Embed Size (px)
Citation preview


UNIVRSIDAD AUTÓNOMA METROPOLITANA UNIDAD IZTAPALAPA DIVISIÓN CIENCIAS BIOLÓGICAS Y DE LA SALUD GRADO LICENCIATURA TÍTULO DEL TRABAJO DE SERVICIO SOCIAL ESTRUCTURA DE LA COMUNIDAD DE PECES EN LA BAHÍA CACALUTA, HUATULCO, OAXACA. PARTICIPANTES BRIAN LUNA MONSIVAIS ASESOR DR. MARGARITO TAPIA GARCÍA LUGAR Y FECHA DE REALIZACIÓN HUATULCO, OAXACA. ENERO 2004 A JULIO 2005

NOMBRE LUNA MONSIVAIS BRIAN MATRÍCULA 99222361 TELÉFONO 56064934 LICENCIATURA HIDROBIOLOGÍA DIVISIÓN CIENCIAS BIOLÓGICAS Y DE LA SALUD UNIDAD IZTAPALAPA TRIMESTRE LECTIVO 03-O TÍTULO DEL PROYECTO DIAGNÓSTICO DE LA BIODIVERSIDAD DEL SISTEMA ZANJÓN-ESTERO LA VENTOSA Y EL RÍO TEHUANTEPEC, OAXACA, MÉXICO. TÍTULO DEL TRABAJO DE SERVICIO SOCIAL ESTRUCTURA DE LA COMUNIDAD DE PECES EN LA BAHÍA CACALUTA, HUATULCO, OAXACA. ASESOR DR. MARGARITO TAPIA GARCÍA LUGAR DE REALIZACIÓN HUATULCO, OAXACA CLAVE DE REGISTRO H.024.03

Nombre: Luna Monsivais Brian. Matrícula: 99222361. Licenciatura: Hidrobiología. Título del trabajo: ESTRUCTURA DE LA COMUNIDAD DE PECES EN LA BAHÍA CACALUTA, HUATULCO, OAXACA. Clave de registro: H.024.03 Fecha de entrega: 1-07-2005 Asesor: Dr. Margarito Tapia García. Área de Diagnóstico Ecológico y Gestión Ambiental. Resumen Se determinó la estructura y función de la comunidad de peces en la Bahía Cacaluta; la composición, distribución, abundancia, diversidad, dominancia, asociaciones ictiofaunísticas y la variación espacio-temporal de la ictiofauna en tres meses de muestreo correspondientes a dos épocas climáticas diferentes (secas y lluvias) con relación a los diferentes tipos de ambientes presentes en esta bahía cuales son el fondo de arrecife de coral, fondo rocoso y fondo arenoso. Se registraron 1,443 organismos correspondientes a 29 especies, 27 géneros, 18 familias y 2 subfamilias. La mayor abundancia (densidad) y riqueza de especies en los tres meses de muestreo la presentó el ambiente de arrecife de coral debido a su alta irregularidad y complejidad morfológica; mientras que el ambiente rocoso debido a su menor extensión, irregularidad y complejidad, así como por su mayor incidencia de oleaje presentó valores menores en estos parámetros. Por su parte la diversidad y dominancia parecen alternarse entre el ambiente arrecifal y el rocoso dependiendo de la época climática ya que durante la temporada de secas la mayor diversidad la presentó el ambiente rocoso, mientras que en la época de lluvias el ambiente de arrecife de coral es el que presenta la mayor diversidad; de manera semejante se comportan los valores de dominancia pues durante secas su mayor valor se registró en el ambiente de arrecife de coral, en tanto que durante lluvias la mayor dominancia se da en el ambiente rocoso. Lo cual se atribuye a la influencia que ejerce la abundancia y disponibilidad del plancton sobre la ictiofauna planctivora durante las dos épocas ya que durante secas hay gran abundancia de este recurso en el lugar y por las condiciones que ofrece el arrecife en cuanto a resguardo y protección es que hay la presencia de cardúmenes de especies que se alimentan del plancton incrementandose la dominacia en la zona, mientras que durante lluvias la abundancia del plancton disminuye teniendo como consecuencia que la dominancia disminuya en la zona de coral y esta se incremente en el ambiente rocoso debido a las condiciones más difíciles que en este ambiente imperan. Esta domianacia se encuentra dada principalmente por las especies Thalassoma lucasanum, Stegastes acapulcoensis y Microspathodon dorsalis; caracterizadas por presentar una alta temporada reproductiva, alta fecundidad y alta frecuencia reproductiva. En el ambiente arenoso solo se registró la presencia de un individuo de la especie Diodon holocanthus durante los tres censos realizados lo cual se atribuye a su nula irregularidad y complejidad morfológica, así como a su fuerte incidencia de oleaje. La presencia y ausencia de especies esta en función del tipo de ambiente y la complejidad e irregularidad que presente su sustrato, además de la disponibilidad de alimento y áreas de resguardo y protección tanto para individuos adultos, juveniles y reclutas que permitan mayores posibilidades de sobrevivencia contra posibles depredadores, factores ambientales y humanos.

1
ESTRUCTURA DE LA COMUNIDAD DE PECES EN LA BAHÍA CAC ALUTA, HUATULCO, OAXACA.
INTRODUCCIÓN
Este trabajo tiene como finalidad principal hacer una aportación en cuanto al conocimiento de la estructura de las comunidades de peces de la bahía Cacaluta, sobre lo cual no existen estudios, es por esta razón que es de vital importancia el conocer como y en que estado se encuentran las comunidades de peces en la bahía Cacaluta para así poder implementar eficientes estrategias de manejo a los mismos, ya que dependiendo del buen cuidado y aprovechamiento que demos a estos se verá beneficiado el desarrollo ecológico y económico del país. Sin duda los ecosistemas acuáticos más diversos y de alta producción fueron los que dieron origen al nacimiento de la vida que hoy día habita el planeta, su formación data de hace más de 200 millones de años y aunque no ocupan más del 0.17% de la superficie marina, constituyen el hábitat de un 25% de las especies marinas (Sorokin, 1995), pues un arrecife de unos cuantos kilómetros puede abrigar unas 400 especies de peces, sin contar las decenas de especies de corales, moluscos, poliquetos, crustáceos y equinodermos. En términos de phyla y clases, los arrecifes coralinos tienen por mucho la mayor diversidad por hectárea que cualquier ecosistema de los océanos y se piensa que la diversidad actual de peces podría ser incluso hasta 3 o 4 veces mayor a la que se conoce (Birkeland, 1997), y albergan a una gran diversidad de organismos zooplanctónicos, bentónicos y nectónicos. Con relación a las comunidades de peces en estos sistemas se pueden encontrar especies de peces que residen permanentemente, en tanto que el resto solo pasan una parte de su ciclo de vida en ellos, tal es el caso de juveniles y especies cosmopolitas que solo llegan a estos a alimentarse, las cuales también se pueden encontrar en las zonas rocosas y de algas cercanas. También hay especies de hábitos diurnos y nocturnos o que son activas las 24 horas del día, así como las que tienen ritmos de vida indefinidos (Helfman, 1978). Este tipo de estrategias para aprovechar el hábitat ha creado una gran diversidad de especies dentro de la comunidad (Hobson, 1974) lo cual le confiere una gran estabilidad; sin embargo la más mínima alteración del hábitat generaría una reacción en cadena que tendría como consecuencia el cambio radical de la comunidad de peces, con lo cual desaparecerían las especies presentes y entrarían nuevas, como lo describe Birkeland (1997). Estas estrategias de aprovechamiento del hábitat a su vez han favorecido la presencia de comunidades locales de peces aisladas las cuales prácticamente no se mezclan con otras, y que se asocian a un tipo de lugar y morfología estructural ya sea rocosa, arenosa o arrecifal como lo indican Sorokin (1995) y Hobson (1974) Los arrecifes de coral además de ser un importante refugio para un gran número de especies tiene un potencial pesquero anual entre 0.2 y 40 ton/K2/año (Sale, 1991)
Hay varios factores que tienen influencia en la distribución geográfica, abundancia y riqueza especifica de los peces arrecifales, dentro de los cuales se pueden citar la disponibilidad de espacio, refugio, alimento, características fisicoquímicas del ambiente, el grado de exposición de un lugar con respecto a corrientes y oleaje, la topografía del sustrato y las relaciones intraespecíficas e

