Embed Size (px)
Citation preview

Folia Medica Lodziensia, 2012, 39/2: 293-326
Adres do korespondencji: dr hab. Magdalena Bryś, prof. nadzw. UŁ; Katedra Cytobiochemii, Uniwersytet Łódzki; 90-236 Łódź; ul. Pomorska 141/143; e-mail: [email protected] Tel.: + 48 42 6354371; Fax: +48 42 6354484
Telomeraza – struktura i funkcja
oraz regulacja ekspresji genu
Telomerase – structure, function and the regulation of gene expression
MAGDALENA BRYŚ, MAGDALENA LASKOWSKA, EWA FORMA, ANNA KRZEŚLAK
Katedra Cytobiochemii, Wydział Biologii i Ochrony Środowiska,
Uniwersytet Łódzki
Streszczenie Telomer jest to fragment chromosomu zlokalizowany na jego końcu, który
zabezpiecza go przed uszkodzeniem podczas kopiowania. Telomery są także
czynnikami kontrolującymi liczbę podziałów komórkowych i dlatego uważane są
za supresory transformacji nowotworowej, ponieważ ograniczona, ściśle
kontrolowana liczba podziałów zapobiega ewentualnemu kumulowaniu się
mutacji w komórce. Przyjęto, że obecność 4-6 mutacji w materiale genetycznym
jest czynnikiem karcynogennym, a po granicznej liczbie podziałów (około 60-70)
komórka wchodzi w fazę spoczynku M1. Enzymem, którego zadaniem jest
dobudowanie 3'-końcowego odcinka nici DNA i tym samym wydłużanie
sekwencji telomerowych jest enzym telomeraza. Białko to jest polimerazą DNA
zależną od RNA, która syntetyzuje telomery na zasadzie odwrotnej transkrypcji.
Unikalną cechą telomerazy jest to, że jej integralnym składnikiem jest matryca
RNA służąca do syntezy DNA. Telomeraza występuje w intensywnie dzielących
się komórkach, a jej aktywność zmniejsza się wraz z wiekiem. W komórkach
prawidłowych zwykle nie stwierdza się aktywności telomerazy, natomiast
w nowotworowych aktywność tego enzymu zwykle jest podwyższona. W pracy
omówiono strukturę sekwencji telomerowych oraz udział białek zaangażowa-
nych w jej utrzymanie. Szczegółowo przedstawiono także strukturę i funkcję
genu/białka telomerazy, z uwzględnieniem regulacji ekspresji genu na poziomie
transkrypcji. Scharakteryzowano ponadto udział telomerazy w procesach
transformacji nowotworowej.

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 294
Słowa kluczowe: telomeraza, gen, białko, ekspresja genu, czynniki
transkrypcyjne.
Abstract
A telomere is a fragment localized at the end of chromosome which protects the chromosome from damage during replication. Telomeres are also factors that control number of cell divisions and are thought to be a suppressors of carcinogenesis since limited, strictly determined number of cell divisions protects from accumulation of mutations in cell. It is assumed that presence of 4-6 mutation in genetic material is a carcinogenic factor and after about 60-70 divisions, the cell enter the resting phase. Telomerase is an enzyme which adds DNA sequence to the 3’ end of DNA and extends the telomere region. This protein is a DNA polymerase dependent on RNA, which syntheses telomere by reverse transcription. The unique characteristics of telomerase is that RNA matrix for DNA synthesis is an integral component of this enzyme. Telomerase is present in intensively dividing cells and its activity is decreasing with age. In normal cells usually activity of telomerase is undetectable but in cancer cells activity of this enzyme is high. The aim of this work is to present the structure of telomeres and the role of proteins involved in maintaining the structure. In details, the structure and function of the telomerase gene/protein is described, including the regulation of gene expression at the transcriptional level. The involvement of telomerase in the neoplastic transformation has been also characterized.
Key words: telomerase, gene, protein, gene expression, transcription factors. Wstęp
Opisy struktury i funkcji telomeru pojawiły się w literaturze naukowej
w latach 30. XX wieku. Po raz pierwszy w 1978 roku sekwencja telomeru
została określona u jednokomórkowego orzęska Tetrahymena pyriformis [1].

M. Bryś i wsp. 295
W latach 90. XX wieku Jerry Shay i współpracownicy wykryli telomerazę
w 90 na 101 preparatach komórek pobranych z różnych nowotworów człowieka
i nie wykazali obecności telomerazy w żadnej z 50 różnych, prawidłowych
komórek somatycznych. Wyniki tych badań sugerowały, że telomeraza pełni
istotną funkcję w procesie transformacji nowotworowej [2]. W 1997 r. Robert A.
Weinberg wraz z zespołem badawczym sklonowali gen kodujący podjednostkę
telomerazy o aktywności odwrotnej transkryptazy [3]. W 2009 roku Nagrodę
Nobla w dziedzinie fizjologii lub medycyny za odkrycie, w jaki sposób telomery
i enzym telomeraza chronią chromosomy, otrzymała trójka amerykańskich
uczonych - Elizabeth H Blackburn, Jack W Szostak i Carol W Greiner [4].
Zgodnie z prawami natury, celem nadrzędnym wszystkich organizmów jest
utrzymanie ciągłości gatunku i dokonuje się to poprzez przekazywanie materiału
genetycznego organizmom potomnym. Aby organizm mógł prawidłowo
funkcjonować, w czasie jego rozwoju osobniczego muszą zachodzić w ściśle
określonym czasie procesy syntezy i naprawy uszkodzeń materiału
genetycznego oraz programowanej śmierci komórki, czyli apoptozy. Zgodnie
z teorią Kirkwooda oraz w związku z faktem, iż średnia długość życia człowieka
wydłuża się, korzystniejsze jest wydatkowanie większej części energii na
procesy naprawcze organizmu niż reprodukcyjne. Wraz z długością życia
zwiększeniu ulega ryzyko związane z powstawaniem nowotworów, co
bezpośrednio wynika z wydłużenia okresu działania czynników karcynogennych
na komórki oraz z gromadzenia się nieprawidłowości na poziomie
molekularnym [5].
Odmienna specyfika aktywności telomerazy w materiale prawidłowym
i nowotworowym stała się podstawą do rozważania tego enzymu w kontekście
terapii przeciwnowotworowej. Komórki nowotworowe charakteryzują się

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 296
znacznie krótszymi telomerami niż komórki prawidłowe i zależne są od
aktywności telomerazy. Zablokowanie aktywności tego enzymu może wpływać
na zahamowanie transformacji nowotworowej bez równoczesnego ingerowania
w funkcjonowanie komórek prawidłowych [6, 7]
Odmienna specyfika aktywności telomerazy w materiale prawidłowym
i nowotworowym stała się podstawą do rozważania tego enzymu w kontekście
terapii przeciwnowotworowej. Komórki nowotworowe charakteryzują się
znacznie krótszymi telomerami niż komórki prawidłowe i zależne są od
aktywności telomerazy. Zablokowanie aktywności tego enzymu może wpływać
na zahamowanie transformacji nowotworowej bez równoczesnego ingerowania
w funkcjonowanie komórek prawidłowych [6, 7].
Struktura i funkcje telomerów
Telomery (gr. telos – koniec i meros – część) są to wyspecjalizowane
kompleksy nukleoproteinowe, które chronią zakończenia prawidłowych
chromosomów przed degradacją i fuzją, co w konsekwencji przekłada się na
utrzymanie stabilności genomu [1, 8, 9]. W komórkach człowieka występują
92 telomery, po jednym na każdym końcu chromosomu. Wysoce
konserwatywne sekwencje telomerowe są kilkutysięcznymi, nie kodującymi,
powtarzającymi się motywami nukleotydowymi, bogatymi w reszty guaniny
i tyminy [10]. U człowieka i większości kręgowców powtarzającym się
motywem jest sekwencja TTAGGG [11, 12]. Sekwencje telomerowe wybranych
organizmów zawarte zostały w Tabeli 1.

