Embed Size (px)
Citation preview

Cytotherapy, 2014; 16: 191e202
Human mesenchymal stromal cells transiently increase cytokineproduction by activated T cells before suppressing T-cell proliferation:effect of interferon-g and tumor necrosis factor-a stimulation
JESSICA CUERQUIS1,*, RAPHAËLLE ROMIEU-MOUREZ1,*, MOÏRA FRANÇOIS1,JEAN-PIERRE ROUTY2, YOON KOW YOUNG1, JING ZHAO1 &NICOLETTA ELIOPOULOS1,3,4
1Lady Davis Institute for Medical Research and Sir Mortimer B. Davis Jewish General Hospital, Montreal, Quebec,Canada, 2Division of Hematology and Chronic Viral Illness Service, Royal Victoria Hospital, Montreal, Quebec,Canada, and 3Department of Surgery, Division of Surgical Research, and 4Department of Oncology, McGill University,Montreal, Quebec, Canada
AbstractBackground aims. Mesenchymal stromal cells (MSCs) suppress T-cell proliferation, especially after activation with in-flammatory cytokines. We compared the dynamic action of unprimed and interferon (IFN)-g plus tumor necrosis factor(TNF)-aepretreated human bone marrowederived MSCs on resting or activated T cells. Methods. MSCs were co-cultured with allogeneic peripheral blood mononuclear cells (PBMCs) at high MSC-to-PBMC ratios in the absence orpresence of concomitant CD3/CD28-induced T-cell activation. The kinetic effects of MSCs on cytokine production andT-cell proliferation, cell cycle and apoptosis were assessed. Results. Unprimed MSCs increased the early production ofIFN-g and interleukin (IL)-2 by CD3/CD28-activated PBMCs before suppressing T-cell proliferation. In non-activatedPBMC co-cultures, low levels of IL-2 and IL-10 synthesis were observed with MSCs in addition to low levels of CD69expression by T cells and no T-cell proliferation. MSCs also decreased apoptosis in resting and activated T cells andinhibited the transition of these cells into the sub-G0/G1 and the S phases. With inhibition of indoleamine 2,3 dioxy-genase, MSCs increased CD3/CD28-induced T-cell proliferation. After priming with IFN-g plus TNF-a, MSCs were lesspotent at increasing cytokine production by CD3/CD28-activated PBMCs and more effective at inhibiting T-cell pro-liferation but had preserved anti-apoptotic functions. Conclusions. Unprimed MSCs induce a transient increase in IFN-gand IL-2 synthesis by activated T cells. Pre-treatment of MSCs with IFN-g plus TNF-a may increase their effectivenessand safety in vivo.
Key Words: cytokines, human, mesenchymal stromal cells, T cells
Introduction
Mesenchymal stromal cells (MSCs) are deemed asimmunosuppressive because they produce severalanti-inflammatory and immune-suppressive factors.Immune suppression by MSCs appears as a multi-factorial process requiring contact-dependent as wellas contact-independent signals, among them inter-leukin (IL)-1 receptor antagonistic, tumor necrosisfactor (TNF)-aestimulated gene/protein 6, prosta-glandin E2 (PGE2), transforming growth factor-b,hepatocyte growth factor, human leukocyte antigen-G, galectins, heat shock protein 32/HO-1, produc-tion of matrix metalloproteases, and indoleamine 2,3
*These authors contributed equally to this work.Correspondence: Nicoletta Eliopoulos, PhD, Departments of Surgery (DivisiInstitute for Medical Research, Jewish General Hospital, 3755 [email protected]
(Received 6 March 2013; accepted 16 November 2013)
ISSN 1465-3249 Copyright � 2014, International Society for Cellular Therapy. Phttp://dx.doi.org/10.1016/j.jcyt.2013.11.008
dioxygenase (IDO) in humans or nitric oxide inmice (1). The expression of several of these immune-suppressive molecules as well as of chemokines andadhesion molecules is upregulated in MSCs byinterferon (IFN)-g in concert with IL-1a, IL-1b orTNF-a, and it has been implicated that full sup-pressive activity depends on a pro-inflammatoryactivation or “licensing” of MSCs with exposure toan inflammatory or Th1-oriented immune environ-ment (2,3). In vitro, the suppression of T-cell pro-liferation by MSCs was reported as more effectivewhen MSCs are pretreated with IFN-g or IFN-gplus TNF-a (4e6). This corroborates with in vivo
on of Surgical Research) and Oncology, McGill University, Lady Davisatherine Road, Montreal, Quebec H3T-1E2, Canada. E-mail: nicoletta.
ublished by Elsevier Inc. All rights reserved.

192 J. Cuerquis et al.
experiments in the graft-versus-host disease modeldemonstrating that MSCs pretreated with IFN-g aremore potent than are non-treated MSCs at control-ling graft-versus-host disease (3).
Some detailed analysis nevertheless suggested thatMSCs can support immune cell activation in an anti-gen-independent fashion. In certain experimentalco-culture settings with PBMCs, human MSCs wereobserved to support in vitro allogeneic T-cell prolifer-ation, lipopolysaccharide- or antigen-induced immu-noglobulin G secretion by spleen B cells or to suppressapoptosis of neutrophils. These outcomes were moreevident in MSC and PBMC co-cultures at low ratiosof MSCs to immune cells (4,7) or with a weak or noimmune activation (2,8e10). The mechanisms sup-porting the immune-stimulatory functions of MSCshave not yet been formally identified. It is suspectedthat MSCs sustain the survival of lymphocytes. In thebone marrow, several MSC populations of distinctphenotypes participate in the maintenance of hemato-poietic stem cells, plasma cells or memory T- andB-cellniches (11). Invitro,dataontheeffectsofMSCsonT-cell apoptosis are conflicting because some authorshave suggested that MSCs protect T cells from basaloractivation-inducedcelldeath (12,13),maintainTcellsin a state of quiescence (12e14) that can be partiallyreversed by IL-2 stimulation (15), whereas othershave reported that MSCs have no effect on activation-induced apoptosis of T cells (14,15) or increase it (16).
To address the dynamic action of MSCs onT cells and the effect of MSC priming with IFN-gplus TNF-a, peripheral blood mononuclear cells(PBMCs) were co-cultured with allogeneic bonemarrowederived MSCs that were pretreated or notpretreated with IFN-g plus TNF-a. UnfractionedPBMCs and direct co-cultures with MSCs wereused throughout the study because MSCs wereshown to act directly on T cells and other PBMCsubsets such as monocytes through cell contact andsoluble factors for an optimal suppression of T-cellactivation (6,17e19). The kinetics of IFN-g, IL-2and IL-10 production in co-cultures, T-cell prolif-eration, entry into the cell cycle and induction ofapoptosis were investigated in the absence andpresence of concomitant CD3/CD28-induced T-cell activation. Our results suggest that unprimedMSCs increase the early production of IFN-g andIL-2 by CD3/CD28-activated PBMCs before sup-pressing T-cell proliferation, an effect that does notoccur in non-activated PBMC co-cultures. Treat-ment of MSCs with IFN-g plus TNF-a diminishestheir potency to trigger the increase in IL-2 andIFN-g production by CD3/CD28-activated PBMCsand results in a more effective suppression of acti-vated T-cell proliferation while preserving theiranti-apoptotic functions toward T cells.
