Embed Size (px)
Citation preview

1
AISLAMIENTO Y SELECCIÓN DE RIZOBACTERIAS PROMOTORAS DE CRECIMIENTO
VEGETAL EN CULTIVOS DE UCHUVA (Physalis peruviana L.) CON CAPACIDAD
ANTAGÓNICA FRENTE A Fusarium sp.
CRIS VENNER RODRÍGUEZ MARÍA JOSÉ MARTIN HERNÁNDEZ
TRABAJO DE GRADO Presentado como requisito parcial para optar al título de
MICROBIÓLOGO AGRÍCOLA Y VETERINARIO y MICROBIÓLOGO INDUSTRIAL
DIRECTOR
María Ximena Rodríguez Bocanegra Ph.D.
PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS CARRERA DE MICROBIOLOGÍA AGRÍCOLA Y VETERINARIA
CARRERA DE MICROBIOLOGÍA INDUSTRIAL BOGOTÁ, D.C.
DICIEMBRE, 2009

2
AISLAMIENTO Y SELECCIÓN DE RIZOBACTERIAS PROMOTORAS DE CRECIMIENTO
VEGETAL EN CULTIVOS DE UCHUVA (Physalis peruviana L.) CON CAPACIDAD
ANTAGÓNICA FRENTE A Fusarium sp.
APROBADO
______________________ ________________________
María Ximena Rodríguez Ph.D Pedro Jimenez Ph.D
Directora Jurado

3
AISLAMIENTO Y SELECCIÓN DE RIZOBACTERIAS PROMOTORAS DE CRECIMIENTO
VEGETAL EN CULTIVOS DE UCHUVA (Physalis peruviana L.) CON CAPACIDAD
ANTAGÓNICA FRENTE A Fusarium sp.
APROBADO
_______________________ ________________________
INGRID SHULER JANETH ARIAS
Decana Académica Directora de Carrera
Facultad de Ciencias Facultad de Ciencias

4
NOTA DE ADVERTENCIA Artículo 23 de la Resolución N° 13 de Julio de 1946 ―La Universidad no se hace responsable por los conceptos emitidos por sus
alumnos en sus trabajos de tesis. Solo velará por que no se publique nada contrario
al dogma y a la moral católica y por que las tesis no contengan ataques personales
contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la
justicia‖.

5
A Dios y a mis padres, por su apoyo incondicional y su amor inmenso
María José Martin Hernández
A mis padres, por su amor y paciencia, a mi hermano y a mi flaquito por acompañarme durante esta etapa. Cris Venner Rodríguez

6
AGRADECIMIENTOS
A Maria Ximena Rodríguez por su infinita paciencia y su guía durante este largo pero gratificante proceso. A todos los niños de UNIDIA por soportarnos en nuestras eternas tardes de trabajo. A Javi por su apoyo y toda la comida. A Carel y Andrés por cuidar de nuestras lindas uchuvitas. A todas las niñas de monitoria, a Marquitos y a Ruca por sacarnos de apuros tantas veces.

7
Tabla de contenido RESUMEN .................................................................................................................. 11
1. INTRODUCCIÓN ................................................................................................. 12
2. MARCO TEÓRICO .............................................................................................. 12
2.1 Generalidades de la Uchuva .................................................................. 12
2.2 Enfermedades de la Uchuva .................................................................. 13
2.3 Reguladores de crecimiento vegetal ..................................................... 14
2.3.1 Auxinas ................................................................................................................ 15
2.4. Rizobacterias promotoras del crecimiento vegetal (PGPR) ................ 15
2.4.1. Pseudomonas fluorescens .............................................................................. 17
2.4.2. Bacillus sp. ......................................................................................................... 17
2.4.3. Actinomycetes ................................................................................................... 17
3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN ...................................... 18
3.1. Formulación del Problema ..................................................................... 18
3.2. Justificación de la investigación ........................................................... 18
4. OBJETIVOS ............................................................................................................ 19
4.1. Objetivo General ..................................................................................... 19
4.2 Objetivos específicos .............................................................................. 19
5. MATERIALES Y MÉTODOS .................................................................................. 19
5.1. Diseño de Investigación ......................................................................... 19
5.1.1. Población de estudio y muestra de población de estudio ........................... 20
5.1.2. Variables de estudio ........................................................................................ 20
5.2. Metodología............................................................................................. 20
5.2.1. Salida de campo ................................................................................................ 20
5.2.2. Procesamiento de muestras ............................................................................ 20
5.2.3. Aislamiento e identificación de Pseudomonas fluorescens ........................ 21
5.2.4. Aislamiento y selección de Bacillus sp. ......................................................... 21
5.2.5. Aislamiento y selección de Actinomycetes ................................................... 22
5.2.6. Pruebas de germinación para determinar promoción de crecimiento
vegetal de las cepas aisladas .................................................................................... 22
5.2.6.1. Cámara Húmeda ............................................................................................ 22

8
5.2.6.2. Imbibición de las semillas ............................................................................. 22
5.2.6.3. Preparación de inóculos. .............................................................................. 22
5.2.7 Pruebas de antagonismo in vitro frente a Fusarium sp. para determinar la
actividad biocontroladora de los aislamientos. ........................................................ 23
5.2.7.1. Preparación de inóculo ................................................................................. 23
5.2.7.2. Montaje de la prueba de antagonismo ....................................................... 23
5.2.8. Cuantificación de la producción de AIA ......................................................... 24
5.2.8.1. Curva de calibración del reactivo de Salkowski. ....................................... 24
5.2.8.2. Cultivo discontinuo en caldo BT de las cepas aisladas y cuantificación
de AIA a partir de los mismos .................................................................................... 24
5.2.9. Cuantificación de la producción de sideróforos ............................................ 24
5.2.9.1. Curva de calibración Cromo Azurol S (CAS)............................................. 24
5.2.9.2. Cultivo discontinuo en medio mínimo (Simmon y Tessman) y
cuantificación de sideróforos a partir de los mismos .............................................. 24
5.2.10. Pruebas de promoción de crecimiento en invernadero ............................ 24
5.2.11. Desarrollo del banco de germoplasma ........................................................ 25
5.3. Análisis de información.......................................................................... 25
6. RESULTADOS Y DISCUSION............................................................................... 25
6.1. Aislamiento e identificación bioquímica de cepas aisladas ................ 25
6.2. Producción de Acido indol-acético ....................................................... 27
6.3. Promoción de germinación .................................................................... 28
6.4. Promoción de crecimiento en invernadero ........................................... 30
6.5. Cuantificación de sideróforos de las cepas aisladas ........................... 31
6.6. Pruebas de antagonismo frente a Fusarium sp .................................... 34
6.7. Discusión general ................................................................................... 36
7. CONCLUSIONES ................................................................................................... 38
8. RECOMENDACIONES ........................................................................................... 40
9. REFERENCIAS ...................................................................................................... 40
10. ANEXOS ............................................................................................................... 45

9
INDICE DE TABLAS
INDICE DE FIGURAS
Figura 1. Longitud promedio en tallo y raíz de las plántulas de uchuva tratadas en invernadero
30
Figura 2. Peso seco promedio y número de brotes de las plántulas de uchuva tratadas en invernadero.
31
Figura 3. Cultivo discontinuo en medio mínimo Simmon y Tessman.
33
Figura 4. Imágenes de la prueba de Antagonismo 35
INDICE DE ANEXOS
Anexo 1. Agrupación Duncan para producción de AIA
45
Anexo 2. Agrupación Duncan para GCR
46
Anexo 3. Análisis de correlación entre variables AIA y GCR
53
Tabla 1. Resultados de la identificación bioquímica. 1° y 2° muestreo.
26
Tabla 2. Producción promedio de AIA (μg/mL) por las cepas bacterianas aisladas.
27
Tabla 3. Índice de Germinación Relativo promedio presentado en cada estadio en los tratamientos con las diferentes cepas aisladas.
28
Tabla 4. Concentración de sideróforos producidos por las cepas aisladas
32
Tabla 5. Porcentaje de inhibición de las cepas aisladas frente a Fusarium sp.
34

10
Anexo 5. Agrupación Duncan para porcentajes de inhibición.
55
Anexo 6. Correlación producción de Sideróforos y porcentajes de inhibición
57
Anexo 7. Agrupación Duncan para Resultados de promoción de crecimiento en invernadero.
57
Anexo 8. Relación entre las cepas destacadas en las diferentes pruebas, y las identificadas.
59

11
RESUMEN
Con el fin de aislar, identificar y seleccionar rizobacterias promotoras del crecimiento vegetal
que además de promover la germinación y el crecimiento de las plantas de uchuva, también
tuvieran un efecto inhibitorio sobre el patógeno Fusarium oxysporum, se realizaron dos
muestreos, de la zona productora de uchuva en Silvania Cundinamarca, en fincas en donde
se presentaba la enfermedad marchitez vascular, causada por fitopatógeno mencionado
anteriormente, pero a partir de plantas sanas. Estas muestras fueron procesadas para
realizar un aislamiento selectivo de Pseudomonas fluorescens, bacterias del género Bacillus
y de Actinomyces. A partir de estos aislamientos se procedió a realizar pruebas de
germinación, en cámara húmeda las cuales tuvieron un seguimiento de 25 días, también
pruebas de producción de ácido indol acético (AIA) en caldo BT y cuya cuantificación se
realizó mediante el reactivo de Salkowky. También se cuantificó la producción de sideróforos
en el caldo Simmon y Tessman, usando del reactivo de CAS para la detección de los
mismos. Además se realizaron pruebas de antagonismo, mediante la técnica de
enfrentamiento dual, en agar PDA. Por último se realizó una prueba de promoción de
crecimiento en invernadero, las cuales tuvieron un seguimiento de un mes, evaluando al final
variables como peso seco, longitud de tallo y raíz y número de brotes, todo esto con las
cepas aisladas en el primer muestreo. Los resultados obtenidos muestran que las mejores
cepas en cuanto a la producción de AIA, fueron O23, OS5 y OS3, en la producción de
sideróforos se destacaron las cepas, O23, O181, O5, OS17 y OS3. Las pruebas de
antagonismo mostraron que las mejores cepas fueron C2, O24, O26 y SB7. En las pruebas
de germinación se comparó el índice de germinación relativo (GCR), teniendo como
referente a los controles con agua destilada estéril, se estableció que en los estadios 1 y 2
no hay diferencia significativas entre las cepas pero en los estadios 3 y 4 si, siendo la cepa
con mayor GCR la O25. En la prueba en invernadero, se observó que en las variables
evaluadas se destacaron diferentes aislamientos, en peso seco, número de brotes y en
longitud de raíz se destacó C4, en longitud de tallo C7 fue la mejor. Se esperaban encontrar
una correlación entre la producción de AIA y los GCR, pero esta no se observó. Igualmente
se esperaba que la producción de sideróforos y los porcentajes de inhibición estuvieran
relacionados, pero de igual forma tampoco se determinó una relación lineal entre los
resultados de las dos pruebas.

12
1. INTRODUCCIÓN
La uchuva (Physalis preuviana L.) es considerada como una fruta exótica, muy apetecida en
países europeos, ya que se le atribuyen diferentes propiedades medicinales y también por
sus características organolépticas. Por esta razón en esta última década ha pasado de ser
un cultivo silvestre, a producirse en cultivos más organizados y tecnificados aumentando su
área cultivada de 221 hectáreas en el año 1999 a 534 hectáreas en el año 2003 (Sanabria,
2005).
Este aumento en el cultivo es resultado de la demanda de la fruta en Europa, convirtiéndose
en uno de los productos de mayor exportación en nuestro país. Sin embargo las normas que
permiten la entrada y comercialización del producto en estos países son rigurosas y solo se
reciben y se comercializan frutas de una alta calidad con trazas mínimas de químicos, ya
que estos pueden afectar la salud humana a largo plazo.
Estas exigencias internacionales más la preocupación por implementar técnicas agrícolas
con un menor impacto en el ecosistema, constituyen la base de esta investigación, en donde
las rizobacterias promotoras del crecimiento vegetal que también puedan controlar
enfermedades tan limitantes como lo es la marchitez vascular, causada por Fusarium
oxysporum, se presentan como una excelente alternativa para que los productores
colombianos sean competitivos el mercado internacional, disminuyendo el uso de
agroquímicos nocivos y por tanto las trazas de los mismos en las frutas.
El presente trabajo planteó el aislamiento, preselección e identificación de rizobacterias de
cultivos de uchuva, pertenecientes al género Bacillus, Pseudomonas fluorescens y
actinomycetes, que promovieran la germinación de semillas de uchuva y además generaran
antagonismo frente a Fusarium oxysporum. Alguno de los aislamientos preseleccionados
fueron probados en invernadero para observar la promoción de crecimiento en plántulas de
uchuva a fin de determinar parámetros para evaluación posteriores a escala invernadero.
2. MARCO TEÓRICO
2.1 Generalidades de la Uchuva
La uchuva pertenece a la familia Solanaceae, la cual posee cerca de 90 géneros y más de
2.600 especies de distribución cosmopolita pero centrada en la zona tropical. (Estupiñan y
Ossa, 2007). Physalis peruviana L. es el nombre científico de la uchuva, cuyo fruto se

13
caracteriza por ser semiácido, redondo, amarillo, dulce y pequeño, midiendo entre 1,25 y 2
cm de diámetro. El fruto está envuelto en un capuchón protector, que lo protege del daño
mecánico por insectos, pájaros y diversos patógenos, además de las condiciones climáticas
externas (Anónimo, 2001). Esta es la especie más conocida de este género y su
comercialización ha aumentado actualmente en el exterior por las diferentes propiedades
nutricionales y medicinales que este fruto posee. Se caracteriza por ser un fruto azucarado y
con altos contenidos de vitaminas A y C, alcanzando una concentración de 26 mg por cada
100 g de pulpa, además contiene hierro y fósforo, en concentraciones de 1.2 y 38 mg/ 100 g
de pulpa respectivamente, lo cual es una concentración alta comparado con otras frutas
(Florez et al. 2000, citado por Anónimo, 2001).
Colombia, se ubica como el primer productor mundial de uchuva, seguido por Sudáfrica;
siendo la uchuva colombiana la de mayor aceptación por tener una mejor coloración y mayor
contenido de azúcares. En nuestro país la mayor área cultivada se encuentra en el
departamento de Cundinamarca, el cual cuenta con 267 hectáreas, que equivalen al 84.5%
del total de área cosechada en el país. Desde el año 2000 se cuenta con registros de
cultivos de uchuva en los departamentos de Antioquia y Boyacá, cada uno con áreas de 28 y
15 hectáreas respectivamente, que equivalen al 13.6% del área cosechada en el país
(Fischer et al., 2000). Dentro de los municipios productores es importante resaltar la
importancia en la explotación comercial de esta fruta en Granada, Silvania y Fusagasugá en
Cundinamarca, y Villa de Leyva en la región oriental del departamento de Boyacá. Una de
las razones que explican la concentración de los cultivos en estas zonas es su cercanía a
Bogotá, lugar desde donde se exporta al mercado europeo. En el departamento de Antioquia
los principales municipios productores son Rionegro y Sonsón (Anónimo, 2002).
La uchuva también se caracteriza por adaptarse fácilmente a una amplia gama de
condiciones agroecológicas. Fischer (2000) afirma que en Colombia esta fruta crece entre
los 1.500 y los 3.000 msnm, ubicándose los mejores cultivos a una altura entre los 1.800 y
los 2.800 msnm, la cual concuerda con una temperatura promedio entre los 13 y 18°C y
cuya pluviosidad puede ir de 1.000 a 2.000 mm anuales. El cultivo requiere de una humedad
relativa promedio de 70 a 80%. Es importante establecer el cultivo en suelos bien drenados
que mantengan un pH entre 5.5 y 7.0 y sean muy ricos en materia orgánica (Zapata, et al.
2002).
2.2 Enfermedades de la Uchuva
Durante el cultivo y la poscosecha de la uchuva se pueden presentar diversas
enfermedades, que afectan los rendimientos y la calidad del producto final de las frutas.

14
Dentro de las enfermedades más limitantes encontramos las generadas por hongos como lo
son el mal de semilleros causado por Pythium sp., la mancha gris causada por Cercospora
sp., muerte descendente o mal de tierra generado por Phoma sp., esclerotiniosis generado
por Sclerotinia sclerotiorum (Lib.) de Bary., mancha foliar generada por Alternaria sp., el
moho gris causado por Botrytis sp. y la marchitez vascular causada por Fusarium oxysporum
Schelecht; sin mencionar la gran variedad de enfermedades causadas por bacterias y por
nematodos (Zapata, et al. 2002; Casas, 2006).
Dentro de estas enfermedades causadas por hongos una de las más importantes por su
difícil control es la causada por Fusarium oxysporum. La marchitez vascular causada por
este agente, se evidencia por la clorosis, el marchitamiento y la posterior muerte de las
plantas, luego de tres meses de haber sido sembradas. Los vasos conductores presentan
grandes áreas necrosadas, generadas por la colonización del hongo a través de estos. Esta
necrosis se va extendiendo desde la base de la planta hasta la parte donde se ubican los
tejidos más jóvenes de la planta, esto produce la incapacidad para transportar nutrientes en
la misma y por esta razón se producen los síntomas mencionados anteriormente (Agrios,
2002). Estudios de De LaRotta y Quevedo (2005), citado por Congora y Rojas (2006)
encontraron incidencia de esta enfermedad de 10 al 50% y mediante análisis de laboratorio y
aplicando pruebas de patogenicidad se confirmó que el agente causal de la enfermedad es
F. oxysporum Schlecht.
Esta enfermedad es la causal de grandes pérdidas económicas a los productores de uchuva,
además de disminuir la calidad de la fruta. Para su control se ha implementado el uso de
fungicidas con principios activos recalcitrantes para el ambiente, que dejan trazas en las
frutas, las cuales pueden llegar a ser tóxicos para el humano. Esto genera rechazo por parte
de los diferentes mercados internacionales, que son muy rigurosos con las medidas y los
requisitos de salubridad al recibir y liberar el producto.
2.3 Reguladores de crecimiento vegetal
Los reguladores de crecimiento vegetal son sustancias, que en pequeñas concentraciones,
son capaces de regular procesos fisiológicos de la planta (Arteca, 1996). Gracias a su
capacidad de regular el desarrollo vegetal, los inductores de crecimiento vegetal, en especial
los del grupo de las auxinas, son utilizadas en agricultura en frecuentemente. En general las
auxinas son obtenidas por síntesis química, por lo cual la síntesis microbiológica de estas
sustancias resulta de gran importancia pudiendo constituir una alternativa viable en el
contexto de una agricultura ecológica (Castillo et al. 2005). Los reguladores de crecimiento
vegetal o fitohormonas se conocen desde 1937, cuando el término era sinónimo de auxina.

15
Tiempo después, las auxinas en compañía de las giberelinas, las citoquininas, el etileno y el
ácido abscísico fueron atribuidas como ―las cinco clásicas‖. Las fitohormonas son
sintetizadas no solo por plantas, sino por microorganismos, incluidos bacterias, hongos y
actinomycetes (Tudzinzki y Sharon, 2002).
2.3.1 Auxinas
El termino auxina califica una clase de compuestos caracterizados por su capacidad para
inducir elongación en células de la región subapical de los brotes (Arteca, 1996). Estas son
ácidos débiles derivados del indol y son los mayores reguladores de crecimiento vegetal. Las
auxinas se encuentran involucradas en diferentes procesos metabólicos de las plantas como
la dominancia del brote principal e inhibición de la ramificación lateral, la diferenciación de
los vasos conductores (xilema y floema), el fototropismo y geotropismo, la estimulación de
formación de raíces adventicias y el desarrollo de frutos.
Dentro del grupo se puede destacar el ácido indol-acético (AIA), como una de las auxinas
naturales más importantes y se ha demostrado que pueden ser sintetizadas por diferentes
especies de bacterias, hongos y algas. Entre los más destacados encontramos bacterias de
los géneros Azospirillum, Azotobacter, Pseudomonas, Rhyzobium y Bacillus, entre otros
(Patten y Glick, 1996). Aunque la producción de estas fitohormonas por parte de los
diferentes microorganismos está debidamente documentada, aún no se encuentran muchos
estudios acerca de los efectos fisiológicos que estas producen en ellos (Tudzinzki y Sharon,
2002). La ruta de síntesis del ácido indo-acético ha sido tema de controversia. El aminoácido
triptófano se considera el mayor precursor de la síntesis de las auxinas. Sin embargo,
mutantes auxotrófos de tiptófano, mostraron evidencia de una ruta de síntesis independiente
de triptófano en la que se utiliza el indol, o indol-glicerol como precursor. (Tudzinzki y
Sharon, 2002).
2.4. Rizobacterias promotoras del crecimiento vegetal (PGPR)
En respuesta a la necesidad de generar cultivos limpios, con trazas mínimas o nulas de
agroquímicos que afecten la salud humana a largo plazo, se ha venido implementado el uso
de los microorganismos benéficos del suelo, que pueden promover el crecimiento de las
plantas y también evitar la infección del tejido vegetal por patógenos, estos son
denominados PGPR (plant growth promoting rhizobacteria; rizobacterias promotoras del
crecimiento vegetal). Estos microorganismos pueden encontrarse en asociaciones
simbióticas o de vida libre. Estos últimos están asociados a las partículas del suelo
generando interacciones con las raíces de las plantas, en la zona de la rizósfera (Peña y
Reyes, 2007).