M. Bryś i wsp. 297
Tabela1. Sekwencje telomerowego DNA u wybranych zwierząt i roślin [wg 13, 14]
ORGANIZM Sekwencja (5’→3’)
człowiek (łac.Homo sapiens) TTAGGG
drożdże (łac. Saccharomyces cerevisiae) TGTGGGTGTGGTG
glista ludzka (łac. Ascaris lumbricoides) TTAGGC
jedwabnik (łac. Bombyx mori) TTAGG
mysz polna (łac. Apodemus agrarius) TTAGGG
pantofelek (łac. Paramecium caudatum) TTGGG(T/G)
rzodkiewnik pospolity (łac. Arabidopsis thaliana) TTTAGGG
tasiemiec uzbrojony (łac. Taenia solium) TTAGGC
zawłotnia (łac. Chlamydomonas) TTTTAGGG
U człowieka powtórzenia (TTAGGG)n mają długość około 15-20 kpz
w chwili narodzin, ale już u osób dorosłych ulegają skróceniu do 8-10 kpz.
Długość sekwencji telomerowych wykazuje zmienność osobniczą, jak również
może wahać się w zależności od komórki czy tkanki [15, 16].
Obowiązujący aktualnie model struktury telomeru uzyskano dzięki
badaniom z użyciem mikroskopu elektronowego. Jest on zdecydowanie bardziej
złożony w swojej przestrzennej strukturze, niż wynikało to z modelu
klasycznego, który zakładał strukturę liniową. W obecnym modelu jednoniciowe
końce 3’ chromosomów, o długości około 75–200 pz, tworzą strukturę
przestrzenną zwaną kwartetem G (G4-DNA), który uniemożliwia dostęp
egzonukleaz do końcowych odcinków telomerów i tym samym zabezpiecza nić
DNA przed degradacją. Różne struktury przestrzenne kwartetu G zostały
przedstawione na ryc. 1 i 2.
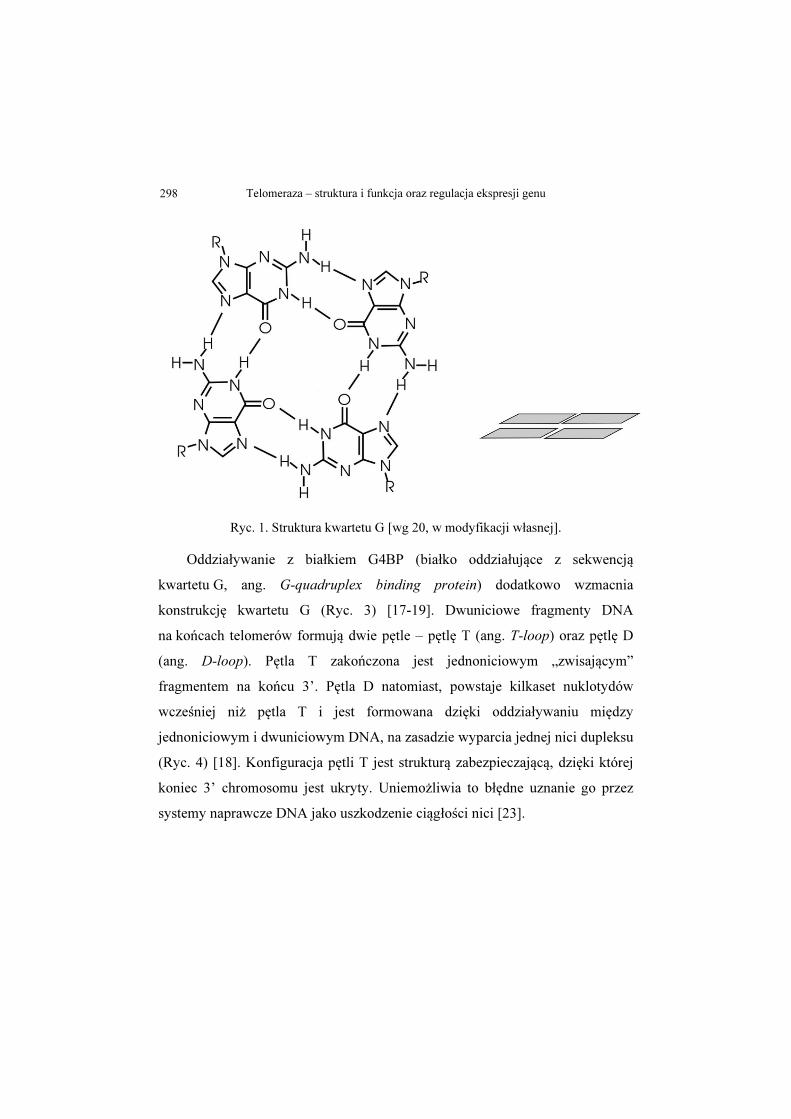
Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 298
Ryc. 1. Struktura kwartetu G [wg 20, w modyfikacji własnej].
Oddziaływanie z białkiem G4BP (białko oddziałujące z sekwencją
kwartetu G, ang. G-quadruplex binding protein) dodatkowo wzmacnia
konstrukcję kwartetu G (Ryc. 3) [17-19]. Dwuniciowe fragmenty DNA
na końcach telomerów formują dwie pętle – pętlę T (ang. T-loop) oraz pętlę D
(ang. D-loop). Pętla T zakończona jest jednoniciowym „zwisającym”
fragmentem na końcu 3’. Pętla D natomiast, powstaje kilkaset nuklotydów
wcześniej niż pętla T i jest formowana dzięki oddziaływaniu między
jednoniciowym i dwuniciowym DNA, na zasadzie wyparcia jednej nici dupleksu
(Ryc. 4) [18]. Konfiguracja pętli T jest strukturą zabezpieczającą, dzięki której
koniec 3’ chromosomu jest ukryty. Uniemożliwia to błędne uznanie go przez
systemy naprawcze DNA jako uszkodzenie ciągłości nici [23].

M. Bryś i wsp. 299
Ryc. 2. Przestrzenne struktury kwartetu G. DNA bogate w reszty guaniny może tworzyć
różne struktury kwartetu G. Pojedyncza nić DNA może składać się ku sobie, tworząc wewnętrzny kwartet G, podczas gdy dwie i cztery nici DNA mogą się łączyć
odpowiednio, w struktury dimeryczne lub tetrameryczne. Strzałki wskazują orientację DNA – równoległą i przeciwrównoległą. Reszty guanozyny przedstawione zostały w
różnych kolorach, według konfiguracji wiązania glikozydowego. Wiązanie glikozydowe typu „anty” – kolor szary, typu „trans” – kolor czarny [wg 21; w modyfikacji własnej].
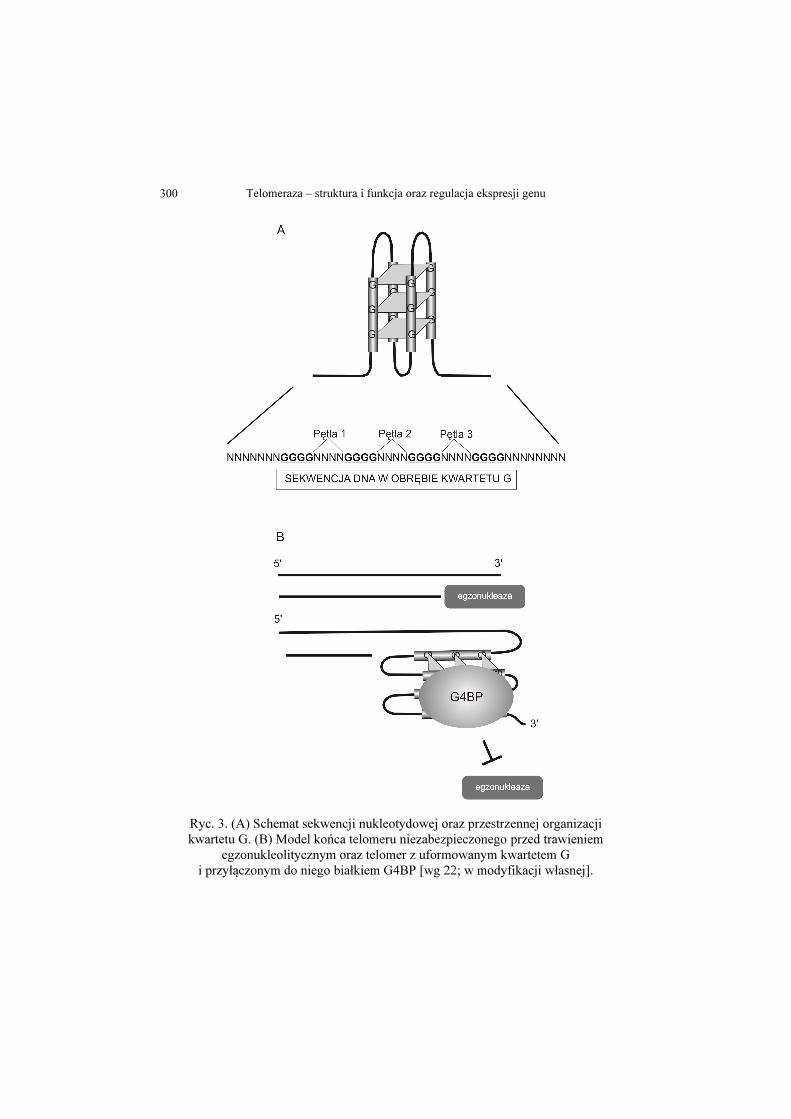
Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 300
Ryc. 3. (A) Schemat sekwencji nukleotydowej oraz przestrzennej organizacji kwartetu G. (B) Model końca telomeru niezabezpieczonego przed trawieniem
egzonukleolitycznym oraz telomer z uformowanym kwartetem G i przyłączonym do niego białkiem G4BP [wg 22; w modyfikacji własnej].

M. Bryś i wsp. 301
Ryc. 4. Schemat obrazujący powstawanie pętli na zakończeniach chromosomów [wg 22; w modyfikacji własnej].
Telomery chronione są przez wyspecjalizowane kompleksy białkowe, które
rozpoznają podwójną nić DNA oraz kwartet G. Taki białkowy kompleks
w komórkach ssaków nazywany jest „shelterin” i składa się z sześciu białek:
TRF1 i TRF2 (czynnik wiążący się do podwójnych powtórzeń TTAGGG 1, 2,
ang. telomeric repeat binding factor 1, 2), RAP1 (białko represorowo-
aktywatorowe, ang. repressor - activator protein 1), TIN2 (czynnik jądrowy
oddziałujący z TRF1 2, ang. TRF1 interacting nuclear factor 2), TPP1
(trójpeptydylopeptydaza 1, ang. tripeptidyl peptidase 1) i POT1 (białko
chroniące telomery 1, ang. protection of telomeres 1) [24]. Białka TRF1 TRF2
łączą się bezpośrednio z telomerowym dwuniciowym DNA, natomiast białka
RAP1, TIN2, TPP1 i POT1 stabilizują strukturę kompleksu „shelterin” [25-27].

