: Ap x aP
F1
F1: AaPp x AaPp
G: AP, Ap, aP, ap x AP, Ap, aP, ap

41
A análise dos resultados obtidos em F1 nos mostram:
9/16 – Aguti
3/16 – Preto
4/16 – Albino
O alelo P condiciona pelagem aguti e o alelo p pelagem preta. O gene A permite
a manifestação de P e p e o seu alelo a é epistático sobre P e p. Assim, indivíduos de
genótipo P_aa não manifestam a cor e aparece como branco. Outro exemplo é a cor
da pelagem em cães labradores. Os cães dessa raça possuem pelagem preta,
chocolate ou dourada. Esses fenótipos são codificados por apenas dois pares de
genes e seus respectivos alelos B/b e E/e. O alelo B determina a produção de um
pigmento preto e seu alelo recessivo b à produção de um pigmento chocolate. O alelo
dominante e condiciona a deposição do pigmento nos pelos e o seu alelo recessivo e
não condiciona a deposição dos pigmentos. Assim, todos os indivíduos que forem ee
não depositarão o pigmento produzido e terão cor dourada. Observe o exemplo do
cruzamento entre um macho dourado e uma fêmea preta:
P: bbee x BBEE
G: be x BE
F1: 100% BbEe (pretos)

42
obtendo F2
G: BE, Be, bE, be x BE, Be, bE, be
Analisando os resultados teremos:
9/16 – Preto
3/16 – Chocolate
4/16 – Dourado
Recentemente foi descoberto mais um gene que participa da determinação da
cor dos olhos em humanos, o EYCL2 ou BEY2, também localizado no cromossomo
15 e que contribui para a coloração castanha do olho. E as pessoas que tem olhos de
cores diferentes, por exemplo, o olho direito é azul e o esquerdo é castanho? Ou
quando no mesmo olho existem regiões de cores diferentes? O que ocorre nesses
casos? Esses fenômenos são chamados de heterocromia e são causados por
diversos motivos. Uma das causas refere-se a um desenvolvimento anormal dos
melanócitos da íris. Para sobreviver, os melanócitos necessitam de impulsos nervosos
que, se por algum motivo forem interrompidos, a cor dos olhos mudará. Além disso,
existem genes de expressividade variável, o que não é assunto novo para nós. Nesse
caso, se os genes forem ativados em apenas um olho teremos olhos com cores
diferentes. Como combinado no início da aula, acredito que agora deu para
compreender a determinação da cor dos olhos em humanos. Caso tenha ficado
alguma dúvida, revise o texto com calma, identificando os principais pontos da aula.

43
9.3 Ligação Gênica
Essas proporções existem por que os genes encontram-se em cromossomos
diferentes e, assim, durante a divisão celular tendem a ir para células diferentes.
Entretanto, muitos genes encontram-se no mesmo cromossomo, sendo que nesse
caso dizemos que os genes estão ligados, de onde deriva o tema da aula de
hoje: Ligação Gênica. Se você se lembra das aulas de citologia, quando estudamos
meiose, existe um fenômeno responsável pela variação genética entre os indivíduos,
que é a recombinação gênica (também chamada de crossing-over). Essa
recombinação gênica ocorre na prófase I da meiose e se dá pela troca de fragmentos
cromossômicos entre os alelos paterno e materno. Assim, em determinados casos
onde consideramos duas ou mais características que fogem da proporção clássica de
Mendel, os genes responsáveis por essas características tendem a ir juntos para o
mesmo gameta, o que altera os resultados esperados. Em Drosophila melanogaster,
podemos observar esse tipo de comportamento na herança da cor do corpo e tamanho
das asas. Essas moscas são encontradas na natureza com uma cor cinzento-
amarelada, sendo determinada por um alelo dominante P. Em laboratório consegue-
se induzir uma mutação nesse gene, inativando-o.
Assim, temos agora um gene recessivo p. No mesmo cromossomo onde se
encontra o gene para a cor do corpo encontra-se também o gene que determina o
tamanho da asa, sendo V o alelo dominante que determina asas alongadas e seu
alelo recessivo v que determina asas vestigiais. Se cruzarmos indivíduos PPVV com
ppvv obteremos uma F1 constituída de 100% dos indivíduos com o genótipo PpVv.
Agora, cruzando-se esses indivíduos com outros do mesmo genótipo deveríamos
esperar, pela lei da segregação independente, 25% dos gametas PV, 25% Pv, 25%
pV e 25% pv. Entretanto não é isso que acontece. Esse cruzamento produz gametas
nas seguintes porcentagens: 41,5% PV, 41,5% pv, 8,5% Pv e 8,5% pV. Esse tipo de
resultado, onde não temos a proporção esperada da segregação independente, nos
mostra que os genes estão localizados no mesmo cromossomo, ou seja, estão
ligados. O primeiro cientista a observar esse fato foi Thomas H.
Morgan, sendo que muito dos termos que usaremos ainda nessa aula são em
sua homenagem. Morgan e seus colaboradores, em 1915, já haviam identificado 85
mutações em D. melanogaster. Ao analisar os cruzamentos, eles observaram que

