Embed Size (px)
Citation preview

TECHNISCHE UNIVERSITÄT MÜNCHEN
Lehrstuhl für Humanmedizin
Positive allosteric modulation of GABAA
receptors in the amygdala, hippocampus and
thalamocortical circuits by XBD173, diazepam
and allopregnanolone: a comparative VSDI
study
Tatiana Engel
2017

TECHNISCHE UNIVERSITÄT MÜNCHEN
Fakultät für Medizin
Positive allosteric modulation of GABAA receptors
in the amygdala, hippocampus and thalamocortical circuits by
XBD173, diazepam and allopregnanolone: a
comparative VSDI study
Tatiana N. Engel
Vollständiger Abdruck der von der Fakultät für Medizin der Technischen Universität
München zur Erlangung des akademischen Grades eines
Doktors der Medizin
genehmigten Dissertation.
Vorsitzender: Prof. Dr. Ernst J. Rummeny
Prüfer der Dissertation: 1. apl. Prof. Dr. Gerhard Rammes
2. Prof. Dr. Johann Förstl
Die Dissertation wurde am 25.10.2017 bei der Technischen Universität München
eingereicht und durch die Fakultät für Medizin am 10.10.2018 angenommen.

Meinen Eltern gewidmet

IV
Table of contents:
Table of figures ............................................................................................................................... VII
Table of Abbreviations ..................................................................................................................... IX
1 Introduction ................................................................................................................................. 1
1.1 Anxiety Disorders ................................................................................................................ 1
1.2 Pharmacological Treatment of Anxiety Disorders .............................................................. 1
1.3 Benzodiazepines ................................................................................................................ 2
1.4 Neurosteroids...................................................................................................................... 2
1.5 Translocator protein 18 kDa ............................................................................................... 4
1.5.1 Location and structure ................................................................................................. 5
1.5.2 TSPO function ............................................................................................................. 5
1.5.3 Ligands to TSPO ......................................................................................................... 7
1.5.4 XBD173 ....................................................................................................................... 7
1.6 GABAA receptor .................................................................................................................. 8
1.7 Amygdala .......................................................................................................................... 10
1.7.1 Anatomy and Neurophysiology ................................................................................. 10
1.7.2 Function ..................................................................................................................... 10
1.8 Hippocampus .................................................................................................................... 11
1.8.1 Neurophysiology ........................................................................................................ 11
1.8.2 Function ..................................................................................................................... 12
1.9 Thalamocortical Connections ........................................................................................... 12
1.9.1 Anatomy and Neurophysiology ................................................................................. 12
1.9.2 Function ..................................................................................................................... 14
1.10 Aim of this dissertation ...................................................................................................... 14

V
2 Material and Methods ............................................................................................................... 16
2.1 Preparation of brain slices ................................................................................................ 16
2.2 Voltage sensitive dye imaging .......................................................................................... 17
2.2.1 Staining of brain slices with Di-4-ANEPPS ............................................................... 17
2.2.2 VSDI Setup ................................................................................................................ 19
2.2.3 Recordings ................................................................................................................ 20
2.3 Data analysis .................................................................................................................... 22
3 Results ..................................................................................................................................... 24
3.1 Alteration of neuronal activity upon stimulus in the Amygdala by diazepam, XBD and
allopregnanolone ........................................................................................................................ 24
3.1.1 Effect of diazepam on neuronal activity upon stimulus in the amygdala .................. 24
3.1.2 Effect of XBD on neuronal activity upon stimulus in the amygdala .......................... 25
3.1.3 Effect of allopregnanolone on neuronal activity upon stimulus in the amygdala ...... 27
3.1.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in the
amygdala ................................................................................................................................ 28
3.2 Alternation of neuronal activity upon stimulus in the hippocampus by diazepam, XBD and
allopregnanolone ........................................................................................................................ 28
3.2.1 Effect of diazepam on neuronal activity upon stimulus in the hippocampus ............ 28
3.2.2 Effect of XBD on neuronal activity upon stimulus in the hippocampus .................... 30
3.2.3 Effect of allopregnanolone on neuronal activity upon stimulus in the hippocampus 31
3.2.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in the
hippocampus .......................................................................................................................... 32
3.3 Alternation on neuronal activity upon stimulus in the thalamocortical connections by
diazepam, XBD and allopregnanolone ...................................................................................... 33
3.3.1 Effect of diazepam on neuronal activity upon stimulus in the thalamocortical
connections ............................................................................................................................ 33

VI
3.3.2 Effect of XBD on neuronal activity upon stimulus in the thalamocortical connections
35
3.3.3 Effect of allopregnanolone on neuronal activity upon stimulus in the thalamocortical
connections ............................................................................................................................ 37
3.3.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in the
thalamocortical connections ................................................................................................... 38
3.3.5 Analyses of diazepam, XBD and allopregnanolone impact on the individual layers of
the cortex ................................................................................................................................ 40
4 Discussion ................................................................................................................................ 42
4.1 Result interpretation .......................................................................................................... 42
4.1.1 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the amygdala
42
4.1.2 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the
hippocampus .......................................................................................................................... 43
4.1.3 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the
thalamocortical connections ................................................................................................... 43
4.2 Clinical relevance, sources of error and outlook .............................................................. 45
4.3 Conclusion ........................................................................................................................ 45
5 Abstract .................................................................................................................................... 47
6 References ............................................................................................................................... 48
7 Appendix .................................................................................................................................. 60
Used Substances ........................................................................................................................... 60
Acknowledgements ........................................................................................................................ 62

VII
Table of figures
Fig. 1: Synthesis of neurosteroids and schematic sketch of allopregnanolone effect ..................... 3
Fig. 2: TSPO relevance in anxiolysis ............................................................................................... 6
Fig. 3: Scheme of XBD’s mode of action and display of GABAA receptor ....................................... 9
Fig. 4: Schematic display of hippocampal formation and the trisynaptic circuit ............................ 11
Fig. 5: Schematic sketch of the thalamocortical connections ........................................................ 13
Fig. 6: Scheme of brain regions associated with benzodiazepine effects and adverse effects .... 15
Fig. 7: Schematic sketch of the principle of voltage sensitive shift of emission ............................ 18
Fig. 8: Schematic sketch of the VSDI-setup and presentation of the processed signal ................ 20
Fig. 9: Display of the slice preparations and electrode and ROI placement .................................. 21
Fig. 10 Excitation transmission in the thalamocortical circuit ........................................................ 24
Fig. 11: Diazepam impact on neuronal activity in the amygdala ................................................... 25
Fig. 12: XBD impact on neuronal activity in the amygdala ............................................................ 26
Fig. 13: allopregnanolone impact on neuronal activity in the amygdala ........................................ 27
Fig. 14 Comparison of substance effects on FDSAUC, FDSmaxInt and area in the amygdala ......... 28
Fig. 15: diazepam impact on neuronal activity in the hippocampus .............................................. 29
Fig. 16: XBD impact on neuronal activity in the hippocampus ...................................................... 30
Fig. 17: allopregnanolone impact on neuronal activity in the hippocampus .................................. 31
Fig. 18 Comparison of substance effects on FDSAUC, FDSmaxInt and area in the hippocampus ... 32
Fig. 19: diazepam impact on neuronal activity in the thalamocortical connections ....................... 34
Fig. 20: XBD impact on neuronal activity upon stimulus in the thalamocortical circuit.................. 36
Fig. 21: allopregnanolone impact on neuronal activity upon stimulus in the thalamocortical
connections ............................................................................................................................. 38
Fig. 22 Comparison of substance effects on FDSAUC FDSmaxInt and area in the thalamocortical
connections ............................................................................................................................. 39

VIII
Fig. 23 Display of cortex division for layer analysis ....................................................................... 40
Fig. 24 Layer analysis of diazepam and XBD impact on normalized maximal Intensity ............... 41

IX
Table of Abbreviations
3,5-THDOC allotetrahydrodeoxycorticosterone, 3,21-Dihydroxy-5-pregnan-20-one
a area [No of pixel]
ACSF artificial cerebrospinal fluid
Allo allopregnanolone, 5-pregnan-3-ol-20-one, 3-5-THP
ANT adenine nucleotide translocase
CCD charged-coupled device
CCK4 cholecystokinin tetrapeptide
Cl- Chloride Ion
CNS central nervous system
Diaz diazepam
DMSO dimethyl sulfoxide
eEPSC electric evoked excitatory postsynaptic current
EPSC excitatory postsynaptic current
F/XBD finasteride-Experiments
FBS fetal bovine serum
FDS fast depolarization signal
Fina finasteride
Fluma flumazenil
GABA γ-Aminobutyric acid
GABAA-R GABAA-Receptor
HR-CCD high-resolution charge-coupled device
Int Intensity [∆F/F]
Intmax maximal change of Intensity (layer analysis)

X
IPSC inhibitorischer postsynaptic current
n number of experiments performed
P450scc Cholesterol side chain cleavage cytochrome P450
PAM positive allosteric modulator
PK 11195 N-butan-2-yl-1-(2-chlorophenyl)-N-methylisoquinoline-3-carboxamide, TSPO Antagonist
PSC Postsynaptic current
RAS reticular activating system
ROI region of interest
SEM standard error of the mean
SSRI selective serotonin reuptake inhibitor
TC thalamocortical neuron, also named relay neuron
TRN Thalamic reticular neuron
TSPO Translocator Protein (18 kDa (kilo Dalton)), formerly called: peripheral benzodiazepine receptor
VDAC voltage-dependent anion channel
XBD XBD-173, also named Emapunil or AC-5216, systematic name: N-benzyl-N-ethyl-2(7-methyl-8-oxo-2-phenylpurin-9-yl)acetamide

Introduction Anxiety Disorders
1
1 Introduction
1.1 Anxiety Disorders
Anxiety disorders are highly prevalent in today’s society. They are amongst the most common
psychiatric illnesses. Prevalence and lifetime risk differ considerably between countries. In
Germany the projected lifetime risk is about 17%, whereas in the USA it is as high as 36% (Kessler
et al. 2007). Anxiety disorders comprise a number of distinct diagnoses. They can further be
categorized into panic disorders, specific phobias, social phobias, post-traumatic stress disorders,
obsessive-compulsive disorders and generalized anxiety disorders amongst others (Baldwin et al.
2014). A multifactorial pathogenesis is assumed, including external factors such as traumatic or
straining experiences as well as internal factors like genetics and neurobiological dysfunction
(Höffler et al. 2003). While symptoms can be minor and temporarily, a lot of patients suffer from
severe and recurring symptoms that impair their quality of life to an extent that requires treatment.
1.2 Pharmacological Treatment of Anxiety Disorders
Therapeutic approaches include psychotherapies and psychopharmacological drugs (Baldwin et
al. 2014). However, current pharmacological treatment options are limited by pharmacodynamics
and undesirable side effects. First line in treatment antidepressants such as selective serotonin
reuptake inhibitors (SSRI) exert their anxiolytic effect only after several weeks of treatment
(Bandelow et al. 2008). Though in general well-tolerated, potential initial adverse effects like
nausea, jitteriness, insomnia and even increase of anxiety symptoms as well as weight gain and
sexual dysfunction that may occur in long-term treatment are known to impair patient compliance
(Bandelow et al. 2008). More rapidly onset drugs commonly used are benzodiazepines.
Unfortunately they are not eligible either, due to their broad range of unfavorable adverse effects,
development of tolerance and danger of addiction and withdrawal symptoms.
A new class of substances with a preferable pharmacological profile is needed. However, more
suitable medication needs yet to be established. Drugs, which dispose of rapidly on setting
anxiolytic effects like benzodiazepines, but lack a broad side effect profile, are much needed.
Satisfying anxiolytic effects of endogenous neurosteroids such as allopregnanolone have been
shown (Bitran et al. 1991, Wieland et al. 1991). Unfortunately, pharmacokinetic obstacles such as
low bioavailability limit their medical use (Reddy 2010). However, there is a way to enhance natural
neurosteroid synthesis: The Translocator Protein 18 kDa (TSPO) and its ligands provide a
promising new option. TSPO is crucially involved in the synthesis of neurosteroids, for it is in charge

Introduction Benzodiazepines
2
of the rate-determining step of neurosteroidogenesis (Papadopoulos et al. 1997a). Therefore
ligands to TSPO, like XBD-173 (XBD; Emapunil), that enhance TSPO’s effects and subsequently
promote neurosteroid synthesis are currently under investigation as a potential new therapeutic
alternative, that has already revealed great anxiolytic capabilities and a favorable side effect profile
(Rupprecht et al. 2009).
1.3 Benzodiazepines
Besides in anxiety disorders Benzodiazepines are used in treatment of sleeping disorders, epilepsy
and schizophrenia (Rote-Liste 2013, 2015). They are also commonly used as premedication before
surgical interventions and for calming effects in emergencies as well as to treat muscle spasms.
Even though they are very potent anxiolytic drugs and exhibit a quick onset of anxiolytic effects
they are not suitable for long-term anxiety treatment due to a broad side effect profile, development
of tolerance and danger of addiction and withdrawal symptoms (Bandelow et al. 2008) . Side effects
include sedation, drowsiness, anterograde amnesia, respiratory depression and central diminution
of tonicity (Rote-Liste 2013, 2015).
Benzodiazepines exert their effects by enhancing γ-Aminobutyric acid (GABA) impact in the central
nervous system (CNS) (Rote-Liste 2013, 2015). Since GABA is the most important inhibitory
transmitter in the CNS, they reduce neuronal activity (Bloom et al. 1971). Benzodiazepines bind to
a specific compound of the GABAA-receptor (GABAA-R) between the α- and -subunit (Möhler
2006). Once connected to the receptor, they exert an allosteric effect that facilitates binding of
GABA to the GABAA-R and increases its opening frequency (see chapter 1.6), thus decreasing
neuronal excitability, which causes anxiolytic effects amongst other things (Walters et al. 2000).
1.4 Neurosteroids
Neuroactive steroids are a specific kind of steroids that, in addition to genomic alternation, the
common course of action of steroids, express the ability to modulate neurotransmitter receptors
(Majewska et al. 1986, Paul et al. 1992, Rupprecht et al. 1999). While gene expression via steroid
activated transcriptional factors take minutes to hours, effects by modulation of neurotransmitter
receptors occur within seconds (McEwen 1991). One of the affected receptors is the GABAA-R
(Majewska et al. 1986).
Neurosteroids are predominantly produced in the brain and independent of peripheral steroid levels
(Corpéchot et al. 1981). The precursor of all neurosteroids is pregnenolone, which is synthesized
out of cholesterol inside the mitochondrion (Lacapère et al. 2003, Rupprecht et al. 2010). Synthesis
of pregnenolone underlies regulation by TSPO (see chapter 1.5.2). Pregnenolone leaves the

Introduction Neurosteroids
3
mitochondrion and is processed further in the cytosol into a variety of other neurosteroids (see Fig.
1A).
A
B
Fig. 1: Synthesis of neurosteroids and schematic sketch of allopregnanolone effect
A Scheme of synthesis of neurosteroids from (Schüle et al. 2014) according to (Nothdurfter et al. 2012a). Neurosteroids are formed out of cholesterol. First step, transformation to pregnenolone, the precursor of all neurosteroids, takes place in the mitochondrion. Further synthesis of diverse neurosteroids out of pregnenolone takes place in the cytoplasm. Utilized enzymes are depicted as arrows. B Scheme of allopregnanolone effect according to (Nothdurfter et al. 2012b). Allopregnanolone binds to the GABAA receptor where it exerts a modulating effect, enhancing GABA binding and subsequent chloride influx, thus potentiating its inhibitory impact on neuronal activity, which leads to Anxiolysis.

