Embed Size (px)
Citation preview

Programa de Pós-Graduação Stricto Sensu Mestrado Profissional em Ciência e Tecnologia de Alimentos
Campus Rio de Janeiro
Angelo Máximo Batista de Amorim
AVALIAÇÃO DA MULTIRRESISTÊNCIA A DROGAS E DA PRODUÇÃO DE OUTROS
FATORES DE VIRULÊNCIA POR ISOLADOS DE ENTEROBACTERIACEAE
PROVENIENTES DE PRODUTOS LÁCTEOS
RIO DE JANEIRO – RJ
2016

Angelo Máximo Batista de Amorim
AVALIAÇÃO DA MULTIRRESISTÊNCIA A DROGAS E DA PRODUÇÃO DE OUTROS
FATORES DE VIRULÊNCIA POR ISOLADOS DE ENTEROBACTERIACEAE
PROVENIENTES DE PRODUTOS LÁCTEOS
Dissertação apresentada como parte dos requisitos necessários para a obtenção do título de Mestre em Ciência e Tecnologia de Alimentos, no Programa de Pós-Graduação Stricto Sensu Mestrado Profissional em Ciência e Tecnologia de Alimentos, do Instituto Federal de Educação, Ciência e Tecnologia (IFRJ).
Orientadora: Profª. Dr.ª Janaína dos Santos Nascimento
RIO DE JANEIRO – RJ
2016

.

Angelo Máximo Batista de Amorim
Avaliação da multirresistência a drogas e da produção de outros fatores de virulência
por isolados de Enterobacteriaceae provenientes de produtos lácteos
Dissertação apresentada como parte dos
requisitos necessários para a obtenção do
título de Mestre em Ciência e Tecnologia de
Alimentos, no Programa de Pós-Graduação
Stricto Sensu Mestrado Profissional em
Ciência e Tecnologia de Alimentos, do
Instituto Federal de Educação, Ciência e
Tecnologia (IFRJ).
Data de aprovação: _____/_____/_____
________________________________________________________________
Prof. Dra. Janaína dos Santos Nascimento
Instituto Federal de Educação, Ciência e Tecnologia do Rio de Janeiro
________________________________________________________________
Prof. Dr. Bárbara Cristina Euzébio Pereira Dias de Oliveira
Instituto Federal de Educação, Ciência e Tecnologia do Rio de Janeiro
________________________________________________________________
Prof. Dra. Jéssica Manya Bittencourt Dias Vieira
Fundação Centro Universitário Estadual da Zona Oeste
Rio de Janeiro – RJ
2016

Dedico esta dissertação à minha esposa Luciane Carvalho dos Santos de Amorim, às minhas filhas Mylena dos Santos de Amorim e Alice dos Santos de Amorim, e, à minha mãe, Jandira Batista da Nóbrega, como um pedido de desculpas pelos momentos ausentes.

AGRADECIMENTOS
Agradeço, primeiramente, a Deus pela oportunidade de transformar este sonho em
realidade.
Agradeço a todos os meus guias espirituais.
Agradeço à Gláucia Motta da Silva Passos Ferreira, pessoa que me apresentou ao
curso e me incentivou a participar do processo seletivo, diante de uma barreira encontrada.
Agradeço à Luciane Martins Medeiros, primeira pessoa a me dar as diretrizes do
projeto de pesquisa e de compartilhar comigo a sua experiência na área de alimentos e
microbiologia.
Agradeço ao pessoal da SEPIN, especialmente à Adriana Marques Frazão, Luciana
Veloso da Costa, Josiane Machado Vieira Mattoso e Vania Maria da Silva Fernandes.
Agradeço ao pessoal da SEMEC, minha antiga seção de trabalho, pela parceria de
sempre.
Agradeço à minha gerente de divisão Claudia Maria Dias.
Agradeço à Érica Louro da Fonseca pelas aulas de biologia molecular.
Agradeço ao meu colega do INCQS Rafael Lawson Ferreira.
Agradeço aos meus colegas de seção (SEPRM).
Agradeço ao meu grupo de trabalho do laboratório de microbiologia do IFRJ:
Brendon Chaves Araújo, Douglas Henrique de Melo, Bárbara Victor Souza, Érica Yang,
assim como as professoras Thaís Souza Silveira Majerowicz, Zilma das Graças Nunes e
Aline dos Santos Garcia Gomes, e, especialmente a professora Bárbara Cristina Euzébio
Pereira Dias de Oliveira, pelas contribuições ao trabalho após a sua participação na banca
de qualificação.
Agradeço aos colegas da turma do mestrado 2015.
Agradeço à coordenadora do mestrado, Márcia Cristina da Silva e a todos os demais
professores do curso.
Um agradecimento especial à Maria Teresa Dias Ferreira, colega de trabalho, amiga
pessoal, conselheira espiritual e pessoa que faz parte da minha vida desde 2004, que
sempre me incentivou e continua incentivando, sem medir esforços.
E, por último, não tenho como deixar de agradecer a esta pessoa tão especial.
Aquela que, até quando os resultados saíram diferentes do esperado, sorriu, confortou-me
dizendo que foi a melhor que podia acontecer, e, sugeriu-me um novo rumo ao trabalho:
Janaína dos Santos Nascimento. Agora entendi porque seus alunos não te deixam em paz!
A todos vocês, meu muito obrigado.

“A verdadeira medida de um homem não se vê na forma como se comporta em momentos de conforto e conveniência, mas em como se mantém em tempos de controvérsia e desafio”.
Martin Luther King

i
AMORIM, A. M. B. Avaliação da multirresistência a drogas e da produção de outros fatores
de virulência por isolados de Enterobacteriaceae provenientes de produtos lácteos.
Dissertação apresentada como parte dos requisitos necessários para a obtenção do
título de Mestre em Ciência e Tecnologia de Alimentos. Programa de Pós-Graduação
Profissional em Ciência e Tecnologia de Alimentos, Instituto Federal de Educação,
Ciência e Tecnologia do Rio de Janeiro (IFRJ), Campus Rio de Janeiro, Rio de Janeiro,
RJ, 2016.
RESUMO
Os produtos lácteos são consumidos mundialmente por indivíduos de todas as classes
sociais. Diante das condições de fabricação, armazenamento, comercialização e consumo,
estes produtos podem carrear diversas bactérias, como alguns membros da família
Enterobacteriaceae que, apesar de possuir representantes com elevado potencial
patogênico, não costumam causar complicações em um indivíduo imunocompetente.
Todavia o número de indivíduos com debilidade no sistema imunológico sobe
continuamente. Representantes da família Enterobacteriaceae podem expressar fenótipos
de multirresistência a drogas (MDR) e, eventualmente, estão associados a infecções e
surtos. Na área de Alimentos, e, mais especificamente em produtos lácteos, os poucos
trabalhos envolvendo bactérias patogênicas da família Enterobacteriaceae estão
relacionados à Escherichia coli e à Salmonella spp. Logo, o objetivo deste trabalho foi
investigar a prevalência de bactérias da família Enterobacteriaceae, diferentes de
Escherichia coli e Salmonella spp., provenientes de amostras de produtos lácteos
comercializados no Rio de Janeiro e caracterizar os isolados quanto à expressão de
multirresistência a drogas e de outros fatores de virulência. A partir de 10 amostras de
conveniência de produtos lácteos, foram obtidos 58 isolados, que foram identificados pelo
sistema VITEK 2. Os resultados do antibiograma revelaram que vinte e quatro isolados
expressaram multirresistência, uma vez que foram resistentes a pelo menos um antibiótico
de três classes distintas. Os isolados MDR foram avaliados quanto à expressão dos
fenótipos de susceptibilidade reduzida aos carbapenens (KPC) e de produção de β-
lactamase de espectro estendido (ESBL) em meios de cultura cromogênicos (CHROMagar®
KPC e CHROMagar® ESBL). Seis deles expressaram fenótipo KPC e 14 expressaram
fenótipos ESBL. A produção qualitativa de biofilme foi avaliada em ágar vermelho Congo e
cinquenta e quatro isolados mostraram ser produtores. Quanto aos demais fatores de
virulência, dezesseis isolados expressaram atividade proteolítica e vinte isolados mostraram
ser lipolíticos. Quarenta e dois isolados apresentaram algum tipo de atividade hemolítica. A
produção de substâncias antimicrobianas (SAM) também foi investigada, porém nenhum
dos isolados produziu SAM contra as estirpes utilizadas como indicadoras. O perfil

ii
plasmideal dos isolados MDR e não MDR do complexo Enterobacter cloacae revelou que
não foi possível identificar um padrão que distinguisse os dois grupos, baseado apenas em
número e tamanho das formas plasmidiais. Este trabalho sugere que alguns produtos
lácteos podem ser veículos de bactérias MDR da família Enterobacteriaceae, algumas
carreadoras de vários fatores de virulência, constituindo uma grave ameaça para a indústria
de laticínios e para os seus consumidores, especialmente aqueles pertencentes a grupos de
risco, como os imunodeprimidos.
Palavras-chave: Enterobacteriaceae. Produtos lácteos. Multirresistência. Fatores de virulência

iii
AMORIM, A. M. B. Avaliação da multirresistência a drogas e da produção de outros
fatores de virulência por isolados de Enterobacteriaceae provenientes de produtos
lácteos. Dissertação apresentada como parte dos requisitos necessários para a obtenção
do título de Mestre em Ciência e Tecnologia de Alimentos. Programa de Pós-Graduação
Profissional em Ciência e Tecnologia de Alimentos, Instituto Federal de Educação, Ciência e
Tecnologia do Rio de Janeiro (IFRJ), Campus Rio de Janeiro, Rio de Janeiro, RJ, 2016.
ABSTRACT
Dairy products are consumed worldwide by individual belonging to all social classes. Under
different manufacturing conditions, storage, commercialization and consumption, these
products may carry various bacteria, as some members of the Enterobacteriaceae family
that, despite having representatives with high pathogenic potential, they do not usually cause
complications in an immunocompetent individual. However, the number of individuals with a
weak immune system rises continuously. Representatives of the family Enterobacteriaceae
can express multidrug resistance phenotypes (MDR) and, possibly, are associated with
outbreaks and infections. In the Food area, and more specifically, in dairy products, the few
studies involving pathogenic bacteria of the Enterobacteriaceae family are related to
Escherichia coli and Salmonella spp. Therefore, the aim of this study was to investigate the
prevalence of bacteria of the Enterobacteriaceae family, non-Escherichia coli and non-
Salmonella spp., from dairy products commercialized in Rio de Janeiro and characterize
isolates with respect to multidrug resistance and other virulence factors. From 10
convenience samples of dairy products were obtained 58 isolates which were identified by
the VITEK 2 system. Results of antibiogram revealed that twenty-four isolates expressed a
MDR phenotype, since they were resistant to antibiotics belonging to at least 3 different
classes. MDR isolates were evaluated for expression of reduced susceptibility to
carbapenems (KPC) and extended spectrum β-lactamase (ESBL) in CHROMogenic culture
media (CHROMagar® KPC and CHROMagar® ESBL). Six of them expressed KPC+
phenotype and 14 expressed ESBL+ phenotypes. Qualitative biofilm production was
evaluated in Congo red agar and fifty-four isolates proved to be producers. Regarding the
other virulence factors, sixteen isolates expressed proteolytic activity and twenty isolates
were lipolytic. Forty-two isolates showed some kind of hemolytic activity. The production of
antimicrobial substances (AMS) has also been investigated, but none of the isolates
produced AMS against the strains used as indicators. The plasmid profile of isolated MDR
and non-MDR from Enterobacter cloacae complex revealed that was not possible to identify
a pattern that distinguished the two groups, based only on number and size of plasmids. This
study suggests that some dairy products can be vehicles of MDR bacteria from

iv
Enterobacteriaceae family, some of them carriers of several virulence factors, constituting a
serious threat to dairy industry and its consumers, especially those belonging to risk groups,
such as immunocompromised.
Keywords: Enterobacteriaceae. Dairy products. Multidrug resistance. Virulence Factors

v
LISTA DE ILUSTRAÇÕES
Figura 1 Etapas da formação do biofilme bacteriano 8
Figura 2 Número de isolados da família Enterobacteriaceae obtidos no presente
estudo 23
Figura 3 Número de isolados produtores de biofilme 30
Figura 4 Exemplo de isolado produtor em biofilme em ágar vermelho congo 30
Figura 5 Exemplos de isolados que apresentaram atividade lipolítica 34
Figura 6 Exemplos de isolados que apresentaram atividade hemolítica 36
Figura 7 Eletroforese em gel de agarose com alguns isolados MDR e não MDR do
complexo Enterobacter cloacae 37

vi
LISTA DE TABELAS
Tabela 1 Antibióticos utilizados no presente estudo 17
Tabela 2 Interpretação dos resultados nos meios cromogênicos CHROMagar® KPC e CHROMagar® ESBL para os representantes da família Enterobacteriaceae 18
Tabela 3 Bactérias utilizadas como indicadoras da produção de substâncias antimicrobianas 20
Tabela 4 Isolados da família Enterobacteriaceae obtidos no presente estudo 21
Tabela 5 Perfil de resistência aos antimicrobianos 25
Tabela 6 Resultado dos testes com os meios cromogênicos CHROMagar® KPC e CHROMagar® ESBL 28
Tabela 7 Isolados que expressaram atividade proteolítica 31
Tabela 8 Isolados que expressaram atividade lipolítica 33
Tabela 9 Isolados que expressaram atividade hemolítica 35

vii
LISTA DE ABREVIATURAS E SIGLAS
AMC Amoxicilina + ácido clavulânico
AMI Amicacina
AMP Ampicilina
ATCC American Type Culture Collection (Coleção norte-americana de micro-
organismos)
ATM Aztreonam
CAZ Ceftazidima
CEP Cefalotina
CHL Cloranfenicol
CIP Ciprofloxacina
CLSI Clinical and Laboratory Standards Institute (Instituto de Padrões Clínicos e
Laboratoriais)
CTX Cefotaxima
DNA Deoxyribonucleic Acid (Ácido Desoxirribonucleico)
DTA Doença Transmitida por Alimento
EDTA Ácido Etilenodiamino Tetra-acético
EMB Eosina Azul de Metileno
ESBL β-lactamase de espectro estendido
GEN Gentamicina
IDSA Infectious Diseases Society of America (Sociedade de Doenças Infecciosas da
América)
IFRJ Instituto Federal de Educação, Ciência e Tecnologia do Rio de Janeiro
IPM Imipenem
HIV Human Immunodeficiency Virus (vírus da imunodeficiência humana)
KPC Sensibilidade reduzida aos carbapenens
LMIFRJ Laboratório de Microbiologia do Instituto Federal de Educação, Ciência e
Tecnologia do Rio de Janeiro.
LPS Lipopolissacarídeo
MDR Multidrug resistant (multirresistente a drogas)

viii
MER Meropenem
MRSA Methicillin-resistant Staphylococcus aureus (Staphylococcus aureus resistente
à meticilina)
NOR Norfloxacina
pH Potencial de Hidrogênio
RDC Resolução da Diretoria Colegiada
SAM Substância(s) antimicrobiana(s)
STR Estreptomicina
TBE Tris/Borato/EDTA
TOB Tobramicina
TRI Trimetoprim
TET Tetraciclina
UFC Unidade(s) formadora(s) de colônia
UHT Ultra High Temperature (Temperatura ultraelevada)

ix
SUMÁRIO
1 INTRODUÇÃO 1
2 REFERENCIAL TEÓRICO 3
2.1 PRODUTOS LÁCTEOS 3
2.1.1 Contaminação microbiológica de produtos lácteos 4
2.2 A FAMÍLIA ENTEROBACTERIACEAE 5
2.3 RESISTÊNCIA AOS ANTIMICROBIANOS 6
2.4 PRODUÇÃO DE FATORES DE VIRULÊNCIA 8
2.4.1 Produção de biofilme 8
2.4.2 Atividade proteolítica 10
2.4.3 Atividade lipolítica 10
2.4.4 Atividade hemolítica 11
2.5 PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM) 12
3 JUSTIFICATIVA 13
4 OBJETIVOS 14
4.1 OBJETIVO GERAL 14
4.2 OBJETIVOS ESPECÍFICOS 14
5 MATERIAIS E MÉTODOS 15
5.1 OBTENÇÃO DAS AMOSTRAS DE PRODUTOS LÁCTEOS 15
5.2 ISOLAMENTO DOS MICRO-ORGANISMOS 15
5.3 MANUTENÇÃO DOS ISOLADOS 15
5.4 IDENTIFICAÇÃO DOS ISOLADOS 16
5.5 PERFIL DE RESISTÊNCIA A ANTIMICROBIANOS 16
5.5.1 Detecção dos fenótipos de ESBL e KPC 17
5.6 AVALIAÇÃO DA PRODUÇÃO DE FATORES DE VIRULÊNCIA 18
5.6.1 Detecção qualitativa da produção de biofilme 18
5.6.2 Detecção de atividade proteolítica 18
5.6.3 Detecção de atividade lipolítica 19
5.6.4 Detecção de atividade hemolítica 19

x
5.7 AVALIAÇÃO DA PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM) 19
5.8 DETERMINAÇÃO E COMPARAÇÃO DO PERFIL PLASMIDEAL DOS ISOLADOS DO COMPLEXO ENTEROBACTER CLOACAE 20
6 RESULTADOS E DISCUSSÃO 21
6.1 PERFIL DE RESISTÊNCIA AOS ANTIMICROBIANOS 24
6.1.1 Detecção dos fenótipos de ESBL e KPC 27
6.2 AVALIAÇÃO DA PRODUÇÃO DE FATORES DE VIRULÊNCIA 29
6.2.1 Detecção qualitativa da produção de biofilme 29
6.2.2 Detecção de atividade proteolítica 31
6.2.3 Detecção de atividade lipolítica 33
6.2.4 Detecção de atividade hemolítica 34
6.3 AVALIAÇÃO DA PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM) 36
6.4 DETERMINAÇÃO DO PERFIL PLASMIDEAL DOS ISOLADOS DO COMPLEXO ENTEROBACTER CLOACAE 36
7 CONCLUSÕES 39
8 CONSIDERAÇÕES FINAIS 41
9 PRODUÇÃO CIENTÍFICA 42
9.1 ARTIGO ACEITO PARA PUBLICAÇÂO (ANEXO 1) 42
9.2 SUBMISSÃO DE ARTIGO: INTERNATIONAL FOOD RESEARCH JOURNAL
(ANEXO 2) 42
9.3 MANUSCRITO EM PREPARAÇÂO 42
9.4 COMUNICAÇÔES EM EVENTOS (ANEXOS 3 E 4) 42
10 REFERÊNCIAS BIBLIOGRÁFICAS 44
11 ANEXOS 59