16
Los primeros trabajos para la inoculación de semillas fueron realizados en Rusia en 1930. A
finales de la década de las 70, Kloepper y colaboradores fueron los pioneros en introducir el
término PGPR para referirse a las rizobacterias capaces de provocar un efecto benéfico en
las plantas. Recientemente, la denominación se ha extendido a microorganismos PGP para
incluir hongos y cualquier organismo afín (Vessey, 2003).
Dentro de los mecanismos para la promoción del crecimiento vegetal encontramos dos tipos,
mecanismos directos e indirectos.
Los mecanismos directos son aquellos en donde los microorganismos actúan sobre la
planta. Dentro de estos encontramos la producción de promotores de crecimiento vegetal
(también llamados fitohormonas), como auxina, giberelinas y citoquininas. La producción de
estos metabolitos, también generan una mejor regulación de estomas, lo que evita su
deterioro el cual está asociado a depresión del crecimiento y síntomas de marchitamiento en
la planta. Ocasionan también un desarrollo radical más ramificado jugando un papel
fundamental en la absorción del agua y en el mejoramiento de la nutrición al aumentar su
acceso a los nutrientes en el suelo (Rivieros, 2008). Adicionalmente, las PGPR pueden
promover el crecimiento vegetal mejorando la disponibilidad de nutrientes en el suelo
mediante solubilización de fosforo, fijación de nitrógenos y producción de sideróforos
(Bobadilla y Rincón, 2008). Igualmente se ha demostrado que las rizobacterias ayudan a
disminuir la resistencia a la conductividad hidráulica, lo cual le da a las plantas una mayor
tolerancia a periodos de sequía (Rivieros, 2008).
Los mecanismos indirectos son aquellos en donde el microorganismo es capaz de inhibir
diferentes patógenos que interfieren con el desarrollo de la planta. Fitopatógenos como
hongos y bacterias son neutralizados por diversos mecanismos, como competencia por
espacio o nutrientes, o también producción de metabolitos antibióticos, secreción de
diversas enzimas hidrolíticas que degradan la pared celular de los patógenos (Reyes et al.
2008). También se ha evidenciado la producción de sideróforos los cuales se definen como
moléculas pequeñas con cadenas laterales y grupos funcionales que les proporcionan alta
afinidad para captar y concentrar iones férricos. Estas moléculas son producidas típicamente
por bacterias, hongos y plantas monocotiledóneas en respuesta al estrés por
concentraciones limitantes de hierro en el ambiente (Perez et al., 2007). Debido a que la
biodisponibilidad de hierro es limitada en el suelo, la producción de sideróforos juega un
papel importante el la competencia por este recurso, ya que, en general, los sideróforos
producidos por bacterias tienen una mayor afinidad a este elemento que los producidos por
hongos, limitando así a posibles patógenos (Compant et al., 2005). Otro mecanismo

17
indirecto es el incremento de la capacidad de respuesta sistémica de la planta frente a los
patógenos (Vessey, 2003).
Aunque los estudios con microorganismos como Pseudomonas fluorescens se habían
basado únicamente en la utilización de su potencial como biocontrolador de fitopatógenos
habitantes del duelo, los esfuerzos ahora comienzan a concentrarse en la capacidad de los
PGPR de promover el desarrollo vegetal, utilizándolos en la obtención de plantas más
saludables y con mejores rendimientos de cultivo. También se incluyen en estos estudios
microorganismos como Azotobacter, Azospirillum, Acetobacter, Burkholderia, Bacillus (Dey
et al. 2004) y actinomycetes (El-Tarabily y Sivasithamparam, 2006).
2.4.1. Pseudomonas fluorescens
Esta bacteria es un cocobacilo Gram negativo, que se encuentra como saprófito en el suelo
y es ampliamente conocido por ser una PGPR. Abundan en la superficie de las raíces, ya
que son versátiles en su metabolismo y pueden utilizar varios sustratos producidos por las
mismas, pero no establecen una relación simbiótica con la planta. Entre sus mecanismos de
acción se encuentran el aumento de la toma de agua y nutrientes por la planta, la solubilización de
fosfatos, la producción de reguladores del crecimiento vegetal y el control biológico de
patógenos, dado fundamentalmente por la producción de sideróforos, la antibiosis y la
inducción de resistencia a la planta, mediante la producción de ácido salicílico, el cual actúa
como una molécula de señalización que activa la ―resistencia sistémica inducida‖ (RSI) que
es muy similar a la ―resistencia sistémica adquirida‖ (RSA) (Zhang et al., 2002).
2.4.2. Bacillus sp.
Estos microorganismos se caracterizan por ser bacterias Gram positivas con forma bacilar,
aerobias estrictos o anaerobias facultativas que en condiciones estresantes forman una
endoespora central, que deforma la estructura de la célula. Esta forma esporulada es
resistente a las altas temperaturas y a los desinfectantes químicos corrientes. Esta bacteria
es capaz de generar un efecto benéfico en el crecimiento de las plantas por diversos
mecanismos, en donde se encuentran la producción de sustancias antibióticas, producción
de lipopéptidos que actúan como biosurfactantes, solubilización de fosfatos y reducción de
enfermedades en las plantas (Kokalis-Burelle et al., 2006).
2.4.3. Actinomycetes
Los actinomycetes son microorganismos ubicuos y se encuentran sobre la gran mayoría de
sustratos naturales corrientes. Entre las propiedades más comunes de estos bacilos Gram-
positivos, se encuentra la tendencia a formar ramificaciones cortas o más desarrolladas; son
quimio-organotrofos, aerobios, mesofílicos y crecen óptimamente en pH cercano a la

18
neutralidad (El-Tarabil y Sivasithampara, 2006). Numerosos estudios demuestran la
importancia de los actinomicetos sobre todo de Streptomyces sp., como controlador de
hongos fitopatógenos del suelo y promoción de crecimiento en plantas; por este motivo, los
métodos comúnmente utilizados para el aislamiento y recuento de cepas de actinomycetes
provenientes de suelo o rizosfera utilizados para biocontrol o promoción de crecimiento en
plantas, tratan exclusivamente con aquellos adecuados para las especies de Streptomyces
(Goodfellow y Williams, 1983 citado por El-Tarabil y Sivasithampara 2006).
3. FORMULACIÓN DEL PROBLEMA Y JUSTIFICACIÓN
3.1. Formulación del Problema
Las grandes pérdidas económicas producidas por enfermedades en la planta de uchuva,
como la marchitez vascular causada por Fusarium oxysporum, no solo por el rechazo por las
instituciones internacionales al disminuir la calidad de este producto, sino también por la
pérdida de hectáreas cultivadas, y por el gran impacto ecológico al tratar de controlar la
enfermedad mediante fungicidas recalcitrantes en el ecosistema, constituyen en la
actualidad una de los principales limitantes del cultivo uchuva en el país.
3.2. Justificación de la investigación
El poder generar una alternativa de control eficiente frente a enfermedades tan agresivas
como la marchitez vascular, que disminuya las pérdidas económicas de los productores de
uchuva, ayudaría a aumentar la producción de los cultivos y además a abrir cada vez más
mercados internacionales, que son mucho más exigentes en cuanto a la calidad fitosanitaria
del producto.
El aislamiento de las PGPR es la primera fase para el desarrollo de un producto efectivo
para la aplicación a los cultivos de la uchuva tendiente a generar un control frente a F.
oxysporum, que sea efectivo y amigable con el ambiente y que promueva el crecimiento de
las plantas produciendo así frutas de una mayor calidad que generen un valor agregado y
una mayor ganancia al productor y al exportador.

19
4. OBJETIVOS
4.1. Objetivo General
Aislar rizobacterias con potencial de control de Fusarium sp., que generen promoción del
proceso de germinación de semillas y promoción de crecimiento de plántulas uchuva en
invernadero.
4.2 Objetivos específicos
Realizar el aislamiento de bacterias del suelo rizosférico en cultivos que presenten
síntomas de marchitamiento vascular.
Seleccionar los aislamientos de rizobacterias de los géneros Bacillus sp. y
Pseudomonas fluorescens.
Determinar la capacidad promotora de crecimiento de los géneros Bacillus y
Pseudomonas con base en el porcentaje de germinación de semillas.
Seleccionar las rizobacterias aisladas por su actividad antagónica in vitro frente a
Fusarium sp. Realizar la selección de las rizobacterias que presenten una mayor
promoción del crecimiento en las plántulas de uchuva en invernadero.
Elaborar una colección de aislamientos de rizobacterias con capacidad promotora de
crecimiento vegetal por crioconservación.
5. MATERIALES Y MÉTODOS
5.1. Diseño de Investigación
El tipo de investigación es experimental aplicada, porque se determinó la acción promotora
del crecimiento vegetal de los aislamientos de las rizobacterias realizando pruebas de
germinación de semillas de uchuva y además evaluando el efecto de algunos de los
aislamientos en plántulas, bajo condiciones de invernadero. También se midió el efecto
antagónico frente al hongo Fusarium sp. de los mismos aislamientos, mediante la
determinación del porcentaje de inhibición de crecimiento radial. Fue una investigación de
tipo observacional, porque se describió si hay o no una acción promotora de crecimiento por
parte de los aislamientos, además de su capacidad antagónica frente a Fusarium sp.

20
5.1.1. Población de estudio y muestra de población de estudio
La población de estudio fueron las rizobacterias aisladas de muestras de raíces y suelo
rizosférico de cultivos localizados en el municipio de Silvania, localizado a los 04° 24´ 19¨ de
latitud norte y 74° 23´26¨ de longitud oeste, a una altura de 1955 m.s.n.m., tiene una
temperatura media de 20°C y la precipitación media anual es de 1955 mm.
5.1.2. Variables de estudio
Para la determinación de la acción promotora de crecimiento de los aislamientos en la
germinación de semillas se midieron las siguientes variables: porcentaje de germinación que
es una variable cuantitativa, continua de razón. Para la medición de la acción promotora de
crecimiento de los aislamientos en el crecimiento de plántulas se midieron las siguientes
variables: altura de la planta, longitud de raíces, peso seco y número de brotes, las cuales
son variables cuantitativas, continuas, de razón. Para la medición de la acción antagónica de
los aislamientos se tuvo en cuenta el porcentaje de inhibición que también es una variable
cuantitativa, continua, de razón.
5.2. Metodología
5.2.1. Salida de campo
Se recolectaron las muestras en diferentes fincas ubicadas en el municipio de Silvania,
teniendo en cuenta que en estas hubiera cultivos afectados con marchitez vascular causada
por F. oxysporum se escogieron plantas sanas en los focos de la enfermedad para tomar
una muestra de raíces secundarias y suelo rizosférico. En cada finca se eligieron tres plantas
separadas entre sí por 50m aproximadamente. Las muestras fueron refrigeradas hasta el
momento de procesamiento en el laboratorio.
5.2.2. Procesamiento de muestras
Luego del traslado de las muestras al laboratorio se procesaron siguiendo el POE de
Corpoica para el aislamiento de Pseudomonas fluorecens y para el aislamiento de bacterias
mesófilas esporuladas.
Se realizó un lavado superficial de las raíces, luego de este lavado se pesaron las raíces y
se llevaron a una dilución 1/10 en una solución de tween 80 al 0.1%. A esta dilución se le
agregaron perlas de vidrio y se llevaron a tres ciclos de agitación en vortex cada uno con
una duración de 10 segundos. Luego se llevaron a agitación durante una hora a 150 rpm,
posteriormente se realizaron nuevamente tres ciclos de agitación en vortex. Seguido a esto,
se realizaron diluciones hasta 10-7
y sembró 0.1 ml de las diluciones 10-4
, 10-5
, 10-6
y 10-7
en
agar King B y en Agar avena suplementado con Nistatina 0,1%; el primero para realizar el
aislamiento selectivo de Pseudomonas fluorescenetes y el segundo para el aislamiento de

21
actinomycetes. Las siembras en agar King B se llevaron a incubar a temperatura ambiente
por 48 horas y se realizó la lectura de las colonias fluorescentes mediante la exposición de
luz UV. Las siembras en agar avena se llevaron a incubar a temperatura ambiente, hasta
que aparezcan colonias características de estos microorganismos. El remanente de la
primera dilución fué llevado a choque térmico, por 10 minutos a una temperatura de 80°C.
Luego se realizaron diluciones hasta 10-7
sembrando 0.1 ml de las diluciones 10-4
, 10-5
, 10-6
y
10-7
en agar para microorganismos esporulados (Corpoica). Estas siembras se llevaron a
incubar de dos a tres días a temperatura ambiente.
5.2.3. Aislamiento e identificación de Pseudomonas fluorescens
A partir de las colonias que presentaron fluorescencia se realizó un aislamiento en agar
nutritivo para purificar las colonias. Posteriormente se identificaron los aislamientos por
pruebas bioquímicas basadas en la matriz de Schaad (1994), en el POE para el aislamiento
de Pseudomonas fluorescens de Corpoica y en Bergey (2000).
Inicialmente se hizo una coloración de Gram. Los cocobacilos Gram-negativos fueron los
seleccionados para las siguientes bioquímicas: producción pioverdina, oxidasa, catalasa,
urea, nitrato, citrato, arginina descarboxilasa y glucosa OF, escogiéndose los aislamientos
que dieran resultados positivos en las anteriores bioquímicas y en esta última, los que dieran
como resultado oxidadores. Estos se sembraron en agar Cetrimide y se escogieron las que
tuvieran crecimiento moderado o crecimiento normal, sin producción de pigmentos verdes y
cafés. Las colonias que coincidieron con los resultados mencionados anteriormente, se
consideraron como Pseudomonas fluorescens. Adicionalmente como pruebas confirmatorias
de género se evalúo la prueba de levano y crecimiento en YDC.
5.2.4. Aislamiento y selección de Bacillus sp.
Las colonias que crecieron en el medio para esporulados, luego de realizar el choque
térmico, se les realizó el siguiente procedimiento basados en la matriz de Schaad (1994) y
en Bergey (2000).
Se realizó una coloración de Gram, para escoger a los bacilos Gram positivos, luego la
coloración de verde malaquita, en donde se observó la presencia de endoesporas. Estos
aislamientos fueron sometidos a una prueba de crecimiento anaeróbico sembrando por
cuadruplicado, en 7mL de medio anaeróbico, incubando dos en anaerobiosis, mediante el
uso de aceite mineral y dos en aerobiosis. El resultado positivo será para aquellos
microrganismos capaces de crecer en anaerobiosis y aerobiosis, o solo en aerobiosis,
evidenciándose en el viraje del medio de azul a amarillo. También se realizaron las
siguientes bioquímicas: catalasa, nitrato y glucosa OF. Se escogieron los aislamientos

22
positivos para las pruebas mencionadas y para esta última los oxidadores y fermentadores.
Se realizó también la prueba lecitinasas, mediante la siembra en agar Mossel, aquí se
descartaron las colonias positivas para esta prueba, ya que este resultado corresponde a B.
cereus, un patógeno humano. Las cepas que coincidieron con los resultados mencionados
anteriormente se consideraron como Bacillus.
5.2.5. Aislamiento y selección de Actinomycetes
Las cepas que crecieron con las características macroscópicas de estos microorganismos,
es decir colonias secas, pulverulentas y con olor característico (olor a suelo por producción
de geosminas), fueron purificadas y se observaron sus características microscópicas, hifas
en forma de espiral y esporas.
5.2.6. Pruebas de germinación para determinar promoción de crecimiento vegetal de las
cepas aisladas
5.2.6.1. Cámara Húmeda
La cámara húmeda fue preparada usando cajas sello pack de tapa alta transparentes, de 16
onzas, en las cuales se ubica un acordeón de papel absorbente (papel para secar manos,
marca Familia). Éste tenía cuatro carriles, en donde se ubicaron 5 semillas por cada uno de
ellos, es decir un total de 20 semillas por caja. El papel se humedeció con 2ml de agua
destilada estéril, por cada carril (Celis y Gallardo, 2008). El montaje se llevó a cabo con
cuatro cajas por microorganismo seleccionado (ISTA, 2006).
5.2.6.2. Imbibición de las semillas
Antes de ubicar las semillas en los carriles de la cámara, fue necesario realizar una
imbibición de 30 minutos, que según Ramos y Valero (2009) es el tiempo suficiente para
estimular la germinación de las semillas de uchuva. Luego de realizar el conteo de las
semillas, estas se ubicaron en bolsitas de velo suizo estériles, se realizó un lavado con agua,
para eliminar químicos y contaminantes para después ser sumergidas totalmente en agua
destilada estéril por el tiempo recomendado. Luego con pinzas estériles se ubicaron en los
carriles de la cámara húmeda para ser inoculadas con los microorganismos aislados.
5.2.6.3. Preparación de inóculos.
A partir de cepas reconstituidas en TSA, e incubadas por 48 horas a temperatura ambiente,
se realizaron suspensiones en solución salina a una concentración de 0.85% por cada cepa
aislada; esta suspensión se llevó a una absorbancia de 0.2 a 540nm, equivalente a
108células/mL. A partir de estas suspensiones se inoculó cada semilla aplicando 100μL de la
suspensión bacteriana.

23
Se tuvieron dos controles, inoculando la misma cantidad mencionada anteriormente con
agua destilada estéril y solución salina estéril.
Se sellaron las cajas y se ubicaron en el fotoperiodo 12h luz/12h oscuridad, realizando
lecturas cada 48 horas, durante 25 días, teniendo en cuenta los estadios de desarrollo de
germinación de la uchuva, denominados de la siguiente forma:
Estadio 1 (E1): Rompimiento de la testa y emergencia de la radícula.
Estadio 2 (E2): Desarrollo del hipocótilo.
Estadio 3 (E3): Desplegamiento de hojas cotiledonares.
Estadio 4 (E4): Emergencia de la primera hoja verdadera.
5.2.7 Pruebas de antagonismo in vitro frente a Fusarium sp. para determinar la actividad
biocontroladora de los aislamientos.
5.2.7.1. Preparación de inóculo
La preparación de los inóculos bacterianos para realizar la prueba de antagonismo se hizo
de igual forma como se describió en el numeral 5.2.6.3. La cepa de Fusarium sp. que se usó
para este ensayo, fue la cepa G1 aislada la Universidad Militar Nueva Granada; esta cepa se
reconstituyó en medio PDA y se dejó crecer por 10 días.
5.2.7.2. Montaje de la prueba de antagonismo
Se realizó la técnica de enfrentamiento dual en medio PDA sembrando el inóculo de las
bacterias antagonistas, preparado como se mencionó en el numeral 5.2.7.1. Se realizó una
siembra masiva de 50μl de la suspensión celular, la cual ocupó 2cm del borde de la caja al
centro. Esta siembra se dejó incubando por 48 horas a temperatura ambiente, para permitir
que el microorganismo liberara metabolitos al medio que inhibieran el crecimiento del
patógeno. Pasadas las 48 horas, se sembró un disco de agar con la cepa G1 a 3cm del
borde de la caja al centro, frente a la cepa antagonista. Se tuvo como testigo, el crecimiento
de la cepa G1 sola, para así poder establecer el porcentaje de inhibición correspondiente a
cada aislamiento. Para ello se usó la siguiente fórmula:
Ecuación 1. Porcentaje de inhibición de Fusarium sp.

24
Se realizó la lectura 10 días después de la siembra del hongo y se sembraron cuatro réplicas
por aislamiento seleccionado.
5.2.8. Cuantificación de la producción de AIA
5.2.8.1. Curva de calibración del reactivo de Salkowski.
El montaje de la reacción y de la curva de calibración, se realizó siguiendo el protocolo de
Celis y Gallardo (2008), y Califa y González (2009).
5.2.8.2. Cultivo discontinuo en caldo BT de las cepas aisladas y cuantificación de AIA a partir
de los mismos
Para inducir la producción de AIA durante el cultivo discontinuo, se usó el medio líquido BT,
con una concentración de triptona de 20g/L (Celis y Gallardo, 2008). Los cultivos se
mantuvieron en agitación constante 120 rpm a 26°C durante 96 horas, protegidos de la luz y
se realizó un muestreo a la hora 96, que según Califa y González (2009) es el pico de
producción de este metabolito; se manejaron tres réplicas por cepa aislada, de igual forma
cada reacción por cepa se hizo por triplicado (Ramos y Valero, 2009). La reacción a partir de
cada cultivo se realizó como lo describen Celis y Gallardo (2008).Como microorganismo de
referencia se usó a una cepa Azotobacter vinelandii ATCC 12518.
5.2.9. Cuantificación de la producción de sideróforos
5.2.9.1. Curva de calibración Cromo Azurol S (CAS)
El montaje de la reacción, asi como el de la curva de calibración se realizó como lo
describen Ramos y Valero (2009), Diaz (2009).
5.2.9.2. Cultivo discontinuo en medio mínimo (Simmon y Tessman) y cuantificación de
sideróforos a partir de los mismos
La inducción para la producción de sideróforos se usó el medio de cultivo líquido Simmon &
Tessman (1963) probando inicialmente la glucosa y el acido succínico como fuentes de
carbono, y nitrato de amonio junto con caseína hidrolizada ácida como fuentes de nitrógeno.
El tiempo de cultivo fue de 96 horas y se realizó la reacción de cada cultivo, como lo
describió Diaz (2009). Como microorganismo control para la producción de sideróforos se
usó una cepa de Pseudomonas fluorescens ATCC BAA 477.
5.2.10. Pruebas de promoción de crecimiento en invernadero
Para el establecimiento de las plántulas inicialmente se debió hacer una germinación en
cámara húmeda como se describió en el numeral 5.2.6., inoculando de igual forma el
microorganismo seleccionado. Para este ensayo se utilizaron únicamente los
microorganismos aislados e identificados en el primer muestreo, a fin de establecer