Methods
Human MSCs and PBMCs
MSCs(donor303:72-year-oldwoman;donor304: 68-year-old woman; donor 305: 62-year-old woman;donor 306: 62-year-old woman; donor 307: 78-year-old woman; donor 311: 70-year-old woman; donor315: 34-year-old woman; donor 316: 57-year-oldman;donor 317: 71-year-old woman; donor 319: 77-year-old man; and donor 320: 58-year-old man) were iso-lated from bone marrow aspirates collected from theiliac crest of consenting individuals undergoing hipreplacement surgery at the Jewish General Hospital(Montreal, QC, Canada) through the use of a protocolapproved by the Research Ethics Committee of theJewish General Hospital. Mononuclear cells were iso-lated by Ficoll gradient and plated in a-minimumessential medium, 20% fetal bovine serum (FBS),20 mmol/L L-glutamine and 100 U/mL penicillin/streptomycin at 4000 cells/cm2 (passage 0). Non-adherent hematopoietic cells were removed by chang-ing the medium after 5 days of culture. Cultures werethen reseeded at 200 cells/cm2 once per week. After thethird passage, MSCs were tested for the absence ofCD31þ endothelial cells and CD45þ hematopoieticcells, the expression of CD44, CD73, CD90 andCD105 and the capacity to differentiate into adipocytesand osteocytes, as described (6). All assays were per-formedwith the use ofMSCs between passages 3 and 5to eliminate any residual hematopoietic cells and avoidsenescence. HumanPBMCswere isolatedwith the useof a Ficoll density gradient from leukapheresis or blooddraws obtained at the Royal Victoria Hospital or theJewishGeneralHospital (Montreal,QC,Canada) from12 consenting healthy volunteers with the use of pro-tocols approved by the Research Ethics Committee ofthe Royal Victoria Hospital or the Jewish GeneralHospital. PBMCs from donors 1e7 and 9 were usedonly fresh.Repeated and separate blooddraws couldbeobtained fromdonors 1, 2 and 3during a 6-month timelapse. PBMCs from donors 8, 10, 11 and 12 werefrozen at 10e20 million cells per vial in FBS with 5%dimethyl sulfoxide, preserved in liquid nitrogen tanks,and thawed in a water bath at 37�C before use. PBMCviability on thawing was >95%, and cells were platedand incubated overnight at 37�C for recovery beforebeing assayed.The numbers of experiments performedwith the different PBMCandMSCdonors are detailedin the figures and legends.
MSC and PBMC co-cultures
All PBMC and MSC co-cultures were allogeneic.PBMCs were cultured in duplicates at 2 � 105 cellsper well in 48-well plates with untreated (naive)MSCs or IFN-g plus TNF-aepretreated MSCs at an

MSCs support cytokine production by activated T cells 193
MSC:PBMC ratio of 1:3 or 1:9 (unless otherwiseindicated) in Roswell Park Memorial Institute me-dium (RPMI)-1640, 10% FBS and 100 U/mL peni-cillin/streptomycin. For cytokine pretreatment ofMSCs, IFN-g and TNF-a (Life Technologies,Carlsbad, CA, USA) were added for 18 h at 10 ng/mL(68 IU/mL) and 3 ng/mL (330 IU/mL), respectively,andMSCs were extensively washed before addition toco-cultures with PBMCs. Treatment of MSCs withIFN-g plus TNF-a at 10 ng/mL and 3 ng/mL,respectively, did not lead to a significant change inMSC numbers over a period of 5 days as determinedby manual cell counting after trypan blue staining(data not shown). Where indicated, stimulation of theT lymphocytes was induced by addition of 1 mg/mL ofsoluble human-specific anti-CD3 and anti-CD28antibodies (BD Biosciences, Franklin Lakes, NJ,USA; clones HIT3a and CD28.2, respectively) to theco-cultures. Where indicated, 10 mmol/L indometh-acin or 1 mmol/L 1-methyl-DL-tryptophan (1-MT)purchased from Sigma-Aldrich (St. Louis, MO,USA) was added at the beginning of the co-cultures.
Analysis of proliferation, cytokine production andapoptosis in T cells in co-cultures with MSCs andPBMCs
Carboxyfluorescein diacetate succinimidyl ester(CFSE)-labeled (Life Technologies) PBMCs werecultured with MSCs as described above, and T-cellproliferation was determined 5 days later. Specifically,non-adherent PBMCs were harvested by gentle pipet-ting at the end of the co-culture, and T cells were sub-jected toanalysis ofCFSEfluorescence intensitybyflowcytometry. For this task, either lymphocytes were gatedon the basis of forward-scattered light (FSC) and side-scattered light (SSC) or CD4þ T cells and/or CD8þ Tcells were stainedwithCD4 (allophycocyanin [APC] orR-phycoerythrin [PE]) or CD8 (APC), respectively.Supernatantswere collected at days 2 and 5 of theMSCandPBMCco-cultures formeasurementof IL-10, IL-2and IFN-g levels. Levels were determined with the useof eBioscience (San Diego, CA, USA) Ready-Set-Goenzyme-linked immunosorbent assay (ELISA) kits,with the exception of samples from non-activated co-cultureswith PBMCs fromdonors 9, 10, 11 and 12 thatwere analyzed with R&D Systems (Minneapolis, MN,USA) Quantikine ELISA kits. Apoptosis in CD4þ Tcells was determined by analysis of the binding of APC-labeled annexin V (BD Biosciences) and CD4 (PE) asdirected by the manufacturer.
Analysis of T-cell cycle and numbers
PBMCs (unlabeled) were cultured with MSCs as indi-cated above. After co-culture, non-adherent PBMCs
were harvested by gentle pipetting, washed in coldphosphate-buffered saline with 5% FBS, labeled withCD4-APC and resuspended in cold hypotonic buffer(0.1% sodium citrate and 0.1% Triton X-100) with50 mg/mL propidium iodide (Sigma) for 20 min onice before cell cycle analysis of CD4þ T cells by flowcytometry. Absolute lymphocyte counts were deter-mined by flow cytometry with the use of Flow-countFluorospheres (Beckman Coulter, Pasenada, CA,USA) as directed by the manufacturer. Specifically,counting was performed on either lymphocytes gatedon an FSC and SSC plot or by staining of live CD4þ
T cells with a LIVE/DEAD Fixable Violet DeadCell Stain Kit (Life Technologies) and CD4-APCantibody.
Flow cytometry staining and analysis
The following antibodies specific for human mole-cules were used for flow cytometry analysis: biotin-coupled anti-CD90 (clone 5E10), CD45 (cloneHI30); fluoresceine-isothiocyanateeconjugated anti-CD105 (clone 8E11; EMD Millipore, Billerica, MA,USA); PE-conjugated anti-CD31 (clone WM-59),CD73 (clone AD2), CD4 (clone RPA-T4); PE-Cy7econjugated CD69 (eBioscience); APC-conju-gated annexin V, anti-CD4 (clone RPA-T4), CD8(clone ICRF44) and CD44 (clone G44-26) and iso-typic controls. Flow cytometry analysis was performedon 20,000 events with the use of a FACS Calibur or aFortessa cytometer, and data were analyzed with theuse of FlowJo software (Tree Star, Ashland, OR,USA). Except where indicated, antibodies, reagentsand apparatus were from BD Biosciences.
