Embed Size (px)
Citation preview

Vietnam J. Agri. Sci. 2017, Vol. 15, No. 12: 1593-1604 Tạp chí Khoa học Nông nghiệp Việt Nam 2017, 15(12): 1593-1604 www.vnua.edu.vn
1593
PHÂN LẬP VÀ ĐÁNH GIÁ KHẢ NĂNG ĐỐI KHÁNG CỦA Trichoderma asperellum ĐỐI VỚI TÁC NHÂN GÂY BỆNH CÂY CÓ NGUỒN GỐC TRONG ĐẤT
Nguyễn Đức Huy1*
, Phạm Quang Nguyên2, Nguyễn Thị Thanh Hồng
1,
Hà Giang3, Nguyễn Văn Viên
1, Nguyễn Tất Cảnh
1
1Khoa Nông học, Học viện Nông nghiệp Việt Nam
2Học viên cao học, Khoa Nông học, Học viện Nông nghiệp Việt Nam
3Trung tâm Bệnh cây nhiệt đới, Học viện Nông nghiệp Việt Nam
Email*: [email protected]
Ngày gửi bài: 10.12.2017 Ngày chấp nhận: 16.01.2018
TÓM TẮT
Nấm đối kháng, Trichoderma spp., đã được ghi nhận như là tác nhân phòng chống sinh học chủ yếu để kiểm soát tác nhân gây bệnh hại cây trồng có nguồn gốc trong đất và có hiệu quả hơn so với hóa học. Trichoderma spp.
được phân bố rộng trong đất và hệ sinh thái vùng rễ cây trồng. Trong nghiên cứu này, chín mẫu đất được thu thập từ đất trồng cây hàng năm và lâu năm ở năm tỉnh của miền Bắc Việt Nam để phân lập nấm Trichoderma spp. Các mẫu nấm đối kháng được xác định dựa vào đặc điểm hình thái và phân tử. Trên cơ sở phân tích trình tự vùng ITS, 8 mẫu được xác định là T. asperellum và 1 mẫu là T. harzianum. Kết quả nghiên cứu cho thấy rằng T. asperellum có tốc độ phát triển nhanh, khả năng sinh bào tử nhiều trên 3 loại cơ chất và có hiệu quả đối kháng mạnh đối với các tác nhân gây bệnh lở cổ rễ do nấm Rhizoctinia solani và thối hạch do nấm Sclerotinia sclerotiorum trong điều kiện in vitro. Hơn nữa, đánh giá tính an toàn sinh học cho thấy nấm T. asperellum an toàn với chuột ở liều lượng 20 g/kg. Nghiên cứu này cho thấy nấm T. asperellum phân bố khá phổ biến ở miền Bắc Việt Nam.
Từ khóa: Trichoderma asperellum, phòng trừ sinh học, Sclerotinia sclerotiorum, Rhizoctonia solani.
Isolation and Evaluation of Antagonistic Ability of Trichoderma asperellum against Soil Borne Plant Pathogens
ABSTRACT
Antagonistic fungi, Trichoderma spp., have been reported as potential bio-control agents to control soil borne plant pathogens more effectively than chemicals. Trichoderma spp. were widely spread in soil and rhizospehre of
plants. In this study, Trichoderma spp. were isolated from nine samples of soils cultivated with annual and perennial crops in five different provinces in the North of Vietnam. Trichoderma spp. were identified based on morphology and molecular characterizations. Based on the sequence analysis of ITS region, the results showed that eight isolates were identified as T. asperellum and one isolate was T. harzianum. The results indicated that T. asperellum exhibited a high growth rate and high sporulation on three different substrates and strong inhibitory effect against pathogens that cause root rot disease by Rhizoctinia solani and Sclerotinia stem rot by Sclerotinia sclerotiorum in vitro. Moreover, biosecurity assessment showed that T. asperellum was safe in rats at 20 g/kg. Our results revealed that T. asperellum is widely distributed in Northern provinces of Vietnam.
Keywords: Trichoderma asperellum, bio-control, Sclerotinia sclerotiorum, Rhizoctonia solani.
1. ĐẶT VẤN ĐỀ
Việt Nam là quốc gia nông nghiệp, đa dạng về giống cây trồng với nhiều loài chủ lực và có giá trị kinh tế cao. Tuy nhiên trong sản xuất nông nghiệp, năng suất và chất lượng của cây trồng luôn bị đe dọa
bởi dịch hại, đặc biệt là bệnh hại. Để bảo vệ năng suất và chất lượng của cây trồng, biện pháp hóa học được sử dụng rộng rãi mặc dù sử dụng nhiều thuốc hóa học sẽ tạo ra tính kháng của dịch hại và gây ô nhiễm môi trường và khó phòng trừ được nhóm nấm gây bệnh hại cây trồng có nguồn gốc trong đất.

Phân lập và đánh giá khả năng đối kháng của Trichoderma asperellum đối với tác nhân gây bệnh cây có nguồn gốc
trong đất
1594
Chính vì thế, hiện nay vi khuẩn và nấm đối kháng được sử dụng rộng trên thế giới và ở Việt Nam để phòng trừ bệnh hại cây trồng nhằm thân thiện với môi trường và phù hợp với sản xuất nông nghiệp theo hướng hữu cơ. Trong số các vi sinh vật đối kháng, nấm Trichoderma spp. được sử dụng nhiều trong phòng trừ bệnh hại cây trồng cả trên thế giới và ở Việt Nam như T. viride, T. harzianum và T. hamatum.
Các loài nấm thuộc chi Trichoderma (Hypocreales, Ascomycota) có phạm vi phân bố rộng khắp ở trong đất và hệ sinh thái vùng rễ cây trồng. Những nghiên cứu trước đây cho thấy nấm Trichoderma spp. có tác dụng đối kháng chống lại ít nhất 18 chi và 29 loài nấm gây bệnh cây, cũng như một loạt các vi khuẩn gây bệnh cây. Cơ chế đối kháng của Trichoderma spp. chủ yếu bao gồm cạnh tranh dinh dưỡng, ký sinh nấm gây bệnh, tạo kháng sinh, tăng sinh tưởng và kích thích phản ứng kháng của cây trồng (Harman, 2006). Nhiều loài Trichoderma spp. có tốc độ phát triển nhanh và khả năng đối kháng một số loài nấm bệnh hại vùng rễ cây trồng có nguồn gốc trong đất như nấm Rhizoctonia solani gây bệnh lở cổ rễ (Anees et al., 2010; Asad et al., 2014), Sclerotinia sclerotiorum gây bệnh thối hạch bắp cải (Troian et al., 2014; Zhang et al., 2016), Sclerotium rolfsii gây bệnh héo rũ gốc mốc trắng cà chua, khoai tây, đậu tương, dưa chuột (Kotasthane et al., 2015), Fusarium solani gây bệnh thối gốc thân lạc (Rojo et al., 2007).