Introduction Translocator protein 18 kDa
4
Neurosteroid biosynthesis is region and neuron specific, depending on the expression of the
respective enzymes for neurosteroid formation (Rupprecht et al. 2010). Region-specific synthesis
of neurosteroids is subsequently responsible for a distinct modulation of neurotransmitter receptors
in certain brain areas, resulting in region-specific alteration of signal transduction. Some of the
neurosteroids formed are 3-reduced neurosteroids, such as allopregnanolone (Rupprecht 1997).
Certain 3-reduced steroids, especially allopregnanolone, have been shown to feature anxiolytic
properties in various stress-models in rodents (Bitran et al. 1991, Picazo et al. 1995, Wieland et al.
1995, Reddy et al. 1997). Furthermore they may play a role in pathology of affective diseases
including anxiety disorders (Strohle et al. 2003, Reddy 2010). Similar to benzodiazepines, 3-
reduced steroids exercise their anxiolytic effects by potentiating inhibitory effects of GABA (see
Fig. 1B) (Gee 1988, Bitran et al. 1991). However they modulate GABAA-receptors using another
binding site (see Fig. 3B) (Reddy et al. 1997, Hosie et al. 2006).
Allopregnanolone stands out with particular effectiveness in reducing stress and anxiety-like
behavior in rodents. Increased allopregnanolone levels were detected in rodents (also
adrenalectomized rodents) exposed to stressors or acute stress paradigms (Purdy et al. 1991,
Barbaccia et al. 1997). Induced panic attacks in patients with panic disorders led to decreasing of
allopregnanolone plasma concentrations (Strohle et al. 2003). While stressors raise
allopregnanolone levels, presumably attempting to prevent anxiety or panic outbreak, panic attacks
reduce allopregnanolone levels assumedly by consumption. Anti-anxiety behavior in rodents has
been reduced by blockage of allopregnanolone formation (Frye et al. 2011, Koonce et al. 2012).
Concordantly, anti-anxiety behavior was reinstated by allopregnanolone injection (Frye et al. 2011).
These findings propose allopregnanolone to be a neurosteroid of particular importance concerning
anxiety treatment (Schüle et al. 2014).
Unfortunately natural neurosteroids like allopregnanolone have a low bioavailability as a result of
rapid inactivation and elimination by glucuronidation and sulfate conjugation, as well as possible
oxidation to ketone (Reddy 2010). Hence, they are not viable for therapeutic use. Fortunately there
are methods to enhance neurosteroid synthesis in the brain as described in the following chapters.
1.5 Translocator protein 18 kDa
The Translocator protein (18 kDa) (TSPO) was first recognized in 1977 in the kidney on the search
for binding sites for benzodiazepines (Braestrup et al. 1977) and thus afterwards named peripheral
benzodiazepine receptor. In consideration of its broad range of function, employment beyond its
receptor capacities and due to the fact that not all benzodiazepines bind to it, but a number of other
substances do and that its structure and function differ tremendously from the central

Introduction Translocator protein 18 kDa
5
benzodiazepine receptor (part of the GABAA receptor complex), it was renamed Translocator
protein (18 kDa) in 2006 (Papadopoulos et al. 2006a).
1.5.1 Location and structure
TSPO has since been detected in various tissues including the brain, though the highest levels of
expression have been found in steroid producing tissues like adrenal and gonad (Anholt et al. 1985,
Gavish et al. 1999). Inside the CNS it is primarily expressed in glia cells (Gavish et al. 1999).
At the subcellular level, TSPO is mainly – though not exceptionally (Olsen et al. 1988, Oke et al.
1992) – located at the outer mitochondrial membrane (Anholt et al. 1986), cumulating at contact
sites of the outer and inner mitochondrial membrane (Culty et al. 1999). Topological analysis of
TSPO revealed it to consist of 5 trans membrane -helices (Joseph-Liauzun et al. 1998). 3D
modeling of TSPO suggested that the helices form a channel, likely used for cholesterol transport
(Bernassau et al. 1993, Culty et al. 1999). However to be accurate TSPO should be characterized
as a cholesterol exchanger rather than a channel, for it actively transports cholesterol instead of
just letting it pass (Lacapère et al. 2003).
TSPO interacts with other mitochondrial membrane proteins such as the 32 kDa voltage dependent
anion channel (VDAC) and the 30 kDa adenine nucleotide translocase (ANT) (McEnery et al.
1992). They form a complex reaching from outer to inner mitochondrial membrane, whereupon
TSPO is the minimal functional unit able to bind cholesterol and drug ligands (Lacapère et al. 2001).
However VDAC is essential for maximizing benzodiazepine and endozepine (endogenous
compounds with likewise effects as benzodiazepines) binding to TSPO (Garnier et al. 1994). The
entire impact of these protein associations is not yet fully understood, but they are believed to alter
drug binding properties and related functions (Golani et al. 2001). Furthermore,
homopolymerization of TSPO influences pharmacological and physiological properties (Delavoie
et al. 2003).
1.5.2 TSPO function
TSPO is expressed of a gene highly conserved throughout evolution, appearing in all sorts of
organisms extending from bacteria to humans (Gavish et al. 1999). It has been found to participate
in many cell regulation and signal transduction mechanism of the mitochondrion. Three main
functions are thought to be primarily responsible for all TSPO-delegated effects (Papadopoulos et
al. 2006a). These include cholesterol (Papadopoulos et al. 1997b, Li et al. 2001), protein (Wright
et al. 1999, Hauet et al. 2005) and porphyrin (Synder et al. 1987, Taketani et al. 1994) binding and
transport into the mitochondrion. Cholesterol translocation is essential for steroid and bile salt
biosynthesis, protein import is necessary for membrane biogenesis and porphyrin import is a part
of the heme biosynthesis pathway (Papadopoulos et al. 2006a). Associated functions might include

Introduction Translocator protein 18 kDa
6
cellular proliferation and differentiation, immunomodulation, cellular respiration and involvement in
oxidative processes, regulation of mitochondrial metabolism and apoptosis (Papadopoulos et al.
2006a, Rupprecht et al. 2010). The importance of TSPO for the organism is demonstrated by the
fetal consequence of functional deactivation in an early embryotic phase in mice (Papadopoulos et
al. 1997a).
As for TSPO involvement in neurosteroid
genesis, cholesterol translocation from the
cytosol to the inner mitochondrial membrane
by TSPO (Papadopoulos et al. 1997a) is the
rate-limiting step in neurosteroid genesis
(Jefcoate 2002). Inside the Mitochondrion is
the cholesterol side chain cleavage
cytochrome enzyme P450 (P450scc), ready to
metabolize cholesterol to pregnenolone, the
precursor of all neurosteroids (Lacapère et al.
2003, Rupprecht et al. 2010). Positive
allosteric neurosteroids such as
allopregnanolone are subsequently formed
and modulate GABAA receptors, thus exerting
anxiolytic effects (Fig. 2) (Papadopoulos et al.
2006b). Since inside the CNS, TSPO is mainly
expressed in glia cells, the acquired
neurosteroids are believed to work in a
paracrine fashion for the most part, operating
at receptors located at surrounding neurons
(Nothdurfter et al. 2012a).
In addition TSPO seems to be a considerable
factor in a number of pathological conditions,
including brain injury, peripheral neuropathy,
neurodegenerative disorders such as Alzheimer’s disease and Parkinson’s disease, epilepsy,
certain kinds of cancer and psychiatric disorders such as anxiety disorders (Papadopoulos et al.
2006a). As a matter of fact a number of studies demonstrated an increase in TSPO density in acute
stress while under chronic stress as well as in anxiety disorders density is usually decreased,
indicating TSPO to be a critical factor in the organism’s way of coping with stress (Gavish et al.
1999). For instance, in patients suffering from generalized anxiety disorder, lower levels of ligand
binding to TSPO have been measured (Weizman et al. 1987). Moreover, levels increased again
Fig. 2: TSPO relevance in anxiolysis
Illustration of TSPO function in anxiolysis according to (Nothdurfter et al. 2012b). Cholesterol is imported to the inside of the mitochondrion by TSPO. Here the first step of neurosteroidogenesis, pregnenolone (precursor of all neurosteroids) formation out of cholesterol, is performed. Subsequent, allopregnanolone is increasingly synthesized and ready to manipulate GABAA-Receptors, resulting in anxiolysis.

Introduction Translocator protein 18 kDa
7
after treatment with diazepam whereupon relief of anxiety was stated (Weizman et al. 1987). Also
in soldiers repeatedly exposed to stress exercises, reduced PBR density was monitored as well as
in civilians during war (Dar et al. 1991, Weizman et al. 1994). Whereas in psychiatric residents
completing their exams significant increase of platelet PBR density was shown, presumably as a
way to cope with stress and prevent anxiety (Karp et al. 1989). Therefore, drugs acting at a
mechanism that appears to be involved in pathology of anxiety formation propose to show great
promise.
1.5.3 Ligands to TSPO
There are a number of substances capable of binding to TSPO. Within high affinity endogenous
ligands are cholesterol and porphyrins. Also endozepines (derivatives of the common polypeptide
precursor diazepam-binding inhibitor (DBI)) are to be mentioned, which are able to displace
benzodiazepines from their binding site. While cholesterol is known to connect to the C-terminus
of TSPO (Li et al. 2001), drug ligands bind to the amino terminus (Rupprecht et al. 2010). This is
consistent with the observation that cholesterol is imported to the inner mitochondrial membrane
by TSPO, while other ligands influence TSPO properties such as cholesterol translocation
(Lacapère et al. 2003). Endogenous drugs are therefore likely to regulate TSPO function. Given
the many TSPO functions and its involvement in pathologies, great therapeutic benefit is to be
expected of synthetic TSPO ligands.
A number of synthetic ligands have been developed. A lot of them initially served as neuroimaging
agents to determine TSPO distribution or for means of identification of TSPO function (Rupprecht
et al. 2010). However certain ligands have been shown to promote neurosteroidogenesis in the
brain and therefore exercise anxiolytic effects in rodent models (Serra et al. 1999, Bitran et al.
2000, Verleye et al. 2005). One of them is XBD173 (XBD) (Rupprecht et al. 2009).
1.5.4 XBD173
XBD’s mode of action
XBD, a phenyl-purine acetamide with promising anxiolytic potential, is a selective ligand to TSPO
with nanomolar affinity (Kita et al. 2004). Its affinity to neurotransmitter receptors including the
GABAA receptor is negligible (Kita et al. 2004). A research group around Rupprecht (2009)
demonstrated that XBD does not exert any direct effects at the GABAA receptor. However,
appliance of XBD to neocortical slices potentiated GABAergic neurotransmission (Rupprecht et al.
2009). This was prevented by finasteride, a 5-reductase inhibitor, indicating 5-reduced
neurosteroids to be responsible for GABA-mediated actions following XBD application (Rupprecht
et al. 2009). Furthermore, significantly elevated levels of allopregnanolone in the brains of rats were
measured after oral medication with XBD (Rupprecht et al. 2009). In addition, anxiolytic properties

Introduction GABAA receptor
8
of XBD during stress tests (social exploration and elevated plus maze test (Pellow et al. 1985)) in
rats were prevented by the TSPO antagonist PK 11195, confirming XBD’s mode of action to be
TSPO-mediated up-regulation of neurosteroidogenesis (Fig. 3) (Rupprecht et al. 2009).
Effect and side-effect profile of XBD
Besides its anxiolytic abilities in stress tests, XBD also counteracted sodium lactate and
cholecystokinin tetrapeptide (CCK4) induced panic attacks in rats (Rupprecht et al. 2009). Hereby
no signs of sedation were detected, different to like-wise application of the benzodiazepine
alprazolam. Moreover, there was no indication for decrease of XBD’s anxiolytic efficiency
(tolerance) in the social exploration stress test under subchronic administration, a phenomenon
well known in benzodiazepine-treatment (Rupprecht et al. 2009, Rote-Liste 2013, 2015). This was
also observed in anxiety models in mice by the research team around Kita (Kita et al. 2009). In
addition, XBD unlike diazepam lacked withdrawal symptoms in special designed withdrawal
experiments they performed on mice (Kita et al. 2009).
In humans, XBD revealed anxiolytic properties as well. Anxiolytic efficiency of XBD proved to be
as potent as alprazolam’s in the CCK-4 challenge (Rupprecht et al. 2009). Occurrence of side
effects in XBD treated subjects was comparable to incidents in subjects treated with placebo.
Alprazolam treated subjects however suffered a much higher incidence of adverse effects,
especially dizziness and somnolence. The same was reported of withdrawal symptoms.
To summarize, the data presented by Rupprecht at al (2009) and Kita at al (2009) indicates that
XBD possesses rapidly onset anxiolytic properties comparable to benzodiazepines, but with a
distinct superior side effect profile. More precisely, as far as up to date studies detected, XBD lacks
adverse effects like sedation, anterograde amnesia, tolerance development, addiction and
withdrawal symptoms. Whereas all of the above have been observed in patients treated with
benzodiazepines (Rote-Liste 2013, 2015). Therefore XBD may be a superior candidate in the
treatment of anxiety disorders.
1.6 GABAA receptor
The diverse side effect profile of XBD and diazepam despite corresponding final modes of action
could be due to different distribution of the various subtypes of GABAA receptors. Though both,
benzodiazepines and certain neurosteroids (5-reduced neurosteroids such as allopregnanolone
and 3,5-THDOC), act as positive allosteric modulators at the GABAA receptor, they occupy
different binding sites (Fig. 3B), which do not always coexist in the same GABAA receptor (Olsen
et al. 2008).

Introduction GABAA receptor
9
A B
Fig. 3: Scheme of XBD’s mode of action and display of GABAA receptor
A: TSPO ligand mediated neurosteroidogenesis; modified according to (Rupprecht et al. 2010). TSPO ligands, like XBD, enhance TSPO function, thus favoring translocation of Cholesterol from the cytosol to the inner mitochondrial membrane, the rate-limiting step in neurosteroid genesis. The cholesterol side-chain-cleaving cytochrome P450 enzyme (P450scc) metabolizes cholesterol to pregnenolone (Pregnenol.), the precursor of all neuroactive steroids. In the cytosol Pregnenolone is metabolized
further. For allopregnanolone formation the enzymes 5-Reductase and 3-Hydroxysteroid
dehydrogenase (3-HSD) amongst others are necessary. Allopregnanolone then exerts anxiolytic effects by modulation of the GABAA receptor. B: systematic display of the GABAA receptor including various binding sites; according to (Nothdurfter et al. 2012a). The GABAA receptor is located at the cell membrane. It is build out of 5 subunits deriving out of 8 subunit classes. Different binding sides for GABA, diazepam (in between α- and γ-subunits) and allopregnanolone (in between α- and β- subunits) can be identified.
-Aminobutyric acid (GABA) is the main inhibitory neurotransmitter receptor in the CNS (Bloom et
al. 1971). The GABAA receptor belongs to the category of ligand-gated ion channels. It is built of 5
subunits, each containing 4 transmembrane domains, forming a central pore (Bormann et al. 1987,
Pirker et al. 2000). Upon binding of two GABA molecules, the pore becomes permeable for chloride
ions (Cl-) and bicarbonate (HCO3-) (Bormann et al. 1987, Macdonald et al. 1994). Influx of negative
charged ions induces hyperpolarization, causing an inhibitory postsynaptic current to occur in the
corresponding neuron, cancelling out potential excitatory postsynaptic currents emerging
simultaneously from another synapse of the same neuron. In short: GABA has a diminishing impact
on neuronal activity e.g. signal transduction.

Introduction Amygdala
10
The subunits forming the GABAA receptor can vary out of a range of 19 different subunits divided
in 8 subunit-classes: 1-6, 1-3, 1-3, , , , , 1-3. (Barnard et al. 1998, Olsen et al. 2008).
Depending on the subunit composition, the GABAA receptor is able to bind a variety of different
drugs and endogenous ligands. Distribution of diverse GABAA receptor subtypes in the brain is
heterogeneous (Pirker et al. 2000).
Benzodiazepines and certain neurosteroids (5-reduced neurosteroids such as allopregnanolone
and THDOC) both facilitate GABA binding by enhancing receptor affinity towards GABA mediated
by allosteric changes of the GABAA receptor (Sigel et al. 1997, Belelli et al. 2005). While the binding
site for benzodiazepines is formed by - and -subunits, neurosteroids occupy another binding site
located in between the - and - subunits (Fig. 3) (Hosie et al. 2006, Möhler 2006). Therefore, not
only availability of benzodiazepines or neurosteroids is necessary for respective effects, but also
the expression of corresponding GABAA receptor subtypes. This might be a significant factor,
leading to the disparity of their side effect profiles.
1.7 Amygdala
The main focus of this dissertation is on the discordance between the side effect profile of XBD
and diazepam. This discordance is most likely due to different effects that these drugs have on
respective parts of the brain. Hence the influence of those substances on the amygdala, the
hippocampus and the thalamocortical connections was measured in the course of this dissertation.
Those cerebral areas will therefore be introduced in this and the following chapters.
1.7.1 Anatomy and Neurophysiology
The corpus amygdaloideum, more commonly known as the amygdala, is localized in the medial
temporal lobe in each hemisphere. The amygdala consists of many distinct, however densely
interconnected, nuclei (Amaral et al. 1992). They can be functionally and anatomically grouped into
three core domains: the basolateral (composed of the lateral and the basal nuclei (Keifer et al.
2015)), the centromedial and the superficial nuclei group (Bzdok et al. 2013). The basolateral
amygdala obtains its information from the sensory organs, the hippocampus and the cortex and is
in charge of assigning emotional values to distinct stimuli (Sah et al. 2003). This will be the nuclei
group this dissertation focuses on.
1.7.2 Function
The amygdala is a part of the limbic system and is known to play a crucial role in processing
emotions and initiating fear and anxiety reactions (Trepel 2008). Functional loss of the amygdala
results in a decrease of negative emotions such as anxiety and rage (Weiskrantz 1956, Zola-

Introduction Hippocampus
11
Morgan et al. 1991). Concordantly, stimulation of the amygdala provokes anxiety and fear (Kaada
1951, Chapman et al. 1954). Furthermore, direct infusion of allopregnanolone into the amygdala
resulted in a decrease of anxiety-patterned behavior (Akwa et al. 1999).
1.8 Hippocampus
The hippocampus is an evolutionary old and well-conserved brain structure. It too is located in the
medial temporal lobe and belongs to the limbic system. It is a part of the hippocampal formation, a
group of functionally and neuronally connected cerebric areas, which in addition to the
hippocampus also includes the entorhinal cortex, the subiculum and the dentate gyrus. The actual
hippocampus equates to the Cornu Ammonis and can be functionally and morphologically
subdivided into 3 domains, the Cornu Ammonis 1 to 3 (CA1-CA3).
1.8.1 Neurophysiology
The hippocampus obtains information from several sensory systems. The entorhinal cortex, as the
main input source of the hippocampus, hereby channels said information (Lomo 1971). The
incoming data is then processed and forwarded back to the cortex. Within the neuronal network of
the hippocampal formation, there are three major successive excitatory projections, which
combined resemble the trisynaptic circuit (Doller et al. 1982, Yeckel et al. 1990). The starting point
is formed by the perforant path, which is made up of fibers emerging from the entorhinal cortex
leading to the dentate gyrus. From here the so-called mossy fibers innervate the CA3 region, which
in turn connects via the Schaffer collaterals to the CA1 region (Amaral 1993). Axons from the CA1
region then project to the subiculum and back to the entorhinal cortex, thus completing the
trisynaptic circuit (see Fig. 4). Within each region there are inhibitory GABAergic interneurons
processing the data (Jones 1993, Pettit et al. 2000). These GABAergic fibers are the working point
of GABAA receptor modulating drugs such as benzodiazepines.
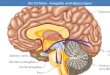
Fig. 4: Schematic display of hippocampal
formation and the trisynaptic circuit
Modified according to (Amaral 1993). The hippocampal formation is made up out of the entorhinal cortex (EC), the dentate gyrus (DG), the hippocampus and the subiculum (SUB). The hippocampus can be subdivided into regions CA1-CA3. The trisynaptic circuit is a successive pathway that is made up of three major excitatory projections: the perforant path (pp) emerging from the EC to innervate the DG, the mossy fibers (mf) that connect the DG with the CA3 region of the hippocampus and the schaffer collaterals (sc) that excite the CA1 region