Statistical analysis
Data are reported as mean � standard deviation. Allcalculations were performed with the use of Prismsoftware (GraphPad Software, La Jolla, CA, USA).Comparisons between groups were made by meansof analysis of variance and Bonferroni post-tests.
Results
IFN-g plus TNF-aepretreated MSCs are more potentthan are naive MSCs in suppressing CD3/CD28-inducedT-cell proliferation
In the present study, we sought to analyze the ki-netics of CD3- and CD28-induced T-cell activationin the absence or presence of third-party (allogeneic)MSCs. T-cell stimulation was obtained with theaddition of 1 mg/mL anti-CD3 plus 1 mg/mL anti-CD28 soluble antibodies, which induced high levelsof CD69 upregulation in CD4þ T cells 24 h later

-
1:9
60
1:2
40
1:2
71:3
1:9
60
1:2
40
1:2
71:3
01
.2
10
06
2.5
10
06
319 320
MSC:PBMC
ratio
MSC donor
Ly
mp
ho
cy
te
co
un
tPBMC
donor 9
-
1:9
1:3
-
1:9
1:3
01
50
00
03
00
00
0
PBMC
donor 7
PBMC
donor 8
320 316
- anti-CD3/CD28
+ anti-CD3/CD28
CD
4+
T c
ell c
ou
nt
PBMC donor 1
-
315
316
319
315
316
319
0
40
80
1:9 1:3
% T
cell p
ro
liferatio
n
PBMC donor 2
-
315
316
317
319
315
316
317
319
0
40
80
1:9 1:3
% T
cell p
ro
liferatio
n
PBMC donor 2
-
304
316
0
40
80
1:9
% T
cell p
ro
liferatio
n
PBMC donor 3
-
316
319
316
319
0
40
80
1:9 1:3
% T
cell p
ro
liferatio
n
PBMC donor 3
-
304
319
304
319
0
40
80
1:9 1:3
% T
cell p
ro
liferatio
n
PBMC donor 4
-
316
319
316
319
0
40
80
1:9 1:3
% T
cell p
ro
liferatio
n
MSC:PBMC ratio
MSC donor
analysis 1
analysis 2
no MSCs IFN- +TNF- -MSCs
PBMC donor 1
-
304
316
319
304
316
319
0
50
100
1:9 1:3
% T
cell p
ro
liferatio
n
MSC:PBMC ratio
MSC donor
A
B
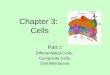
Figure 1. Suppression of CD3/CD28-induced T-cell proliferation by naive or IFN-g plus TNF-aepretreated MSCs. (A) MSCs inhibitCD3/CD28-induced T-cell expansion. PBMCs (2 � 105 cells) were cultured alone (e) or with non-treated (naive) MSCs at MSC:PBMCratios between 1:3 and 1:960 in the presence of 1 mg/mL anti-CD3 and anti-CD28. After 5 days, the absolute numbers of lymphocytesidentified on an FSC and SSC plot (left panel) or by staining live CD4þ T cells (right panel) were assessed by flow cytometry. Controlsincluded unstimulated PBMCs cultured in the absence of MSCs (white bars). (B) IFN-g plus TNF-aepretreated MSCs are more effectivethan are naive MSCs at inhibiting CD3/CD28-induced T-cell proliferation. MSCs were either untreated (naive) or pretreated with IFN-gplus TNF-a for 18 h. MSCs were then cultured with 2 � 105 CFSE-labeled PBMCs in the presence of 1 mg/mL anti-CD3 and anti-CD28antibodies. Tested MSC:PBMC ratios were 1:3 or 1:9. Five days later, lymphocytes were gated on FSC and SSC, and proliferation (startingone cycle of cell division) was analyzed by assessing CFSE levels by means of flow cytometry. Repeated fresh blood draws were obtained fromPBMC donors 1, 2 and 3 over a 6-month time lapse; results obtained with two of these blood draws are shown in analysis 1 and analysis 2.
194 J. Cuerquis et al.
(Supplementary Figure 1). We observed that evenlow numbers of untreated (naive) MSCs in co-culturewith PBMCs inhibited the expansion of activatedlymphocytes or live CD4þ T cells induced by thisstrong immune signal as revealed by flow cytometricabsolute cell counting (Figure 1A). Specifically, withall tested PBMC (n ¼ 4) and MSC (n ¼ 1e3 perPBMC donor) donors, a 14.8 � 7.24efold (P <0.001) and 1.9 � 0.51efold (P < 0.001) decrease inlymphocyte numbers at day 5 of the co-culture wereobserved in the presence of MSCs at MSC:PBMCratios of 1:3 and 1:240, respectively (Figure 1A andSupplementary Figure 2). MSCs had no significanteffects on CD3/CD28-induced lymphocyte expansion
at MSC:PBMC ratios of 1:480 and/or 1:960 (data notshown and Supplementary Figure 2).
To investigate the effect of naive MSCs or IFN-gplus TNF-aepretreated MSCs on T-cell prolifera-tion, MSCs were co-cultured with PBMCs labeledwith CFSE to track cell division. CFSElow prolifer-ating T cells were defined as lymphocytes that un-derwent at least one cycle of cell division. PBMCpreparations from seven donors were tested; someanalyses could be repeated on separate consecutivefresh blood draws from PBMC donors 1, 2 and 3over a 6-month lapse (analyses 1 and 2 in Figure 1B).T-cell proliferation in all PBMC donor preparationscould be induced on T-cell receptor and CD28

MSCs support cytokine production by activated T cells 195
engagement and was inhibited by naive MSCs at anMSC:PBMC ratio of 1:3. Experiments displayed inFigure 1B with individual values obtained withPBMC donors 1e4 showed a 4.0 � 3.4efoldreduction in levels of T-cell proliferation with naiveMSCs at an MSC:PBMC ratio of 1:3 compared withthe absence of MSCs (P < 0.001). The reduction inthe percentages of CFSElow T cells was less signifi-cant at an MSC:PBMC ratio of 1:9 compared with1:3 (P < 0.01, Supplementary Figure 3). However,the analysis of CFSE peaks revealed that activated Tcells still underwent fewer cycles of cell division at anMSC:PBMC ratio of 1:9 compared with the absenceof MSCs (data not shown). This probably accountedfor the low absolute numbers of lymphocytes andCD4þ T cells observed in CD3/CD28-activated co-cultures at MSC:PBMC ratios >1:240 (P < 0.001,Figure 1A). With IFN-g plus TNF-aepretreatedMSCs compared with naive MSCs at anMSC:PBMC ratio of 1:3, T-cell proliferationmeasured also by CFSE intensity was furtherdecreased significantly by 0.9-fold to 31.5-fold (P <0.01, Figure 1B and Supplementary Figure 3), withthe exception of PBMC donor 2. Hence, MSCseffectively inhibit CD3/CD28-induced T-cell prolif-eration, especially when pretreated with IFN-g plusTNF-a.
