Embed Size (px)
Citation preview

Ihunnah et al - 1 -
Estrogen Sulfotransferase (EST/SULT1E1) Promotes Human Adipogenesis 1
2
Chibueze A. Ihunnah, Taira Wada, Brian J. Philips, Sudheer K. Ravuri, Robert B. Gibbs, Levent 3
Kirisci, J. Peter Rubin, Kacey G. Marra, Wen Xie 4
5
Center for Pharmacogenetics (C.A.I., T.W., W.X.) and Department of Pharmaceutical Sciences 6
(C.A.I., T.W., R.B.G., L.K., W.X.), University of Pittsburgh, Pittsburgh, Pennsylvania 15261; 7
Division of Plastic Surgery and McGowan Institute for Regenerative Medicine (B.J.P., S.K.R., 8
J.P.R., K.G.M.), Department of Surgery, University of Pittsburgh, PA 15261; and Department of 9
Pharmacology and Chemical Biology (W.X.), University of Pittsburgh, Pittsburgh, Pennsylvania 10
15261 11
12 *Corresponding author: Dr. Wen Xie, Center for Pharmacogenetics, 633 Salk Hall, University 13
of Pittsburgh, Pittsburgh, PA 15261. Telephone: 412-648-9941; Fax: 412-648-1664; E-mail: 14
16 Running Title: Estrogen sulfotransferase promotes human adipogenesis 17 18 Key Words: estrogen sulfotransferase, adipocyte differentiation, estrogens, estrogen receptor, 19 obesity 20 21 Abbreviations: ACC1, acetyl CoA carboxylase 1; ATGL, adipose triglyceride lipase; ASCs, 22 adipose derived stem cells; aP2, adipocyte protein 2; BMI, body mass index; C/EBP, 23 CCAAT/enhancer-binding protein; ChIP, chromatin immunoprecipitation; DCC-FBS, dextran-24 coated charcoal-stripped fetal bovine serum; DEX, dexamethasone; ER, estrogen receptor; EST, 25 estrogen sulfotransferase; HSL, hormone sensitive lipase; IBMX, isobutylmethylxanthine; IRS-1, 26 insulin receptor substrate-1; LPL, lipoprotein lipase; PPARγ, peroxisome proliferator-activated 27 receptor γ; SREBP-1c, sterol regulatory element binding protein 1c 28
29
MCB Accepts, published online ahead of print on 24 February 2014Mol. Cell. Biol. doi:10.1128/MCB.01147-13Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 2 -
ABSTRACT 30
The estrogen sulfotransferase (EST/SULT1E1) is known to catalyze the sulfoconjugation and 31
deactivation of estrogens. The goal of this study is to determine whether and how EST plays a 32
role in human adipogenesis. By using human primary adipose derived stem cells (ASCs) and 33
whole fat tissues from the abdominal subcutaneous fat of obese and non-obese subjects, we 34
showed that the expression of EST was low in pre-adipocytes but increased upon differentiation. 35
Overexpression and knockdown of EST in ASCs promoted and inhibited differentiation, 36
respectively. The pro-adipogenic activity of EST in humans was opposite to the anti-adipogenic 37
effect of the same enzyme in rodents. Mechanistically, EST promoted adipogenesis by 38
deactivating estrogens. The pro-adipogenic effect of EST can be recapitulated by using an 39
estrogen receptor (ER) antagonist or ERα knockdown. In contrast, activation of ER in ASCs 40
inhibited adipogenesis by decreasing the recruitment of the adipogenic peroxisome proliferator 41
activated receptor γ (PPARγ) onto its target gene promoters, whereas ER antagonism increased 42
the recruitment of PPARγ to its target gene promoters. Linear regression analysis revealed a 43
positive correlation between the expression of EST and body mass index (BMI), as well as a 44
negative correlation between the ERα expression and BMI. We conclude that EST is a pro-45
adipogenic factor, which may serve as a druggable target to inhibit the turnover and 46
accumulation of adipocytes in obese patients. 47
48
49
50
51
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 3 -
INTRODUCTION 52
Obesity is a major health concern with high prevalence. Obesity is defined as having a BMI 53
equal to or greater than 30, whereas a BMI equal to or greater than 25 is considered overweight. 54
An estimated 400 million people worldwide are obese. In the United States alone, obesity 55
related medical complications contribute to an average of 300,000 deaths annually (1). 56
57
The obesity epidemic has generated much research attention toward understanding the 58
biochemical regulation of adipose tissue and the development of adipocytes, known as 59
adipogenesis. Adipogenesis is a multifaceted process that is regulated by temporal and spatial 60
expression of a battery of adipogenic genes. When the pre-adipocytes located in various visceral 61
and subcutaneous fat depots are stimulated by specific mitogenic and adipogenic cues, they 62
begin the differentiation process until reaching maturation (2). This process is accompanied by a 63
dramatic increase in the expression of adipogenic genes, such as the lipoprotein lipase (LPL), 64
fatty acid binding protein 4 (FABP4/aP2), and the CCAATT enhancer binding proteins α, β, and 65
δ (C/EBPα, β, δ) (3). The peroxisome proliferator activated receptor γ (PPARγ) is a nuclear 66
receptor known as the master regulator of adipogenesis. Activation of PPARγ is required for the 67
induction of several aforementioned adipogenic enzymes and transcription factors (3). 68
Differentiation also leads to morphological and biochemical changes in pre-adipocytes that allow 69
them to store lipids and secrete adipokines (4). 70
71
The sex hormones are known to have a marked impact on adipose tissue development, 72
accumulation, and distribution in humans. Men tend to have a more central abdominal 73
accumulation of fat, while women tend to accumulate adipose tissue around the gluteal and 74
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 4 -
femoral area (5). Animal models and human epidemiological studies have shown that in general, 75
loss of estrogen signaling facilitates the increased adipose tissue accumulation. These have been 76
described in ERα and aromatase knockout mice, and documented in post-menopausal women (6-77
9). In contrast, estrogen replacement therapy in older women resulted in reductions in central 78
subcutaneous and visceral adipose tissue (10, 11). 79
80
The homeostasis of estrogens is tightly regulated by balanced synthesis and deactivation. The 81
estrogen sulfotransferase (EST or SULT1E1) is a key enzyme known to catalyze the sulfation of 82
estrogens, leading to their inactivation because of the inability of estrogen sulfates to bind to the 83
estrogen receptor (ER) (12). We recently described a novel role for Est in murine adipogenesis, 84
in which Est functions as a negative regulator of adipogenesis. Est is highly expressed in mouse 85
pre-adipocytes and differentiation attenuates the expression of Est (13). Moreover, Est over-86
expression and ablation inhibited and promoted murine adipogenesis, respectively (13). It is 87
unclear whether the role of EST in adipogenesis is conserved in humans. 88
89
In this study, we found the effect of EST on adipogenesis is highly species specific. EST 90
promoted human adipogenesis by deactivating estrogens. The pro-adipogenic effect of EST was 91