44
algumas das mutações apresentavam segregação independente e outras
apresentavam ligação gênica. Essas 85 mutações foram distribuídas em quatro
grupos, que foram denominados grupos de ligação, onde um dos grupos não
apresentava mutações com ligação gênica. Posteriormente, estudos citológicos
demonstraram que essas moscas apresentam 4 pares de cromossomos (2n = 8).
Assim, Morgan e seus colaboradores notaram uma exata relação entre os 4 grupos
onde foram divididos as mutações e o número de cromossomos, o que era mais um
indício de que os genes se localizavam nos cromossomos. Em 1909 o belga Frans
Alfons Janssen (1863-1924) propôs uma explicação para o entrelaçamento dos
cromossomos que ocorriam durante a meiose e que vinham sendo estudados pelos
cientistas há algum tempo. Janssen considerou que nos entrelaçamentos dos
cromossomos havia trocas de fragmentos cromossômicos, evento que ele chamou de
permuta. O termo quiasma, utilizado para designar o mesmo evento designa do termo
gregoKhiasmós, que significa “posição em cruz”, ou em forma da letra khi, x.
Assim, com base na proposta de Janssen, Morgan criou uma hipótese para
explicar a proporção de gametas que fugiram aos padrões mendelianos. Sua hipótese
sugeria que os genes que estavam localizados no mesmo cromossomo (o da forma
da asa e o da cor do corpo), na prófase I da meiose, sofriam permuta, formando
cromátides recombinantes que iriam formar gametas diferentes. A sua hipótese foi
comprovada em diversas espécies.
9.4 Arranjos “cis” e “trans”
Tomando-se como exemplo o caso da mosca D. melanogaster, podemos
verificar duas possibilidades de arranjo dos alelos P/p e V/v no cromossomo. Veja a
imagem abaixo:

45
No arranjo cis os genes dominantes P/V encontram-se no mesmo cromossomo
e os genes recessivos p/v encontram-se no cromossomo homólogo correspondente.
Já no arranjo trans um cromossomo possui um gene dominante e outro recessivo.
Para se saber na prática qual é o arranjo gênico em questão basta realizar
um cruzamento-teste. Analisando-se a descendência, as classes que aparecem com
maior frequência são as portadoras das combinações parentais e, consequentemente,
as que aparecem em menor frequência são as recombinantes.
9.5 Mapeamento Cromossômico
Relembrando os resultados obtidos por Morgan nos experimentos com D.
melanogaster, observamos que surgiram gametas que não deveriam existir, pois do
cruzamento entre indivíduos PPVV com ppvv os únicos gametas possíveis seriam PV
e pv, respectivamente. Esses gametas surgiram pela recombinação gênica, no
momento em que houve uma quebra e troca de fragmentos dos cromossomos
homólogos, sendo, portanto, Pv e pV os novos gametas. Como um deles teve uma
frequência de 8,5%, ou seja, entre os genes P e V – e entre seus alelos p e v – existe
uma frequência de 17% (8,5% + 8,5%) de recombinação. Vamos imaginar um
cromossomo como uma estrutura linear onde os genes estão dispostos um em
sequência do outro com distâncias variáveis. Observe a imagem abaixo:

46
Entre quais pares de genes é mais fácil a ocorrência de uma quebra? Entre A
e B ou entre B e C? Obviamente entre A e B, pois a distância entre eles é maior, ou
seja, o local onde essa quebra pode ocorrer é maior. Sendo maior a probabilidade de
ocorrer uma quebra entre A e B, maior vai ser também a frequência com que esses
genes irão se recombinar. Assim, a distância entre os genes é igual a sua frequência
de recombinação. Observe o mesmo esquema só que com as distâncias entre os
genes:
Como você pôde notar a distância entre os genes é medida em centimorgans,
em homenagem ao cientista Thomas H. Morgan, entretanto, essa distância também
pode ser chamada de unidades de recombinação (UR) ou ainda, morganídeos. Assim,
como dissemos acima, se o gene A está a 32 UR de distância do gene B, sua
frequência de recombinação será igual a 32%. Com base nesse conceito de
frequência e distância entre os genes nós podemos montar um mapa cromossômico.
Vamos a um exemplo: suponhamos que a cidade A fica a 25 Km de distância da
cidade B. cidade C fica a 10 Km de distância da cidade A. Qual será a ordem em que
as cidades se localizam na estrada? Temos, nesse caso, duas possibilidades: 1) a da
cidade C estar antes da cidade A ou, 2) a da cidade C estar entre a cidade A e B,
assim:

47
Para podermos afirmar qual é a ordem correta, devemos saber qual é a
distância entre a cidade C e B. Desta maneira, se a distância entre C e B for 35 Km,
a primeira possibilidade estará correta. Agora, se a distância entre C e B for de 15 Km,
a segunda possibilidade estará correta. O mesmo princípio aqui mostrado é utilizado
para localizar a posição dos genes no cromossomo. Tendo-se as frequências de
recombinação tem-se também a distância entre os genes. Agora é só colocá-los em
ordem. Para fazer isso, sempre começamos com os genes que apresentam maior
recombinação e vamos até o último de forma decrescente. Observe o exemplo: quatro
genes, A, B, C e D, localizados no mesmo cromossomo apresentam as seguintes
frequências de recombinação:
A-B = 32% A-C = 45 % A-D = 12% B-C = 13% B-D = 20% C-D = 33%
Iniciando com os genes que apresentam maior recombinação, teremos:

48
Inserindo-se o gene que tem a segunda maior frequência de recombinação,
podemos ter:
Para resolver esse problema basta olharmos para a distância entre os genes A
e D. Como demonstrado no enunciado, a distância entre A e D é de 12 UR. Assim, o
gene D deve ficar entre os genes A e C (45 – 33 = 12). Caso ficasse para o lado de
fora, a distância entre A e d deveria ser 78 UR (45 + 33 = 78), portanto:
Para finalizar precisamos saber qual é a posição do gene B. A distância entre
ele e o gene A é de 32 UR. Da mesma forma que fizemos da vez anterior vamos fazer
agora. Vamos considerar o gene B do lado de fora do gene A e entre o gene A e os
demais genes:

49
Ao observarmos novamente o enunciado, constatamos que o gene B está a 13
UR do gene C. Assim, podemos afirmar que ele está do lado de dentro, pois 45
(distância entre os genes A e C) menos 32 (distância entre A e B) é igual a 13, que é
a distância entre B e C.
Concluímos, então, que a ordem dos genes nos cromossomos é ADBC. Espero
que você tenha compreendido o que foi exposto acima.
9.6 Sistemas de Determinação Sexual
A reprodução é uma característica fundamental de qualquer ser vivo. Por meio
dela a vida se expandiu pela Terra há aproximadamente 3 bilhões de anos.
Basicamente, existem dois tipos de reprodução: a reprodução sexuada e a reprodução
assexuada. Infelizmente os termos usados não condizem com verdade nesse caso. À
primeira vista teríamos a impressão de que a reprodução sexuada envolve o ato
sexual, enquanto a reprodução assexuada não envolve o ato sexual. O que ocorre é