Naive MSCs induce an early surge in IFN-g and IL-2but not IL-10 production by CD3/CD28-activatedPBMCs, an effect that is reduced by pretreatment ofMSCs with IFN-g plus TNF-a
The effect of MSCs on the synthesis of IL-2, IFN-gand IL-10 by activated T cells was assessed byELISA analysis of supernatants from unfractionedPBMCs co-cultured with MSCs in the presence ofanti-CD3 plus anti-CD28 at days 2 and 5. Super-natants from naive or IFN-g plus TNF-aepretreatedMSCs cultured alone for 2 or 5 days did not revealdetectable levels of IL-2, IFN-g and IL-10 (data notshown). In PBMCs cultured alone, the activation ofT cells with CD3/CD28 induced the production ofIL-2 and IFN-g at variable levels at days 2 and 5,depending on PBMC donors (Figure 2A). Concor-dant with the observed inhibition of T-cell prolifer-ation, production levels of these cytokines were inmost cases reduced at day 5 in the presence of MSCs(Figure 2A, bottom panel). Nevertheless, at earlytimes of the co-culture, naive MSCs unexpectedlyfurther increased CD3/CD28-induced IL-2 andIFN-g production by PBMCs (Figure 2A top panel).Specifically, a 6.0 � 4.5efold (2.0e14.3) increase inIFN-g levels was observed at day 2 in co-cultureswith naive MSCs and PBMCs from donor 1 (P <0.001), donor 2 (P < 0.01), donor 3 (P < 0.05) and
donor 4 (P < 0.05) at an MSC:PBMC ratio of 1:3compared with levels in the absence of MSCs. A 1.5-fold to 5.4-fold increase in IL-2 levels at day 2 wasobserved in co-cultures of naive MSCs with PBMCsfrom donor 1 (P < 0.001), donor 2 (P < 0.001, withthe exception of MSC donor 317) and donor 4 (P <0.05). Levels of IL-2 in PBMCs from donor 3 wereunaffected by MSCs. Experiments each includingtwo to three MSC donors were repeated with PBMCdonors 1, 2 and 3 at MSC:PBMC ratios of 1:9 or1:3, and data showed an increase in IFN-g and IL-2production by T cells at day 2 in the presence ofnaive MSCs (Supplementary Figure 4 and data notshown). This early increased production of IFN-gand IL-2 by activated PBMCs was on the other handreduced with IFN-g plus TNF-aepretreated MSCsat an MSC:PBMC ratio of 1:3 (Figure 2A) andin most instances at ratio of 1:9 (SupplementaryFigure 4).
Significant differences in the individual levels ofIL-10 produced by CD3/CD28-activated PBMCswere observed. PBMCs from donors 1 and 2 pro-duced low levels of IL-10 at days 2 and 5 on CD3/CD28-mediated activation in the absence or pres-ence of MSCs (data not shown). Activated PBMCsfrom donors 3, 4, 9, 10, 11 and 12 produced sig-nificant levels of IL-10 at days 2 and 5. Naive MSCshad no effect on IL-10 production by PBMCs fromdonor 3 and induced a modest increase of IL-10synthesis by PBMCs from donor 4 at day 5 (data notshown). At days 2 and 5, naive MSCs did not affectCD3/CD28-induced IL-10 production in PBMCsfrom donors 11 and 12 but decreased productionlevels in PBMCs from donors 9 and 10 (Figure 2B).Cytokine-pretreated MSCs did not further decreaseIL-10 levels in CD3/CD28-activated co-culturescompared with naive MSCs at days 2 and 5(Figure 2B).
In brief, naive MSCs appear to effectively inhibitCD3/CD28-induced T-cell proliferation; neverthe-less, this suppression is preceded by a surge in theproduction of IFN-g and IL-2 but not IL-10 byCD3/CD28-activated PBMCs at early times of theco-culture. Priming of MSCs with IFN-g plus TNF-a reduced the early surge in IFN-g and IL-2 pro-duction by CD3/CD28-activated PBMCs.
In non-activated PBMC cultures, naive and IFN-g plusTNF-aepretreated MSCs sustain the survival in vitro oflymphocytes and induce low levels of CD69 expression inT cells and low levels of IL-2 and IL-10 but not IFN-gsynthesis by PBMCs
We next investigated whether or not naive or IFN-gplus TNF-aepretreated MSCs displayed immune-regulatory properties toward T cells in unfractioned

-
315
316
319 -
315
316
317
319 -
316
319 -
316
319
0
350
700
IFN- day 2
PBMC
donor 1
PBMC
donor 2
PBMC
donor 3
PBMC
donor 4
MSC
donor
IFN
- (p
g/m
l)
-
315
316
319 -
315
316
317
319 -
316
319 -
316
319
0
200
400
PBMC
donor 1
PBMC
donor 2
PBMC
donor 3
PBMC
donor 4
MSC
donor
IL-2 day 2
IL-2
(p
g/m
l)
-
319
320 -
319
320
304 -
320
304 -
319
320
304
0
750
1500
PBMC
donor 9
PBMC
donor 10
PBMC
donor 11
PBMC
donor 12
MSC
donor
IL-10 day 2
IL-1
0 (p
g/m
l)
-
315
316
319 -
315
316
317
319 -
316
319 -
316
319
0
350
700
PBMC
donor 1
PBMC
donor 2
PBMC
donor 3
PBMC
donor 4
MSC
donor
IFN- day 5
IFN
- (p
g/m
l)
-
315
316
319 -
315
316
317
319 -
316
319 -
316
319
0
200
400
PBMC
donor 1
PBMC
donor 2
PBMC
donor 3
PBMC
donor 4
MSC
donor
IL-2 day 5
IL-2
(p
g/m
l)
-
319
320 -
319
320
304 -
320
304 -
319
320
304
0
750
1500
PBMC
donor 9
PBMC
donor 10
PBMC
donor 11
PBMC
donor 12
MSC
donor
IL-10 day 5
IL-1
0 (p
g/m
l)
no MSCs naive MSCs IFN- +TNF- -MSCs
no MSCs naive MSCs IFN- +TNF- -MSCs
A
B
Figure 2. Naive MSCs but not IFN-g plus TNF-aepretreated MSCs increase early cytokine production by CD3/CD28-activated PBMCs.MSCs either untreated (naive) or pretreated with IFN-g plus TNF-a for 18 h were cultured with 2 � 105 PBMCs in the presence of 1 mg/mLanti-CD3 and anti-CD28 at an MSC:PBMC ratio of 1:3. Two and 5 days later, levels of (A) IFN-g, IL-2 (PBMCs from donors 1, 2, 3and 4) and (B) IL-10 (PBMCs from donors 9, 10, 11 and 12) in the co-culture supernatant were quantified by means of eBioscience Ready-Set-Go ELISA systems. Figure 2A shows IFN-g and IL-2 levels in co-cultures from the experiments displayed as analysis 1 in Figure 1B.