1
1 INTRODUÇÃO
Os produtos lácteos são consumidos mundialmente por indivíduos de todas as
classes sociais, e geralmente, in natura, ou seja, sem qualquer processamento prévio até a
sua ingestão (DAMACENO et al., 2015; CASALINUOVO et al., 2014), o que exige destes
produtos uma qualidade microbiológica satisfatória. Apesar deste consumo abrangente, em
muitos locais, a produção, o armazenamento e a comercialização destes produtos ainda são
carentes de Boas Práticas de Fabricação e Higiene, podendo torná-los potenciais veículos
de transmissão de micro-organismos, muitos deles com elevado potencial patogênico, como
alguns representantes da família Enterobacteriaceae (LAGRANGE et al., 2015; CAINE et al.,
2014).
Os micro-organismos da família Enterobacteriaceae não costumam gerar grandes
transtornos a indivíduos imunocompetentes, tanto que alguns micro-organismos desta
família fazem parte da flora intestinal humana (FAKRUDDIN et al., 2014). Todavia, o número
de indivíduos com debilidade no sistema imunológico sobe continuamente, como nos casos
de imunodepressão (HIV, câncer, tuberculose entre outras) e imunossupressão (indivíduos
transplantados, submetidos à quimioterapia e portadores de doenças autoimunes)
(REZENDE, 2011). As doenças autoimunes são muito comuns na sociedade atual,
principalmente às ligadas a fatores reumatológicos. Estas doenças predispõem ainda mais
os indivíduos acometidos a doenças infecciosas (LUZ et al., 2007). Com isso, o contato
destes indivíduos com enterobactérias multirresistentes pode ser de suma criticidade.
Representantes da família Enterobacteriaceae podem expressar multirresistência
(MDR; do inglês multidrug resistant), e também fenótipos de susceptibilidade reduzida ao
carbapenens (KPC) e de β-lactamase de espectro estendido (ESBL), uma enzima que
hidrolisa o anel β-lactâmico dos antibióticos lactâmicos. Ambas as classes de antibióticos
são amplamente utilizadas no tratamento de infecções com bactérias gram-negativas
(PFEIFER et al., 2010; KONG et al., 2010).
Artigos evidenciando e/ou investigando o relacionamento de bactérias
multirresistentes com surtos hospitalares estão cada vez mais disponíveis nos bancos de
dados científicos (FERNÁNDEZ et al., 2015; CORNEJO-JUÁREZ et al., 2015; STOESSER
et al., 2015; GEFFERS e GASTMEIER, 2011). Entretanto, há poucos trabalhos relacionando
bactérias patogênicas da família Enterobacteriaceae a produtos lácteos. Os poucos
trabalhos disponíveis, são referentes à espécie Escherichia coli e ao gênero Salmonella.
(KOBAYASHI et al., 2016; LIU et al., 2016b; NUESCH-INDERBINEN et al., 2015; DEVAUX
et al., 2015; LAUNDERS et al., 2013; FARROKH et al., 2013).
Além da multirresistência aos antimicrobianos, outros fatores relacionados a

2
membros da família Enterobacteriaceae destacam-se na literatura, tais como:
O número de bactérias capazes de sobreviverem a ambientes totalmente
desfavoráveis, como em superfícies e equipamentos da indústria de alimentos, devido à
capacidade de produção de biofilme (OLIVEIRA et al., 2013c; OTTER et al., 2011);
A produção de enzimas proteolíticas (proteinases ou proteases), responsáveis pela
deterioração do leite e seus derivados (ZAJÁC et al., 2015), e pela desestabilização das
micelas de caseína, impedindo a coagulação do leite, logo a formação do queijo (CALDERA
et al., 2015; PINTO et al., 2007). Além disso, ainda afetam consideravelmente as
propriedades sensoriais dos produtos lácteos (CALDERA et al., 2015; HANTSIS-
ZACHAROV e HALPERN, 2007);
A produção de enzimas lipolíticas que influenciam diretamente nas propriedades
sensoriais e desenvolvendo no produto um sabor acre e um odor desagradável (CARPINÉ
et al., 2010; SANTOS et al., 2007);
A produção de hemolisina, enzima capaz de causar doença nas glândulas mamárias
dos animais produtores de leite, podendo, assim, afetar o leite delas extraído (BURGOS e
BEUTIN, 2010). No ser humano, afetam os mecanismos de modulação da morte celular
induzida e as vias de sinalização inflamatória (MILHAS et al., 2010);
A produção de substâncias antimicrobianas, que em um nicho de competição,
proporcionam vantagens às bactérias que as produzem, pois servem de ferramenta contra
as suas concorrentes.
A presença de plasmídeos relacionados aos mecanismos de resistência aos
antimicrobianos e a outros fatores de virulência
Diante de todos estes fatores, ressaltou-se a importância de se investigar a
prevalência de bactérias da família Enterobacteriaceae, diferentes de Escherichia coli e
Salmonella spp., em produtos lácteos comercializados no Rio de Janeiro, caracterizando-os
quanto à expressão de multirresistência a drogas e de outros fatores de virulência.

3
2 REFERENCIAL TEÓRICO
2.1 PRODUTOS LÁCTEOS
O leite e seus derivados são a base da alimentação de muitas famílias no Brasil e no
mundo em todas as classes sociais (LAGRANGE et al., 2015). Assim sendo, o leite, uma
das fontes mais acessíveis de energia e nutriente, principalmente para recém-nascidos,
voltou a ocupar posição de destaque no cenário mundial devido aos seus lipídios, proteínas
e polipeptídios com funções fisiológicas. Estes fatores estão presentes na membrana do
glóbulo de gordura do leite, que dentre muitas funções destacam-se no auxílio à inibição do
câncer do colo do útero; na supressão à esclerose múltipla; na ação bactericida e na
inibição do crescimento celular de micro-organismos (ZHOU et al., 2014), assim como há
relatos de sua atuação anti-hipertensiva, antioxidante, anti-inflamatória e
hipocolesterolêmica (HSIEH et al., 2015).
Um dos principais produtos derivados do leite é o queijo, estando o Brasil entre as
principais elites mundiais na sua produção, destacando-se a produção de queijo mussarela,
queijo prato e queijo minas frescal (queijo de alta umidade), os mais consumidos no país,
principalmente após as campanhas de redução de sódio que envolve estes produtos. Para
alguns autores, a redução do sódio nestes produtos acaba deixando-os mais vulneráveis,
pois este é eficaz no controle de alguns micro-organismos patogênicos e/ou deteriorantes,
afetando diretamente a vida de prateleira, além da possibilidade de alterações reológicas e
sensoriais (DAMACENO et al., 2015; CRUZ et al., 2011; DOYLE e GLASS, 2010). Outros
pontos importantes incluem o fato de que os queijos podem não ser fabricados através de
leite pasteurizado e esta produção, muitas vezes, é realizada de forma artesanal em
fazendas, o que pode ser uma fonte em potencial para micro-organismos como os da família
Enterobacteriaceae (ZHANG et al., 2015b). Até mesmo nos produtos que sofrem processo
de pasteurização, Silva e colaboradores (2010) destacaram que não é garantida a ausência
de contaminantes, devido a possíveis falhas no processo ou na pós-pasteurização.
Jeyakumar e Lawrence (2014), por exemplo, conseguiram isolar bactérias entéricas
patogênicas, como E. coli, Salmonella sp., Klebsiella pneumoniae, Proteus vulgaris e
Shigella, em 64% das amostras de leite pasteurizado do seu estudo. Até em fórmulas
infantis e leite em pó, que sofrem processo de retirada de toda umidade, há relatos da
presença de representantes desta família (ARAÚJO et al., 2015; FAKRUDDIN et al., 2014;
OLADIPO e OMO-ADUA, 2011a).

4
2.1.1 Contaminação microbiológica de produtos lácteos
O consumo in natura de alimentos contaminados está entre as principais causas de
doenças transmitidas por alimentos (DTA) nos países em desenvolvimento, e até mesmo
em países desenvolvidos, configurando-se, assim, um grave problema de saúde pública.
Nesse âmbito, o leite e seus derivados merecem destaque (DAMACENO et al., 2015;
CASALINUOVO et al., 2014). Segundo o Committee on Infectious Diseases e o Committee
on Nutrition (2014), nos Estados Unidos, por exemplo, as doenças transmitidas pela
ingestão de produtos lácteos deveriam ter menor impacto, pois em grande parte do país a
legislação não permite a comercialização de produtos lácteos sem processamento térmico,
porém, segundo este comitê, as informações equivocadas de que o consumo in natura
destes produtos traz benefícios à saúde dos consumidores, influenciam negativamente
nesta estatística.
Durante a sua produção, processamento, armazenamento, distribuição e
comercialização, os produtos lácteos podem estar sujeitos a condições de higiene
inadequadas, que podem promover a contaminação por micro-organismos patogênicos e/ou
deteriorantes (CUSATO et al., 2013; FREITAS et al., 2013). Galinari e colaboradores (2014),
por exemplo, avaliaram bancadas e utensílios de madeiras utilizados na produção artesanal
de queijos. Estes utensílios de madeira haviam sido substituídos pela legislação local por
suspeitas de contaminarem os produtos. Todavia, apenas alguns utensílios de madeira
apresentaram contaminação. Em contrapartida, com a nova metodologia de produção ainda
foi possível encontrar contagem de micro-organismos contaminantes, inclusive acima da
contagem permitida pela legislação, sugerindo que o problema possa estar relacionado com
más práticas na produção.
O queijo de alta umidade é um dos exemplos mais clássicos de alimentos
consumidos crus. Em muitos países, este tipo de queijo é obtido através do processo
fermentativo de leite de vaca, soro de leite e adição de água. Esta água, muitas vezes é
utilizada sem tratamento prévio, o que pode aumentar consideravelmente a carga
microbiana durante o tempo de estocagem (BARUZZI et al., 2012). Mesmo em produtos
com ausência ou baixa umidade, os representantes da família Enterobacteriaceae podem
ser encontrados, como relataram Yang e colaborares (2015) em fórmulas lácteas infantis.
O alto teor de nutrientes do leite, e, consequentemente, dos seus derivados, além do
pH próximo ao neutro e elevada atividade de água, fornecem um ambiente ideal para o
desenvolvimento de diversos micro-organismos (OLADIPO e OMO-ADUA, 2011a).
Populações de micro-organismos psicrotróficos que se estabelecem durante o
armazenamento em baixas temperaturas são os principais componentes da microbiota
desses produtos (QUIGLEY et al., 2013). Ao longo dos últimos 50 anos, inúmeros estudos

5
sobre a contaminação de produtos lácteos têm sido realizados, embora muitas questões
relacionadas com as populações de micro-organismos psicrotróficos no leite e seus
derivados ainda permaneçam sem resposta (HANTSIS-ZACHAROV e HALPERN, 2007).
Esses grupos de micro-organismos constituem um problema econômico significativo para a
indústria de produtos lácteos, uma vez que são os principais agentes causadores de
deterioração microbiana nesses produtos, e várias espécies consideradas patogênicas e
muitas vezes resistentes a antibióticos já foram relatadas (ZHANG et al., 2015b;
SAMARŽIJA et al., 2012). A deterioração destes produtos é facilmente percebida, pois os
micro-organismos deteriorantes geram compostos voláteis como, por exemplo, os
compostos voláteis de enxofre, que alteram o odor, a cor e o sabor destes alimentos
(BÖHME et al., 2010).
Já Zhang e colaboradores (2015b) destacaram como ponto importante a alimentação
de animais voltados para a produção de leite, que pode causar distúrbios metabólicos,
levando a acidose ruminal, podendo resultar na translocação de LPS e outros fatores de
virulência para as glândulas mamárias e, consequentemente, para o leite delas extraído.
2.2 A FAMÍLIA ENTEROBACTERIACEAE
As bactérias da família Enterobacteriaceae possuem as seguintes características:
são bastonetes retos gram-negativos, oxidase-negativos, catalase-positivos, não formam
esporos, são anaeróbios facultativos, movimentam-se por flagelos peritríquios, exceto
alguns representantes imóveis (como, por exemplo, Klebsiella spp. e Shigella spp.),
temperatura ideal de crescimento 37ºC, com algumas exceções, a maioria produz ácidos e
gases durante a fermentação de carboidratos (PUBLIC HEALTH ENGLAND, 2015;
TORTORA et al., 2012; MADIGAN et al., 2010; OWEN et al., 2010).
Esta família de micro-organismos compreende um grupo de bactérias entéricas que
possui entre suas representantes, bactérias de baixo potencial patogênico, mas também
bactérias de grande relevância clínica; como as espécies Klebsiella pneumoniae e
Escherichia coli e os gêneros Salmonella e Enterobacter, capazes de gerar, desde leves
desconfortos intestinais a graves infecções em seres humanos e demais animais de sangue
quente. Atualmente, cinquenta e três gêneros estão descritos, sendo que aproximadamente
vinte e seis destes possuem relatos de infecções em humanos (PUBLIC HEALTH
ENGLAND, 2015). Além de infecções intestinais, também estão relacionadas com outros
tipos de infecções como as do trato respiratório, urogenital, cerebral (meningite neonatal
bacteriana) e, ainda, infecção generalizada e pós-cirúrgicas (HAWKEY e CHOY, 2015;
PHILIPPE et al., 2015; COSKUN et al.; 2014; FAKRUDDIN et al., 2014). Dos 26 gêneros
supracitados, algumas bactérias merecem destaque, principalmente as dos gêneros

6
Salmonella e Enterobacter; e as espécies Escherichia coli e Klebsiella pneumoniae.
Na literatura, estão disponíveis diversos trabalhos tanto para a Salmonella spp.
quanto para a Escherichia coli (KOBAYASHI et al., 2016; LIU et al., 2016b; NUESCH-
INDERBINEN et al., 2015; DEVAUX et al., 2015; LAUNDERS et al., 2013; FARROKH et al.,
2013), porém há um número bem limitado de trabalhos evidenciando os gêneros
Enterobacter, Klebsiella e Serratia, principalmente no que se refere a presença destes
gêneros em alimentos, e mais especificamente, em produtos lácteos. Além de poucos, os
trabalhos disponíveis na literatura relacionando estas bactérias a produtos lácteos e/ou
ambientes de produção, armazenamento e comercialização, limitam-se à quantificação, à
identificação e ao perfil de resistência a alguns antimicrobianos. Mas este quadro vem
mudando e já há alguns trabalhos explorando e evidenciando outros fatores de virulência
como, a produção de biofilme (CHERIF-ANTAR et al., 2016), a produção de enzimas
proteolíticas (CHOVE et al.; 2013), a produção de enzimas lipolíticas (MASIELLO et al.,
2016) e a produção de substâncias antimicrobianas (DAMACENO et al., 2015).
2.3 RESISTÊNCIA AOS ANTIMICROBIANOS
Algumas classes de antibióticos são mais eficientes no combate às infecções por
micro-organismos patogênicos da família Enterobacteriaceae. Dentre estas classes,
destacam-se as ciclosporinas de 1ª a 4ª gerações (ceftazidima, cefepima, cefotaxima,
ceftriaxona) e os carbapenens (doripenem, ertapenem, imipenem, meropenem), logo a
utilização destes antibióticos em animais voltados para a obtenção de leite pode levar à
geração de bactérias resistentes. Isto porque, muitas vezes, os antibióticos são utilizados
em demasia e indiscriminadamente nestes animais, objetivando a prevenção e o tratamento
de infecções e o desenvolvimento destes animais (MURPHY et al., 2016; GUIMARÃES et
al., 2012; BOSCO et al., 2012; FLEMING et al., 2010) e esta rotina pode estar causando
grande impacto para a indústria alimentícia (ROLAIN, 2013). O leite é considerado um
excelente meio de cultura para a transferência gênica por conjugação, tendo relato na
literatura de uma eficiência 10 vezes maior do que meios de cultura laboratoriais (VERRAES
et al., 2013). O uso de antibióticos visando o desenvolvimento do animal ainda é muito
discutido, pois alguns autores defendem que o consumo de produtos oriundos destes
animais não traz riscos à saúde humana (PHILLIPS et al., 2004), mas outros questionam a
possibilidade de bactérias com genes de resistência serem transmitidas pelos alimentos
advindos destes animais (GUIMARÃES et al., 2012). Outro ponto importante foi relatado por
Zhang e colaboradores (2009a): em uma fazenda aonde na agricultura era utilizado o
antibiótico Apramicina. Tanto os trabalhadores do local, quanto os animais, possuíam genes
de resistência a este antibiótico, muito semelhantes em suas floras intestinais, que foi

7
evidenciado através de estudos de conjugação gênica.
As diferentes estratégias que as bactérias têm desenvolvido para alcançarem
resistência aos antibióticos, seja por transdução, transformação e/ou conjugação, carece
cada vez mais de estudos, no intuito de se descobrir o papel dos antibióticos na evolução
dos ecossistemas (ROLAIN, 2013). Alterações na permeabilidade das membranas e
paredes celulares, o papel das porinas e, principalmente, a participação das bombas de
efluxo, constam na literatura como alguns dos principais responsáveis por este processo de
resistência (PHILIPPE et al., 2015; BECEIRO et al., 2013). As bombas de efluxo são
mecanismos de bombeamento de solutos para fora do citoplasma celular, como substâncias
tóxicas à célula, dentre elas os antimicrobianos (SOTO, 2013). Por estarem relacionadas à
resistência aos antibióticos e à produção de biofilme, as bombas de efluxo estão entre os
mecanismos mais visados pelos pesquisadores na busca de elucidar a sua contribuição nos
processos biológicos e avaliação da eficácia de agentes terapêuticos (BLAIR e PIDDOCK,
2016).
Porém, dos fatores relacionados à aquisição de resistência aos antimicrobianos
pelos representantes da família Enterobacteriaceae, o que parece estar preocupando mais a
comunidade científica é a capacidade dessas bactérias produzirem β-lactamases de
espectro estendido (ESBL), que são enzimas mediadas por plasmídeos, assim chamadas
porque lisam a estrutura do anel β-lactâmico presente nos antibióticos lactâmicos: as
penicilinas, as cefalosporinas, os carbapenêmicos e os monobactâmicos (TEKINER e
ÖZPINAR, 2016; THENMOZHI et al., 2014). Estes antibióticos são amplamente utilizados
para o tratamento de diversas infecções bacterianas, em especial as relacionadas a
bactérias Gram-negativas, logo conferindo resistência a uma quantidade considerável de
antibióticos destas classes. Outro ponto preocupante é que estas bactérias têm também se
mostrado resistentes aos antibióticos inibidores de β-lactamases (THAM et al., 2012).
Vale ressaltar que estes fenômenos podem ocorrer, inclusive, no lúmen dos
intestinos humano e/ou animal, através de processos lisogênicos, influenciando na evolução
de novas cepas patogênicas (GROTIUZ et al., 2006).
As opções de antimicrobianos estão cada vez mais escassas devido à aquisição de
resistência bacteriana, atingindo um ponto preocupante. A indústria farmacêutica não
encontra incentivos para buscar novos princípios ativos, visto que, de certo ponto, este
tipo de produto não se torna rentável, pois o desenvolvimento de novos medicamentos
envolve anos de pesquisa por um produto de uso relativamente curto quando comparado
a medicamentos que os indivíduos utilizam por longos períodos (BROGAN e
MOSSIALOS, 2013). O que se tem visto é a tentativa de a indústria farmacêutica produzir
compostos eficazes através da modificação de antibióticos já existentes (ZIPPERER et al.,
2016).

8
2.4 PRODUÇÃO DE FATORES DE VIRULÊNCIA
2.4.1 Produção de biofilme
A formação do biofilme na indústria de produtos lácteos pode ocorrer em poucas
horas após um processamento (aproximadamente 8 a 12 horas) (MOGHA et al.,2014).
Para que ocorra a formação de biofilme em uma determinada superfície,
primeiramente deve ocorrer adesão à superfície, seguido da formação da matriz polimérica,
replicação celular e, finalmente, a separação celular (BAYOUMI et al., 2012). A Figura 1
demonstra um esquema básico para a formação do biofilme.
Figura 1: etapas da formação do biofilme bacteriano
Fonte: adaptado de Rendueles e Ghigo (2012)
O biofilme é uma estrutura porosa complexa, composta de matriz polimérica
extracelular (polissacarídeos, proteínas, lipídeos), DNA, umidade e nutrientes, aonde
conjuntos de subpopulações heterogêneas de micro-organismos, com comportamentos
diversificados e características fisiológicas específicas, formam um ambiente de cooperação

9
ecológica altamente eficiente e resistente (BRIDIER et al., 2015; MARQUES et al., 2013;
SOTO, 2013). Neste ambiente de cooperação, alguns indivíduos são mais resistentes,
enquanto outros dependem fortemente desta interação, numa diversidade de características
fenotípicas e heterogeneidade fisiológica (OTTER et al., 2011). Mas, apesar de estes
indivíduos viverem em cooperação, não estão livres do estresse oxidativo, que pode levar a
erros e mutações no momento da replicação do DNA bacteriano. Segundo Høiby e
colaboradores (2010), estas mutações estão diretamente relacionadas ao crescimento do
biofilme.
Beceiro e colaboradores (2013) sugeriram uma forte relação entre a produção de
biofilme e a transferência horizontal de genes. Para os autores, numa população que forma
biofilme, estirpes presentes nesta biomassa podem apresentar resistência a antibióticos e
outros fatores de virulência, e este “pool de genes” pode ser transferido entre elas,
principalmente por conjugação.
Assim como ocorre com os fatores de resistência a antibióticos e sanitizantes, a
formação de biofilme também tem forte relação com as bombas de efluxo. O papel das
bombas de efluxo na formação de biofilme carece de mais estudos. Baugh e colaboradores
(2014) relataram que a inativação das bombas de efluxo é capaz de reprimir a transcrição
de componentes da matriz polimérica do biofilme. Por outro lado, Schlisselberg e
colaboradores (2015) avaliaram estirpes de Salmonella enterica tratadas com o antibiótico
cloranfenicol e verificaram que estas bactérias, mesmo com os mecanismos das bombas de
efluxo inativados, foram capazes de produzir biofilme.
Além disso, uma diversidade de enzimas pode ser produzida pelas bactérias dentro
do ambiente formado pelo biofilme, podendo causar a deterioração dos produtos lácteos
(TEH et al., 2014b).
Apesar do controle mais rígido exigido pela indústria de alimentos, os micro-
organismos podem produzir biofilme, nas bancadas, equipamentos, esteiras, tubulações,
entre outros (VERRAES et al., 2013). Bayoumi e colaboradores (2012) relataram a presença
de biofilme constituído por indivíduos do gênero Cronobacter em superfícies de polipropileno
(5,48 logs UFC/cm2) e aço inoxidável (5,44 logs UFC/cm2). Segundo Simões e
colaboradores (2010), as reações químicas e biológicas ocorridas dentro do biofilme geram
corrosões nas superfícies que, podem reduzir a eficácia da transferência de calor nos
processos térmicos. Estas superfícies, se não sofrerem processos eficazes de sanitização,
podem permitir a presença destas bactérias por meses e até anos (ČABARKAPA et al.,
2015). O biofilme pode acumular-se após cada processo de lavagem e sanitização,
sugerindo que, no biofilme, as bactérias estão protegidas contra danos físicos, químicos e
condições de estresse, e que o tempo entre os processos de sanitização deve ser curto,
mas isto pode favorecer a formação de indivíduos mais resistentes (LIPSKI, 2005).