50
que o ato sexual não é necessariamente obrigatório na reprodução sexuada. Um
exemplo claro desse fato é a reprodução dos sapos. Não há penetração do macho na
fêmea, mas apenas um estímulo sexual gerado pelo “apertão”, por assim dizer, do
macho sobre o ventre da fêmea, que faz com que ela libere seus gametas na água,
onde o macho irá liberar seus espermatozoides posteriormente, para que haja
fecundação. Portanto, reprodução sexuada NÃO envolve necessariamente sexo, mas
obrigatoriamente gametas. Já a reprodução assexuada não envolve gametas. os
gametas são células especializadas na reprodução. Elas são produzidas nas gônadas
por um tipo de divisão celular chamada de meiose. Em humanos, a gônada masculina
é o testículo, o qual produz o espermatozoide, e a gônada feminina é o ovário, que
produz o ovócito secundário. Talvez você esteja estranhando o termo “ovócito
secundário”. Na verdade, o gameta feminino, na maioria dos vertebrados, é o ovócito
secundário, o qual somente completará sua divisão celular se for fecundado,
transformando-se em óvulo logo em seguida.
Em algumas espécies os mesmos indivíduos são capazes de originar tanto o
gameta masculino quanto o gameta feminino. Nesse caso estamos falando de
hermafroditismo, ou espécies monoicas (do grego mono, um, e oikos, casa). Quando
existem indivíduos especializados na produção de um tipo de gameta e outros no outro
tipo, estamos falando de espécies dioicas (do grego di, dois, e oikos, casa), a exemplo
da espécie humana. Como mencionado anteriormente, a determinação do sexo nas
espécies é caracterizada pela presença dos cromossomos sexuais. Entretanto,
algumas espécies animais, como jacarés, crocodilos e muitas espécies de tartarugas
e alguns lagartos têm o sexo determinado por condições ambientais. Nos crocodilos,
por exemplo, a temperatura de incubação dos ovos é fator determinante para o sexo
do embrião. Se o embrião for incubado em locais onde a temperatura é relativamente
baixa nascerão somente indivíduos do sexo feminino; já os machos somente nascerão
onde o local de incubação dos ovos possuírem temperaturas relativamente altas. Em
tartarugas ocorre exatamente o contrário: em locais de temperatura alta nascem
fêmeas e em locais de temperatura baixa nascem somente machos. Na natureza
encontramos 5 sistemas de determinação do sexo, o sistema XY, o sistema X0 (leia-
se xis-zero), o sistema ZW e o sistema Z0 (leia-se zê-zero) e o sistema
haploide/diploide. No sistema XY as fêmeas possuem um par de cromossomos
sexuais idênticos, os quais são denominados XX.