Một loài nấm đối kháng mới, T. asperellum, đã được phát hiện trong những năm gần đây. T. asperellum có tốc độ phát triển nhanh, hình thành số lượng bào tử phân sinh lớn và có khả năng đối kháng cao đối với các tác nhân gây bệnh thối rễ do nấm Pythium myriotylum (Mbaga et al., 2012), bệnh thán thư xoài (Santos-Villalobos et al., 2013), bệnh héo fusarium trên cây cà chua (Komy et al., 2015), bệnh héo fusarium trên cây dưa chuột và thối rễ cây ngô (Wu et al., 2017). Hơn nữa, T. asperellum tiết ra chitinase, glucanase và protease, có thể làm phá hủy vách tế bào của nấm và ký sinh trong sợi nấm gây bệnh. Ngoài ra, chất xylanases do T. asperellum tiết ra có tác dụng làm tính kháng và miễn dịch của cây trồng đối với tác nhân gây bệnh (Wu et al., 2017).
Trong đất nông nghiệp, nấm Trichoderma có nhiều loài và phân bố rộng ở các vùng sinh thái. Ở Việt Nam, trước đây, phân lập và giám định nấm Trichoderma spp. dựa vào hình thái cho thấy chủ yếu là loài T. harzianum và T. viride. Các thử nghiệm trong phòng và ứng dụng ngoài đồng ruộng để phòng trừ nấm gây bệnh hại vùng rễ cây trồng chủ yếu sử dụng nấm T. harzianum và T. viride (Trần Thị Thuần, 1999; Trần Kim Loang và cs., 2009; Nguyễn Văn Viên và cs., 2012). Gần đây, nấm T. asperelum cũng đã được phát hiện ở Tây Nguyên (Phạm Ngọc Dung và
cs., 2012), ở Quảng Trị và Thừa Thiên Huế (Loc et al., 2011; Trần Thị Thu Hà và Phạm Văn Hòa, 2012). Mục đích của nghiên cứu này nhằm i) thu thập, phân lập và xác định nấm Trichoderma spp. ở năm tỉnh phía Bắc của Việt Nam dựa vào hình thái kết hợp với giải trình tự gen vùng rDNA-ITS, ii) đánh giá hiệu lực ức chế của loài T. asperellum lần đầu tiên phát hiện tại miền Bắc Việt Nam đối với nấm R. solani và S. sclerotiorum và iii) đánh giá an toàn sinh học của loài T. asperellum đối với chuột.
2. PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Thu thập mẫu đất và phân lập nấm Trichoderma spp.
Các mẫu đất để phân lập nấm Trichoderma spp. được thu thập ở độ sâu 5 - 20 cm (300 g/mẫu) xung quanh gốc cây trồng khỏe ở đất trồng cây hàng năm và lâu năm của năm tỉnh miền Bắc Việt Nam gồm Hà Nội, Hải Dương, Bắc Ninh, Cao Bằng và Hòa Bình (Bảng 1). Các mẫu đất được đựng trong túi nilon, dán nhãn, mang về phòng thí nghiệm và bảo quản ở 4oC cho tới khi sử dụng.
Nấm Trichoderma spp. được phân lập theo phương pháp của Kumar et al. (2012). Theo đó, mẫu đất được pha loãng 5 lần trong nước cất vô trùng. Tiếp theo, 0,5 ml dung dịch pha loãng được nhỏ lên 5 đĩa môi trường TSM (Trichoderma Specific Medium). Các đĩa được ủ ở 28 ± 2°C trong 96 giờ. Tản nấm khác nhau về hình thái được cấy chuyển sang môi trường Potato Dextrose Agar (PDA). Nấm Trichoderma spp. được giám định sơ bộ dựa vào hình thái. Sau cùng, nấm Trichoderma spp. được phân lập thuần bằng kỹ thuật cấy đơn bào tử sử dụng kim thủy tinh với sự hỗ trợ của kính hiển vi quang học. Các mẫu nấm thuần được bảo quản ở 4°C và được sử dụng trong quá trình nghiên cứu.
2.2. Định danh nấm Trichoderma spp. bằng PCR và giải trình tự gen vùng ITS
2.2.1. Chiết DNA và giải trình tự gen vùng ITS
DNA được chiết từ các mẫu nấm Trichoderma spp. thuần sau 7 ngày nuôi cấy trên môi trường PDA theo phương pháp CTAB (Cetyltrimethyl ammonium bromide) của Doyle & Doyle (1987). Cặn DNA được hòa trong 50 µl đệm TE và bảo quản ở -20oC. Phản ứng PCR được thực hiện với cặp mồi ITS4 và ITS5 (White et al., 1990) để nhân toàn bộ vùng gen ITS của các mẫu nấm. Phản ứng PCR được thực hiện với tổng thể tích của phản ứng là 25 µl trong đó 12,5 µl 2x PCR Master mix (INtRON), 0,5 µl mồi ITS5, 0,5 µl mồi ITS4, 0,5 µl DNA khuôn và 11,0 µl dH2O. Phản ứng PCR được thực hiện theo chu trình: (94oC trong 5 phút) x 1 chu kỳ, (94o trong 40 giây, 52o trong 40 giây, 72oC trong 1 phút) x 35 chu kỳ, (72oC trong 5

Nguyễn Đức Huy, Phạm Quang Nguyên, Nguyễn Thị Thanh Hồng, Hà Giang, Nguyễn Văn Viên, Nguyễn Tất Cảnh
1595
phút) x 1 chu kỳ, dừng phản ứng ở nhiệt độ phòng. Sản phẩm PCR được tinh chiết từ agarose gel dùng kít tinh chiết thương mại DNA Gel Extraction Kit (NORGEN) theo hướng dẫn của nhà sản xuất. Hàm lượng DNA được ước lượng nồng độ bằng điện di agarose. Sản phẩm PCR được giải trình tự trực tiếp với cặp mồi dùng trong phản ứng PCR (ITS4 và ITS5) với kít BigDye® Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems) trên máy PCR (gửi thực hiện phản ứng giải trình tự tại hãng Macrogen, Hàn Quốc).