10
2.4.2 Atividade proteolítica
Bactérias capazes de sintetizar enzimas proteolíticas são as grandes responsáveis
pela deterioração do leite e seus derivados, podendo causar diversos transtornos à indústria
de produtos lácteos (ZAJÁC et al., 2015). Estas enzimas podem, ainda, ser responsáveis
pela etapa inicial da produção de biofilme (PONCE et al.; 2012).
A atividade proteolítica é um importante fator de virulência, capaz de propiciar a
sobrevivência de muitos patógenos dentro do organismo hospedeiro e, assim, trazer sérios
riscos à saúde dos consumidores (IQBAL et al., 2013).
Quando relacionadas aos produtos lácteos, na produção de queijos, por exemplo,
estas enzimas desestabilizam as micelas de caseína e podem alterar ou até mesmo impedir
a coagulação do leite, podendo afetar diretamente a formação do produto (CALDERA et al.,
2015; PINTO et al., 2007). Outro grande problema é que estas bactérias levam a um
processo conhecido como off-flavour, ou seja, afetam consideravelmente as propriedades
sensoriais dos alimentos como, a cor, o odor, o sabor, e a textura (CALDERA et al., 2015;
HANTSIS-ZACHAROV e HALPERN, 2007), o que afeta diretamente a aceitação ou a
rejeição do produto pelo consumidor (TEIXEIRA, 2009). Além disso, muitas destas enzimas
são termorresistentes, ou seja, capazes de resistirem às temperaturas baixas de
refrigeração e às temperaturas elevadas dos processos de esterilização (UHT e
pasteurização) (BAUR et al., 2015).
Não são apenas os fatores negativos que colocam a produção de proteinases em
evidência. Algumas proteinases produzidas por bactérias são de grande interesse para os
processos biotecnológicos, podendo ser obtidas em larga escala e em um curto período de
tempo (ABED et al., 2016), e as colônias de bactérias que as produzem são de fácil
manipulação e manutenção (RODARTE et al. 2011). Estas enzimas conseguem hidrolisar a
caseína do leite e, como consequência, liberam peptídeos bioativos, com propriedades
funcionais e farmacêuticas, dentre elas, ação antioxidante, antimicrobiana, anticancerígena
e transportadora de cálcio, melhorando a biodisponibilidade deste micronutriente, logo se
tem buscado produzir alimentos, especialmente os nutracêuticos, que são considerados por
muitos especialistas como alimentos com ação de medicamento (MUHAMMAD et al., 2016;
LI et al., 2013; SALAMI et al., 2011; PHELAN e KERINS, 2011; MAO et al., 2011).
2.4.3 Atividade lipolítica
A síntese bacteriana de enzimas lipolíticas tem se mostrado um processo ao mesmo
tempo preocupante e importante para a indústria alimentícia, porque exercem influência
direta nas propriedades sensoriais destes produtos, principalmente no aroma e na textura

11
(KREWINKEL et al., 2016; CARPINÉ et al., 2010). A lipólise pode levar a um processo
denominado rancidez hidrolítica, desenvolvendo no produto um sabor acre e um odor
desagradável (SANTOS et al., 2007).
A alimentação dos ruminantes é composta por ácidos graxos poliinsaturados.
Todavia, devido à ação de enzimas lipolíticas, produzidas por bactérias, dentro do
organismo animal, o leite obtido destes animais, assim como os produtos lácteos deles
obtidos, são compostos por ácidos graxos saturados, sabidamente mais maléficos à saúde,
podendo causar, principalmente, doenças coronarianas (LOURENÇO et al., 2010), formação
de tumores e outras doenças em humanos (ZECKNER et al., 2012).
O interesse por enzimas catalizadoras sintetizadas por micro-organismos vem
crescendo constantemente, pois apresentam inúmeras vantagens quando comparadas às
enzimas de origem animal e vegetal, dentre elas, o baixo custo e a capacidade de larga
escala de produção; a modificação de gorduras e óleos; o realce no sabor e até mesmo a
utilização no tratamento de resíduos industriais (ROVEDA et al., 2010).
2.4.4 Atividade hemolítica
Algumas bactérias são capazes de expressar atividade hemolítica devido à produção
de hemolisinas, bem como podem, ainda, por transferência gênica horizontal, estar
transferindo plasmídeos responsáveis pela produção destas hemolisinas, como alguns
representantes da família Enterobacteriaceae. O principal mecanismo de ação das
hemolisinas é a criação de poros na membrana celular causando, por exemplo, doença nas
glândulas mamárias dos animais utilizados pela indústria alimentícia, podendo assim afetar
o leite delas extraído (BURGOS e BEUTIN, 2010). Dois tipos de hemolisina destacam-se: a
α-hemolisina e a β-hemolisina, sendo que a β-hemolisina é mais preocupante, pois atua na
esfingomielina da membrana celular (MARQUES et al.,2013). Esta ação, neste importante
componente da membrana plasmática, pode afetar a sinalização celular, influenciar nos
mecanismos de modulação da morte celular induzida e nas vias de sinalização inflamatória
(MILHAS et al., 2010).
Além disso, a sobrevivência de muitas bactérias no hospedeiro e a sua
patogenicidade acabam dependendo do sequestro do ferro das proteínas de ligação e de
transporte deste nutriente no hospedeiro, mas pouco se sabe sobre este mecanismo de
sequestro do ferro, por estas bactérias, e as estratégias utilizadas por estas para escapar do
processo denominado imunidade nutricional. O ferro é essencial para o metabolismo
humano, mas também para o processo de replicação bacteriana (ATANASSOVA et al.,
2015; BARBER e ELDE, 2014; CASSAT e SKAAR, 2013).

12
2.5 PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM)
Algumas bactérias patogênicas, como alguns representantes da família
Enterobacteriaceae, são capazes de produzir compostos biologicamente ativos, conhecidos
como substâncias antimicrobianas, atuando, num nicho de competição, contra suas
concorrentes. Estas substâncias, através de processos de purificação, vêm sendo utilizadas
pela indústria alimentícia como ferramenta contra bactérias que causam deterioração em
seus produtos, logo aumentando a sua vida de prateleira (DAMACENO et al., 2015;
VERRAES et al., 2013). Nesse sentido, atuam na manutenção das características destes
produtos, visto que possuem ação bactericida e/ou bacteriostática sem alterar as
propriedades sensoriais dos alimentos (OLIVEIRA et al., 2012b). Estas substâncias têm se
mostrado essenciais à indústria alimentícia, visto que os antibióticos não podem ser
utilizados em alimentos (OLIVEIRA et al., 2011a).
Uma das principais substâncias antimicrobianas produzidas por bactérias são as
bacteriocinas. Seu mecanismo de ação em uma célula-alvo é a produção de poros na
membrana citoplasmática desta célula, provocando a saída de compostos celulares e
alterando o gradiente eletroquímico (força próton-motora), fundamental para a geração de
energia e a síntese de proteínas, podendo, com isso, levar a célula-alvo à morte (ZHENG et
al., 2015; OLIVEIRA et al., 2012b; BAYOUB et al., 2011).
As bacteriocinas produzidas por bactérias comensais do trato intestinal animal
podem ter um papel importante na eliminação de micro-organismos patogênicos, sem
alterações significativas na flora intestinal (KOMMINENI et al., 2015), pois estas substâncias
são degradas por enzimas do sistema digestivo e ainda possuem propriedades probióticas
(ROSA et al., 2016).
É importante ressaltar que o número de estudos relacionados às substâncias
antimicrobianas ainda é muito reduzido, mas este cenário está começando a mudar, na
busca de se elucidar e/ou aprimorar os seus mecanismos de ação antimicrobiana e utilizá-
los não só em alimentos, mas também no tratamento de doenças em seres humanos e
animais (FLEMING et al., 2010).

13
3 JUSTIFICATIVA
Alguns representantes da família Enterobacteriaceae, eventualmente, estão
relacionados a surtos hospitalares, especialmente em unidades de terapia intensiva, aonde
na maioria dos casos os indivíduos encontram-se imunocomprometidos, estando, assim,
mais vulneráveis a estes micro-organismos. Entretanto, o número de indivíduos com
debilidade no sistema imunológico fora do ambiente hospitalar sobe continuamente em todo
o mundo (BERG et al., 2014).
Diante do exposto, é necessário que se evidencie potenciais fontes extra-
hospitalares veiculadoras destes micro-organismos e neste contexto, os alimentos possuem
grande probabilidade. É consenso que os produtos lácteos são muito consumidos
mundialmente e por todas as classes sociais, mas que pelas condições de fabricação,
comercialização e consumo, podem ser considerados potenciais reservatórios extra-
hospitalares de representantes da família Enterobacteriaceae.
Há um número extremamente reduzido de trabalhos associando os produtos lácteos
a patógenos desta família e, geralmente, a pesquisa é mais direcionada para a presença da
espécie Escherichia coli e do gênero Salmonella. Dessa forma, fazem-se necessários mais
estudos evidenciando e caracterizando os demais micro-organismos desta família, nestes
alimentos, principalmente a espécie Klebsiella pneumoniae e o gênero Enterobacter,
eventualmente relacionados a surtos hospitalares (LIU et al.; 2016a; THOMAS et al., 2013;
MSHANAA et al., 2011; KONTOPOULOU et al., 2010).

14
4 OBJETIVOS
4.1 OBJETIVO GERAL
O objetivo deste trabalho foi investigar a presença de bactérias da família
Enterobacteriaceae, diferentes de Escherichia coli e Salmonella spp., provenientes de
amostras de produtos lácteos comercializados na cidade do Rio de Janeiro e caracterizar os
isolados quanto à expressão de multirresistência aos antimicrobianos e de outros fatores de
virulência, além de investigar o possível papel desses alimentos como potenciais
veiculadores destas bactérias fora do ambiente hospitalar.
4.2 OBJETIVOS ESPECÍFICOS
Isolar, identificar e caracterizar estirpes da família Enterobacteriaceae, diferentes de
Escherichia coli e Salmonella spp., a partir de produtos lácteos comercializados na
cidade do Rio de Janeiro;
Determinar o perfil de resistência a antimicrobianos dos isolados;
Avaliar a expressão de fenótipos de susceptibilidade reduzida aos carbapenens
(KPC) e de produção de β-lactamase de espectro estendido (ESBL) dos isolados;
Caracterizar os isolados obtidos quanto à produção qualitativa de biofilme e quanto à
expressão de atividade proteolítica; atividade lipolítica e atividade hemolítica;
Investigar a produção de substâncias antimicrobianas (SAM) pelos isolados;
Determinar e comparar o perfil plasmideal dos isolados MDR e não MDR do
complexo Enterobacter cloacae.

15
5 MATERIAIS E MÉTODOS
5.1 OBTENÇÃO DAS AMOSTRAS DE PRODUTOS LÁCTEOS
As amostras de produtos lácteos (queijo, leite e bebida láctea fermentada) utilizadas
neste trabalho foram obtidas em diferentes estabelecimentos comerciais da cidade do Rio
de Janeiro como supermercados, padarias e queijarias. Cada amostra foi acondicionada
numa caixa térmica, contendo gelo reutilizável, dentro da embalagem original de cada
produto e conduzida ao Laboratório de Microbiologia do IFRJ no máximo duas horas após a
sua aquisição. No presente estudo utilizou-se amostragem de conveniência.
5.2 ISOLAMENTO DOS MICRO-ORGANISMOS
Após assepsia da embalagem, foram pesados 25g (ou 25 ml) de cada amostra e
colocados em 225 ml de água peptonada estéril 0,1% (BIOCEN do Brasil, São Paulo).
Foram preparadas diluições seriadas, até 10-2, em tubos de ensaio com rosca contendo o
mesmo diluente, sem considerar o processo de colocação dos 25g/25 ml em 225 ml de água
peptonada como primeira diluição.
Alíquotas das diluições foram inoculadas, pelo método spread plate, em placas
contendo os meios indicadores seletivos ágar EMB (Eosina Azul de Metileno; Himedia,
Índia) e ágar MacConkey (Merck, Alemanha). As placas foram incubadas a 37°C por 18-24h.
Após o prazo estipulado, as placas foram avaliadas e cerca de 50% do número de colônias
obtidas em cada placa foram selecionadas e passadas individualmente para placas
contendo ágar Casoy (BIOCEN do Brasil, São Paulo). Colônias sugestivas de Escherichia
coli, com brilho verde metálico no ágar EMB, não foram selecionadas.
Para confirmação da seleção de bactérias Gram-negativas, realizou-se a coloração
de Gram dos isolados pré-selecionados e, em seguida, foi realizado o teste da oxidase.
Todos os representantes da família Enterobacteriaceae apresentam-se negativos para o
teste da oxidase. Realizou-se o estriamento em placas contendo ágar Casoy. Com uma alça
descartável, coletou-se uma colônia e colocou-se na fita do teste da oxidase (Laborclin,
Paraná). Os isolados que, em até 20 segundos, apresentaram coloração púrpura, indicando
positividade para o teste de oxidase, foram descartados.
5.3 MANUTENÇÃO DOS ISOLADOS
Após a purificação da cultura, o crescimento em massa de cada isolado foi

16
coletado e transferido para criotubos contendo 1,5 mL de caldo Casoy (p v-1; Merck,
Alemanha), acrescidos de glicerol 40% (v v-1; Merck, Alemanha), devidamente identificados
e armazenados em criotubos a -20°C ± 2°C até o momento do uso.
5.4 IDENTIFICAÇÃO DOS ISOLADOS
A identificação dos isolados foi realizada através da tecnologia de espectrometria de
massas, utilizando-se o equipamento VITEK 2, modelo Compact (BioMèrieux, França).
5.5 PERFIL DE RESISTÊNCIA A ANTIMICROBIANOS
A determinação do perfil de resistência a antimicrobianos das estirpes produtoras e
indicadoras foi realizada utilizando-se a técnica de difusão em disco, conforme
recomendações do Clinical and Laboratory Standards Institute (CLSI, 2015) para bactérias
da família Enterobacteriaceae.
A partir do isolamento em ágar Casoy, algumas colônias foram transferidas para
tubos de ensaio contendo 5 mL de água peptonada 0,1% até a obtenção da turvação
correspondente a 0,5 da escala de McFarland (aproximadamente 1,5 x 108 UFC/mL).
Após obter a turvação ideal, mergulhou-se um swab estéril no tubo, pressionou-se
levemente contra a parede do mesmo, para que o excesso fosse retirado. O material foi
semeado sobre a superfície de uma placa contendo ágar Müeller Hinton (Himedia, Índia),
em pelo menos três direções diferentes, para a obtenção de um crescimento homogêneo e
confluente. Com o auxílio de uma pinça estéril, os discos de antibióticos foram dispostos de
forma equidistante na placa, perfazendo um total de, no máximo, cinco discos por placa. As
placas foram incubadas a 37°C por 18-24h. Após este período, o diâmetro (em mm) dos
halos de inibição formados foi medido e a interpretação dos resultados foi realizada segundo
as recomendações da CLSI (2015). Foram consideradas multirresistentes as bactérias que
apresentaram resistência a, no mínimo, um antibiótico em, pelo menos, três classes
diferentes. Foram utilizados dezessete antibióticos (Laborclin, Paraná), distribuídos em onze
classes que estão apresentados na Tabela 1.

17
Tabela 1: Antibióticos utilizados no presente estudo
Classe Antibiótico
Penicilinas AMP 10 µg
Inibidores de β-Lactamases AMC 20/10 µg
Cefalosporinas (3ª geração) CTX 30µg
CAZ 30 µg
Cefalosporinas (1ª geração) CEP 30µg
Quinolonas CIP 5µg
NOR 10µg
Aminoglicosídeos
AMI 30µg
GEN 10µg
STR 10µg
TOB 10µg
Fenicóis CHL 30 µg
Carbapenêmicos IPM 10µg
MER 10µg
Monobactâmicos ATM 30µg
Análogos do Ácido Fólico TRI 5µg
Tetraciclinas TET 30µg
Legenda: AMP, ampicilina; AMC, amoxicilina + ácido clavulânico; CTX, cefotaxima; CAZ, ceftazidima;
CEP, cefalotina; CIP, ciprofloxacina; NOR, norfloxacina; AMI, amicacina; GEN, gentamicina; STR,
estreptomicina; TOB, tobramicina; CHL, cloranfenicol; IPM, imipenem; MER, meropenem; ATM,
aztreonam; TRI, trimetoprim; TET, tetraciclina; µg, micrograma.
5.5.1 Detecção dos fenótipos de ESBL e KPC
Para fins de detecção dos fenótipos KPC e ESBL dos isolados multirresistentes,
através de um método prático e rápido, foram utilizados dois meios de cultura cromogênicos:
CHROMagar®KPC e CHROMagar®ESBL.
Para cada isolado multirresistente, obtido após isolamento em placa contendo ágar
Casoy, foi estriada uma colônia em cada um dos meios cromogênicos. As placas foram
incubadas a 37°C por 18-24h.
A Tabela 2 apresenta o modo de interpretação dos resultados dos testes com os
meios cromogênicos utilizados, mas apenas para os gêneros da família Enterobacteriaceae,
alvos do presente estudo.

18
Tabela 2: Interpretação dos resultados nos meios cromogênicos CHROMagar® KPC e
CHROMagar® ESBL para os representantes da família Enterobacteriaceae.
Resultado positivo
Gênero CHROMagar® KPC
Estirpes resistentes aos carbapenens
CHROMagar® ESBL Estirpes produtoras de β-lactamase
de espectro estendido
Citrobacter sp - Colônias azuis metálicas
Enterobacter sp Colônias azuis metálicas Colônias azuis metálicas
Klebsiella sp Colônias azuis metálicas Colônias azuis metálicas
Serratia sp N/A Colônias azuis metálicas
Legenda: N/A - Não aplicável; KPC - Susceptibilidade reduzida aos carbapenens; ESBL - β-lactamases de espectro estendido (Fonte: http://www.CHROMagar.com).
5.6 AVALIAÇÃO DA PRODUÇÃO DE FATORES DE VIRULÊNCIA
5.6.1 Detecção qualitativa da produção de biofilme
Os isolados foram inoculados sobre a superfície de placas de ágar Vermelho Congo
[(15 g/l de ágar nutriente; p v-1; Merck, Alemanha); (37 g/l de sacarose; p v-1; Pro Analysi,
São Paulo), e, após autoclavação, adicionado solução de vermelho congo (0,8 g/l; p v-1;
Vetec, Rio de Janeiro)], conforme descrito por Freeman e colaboradores (1989), com
algumas modificações. As placas foram incubadas a 37°C por 24h. Foi utilizada como
controle positivo a cepa padrão Salmonella enterica ATCC14028.
Nesta técnica, os isolados produtores de biofilme apresentam-se como colônias de
coloração preta, enquanto que os isolados não produtores se apresentam despigmentados
ou avermelhados.
5.6.2 Detecção de atividade proteolítica
Para esta análise, foi utilizado ágar leite [(leite em pó integral a 10%; p v-1; Nestlé,
São Paulo), ágar-ágar a 2% (p v-1; Merck, Alemanha)], conforme descrito por Minotto e
colaboradores (2014).
O ágar leite foi autoclavado por 5 minutos e vertido em placas. Os isolados foram
inoculados e incubados por 18-24h a 37ºC. Após a incubação, a hidrólise da caseína no
ágar leite foi observada pela presença de halo transparente ao redor das colônias.