25
condiciones apropiadas para el ensayo. Cuando las plántulas tuvieron un mes y un par de
hojas verdaderas, se trasplantaron a semilleros de 72 alveolos (Riveros, 2008) en suelo con
cascarilla, luego las plántulas se inocularon con 500μl de la suspensión de las cepa
seleccionadas, preparada como se describió en el numeral 5.2.6.3. Se realizó un
seguimiento durante un mes, evaluando al final: altura, longitud de raíz, número de brotes y
peso seco (Riveros, 2008).
5.2.11. Desarrollo del banco de germoplasma
A partir de las cepas aisladas y puras sembradas en un medio nutritivo por 48 horas, se
realizó el banco. El medio usado para la criopreservación tiene glicerol al 30%, peptona 5g/l
y extracto de carne 3g/l. Se depositó 1.2mL de este medio en tubos eppendorf estériles y
luego se suspendió una colonia de la cepa a conservar, posteriormente se conservó en
temperatura de -20°C.
5.3. Análisis de información
Para las variables de tipo continuo se realizó un análisis estadístico de varianza ANOVA,
para así identificar si hay o no diferencias significativas entre los resultados obtenidos con
cada cepa evaluada. Todas las herramientas estadísticas de análisis se aplicaron con una
confianza mínima del 95% y se utilizó el software SPSS.
6. RESULTADOS Y DISCUSION
6.1. Aislamiento e identificación bioquímica de cepas aisladas
En el primer y segundo muestreos realizados se hizo la selección de las rizobacterias de
interés, mediante un aislamiento selectivo, por medio de protocolos y medios de cultivo que
permitían la diferenciación fenotípica de los géneros Bacillus y Pseudomonas, así como de
los actinomycetes. Sin embargo hubo diferencias en cuanto al seguimiento de las pruebas
de identificación, ya que en el primer muestreo, se optó por identificar bioquímicamente las
cepas aisladas de forma selectiva, para así trabajar con los géneros de interés y descartar lo
que no perteneciera a estos grupos; mientras que en el segundo muestreo a partir del
aislamiento selectivo se trabajaró con pruebas screening que permitieran identificar cuales
cepas eran presentaban características PGPR en una población mucho mayor, y luego
identificarlas. Los resultados se muestran en la tabla 1, en donde se observan las cepas
identificadas dentro de los grupos de interés; en el primer muestreo a partir de 35 cepas de
cocobacilos Gram negativos y de 15 cepas de Bacilos Gram positivos esporulados ambos
aislados de forma selectiva, se identificaron dentro de la especie Pseudomonas fluorescens
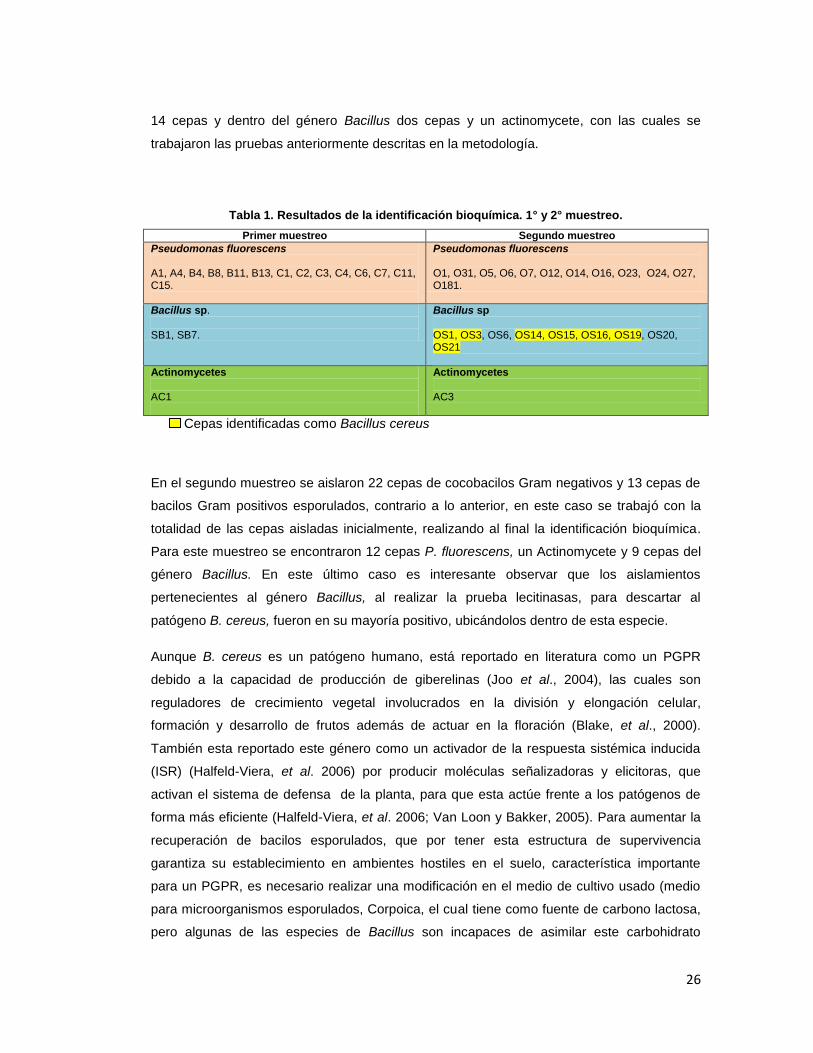
26
14 cepas y dentro del género Bacillus dos cepas y un actinomycete, con las cuales se
trabajaron las pruebas anteriormente descritas en la metodología.
Tabla 1. Resultados de la identificación bioquímica. 1° y 2° muestreo.
Primer muestreo Segundo muestreo
Pseudomonas fluorescens A1, A4, B4, B8, B11, B13, C1, C2, C3, C4, C6, C7, C11, C15.
Pseudomonas fluorescens O1, O31, O5, O6, O7, O12, O14, O16, O23, O24, O27, O181.
Bacillus sp. SB1, SB7.
Bacillus sp OS1, OS3, OS6, OS14, OS15, OS16, OS19, OS20, OS21
Actinomycetes AC1
Actinomycetes AC3
Cepas identificadas como Bacillus cereus
En el segundo muestreo se aislaron 22 cepas de cocobacilos Gram negativos y 13 cepas de
bacilos Gram positivos esporulados, contrario a lo anterior, en este caso se trabajó con la
totalidad de las cepas aisladas inicialmente, realizando al final la identificación bioquímica.
Para este muestreo se encontraron 12 cepas P. fluorescens, un Actinomycete y 9 cepas del
género Bacillus. En este último caso es interesante observar que los aislamientos
pertenecientes al género Bacillus, al realizar la prueba lecitinasas, para descartar al
patógeno B. cereus, fueron en su mayoría positivo, ubicándolos dentro de esta especie.
Aunque B. cereus es un patógeno humano, está reportado en literatura como un PGPR
debido a la capacidad de producción de giberelinas (Joo et al., 2004), las cuales son
reguladores de crecimiento vegetal involucrados en la división y elongación celular,
formación y desarrollo de frutos además de actuar en la floración (Blake, et al., 2000).
También esta reportado este género como un activador de la respuesta sistémica inducida
(ISR) (Halfeld-Viera, et al. 2006) por producir moléculas señalizadoras y elicitoras, que
activan el sistema de defensa de la planta, para que esta actúe frente a los patógenos de
forma más eficiente (Halfeld-Viera, et al. 2006; Van Loon y Bakker, 2005). Para aumentar la
recuperación de bacilos esporulados, que por tener esta estructura de supervivencia
garantiza su establecimiento en ambientes hostiles en el suelo, característica importante
para un PGPR, es necesario realizar una modificación en el medio de cultivo usado (medio
para microorganismos esporulados, Corpoica, el cual tiene como fuente de carbono lactosa,
pero algunas de las especies de Bacillus son incapaces de asimilar este carbohidrato
27
(Bergey, 2000), por lo cual se propone el uso de glucosa en el medio, para que luego de
realizar el choque térmico y eliminar durante este los microorganismos no esporulados, se
recuperen con mayor facilidad las bacterias de interés.
6.2. Producción de Acido indol-acético
Los reguladores de crecimiento vegetal (PGRs) son sustancias orgánicas que influencian la
fisiología y desarrollo de la planta a concentraciones muy bajas (actúan a concentraciones
internas menores a 1μM) (García et al., 2005). Las bacterias habitantes de la rizósfera
pueden influenciar el crecimiento de las plantas contribuyendo con el pool endógeno de
PGRs en estas, como las auxinas, entre las que se encuentra el Acido Indol-Acético (AIA)
(Patten y Glick, 2002). La producción de AIA por las cepas aisladas fue evaluada en medio
mínimo con triptona, inductor de la producción de AIA debido a su alto contenido de
triptófano, precursor del este metabolito, cuya concentración fue cuantificada por medio de
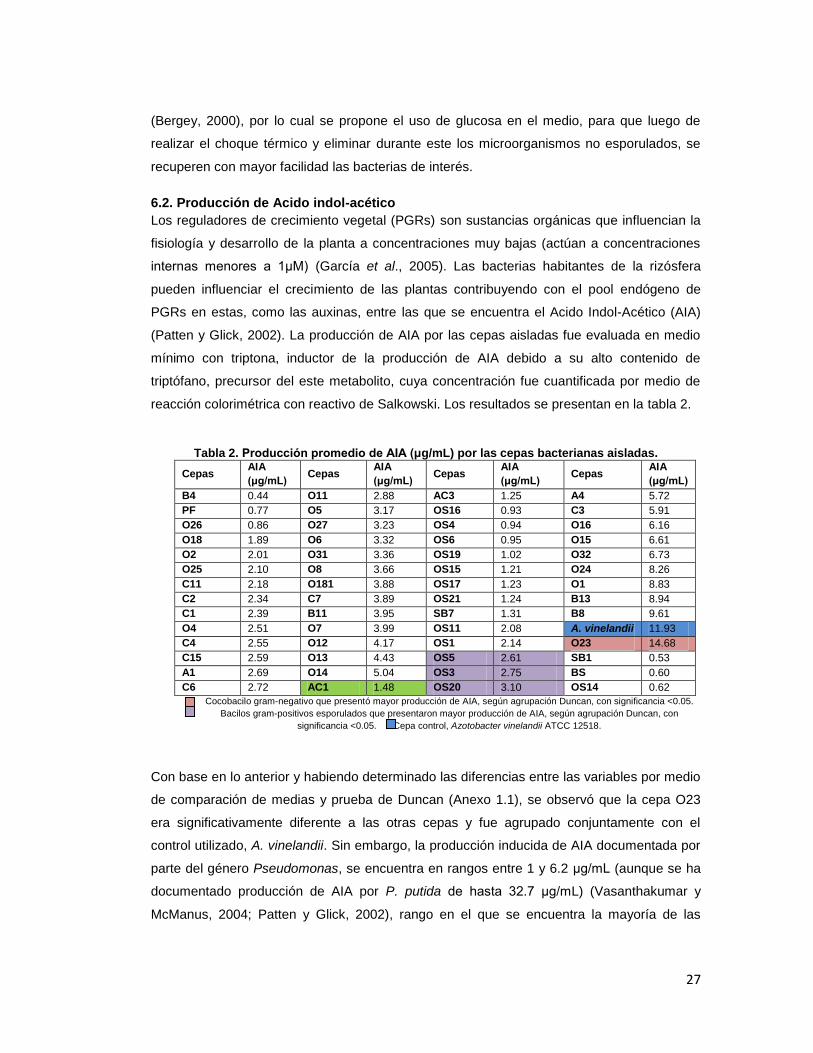
reacción colorimétrica con reactivo de Salkowski. Los resultados se presentan en la tabla 2.
Tabla 2. Producción promedio de AIA (μg/mL) por las cepas bacterianas aisladas.
Cepas AIA
(μg/mL) Cepas
AIA
(μg/mL) Cepas
AIA
(μg/mL) Cepas
AIA
(μg/mL)
B4 0.44 O11 2.88 AC3 1.25 A4 5.72
PF 0.77 O5 3.17 OS16 0.93 C3 5.91
O26 0.86 O27 3.23 OS4 0.94 O16 6.16
O18 1.89 O6 3.32 OS6 0.95 O15 6.61
O2 2.01 O31 3.36 OS19 1.02 O32 6.73
O25 2.10 O8 3.66 OS15 1.21 O24 8.26
C11 2.18 O181 3.88 OS17 1.23 O1 8.83
C2 2.34 C7 3.89 OS21 1.24 B13 8.94
C1 2.39 B11 3.95 SB7 1.31 B8 9.61
O4 2.51 O7 3.99 OS11 2.08 A. vinelandii 11.93
C4 2.55 O12 4.17 OS1 2.14 O23 14.68
C15 2.59 O13 4.43 OS5 2.61 SB1 0.53
A1 2.69 O14 5.04 OS3 2.75 BS 0.60
C6 2.72 AC1 1.48 OS20 3.10 OS14 0.62
Cocobacilo gram-negativo que presentó mayor producción de AIA, según agrupación Duncan, con significancia <0.05.
Bacilos gram-positivos esporulados que presentaron mayor producción de AIA, según agrupación Duncan, con
significancia <0.05. Cepa control, Azotobacter vinelandii ATCC 12518.
Con base en lo anterior y habiendo determinado las diferencias entre las variables por medio
de comparación de medias y prueba de Duncan (Anexo 1.1), se observó que la cepa O23
era significativamente diferente a las otras cepas y fue agrupado conjuntamente con el
control utilizado, A. vinelandii. Sin embargo, la producción inducida de AIA documentada por
parte del género Pseudomonas, se encuentra en rangos entre 1 y 6.2 μg/mL (aunque se ha
documentado producción de AIA por P. putida de hasta 32.7 μg/mL) (Vasanthakumar y
McManus, 2004; Patten y Glick, 2002), rango en el que se encuentra la mayoría de las
28
cepas evaluadas en su mayoría pertenecientes al género. Por otro lado, los bacilos gram-
positivos y actinomicetos evaluados presentaron resultados mucho menores a comparación
de los cocobacilos gram-negativos, siendo significativamente mejores según la comparación
de medias y prueba de Duncan las cepas OS5, OS3 y OS20 (Anexo 1.2.). Se ha encontrado
que la producción de AIA en el género Bacillus es generalmente baja aunque se ha
documentado producciones de hasta 55μg/mL (Felici et al., 2008; Tsavkelova, 2006). Es de
mencionar que la literatura en cuanto a la producción de AIA por actinomicetostiene pocos
reportes, pero se encuentra documentado actinomicetos del género Streptomyces como
productores de AIA, cuya capacidad de producción alcanza los 5μg/mL (Dimkpa et al.,
2008), valores más altos comparados con los obtenidos por las cepas evaluadas.
6.3. Promoción de germinación
La capacidad de promoción de germinación por parte de las cepas aisladas fue evaluada
mediante montaje de semillas inoculadas en cámara húmeda, registrando los estadios de
germinación de las semillas durante 25 días. Los resultados obtenidos se analizaron para
obtener los índices de germinación relativos (GCR, cociente de capacidad de germinación
máxima del tratamiento y el control (agua), que fueron analizados por comparación de
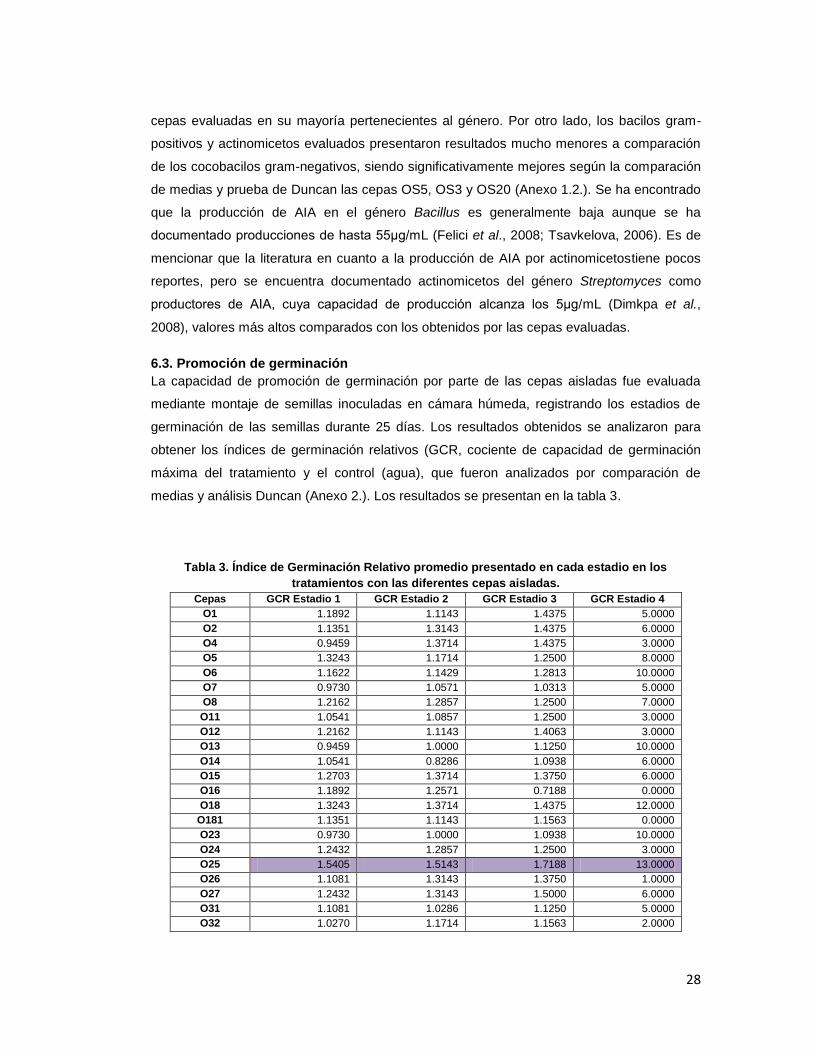
medias y análisis Duncan (Anexo 2.). Los resultados se presentan en la tabla 3.
Tabla 3. Índice de Germinación Relativo promedio presentado en cada estadio en los
tratamientos con las diferentes cepas aisladas.
Cepas GCR Estadio 1 GCR Estadio 2 GCR Estadio 3 GCR Estadio 4
O1 1.1892 1.1143 1.4375 5.0000
O2 1.1351 1.3143 1.4375 6.0000
O4 0.9459 1.3714 1.4375 3.0000
O5 1.3243 1.1714 1.2500 8.0000
O6 1.1622 1.1429 1.2813 10.0000
O7 0.9730 1.0571 1.0313 5.0000
O8 1.2162 1.2857 1.2500 7.0000
O11 1.0541 1.0857 1.2500 3.0000
O12 1.2162 1.1143 1.4063 3.0000
O13 0.9459 1.0000 1.1250 10.0000
O14 1.0541 0.8286 1.0938 6.0000
O15 1.2703 1.3714 1.3750 6.0000
O16 1.1892 1.2571 0.7188 0.0000
O18 1.3243 1.3714 1.4375 12.0000
O181 1.1351 1.1143 1.1563 0.0000
O23 0.9730 1.0000 1.0938 10.0000
O24 1.2432 1.2857 1.2500 3.0000
O25 1.5405 1.5143 1.7188 13.0000
O26 1.1081 1.3143 1.3750 1.0000
O27 1.2432 1.3143 1.5000 6.0000
O31 1.1081 1.0286 1.1250 5.0000
O32 1.0270 1.1714 1.1563 2.0000
29
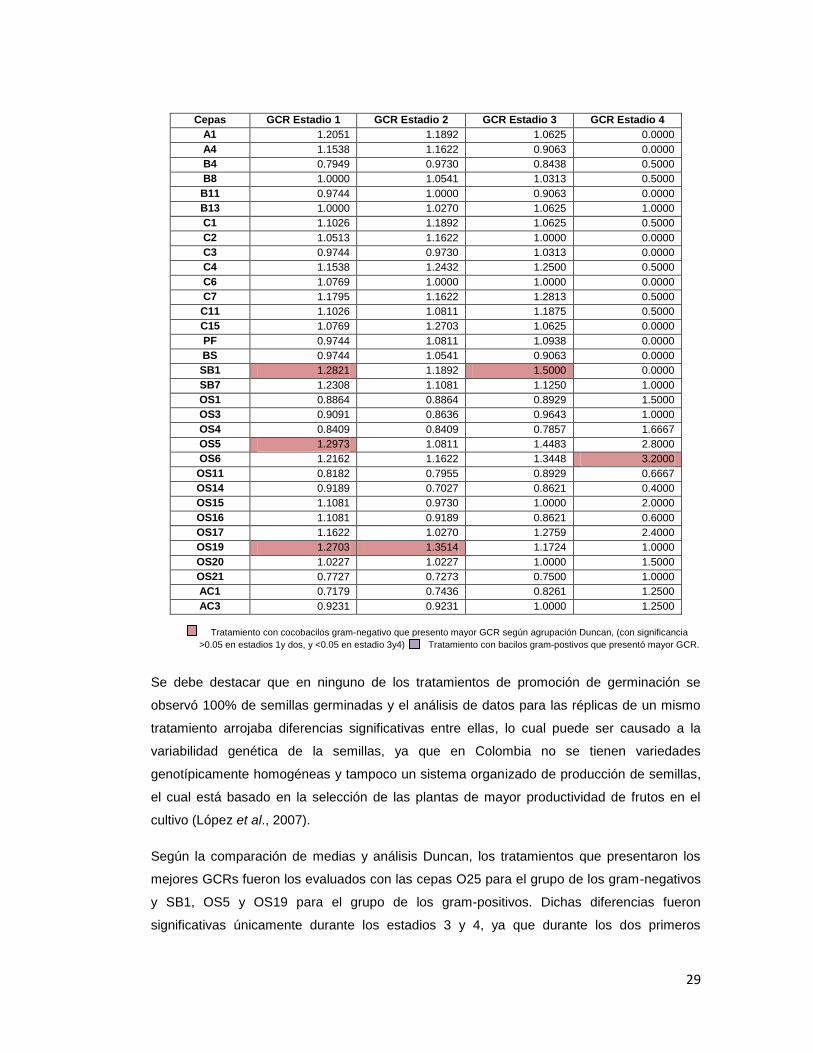
Cepas GCR Estadio 1 GCR Estadio 2 GCR Estadio 3 GCR Estadio 4
A1 1.2051 1.1892 1.0625 0.0000
A4 1.1538 1.1622 0.9063 0.0000
B4 0.7949 0.9730 0.8438 0.5000
B8 1.0000 1.0541 1.0313 0.5000
B11 0.9744 1.0000 0.9063 0.0000
B13 1.0000 1.0270 1.0625 1.0000
C1 1.1026 1.1892 1.0625 0.5000
C2 1.0513 1.1622 1.0000 0.0000
C3 0.9744 0.9730 1.0313 0.0000
C4 1.1538 1.2432 1.2500 0.5000
C6 1.0769 1.0000 1.0000 0.0000
C7 1.1795 1.1622 1.2813 0.5000
C11 1.1026 1.0811 1.1875 0.5000
C15 1.0769 1.2703 1.0625 0.0000
PF 0.9744 1.0811 1.0938 0.0000
BS 0.9744 1.0541 0.9063 0.0000
SB1 1.2821 1.1892 1.5000 0.0000
SB7 1.2308 1.1081 1.1250 1.0000
OS1 0.8864 0.8864 0.8929 1.5000
OS3 0.9091 0.8636 0.9643 1.0000
OS4 0.8409 0.8409 0.7857 1.6667
OS5 1.2973 1.0811 1.4483 2.8000
OS6 1.2162 1.1622 1.3448 3.2000
OS11 0.8182 0.7955 0.8929 0.6667
OS14 0.9189 0.7027 0.8621 0.4000
OS15 1.1081 0.9730 1.0000 2.0000
OS16 1.1081 0.9189 0.8621 0.6000
OS17 1.1622 1.0270 1.2759 2.4000
OS19 1.2703 1.3514 1.1724 1.0000
OS20 1.0227 1.0227 1.0000 1.5000
OS21 0.7727 0.7273 0.7500 1.0000
AC1 0.7179 0.7436 0.8261 1.2500
AC3 0.9231 0.9231 1.0000 1.2500
Tratamiento con cocobacilos gram-negativo que presento mayor GCR según agrupación Duncan, (con significancia
>0.05 en estadios 1y dos, y <0.05 en estadio 3y4) Tratamiento con bacilos gram-postivos que presentó mayor GCR.
Se debe destacar que en ninguno de los tratamientos de promoción de germinación se
observó 100% de semillas germinadas y el análisis de datos para las réplicas de un mismo
tratamiento arrojaba diferencias significativas entre ellas, lo cual puede ser causado a la
variabilidad genética de la semillas, ya que en Colombia no se tienen variedades
genotípicamente homogéneas y tampoco un sistema organizado de producción de semillas,
el cual está basado en la selección de las plantas de mayor productividad de frutos en el
cultivo (López et al., 2007).
Según la comparación de medias y análisis Duncan, los tratamientos que presentaron los
mejores GCRs fueron los evaluados con las cepas O25 para el grupo de los gram-negativos
y SB1, OS5 y OS19 para el grupo de los gram-positivos. Dichas diferencias fueron
significativas únicamente durante los estadios 3 y 4, ya que durante los dos primeros
30
estadios se presentaba solapamiento en los grupos y significancias mayores a 0.05 (Anexo
2.).
Los actinomicetos evaluados presentaron GCR menores o iguales a 1 en los estadios 1, 2 y
3 lo que indica que sus índices de germinación fueron menores a los que control con agua,
es decir, no presentaron promoción de germinación, mientras que en el estadio 4 fueron
mayores, aunque no fueron tan altos como los presentados presentados por los tratamientos
con gram-negativos en los que se observan GCRs mayores a 5 en el estadio 4 con muy
buena promoción de germinación.
6.4. Promoción de crecimiento en invernadero
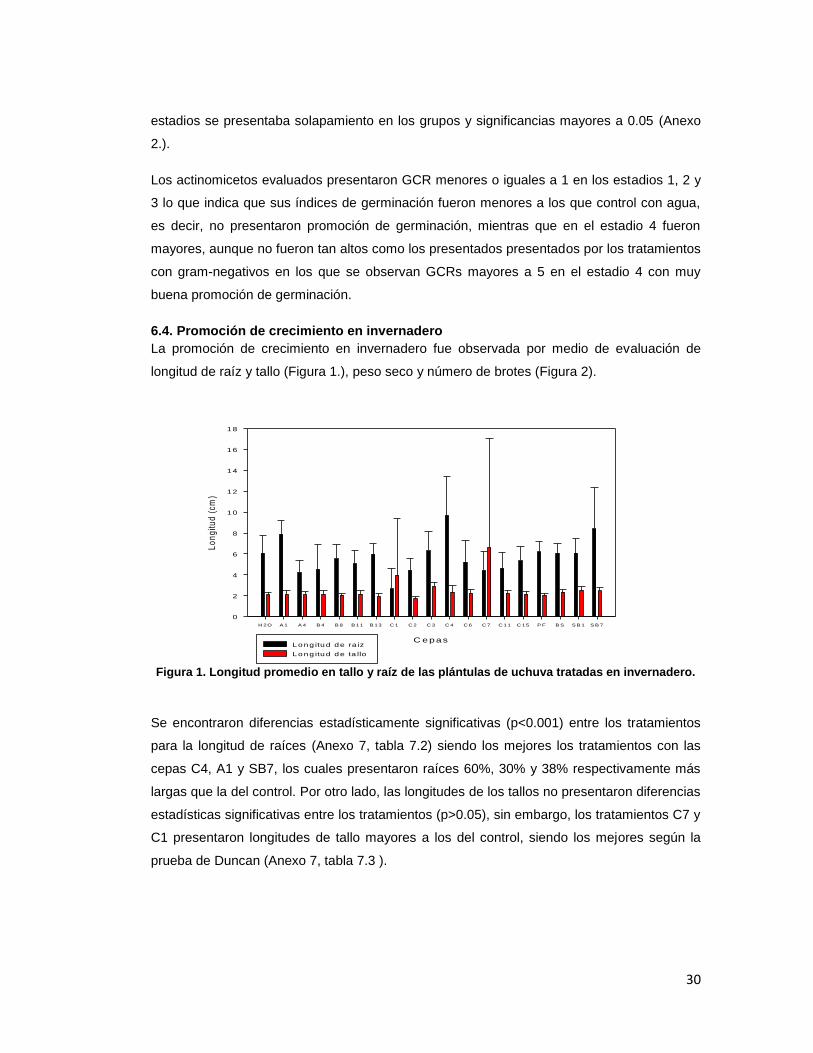
La promoción de crecimiento en invernadero fue observada por medio de evaluación de
longitud de raíz y tallo (Figura 1.), peso seco y número de brotes (Figura 2).
C epas
H 2O A 1 A 4 B 4 B 8 B 11 B 13 C 1 C 2 C 3 C 4 C 6 C 7 C 11 C 15 P F B S S B 1 S B 7
Lo
ng
itud
(cm
)
0
2
4
6
8
10
12
14
16
18
Long itud de ra iz
Long itud de ta llo
Figura 1. Longitud promedio en tallo y raíz de las plántulas de uchuva tratadas en invernadero.
Se encontraron diferencias estadísticamente significativas (p<0.001) entre los tratamientos
para la longitud de raíces (Anexo 7, tabla 7.2) siendo los mejores los tratamientos con las
cepas C4, A1 y SB7, los cuales presentaron raíces 60%, 30% y 38% respectivamente más
largas que la del control. Por otro lado, las longitudes de los tallos no presentaron diferencias
estadísticas significativas entre los tratamientos (p>0.05), sin embargo, los tratamientos C7 y
C1 presentaron longitudes de tallo mayores a los del control, siendo los mejores según la
prueba de Duncan (Anexo 7, tabla 7.3 ).
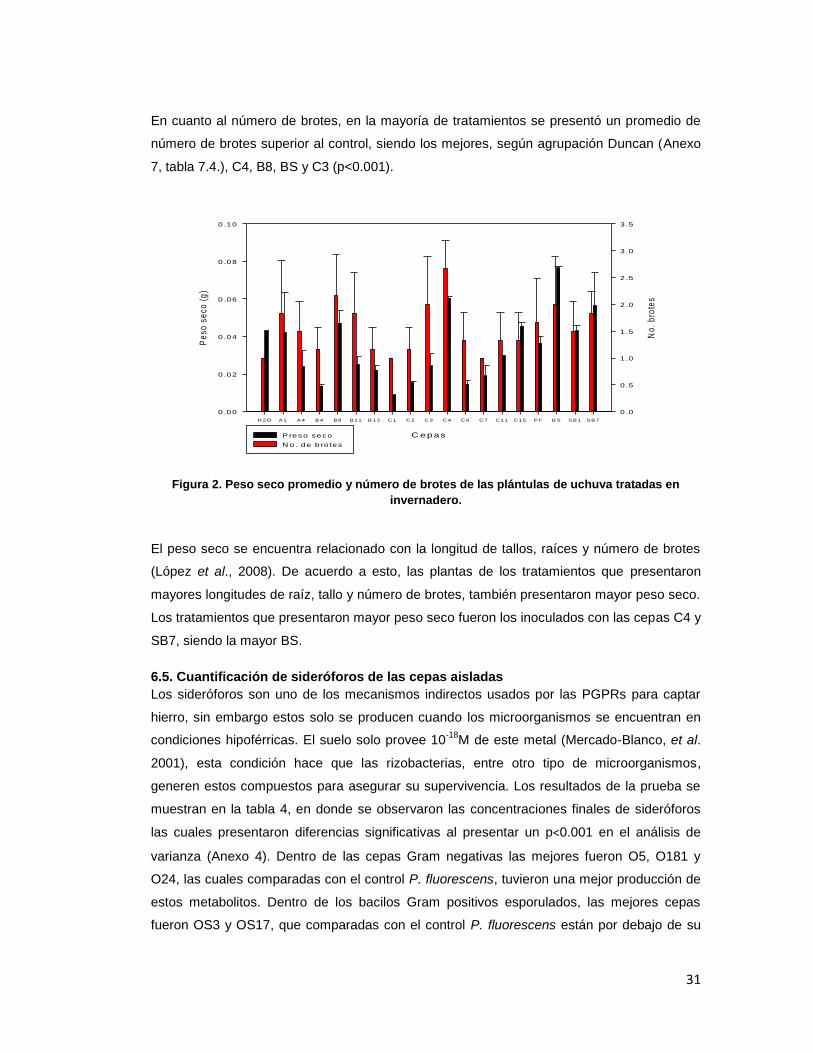
31
En cuanto al número de brotes, en la mayoría de tratamientos se presentó un promedio de
número de brotes superior al control, siendo los mejores, según agrupación Duncan (Anexo
7, tabla 7.4.), C4, B8, BS y C3 (p<0.001).
C e p a s
H 2 O A 1 A 4 B 4 B 8 B 1 1 B 1 3 C 1 C 2 C 3 C 4 C 6 C 7 C 1 1 C 1 5 P F B S S B 1 S B 7
Pe
so s
eco
(g
)
0 .00
0 .02
0 .04
0 .06
0 .08
0 .10
No
. b
rote
s
0 .0
0 .5
1 .0
1 .5
2 .0
2 .5
3 .0
3 .5
P reso seco
N o . de b ro tes
Figura 2. Peso seco promedio y número de brotes de las plántulas de uchuva tratadas en
invernadero.
El peso seco se encuentra relacionado con la longitud de tallos, raíces y número de brotes
(López et al., 2008). De acuerdo a esto, las plantas de los tratamientos que presentaron
mayores longitudes de raíz, tallo y número de brotes, también presentaron mayor peso seco.
Los tratamientos que presentaron mayor peso seco fueron los inoculados con las cepas C4 y
SB7, siendo la mayor BS.
6.5. Cuantificación de sideróforos de las cepas aisladas
Los sideróforos son uno de los mecanismos indirectos usados por las PGPRs para captar
hierro, sin embargo estos solo se producen cuando los microorganismos se encuentran en
condiciones hipoférricas. El suelo solo provee 10-18
M de este metal (Mercado-Blanco, et al.
2001), esta condición hace que las rizobacterias, entre otro tipo de microorganismos,
generen estos compuestos para asegurar su supervivencia. Los resultados de la prueba se
muestran en la tabla 4, en donde se observaron las concentraciones finales de sideróforos
las cuales presentaron diferencias significativas al presentar un p<0.001 en el análisis de
varianza (Anexo 4). Dentro de las cepas Gram negativas las mejores fueron O5, O181 y
O24, las cuales comparadas con el control P. fluorescens, tuvieron una mejor producción de
estos metabolitos. Dentro de los bacilos Gram positivos esporulados, las mejores cepas
fueron OS3 y OS17, que comparadas con el control P. fluorescens están por debajo de su
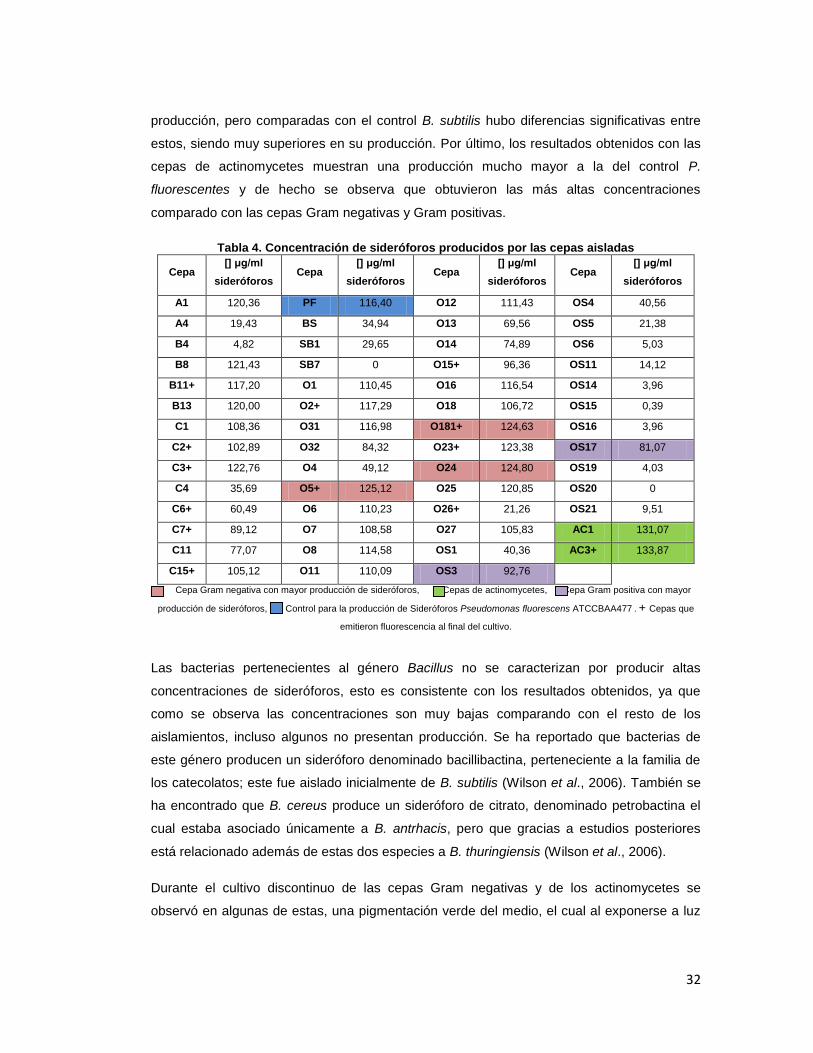
32
producción, pero comparadas con el control B. subtilis hubo diferencias significativas entre
estos, siendo muy superiores en su producción. Por último, los resultados obtenidos con las
cepas de actinomycetes muestran una producción mucho mayor a la del control P.
fluorescentes y de hecho se observa que obtuvieron las más altas concentraciones
comparado con las cepas Gram negativas y Gram positivas.
Tabla 4. Concentración de sideróforos producidos por las cepas aisladas
Cepa [] μg/ml
sideróforos Cepa
[] μg/ml
sideróforos Cepa
[] μg/ml
sideróforos Cepa
[] μg/ml
sideróforos
A1 120,36 PF 116,40 O12 111,43 OS4 40,56
A4 19,43 BS 34,94 O13 69,56 OS5 21,38
B4 4,82 SB1 29,65 O14 74,89 OS6 5,03
B8 121,43 SB7 0 O15+ 96,36 OS11 14,12
B11+ 117,20 O1 110,45 O16 116,54 OS14 3,96
B13 120,00 O2+ 117,29 O18 106,72 OS15 0,39
C1 108,36 O31 116,98 O181+ 124,63 OS16 3,96
C2+ 102,89 O32 84,32 O23+ 123,38 OS17 81,07
C3+ 122,76 O4 49,12 O24 124,80 OS19 4,03
C4 35,69 O5+ 125,12 O25 120,85 OS20 0
C6+ 60,49 O6 110,23 O26+ 21,26 OS21 9,51
C7+ 89,12 O7 108,58 O27 105,83 AC1 131,07
C11 77,07 O8 114,58 OS1 40,36 AC3+ 133,87
C15+ 105,12 O11 110,09 OS3 92,76
Cepa Gram negativa con mayor producción de sideróforos, Cepas de actinomycetes, Cepa Gram positiva con mayor
producción de sideróforos, Control para la producción de Sideróforos Pseudomonas fluorescens ATCCBAA477 . + Cepas que
emitieron fluorescencia al final del cultivo.
Las bacterias pertenecientes al género Bacillus no se caracterizan por producir altas
concentraciones de sideróforos, esto es consistente con los resultados obtenidos, ya que
como se observa las concentraciones son muy bajas comparando con el resto de los
aislamientos, incluso algunos no presentan producción. Se ha reportado que bacterias de
este género producen un sideróforo denominado bacillibactina, perteneciente a la familia de
los catecolatos; este fue aislado inicialmente de B. subtilis (Wilson et al., 2006). También se
ha encontrado que B. cereus produce un sideróforo de citrato, denominado petrobactina el
cual estaba asociado únicamente a B. antrhacis, pero que gracias a estudios posteriores
está relacionado además de estas dos especies a B. thuringiensis (Wilson et al., 2006).
Durante el cultivo discontinuo de las cepas Gram negativas y de los actinomycetes se
observó en algunas de estas, una pigmentación verde del medio, el cual al exponerse a luz