2
interespecíficas, cuales generan que la estructura de las comunidades arrecifales sea el resultado de las interacciones entre dichos procesos a través del tiempo (Barrientos-Villalobos, 2000).
Por la importancia económica de los arrecifes coralinos millones de personas en el mundo dependen de ellos a través de la pesca artesanal y el turismo. Los arrecifes de coral proveen del 20 al 25% del total de peces capturados en los países en desarrollo (Sánchez y Guzmán, 1990). El conocimiento de la diversidad de especies y de las densidades de sus comunidades para los arrecifes coralinos hoy día se encuentra en desarrollo y es de vital importancia el preservar las condiciones naturales de estos sistemas para su adecuada utilización humana actual y a futuro. Bahía Cacaluta se localiza al suroeste de Santa Cruz. Tiene forma abierta parecida a un corazón y enfrente de ella se encuentra isla Cacaluta cuyas dimensiones no son muy grandes teniendo aproximadamente 16 ha de extensión. Esta bahía tiene un gran valor natural y ecológico, presenta clima sub-húmedo con una temperatura media anual de 28 grados centígrados y una vegetación integrada por selva baja caducifolia y manglares. En la franja marina existen zonas rocosas principalmente conformadas por escarpes de este tipo, zonas arenosas y comunidades coralinas. Así como suelos con texturas más finas donde el grado de arcillas es más variable como la vega de los arroyos Cacaluta y Cacalutilla. Esta bahía se encuentra integrada al Parque Nacional como área protegida ecológicamente, políticamente la parte terrestre pertenece al municipio de Santa María Huatulco y esta puesto a disposición del fideicomiso Fondo Nacional de Fomento al Turismo (FONATUR) (SEMARNAT, 2003).
ANTECEDENTES
Los estudios sobre comunidades de peces de arrecifes de coral, son todavía muy pocos, ya que inicialmente se realizaron en los años 50, 60 y 70; los cuales eran llevados a cabo junto con el desarrollo del buceo SCUBA el cual abrió una pauta para la exploración de los ambientes acuáticos y así poder realizar estudios ecológicos, por ejemplo los hechos por Sale (1980) y Sorokin (1995) Los estudios más diversos son los relacionados a la dinámica de peces en arrecifes y sus variaciones espacio-temporal. Como ejemplo están los estudios de patrones de diversidad y conjunto ictiofaunístico en arrecifes del Caribe Mexicano realizados por Caley (1993). En lo referente al impacto provocado por el esfuerzo pesquero en zonas arrecifales y por el asentamiento humano se han realizado muy pocos estudios como los de (Luna,1993). Diversas técnicas han sido aplicadas para el estudio de peces de arrecifes, tal es el caso de sistemas de vídeo y estéreo. En las Bahías de Huatulco existen pocos antecedentes ecológicos de evaluaciones de la comunidad de peces entre los que destacan los de Leyte-Morales (1995), Barrientos-Villalobos (2000), Ramos-Santiago (2004), Juárez-Hernández (2005), Martínez-Morgan (2004) y Ramírez-Ramírez (2004); ya que la mayor parte de los estudios que se han realizado van destinados a otros propósitos como lo son las asociaciones de organismos bentónicos al coral, así como también se han enfocado a aspectos biogeográficos y ecológicos en arrecifes del Pacífico

3
Mexicano. En lo referente al desarrollo turístico de Huatulco y su problemática ambiental varios autores han dado sus opiniones e identificado problemas de tipo sanitario, servicios urbanos y desechos sólidos para lo cual recomiendan hacer continuos monitoreos a fin de reconocer el daño en comunidades biológicas y terrestres. (SEMARNAT, 2003).
OBJETIVOS
Conocer la estructura y función de la comunidad de peces de la Bahía Cacaluta, Oaxaca. Objetivos particulares 1.- Determinar la composición, diversidad, dominancia, distribución y abundancia de la ictiofauna de la bahía Cacaluta, con relación a los diferentes subsistemas que presenta (coral, fondo arenoso y fondo rocoso). 2.- Conocer la variación espacio-temporal de la ictiofauna. 3.- Determinar las poblaciones dominantes. 4.- Determinar las asociaciones ictiofaunísticas.
METODOLOGÍA
Área de estudio Respecto a las bahías de Huatulco, estas se localizan en la Costa de Oaxaca, ubicadas en las estribaciones finales de la Sierra Madre del Sur. Se extienden por 35 Kilómetros de la costa Mexicana del océano pacifico y están delimitadas por la desembocadura de los ríos Coyula y Copalita. Poseen un clima cálido subhúmedo con una temperatura media anual de 28°C y lluvias en verano (CONABIO, 2003). La topografía de la zona es accidentada por montañas, valles y laderas, a través de donde se abren paso los ríos Coyula, San Agustín y Copalita. Son 9 las bahías que se ubican en la región de Huatulco, y estas son: Bahía Cacaluta, Bahía Santa Cruz, Bahía la Entrega, Bahía Tangolunda, Bahía Organo y Maguey, Bahía Chachacual, Bahía Chahué, Bahía de San Agustín y Bahía Conejos (CONABIO, 2003). Bahía Cacaluta se localiza al suroeste de la playa Principal de la bahía de Santa Cruz, sus coordenadas son 15°43.3´LN 96°09.4´ LW, exactamente se ubica entre la punta oeste, cuyas coordenadas son 15°43´09”LN 96°09´59”LW, y la punta este, cuyas coordenadas son 15°43´20.5”L N 96°09´11”LW; (Figura 1). Esta bahía tiene un gran valor natural y ecológico. Presenta clima sub-húmedo con una temperatura media anual de 28 grados centígrados y una vegetación integrada por selva baja caducifolia y manglares. La bahía Cacaluta tiene como principal característica su forma abierta que semeja un corazón. Frente a ella se encuentra isla Cacaluta la cual la protege totalmente de los

4
vientos dominantes. Esta bahía se encuentra integrada al Parque Nacional por lo cual es un área protegida ecológicamente, así como posee uno de los aguajes (de agua dulce) de suma importancia para el entorno natural del mismo parque (CONABIO, 2003) (SEMARNAT, 2003). Bahía Cacaluta esta conformada por dos playas: Cacaluta y Arroyo: • Playa Cacaluta: Se localiza en el extremo oriente de la bahía. La arena es
gruesa y blanca con pendientes planas y convexas, dentro de un área de 1 km. de longitud por 50 metros de ancho aproximadamente. El agua es templada y tiene tonalidades verdes y azules, es profunda y de oleajes fuertes. En su entorno existe vegetación tropical y formaciones rocosas. (CONABIO, 2003) (SEMARNAT, 2003)
• Playa Arroyo: Está ubicada al poniente de la Bahía de Cacaluta en la misma
dirección de la Bahía de Santa Cruz Huatulco. Es de pequeñas dimensiones; mide 70 metros de largo y 15 metros de ancho. No es muy concurrida, debido a que en su arena blanca se haya demasiada grava y restos de coral blanco. Su pendiente es moderada y sus oleajes tranquilos. El agua es cristalina y adquiere tonalidades verdes y azules, debido a la vegetación circundante y a la profundidad que poco a poco varía. (CONABIO, 2003) (SEMARNAT, 2003)
Figura 1. Bahías de Huatulco. Se proyecta la bahía Cacaluta donde se indican los puntos de muestreo.
BAHÍA SANAGUSTIN
BAHÍA RISCALILLO
BAHÍAJICARAL
BAHÍACHACHACUAL
BAHÍACACALUTA
BAHÍAMAGUEY
BAHÍAORGANO
BAHÍA SANTA CRUZ
BAHÍACHAHUÉ
BAHÍATANGOLUNDA
BAHÍACONEJOS
Playa Cacaluta
Pla ya Arroyo
Isla Cacaluta
T1
T2
T3
T4
T5
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
OAXACA
BAHÍAS DE HUATULCO
AMBIENTE ROCOSO
AMBIENTE DEARRECIFE DE CORAL