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 302
Układ przestrzenny i charakterystyka wyżej wymienionych białek została
przedstawiona na rycinie 5 oraz w Tabeli 2.
Ryc. 5. Układ przestrzenny kompleksu „shelterin” oraz konfiguracja białek podtrzymująca strukturę pętli T i pętli D telomeru [wg 28, 29; w modyfikacji własnej].
Białko POT1 łącząc się z pojedynczą nicią telomerowego DNA oddziałuje
z białkiem TPP1. Guanina na końcu 3' telomerowego DNA jest „zanurzona”
w białku POT1, co powoduje niedostępność substratu dla telomerazy
i najprawdopodobniej hamuje wydłużanie nici DNA. Z kolei białko Rap1 łączy
się z białkiem TIN2 oraz białkami TRF1 i TRF2. Białko TIN2 oddziałuje
również zarówno z białkiem TRF1, jak i TRF2, a jego zadaniem jest rekrutacja
białek POT1 oraz TPP1 do dwuniciowego fragmentu telomeru [30, 31]
M. Bryś i wsp. 303
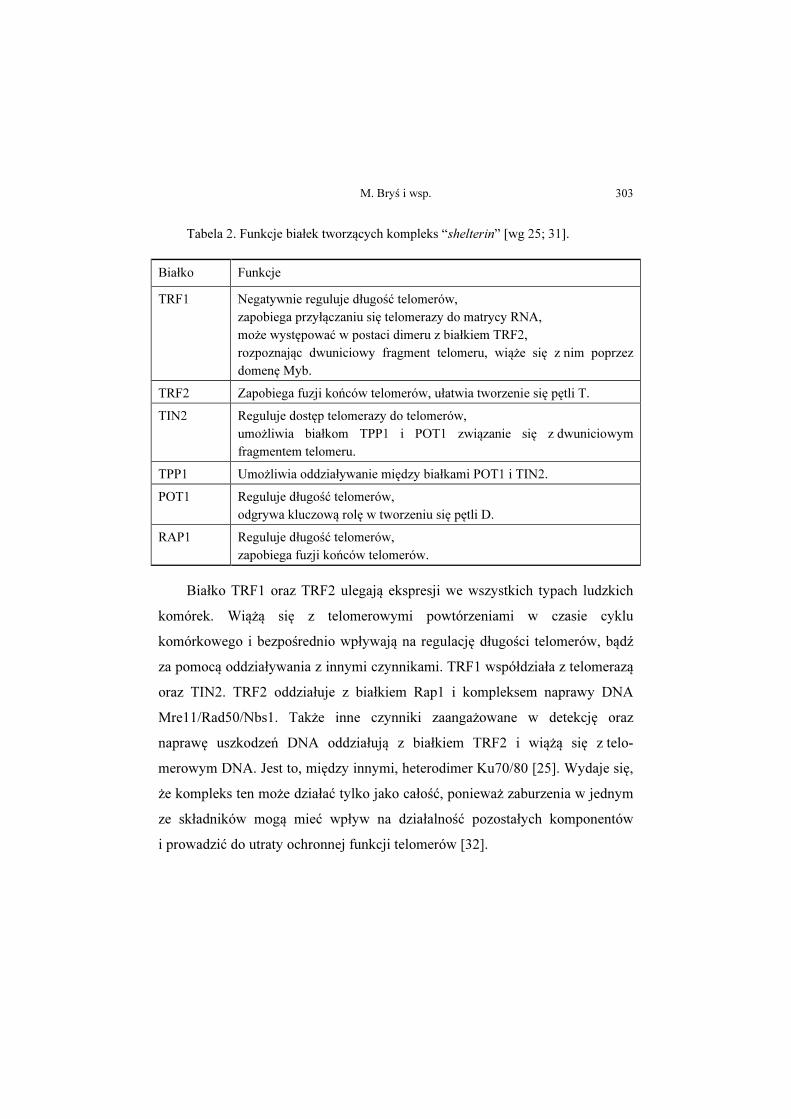
Tabela 2. Funkcje białek tworzących kompleks “shelterin” [wg 25; 31].
Białko Funkcje
TRF1 Negatywnie reguluje długość telomerów, zapobiega przyłączaniu się telomerazy do matrycy RNA, może występować w postaci dimeru z białkiem TRF2, rozpoznając dwuniciowy fragment telomeru, wiąże się z nim poprzez domenę Myb.
TRF2 Zapobiega fuzji końców telomerów, ułatwia tworzenie się pętli T.
TIN2 Reguluje dostęp telomerazy do telomerów, umożliwia białkom TPP1 i POT1 związanie się z dwuniciowym fragmentem telomeru.
TPP1 Umożliwia oddziaływanie między białkami POT1 i TIN2.
POT1 Reguluje długość telomerów, odgrywa kluczową rolę w tworzeniu się pętli D.
RAP1 Reguluje długość telomerów, zapobiega fuzji końców telomerów.
Białko TRF1 oraz TRF2 ulegają ekspresji we wszystkich typach ludzkich
komórek. Wiążą się z telomerowymi powtórzeniami w czasie cyklu
komórkowego i bezpośrednio wpływają na regulację długości telomerów, bądź
za pomocą oddziaływania z innymi czynnikami. TRF1 współdziała z telomerazą
oraz TIN2. TRF2 oddziałuje z białkiem Rap1 i kompleksem naprawy DNA
Mre11/Rad50/Nbs1. Także inne czynniki zaangażowane w detekcję oraz
naprawę uszkodzeń DNA oddziałują z białkiem TRF2 i wiążą się z telo-
merowym DNA. Jest to, między innymi, heterodimer Ku70/80 [25]. Wydaje się,
że kompleks ten może działać tylko jako całość, ponieważ zaburzenia w jednym
ze składników mogą mieć wpływ na działalność pozostałych komponentów
i prowadzić do utraty ochronnej funkcji telomerów [32].

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 304
Telomery jako struktury dynamiczne, ochraniające końce chromosomów
przed degradacją, odpowiadają między innymi za umożliwienie systemom
naprawczym rozpoznania prawidłowych i uszkodzonych zakończeń
chromosomów, przestrzenną organizację jądra komórkowego oraz regulację
transkrypcji genów zlokalizowanych w pobliżu telomerów [33, 34]. Ponadto,
telomery zapobiegają translokacjom, amplifikacjom i delecjom materiału
genetycznego. Zabezpieczając przed fuzją chromosomów, zapewniają
prawidłowy przebieg procesu rekombinacji i zapobiegają powstawaniu
chromosomów dicentrycznych. W przypadku uszkodzenia telomeru często
dochodzi do procesu rekombinacji. W zależności od typu tego zjawiska,
dochodzi do fuzji ramion chromosomów typu p-p, p-q i q-q [35, 36].
Po osiągnięciu krytycznego stanu skrócenia telomerów (3 kpz), przy którym
zakończenia chromosomów nie są w stanie utrzymać charakterystycznej dla
telomerów struktury pętli, telomery nie mogą chronić chromosomów przed
utratą materiału genetycznego i niestabilnością genomu, co prowadzi do śmierci
komórki. Jest to jeden z mechanizmów inicjujących apoptozę i naturalna
konsekwencja starzenia się komórek [37].
Aż do wczesnych lat 60 uważano, że hodowane prawidłowe komórki
człowieka, mogą dzielić się nieskończenie wiele razy, tak długo jak dobre
warunki hodowli zostaną utrzymane. Przełom nastąpił w 1965 roku za sprawą
Leonarda Hayflick’a, który obalił dotychczasową koncepcję i wykazał, iż
prawidłowe komórki człowieka mają ograniczoną liczbę podziałów. To zjawisko
starzenia replikacyjnego prawidłowych komórek ludzkich bardzo często
nazywane jest „limitem Hayflicka” (około 60 podziałów). Każdy podział
komórki związany jest z nieodwracalnym skracaniem sekwencji telomerowej
o ok. 50-150 pz., co jest związane z problem replikacji końców nici opóźnionej.

M. Bryś i wsp. 305
Polimeraza DNA replikuje tylko w kierunku 5’→3’, a do uruchomienia
replikacji wymaga starterowych fragmentów RNA, co powoduje niepełną
replikację nici DNA przy końcu 5’ [38, 39]. Telomer jest tzw. molekularnym
zegarem biologicznym wyznaczającym ściśle określoną liczbę podziałów
komórkowych. Komórki mają więc ograniczony potencjał proliferacyjny,
a co za tym idzie po pewnej liczbie podziałów wkraczają w etap starzenia się.
Tracą wtedy zdolność podziałów, ale nadal są aktywne metabolicznie [40].
Telomery dzięki określonej liczbie podziałów komórkowych stają się
supresorami procesu nowotworzenia, gdyż w ten sposób zapobiegają nagro-
madzeniu się mutacji prowadzących do rozwoju nowotworu [35, 36, 41, 42].
Telomeraza – struktura i funkcje
U większości ssaków za utrzymanie odpowiedniej długości telomerów
odpowiada specyficzna odwrotna tranksyptaza – telomeraza. Jest to enzym
odgrywający kluczową rolę także w takich procesach jak proliferacja komórki
i transformacja nowotworowa [43]. Unikalną cechą telomerazy jest to, że jej
integralnym składnikiem jest matryca RNA służąca do syntezy DNA.
Telomeraza jest polimerazą DNA zależną od RNA, która syntetyzuje telomery
na zasadzie odwrotnej transkrypcji. Jest to duży kompleks rybonukleo-
proteinowy - masa cząsteczkowa holoenzymu telomerazy przekracza 500 kDa
[44]. Telomeraza człowieka składa się z fragmentu nici RNA (hTERC/hTR) oraz
podjednostki białkowej o aktywności odwrotnej transkryptazy (hTERT) [45].
Struktura genu TERC u różnych organizmów różni się pod względem rozmiaru,
jak i sekwencji nukleotydowej. Dojrzała struktura genu TERC u Saccharomyces
cerevisiae składa się z 1167 nukleotydów, podczas, gdy u pierwotniaków i

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 306
ssaków jest ona znacząco krótsza i wynosi odpowiednio około 150 i 450
nukleotydów [46-49].
Struktura TERC zawiera:
• domenę centralną
• miejsce wiązania białek odpowiedzialnych za dojrzewanie i stabilizację
domeny
• miejsce wiązania z TERT
Ryc. 6. Schemat ludzkiego chromosomu 3 z zaznaczoną lokalizacją genu hTERC (q26).
Gen hTERC zlokalizowany jest na chromosomie 3q26 (Rys 6). Jest on
transkrybowany przez polimerazę RNA II. Składa się z 445 nukleotydów,
z powtarzającą się sekwencją 11 nukleotydową: 5’-(CUAACCCUAAC)-3’,
która jest komplementarna do sekwencji telomeru (TTAGGG) [50]. Sekwencja
nukleotydowa oraz konformacja przestrzenna podjednostki TR przedstawiona
została na rycinie 7.
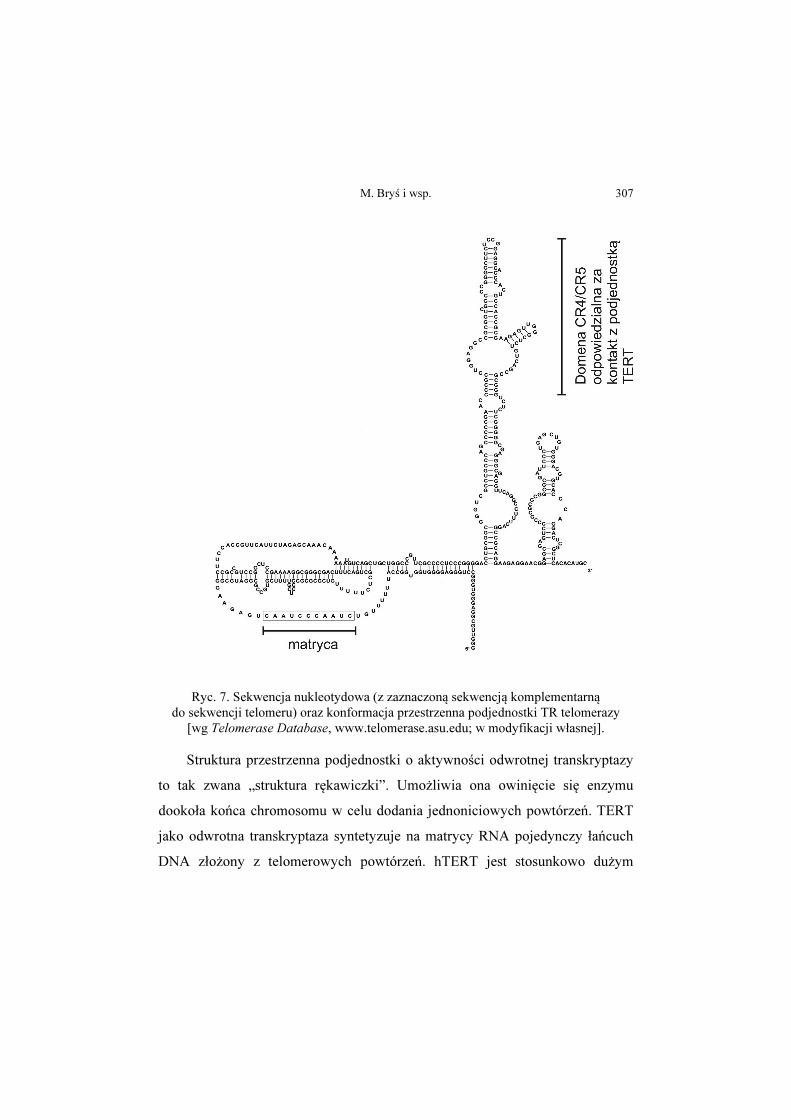
M. Bryś i wsp. 307
Ryc. 7. Sekwencja nukleotydowa (z zaznaczoną sekwencją komplementarną do sekwencji telomeru) oraz konformacja przestrzenna podjednostki TR telomerazy
[wg Telomerase Database, www.telomerase.asu.edu; w modyfikacji własnej].
Struktura przestrzenna podjednostki o aktywności odwrotnej transkryptazy
to tak zwana „struktura rękawiczki”. Umożliwia ona owinięcie się enzymu
dookoła końca chromosomu w celu dodania jednoniciowych powtórzeń. TERT
jako odwrotna transkryptaza syntetyzuje na matrycy RNA pojedynczy łańcuch
DNA złożony z telomerowych powtórzeń. hTERT jest stosunkowo dużym