33
UV emitía fluorescencia (Figura 3.), esto se debe a la producción de pioverdina que es el
sideróforo más comúnmente secretado por las cepas fluorescentes de Pseudomonas
Adicionalmente, se observó que las cepas que emitieron fluorescencia también presentaron
los valores más altos de producción de sideróforos (60-124µg/mL), exceptuando a la cepa
O26 (21,26µg/mL). Se ha reportado también que este género es capaz de producir otros
tipos de sideróforos, como pseudomonina, quinolobactina, corrugativa, nocardamina y ácido
piridina-2,6-ditiocarboxílico (Cornelis y Matthijs, 2002 citado por Luque, 2007; Budzikiewiez,
1997). Esta razón puede explicar el por qué cepas como la O24 no presentaron
fluorescencia pero si registraron alta la producción de sideróforos, al igual que otras cepas
evaluadas que tampoco emitieron fluorescencia al exponerlas a luz UV.
a. b.
Figura 3. Cultivo discontinuo en medio mínimo Simmon y Tessman.
Cepa C2, a las 96 horas de cultivo. a. Se observa la pigmentación verde por producción de pioverdinas; b. Se observa la fluorescencia al exponerse a luz UV.
Los resultados obtenidos con las cepas de actinomicetos son consistentes con los
resultados de Díaz (2009), quien reporta en una de sus cepas valores de hasta 237μg/mL y
por Franco (2008) quien reportó un máximo de producción de 142μg/ml al día 11 de cultivo.
En estos cultivos también se observó la producción de pioverdina.
Es importante que el cultivo discontinuo para la producción de sideróforos se haga en un
medio que permita el crecimiento y estimule la producción de estos metabolitos, en este
caso se usó como fuente de carbono ácido succínico y caseína hidrolizada con ácido como
una fuente orgánica de nitrógeno. El primero en condiciones normales, mediante una ruta
metabólica corta a partir de el ciclo de Krebs, es un precursor de la formación de pioverdina
(Diaz, 2009; Budzikiewiez, 1997). Esta condición se confirmó con los resultados de Ramos y
Valero (2009) quienes al usar glucosa como fuente de carbono y solo una fuente inorgánica
de nitrógeno, reportaron que no hubo crecimiento, ni producción de sideróforos por parte de
las cepas aisladas.
34
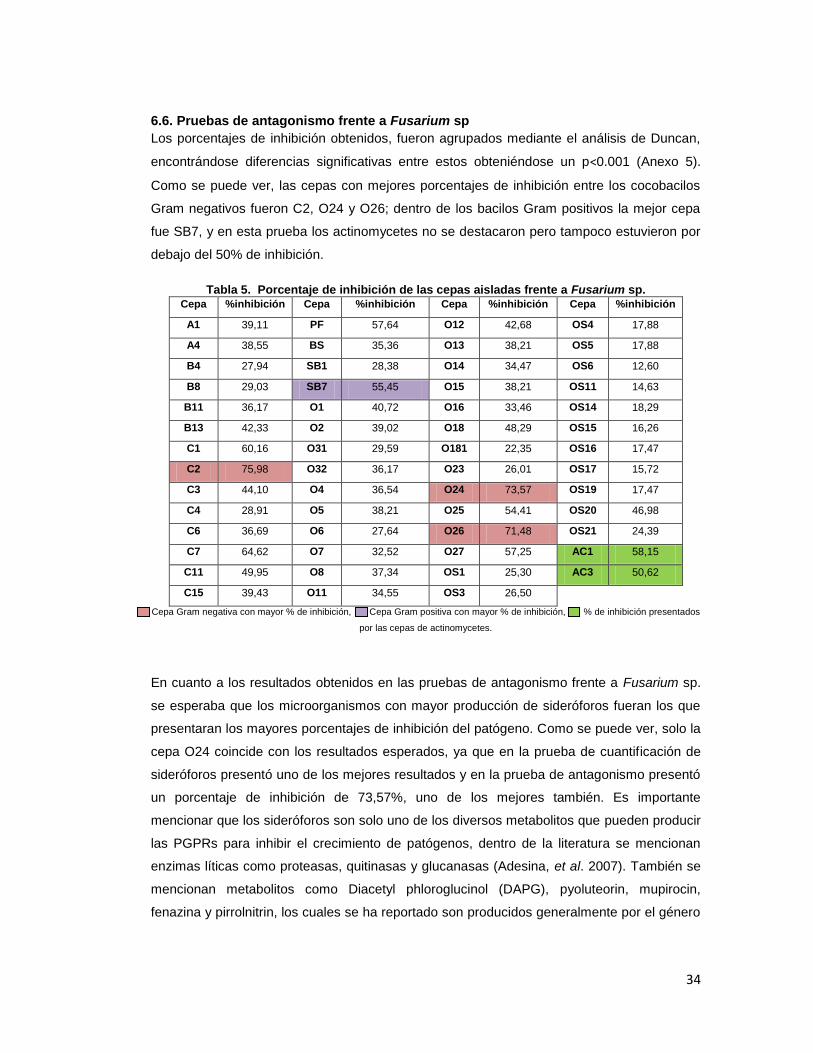
6.6. Pruebas de antagonismo frente a Fusarium sp
Los porcentajes de inhibición obtenidos, fueron agrupados mediante el análisis de Duncan,
encontrándose diferencias significativas entre estos obteniéndose un p<0.001 (Anexo 5).
Como se puede ver, las cepas con mejores porcentajes de inhibición entre los cocobacilos
Gram negativos fueron C2, O24 y O26; dentro de los bacilos Gram positivos la mejor cepa
fue SB7, y en esta prueba los actinomycetes no se destacaron pero tampoco estuvieron por
debajo del 50% de inhibición.
Tabla 5. Porcentaje de inhibición de las cepas aisladas frente a Fusarium sp. Cepa %inhibición Cepa %inhibición Cepa %inhibición Cepa %inhibición
A1 39,11 PF 57,64 O12 42,68 OS4 17,88
A4 38,55 BS 35,36 O13 38,21 OS5 17,88
B4 27,94 SB1 28,38 O14 34,47 OS6 12,60
B8 29,03 SB7 55,45 O15 38,21 OS11 14,63
B11 36,17 O1 40,72 O16 33,46 OS14 18,29
B13 42,33 O2 39,02 O18 48,29 OS15 16,26
C1 60,16 O31 29,59 O181 22,35 OS16 17,47
C2 75,98 O32 36,17 O23 26,01 OS17 15,72
C3 44,10 O4 36,54 O24 73,57 OS19 17,47
C4 28,91 O5 38,21 O25 54,41 OS20 46,98
C6 36,69 O6 27,64 O26 71,48 OS21 24,39
C7 64,62 O7 32,52 O27 57,25 AC1 58,15
C11 49,95 O8 37,34 OS1 25,30 AC3 50,62
C15 39,43 O11 34,55 OS3 26,50
Cepa Gram negativa con mayor % de inhibición, Cepa Gram positiva con mayor % de inhibición, % de inhibición presentados
por las cepas de actinomycetes.
En cuanto a los resultados obtenidos en las pruebas de antagonismo frente a Fusarium sp.
se esperaba que los microorganismos con mayor producción de sideróforos fueran los que
presentaran los mayores porcentajes de inhibición del patógeno. Como se puede ver, solo la
cepa O24 coincide con los resultados esperados, ya que en la prueba de cuantificación de
sideróforos presentó uno de los mejores resultados y en la prueba de antagonismo presentó
un porcentaje de inhibición de 73,57%, uno de los mejores también. Es importante
mencionar que los sideróforos son solo uno de los diversos metabolitos que pueden producir
las PGPRs para inhibir el crecimiento de patógenos, dentro de la literatura se mencionan
enzimas líticas como proteasas, quitinasas y glucanasas (Adesina, et al. 2007). También se
mencionan metabolitos como Diacetyl phloroglucinol (DAPG), pyoluteorin, mupirocin,
fenazina y pirrolnitrin, los cuales se ha reportado son producidos generalmente por el género

35
Pseudomonas y Streptomyces (Fernando et al., 2005). En cuanto a las bacterias del género
Bacillus, se reportan diversos lipopéptidos con propiedades antifúngicas como iturina,
bacillomicina, plipastatina, surfactina y aminopolioles (Fernando et al., 2005). Es posible que
en cepas como C2, que presentó una baja producción de sideróforos, inhibió al patógeno
mediante la producción no solo de estos metabolitos, sino que también mediante la
producción de enzimas o antibióticos, como los que se mencionaron anteriormente. Este
también puede ser el caso de los bacilos Gram positivos y de los actinomycetes. En estos
últimos se observó la producción de pigmentos que se difundieron en el medio de cultivo
(Figura 4).
a. b. c.
d.
Figura 4. Imágenes de la prueba de Antagonismo
Se observa el crecimiento influenciado y no influenciado de Fusarium sp., frente a las bacterias antagonistas, a los 10 días de incubación. a. cepa O24 se observa la inhibición del crecimiento del hongo, b. cepa OS14 no hay inhibición del crecimiento del
hongo, c. Control, d. cepa de actinomycetes AC3.
Estos resultados son consistentes con los reportados por Franco (2008) quien reportó un
porcentaje de inhibición de 68% frente a F. oxysporum, solo con uno de sus aislamientos.
Estos resultados, en general, superan los obtenidos por Ramos y Valero (2009), cuyos
aislamientos de Gram negativos y Gram positivos, no superaron el 40% de inhibición frente a
Fusarium sp. Es posible que los porcentajes de inhibición obtenidos con nuestros
aislamientos hayan sido mejores gracias a que 48 horas antes de haberlas sembrado en el
PDA para el enfrentamiento dual, estas fueron reconstituidas en agar King B para que de
esta forma hubiera una inducción previa de sideróforos.