51
Os machos possuem um cromossomo igual ao das fêmeas e um cromossomo
caracteristicamente masculino, o cromossomo Y, sendo constituído, portanto, por um
par XY. Dessa forma, dizemos que o sexo feminino é homogamético (do grego homo,
igual), pois todos os gametas conterão o cromossomo sexual X. O sexo masculino é
classificado como heterogamético (do grego hetero, diferente), pois 50% dos seus
gametas conterão o cromossomo X e 50% conterão o cromossomo Y. Em mamíferos,
um gene localizado no cromossomo Y é o responsável pela determinação direta do
sexo masculino. Nele encontra-se um gene chamado SRY, cuja sigla em inglês
significa sex-determining region Y. A proteína codificada por esse gene induz a
formação de testículos no embrião, por meio da ativação de outros genes em
diferentes cromossomos. O sistema X0 é muito parecido com o sistema XY. As
fêmeas apresentam dois cromossomos sexuais X (XX), entretanto, nos machos não
existem o cromossomo sexual Y e dessa forma eles apresentam apenas um
cromossomo sexual, o próprio X. Assim, o que determina ser um macho é a ausência
do cromossomo Y.
Um exemplo de animal que possui esse tipo de determinação sexual é o
gafanhoto. O sistema ZW, o qual ocorre em diversos répteis e em algumas espécies
de insetos (mariposas e borboletas, por exemplo) e de peixes, é caracterizado pelos
machos serem classificados como homogaméticos, ou seja, nesse caso eles possuem
dois cromossomos sexuais iguais, que são chamados de ZZ. As fêmeas possuem um
cromossomo sexual diferente, sendo, portanto, chamadas de sexo heterogamético,
possuindo os cromossomos ZW. Ou seja, nessas espécies ocorre o inverso do
sistema XY. Como podemos perceber no sistema XY quem determina o sexo da prole
é o macho e no sistema ZW quem determina o sexo da prole é a fêmea. O sistema Z0
restringe-se a galinha doméstica e algumas espécies de répteis, as quais, a exemplo
das espécies pertencentes ao sistema X0, também não possuem um cromossomo
sexual, nesse caso o cromossomo um caso muito interessante de determinação do
sexo ocorre em abelhas e formigas (insetos himenópteros). Nesses insetos, ocorre o
sistema haploide/diploide, nos quais os machos possuem apenas um lote
cromossômico e têm, portanto, apenas n cromossomos. Já as fêmeas são
constituídas por dois lotes cromossômicos e são, portanto, 2n. Vamos tomar como
exemplo as abelhas. Os zangões originam-se por meio de um óvulo não fecundado,
por um processo conhecido como partenogênese (do grego partenos, virgem, e

52
genesis, origem). A partenogênese é, portanto, um processo de reprodução na qual
não há fecundação do óvulo por um gameta masculino. Assim, como o zangão
desenvolve-se de um óvulo não fecundado ele é haploide, possuindo apenas n
cromossomos. As abelhas (fêmeas) se originam dos óvulos que foram fecundados e,
portanto, são diploides, possuindo 2n cromossomos. A diferença entre a rainha e as
abelhas operárias está na alimentação durante a sua fase inicial de vida, a fase larval.
Se durante a fase larval elas forem nutridas com a geleia real elas tornam-se
indivíduos adultos férteis, as quais serão responsáveis apenas pela reprodução e por
isso são chamadas de rainhas. Se, por outro lado, elas não forem nutridas com a
geleia real, elas tornam-se indivíduos adultos estéreis e farão todo o serviço da
colmeia, sendo, assim, chamadas de operárias. Mais recentemente foram feitas novas
descobertas em relação à determinação do sexo masculino em abelhas, mais
precisamente nas abelhas do gênero Apis. Segundo esses novos estudos, o sexo não
é determinado pelo número de lotes cromossômicos. Os óvulos haploides possuem
apenas uma única versão de um gene chamado csd, palavra que em inglês significa
complementary sex determiner, cuja tradução é “determinante complementar do
sexo”. O csd possui 19 formas alélicas. Se um indivíduo possuir apenas uma versão
do gene ele será do sexo masculino, mas, se possuir 2 versões do gene será do sexo
feminino. Como existem 19 formas alélicas, a maioria das fêmeas são heterozigotas,
pelo grande número de possíveis combinações entre elas. Muito raramente ocorre a
formação de um indivíduo homozigoto para o gene csd, o qual acaba por se
desenvolver em um macho diploide. Em diversos tipos de plantas o sexo é
determinado da mesma forma como ocorre em animais. O cânhamo e o espinafre, por
exemplo, têm a determinação do sexo de acordo com o sistema XY; já no morango
silvestre a determinação do sexo ocorre de acordo com o sistema ZW.
9.7 Herança Sexual
Podemos notar que o cromossomo X do filho foi herdado da mãe, enquanto o
pai forneceu o cromossomo Y. Como o daltonismo é uma herança recessiva
relacionada ao cromossomo X, o pai, por ser normal possui o alelo XDY. A mãe,
também normal, possui um alelo XD, mas, para ter um filho daltônico, ela deve ser
portadora do alelo Xd, sendo, portanto, XDXd. Observe:

53
Assim, do cruzamento entre XDY com XDXd, obtemos as seguintes
proporções:
P: XDY x XDXd
G: XD, Y x XD, Xd
F1: XDXD – 25%
XDXd – 25%
XDY – 25%
XdY – 25%
Proporção fenotípica:
50% de chances de mulher normal (XDXD e XDXd)
25% de chances de um homem normal (XDY)
25% de chances de um homem daltônico (XdY)
O mesmo vale para os casos de hemofilia. Essa doença é causada por um alelo
recessivo h localizado no cromossomo X. Assim, indivíduos XH_ serão normais e
indivíduos XhXh ou XhY serão afetados pela doença. O daltonismo é uma doença que
atinge cerca de 5% a 8% dos homens e 0,04% das mulheres. Essa doença
caracteriza-se pela incapacidade de distinguir as cores. O termo daltonismo deriva do
nome do físico John Dalton, o qual apresentava essa característica. Basicamente
existem três tipos de daltonismo. No primeiro, as pessoas afetadas não distinguem a
cor púrpura da cor vermelha; no segundo, as pessoas veem a cor vermelha como se
fosse a cor verde; e no terceiro as pessoas não conseguem separar a cor verde da
vermelha, vendo as duas cores como avermelhadas. O primeiro tipo de daltonismo
discutido acima é condicionado por um gene recessivo localizado num cromossomo
autossomo, não tendo, portanto, o mesmo padrão de herança que discutimos acima.
Os segundos e o terceiro tipos são condicionados por genes recessivos localizados
no cromossomo X, seguindo o padrão acima estudado. A hemofilia é uma doença na
qual, na pessoa afetada, o sistema de coagulação sanguínea não funciona, e por isso
essas pessoas apresentam hemorragias abundantes, mesmo sendo pequeno o
ferimento. O processo de coagulação sanguínea envolve uma série de reações entre
proteínas presentes no sangue, as quais são codificadas por diversos genes
localizados no cromossomo X. Dentre essas proteínas, está o fator VIII de coagulação

54
sanguínea, cuja mutação no gene responsável pela sua codificação causa o tipo mais
grave de hemofilia. Pessoas hemofílicas são tratadas com injeções de fator VIII de
coagulação sanguínea extraídas do sangue de pessoas normais. A distrofia muscular
de Duchenne é uma doença genética progressiva e letal causada por uma mutação
no gene que codifica a proteína distrofina, localizado no cromossomo X. Essa doença
afeta um a cada 3500 homens e a pessoa afetada sofre uma deficiência no suporte
muscular, fazendo com que o desenvolvimento do músculo não acompanhe o do
corpo. Os pacientes, quando atingem os 12 anos de idade, comumente necessitam
de cadeira de rodas e morrem de parada respiratória ao final da adolescência ou
pouco depois dos 20 anos de idade. Muitos garotos também mostram
eletrocardiograma anormal, indicando que o músculo cardíaco e o diafragma também
estão envolvidos nessa patologia. Embora tenhamos falado até o momento apenas
nas heranças relacionadas ao cromossomo X, existentes na espécie humana, uns
poucos genes estão localizados no cromossomo Y.
Quando alguns desses genes sofrem uma mutação ele é passado de pai para
filho, uma vez que as mulheres não possuem o cromossomo Y. Assim, os genes
localizados nos cromossomos Y são chamados de genes holândricos (do grego holos,
totalmente, e andros, masculino), embora alguns autores também se refiram a esse
tipo de herança como “herança restrita ao sexo”. A hipertricose auricular (do grego
hiper, excesso, e trichos, pelos) é caracterizada pela presença de pelos longos nas
orelhas, manifestando-se apenas em homem. Até pouco tempo acreditava-se que a
hipertricose auricular era causada por um gene localizado no cromossomo Y, mas
estudos recentes têm demonstrado que esse gene se localiza em um cromossomo
autossomo. Assim, passamos a classificar a hipertricose auricular como “expressão
limitada ao sexo”. Algumas outras características têm “expressão influenciada pelo
sexo”, como o caso da calvície. A calvície manifesta-se como uma diminuição
generalizada de cabelos, o que é diferente da perda total de cabelos que ocorre no
topo do couro cabeludo. Alguns estudos têm levantado a possibilidade da calvície ser
causada por um gene dominante autossômico no homem e um gene recessivo na
mulher.