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 308
białkiem, jego masa cząsteczkowa wynosi 127 kDa. Gen dla tej podjednostki
zlokalizowany jest na krótszym ramieniu chromosomu 5 (p15.33) (Ryc.8).
Obejmuje on 37 pz genomowego DNA i składa się z 16 eksonów [43].
Ryc. 8. Schemat ludzkiego chromosomu 5 z zaznaczoną lokalizacją genu hTERT (p15.33).
Chociaż struktura krystaliczna dla białka TERT nie została jeszcze w pełni
poznana, sugeruje się istnienie trzech domen:
• N-końowej (NTE, ang. N-terminal extension) – zawierającej motyw
wiążący RNA oraz DNA
• centralnej (RT, ang. reverse transcriptase) o aktywności katalitycznej
• C-końcowej (CTE, ang. C-terminal extension) [51-53].
Na rycinie 9 przedstawiony został schemat organizacji strukturalnej
podjednostki TERT telomerazy.

M. Bryś i wsp. 309
Ryc. 9. Schemat organizacji strukturalnej podjednostki TERT telomerazy. U większości organizmów domena TERT składa się z długiego N-końca (NTE), domeny centralnej
o aktywności katalitycznej (RT) oraz krótkiego C-końca (CTE). Na schemacie zaznaczono motywy CP, QFP i TS, wchodzące w skład odcinka TRBD
oraz telomerazowo specyficzną domenę IFD (pole zakreślone ukośnie). W obrębie N-końca znajduje się region „linkerowy” łączący domenę TEN i TRBD
[wg 55; w modyfikacji własnej].
Domena N-końcowa zawiera dwa motywy - TEN (N-końcowy motyw
telomerazy, ang. telomerase essential N-terminal) i TRBD (motyw wiążący się
z RNA, ang. telomerase RNA-binding domain). Oba odpowiedzialne są za
fałdowanie białka zaangażowanego w wiązanie się z jednoniciowym kwasem
nukleinowym. Pomiędzy tymi regionami znajduje się stosunkowo długi region
łącznikowy, który może mieć znaczenie w utrzymaniu elastyczności
holoenzymu. N-końcowa część białka TERT zawiera również kilka
specyficznych dla telomerazy motywów, w tym CP, QFP i TS. Regiony te
odgrywają rolę w oddziaływaniach TERT-TERC, a także mają wpływ na
szybkość kopiowania matrycy podczas syntezy telomerów [44]. Katalityczna
domena białka TERT zawiera siedem ewolucyjnie konserwatywnych motywów
RT, istotnych dla aktywności enzymatycznej telomerazy. Domena RT
zorganizowana jest w dwie poddomeny, które przypominają „palce” i „dłoń”.
Regiony te połączone są w pętlę, która zawiera konserwatywny region

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 310
„primer grip”, determinujący aktywność enzymatyczną. Pętla ta umożliwia
bezpośredni kontakt hybrydy RNA-DNA z końcem 3’ jednoniciowego DNA, co
sugeruje, że region „primer grip” może brać udział w lokowaniu końca 3’ DNA
w centrum aktywnym enzymu. Unikatową cechą domeny RT jest obecność
poddomeny IFD (domena odcinka RT białka TERT, ang. insertion in fingers).
Składa się ona z dwóch antyrównoległych helis α, które umieszczone są
pomiędzy „palcami” a „dłonią”. Poddomena IFD zaangażowana jest
w wewnątrzcząsteczkowe oddziaływania białko-białko, które warunkują
i stabilizują konformację przestrzenną tego regionu domeny RT [44, 54].
Domena C-końcowa w przeciwieństwie do domeny N-końcowej i domeny RT,
jest słabo zachowana ewolucyjnie. Pozwala to wnioskować, iż może ona
spełniać specyficzne gatunkowo funkcje. Jak dotąd nie wyjaśniono kwestii
dlaczego domena C-końcowa jest niezbędna dla aktywności telomerazy u, na
przykład orzęska i człowieka, ale nie jest wymagana dla aktywności tego
enzymu u drożdży i w ogóle nie występuje u innych organizmów [44].
Z wymienionymi podjednostkami telomerazy zasocjowane są liczne białka, jak
na przykład TP1 (białko związane z telomerazą 1, ang. telomerase associated
protein 1), Hsp23, Hsp90 (białka szoku cieplnego 23 i 90, ang. heat shock
protein 23, 90), hStau (białko łączące się z dwuniciowym RNA, ang. human
Staufen), L22 i dyskeryna. Ich obecność nie jest jednak konieczna dla
zachowania aktywności telomerazy [56-59]. Wydaje się, że domena hTERT jest
głównym wyznacznikiem aktywności telomerazy. Zgodnie z danymi
literaturowymi, gen hTERC ulega ekspresji we wszystkich typach tkanek,
natomiast ekspresja genu hTERT ograniczona jest do tak zwanych tkanek
telomerazowo-dodatnich, co sugeruje, że hTERT jest czynnikiem
odpowiedzialnym za enzymatyczną aktywność telomerazy.
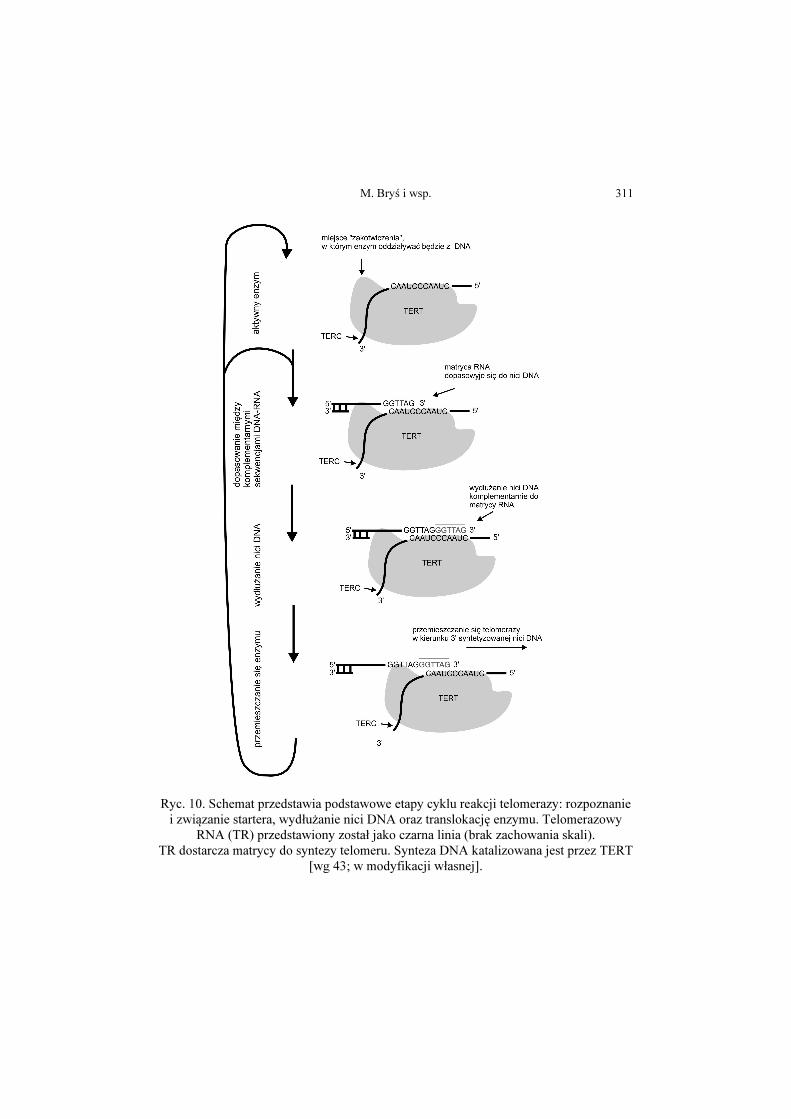
M. Bryś i wsp. 311
Ryc. 10. Schemat przedstawia podstawowe etapy cyklu reakcji telomerazy: rozpoznanie i związanie startera, wydłużanie nici DNA oraz translokację enzymu. Telomerazowy
RNA (TR) przedstawiony został jako czarna linia (brak zachowania skali). TR dostarcza matrycy do syntezy telomeru. Synteza DNA katalizowana jest przez TERT
[wg 43; w modyfikacji własnej].