2.2.2. Phân tích trình tự và xác định tên loài nấm Trichoderma spp.
Dựa trên các trình tự thu được, tìm kiếm trên cơ sở dữ liệu trên ngân hàng gen (Genbank) bằng phần mềm trực tuyến BLAST tại NCBI (the National Center for Biotechnology Information). Trình tự các mẫu nấm thu được và các mẫu tương đồng trên ngân hàng gen được sử dụng để phân tích trình tự và phả hệ bằng các phần mềm BioEdit 7.0, ClustalX2 và MEGA 7.0.
2.3. Đánh giá hiệu lực đối kháng của Trichoderma asperellum. đối với nấm Rhizoctonia solani gây bệnh lở cổ rễ và nấm Sclerotinia sclerotiorum gây bệnh thối hạch
Các nguồn nấm gây bệnh lở cổ rễ (R. solani) được phân lập từ cây bắp cải con (2 lá thật) bị bệnh ở vườn ươm tại Bảo Thắng, Lào Cai và nấm gây bệnh thối hạch (S. sclerotiorum) được phân lập từ cây bắp cải ở giai đoạn đóng bắp tại Văn Đức, Gia Lâm, Hà Nội. Những nguồn nấm này đã được phân lập thuần, nuôi cấy và bảo quản tại phòng thí nghiệm của Bộ môn Bệnh cây, Khoa Nông học. Học viện Nông nghiệp Việt Nam. Nguồn nấm T. asperellum đã được xác định.
Thí nghiệm được thiết kế gồm 3 công thức (T) và đối chứng (C). Đối chứng, nấm gây bệnh và nấm T. asperellum được cấy riêng rẽ. Công thức 1 (T1) nấm gây bệnh và nấm T. asperellum được cấy cùng nhau và cách nhau 5 cm, công thức 2 (T2) nấm T. asperellum được cấy trước nấm R. solani (R.s) hoặc nấm S. sclerotiorum (S.s) 24 giờ và công thức 3 (T3) nấm T. asperellum được cấy sau nấm R. solani 24 giờ. Mỗi công thức được nhắc lại 3 lần và theo dõi sự phát triển của nấm T. asperellum, R. solani và S. sclerotiorum. Thí nghiệm được thực hiện theo phương pháp của Komy et al. (2015) có sửa đổi để phù hợp trong nghiên cứu này. Toàn bộ thí nghiệm được tiến hành trên đĩa Petri đường kính 8,0 cm chứa môi trường PDA và giữ ở nhiệt độ 25 ± 2oC trong 5 - 7 ngày. Sự phát triển của nấm gây bệnh được đo đường kính tản nấm hàng ngày ở cả công thức thí nghiệm và đối chứng. Hiệu lực ức chế được
tính theo công thức I = (C-T/C) x 100. Trong đó, I là phần trăm hiệu lực hực chế, C là đường kính tản nấm gây bệnh ở công thức đối chứng và T là đường kính tản nấm ở công thức thí nghiệm.
2.4. Đánh giá khả năng phát triển và hình thành bào tử phân sinh của nấm Trichoderma asperellum trên một số cơ chất
Nấm T. asperellum được sử dụng nhiều trong phòng trừ bệnh hại cây trồng có nguồn gốc trong đất. Để có số lượng sinh khối lớn nấm trong sản xuất chế phẩm sinh học, nấm T. asperellum thường được nhân nuôi trên các cơ chất. Trong nghiên cứu này, cơ chất được sử dụng để khảo sát sự phát triển và hình thành bào tử phân sinh của nấm T. asperellum là bã mía, thóc luộc, bột ngô, trấu - cám gạo và vỏ hoa diêm mạch. Khả năng hình thành sinh khối được theo dõi sau các ngày nuôi cấy và đếm số lượng bào tử phân sinh hình thành trong 1 gam cơ chất sử dụng buồng đếm hồng cầu.
2.5. Thử nghiệm tính an toàn sinh học của nấm đối khối Trichoderma asperellum
Thử nghiệm tính an toàn sinh học của nấm đối kháng T. asperellum được tiến hành trên chuột nhắt trắng giống Swiss với số lượng 40 con, cân nặng 18 - 20 gam/con, do Viện Vệ sinh dịch tễ Trung ương cung cấp. Phương pháp thử theo thường quy kỹ thuật Thử nghiệm độc tính cấp của Viện Kiểm nghiệm thuốc Trung ương VKN/TQKT-DL/03. Toàn bộ thí nghiệm được tiến hành bởi Khoa Dược lý, Viện Kiểm nghiệm thuốc Trung ương.
2.6. Xử lý số liệu
Số liệu được trình bày dưới dạng giá trị trung bình cộng trừ độ lệch chuẩn (mean ± SD) và được xử lý thống kê bằng phép phân tích biến 1 chiều (one-way ANOVA) với hậu kiểm (post-hoc) Newman-Keuls test hoặc được xử lý thống kê bằng trắc nghiệm Student sử dụng phần mềm Prism phiên bản 6.0 (Graph Pad Software). Giá trị P < 0,05 được coi là có ý nghĩa thống kê.
3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Kết quả thu thập mẫu đất để phân lập nấm đối kháng Trichoderma spp. tại năm tỉnh miền Bắc của Việt Nam
Nấm Trichoderma spp. có nhiều trong đất tự nhiên, trong đó nhiều loài như Trichoderma viride, T. harzianum, T. aperellum,... có khả năng đối kháng lại các tác nhân gây bệnh hại cây trồng có nguồn gốc trong đất như nấm Sclerotium rolfsii, Rhizoctonia solani, Fusarium solani, F. oxysporum, Pythium sp., Phytophthora nicotiana. Trong nghiên cứu này, chín mẫu đất ở các cánh đồng trồng cây hàng năm và lâu

Phân lập và đánh giá khả năng đối kháng của Trichoderma asperellum đối với tác nhân gây bệnh cây có nguồn gốc
trong đất
1596
năm (Bảng 1) đã được thu thập ở năm tỉnh miền Bắc của Việt Nam làm vật liệu để phân lập nấm Trichoderma spp.
Các mẫu đất đã thu thập được đựng trong túi nilon, dán nhãn và mang về phòng thí nghiệm bảo quản ở điều kiện 4oC cho tới khi sử dụng để phân lập nấm Trichoderma spp.