Introduction Thalamocortical Connections
12
1.8.2 Function
A core function of the hippocampus is the consolidation of the declarative memory. Scoville and
his team were the first to discover, that upon bilateral removal of the hippocampus, patients suffered
from anterograde amnesia (Scoville et al. 1957). Afterwards it was repeatedly confirmed that the
hippocampus is indeed crucial for forming new memories (Moss et al. 1981, Eichenbaum et al.
1992, Squire 1992). Close evaluation showed that hereby only the declarative memory was
compromised, whilst the procedural memory was not impaired (Eichenbaum et al. 1992). Since the
hippocampus plays such a leading role in memory formation it can be supposed, that drugs with
amnestic side effects interfere within some point of hippocampal data processing. For instance,
benzodiazepines might act at hippocampal GABAA receptors, enhancing inhibitory interneurons
that subsequently reduce data transfer and therefore prevent memory consolidation.
1.9 Thalamocortical Connections
The thalamus is densely connected to the cortex and together with the latter shapes up to form the
thalamocortical system (Guillery et al. 2002).
1.9.1 Anatomy and Neurophysiology
The thalamus is located in the diencephalon, can be anatomically split into a ventral and dorsal
thalamus and consists of numerous nuclei. The nuclei can be divided according to their
connections. While some nuclei (so called specific nuclei) send their efferent nerve fibers quite
distinct to each respective corresponding area of the cerebral cortex and gather information from
sensory and sensitive organs for the most part, others have rather fewer direct efferent nerve fibers
to the cortex but connect mainly to the first mentioned sort of thalamic nuclei (Trepel 2008). The
second kind of nuclei, also called unspecific nuclei, receive their afferent input from the cerebellum,
the basal ganglia, the cortex and above all from the reticular formation. Their efferent fibers to
specific nuclei serve to modulate the activity and therefore output of the specific nuclei (Trepel
2008).
A certain nucleus of the ventral thalamus needs to be emphasized: the thalamic reticular nucleus
(TRN) that wraps around the remaining thalamus (Fig. 5A). As indicated by the name, it mostly
consists of reticular cell that converse with diverse specific nuclei in an inhibitory manner using
GABAergic connections (Lubke 1993). It is reached by collaterals of thalamocortical and
corticothalamic fibers and regulates activity in the dorsal thalamus accordingly (Fig. 5B) (Jones
1975, Shosaku et al. 1989). The TRN is regulated itself by the reticular formation (located in the
brainstem) via inhibitory afferent fibers, which enables activity of thalamocortical fibers by
disinhibition, thus allowing efferent nerve fibers of specific thalamic nuclei to excite the cerebral

Introduction Thalamocortical Connections
13
cortex (Fig. 5B) (McCormick et al. 1997, Trepel 2008). While the excitatory cortex afferent fibers
quite distinguished and selective determine which specific nuclei will be increasingly inhibited by
the TRN, the inhibitory reticular formation afferents inhibit the TRN in an undirected manner (Trepel
2008).
The cerebral cortex plays a vital role in perception, awareness, attention, thought, intended actions
and language (Trepel 2008). The most part of the cortex is amounted by neocortex, the evolutionary
youngest developed component of the brain (Trepel 2008). Due to distribution of different cellular
types, it can be sectioned into six layers, also called laminas, which each play their role in neuronal
processing (Mountcastle 1997). Layer 4 is known to be the main entrance of external information
to the cerebral cortex, basically where specific thalamic nuclei efferent nerve fibers end (Trepel
2008). Neurons of the 5th layer project to diverse subcortical destinations except the thalamus
(Molnár et al. 2006). Lamina 6 is made up out of many different cells, whereupon the pyramidal
cells of this layer sent their axons to the specific nuclei of the thalamus (Trepel 2008). Layer 1 to 3
serve to modulate lower layers and to interconnect cortex areal units (Trepel 2008).
Fig. 5: Schematic sketch of the thalamocortical connections
A: Sketch of a coronary brain slice depicting the location of the thalamus and the cortex. Changed according to (Wahl-Schott et al. 2009). B: Simplified display of the thalamocortical connections. Hereby the + indicates glutamatergic fibers while the – represents GABAergic contacts. Ascending sensory information activates neurons of the specific thalamic nuclei, which then in turn excite the cortex. Thalamoreticular neurons modulate relay neuron function via GABAergic synapses. When the reticular formation as a part of the reticular activation system (RAS) is active, it inhibits inhibitory thalamoreticular neurons, disinhibiting negative influence of thalamoreticular neurons on thalamocortical (TC) neurons. Changed according to (Wahl-Schott et al. 2009, Mattusch 2012)
TC neuron
Thalamoreticular
neuron
Pyramidal
cell of cortex
RAS Sensory input
Cortex
Ncl. reticularis thalami
Thalamus
B

Introduction Aim of this dissertation
14
1.9.2 Function
In means of perception of one’s environment the thalamus plays the role of an operator. All sensory
information gathered, apart from olfaction, passes through the diverse thalamic nuclei, where it is
sorted and selected before being rendered to the respective cortex areas (McCormick et al. 1997,
Huguenard et al. 2007). This switchover serves as a filter, deciding which part of the incoming
information flow is relevant for the individual at the moment and therefore passed on to the cortex,
i.e. to consciousness, and vice versa. Thus did the thalamus acquire its reputation as the gate to
consciousness. In addition to its gate-like function, the thalamus also portrays a regulatory function
in alternating states of consciousness, such as sleeping and being awake (McCormick et al. 1997,
Steriade 2005). Therefore benzodiazepine effect on the thalamocortical connections may be
responsible for side effects such as drowsiness and sedation.
1.10 Aim of this dissertation
Benzodiazepines such as diazepam and the TSPO ligand XBD elicit similar anxiolytic properties
but feature diverging side effect profiles. While diazepam exposes amnestic and sedative adverse
effects as well as tolerance development and risk of addiction, XBD lacks any of those, as far as
experiments on humans and rodents have surveyed (Kita et al. 2009, Rupprecht et al. 2009, Rote-
Liste 2013, 2015). XBD enhances the synthesis of neurosteroids such as allopregnanolone, which
are presumably responsible for XBD effects (Rupprecht et al. 2009). Both, allopregnanolone and
diazepam are positive allosteric modulators (PAMs) at the GABAA receptor, though they occupy
different binding sides (Hosie et al. 2006, Möhler 2006). Due to diverse GABAA receptor subunit
expression, GABAA receptor binding abilities vary throughout different brain areas, probably
accounting for the discrepancy of diazepam and XBD side effect profile (see Fig. 6) (Pirker et al.
2000).
The intention of this study is to investigate the diverging side effect profile of diazepam and XBD
on a neurophysiological level using the imaging technique voltage sensitive dye imaging (VSDI).
Therefore, a series of experiments was designed and conducted to determine neuronal activity
propagation under influence of diazepam, as a representative of benzodiazepines, and XBD.
According to the different side effect profile of these drugs, VSDI experiments in areas of the brain
crucial for neuronal processing related to anxiety, memory and sedation, namely the amygdala,
hippocampus and thalamocortical region respectively were performed. To verify diazepam and
XBD effects, measurements with their antagonists (flumazenil and finasteride) were performed as
well.

Introduction Aim of this dissertation
15
Additionally, experiments with allopregnanolone, a neurosteroid acting as a PAM at GABAA
receptors and producing anxiolysis, were performed (Bitran et al. 1995). It has been shown that
allopregnanolone synthesis increases upon XBD-induced TSPO activation (Rupprecht et al. 2009).
Fig. 6: Scheme of brain regions associated with benzodiazepine effects and adverse effects
According to (Nothdurfter et al. 2012b). Depicted are the brain areas relevant for the adverse effect profile of benzodiazepines and neurosteroids based on the specific GABAA receptor selectivity (red: receptive for benzodiazepines, blue: receptive for neurosteroids) and GABAA receptor subunit distribution in the brain. The color coded chart indicates in which brain region the side effects origin:
Anxiety: cerebral cortex, amygdala (AM), bed nucleus of stria terminalis (BNST), hippocampus (HIP) (Charney et al. 1996, Brandao et al. 2003) Sedation: AM, formatio reticularis (FR) (Bonin et al. 2008) Hypnosis: ventrobasal nucleus (VB), reticular nucleus (RTN), locus coeruleus (LC), Raphe nuclei (RN), substantia nigra (SN), ventrolateral preoptic nucleus (VLPO), tuberomamillary nucleus (TMN) (Bonin et al. 2008) Abuse: N. accumbens (NA), ventral tegmental area (VTA) (Di Chiara et al. 1993) Physical dependence: NA, VTA (Lingford-Hughes et al. 2003, Wafford 2005) Tolerance: HIP, NA, AM (Wafford 2005) Withdrawal: HIP, NA, AM (Wafford 2005) Amnesia: cerebral cortex, HIP (Savic et al. 2005, Bonin et al. 2008)
Abbreviations: extra - extrasynaptical receptor, syn - synaptical receptor, IN – interneuron, PN - pyramidal neuron
The diagram is not suitable to compare the absolute levels of various subunits in either brain region. The distribution of GABA receptor mRNA is not indicated. (Nothdurfter et al. 2012b)

Material and Methods Preparation of brain slices
16
2 Material and Methods
To observe the effects of XBD, diazepam and allopregnanolone in the desired brain regions, brain
slices of mice were prepared. The respective slices were subsequently stained with a voltage
sensitive fluorescent dye, that allowed monitoring changes in signal transduction of neuronal
networks due to substance application (enhancement of GABAergic inhibition) using VSDI.
A detailed description of all applied solutions is listed in the appendix.
2.1 Preparation of brain slices
All experiments were performed on 3 to 8 week old male C57Bl6 mice. The rodents were
decapitated under sedation with the inhalation anesthetic Isofluran. All steps of the preparation
henceforth were realized in ice-cold sucrose-based-ACSF (artificial cerebrospinal fluid) saturated
with carbogen (95% oxygen / 5% carbon dioxide). Access to the brain was achieved after stripping
the skull by two transversal cuts from caudal and a shallow guided cut along the crown’s midline.
The divided cranium halves were bent upwards with forceps, the Nervi optici torn and the brain
carefully removed from the cranial cavity using a rounded spatula. Depending on the desired slice
the brain was prepared further differently. The obtained plain surface was then glued to a metal
block with a cyanoacrylate-glue (Histoacryl, B. Braun, Melsungen, Germany).
Coronary brain slices
In order to get a preparation of the amygdala, the cerebellum was cut off with a razor blade in a
coronary manner.
Sagittal brain slices
For preparation of sagittal brain slices to reach the hippocampus the brain was divided into its
hemispheres via a sagittal cut after removing the cerebellum.
Thalamocortical brain slices
The thalamocortical connections were obtained using a method developed by Agmon and Connors
(Agmon et al. 1991). This preparation allows preservation of the ventrobasal nucleus of the
thalamus, the barrel cortex (somatosensory cortex of rodents) and the fiber tract connecting them.
To achieve intact connections the forebrain needs to be cut off at specific angles (55° sagittal, 10°
horizontal).

Material and Methods Voltage sensitive dye imaging
17
Subsequently, regardless of the desired preparation, the metal block with the glued on brain was
integrated into a vibratome (HM 650 V, Microm International, Walldorf, Germany). The brain has
been cut into slices of 350 µm (amygdala, hippocampus) and of 550 µm thickness (thalamocortical
slices; Fig. 9, p. 21). With the wide end of a Pasteur pipette the slices were transferred into a
transportable chamber filled with general ACSF continuously saturated with carbogen. For the
following 30 minutes they were incubated in a 34°C hot water bath, before being dyed, to allow the
slices to rest and recover from the stress of slice preparation.
2.2 Voltage sensitive dye imaging
VSDI is an imaging technique that allows monitoring of neuronal electric networks in high temporal
and spatial resolution imaging down to 20-50 µm and a millisecond (Chemla et al. 2010). Offering
the great advantage of visualizing the processing of neuronal activity in real time, VSDI provides a
new perspective in the field of neuroscience. Key element to this technique is the voltage sensitive
dye (VSD), which infiltrates organic tissue and binds to the external surface of cell membranes
without interrupting their usual function (Loew 2015). In addition to fluorescent properties, voltage-
sensitive-dyes feature the ability to change its emission spectrum according to the surrounding
charge. Therefore changes in membrane voltage are displayed as changes in fluorescence. This
feature allows the fast monitoring of neuronal signal transduction. Validation and accuracy of VSDI
has been confirmed by close correlation of the optical signal of VSDI recordings and changes in
membrane potential measured via conventional electrophysiological means (Davila et al. 1973,
Salzberg et al. 1973, Morad et al. 1979, Windisch et al. 1985, Chien et al. 1991). Since VSDI was
first introduced in 1968 (Tasaki et al. 1968) it has further improved thanks to the development of
new and superior dyes and progress in technology.
2.2.1 Staining of brain slices with Di-4-ANEPPS
In this dissertation brain slices were stained with Di-4-ANEPPS ((4-(2-(6-(Dibutylamino)-2-
naphthalenyl)ethenyl)-1-(3-sulfopropyl)pyridinium hydroxide inner salt; Sigma-Aldrich Chemie
GmbH; Schnelldorf; Germany), a well-established and commonly used fast acting voltage sensitive
dye (Fluhler et al. 1985, Thiele 2008, Stepan et al. 2012), which allows the monitoring of voltage
changes within a millisecond range. When excited with light of wavelengths between 350 to 550
nm it emits light with longer wavelengths (about 500 to 800 nm) (stokes shift). If the surrounding
tissue is electrically excited the fluorescence shifts to lower wavelength due to a shift of the
absorption, excitation and emission spectrum caused by the dyes electrochromism characteristic
(Loew et al. 1992). Therefore differences of tissue charge can be measured by assessing the
intensity of emission equal and higher than a certain wavelength as demonstrated in the figure
below (Fig. 7). In addition due to the change of excitation spectrum when the surrounding tissue is

Material and Methods Voltage sensitive dye imaging
18
depolarized, the dye will be less excited (presupposed the same spectrum of light enlightens the
dye) and consequently a further reduction of emission of fluorescence occurs (see Fig. 7).
Fig. 7: Schematic sketch of the principle of voltage sensitive shift of emission
Modified according to (Loew et al. 1992). Depending on the voltage of the surrounding tissue the emission spectrum varies due to the electrochromism characteristic of voltage sensitive dyes. As depolarization advances the excitation and emission spectrum of the dye changes (depicted by the arrows in the middle diagram). In addition due to the change of excitation spectrum when the surrounding tissue is depolarized, the dye will be less excited (presupposed the same spectrum of light enlightens the dye) and consequently a further reduction of emission of fluorescence occurs (depicted by the arrow in the last diagram). Therefore changes in membrane voltage of the surrounding tissue can be calculated by considering the intensity of the fluorescence greater or equal of a certain wavelength (represented as the green area).
Depending on the investigated region of the brain and therefore variable necessity of tissue
magnification, the setup was build up differently, with or without a Barlow lens. Since the Barlow
lens also affected exposure to light, which in turn affected excitation and emission of the dye, two
different dyeing methods were applied.
Staining procedure for slices of the amygdala and thalamocortical connections
Di-4-ANEPPS, aliquoted in DMSO (10 mg/ml) was stored at -20°C. Before staining the dye was
incubated at 50° for 5 minutes and thoroughly vortexed. 7.5 µl were properly mixed in with 10 ml
-90 mV
0 mV

Material and Methods Voltage sensitive dye imaging
19
ACSF (final concentration of about 0.0075 mg/ml) and filled into a petri dish continually aerated
with carbogen. Here the slices were stained for 15 minutes in a shaded room.
Staining procedure for hippocampal slices
Initially 5 mg of Di-4-ANEPPS were aliquoted in 1 ml DMSO and 0.5 ml 10-%-El-Cremophor-
solution and kept at -20°C. For staining 40 µl of the aliquot were blended in with 0.5 ml ACSF and
0.5 ml fetal bovine serum. The slices were dyed in the resulting staining solution under continuous
aeration with carbogen for about 20 minutes in a darkened room.
After either staining procedure excessive dye was rinsed off with fresh ACSF. Following the slices
were transferred to a shaded, ACSF-filled, continuously carbogen-aerated holding chamber where
they were allowed to rest for at least 30 minutes before starting measurements. From this point on
and forward Bicuculline (GABAA-R-antagonist) was always added to the ACSF to reach a
concentration of 0.6 µMolar. This step was necessary to assure an adequate signal for two reasons
(Stepan et al. 2012). First, Di-4-ANEPPS potentiates GABA function at the GABAA-receptor
(Mennerick et al. 2010) and second due to their in comparison longer range, glutamatergic axons
are cut to a greater extend than the usually relatively shorter ranged axons of GABAergic
interneurons during brain slice preparation (Stepan et al. 2012).
2.2.2 VSDI Setup
The VSDI-Setup including recording chamber, microscope and electrodes was safely positioned
on a vibration-cushioned table (via pressurized air). To avoid electric noise it was encased by a
Faraday cage and earthed. The employed epifluorescence microscope (BX51 WI, Olympus,
Hamburg, Germany) was equipped with a MiCAM02 HR-CCD camera and processor (BrainVision
Inc., Tokyo, Japan) and an XLFluor4X/340 objective lens (NA 0.28; Olympus, Hamburg, Germany).
A motorized micromanipulator (Luigs and Neumann, Ratingen, Germany) was enabled to manage
movement of the microscope, making it possible to arrange it in all three dimensions without
affecting the recording chamber.
Source of light was a halogen bulb (MHAB-150 W; Moritex corp., China). A software-triggered
shutter controlled light exposure. A band-pass excitation filter ensured that only light of 480-550
nm wavelengths illuminated the slice. A dichroitic mirror (570 nm) reflected the light, projecting it
onto the slice. The fluorescence however, being of longer wavelength, passes through the dichroitic
mirror and through a following long-pass emission filter (590 nm). Thus ensuring that only the dye-
emitted fluorescence is detected by the CCD sensor chip and that the resulting signal is unaltered
by excitation radiation and potential other fluorescence (Fig. 8A). The recorded signal is then
processed as normalized differences of fluorescence intensity [∆F/F] in each pixel over time and
presented as a color-coded sequence-film on the monitor (Fig. 8B).