5
La fauna del lugar está formada por aves acuáticas: (gaviotas, garzas, zopilote, tijerillas, patos buzo), peces (marlin, pez vela, dorado, atún, pez gallo, etc.). Con menor frecuencia y entre la selva baja, se encuentran aves silvestres como chachalacas, andapiés, codornices, palomas, tórtola, existen también algunas especies de mamíferos: tejón, armadillo, zorrillo, mapache, puerco espín, conejo, jabalí y venado. Así como en cuanto a reptiles encontramos iguanas y tortugas (CONABIO, 2003) (SEMARNAT, 2003) Los ambientes que predominan en esta bahía y en los cuales fueron ubicados los transectos, son tres, el primero correspondiente al ambiente rocoso, el segundo al ambiente arenoso y el tercero al ambiente de arrecife de coral. Ambiente rocoso Se presenta en el lado este de playa Arroyo, presenta un relieve complejo el cual no es muy amplio en área de extensión, su pendiente es pronunciada desde el inicio del acantilado hasta el fondo marino con una profundidad aproximada de 7 a 11 metros, apreciandose la presencia de conglomerados de roca ígnea y metamórfica de diversos tamaños yendo desde guijarros y rocas hasta muy grandes peñascos los cuales presentan grietas, cavidades y fracturas tanto grandes como pequeñas; la morfología que presentan estas masas de roca es variada, la cual va desde formas redondeadas, semiredondeadas y cúspides desde la superficie hasta el fondo, siendo las formas redondeadas y semiredondeadas las más visibles debido a la fuerte influencia que ejercen el oleaje el viento y las corrientes en esta zona. La región bentónica se encuentra conformada tanto por roca como por arena haciendo que este sistema sea más complejo. El oleaje es fuerte debido a que la bahía es abierta y a pesar de tener la presencia de la isla Cacaluta la cual aminora los efectos de éste. Ambiente arenoso Presenta una muy amplia extensión litoral con presencia de playa, cuya pendiente es poco pronunciada con una profundidad de 3 a 6 metros. No se encuentra en colindancia con los otros ambientes debido a su basta extensión y a la posición en la que se encuentra. Presenta algunos fragmentos de coral muerto arrastrados por las corrientes e incluso por la acción del oleaje, materia orgánica e inorgánica en suspensión en la columna de agua, sin evidencia de la presencia de basura y otros tipos de contaminantes de origen humano. Con respecto al oleaje este es intenso, de manera semejante al ambiente anterior. Ambiente de arrecife de coral Se encuentra conformado por corales hermatípicos. Se ubica en la parte interna de la isla hacia la bahía, con una profundidad de 1.5 a 11.6 metros; el cual posee a 6 de las especies de corales de la región siendo la gran mayoría pertenecientes al género Pocillopora (Glynn & Leyte, 1997). El arrecife en su gran mayoría se encuentra en buenas condiciones, siendo pocos los fragmentos de coral muerto y en vias de rompimiento aunque ya existe el proceso de blanqueamiento de coral que es originado por la resuspensión del sedimento arenoso por efecto del oleaje, el cual es acarreado y depositado sobre los pólipos de coral propiciando la muerte de los mismos.. Dada la morfología irregular que posee este ambiente, existe la formación de gran

6
cantidad de oquedades, ranuras, fisuras y espacios de diversos tamaños que ofrecen zonas de refugio y alimentación para organismos juveniles y adultos. Se realizaron tres visitas de muestreo repartidas a lo largo de seis meses con la finalidad de obtener la información necesaria correspondiente a las diferentes épocas climáticas presentes en la región, lluvias y secas, con el objeto de observar la variabilidad espacio temporal de la comunidad de peces en estos dos periodos. Actividades de campo
Se hicieron tres salidas a campo durante el año 2004 correspondientes a dos épocas climáticas diferentes (lluvias y secas), con el fin de obtener los datos necesarios para determinar la estructura de la comunidad de peces, diversidad, distribución y abundancia en la Bahía Cacaluta. En cada salida, una vez estando en el lugar en el cual se realizaron los censos visuales, lo primero que se realizó fue la ubicación de cada uno de los transectos a través de las coordenadas geográficas obtenidas en la salida prospectiva. Ya ubicados cada uno de los transectos se procedió al tendido de los mismos, y se comenzó a realizar el censo visual; todo lo anterior se realizó para cada uno de los transectos en las tres salidas de campo. Los censos visuales aún cuando presentaron ciertos inconvenientes y desventajas como por ejemplo una identificación errónea de las especies y una estimación de las densidades imprecisa (Sale, 1980; Brock, 1982; Sale y Sharp, 1983; Buckley y Hueckel, 1989; Greene y Alevizon, 1989; Elorduy-Garay y Jímenez-Gutiérrez, 2000), fueron menos nocivos que el uso de ictiocidas y explosivos. Con la ayuda de equipo de buceo libre o snorkel, y sobre una hoja de campo de papel polypap, con los datos pertinentes se registraron las especies, número de individuos por especie, y observaciones generales, considerando lo propuesto por Sale y Douglas (1981). Se realizaron transectos que consisten en cuerdas de polipropileno de 5mm de diámetro y diez metros de largo, ambos del mismo tamaño para evitar errores de comparación, graduados cada metro para obtener la profundidad. En un extremo de la cuerda poseía un plomo para darle estabilidad, y en el otro una boya de unicel con la señalización de buceo. Los transectos se ubicaron en los diferentes ambientes del sistema y las coordenadas de cada uno se tomaron con la ayuda de un geoposicionador satelital (GPS) (Tabla 1), evitando confusión en la colocación de los transectos durante cada una de las diferentes visitas realizadas. Tabla 1. Coordenadas geográficas para cada uno de los transectos en la Bahía Cacaluta.
Transecto LN LW
T1 15°43’14’’ 96°09’59’’
T2 15°43’07’’ 96°09’49’’
T3 15°43’’09’’ 96°09’46’’
T4 15°43’’11’’ 96°09’44’’
T5 15°43’’18’’ 96°09’39’’

7
Figura 2. Bahía Cacaluta. Ubicación de ambientes y transectos, en los cuales se realizaron los censos visuales. La técnica de muestreo que se empleo fue la misma para cada uno de los diferentes ambientes. La ubicación de los transectos se muestra en la figura 2. Muestreo de la ictiofauna Posterior a la colocación del transecto de 10 m de longitud, sobre el transecto se consideró un campo de visión de 3 m de distancia a cada lado. El recorrido de los censos visuales se hizo en 5 minutos y se realizó otro recorrido hacia el punto de partida para buscar entre los recovecos especies crípticas y registrarlas. Todos los muestreos se llevaron a cabo por las mismas personas para reducir errores de identificación (Brock, 1982; Buckley y Hueckel, 1989; Sale, 1980). Actividades de laboratorio Al regreso de cada una de las salidas a campo, se procedió en el laboratorio a procesar y analizar los datos obtenidos de cada uno de los transectos pertenecientes a los censos visuales realizados, así como también a la redacción del reporte de servicio social. Análisis general de datos Las especies se determinaron taxonómicamente, y se elaboró un listado sistemático para la determinación de la composición ictiofaunística del área de estudio con base en Nelson (1994).
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
T1
T2
T3
T4
T5
96°09’59’’LW 96°09’11’’LW 15°43’20.5’’LN
15°43’09’’LN

8
La abundancia se calculó en densidad de acuerdo a la siguiente expresión: N D = ------- A donde: D = número de individuos por m2 N = número total de individuos y A = área muestreada. A partir de los datos obtenidos de las comunidades estudiadas se elaboraron matrices en excel en las cuales se realizaron los análisis conducentes a la interpretación de los datos y se expresaron en forma de gráficos. Posteriormente se realizó un análisis de la diversidad y similitud utilizando los siguientes índices. Índices de diversidad Para el análisis de diversidad, además de la riqueza de especies, se utilizaron: índice de riqueza de especies (D) de Margalef (1958) con la expresión S - 1 D = ----------- log(N) Este índice se basa en la relación entre el número total de especies en una comunidad S y el número total de individuos observados N. índice de Shannon y Wiener (1963) de acuerdo a la expresión: N log2 N - ∑ ni log2 ni
H´n = ------------------------------ N donde N = número total de individuos observados y ni = número de individuos de la especie i. Diversidad máxima (H´max). La regularidad de la distribución del total de individuos en el total de especies se puede expresar entonces como la relación de la diversidad según los datos observados y el índice de máxima diversidad.
H’max = 3.3219 (log10 N – 1/N ∑ ni log10 ni)
Donde: N= número total de individuos