Bảng 1. Danh mục các mẫu đất thu thập để phân lập nấm Trichoderma spp. tại năm tỉnh của miền Bắc Việt Nam
Ký hiệu mẫu Địa điểm thu thập Đất trồng Giai đoạn sinh trưởng của cây
Năm Số mẫu
Vn.01 Gia Lâm, Hà Nội Tỏi 4 lá thật 2015 01
Vn.02 Lương Sơn, Hòa Bình Mía Vươn lóng 2015 01
Vn.03 Thuận Thành, Bắc Ninh Đậu tương Ra hoa 2015 01
Vn.04 Đông Anh, Hà Nội Cà chua Ra hoa 2015 01
Vn.05 Cao Bằng Diêm mạch 6 lá thật 2016 01
Vn.06 Gia Lâm, Hà Nội Tỏi 4 lá thật 2016 01
Vn.07 Kinh Môn, Hải Dương Ổi 4 năm tuổi 2016 01
Vn.08 Đông Anh, Hà Nội Rau cải Sau gieo 20 ngày 2017 01
Vn.09 Võ Cường, Bắc Ninh Cà chua Ra hoa 2017 01
3.2. Kết quả phân lập nấm đối kháng Trichoderma spp. từ các mẫu đất đã thu thập tại một số tỉnh miền Bắc
Các mẫu đất sau khi thu thập về được phơi khô, nghiền mịn và phân lập theo phương pháp của Kumar et al. (2012) đã được trình bày trong mục 2.1. Sau cùng, để làm thuần nấm Trichoderma sp., kỹ thuật cấy đơn bào tử sử dụng kim thủy tinh dưới sự hỗ trợ của kính hiển vi quang học và kỹ thuật cấy ria được sử dụng. Kết quả bước đầu cho thấy các mẫu đất thu thập được đều có nấm Trichoderma sp. dựa vào đặc điểm hình thái trên môi trường PDA (Bảng 2).
Quan sát các mẫu nấm Trichoderma sp. trên môi trường PDA cho thấy rằng lúc đầu tản nấm có màu trắng, hơi xốp và hình thành cành bào tử phân sinh. Sau 2 ngày nuôi cấy, tản nấm chuyển dần sang màu xanh, hình thành nhiều bào cành tử phân sinh. Bào tử phân sinh của nấm có dạng hình cầu hoặc hình trứng, bề mặt nhẵn. Thể bình có dạng hình trụ. Đa số các mẫu thu thập được hình thành nhiều bào tử phân sinh, sau 3 ngày nuôi cấy và đến ngày thứ 4 và thứ 5 thì nấm mọc kín đĩa và chuyển mầu xanh hoàn toàn. Ngoài ra, khi quan sát đặc điểm phát triển của nấm trên môi trường PDA, mẫu Vn.Tri08 có đặc điểm phát triển và cành bào tử phân sinh của mẫu nấm này khác so với các mẫu nấm còn lại. Vì vậy, dựa vào hình thái khó
để định danh tên loài nấm Trichoderma sp.
Như vậy, dựa vào các đặc điểm phát triển của tản nấm trên môi trường PDA, đặc điểm cành bào tử phân sinh và thể bình hình thành sau nuôi cấy trên môi trường PDA, 2 loài nấm Trichoderma được phân lập từ các mẫu đất đã thu thập. Tiếp theo, kỹ thuật giải trình tự dựa trên vùng gen ITS được sử dụng để xác định chính xác tên loài.
3.3. Kết quả xác định các mẫu nấm đối kháng dựa vào giải trình tự gen vùng rDNA-ITS
Các mẫu nấm sau khi được xác định là Trichoderma sp. dựa vào hình thái đã được sử dụng tiếp tục để định danh chính xác tên loài dựa vào giải trình tự vùng ITS (internal transcribed spacer). Chiết DNA của nấm và thực hiện phản ứng PCR như đã mô tả trong phương pháp nghiên cứu (mục 2.2). Kết quả điện di sản phẩm PCR cho thấy tất cả 8 mẫu Trichoderma sp. đều xuất hiện băng sản phẩm với kích thước khoảng 600 bp (Hình 1).
Tám băng sản phẩm PCR đã được cắt và thôi gel để tinh sạch DNA và giải trình tự gen theo mô tả ở mục 2.2 và 2.3 phần phương pháp nghiên cứu. Kết quả giải trình tự gen cho thấy 8 mẫu nấm đều có chất lượng trình tự tốt, rõ nét, không bị nhiễu và trình tự đọc được là 500 - 550 nts (nucleotides). Các trình tự này được tìm kiếm
Bảng 2. Kết quả phân lập nấm đối kháng Trichoderma sp. từ các mẫu đất đã thu thập
Ký hiệu mẫu
Địa điểm thu thập Đặc điểm tản nấm trên môi trường PDA Kết quả phân lập
Vn.Tri01 Gia Lâm, Hà Nội Xanh, nhiều BTPS*, thể bình mọc thành chùm 2 - 3 cái
(Hình 3b) Trichoderma sp.
Vn.Tri02 Lương Sơn, Hòa Bình Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp.

Nguyễn Đức Huy, Phạm Quang Nguyên, Nguyễn Thị Thanh Hồng, Hà Giang, Nguyễn Văn Viên, Nguyễn Tất Cảnh
1597
Vn.Tri03 Thuận Thành, Bắc Ninh Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp. Vn.Tri04 Đông Anh, Hà Nội Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp. Vn.Tri05 Cao Bằng Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp. Vn.Tri06 Gia Lâm, Hà Nội Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp. Vn.Tri07 Kinh Môn, Hải Dương Xanh, nhiều BTPS¸ thể bình mọc thành chùm 2 - 3 cái Trichoderma sp. Vn.Tri08 Đông Anh, Hà Nội Xanh, nhiều BTPS, thể bình mọc đỗi xứng (Hình 3c) Trichoderma sp. Vn.Tri09 Võ Cường, Bắc Ninh Xanh, nhiều BTPS, thể bình mọc thành chùm 2 - 3 cái Trichoderma sp.
Ghí chú: *Bào tử phân sinh
Hình 1. Sản phẩm điện di 08 mẫu nấm đối kháng Trichoderma sp. Vn.Tri02-Vn.Tri09
Ghi chú: Kích thước sản phẩm PCR khoảng 600 bp. Thang DNA 1 kb. Mẫu Vn.Tri01 được thực hiện tại Đại học Shimane, Nhật Bản
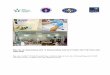
kiếm và so sánh với các chuỗi tương đồng trên Ngân hàng Gen (GenBank) bằng sử dụng phần mềm trực tuyến BLAST của NCBI. Kết quả tìm kiếm các trình tự tương đồng cho thấy rằng trình tự của 7 mẫu nấm đã thu thập đều là các chuỗi mã hóa cho vùng gen ITS của nấm T. asperellum, 1 mẫu nấm là T. harzianum. Phần trăm đoạn so sánh là 100% có mức đồng nhất trình tự là 99 - 100% với các mẫu T. asperellum và T. harzianum (Hình 2). Mẫu Vn.Tri01 được thực hiện tại Đại học Shimane của Nhật Bản cũng cho kết quả là T. asperellum.Phân tích phả hệ các mẫu nấm Trichoderma spp. đã thu thập cho thấy mẫu nấm phân lập ở 5 tỉnh miền Bắc thuộc 2 cụm T. asperellum và T. harzianum (chữ in đậm trong hình 2).