Material and Methods Voltage sensitive dye imaging
20
A B
Fig. 8: Schematic sketch of the VSDI-setup and presentation of the processed signal
A: schematic sketch of the VSDI-setup, slightly modified according to (Mühlpfordt et al. 2008). Bulb light was filtered by a band-pass excitation filter and projected onto the slice by a dichroitic mirror. Fluorescence emerging from the stained slice passed through the dichroitic mirror and through a following long pass emission filter to the CCD-sensor (detector). B: Color-coded presentation in BrainVision of the processed CCD-camera-recorded signal projected onto a live-picture of the employed brain slice.
2.2.3 Recordings
The slices were carefully positioned in the recording chamber and fixed by a grid (thin nylon threads
tensely attached to a platinum frame). Continuously carbonated ACSF perfused the recording
chamber throughout the entire experiment at a flow rate of about 5-8 ml/min. Experiments were
conducted at room temperature to increase tolerance of hypoxia. To monitor changes in signal
transduction of neuronal networks by GABAergic inhibition under substance influence, the slices
were artificially electrically excited. For excitation of the to examine brain regions a bipolar tungsten
electrode was carefully placed visually. For analysis only the desired regions were considered.
Therefore a ROI (region of interest) was placed to include the area under investigation.
Basolateral amygdala:
The electrode was cautiously positioned inside the amygdala right next to the branching point of
the external capsule that encloses the amygdala (Fig. 9A). The ROI was put to include the
basolateral amygdala.
CA1 region of the hippocampus:
In this preparation the Schaffer collateral was stimulated and the signal obtained from the CA1
region of the hippocampus innervated by it (Fig. 9B).
dF/Fmax (%)

Material and Methods Voltage sensitive dye imaging
21
Thalamocortical preparation:
In this slice preparation two electrodes were positioned (Fig. 9C), one in the ventrobasal thalamus
and another one where layer 4 of the cortex was suspected to be and a total of three areas were
investigated, including the thalamus, the thalamocortical connections and the cortex itself to not
only examine whether the substances have an effect but also to distinguish at which point the
thalamocortical connectivity is being manipulated. Therefore the ROI was positioned to include the
cortex in both stimulation techniques and the thalamus as a second area of interest when
stimulated inside the thalamus.
A B C
Fig. 9: Display of the slice preparations and electrode and ROI placement
White arrows indicate electrode placement. A: Amygdala preparation, the electrode was placed next to the branching point of the external capsule to stimulate the basolateral and lateral amygdala (yellow frame). B: Hippocampus preparation, the electrode was placed to excite the Schaffer collateral, which stimulate the CA1 region of the hippocampus (green). C: Thalamocortical preparation, two electrodes were placed, one in the thalamus to stimulate thalamocortical connections and one in the cortex. Region of interests were the cortex (blue) stimulated both ways and the thalamus (red) stimulated by the electrode placed in the thalamus.
The intensity and duration of pulse stimulation was adjusted via isolated stimulators (type 2533,
Devices Instruments, Welwyn Garden City, England). Single square pulses were applied to
determine the necessary stimulus intensity (0.1-100 V) and length (0.05-0.2 ms). They were set for
each experiment individually in order to generate a similar intense signal throughout all
experiments.
Recording settings were adjusted by BrainVision software (BrainVision 13.04.20, BrainVision Inc.,
Tokyo, Japan). 512 frames were recorded with a sample rate of 2.2 ms per frame, allowing a spatial
resolution of 88*60 pixel with a pixel size of 8.4*9.8 µm in the hippocampus preparations and
36.4*40.0 µm in the amygdala and thalamocortical slices. The resulting 512 images were put
together as a sequence to a film of 1.126 seconds. All electric stimuli were applied in the 103rd
Frame, except for the pulses set in the thalamus, which were released in the 48th Frame.
Recordings were set to record 16 sequences of 512 frames every 6.5 minutes. To improve signal
to noise ratio 16 recorded sequences were averaged, resulting in one averaged sequence every

Material and Methods Data analysis
22
6.5 minutes. This recording protocol was altered for thalamocortical slice measurements, since
there were two electrodes placed in this preparation. Alternating they stimulated the slice,
producing an averaged sequence made of 16 recordings each in a time frame of 11.5 minutes. The
protocol was controlled by WinLTP 1.11b software (Anderson and Collingridge, University Bristol,
UK), which triggered the stimulators and the shutter (controlling light exposure) according to the
set protocol. To spatially and temporally smooth ∆F/F values a 3x3x3 average filter was applied for
all quantifications in addition to an interpolation function to reduce pixilation of images.
As soon as all settings were adjusted measurements started. Once there has been a steady signal
in the desired region for at least 19 minutes (3 control movies), the investigated substance was
applied (XBD: 3 µM, diazepam: 1 µM, allopregnanolone: 1 µM) and recordings continued for about
an hour. Therefore, in addition to the 3 control recordings, 8 more recordings of substance influence
were achieved in amygdala and hippocampal slices or in case of thalamocortical preparation 5
recordings of substance influence. In experiments with diazepam flumazenil (10 µM) was added
afterwards to antagonize diazepam. In addition experiments with finasteride (10 µM) and XBD were
performed, to inhibit XBD effects. Different from flumazenil, finasteride needed to be applied at the
same time as XBD. XBD’s mode of action is via enhancement of neurosteroid genesis, which then
in turn carry out the actual impact. As a 5α-reductase-inhibitor, finasteride prevents neurosteroids
from being synthesized, but cannot antagonize effects of already synthesized neurosteroids. To
avoid data interference by possible effects of finasteride itself it was already added to the ACSF
during control recordings. Finally, to avoid interference by potential (though unlikely, considering
the small amounts) cell damage caused by the Alcohol (70%) used for XBD aliquotation, the same
amount of Alcohol was also added to the ACSF during control recordings in all experiments
containing XBD medication.
2.3 Data analysis
The measured data was presented in BrainVision as a color-coded signal in a sequence-film of
1.126 seconds. This signal can be broken down into three main components: Intensity, spread and
duration of the excitation. Therefore BrainVision-data was processed with a custom programmed
macro in Matlab (Matlab R2008b, The MathWorks, Natick, USA). Hereby the BrainVision acquired
fluorescence inside the ROI representing the fast depolarization signal (FDS) was broken down
into descriptive and comparable numerical values as described below:
The ‘area’ is a numeric value counting all active pixel. An active pixel is defined as a pixel inside
the ROI where the change of fluorescence (∆F/F) was at least three times higher than the standard
deviation of random background noise at anytime within the 512 frames. It is a parameter to
determine the spread of the excitation upon stimulation.

Material and Methods Data analysis
23
The ‘FDSmaxInt’ [∆F/F] is a value describing the mean of the maximal change of fluorescence
throughout all frames of every active pixel within a ROI.
The value ‘FDSareaInt’ [∆F/F] represents the sum of the intensities of every active pixel for each of
the 512 frames within a defined ROI. It will be displayed as a graph of FDSareaInt values over time.
Since neuronal activity propagation upon stimulus only lasts for about 50-300 ms, depending on
the brain area, and since the dye bleaches over time, only the first 228 frames (500 ms) will be
presented in this dissertation.
The ‘amplitude of FDSareaInt’ is a parameter that amounts to the maximal FDSareaInt value under
substance influence normalized to the maximal FDSareaInt value of the respective control recording,
basically representing the maximal amplitude of the FDSareaInt graph (peak of neuronal activity)
over the 550 frames. This parameter depicts the effect the substances have on propagation and
intensity of neuronal activity upon stimulation.
The product ‘FDSAUC’ [∆F/F] is a value designed to give an impression of a summarized overall
effect. It includes the compounds propagation (area), Intensity (Int) and duration of excitation. It is
calculated by adding the values of ‘FDSareaInt’ [∆F/F] of all frames from pulse release till excitation
fade-out, basically resembling the area under the curve (AUC) of the graph depicting the FDSareaInt.
To allow comparability the data was normalized to its respective control (all values were divided by
the mean value of the last 19.8 ms before electric stimulation of the respective three controls). The
results are present by comparing the last recording of the substance to the last control recording.
They will be featured as mean ± SEM (standard error of the mean). Diagrams were created in Excel
(Excel 2003, Microsoft Corporation, Redmond, USA).
Statistic significance was investigated with either the Wilcoxon signed rank test or the Man-
Whitney-U test depending on pairing. Level of significance was defined as α = 0.05 and is indicated
by asterisks as followed: * for p < 0.05, ** for p < 0.02 and *** for p < 0.01.

Results Alteration of neuronal activity upon stimulus in the Amygdala by diazepam, XBD and
allopregnanolone
24
3 Results
Several rows of experiments in the amygdala, the hippocampus and the thalamocortical
connections were performed as described above. In the figure below an exemplary excitation
transmission of neuronal activity in the cortex upon thalamic stimulation is depicted.
Fig. 10 Excitation transmission in the thalamocortical circuit
Depicted is the thalamocortical cut, the first picture is before stimulation (A), then stimulus is released by the electrode in the thalamus (B). The stimulus spreads throughout the thalamus (C) before exciting the thalamocortical connections and hitting the cortex (D) where it spreads (E-F) before fading away again (G-H).
3.1 Alteration of neuronal activity upon stimulus in the Amygdala by diazepam,
XBD and allopregnanolone
Since the amygdala is the crucial area for those information processing related to fear and anxiety
and hence, initiates referring actions (see ch. 1.7) a set of experiments was performed in this brain
region.
3.1.1 Effect of diazepam on neuronal activity upon stimulus in the amygdala
Diazepam application to amygdala brain slices resulted in a decrease of the stimulated excitation
transmission in the basolateral amygdala. As depicted in the graph below (Fig. 11A), FDSareaInt
showed a decrease throughout stimulated excitation transmission by diazepam (amplitude of
FDSareaInt 81.0 ± 8.2 %, n=6). FDSAUC showed a distinct though not significant decrease to 85.0 ±
12.8 % (n=6) in average. Hereby especially the FDSmaxInt of neuronal activity upon stimulus was
reduced (82.2 ± 0.05 %, n=6, p<0.05) (Fig. 11B), while the area remained quite stable (98.2 ± 6.6
%, n=6) (Fig. 11C). The subsequent application of the benzodiazepine antagonist flumazenil
A B C D
E F G H
dF/Fmax
(%)

Results Alteration of neuronal activity upon stimulus in the Amygdala by diazepam, XBD and
allopregnanolone
25
resulted in an increase of the diminished neuronal activity (FDSAUC 100.6 ± 12.8 %, n=6),
successfully though not significantly antagonizing diazepam effect.
A
B C
Fig. 11: Diazepam impact on neuronal activity in the amygdala
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and diazepam (red) recordings. On the right, the maximal normalized FDSareaInt values of control and diazepam (diaz, 0.81 ± 0.08, n=6) and 10 µg flumazenil (fluma, 0.92 ± 0.08, n=6) recordings in the amygdala are depicted, representing the normalized amplitude of their peak values. B: Effect of 1 µM diazepam (red) on FDSmaxInt (∆F/F) of the VSDI-signal in the amygdala. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. Distinct and significant decrease of FDSmaxInt by diazepam 0.82 ± 0.05 (n=6, p<0.5). C: No significant alteration of area in the amygdala under 1 µM diazepam 0.98 ± 0.07 (n=6).
3.1.2 Effect of XBD on neuronal activity upon stimulus in the amygdala
XBD application to amygdala brain slices resulted in a decrease of the stimulated excitation
transmission in the basolateral amygdala. As depicted in the graph below (Fig. 12A), FDSareaInt
showed a decrease throughout stimulated excitation transmission by XBD (amplitude of FDSareaInt
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
FD
S max
Int
0
5
10
15
20
25
30
35
40
45
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Amygdala
0,81 0,920
0,2
0,4
0,6
0,8
1
1,2
Control Diaz Fluma
no
rm. a
mp
litu
de
of
FDS a
reaI
nt
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
are
a

Results Alteration of neuronal activity upon stimulus in the Amygdala by diazepam, XBD and
allopregnanolone
26
83.3 ± 4.8 %, n=6, p<0.05). FDSAUC showed a distinct though not significant decrease to 86.3 ± 8.5
% (n=6) in average. Hereby especially the FDSmaxInt of neuronal activity upon stimulation was
reduced (87.4 ± 2.3 %, n=6, p<0.05) (Fig. 12B), while the area remained quite stable (95.6 ± 2.9%,
n=6) (Fig. 12C). XBD effect could not be significantly antagonized by joint application with
finasteride (amplitude of FDSareaInt 87.9 ± 7.0, n=6).
A
B C
Fig. 12: XBD impact on neuronal activity in the amygdala
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and XBD (green) recordings. On the right, the maximal normalized FDSareaInt values of, XBD (0.83 ± 0.05, n=6, p<0.05) and XBD plus finasteride (x/fina, 0.88 ± 0.07, n=6) recordings in the amygdala are depicted, representing the normalized amplitude of peak values. The black bar represents both control recordings. B: Effect of 3 µM XBD on FDSmaxInt (∆F/F) of the VSDI-signal in the amygdala. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. Distinct and significant decrease of FDSmaxInt by XBD 0.87 ± 0.03 (n=6, p<0.05). C: No significant alteration of area in the amygdala under 3 µM XBD influence 0.96 ± 0.03 (n=6).
0,4
0,6
0,8
1
1,2
1,4
Control XBD
no
rmal
ized
are
a
0,4
0,6
0,8
1
1,2
1,4
Control XBD
no
rmal
ized
FD
S max
Int
0
5
10
15
20
25
30
35
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Amygdala
0,83 0,880
0,2
0,4
0,6
0,8
1
1,2
Control XBD X/Finan
orm
. am
plit
ud
e o
f FD
S are
aIn
t

Results Alteration of neuronal activity upon stimulus in the Amygdala by diazepam, XBD and
allopregnanolone
27
3.1.3 Effect of allopregnanolone on neuronal activity upon stimulus in the amygdala
Allopregnanolone application to amygdala brain slices resulted in a decrease of the stimulated
excitation transmission in the basolateral amygdala (Fig. 13A). As depicted in the graph below (Fig.
13A), FDSareaInt showed a decrease throughout stimulated excitation transmission by
allopregnanolone (amplitude of FDSareaInt 88.9 ± 3.8 %, n=6, p<0.05). FDSAUC showed a distinct
though not significant decrease to 80.5 ± 6.0 % (n=6) in average. Hereby especially the FDSmaxInt
of neuronal activity upon stimulus was reduced (87.6 ± 4.3 %, n=6, p<0.05) (Fig. 13B), while the
area remained quite stable (99.1 ± 5.1 %, n=6) (Fig. 13C).
A
B C
Fig. 13: allopregnanolone impact on neuronal activity in the amygdala
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and allopregnanolone (yellow) recordings. On the right, the maximal normalized FDSareaInt values of control and allopregnanolone (allo, 0.89 ± 0.04, n=6, p<0.05) recordings in the amygdala are depicted, representing the normalized amplitude of peak values. B: Effect of 1 µM allopregnanolone on FDSmaxInt (∆F/F) of the VSDI-signal in the amygdala. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. Distinct and significant decrease of FDSmaxInt by allopregnanolone 0.88 ± 0.04 (n=6, p<0.05). C: No significant alteration of area in the amygdala under 1 µM allopregnanolone 0.99 ± 0.05 (n=6).
0,4
0,6
0,8
1
1,2
1,4
Control Allo
no
rmal
ized
FD
S max
Int
0,4
0,6
0,8
1
1,2
1,4
Control Allo
no
rmal
ized
are
a
0
5
10
15
20
25
30
35
2,2 222,2 442,2
no
rmal
ized
FD
S aea
Int
time [ms]
Amygdala
0,890
0,2
0,4
0,6
0,8
1
1,2
Control Allo
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results Alternation of neuronal activity upon stimulus in the hippocampus by diazepam, XBD and
allopregnanolone
28
3.1.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in
the amygdala
A
B
C
Fig. 14 Comparison of substance effects on FDSAUC, FDSmaxInt and area in the amygdala
Bar chart depicting diazepam (red), XBD (green) and allopregnanolone (yellow) effect on FDSmaxInt (A), area (B) and FDSAUC (C) in the amygdala. The rounded values are displayed on the bottom of the bar chart, since they are each normalized to their respective controls, the value of each control is set at 1.00 and only displayed once, representing all controls. Fault indicators demonstrate standard error of the mean, for exact values see chapter 3.1. Asterisks illustrate significant change between the substance and it’s respective control.
3.2 Alternation of neuronal activity upon stimulus in the hippocampus by
diazepam, XBD and allopregnanolone
As the brain area that is essentially involved in memory formation, the hippocampus portrays the
brain area the next set of experiments was realized in.
3.2.1 Effect of diazepam on neuronal activity upon stimulus in the hippocampus
Diazepam application to hippocampal brain slices resulted in a decrease of the stimulated exci-
tation transmission in the basolateral amygdala (FDSAUC 82.2 ± 7.90 %, n=7). As depicted in the
1,00 0,82 0,87 0,880
0,2
0,4
0,6
0,8
1
1,2
Control Diaz XBD Allo
no
rmal
ized
FD
S max
Int
1,00 0,98 0,96 0,990
0,2
0,4
0,6
0,8
1
1,2
Control Diaz XBD Allo
no
rmal
ized
are
a
1,00 0,85 0,86 0,810
0,2
0,4
0,6
0,8
1
1,2
Co Di XBD Al
no
rmal
ized
FD
S AU
C