recapitulated in pre-adipocytes whose ER was pharmacologically or genetically inhibited. In 92
contrast, a pharmacological activation of ER inhibited adipogenesis. We propose EST as a 93
druggable target whose inhibition may be used to inhibit the turnover of adipocytes in obese 94
patients. 95
96
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 5 -
MATERIALS AND METHODS 97
Adipose Derived Stem Cell (ASC) and Adipose Tissue Collection 98
Human adipose derived stem cells (primary pre-adipocytes) and whole fat/lipoaspirate were 99
obtained through the Adipose Stem Cell Center, Department of Plastic Surgery, University of 100
Pittsburgh. Pre-adipocytes were isolated from the abdominal subcutaneous fat of 15 obese 101
patients and 3 non-obese (lean) patients whom had undergone liposuction or whole fat removal 102
surgery. All experiments were performed on cells from the obese patients except those 103
otherwise specified. The names of the patients were kept anonymous and all patients used for 104
pre-adipocyte differentiation experiments were female, non-diabetic, non-smokers, and ranged in 105
age from 25 to 56. The demographic information of the 18 patients is shown in the 106
Supplementary Table 1 (available upon request). The whole fats or liposuction aspirates were 107
collected from the abdominal subcutaneous fat of 16 female patients who were non-diabetic, 108
non-smokers, and ranged in age from 32-59. The demographic information of these 16 patients 109
is shown in the Supplementary Table 2 (available upon request). The cells were cultured in 110
standard conditions as reported (14). Briefly, cells were cultured in DMEM F-12 containing 111
10% standard fetal bovine serum (FBS) and 1% penicillin-streptomycin. Each cell line was 112
cultured for no more than four passages. Patient sample and data collection were performed in 113
accordance with the University of Pittsburgh Institutional Review Board Protocol PRO12050016. 114
115
Lentivirus Generation and Production 116
To generate the EST and EST AAK mutant expressing lentivirus, the EST cDNA, cloned by 117
RT-PCR using cDNA derived from differentiated human adipocytes, was cloned into the 118
lentiviral expression plasmid (pWPI) via the Pme1 restriction sites. Lentiviral particles were 119
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 6 -
generated using the 2nd generation system that contained three plasmids: the transgene expression 120
plasmid (pWPI-EST), a packaging plasmid (psPAX2) and an envelope plasmid (pMDG.2). All 121
three plasmids were transfected simultaneously into 293T cells for viral particle packaging, 122
assembly, and amplification using the Trans-IT transfection reagent from Mirus (Madison, WI). 123
Viral lysates were collected every 24 h after transfection, filtered with 0.45 μM Millex GV 124
syringe filter units from Millipore (Billerica, MA), pooled, and concentrated with Lenti-X-125
Concentrator from Clonetech (Mountain View, CA). Titer concentrations were assessed with 126
Lenti-X-Stix from Clonetech and by fluorescent examination of the green fluorescence protein 127
that was engineered in the lentiviral vector. Viral lysates were aliquoted and stored at -80 °C 128
until use. The cDNA for the EST AAK mutant was cloned by overlap extension PCR 129
mutagenesis (15). Pre-adipocytes were infected at an MOI of 3 before the puromycin selection. 130
Based on our observations, there was no noticeable effect of lentiviral infection and puromycin 131
selection alone on the differentiation. 132
133
To generate the shEST and shERα knockdown lentivirus, the expression plasmids containing 134
shEST and shERα were purchased from Open Biosystems (Pittsburgh, PA). For each 135
knockdown, five sequences were purchased and tested in transient transfection assays and the 136
sequence with the most efficient knockdown was chosen for lentiviral production. The targeting 137
sequences for EST and ERα are ATGAGTCTTCACAATTCTAGG (Product # 138
TRCN0000035880) and TTCCAGAGACTTCAGGGTGCT (Product # TRCN0000003299), 139
respectively. A scrambled shRNA (shSCR) plasmid was purchased to serve as a control. 140
141
Adipocyte Differentiation 142
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 7 -
In all experiments except those in which exogenous estrogen (E2) was added, the differentiation 143
media 1 (DM1) consisted of DMEM F-12, 10% standard FBS, 1% Pen-Strep, 33 μM biotin, 100 144
nM insulin, 17 μM pentothenic acid, 0.5 mM methylisobutylxanthine, 1 μM dexamethasone and 145
1 μM rosiglitazone. Differentiation media 2 (DM2) consisted of DMEM F-12, 10% FBS, 1% 146
Pen-Strep, 1 μM dexamethasone, and 100 nM insulin. Confluent pre-adipocytes were cultured 147
in DM1 for 3 days before switching to DM2 for 2 weeks to reach terminal differentiation. 148
Culture media was changed every other day. For differentiation experiments in which E2 was 149
exogenously added, the cells were cultured in phenol red free DMEM F12 and dextran coated 150
charcoal (DCC) stripped FBS before adding E2 at a final concentration of 10 nM. 151
152
Quantitative Real-Time RT-PCR 153
The cDNA was synthesized from 1.0 μg of total RNA by Iscript from Bio-Rad (Hercules, CA). 154
Aliquots of cDNA were amplified on an ABI 7300 Real-Time PCR System from Applied 155
Biosystems (Foster City, CA) using the SYBR Green PCR master mix. The mRNA expression 156
was normalized against the cyclophilin or glyeraldehyde 3-phosphate dehydrogenase (GAPDH) 157
expression. 158
159
Oil-Red O Staining and Quantification 160
Differentiated 6-well culture dishes were washed twice with cold PBS and then pre-fixed for 1 hr 161
with 10% formaldehyde in PBS. After 1 hr, fresh 10% formaldehyde was added and the cells 162
were incubated overnight at room temperature. The next day, the cells were washed twice with 163
PBS and then incubated in 60% isopropanol for 5 min, followed by drying at RT. Cells were 164
incubated with Oil-red O working solution for 1 min and then washed 5 times with deionized 165
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 8 -
water. Images were acquired microscopically. For quantification of Oil-red O staining, cells 166
were differentiated in triplicate in 24-well plates, stained with Oil-red O, eluted with 100% 167
isopropanol, and 100 μl of elute was loaded onto 96-well plates. Absorbance was measured at 168
500 nm using a Perkin Elmer plate reader. 169
170
Western Blot Analysis 171
Cells were lysed with NP-40 lysis buffer containing protease inhibitors, and then quantified for 172
protein concentrations by the BCA assay kit from Pierce (Rockford, IL). Protein samples were 173
resolved by electrophoresis on 10% SDS-polyacrylamide gels. For the detection of IRS1 and its 174
phosphorylation, cell lysates were immunoprecipitated with an IRS1 antibody before subjecting 175
to Western blotting using the IRS1 antibody and phospho-tyrosine antibody. After transfer of 176