9
ni = número de individuos de la especie i. 3.3219 = Factor de conversión del log10 a ln. Índice de dominancia de Simpson (Zar,1996). Este índice se empleo para estimar la dominancia, sin embargo, a diferencia del índice de Margalef en éste si se toma en cuenta la abundancia relativa de cada especie. Su rango de valores va de 0 a 1, siendo aquellos valores más cercanos a 0 los que presentan menor dominancia y entre más se acerquen a 1 presentaran mayor dominancia.
S = N (N-1) / ∑ ni (ni – 1) Donde: N = Total de individuos de todas las especies. ni = Número de individuos de la especie i. índice de equitatividad (J´) de Pielou (1975, 1977), con: H´ H´ J´= ---------- = ----------- log(S) H´max Que expresa H´ corresponde a la diversidad obtenida en el muestreo y H´max es relativa al valor máximo que puede obtener cuando todas las especies en las muestras están perfectamente uniformes con un individuo por especie. Similitud de la comunidad y clasificación Se realizó un análisis de los patrones de distribución y de afinidad de los grupos bióticos estudiados, para lo cual se utilizó el análisis de factores a través del método de componentes principales y el análisis de clasificación por conglomerados por medio del método de Ward (1963) considerando la abundancia de las especies (Pielou, 1984; Ludwing y Reynolds, 1988; Magurran, 1998; Gauch, 1989), para lo cual se utilizó el paquete estadístico “STATISTICA” para Windows. El método de clasificación propuesto por Ward (1963) consiste en la agrupación inicial de N grupos, los que contiene un solo individuo; en cada ciclo de agrupación N-1, se unen los grupos más similares entre sí, a la vez que se calcula una matriz de similaridad que produce una agrupación jerárquica por medio de una transformación combinatoria de los coeficientes de similaridad; en la transformación de grupos P y Q son fusionados. La similitud S(R, P+Q) entre cualquier grupo R y el nuevo grupo (P+Q) es obtenida de la transformación: S(R, P+Q) = AP * S(R, P) + AQ * S(R, Q) + B * S(P, Q) donde:

10
AP = (NR + NP)/(NR + NP + NQ) AQ = (NR + NQ)/(NR * NP * NQ) B = NR / (NR + NP + NQ) NR, NP y NQ son tamaños de los grupos.
ACTIVIDADES REALIZADAS Primero.- Se realizó una salida de campo prospectiva en el mes de noviembre del año 2003 con el fin de conocer las condiciones morfológicas y biológicas de la Bahía Cacaluta; como lo son el tipo de coral, las condiciones que este presenta, presencia de zona rocosa y zona arenosa así como para hacer el reconocimiento de la zona de muestreo y establecer los transectos determinando su posición geográfica a la vez de poner en practica la técnica de censos visuales. Segundo.- Se llevo a cabo la primera salida de campo a la Bahía Cacaluta en el mes de febrero del año 2004 cual corresponde a la temporada de secas; se realizaron los censos visuales correspondientes a cada tipo de ambiente. Durante este mes y hasta abril se procesaron y analizaron los datos obtenidos en esta salida. Tercero.- Se realizó la segunda salida de campo en el mes de abril del 2004 correspondiente a la época de secas. Los datos obtenidos se analizaron en mayo, junio y principios del mes de julio. Cuarto.- Se efectuó la última salida de campo, considerada en el servicio social, a principios del mes de julio la cual corresponde a la temporada de lluvias. En enero, febrero, marzo y parte del mes de abril de 2005 se analizaron los datos de esta salida y de las 2 salidas de campo anteriores para proceder con la elaboración del reporte final de servicio social.
OBJETIVOS Y METAS ALCANZADOS
En lo que respecta a las tres salidas de campo, incluyendo la salida prospectiva, fue posible realizar todos los censos visuales programados para la Bahía Cacaluta, de acuerdo a los objetivos establecidos, lo que permitió conocer la composición, distribución, abundancia, diversidad, dominancia y equidad de la ictiofauna característica de esta bahía. De igual manera, se conocieron las fluctuaciones en la composición ictiofaunística durante las dos épocas climáticas durante las que se llevó a cabo el estudio, conociendo de esta forma la variación espacio-temporal presente en el sistema. La información que se logró obtener contribuirá, para apoyar las estrategias de manejo del Parque Nacional Huatulco, así como ayudará a establecer un mayor control de las actividades turísticas, deportivas, pesqueras y antropogénicas presentes en el lugar.

11
Resultados y Discusión.
Caracterización de la comunidad Composición taxonómica Se logro censar un total de 1,443 organismos que son correspondientes a 29 especies, 27 géneros, 18 familias y 2 subfamilias; los cuales se muestran a continuación en la siguiente lista sistemática de acuerdo a Nelson (1994).
Lista sistemática de peces en la bahía Cacaluta
Phylum Chordata Subphylum Vertebrata Superclase Gnatostomata Clase Actinopterygii Subclase Neopterygii División Teleostei Subdivisión Elopomorpha Orden Perciformes Familia Pomacentridae Género Abudefduf Abudefduf troschelii (Gill, 1862) Orden Tetraodontiformes Familia Tetraodontidae Género Arothron Arothron meleagris (Lacepede, 1798) Orden Tetraodontiformes Familia Balistidae Género Balistes Balistes polylepis Steindachner, 1876 Orden Perciformes Familia Labridae Subfamilia Bodianidae Género Bodianus Bodianus diplotaenia (Gill, 1862) Orden Perciformes Suborden Percoidei Familia Carangidae Género Caranx Caranx caballus Günther, 1868 Orden Perciformes Suborden Percoidei Familia Cirrhitidae Género Cirrhitus Cirrhitus rivulatus Valenciennes, 1846 Orden Perciformes Familia Chaetodontidae Género Chaetodon Chaetodon humeralis Günther, 1860

12
Orden Perciformes Familia Pomacentridae Género Chromis Chromis atrilobata Gill, 1862 Orden Tetraodontiformes Familia Diodontidae Género Diodon Diodon holocanthus Linnaeus, 1758 Orden Perciformes Suborden Percoidei Familia Serranidae Género Epinephelus Epinephelus labriformis (Jenyns, 1840) Orden Perciformes Familia Haemulidae Género Haemulon Haemulon sexfasciatum Gill, 1862 Orden Perciformes Familia Labridae Género Halichoeres Halichoeres notospilus (Günther, 1864) Orden Beloniformes Suborden Belonoidei Familia Hemiramphidae Género Hemirhamphus Hemirhamphus saltator Gilbert & Starks, 1904 Orden Perciformes Familia Pomacanthidae Género Holacanthus Holacanthus passer Valenciennes, 1846 Orden Perciformes Familia Chaetodontidae Género Jhonrandallia Jhonrandallia nigrirostris (Gill, 1862) Orden Perciformes Familia Kyphosidae Género Kyphosus Kyphosus elegans (Peters, 1869) Orden Perciformes Familia Lutjanidae Género Lutjanus Lutjanus argentiventris (Peters, 1869) Orden Perciformes Familia Pomacentridae Género Microspathodon Microspathodon bairdii (Gill, 1862) Orden Perciformes Familia Pomacentridae Género Microspathodon Microspathodon dorsalis (Gill, 1862)

13
Orden Perciformes Familia Mugilidae Género Mugil Mugil curema Valenciennes, 1836 Orden Perciformes Familia Labridae Género Novaculichthys Novaculichthys taeniourus (Lacepede, 1801) Orden Perciformes Familia Blenniidae Género Ophioblennius Ophioblennius steindachneri Jordan y Evermann, 1898 Orden Tetraodontiformes Familia Ostraciidae Subfamilia Ostraciinae Género Ostracion Ostracion meleagris Shaw, 1796 Orden Perciformes Familia Cirrhitidae Género Oxycirrhites Oxycirrhites typus Bleeker, 1857 Orden Perciformes Familia Acanthuridae Género Prionurus Prionurus punctatus Gill, 1862 Orden Perciformes Familia Pomacentridae Género Stegastes Stegastes acapulcoensis (Fowler, 1944) Orden Perciformes Familia Pomacentridae Género Stegastes Stegastes flavilatus (Gill, 1862) Orden Tetraodontiformes Familia Balistidae Género Sufflamen Sufflamen verres (Gilbert & Starks, 1904) Orden Perciformes Familia Labridae Género Thalassoma Thalassoma lucasanum (Gill, 1862) No se descarta la probabilidad de que algunas otras especies no hayan sido censadas, esto principalmente debido a que en la técnica de censo visual los resultados dependen de la capacidad y del campo visual del observador y a pesar de que no es la más exacta en comparación con otras técnicas, se utilizó por ser la más benéfica para el sistema, pues el daño y estrés que se genera es mínimo (Brock, 1954).