Results Alternation of neuronal activity upon stimulus in the hippocampus by diazepam, XBD and
allopregnanolone
29
graph below (Fig. 15A), FDSareaInt showed a distinct decrease throughout stimulated excitation
transmission by diazepam (amplitude of FDSareaInt 84.4 ± 4.5 %, n=7, p<0.02). Hereby intensity of
neuronal activity upon stimulation was distinctly though not significantly reduced in average to 90.4
± 4.2 % (n=7) (Fig. 15B). The decrease in area was significant and averaged to 88.0 ± 4.1 % (n=7,
p<0.02) of initial stimulus propagation (Fig. 15C). Subsequent admission of benzodiazepine
antagonist flumazenil resulted in significant increase of the diminished excitation transmission (area
= 101.3 ± 2.7 %, n=7, p<0.05), however increase of iazepam-diminished FDSmaxInt (91.6 ± 6.4 %,
n=7) and FDSAUC (85.6 ± 8.0, n=7) by flumazenil was only slight and not significant.
A
B C
Fig. 15: diazepam impact on neuronal activity in the hippocampus
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and diazepam (red) recordings. On the right, the maximal normalized FDSareaInt values of control and diazepam (diaz, 0.84 ± 0.04, n=7, p<0.02) and 10 µg flumazenil (fluma, 0.91 ± 0.07, n=7) recordings in the hippocampus are depicted, representing the normalized amplitude of their peak values. B: Effect of 1 µM diazepam (red) on FDSmaxInt (∆F/F) of the VSDI-signal in the hippocampus. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. Distinct decrease of intensity by diazepam 0.90 ± 0.04 (n=7). C: Effect on the area in the hippocampus. Significant reduction of area under 1 µM diazepam 0.88 ± 0.04 (n=7, p<0.02).
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
FD
S max
Int
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
are
a
0
5
10
15
20
25
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time[ms]
Hippocampus
0,84 0,910
0,2
0,4
0,6
0,8
1
1,2
Control Diaz Fluma
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results Alternation of neuronal activity upon stimulus in the hippocampus by diazepam, XBD and
allopregnanolone
30
3.2.2 Effect of XBD on neuronal activity upon stimulus in the hippocampus
XBD application to hippocampal brain slices did not result in a significant alteration of the stimulated
excitation transmission in the basolateral amygdala (FDSAUC = 96.1 ± 7.8 %, n=7) (Fig. 16A).
Neither amplitude of FDSareaInt (93.2 ± 4.2 %, n=7), FDSmaxInt (93.5 ± 4.8 %, n=7) nor the area
(103.5 ± 4.8 %, n=7) of neuronal activity upon stimulation showed significant alteration under XBD
influence (Fig. 16B,C).
A
B C
Fig. 16: XBD impact on neuronal activity in the hippocampus
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 frames of control (black) and XBD (green) recordings. On the right, the maximal normalized FDSareaInt values of control and XBD (0.93 ± 0.04, n=7) recordings in the hippocampus are depicted, representing the normalized amplitude of their peak values. B: Effect of 3 µM XBD (green) on FDSmaxInt (∆F/F) of the VSDI-signal in the hippocampus. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. No significant decrease of FDSmaxInt by XBD 0.94 ± 0.05 (n=7). C No significant alteration of area under 3 µM XBD influence in the hippocampus 1.04 ± 0.05 (n=7).
0,4
0,6
0,8
1
1,2
1,4
Control XBD
no
rmal
ized
FD
S max
Int
0,4
0,6
0,8
1
1,2
1,4
Control XBD
no
rmal
ized
are
a
0
2
4
6
8
10
12
14
16
18
20
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Hippocampus
0,930
0,2
0,4
0,6
0,8
1
1,2
Control XBD
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results Alternation of neuronal activity upon stimulus in the hippocampus by diazepam, XBD and
allopregnanolone
31
3.2.3 Effect of allopregnanolone on neuronal activity upon stimulus in the hippocampus
Allopregnanolone application to hippocampal brain slices resulted in a decrease of the stimulated
excitation transmission in the basolateral amygdala (Fig. 17A). As depicted in the graph below (Fig.
17A), FDSareaInt showed a distinct decrease throughout stimulated excitation transmission by
allopregnanolone (amplitude of FDSareaInt 79.5 ± 4.1 %, n=6, p<0.05). FDSAUC showed a distinct
and significant decrease to 77.2 ± 5.0 % (n=6, p<0.05) in average. Hereby only the intensity of
neuronal activity upon stimulation was reduced (82.0 ± 3.0 %, n=6, p<0.05) (Fig. 17B), while the
area of activity propagation remained quite stable (97.1 ± 2.2 %, n=6) (Fig. 17C).
A
B C
Fig. 17: allopregnanolone impact on neuronal activity in the hippocampus
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 frames of control (black) and allopregnanolone (yellow) recordings. On the right, the maximal normalized FDSareaInt values of control and allopregnanolone (allo, 0.80 ± 0.04, n=6, p<0.05) recordings in the hippocampus are depicted, representing the normalized amplitude of their peak values. B: Effect of 1 µM allopregnanolone (yellow) on FDSmaxInt (∆F/F) of the VSDI-signal in the hippocampus. Control recordings are illustrated as black squares. The grey arrow indicates the average effect. Distinct and significant decrease of FDSmaxInt by allopregnanolone 0.82 ± 0.03 (n=6, p<0.05). C: Effect of 1 µM allopregnanolone on area of stimulus propagation in the amygdala. No significant alteration of area under allopregnanolone 0.97 ± 0.02 (n=6).
0,4
0,6
0,8
1
1,2
1,4
Control Allo
no
rmal
ized
FD
S max
Int
0,4
0,6
0,8
1
1,2
1,4
Control Allo
no
rmal
ized
are
a
0
5
10
15
20
25
30
35
2,2 222,2 442,2
no
mra
lized
FD
S are
aIn
t
time [ms]
Hippocampus
0,800
0,2
0,4
0,6
0,8
1
1,2
Control Allo
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results
32
3.2.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in
the hippocampus
A
B
C
Fig. 18 Comparison of substance effects on FDSAUC, FDSmaxInt and area in the hippocampus
Bar chart depicting diazepam (red), XBD (green) and allopregnanolone (yellow) effect on FDSmaxInt (A), area (B) and FDSAUC (C) in the hippocampus. The rounded values are displayed on the bottom of the bar chart, since they are each normalized to their respective controls, the value of each control is set at 1.00 and only displayed once, representing all controls. Fault indicators demonstrate standard error of the mean, for exact values see chapter 3.2. Asterisks illustrate significant change between the substance and it’s respective control (one asterisks indicates p<0.05, double asterisks indicate p<0.02).
1,00 0,90 0,94 0,820
0,2
0,4
0,6
0,8
1
1,2
Control Diaz XBD Allo
no
rmal
ized
FD
S max
Int
1,00 0,88 1,04 0,970
0,2
0,4
0,6
0,8
1
1,2
Control Diaz XBD Allo
no
rmal
ized
are
a
1,00 0,82 0,96 0,770
0,2
0,4
0,6
0,8
1
1,2
Co Di XBD Al
no
rmal
ized
FD
S AU
C

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
33
3.3 Alternation on neuronal activity upon stimulus in the thalamocortical
connections by diazepam, XBD and allopregnanolone
Two questions were addressed with the measurement in the thalamocortical connections. First, is
there an effect of the corresponding substances and second, if so, at which point within the
thalamocortical system does the effect occur: right in the thalamus, the connecting TC neurons or
within the cortex? Therefore, measurements were designed to detected neuronal activity in the
thalamus (intrathalamic measurements) as well as in the cortex (thalamocortical measurements)
upon stimulation in the ventrobasal thalamus. In addition FDS propagation in the cortex was
measured upon intracortical stimulation (intracortical measurements). Therefore an electrode was
additionally placed in layer 4 of the cortex.
3.3.1 Effect of diazepam on neuronal activity upon stimulus in the thalamocortical
connections
Diazepam application to thalamocortical brain slices resulted in a decrease of the FDS propagation.
As depicted in the graph below (Fig. 19A), FDSareaInt showed a distinct decrease of neuronal activity
in the cortex upon thalamic stimulation (amplitude of FDSareaInt 79.5 ± 3.2 %, n=7, p<0.02; FDSAUC
= 76.3 ± 4.9, n=7, p<0.02). Hereby especially FDSmaxInt of neuronal activity upon stimulation was
significantly reduced in average to 84.4 ± 3.11 % (n=7, p<0.02) (Fig. 19B). There was no significant
alteration in area (94.6 ± 3.01 %, n=7) (Fig. 19C). Subsequent admission of benzodiazepine
antagonist flumazenil resulted in distinct increase of the reduced area (FDSmaxInt = 96.0 ± 6.3 %,
n=7; area= 98.9 ± 2.9 %, n=7) (Fig. 19A). Intrathalamic (amplitude of FDSareaInt 87.7 ± 2.4 %, n=7,
p<0.02; FDSAUC = 86.4 ± 4.0 %, n=7) as well as intracortical (amplitude of FDSareaInt 83.3 ± 3.6 %,
n=6, p<0.05; FDSAUC = 81.4 ± 4.6 %, n=6, p<0.05) experiments with diazepam showed similar
results (Fig. 19B,C). Hereby diazepam showed equal impact on thalamocortical and intracortical
excitation propagation, while diazepam effect on the intrathalamic connections was slighter in
comparison. Intrathalamic diazepam impact could not be antagonized by subsequent flumazenil
application, in contrast to thalamocortical and intracortical diazepam effect.

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
34
A
B
C
Fig. 19: diazepam impact on neuronal activity in the thalamocortical connections
A: On the left averaged normalized FDSareaInt values of the first 500 ms of control (black) and diaze-pam (red) recordings are depicted. On the right, the maximal normalized FDSareaInt values of control and diazepam (diaz, 0.80 ± 0.03, n=7, p<0.02) and 10 µg flumazenil (fluma, 0.93 ± 0.07, n=7) recordings in the thalamocortical connections are depicted, representing the normalized amplitude of their peak values. B: Effect of 1 µM diazepam (red) on FDSmaxInt (∆F/F) of the VSDI detected signal in the thalamus (0.90 ± 0.02, n=7, p<0.02), the thalamocortical connections (0.84 ± 0.03, n=7, p<0.02) and intracortical (0.86 ± 0.03, n=6, p<0.05). Control recordings are illustrated as black squares. The grey arrow indicates the average effect. C: Effect of 1 µM diazepam on the area of stimulus propagation in the thalamus (0.94 ± 0.02, n=7, p<0.05), the thalamocortical connections (0.95 ± 0.03, n=7) and intracortical (0.92 ± 0.05, n=6).
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
FD
S max
Int Intrathalamic
Control Diazepam
Thalamocortical
Control Diazepam
Intracortical
0,4
0,6
0,8
1
1,2
1,4
Control Diazepam
no
rmal
ized
are
a
Intrathalamic
Control Diazepam
Thalamocortical
Control Diazepam
Intracortical
0
5
10
15
20
25
30
35
40
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Thalamocortical
0,80 0,930
0,2
0,4
0,6
0,8
1
1,2
Control Diaz Fluma
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
35
3.3.2 Effect of XBD on neuronal activity upon stimulus in the thalamocortical connections
XBD application to thalamocortical brain slices resulted in a decrease of the stimulated excitation
transmission. As depicted in the graph below (Fig. 20A), FDSareaInt showed a distinct decrease of
neuronal activity in the cortex upon thalamic stimulation (amplitude of FDSareaInt 83.5 ± 3.2 %, n=8,
p<0.01; FDSAUC 77.6 ± 8.1%, n=8, p<0.02). Hereby especially FDSmaxInt of neuronal activity upon
stimulation was significantly reduced in average to 86.1 ± 3.3 % (n=8, p<0.01) (Fig. 20B), while
there was no significant alteration in area (102.0 11.5 %, n=8) (Fig. 20C). In experiments with
simultaneous admission of XBD and finasteride, no alteration of excitation propagation was
detectable (FDSmaxInt 101.8 ± 5.3 %, n=7; area 98.9 ± 2.9 %, n=7). Intrathalamic (amplitude of
FDSareaInt 86.2 ± 3.0 %, n=8, p<0.01; FDSAUC 84.2 ± 7.0 %, n=8) as well as intracortical experiments
(amplitude of FDSareaInt 87.4 ± 2.3 %, n=8, p<0.01; FDSAUC 84.6 ± 3.3 %, n=8, p<0.01) with XBD
showed similar results (Fig. 20B,C). Hereby XBD showed equal impact on thalamocortical and
intrathalamic neuronal activity upon stimulation, while intracortical XBD effect was slighter in
comparison. Intrathalamic and intracortical XBD impact could not be antagonized by additional
finasteride application, in contrast to thalamocortical XBD effect.

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
36
A
B
C
Fig. 20: XBD impact on neuronal activity upon stimulus in the thalamocortical circuit
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and XBD (green) recordings. On the right, the maximal normalized FDSareaInt values of XBD (0.83 ± 0.03, n=8, p<0.01) and XBD plus finasteride (X/Fina, 1.00 ± 0.05, n=7) recordings in the thalamocortical connections are depicted, representing the normalized amplitude of their peak values. The black bar represents both their respective control recordings. Additional finasteride application significantly antagonized XBD effects (p<0.05). B: Effect of 3 µM XBD (green) on FDSmaxInt of the VSDI-signal in the thalamus (0.89 ± 0.03, n=8, p<0.02), the thalamocortical connections (0.86 ± 0.03, n=8, p<0.01) and intracortical (0.93 ± 0.03, n=8). Control recordings are illustrated as black squares. The grey arrow indicates the average effect. C: Effect of 3 µM XBD on the area in the thalamus (0.94 ± 0.05, n=8), the thalamocortical connections (1.02 ± 0.11, n=8) and intracortical (0.96 ± 0.04, n=8).
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
Control XBD
no
rmal
ized
are
a
Intrathalamic
Control XBD
Thalamocortical
Control XBD
Intracortical
0,4
0,6
0,8
1
1,2
1,4
Control XBD
no
rmal
ized
FD
S max
Int
Intrathalamic
Control XBD
Thalamocortical
Control XBD
Intracortical
0
1
2
3
4
5
6
7
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Thalamocortical
0,83 1,000
0,2
0,4
0,6
0,8
1
1,2
Control XBD X/Fina
no
rm. a
mp
litu
de
of
FDS a
reaI
nt

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
37
3.3.3 Effect of allopregnanolone on neuronal activity upon stimulus in the thalamocortical
connections
As Fig. 21A demonstrates, allopregnanolone affected the duration of neuronal activity in the cortex
upon thalamic stimulation, as quantified by the parameter FDSAUC = 87.5 ± 9.4 % (n=5) in
comparison to amplitude of FDSareaInt (1.01 ± 6.5 %, n=5). Allopregnanolone application did not
affect FDSmaxInt of intrathalamic (97.6 ± 4.7 %, n=4) and thalamocortical (97.5 ± 5.4 %, n=5)
excitation transmission (Fig. 21B). However, allopregnanolone exertion resulted in decrease of
FDSmaxInt of intracortical neuronal activity upon cortical stimulation to about 81.5 ± 6.7 % (n=5).
Intrathalamic no alteration of area could be detected under allopregnanolone influence (93.8 ± 5.4
%, n=4). Allopregnanolone showed a tendency to increased thalamocortical (113.1 ± 13.3 %, n=5)
and intracortical stimulus propagation (area = 118.6 ± 10.8 %, n=5) (Fig. 21C). None of the
allopregnanolone experiments performed on thalamocortical slices provided significant results.
A
B
0
2
4
6
8
10
12
14
2,2 222,2 442,2
no
rmal
ized
FD
S are
aIn
t
time [ms]
Thalamocortical
1,010
0,2
0,4
0,6
0,8
1
1,2
Control Allo
no
rm. a
mp
litu
de
of
FDS a
reaI
nt
0,4
0,6
0,8
1
1,2
1,4
Control Allo
no
rmal
ized
FD
S max
Int
Intrathalamic
Control Allo
Thalamocortical
Control Allo
Intracortical

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
38
C
Fig. 21: allopregnanolone impact on neuronal activity upon stimulus in the thalamocortical connections
A: The diagram on the left shows averaged normalized FDSareaInt values of the first 500 ms of control (black) and allopregnanolone (yellow) recordings. On the right, the maximal normalized FDSareaInt values of control and allopregnanolone (allo, 1.01 ± 0.07, n=5) recordings in the thalamocortical connections are depicted, representing the normalized amplitude of their peak values. B: Effect of 1 µM allopregnanolone (yellow) on FDSmaxInt (∆F/F) of the VSDI-signal in the intrathalamic (0.98 ± 0.05, n=4), the thalamocortical connections (0.97 ± 0.05, n=5) and intracortical (0.81 ± 0.07, n=5). Control recordings are illustrated as black squares. The grey arrow indicates the average effect C: Effect of 1 µM allopregnanolone on the area of the stimulus propagation in the thalamus (0.94 ± 0.05, n=4), the thalamocortical connections (1.13 ± 0.13, n=5) and intracortical (1.19 ± 0. 11, n=5).
3.3.4 Comparison of diazepam, XBD and allopregnanolone effect on neuronal activity in
the thalamocortical connections
A
Control Allo
Thalamocortical
0,4
0,6
0,8
1
1,2
1,4
1,6
Control Allo
no
rmal
ized
are
aIntrathalamic
Control Allo
Intracortical
1,00 0,90 0,89 0,980
0,2
0,4
0,6
0,8
1
1,2
Co Di XBD Al
no
rmal
ized
FD
S max
Int
Intrathalamic
1,00 0,84 0,86 0,97
Co Di XBD Al
Thalamocortical
1,00 0,86 0,93 0,81
Co Di XBD Al
Intracortical

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
39
B
C
Fig.
22 Comparison of substance effects on FDSAUC FDSmaxInt and area in the thalamocortical connections
The bar charts serve to ease comparison of average effect of diazepam (red), XBD (green) and allopregnanolone (yellow) on FDSmaxInt (A), area (B) and FDSAUC (C) in the thalamocortical connections. The rounded values are displayed on the bottom of the bar chart, since they are each normalized to their respective controls, the value of each control is set at 1.00 and only displayed once, representing all controls. Fault indicators demonstrate standard error of the mean, for exact values see chapter 3.3. Asterisks illustrate significant change between the substance and it’s respective control (one asterisks indicates p<0.05, double asterisks indicate p<0.02).
1,00 0,94 0,94 0,940
0,2
0,4
0,6
0,8
1
1,2
1,4
Co Di XBD Al
no
rmal
ized
are
a
Intrathalamic
1,00 0,95 1,02 1,13
Co Di XBD Al
Thalamocortical
1,00 0,92 0,96 1,19
Co Di XBD Al
Intracortical
1,00 0,76 0,78 0,870
0,2
0,4
0,6
0,8
1
1,2
Co Di XBD Al
Thalamocortical
1,00 0,86 0,84 0,980
0,2
0,4
0,6
0,8
1
1,2
Co Di XBD Al
no
rmal
ized
FD
S AU
C
Intrathalamic
1,00 0,81 0,85 0,81
Co Di XBD Al
Intracortical