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 312
Wykazano, że sztucznie wymuszona ekspresja genu TERT wydłuża
żywotność komórki w ponad dwudziestu różnych typach komórek [50, 60, 61].
Rzeczywisty mechanizm reakcji telomerazy można opisać w trzech etapach.
W pierwszym etapie, krótka telomerowa sekwencja na końcu 3’ chromosomu
wiąże się z matrycowym RNA telomerazy. Kolejny etap stanowi wydłużanie
nici, gdzie podjednostka TERT wykorzystuje fragment RNA jako matrycę do
produkcji nowego zakończenia telomeru. Ostatnim etapem jest translokacja bądź
dysocjacja, gdzie telomeraza przenosi się na nowy koniec 3’ chromosomu w
celu dodawania kolejnych powtórzeń lub opuszcza obszar telomeru [50]. Rycina
10 przedstawia mechanizm telomerazy.
Regulacja ekspresji genu TERT na poziomie mRNA i białka odbywać się
może na poziomie transkrypcji genu poprzez udział czynników transkryp-
cyjnych oraz modyfikacje potranskrypcyjne i potranslacyjne [62, 63]. Ścisła
korelacja między ekspresją genu hTERT na poziomie mRNA a aktywnością
telomerazy sugeruje, że regulacja transkrypcji tego genu jest jednym
z podstawowych mechanizmów regulujących aktywność enzymu [64; 65].
Analiza promotora genu hTERT wykazała obecność szeregu potencjalnych
miejsc dla oddziaływania z czynnikami transkrypcyjnymi, zarówno
wzmacniającymi, jak i wyciszającymi transkrypcję genu. Schemat obszaru
promotorowego genu TERT z zaznaczonymi miejscami oddziaływania
z wybranymi czynnikami transkrypcyjnymi, przedstawia rycina 11. Dwie
sekwencje oddziałujące z białkami c-Myc i Mad/Max oznaczane są jako dalszy
i bliższy motyw E-box (ang. distal/proximal E-box) [58, 65-67].
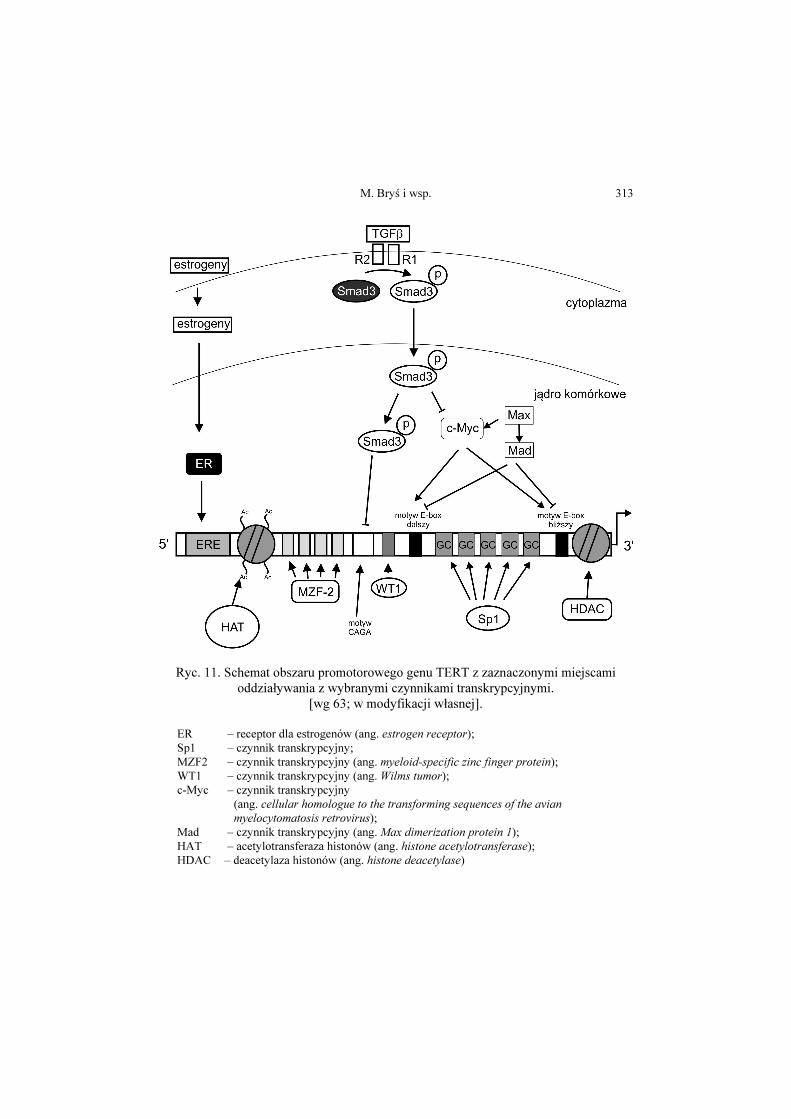
M. Bryś i wsp. 313
Ryc. 11. Schemat obszaru promotorowego genu TERT z zaznaczonymi miejscami oddziaływania z wybranymi czynnikami transkrypcyjnymi.
[wg 63; w modyfikacji własnej].
ER – receptor dla estrogenów (ang. estrogen receptor); Sp1 – czynnik transkrypcyjny; MZF2 – czynnik transkrypcyjny (ang. myeloid-specific zinc finger protein); WT1 – czynnik transkrypcyjny (ang. Wilms tumor); c-Myc – czynnik transkrypcyjny
(ang. cellular homologue to the transforming sequences of the avian myelocytomatosis retrovirus);
Mad – czynnik transkrypcyjny (ang. Max dimerization protein 1); HAT – acetylotransferaza histonów (ang. histone acetylotransferase); HDAC – deacetylaza histonów (ang. histone deacetylase)
Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 314
Tabela 3. Zestawienie czynników regulujących ekspresję genu TERT [wg 68].
Czynnik regulujący i szlak komórkowy, w którym bierze udział
Funkcja związane z regulacją
transkrypcji genu TERT
Cykl komórkowy
CDK2 – kinaza zależna od cyklin (ang. cyclin dependent kinase) CDK4 - kinaza zależna od cyklin (ang. cyclin dependent kinase) cyklina D cyklina E p21 p53 p27 Rb (retinoblastoma; ang retinoblastoma protein) E2F1 (ang. E2F transcriptiom factor 1) p14 MDM2 (mysie biało MDM2; ang. murine double minute 2) p16
aktywator aktywator aktywator aktywator represor represor represor represor represor represor
aktywator represor
Szlak TGF-β
TGF-β (transformujący czynnik wzrostu β; ang. transforming
growth factor beta) TGF-βR1 (receptor 1 dla transformującego czynnika wzrostu β;
ang. transforming growth factor beta receptor 1) TGF-βR2 (receptor 2 dla transformującego czynnika wzrostu β;
ang. transforming growth factor beta receptor 2) SMAD3 (Sma i MAD; MAD ang. mothers against
decapentaplegic 3) SMAD4 (Sma i MAD; MAD ang. mothers against
decapentaplegic 4) E2F4 (ang. E2F transcriptiom factor 4) E2F5 (ang. E2F transcriptiom factor 5)
represor
represor
represor
represor
represor represor represor
Szlak PI3K/Akt
PI3K (kinaza fosfatydyloinozytolu 3; ang. phosphatidylinositide 3-kinases)
Akt PTEN (ang. phosphatase and tensin homolog) mTOR (ang. mammalian target of rapamycin) IKK (inhibitor kinazy kappaB; ang. inhibitor of kappaB kinase)
aktywator aktywator represor
aktywator aktywator

M. Bryś i wsp. 315
Tabela 3. Zestawienie czynników regulujących ekspresję genu TERT [wg 68]. (cd)
Czynnik regulujący i szlak komórkowy, w którym bierze udział
Funkcja związane z regulacją
transkrypcji genu TERT
Szlak NF-κB (kanoniczny) NF-κB (p50) (ang. nuclear factor kappa-light-chain-enhancer of
activated B cells) NF-κB (p65) IKK (ang. inhibitor of kappaB kinase) IκB (ang. inhibitor of kappa B)
aktywator aktywator aktywator represor
Szlak NF-κB (niekanoniczny) NIK (ang. NF-kappaB-inducing kinase) IKK NF-κB (p100) NF-κB (RelB)
aktywator aktywator aktywator aktywator
Szlak kinaz MAP
RAS (ang. rat sarcoma) MEK (ang. mitogen extracellular kinase) ERK (ang. extracellular signal-regulated kinase)
aktywator aktywator aktywator
Szlak erbB
EGF (ang. epidermal growth factor) EGFR (ang. epidermal growth factor receptor) erB-2 (ang. human epidermal growth factor 2) PI3K
aktywator aktywator aktywator aktywator
Inne
AP2 (ang. activator protein 2) AP4 (ang. activating enhancer-binding protein 4) BRCA1 (ang. breast cancer type 1 susceptibility protein 1) c-Ets-2 (ang. v-ets erythroblastosis virus E26 oncogene
homolog 2) c-Myb (ang. myeloblastosis) CREB/ATF (ang. cAMP responsive element binding protein/
activating transcription factors) CTCF (ang. CCCTC-binding factor) AP1 (ang. activator protein 1) ERβ (ang. estrogen receptor beta) IK2 (ang. I kappaB kinase-like 2)
aktywator aktywator represor
aktywator aktywator
aktywator represor
aktywator/represor aktywator aktywator

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 316
Tabela 3. Zestawienie czynników regulujących ekspresję genu TERT [wg 68]. (cd)
Czynnik regulujący i szlak komórkowy, w którym bierze udział
Funkcja związane z regulacją
transkrypcji genu TERT
Inne (cd)
INHBC (ang. inhibin beta C chain) MAD3 (ang. mitotic spindle assembly checkpoint protein MAD3) MAZ (ang. Myc-associated zinc finger protein) Menin MNT (ang. MAX binding protein) MyoD (ang. myoblast determination protein) MZF-2 (ang. myeloid zinc finger protein 2) NF1 (ang. nuclear factor 1) NFAT (ang. nuclear factor of activated T-cells) NF-E2 [ang. nuclear factor (erythroid-derived 2)] PITX1 (ang. paired-like homeodomain 1) Sp1 (ang. specificity protein 1) USF1/USF2 (ang. upstream stimulatory factor 1/2) WT1 (ang. Wilms tumor)
aktywator represor
aktywator represor represor
aktywator/represor represor
aktywator aktywator aktywator represor
aktywator aktywator/represor
represor
W Tabeli 3 zestawiono czynniki oddziałujące z sekwencjami regulatoro-
wymi genu TERT, wzmacniające oraz wyciszające jego ekspresję. Czynniki
podzielono na grupy odnośnie do ich udziału w szlakach komórkowych [68].
Szczegółowo opisany został mechanizm regulacji ekspresji genu TERT z udzia-
łem czynnika transkrypcyjnego c-Myc. Białko to jest produktem protoonkogenu
c-MYC i należy do klasy białek posiadających domenę typu helisa-pętla-helisa
(HLH; ang. helix-loop-helix). Czynnik ten oddziałuje z DNA wyłącznie w po-
staci kompleksu z białkiem Max. W przypadku negatywnej regulacji trans-
krypcji z udziałek białka c-Myc, konieczne jest dodatkowe oddziaływanie czyn-
nika transkrypcyjnego Max (ang. myc-associated factor X) z białkiem jądrowym
Mad (ang. MAX dimerization protein). W komórkach nowotworowych