36
6.7. Discusión general
Evaluando el porcentaje de cepas promisorias para PGPR obtenido con respecto al total
aislado durante el primer y segundo muestreo, se observó que el porcentaje de cepas
promisorias obtenido durante el segundo muestreo (Anexo 8), por lo cual se estableció que
es mejor trabajar con todos los aislamientos durante los ensayos para establecer las cepas
promisorias para promoción de crecimiento, de tal manera que no se sesguen los resultados,
descartando cepas que aunque no pertenezcan a los géneros de interés, puedan presentar
buenos resultados en los ensayos realizados, y por tanto presentar un buen perfil como
rizobacterias promotoras de crecimiento vegetal.
El ácido indol acético promueve el crecimiento radicular y es uno de los parámetros más
utilizados para medir los efectos beneficiosos de las bacterias promotoras de crecimiento, ya
que ayudan al rápido establecimiento de raíces y anclaje de la planta al suelo, mejorando la
capacidad de obtención de agua y nutrientes del medio y por tanto aumentando sus
posibilidades de sobrevivencia (Patten y Glick, 2002). Sin embargo, la producción de AIA
parece no ser proporcional al efecto de promoción de crecimiento que pueda tener una
PGPR sobre una planta evaluado por medio de índice de germinación relativo, ya que la
cepa O23, mayor productora de AIA, no presentó GCR altos en los primeros estadios de
germinación como se esperaría; mientras que el tratamiento con la cepa O25 presentó los
mejores GCR en todos los estadios de geminación aunque esta no se encontró entre los
mejores productores de AIA (Anexo 2). Adicionalmente, los índices de correlación Pearson
evaluados entre la producción de AIA y el GCR relativo para cada estadío de germinacón
son menores a 0,1 para los estadios 1, 2 y 3, mostrando que no se presenta correlación
entre estas dos variables, y aunque en el estadio 4 el valor de correlación aumenta
(r=0.240), no es representativo. El mismo comportamiento se observó en las pruebas de
promoción de crecimiento en invernadero en relación a la producción de ácido indol acético.
Los mayores valores de longitud de raíz, tallo, número de brotes y peso seco (cepas C4,
SB7, BS, A1, C4, C7 y C1), no corresponden con las mejores cepas productoras de AIA (1 -
4μg/mL), mientras que las cepas B13 y B11 presentaron alta producción de AIA (8.94 y 9.61
μg/mL respectivamente), no presentaron promoción en crecimiento de raíz y tallo (Figura 2).
Respecto a los resultados mencionados anteriormente se presume que las concentraciones
de AIA en algunas cepas, aunque bajas, pueden ser suficientes para tener una influencia
directa sobre la germinación, teniendo en cuenta que los reguladores de crecimiento vegetal
actúan en bajas concentraciones (menores a 1μM) (García et al., 2005). Adicionalmente se
debe tener en cuenta que la promoción de crecimiento exitosa por parte de una cepa no está
dada únicamente por el potencial de producción de AIA, sino la colonización competitiva en

37
la rizosfera durante un periodo de tiempo considerable para que esta tenga efecto (Compant
et al., 2005). Estos resultados también podrían confirmar lo propuesto por Glick y
colaboradores (1998), en donde una alta concentración de AIA producido por las
rizobacterias sumado al AIA producido endógenamente por la planta pueden activar la
producción de 1-Acido 1-aminociclopropano 1-carboxilico (ACC), precursor del etileno, el
cual puede anular la acción del AIA inhibiendo la elongación celular en la planta.
Por otro lado, un único mecanismo puede ser responsable por algún porcentaje de la
promoción de crecimiento observada, pero el ciento por ciento de la promoción de
crecimiento debe ser atribuida a los muchos mecanismos que actúan sinérgicamente
durante la etapa de desarrollo de la planta (Kloepper, 2003). Entonces, además de la
producción de AIA, otros mecanismos se encuentran involucrados en la promoción de
crecimiento de las plantas por parte de la PGPRs, entre estos la colonización competitiva de
raíces, la producción de sideróforos, la solubilización de nutrientes en el suelo y la
producción de antibióticos y enzimas líticas para combatir patógenos (Patten y Glick, 2002;
Antoun y Prevost, 2005; Compant et. al, 2005).
Los sideróforos y la capacidad antagonista están relacionados, ya que la capacidad de
producir estos metabolitos es una ventaja competitiva de las PGPRs frente a
microorganismos que no son capaces de producirlos, inhibiendo el crecimiento de patógenos
en condiciones limitantes de hierro. Sin embargo, al realizar el análisis de correlación entre
estas dos variables no se observa una relación significativa entre los dos (r: 0.409). En la
literatura se observa que en las pruebas de antagonismo para estimular la producción de
sideróforos y así identificar a los posibles antagonistas, la técnica de enfrentamiento en
placa es montada en un medio en donde las concentraciones de hierro son limitantes,
asegurando de esta forma la secreción de estos metabolitos. Con este fin se usa
generalmente el medio King B para este tipo de pruebas (Sindhu et al., 1999; Mercado-
Blanco et al., 2004; Adesina et al., 2007), también agar Martin desferrizado (Kurek y
Jaroszuk-SScisel, 2003). De esta forma se asume que se podrán obtener resultados
directamente relacionados con la producción de sideróforos, ya que como se sabe el medio
PDA es complejo y puede traer consigo trazas de hierro que inhiban la producción de los
mismos.
Para el aislamiento de PGPRs es importante que las muestras de suelo rizosférico
provengan de suelos supresivos, estos son aquellos que tienen capacidad para controlar
enfermedades de origen edáfico, existen de forma natural y deben ser buscados en aquellas
zonas en donde esté el patógeno y ocurra la enfermedad. Por su modo de acción estos

38
suelos pueden ser clasificados en suelos que impiden el establecimiento del patógeno o que
permitiendo su establecimiento impiden su total o parcial manifestación (Florez et al., 2007).
Según lo mencionado anteriormente, las muestras usadas en este estudio provienen de un
suelo supresivo, ya que se tomaron de zonas en donde está la enfermedad, pero de la
rizósfera de plantas sanas; por esta razón se espera encontrar microorganismos
antagónicos, que logren inhibir al patógeno in vitro, como lo hacen in vivo, como es el caso
de Adesina y colaboradores (2007) quienes realizaron un secreening de microorganismos
antagonistas de diversos fitopatógenos en suelos supresivos europeos, encontrando de
1788 aislamientos 327 antagonistas, entre Pseudomonas, Bacillus y Streptomyces. Se ha
reportado que la producción de sideróforos como pseudomonina y pioquelina, tiene como
intermediario ácido salicílico (AS) (Mercado-Blanco, et al. 2001), la síntesis de esta molécula
podría ser considerada como un mecanismo indirecto de promoción de crecimiento vegetal
ya que actúa como señalizador para la inducción de resistencia sistémica en plantas
(Halfeld-Viera et al., 2006). La producción de ácido salicílico podría establecerse como un
criterio de selección para considerar a una cepa PGPR como promisoria.
7. CONCLUSIONES
A partir de las muestras trabajadas se obtuvieron en el primer y segundo muestreo 88
aislamientos de los cuales se identificaron como Pseudomonas fluorescens 26 aislamientos,
Bacillus 11 aislamientos y Actinomycetes 2 aislamientos.
Los resultados obtenidos durante el desarrollo de las pruebas y la identificación indican que
es mejor trabajar primero con el total de los aislamientos iniciales las pruebas de selección
de promotores de crecimiento y por ultimo identificar los mejores, ya que de esta forma se
disminuye la probabilidad de descartar cepas promisorias durante la identificación, las cuales
no estén dentro de las especies de interés.
Se encontraron diferencias significativas en cuanto a la producción de AIA por parte de las
rizobacterias evaluadas, determinando que el aislamiento O23 fue el mejor productor y fue
agrupado conjuntamente con el control utilizado, A. vinelandii en el grupo de los Gram-
negativos, OS3, OS5 y OS20 en el grupo de los Gram-positivos y AC1 en el grupo de los
actinomycetes.
No se encontraron diferencias significativas en el GCR en los estadios de germinación 1 y 2
entre los tratamientos con cocobacilos gram-negativos, siendo la mejor cepa según

39
agrupación Duncan la O25 al igual que en los estadios 3 y 4, donde si hubo diferencias
significativas.
Se encontraron diferencias significativas en el GCR de todos los estadios de germinación
entre los tratamientos con bacilos Gram-positivos esporulados, siendo las mejores cepas
SB1, OS5 y OS19.
No se observó una correlación fuerte entre la producción de AIA y la promoción de
germinación, ya que los mayores productores de AIA no presentaron los mayores GCR, por
lo cual se presume que en algunas cepas la concentraciones de AIA bajas pueden ser
suficientes para tener una influencia sobre la germinación.
La promoción de crecimiento exitosa por parte de una cepa no está dada únicamente por el
potencial de producción de AIA, mas debe ser atribuida a los muchos mecanismos que
actúan sinérgicamente durante la etapa de desarrollo de la planta.
Se observaron diferencias significativas en cuanto a la producción de sideróforos por parte
de las cepas aisladas, encontrándose los mejores resultados en las cepas O5, O181 y O24
de los aislamientos Gram negativos, y OS3 y OS17 las mejores cepas Gram positivas;
siendo las cepas Gram negativas superiores en la producción con valore mayores a los de
P. fluorescens ATCCBAA477, la cepa de referencia para esta prueba. Los actinomycetes
obtuvieron las concentraciones de producción más altas superando a las cepas Gram
negativas.
Los resultados en las pruebas de antagonismo demostraron que las mejores cepas de los
aislamientos fueron C2, O24 y O26 de las cepas Gram negativas alcanzado porcentajes de
inhibición que van de 71 al 75%; dentro de las cepas Gram positivas la mejor fue la SB7 con
un porcentaje de 55.4%, siendo superiores las cepas Gram negativas. Los actinomycetes
presentaron una inhibición que no superó el 58%.
Con los resultados obtenidos no se observó una relación proporcional entre la producción de
sideróforos y la capacidad antagónica frente a la cepa de Fusarium sp., esto es posible
gracias a la capacidad que tienen los PGPRs de producir diversos metabolitos con potencial
antibiótico que pueden inhibir el crecimiento del patógeno.

40
8. RECOMENDACIONES
Se recomienda en las pruebas de germinación, que el seguimiento de las semillas se
extienda a 30 días, ya que estas tienen un crecimiento muy lento y así es posible observar
una mayor cantidad de plántulas en estadio cuatro. Lo cual puede hacer variar los valores
GCR.
En las pruebas de antagonismo se sugiere el uso de un medio con un contenido de hierro
mínimo para que de esta forma, se induzca la producción de sideróforos. De esta forma se
observará que la inhibición si está relacionada con la producción de estos metabolitos.
También se recomienda realizar otras pruebas para la selección de las PGPR como fosfato
solubilización, producción de enzimas líticas como quitinasas, proteasas y glucanasas, y la
medición de la producción de ácido salicílico, ya que todos estos son mecanismos usados
por estas rizobacterias para promover el crecimiento de las plantas.
Se sugiere que las pruebas de invernadero se realicen con plántulas sin inoculación, durante
su germinación, ya que como se sabe hay cepas que pueden estimular los diferentes
estadios de germinación y hay otras que estimulan el crecimiento en el suelo, así que de
esta forma usando plantas homogéneas se observará el efecto real de cada cepa.
9. REFERENCIAS
ADESINA,M.F.; LEMBKE,A.; COSTA, R.; SPEKSNIJDER, A.; SMALLA, K. 2007. Screening of bacterial isolates from various European soils for in vitro antagonistic activity towards Rhizoctonia solani and Fusarium oxysporum: Site-dependent composition and diversity revealed. Soil Biology & Biochemistry, 39:2818–2828.
AGRIOS, GN. Fitopatología. Segunda edición, Noriega editores. México 2002.
AHMAD, S.; MALIK, A.; YASMIN, R.; ULLAH, N.; GUL, W.; KHAN, M.P.; NAWAZ, R.H. Y AFZA, N. 1999. Withanolides from Physalis peruvia. Phytochemistry, 50: 647-651.
ANÓNIMO. 2001. Boletín SIM. Perfil de Producto No. 13. Julio–Septiembre del 2001 Corporación Colombia Internacional. Documento en línea: www.cci.org.co. Fecha: 13/07/09.
ANÓNIMO. 2002. Asesoria integral en comercio exterior. Fenalcomex. Documento en Línea: www.fenalcotolima.com/fenalcomex. Fecha: 13/07/09.
ANTOUN, H.; PREVOST, D. 2005. Ecology of plant growth promoting rhizobacteria. PGPR: Biocontrol and fertilization. Pg 1-38.

41
ARTECA, R. N. 1996. Plant growth substances: principles and applications. Primera edición. Chapman & Hall. Pennsylvania, USA. 332p.
BERGEY, D.H. 2000. Bergey's manual of determinative bacteriology. Novena edición. Lippincott Williams & Wilkins. Philadelphia, USA. Página 787.
BLAKE, P.; MANDER, L.N.; TAYLOR, D.R.; CRISP, C.M. Y OWEN, C.J. 2000. Identification of endogenous gibberellins in strawberry, including the novel gibberellins GA123, GA124 and GA125. Phytochemistry, 55: 887-890.
BOBADILLA, C. Y RINCÓN, S.C. 2008. Aislamienot y producción de bacterias fosfatos solubilizadoras a partir de compost obtenido de residuos de plaza. Microbiologo Industrial. Pontificia Universidad Javeriana. Facultad de Ciencias. Pagina 16.
BUDZIKIEWIEZ, H. 1997. Siderophores from Fluorescent Pseudomonas. Studies in Natural Products Chemistry, Vol.19. 793 – 835.
CALIFA, C. Y GONZALES, A. 2009. Estandarización de un bioensayo en modelos vegetales para la detección de reguladores vegetal (ácido indol acético y giberelinas) en cultivos microbianos. Microbiologo Industrial, Microbiologo Agrícola y veterianrio. Pontificia Universidad Javeriana. Facultad de Ciencias.
CASAS, A.G. 2006. Parámetros de calidad en uchuva (Physalis peruviana L) con énfasis en EUREP – GAP. Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Agronomía. Bogotá. Páginas 6-20.
CASTILLO, G.; ALTUNA, B.; MICHELENA, G.; SÁNCHEZ-BRAVO, J. Y ACOSTA, M. 2005. Cuantificación del contenido de ácido indolacético (AIA) en un caldo de fermentación microbiana. Anales de Biologia, 27: 137-142.
CELIS, L.X. Y GALLARDO, I.R. 2008. Estandarización de métodos de detección para promotores de crecimiento vegetal (Ácido indol acético y Giberelinas) en cultivos microbianos. Microbiologo Agrícola y Veterinario. Pontificia Universidad Javeriana. Facultad de ciencias. Bogotá. Página: 159.
COMPANT, S.; DUFFY, B.; NOWAK, J.; CLE´MENT, C.; BARKA, E.A. 2005. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Applied and environmental microbiology, 71(9): 4951–4959.
CÓNGORA, A.C. Y ROJAS, P. 2006. Incidencia de las enfermedades en uchuva Physalis peruviana L por estado fenológico y de acuerdo con la ubicación en los diferentes estratos de la planta, en el departamento de Cundinamarca. Microbiologo agrícola y veterinario. Pontificia Universidad Javeriana. Facultad de Ciencias. Página 18.
De La ROTTA, M.C. Y QUEVEDO, K. 2005. Marchitamiento vascular de Physalis floridiana L causada por Fusarium oxysporum. En: resúmenes del XXVI congreso Ascolfi. Bogotá. Octubre 5-7 páginas 70.
DEY, R.; PAL, K.K.; BHATT, D.M. Y CHAUHAN, S.M. 2006. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiological Research, 159: 371—394

42
DIAZ, S. 2009. Detección y cuantificación de sideroforos en aislamientos de actinos provenientes de suelo Cundiboyacense. Microbiologo Industrial. Pontificia Universidad Javeriana. Facultad de ciencias. Bogotá.
DIMKPA A, C.O.; SVATOŠ B, A.; DABROWSKA C, P.;SCHMIDT A, A.; BOLAND C, W.; KOTHE, E. 2008. Involvement of siderophores in the reduction of metal-induced inhibition of auxin synthesis in Streptomyces spp. Chemosphere, 74:19–25.
EL-TARABIL K.A. Y SIVASITHAMPARA, K. 2006. Non-streptomycete actinomycetes as biocontrol agents of soil-borne fungal plant pathogens and as plant growth promoters. Soil Biology & Biochemistry, 38: 1505–1520
ESTUPIÑAN, H. Y OSSA, J.A. 2007. Efecto del agente causal de la marchitez vascular de la uchuva (Physalis peruviana L.) el hongo Fusarium oxysporum Schlecht, sobre algunas solanáceas y otras especies cultivadas afectadas por formas especiales del microorganismo. Microbiologo Agrícola y veterinario. Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá. Página 20.
FELICI, C.; VETTORI, L.; GIRALDI, E.; FORINO, L.A.; TOFFANIN, L.; TAGLIASACCHI, L.A.; NUTI, M. 2008. Single and co-inoculation of Bacillus subtilis and Azospirillum brasilense on Lycopersicon esculentum: Effects on plant growth and rhizosphere microbial community. Applied Soil Ecology, 40:260–270.
FERNANDO, W.G.; NAKEERAN, S.; ZHANG, Y. 2005. Biosynthesis of antibiotics by PGPR and its relation in biocontrol of plant disease. PGPR: Biocontrol and Biofertilization: 67- 109.
FLORES, C.; FERNANDEZ, B.A.; AGUIRRE, C.; RIDAO, A. 2007. Supresión de la podredumbre radical del palto en un suelo de jujuy, Argentina. Actas VI Congreso Mundial del Aguacate. Viña Del Mar, Chile. 12 – 16 Nov.
FLOREZ, V.; FISCHER, G. Y SORA, A. 2000. Producción, Poscosecha y Exportación de la Uchuva (Physalis peruviana L.). Unibiblos. Universidad Nacional de Colombia, Facultad de Agronomía, pág. 11, 131.
GARCIA, I.N.; HYNES, R.K.; NELSON, L.M. 2005. Role of cytokinins in plant growth by rhizosphere bacteria. PGPR: Biocontrol and fertilization. Pg 173-195.
GLICK, B.R.; PENROSE, D.M.; LI J.1998. A model for the lowering of plant ethylene concentration by plant growth-promoting bacteria. Journal of Theoretical biology. 190:63-68.
HALFELD-VIEIRA, B.A.; VIEIRA, J.R.; ROMEIRO, R.S.; ALVES SILVA, H.S.; BARACAT-PEREIRA, M.C. 2006. Induction of systemic resistance in tomato by the autochthonous phylloplane resident Bacillus cereus. Pesquisa agropecuaria brasileira, 41-8:1247-1252.
HERRERA J., ALIZAGA R., GUEVARA E. y JIMENEZ E. 2006. Germinación y crecimiento de la planta. Fisiología de productos tropicales. Primera Edición. Editorial Universidad de Costa Rica. San José Costa Rica. 60 p.
INTERNATIOAL SEED TESTING ASSOCIATION. 2006. International rules for seed
testing. Bassersdorf, Suiza. ISTA.

43
JOO, G.J.; KIM, Y.M.; LEE, I.J.; SONG, K.S.; RHEE, I.K. 2004. Growth promotion of red pepper plug seedlings and the production of gibberellins by Bacillus cereus, Bacillus macroides and Bacillus pumilus. Biotechnology Letters 26: 487–491.
KLOEPPER, J.W. 2003. A review of mechanisms for plant growth promotion by pgpr. 6th international pgpr workshop. Calicut, india. Session vi - mechanisms of growth promotion. Pg 81-92.
KOKALIS-BURELLE, N.; KLOEPPER, J.W. y Reddy, M.S. 2006. Plant growth-promoting rhizobacteria as transplant amendments and their effects on indigenous rhizosphere microorganisms. Applied Soil Ecology, 31: 91–100.
KUREK, E.; JAROSZUK-SSCISEL, J. 2003. Rye (Secale cereale) growth promotion by Pseudomonas fluorescens strains and their interactions with Fusarium culmorum under
various soil conditions. Biological Control 26: 48–56
LOPEZ, F.J.; GUIO, N.R.; FISCHER, G.; MIRANDA, D. 2007. Propagación de uchuva (Physalis peruviana l.) Mediante diferentes tipos de esquejes y sustratos. Revista
Facultad Nacional de Agronomia. 61(1):4347-4357.
LUQUE, V. 2007. Metabolismo del cianuro y del cianato en Pseudomonas pseudoalcaligenes CECT5344. Aplicaciones biotecnológicas. Capitulo IV, Discusión. Documento en línea: http://www.economiaandaluza.es/archivos/capitulos/cap295.pdf. Fecha: 21/11/09.
MERCADO-BLANCO, J.; DRIFT VAN DER, K.G.; OLSSON, P.;THOMAS-OATES, J.; LOON VAN, L.; BAKKER, P. 2001. Analysis of the pmsCEAB Gene Cluster Involved in Biosynthesis of Salicylic Acid and the Siderophore Pseudomonine in the Biocontrol Strain Pseudomonas fluorescens WCS374. Journal of Bacteriology, Pg. 1909–1920
MERCADO-BLANCO, J.; RODRÍGUEZ-JURADO, D.; HERVAS, A.; JIMENEZ-DIAZ, R. 2004. Suppression of Verticillium wilt in olive planting stocks by root-associated fluorescent Pseudomonas spp. Biological Control 30: 474–486.
PATTEN C. Y GLICK B.R. 1996. Bacterial biosynthesis of Indole-3-acetic acid (review). Canadian Journal of Microbiology, 42: 207-220.
PATTEN, C.L.; GLICK. B.L. 2002. Role of Pseudomonas putida Indoleacetic Acid in Development of the Host Plant Root System. Applied and environmental microbiology, 68:3795–380.
PEÑA, H. Y REYES, I. 2007. Aislamiento y evaluación de bacterias fijadoras de nitrógeno y disolventes de fosfatos en la promoción del crecimiento de la lechuga (Lactuca sativa l.) Interciencia vol. 32 n.8.
PEREZ, S.; CABIROL, N.; GEORGE, R.; ZAMUDIO, L.S. y FERNÁNDEZ, F.J. 2007. O-CAS, a fast and universal method for siderophore detection. Journal of Microbiological Methods. 70:127–131
RAMOS, M.; VALERO, C. 2008. Aislamiento de rizobacterias promotores de germinación de uchuva (Physalis peruviana L.) Y antagónicas frente a Fusarium oxysprum. Microbiología industrial y Microbiología Agrícola y Veterinaria. Pontificia Universidad Javeriana. Facultad de Ciencias. Departamento de Microbiologia. Bogotá, Colombia. 128 páginas.