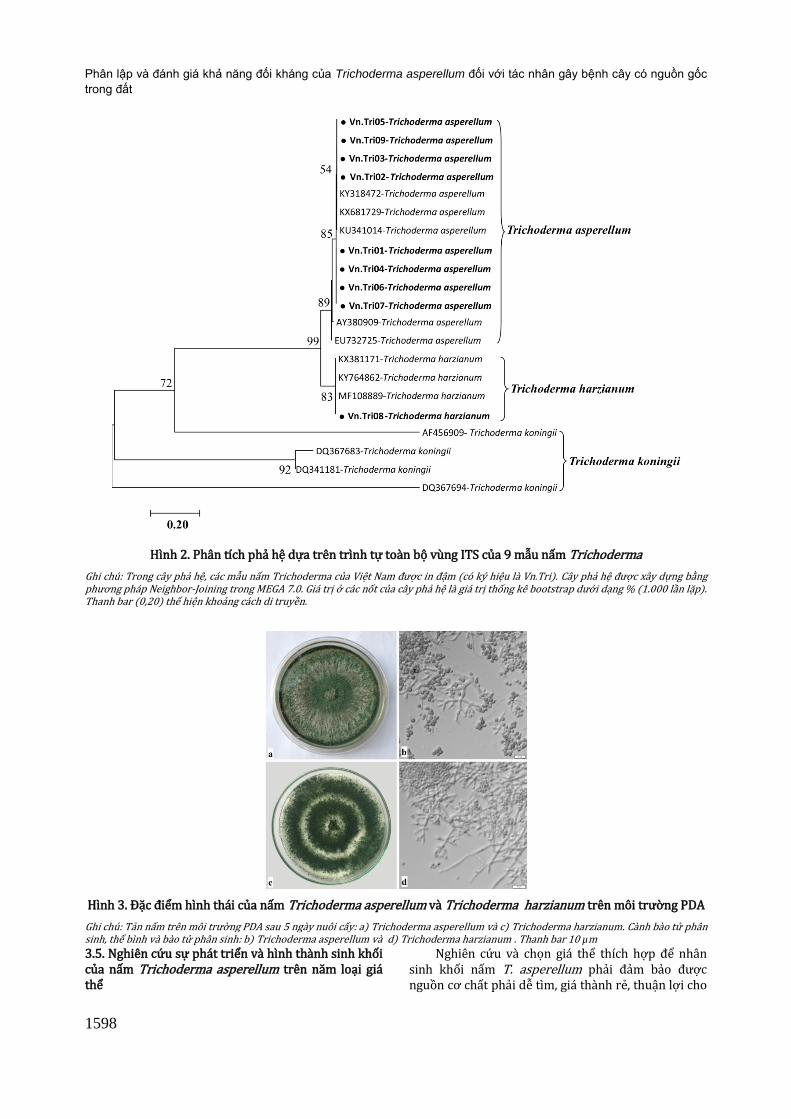
Nghiên cứu này đã xác định được 8 mẫu là T. asperellum, 1 mẫu là T. harzianum. Trên môi trường PDA, mẫu Vn.Tri08 (T. harzianum) có tản nấm phát triển mịn và thưa hơn so với tản nấm của các mẫu Vn.Tri01-07, 09 (T. asperellum) (Hình 3a và c), thể bình hình thành trên cành bào tử phân sinh mọc không thành chùm 2 - 3 thể bình như T. asperellum và thường đối xứng nhau (Hình 3b và d). Nấm T. asperellum cũng được phát hiện trên đất trồng cao su của Tây Nguyên (Phạm Ngọc Dung và cs., 2012), Quảng Trị, Thừa Thiên Huế (Nguyễn Hoàng Lộc và cs., 2011) và một số tỉnh miền Đông Nam Bộ (mới chỉ công bố trình tự trên Ngân hàng gen, NCBI). Như vậy, nấm T. asperellum phân bố khá phổ biến ở Việt
Nam và là lần đầu tiên được phát hiện trong đất trồng cây hàng năm và cây lâu năm ở một số tỉnh thuộc đồng bằng sông Hồng và miền núi phía Bắc của Việt Nam. Chính vì vậy, các nghiên cứu tiếp theo trong nghiên cứu này, chúng tôi sử dụng mẫu nấm T. asperellum Vn.Tri01.
3.4. Đặc điểm hình thái nấm Trichoderma asperellum và Trichoderma harzianum
Nấm T. asperellum được nuôi cấy trên môi trường PDA ở nhiệt độ 25 ± 2oC, quan sát sự phát triển của nấm, đặc điểm của tản nấm, cành bào tử phân sinh và bào tử phân sinh. Tản nấm T. asperellum phát triển nhanh, dầy và xốp hơn của T. harzianum (Hình 3a và c), sau 4 - 5 ngày nuôi cấy, nấm phát triển được 9,0 cm và hình thành nhiều bào tử phân sinh. Thể bình của nấm T. asperellum mọc đối xứng và thành chùm 2 - 3 thể bình trên đầu cành bào tử phân sinh (Hình 3a) so với của T. harzianum (Hình 3d). Từ thể bình hình thành các bào tử phân sinh, bào tử phân sinh đều tròn tròn hoặc hình trứng. Kích thước bào tử trung bình 2,9 - 3,1 x 2,3 - 3,3 μm (Hình 3b).
Với các đặc điểm phát triển nhanh và hình thành số lượng bào tử phân sinh lớn, nấm T. asperellum có thể được ứng dụng để phòng trừ nhiều nấm gây bệnh cây quan trọng có nguồn gốc trong đất. Để ứng dụng được ngoài sản xuất, việc thử nghiệm sự phát triển của nấm trên các giá thể để nhân nhanh sinh khối nấm là cần thiết.