196 J. Cuerquis et al.
non-activated PBMC cultures by assessing T-cellnumbers, the expression of the early T-cell activationmarker CD69 and cytokine production at anMSC:PBMC ratio of 1:3. After 2 and 5 days of co-culture of non-activated PBMCs (n ¼ 3 donors) withnaive or IFN-g- plus TNF-aepretreated MSCs(n ¼ 3 donors), the analysis of CFSE levels in
CFSE-labeled PBMCs, manual cell counting ofnon-adherent live PBMCs by trypan blue stainingand flow cytometryebased counting of live CD4þ
T cells revealed no significant change in lymphocyteproliferation or absolute numbers (data not shown).At days 11e15, decreased loss of lymphocytes orCD4þ T cells from 3 of 4 tested donor PBMCs were

-
319
320 -
304
320 -
304
319
320
0
30000
60000
MSC donor
PBMC
donor 11
PBMC
donor 12
PBMC
donor 9
Lym
ph
ocyte c
ou
nt
-
316 -
304
0
25000
50000
no MSCs
naive MSCs
IFN- +TNF- -MSCs
MSC donor
PBMC
donor 9
PBMC
donor 10
CD
4+ T
ce
ll c
ou
nt
IFN- day 2
no M
SCs
MSCs
I+T M
SCs
0.0
12.5
25.0
IFN
- (
pg
/ml)
IL-2 day 2
no M
SCs
MSCs
I+T M
SCs
0.0
12.5
25.0
IL-2
(p
g/m
l)
IL-10 day 2
no M
SCs
MSCs
I+T M
SCs
0
20
40
IL-1
0 (
pg
/ml)
IFN- day 5
no M
SCs
MSCs
I+T M
SCs
0.0
12.5
25.0
IFN
- (
pg
/ml)
IL-2 day 5
no M
SCs
MSCs
I+T M
SCs
0.0
12.5
25.0
IL-2
(p
g/m
l)
IL-10 day 5
no M
SCs
MSCs
I+T M
SCs
0
20
40
IL-1
0 (
pg
/ml)
-
MSCs
I+T M
SCs
CD3/C
D28
0
50
100
CD
4+ T
ce
lls
exp
ressin
g C
D69 (
%)
no M
SCs
MSCs
I+T M
SCs
CD3/C
D28
0
500
2000
7000
CD
69
ex
pre
ss
ion
le
ve
ls
(M
FI)
A B
C
Figure 3. In non-activated PBMC cultures, naive and IFN-g plus TNF-aepretreated MSCs sustain the survival in vitro of lymphocytesand induce low levels of CD69 expression in T cells and low levels of IL-2 and IL-10 but not IFN-g synthesis by PBMCs. MSCs eitheruntreated (naive) or pretreated with IFN-g plus TNF-a for 18 h were cultured with 2 � 105 unlabeled PBMCs in the absence of anti-CD3and -CD28 antibodies unless otherwise indicated. (A) MSCs reduce lymphocyte loss in culture. PBMCs were co-cultured with MSCs atan MSC:PBMC ratio of 1:3 with the exception of PBMCs from donor 10 (ratio 1:9). Eleven to 15 days later, the absolute numbers oflymphocytes gated on the basis of FSC and SSC or live CD4þ T cells were assessed by flow cytometry. (B) MSCs induce low levels ofCD69 expression in T cells. PBMCs from donor 8 (tested with MSCs from donor 320), PBMCs from donor 10 (tested with MSCs fromdonors 304 and 320), PBMCs from donor 11 (tested with MSCs from donors 304 and 320) and PBMCs from donor 12 (tested withMSCs from donors 320, 304 and 319) were cultured alone, with naive MSCs or with IFN-g plus TNF-a-pretreated MSCs (I/T MSCs) atan MSC:PBMC ratio of 1:3 for 24 h. The percentage of CD4þ T cells expressing CD69 (top panel) and CD69 median fluorescenceintensity in CD4þ T cells (bottom panel) were determined by flow cytometry. Positive controls for CD69 expression in CD4þ T cellsconsisted of PBMCs activated with 1 mg/mL anti-CD3 and anti-CD28 (CD3/CD28) in the absence of MSCs. (C) IFN-g, IL-2 and IL-10levels in non-activated co-cultures. PBMCs from donor 9 (tested with MSCs from donor 320 for IL-2 and IL-10 and donor 319 for allcytokines) and PBMCs from donors 10, 11 and 12 (tested with MSCs from donors 304 and 320 for all cytokines) were co-cultured or notwith naive MSCs or IFN-g plus TNF-aepretreated MSCs (I/T MSCs) at an MSC:PBMC ratio of 1:3 for 2 or 5 days. Supernatants wereharvested and quantified for levels of IFN-g, IL-2 and IL-10 by Quantikine ELISA assays (R&D systems). Positive controls for cytokineproduction consisted of supernatants from PBMCs from donors 9e12 activated for 2 days with 1 mg/mL anti-CD3 and anti-CD28 in theabsence of MSCs that displayed 787 � 517 pg/mL, 131 � 75 pg/mL and 872 � 373 pg/mL IFN-g, IL-2 and IL-10, respectively.
MSCs support cytokine production by activated T cells 197

Figure 4. Naive and IFN-g plus TNF-aepretreated MSCs decrease apoptosis in non-activated and CD3/CD28-activated T cells and retainT cells in the G1 phase of the cell cycle. (A) MSCs inhibit apoptosis in T cells. Naive or IFN-g plus TNF-aepretreated MSCs were culturedwith CFSE-labeled PBMCs as explained in Figure 1B legend at an MSC:PBMC ratio of 1:3. Co-cultures were then activated (left panel) ornot activated (right panel) with 1 mg/mL anti-CD3 and anti-CD28 antibodies. Five days later, apoptosis in CD4þ T cells was analyzed byassessing Annexin V binding by flow cytometry. The experiment was also repeated with PBMCs from donor 1 and MSCs from donors 316or 319, and similar trends of responses were observed. (B) Flow cytometry profiles of Annexin V and CFSE staining in non-activated orCD3/CD28-activated CD4þ T cells from PBMCs from donor 1 after 5 days of culture with MSCs from donor 304 either untreated (naive)or pretreated with IFN-g plus TNF-a (I/T MSCs) at MSC:PBMC ratios of 1:3 or 1:9. (C) Cell cycle analysis. MSCs from donor 319, eithernaive (319) or pretreated with IFN-g plus TNF-a (I/T 319), were cultured with PBMCs from donor 3. Alternatively, MSCs from donors
198 J. Cuerquis et al.

MSCs support cytokine production by activated T cells 199
observed in co-cultures with naive and/or IFN-g plusTNF-aepretreated MSCs (P < 0.05, Figure 3A).
Analysis of CD69 expression on CD4þ T cellsfrom non-activated co-cultures was performed byflow cytometry on four donor PBMC preparations,each tested with one to three donor MSC prepara-tions. IFN-g plus TNF-aepretreated MSCs and to alesser extent naive MSCs co-cultured with restingPBMCs induced CD69 expression in 47.0% � 14.2%and 17.7% � 7.4% of CD4þ T cells, respectively, asearly as day 1 (Figure 3B, top panel) and maintainedat day 3 (data not shown). Expression of CD69 wasalso observed as upregulated in lymphocytes that didnot express CD4 (data not shown). However, levelsof CD69 expression in T cells from non-activatedPBMCs co-cultured with MSCs were significantlylower than levels induced by CD3/CD28-mediatedactivation (P < 0.001, Figure 3B, bottom panel).Specifically, the median fluorescence intensity (MFI)of CD69 staining in CD4þ T cells at day 1 were 8� 2,100 � 26, 237 � 80 and 3872 � 1542 in non-activated PBMC cultures alone, with naive MSCs,with IFN-g plus TNF-aepretreated MSCs or acti-vated anti-CD3/CD28 PBMCs alone, respectively.