M. Bryś i wsp. 317
konsekwencją tego procesu jest przesunięcie równowagi w kierunku kompleksu
Myc-Max, co prowadzi do ciągłych podziałów komórkowych [69].
Aktywacja ekspresji genu TERT wymaga oddziaływania z DNA, w obrębie
regionu promotorowego, zarówno czynników modyfikujących strukturę
chromatyny, jak i podstawowych czynników transkrypcyjnych. W przypadku
regulacji ekspresji genu TERT za pośrednictwem białka c-Myc,
zidentyfikowano poza kompleksem Mad-Max, białka koregulatorowe takie jak,
koaktywatory transkrypcji o aktywności acetylotransferazy histonów
(p300/CBP; (ang. CREB binding protein and binding protein p300), kompleks
STAGA (SPT3-TAF9-GCN5-acetylaza; SPT3; ang. suppressor of Ty 3, TAF9;
ang. TBP-associated factor 9, GCN5, acetylase) pełniący funkcję koaktywatora
i kompleks Mediator (kompleks białkowy oddziałujący z polimerazą RNA II)
[70-74]. Szczegółowy model składania kompleksu preinicjacyjnego na
promotorze genu TERT, przy udziale czynnika transkrypcyjnego c-Myc,
przedstawia rycina 12.
Czynniki transkrypcyjne, których udział w regulacji ekspresji genu TERT
został stwierdzony, podlegają wzajemnym oddziaływaniom (ang. cross talk).
W wyniku tych procesów mogą one osłabiać lub wzmacniać swoją aktywność
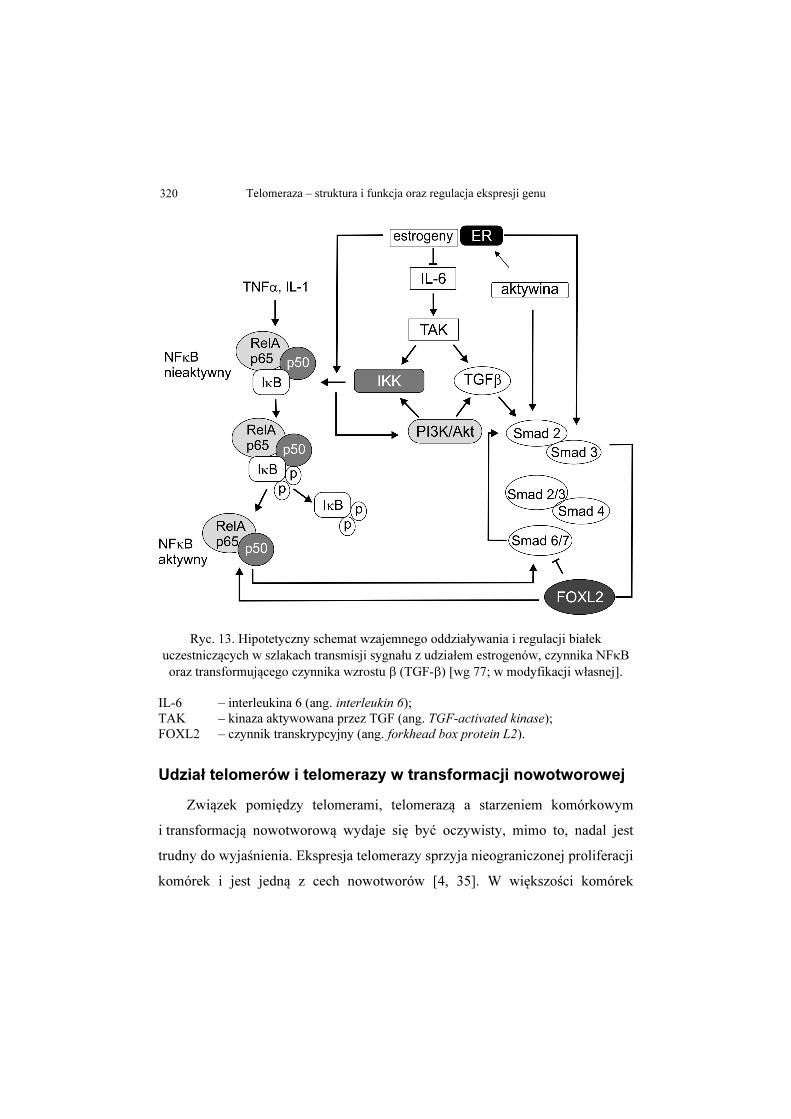
[77]. Rycina 13 przedstawia hipotetyczny schemat wzajemnego oddziaływania
i regulacji białek uczestniczących w szlakach transmisji sygnału z udziałem
estrogenów, czynnika NFkB oraz transformującego czynnika wzrostu β
(TGF-β).
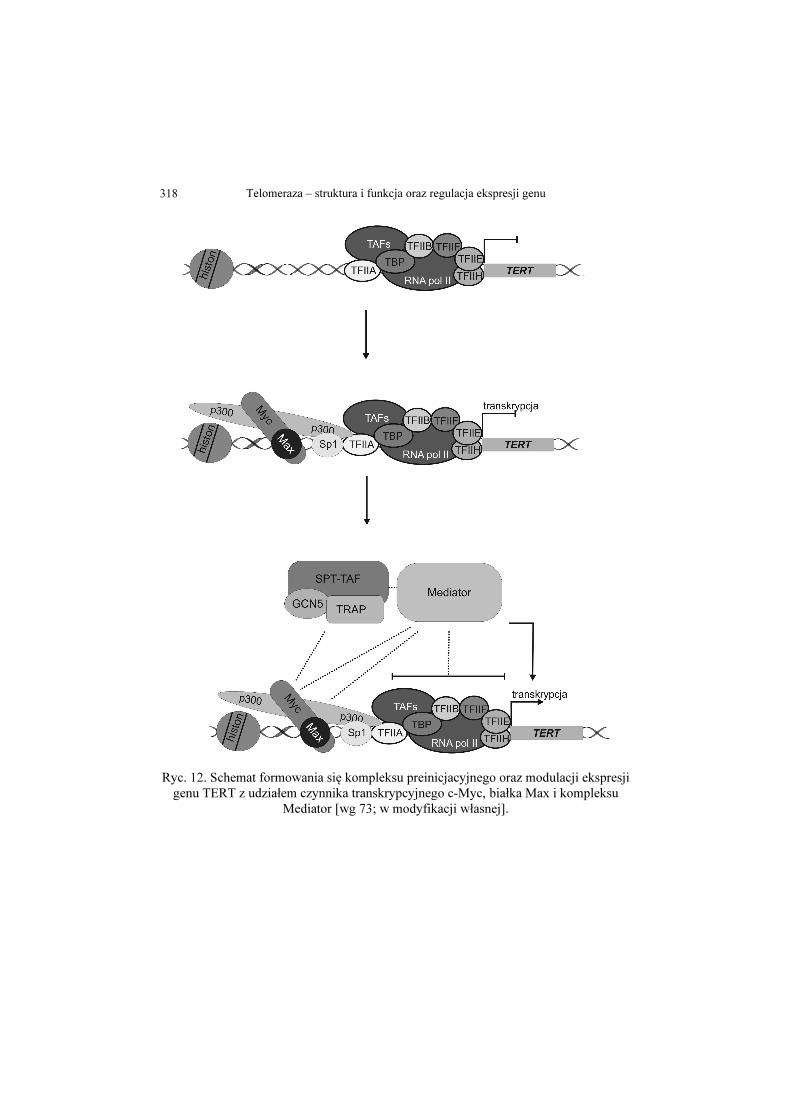
Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 318
Ryc. 12. Schemat formowania się kompleksu preinicjacyjnego oraz modulacji ekspresji genu TERT z udziałem czynnika transkrypcyjnego c-Myc, białka Max i kompleksu
Mediator [wg 73; w modyfikacji własnej].

M. Bryś i wsp. 319
CBP/p300 - koaktywatory transkrypcji o aktywności acetylotransferazy histonów
(ang. CREB binding protein and binding protein p300); c-Myc - czynnik transkrypcyjny
(ang. cellular homologue to the transforming sequences of the avian myelocytomatosis retrovirus);
CREB - czynnik transkrypcyjny oddziałujący z sekwencją CRE aktywowany w wyniku fosforylacji przez PKA (ang. cAMP response element binding);
GCN5 - acetylotransferaza histonów; Max - czynnik transkrypcyjny (ang. myc-associated factor X); p300/CBP - koaktywatory transkrypcji o aktywności acetylotransferazy histonów (ang. CREB
binding protein and binding protein p300); SPT3 - białko wchodzące w skład kompleksu STAGA (ang. suppressor of Ty 3); STAGA - kompleks białkowy o funkcji acetylazy histonów i aktywatora transkrypcji (ang.
SPT-ADA-GCN5 acetylase); TAF - czynnik oddziałujący z białkiem TBP (ang. TBP-associated factor); TATA - sekwencja DNA znajdująca się obszarze promotorowym genów eukariotycznych
(ang. TATA-box); TBP - białko łączące się z sekwencją TATA (ang. TATA-box binding protein); TF - podstawowy czynnik transkrypcyjny (ang. transcription factor); TRRAP - białko o aktywności fosfatazy (ang. tartrate-resistant acid phosphatase).
Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 320
Ryc. 13. Hipotetyczny schemat wzajemnego oddziaływania i regulacji białek uczestniczących w szlakach transmisji sygnału z udziałem estrogenów, czynnika NFκB oraz transformującego czynnika wzrostu β (TGF-β) [wg 77; w modyfikacji własnej].
IL-6 – interleukina 6 (ang. interleukin 6); TAK – kinaza aktywowana przez TGF (ang. TGF-activated kinase); FOXL2 – czynnik transkrypcyjny (ang. forkhead box protein L2).
Udział telomerów i telomerazy w transformacji nowotworowej
Związek pomiędzy telomerami, telomerazą a starzeniem komórkowym
i transformacją nowotworową wydaje się być oczywisty, mimo to, nadal jest
trudny do wyjaśnienia. Ekspresja telomerazy sprzyja nieograniczonej proliferacji
komórek i jest jedną z cech nowotworów [4, 35]. W większości komórek