14
Se realizaron 3 matrices de datos correspondientes una por cada época de censo con la finalidad de determinar los índices de diversidad, dominancia, equidad, así como también la densidad, el número total de individuos y la riqueza de especies que se encuentra por transecto.
Tabla 2. Se presenta el número de individuos por especie y los valores de los índices de diversidad, dominancia, equidad y densidad por transecto obtenidos en febrero de 2004. Los valores correspondientes al transecto número 5 no se presentan debido a que no se registraron organismos en el fondo arenoso. Densidad. En el caso del censo de febrero (Tabla 2, Figuras 3a y 4a) la mayor abundancia o densidad de organismos se presentó en los transectos correspondiente a la zona arrecifal (T2, T4 y T3) enlistados en orden decreciente de acuerdo a su valor de densidad. Para el caso de la región rocosa el valor de densidad es menor a los anteriores. Presenta una riqueza de especies relativamente alta a pesar de que su abundancia no es muy alta. Esto es posible a que en esta zona se da una transición entre el ambiente arrecifal y rocoso lo que permite la presencia de muchas especies en busca de alimento, principalmente especies no territoriales (DeMartini y Roberts, 1982). La zona coralina tiene mayor abundancia debido a que la disponibilidad de hábitats es mayor por la irregularidad del substrato, así como por la protección que brinda la isla contra el oleaje, Esto determina que una mayor cantidad de especies territoriales como Microspathodon dorsalis y Stegastes acapulcoensis, logren establecerse. Este mismo patrón se observa en Thalassoma lucasanum cuyo número se ve enormemente incrementado en la zona arrecifal, especie que
Especie T1 T2 T3 T4Abudefduf troschelii 17 20Arothron meleagris 1Bodianus diplotaenia 1 1 1 3Chromis atrilobata 50 30Diodon holocanthus 1 3Epinephelus labriformis 1Haemulon sexfasciatum 2Halichoeres notospilus 1 2Hemirhamphus saltator 5 1Microspathodon bairdii 1Microspathodon dorsalis 10 9.5 15.5 12.5Mugil curema 30Ophioblennius steindachneri 1Ostracion meleagris 1Prionurus punctatus 7Stegastes acapulcoensis 3.5 8.5 18 35Stegastes flavilatus 15Thalassoma lucasanum 4.5 150 125 115NUMERO TOTAL (S) 93 254 162.5 192.5RIQUEZA DE ESPECIES 10 9 6 10ÍNDICE DE MARGALEF 1.376 1.001 0.681 1.186SIMPSON 0.327 0.382 0.611 0.047H'max 3.322 3.170 2.585 3.322H'n 2.198 1.985 1.135 1.832J' 0.662 0.626 0.439 0.551Densidad 1.550 4.233 2.708 3.208

15
aporta alta abundancia en la zona coralina. En la zona arenosa no se encontraron organismos lo cual podría estar muy relacionado con el hecho de que la bahía de Cacaluta presenta una fuerte incidencia del oleaje y corrientes las cuales llegan con toda intensidad por la ausencia de alguna barrera física que las aminore (McGehee, 1994). Los valores de la riqueza de especies (Tabla 2, Figuras 3b y 4b) son muy similares para los dos ambientes (rocoso y coralino) pues como anteriormente se mencionó, la zona rocosa esta relativamente en colindancia con la región arrecifal, lo que la haría una zona de transición de organismos que a pesar de no ser muchos si son muy variados obteniendo con esto una cantidad similar de especies a la zona coralina. Otra situación es que esta región no se encuentra protegida de la incidencia del oleaje por la isla Cacaluta, en tanto que la zona arrecifal si lo cual hace que ambas regiones sean muy diferentes. No obstante aún cuando la irregularidad del arrecife no es mayor que la de la zona rocosa es por la presencia de la isla que este les brinda resguardo, protección y alimento a las especies por eso aquí la abundancia de individuos es mucho mayor. En tanto que en el transecto 5 no se dan estas mismas características, pues este transecto se encuentra ubicado en una región donde el oleaje es más fuerte por no estar protegido por una barrera física y por ser un fondo arenoso no hay disponibilidad de alimento ni resguardo para la ictiofauna por lo que no se observó ninguna especie. Diversidad. Los resultados obtenidos mediante los índices de diversidad (Margalef y Shannon) son muy semejantes, en ambos la mayor diversidad se encuentra en el transecto 1 correspondiente a la zona rocosa, esto porque su riqueza de especies es alta a pesar de no tener un alto número de individuos característica en la que se basa el índice de Margalef (Figuras 3e y 4e), en tanto que Shannon (Figuras 3g y 4g) toma en cuenta también la abundancia relativa de las especies, es decir, que tan heterogénea es la distribución, de aquí que esto hace que el transecto rocoso sea el más diverso. (DeMartini y Roberts, 1982), determinado por la baja presencia de especies territoriales. En cuanto a los transectos 2 y 4 correspondiente a zona arrecifal hay una diferencia entre los dos índices, Margalef indica que la mayor diversidad la posee el transecto 4, en tanto que Shannon indica que la posee el transecto 2, lo que se atribuye a que el primero se basa en la riqueza de especies, en tanto que el segundo evalúa también la abundancia relativa, o en otras palabras, a pesar de que el transecto 4 tiene más especies, una de ellas ocupa más del 60% la especie Thalassoma lucasanum cual no es territorial lo que hace que sea menos equitativo dicho transecto con relación al transecto 2 que a pesar de que también presenta a dicha especie y que posee una menor riqueza de especies, tiene una mayor equitatividad debido a la cantidad de hábitats disponibles para las especies territoriales por la baja irregularidad del arrecife y a la intensidad del oleaje cual es más fuerte en el transecto 4. Mientras que el transecto 3 posee una baja diversidad por presentar pocas especies y poca equitatividad ya que hay muchos individuos los cuales corresponden a solo dos especies las cuales dominan la zona por las condiciones del lugar y como consecuencia impiden la colonización de otras especies (Thomson et al., 1979).

16
Figura 3. Abundancia y diversidad de la ictiofauna por transecto en febrero de 2004.
Riqueza de especies
(b) Número de especies
T1
T2
T3
T4
T5
0 2 4 6 8 10 12
Número de individuos
(c) Individuos
T1
T2
T3
T4
T5
0 50 100 150 200 250 300
Simpson
(d) Dominancia S
T1
T2
T3
T4
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Indice de Margalef
(e) Diversidad D
T1
T2
T3
T4
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6
Diversidad máxima
(f) H´max
T1
T2
T3
T4
0 0.5 1 1.5 2 2.5 3 3.5
Indice de Shannon
(g) Diversidad H´n
T1
T2
T3
T4
0 0.5 1 1.5 2 2.5
Pielou
(h) Equidad J
T1
T2
T3
T4
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Densidad
(a) Número de individuos
T1
T2
T3
T4
T5
0 1 2 3 4 5

17
Figura 4. Abundancia y diversidad de la ictiofauna en febrero de 2004.
(c)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
93
254163
193
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Número de individuos
(a)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
1.5
4.232.7
3.2
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Densidad g/m2
(b)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
10
96
10
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Riqueza de especies
(d)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.32
0.380.61
0.04
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Dominancia (S)
(e)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
1.37
1.000.68
1.18
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Margalef
(f)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
3.32
3.162.58
3.32
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad Máxima (H’max)
(g)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
2.19
1.981.13
1.83
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Shannon
(h)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.66
0.620.43
0.55
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Equidad (J’)