Analysis of IFN-g, IL-2 and IL-10 levels in su-pernatants from co-cultures with non-activatedPBMCs from donors 1e4 (data not shown) anddonors 9e12 (Figure 3C) revealed undetectable orlow cytokine levels by ELISA. Compared with non-activated PBMCs alone, naive MSCs and IFN-gplus TNF-aepretreated MSCs had no effect onIFN-g production but similarly increased the pro-duction of IL-10 at days 2 and 5 (P < 0.05), whereasonly naive MSCs increased the production of IL-2 atday 5 (P < 0.05). However, levels of IL-2 and IL-10induced by MSCs (11 � 7 pg/mL and 19 � 5 pg/mL,respectively, at day 5 with naive MSCs, Figure 3C)were significantly lower than levels produced by fullyactivated T cells from the same donors. Specifically,CD3/CD28-activated PBMCs cultured alone andharvested at day 2 from the same experiments dis-played a production of 131 � 75 pg/mL and 872 �373 pg/mL of IL-2 and IL-10, respectively. IFN-g,IL-2 and IL-10 were not detected in supernatantsfrom naive or IFN-g plus TNF-aepretreated MSCdonors cultured alone for 2 or 5 days (data notshown).
Hence, in initially non-activated PBMCs, naiveand IFN-g plus TNF-aepretreated MSCs induce lowlevels of CD69 expression in T cells in the absence oflymphocyte proliferation or IFN-g secretion, whereas
304, 316 or 319 were cultured with PBMCs from donor 1. MSC:PBMCactivated (top panels) with 1 mg/mL anti-CD3 and anti-CD28 antibodiesby means of flow cytometry.
the synthesis of IL-2 and IL-10 by PBMCs is in-creased at low levels.
Naive and IFN-g plus TNF-aepretreated MSCs exhibitcomparable ability to inhibit apoptosis in resting andproliferating T cells and to retain quiescent T cells into theG0/G1 phase of the cell cycle
We compared the capacity of naive and IFN-g plusTNF-aepretreated MSCs to regulate apoptosis in Tcells at rest or on T-cell receptor and CD28 engage-ment. For this task, we measured annexin V binding inCFSE-labeled T cells by flow cytometry after 5 days inco-culture. The percentage of apoptosis in both restingand CD3/CD28-activated CD4þ T cells (Figure 4A)or CD8þ T cells (data not shown) was reduced in co-cultures with MSCs at MSC:PBMC ratios of 1:3 (P <0.001, Figure 4A) or 1:9 (data not shown), and nostatistical differences were observed between naive andIFN-g plus TNF-aepretreated MSCs for the reduc-tion of T-cell apoptosis. Reduction of annexin Vþ
CD4þ T cells was observed in both CFSElow dividingcells and CFSEhi non-dividing cells (Figure 4B).These results suggest that MSCs, naive or pretreatedwith IFN-g plus TNF-a, inhibited apoptosis in restingand dividing T cells. Cell cycle analysis performed atdays 3 and 5 or 6 revealed that in the absence ofMSCs, resting CD4þ T cells entered progressivelyinto the sub-G0/G1 phase from the G0/G1 phase,which suggests the induction of apoptosis during invitro culture (Figure 4C). In CD3/CD28-activatedCD4þ T cells, entry into the S phase was detected atday 3, and, at day 5 or 6 the percentage of apop-totic cells in the sub-G0/G1 phase was significantlyincreased. At day 5 or 6 and in the absence of MSCs,only 26e38% of CD3/CD28-activated CD4þ T cellswere in G0/G1 phase, compared with the 63e89% ofresting CD4þ T cells. In contrast, at day 3 of the co-culture, naive and IFN-g plus TNF-aepretreatedMSCs effectively reduced the entry of CD3/CD28-activated CD4þ T cells into the S phase. In addition,MSCs inhibited the entry of CD3/CD28-activated andto a lesser extent of resting CD4þ T cells into the sub-G0/G1 phase at day 5 or 6 of the co-culture.
Altogether, these data suggest that MSCs retainactivated T cells into the G0/G1 phase and inhibitapoptosis in resting and proliferating T cells. Treat-ment of MSCs with IFN-g plus TNF-a further in-creases their capacity to inhibit transition of activatedT cells into the S phase while preserving their anti-apoptotic functions.
ratios were 1:3. Co-cultures were activated (bottom panels) or not. Cell cycle analysis of CD4þ T cells was performed 3e6 days later

-
303
304
305
306
307
311
0
60
120
no inhibitor
MSC
donor
no MSCs
naive MSCs
IFN- +TNF- -MSCs
-
303
304
305
306
307
311
0
60
1201-MT
MSC
donor
Indomethacin
200 J. Cuerquis et al.
In the absence of IDO activity, naive and IFN-g plusTNF-aepretreated MSCs can support CD3- andCD28-induced T-cell proliferation
Several authors have shown that IDO and PGE2 pro-duced by IFN-geactivated MSCs are essential forsuppression of CD3-mediated T-cell proliferation(5,20,21). Herein, we tested effects of blockage of IDOor PGE2 activity in co-cultures of PBMCs and naive orIFN-g plus TNF-aeactivated MSCs in three donorPBMC preparations (donors 5, 6 and 7). We observedthat CD3/CD28-stimulated T cells were sensitive toMSC-mediated immune suppression by IDO and notPGE2 activity because reversionof immune suppressionby MSCs was observed with the IDO inhibitor 1-MT(P< 0.001with either naiveMSCsor IFN-gplusTNF-aeactivated MSCs) and not the COX2 and PGE2 in-hibitor indomethacin (PBMCs fromdonor 5, Figure5).In the presence of 1-MT, levels of T-cell proliferation inco-cultures with naive MSCs or IFN-g plus TNF-aeactivated MSCs were higher than levels observed intheabsenceofMSCs.Similar resultswereobservedwithtwo other PBMC donors (donors 6 and 7, data notshown).No significant cell toxicity was reported with 1-MT or indomethacin with either MSCs or PBMCs, astested with a cell viability assay with the use of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide(MTT) (data not shown).These results show thatwhenIDO activity is blocked, naive and IFN-g plus TNF-aeactivated MSCs can increase T-cell proliferation.
-
303
304
305
306
307
311
0
60
120
MSC
donor
Figure 5. Inhibition of IDO enables naive or IFN-g plus TNF-aepretreated MSCs to increase CD3/CD28-induced T-cell pro-liferation. Naive or IFN-g plus TNF-aepretreated MSCs werecultured with CFSE-labeled PBMCs from donor 5 at anMSC:PBMC ratio of 1:3 in the presence or absence of 1-MT orindomethacin, and T-cell proliferation was analyzed 5 days later.Co-cultures were activated with 1 mg/mL anti-CD3 and anti-CD28antibodies. This experiment was also performed at anMSC:PBMCratio of 1:9 and repeated with the same MSC donors and PBMCdonors 6 and 7 at MSC:PBMC ratios of 1:9 and 1:3, and similartrends of responses were obtained.