M. Bryś i wsp. 321
somatycznych człowieka stwierdza się aktywność telomerazy poniżej granicy
wykrywalności, podczas gdy nowotwory złośliwe cechują się wysoką
aktywnością tego enzymu. Ulega ona ekspresji w prawie 90% nowotworów
człowieka (Tabela 4.).
Tabela 4. Aktywność telomerazy w wybranych nowotworach człowieka [wg 75].
UKŁAD NERWOWY
mózg (prawidłowy) 0% siatkówczak (łac. retinoblastoma) 50% glejak wielopostaciowy (łac. glioblastoma multiforme) 75% skąpodrzewiak (łac. oligodendroglioma) 100% gwiaździak anaplastyczny (łac. astrocytoma anaplasticum) 100% oponiaki (łac. meningiomas) 17-100% nerwiak płodowy (łac. neuroblastoma) 94%
GŁOWA I SZYJA
błona śluzowa jamy ustnej (prawidłowa) 3% rak kolczystokomórkowy skóry (łac. carcinoma spinocellulare) 86%
PŁUCA
rak płaskonabłonkowy drobnokomórkowy (łac. carcinoma planoepitheliale microcellulare) 100% rak płaskonabłonkowy niedrobnokomórkowy (łac. carcinoma planoepitheliale non microcellulare) 86%
GRUCZOŁ PIERSIOWY
tkanka prawidłowa 0% gruczolakowłókniak (łac. fibroadenoma) 28% rak nieinwazyjny przewodowy (łac. carcinoma ductale in situ) 75% rak przewodowy lub zrazikowy (łac. carcinoma ductale or lobulare) 88%
WĄTROBA
tkanka prawidłowa 0% nowotwory łagodne 29% rak pierwotny (łac. carcinoma hepatocellulare) 86%

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 322
Tabela 4. Aktywność telomerazy w wybranych nowotworach człowieka [wg 75]. (cd)
UKŁAD POKARMOWY
gruczolak żołądka (łac. adenoma ventriculi,) 27% rak żołądka (łac. carcinoma ventriculi,) 86% polip gruczolakowaty (łac polypus adenomatosus) 48%
rak jelita grubego (łac. carcinoma coli et recti) 85% rak trzustki (łac. carcinoma pancreatis) 95%
UKŁAD ROZRODCZY - KOBIETY
jajniki 33% błona śluzowa trzonu macicy (prawidłowa) 0% mięśniak gładkokomórkowy (łac. leiomyoma) 0% mięsak gładkokomórkowy (łac. leiomyosarcoma) 100% rak endometrium (łac. carcinoma endometriale) 100% rak jajnika (łac. carcinoma ovarii) 91%
UKŁAD ROZRODCZY - MĘŻCZYŹNI
jądra (łac. testis) 100% gruczoł krokowy (łac. prostata) 0% łagodny rozrost gruczołu krokowego (łac. hyperplasia prostatae) 5% rak gruczołu krokowego (łac. carcinoma prostatae) 90%
SKÓRA
naskórek (łac. epidermis) 44% czerniak (łac. melanoma malignum) 86%
Wysoka aktywność telomerazy może prowadzić do tzw. „unieśmiertel-
nienia” komórki i/lub umożliwić jej nieograniczoną proliferację. W przypadku
braku aktywności tego enzymu, telomery ulegają skracaniu w każdej rundzie
podziału komórkowego, aż do osiągnięcia długości progowej [76].
Nowotwory złośliwe są chorobami charakteryzującymi się niekontrolowaną
proliferacją oraz naciekaniem otaczających tkanek lub odległych narządów.
Większość komórek somatycznych człowieka nie wykazuje aktywności
telomerazy, ma ograniczoną żywotność i w celu wydłużenia długości ich życia
konieczne jest uaktywnienie telomerazy. Doniesienia literaturowe sugerują, że
telomeraza może sprzyjać procesowi transformacji nowotworowej niezależnie

M. Bryś i wsp. 323
od długości telomerów [33, 78]. Poprzez stabilizację długości telomerów
i stymulowanie nieograniczonych podziałów komórek nowotworowych,
telomeraza odgrywa kluczową rolę w etiologii i progresji nowotworów. Uważa
się, iż telomeraza może być aktywowana na różnych etapach procesu
nowotworowego. W niektórych przypadkach ekspresja tego enzymu może być
obserwowana w stanach przednowotworowych, podczas gdy w innych
przypadkach telomeraza może być aktywowany dopiero na etapie progresji
procesu nowotworowego. Różnice te mogą w bardzo istotny sposób rzutować na
kliniczną użyteczność telomerazy jako markera nowotworowego oraz określenie
czy enzym ten może być wykorzystany w przesiewowych badaniach pacjentów
z grupy wysokiego ryzyka, w diagnostyce wczesnego wykrywania, czy też do
celów prognostycznych [79]. Pomimo, iż w większości komórek somatycznych
obserwuje się brak aktywności telomerazy, to jednak niektóre tkanki posiadają
wyspecjalizowane komórki, takie jak limfocyty, komórki rozrodcze, komórki
nabłonkowe, które wykazują niewielki poziom aktywności tego enzymu.
W tkankach zawierających wymienione wyżej komórki, wykrywanie
aktywności telomerazy jest utrudnione i wymaga precyzyjnego określenia czy
aktywność enzymu pochodzi z prawidłowych, telomerazowo-dodatnich
komórek, czy też z komórek zmienionych nowotworowo. Jak już zostało
wspomniane, aktywność ludzkiej telomerazy uwarunkowana jest obecnością
dwóch głównych składowych - fragmentu nici RNA (hTERC) oraz podjednostki
o aktywności odwrotnej tranksryptazy (hTERT). Prowadzone obecnie badania
naukowe i kliniczne koncentrują się na osobnej analizie ekspresji tych dwóch
elementów, jako doskonalszych zamienników w stosunku do oznaczania
aktywności telomerazy [52, 80].

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 324
Podsumowanie
Poznanie struktury i funkcji telomerów oraz telomerazy przyczyniło się do
opracowania nowych form leczenia przeciwnowotworowego. Przewiduje się, że
blokowanie aktywności telomerazy zaowocuje w niedalekiej przyszłości
opracowaniem skuteczniejszych leków dla pacjentów cierpiących z powodu
chorób nowotworowych, jak również rozszerzy możliwość ich chemoprewencji.
Najbardziej efektywne wydaje się zastosowanie kombinacji klasycznych
chemioterapeutyków z bezpośrednimi inhibitorami aktywności telomerazy, czy
związkami blokującymi oddziaływania telomeraza-telomer. Dzięki takiemu
postępowaniu terapeutycznemu być może uda się zmniejszyć dawki
stosowanych leków, a tym samym zmniejszyć skutki uboczne terapii.
Piśmiennictwo
1. Zvereva MI, Shcherbakova DM, Dontsova OA. Telomerase: structure, functions, and activity regulation. Biochemistry. 2010; 75: 1563-1583.
2. Kim NW, Piatyszek MA, Prowse KR, Harley CB, West MD, Ho PL i wsp. Specific association of human telomerase activity with immortal cells and cancer. Science. 1994; 266: 2011-2015.
3. Meyerson M, Counter CM, Eaton EN, Ellisen LW, Steiner P, Caddle SD i wsp. Hest2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell. 1997; 90:785–795.
4. Sikora E. Telomery i telomeraza — starzenie komórkowe nagroda nobla z fizjologii lub medycyny. Kosmos. 2009; 59: 286–287.
5. Wright WE, Shay JW. Telomere biology in aging and cancer. J Am Geriatr Soc. 2005; 53: 292-294.
6. Kowalska A, Kowalik A. Telomer i telomeraza w ontogenezie. Współczesna Onkologia. 2006; 10: 485–496.
7. Shay JW, Wright WE. Telomerase therapeutics for cancer: challenges and new directions. Nat Rev Drug Discov. 2006; 5: 577-584.
8. Cheung AL, Deng W. Telomere dysfunction, genome instability and cancer. Front Biosci. 2008; 13: 2075–2090.

M. Bryś i wsp. 325
9. McGrath M, Wong JY, Michaud D, Hunter DJ, De Vivo I. Telomere length, cigarette smoking, and bladder cancer risk in men and women. Cancer Epidemiol Biomarkers Prev. 2007; 16:815–819.
10. Allshire RC, Dempster M, Hastie ND. Human telomeres contain at least three types of G-rich repeat distributed non-randomly. Nucleic Acids Res. 1989; 17: 4611–4627.
11. Blackburn EH, Gall JG. A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J Mol Biol. 1978; 120:33–53.
12. Griffith JD, Comeau L, Rosenfield S, Stansel RM, Bianchi A, Moss H i wsp. Mammalian telomeres end in a large duplex loop. Cell. 1999; 97: 503–514.
13. Moyzis RK, Buckingham JM, Cram LS, Dani M, Deaven LL, Jones MD i wsp. A highly conserved repetitive DNA sequence, (TTAGGG)n, present at the telomeres of human chromosomes. Proc Natl Acad Sci U S A. 1988; 85: 6622-6626.
14. Witzany G. Viral origins of telomeres and telomerases. And their important role in eukaryogenesis and genome maintenance. Biosemiotics. 2008; 1: 191.
15. Aubert G, Lansdorp PM. Telomeres and aging. Physiol Rev. 2008; 88: 557-579. 16. Marión RM, Blasco MA. Telomeres and telomerase in adult stem cells and
pluripotent embryonic stem cells. Advexpmed Biol. 2010; 695:118-131. 17. Bryan TM, Baumann P. G-quadruplexes: from guanine gels to chemo-
therapeutics. Mol Biotechnol. 2011; 49: 198-208. 18. Oeseburg H, de BoerR A, van Gilst WH, van der Harst P. Telomere biology in
healthy aging and disease. Eur J Physiol. 2010; 459: 259–268. 19. Songyang Z. Introduction to telomeres and telomerase. Methods Mol Biol. 2011;
735: 1-11. 20. Oganesian L, Moon IK, Bryan TM, Jarstfer MB. Extension of G-quadruplex
DNA by ciliate telomerase. EMBO J. 2006; 25: 1148-1159. 21. Oganesian L, Karlseder J. Telomeric armor: the layers of end protection. J Cell
Sci. 2009; 122: 4013-4025. 22. Olaussen KA, Dubrana K, Domont J, Spano JP, Sabatier L, Soria JCh. Telomeres
and telomerase as a targets for anticancer drug development. Crit Rev in Oncol/Hematol. 2006; 57:191-214.
23. De Lange T. T-loops and the origin of telomeres. Nat Rev Mol Cell Biol. 2004; 5: 323–329.