18
Con respecto a la equidad de los transectos (Tabla 2, Figuras 3h y 4h), los valores más altos los presenta el transecto 1 y 2 ya que la gran mayoría de las especies que los conforman presentan valores no muy separados en el número total de individuos de unas con otras, en donde como en el caso del T1 la especie que más se aleja del resto es Chromis atrilobata ya que su número de individuos es el más alto para este transecto, lo cual se atribuye a que en esta zona la influencia del oleaje es mas fuerte lo que limita el establecimiento de muchas especies en el lugar (McGehee, 1994), en tanto que en el caso del transecto 2 de la zona de coral hay una equidad alta debido principalmente al incremento en el número de individuos de las especies en general a causa de la protección y mayor disponibilidad de hábitats por la irregularidad del substrato así como por la protección que brinda la isla contra el oleaje lo que favorece el establecimiento de la ictiofauna en el lugar, también muy posiblemente porque la relativa cercanía entre los dos transectos permite la transición de las especies entre ambos (DeMartini y Roberts, 1982). Por su parte el valor más bajo de equidad lo presenta el transecto 3 de la zona de coral en el cual hay la presencia de pocas especies y los valores en su número de individuos son muy contrastantes ya que algunas tienen muchos representantes en tanto que otras solo presentan un individuo, esto es debido a que este transecto es el más protegido por la isla Cacaluta contra el fuerte oleaje por lo que se favorece el establecimiento de las especies territoriales en lugar, las cuales se encargan de alejar e impedir el establecimiento de otras, las cuales muy posiblemente tienen un menor grado de dominancia porque quizá su frecuencia reproductiva y taza de fertilidad son mucho más bajas. Sin embargo, no es una equidad muy marcada ya que el resto de los transectos también presentan a muchas de estas especies tales como Bodianus diplotaenia, Microspathodon dorsalis, Stegastes acapulcoensis y Thalassoma lucasanum; las cuales por ser especies que se establecen en un hábitat y otras que se desplazan de un lugar a otro en busca de alimento permite que se encuentran en ambos ambientes (rocoso y arrecifal) dependiendo en que lugar las características son las más favorables para su subsistencia como las preferencias por el tipo de ambiente derivadas de los factores físicos, biológicos y morfológicos de estos (Alevizon et al., 1985). En cuanto a lo que se refiere a la dominancia (Tabla 2, Figuras 3d y 4d) tenemos que el valor más alto de esta se encuentra en el transecto 3, cual corresponde a la zona de coral y el cual esta protegido por la isla de forma más favorable, ya que este se ubica a la mitad de ésta, por lo que tiene como consecuencia que muchas de las especies territoriales se ubiquen en esta zona porque la acción del oleaje y corrientes no es tan fuerte de ahí que en éste transecto se encuentran muchos individuos de las especies dominantes Stegastes acapulcoensis, Microspathodon dorsalis y Thalassoma lucasanum, Siendo aquí también donde se encuentra la mayor cantidad de individuos de Microspathodon de todos los transectos en las oquedades y filones rocosos que se presentan en algunas partes de ésta zona coralina ya que este tipo de microambientes favorecen su alimentación debido a la presencia de algas (también hay la presencia de erizos) lo que les permite seleccionar a estos sitios como zonas de reproducción en donde los machos giran en círculos para defender sus territorios y atraer a las hembras (Williams, 1980), En el caso de

19
los individuos de Stegastes su cantidad es mucho menor ya que la zona coralina no es muy extensa y dado a sus necesidades de hábitat de esta especie es por lo que no se presentaron en muy alta cantidad sucediendole lo mismo a otros peces. Por su parte los individuos de Thalassoma son muy abundantes ya que forman grandes agrupaciones que se desplazan a través de la columna de agua alimentándose de plancton, algas y corales sin interactuar con las otras dos especies, de ahí que estos se encuentran prácticamente en todos los ambientes ya que no dependen de un territorio especifico (Thomson et al., 1979; Warner y Hoffman, 1980). En el resto de los transectos los valores de dominancia fueron menores. Riqueza de especies por tipo de ambiente (Figura 5). El ambiente que más riqueza de especies presenta es el ambiente rocoso, aún cuando la extensión de este ambiente no es muy amplia muchas especies fueron registradas aquí ya que estas transitan a través de este en busca de alimento y protección, sin embargo debido a la fuerte incidencia de oleaje que presenta es por lo que hay pocas especies territoriales, siendo una de las más abundantes Microspathodon dorsalis cual es característica de este tipo de ambiente por su alimentación a base de algas con lo que mantiene ocupados los hábitats disponibles de esta zona. El resto de las especies son organismos que conforman cardúmenes como Chromis atrilobata el cual se alimenta de zooplancton desplazándose a través de la columna de agua en busca de este alimento, Hemiramphus saltator que es una especie oceánica que se alimenta de pequeños peces cuales se encuentran es estas zonas y algunas otras especies de hábitos solitarios como Bodianus diplotaenia, Diodon holocanthus y Halichoeres notospilus que se alimentan de pequeños crustáceos, moluscos y equinodermos que se encuentran asociados principalmente al substrato rocoso y algas (Froese y Pauly, 2004). Por su parte la zona arrecifal a pesar de tener una mayor abundancia presenta una menor diversidad ya que muchos de los organismos son especies territoriales que ocupan los hábitats disponibles y no permiten la colonización de otras especies, así como la presencia de algas, crustáceos, moluscos y equinodermos es menor por lo que la ictiofauna que se alimenta de estos organismos se desplaza a otras áreas en busca de estos. Finalmente la zona arenosa no muestra la presencia de ictiofauna ya que esta es un área muy extensa con una fuerte incidencia de oleaje y totalmente desierta por lo que el único alimento disponible es el plancton suspendido en la columna de agua.