Discussion
We report for the first time that human bone marrowederived MSCs induce a surge in IFN-g and IL-2 pro-duction by CD3/CD28-activated PBMCs at earlystages of their interaction with PBMCs before sup-pressing activated T-cell proliferation. Specifically, 2.0-fold to14.3-fold and1.5-fold to5.4-fold increases in theproduction of IFN-g and IL-2, respectively, wereobserved in supernatants after 2 days in co-cultures ofPBMCs with all MSC donors at high MSC-to-PBMCratios, an effect that was significantly less pronouncedafter pretreatment of MSCs with IFN-g and TNF-a.We further demonstrate that pretreatment of MSCswith IFN-g plus TNF-a increases their anti-prolifera-tive capacity toward activated T cells in a mechanismthat preserves their anti-apoptotic functions. These re-sults suggest that unprimed MSCs induce a transientincrease in IFN-g and IL-2 synthesis by activatedTcellsbefore immune licensing by activated PBMC-derivedcytokines that induce the expression of immune sup-pressive molecules by MSCs such as IDO that blockT-cell proliferation. In support of this hypothesis, inhi-bition of IDO activity in co-cultures of activated T cellsandMSCs,whethernaiveorpretreatedwith IFN-gplus
TNF-a, results in levels of T-cell proliferation higherthan levels observed in the absence of MSCs.
In non-activated PBMC cultures, low levels ofIL-2 and IL-10 synthesis were observed with MSCs

MSCs support cytokine production by activated T cells 201
in addition to low levels of CD69 expression by Tcells in the absence of T-cell proliferation. Thissuggests that MSCs support a modest activation ofinitially resting PBMCs rather than a primary stim-ulatory full immune response followed with aninhibitory response. Crop et al. (10) showed thatadipose tissueederived MSCs express the immunesuppressive factor IDO after 7 days of co-culturewith unstimulated PBMCs and that their separationresults in a phase of active T-cell proliferation. It islikely that MSCs and initially resting PBMCs displaycomplex reciprocal actions, and therefore furtherstudies are required for clarification.
Independent studies reported that MSCs inducethe differentiation of monocytes, dendritic cellsand macrophages and into IL-10esecreting cells(6,22,23). This process appears to be central to thesuppression of T-cell activation (6) and the reductionof inflammation (23). Accordingly, we observed thatnaive MSCs support low levels of IL-10 production innon-activated PBMC cultures; interestingly this pro-priety was not lost on IFN-g plus TNF-a pretreat-ment. The effect ofMSCs on IL-10 synthesis byT cellsis less apprehended. In vitro, MSCs were shown toincrease IL-10 secretion by PBMCs activated withanti-CD3/CD28 antibodies or in allogeneic reactionsbut also to decrease IL-10 production by IL-2/CD3-activated PBMCs (24,25), whereas they augment thefrequency of IL-10eproducing T cells in vivo (26). InCD3/CD28-activated PBMC cultures, the overall IL-10 levels were observed to be unaffected or reduced bynaive MSCs. Of note, cytokine-pretreated MSCs ten-ded to preserve IL-10 production by CD3/CD28-activated PBMCs compared with naive MSCs.
The early increased activation of T cells byMSCsmay be caused by factors secreted by MSCs and/orthe recognition of allogeneic MHC molecules onMSCs. In addition, distinct factors produced byimmune-licensed MSCs may be involved in the lateinhibition of proliferation and cytokine productionof activated T cells. MSCs produce a variety of pro-survival factors such as IL-6, vascular endothelialgrowth factor (VEGF), basic fibroblast growth fac-tor, hepatocyte growth factor, insulin-like growthfactor-1, stromal cellederived factor-1, and platelet-derived growth factor. Some of these molecules acton both the activation and survival of immune cells.As an example, IL-6 is a well-known immunemodulator that also inhibits apoptosis in antigen-stimulated T cells (27) and resting T cells by sus-taining the expression of the anti-apoptotic moleculeBcl-2 (28). Inhibition of IL-6 produced by MSCsresults in an additional decrease in the proliferationof activated T lymphocytes in vitro in co-cultureswith MSCs (8,29) and increases apoptosis in neu-trophils (7). IFN-g and TNF-a were described as
major regulators for the expression of chemokines(IL-8, chemokine [C-C motif] ligand [CCL]2,CCL3, CCL5, CCL7 and CCL8), interleukins (IL-1b, IL-6 and IL-7), immune suppressive molecules(PGE2, IDO) and factors regulating angiogenesisand apoptosis (VEGF) (4,6,30,31). Whereasincreased expression of IDO followed by tryptophandepletion and accumulation of its catabolites wereshown to sensitize lymphocytes to apoptosis (32),other molecules with increased expression in cyto-kine-activated MSCs, such as PGE2, VEGF or IL-6,may counteract these pro-apoptotic effects.
Animal models of immune ailments have shownthat the timing ofMSC administration influences theiroutcomes (33). MSCs appear to act optimally ifinjected during a narrow window of disease develop-ment when sufficient concentration of inflammatorycytokines are present for the immune licensing ofMSCs but also when damages are not too advanced fortherapeutic intervention (3,34e36).We showed hereinthat MSCs exert dynamic actions on T-cell activationand that pretreatment of MSCs with IFN-g and TNF-a results in increased expression of factors that blockT-cell proliferation and counteract the boosting effectsof MSCs on the early IL-2 and IFN-g production byactivated T cells. In this regard, priming ofMSCs withIFN-g plus TNF-a may increase their effectivenessand safety in vivo at the early disease stage.
Acknowledgments
The authors thank John Antoniou and FacksonMwale (McGill University, Jewish General Hospital)for the bone marrow biopsies from individuals un-dergoing hip surgery and Jacques Galipeau (EmoryUniversity) for helpful discussions. This work wassupported by the Canadian Institutes of HealthResearch-CIHR Operating Grant MOP-15017.
Disclosure of interests: The authors have nocommercial, proprietary, or financial interest in theproducts or companies described in this article.
References
1. Newman RE, Yoo D, LeRoux MA, Danilkovitch-Miagkova A.Treatment of inflammatory diseases with mesenchymal stemcells. Inflamm Allergy Drug Targets. 2009;8:110e23.
2. Renner P, Eggenhofer E, Rosenauer A, Popp FC,Steinmann JF, Slowik P, et al. Mesenchymal stem cells requirea sufficient, ongoing immune response to exert their immu-nosuppressive function. Transplant Proc. 2009;41:2607e11.
3. Polchert D, Sobinsky J, Douglas G, Kidd M, Moadsiri A,Reina E, et al. IFN-gamma activation of mesenchymal stemcells for treatment and prevention of graft versus host disease.Eur J Immunol. 2008;38:1745e55.

202 J. Cuerquis et al.
4. Prasanna SJ, Gopalakrishnan D, Shankar SR, Vasandan AB.Pro-inflammatory cytokines, IFNgamma and TNFalpha, in-fluence immune properties of human bone marrow andWharton jelly mesenchymal stem cells differentially. PLoSOne. 2010;5:e9016.
5. Ren G, Su J, Zhang L, Zhao X, Ling W, L’Huillie A, et al.Species variation in the mechanisms of mesenchymal stem cell-mediated immunosuppression. Stem Cells. 2009;27:1954e62.
6. Francois M, Romieu-Mourez R, Li M, Galipeau J. HumanMSC suppression correlates with cytokine induction of indo-leamine 2,3-dioxygenase and bystander M2 macrophage dif-ferentiation. Mol Ther. 2012;20:187e95.
7. Raffaghello L, Bianchi G, Bertolotto M, Montecucco F,Busca A, Dallegri F, et al. Human mesenchymal stem cellsinhibit neutrophil apoptosis: a model for neutrophil preserva-tion in the bone marrow niche. Stem Cells. 2008;26:151e62.