proteins to PVDF membranes, the membranes were probed with antibodies against total ERK1/2 177
(Cat # sc94, Santa Cruz), phospho-ERK1/2 (Cat #sc7383, Santa Cruz), total AKT (Cat # 9272, 178
Cell Signaling), phospho-AKT (Cat # 9215, Cell Signaling), phospho-CREB (Cat # 87G3, Cell 179
Signaling), total CREB (Cat # 48H2, Cell Signaling), human EST (Cat # SAB1400267, Sigma), 180
ERα (Cat # sc7207, Santa Cruz), IRS1 (Cat # 2382S, Cell Signaling), and phospho-tyrosine (Cat 181
# ab10321, Abcam). Detection was achieved by using the ECL system from Amersham 182
(Piscataway, NJ). Quantification was performed using the NIH Image J software. 183
184
MTT Cell Proliferation Assay 185
MTT assay was performed using an assay kit from ATCC (Manassas, VA). Briefly, pre-186
adipocytes were grown in 150-cm dishes and treated with either EST or Vector lentivirus at an 187
MOI of 3. Five days later, cells were plated in triplicate at a density of 3 x 10^3 cells per well in 188
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 9 -
96-well plates for 1, 3, and 5 days. On the appropriate day, 10 μl of MTT reagent was added to 189
each well and the cells were incubated for 3 hr at 37 °C. Subsequently, 100 μl of detergent was 190
added and the cells were incubated overnight at RT before a colorimetric assessment using a 191
Perkin Elmer plate reader at 570 nm. 192
193
Transient Transfection and Luciferase Assays 194
HepG2 cells or 293T cells were plated at a density of 2 x 10^6 per 48-well plate and incubated 195
overnight. Transfection was performed using Trans-IT reagent from Mirus. Plasmids that were 196
used in triplicate at an amount of 300 ng included pCMX-EST, pCMX-EST AAK, pCMX-ERα, 197
pCMX-PPARγ and pCMX. The triplicate plasmid amounts for pCMX-CBP, pCMX-β-gal, and 198
tk-ERE-Luc or tk-PPRE-Luc were 50 ng, 200 ng, and 600 ng, respectively. Cells were 199
transfected and incubated for 24 hr. Transfected cells were treated with the appropriate ligand 200
using DMEM without phenol red and DCC FBS, followed by lysis and assays for luciferase and 201
β-gal activities. The luciferase activities were normalized to β-gal activities. 202
203
Chromatin Immunoprecipitation (ChIP) Assay 204
Chromatin Immunoprecipitation was performed according to a standard protocol (16). In brief, 205
pre-adipocytes were plated in duplicate 6-well plates, infected with either the vector or EST virus, 206
grown to confluence, and differentiated. Crosslinking was performed by adding formaldehyde, 207
followed by sonication to shear the DNA. Immunoprecipitation was performed using an anti-208
PPARγ antibody (Cat # ab45036) from Abcam, followed by elution using Protein A magnetic 209
beads (Cat # S1425S) from NEB (Ipswich, MA). Duplicate eluates, and 2% of the input DNA 210
were amplified by PCR and the PCR products were resolved on 1% agarose gel. Quantification 211
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 10 -
was performed by using the NIH Image J software. Fold enrichment was calculated as 212
precipitated DNA vs. input DNA. 213
214
Statistical Analysis 215
When applicable, results are presented as means ± SD. The Student's t test was used to calculate 216
the P values. P values of less than 0.05 are considered to be significant. Linear regression 217
analysis was performed using the GRAPHPAD Prism software. 218
219
220
221
222
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 11 -
RESULTS 223
The expression of EST was induced during adipogenesis 224
We have previously reported that the expression of the mouse Est gene was high in pre-225
adipocytes and the expression decreased upon differentiation (13). In an effort to determine 226
whether the human EST affects the differentiation of human pre-adipocytes, we were surprised 227
to find that in four independent cases of human pre-adipocytes isolated from obese patients, the 228
mRNA expression of EST in pre-adipocytes was low, and differentiation led to a marked 229
increase in EST expression (Fig. 1A). The induction of EST coincided with the induction of 230
adipocyte differentiation maker genes, such as PPARγ (Fig. 1B), LPL (Fig. 1C), and aP2 (Fig. 231
1D). Western blotting result showed that the expression of ERα remained constant from pre-232
adipocytes to differentiated adipocytes regardless of the EST overexpression (Fig. 1E). 233
234
Overexpression of EST promoted adipogenesis 235
To determine the functional relevance of EST induction during adipogenesis, we overexpressed 236
EST in pre-adipocytes before subjecting them to differentiation. Four cases of pre-adipocytes 237
were transduced with lentivirus overexpressing EST or the vector control virus, and then induced 238
to terminal differentiation for 2 weeks. The overexpression of EST was confirmed by real-time 239
PCR (Fig. 2A) and Western blotting (Fig. 2B). Overexpression of EST promoted adipogenesis, 240
as confirmed by both Oil-red O staining (Fig. 2C) and gene expression analysis (Fig. 2D). The 241
gene expression changes included the induction of PPARγ, LPL, aP2, and C/EBPα. The C/EBPs 242
are vital for adipogenesis (3, 4, 17). C/EBPα has been shown to facilitate the activation of 243
PPARγ target genes, whereas C/EBPβ and C/EBPδ are important for the transactivation of the 244
PPARγ gene expression (18-23). Interestingly, the enhanced adipogenesis in EST infected cells 245
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 12 -
was associated with the induction of C/EBPα (Fig. 2D), but the expression of C/EBPβ and 246
C/EBPδ was unchanged (data not shown). The expression of lipogenic genes, including sterol 247
regulatory element binding protein 1c (SREBP-1c), acetyl CoA carboxylase 1 (ACC1) (Fig. 2D), 248
fatty acid synthase (FAS), and stearoyl CoA desaturase 1 (SCD1) (data not shown), was 249
increased in EST transduced cells. In contrast, the expression of lipolytic genes adipose 250
triglyceride lipase (ATGL) and hormone sensitive lipase (HSL) was not affected by EST 251
overexpression (data not shown). 252
253
The enhancement of differentiation in EST infected cells was associated with an increased 254
phosphorylation of AKT and CREB in terminally differentiated cells (Fig. 2E), which was 255
suggestive of increased insulin signaling. However, when the acute insulin response was 256
evaluated in pre-adipocytes treated with insulin for 10 min, we found the phosphorylation of the 257
insulin receptor substrate 1 (IRS1) was similarly increased in vector infected and EST infected 258
cells (Fig. 2F). These results suggested that overexpression of EST alone did not confer 259
increased acute insulin sensitivity. The increased AKT and CREB phosphorylation in terminally 260
differentiated cells might have been secondary to the enhanced differentiation when EST was 261
overexpressed. The phosphorylation of ERK1/2 was also increased in EST infected cells (Fig. 262