Cụm loài
Cụm loài
Phân lập và đánh giá khả năng đối kháng của Trichoderma asperellum đối với tác nhân gây bệnh cây có nguồn gốc
trong đất
1598
Hình 2. Phân tích phả hệ dựa trên trình tự toàn bộ vùng ITS của 9 mẫu nấm Trichoderma
Ghi chú: Trong cây phả hệ, các mẫu nấm Trichoderma của Việt Nam được in đậm (có ký hiệu là Vn.Tri). Cây phả hệ được xây dựng bằng phương pháp Neighbor-Joining trong MEGA 7.0. Giá trị ở các nốt của cây phả hệ là giá trị thống kê bootstrap dưới dạng % (1.000 lần lặp). Thanh bar (0,20) thể hiện khoảng cách di truyền.
Hình 3. Đặc điểm hình thái của nấm Trichoderma asperellum và Trichoderma harzianum trên môi trường PDA
Ghi chú: Tản nấm trên môi trường PDA sau 5 ngày nuôi cấy: a) Trichoderma asperellum và c) Trichoderma harzianum. Cành bào tử phân sinh, thể bình và bào tử phân sinh: b) Trichoderma asperellum và d) Trichoderma harzianum . Thanh bar 10 µm
3.5. Nghiên cứu sự phát triển và hình thành sinh khối của nấm Trichoderma asperellum trên năm loại giá thể
Nghiên cứu và chọn giá thể thích hợp để nhân sinh khối nấm T. asperellum phải đảm bảo được nguồn cơ chất phải dễ tìm, giá thành rẻ, thuận lợi cho
Nguyễn Đức Huy, Phạm Quang Nguyên, Nguyễn Thị Thanh Hồng, Hà Giang, Nguyễn Văn Viên, Nguyễn Tất Cảnh
1599
sự sinh trưởng, phát triển và sinh sản của nấm. Trong nghiên cứu này, năm loại giá thể: bã mía, vỏ hoa cây diêm mạch, bột ngô, trấu - cám và thóc luộc được sử dụng để nhân sinh khối nấm T. asperellum (Bảng 3, Hình 4).
Kết quả cho thấy rằng sau 3 - 4 ngày các sợi nấm màu trắng đã phát triển trên bề mặt giá thể, sau đó chúng chuyển dần sang màu xanh nhạt (hình thành bào tử phân sinh) và màu xanh đậm (lượng bào tử phân sinh được hình thành lớn nhất). Giá thể thóc luộc có độ xốp cao, thoáng khí hơn và hàm lượng B1 cao nên mật độ bào tử hình thành cao nhất (7,8 x 109 cfu/g) sau 11 ngày nhân nuôi. Tiếp đến là giá thể bột ngô và trấu
cám có mật độ bào tử lần lượt là 7,2 x 108 cfu/g và 7,0 x 108 cfu/g. Nấm T. asperellum không phát triển được trên bã vỏ hoa cây diêm mạch, nguyên nhân có thể do vỏ hoa cây diêm mạch có chứa chất ức chế sự phát triển của nấm.
3.6. Đánh giá hiệu lực đối kháng của nấm Trichoderma asperellum đối với nấm Rhizoctonia solani gây bệnh lở cổ rễ và Sclerotinia sclerotiorum gây bệnh thối hạch cải bắp
Một trong những biện pháp hiệu quả để phòng trừ nhóm nấm gây bệnh hại cây trồng có nguồn gốc trong đất là sử dụng các vi sinh vật đối kháng. Trong nghiên cứu này, nấm T. asperellum được sử dụng để đánh giá khả năng ức chế sự phát triển của nấm R. solani gây bệnh lở cổ rễ cây và nấm S. sclerotiorum gây bệnh thối hạch cải bắp trong điều kiện in vitro (Bảng 4, 5 và Hình 5a-b).
Bảng 3. Sự phát triển và hình thành bào tử (cfu/g) của nấm Trichoderma asperellum trên năm loại giá thể
Ngày theo dõi Giá thể
Bã mía Vỏ hoa cây diêm mạch Bột ngô Trấu - cám Thóc luộc
3 - - - - -
5 - - 2,4 x 104
1,9 x 103
2,3 x 104
7 1,1 x 102 - 4,5 x 10
5 4,3 x 10
6 4,9 x 10
6
9 1,8 x 103 - 5,5 x 10
7 5,1 x 10
8 5,3 x 10
8
11 2,5 x 103 - 7,2 x 10
8 7,0 x 10
8 7,8 x 10
9
Ghi chú: - chưa/không phát triển
Bảng 4. Hiệu lực đối kháng của nấm Trichoderma asperellum đối với nấm Rhizoctonia solani trên môi trường PDA
Ngày theo dõi
Đường k nh tản nấm (mm)
C T1 T2 T3
Tri R.s Tri R.s HLĐK Tri R.s HLĐK Tri R.s HLĐK
1 31,0 30,3 35,0 23,0 24,2 35,0 0 100 0 29,3 3,40
2 50,0 56,5 46,3 10,7 81,1 49,3 10,0 82,3 22,0 50,3 11,0
3 80,0 80,0 74,0 6,0 92,5 71,5 0 100 34,5 45,5 43,1
4 - - 80,0 0 100 80,0 - - 44,0 36,0 55,0
5 - - - - - - - - 45,0 35,0 56,3
6 - - - - - - - - 49,0 31,0 61,3
7 - - - - - - - - 75,0 5,0 93,8
Ghi chú: HLĐK: Hiệu lực đối kháng, T1: công thứ thí nghiêm 1 (nấm T. asperellum (Tri) và nấm R. solani (R.s) được cấy cùng thời gian), T2: công thức thí nghiệm 2 (nấm Tri được cấy trước nấm R.s 24 giờ) và T3: công thức thí nghiệm 3 (nấm Tri được cấy sau nấm R.s 24 giờ).

Phân lập và đánh giá khả năng đối kháng của Trichoderma asperellum đối với tác nhân gây bệnh cây có nguồn gốc
trong đất
1600
Hình 4. Nhân nuôi nấm Trichoderma asperellum trên năm loại giá thể
Ghi chú: a) bã mía, b) vỏ hoa cây diêm mạch, c) trấu - cám, d) bột ngô và e) thóc luộc
Kết quả cho thấy nấm T. asperellum cấy trước nấm R. solani 24 thì hiệu quả đối kháng đạt cao nhất (100%) sau 4 ngày nuôi cấy (Bảng 4, Hình 5a). Nấm T. asperellum lấn át và phát triển lan sang cả nấm R. solani. Như vậy, khi xử lý chế phẩm nấm T. asperellum ngoài đồng ruộng nên xử lý hoặc bón vào đất trước khi gieo hạt/trồng cây 2 - 3 ngày, nấm T. asperellum sẽ phát triển và ngăn chặn nấm gây bệnh khi hạt được gieo hoặc cây được trồng vào đất và sẽ cho hiệu quả phòng trừ bệnh cao. Những nghiên cứu trước đây cũng cho thấy rằng nấm T. asperellum có hiệu quả đối kháng cao đối với nấm R. solani (Asad et al., 2014; Yang., 2014). Theo đó, T. asperellum có thể tiết enzyme phá hủy vách tế bào
sợi nấm R. solani hoặc cũng có thể ký sinh trực tiếp và giết chết sợi nấm R. solani.