8. Najar M, Rouas R, Raicevic G, Boufker HI, Lewalle P,Meuleman N, et al. Mesenchymal stromal cells promote orsuppress the proliferation of T lymphocytes from cord bloodand peripheral blood: the importance of low cell ratio and roleof interleukin-6. Cytotherapy. 2009;11:570e83.
9. Rasmusson I, Le Blanc K, Sundberg B, Ringden O. Mesen-chymal stem cells stimulate antibody secretion in human Bcells. Scand J Immunol. 2007;65:336e43.
10. Crop MJ, Baan CC, Korevaar SS, Ijzermans JN, Weimar W,Hoogduijn MJ. Human adipose tissue-derived mesenchymalstem cells induce explosive T-cell proliferation. Stem CellsDev. 2010;19:1843e53.
11. Tokoyoda K, Hauser AE, Nakayama T, Radbruch A. Orga-nization of immunological memory by bone marrow stroma.Nat Rev Immunol. 2010;10:193e200.
12. Glennie S, Soeiro I, Dyson PJ, Lam EW, Dazzi F. Bonemarrow mesenchymal stem cells induce division arrest anergyof activated T cells. Blood. 2005;105:2821e7.
13. Benvenuto F, Ferrari S, Gerdoni E, Gualandi F, Frassoni F,Pistoia V, et al. Human mesenchymal stem cells promotesurvival of T cells in a quiescent state. Stem Cells. 2007;25:1753e60.
14. Giuliani M, Fleury M, Vernochet A, Ketroussi F, Clay D,Azzarone B, et al. Long-lasting inhibitory effects of fetal livermesenchymal stem cells on T-lymphocyte proliferation. PLoSOne. 2011;6:e19988.
15. Zappia E, Casazza S, Pedemonte E, Benvenuto F, Bonanni I,Gerdoni E, et al. Mesenchymal stem cells ameliorate experi-mental autoimmune encephalomyelitis inducing T-cellanergy. Blood. 2005;106:1755e61.
16. Plumas J, Chaperot L, Richard MJ, Molens JP, Bensa JC,Favrot MC. Mesenchymal stem cells induce apoptosis ofactivated T cells. Leukemia. 2005;19:1597e604.
17. Cutler AJ, Limbani V, Girdlestone J, Navarrete CV. Umbilicalcord-derived mesenchymal stromal cells modulate monocytefunction to suppress T cell proliferation. J Immunol. 2010;185:6617e23.
18. Wang D, Chen K, Du WT, Han ZB, Ren H, Chi Y, et al.CD14þ monocytes promote the immunosuppressive effect ofhuman umbilical cord matrix stem cells. Exp Cell Res. 2010;316:2414e23.
19. Hof-Nahor I, Leshansky L, Shivtiel S, Eldor L, Aberdam D,Itskovitz-Eldor J, et al. Human mesenchymal stem cells shiftCD8þ T cells towards a suppressive phenotype by inducingtolerogenic monocytes. J Cell Sci. 2012;125:4640e50.
20. Meisel R, Zibert A, Laryea M, Gobel U, Daubener W,Dilloo D. Human bone marrow stromal cells inhibit alloge-neic T-cell responses by indoleamine 2,3-dioxygenase-medi-ated tryptophan degradation. Blood. 2004;103:4619e21.
21. Ryan JM, Barry F, Murphy JM, Mahon BP. Interferon-gamma does not break, but promotes the immunosuppressive
capacity of adult human mesenchymal stem cells. Clin ExpImmunol. 2007;149:353e63.
22. Aggarwal S, Pittenger MF. Human mesenchymal stem cellsmodulate allogeneic immune cell responses. Blood. 2005;105:1815e22.
23. Nemeth K, Leelahavanichkul A, Yuen PS, Mayer B,Parmelee A, Doi K, et al. Bone marrow stromal cells attenuatesepsis via prostaglandin E(2)-dependent reprogramming ofhost macrophages to increase their interleukin-10 production.Nat Med. 2009;15:42e9.
24. Gieseke F, Bohringer J, Bussolari R, Dominici M,Handgretinger R, Muller I. Human multipotent mesenchymalstromal cells employ galectin-1 to inhibit immune effectorcells. Blood. 2010;116:3770e9.
25. Batten P, Sarathchandra P, Antoniw JW, Tay SS,Lowdell MW, Taylor PM, et al. Human mesenchymal stemcells induce T cell anergy and downregulate T cell allo-re-sponses via the TH2 pathway: relevance to tissue engineeringhuman heart valves. Tissue Eng. 2006;12:2263e73.
26. Jitschin R, Mougiakakos D, Von Bahr L, Volkl S, Moll G,Ringden O, et al. Alterations in the cellular immunecompartment of patients treated with third-party mesen-chymal stromal cells following allogeneic hematopoietic stemcell transplantation. Stem Cells. 2013;31:1715e25.
27. Rochman I, Paul WE, Ben-Sasson SZ. IL-6 increases primedcell expansion and survival. J Immunol. 2005;174:4761e7.
28. Teague TK, Marrack P, Kappler JW, Vella AT. IL-6 rescuesresting mouse T cells from apoptosis. J Immunol. 1997;158:5791e6.
29. Francois M, Copland IB, Yuan S, Romieu-Mourez R,Waller EK, Galipeau J. Cryopreserved mesenchymal stromalcells display impaired immunosuppressive properties as aresult of heat-shock response and impaired interferon-gammalicensing. Cytotherapy. 2012;14:147e52.
30. Croitoru-Lamoury J, Lamoury FM, Zaunders JJ, Veas LA,Brew BJ. Human mesenchymal stem cells constitutively ex-press chemokines and chemokine receptors that can be upre-gulated by cytokines, IFN-beta, and Copaxone. J InterferonCytokine Res. 2007;27:53e64.
31. Liu Y, Han ZP, Zhang SS, Jing YY, Bu XX, Wang CY, et al.Effects of inflammatory factors on mesenchymal stem cellsand their role in the promotion of tumor angiogenesis in coloncancer. J Biol Chem. 2011;286:25007e15.
32. Mulley WR, Nikolic-Paterson DJ. Indoleamine 2,3-dioxyge-nase in transplantation. Nephrology. 2008;13:204e11.
33. Krampera M. Mesenchymal stromal cell ‘licensing’: a multi-step process. Leukemia. 2011;25:1408e14.
34. Yanez R, Lamana ML, Garcia-Castro J, Colmenero I,Ramirez M, Bueren JA. Adipose tissue-derived mesenchymalstem cells have in vivo immunosuppressive properties appli-cable for the control of the graft-versus-host disease. StemCells. 2006;24:2582e91.
35. Le Blanc K, Frassoni F, Ball L, Locatelli F, Roelofs H,Lewis I, et al. Mesenchymal stem cells for treatment ofsteroid-resistant, severe, acute graft-versus-host disease: aphase II study. Lancet. 2008;371:1579e86.
36. Chang YS, Choi SJ, Ahn SY, Sung DK, Sung SI, Yoo HS,et al. Timing of umbilical cord blood derived mesenchymalstem cells transplantation determines therapeutic efficacy in theneonatal hyperoxic lung injury. PLoS One. 2013;8:e52419.
Supplementary data
Supplementary data related to this article can befound online at http://dx.doi.org/10.1016/j.jcyt.2013.11.008.