44
REYES, I. ALVARES, L. EL-AYUOBI H. VALERY A. 2008. Selección y evaluación de
rizobacterias promotoras del crecimiento en pimentón y maíz. Bioagro vol 20(1). Páginas 37-
48.
RIVIEROS, J.A. 2008. Evaluacion de microorganismos con potencial en biofertilizacion para un cultivo de uchuva (Physalis peruviana L.) en etapas de semillero y vivero. Ingeniero Agrónomo. Universidad Nacional de Colombia. Facultad de Agronomía. Página 11 – 12.
SANABRIA, S. 2005. Situación actual de la uchuva en Colombia. Avances en cultivo,
poscosecha y exportación de la uchuva Physalis peruviana L. en Colombia. Unibiblos.
Universidad Nacional. Facultad de Agronomía. Página 20.
SCHAAD, N.W. 1994. Laboratory guide for identification of plant pathogenic bacteria. Second Edition. St. Paul, Minnesota: American Phytopathological Society, página 157
SINDHU, S.; GUPTA, K.; DADARWAL, K.R. 1999. Antagonistic effect of Pseudomonas spp. on pathogenic fungi and enhancement of growth of green gram (Vigna radiata). Biol Fertil Soils 29: 62–68
TSAVKELOVA, E.; CHERDYNTSEVA, T.A.; BOTINA, S.G.; NETRUSOV, A.I. 2006. Bacteria associated with orchid roots and microbial production of auxin. Microbiological Research, 162:69—76.
TUDZINSKI, B. y SHARON, A. 2002. Byosinthesis, biological role and application of fungal phytohormones. The mycota. 183-205
VAN LOON, L.C.; BAKKER, P.A. 2005. Induced systemic resstance as a mechanism of disease suppression by rhizobacteria. PGPR: Biocontrol and fertilization. Pg 39-66.
VASANTHAKUMAR, A.; MCMANUS, P.S. 2004 Indole-3-Acetic Acid-Producing Bacteria Are Associated with Cranberry Stem Gall. Phytopathology,94(11):1164-1171.
VESSEY, J.K. 2003. Plant growth promoting rhizobacteria as biofertilizers. Plant and Soil, 255: 571–586.
WILSON, M.; ABERGEL, R.J.; RAYMOND, K.N.; ARCENEAUX, J.; BYERS, B.R.
2006. Siderophores of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis.
Biochemical and Biophysical Research Communications 348: 320–325.
ZAPATA, J.L.; SALDARRIAGA, A.; LONDOÑO, M. Y DÍAZ, C. 2002. Manejo del cultivo de la uchuva en Colombia, Boletín técnico. Corpoica, Documento en línea: http://www.agronet.gov.co/www/docs_si2/Manejo%20del%20cultivo%20de%20la%20ucuchu.pdf. Fecha: 20/07/09.
ZHANG , S.; MOYNE, A.L.; REDDY, M.S. Y KLOEPPER, J.W. 2002. The role of salicylic acid in induced systemic resistance elicited by plant growth-promoting rhizobacteria against blue mold of tobacco. Biological Control, 25: 288–296.
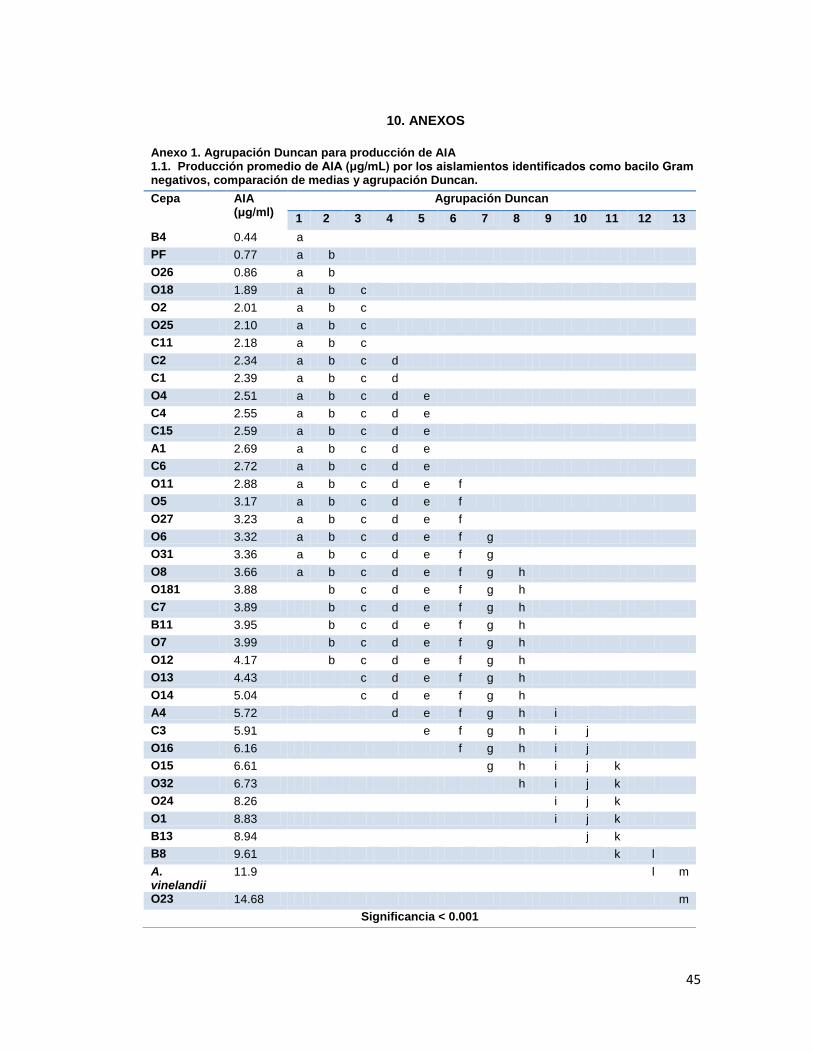
45
10. ANEXOS
Anexo 1. Agrupación Duncan para producción de AIA 1.1. Producción promedio de AIA (μg/mL) por los aislamientos identificados como bacilo Gram negativos, comparación de medias y agrupación Duncan.
Cepa AIA (μg/ml)
Agrupación Duncan
1 2 3 4 5 6 7 8 9 10 11 12 13
B4 0.44 a
PF 0.77 a b
O26 0.86 a b
O18 1.89 a b c
O2 2.01 a b c
O25 2.10 a b c
C11 2.18 a b c
C2 2.34 a b c d
C1 2.39 a b c d
O4 2.51 a b c d e
C4 2.55 a b c d e
C15 2.59 a b c d e
A1 2.69 a b c d e
C6 2.72 a b c d e
O11 2.88 a b c d e f
O5 3.17 a b c d e f
O27 3.23 a b c d e f
O6 3.32 a b c d e f g
O31 3.36 a b c d e f g
O8 3.66 a b c d e f g h
O181 3.88 b c d e f g h
C7 3.89 b c d e f g h
B11 3.95 b c d e f g h
O7 3.99 b c d e f g h
O12 4.17 b c d e f g h
O13 4.43 c d e f g h
O14 5.04 c d e f g h
A4 5.72 d e f g h i
C3 5.91 e f g h i j
O16 6.16 f g h i j
O15 6.61 g h i j k
O32 6.73 h i j k
O24 8.26 i j k
O1 8.83 i j k
B13 8.94 j k
B8 9.61 k l
A. vinelandii
11.9 l m
O23 14.68 m
Significancia < 0.001
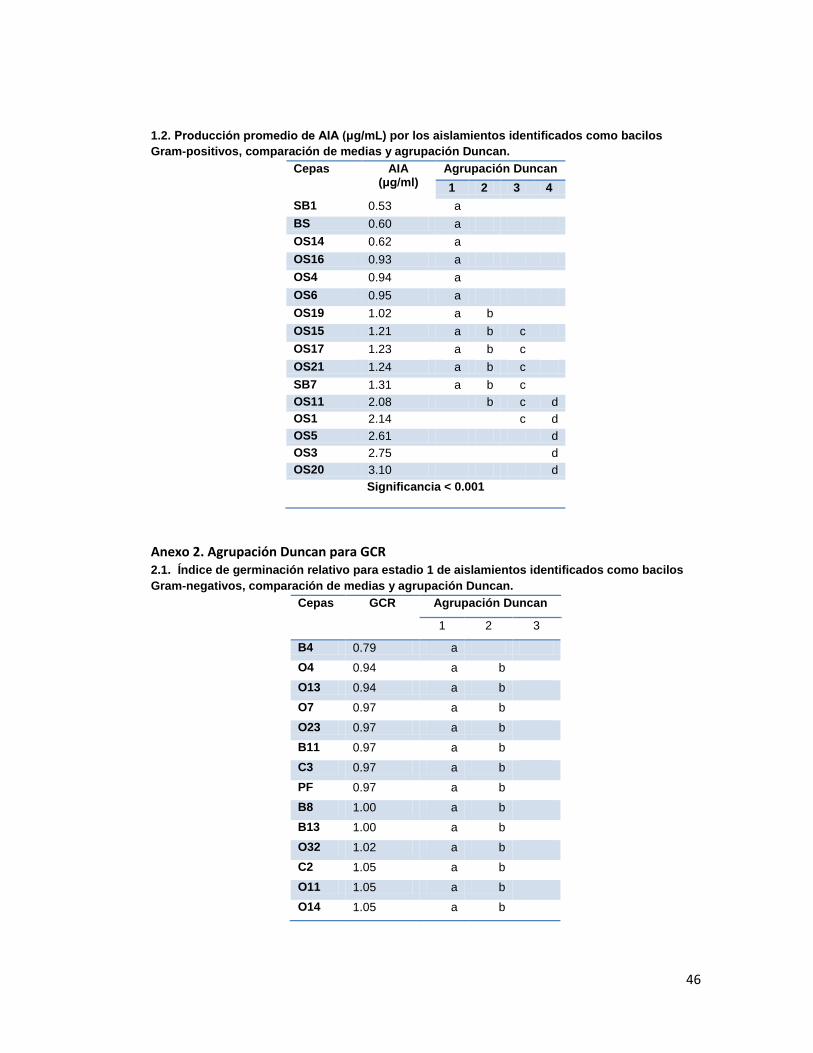
46
1.2. Producción promedio de AIA (μg/mL) por los aislamientos identificados como bacilos
Gram-positivos, comparación de medias y agrupación Duncan.
Cepas AIA (μg/ml)
Agrupación Duncan
1 2 3 4
SB1 0.53 a
BS 0.60 a
OS14 0.62 a
OS16 0.93 a
OS4 0.94 a
OS6 0.95 a
OS19 1.02 a b
OS15 1.21 a b c
OS17 1.23 a b c
OS21 1.24 a b c
SB7 1.31 a b c
OS11 2.08 b c d
OS1 2.14 c d
OS5 2.61 d
OS3 2.75 d
OS20 3.10 d
Significancia < 0.001
Anexo 2. Agrupación Duncan para GCR 2.1. Índice de germinación relativo para estadio 1 de aislamientos identificados como bacilos
Gram-negativos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3
B4 0.79 a
O4 0.94 a b
O13 0.94 a b
O7 0.97 a b
O23 0.97 a b
B11 0.97 a b
C3 0.97 a b
PF 0.97 a b
B8 1.00 a b
B13 1.00 a b
O32 1.02 a b
C2 1.05 a b
O11 1.05 a b
O14 1.05 a b
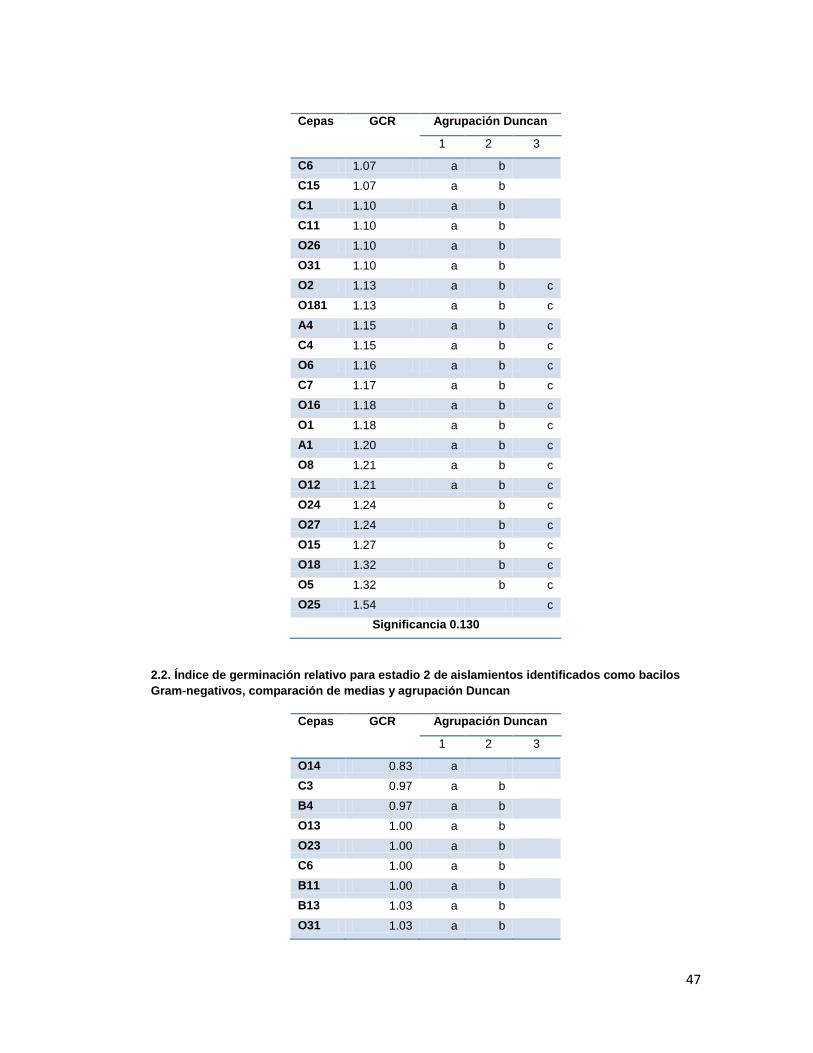
47
Cepas GCR Agrupación Duncan
1 2 3
C6 1.07 a b
C15 1.07 a b
C1 1.10 a b
C11 1.10 a b
O26 1.10 a b
O31 1.10 a b
O2 1.13 a b c
O181 1.13 a b c
A4 1.15 a b c
C4 1.15 a b c
O6 1.16 a b c
C7 1.17 a b c
O16 1.18 a b c
O1 1.18 a b c
A1 1.20 a b c
O8 1.21 a b c
O12 1.21 a b c
O24 1.24 b c
O27 1.24 b c
O15 1.27 b c
O18 1.32 b c
O5 1.32 b c
O25 1.54 c
Significancia 0.130
2.2. Índice de germinación relativo para estadio 2 de aislamientos identificados como bacilos
Gram-negativos, comparación de medias y agrupación Duncan
Cepas GCR Agrupación Duncan
1 2 3
O14 0.83 a
C3 0.97 a b
B4 0.97 a b
O13 1.00 a b
O23 1.00 a b
C6 1.00 a b
B11 1.00 a b
B13 1.03 a b
O31 1.03 a b
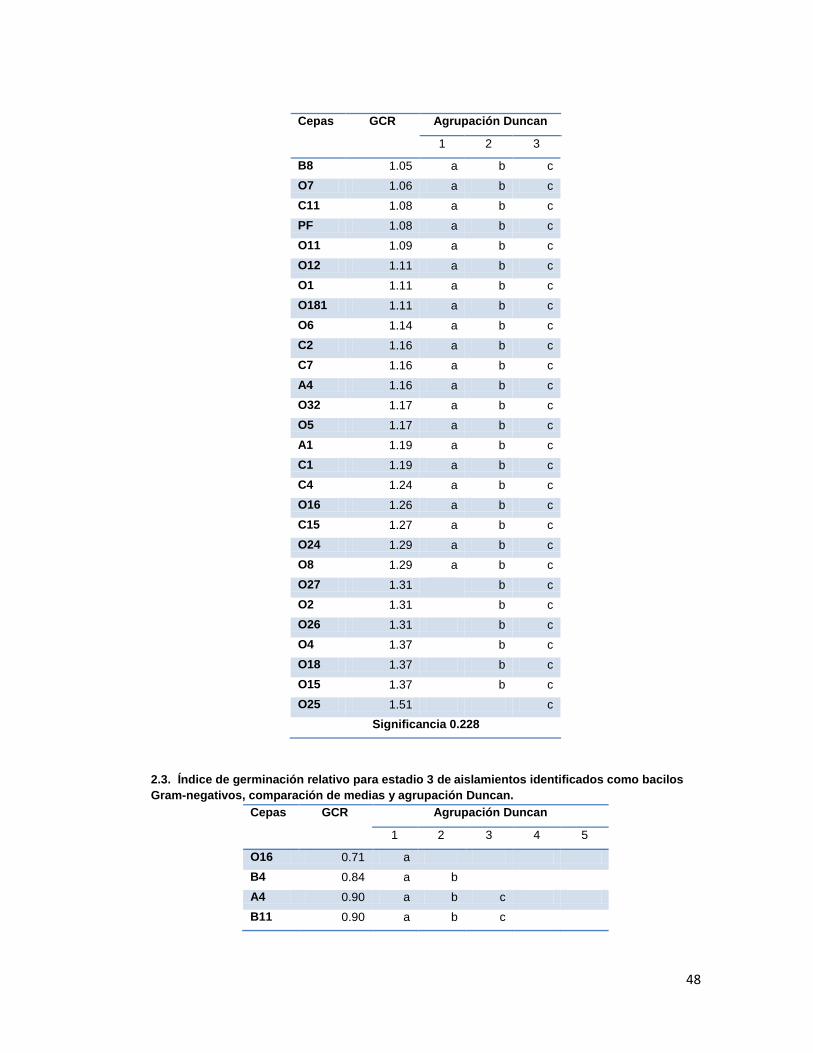
48
Cepas GCR Agrupación Duncan
1 2 3
B8 1.05 a b c
O7 1.06 a b c
C11 1.08 a b c
PF 1.08 a b c
O11 1.09 a b c
O12 1.11 a b c
O1 1.11 a b c
O181 1.11 a b c
O6 1.14 a b c
C2 1.16 a b c
C7 1.16 a b c
A4 1.16 a b c
O32 1.17 a b c