2G). ERK1/2 activation is often linked to cell proliferation. During adipocyte differentiation, 263
ERK1/2 activation was required during the phase of clonal expansion, whereas persistent 264
activation of ERK1/2 may inhibit adipocyte differentiation (24, 25). Consistent with the 265
increased ERK1/2 phosphorylation, the proliferation of EST infected cells was increased 266
compared to the vector infected cells in the early phase of growth, but the difference became 267
insignificant after three days of culture when the cells were more confluent (Fig. 2H). Addition 268
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 13 -
of the ERK1/2 inhibitor PD98059 had little effect on the differentiation of either the vector or 269
EST infected cells (data not shown), suggesting that difference in proliferation rate in the early 270
phase might not be the key factor responsible for the difference in differentiation. 271
272
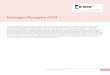
The enzymatic activity of EST was required for the adipogenic effect of EST 273
EST catalyzes the transfer of a sulfonate group from the universal sulfonate donor 3'-274
phosphoadenosine 5'-phosphosulfate (PAPS) to the estrogens. To assess whether the enzymatic 275
activity of EST is necessary for the pro-adipogenic effect, we generated a lentiviral vector 276
expressing a mutant human EST lacking its enzymatic activity. We and others reported that by 277
mutating a conserved domain in the P-loop region of the PAPS-binding domain from GxxGxxK 278
(GGK) to AxxAxxK (AAK), the enzymatic activity of EST was completely abolished (13, 26). 279
The lack of enzymatic activity of the EST AAK mutant was validated in a transfection and 280
reporter gene assay, in which the estrogen-deactivating activity of wild type EST was abolished 281
in EST AAK mutant transfected cells (Fig. 3A). The expression of EST AAK in transduced pre-282
adipocytes was confirmed by real-time PCR (data not shown) and Western blotting (Fig. 3B). 283
Oil-red O staining showed that overexpression of EST AAK failed to promote differentiation 284
(Fig. 3C). The lack of adipogenic activity of EST AAK was also supported by the lack of 285
induction of adipogenic gene expression (Fig. 3D) and phosphorylation of AKT and ERK (Fig. 286
3E). 287
288
Genetic knockdown or pharmacological inhibition of EST inhibited adipogenesis 289
Having demonstrated that a forced expression of EST promoted adipocyte differentiation, we 290
went on to determine whether a down-regulation of EST inhibited adipogenesis. In this 291
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 14 -
experiment, two cases of human pre-adipocytes were infected with lentivirus expressing an 292
shRNA against EST (shEST) or the control shScrambled (shSCR) before subjecting them to 293
differentiation for 2 weeks. The efficiency of EST knockdown was confirmed by real-time PCR 294
(Fig. 4A). Indeed, down-regulation of EST inhibited differentiation, as supported by the 295
markedly decreased Oil-red O staining (Fig. 4B). At the gene expression level, the expression of 296
PPARγ, LPL, aP2 and C/EBPα was decreased in shEST infected cells (Fig. 4C). The expression 297
of lipogenic genes SREBP-1c, ACC1, FAS, and SCD-1 was decreased in shEST infected cells 298
(Fig. 4C and data not shown). The expression of lipolytic genes ATGL and HSL was also 299
decreased in shEST knockdown cells (Fig. 4C). The pattern of lipogenic and lipolytic gene 300
expression was consistent with the suppression of adipogenesis. The phosphorylation of AKT 301
and CREB was also decreased in shEST infected cells (Fig. 4D). 302
303
The inhibition of differentiation was also observed when the pre-adipocytes were differentiated 304
in the presence of the EST inhibitor Triclosan. Triclosan, a proposed antimicrobial agent used in 305
many commercial products, inhibits EST from transferring a sulfuryl moiety from PAPS onto E2 306
by binding to the E2-binding site on EST, causing the formation of Triclosan-sulfate conjugates 307
instead of the E2-sulfate conjugates (27, 28). The inhibition by Triclosan appeared to be EST 308
specific (27). Our own results showed Triclosan inhibited EST, but had little effect on the 309
activity of the hydroxysteroid sulfotransferase SULT2A1 (data not shown). We showed that 310
treatment with Triclosan decreased Oil-red O staining (Fig. 4E), suppressed the expression of 311
differentiation marker genes PPARγ, LPL, and aP2, but had little effect on the expression of the 312
endogenous EST (Fig. 4F). 313
314
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 15 -
The effects of EST overexpression and knockdown were conserved in pre-adipocytes 315
isolated from non-obese patients 316
The above experiments were performed on pre-adipocytes isolated from overweight or obese 317
patients. To determine whether the EST effect on adipogenesis was conserved in non-obese 318
subjects, the EST overexpression and shEST knockdown experiments were repeated in three 319
cases of pre-adipocytes isolated from non-obese patients (BMI 21.4 - 23.7). As shown in Fig. 320
5A, lentiviral overexpression of EST increased adipogenic and lipogenic gene expression. In 321
contrast, shEST infected cells showed decreased expression of adipogenic genes, lipogenic genes 322
(Fig. 5B), and lipolytic genes (data not shown). The respective promotion and inhibition of 323
adipogenesis by EST overexpression (Fig. 5C) and knockdown (Fig. 5D) were confirmed by Oil-324
red O staining. 325
326
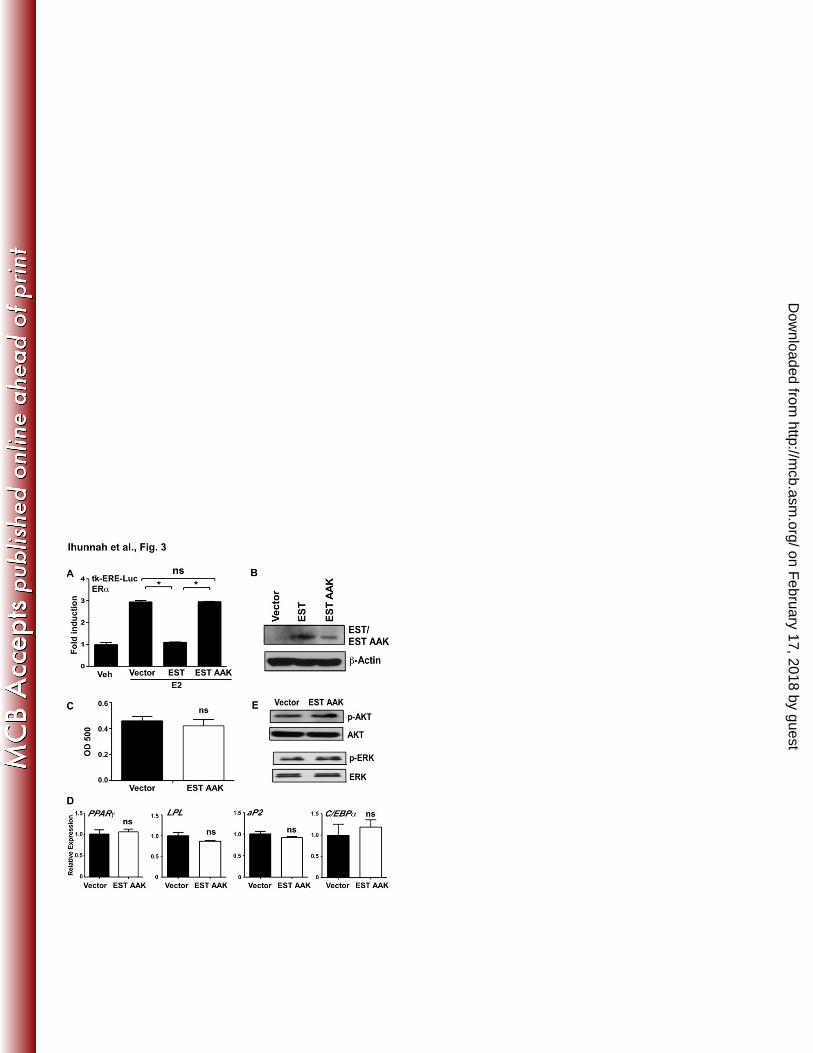
Pharmacological antagonism or genetic knockdown of ERα enhanced adipogenesis 327
A primary function of EST is to sulfonate and deactivate estrogens. The pro-adipogenic activity 328
of EST and the requirement of the enzymatic activity for the pro-adipogenic effect of EST led to 329
our hypothesis that EST may have promoted adipogenesis by antagonizing the estrogen activities. 330
Indeed, we showed that treatment of vector infected human pre-adipocytes maintained in DCC 331
serum with exogenously added E2 inhibited differentiation (Fig. 6A), which was in contrast to 332
the previously reported lack of estrogen effect on the differentiation of the mouse pre-adipocytes 333
(13). The inhibitory effect of E2 was abolished in EST-transduced cells, presumably due to the 334
enhanced estrogen deactivation. Interestingly, overexpression of EST alone in the absence of 335
exogenously added E2 was still efficient to increase the expression of PPARγ and aP2, although 336
the induction of LPL was not significant (Fig. 6A). The effectiveness of estrogen treatment in 337
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 16 -
vector infected cells and loss of estrogen effect in EST infected cells was confirmed by the 338
measurement of the expression of estrogen-responsive genes insulin-like growth factor binding 339
proteins 2 and 4 (IGFBP2 and IGFBP4) and glutathione peroxidase 3 (GPX3) (Fig. 6B). 340