55
9.8 Composição do Material Genético
A história do DNA teve início no final da década de 1860 com os estudos do
médico suíço Friedrich Miescher (1844-1895), na Universidade de Tübingen. Miescher
trabalhava com células do pus e reparou que em alguns dos seus experimentos
formou-se um precipitado quimicamente diferente das outras substâncias proteicas
conhecidas. A substância que formava esse precipitado estava presente no núcleo
celular, que, na época, era tido como uma estrutura de pouca importância na célula.
Uma análise química mais detalhada mostrou que a substância desse precipitado era
composta de hidrogênio, carbono, oxigênio, nitrogênio e fósforo. A identificação desse
precipitado como uma nova substância se deu pelo fato de que as proteínas não
possuem fósforo em sua composição, além de que as quantidades relativas dos outros
elementos químicos não eram equivalentes ao encontrados nas proteínas. Assim,
Miescher, pelo fato de a substância estar concentrada no núcleo celular, deu o nome
de nucleína a essa nova substância.
No ano de 1889, Richard Altmann (1852-1900) obteve preparações altamente
purificadas de nucleína e verificou o seu caráter ácido, sugerindo que essa substância
passasse a ser chamada de ácido nucleico. Albert Kossel (1853-1927), outro
pesquisador de grande importância na história do DNA, em 1877, juntou-se ao grupo
de pesquisas de Hoppe-Seyler, que trabalhava na Universidade de Estrasburgo, na
França, onde iniciou seus estudos da composição química das diferentes nucleínas
encontradas em diferentes células. Nesses estudos ele identificou as bases
nitrogenadas adenina e guanina, já conhecidas anteriormente. Em 1893, Kossel
descobriu uma nova base nitrogenada em células provenientes do timo, dando o nome
de timina a essa base. Em seguida, descobriu a existência de uma quarta base
nitrogenada na nucleína, a qual ele chamou de citosina. No ano seguinte, Kossel e
seu grupo de pesquisa descobriram que os ácidos nucléicos também continham
pentose (um açúcar de 5 átomos de carbono). Em 1930, Phoebis Levine (1869-940)
e seus colaboradores identificaram a pentose do ácido nucleico como 2-desoxi-D-
ribose. Essa pentose diferia da ribose (já conhecida na época) por possuir um átomo
de oxigênio a menos no carbono 2. Com essa identificação ficaram então
caracterizados dois tipos de ácidos nucleicos: o ácido ribonucléico (RNA, em inglês
ribonucleic acid) e o ácido desoxirribo nucléico (DNA, em inglês deoxyribose nucleic