19
5.6.3 Detecção de atividade lipolítica
Para esta metodologia foi utilizado o meio de cultura ágar Spirit Blue [1% de digerido
de caseína (p v-1; Himedia, Índia), 0,05% de extrato de levedura (p v-1; Becton, Dickinson
and Company, Estados Unidos da América), 2% de ágar-ágar (p v-1; Merck, Alemanha),
0,0002% de Spirit Blue (v v-1; Mago, São Paulo), 970 ml de água destilada] com adição de
3% de reagente lipase [0,12 ml de Tween 80 (v v-1; Proquimico, Brasil)], 12 ml de azeite de
oliva (v v-1; Natural Alimentos, São Paulo), 17,9 ml de água destilada], conforme descrito em
Becton, Dickinson and Company (2009), com algumas modificações.
O ágar Spirit Blue e o reagente lipase foram autoclavados por 15 minutos, em um
mesmo ciclo, porém em recipientes separados. Após a autoclavação, ambos os recipientes
foram resfriados até que se atingisse a temperatura de 55°C. No ambiente estéril de uma
cabine de segurança biológica, o reagente lipase foi adicionado cuidadosamente ao
recipiente contendo o ágar Spirit Blue; homogeneizado sob forte agitação; em seguida, o
material obtido foi vertido em placas de petri estéreis. Os isolados foram inoculados na
superfície do ágar e incubados por 18-24h a 37ºC. As culturas foram avaliadas quanto à
formação de halo opaco ao redor das colônias, diferenciando-se da coloração azulada do
meio de cultura utilizado.
5.6.4 Detecção de atividade hemolítica
A atividade hemolítica dos isolados foi verificada através do crescimento em placas
contendo ágar sangue base com 5% de sangue de carneiro desfibrinado (p v-1; BIOCEN do
Brasil, São Paulo). As placas foram incubadas a 37°C por 48 h e as culturas foram avaliadas
quanto à formação de zona de hemólise.
5.7 AVALIAÇÃO DA PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM)
Este método foi realizado conforme descrito por Giambiagi-Marval e colaboradores
(1990), com algumas modificações.
A partir do estoque, as bactérias foram crescidas em caldo Casoy por 18h a 37 ºC.
Cinco (5) µl deste material foi inoculado sob a forma de ponto na superfície de uma placa
contendo ágar Casoy (p v-1; Merck, Alemanha). As placas foram incubadas a 37ºC por 18 h
e, em seguida, as bactérias foram mortas por exposição a vapores de clorofórmio por 30
min. Após a evaporação do clorofórmio, verteu-se 3 mL de Casoy semissólido acrescido de
uma alíquota de 0,3 mL da estirpe indicadora (previamente crescida em 3 mL de caldo

20
Casoy a 37°C por 18h). As placas foram incubadas a 37ºC por 18 h.
Neste teste, a produção de SAM é avaliada pela formação de zonas de inibição ao
redor dos pontos de crescimento das estirpes produtoras. As bactérias utilizadas como
indicadoras nos testes estão descritas na Tabela 3.
Tabela 3: Bactérias utilizadas como indicadoras da produção de substâncias antimicrobianas
Identificação Referência
Escherichia coli Escherichia coli
Klebsiella pneumoniae Proteus mirabilis Proteus vulgaris
Pseudomonas aeruginosa Pseudomonas fluorescens
Salmonella enterica sorotipo Typhi Salmonella spp. I
Salmonella spp. VI Yersinia enterocolitica
ATCC 25922 LMIFRJ
ATCC 4352 LMIFRJ LMIFRJ
ATCC 27853 ATCC 13525 ATCC 19214
LMIFRJ LMIFRJ
ATCC 9610
Legenda: ATCC - American Type Culture Collection; LMIFRJ – Laboratório de Microbiologia do Instituto Federal de Educação, Ciência e Tecnologia do Rio de Janeiro.
5.8 DETERMINAÇÃO E COMPARAÇÃO DO PERFIL PLASMIDEAL DOS ISOLADOS DO
COMPLEXO ENTEROBACTER CLOACAE
Para fins de detecção do perfil plasmideal, todos os isolados do complexo
Enterobacter cloacae foram submetidos à extração de DNA.
A extração de DNA plasmideal foi realizada por lise alcalina conforme descrito por
Birnboim e Doly (1979), com algumas modificações. O perfil plasmideal foi analisado através
de géis de agarose a 0,8 % (p v-1) preparados em tampão TBE (Tris 89 mM, ácido bórico 89
mM, EDTA 2,5 mM; pH 8,2) e contendo o corante GelRed (Biotium, USA). As preparações
de DNA foram submetidas à eletroforese horizontal em tampão TBE a 80 V por 5 h. Após a
eletroforese, os géis foram visualizados e fotografados através do sistema de captura de
imagens L-PIX (Loccus Biotecnologia, São Paulo).
.

21
6 RESULTADOS E DISCUSSÃO
Para este estudo, foram analisadas 5 amostras de queijo de alta umidade (queijo
minas frescal), 3 amostras de bebida láctea fermentada e 2 amostras de leite pasteurizado
(comercializados em sacos plásticos), totalizando 10 amostras de conveniência, e
representando os produtos lácteos mais consumidos.
Foram encontrados, no total, 89 isolados, entretanto, 31 deles foram descartados
após a identificação no sistema VITEK 2, sendo 10 isolados da família Moraxellaceae
(Acinetobacter spp.), 6 da família Pseudomonaceae (Pseudomonas spp.), 4 da família
Neisseriaceae (Chromobacterium spp.), 3 da família Burkseudomonaceae (Burkholderia
spp) e 6 da família Enterobacteriaceae com baixa relevância na área clínica e de alimentos
[Kluyvera spp. (3), Tatumella spp. (1), Rahnella spp. (1), Escherichia hermannii (1)].
Dessa forma, deu-se prosseguimento ao trabalho com 58 isolados da família
Enterobacteriaceae (Tabela 4) sendo, 21 (36%) do complexo Enterobacter cloacae, 20
(35%) da espécie Serratia marcescens; 8 (14%) da espécie Raoutella planticola; 3 (5%) da
espécie Citrobacter freundii; 2 (3%) da espécie Klebsiella pneumoniae; 2 (3%) da espécie
Citrobacter braakii; 1 (2%) da espécie Klebsiella oxytoca; 1 (2%) do gênero Raoutella (2%).
Recentemente, Masiello e colaboradores (2016), a partir de 21 amostras de leite
pasteurizado, também isolaram uma porcentagem considerável de bactérias da família
Enterobacteriaceae, e assim como no presente trabalho, o maior número de isolados foi do
gênero Enterobacter (42%), seguidos dos gêneros Citrobacter (12%), Serratia (10%) e
Raoutella (9%).
Tabela 4: Isolados da família Enterobacteriaceae obtidos no presente estudo.
Identificação Isolados Origem
Q1C2
Q1C3
Q1L2
Q1R1
Q1R2
Q1R4
Q2E1
Complexo Enterobacter cloacae Q2L1 Queijo
Q2M1
Q2R2
Q2R3
Q11
Q12
Q15
Q16
Continua

22
Continuação da Tabela 4
R4
E1
E2 Leite
E3
L3
BLL1 Bebida Láctea Fermentada
Q1E2
R7 Queijo
R9
R19
E4
E5
E6
E7
E8
Serratia marcescens E9
E10
E12 Leite
E13
E14
E15
E16
E17
E19
E20
L6
Q1L1
Q1R3
Q2E2
Raoutella planticola Q2E3 Queijo
Q2R4
QJ3
QJ5
E11 Leite
Raoutella sp Q1M1 Queijo
Q1M3
Citrobacter freundii Q1R5 Queijo
Q2R1
Citrobacter braakii Q1C1 Queijo
Q1E1
Klebsiella pneumoniae Q2C3 Queijo
R2
Klebsiella oxytoca Q1E3 Queijo
O complexo Enterobacter cloacae, que constituiu o maior número de isolados
obtidos, como apresentado na Figura 2 é composto por 6 espécies (Enterobacter cloacae,
Enterobacter asburiae, Enterobacter hormaechei, Enterobacter kobei, Enterobacter ludwigii
e Enterobacter nimipressuralis). A diferenciação dessas espécies por meio de identificação
fenotípica é muito difícil, necessitando, assim, do uso de métodos moleculares

23
(MEZZATESTA et al., 2012). Desde 1997 vem surgindo relatos da associação de alguns
representantes deste grupo com infecções hospitalares (DAVIN-REGLI e PAGÈS, 2015;
MEZZATESTA et al., 2012), especialmente a espécies Enterobacter cloacae e Enterobacter
hormaechei, porém em produtos lácteos os trabalhos são mais escassos.
21
20
8
3
2
2
1
1
0 5 10 15 20 25
Complexo Enterobacter cloacae
Serratia marcescens
Raoutella planticola
Citrobacte freundii
Citrobacter braakii
Klebsiella pneumoniae
Klebsiella oxytoca
Raoutella sp
Número de isolados
Ide
ntifica
çã
o
Figura 2: Número de isolados da família Enterobacteriaceae obtidos no presente estudo.
Masiello e colaboradores (2016), em um estudo com 29 amostras de leite
pasteurizado obtidas nos Estados Unidos, em suas embalagens originais, conseguiram
isolar 100 representantes do gênero Enterobacter, 28 do gênero Cronobacter, 24 do gênero
Serratia e 22 do gênero Raoutella. A diversidade de gêneros encontrada foi muito próxima
da obtida no presente estudo.
Fakruddin e colaboradores (2014), num estudo semelhante ao presente trabalho,
porém com amostras mais heterogêneas, entre elas, leite em pó, conseguiram 3 isolados do
gênero Enterobacter, 3 do gênero Citrobacter e 2 do gênero Klebsiella. Já Jeyakumar e
Lawrence (2014), num estudo com 50 amostras de leite cru, isolaram 5 representantes da
espécie Klebsiella pneumoniae. Em nosso estudo, entretanto, nas duas amostras de leite
utilizadas, não foram encontrados isolados da espécie Klebsiella pneumoniae, mas esta
espécie foi identificada a partir das amostras de queijo utilizadas.
No nosso trabalho, foram encontrados 20 isolados da espécie Serratia marcescens,
o que não é muito relatado na literatura. Um dos poucos trabalhos que descreve a detecção

24
dessas bactérias em produtos lácteos foi o publicado por Teh e colaboradores (2011a), que
citam o isolamento de 17 representantes desse gênero a partir de amostras obtidas de 2
tanques de leite cru.
Yao e colaboradores (2012) em amostras de 185 embalagens de fórmulas infantis
conseguiram isolar 8 representantes da espécie Klebsiella pneumoniae, 1 da espécie
Citrobacter freundii, 1 da espécie Enterobacter cloacae, comuns ao presente estudo. Sani e
Yi (2011), também em amostras de fórmulas infantis, isolaram 2 representantes da espécie
Klebsiella pneumoniae, semelhante ao presente estudo e, 4 representantes do complexo
Enterobacter cloacae. Já Oonaka e colaboradores (2010), isolaram 2 representantes da
espécie Klebsiella pneumoniae, além de 6 representantes da espécie Enterobacter cloacae
em fórmulas infantis. Em um trabalho anterior do nosso grupo de pesquisa, com amostras
de fórmulas infantis, foram isolados 12 representantes do complexo Enterobacter cloacae
(ARAÚJO et al., 2015). Esses estudos citados demonstram que as bactérias da espécie
Enterobacter cloacae são frequentemente isoladas de produtos lácteos, mas estudos de
fatores de virulência e de seu potencial MDR são escassos.
6.1 PERFIL DE RESISTÊNCIA AOS ANTIMICROBIANOS
Os cinquenta e oito isolados expressaram resistência a, pelo menos, um antibiótico
dos 17 testados. Os isolados expressaram maior resistência (78%) ao antibiótico cefalotina.
Todos os isolados da espécie Serratia marcescens expressaram resistência a este
antibiótico. Já no complexo Enterobacter cloacae, apenas o isolado Q2R2 foi sensível a este
antibiótico, assim como o isolado E11 (Raoutella planticola).
Todos os isolados do gênero Raoutella planticola e do gênero Klebsiella
expressaram resistência à ampicilina. A maior parte dos isolados (85%) da espécie Serratia
marcescens também expressaram resistência a este antibiótico, assim como 48% dos
isolados do complexo Enterobacter cloacae.
Todos os 58 isolados foram sensíveis aos antibióticos tetraciclina, meropenem e
cloranfenicol.
No presente trabalho, 24 isolados foram considerados multirresistentes a drogas: 12
(50%) do complexo Enterobacter cloacae; 11 (46%) da espécie Serratia marcescens; 1 (4%)
da espécie Raoutella planticola. Para ser considerada multirresistente (MDR, de multidrug
resistant), uma bactéria deve ser resistente a um antibiótico de, pelo menos, três classes
diferentes (HEIZMANN et al., 2013). Das bactérias MDR encontradas no trabalho, a
quantidade de antibióticos ao qual as mesmas apresentaram resistência oscilou entre 3 e 4
em, pelo menos, 3 classes diferentes, porém a bactéria E8 (Serratia marcescens)
apresentou resistência a 6 antibióticos distribuídos em 5 classes diferentes. Os resultados

25
estão dispostos na Tabela 5.
Tabela 5: Perfil de resistência aos antimicrobianos.
Identificação Isolados Resistência
Complexo Enterobacter cloacae
Q1C2* CEP / AMC / AMP
Q1C3 CEP / AMC
Q1L2* CEP / NOR / AMC / AMP / TRI
Q1R1 CEP / AMC
Q1R2 CEP / AMC
Q1R4* CEP / AMC / AMP
Q2E1* CEP / AMC / AMP / TOB
Q2L1* IPM / CEP / AMC / TOB
Q2M1* CEP / CTX / AMP
Q2R2 AMC / AMP
Q2R3* IPM / CEP / AMC
Q11 CEP / AMC
Q12 CEP / AMC
Q15 CEP / AMC
Q16 CEP / AMC
R4 CEP / STR
E1* CEP / AMC / AMP
E2* CEP / AMC / AMP
E3* CEP / AMC / AMP
L3* CEP / AMC / AMP
BLL1* ATM / CEP / CAZ
Q1E2* CEP / AMC / AMP
R7 CEP
R9 CEP / STR
R19 CEP
E4* CEP / AMC / AMP
E5* CEP / AMC / AMP
E6* CEP / AMC / AMP / STR
E7* CEP / AMC / AMP / STR
E8* CEP / CAZ / AMC / AMP / AMI / TOB
E9* CEP / AMC / AMP / GEN
Serratia marcescens E10 CEP / AMP
E12 CEP / AMP
E13 CEP / AMP
E14* CEP / AMP / TOB
E15 CEP / AMP
E16 CEP / AMP
E17* CEP / AMC / AMP / STR
E19 CEP / AMP
E20* CEP / CIP / AMC / AMP
L6* CEP / AMC / AMP
Q1L1* CTX / AMP / GEN
Q1R3 AMP
Raoutella planticola Q2E2 AMP
Q2E3 AMP / GEN / TOB
Continua

26
Continuação da Tabela 5
Q2R4 AMP
QJ3 AMP
QJ5 AMP
Raoutella sp E11 AMP
Q1M1 AMP
Q1M3 CEP
Citrobacter freundii Q1R5 CEP / AMC
Q2R1 CEP / AMP
Citrobacter braakii Q1C1 CEP
Q1E1 TRI
Klebsiella pneumoniae Q2C3 AMP
R2 AMP
Klebsiella oxytoca Q1E3 AMP
Legenda: AMC, amoxicilina + ácido clavulânico; AMI, amicacina; AMP, ampicilina; ATM, aztreonam; CAZ, ceftazidima; CEP, cefalotina; CIP, ciprofloxacina; CTX, cefotaxima; GEN, gentamicina; IPM, imipenem; MER, meropenem; NOR, norfloxacina; STR, estreptomicina; TOB, tobramicina; TRI, trimetoprim; *Isolados multirresistentes (MDR).
Merece destaque, neste trabalho, o complexo Enterobacter cloacae, que apresentou
12 representantes multirresistentes (50%). Segundo Davin-Regli e Pagès (2015), neste
complexo, os principais fatores responsáveis por conferirem maior resistência aos
antibióticos são a permeabilidade seletiva da membrana celular, somados à capacidade de
adquirir componentes móveis, que os permite colonizarem uma diversidade maior de áreas
nos organismos infectados, além da facilidade de alterar seu metabolismo para adaptar-se
às pressões ambientais.
Fakruddin e colaboradores (2014), em amostras de alimentos, dentre elas leite em
pó, encontraram isolados multirresistentes dos gêneros Enterobacter (3); Citrobacter (3) e
Klebsiella (2). Destes oito isolados apenas dois do gênero Citrobacter foram sensíveis ao
antibiótico ampicilina. No nosso estudo, dos cinco isolados do gênero Citrobacter
encontrados, apenas um apresentou resistência à ampicilina. Já os três isolados do gênero
Klebsiella encontrados foram resistentes à ampicilina. A maior parte (52%) dos isolados do
complexo Enterobacter cloacae do nosso estudo expressaram-se sensíveis ao antibiótico
em questão. Além disso, todos os oito isolados do estudo de Fakruddin e colaboradores
(2014) expressaram resistência ao antibiótico imipenem, enquanto que no presente trabalho
apenas dois isolados (complexo Enterobacter cloacae: Q2L1 e Q2R3) foram resistentes a
este antibiótico.
Damaceno e colaboradores (2015), em amostras de queijo minas frescal,
conseguiram isolar onze representantes da família Enterobacteriaceae, sendo dois isolados
do gênero Enterobacter. Estes expressaram resistência a quatro antibióticos de classes
diferentes, assim como os isolados Q2E1 e Q2L1 (complexo Enterobacter cloacae) do