Estrogens function through the estrogen receptors. Both ERα and ERβ are expressed in adipose 341
tissues, but ERα is the predominant ER isoform in the human subcutaneous fat (29). We then 342
used the ER antagonist Fulvestrant (ICI 182,780) and ERα knockdown to further determine the 343
role of estrogen-ER signaling in human adipocyte differentiation. Fulvestrant is a potent 344
ERα/β antagonist clinically used to treat estrogen dependent breast cancer (30). The 345
ERα knockdown was achieved by using a lentivirus expressing shRNA against ERα (shERα). 346
The efficiency of ERα knockdown was confirmed by real-time PCR (Fig. 6C, left panel) and 347
Western blotting (Fig. 6C, right panel). Knockdown of ERα did not affect the expression of 348
ERβ (Fig. 6C, left panel). 349
350
When subjecting the vector or EST infected pre-adipocytes to differentiation in the presence or 351
absence of Fulvestrant, or in the presence or absence of shERα, we found that treatment of the 352
vector cells with Fulvestrant or knockdown of ERα in the vector cells promoted adipogenesis to 353
a level similar to what was observed in vehicle-treated EST cells, as judged by Oil-red O staining 354
(Fig. 6D), adipogenic and lipogenic gene expression (Fig. 6E), and phosphorylation of AKT and 355
CREB (Fig. 6F). In contrast, treatment of the EST infected cells with Fulvestrant or knockdown 356
of ERα in EST infected cells showed little effect in further enhancing adipogenesis (Fig. 6D-6F). 357
These results demonstrated that adipogenesis was enhanced when the estrogen-ER signaling was 358
down-regulated through the use of ER antagonist or ERα knockdown, supporting that the pro-359
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 17 -
adipogenic effect of EST may have been mediated through the inhibition of the estrogen-ER 360
signaling pathway. 361
362
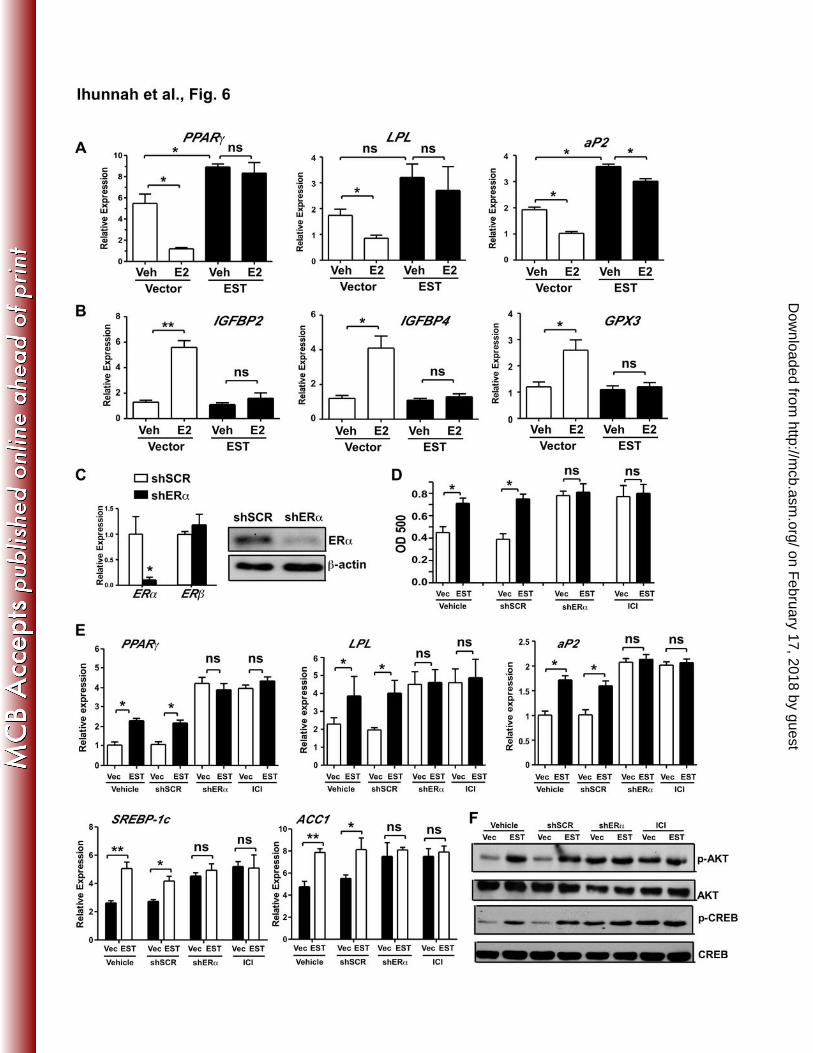
Molecular mechanism for the inhibitory effect of the estrogen-ERα signaling pathway on 363
adipogenesis 364
Crosstalk between nuclear receptors is an important regulatory mechanism to fine tune gene 365
expression. The heart of this crosstalk could lie in the limited supply of coactivators or 366
corepressors that are shared by two or more nuclear receptors within the same cells. PPARγ is a 367
critical adipogenic nuclear receptor (31, 32). CBP/p300 is an important coactivator shared by 368
both ERα and PPARγ. We then hypothesized that ERα may antagonize adipogenesis by trans-369
suppressing PPARγ. Indeed, we showed that in transient transfection and luciferase reporter 370
gene assays, ERα inhibited the PPARγ-mediated activation of a PPAR responsive reporter gene 371
tk-PPRE-Luc in a ligand dependent manner, whereas this inhibition was attenuated by the co-372
transfection of CBP (Fig. 7A). Reciprocally, PPARγ inhibited the ERα-mediated activation of 373
an ER responsive reporter gene tk-ERE-Luc in a ligand dependent manner, and this inhibition 374
was abolished by the co-transfection of CBP (Fig. 7B). 375
376
We then used chromatin immunoprecipitation (ChIP) assay on the vector and EST infected pre-377
adipocytes to determine whether ER activation or antagonism affected the recruitment of 378
PPARγ onto its adipogenic target gene promoters. As shown in Fig. 7C, overexpression of EST 379
led to an increased recruitment of PPARγ onto the LPL and aP2 gene promoters, which was 380
consistent with the phenotype of enhanced adipogenesis. Treatment of cells with Fulvestrant 381
increased the recruitment of PPARγ in vector cells to a level equal to that seen in the EST 382
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 18 -
infected cells. In contrast, treatment with E2 inhibited the recruitment of PPARγ in the vector 383
cells, and this inhibition was normalized in EST infected cells, likely due to the enhanced 384
estrogen deactivation. 385
386
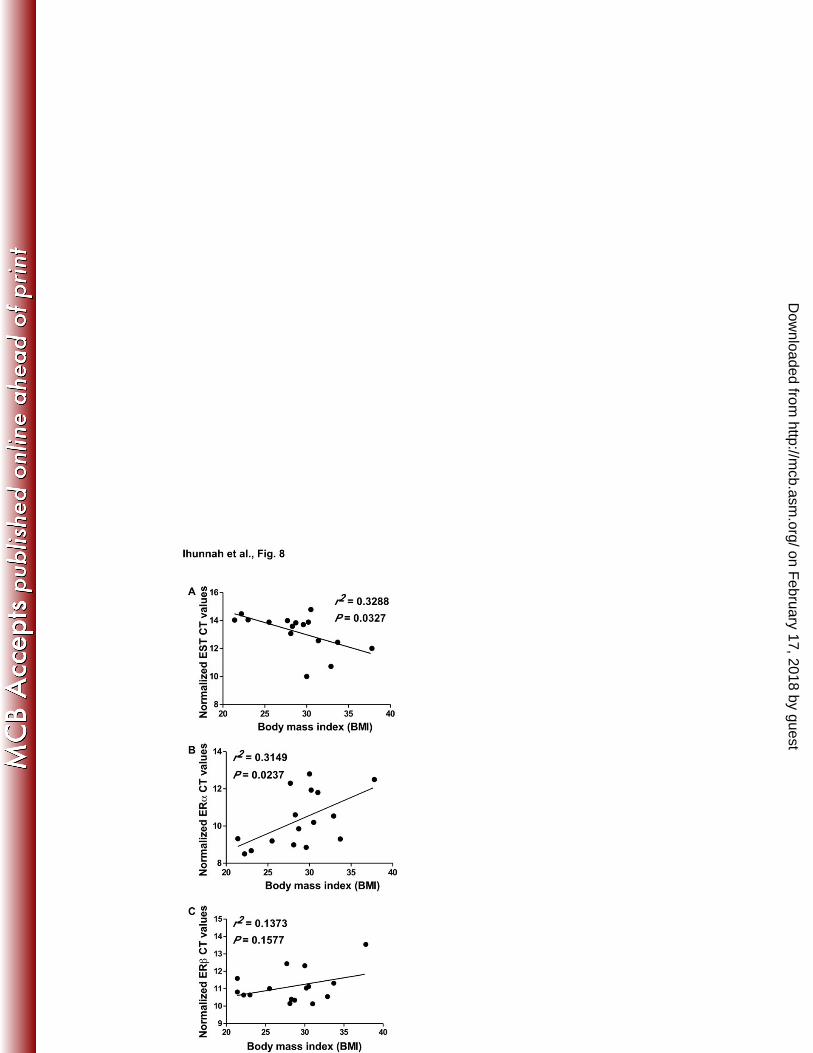
The respective expression of EST and ERα was positively and negatively correlated with 387
adiposity in human patients 388
The pro-adipogenic activity of EST and the anti-adipogenic activity of the estrogen-ER signaling 389
pathway prompted us to determine whether the expression of EST and ERα was positively and 390
negatively correlated with adiposity, respectively. In this experiment, total RNAs were isolated 391
from abdomen whole fat or liposuction aspirate from 16 patients, and the gene expression was 392
analyzed by real-time PCR. The correlation between gene expression and BMI was analyzed by 393
linear regression. We found a positive and significant correlation between EST expression and 394
BMI (Fig. 8A), as well as an inverse and significant correlation between ERα expression and 395
BMI (Fig. 8B). The correlation between ERβ expression and BMI did not reach a statistical 396
significance (Fig. 8C). 397
398
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 19 -
DISCUSSION 399
In this study, we have uncovered a novel role for EST in human adipogenesis. By using pre-400
adipocytes isolated from obese and non-obese patients and lentivirus overexpressing or 401
knocking-down EST, we showed that EST functioned as a positive regulator of adipogenesis. 402
Moreover, a positive correlation was found between the expression of EST and BMI, whereas a 403
negative correlation was found between the expression of ERα and BMI in human patients. 404
These results were consistent with the clinical observations that loss of estrogen signaling led to 405
increased adiposity (6, 7, 9-11, 33, 34) and there was an inverse correlation between the ERα 406
expression and BMI (35). Mechanistically, EST promoted adipogenesis by deactivating 407
estrogens, and the gain of function EST effect can be recapitulated by the use of an ER 408
antagonist or genetic knockdown of ERα. The inhibitory effect of the estrogen-ERα signaling 409
pathway on adipogenesis may have been achieved through the trans-suppression of PPARγ by 410
ERα, in which the activation of ERα inhibited the recruitment of PPARγ onto the promoters of 411
PPARγ target genes. In contrast, treatment with an ER antagonist increased the recruitment of 412
PPARγ onto its target gene promoters. 413
414
It is interesting to note that the effect of EST on adipogenesis in humans was opposite to what we 415
reported in mice. Using the mouse 3T3-L1 pre-adipocytes and mouse primary pre-adipocytes, 416
we found that Est levels were high in pre-adipocytes, and differentiation led to a decreased 417
expression of Est (13). Moreover, a forced expression of Est in 3T3-L1 cells inhibited 418
differentiation, whereas the pre-adipocytes isolated from the Est null mice showed enhanced 419
adipogenesis. In mouse pre-adipocytes, overexpression of Est prevented a molecular switch 420
from Erk1/2 activation in to Akt activation needed to facilitate adipocyte differentiation. 421