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
40
3.3.5 Analyses of diazepam, XBD and allopregnanolone impact on the individual layers of
the cortex
To get more insight into the diazepam, XBD and allopregnanolone mediated impact on
thalamocortical excitation propagation upon stimulation, the change in neuronal activity of each
layer of the cortex inside the ROI upon thalamic stimulation was investigated. For this purpose the
following parameters were analyzed in each layer individually: the maximal change of intensity
(Intmax) and the time till Intmax (delay of Intmax).
The reduction of Intmax by diazepam, XBD and allopregnanolone was distinct throughout all three
substances, however different amongst them in regard of the extend with which each layer was
affected (see
Fig. 24). Diazepam elicited most effect in layer 1 to 3 and was successfully and partially significantly
antagonized by flumazenil. XBD affected layer 1,2,5 and 6 the most. Allopregnanolone seemed to
diminish the maximal intensity of thalamocortical stimulus propagation least of all three substances
and only showed consistent decrease in layer 3 and 4.
Diazepam significantly decreased the delay of Intmax in the 2nd layer (74.7 ± 7.2 %, n=7, p<0.05)
and 4th layer (86.6 ± 4.8 %, n=7, p<0.05) and showed a tendency to decrease delay of Intmax in the
1st layer (89.5 ±7.7 %, n=7) without significance. Flumazenil did not significantly antagonize
diazepam effect on the delay of Intmax. XBD significantly decreased the delay of Intmax in the 4th
layer of the cortex (to 73.4 ± 13.4%, n=8, p<0.05). Simultaneous application of XBD and finasteride
showed to the tendency to but did not significantly prevent XBD effect in the 4th layer of the cortex.
Allopregnanolone application showed the same tendency in the 4th layer without significance (76.8
± 27.9 %, n=5). Apart form the 4th layer and in the case of diazepam also in the 1st and 2nd layer of
the cortex, the evaluation of delay of Intmax did not offer valuable information due to discrepancies
in the results.
Fig. 23 Display of cortex division
for layer analysis
Depiction of the roi (yellow outlining) and the 6 areas (I-VI) presenting the 6 lawyers of the cortex.

Results Alternation on neuronal activity upon stimulus in the thalamocortical connections by diazepam,
XBD and allopregnanolone
41
Fig. 24 Layer analysis of diazepam and XBD impact on normalized maximal Intensity
Depicted is a bar chart of the effect of diazepam (red), XBD (green) and allopregnanolone (yellow) on maximal Intensity normalized to their respective control recordings (black) in each layer (L1-6) of the cortex upon thalamic stimulation. Diazepam and XBD effects are depicted with their respective antagonists. The values are depicted inside the bars. Fault indicators demonstrate standard error of the mean. Asterisks represent significant results (one asterisks: p<0.05, two asterisks: p<0.02).
0,931
0,938
0,848
0,910
0,981
0,984
0 0,2 0,4 0,6 0,8 1 1,2
L1
L2
L3
L4
L5
L6
Kontrolle 1 µM AllopregnanoloneControl
normalized maximal Intensity
0,696
0,795
0,831
0,924
0,890
0,852
0,953
0,957
0,962
1,039
0,902
0,881
0 0,5 1 1,5
L1
L2
L3
L4
L5
L6
Kontrolle 1 µM Diazepam 10 µM FlumazenilControl
0,794
0,803
0,960
0,934
0,859
0,822
0,835
0,905
0,905
1,026
0,953
1,012
0 0,5 1 1,5
L1
L2
L3
L4
L5
L6
Kontrolle 3 µM XBD 10 µM FinasterideControl
normalized maximal Intensity normalized maximal Intensity

Discussion Result interpretation
42
4 Discussion
4.1 Result interpretation
Benzodiazepines such as diazepam and XBD, a ligand of TSPO, elicit similar anxiolytic properties
but dispose of diverging side effect profiles. While diazepam exposes amnestic and sedative
adverse effects as well as tolerance development and risk of addiction, XBD lacks any of those, as
far as experiments on humans and rodents have surveyed (Kita et al. 2009, Rupprecht et al. 2009,
Rote-Liste 2013, 2015). XBD enhances neurosteroid genesis, e.g. allopregnanolone synthesis,
which is responsible for XBD effects (Rupprecht et al. 2009). Both, allopregnanolone and diazepam
are positive allosteric modulators at the GABAA receptor, though they occupy different binding sides
(Hosie et al. 2006, Möhler 2006). Due to diverse GABAA receptor subunit expression, GABAA
receptor binding abilities vary throughout different brain areas, probably accounting for the
discrepancy of diazepam and XBD side effect profile (Pirker et al. 2000). To investigate diazepam,
XBD and allopregnanolone impact on neuronal activity in brain areas relevant to their effects,
alteration of excitation propagation upon stimulation under their influence was detected using VSDI
in the course of this dissertation. Brain areas included in the experiments are the amygdala as the
anxiety-center of the brain (see ch. 1.7), the hippocampus as the brain area where memory
formation occurs (see ch. 1.8) and the thalamocortical system, indispensable for awareness (see
ch. 1.9). Corresponding mice brain slices were prepared for VSDI and signal propagation upon
stimulus under the influence of diazepam, allopregnanolone and XBD was determined.
4.1.1 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the amygdala
The amygdala has been found to be crucially involved in processing emotions, especially anxiety
(see chapter 1.7.2). Diazepam, XBD and allopregnanolone displayed significant negative impact
on neuronal excitability in the amygdala. Hereby all three substances demonstrated similar
inhibition of signal transduction in the amygdala, whereupon especially the Intensity (FDSmaxInt)
rather than the area of signal propagation was affected, indicating that the amount of neurons
activated upon stimulation remained the same but the level of their excitation was reduced by
diazepam, XBD and allopregnanolone.
These results are in line with clinical observations and animal models determining their anxiolytic
potential. Similar anxiolytic properties of XBD and benzodiazepines have been observed in rodents
as well as in humans (Kita et al. 2009, Rupprecht et al. 2009). Comparable anxiolytic properties of
allopregnanolone have been shown in rodents as well (Bitran et al. 1991, Picazo et al. 1995).
Taken together, the anxiolytic effect of all three substances probably origins in their diminishing
effect on neuronal activity in the amygdala. In terms of similarity of XBD and allopregnanolone

Discussion Result interpretation
43
results, it is likely that the impact of XBD on the amygdala is due to allopregnanolone formation.
These results therefore support the data of Rupprecht at al (Rupprecht et al. 2009), which proposes
that XBD effects are mediated by allopregnanolone.
4.1.2 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the
hippocampus
Diazepam displayed distinct impact on neuronal activity in the hippocampus, whereupon the area
of the stimulation was significantly reduced and the intensity distinctly diminished, indicating that
diazepam did not only affect the amount of neurons excited upon stimulation but also reduced the
level of excitation. This appears to explain diazepam’s known adverse effect on memory resulting
in anterograde amnesia (Rote-Liste 2013, 2015). XBD however, did not significantly disturb
neuronal activity upon stimulus in the hippocampus, indicating that XBD administration to subjects
would not result in affection of memory. Just as no affection in memory upon XBD application has
been observed in clinical studies so far (Rupprecht et al. 2009).
Interestingly, allopregnanolone had a significant negative effect on the intensity of signal
transduction in the hippocampus even though XBD showed no significant or even relevant effect
here. However, glia cells located in the hippocampus might lack expression of enzymes needed to
form allopregnanolone out of pregnenolone. If this was the case, allopregnanolone may exert a
negative impact on signal propagation in the hippocampus, but would not be increasingly
synthesized in the hippocampus following XBD administration. Consequently, XBD would not result
in inhibition of signal transduction in the hippocampus and thus not effect memory (assuming other
neurosteroids that positive allosteric modulate at GABAA receptors are not promoted either, which
could be the case, for XBD experiments in the amygdala did not result in suppression of neuronal
activity). XBD would still enhance pregnenolone synthesis and following other neurosteroids in the
hippocampus, but not allopregnanolone or any other positive allosteric GABAA receptor modulating
neurosteroids or XBD promotes diverse neurosteroids, which cancel out each others effects.
Supporting this theory, allopregnanolone has been observed to have a negative impact on memory
(Mayo et al. 1993, Zimmerberg et al. 1995). Further affirming this theory, enzymes for synthesis of
diverse neurosteroids have been determined to be region and neuron specific (Rupprecht et al.
2010). For instance, enzymes needed for formation of 5α-reduced neurosteroids have been found
to be almost absent in GABAergic hippocampal interneurons (Agís-Balboa et al. 2006).
4.1.3 Impact of diazepam, XBD and allopregnanolone on neuronal activity in the
thalamocortical connections
XBD impact on neuronal activity in the thalamocortical system was overall similar to diazepam
effect, with some distinct differences though. While XBD’s effect focused on merely diminishing the
intensity of intrathalamic and thalamocortical activity propagation, diazepam additionally affected

Discussion Result interpretation
44
the intensity in the intracortical experiment and the area of neuronal stimulus propagation in the
intrathalamic neuronal network. Ergo diazepam impact on the thalamocortical connections
exceeded XBD impact.
However, like diazepam, XBD clearly and significantly had an impact on intensity of signal
propagation in the thalamocortical connections arising from the ventrobasal nucleus of the
thalamus leading to the somatosensory cortex. Kita and his research group could not observe any
signs of sedative adverse effects upon XBD application to rodents (Kita et al. 2009). Furthermore,
subjects exposed to XBD intake did not report sedative effects (Rupprecht et al. 2009). In hindsight
it can be debated that a decline in the signal transduction in the thalamocortical connections might
be an additional reason for anxiety dissolution and not a cause for sedation. Hereby the train of
thought is that impairment of the thalamocortical connections might lead to a decrease in
awareness, but not on the physical condition of being awake. If this is the case, XBD might affect
alertness but would not sedate subjects. Encouraging this theory is the fact that the reticular
formation (as a part of the Reticular Activating System) is essentially involved in physical condition
states such as wakefulness and sleep-wake transitions (Trepel 2008) and might therefore be the
working point of the adverse effect of sedation upon diazepam application instead of the
thalamocortical system. To get more insight in this matter it would be interesting to perform clinical
studies investigating possible impact of XBD on awareness, concentration and reaction time. It also
would be interesting to see if XBD and diazepam show an effect in the Ascending Reticular
Activating System (RAS). Since up to date a slice preparation to properly inspect the RAS has not
been developed, there was no way to include this brain area in this dissertation.
The experiments with allopregnanolone in the thalamocortical system showed quite different
results. Allopregnanolone did not significantly affect the intrathalamic neuronal network. It did
however increase the area in the thalamocortical connections. Additionally, allopregnanolone
decreased the intensity of the stimulus propagation intracortical, however it did not in the
thalamocortical connections. The tendency of allopregnanolone to increase area in the
thalamocortical experiments might be considered as scattering of stimulus propagation, implying
that the once accurate and specific projection of excitation from the thalamus to a certain area of
the cortex is now affected and stimulus propagation left unguided.
The discrepancy in allopregnanolone and XBD results in the thalamocortical connections might
indicate that allopregnanolone is not the only neurosteroid promoted upon XBD application in the
thalamocortical connections. Since XBD rendered a quite different outcome, it is to be supposed
that other neurosteroids besides allopregnanolone conducted its actions, which might be due to a
different enzymatic equipment for NS synthesis in the thalamocortical system compared to the
hippocampus.

Discussion Clinical relevance, sources of error and outlook
45
For further insight into substance effect on the thalamocortical connections an analysis of alteration
of neuronal activity in each cortex layer upon thalamic stimulation was additionally undertaken.
While a few tendencies can be derived from this layer analysis (see chapter 4.1.3), due to
discrepancies in the results no ascertained conclusions can be drawn.
4.2 Clinical relevance, sources of error and outlook
While XBD and diazepam concentrations correspond with clinically used dosage, allopregnanolone
concentration was quite elevated in comparison to endogenous found concentrations. Little is
known about the XBD-induced synthesis of other neurosteroids than allopregnanolone. However,
the experiments with allopregnanolone served to determine general discrepancies to XBD. Even
though allopregnanolone has been applied at low doses, the effects of allopregnanolone on
neuronal activity might change remarkably when concentrations are adapted to physiological
levels.
Possible source of interference could have been the voltage sensitive dye di-4-ANEPPS, which is
known to have a modulating effect on GABA-receptors itself (Mennerick et al. 2010). However,
these effects can be considered negligible, since control recordings were equally affected as
recordings in the presence of substances.
Another TSPO ligand, alpidem, has already been approved for anxiety treatment in France in 1991
(Langer et al. 1990). Due to severe liver damage with profound and even lethal consequences it
was withdrawn in 1994 (Barki et al. 1993, Baty et al. 1994, Ausset et al. 1995). However, different
to XBD, alpidem also binds to GABAA receptors directly, which could also be the cause for liver
dysfunction (Langer et al. 1990). Nevertheless, further assessment of possible adverse effects and
tolerance in long-term clinical studies, including studies on diseased patients, are necessary to
investigate long-term tolerability and confirm clinical safety and relevance before XBD can be
introduced as a treatment option in anxiety disorders.
4.3 Conclusion
According to the data collected, XBD reduces neuronal activity in the amygdala, correspondingly
counteracting anxiety, while not significantly modulating neuronal networks in the hippocampus,
explaining the lack of anterograde amnesia. As for its alteration of neuronal activity in the
thalamocortical system, further investigations such as clinical studies determining XBD effect on
alertness need to be realized previous to final conclusions. To further elucidate the discrepancy
concerning the adverse sedative effect, the neurophysiological impact of diazepam and XBD on

Discussion Conclusion
46
brainstem areas involved in sleep-wake rhythms such as the reticular formation are of considerable
interest.
Similar to XBD, diazepam affects neuronal activity in the amygdala and the thalamocortical circuits,
but unlike XBD, also demonstrated diminishing effects on neuronal activity in the hippocampus,
which is in line with clinical observation of anterograde amnesia upon benzodiazepine application
(Rote-Liste 2013, 2015). In comparison, the presented data provides further evidence for XBD as
a promising anxiolytic drug to be superior to diazepam, regarding its side effect profile (Kita et al.
2009, Rupprecht et al. 2009).
The acquired data suggests that allopregnanolone is not the only neurosteroid formed upon XBD
application. This can be drawn from the fact that XBD and allopregnanolone utilization in the
thalamocortical connections resulted in different change of stimulation propagation. Furthermore,
in contrast to XBD, allopregnanolone reduced stimulation propagation in the hippocampus. This
indicates that, while allopregnanolone could affect neuronal networks in the amygdala, XBD
application does not lead to allopregnanolone formation in the hippocampus. This is in line with the
observation that neurosteroid biosynthesis is region and neuron specific and depends on TSPO
abundance and expression of the respective enzymes, responsible for genesis of the diverse
neurosteroids (Rupprecht et al. 2010). Region-specific synthesis of neurosteroids is subsequently
responsible for distinct modulation of neurotransmitter receptors in certain brain areas, resulting in
region-specific alteration of signal transduction. Additionally, distribution of GABAA receptor
subtypes adds to region specific alteration of signal transduction, since they vary in binding
capabilities (Hosie et al. 2006). Therefore, there are diverse parameters relevant for the
pharmacological profile of XBD, including TSPO abundance, distribution of the respective enzymes
needed for neurosteroid syntheses and GABAA receptor subtypes.

Abstract
47
5 Abstract
Despite the high prevalence of anxiety disorders in today’s society, suitable anxiolytic medication
needs yet to be established. Frequently used benzodiazepines show rapidly on-setting anxiolytic
effects. However, they elicit a broad range of adverse effects such as sedation and anterograde
amnesia. XBD, a ligand to the Translocator Protein 18 kDa (TSPO), is a promising new option.
XBD enhances TSPO mediated syntheses of neurosteroids, which have been shown to exert
anxiolytic effects. Clinical studies suggest XBD to lack unfavorable side effects commonly known
in benzodiazepine treatment. Like benzodiazepines, neurosteroids e.g. allopregnanolone act as
positive allosteric modulators at the GABAA receptor, though they occupy another binding site. The
discordance in the side effect profile is thought to origin in GABAA subtype distribution in the brain.
This present study investigates effects of XBD and diazepam on diverse brain areas using an
imaging technique, VSDI, that enables monitoring of signal propagation in the brain. The effects of
XBD and diazepam in diverse brain areas were monitored and compared. While both substances
showed considerable and similar impact on stimulus propagation in the amygdala, the anxiety
center of the brain, only diazepam affected neuronal activity in the hippocampus, the essential brain
area for memory consolidation. The collected data suggest similar effects on neuronal activity in
the amygdala (development of anxiety), while XBD lacks the negative effect of benzodiazepines
on neuronal activity in the hippocampus (known for memory formation). This is concordant with
clinical studies where XBD lacks the adverse effect of anterograde amnesia much known in
benzodiazepine treatment, while exerting similar anxiolytic properties. Like diazepam, XBD
affected neuronal activity in the thalamocortical system, which might be an indicator that XBD could
impair alertness and concentration. To investigate the mechanism of action of XBD, an additional
set of experiments was performed with allopregnanolone as presumably the most important
neurosteroid promoted upon XBD application. The discrepancies in neuronal activity observed after
XBD or allopregnanolone application in different brain regions may be explained by the fact that it
is highly probable that beside allopregnanolone, XBD enhances the syntheses of other
neurosteroids which are also pharmacological active. Furthermore, neurosteroid synthesis upon
TSPO activation depends on the specific enzymatic machinery needed for neurosteroid formation.
Which, at least in the hippocampus, does not allow the synthesis of allopregnanolone. Altogether,
the data collected provides further evidence that XBD promises to be superior to diazepam in
pharmacological treatment of anxiety disorders.