27
presente estudo.
Em outro trabalho, Chauhan e colaboradores (2013) conseguiram isolar 27 bactérias
da espécie Klebsiella pneumoniae, a partir de 100 amostras de leite cru, sendo que 26
destas foram multirresistentes, diferente do presente estudo, onde os dois isolados desta
espécie (Q2C3 e R2) expressaram resistência somente à ampicilina, sendo sensíveis aos
outros dezesseis antibióticos utilizados.
No presente trabalho, obteve-se um índice alto de estirpes MDR, levando em
consideração a amostragem utilizada e o tipo de alimento (pronto para consumo). Assim,
decidiu-se investigar a expressão de outros fenótipos importantes relacionados à
resistência, como o KPC e o ESBL.
6.1.1 Detecção dos fenótipos de ESBL e KPC
Os meios de cultura cromogênicos CHROMagar KPC® e CHROMagar ESBL® são
considerados bons e rápidos indicadores/confirmadores da expressão de fenótipos de
resistência, facilitando a identificação e a triagem de estirpes resistentes (RAMESHKUMAR
et al., 2015; HORNSEY et al., 2013; SAITO et al., 2010).
Os vinte e quatro isolados multirresistentes foram submetidos aos testes com os
meios cromogênicos. Seis isolados, sendo um do complexo Enterobacter cloacae e cinco da
espécie Serratia marcescens, expressaram fenótipo de susceptibilidade reduzida aos
carbapenens (KPC), e quatorze isolados (4 do complexo Enterobacter cloacae e 10 da
espécie Serratia marcescens) expressaram fenótipos de produção de β-lactamase de
espectro estendido (ESBL). Os resultados estão dispostos na Tabela 6.
Apesar de a detecção dos fenótipos KPC e/ou ESBL ter sido evidenciada nos testes
com os meios cromogênicos, apenas o isolado E8 (Serratia marcescens) expressou fenótipo
de produção de ESBL e resistência a um dos antibióticos da classe das cefalosporinas de 3ª
geração (ceftazidima), concomitantemente, podendo sugerir uma relação direta deste
fenótipo com o antibiótico testado, o que não ocorreu com os demais isolados. Isto sugere
que estes isolados possam ser resistentes a outro(s) antibiótico(s) das classes estudadas,
porém não testado(s) neste trabalho. Situação inversa ocorreu com os isolados Q2L1 e
Q2R3 (complexo Enterobacter cloacae), que expressaram, no antibiograma, resistência ao
antibiótico IPM, da classe dos carbapenens, e os isolados Q2M1 e BLL1, que expressaram
resistência aos antibióticos da classe das cefalosporinas de 3ª geração cefotaxima e
ceftazidima, mas não expressaram fenótipos KPC e ESBL, respectivamente. Além disso,
90% dos representantes do complexo Enterobacter cloacae foram resistentes à combinação
de antibióticos inibidores de β-lactamase amoxicilina- ácido clavulânico.
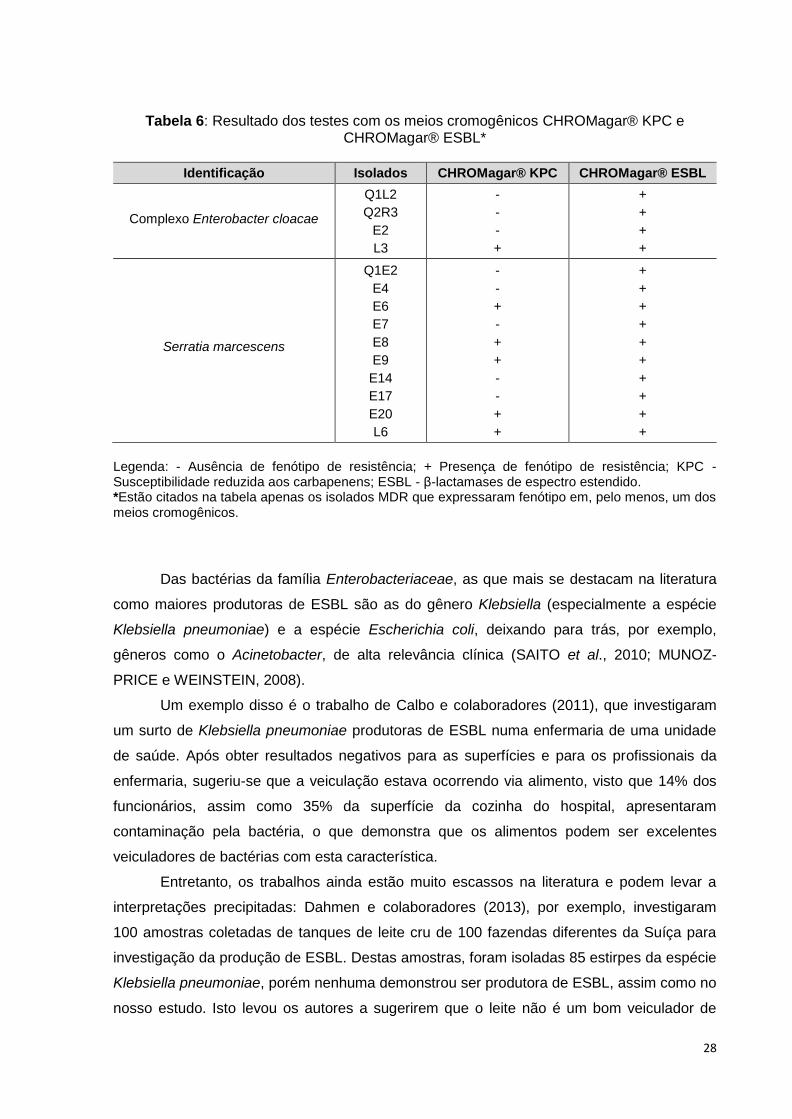
28
Tabela 6: Resultado dos testes com os meios cromogênicos CHROMagar® KPC e CHROMagar® ESBL*
Identificação Isolados CHROMagar® KPC CHROMagar® ESBL
Complexo Enterobacter cloacae
Q1L2
Q2R3
E2
L3
-
-
-
+
+
+
+
+
Serratia marcescens
Q1E2
E4
E6
E7
E8
E9
E14
E17
E20
L6
-
-
+
-
+
+
-
-
+
+
+
+
+
+
+
+
+
+
+
+
Legenda: - Ausência de fenótipo de resistência; + Presença de fenótipo de resistência; KPC - Susceptibilidade reduzida aos carbapenens; ESBL - β-lactamases de espectro estendido. *Estão citados na tabela apenas os isolados MDR que expressaram fenótipo em, pelo menos, um dos meios cromogênicos.
Das bactérias da família Enterobacteriaceae, as que mais se destacam na literatura
como maiores produtoras de ESBL são as do gênero Klebsiella (especialmente a espécie
Klebsiella pneumoniae) e a espécie Escherichia coli, deixando para trás, por exemplo,
gêneros como o Acinetobacter, de alta relevância clínica (SAITO et al., 2010; MUNOZ-
PRICE e WEINSTEIN, 2008).
Um exemplo disso é o trabalho de Calbo e colaboradores (2011), que investigaram
um surto de Klebsiella pneumoniae produtoras de ESBL numa enfermaria de uma unidade
de saúde. Após obter resultados negativos para as superfícies e para os profissionais da
enfermaria, sugeriu-se que a veiculação estava ocorrendo via alimento, visto que 14% dos
funcionários, assim como 35% da superfície da cozinha do hospital, apresentaram
contaminação pela bactéria, o que demonstra que os alimentos podem ser excelentes
veiculadores de bactérias com esta característica.
Entretanto, os trabalhos ainda estão muito escassos na literatura e podem levar a
interpretações precipitadas: Dahmen e colaboradores (2013), por exemplo, investigaram
100 amostras coletadas de tanques de leite cru de 100 fazendas diferentes da Suíça para
investigação da produção de ESBL. Destas amostras, foram isoladas 85 estirpes da espécie
Klebsiella pneumoniae, porém nenhuma demonstrou ser produtora de ESBL, assim como no
nosso estudo. Isto levou os autores a sugerirem que o leite não é um bom veiculador de

29
bactérias produtoras de ESBL. Mas, esta sugestão contrasta com Chauhan e colaboradores
(2013), por exemplo, aonde num estudo, também com 100 amostras de leite cru, coletados
de fazendas em um vale da Índia, os autores conseguiram isolar 27 estirpes de Klebsiella
pneumoniae, e todas se apresentaram como produtoras de ESBL. Também contrasta com o
trabalho de Sudarwanto e colaboradores (2015), em um estudo realizado em 80 tanques de
leite cru de fazendas na Indonésia, quando 7 isolados produtores de ESBL da espécie
Klebsiella pneumoniae foram confirmados fenotipicamente. Vários fatores podem ter
influenciado para esta divergência de resultados entre o primeiro e os dois outros estudos,
como questões socioeconômicas (mão-de-obra, controle de qualidade, questões
infraestruturais), legislativas/normativas e até mesmo culturais. No presente estudo, apesar
dos isolados do gênero Klebsiella não expressarem fenótipos ESBL, e, de se trabalhar com
um número menor de amostras de leite, foram isoladas, nestas amostras, onze bactérias
com esta característica (2 do complexo Enterobacter cloacae e 9 da espécie Serratia
marcescens), demonstrando que o leite pode ser considerado um bom veiculador de
bactérias produtoras de ESBL.
Nesta última década, porém, o gênero Enterobacter vem surgindo como um fator
preocupante, principalmente a espécie Enterobacter cloacae, pertencente ao complexo
Enterobacter cloacae, que começa a se destacar neste grupo e já aparece como a terceira
maior produtora de ESBL da família Enterobacteriaceae, segundo Arnaud e colaboradores
(2014).
6.2 AVALIAÇÃO DA PRODUÇÃO DE FATORES DE VIRULÊNCIA
6.2.1 Detecção qualitativa da produção de biofilme
Dos 58 micro-organismos isolados, 54 expressaram a produção de biofilme (93%),
sendo: 20 (37%) do complexo Enterobacter cloacae, 17 (31%) da espécie Serratia
marcescens, 8 (15%) da espécie Raoutella planticola, 3 (5%) da espécie Citrobacter freundii,
2 (4%) da espécie Klebsiella pneumoniae, 2 (4%) da espécie Citrobacter braakii, 1 (2%) da
espécie Klebsiella oxytoca, 1 (2%) do gênero Raoutella. Apenas quatro isolados (R4, R7, R9
e R19) não expressaram produção de biofilme. Esses resultados estão apresentados na
Figura 3. A Figura 4 ilustra um isolado que expressou produção de biofilme.

30
20
17
8
3
2
2
1
1
0 5 10 15 20 25
Complexo Enterobacter cloacae
Serratia marcescens
Raoutella planticola
Citrobacter freundii
Citrobacter braakii
Klebsiella pneumoniae
Klebsiella oxytoca
Raoutella sp
Número de isolados
Ide
ntifica
çã
o
Figura 3: Número de isolados produtores de biofilme.
Figura 4: Exemplo de isolado produtor de biofilme em ágar vermelho Congo (Q1L2;
complexo Enterobacter cloacae)
Outros trabalhos relataram membros da família Enterobacteriaceae produtores de
biofilme e associados à produtos lácteos. Cherif-Antar e colaboradores (2016) encontraram,
aderidas às superfícies de aço inoxidável das tubulações de uma fábrica de processamento
de leite, 32 bactérias Gram-negativas: 13 pertenciam à espécie Klebsiella pneumoniae, 6 à
espécie Serratia marcescens, 5 ao gênero Enterobacter.
Malek e colaboradores (2012) realizaram coleta de amostras de 5 fazendas
produtoras de leite pasteurizado e de leite em pó desnatado na Argélia. Toda a linha de
produção passava por processos constantes de sanitização com produtos à base de
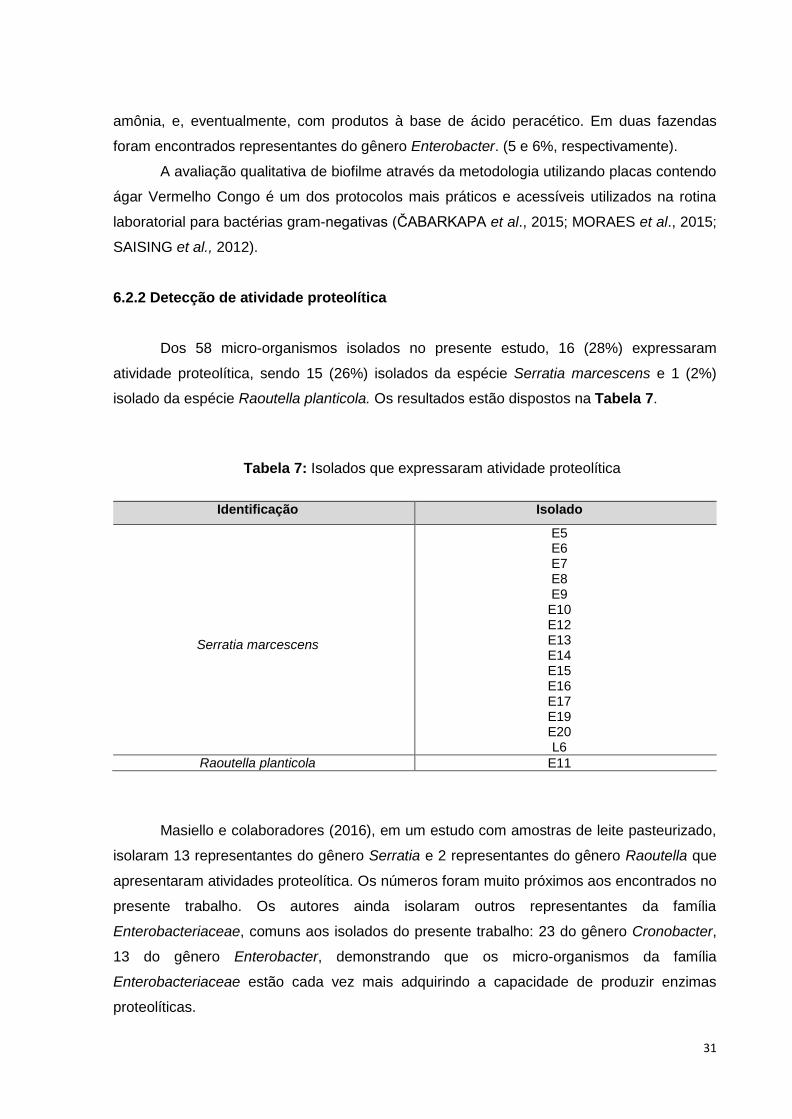
31
amônia, e, eventualmente, com produtos à base de ácido peracético. Em duas fazendas
foram encontrados representantes do gênero Enterobacter. (5 e 6%, respectivamente).
A avaliação qualitativa de biofilme através da metodologia utilizando placas contendo
ágar Vermelho Congo é um dos protocolos mais práticos e acessíveis utilizados na rotina
laboratorial para bactérias gram-negativas (ČABARKAPA et al., 2015; MORAES et al., 2015;
SAISING et al., 2012).
6.2.2 Detecção de atividade proteolítica
Dos 58 micro-organismos isolados no presente estudo, 16 (28%) expressaram
atividade proteolítica, sendo 15 (26%) isolados da espécie Serratia marcescens e 1 (2%)
isolado da espécie Raoutella planticola. Os resultados estão dispostos na Tabela 7.
Tabela 7: Isolados que expressaram atividade proteolítica
Identificação Isolado
Serratia marcescens
E5 E6 E7 E8 E9 E10 E12 E13 E14 E15 E16 E17 E19 E20 L6
Raoutella planticola E11
Masiello e colaboradores (2016), em um estudo com amostras de leite pasteurizado,
isolaram 13 representantes do gênero Serratia e 2 representantes do gênero Raoutella que
apresentaram atividades proteolítica. Os números foram muito próximos aos encontrados no
presente trabalho. Os autores ainda isolaram outros representantes da família
Enterobacteriaceae, comuns aos isolados do presente trabalho: 23 do gênero Cronobacter,
13 do gênero Enterobacter, demonstrando que os micro-organismos da família
Enterobacteriaceae estão cada vez mais adquirindo a capacidade de produzir enzimas
proteolíticas.

32
Hantsis-Zacharov e Halpern (2007) conseguiram isolar, em amostras de leite cru de 4
fazendas do norte de Israel, 2 bactérias da espécie Serratia marcescens que expressaram
atividade proteolítica. Em outro trabalho, Teh e colaboradores (2011a), a partir de amostras
retiradas da superfície interna de tanques de leite, isolaram 17 representantes do gênero
Serratia que apresentaram produção de enzimas proteolíticas. No presente estudo, 75% dos
isolados da espécie Serratia marcescens expressaram atividade proteolítica.
No trabalho de Nörnberg e colaboradores (2010), a partir de amostras de leite cru,
foram isolados 2 representantes da espécie Klebsiella oxytoca com alta capacidade
proteolítica. Amostras de leite UHT deste trabalho, quando expostas a estas cepas, em
temperatura ambiente, coagularam após 5 dias de exposição. Oladipo e Omo-Adua (2011a)
conseguiram obter, a partir de 10 amostras de leite em pó enlatado, um isolado da espécie
Klebsiella pneumoniae que expressou considerável atividade proteolítica. No presente
estudo, nenhum dos isolados do gênero Klebsiella expressou atividade proteolítica, porém
merece destaque o isolado E11 (Raoutella planticola), que até a alguns anos pertencia ao
gênero Klebsiella e expressou atividade proteolítica.
Bactérias com a capacidade de produzir enzimas proteolíticas podem ser
responsáveis pela degradação do leite e seus derivados, mesmo em produtos que sofrem
processamento térmico (MASIELLO et al., 2016), visto que muitas destas enzimas são
termorresistentes (ZAJÁC et al., 2015). Chove e colaboradores (2013), por exemplo,
analisaram a atividade proteolítica em amostras de leite cru após processamento térmico
nas temperaturas de 85, 110, 120, 130 e 142° C. Os autores evidenciaram nas amostras
que sofreram processamento térmico até 85° C/ 15 segundos e nas amostras sem
processamento, considerável atividade proteolítica. Nas demais temperaturas as enzimas
foram inativadas.
O exemplo mais prático da ação de proteinases sobre a caseína, influenciando
negativamente os produtos lácteos, está na produção de queijos: as proteases podem
desestabilizar as micelas de caseína, e, quando isto ocorre, o leite não coagula,
consequentemente o produto não adquire a consistência padrão (CALDERA et al., 2015;
PINTO et al., 2007).
Além disso, bactérias que sintetizam proteinases afetam consideravelmente as
propriedades sensoriais dos alimentos como, a cor, o odor, o sabor, e a textura (CALDERA
et al., 2015; HANTSIS-ZACHAROV e HALPERN, 2007), o que pode acarretar na rejeição do
produto pelo consumidor (TEIXEIRA, 2009).
Por outro lado, nesta última década, surgiu um grande interesse nas pesquisas
visando à utilização de proteinases na promoção de hidrólise da caseína para a fabricação
de alimentos com propriedades funcionais e farmacêuticas, como os nutracêuticos (LI et al.,
2013; SALAMI et al., 2011; PHELAN e KERINS, 2011). Mao e colaboradores (2011), por

33
exemplo, demostraram que produtos fabricados através de caseína hidrolisada por
proteases possuem capacidade de liberar peptídeos bioativos que atuam como
antioxidantes, entre outros benefícios à saúde. Um estudo de Muhammad e colaboradores
(2016) sugeriu, ainda, que estes peptídeos podem atuar no transporte de cálcio, melhorando
a sua biodisponibilidade.
6.2.3 Detecção de atividade lipolítica
Dos 58 micro-organismos isolados no presente estudo, 20 (34%) expressaram
atividade lipolítica, sendo 18 (31%) da espécie Serratia marcescens, 1 (2%) do complexo
Enterobacter cloacae, 1 (2%) da espécie Raoutella planticola. Os resultados estão dispostos
na Tabela 8.
Tabela 8: Isolados que expressaram atividade lipolítica.
Identificação Isolado
Serratia marcescens
E4 E5 E6 E7 E8 E9 E10 E12 E13 E14 E15
E16 E17 E19 E20 R7 R9 R19
Complexo Enterobacter cloacae Q1R4
Raoutella planticola E11
A Figura 5 ilustra uma placa com meio de cultura Spirit Blue apresentando 4 isolados
da espécie Serratia marcescens que expressaram atividade lipolítica.

34
Figura 5: Exemplos de isolados que apresentaram atividade lipolítica
Masiello e colaboradores (2016), em amostras de leite pasteurizado, conseguiram
isolar alguns representantes da família Enterobacteriaceae, comuns ao presente estudo,
que expressaram atividade lipolítica no meio ágar Spirit blue: 17 isolados do gênero Serratia,
14 do gênero Enterobacter e 1 do gênero Raoutella, números muito próximos ao do
presente estudo. Apesar de nenhum dos isolados do gênero Citrobacter do presente estudo
ter expressado atividade lipolítica, vale ressaltar que no estudo de Masiello e colaboradores
(2016) 5 representantes expressaram.
Se por um lado as bactérias que expressam atividade lipolítica podem gerar
prejuízos à indústria de produtos lácteos com a deterioração de seus produtos, por outro, há
a possibilidade da utilização destas lipases como catalisadoras nos processos de
produção de biodiesel (WINAYANUWATTIKUN et al., 2011), lipases estas obtidas através
de fontes como os resíduos sólidos das indústrias, que são ricos em gorduras, promovendo,
assim, destinação a estes resíduos (SALIHU et al., 2012).
6.2.4 Detecção de atividade hemolítica
Dos 58 micro-organismos isolados no presente estudo, 42 (72%) expressaram
atividade hemolítica, sendo 20 (35%) do complexo Enterobacter cloacae, 8 (14%) da
espécie Serratia marcescens, 6 (10%) da espécie Raoutella planticola, 3 (5%) da espécie
Citrobacter freundii, 2 (3%) da espécie Citrobacter braakii, 2 (3%) da espécie Klebsiella
pneumoniae e 1 (2%) da espécie Klebsiella oxytoca. Os resultados estão dispostos na
Tabela 9. A Figura 6 ilustra a atividade hemolítica de alguns isolados.