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 20 -
Treatment with the Erk1/2 inhibitor PD98059 partially restored the ability of Est-overexpressing 422
cells to differentiate (13). In human pre-adipocytes, we found that overexpression of EST caused 423
sustained activation of both ERK1/2 and AKT, and the final outcome is a promotion of 424
adipogenesis. Moreover, treatment of human pre-adipocytes with the ERK1/2 inhibitor 425
PD98059 had little effect on the differentiation of either the vector or EST infected cells. 426
427
The species-specific effect of EST may have been due to the differential effect of estrogens on 428
adipogenesis in mice and humans. Although the Est enzymatic activity was required for the 429
inhibitory effect Est on rodent adipogenesis, treatment of vector or Est overexpressing 3T3-L1 430
cells with estrogens failed to affect the adipogenesis (13), suggesting that Est substrate(s) other 431
than the estrogens may have been responsible for the effect of Est on mouse adipogenesis. In 432
contrast, in human pre-adipocytes, treatment with estrogens efficiently suppressed adipogenesis 433
and the inhibitory effect of estrogens was abolished in EST overexpressing cells, presumably due 434
to the enhanced estrogen deactivation. The mechanism for the species-specific effect of 435
estrogens and EST on adipogenesis remains to be clearly defined. Nevertheless, our 436
observations underscore the importance of using human cells and human models in 437
understanding the human adipogenesis. 438
439
The identification of EST as a positive regulator of human adipogenesis may have its clinical and 440
therapeutic implications. In humans, adipocytes from white adipose tissue replace themselves at 441
a rate of approximately 10% each year (36, 37). Additionally, it is believed that the human body 442
establishes a fixed number of adipocytes during early childhood development, adolescence, and 443
puberty. That number is maintained tightly throughout an individual’s adult life, despite long-444
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 21 -
term changes in diet and exercise that may result in a dramatic weight loss. It was reported that 445
even after lipectomy in non-obese female patients, body fat regenerates and redistributes 446
particularly to the abdominal region (38). This phenomenon was also recapitulated in mouse 447
models (39). It is conceivable that adipocyte differentiation and replenishment play an essential 448
role in maintaining the adipose tissue homeostasis. This provides a novel concept of anti-obesity 449
therapy, that is the need to limit adipocyte turnover, either through an increased apoptosis of 450
adipocytes or through the inhibition of adipocyte differentiation. One caveat that might hinder 451
the attempt to increase apoptosis is that increased cell death may cause an inflammatory response 452
that could actually worsen the metabolic syndrome. Our results showed that a pharmacological 453
inhibition or genetic knockdown of EST attenuated the adipogenic process, suggesting that 454
inhibition of EST may be an effective weight reduction strategy in the future. It is encouraging 455
that major progress has been made in the identification and characterization of chemical EST 456
inhibitors (27, 40). 457
458
Among limitations, it is noted that the age range of our human subjects was quite varied and all 459
samples were from the abdominal subcutaneous fat. Although the effects of EST overexpression 460
and knockdown were observed in pre-adipocytes isolated from both the obese and non-obese 461
patients, we cannot exclude the possibility that age, menopausal status, steroid hormone level, 462
obesity, and origin of the fat depot may affect the phenotypic exhibition. In addition, since 463
overexpression of EST in the “estrogen-free” DCC cell culture condition can still increase the 464
expression of certain adipogenic marker genes (Fig. 6A), we cannot exclude the possibility of 465
off-target effect due to the overexpression of EST, as well as the existence of additional EST 466
substrates that may also have an effect on adipogenesis. 467
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 22 -
468
In summary, we have established that EST is an important positive regulator of adipogenesis in 469
humans. We propose that EST is a druggable target whose inhibition can be used to inhibit the 470
turnover of adipocytes in obese patients. 471
472
473
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 23 -
ACKNOWLEDGEMENTS 474
The authors want to thank Dr. Donald DeFranco (University of Pittsburgh) for his insightful 475
comments on this study and Dr. Jiang Li (University of Pittsburgh) for some of the data analysis. 476
This work was supported in part by NIH grants DK083953 and HD073070 (to W.X.), and 477
CA114246 (to J.P.R.). C.A.I was supported by the NIH Ruth L. Kirschstein National Research 478
Service Awards for Individual Predoctoral Fellowships to Promote Diversity in Health-Related 479
Research F31-DK-095589. W.X. is the Joseph Koslow Endowed Chair in Pharmaceutical 480
Sciences at University of Pittsburgh School of Pharmacy. 481
482
483
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 24 -
FIGURE LEGENDS 484
Figure 1. The expression of EST was induced during adipogenesis. (A to D) Pre-adipocytes 485
were cultured and differentiated in standard media for 14 days. Total RNA was extracted before 486
and after differentiation and the expression of EST (A), PPARγ (B), LPL (C) and aP2 (D) was 487
measured by real-time PCR analysis. *, P < 0.05. (E) The expression of ERα was measured by 488
Western blotting. NT, non-transfected. 489
490
Figure 2. Overexpression of EST promoted adipogenesis. (A and B) Pre-adipocytes were 491
transduced with vector or EST expressing lentivirus. The expression of the transduced EST was 492
confirmed by real-time PCR (A) and Western blotting (B). (C) Vector and EST cells were 493
induced to differentiate for 14 days in standard differentiation media before being examined by 494
Oil-Red O staining microscopically (left panel) and quantitatively (right panel). (D) Total RNA 495
was extracted before and after differentiation and the expression of indicated genes was 496
measured by real-time PCR. (E) Lysates from differentiated cells were subjected to Western 497
blot analysis to detect the protein expression of total AKT, phospho-AKT, total CREB, and 498
phospho-CREB. The signals were quantified by using NIH Image J software. N=3. (F) Pre-499
adipocytes were treated with insulin (100 nM) for 10 min before being evaluated for the protein 500
expression of total IRS1 and phospho-IRS1 by immunoprecipitation and Western blotting. (G) 501
Lysates from differentiated cells were subjected to Western blot analysis to detect the protein 502
expression of total ERK1/2 and phospho-ERK1/2. (H) Pre-adipocytes were transduced with 503