20
Figura 5. Riqueza de especies por ambiente en febrero de 2004 correspondiente a la época de secas.
0
2
4
6
8
10R
ique
za
Roca Arrecife decoral
Arena
Tipo de ambiente
Riqueza de especies por ambiente
Roca
Arrecife de coral
Arena
21
Abril 2004.
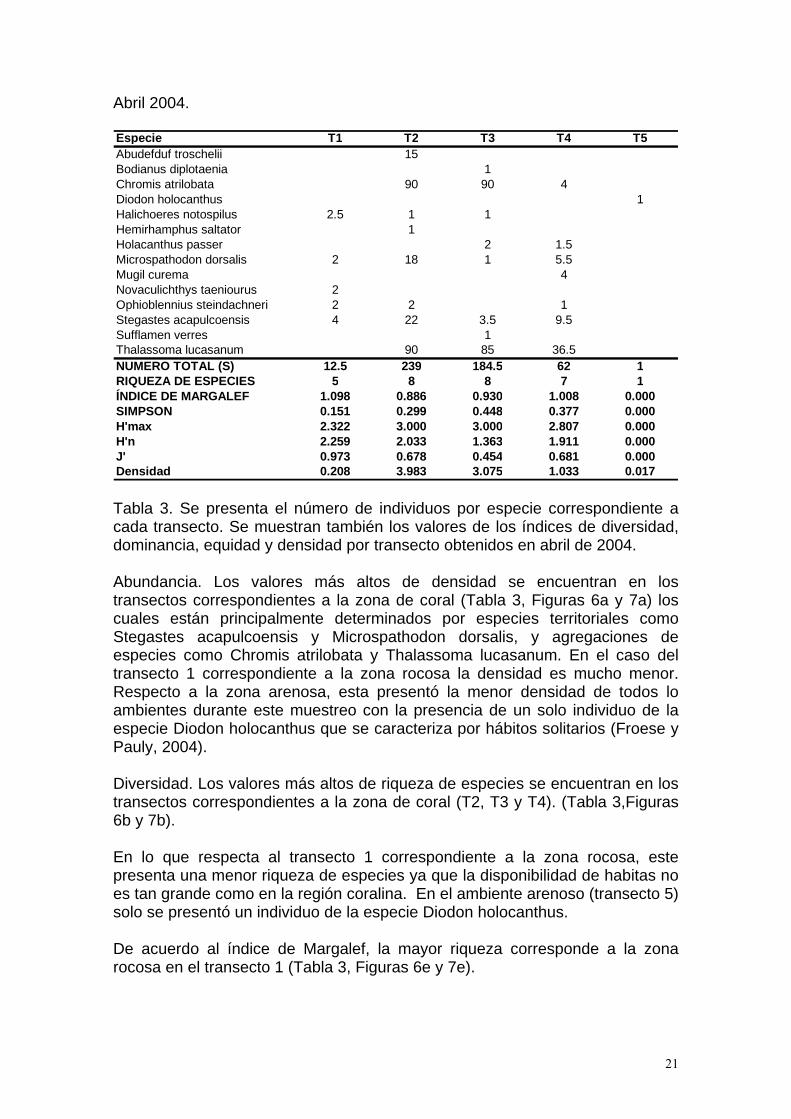
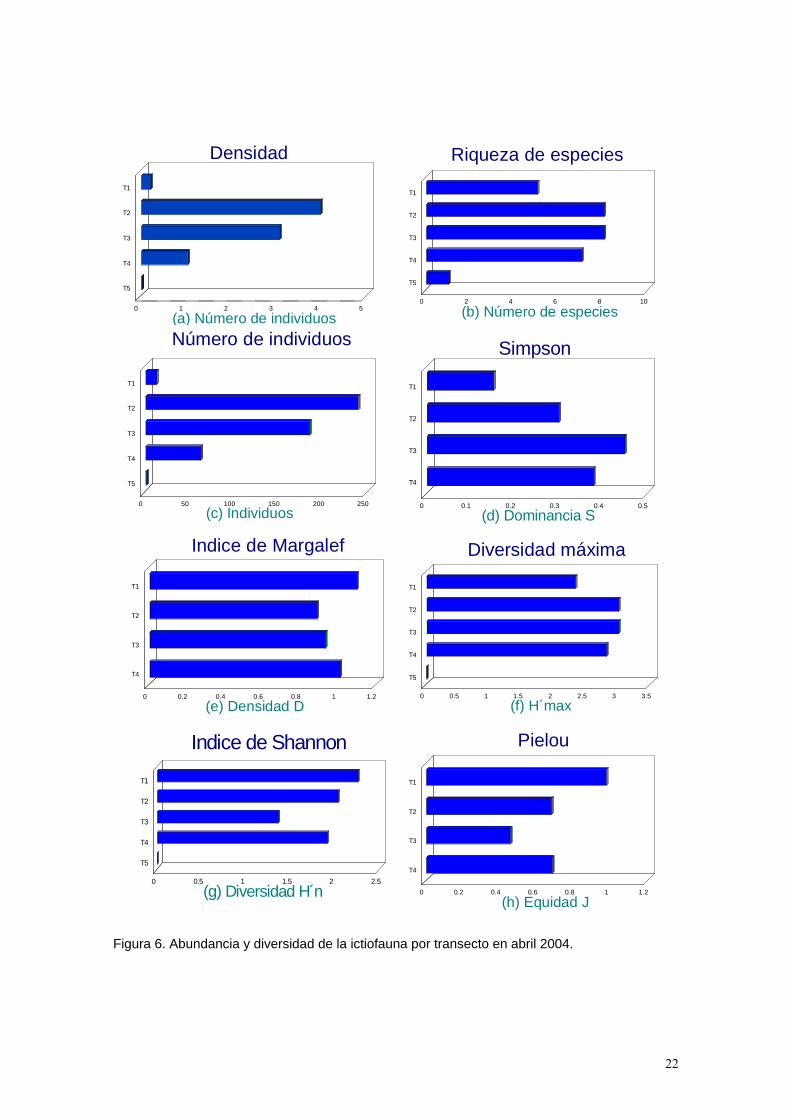
Tabla 3. Se presenta el número de individuos por especie correspondiente a cada transecto. Se muestran también los valores de los índices de diversidad, dominancia, equidad y densidad por transecto obtenidos en abril de 2004. Abundancia. Los valores más altos de densidad se encuentran en los transectos correspondientes a la zona de coral (Tabla 3, Figuras 6a y 7a) los cuales están principalmente determinados por especies territoriales como Stegastes acapulcoensis y Microspathodon dorsalis, y agregaciones de especies como Chromis atrilobata y Thalassoma lucasanum. En el caso del transecto 1 correspondiente a la zona rocosa la densidad es mucho menor. Respecto a la zona arenosa, esta presentó la menor densidad de todos lo ambientes durante este muestreo con la presencia de un solo individuo de la especie Diodon holocanthus que se caracteriza por hábitos solitarios (Froese y Pauly, 2004). Diversidad. Los valores más altos de riqueza de especies se encuentran en los transectos correspondientes a la zona de coral (T2, T3 y T4). (Tabla 3,Figuras 6b y 7b). En lo que respecta al transecto 1 correspondiente a la zona rocosa, este presenta una menor riqueza de especies ya que la disponibilidad de habitas no es tan grande como en la región coralina. En el ambiente arenoso (transecto 5) solo se presentó un individuo de la especie Diodon holocanthus. De acuerdo al índice de Margalef, la mayor riqueza corresponde a la zona rocosa en el transecto 1 (Tabla 3, Figuras 6e y 7e).
Especie T1 T2 T3 T4 T5Abudefduf troschelii 15Bodianus diplotaenia 1Chromis atrilobata 90 90 4Diodon holocanthus 1Halichoeres notospilus 2.5 1 1Hemirhamphus saltator 1Holacanthus passer 2 1.5Microspathodon dorsalis 2 18 1 5.5Mugil curema 4Novaculichthys taeniourus 2Ophioblennius steindachneri 2 2 1Stegastes acapulcoensis 4 22 3.5 9.5Sufflamen verres 1Thalassoma lucasanum 90 85 36.5NUMERO TOTAL (S) 12.5 239 184.5 62 1RIQUEZA DE ESPECIES 5 8 8 7 1ÍNDICE DE MARGALEF 1.098 0.886 0.930 1.008 0.000SIMPSON 0.151 0.299 0.448 0.377 0.000H'max 2.322 3.000 3.000 2.807 0.000H'n 2.259 2.033 1.363 1.911 0.000J' 0.973 0.678 0.454 0.681 0.000Densidad 0.208 3.983 3.075 1.033 0.017
22
Figura 6. Abundancia y diversidad de la ictiofauna por transecto en abril 2004.
Riqueza de especies
(b) Número de especies
T1
T2
T3
T4
T5
0 2 4 6 8 10
Número de individuos
(c) Individuos
T1
T2
T3
T4
T5
0 50 100 150 200 250
Simpson
(d) Dominancia S
T1
T2
T3
T4
0 0.1 0.2 0.3 0.4 0.5
Indice de Margalef
(e) Densidad D
T1
T2
T3
T4
0 0.2 0.4 0.6 0.8 1 1.2
Diversidad máxima
(f) H´max
T1
T2
T3
T4
T5
0 0.5 1 1.5 2 2.5 3 3.5
Indice de Shannon
(g) Diversidad H´n
T1
T2
T3
T4
T5
0 0.5 1 1.5 2 2.5
Pielou
(h) Equidad J
T1
T2
T3
T4
0 0.2 0.4 0.6 0.8 1 1.2
Densidad
(a) Número de individuos
T1
T2
T3
T4
T5
0 1 2 3 4 5

23
Figura 7. Abundancia y diversidad de la ictiofauna en abril de 2004.
(f)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
2.32
33
2.80
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad Máxima (H´max)
(a)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.20
3.983.07
1.03
0.017
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Densidad g/m2
(b)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
5
88
7
1
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Riqueza de especies
(c)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
13
239185
62
1
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Número de individuos
(d)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.15
0.290.44
0.37
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Dominancia (S)
(e)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
1.09
0.880.92
1.00
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Margalef
(g)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
2.25
2.031.36
1.91
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Shannon
(h)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.97
0.670.45
0.68
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Equidad (J´)