35
Tabela 9: Isolados que expressaram atividade hemolítica.
Identificação Isolado Tipo de hemólise
Complexo Enterobacter cloacae
BLL1 E1 E3 L3
Q11 Q12 Q15 Q16
Q1C2 Q1C3 Q1L2 Q1R1 Q1R2 Q1R4 Q2E1 Q2L1 Q2M1 Q2R2 Q2R3
R4
α α α α α α β α α α α α
α α α α α α β β
Serratia marcescens
E7 E13 E14 E15 E17 E19 L6
R19
α α α α α α α α
Raoutella planticola
QJ3 QJ5
Q1R3
α α α
Q2E2 Q2E3 Q2R4
α α α
Citrobacter freundii
Q1M3 Q1R5 Q2R1
α α α
Citrobacter braakii Q1C1 Q1E1
α α
Klebsiella pneumoniae Q2C3
R2 α α
Klebsiella oxytoca Q1E3 α
Rajani e colaboradores (2016), em amostras de leite cru, conseguiram isolar um
representante da espécie Enterobacter sakazakii que expressou atividade β-hemolítica. No
presente estudo, três isolados do complexo Enterobacter cloacae expressaram atividade β-
hemolítica. Até a publicação do artigo de Rajani e colaboradores (2016) não havia, na
literatura, trabalhos relacionando bactérias hemolíticas a produtos lácteos.

36
Figura 6: Exemplos de isolados que apresentaram atividade hemolítica. À esquerda, α-
hemólise apresentada pelo Q1E3 (Klebsiella oxytoca) e à direita, β-hemólise apresentada
pelo isolado R4 (do complexo Enterobacter cloacae).
6.3 AVALIAÇÃO DA PRODUÇÃO DE SUBSTÂNCIAS ANTIMICROBIANAS (SAM)
Nenhum dos micro-organismos isolados neste trabalho produziu algum tipo de SAM
contra as estirpes indicadoras utilizadas. É importante ressaltar que os micro-organismos
produtores de SAM possuem uma grande vantagem na competição em um determinado
nicho ecológico.
Diante disto, a indústria está começando a demonstrar interesse no potencial de
prospecção biotecnológica dos micro-organismos produtores de SAM. Segundo Oliveira e
colaboradores (2011a), como os antibióticos não podem ser utilizados nos alimentos, estas
substâncias podem ser de suma importância na prevenção e controle de contaminantes em
alimentos. Em um trabalho anterior realizado pelo nosso grupo de pesquisa, por exemplo,
dois representantes do gênero Enterobacter e outros nove representantes da família
Enterobacteriaceae foram capazes de produzir substâncias antimicrobianas contra as cepas
E. coli ATCC25922 e S. enterica ATCC19214 utilizadas como indicadoras da produção de
SAM (DAMACENO et al., 2015). Estas duas espécies de bactérias estão entre as maiores
causadoras de doenças transmitidas por alimentos (FLEMING et al., 2010).
6.4 DETERMINAÇÃO DO PERFIL PLASMIDEAL DOS ISOLADOS DO COMPLEXO
ENTEROBACTER CLOACAE
Por constituírem a maior parte das bactérias detectadas neste trabalho, todos os
isolados do complexo Enterobacter cloacae foram submetidos à extração de DNA e
eletroforese em gel de agarose para a determinação dos seus perfis

37
plasmideais. A maioria dos isolados possuía mais de uma forma plasmideal. Alguns isolados
não MDR expressaram o mesmo perfil plasmideal dos isolados MDR e, além disso, as suas
formas plasmideais possuíam tamanho molecular entre 2 e 10 Kb. Alguns representantes
estão ilustrados na Figura 7.
Figura 7: Eletroforese em gel de agarose com alguns isolados MDR e não MDR do
complexo Enterobacter cloacae. A, Supercoilled; B, Q1C2; C, Q1C3; D, Q1L2; E, Q2M1; F,
Q1R1; G, Q1R2; H, Q2L1; I, Q2R3; J, L3; Os asteriscos indicam os isolados MDR.
Excetuando-se o isolado Q2R2, todos os demais isolados com o perfil plasmideal
investigado expressaram resistência ao antimicrobiano CEP, porém nem todos os isolados
apresentaram formas plasmideais semelhantes. Já a resistência ao antimicrobiano AMC só
não ocorreu nos isolados Q2M1, BLL1 e Q2R2, mas também não apresentaram formas
plasmideais semelhantes (dados não mostrados). Por outro lado, os isolados Q2L1 e Q2R3
expressaram resistência ao antimicrobiano IPM e apresentaram algumas formas
plasmideais com tamanhos moleculares muito semelhantes. Entretanto, segundo Huang e

38
colaboradores (2012), os plasmídeos que contribuem para a resistência aos antimicrobianos
costumam ser de grande peso molecular. Davin-Regli e Pagès (2015), por exemplo,
relataram que micro-organismos da espécie Enterobacter aerogenes integraram um gene de
origem cromosssômica em um plasmídeo de grande tamanho (130 kb).
Em um trabalho realizado por nosso grupo, Damaceno e colaboradores (2015)
conseguiram isolar um representante da espécie Enterobacter cloacae a partir de uma
amostra de queijo minas frescal, e este expressou várias formas plasmidiais de tamanho
inferior a 6 kb, semelhante a alguns dos isolados do complexo Enterobacter cloacae do
presente trabalho. Vale ressaltar que o isolado do trabalho de 2015 também apresentava
resistência ao imipenem, assim como o isolado Q2L1.
Os resultados apontam que não existe uma fórmula padrão de relação entre o peso
molecular de um plasmídeo e sua ligação com determinada característica em um gênero
bacteriano como, por exemplo, a produção de substâncias antimicrobianas (SAM). Içgen e
colaboradores (2002) compartilham da ideia que as bactérias com plasmídeos de alto peso
molecular tendem a possuir um grau maior de patogenicidade que bactérias com
plasmídeos de baixo peso molecular, mas os próprios autores também relataram que isto
não é uma regra. A única regra é a de que, em bactérias sem plasmídeos, os genes
responsáveis por conferir resistência a antibióticos e outros fatores de virulência encontram-
se no cromossomo. A confirmação da associação de um plasmídeo com determinada
característica somente seria possível de ser comprovada através de experimentos de cura
plasmidial e/ou de transferência gênica.

39
7 CONCLUSÕES
Neste trabalho, pode-se considerar que as amostras de conveniência dos produtos
lácteos investigados são potenciais veículos de micro-organismos da família
Enterobacteriaceae que:
Podem expressar multirresistência aos antimicrobianos disponíveis, visto que 24
isolados expressaram resistência a, pelo menos, 1 antibiótico em 3 classes
diferentes. Vale ressaltar que os antimicrobianos usados no combate a infecções
causadas por estas bactérias estão cada vez mais escassos e a busca por novos
compostos não segue o mesmo ritmo da aquisição de resistência aos antibióticos por
estes micro-organismos;
Podem expressar fenótipos de susceptibilidade reduzida ao carbepenens (KPC) e
fenótipos de produção de β-lactamase de espectro estendido (ESBL): estes fenótipos
conferem resistência as principais classes de antibióticos utilizados no tratamento de
infecções causadas por bactérias Gram-negativas. Nesta última década, a espécie
Enterobacter cloacae, pertencente ao complexo Enterobacter cloacae, está se
destacando como a terceira maior produtora de ESBL da família Enterobacteriaceae.
Podem produzir biofilme: uma das características mais preocupantes para a indústria
de alimentos, pois bactérias com os perfis encontrados, aderidos às superfícies das
cadeias produtivas, podem trazer muitos prejuízos;
Podem produzir enzimas proteolíticas: capazes de gerar a deterioração dos produtos
lácteos e/ou causar danos às propriedades sensoriais dos alimentos, induzindo à
rejeição do produto por parte do consumidor. Além disso, podem propiciar a
sobrevivência de muitos patógenos dentro do organismo hospedeiro, logo podem
trazer sérios riscos à saúde dos consumidores;
Podem produzir enzimas lipolíticas: estas enzimas podem levar a um processo
denominado rancidez hidrolítica, que desenvolve no produto um sabor acre e um
odor desagradável. Podem, ainda, estar relacionadas à formação de tumores e
doenças coronarianas nos consumidores.
Podem produzir enzimas hemolíticas: estas enzimas podem provocar doenças nas
glândulas mamárias de animais leiteiros, e, possível contaminação do leite extraído.
No organismo hospedeiro, são capazes de afetar a sinalização celular, os
mecanismos de modulação da morte celular induzida, as vias de sinalização
inflamatória e levar à carência nutricional de ferro, já que atuam no sequestro de
ferro.

40
Apesar de nenhum dos isolados ter produzido substâncias antimicrobianas contra as
cepas indicadoras utilizadas, é importante ressaltar que esta característica é uma vantagem
para as bactérias em um nicho de competição.
O perfil plasmideal dos isolados não foi suficiente para associar determinadas
características às formas plasmidiais expressas, e, isso somente seria possível de ser
comprovado através de experimentos de cura plasmidial e/ou de transferência gênica.
Todos estes fatores representam riscos potenciais para a saúde dos consumidores
de produtos lácteos, especialmente os consumidores imunocomprometidos, situação que
vem se agravando na sociedade atual.
Diante do exposto, cabe a toda cadeia produtiva de produtos lácteos, em todas as
etapas (linhas de produção, transporte e armazenamento), seguirem as Boas Práticas de
Fabricação e Higiene, assim como boas práticas nos locais de comercialização,
principalmente para produtos consumidos sem qualquer processamento prévio. Além disso,
de acordo com a legislação vigente no Brasil, que preconiza os patógenos que devem ser
analisados e seus limites nos produtos lácteos [RDC nº 12 de 2001 (BRASIL, 2001)], apenas
os coliformes termotolerantes e o gênero Salmonella são destacados. Diante dos achados
deste trabalho, sugere-se uma revisão dos grupos de micro-organismos e dos limites de
aceitação dessa resolução.

41
8 CONSIDERAÇÕES FINAIS
A presente dissertação gerou algumas produções para o nosso grupo de pesquisa do
Laboratório de Microbiologia, dentre estas: quatro comunicações em eventos, a publicação
de um artigo em revista internacional, a submissão de outro artigo para outra revista
internacional e um manuscrito de artigo, conforme descritos no item 9 (Produção Científica).
O manuscrito depende da realização de alguns testes complementares, que estão
ocorrendo, porém com previsão de serem finalizados somente após a defesa desta
dissertação. Os testes complementares são: a exposição dos isolados ao hipoclorito de
sódio para avaliar a resistência destes a um sanitizante amplamente utilizado para a
desinfecção de alimentos e a exposição ao suco gástrico simulado (SGS), mimetizando a
passagem dos isolados pelo estômago.

42
9 PRODUÇÃO CIENTÍFICA
9.1 ARTIGO ACEITO PARA PUBLICAÇÂO (ANEXO 1)
AMORIM, A. M. B. & NASCIMENTO, J. S. Acinetobacter: an underrated foodborne
pathogen? The Journal of Infection in Developing Countries, 2016. In press.
9.2 SUBMISSÃO DE ARTIGO: INTERNATIONAL FOOD RESEARCH JOURNAL (ANEXO 2)
AMORIM, A. M. B., MELO, D. H., SOUZA, B. V., MEDEIROS, L. M.; MATTOSO, J.
M. V.; NASCIMENTO, J. S. A reddish problem: antibiotic-resistant Serratia marcescens in
dairy food commercialized in Rio de Janeiro.
9.3 MANUSCRITO EM PREPARAÇÂO
AMORIM, A. M. B., ARAÚJO, B. C. A., MELO, D. H., FRAZÃO, A. M.; COSTA, L. V.;
NASCIMENTO, J. S. Multidrug resistance and viability in simulated gastric fluid exhibited by
Enterobacter cloacae complex isolated from dairy products.
9.4 COMUNICAÇÔES EM EVENTOS (ANEXOS 3 E 4)
AMORIM, A. M. B.; ARAÚJO, B. C.; YANG, E.; SOUZA, B. V.; NASCIMENTO, J. S.
Avaliação da significância de produtos lácteos como potenciais fontes de
transmissão de Acinetobacter spp. multirresistentes. XXXV Semana da Química - Rio
de Janeiro - Produzindo Ciências há 450 anos / VIII Jornada da Pós-Graduação em
Alimentos. Instituto Federal do Rio de Janeiro / Campus Rio de Janeiro – 19 a 24 de outubro
de 2015. Disponível em: http://www.ifrj.edu.br/sites/default/files/webfm/images/JPAL-15.pdf
AMORIM, A. M. B.; MELO, D. H.; ARAÚJO, B. C.; SOUZA, B. V.; NEVES, H. V. M.;
NASCIMENTO, J. S. Avaliação do papel de produtos lácteos como possíveis veículos
de transmissão de bactérias multirresistentes da família Enterobacteriaceae. X
Jornada Interna de Iniciação Científica e Tecnológica” (X JIT) / V Fórum de Inovação,
Tecnologia e Educação” (V Fórum ITE). Instituto Federal do Rio de Janeiro / Campus
Nilópolis – 21 e 22 de junho de 2016.
AMORIM, A. M. B.; MELO, D. H.; SOUZA, B. V.; COSTA, L. V., FRAZÃO, A. M.;

43
NASCIMENTO, J. S. Avaliação da multirresistência a drogas por isolados do complexo
Enterobacter cloacae obtidos de produtos lácteos comercializados no Rio de Janeiro.
IV Seminário em Inovação e Tecnologia na área de Alimentos. Instituto Federal do Rio de
Janeiro / Campus Rio de janeiro – 06 e 07 de outubro de 2016.
AMORIM, A. M. B.; ARAÚJO, B. C.; SOUZA, B. V.; NASCIMENTO, J. S. Avaliação
da produção de enzimas proteolíticas, lipolíticas e hemolíticas por isolados da
espécie Serratia marcescens obtidos de produtos lácteos comercializados no Rio de
Janeiro. IV Seminário em Inovação e Tecnologia na área de Alimentos. Instituto Federal do
Rio de Janeiro / Campus Rio de janeiro – 06 e 07 de outubro de 2016.

44
10 REFERÊNCIAS BIBLIOGRÁFICAS
ABED, B. K.; AUTHMAN, S. H.; YASSEIN, K. H. Optimization of extracellular protease extracted from Escherichia coli. European journal of pharmaceutical and medical research, v. 3, n. 2, p. 113-118, 2016. ARAÚJO, B. C.; MORAES, M. S.; COSTA, L. E. O.; NASCIMENTO, J. S. Short communication: multidrug-resistant Acinetobacter baumannii-calcoaceticus complex isolated from infant milk formula and utensils in a nursery in Rio de Janeiro, Brazil. Journal of Dairy Science, v. 98, n. 4, p. 2303-2306, 2015. ARNAUD, I., JARLIER V., Groupe de travail BMR-Raisin. Surveillance des bactéries multiresistantes dans les établissements de santé français. Données 2012. Saint-Maurice: Institut de Veille Sanitaire, 100p, 2014. ATANASSOVA, M.; FERNÁNDEZ-OTERO, C.; CENTENO, J. A.; GARABAL, J. I. Alcohol-mediated hemolysis in dairy yeast isolates and hemolytic activities on blood agar media containing milk and cheese. Journal of Food Safety, v. 35, p. 237-247, 2015. BARBER, M. F.; ELDE, N. C. Escape from bacterial iron piracy through rapid evolution of transferrin. Science, v. 346, n. 6215, p. 1362-1366, 2014. BARUZZI, F.; LAGONIGRO, R.; QUINTIERI, L.; MOREA, M.; CAPUTO, L. Occurrence of non-lactic acid bacteria populations involved in protein hydrolysis of cold-stored high moisture Mozzarella cheese. Food Microbiology, v. 30, n. 1, p. 37-44, 2012. BAUGH, S.; PHILLIPS, C. R.; EKANAYAKA, A. S.; PIDDOCK, L. J. V.; WEBBER, M. A. Inhibition of multidrug efflux as a strategy to prevent biofilm formation. The Journal of Antimicrobial Chemotherapy, v. 69, n. 3, p. 673-681, 2014. BAUR, C.; KREWINKEL, M.; KRANZ, B.; VON NEUBECK, M.; WENNING, M.; SCHERER S.; STOECKEL, M.; HINRICHS, J.; STRESSLER, T.; FISCHER, L. Quantification of the proteolytic and lipolytic activity of microorganisms isolated from raw milk. International Dairy Journal, v. 49, p. 23-29, 2015. BAYOUB, K.; MARDAD, I.; AMMAR, E.; SERRANO, A.; SOUKRI, A. Isolation and purification of two bacteriocins 3D produced by Enterococcus faecium with inhibitory activity against Listeria monocytogenes. Current Microbiology, v. 62, n. 2, p 479-485, 2011. BAYOUMI, M. A.; KAMAL, R. M.; ABD EL AAL S. F.; AWAD, E. I. Assessment of a regulatory sanitization process in Egyptian dairy plants in regard to the adherence of some food-borne pathogens and their biofilms. International Journal of Food Microbiology, v. 158, n. 3, p. 225-231, 2012.

45
BECEIRO, A.; TOMÁS, M.; BOU, G. Antimicrobial resistance and virulence: a successful or deleterious association in the bacterial world? Clinical Microbiology Reviews, v. 26, n. 2, p. 185-230, 2013. BECTON, DICKINSON AND COMPANY. Difco™ & BBL™ Manual: Manual of Microbiological Culture Media. 2 ed. Maryland, Estados Unidos da América, 2009. p. 513-514. BERG, G.; ERLACHER, A.; SMALLA, K.; KRAUSE, R. Vegetable microbiomes: is there a connection among opportunistic infections, human health and our “gut feeling”? Microbial Biotechnology, v. 7, n. 6, p. 487-495, 2014. BIRNBOIM, H. C.; DOLY, J. A rapid alkaline extraction procedure for screening recombinant plasmid DNA. Nucleic acids research, v. 7, n. 6, p. 1513-1523, 1979. BLAIR, J. M. A.; PIDDOCK, L. J. V. How to measure export via bacterial multidrug resistance efflux pumps. mBio, v. 7, n. 4, p. 1-6, 2016. BÖHME, K.; FERNÁNDEZ-NO, I. C.; BARROS-VELÁZQUEZ, J.; GALLARDO, J. M.; CALO-MATA, P.; CAÑAS, B. Species identification of food spoilage and pathogenic bacteria by MALDI-TOF mass fingerprinting. Journal of Proteome Research, v. 9, n. 6, p. 3169-3183, 2010. BOSCO, K. J.; KADDU-MULINDWA, D. H.; ASIIMWE, B. B. Antimicrobial drug resistance and plasmid profiles of Salmonella isolates from humans and foods of animal origin in Uganda. Advances in Infectious Diseases, v. 2, n. 4, p. 151-155, 2012. BRASIL. Resolução RDC nº 12, de 2 de janeiro de 2001. Aprova o Regulamento técnico sobre padrões microbiológicos para alimentos. Órgão emissor: ANVISA – Agência Nacional de Vigilância Sanitária. Disponível em: www.anvisa.com.br. Acesso em: 06 de novembro de 2016. BRIDIER, A.; SANCHEZ-VIZUETE, P.; GUILBAUD, M.; PIARD, J. C.; NAÏTALI, M.; BRIANDET, R. Biofilm-associated persistence of food-borne pathogens. Food Microbiology, v. 45 (Part B), p. 167-178, 2015. BROGAN, D. M.; MOSSIALOS, E. Incentives for new antibiotics: the Options Market for Antibiotics (OMA) model. Globalization and Health, v. 9, n. 58, p. 1-10, 2013. BURGOS, Y.; BEUTIN, L. Common origin of plasmid encoded alpha-hemolysin genes in Escherichia coli. BMC Microbiology, v. 10, n. 193, 2010.