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 326
24. Akbay EA, Peña CG, Ruder D, Michel JA, Nakada Y, Pathak S i wsp. Cooperation between p53 and the telomere-protecting shelterin component Pot1a in endometrial carcinogenesis. Oncogene. 2012; 11. doi: 10.1038/onc.2012.232.
25. O’Sullivan RJ, Karlseder J. Telomeres: protecting chromosomes against genome instability. Nat Rev Mol Cell Biol. 2010; 11: 171–181.
26. Stewart JA, Chaiken MF, Wang F, Price CM. Maintaining the end: roles of telomere proteins in end-protection, telomere replication and length regulation. Mutat Res. 2012; 730: 12-19.
27. Xin H, Liu D, Songyang Z. The telosome/shelterin complex and itsfunctions. Genome Biol. 2008; 9:232.1-232.7.
28. Bayne S, Liu JP. Hormones and growth factors regulate telomerase activity in ageing and cancer. Mol Cell Endocrinol. 2005; 240:11–22.
29. Martinez P, Blasco MA. Telomeric and extra-telomeric roles for telomerase and the telomere-binding proteins. Nat Rev Cancer. 2011; 11: 161-176.
30. Bianchi A, de Lange T. Ku binds telomeric DNA in vitro. J Biol Chem. 1999; 274: 21223-21227.
31. Van Steensel B, de Lange T. Control of telomere length by the human telomeric protein TRF1. Nature. 1997; 385:740-743.
32. De Lange T. How telomeres solve the end-protection problem. Science. 2009; 326: 948-952.
33. Deng Y, Chang S. Role of telomeres and telomerase in genomic instability, senescence, and cancer. Lab Invest. 2007; 87: 1071–1076.
34. Urquidi V, Tarin D, Goodison S. Role of telomerase.in cell senescense and oncogenesis. Annu Rev Med. 2000; 51: 65-79.
35. Artandi SE, DePinho RA. Telomeres and telomerase in cancer. Carcinogenesis. 2010; 31: 9–18.
36. McNees CJ, Tejera AM, Martinez P, Murga M, Mulero F, Fernandez-Capetillo O i wsp. ATR suppresses telomere fragility and recombination but is dispensable for elongation of short telomeres by telomerase. J Cell Biol. 2010; 188: 639–652.
37. Zhu H, Belcher M, van der Harst P. Healthy aging and disease: role for telomere biology? Clinical Science. 2011; 120: 427–440.
38. Lingner J, Cooper JP, Cech TR. Telomerase and DNA end replication: no longer a lagging strand problem? Science. 1995; 269: 1533–1534.
39. Zhong Z-H, Jiang W-Q, Cesare AJ, Neumann AA, Wadhwa R, Reddel R R. Disruption of telomere maintenance by depletion of the MRE11/RAD50/NBS1 complex in cells that use alternative lengthening of telomeres. J Biol Chem. 2007; 282: 29314-29322.

M. Bryś i wsp. 327
40. Georgin-Lavialle S, Aouba A, Mouthon L, Londono-Vallejo JA, Lepelletier Y, Gabet AS i wsp. The telomere/telomerase system in autoimmune and systemic immune-mediated diseases. 2010; 9: 646-651.
41. Azzalin CM, Reichenbach P, Khoriauli L, Giulotto E, Lingner J. Telomeric repeat containing RNA and RNA surveillance factors at mammalian chromosome ends. Science. 2007; 318:798–801.
42. Baur JA, Zou Y, Shay JW, Wright WE. Telomere position effect in human cells. Science. 2001; 292: 2075–2077.
43. Podlevsky JD, Chen JJL. It all comes together at the ends: Telomerase structure, function, and biogenesis. Mutat Res. 2012; 730:3-11.
44. Wyatt HD, West SC, Beattie TL. In TERT preting telomerase structure and function. Nucleic Acids Res. 2010; 38: 5609-5622.
45. Philippi C, Loretz B, Schaefer UF, Lehr CM. Telomerase as an emerging target to fight cancer — Opportunities and challenges for nanomedicine. J Control Release. 2010; 146: 228–240.
46. Chen JL, Blasco MA, Greider CW. Secondary structure of vertebrate telomerase RNA. Cell. 2000; 100: 503-514.
47. Dandjinou AT, Levesque N, Larose S, Lucier JF, Abou Elela S, Wellinger RJ. A phylogenetically based secondary structure for the yeast telomerase RNA. Curr Biol. 2004; 14: 1148-1158.
48. Romero DP, Blackburn EH. A conserved secondary structure for telomerase RNA. Cell. 1991; 67:343-353.
49. Zappulla DC, Cech TR. Yeast telomerase RNA: a flexible scaffold for protein subunits. Proc Natl Acad Sci USA. 2004; 101: 10024-10029.
50. Feng J, Funk WD, Wang SS, Weinrich SL, Avilion AA, Chiu CP i wsp. The RNA component of human telomerase. Science. 1995; 269: 1236–1241.
51. Autexier C, Lue NF. The structure and function of telomerase reverse transcriptase. Annu Rev Biochem. 2006; 75: 493–517.
52. Avendaño C, Menéndez JC. Medicinal Chemistry of Anticancer Drugs. Elsevier, Amsterdam, 2008.
53. Collins K. The biogenesis and regulation of telomerase holoenzymes. Nat Rev Mol Cell Biol. 2006; 7: 484–494.
54. Wyatt HD, Lobb DA, Beattie TL. Characterization of physical and functional anchor site interactions in human telomerase. Mol Cell Biol. 2007; 27:3226-3240.
55. Nicholls C, Li H, Wang JQ, Liu JP. Molecular regulation of telomerase activity in aging. Protein Cell. 2011; 2:726–738.

Telomeraza – struktura i funkcja oraz regulacja ekspresji genu 328
56. Blackburn EH. Switching and signaling at the telomere. Cell. 2001; 106: 661-673.
57. Horikawa L, Oshimura M, Barrett JC. Repression of the telomerase catalytic subunit by a gene on human chromosome 3 that induces cellular senescence. Mol Carcinog. 1998; 22: 65-72.
58. Kyo S, Takakura M, Kanaya T, Zhuo W, Fujimoto K, Nishio Y i wsp. Estrogen activates telomerase. Cancer Res. 1999; 59: 5917–5922.
59. Nakamura TM, Morin GB, Chapman KB, Weinrich SL, Andrews WH, Lingner J i wsp. Telomerase catalytic subunit homologs from fission yeast and human. Science. 1997; 277: 955–959.
60. Nakayama J, Tahara H, Tahara E, Saito M, Ito K, Nakamura H i wsp. Telomerase activation by hTRT in human normal fibroblasts and hepatocellular carcinomas. Nat Genet. 1998; 18: 65–68.
61. Weinrich Sl, Pruzan R, Ma L, Ouellette M, Tesmer Vm, Holt Se i wsp. Reconstitution of human telomerase with template RNA component hterc and the catalytic proteinsubunit htert. Nat Genet. 1997; 17: 498–502.
62. Liu JP. Studies of the molecular mechanisms in the regulation of telomerase activity. FASEB J. 1999; 13: 2091–2104.
63. Liu JP. Telomerase: Not just black and white, but shades of gray. Mol Cell Biol Res Commun. 2000; 3: 129–135.
64. Bodnar AG, Ouellette M, Frolkis M, Holt SE, Chiu CP, Morin GB i wsp. Extension of life-span by introduction of telomerase into normal human cells. Science. 1998; 279: 349–352.
65. Xu D, Gruber A, Bjorkhold M, Peterson C, Pisa P. Suppression of telomerase reverse transcriptase (hTERT) expression in differentiated HL-60 cells: regulatory mechanisms. Br J Cancer. 1999; 80: 1156–1161.
66. Horikawa I, Cable PL, Afshari C, Barrett JC. Cloning and characterization of the promoter region of human telomerase reverse transcriptase gene. Cancer Res. 1999; 59: 826–830.
67. Misiti S, Nanni S, Fontemaggi G, Cong YS, Wen J, Hirte HW i wsp. Induction of hTERT expression and telomerase activity by estrogens in human ovary epithelium cells. Mol Cell Biol. 2000; 20: 3764–3771.
68. Daniel M, Peek GW, Tollefsbol TO. Regulation of the human catalytic subunit of telomerase (hTERT). Gene. 2012; 498: 135-46.
69. Ścibior-Bentkowska D, Czeczot H. Komórki nowotworowe a stres oksydacyjny. Postępy Hig Med Dośw. 2009; 63: 58-72.

M. Bryś i wsp. 329
70. Conaway RC, Conaway JW. Origins and activity of the Mediator complex. Semin Cell Dev Biol. 2011; 22: 729-734.
71. Gamper AM, Kim J, Roeder RG. The STAGA subunit ADA2b is an important regulator of human GCN5 catalysis. Mol Cell Biol. 2009; 29: 266-280.
72. Holmlund T, Lindberg MJ, Grander D, Wallberg AE. GCN5 acetylates and regulates the stability of the oncoprotein E2A-PBX1 in acute lymphoblastic leukemia. Leukemia. 2012; doi: 10.1038/leu.2012.265.
73. Liu X, Vorontchikhina M, Wang YL, Faiola F, Martinez E. STAGA recruits Mediator to the MYC oncoprotein to stimulate transcription and cell proliferation. Mol Cell Biol. 2008; 28: 108-121.
74. Taatjes DJ. The human Mediator complex: a versatile, genome-wide regulator of transcription. Trends Biochem Sci. 2010; 35: 315-322.
75. Granger MP, Wright WE, Shay JW. Telomerase in cancer and aging. Crit Rev Oncol Hematol. 2002; 41: 29-40.
76. .Biroccio A, Leonetti C. Telomerase as a new target for the treatment of hormone-refractory prostate cancer. Endocr Relat Cancer. 2004; 11:407-421.
77. Drummond AE, Fuller PJ. Activin and inhibin, estrogens and NFκB, play roles in ovarian tumourigenesis is there crosstalk? Mol Cell Endocrinol. 2012; 359: 85-91.
78. Heaphy CM, Meeker AK. The potential utility of telomere-related markers for cancer diagnosis. J Cell Mol Med. 2011; 15: 1227-1238.
79. Wanat JJ, Johnson FB. Telomere stability and carcinogenesis: an off-again, on-again relationship. J Clin Invest. 2012; 122: 1962-1965.
80. Elmore LW, Forsythe R, Forsythe H, Bright AT, Nasim S, Endo K i wsp. Overexpression of telomerase-associated chaperone proteins in prostatic intraepithelial neoplasia and carcinomas. Oncol Rep. 2008; 20:613-617.





