O5 1.17 a b c
A1 1.19 a b c
C1 1.19 a b c
C4 1.24 a b c
O16 1.26 a b c
C15 1.27 a b c
O24 1.29 a b c
O8 1.29 a b c
O27 1.31 b c
O2 1.31 b c
O26 1.31 b c
O4 1.37 b c
O18 1.37 b c
O15 1.37 b c
O25 1.51 c
Significancia 0.228
2.3. Índice de germinación relativo para estadio 3 de aislamientos identificados como bacilos
Gram-negativos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3 4 5
O16 0.71 a
B4 0.84 a b
A4 0.90 a b c
B11 0.90 a b c
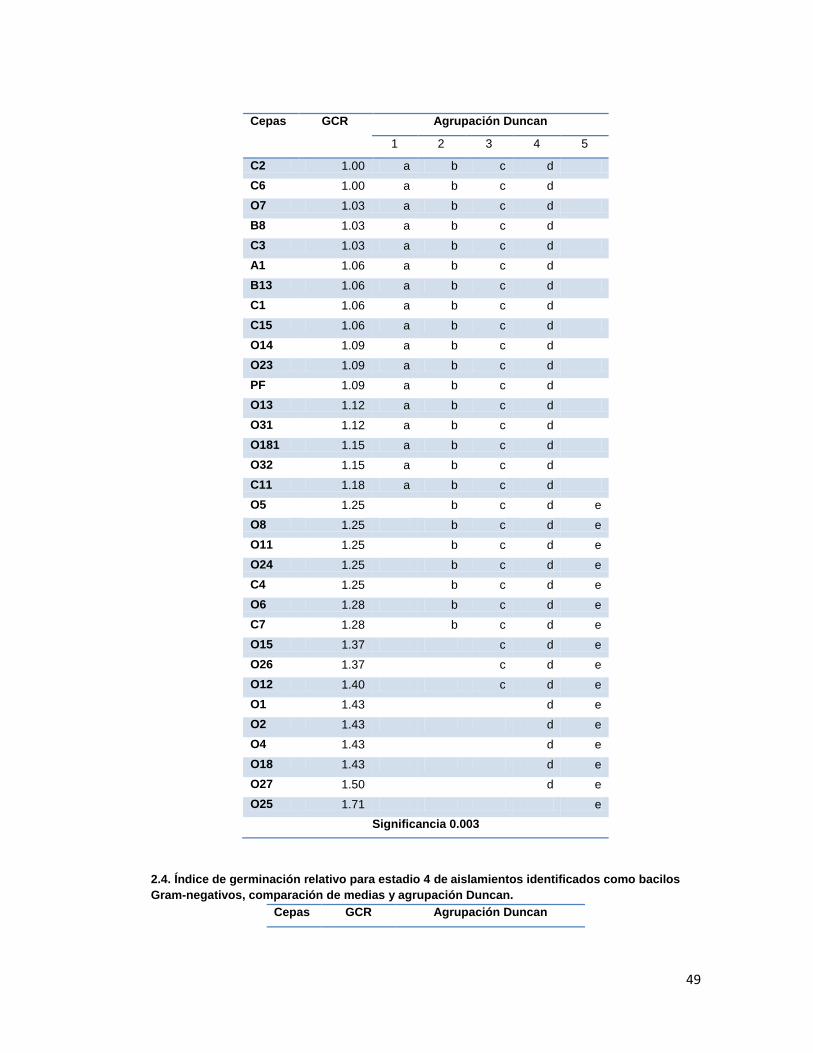
49
Cepas GCR Agrupación Duncan
1 2 3 4 5
C2 1.00 a b c d
C6 1.00 a b c d
O7 1.03 a b c d
B8 1.03 a b c d
C3 1.03 a b c d
A1 1.06 a b c d
B13 1.06 a b c d
C1 1.06 a b c d
C15 1.06 a b c d
O14 1.09 a b c d
O23 1.09 a b c d
PF 1.09 a b c d
O13 1.12 a b c d
O31 1.12 a b c d
O181 1.15 a b c d
O32 1.15 a b c d
C11 1.18 a b c d
O5 1.25 b c d e
O8 1.25 b c d e
O11 1.25 b c d e
O24 1.25 b c d e
C4 1.25 b c d e
O6 1.28 b c d e
C7 1.28 b c d e
O15 1.37 c d e
O26 1.37 c d e
O12 1.40 c d e
O1 1.43 d e
O2 1.43 d e
O4 1.43 d e
O18 1.43 d e
O27 1.50 d e
O25 1.71 e
Significancia 0.003
2.4. Índice de germinación relativo para estadio 4 de aislamientos identificados como bacilos
Gram-negativos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
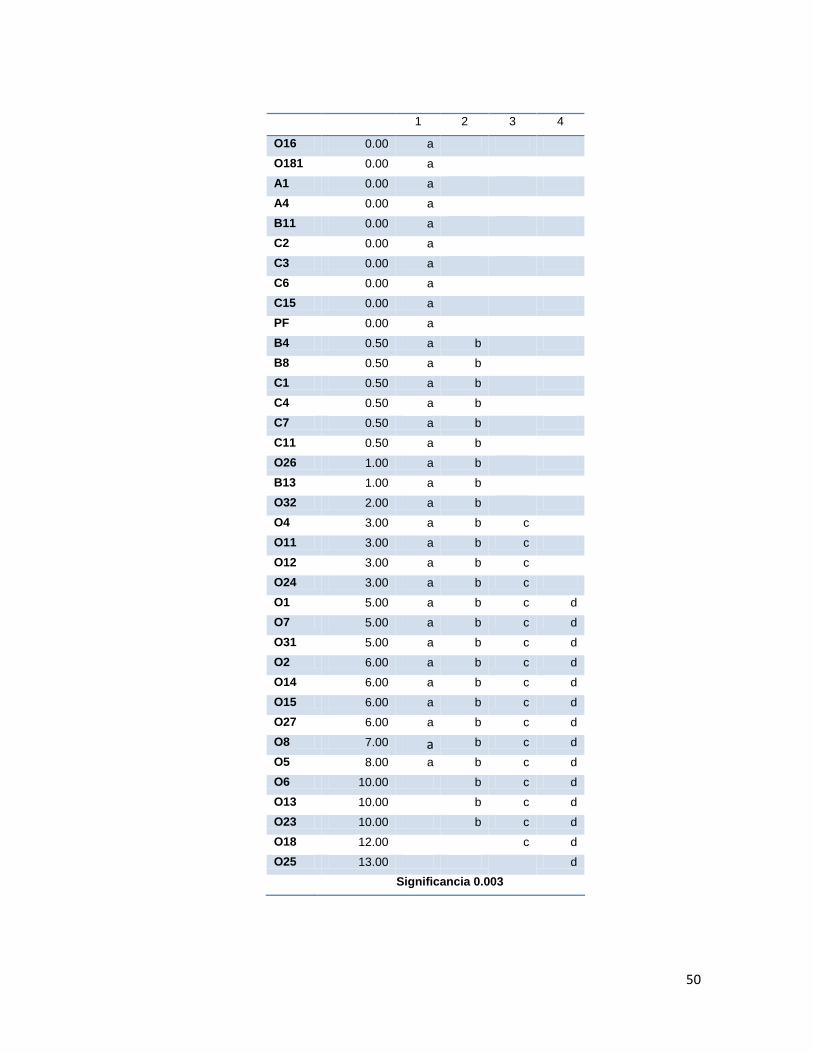
50
1 2 3 4
O16 0.00 a
O181 0.00 a
A1 0.00 a
A4 0.00 a
B11 0.00 a
C2 0.00 a
C3 0.00 a
C6 0.00 a
C15 0.00 a
PF 0.00 a
B4 0.50 a b
B8 0.50 a b
C1 0.50 a b
C4 0.50 a b
C7 0.50 a b
C11 0.50 a b
O26 1.00 a b
B13 1.00 a b
O32 2.00 a b
O4 3.00 a b c
O11 3.00 a b c
O12 3.00 a b c
O24 3.00 a b c
O1 5.00 a b c d
O7 5.00 a b c d
O31 5.00 a b c d
O2 6.00 a b c d
O14 6.00 a b c d
O15 6.00 a b c d
O27 6.00 a b c d
O8 7.00 a b c d
O5 8.00 a b c d
O6 10.00 b c d
O13 10.00 b c d
O23 10.00 b c d
O18 12.00 c d
O25 13.00 d
Significancia 0.003
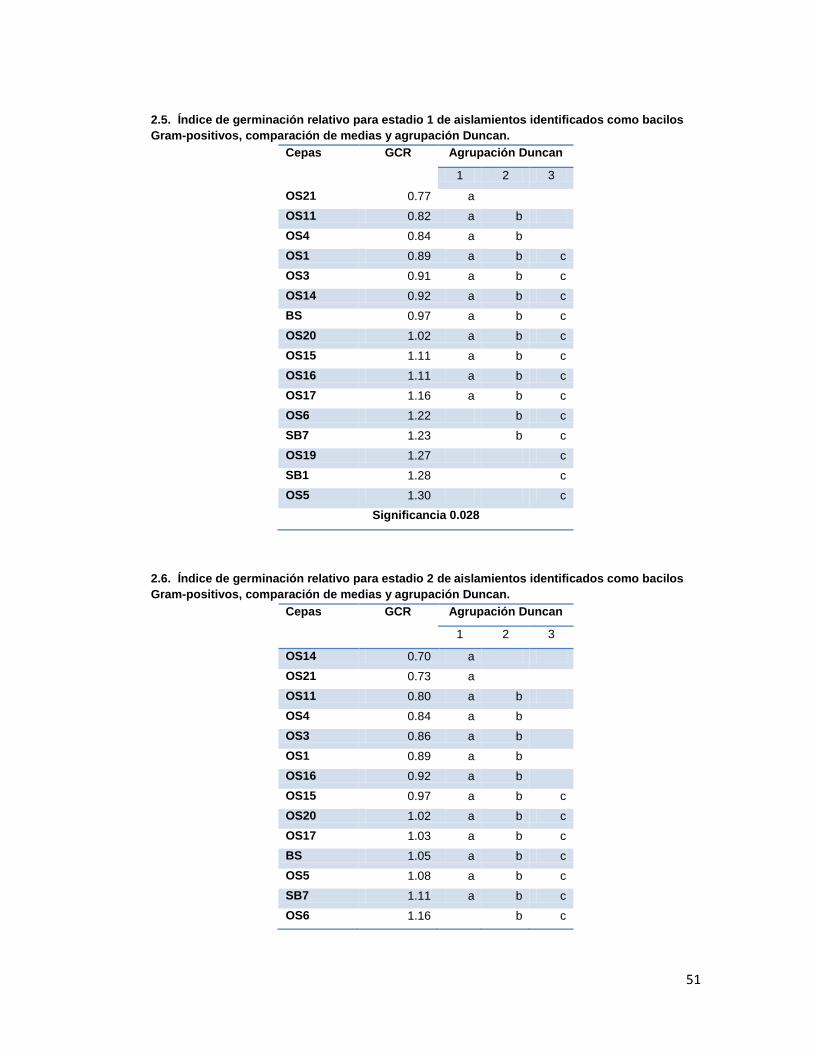
51
2.5. Índice de germinación relativo para estadio 1 de aislamientos identificados como bacilos
Gram-positivos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3
OS21 0.77 a
OS11 0.82 a b
OS4 0.84 a b
OS1 0.89 a b c
OS3 0.91 a b c
OS14 0.92 a b c
BS 0.97 a b c
OS20 1.02 a b c
OS15 1.11 a b c
OS16 1.11 a b c
OS17 1.16 a b c
OS6 1.22 b c
SB7 1.23 b c
OS19 1.27 c
SB1 1.28 c
OS5 1.30 c
Significancia 0.028
2.6. Índice de germinación relativo para estadio 2 de aislamientos identificados como bacilos
Gram-positivos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3
OS14 0.70 a
OS21 0.73 a
OS11 0.80 a b
OS4 0.84 a b
OS3 0.86 a b
OS1 0.89 a b
OS16 0.92 a b
OS15 0.97 a b c
OS20 1.02 a b c
OS17 1.03 a b c
BS 1.05 a b c
OS5 1.08 a b c
SB7 1.11 a b c
OS6 1.16 b c
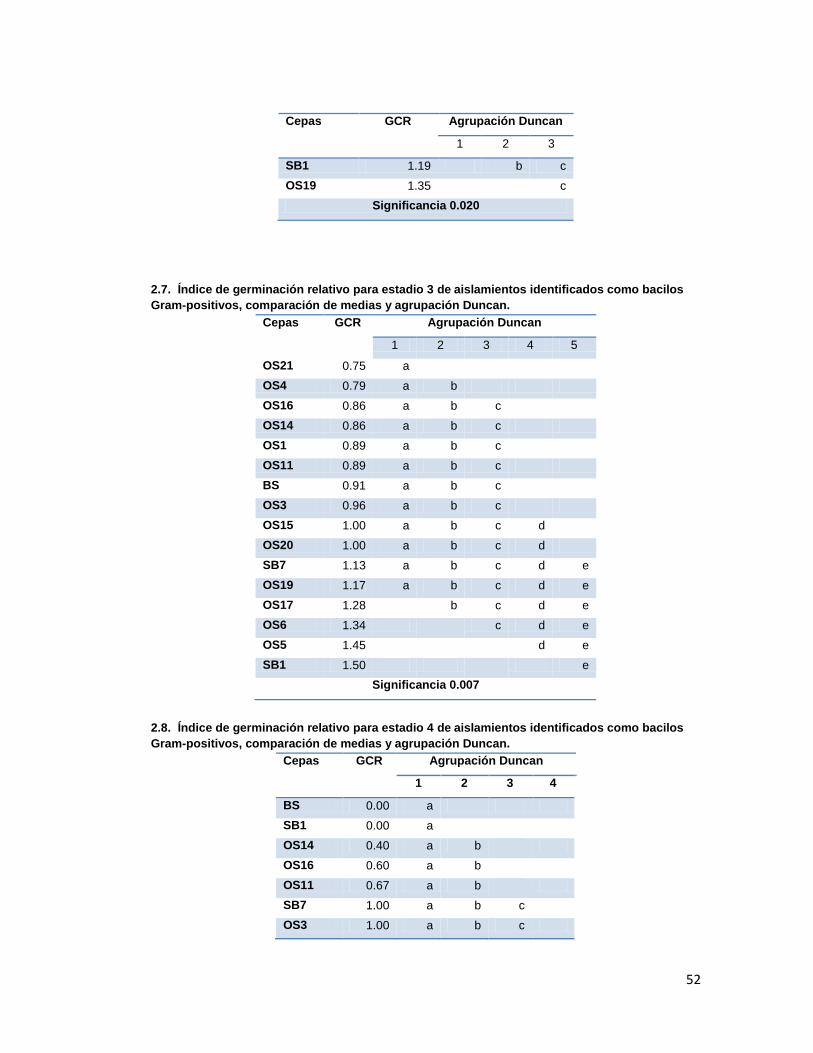
52
Cepas GCR Agrupación Duncan
1 2 3
SB1 1.19 b c
OS19 1.35 c
Significancia 0.020
2.7. Índice de germinación relativo para estadio 3 de aislamientos identificados como bacilos
Gram-positivos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3 4 5
OS21 0.75 a
OS4 0.79 a b
OS16 0.86 a b c
OS14 0.86 a b c
OS1 0.89 a b c
OS11 0.89 a b c
BS 0.91 a b c
OS3 0.96 a b c
OS15 1.00 a b c d
OS20 1.00 a b c d
SB7 1.13 a b c d e
OS19 1.17 a b c d e
OS17 1.28 b c d e
OS6 1.34 c d e
OS5 1.45 d e
SB1 1.50 e
Significancia 0.007
2.8. Índice de germinación relativo para estadio 4 de aislamientos identificados como bacilos
Gram-positivos, comparación de medias y agrupación Duncan.
Cepas GCR Agrupación Duncan
1 2 3 4
BS 0.00 a
SB1 0.00 a
OS14 0.40 a b
OS16 0.60 a b
OS11 0.67 a b
SB7 1.00 a b c
OS3 1.00 a b c
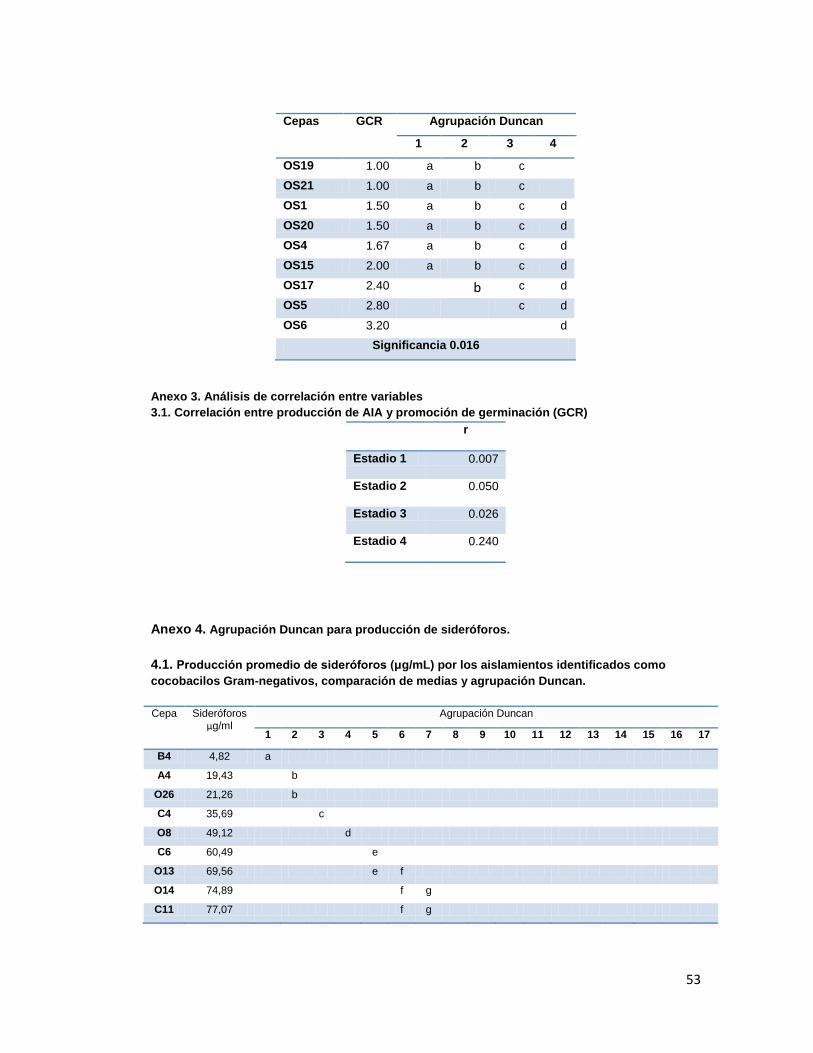
53
Cepas GCR Agrupación Duncan
1 2 3 4
OS19 1.00 a b c
OS21 1.00 a b c
OS1 1.50 a b c d
OS20 1.50 a b c d
OS4 1.67 a b c d
OS15 2.00 a b c d
OS17 2.40 b c d
OS5 2.80 c d
OS6 3.20 d
Significancia 0.016
Anexo 3. Análisis de correlación entre variables
3.1. Correlación entre producción de AIA y promoción de germinación (GCR)
r
Estadio 1 0.007
Estadio 2 0.050
Estadio 3 0.026
Estadio 4 0.240
Anexo 4. Agrupación Duncan para producción de sideróforos.
4.1. Producción promedio de sideróforos (μg/mL) por los aislamientos identificados como
cocobacilos Gram-negativos, comparación de medias y agrupación Duncan.
Cepa Sideróforos µg/ml
Agrupación Duncan
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
B4 4,82 a
A4 19,43 b
O26 21,26 b
C4 35,69 c
O8 49,12 d
C6 60,49 e
O13 69,56 e f
O14 74,89 f g
C11 77,07 f g
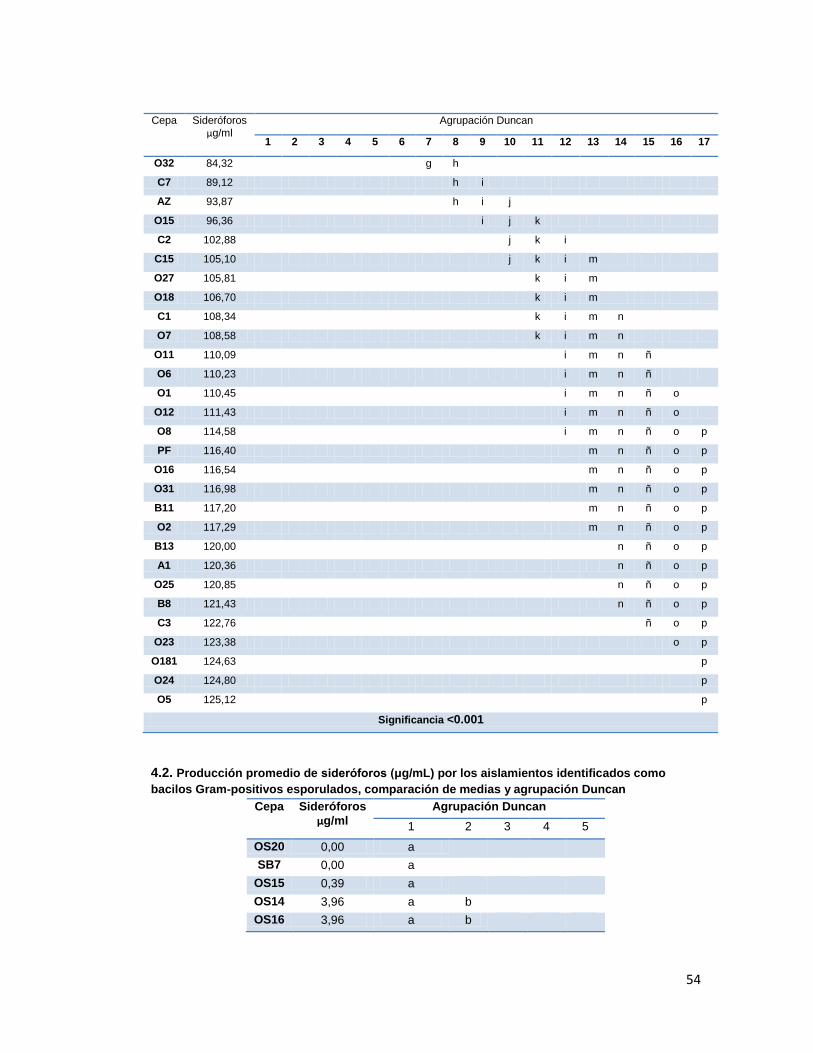
54
Cepa Sideróforos
µg/ml
Agrupación Duncan
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
O32 84,32 g h
C7 89,12 h i
AZ 93,87 h i j
O15 96,36 i j k
C2 102,88 j k i
C15 105,10 j k i m
O27 105,81 k i m
O18 106,70 k i m
C1 108,34 k i m n
O7 108,58 k i m n
O11 110,09 i m n ñ
O6 110,23 i m n ñ
O1 110,45 i m n ñ o
O12 111,43 i m n ñ o
O8 114,58 i m n ñ o p
PF 116,40 m n ñ o p
O16 116,54 m n ñ o p
O31 116,98 m n ñ o p
B11 117,20 m n ñ o p
O2 117,29 m n ñ o p
B13 120,00 n ñ o p
A1 120,36 n ñ o p
O25 120,85 n ñ o p
B8 121,43 n ñ o p
C3 122,76 ñ o p
O23 123,38 o p
O181 124,63 p
O24 124,80 p
O5 125,12 p
Significancia <0.001
4.2. Producción promedio de sideróforos (μg/mL) por los aislamientos identificados como
bacilos Gram-positivos esporulados, comparación de medias y agrupación Duncan
Cepa Sideróforos
µg/ml
Agrupación Duncan
1 2 3 4 5
OS20 0,00 a
SB7 0,00 a
OS15 0,39 a
OS14 3,96 a b
OS16 3,96 a b
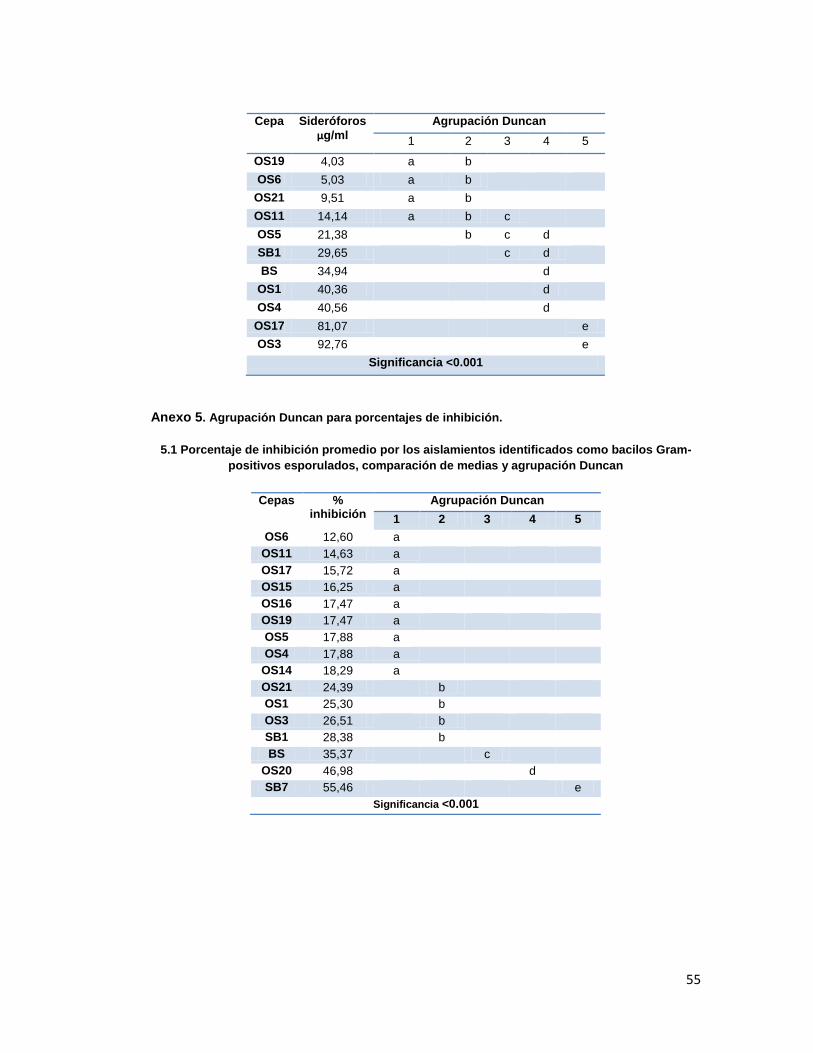