46
CAINE, C. L.; NWODO, U.; OKOH, A.; NDIP, R. N.; GREEN, E. Occurrence of virulence genes associated with diarrheagenic Escherichia coli isolated from raw cow’s milk from two commercial dairy farms in the Eastern Cape Province, South Africa. International Journal of Environmental Research and Public Health, v. 11, n. 11, p. 11950-11963, 2014. ČABARKAPA, I.; ŠKRINJAR, M.; LEVIĆ, J.; KOKIĆ, B.; LAGOJEV, N.; MILANOV, D.; SUVAJDŽIĆ, L. Biofilm forming ability of Salmonella enteritidis in vitro. Acta Veterinaria-Beograd: The Journal of University of Belgrade, v. 65, n. 3, p. 371-389, 2015. CALBO, E.; FREIXAS, N.; XERCAVINS, M.; RIERA, M.; NICOLÁS, C.; MONISTROL, O.; DEL MAR SOLÉ, M.; SALA, M. R.; VILA, J.; GARAU, J. Foodborne nosocomial outbreak of SHV1 and CTX-M-15–producing Klebsiella pneumoniae: epidemiology and control. Clinical Infectious Diseases, v. 52, n. 6, p. 743-749, 2011. CALDERA, L.; ARIOLI, S.; STUKNYTĖ, M.; SCARPELLINI, M.; FRANZETTI, L. Setup of a rapid method to distinguish among dead, alive, and viable but not cultivable cells of Pseudomonas spp. in mozzarella cheese. Journal of Dairy Science, vol. 98, n. 2, p. 8368-8374, 2015. CARPINÉ, D.; DAGOSTIN, J. L. A.; SANTA, H. S. D.; ALVAREZ, D. C.; TERRA, N. N.; SANTA. O. R. D. Atividade proteolítica e lipolítica de bactérias lácticas isoladas de salames artesanais. Ambiência Guarapuava, v. 6, n. 1, p. 125-132, 2010. CASALINUOVO, F.; RIPPA, P.; BATTAGLIA, L.; PARISI, N. Isolation of Cronobacter spp. (Enterobacter sakazakii) from artisanal mozzarella. Italian Journal of Food Safety, v. 3, p. 1526, 2014. CASSAT, J. E.; SKAAR, E. P. Iron in Infection and Immunity. Cell Host & Microbe, v. 13, p. 509-519, 2013. CHAUHAN, S.; FAROOQ, U.; SINGH, V.; KUMAR, A. Determination of prevalence and antibacterial activity of ESBL (Extended Spectrum Beta-lactamases) producing Klebsiella species isolated from raw milk of Doon Valley in India. International Journal of Pharma and Bio Sciences, v. 4, n. 1, p.417-423, 2013. CHERIF-ANTAR, A.; MOUSSA-BOUDJEMÂA, B.; DIDOUH, S.; MEDJAHDI, K.; MAYO, B.; FLÓREZ, A. B. Diversity and biofilm-forming capability of bacteria recovered from stainless steel pipes of a milk-processing dairy plant. Dairy Science & Technology, v. 96, p. 27-38, 2016. CHOVE, L. M.; ISSA- ZACHARIA, A.; GRANDISON, A. S.; LEWIS, M. J. Proteolysis of milk heated at high temperatures by native enzymes analysed by trinitrobenzene sulphonic acid (TNBS) method. African Journal of Food Science, v. 7, n. 8, p. 232-237, 2013.

47
CLSI - Performance standards for antimicrobial susceptibility testing: twenty-fifth Informational Supplement. CLSI Document M100-S25. CLSI, Wayne, PA: Clinical and Laboratory Standards Institute, 2015. COMMITTEE ON INFECTIOUS DISEASES AND COMMITTEE ON NUTRITION. Consumption of raw or unpasteurized milk and milk products by pregnant women and children. Pediatrics, v. 133, n. 1, 2014. CORNEJO-JUÁREZ, P.; VILAR-COMPTE, D.; PÉREZ-JIMÉNEZ, C.; ÑAMENDYS-SILVA, S. S.; SANDOVAL-HERNÁNDEZ, S.; VOLKOW-FERNÁNDEZ, P. The impact of hospital-acquired infections with multidrug-resistant bacteria in an oncology intensive care unit. International Journal of Infectious Diseases, v. 31, p. 31-34, 2015. COSKUN, Y.; DEMIREL, U.; CENGIZ, M.; AKMAN, M.; AKMAN, I. Nosocomial infections due to multidrug-resistant microorganisms in neonates. Archives of Disease in Childhood, v. 99 (Suppl 2), p. A192, 2014. CRUZ, A. G.; FARIA, J. A. F.; POLLONIO, M. A. R.; BOLINI, H. M. A.; CELEGHINI, R. M. S.; GRANATO, D.; SHAH, N. G. Cheeses with reduced sodium content: effects on functionality, public health benefits and sensory properties. Trends in Food Science & Technology, v. 22, n. 6, p. 276-291, 2011. CUSATO, S.; GAMEIRO, A. H.; CORASSIN, C. H.; SANT’ANA, A. S.; CRUZ, A. G.; FARIA, J. A.; OLIVEIRA, C. A. Food safety systems in a small dairy factory: implementation, major challenges, and assessment of systems performances. Foodborne Pathogens and Disease, v. 10, n. 1, p. 6-12, 2013. DAHMEN, S.; MÉTAYER, V.; GAY, E.; MADEC, J-Y.; HAENNI, M. Characterization of extended-spectrum beta-lactamase (ESBL)-carrying plasmids and clones of Enterobacteriaceae causing cattle mastitis in France. Veterinary Microbiology, v. 162, p. 793-799, 2013. DAMACENO, H. F. B.; FREITAS JUNIOR, C. V.; MARINHO, I. L.; CUPERTINO, T. R.; COSTA, L. E. O.; NASCIMENTO, J. S. Antibiotic resistance versus antimicrobial substances production by gram-negative foodborne pathogens isolated from minas frescal cheese: heads or tails? Foodborne Pathogens and Disease, v. 12, n. 4, p. 297-301, 2015. DAVIN-REGLI, A.; PAGÈS, J-M. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Frontiers in microbiology, v. 6, p. 392, 2015. DEVAUX, I.; VARELA-SANTOS, C.; PAYNE-HALLSTRÖM, L.; TAKKINEN, J.; BOGAARDT, C.; COULOMBIER, D. Investigation of travel-related cases in a multinational outbreak: example of the Shiga-toxin producing E. coli outbreak in Germany, May-June 2011. Epidemiology and Infection, v. 143, n. 16, p. 3468-3474, 2015.

48
DOYLE, M. E.; GLASS, K. A. Sodium reduction and its effect on food safety, food quality, and human health. Comprehensive Reviews in Food Science and Food Safety, v. 9, n. 1, p. 44-56, 2010. FAKRUDDIN; RAHAMAN, M.; AHMED, M. M.; HOQUE, M. Antimicrobial resistance and virulence factors of Enterobacteriaceae isolated from food samples of Bangladesh. International Journal of Microbiology and Immunology Research, v. 3, n. 1, p. 12-18, 2014. FARROKH, C.; JORDAN, K.; AUVRAY, F.; GLASS, K.; OPPEGAARD, H.; RAYNAUD, S.; THEVENOT, D.; CONDRON, R.; DE REU, K.; GOVARIS, A.; HEGGUM, K.; HEYNDRICKX, M.; HUMMERJOHANN, J.; LINDSAY, D.; MISZCZYCHA, S.; MOUSSIEGT, S.; VERSTRAETE, K.; CERF, O. Review of Shiga-toxin-producing Escherichia coli (STEC) and their significance in dairy production. International Journal of Food Microbiology, v. 162, n. 2, p.190-212, 2013. FERNÁNDEZ, J.; MONTERO, I.; MARTÍNEZ, O.; FLEITES, A.; POIREL, L.; NORDMANN, P.; RODICIO, M. R. Dissemination of multiresistant Enterobacter cloacae isolates producing OXA-48 and CTX-M-15 in a Spanish hospital. International Journal of Antimicrobial Agents, v. 46, n. 4, p. 469-474, 2015. FLEMING, L. R.; BOLZAN, D. N.; NASCIMENTO, J. S. Antimicrobial substances produced by coliform strains active against foodborne pathogens. Foodborne Pathogens and Disease, v. 7, n. 3, p. 243-247, 2010. FLINT, S.; JAMALUDIN, N. M.; SOMERTON, B.; PALMER, J.; BROOKS, J. The effect of milk composition on the development of biofilms. Biofilms in the Dairy Industry, ed. 1, Chichester, UK, John Wiley & Sons, Ltd, 2015 FREEMAN, D. J.; FALKINER, F. R.; KEANE, C. T. New method for detecting slime production by coagulase negative staphylococci. Journal of Clinical Pathology, v. 42, n. 8, p. 872-874, 1989. FREITAS, R.; BRITO, M. A.; NERO, L. A.; DE CARVALHO, A. F. Microbiological safety of Minas Frescal Cheese (MFC) and tracking the contamination of Escherichia coli and Staphylococcus aureus in MFC processing. Foodborne Pathogens and Disease, v. 10, n. 11, p. 951-955, 2013. GALINARI, E.; NÓBREGA, J. E.; ANDRADE, N. J.; FERREIRA, C. L. L. F. Microbiological aspects of the biofilm on wooden utensils used to make a Brazilian artisanal cheese. Brazilian Journal of Microbiology, v. 45, n. 2, p. 713-720, 2014. GEFFERS, C.; GASTMEIER, P. Nosocomial infections and multidrug-resistant organisms in Germany. Deutsches Ärzteblatt International, v. 108, n. 6, p. 87-93, 2011.

49
GIAMBIAGI-MARVAL, M.; MAFRA, M. A.; PENIDO, E. G.; BASTOS, M. C. Distinct groups of plasmids correlated with bacteriocin production in Staphylococcus aureus. Journal of General Microbiology, v. 136, n. 8, p. 1591-1599, 1990. GROTIUZ, G.; SIROK, A.; GADEA, P.; VARELA, G.; SCHELOTTO, F. Shiga toxin 2-producing Acinetobacter haemolyticus associated with a case of bloody diarrhea. Journal of Clinical Microbiology, v. 44, n. 10, p. 3838-3841, 2006. GUIMARÃES, A. G.; CARDOSO, R. C. V.; AZEVÊDO, P. F.; MENESES, R. B. Perfil de susceptibilidade antimicrobiana de bactérias isoladas de queijos coalho. Revista do Instituto Adolfo Lutz (Impresso), v. 71, n. 2, p. 259-265, 2012. HANTSIS-ZACHAROV, E.; HALPERNM, M. Chryseobacterium haifense sp. nov., a psychrotolerant bacterium isolated from raw milk. International Journal of Systematic and Evolutionary Microbiology, v. 57, p. 2344-2348, 2007. HAWKEY, S.; CHOY, A. M. Serratia marcescens: A rare cause of recurrent implantable cardioverter defibrillator site infection. Case Reports in Cardiology, v. 2015, p. 1-3, 2015. HEIZMANN, W. R.; DUPONT, H.; MONTRAVERS, P.; GUIRAO, X.; ECKMANN, C.; BASSETTI, M.; GARCIA, M. S.; CAPPARELLA, M. R.; SIMONEAU, D.; BODMANN, K. F. Resistance mechanisms and epidemiology of multiresistant pathogens in Europe and efficacy of tigecycline in observational studies. The Journal of Antimicrobial Chemotherapy, v. 68, supl. 2, p. ii45-ii55, 2013. HØIBY, N.; BJARNSHOLT, T.; GIVSKOV, M.; MOLIN, S.; CIOFU, O. Antibiotic resistance of bacterial biofilms. International Journal of Antimicrobial Agents, v. 35, n. 4, p. 322-332, 2010. HORNSEY, M., PHEE, L., WOODFORD, N., TURTON, J., MEUNIER, D., THOMAS, C., WAREHAM, D. W. Evaluation of three selective CHROMogenic media, CHROMagar ESBL, CHROMagar CTX-M and CHROMagar KPC, for the detection of Klebsiella pneumoniae producing OXA-48 carbapenemase. Journal of Clinical Pathology, v. 66, n. 4, p. 348-350, 2013. HSIEH, C-C.; HERNÁNDEZ-LEDESMA, B.; FERNÁNDEZ-TOMÉ, S.; WEIBORN, V.; BARILE, D.; BELL, J. M. L. N. M. Milk proteins, peptides, and oligosaccharides: effects against the 21st century disorders. BioMed Research International, v. 2015, p. 1-16, 2015. HUANG, X-Z.; FRYE, J. G.; CHAHINE, M. A.; GLENN, L. M.; AKE, J. A.; SU, W.; NIKOLICH, M. P.; LESHO, E. P. Characteristics of plasmids in multi-drug-resistant Enterobacteriaceae isolated during prospective surveillance of a newly opened hospital in Iraq. PLOSone, v. 7, n. 7, p. 1-8, 2012.

50
IÇGEN, B.; GÜRAKAN, G. C.; ÖZCENGIZ, G. Characterisation of local isolates of Enterobacteriaceae from Turkey. Microbiological research, v. 157, n. 3, p. 233-238, 2002. IQBAL, J.; RAJANI, M.; SIDDIQUI, R.; KHAN, N. A. Neuropathogenic Escherichia coli K1 does not exhibit proteolytic activities to exert its pathogenicity. Journal of negative results in biomedicine, v. 12, n. 8, 2013. JEYAKUMAR, E.; LAWRENCE, R. Prevalence of multi-drug resistant (MDR) enteric bacteria in milk and milk products. Indian Journal of Dairy Science, v. 67, n. 5, 2014. JOHNSON, E. E., WESSLING-RESNICK, M. Iron metabolism and the innate immune response to infection. Microbes and Infection, v. 14, n. 3, p. 207-216, 2012. KOBAYASHI, T.; KUTSUNA, S.; HAYAKAWA, K.; KATO, Y.; OHMAGARI, N.; URYU, H.; YAMADA, R.; KASHIWA, N.; NEI, T.; EHARA, A.; TAKEI, R.; MORI, N.; YAMADA, Y.; HAYASAKA, T.; KAGAWA, N.; SUGAWARA, M.; SUZAKI, A.; TAKAHASHI, Y.; NISHIYAMA, H.; MORITA, M.; IZUMIYA, H.; OHNISHI, M. Case report: an outbreak of food-borne typhoid fever due to Salmonella enterica serotype Typhi in Japan reported for the first time in 16 years. The American Journal of Tropical Medicine and Hygiene, v. 94, n. 2, p. 289-291, 2016. KOMMINENI, S.; BRETL, D. J.; LAM, V.; CHAKRABORTY, R.; HAYWARD M.; SIMPSON P.; CAO Y.; BOUSOUNIS P.; KRISTICH C. J.; SALZMAN N. H. Bacteriocin production augments niche competition by enterococci in the mammalian gastrointestinal tract. Nature, v. 526, p. 719-722, 2015. KONG, K-F.; SCHNEPER, L.; MATHEE, K. Beta-lactam antibiotics: from antibiosis to resistance and bacteriology. Acta pathologica, microbiologica, et immunologica Scandinavica, v. 118, n. 1, p. 1-36, 2010. KONTOPOULOU, K.; PROTONOTARIOU, E.; VASILAKOS, K.; KRITI, M.; KOTELI, A.; ANTONIADOU, E.; SOFIANOU, D. Hospital outbreak caused by Klebsiella pneumoniae producing KPC-2 β-lactamase resistant to colistin. The Journal of hospital infection, v. 76, n. 1, p. 70-73, 2010. KREWINKEL, M.; BAUR, C.; KRANZ, B.; VON NEUBECK, M.; WENNING, M.; SCHERER, S.; STOECKEL, M.; HINRICHS, J.; FISCHER, L. A sensitive and robust method for direct determination of lipolytic activity in natural milk environment. Food Analytical Methods, v. 9, n. 3, p. 646-655, 2016. LAGRANGE, V.; WHITSETT, D.; BURRIS, C. Global market for dairy proteins. Journal of Food Science, v. 80, supl. 1, p. A16-A22, 2015.

51
LAUNDERS, N.; BYRNE, L.; ADAMS, N.; GLEN, K.; JENKINS, C.; TUBIN-DELIC, D.; LOCKING, M.; WILLIAMS, C.; MORGAN, D.; OUTBREAK CONTROL TEAM. Outbreak of Shiga toxin producing E. coli O157 associated with consumption of watercress, United Kingdom, august to september 2013. Euro surveillance: bulletin Europeen sur les maladies transmissibles= European communicable disease bulletin, v. 18, n. 44, p. 1-5, 2013. LI, Z.; JIANG, A.; YUE, T.; WANG, J.; WANG, Y.; SU, J. Purification and identification of five novel antioxidant peptides from goat milk casein hydrolysates. Journal of Dairy Science, v. 96, n. 7, p. 4242-4251, 2013. LIPSKI, A. New focus in combating biofims: the pioneer organisms on cleaned surfaces. Brauerei Forum, v. 20, n. 7, p. 178-179, 2005. LIU, C.; ZHANG, P.; LI, X.; YANG, J.; WANG, B. An outbreak of carbapenem-resistant Enterobacter cloacae in pediatric intensive care unit of a Chinese teaching hospital International Journal of Clinical and Experimental Medicine, v. 9, n. 2, p. 3565-3571, 2016a. LIU, L. G.; ZHOU, X. Y.; LAN, Z.; LI, L.; LI, Z.; CHEN, W.; WANG, J. Y.; ZHANG, L. J. Salmonella Typhimurium outbreak associated with a contaminated container in a school in Sichuan Province, China. Epidemiology and Infection, v. 144, n. 2, p. 285-290, 2016b. LOURENÇO, M.; RAMOS-MORALES, E.; WALLACE, R. J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal, v. 4, n. 7 (special issue), p. 1008-1023, 2010. LUZ, K. R.; SOUZA, D. C. C.; CICONELLI, R. M. Vacinação em pacientes imunossuprimidos e com doenças reumatológicas auto-imunes. Revista Brasileira de Reumatologia, v.47, n. 2, p. 106-113, 2007. MADIGAN, M. T.; MARTINKO, J. M.; DUNLAP, P. V.; CLARK, D. P. Microbiologia de Brock. 12. ed., Porto Alegre: Artmed, 1160p, 2010. MALEK F.; MOUSSA-BOUDJEMÂA, B.; KHAOUANI-YOUSFI, F.; KALAI, A.; KIHEL, M. Microflora of biofilm on Algerian dairy processing lines: an approach to improve microbial quality of pasteurized milk. African Journal of Microbiology Research, v. 6, n. 17, p. 3836-3844, 2012. MAO, X-Y.; CHENG, X.; WANG, X.; WU, S. J. Free-radical-scavenging and anti-inflammatory effect of yak milk casein before and after enzymatic hydrolysis. Food Chemistry, v. 26, n. 2, p. 484-490, 2011.