56
acid). Somente a partir da década de 1940 alguns indícios apontavam para o DNA
como material genético, o que levou inúmeros pesquisadores a voltarem sua atenção
para essa substância. Até o final da década de 1940 todas as informações sobre o
DNA estavam dispersas, o que não levava os cientistas à conclusão alguma. Foi então
que o biólogo James D. Watson e o físico Francis H. C. Crick decidiram juntar todos
os dados disponíveis até o momento e determinar a estrutura da molécula de DNA.
Para realizar tal fato, Watson e Crick usaram difração de raio-X para analisar a
molécula e, ao final dos estudos, propuseram um modelo de dupla-hélice, o qual
explicava ao menos três características fundamentais do material genético: a
capacidade de conter informações para a síntese de proteínas, capacidade de sofrer
mutações e capacidade de duplicação.
Esse modelo, proposto em 1953, foi prontamente aceito por toda a comunidade
científica, que junto com muito esforço de milhares de cientistas de todas as partes do
mundo, nos levou a esse enorme avanço na área da biotecnologia que vivemos hoje.
Foi em meados da década de 1930 que surgiram os primeiros indícios experimentais
de que os genes atuam por meio do controle da síntese de proteínas. Um experimento
realizado por George W. Beadle (1903-1989), Boris Ephurussi (1901-1979) e Edward
L. Tatum (1909-1975) demonstrou que a cor do olho alterada em um mutante de D.
melanogaster se devia à incapacidade da mosca realizar uma reação química de uma
determinada via metabólica da síntese do pigmento normal. Uma vez que o inseto D.
melanogaster é um organismo muito complexo para poder afirmar os resultados
obtidos inicialmente, Beadle e Tatum resolveram utilizar o fungo Neurospora crassa,
conhecido popularmente como bolor rosado do pão, que é um organismo mais simples
quando comparado a D. melanogaster. Os resultados dos experimentos de Beadle e
Tatum consolidaram a teoria um gene – uma enzima, a qual logo em seguida foi
alterada para teoria um gene – uma proteína. Tempos depois foi descoberto que uma
proteína pode ser formada por mais de uma cadeia polipeptídica (proteínas que
possuem mais de uma cadeia polipeptídica possuem uma estrutura quaternária, a
qual se dá pelo arranjo dessas cadeias) e passou então a se falar em teoria um gene
– um polipeptídio. Todas essas descobertas da primeira metade do século XX
conduziram os pesquisadores aos estudos da genética molecular. Os primeiros
estudos mostraram que o DNA atua diretamente na síntese de proteínas.

57
Assim, o DNA contém as instruções para a síntese de uma dada proteína
codificada na sequência de suas bases nitrogenadas, as quais são transcritas para
uma molécula de RNA e, a partir destas, as instruções serão traduzidas na forma de
proteína. Existem três tipos de RNA envolvido na síntese de proteínas. O primeiro
deles é o RNA mensageiro, abreviado como mRNA. Ele nada mais é do que uma
cópia do gene de interesse na forma de RNA, uma vez que o DNA não sai do núcleo
celular. Cada sequência de 3 bases nitrogenadas corresponde ao que chamamos de
códon.

58
10 BIBLIOGRAFIA BÁSICA
PIERCE, Benjamin A. Genética: um enfoque conceitual. Rio de Janeiro: Guanabara
Koogan, c2004. xxvi, 758p., il. (algumas col.), 29 cm. Inclui bibliografia e índice.
RINGO, John. Genética básica. Rio de Janeiro: Guanabara Koogan, 2005. 390p., il.,
28 cm.
SNUSTAD, D. Peter. Fundamentos de genética. Rio de Janeiro: Guanabara
Koogan, c2008. xviii, 756p., il. col., 29 cm. Inclui bibliografia e índice.
AMABIS, José Mariano. Biologia: volume 3: genética, evolução e ecologia. 2 ed. São
Paulo: Moderna, 1979. 469p., il., 22 cm. Inclui índice.
BIBLIOGRÁFIA COMPLEMENTAR STRACHAN, T. Genética molecular humana. Porto Alegre: Artmed, 2002. xxiii,
576p., il., 28 cm. Inclui bibliografia e índice.
BURNS, George W. Genética. Rio de Janeiro: Guanabara Koogan, c1998. 381p., il.,
28 cm. Bibliografia: p. [359]-368.
LIMA, Celso Piedemonte de. Genética humana. 3ed. São Paulo: Harbra, c2004. xiv,
442p., il. (algumas col.), 24cm.
GELBART, W. M. Introdução à genética. Rio de Janeiro: Guanabara Koogan, 2006.
xviii, 743p., il. (algumas col.), 28 cm.
AMABIS, José Mariano. Biologia: volume 3 : genética, evolução e ecologia. 2 ed. São
Paulo: Moderna, 1979. 469p., il., 22 cm. Inclui índice.