55
Cepa Sideróforos
µg/ml
Agrupación Duncan
1 2 3 4 5
OS19 4,03 a b
OS6 5,03 a b
OS21 9,51 a b
OS11 14,14 a b c
OS5 21,38 b c d
SB1 29,65 c d
BS 34,94 d
OS1 40,36 d
OS4 40,56 d
OS17 81,07 e
OS3 92,76 e
Significancia <0.001
Anexo 5. Agrupación Duncan para porcentajes de inhibición.
5.1 Porcentaje de inhibición promedio por los aislamientos identificados como bacilos Gram-
positivos esporulados, comparación de medias y agrupación Duncan
Cepas % inhibición
Agrupación Duncan
1 2 3 4 5
OS6 12,60 a
OS11 14,63 a
OS17 15,72 a
OS15 16,25 a
OS16 17,47 a
OS19 17,47 a
OS5 17,88 a
OS4 17,88 a
OS14 18,29 a
OS21 24,39 b
OS1 25,30 b
OS3 26,51 b
SB1 28,38 b
BS 35,37 c
OS20 46,98 d
SB7 55,46 e
Significancia <0.001
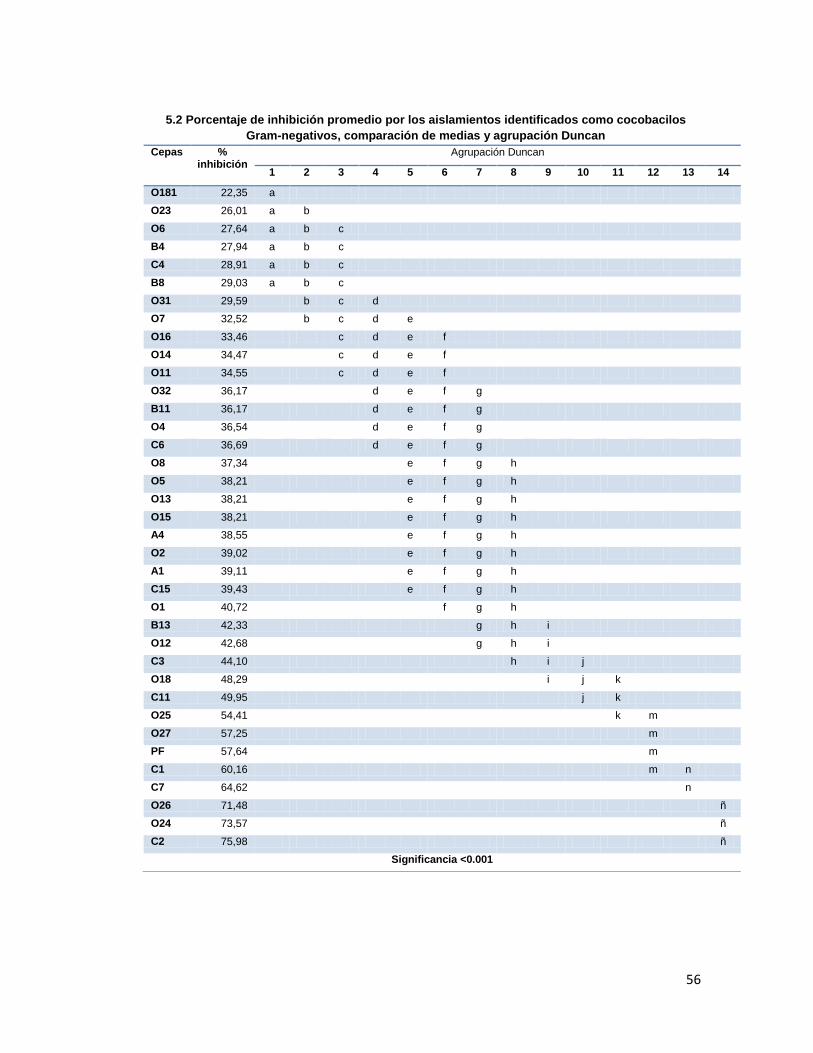
56
5.2 Porcentaje de inhibición promedio por los aislamientos identificados como cocobacilos
Gram-negativos, comparación de medias y agrupación Duncan
Cepas % inhibición
Agrupación Duncan
1 2 3 4 5 6 7 8 9 10 11 12 13 14
O181 22,35 a
O23 26,01 a b
O6 27,64 a b c
B4 27,94 a b c
C4 28,91 a b c
B8 29,03 a b c
O31 29,59 b c d
O7 32,52 b c d e
O16 33,46 c d e f
O14 34,47 c d e f
O11 34,55 c d e f
O32 36,17 d e f g
B11 36,17 d e f g
O4 36,54 d e f g
C6 36,69 d e f g
O8 37,34 e f g h
O5 38,21 e f g h
O13 38,21 e f g h
O15 38,21 e f g h
A4 38,55 e f g h
O2 39,02 e f g h
A1 39,11 e f g h
C15 39,43 e f g h
O1 40,72 f g h
B13 42,33 g h i
O12 42,68 g h i
C3 44,10 h i j
O18 48,29 i j k
C11 49,95 j k
O25 54,41 k m
O27 57,25 m
PF 57,64 m
C1 60,16 m n
C7 64,62 n
O26 71,48 ñ
O24 73,57 ñ
C2 75,98 ñ
Significancia <0.001
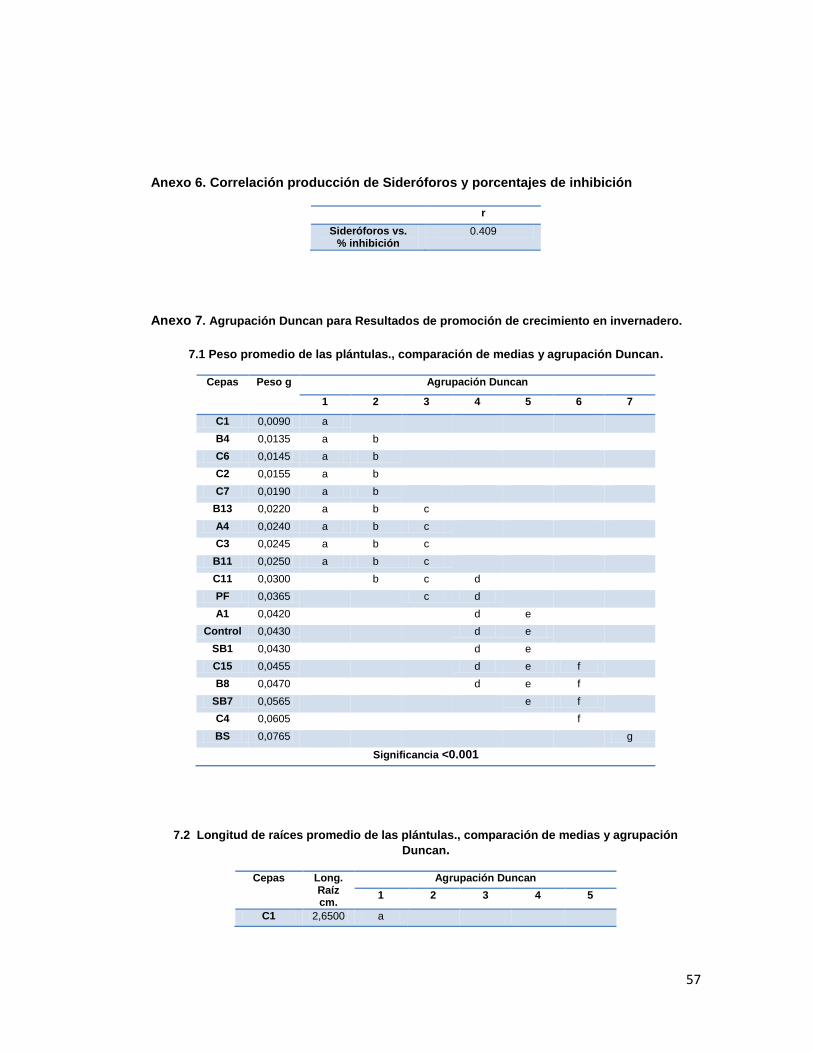
57
Anexo 6. Correlación producción de Sideróforos y porcentajes de inhibición
r
Sideróforos vs. % inhibición
0.409
Anexo 7. Agrupación Duncan para Resultados de promoción de crecimiento en invernadero.
7.1 Peso promedio de las plántulas., comparación de medias y agrupación Duncan.
Cepas Peso g Agrupación Duncan
1 2 3 4 5 6 7
C1 0,0090 a
B4 0,0135 a b
C6 0,0145 a b
C2 0,0155 a b
C7 0,0190 a b
B13 0,0220 a b c
A4 0,0240 a b c
C3 0,0245 a b c
B11 0,0250 a b c
C11 0,0300 b c d
PF 0,0365 c d
A1 0,0420 d e
Control 0,0430 d e
SB1 0,0430 d e
C15 0,0455 d e f
B8 0,0470 d e f
SB7 0,0565 e f
C4 0,0605 f
BS 0,0765 g
Significancia <0.001
7.2 Longitud de raíces promedio de las plántulas., comparación de medias y agrupación
Duncan.
Cepas Long. Raíz cm.
Agrupación Duncan
1 2 3 4 5
C1 2,6500 a
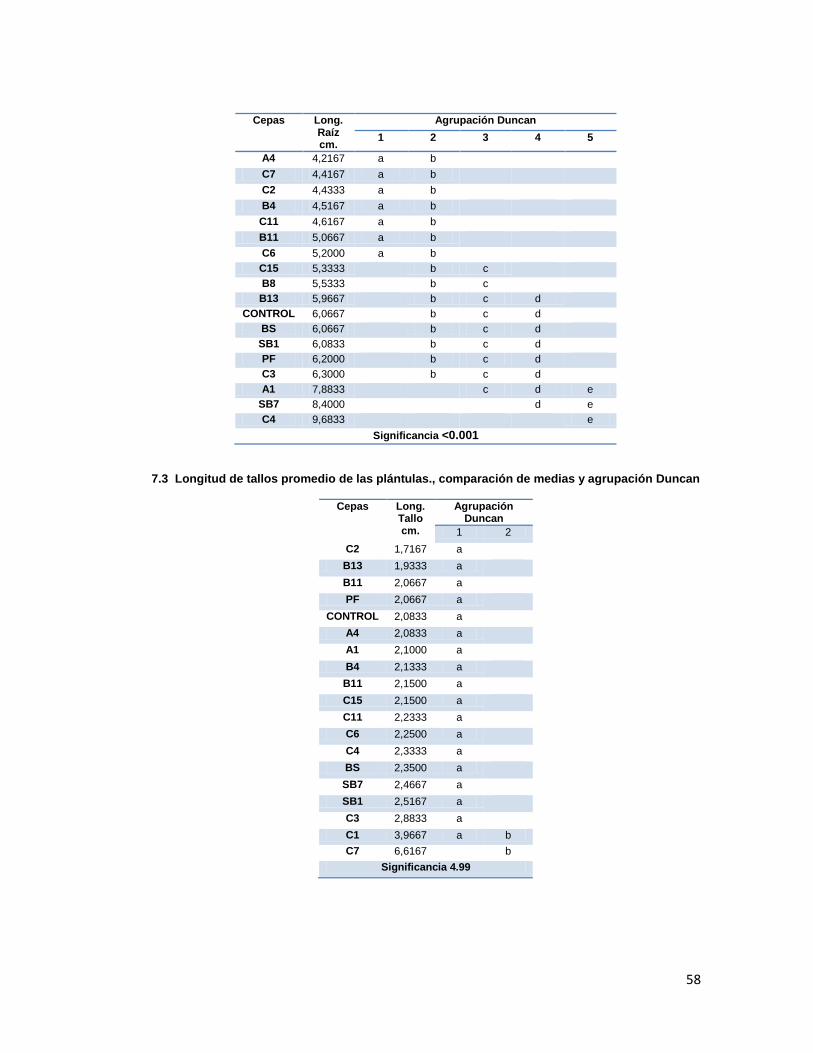
58
Cepas Long. Raíz cm.
Agrupación Duncan
1 2 3 4 5
A4 4,2167 a b
C7 4,4167 a b
C2 4,4333 a b
B4 4,5167 a b
C11 4,6167 a b
B11 5,0667 a b
C6 5,2000 a b
C15 5,3333 b c
B8 5,5333 b c
B13 5,9667 b c d
CONTROL 6,0667 b c d
BS 6,0667 b c d
SB1 6,0833 b c d
PF 6,2000 b c d
C3 6,3000 b c d
A1 7,8833 c d e
SB7 8,4000 d e
C4 9,6833 e
Significancia <0.001
7.3 Longitud de tallos promedio de las plántulas., comparación de medias y agrupación Duncan
Cepas Long. Tallo cm.
Agrupación Duncan
1 2
C2 1,7167 a
B13 1,9333 a
B11 2,0667 a
PF 2,0667 a
CONTROL 2,0833 a
A4 2,0833 a
A1 2,1000 a
B4 2,1333 a
B11 2,1500 a
C15 2,1500 a
C11 2,2333 a
C6 2,2500 a
C4 2,3333 a
BS 2,3500 a
SB7 2,4667 a
SB1 2,5167 a
C3 2,8833 a
C1 3,9667 a b
C7 6,6167 b
Significancia 4.99
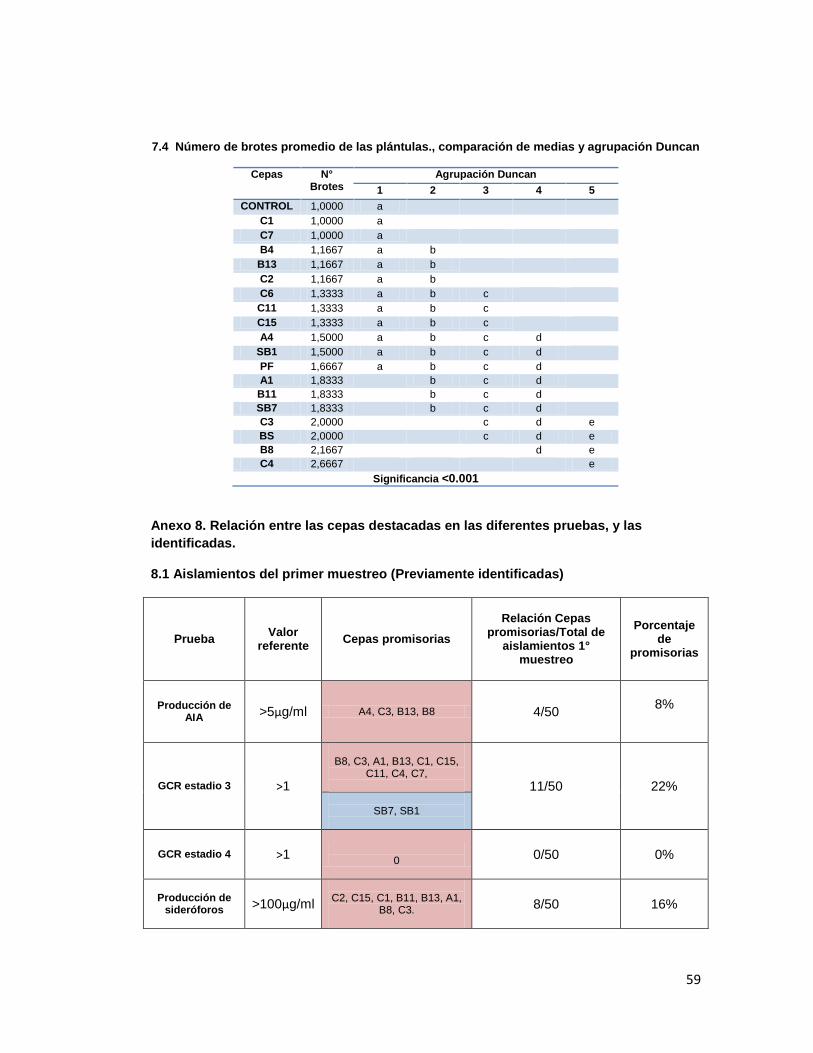
59
7.4 Número de brotes promedio de las plántulas., comparación de medias y agrupación Duncan
Cepas N° Brotes
Agrupación Duncan
1 2 3 4 5
CONTROL 1,0000 a
C1 1,0000 a
C7 1,0000 a
B4 1,1667 a b
B13 1,1667 a b
C2 1,1667 a b
C6 1,3333 a b c
C11 1,3333 a b c
C15 1,3333 a b c
A4 1,5000 a b c d
SB1 1,5000 a b c d
PF 1,6667 a b c d
A1 1,8333 b c d
B11 1,8333 b c d
SB7 1,8333 b c d
C3 2,0000 c d e
BS 2,0000 c d e
B8 2,1667 d e
C4 2,6667 e
Significancia <0.001
Anexo 8. Relación entre las cepas destacadas en las diferentes pruebas, y las
identificadas.
8.1 Aislamientos del primer muestreo (Previamente identificadas)
Prueba Valor
referente Cepas promisorias
Relación Cepas
promisorias/Total de aislamientos 1°
muestreo
Porcentaje de
promisorias
Producción de AIA >5μg/ml A4, C3, B13, B8
4/50
8%
GCR estadio 3 >1
B8, C3, A1, B13, C1, C15,
C11, C4, C7, 11/50 22%
SB7, SB1
GCR estadio 4 >1
0 0/50 0%
Producción de sideróforos >100μg/ml
C2, C15, C1, B11, B13, A1,
B8, C3.
8/50 16%
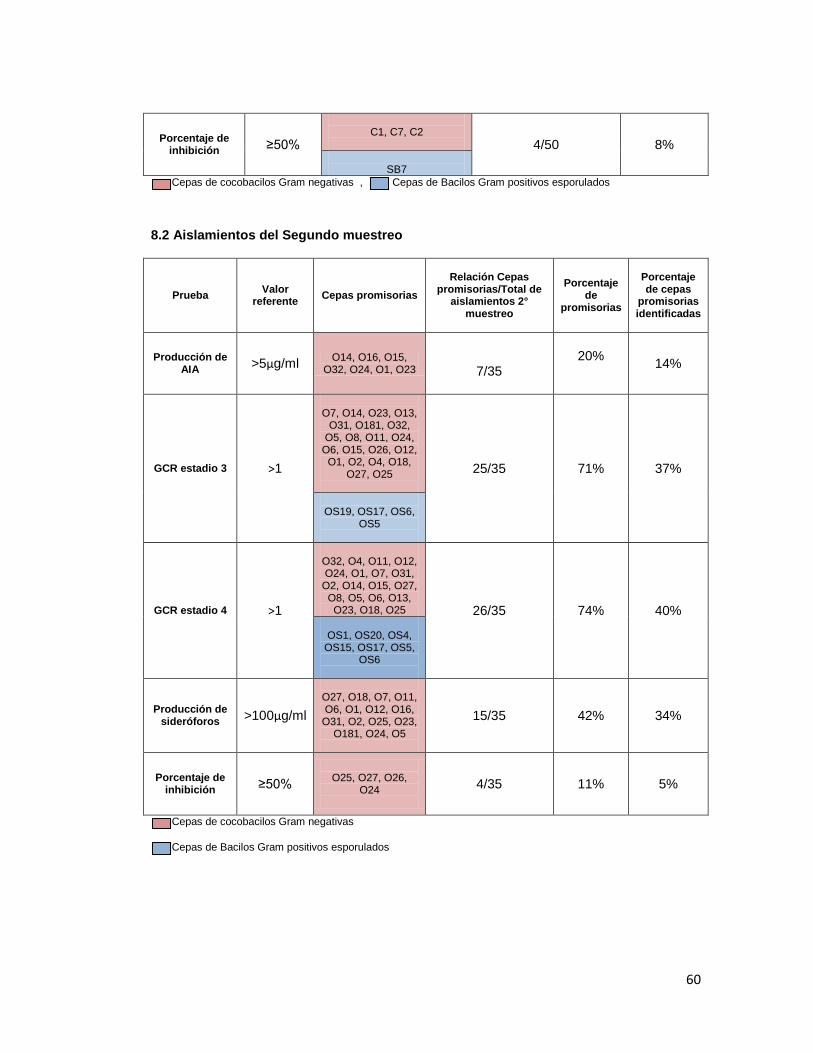
60
Porcentaje de inhibición ≥50%
C1, C7, C2
4/50 8%
SB7
Cepas de cocobacilos Gram negativas , Cepas de Bacilos Gram positivos esporulados
8.2 Aislamientos del Segundo muestreo
Prueba Valor
referente Cepas promisorias
Relación Cepas
promisorias/Total de aislamientos 2°
muestreo
Porcentaje de
promisorias
Porcentaje de cepas
promisorias identificadas
Producción de AIA
>5μg/ml O14, O16, O15,
O32, O24, O1, O23
7/35
20%
14%
GCR estadio 3 >1
O7, O14, O23, O13,
O31, O181, O32, O5, O8, O11, O24,
O6, O15, O26, O12, O1, O2, O4, O18,
O27, O25
25/35 71% 37%
OS19, OS17, OS6,
OS5
GCR estadio 4 >1
O32, O4, O11, O12, O24, O1, O7, O31,
O2, O14, O15, O27, O8, O5, O6, O13, O23, O18, O25 26/35 74% 40%
OS1, OS20, OS4,
OS15, OS17, OS5, OS6
Producción de sideróforos
>100μg/ml
O27, O18, O7, O11, O6, O1, O12, O16,
O31, O2, O25, O23, O181, O24, O5
15/35 42% 34%
Porcentaje de inhibición ≥50%
O25, O27, O26,
O24
4/35 11% 5%
Cepas de cocobacilos Gram negativas
Cepas de Bacilos Gram positivos esporulados

61





