Cùng với nấm R. solani, nấm S. sclerotiorum cũng gây bệnh thối hạch trên nhiều cây trồng và gây thiệt hại có ý nghĩa kinh tế như cây cải bắp. Trong nghiên cứu này, hiệu lực đối kháng của nấm T. asperellum cũng được thử nghiệm đối với nấm S. sclerotiorum (Bảng 5, Hình 5b). Nghiên cứu cũng cho thấy kết quả tương tự, khi nấm T. asperellum được cấy trước nấm S. sclerotiorum 24 giờ thì liệu lực ức chế đạt 100% sau 3 ngày nuôi cấy. Tản nấm T. asperellum cũng lấn át và xâm lấn hết tản nấm S. sclerotiorum (Hình 5b). Nghiên cứu của Lopes et al. (2012) cũng cho thấy nấm T. asperellum có khả năng tiêu diệt nấm S. sclerotiorum.
Bảng 5. Hiệu lực đối kháng của nấm Trichoderma asperellum đối với nấm Sclerotinia sclerotiorum trên môi trường PDA
Ngày theo dõi
Đường k nh tản nấm (mm)
C T1 T2 T3
Tri S.s Tri S.s HLĐK Tri S.s HLĐK Tri S.s HLĐK
1 31,0 20,0 32,0 24,0 - 30,0 0 - 0 20,1 79,9

Nguyễn Đức Huy, Phạm Quang Nguyên, Nguyễn Thị Thanh Hồng, Hà Giang, Nguyễn Văn Viên, Nguyễn Tất Cảnh
1601
2 50,0 48,0 60,0 20,0 58,3 57,0 11,0 81,1 38,0 43,7 46,1 3 80 67,0 80 0 100 80 0 100 39,5 36,3 63,7 4 - 80 - - - - - - 46,0 30,7 69,3
5 - - - - - - - - 64,0 18,3 81,7
Ghi chú: HLĐK: Hiệu lực đối kháng, C: công thức đối chứng (cấy T. asperellum (Tri) và nấm S. sclerotiorum (S.s) riêng. T1: công thứ thí nghiêm 1 (nấm Tri và nấm S.s được cấy cùng thời gian), T2: công thức thí nghiệm 2 (nấm Tri được cấy trước nấm S.s 24 giờ) và T3: công thức thí nghiệm 3 (nấm Tri được cấy sau nấm S.s 24 giờ).
a
b
Hình 5. a) Khả năng đối kháng của nấm Trichoderma asperellum đối với nấm Rhizoctonia solani trên môi trường PDA và b) Khả năng đối kháng
của nấm T. asperellum đối với nấm Sclerotinia sclerotiorum trên môi trường PDA
Ghi chú: Từ trái quá phải, từ trên xuống dưới: a) đối chứng (nấm R. solani), đối chứng (nấm T. asperellum), T. asperellum cấy trước nấm R. solani 24 giờ, T. asperellum cấy cùng nấm R. solani và T. asperellum cấy sau nấm R. solani 24 giờ; b) đối chứng (nấm T. asperellum), đối chứng (nấm S. sclerotiorum), T. asperellum cấy trước nấm S. sclerotiorum 24 giờ, T. asperellum cấy sau nấm S. sclerotiorum 24 giờ và T. asperellum cấy cùng nấm S. sclerotiorum.
3.7. Đánh giá an toàn sinh học của nấm Trichoderma asperellum đối với chuột
Căn cứ vào kết quả đánh giá hiệu lực đối kháng của nấm T. asperellum đối với hai nấm R. solani và S. sclerotiorum cho thấy hiệu lực đối kháng đạt 100% (Bảng 4, 5). Các nghiên cứu trên thế giới cũng cho thấy T. asperellum có khả năng tiêu diệt nhiều nấm bệnh hại cây trồng có nguồn gốc trong đất. Như vậy, có thể ứng dụng T. asperellum để phòng trừ nhiều bệnh nấm hại cây trồng quan trọng trong sản xuất. Khi ứng dụng ngoài sản xuất, việc đánh giá an toàn sinh học của nấm này đối với động vật là cần thiết.
Thí nghiệm được thực hiện trên chuột nhắt trắng như đã mô tả ở phương pháp nghiên cứu. Kết quả thí nghiệm trình bày ở bảng 6.
Kết quả theo dõi khối lượng trung bình của chuột trong quá trình thử nghiệm 7 ngày cho thấy: Trước khi uống mẫu thử, khối lượng trung bình của chuột không có sự khác biệt có ý nghĩa thống kê so với nhau và so với nhóm đối chứng (PANOVA trước > 0,05; P (T-C) trước > 0,05; P (T-T) trước > 0,05). Sau uống mẫu thử 7 ngày, chuột ở nhóm đối chứng và nhóm thử đều tăng cân. Có sự khác
Bảng 6. Kết quả so sánh khối lượng chuột giữa nhóm đối chứng và các nhóm thử nghiệm cho uống nấm đối kháng Trichoderma asperellum
Nhóm Trước thử nghiệm Sau thử nghiệm Tăng khối
lượng (%) Ptrước-sau
Khối lượng (g) Ptrước thử nghiệm Khối lượng (g) Psau thử nghiệm
Đối chứng (C) 20,8 ± 0,8 PANOVA > 0,05 31,9 ± 1,8 PANOVA > 0,001 153,6 P < 0,001 Thử 1 (T1) 20,4 ± 0,6 PT1-C > 0,05
PT1-T2 > 0,05 PT1-T3 > 0,05
30,4 ± 1,9 PT1-C > 0,05 PT1-T2 > 0,05 PT1-T3 < 0,001
149,4 P < 0,001
Thử 2 (T2) 20,7 ± 0,8 PT2-C > 0,05 PT2-T3 > 0,05
30,6 ± 1,9 PT2-C > 0,05 PT2-T3 < 0,001
147,5 P < 0,001
Thử 3 (T3) 20,5 ± 0,8 PT3-C > 0,05 30,1 ± 2,5 PT3-C < 0,001 129,3 P < 0,001
khác biệt có ý nghĩa thống kê về khối lượng của chuột khi so sánh với trước thử nghiệm ở mỗi nhóm
(Ptrước-sau < 0,001). Đồng thời có sự khác biệt có ý nghĩa thống kê về cân nặng trung bình sau thử

Phân lập và đánh giá khả năng đối kháng của Trichoderma asperellum đối với tác nhân gây bệnh cây có nguồn gốc
trong đất
1602
nghiệm giữa nhóm thử liều cao so với 2 nhóm liều thấp hơn và so với nhóm đối chứng (PANOVA sau < 0,001; P (T3-C) sau < 0,001; P (T3-T1) sau < 0,001, P (T3-T2)
sau < 0,001). Không có sự khác biệt về cân nặng trung bình giữa 2 nhóm thử liều thấp (T2 và T3) với nhau và so với nhóm đối chứng (P (T-C) sau > 0,05; P
(T2-T3) sau > 0,05).