References
48
6 References
Agís-Balboa, R. C., G. Pinna, A. Zhubi, E. Maloku, M. Veldic, E. Costa and A. Guidotti (2006). "Characterization of brain neurons that express enzymes mediating neurosteroid biosynthesis." Proc Natl Acad Sci U S A 103: 14602-14607.
Agmon, A. and B. W. Connors (1991). "Thalamocortical responses of mouse somatosensory (barrel) cortex in vitro." Neuroscience 41: 365-379.
Akwa, Y., R. H. Purdy, G. F. Koob and K. T. Britton (1999). "The amygdala mediates the anxiolytic-like effect of the neurosteroid allopregnanolone in rat." Behav Brain Res 106: 119-125.
Amaral, D. G. (1993). "Emerging principles of intrinsic hippocampal organization." Curr Opin Neurobiol 3: 225-229.
Amaral, D. G., J. L. Price, A. Pitkanen and S. T. Carmichael (1992). he Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. Anatomical organization of the primate amygdaloid complex. New York, Wiley-Liss: 1-66.
Anholt, R. R. H., E. B. De Souza, M. L. Oster-Granite and S. H. Snyder (1985). "Peripheral-type benzodiazepine receptors: autoradiographic localization in whole-body sections of neonatal rats." J Pharmacol Exp Ther 233: 517-526.
Anholt, R. R. H., P. L. Pedersen, E. B. De Souza and S. H. Snyder (1986). "The Peripheral-type Benzodiazepine Receptor - Localization to the mitochondrial outer membrane." J Biol Chem 261: 576-583.
Ausset, P., P. Malavialle, A. Vallet, G. Miremont, B. Le Bail, F. Dumas, J. Saric and S. Winnock (1995). "Subfulminant hepatitis caused by alpidem and treated by liver transplantation." Gastroenterol Clin Biol 19(2): 222-223.
Baldwin, D., I. M. Anderson, D. J. Nutt, C. Allgulander, B. Bandelow, J. A. den Boer, D. M. Christmas, S. Davies, N. Fineberg, N. Lidbetter, A. Malizia, P. McCrone, D. Nabarro, C. O'Neill, J. Scott, N. van der Wee and H. U. Wittchen (2014). "Evidence-based pharmacological treatment of anxiety disorders, post-traumatic stress disorder and obsessive-compulsive disorder: a revision of the 2005 guidelines from the British Association for Psychopharmacology." J Psychopharmacol 28: 403-439.
Bandelow, B., J. Zohar, E. Hollander, S. Kasper, H. J. Möller, C. Allgulander, J. Ayuso-Gutierrez, D. S. Baldwin, R. Buenvicius, G. Cassano, N. Fineberg, L. Gabriels, I. Hindmarch, H. Kaiya, D. F. Klein, M. Lader, Y. Lecrubier, J. P. Lépine, M. R. Liebowitz, J. J. Lopez-Ibor, D. Marazziti, E. C. Miguel, K. S. Oh, M. Preter, R. Rupprecht, M. Sato, V. Starcevic, D. J. Stein, M. van Ameringen and J. Vega (2008). "World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the pharmacological treatment of anxiety, obsessive-compulsive and post-traumatic stress disorders - first revision." World J Biol Psychiatry 9: 248-312.

References
49
Barbaccia, M. L., G. Roscetti, M. Trabucchi, R. H. Purdy, M. C. Mostallino, A. Concas and G. Biggio (1997). "The effects of inhibitors of GABAergic transmission and stress on brain and plasma allopregnanolone concentrations." Br J Pharmacol 120: 1582-1588.
Barki, J., D. Larrey, G. Pageaux, G. Lamblin, J. L. Gineston and H. Michel (1993). "Fatal subfulminant hepatitis during treatment with alpidem (Ananxyl)." Gastroenterol Clin Biol 17(11): 872-874.
Barnard, E. A., P. Skolnick, R. W. Olsen, H. Mohler, W. Sieghart, G. Biggio, C. Braestrup, A. N. Bateson and S. Z. Langer (1998). "International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function." Pharmacol Rev 50(3): 291-313.
Baty, V., B. Denis, C. Goudot, V. Bas, P. Renkes, M. A. Bigard, P. Boissel and P. Gaucher (1994). "Hepatitis induced by alpidem (Ananxyl). Four cases, one of them fatal." Gastroenterol Clin Biol 18(12): 1129-1131.
Belelli, D. and J. J. Lambert (2005). "Neurosteroids: endogenous regulators of the GABA(A) receptor." Nat Rev Neurosci 6(7): 565-575.
Bernassau, J. M., J. L. Reversat, P. Ferrara, D. Caput and G. Lefur (1993). "A 3D model of the peripheral benzodiazepine receptor and its implication in intra mitochondrial cholesterol transport." J Mol Graph 11: 236-244.
Bitran, D., M. Foley, D. Audette, N. Leslie and C. A. Frye (2000). "Activation of peripheral mitochondrial benzodiazepine receptors in the hippocampus stimulates allopregnanolone synthesis and produces anxiolytic-like effects in the rat." Psychopharmacology (Berl) 151: 64-71.
Bitran, D., R. J. Hilvers and C. K. Kellogg (1991). "Anxiolytic effects of 3a-hydroxy-5a[IZ]-pregnan-20-one: endogenous metabolites of progesterone that are active at the GABAAreceptor." Brain Res 561: 157-161.
Bitran, D., M. Shiekh and M. McLeod (1995). "Anxiolytic Effect of Progesterone is Mediated by the Neurosteroid Allopregnanolone at Brain GABAAReceptors." J Neuroendocrinol 7: 171-177.
Bloom, F. E. and L. L. Iversen (1971). "Localizing 3H-GABA in nerve terminals of rat cerebral cortex by electron microscopic autoradiography." Nature 229(5287): 628-630.
Bonin, R. P. and B. A. Orser (2008). "GABA(A) receptor subtypes underlying general anesthesia." Pharmacol Biochem Behav 90: 105-112.
Bormann, J., O. P. Hamill and B. Sakmann (1987). "Mechanism of anion permeation through channels gated by glycine and gamma-aminobutyric acid in mouse cultured spinal neurones." J Physiol 385: 243-286.

References
50
Braestrup, C., R. Albrechtsen and R. F. Squires (1977). "High densities of benzodiazepine receptors in human cortical areas." Nature 269: 702–704.
Brandao, M. L., A. C. Troncoso, M. A. de Souza Silva and J. P. Huston (2003). "The relevance of neuronal substrates of defense in th emidbrain tectum to anxiety and stress: empirical and conceptual considerations." Eur J Pharmacol 463: 225-233.
Bzdok, D., A. R. Laird, K. Zilles, P. T. Fox and Eickhoff (2013). "An investigation of the structural, connectional, and functional subspecialization in the human amygdala." Hum Brain Mapp 34(12): 3247-3266.
Chapman, W. P., H. R. Schroeder, G. Geyer, M. A. Brazier, C. Fager, J. L. Poppen, H. C. Solomon and P. I. Yakovlev (1954). "Physiological evidence concerning importance of the amygdaloid nuclear region in the integration of circulatory function and emotion in man." Science 120: 949-950.
Charney, D. S. and A. Deutch (1996). "A functional neuroanatomy of anxiety and fear: implications for the pathophysiology and treatment of anxiety disorders." Crit Rev Neurobiol 10: 419-446.
Chemla, S. and F. Chavane (2010). "Voltage-sensitive dye imaging: Technique review and models." J Physiol Paris 104: 40-50.
Chien, C. B. and J. Pine (1991). "Voltage-sensitive dye recording of action potentials and synaptic potentials from sympathetic microcultures." Biophys J 60: 697-711.
Corpéchot, C., P. Robel, M. Axelson, J. Sjövall and E. E. Baulieu (1981). "Characterization and measurement of dehydroepiandrosterone sulfate in rat brain." Proc Natl Acad Sci U S A 78: 4704-4707.
Culty, M., H. Li, N. Boujrad, H. Amri, B. Vidic, J. M. Bernassau, J. L. Reversat and V. Papadopoulos (1999). "In vitro studies on the role of the peripheral-type benzodiazepine receptor in steroidogenesis." J Steroid Biochem Mol Biol 69: 123-130.
Dar, D. E., A. Weizman, L. Karp, A. Grinshpoon, M. Bidder, M. Kotler, S. Tyano, A. Bleich and M. Gavish (1991). "Platelet peripheral benzodiazepine receptors in repeated stress." Life Sci 48(4): 341-346.
Davila, H. V., B. M. Salzberg, L. B. Cohen and A. S. Waggoner (1973). "A large change in axon fluorescence that provides a promising method for measuring membrane potential." Nat New Biol 241: 159-160.
Delavoie, F., H. Li, M. Hardwick, J. C. Robert, C. Giatzakis, G. Péranzi, Z. X. Yao, J. Maccario, J. J. Lacapère and V. Papadopoulos (2003). "In vivo and in vitro peripheral-type benzodiazepine receptor polymerization: functional significance in drug ligand and cholesterol binding." Biochemistry 42: 4506–4519.

References
51
Di Chiara, G., E. Acquas, G. Tanda and C. Cadoni (1993). "Drugs of abuse: biochemical surrogates of specific aspects of natural reward?" Biochem Soc Symp 59: 65-81.
Doller, H. J. and F. F. Weight (1982). "Perforant pathway activation of hippocampal CA1 stratum pyramidale neurons: electrophysiological evidence for a direct pathway." Brain Res 237: 1-13.
Eichenbaum, H., T. Otto and N. J. Cohen (1992). "The hippocampus--what does it do?" Behav Neural Biol 57(1): 2-36.
Fluhler, E., V. G. Burnham and L. M. Loew (1985). "Spectra, membrane binding, and potentiometric responses of new charge shift probes." Biochemistry 24: 5749-5755.
Frye, C. A. and J. J. Paris (2011). "Progesterone turnover to its 5α-reduced metabolites in the ventral tegmental area of the midbrain is essential for initiating social and affective behavior and progesterone metabolism in female rats." J Endocrinol Invest 34: 188-199.
Garnier, M., A. B. Dimchev, N. Boujrad, J. M. Price, N. A. Musto and V. Papadopoulos (1994). "In vitro reconstitution of a functional peripheral-type benzodiazepine receptor from mouse Leydig tumor cells." Mol Pharmacol 45: 201-211.
Gavish, M., I. Bachman, R. Shoukrun, Y. Katz, L. Veenman, G. Weisinger and A. Weizman (1999). "Enigma of the peripheral benzodiazepine receptor. ." Pharmacol Rev 51: 629–650.
Gee, K. W. (1988). "Steroid modulation of the GABA/benzodiazepine receptor-linked chloride ionophore." Mol Neurobiol. 2: 291-317.
Golani, I., A. Weizman, S. Leschiner, I. Spanier, N. Eckstein, R. Limor, J. Yanai, K. Maaser, H. Scherübl, G. Weisinger and M. Gavish (2001). "Hormonal Regulation of Peripheral Benzodiazepine Receptor Binding Properties Is Mediated by Subunit Interaction." Biochemistry 40: 10213–10222.
Guillery, R. W. and S. M. Sherman (2002). "Thalamic relay functions and their role in corticocortical communication: generalizations from the visual system." Neuron 33: 163-175.
Hauet, T., Z. X. Yao, H. S. Bose, C. T. Wall, Z. Han, W. Li, D. B. Hales, W. L. Miller, M. Culty and V. Papadopoulos (2005). "Peripheral-Type Benzodiazepine Receptor-Mediated Action of Steroidogenic Acute Regulatory Protein on Cholesterol Entry into Leydig Cell Mitochondria." Mol Endocrinol 19: 540-554.
Höffler, D., R. Lasek, H. K. Berthold and J. D. Tiaden (2003). Therapieempfehlungen der Arzneimittelkommission der deutschen Ärzteschaft - Angst- und Zwangstörungen. Arzneiverordnung in der Praxis. Kölon, Arzneimittelkommission der deutschen Ärzteschaft.
Hosie, A. M., M. E. Wilkins, H. M. da Silva and T. G. Smart (2006). "Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites." 444: 486-489.

References
52
Huguenard, J. R. and D. A. McCormick (2007). "Thalamic synchrony and dynamic regulation of global forebrain oscillations." Trends Neurosci 30: 350-356.
Jefcoate, C. (2002). "High-flux mitochondrial cholesterol trafficking, a specialized function of the adrenal cortex." J Clin Invest 11ß: 81–890.
Jones, E. G. (1975). "Some aspects of the organization of the thalamic reticular complex." J Comp Neurol 162(3): 285-308.
Jones, R. S. (1993). "Entorhinal-hippocampal connections: a speculative view of their function." Trends Neurosci 16(2): 58-64.
Joseph-Liauzun, E., P. Delmas, D. Shire and P. Ferrara (1998). "Topological analysis of the peripheral benzodiazepine receptor in yeast mitochondrial membranes supports a five-transmembrane structure." J Biol Chem 273: 2146-2152.
Kaada, B. R. (1951). "Somato-motor, autonomic and electrocorticographic responses to electrical stimulation of rhinencephalic and other structures in primates, cat, and dog; a study of responses from the limbic, subcallosal, orbito-insular, piriform and temporal cortex, hippocampus-fornix and amygdala." Acta Physiol Scand Suppl 24(83): 1-262.
Karp, L., A. Weizman, S. Tyano and M. Gavish (1989). "Examination stress, platelet peripheral benzodiazepine binding sites, and plasma hormone levels." Life Sci 44(16): 1077-1082.
Keifer, O. P., R. C. Hurt, K. J. Ressler and P. J. Marvar (2015). "The Physiology of Fear: Reconceptualizing the Role of the Central Amygdala in Fear Learning." Physiology (Bethesda) 30(5): 389-401.
Kessler, R. C., M. Angermeyer, J. C. Anthony, R. Graaf, K. Demyttenaere, I. Gasquet, G. Girolamo, S. Gluzman, O. Gureje, J. M. Haro, N. Kawakami, A. Karam, D. Levinson, M. E. Medina Mora, M. A. Oakley Browne, J. Posada-Villa, D. J. Stein, C. H. Adley Tsang, S. Aguilar-Gaxiola, J. Alonso, S. Lee, S. Heeringa, B. E. Pennell, P. Berglund, M. J. Gruber, M. Petukhova, S. Chatterji and T. Ustün (2007). "Lifetime prevalence and age-of-onset distributions of mental disorders in the World Health Organization’s World Mental Health Survey Initiative." World Psychiatry 6: 168-176.
Kita, A., T. Kinoshita, H. Kohayakawa, K. Furukawa and A. Akaike (2009). "Lack of tolerance to anxiolysis and withdrawal symptoms in mice repeatedly treated with AC-5216, a selective TSPO ligand." Prog Neuropsychopharmacol Biol Psychiatry 33: 1040–1045.
Kita, A., H. Kohayakawa, T. Kinoshita, Y. Ochi, K. Nakamichi, S. Kurumiya, K. Furukawa and M. Oka (2004). "Antianxiety and antidepressant-like effects of AC-5216, a novel mitochondrial benzodiazepine receptor ligand." Br J Pharmacol 142: 1059-1072.
Koonce, C. J., A. A. Walf and C. A. Frye (2012). "Type 1 5α-reductase may be required for estrous cycle changes in affective behaviors of female mice." Behav Brain Res 226: 376-380.

References
53
Lacapère, J. J., F. Delavoie, H. Li, G. Péranzi, J. Maccario, V. Papadopoulos and B. Vidic (2001). "Structural and Functional Study of Reconstituted Peripheral Benzodiazepine Receptor." Biochem Biophys Res Commun 284: 536–541.
Lacapère, J. J. and V. Papadopoulos (2003). "Peripheral-type benzodiazepine receptor: structure and function of a cholesterol-binding protein in steroid and bile acid biosynthesis." Steroids 68: 569–585.
Langer, S. Z., S. Arbilla, J. Benavides and B. Scatton (1990). "Zolpidem and alpidem: two imidazopyridines with selectivity for omega 1- and omega 3-receptor subtypes." Adv Biochem Psychopharmacol 46: 61-72.
Li, H., Z. X. Yao, B. Degenhardt, G. Teper and V. Papadopoulos (2001). "Cholesterol binding at the cholesterol recognition/ interaction amino acid consensus (CRAC) of the peripheral-type benzodiazepine receptor and inhibition of steroidogenesis by an HIV
TAT-CRAC peptide." Proc Natl Acad Sci U S A 98: 1267-1272.
Lingford-Hughes, A. R., S. J. Davies, S. McIver, T. M. Williams, M. R. Daglish and D. J. Nutt (2003). "Addiciton." Br Med Bull 65: 209-222.
Loew, L. M. (2015). Design and Use of Organic Voltage Sensitive Dyes., Springer.
Loew, L. M., L. B. Cohen, J. Dix, E. N. Fluhler, V. Montana, G. Salama and J. Y. Wu (1992). "A naphthyl analog of the aminostyryl pyridinium class of potentiometric membrane dyes shows consistent sensitivity in a variety of tissue, cell, and model membrane preparations." J Membr Biol 130: 1-10.
Lomo, T. (1971). "Patterns of activation in a monosynaptic cortical pathway: the perforant path input to the dentate area of the hippocampal formation.y<." Exp Brain Res 12(1): 18-45.
Lubke, J. (1993). "Morphology of neurons in the thalamic reticular nucleus (TRN) of mammals as revealed by intracellular injections into fixed brain slices." J Comp Neurol 329: 458-471.
Macdonald, R. L. and R. W. Olsen (1994). "GABAA receptor channels." Annu Rev Neurosci 17: 569-602.
Majewska, M. D., N. L. Harrison, R. D. Schwartz, J. L. Barker and S. M. Paul (1986). "Steroid hormone metabolites are barbiturate-like modulators of the GABA receptor." Science 232: 1004-1007.
Mattusch, C. (2012). Der Einfluss des Inhalationsanästhetikums Xenon auf die neuronale Aktivität im hippocampalen und thalamokortikalen Schnittpräparat. Humanbiologie. München, Technische Universität München. Doctor of Science.