vector or EST lentivirus and grown in triplicate for each time point and examined by MTT 504
proliferation assay. *, P < 0.05; **, P < 0.01; ns, statistically not significant. 505
506
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 25 -
Figure 3. The enzymatic activity of EST was required for the adipogenic effect of EST. (A) 507
The activity of the EST AAK mutant was compared to the wild type EST. 293T cells were 508
transfected with tk-ERE-Luc reporter gene and ERα, along with EST, EST AAK, or empty 509
vector as indicated. The cells were either treated with 1 nM E2 or vehicle in phenol red-free 510
DMEM containing 10% DCC FBS. The luciferase activities are normalized against β-gal 511
activities from the co-transfected CMX-β-gal vector. Results are shown as fold induction over 512
vehicle treated triplicate. (B) Pre-adipocytes were infected with EST or EST AAK lentivirus. 513
The expression of EST and EST AAK was measured by Western blotting. (C to E) Pre-514
adipocytes were transduced with vector or AAK lentivirus and then induced to differentiate for 515
14 days before being evaluated for Oil-red O staining (C), gene expression analysis by real-time 516
PCR (D), and Western blot analysis to detect total AKT, phospho-AKT, total ERK1/2 and 517
phospho-ERK1/2 (E). *, P < 0.05; ns, statistically not significant. 518
519
Figure 4. Genetic knockdown or pharmacological inhibition of EST inhibited adipogenesis. 520
(A to D) Pre-adipocytes were transduced with either shEST or shScrambled (shSCR) lentivirus 521
and then induced to differentiate for 2 weeks before being evaluated for the efficiency of EST 522
knockdown by real-time PCR analysis (A), Oil-red O staining and quantification (B), gene 523
expression analysis by real-time PCR (C), Western blot analysis to detect the expression of total 524
AKT, phospho-AKT, total CREB and phosphor-CREB (D). (E and F) Pre-adipocytes were 525
induced to differentiate for 2 weeks in the presence of 10 μM Triclosan or vehicle before being 526
evaluated for Oil-red O staining (E) and the expression of adipogenic genes and EST (F). *, P < 527
0.05; **, P < 0.01; ns, statistically not significant. 528
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 26 -
Figure 5. The effects of EST overexpression and knockdown were conserved in pre-529
adipocytes isolated from non-obese patients. The BMI rage for these three patients is 21.4 to 530
23.7. (A and B) Vector and EST infected cells (A) or shSCR and shEST infected cells (B) were 531
induced to differentiate for 2 weeks before gene expression profiling by real-time PCR. (C and 532
D) The differentiation of EST infected cells (C) and shEST infected cells (D) was evaluated by 533
the quantification of Oil-red O staining. N=3. *, P < 0.05; **, P < 0.01. 534
535
Figure 6. Pharmacological antagonism or genetic knockdown of ERα enhanced 536
adipogenesis. (A and B) Pre-adipocytes were transduced with either vector or EST virus and 537
induced to differentiate, in the presence or absence of 10 nM E2 for 2 weeks before being 538
evaluated for the expression of adipogenic genes (A) and estrogen responsive genes (B) by real-539
time PCR. (C) The efficiency of lentiviral shRNA knockdown of ERα was confirmed by real-540
time PCR (left panel) and Western blotting (right panel). (D to F) The vector or EST infected 541
pre-adipocytes were treated with 100 nM Fulvestrant (ICI) or transduced with shSCR or shERα 542
lentivirus and then induced to differentiate for 2 weeks before being evaluated for Oil-red O 543
staining and quantification (D), expression of adipogenic and lipogenic genes by real-time PCR 544
(E), and Western blot analysis to detect the expression of total AKT, phospho-AKT, total CREB 545
and phospho-CREB (F). *, P < 0.05; **, P < 0.01; ns, statistically not significant. 546
547
Figure 7. Molecular mechanism for the inhibitory effect of the estrogen-ER signaling 548
pathway on adipogenesis. (A and B) 293T cells were transfected with either the tk-ERE-Luc 549
reporter gene together with ERα and/or CBP (A) or the tk-PPRE-Luc reporter gene together with 550
PPARγ and/or CBP (B). Transfected cells were treated with E2 (10 nM) and/or rosiglitazone 551
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 27 -
(Rosi, 1uM) for 24 h before luciferase assay. The luciferase activities were normalized 552
against β-gal activities from the co-transfected β-gal vector. Results are shown as fold induction 553
over vehicle treated triplicates. (C) The vector or EST transduced pre-adipocytes were treated 554
with 100 nM Fulvestrant (ICI) or 10 nM E2 and then induced to differentiate for 2 weeks before 555
being evaluated for the recruitment of PPARγ onto the LPL (left panel) and aP2 (right panel) 556
gene promoters by chromatin immunoprecipitation (ChIP) assay. Quantification was performed 557
by using NIH Image J software. *, P < 0.05; ns, statistically not significant. 558
559
Figure 8. The respective expression of EST and ERα was positively and negatively 560
correlated with adiposity in human patients. (A-C) Total RNAs extracted from the whole fat 561
or lipoaspirate of a cohort of 16 patients were subjected to gene expression analysis by real-time 562
PCR. The correlation between body mass index (BMI) and EST (A), ERα (A) or ERβ (C) gene 563
expression was analyzed by linear regression analysis. The expression of EST and ERs was 564
presented as normalized CT values, in which a higher CT value indicates a low gene expression. 565
The expression is normalized to GAPDH. 566
567
568
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 28 -
REFERENCES 569
1. Zammit, C., H. Liddicoat, I. Moonsie, and H. Makker. 2010. Obesity and respiratory 570 diseases. Int J Gen Med 3:335-343. 571
2. Rayalam, S., M.A. Della-Fera, and C.A. Baile. 2008. Phytochemicals and regulation of 572 the adipocyte life cycle. J Nutr Biochem 19:717-726. 573
3. Gregoire, F.M., C.M. Smas, and H.S. Sul. 1998. Understanding adipocyte 574 differentiation. Physiol Rev 78:783-809. 575
4. Gregoire, F.M. 2001. Adipocyte differentiation: from fibroblast to endocrine cell. Exp 576 Biol Med (Maywood) 226:997-1002. 577
5. Mayes, J.S. and G.H. Watson. 2004. Direct effects of sex steroid hormones on adipose 578 tissues and obesity. Obes Rev 5:197-216. 579
6. Mueller, S.O. and K.S. Korach. 2001. Estrogen receptors and endocrine diseases: 580 lessons from estrogen receptor knockout mice. Curr Opin Pharmacol 1:613-619. 581
7. Couse, J.F. and K.S. Korach. 1999. Estrogen receptor null mice: what have we learned 582 and where will they lead us? Endocr Rev 20:358-417. 583
8. Jones, M.E., A.W. Thorburn, K.L. Britt, K.N. Hewitt, N.G. Wreford, J. Proietto, 584 O.K. Oz, B.J. Leury, K.M. Robertson, S. Yao, and E.R. Simpson. 2000. Aromatase-585 deficient (ArKO) mice have a phenotype of increased adiposity. Proc Natl Acad Sci U S 586 A 97:12735-12740. 587