Như vậy, cho chuột uống mẫu thử với mức liều 10 g và 20 g mẫu thử/kg chuột không nhận thấy biểu hiện khác thường so với nhóm đối chứng, không nhận thấy có biểu hiện ngộ độc trên chuột thí nghiệm trong thời gian theo dõi. Tất cả chuột đều ăn uống, hoạt động bình thường.
4. KẾT LUẬN
Đã xác định được hai loài nấm đối kháng T. asperllum và T. harzianum dựa vào đặc điểm hình thái và trình tự vùng gen ITS. Nấm T. asperllum phân bố phổ biến ở miền Bắc Việt Nam. Nấm T. asperllum phát triển nhanh và hình thành sinh khối bào tử phân sinh nhiều nhất trên cơ chất thóc luộc (7,8 x 109 cfu/g), bột ngô (7,2 x 108 cfu/g) và trấu cám (7,0 x 108 cfu/g). Nấm T. asperllum có khả năng ức chế tốt sự phát triển của nấm R. solani gây bệnh lở cổ rễ và nấm S. sclerotiorum gây bệnh thối hạch bắp cải trong điều kiện invitro, mức độ ức chế là 100% sau 3 - 4 ngày nuôi cấy) Nấm T. asperellum an toàn đối với chuột ở liều lượng 20 g chế phẩm/kg chuột.
LỜI CẢM ƠN
Nghiên cứu này được tài trợ bởi đề tài trọng điểm cấp Học viện mã số T2015-01-01TĐ. Chúng tôi cũng xin chân thành cảm ơn GS. TS. Makoto Ueno, Trường đại học Shimane, Nhật Bảnđã giúp đỡ xác định mẫu Vn.Tri01 và Khoa Dược lý, Viện Kiểm nghiệm thuốc Trung ương đã hỗ trợ kỹ thuật đánh giá tính an toàn sinh học của nấm T. asperellum đối với chuột.
TÀI LIỆU THAM KHẢO
Anees M., Tronsmo A., Edel-Hermann V., Hjeljord
L.G., Héraud C. and Steinberg C. (2010).
Characterization of field isolates of Trichoderma
antagonistic against Rhizoctonia solani. Fungal
Biol., 114(9): 691-701.
Asad S.A., Ali N., Hameed A., Khan S.A., Ahmad R.,
Bilal M., Shahzad M. and Tabassum A. (2014).
Biocontrol efficacy of different isolates of
Trichoderma against soil borne pathogen Rhizoctonia
solani. Pol. J. Microbiol., 63(1): 95-103.
Doyle J.J. and Doyle J.L. (1987). A rapid DNA
isolation procedure for small quantities of fresh
leaf tissue. Phytochem Bull., 19: 11-15.
Harman G.E. (2006). Overview of mechanisms and uses
of Trichoderma spp. Phytopathol., 96: 190-194.
Komy M.H.E., Saleh A.A., Eranthodi A. and Molan
Y.Y. (2015). Characterization of
Novel Trichoderma asperellum Isolates to Select
Effective Biocontrol Agents Against Tomato
Fusarium Wilt. Plant Pathol J., 31(1): 50-60.
Kotasthane A., Agrawal T., Kushwah R. and Rahatkar
O.V. (2015). In-vitro antagonism of Trichoderma
spp. against Sclerotium rolfsii and Rhizoctonia
solani and their response towards growth of
cucumber, bottle gourd and bitter gourd.
European J. of Plant Pathol., 141(3): 523-543.
Kumar K., Amaresan N., Bhagat S., Madhuri K. and
Srivastava R.C. (2012). Isolation and
Characterization of Trichoderma spp. for
Antagonistic Activity Against Root Rot and Foliar
Pathogens. Indian J. Microbiol., 52(2): 137-144.
Lopes F.A., Steindorff A.S., Geraldine A.M., Brandão
R.S., Monteiro V.N., Lobo M. Jr., Coelho A.S.,
Ulhoa C.J. and Silva R.N. (2012). Biochemical and
metabolic profiles of Trichoderma strains isolated
from common bean crops in the Brazilian Cerrado,
and potential antagonism against Sclerotinia
sclerotiorum. Fungal Biol., 116(7): 15-24.
Mbarga J.B., Hoopen G.M.T, Kuaté J., Adiobo A.,
Ngonkeu M.E.L., Ambang Z., Akoa A., Tondje
P.R. and B.A.D. Begoude. (2012). Trichoderma
asperellum: A potential biocontrol agent
for Pythium myriotylum, causal agent of cocoyam
(Xanthosoma sagittifolium) root rot disease in
Cameroon. Crop Protection, 36: 18-22.
Nguyen Hoang Loc, Hoang Tan Quang, Nguyen Bao
Hung, Nguyen Duc Huy, Truong Thi Bich Phuong
and Tran Thi Thu Ha (2011). Trichoderma
asperellum Chi42 Genes Encode Chitinase.
Mycobiology, 39(3): 182-186.
ùi V n Công
(2012) cứ sả x ất sử dụ c ế p ẩm
ấm đố k á Trichoderma viride p ò trừ một số
bệ ấm ạ ù r câ khoai tây, lạc đậ
tươ . ạp c í K oa ọc P át tr ể , 10(1): 95-102.
P ạm ọc ết ư ao
a r ư oa
ạ (2012). cứ sử dụ
ấm đố k á Trichoderma asperellum trong
p ò trừ ấm Phytophthora spp. â bệ tr
câ cao s . ạp c í Bảo ệ t ực ật ộ ô
ệp P át tr ể ô t ô 12.
Rojo F.G., Reynoso M.M., Ferez M. and Chulze S.N.
(2007). Biological control by Trichoderma species