References
54
Mayo, W., F. Dellu, P. Robel, J. Cherkaoui, M. Le Moal, E. E. Baulieu, H. Simon and D. F. Mayo W, Robel P, Cherkaoui J, Le Moal M, Baulieu EE, Simon H. Infusion of neurosteroids into the nucleus basalis magnocellularis affects cognitive processes in the rat. Brain Research. 1993; 607:324–328. [PubMed: 8386975] (1993). "Infusion of neurosteroids into the nucleus basalis magnocellularis affects cognitive processes in the rat. ." Brain Res 607: 324-328.
McCormick, D. A. and T. Bal (1997). "Sleep and arousal: thalamocortical mechanisms." Annu Rev Neurosci 20: 185-215.
McEnery, M. W., A. M. Snowman, R. R. Trifiletti and S. H. Snyder (1992). "Isolation of the mitochondrial benzodiazepine receptor: Association with the voltage-dependent anion channel and the adenine nucleotide carrier." Proc Natl Acad Sci U S A 89: 3170-3174.
McEwen, B. S. (1991). "Non-genomic and genomic effects of steroids on neural activity." Trends Pharmacol Sci 12: 141-147.
Mennerick, S., M. Chisari, H. J. Shu, A. Taylor, M. Vasek, L. N. Eisenman and C. F. Zorumski (2010). "Diverse voltage-sensitive dyes mod- ulate GABAA receptor function." J. Neurosci. 30: 2871–2879.
Möhler, H. (2006). "GABA(A) receptor diversity and pharmacology." Cell Tissue Res 326(2): 505-516.
Molnár, Z. and A. F. Cheung (2006). "Towards the classification of subpopulations of layer V pyramidal projection neurons." Neurosci Res 55(2): 105-115.
Morad, M. and G. Salama (1979). "Optical probes of membrane potential in heart muscle." J Physiol 292: 267-295.
Moss, M., H. Mahut and S. Zola-Morgan (1981). "Concurrent discrimination learning of monkeys after hippocampal, entorhinal, or fornix lesions." J Neurosci 1(3): 227-240.
Mountcastle, V. B. (1997). "The columnar organization of the neocortex." Brain 120: 701-722.
Mühlpfordt, H. and K. Blachnicki (2008). "Schematic of a fluorescence microscope.". Retrieved 25.04.2015, from http://commons.wikimedia.org/wiki/File:FluorescenceFilters_2008-09-28.svg.
Nothdurfter, C., G. Rammes, T. C. Baghai, C. Schüle, M. Schumacher, V. Papadopoulos and R. Rupprecht (2012a). "Translocator protein (18 kDa) as a target for novel anxiolytics with a favourable side-effect profile." J Neuroendocrinol 24: 82-92.
Nothdurfter, C., R. Rupprecht and G. Rammes (2012b). "Recent developments in potential anxiolytic agents targeting GABAA/BzR complex or the translocator protein (18kDa) (TSPO)." Curr Top Med Chem 12(4).

References
55
Oke, B. O., C. A. Suarez-Quian, J. Riond, P. Ferrara and V. Papadopoulos (1992). "ll surface localization of the peripheral-type benzodiazepine receptor (PBR) in adrenal cortex." Mol Cell Endocrinol 87: R1-R6.
Olsen, J. M. M., B. J. Ciliax, W. R. Mancini and A. B. Young (1988). "Presence of peripheral-type benzodiazepine binding sites on human erythrocyte membranes." Eur J Pharmacol 152: 47-53.
Olsen, R. W. and W. Sieghart (2008). "International Union of Pharmacology. LXX. Subtypes of gamma-aminobutyric acid(A) receptors: classification on the basis of subunit composition, pharmacology, and function. Update." Pharmacol Rev 60(3): 243-260.
Papadopoulos, V., H. Amri, N. Boujrad, C. Cascio, M. Culty, M. Garnier, M. Hardwick, H. Li, B. Vidic, A. S. Brown, J. L. Reversa, J. M. Bernassau and K. Drieu (1997a). "Peripheral benzodiazepine receptor in cholesterol transport and steroidogenesis." Steroids 62: 21-28.
Papadopoulos, V., H. Amri, H. Li, N. Boujrad, B. Vidic and M. Garnier (1997b). "Targeted disruption of the peripheral-type benzodiazepine receptor gene inhibits steroidogenesis in the R2C Leydig tumor cell line." J Biol Chem 272: 32129-32135.
Papadopoulos, V., M. Baraldi, T. R. Guilarte, T. B. Knudsen, J. J. Lacapère, P. Lindemann, N. M. D., D. Nutt, A. Weizman, M. R. Zhang and M. Gavish (2006a). "Translocator protein (18kDa): new nomenclature for the peripheral-type benzodiazepine receptor based on its structure and molecular function." Trends Pharmacol Sci 27: 402-409.
Papadopoulos, V., L. Lecanu, R. C. Brown, Z. Han and Z.-X. Yao (2006b). "Peripheral-type benzodiazepine receptor in neurosteroid biosynthesis, neuropathology and neurological disorders." Neuroscience 138: 749–756.
Paul, S. M. and R. H. Purdy (1992). "Neuroactive steroids." FASEB J 6: 2311-2322.
Pellow, S., P. Chopin, S. E. File and M. Briley (1985). "Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat." J Neurosci Methods 14: 149-167.
Pettit, D. L. and G. J. Augustine (2000). "Distribution of functional glutamate and GABA receptors on hippocampal pyramidal cells and interneurons." J Neurophysiol 84(1): 28-38.
Picazo, O. and A. Fernández-Guasti (1995). "Anti-anxiety effects of progesterone and some of its reduced metabolites: an evaluation using the burying behavior test." Brain Res 680: 135-141.
Pirker, S., C. Schwarzer, A. Wieslthaler, W. Sieghart and G. Sperk (2000). "GABA(A) receptors: immunocytochemical distribution of 13 subunits in the adult rat brain." Neuroscience 101(4): 815-850.
lt Coco: GABA-Rezeptor

References
56
Purdy, R. H., A. L. Morrow, P. H. J. Moore and S. M. Paul (1991). "Stress-induced elevation of γ-aminobutyric acid type A receptor-active steroids in the rat brain. ." Proc Natl Acad Sci U S A 88: 4553-4557.
Reddy, D. S. (2010). Neurosteroids: endogenous role in the human brain and therapeutic potentials. Prog Brain Res. 186: 113-137.
Reddy, D. S. and S. K. Kulkarni (1997). "Differential anxiolytic effects of neurosteroids in the mirrored chamber behavior test in mice." Brain Res 752: 61-71.
Rote-Liste (2013, 2015). Fachinformation Diazepam AbZ (009726-4178), Fachinformation Midazolam rotexmedica (020770-14688), Fachinformation Diazepam-ratiopharm® (011210-5074). R. L. S. GmbH, Rote Liste Service GmbH.
Rupprecht, R. (1997). "The neuropsychopharmacological potential of neuroactive steroids." J Psychiatr Res 31: 297-314.
Rupprecht, R. and F. Holsboer (1999). "Neuroactive steroids: mechanisms of action and neuropsychopharmacological perspectives." Trends Neurosci 22: 410-416.
Rupprecht, R., V. Papadopoulos, G. Rammes, T. C. Baghai, J. Fan, N. Akula, G. Groyer, D. Adams and M. Schumacher (2010). "Translocator protein (18kDa) (TSPO) as a therapeutic target for neurological and psychiatric disorders." Nat Rev Drug Discov 9: 971-988.
Rupprecht, R., G. Rammes, D. Eser, T. C. Baghai, C. Schule, C. Nothdurfter, T. Troxler, C. Gentsch, H. O. Kalkman, F. Chaperon, V. Uzunov, K. H. McAllister, V. Bertaina-Anglade, C. D. La Rochelle, D. Tuerck, A. Floesser, B. Kiese, M. Schumacher, R. Landgraf, F. Holsboer and K. Kucher (2009). "Translocator Protein (18 kD) as Target for Anxiolytics Without Benzodiazepine-Like Side Effects." Science 325: 490-493.
Sah, P., E. S. Faber, M. Lopez De Armentia and J. Power (2003). "The amygdaloid complex: anatomy and physiology." Physiol Rev 83: 803-834.
Salzberg, B. M., H. V. Davila and L. B. Cohen (1973). "Optical recording of impulses in individual neurones of an invertebrate central nervous system." Nature 246: 508-509.
Savic, M. M., D. I. Obradovic, N. D. Ugresic and D. R. Bokonjic (2005). "Memory effects of benzodiazepines: memory stages and types versus binding-site subtypes." Neural Plast 12: 289-298.
Schüle, C., C. Nothdurfter and R. Rupprecht (2014). "The role of allopregnanolone in depression and anxiety." Prog Neurobiol 113: 79–87.

References
57
Scoville, W. B. and B. Milner (1957). "Loss of recent memory after bilateral hippocampal lesions." J Neurol Neurosurg Psychiatry 20: 11-21.
Serra, M., P. Madau, M. F. Chessa, M. Caddeo, E. Sanna, G. Trapani, M. Franco, G. Liso, R. H. Purdy, M. L. Barbaccia and G. Biggio (1999). "2-Phenyl-imidazo[1,2-a]pyridine derivatives as ligands for peripheral benzodiazepine receptors: stimulation of neurosteroid synthesis and anticonflict action in rats." Br J Pharmacol 127: 177-187.
Shosaku, A., Y. Kayama, I. Sumitomo, M. Sugitani and K. Iwama (1989). "Analysis of recurrent inhibitory circuit in rat thalamus: neurophysiology of the thalamic reticular nucleus." Prog Neurobiol 32(2): 77-102.
Sigel, E. and A. Buhr (1997). "The benzodiazepine binding site of GABAA receptors." Trends Pharmacol Sci 18(11): 425-429.
Squire, L. R. (1992). "Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans." Psychol Rev 99(1): 195-231.
Stepan, J., J. Dine, T. Fenzl, S. Polta, G. von Wolff, C. T. Wotjak and M. Eder (2012). "Entorhinal theta-frequency input to the dentate gyrus trisynaptically evokes hippocampal CA1 LTP." Front. Neural Circuits 6: 1-13.
Steriade, M. (2005). "Sleep, epilepsy and thalamic reticular inhibitory neurons." Trends Neurosci 28: 317-324.
Strohle, A., E. Romeo, F. di Michele, B. Hermann, G. Gajewsky, F. Holsboer and R. Rupprecht (2003). "Induced panic attacks shift gamma-aminobutyric acid type A receptor modulatory neuroactive steroid composition in patients with panic disorder: preliminary results." Arch Gen Psychiatry 60: 161-168.
Synder, S. H., A. Verma and R. R. Trifiletti (1987). "The peripheral-type benzodiazepine receptor: a protein of mitochondrial outer membranes utilizing porphyrins as endogenous ligands." FASEB J 1: 282-288.
Taketani, S., H. Kohno, M. Okuda, T. Furukawa and R. Tokunaga (1994). "Induction of peripheral-type benzodiazepine receptors during differentiation of mouse erythroleukemia cells. A possible involvement of these receptors in heme biosynthesis." J Biol Chem 269: 7527-7531.
Tasaki, I., A. Watanabe, R. Sandlin and L. Carnay (1968). "Changes in fluorescence, turbidity, and birefringence associated with nerve excitation." Proc Natl Acad Sci U S A 61: 883-888.
Thiele, J. (2008). Optische und mechanische Messungen von elektrophysiologischen Vorgangen im Myokardgewebe. Fakultat fur Elektrotechnik und Informationstechnik, Universitat Karlsruhe.
Trepel, M. (2008). Neuroanatomie - Struktur und Funktion. München, Urban & Fischer.

References
58
Verleye, M., Y. Akwa, P. Liere, N. Ladurelle, A. Pianos, B. Eychenne, M. Schumacher and J. M. Gillardin (2005). "The anxiolytic etifoxine activates the peripheral benzodiazepine receptor and increases the neurosteroid levels in rat brain." Pharmacol Biochem Behav 82: 712-720.
Wafford, K. A. (2005). "GABAA receptor subtypes: any clues to the mechanism of benzodiazepine dependence?" Curr Opin Pharmacol 5: 47-52.
Wahl-Schott, C. and M. Biel (2009). "HCN channels: Structure, cellular regulation and physiological function." Cell Mol Life Sci 66: 470-494.
Walters, R. J., S. H. Hadley, K. D. Morris and J. Amin (2000). "Benzodiazepines act on GABAA receptors via two distinct and separable mechanisms." Nat Neurosci 3(12): 1274-1281.
Weiskrantz, L. (1956). "Behavioral changes associated with ablation of the amygdaloid complex in monkeys." J Comp Physiol Psychol 49: 381-391.
Weizman, R., N. Laor, L. Karp, E. Dagan, A. Reiss, D. E. Dar, L. Wolmer and M. Gavish (1994). "Alteration of platelet benzodiazepine receptors by stress of war." Am J Psychiatry 151: 766-767.
Weizman, R., Z. Tanne, M. Granek, L. Karp, M. Golomb, S. Tyano and M. Gavish (1987). "Peripheral benzodiazepine binding sites on platelet membranes are increased during diazepam treatment of anxious patients." Eur J Pharmacol 138: 289-292.
Wieland, S., J. D. Belluzzi, L. Stein and N. C. Lan (1995). "Comparative behavioral characterization of the neuroactive steroids 3 alpha-OH,5 alpha-pregnan-20-one and 3 alpha-OH,5 beta-pregnan-20-one in rodents." Psychopharmacology (Berl) 118: 65-71.
Wieland, S., N. C. Lan, S. Mirasedeghi and K. W. Gee (1991). "Anxiolytic activity of the progesterone metabolite 5 alpha-pregnan-3 alpha-o1-20-one." Brain Res 565: 263-268.
Windisch, H., W. Müller and H. A. Tritthart (1985). "Fluorescence monitoring of rapid changes in membrane potential in heart muscle." Biophys J 48: 877-884.
Wright, G. and V. Reichenbecher (1999). "The Effects of Superoxide and the Peripheral Benzodiazepine Receptor Ligands on the Mitochondrial Processing of Manganese-Dependent Superoxide Dismutase." Exp Cell Res 246: 443-450.
Yeckel, M. F. and T. W. Berger (1990). "Feedforward excitation of the hippocampus by afferents from the entorhinal cortex: redefinition of the role of the trisynaptic pathway." Proc Natl Acad Sci U S A 87: 5832-5836.
Ying, S.-W. and P. A. Goldstein (2005). "Propofol-Block of SK Channels in Reticular Thalamic Neurons Enhances GABAergic Inhibition in Relay Neurons." J Neurophysiol 93: 1935-1948.

References
59
Zimmerberg, B., P. C. Drucker and J. M. Weider (1995). "Differential behavioral effects of the neuroactive steroid allopregnanolone on neonatal rats prenatally exposed to alcohol." Pharmacol Biochem Behav 51: 463-468.
Zola-Morgan, S., L. R. Squire, P. Alvarez-Royo and R. P. Clower (1991). "Independence of memory functions and emotional behavior: separate contributions of the hippocampal formation and the amygdala." Hippocampus 1: 207-220.

Appendix
60
7 Appendix
Used Substances
Sucrose-based-ACSF: pH ≈ 7,37 (saturated with carbogen) by Ying und Goldberg (Ying et al.
2005)
Substance Concentration (mM) Manufacturer
CaCl2 0,5 Sigma Aldrich
Glucose 11 Sigma Aldrich
Sucrose 234 Sigma Aldrich
KCl 2,5 Sigma Aldrich
MgSO4 10 Sigma Aldrich
NaHCO3 24 Sigma Aldrich
NaH2PO4 1,25 Sigma Aldrich
General ACSF: pH ≈ 7,37 (saturated with carbogen)
Substance Concentration (mM) Manufacturer
CaCl2 2 Sigma Aldrich
D(+)-Glucose 25 Sigma Aldrich
KCl 2,5 Sigma Aldrich
MgCl2 1 Sigma Aldrich
NaCl 125 Sigma Aldrich
NaHCO3 25 Sigma Aldrich
NaH2PO4 1,25 Sigma Aldrich

Appendix
61
Further Chemicals:
Substance Concentrations used Manufacturer
Allopregnanolone 1 µM (DMSO) Sigma Aldrich
Bicucullin 0,6 µM (DD) Sigma Aldrich
Di-4-Annepps see ch. 2.2.1 Sigma Aldrich
Diazepam 1 µM (DMSO) Sigma Aldrich
DMSO - Sigma Aldrich
Finasterid 10 µM (DMSO) Sigma Aldrich
Flumazenil 10 µM (DMSO) Sigma Aldrich
XBD 173 3 µM (70% Alcohol) Novartis

Appendix
62
Acknowledgements
Sincere thanks are given to everybody making this dissertation possible, especially:
My doctoral advisor, Prof. Dr. Gerhard Rammes for awarding me the topic and presenting me with
the opportunity to finalize my dissertation at the clinic of anesthesiology of the Klinikum Rechts der
Isar, to work in his charming research group, the excellent supervision as well as for reviewing my
thesis.
Dr. C. Mattusch for the splendid introduction to scientific working, outstanding mentoring and
helpfulness in all concerns.
As well as Martina Bürge, for her patients and utterly helpful guidance.
Dr. Matthias Eder, for letting me use the premises of his working group in the Max-Planck-Institute
for psychiatry as well as academic council.
The entire working group of AG Eder at the Max-Planck-Institute for the charming collaboration. To
be individually mentioned are Konrad Straub, Stefan Kratzer, Julien Dien, Jens Stephan and
Florian Hladky.
Last but certainly not least I want to thank Matthias Kreuzer for designing the Matlab-macros and
even more for his patience and encouraging words.