9. D'Eon, T.M., S.C. Souza, M. Aronovitz, M.S. Obin, S.K. Fried, and A.S. Greenberg. 588 2005. Estrogen regulation of adiposity and fuel partitioning. Evidence of genomic and 589 non-genomic regulation of lipogenic and oxidative pathways. J Biol Chem 280:35983-590 35991. 591
10. Salpeter, S.R., J.M.E. Walsh, T.M. Ormiston, E. Greyber, N.S. Buckley, and E.E. 592 Salpeter. 2006. Meta-analysis: effect of hormone-replacement therapy on components of 593 the metabolic syndrome in postmenopausal women. Diabetes Obes Metab 8: 538-554. 594
11. Davis, S.R., C. Castelo-Branco, P. Chedraui, M.A. Lumsden, R.E. Nappi, D. Shah, P. 595 Villaseca, and Writing Group of the International Menopause Society for World 596 Menopause Day 2012. 2012. Understanding weight gain at menopause. Climacteric 597 15:419-429. 598
12. Song, W.C. 2001. Biochemistry and reproductive endocrinology of estrogen 599 sulfotransferase. Ann N Y Acad Sci. 948:43-50. 600
13. Wada, T., C.A. Ihunnah, J. Gao, X. Chai, S. Zeng, B.J. Philips, J.P. Rubin, K.G. 601 Marra, and W. Xie. 2011. Estrogen sulfotransferase inhibits adipocyte differentiation. 602 Mol Endocrinol 25:1612-1623. 603
14. Gerlach, J.C., Y.C. Lin, C.A. Brayfield, D.M. Minteer, H. Li, J.P. Rubin, and K.G. 604 Marra. 2012. Adipogenesis of human adipose-derived stem cells within three-605 dimensional hollow fiber-based bioreactors. Tissue Eng Part C Methods. 18:54-61. 606
15. Ho, S.N., H.D. Hunt, R.M. Horton, J.K. Pullen, and L.R. Pease. 1989. Site-directed 607 mutagenesis by overlap extension using the polymerase chain reaction. Gene 77:51-59. 608
16. Nelson, J.D., O. Denisenko, and K. Bomsztyk. 2006. Protocol for the fast chromatin 609 immunoprecipitation (ChIP) method. Nat Protoc 1:179-185. 610
17. Rangwala, S.M. and M.A. Lazar. 2000. Transcriptional control of adipogenesis. Annu 611 Rev Nutr 20:535-559. 612
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 29 -
18. Cao, Z., R.M. Umek, and S.L. McKnight. 1991. Regulated expression of three C/EBP 613 isoforms during adipose conversion of 3T3-L1 cells. Genes Dev 5:1538-1552. 614
19. Hwang, C.S., S. Mandrup, O.A. MacDougald, D.E. Geiman, and M.D. Lane. 1996. 615 Transcriptional activation of the mouse obese (ob) gene by CCAAT/enhancer binding 616 protein alpha. Proc Natl Acad Sci U S A 93:873-877. 617
20. Ross, S.R., R.A. Graves, A. Greenstein, K.A. Platt, H.L. Shyu, B. Mellovitz, and B.M. 618 Spiegelman. 1990. A fat-specific enhancer is the primary determinant of gene expression 619 for adipocyte P2 in vivo. Proc Natl Acad Sci U S A 87:9590-9594. 620
21. Wu, Z., N.L. Bucher, and S.R. Farmer. 1996. Induction of peroxisome proliferator-621 activated receptor gamma during the conversion of 3T3 fibroblasts into adipocytes is 622 mediated by C/EBPbeta, C/EBPdelta, and glucocorticoids. Mol Cell Biol 16:4128-4136. 623
22. Wu, Z., Y. Xie, N.L. Bucher, and S.R. Farmer. 1995. Conditional ectopic expression 624 of C/EBP beta in NIH-3T3 cells induces PPAR gamma and stimulates adipogenesis. 625 Genes Dev 9:2350-2363. 626
23. Miki, H., T. Yamauchi, R. Suzuki, K. Komeda, A. Tsuchida, N. Kubota, Y. Terauchi, 627 J. Kamon, Y. Kaburagi, J. Matsui, Y. Akanuma, R. Nagai, S. Kimura, K. Tobe, and 628 T. Kadowaki. 2001. Essential role of insulin receptor substrate 1 (IRS-1) and IRS-2 in 629 adipocyte differentiation. Mol Cell Biol 21:2521-2532. 630
24. Font de Mora, J., A. Porras, N. Ahn, and E. Santos. 1997. Mitogen-activated protein kinase 631 activation is not necessary for, but antagonizes, 3T3-L1 adipocytic differentiation. Mol Cell Biol 632 17:6068-75. 633
25. Sale, E. M., P. G. Atkinson, and G. J. Sale. 1995. Requirement of MAP kinase for 634 differentiation of fibroblasts to adipocytes, for insulin activation of p90 S6 kinase and for insulin 635 or serum stimulation of DNA synthesis. EMBO J 14:674-84. 636
26. Komatsu, K., W.J. Driscoll, Y.C. Koh, and C.A. Strott. 1994. A P-loop related motif 637 (GxxGxxK) highly conserved in sulfotransferases is required for binding the activated 638 sulfate donor. Biochem Biophys Res Commun 204:1178-1185. 639
27. Wang, L.Q., C.N. Falany, and M.O. James. 2004. Triclosan as a substrate and inhibitor 640 of 3'-phosphoadenosine 5'-phosphosulfate-sulfotransferase and UDP-glucuronosyl 641 transferase in human liver fractions. Drug Metab Dispos 32:1162-1169. 642
28. Zhang, H., O. Varlamova, F.M. Vargas, C.N. Falany, and T.S. Leyh. 1998. Sulfuryl 643 transfer: the catalytic mechanism of human estrogen sulfotransferase. J Biol Chem 273: 644 10888-10892. 645
29. Joyner, J.M., L.J. Hutley, and D.P. Cameron. 2001. Estrogen receptors in human 646 preadipocytes. Endocrine 15:225-230. 647
30. Cardoso, F., J. Bischoff, E. Brain, A.G. Zotano, H.-J. Luck, V.C. Tjan-Heijnen, M. 648 Tanner, and M. Aapro. 2013. A review of the treatment of endocrine responsive 649 metastatic breast cancer in postmenopausal women. Cancer Treat Rev 39:457-465. 650
31. Anghel, S.I. and W. Wahli. 2007. Fat poetry: a kingdom for PPAR gamma. Cell Res 17: 651 486-511. 652
32. Spiegelman, B.M. 1998. PPAR-gamma: adipogenic regulator and thiazolidinedione 653 receptor. Diabetes 47:507-514. 654
33. Cooke, P.S. and A. Naaz. 2005. Effects of estrogens and the phytoestrogen genistein on 655 adipogenesis and lipogenesis in males and females. Birth Defects Res A Clin Mol Teratol 656 73:472-473. 657
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Ihunnah et al - 30 -
34. Heine, P.A., J.A. Taylor, G.A. Iwamoto, D.B. Lubahn, and P.S. Cooke. 2000. 658 Increased adipose tissue in male and female estrogen receptor-alpha knockout mice. Proc 659 Natl Acad Sci U S A 97:12729-12734. 660
35. Lundholm, L., H. Zang, A.L. Hirschberg, J.-A. Gustafsson, P. Arner, and K. 661 Dahlman-Wright. 2008. Key lipogenic gene expression can be decreased by estrogen in 662 human adipose tissue. Fertil and Steril, 90: 44-48. 663
36. Arner, P. and K.L. Spalding. 2010. Fat cell turnover in humans. Biochem Biophys Res 664 Commun 396:101-104. 665
37. Spalding, K.L., E. Arner, P.O. Westermark, S. Bernard, B.A. Buchholz, O. 666 Bergmann, L. Blomqvist, J. Hoffstedt, E. Naslund, T. Britton, H. Concha, M. 667 Hassan, M. Ryden, J. Frisen, and P. Arner. 2008. Dynamics of fat cell turnover in 668 humans. Nature 453:783-787. 669
38. Hernandez, T.L., J.M. Kittelson, C.K. Law, L.L. Ketch, N.R. Stob, R.C. Lindstrom, 670 A. Scherzinger, E.R. Stamm, and R.H. Eckel. 2011. Fat redistribution following 671 suction lipectomy: defense of body fat and patterns of restoration. Obesity (Silver Spring) 672 19:1388-1395. 673
39. Rigamonti, A., K. Brennand, F. Lau, and C.A. Cowan. 2011. Rapid cellular turnover 674 in adipose tissue. PLoS One 6:e17637. 675
40. James, M.O., W. Li, D.P. Summerlot, L. Rowland-Faux, and C.E. Wood. 2010. 676 Triclosan is a potent inhibitor of estradiol and estrone sulfonation in sheep placenta. 677 Environ Int 36:942-949. 678
679 680
on February 17, 2018 by guest
http://mcb.asm
.org/D
ownloaded from