52
MARQUES, V. F.; SOUZA, M. M. S.; MENDONÇA, E. C. L.; ALENCAR, T. A.; PRIBUL, B. R.; COELHO, S. M. O.; LASAGNO, M.; REINOSO, E. B. Análise fenotípica e genotípica da virulência de Staphylococcus spp. e de sua dispersão clonal como contribuição ao estudo da mastite bovina. Pesquisa Veterinária Brasileira, v. 33, n. 2, p. 161-170, 2013. MASIELLO, S. N.; MARTIN, N. H.; TRMČIĆ, A.; WIEDMANN, M.; BOOR, K. J. Identification and characterization of psychrotolerant coliform bacteria isolated from pasteurized fluid milk. Journal of Dairy Science, v. 99, n. 1, p. 130-140, 2016. MEZZATESTA, M. L.; GONA; F.; STEFANI, S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance. Future Microbiology, v. 7, n. 7, p. 887-902, 2012. MILHAS, D.; CLARKE, C. J.; HANNUN, Y. A. Sphingomyelin metabolism at the plasma membrane: implications for bioactive sphingolipids. FEBS letters, v. 584, n. 9, p. 1887-1894, 2010. MINOTTO, E.; MILAGRE, L. P.; OLIVEIRA, M. T.; VAN DER SAND, S. T. Enzyme characterization of endophytic actinobacteria isolated from tomato plants. Journal of Advanced Scientific Research, v. 5, n. 2, p. 16-23, 2014. MOGHA, K. V.; SHAH, N. P.; PRAJAPATI, J. B.; CHAUDHARI, A. R. Biofilm - A threat to dairy industry. Indian Journal Dairy Science, v. 67, n. 6, 2014. MORAES, M. S.; ARAÚJO, B. C.; COSTA, L. E. O.; NASCIMENTO, J. S. Shigella in baby bottles of a Brazilian newborn nursery. The Journal of Infection Developing Countries, v. 9, n. 6, p. 679-681, 2015. MSHANAA, S.; GERWINGA, S.; MINDEA, M.; HAINB, T.; DOMANNB, E.; LYAMUYAC, E.; CHAKRABORTYB, T.; IMIRZALIOGLU, C. Outbreak of a novel Enterobacter sp. carrying blaCTX-M-15 in a neonatal unit of a tertiary care hospital in Tanzania. International Journal of Antimicrobial Agents, v. 38, n. 3, p. 265-269, 2011. MUHAMMAD, A. N.; KIMIA, A. I.; MARYAM, K.; DÉRICK, R. Disorder in milk proteins: formation, structure, function, isolation and applications of casein phosphopeptides. Current Protein and Peptide Science, v. 17, n. 4, p. 368-379, 2016. MUNOZ-PRICE, L. S.; WEINSTEIN, R. A. Acinetobacter infection. The New England Journal of Medicine, v. 358, n. 12, p. 1271-1281, 2008. MURPHY, C. P.; FAJT, V. R.; SCOTT, H. M.; FOSTER, M. J.; WICKWIRE, P.; MCEWEN, S. A. Scoping review to identify potential non-antimicrobial interventions to mitigate antimicrobial resistance in commensal enteric bacteria in North American cattle production systems. Epidemiology and Infection, v. 144, n. 1, p. 1-18, 2016.

53
NÖRNBERG, M. F. B. L.; FRIEDRICH, R. S. C.; WEISS, R. D. N.; TONDO, E. C.; BRANDELLI, A. Proteolytic activity among psychrotrophic bacteria isolated from refrigerated raw milk. International Journal of Dairy Technology, v. 63, n. 1, p. 41-46, 2010. NUESCH-INDERBINEN, M.; CEMELA, N.; ALTHAUS, D.; HÄCHLER, H.; STEPHAN, R. Salmonella enterica serovar szentes, a rare serotype causing a 9-month outbreak in 2013 and 2014 in Switzerland. Foodborne Pathogens and Disease, v. 12, n. 11, p. 887-890, 2015. OLADIPO, I. C.; OMO-ADUA, R. O. Antibiotics resistance among bacteria isolated from evaporated milk. Asian Journal of Biological Sciences, v. 4, n. 1, p. 77-83, 2011a. OLIVEIRA, V. F.; FLEMING, L. R.; FERREIRA, P. S.; NASCIMENTO, J. S. Substâncias antimicrobianas produzidas por Bacillus spp. isolados de frutas. Boletim do Centro de Pesquisa de Processamento de Alimentos, v. 29, n. 1, p. 57-62, 2011a. OLIVEIRA, C. P.; JÚNIOR, J. P. S.; SILVA, J. A. Bacteriocinas como alternativa na conservação de alimentos. Revista Verde, v. 7, n. 1, p. 09-15, 2012b. OLIVEIRA, M. M. M.; BRUGNERA, D. F.; PICOLLI, R. H. Biofilmes em indústrias de laticínios: aspectos gerais e uso de óleos essenciais como nova alternativa de controle. Revista do Instituto de Laticínios Cândido Tostes, v. 68, n. 390, p. 65-73, 2013c. OONAKA, K.; FURUHATA, K.; HARA, M.; FUKUYAMA, M. Powder infant formula milk contaminated with Enterobacter sakazakii. Japanese journal of infectious diseases, v. 63, p. 103-107, 2010. OTTER, J. A.; YEZLI, S.; FRENCH, G. L. The role played by contaminated surfaces in the transmission of nosocomial pathogens. Infection Control and Hospital Epidemiology, v. 32, n. 7, p. 687-699, 2011. OWEN, M.; WILLIS, C.; LAMPH, D. Evaluation of the TEMPO (®) most probable number technique for the enumeration of Enterobacteriaceae in food and dairy products. Journal of Applied Microbiology, v .109, n. 5, p. 1810-1816, 2010. PFEIFER, Y.; CULLIK, A.; WITTE, W. Resistance to cephalosporins and carbapenems in Gram-negative bacterial pathogens. International Journal of Medical Microbiology, v. 300, n. 6, p. 371-379, 2010. PHELAN, M.; KERINS, D. The potential role of milk-derived peptides in cardiovascular disease. Food & Function, v. 2, n. 3-4, p. 153-167, 2011.

54
PHILIPPE, N.; MAIGRE, L.; SANTINI, S.; PINET, E.; CLAVERIE, J-M.; DAVI-RÉGLI, A-V.; PAGÈS, J-M.; MASI, M. In vivo evolution of bacterial resistance in two cases of Enterobacter aerogenes infections during treatment with imipenem. PLoS ONE, v. 10, n. 9, p. 1-19, 2015. PHILLIPS, I.; CASEWELL, M.; COX, T.; DE GROOT, B.; FRIIS, C.; JONES, R.; NIGHTINGALE, C.; PRESTON, R.; WADDELL. Does the use of antibiotics in food animals pose a risk to human health? A critical review of published data. Journal of Antimicrobial Chemotherapy, v. 53, p. 28-52, 2004. PINTO, C. L. O.; MARTINS, M. L.; VANETTI, M. C. D. Bactérias psicrotróficas e importância de seu controle na cadeia produtiva do leite. Informe Agropecuário, v. 28, n. 238, p .29-37, 2007. PONCE, A. R.; MARTINS, M. L.; ARAUJO, E. F.; MANTOVANI, H. C.; VANETTI, M. C. D. AiiA quorum-sensing quenching controls proteolytic activity and biofilm formation byEnterobacter cloacae. Current microbiology, v. 65, n. 6, p. 758-763, 2012. PUBLIC HEALTH ENGLAND. Identification of Enterobacteriaceae. UK Standards for Microbiology Investigations, v. 16, n. 4, 2015. QUIGLEY, L.; O’SULLIVAN, O.; STANTON, C.; BERESFORD, T. P.; ROSS, R. P.; FITZGERALD, G. F.; COTTER, P. D. The complex microbiota of raw milk. FEMS microbiology reviews, v. 37, n. 5, p. 664-698, 2013. RAJANI C. S. R.; CHAUDHARY A.; SWARNA, A.; PUNIYA A. K. Identification and virulence of Enterobacter sakazakii. Journal of Food & Industrial Microbiology, v. 2, n. 1, 2016 RAMESHKUMAR, M. R., VIGNESH, R., SWATHIRAJAN, C. R., BALAKRISHNAN, P., ARUNAGIRINATHAN, N. Chromogenic agar medium for rapid detection of extended-spectrum β-lactamases and Klebsiella pneumoniae carbapenemases producing bacteria from human immunodeficiency virus patients. Journal of Research in Medical Sciences, v. 20, n. 12, p. 1219-1220, 2015. RENDUELES, O.; GHIGO, J-M. Multi-species biofilms: how to avoid unfriendly neighbors. FEMS microbiology reviews, v. 36, n. 5, p. 972-989, 2012. REZENDE, J. M. Imunodepressão, Imunossupressão. Linguagem Médica, v. 40, n. 2, p. 199-201, 2011. RODARTE, M. P.; DIAS, D. R.; VILELA, D. M.; SCHWAN, R. F. Proteolytic activities of bacteria, yeasts and filamentous fungi isolated from coffee fruit (Coffea arabica L.). Acta Scientiarum. Agronomy, v. 33, n. 3, p. 457-464, 2011.

55
ROLAIN, J. Food and human gut as reservoirs of transferable antibiotic resistance encoding genes. Frontiers in Microbiology, v.4, art. 173, p. 1-10, 2013. ROSA, L. J. B.; ESPER, L. M. R.; CABRAL, J. P. L. G.; FRANCO, R. M.; CORTEZ, M. A. S. Viability of probiotic micro-organism Lactobacillus acidophilus in dairy chocolate dessert and its action against foodborne pathogens. Ciência Rural, v. 46, n. 2, p. 368-374, 2016. ROVEDA, M.; HEMKEMEIER, M.; COLLA, L. M. Avaliação da produção de lipases por diferentes cepas de microrganismos isolados em efluentes de laticínios por fermentação submersa. Ciência e Tecnologia de Alimentos, v. 30, n. 1, p. 126-131, 2010 SAISING, J.; SINGDAM, S.; ONGSAKUL, M.; VORAVUTHIKUNCHAI, S. P. Lipase, protease, and biofilm as the major virulence factors in staphylococci isolated from acne lesions. Bioscience Trends, v. 6, n. 4, p. 160-164, 2012. SAITO, R.; KOYANO, S.; NAGAI, R.; OKAMURA, N.; KOIKE, K. Evaluation of a Chromogenic medium for extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae. Letters in applied microbiology, v. 51, n. 6, p. 704-706, 2010. SALAMI, M.; MOOSAVI-MOVAHEDI, A. A.; MOOSAVI-MOVAHEDI, F.; EHSANI, M. R.; YOUSEFI, R.; FARHADI, M.; NIASARI-NASLAJI, A.; SABOURY, A. A.; CHOBERT, J-M.; HAERTLÉ, T. Biological activity of camel milk casein following enzymatic digestion. Journalof Dairy Research, v. 78, n. 4, p. 471-478, 2011. SALIHU, A.; ALAM, Z.; ABDULKARIM, I.; SALLEH, H. M. Lipase production: An insight in the utilization of renewable agricultural residues. Resources, Conservation and Recycling, v. 58, p. 36-44, 2012. SAMARŽIJA, D.; ZAMBERLIN, S.; POGAČIĆ, T. Psychrotrophic bacteria and their negative effects on milk and dairy products quality. Mljekarstvo, v. 62, n. 2, p. 77-95, 2012. SANI, N. A.; YI, L. Y. Enterobacteriaceae, Cronobacter (Enterobacter) sakazakii and microbial population in infant formula products in the Malaysian market. Sains Malaysiana, v. 40, n. 4, p. 345-351, 2011. SANTOS, M. V.; OLIVEIRA, C. A. F.; AUGUSTO, L. F. B.; AQUINO, A. A. Atividade lipolítica do leite com células somáticas ajustadas para diferentes níveis. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 59, n. 4, 2007. SCHLISSELBERG, D. B.; KLER, E.; KISLUK, G.; SHACHAR, D.; YARON, S. Biofilm formation ability of Salmonella enterica serovar Typhimurium acrAB mutants. International Journal of Antimicrobial Agents, v. 46, n. 4, p. 456-459, 2015.

56
SILVA, V. A. M.; RIVAS, P. M.; ZANELA, M. B.; PINTO, A. T.; RIBEIRO, M. E. R.; SILVA, F. F. P.; MACHADO, M. Avaliação da qualidade físico-química e microbiológica do leite cru, do leite pasteurizado tipo A e de pontos de contaminação de uma Granja Leiteira no RS. Acta Scientiae Veterinariae, v. 38, n. 1, p. 51-57, 2010. SIMÕES, M.; SIMÕES, L. C.; VIEIRA, M. J. A review of current and emergent biofilm control strategies. LWT - Food Science and Technology, v. 43, p. 573-583, 2010. SOTO, S. M. Role of efflux pumps in the antibiotic resistance of bacteria embedded in a biofilm. Virulence, v. 4, n. 3, p. 223-229, 2013. STOESSER, N.; SHEPPARD, A. E.; SHAKYA, M.; STHAPIT, B.; THORSON, S.; GIESS, A.; KELLY, D.; POLLARD, A. J.; PETO, T. E.; WALKER, A. S.; CROOK, D. W. Dynamics of MDR Enterobacter cloacae outbreaks in a neonatal unit in Nepal: insights using wider sampling frames and next-generation sequencing. Journal of Antimicrobial Chemotherapy, v. 70, n. 4, p. 1008-1015, 2015. SUDARWANTO, M., AKINEDEM, O., ODENTHAL, S.; GROSS, M., USLEBER, E. Extended-spectrum β-lactamase (ESBL)–producing Klebsiella pneumoniae in bulk tank milk from dairy farms in Indonesia. Foodborne Pathogens and Disease, v. 12, n. 7, p. 585-590, 2015. TEH, K. H.; FLINT, S.; PALMER, J.; LINDSAY, D.; ANDREWES, P.; BREMER, P. Thermo-resistant enzyme-producing bacteria isolated from the internal surfaces of raw milk tankers International Dairy Journal, v. 21, p. 742-747, 2011a. TEH, K. H.; FLINT, S.; PALMER, J.; ANDREWES, P.; BREMER, P.; LINDSAY, D. Biofilm − An unrecognised source of spoilage enzymes in dairy products? International Dairy Journal, v. 34, n. 1, p. 32-40, 2014b. TEIXEIRA, L. V. Análise sensorial na indústria de alimentos. Revista do Instituto de Laticínios Cândido Tostes, v. 64, n. 366, p. 12-21, 2009. TEKINER, I. H.; ÖZPINAR, H. Occurrence and characteristics of extended spectrum beta-lactamases-producing Enterobacteriaceae from foods of animal origin. Brazilian journal of microbiology, v. 47, n. 2, p. 444-451, 2016. THAM, J.; WALDER, M.; MELANDER, E.; ODENHOLT, I. Prevalence of extended-spectrum beta-lactamase-producing bacteria in food. Infection and drug resistance, v. 5, p. 143-147, 2012. THENMOZHI, S.; MOORTHY, K.; SURESHKUMAR, B. T.; SURESH, M. Antibiotic resistance mechanism of ESBL producing Enterobacteriaceae in clinical field: a review. International Journal of Pure & Applied Bioscience, v. 2, n. 3, p. 207-226, 2014.

57
THOMAS, C. P.; MOORE, L. S.; ELAMIN, N.; DOUMITH, M.; ZHANG, J.; MAHARJAN, S.; WARNER, M.; PERRY, C.; TURTON, J. F.; JOHNSTONE, C.; JEPSON, A.; DUNCAN, N. D. C.; HOLMES, A. H.; LIVERMORE, D. M.; WOODFORD, N. Early (2008–2010) hospital outbreak of Klebsiella pneumoniae producing OXA-48 carbapenemase in the UK. International Journal of Antimicrobial Agents, v. 42, n. 6, p. 531-536, 2013. TORTORA, G. J.; FUNKE, B. R.; CASE, C. L. Microbiologia. 10. ed., Porto Alegre: Artmed, 2012. VERRAES, C.; VAN BOXSTAEL, S.; VAN MEERVENNE, E.; VAN COILLIE, E.; BUTAYE, P.; CATRY, B.; DE SCHAETZEN, M. A.; VAN HUFFEL, X.; IMBERECHTS, H.; DIERICK, K.; DAUBE, G.; SAEGERMAN, C.; DE BLOCK, J.; DEWULF, J.; HERMAN, L. Antimicrobial resistance in the food chain: a review. International journal of environmental research and public health, v. 10, n. 7, p. 2643-2669, 2013. WINAYANUWATTIKUN, P.; KAEWPIBOON, C.; PIRIYAKANANON, K.; CHULALAKSANANUKUL, W.; YONGVANICH, T.; SVASTI, J. Immobilized lipase from potential lipolytic microbes for catalyzing biodiesel production using palm oil as feedstock. African Journal of Biotechnology, v. 10, n. 9, p. 1666-1673, 2011. YANG, H-Y.; KIM, S-K.; CHOI, S-Y.; YOU, D-H.; LEE, S-C.; BANG, W-S.; YUK, H-G. Effect of acid, desiccation and heat stresses on the viability of Cronobacter sakazakii during rehydration of powdered infant formula and in simulated gastric fluid. Food Control, v. 50, p. 336-341, 2015. YAO, K.; ZINZENDORF, N. Y.; BOHOUA, G.; KOUASSI, K. A.; KOUA, A.; KOUADIO, L. K.; KOUAMÉ, P.; LOUKOU, G. Y. Occurrence of Enterobacter sakazakii (Cronobacter) and Other Enterobacteriaceae in commercial powdered infant formula in Abidjan, Ivory Coast Food and Nutrition Sciences, v. 3, n. 6, 2012. ZAJÁC, P.; ČAPLA, J.; VIETORIS, V.; ZUBRICKÁ, S.; ČURLEJ, J. Effects of storage on the major constituents of raw milk. Potravinarstvo® Scientific Journal for Food Industry, v. 9, n. 1, p. 375-381, 2015. ZECKNER, R.; ZIMMERMANN, R.; EICHMANN, T. O.; KOHLWEIN, S. D.; HAEMMERLE, G., LASS, A.; MADEO, F. Fat signals - Lipases and Lipolysis in Lipid Metabolism and Signaling. Cell Metabolism, v. 15, n. 3, p. 279-291, 2012 ZHANG, X. Y.; DING, L. J.; FAN, M. Z. Resistance patterns and detection of aac(3)-IV gene in apramycin-resistant Escherichia coli isolated from farm animals and farm workers in northeastern of China. Research in veterinary science, v. 87, p.449-454, 2009a. ZHANG, R.; HUO, W.; ZHU, W.; MAO, S. Characterization of bacterial community of raw milk from dairy cows during subacute ruminal acidosis challenge by high-throughput sequencing. Journal of the Science of Food and Agriculture, v. 95, p. 1072-1079, 2015b.

58
ZHENG, J.; GÄNZLE M. G.; LIN X. B.; RUAN L.; SUN M. Diversity and dynamics of bacteriocins from human microbiome. Environmental microbiology, v. 17, n. 6, p. 2133-2143, 2015. ZHOU, P.; LIU, D.; CHEN, Y.; LABUZA, T. P. Stability of whey protein hydrolysate powders: effects of relative humidity and temperature. Food Chemistry, v. 150, p. 457-462, 2014. ZIPPERER, A.; KONNERTH, M. C.; LAUX, C.; BERCHEID, A.; JANEK, D.; WEIDENMAIER, C.; BURIAN, M.; SCHILLING, N.; SLAVETINSKY, C.; MARSCHAL, M.; WILLMANN, M.; KALBACHER, H.; SCHITTEK, B.; BRÖTZ-OESTERHELT, H.; GROND, S.; PESCHEL, A.; KRISMER, B. Human commensals producing a novel antibiotic impair pathogen colonization. Nature, v. 535, p. 511-516, 2016.
59
11 ANEXOS
ANEXO 1
E-MAIL DE ACEITE DO ARTIGO PELO PERIÓDICO THE JOURNAL OF INFECTION IN
DEVELOPING COUNTRIES

60
ANEXO 2
COMPROVANTE DE SUBMISSÃO DE ARTIGO PELO PERIÓDICO INTERNATIONAL
FOOD RESEARCH JOURNAL

61
ANEXO 3
CERTIFICADO DA APRESENTAÇÃO DE TRABALHO NA XXXV SEMANA DA QUÍMICA -
RIO DE JANEIRO - PRODUZINDO CIÊNCIAS HÁ 450 ANOS / VIII JORNADA DA PÓS-
GRADUAÇÃO EM ALIMENTOS.

62
ANEXO 4
CERTIFICADO DA X JORNADA INTERNA DE INICIAÇÃO CIENTÍFICA E
TECNOLÓGICA” (X JIT) / V FÓRUM DE INOVAÇÃO, TECNOLOGIA E EDUCAÇÃO” (V
FÓRUM ITE)





