24
Los transectos 4, 3 y 2 correspondiente a la zona de coral presentan una menor riqueza, debido a que a pesar de que en esta zona hay una mayor disponibilidad de hábitats, muchos de estos ya se encuentran ocupados por especies territoriales tales como Microspathodon dorsalis y Stegastes acapulcoensis los cuales impiden que otras especies lleguen a establecerse en el lugar. En cuanto a la diversidad obtenida por el índice de Shannon (H´n), el valor más alto de diversidad se da en el transecto 1 (Tabla 3, Figuras 6g y 7g), ya que a pesar de que el número de especies es menor que en relación a las que hay en el arrecife, estas son más equitativas entre sí en cuanto a su número de individuos. En el resto de los transectos, los cuales corresponden a la zona de coral, presentan valores de diversidad menores ya que en esta zona a pesar de haber más especies y hábitats muchos de estos ya están ocupados por las especies más territoriales (Tabla 3, Figuras 6f y 7f). El valor más alto de equidad se presenta en el transecto 1 (Tabla 3, Figuras 6h y 7h), el cual corresponde a la zona rocosa, esto debido a que en este lugar la relación que hay entre las especies es más equitativa en cuanto al número de individuos de cada una, lo cual se atribuye a que en esta región posiblemente no hay gran disponibilidad de hábitats debido a que el área de este sistema no es muy extensa. Los transectos 2 y 4 son los siguientes en valor de equidad debido a que en esta región la relación entre el número de los organismos no es tan equitativa, debido a que aquí la disponibilidad de habitas es mucho mayor, lo cual permite que especies más territoriales se ubiquen en esta región, no obstante no presenta los valores más bajos de equidad. Dominancia. La mayor dominancia se presenta en el transecto 3 el cual corresponde a la zona de coral, (Tabla 3, Figuras 6d y 7d), y dada principalmente por las especies Chromis atrilobata y Thalassoma lucasanum, especies que no son territoriales con hábitos gregarios. En lo que respecta a los transectos 4 y 2 también de la zona de coral, son los que siguen en dominancia después del transecto 3, la cual esta dada prácticamente por las dos mismas especies. En el caso del transecto 1 correspondiente a la zona de roca, este fue el que presento la menor dominancia, debido a que a pesar de presentar pocas especies, estas son relativamente equitativas en su número de individuos. Riqueza de especies por tipo de ambiente (Figura 8). La mayor riqueza de especies registrada se encuentra en el ambiente arrecifal, lo cual sea atribuye a que es aquí donde se da la mayor protección para la ictiofauna contra la influencia del oleaje, así como también por la irregularidad del ambiente es que hay una mayor disponibilidad de hábitats que brindan refugio, protección y alimento a estas (Alevizon et al., 1985). Aquí también se presenta un gran número de individuos de las especies Thalassoma lucasanum y Chromis atrilobata, lo cual puede deberse a que en esta época es alto el pico del crecimiento del plancton y las dos especies que presenta mayor abundancia se alimentan de este recurso. El segundo ambiente en riqueza de especies es el rocoso, el cual no brinda una gran protección a los organismos y debido a esto es que su disponibilidad de hábitats es muy poca por lo que algunos cuantos

25
organismos son encontrados aquí estando presentes principalmente aquellos de hábitat rocoso como Microspathodon dorsalis y Novaculichthys taeniourus, entre otros. Finalmente el ambiente que menos riqueza de especies presenta es el arenoso, esto debido a que en esta zona no hay sitios de resguardo para las especies (McGehee, 1994), lo cual hace que la fuerza hidráulica presente en esta zona impida el crecimiento de algas y otros organismos lo cual limita que la ictiofauna se establezca en esta región, donde por lo general los organismos encontrados muy posiblemente estén de paso o en busca de alimento enterrado en la arena.
Figura 8. Riqueza de especies por ambiente en abril de 2004 correspondiente a la época de secas.
012345678
Riq
ueza
Roca Arrecife decoral
Arena
Tipo de ambiente
Riqueza de especies por ambiente
Roca
Arrecife de coral
Arena

26
Julio 2004.
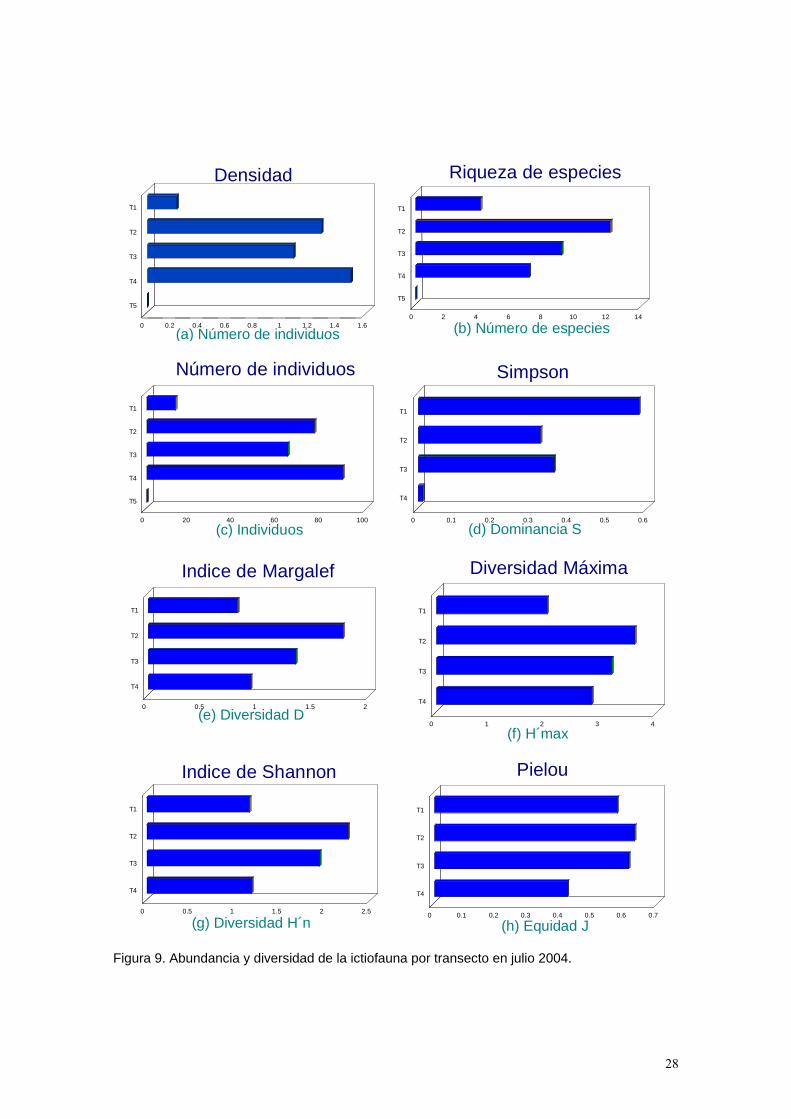
Tabla 4. Se presenta el número de individuos por especie correspondiente a cada transecto. Se muestran también los valores de los índices de diversidad, dominancia, equidad y densidad por transecto obtenidos en julio de 2004. Los valores del transecto número 5 correspondiente a la parte arenosa no se presentan debido a que no se registraron organismos en la zona. Abundancia. La mayor densidad la presenta el transecto 4 correspondiente a la zona de coral (Tabla 4, Figuras 9a y 10a), determinado principalmente por el gran número de individuos de la especie Thalassoma lucasanum. El transecto 2 y 3 son los que siguen en valor decreciente de densidad determinados por la misma especie a la del transecto 4, sin embargo el número de organismos de especies territoriales es mayor posiblemente debido a la disponibilidad de hábitats para los organismos así como también por la menor influencia que ejerce el oleaje en esta zona por la protección que le brinda la isla. El transecto 1 presenta la más baja abundancia de organismos ya que en esta zona la incidencia del oleaje es mucho mayor a la que se presenta en los transectos de coral. Finalmente el transecto 5 correspondiente a la zona de arena, presentó cero individuos posiblemente debido a que es una zona muy extensa de arena y es donde se da una de las más fuertes incidencias de oleaje. Diversidad. Los valores más altos de riqueza de especies (Tabla 4, Figuras 9b y 10b) se encuentran en los transectos correspondientes a la zona de coral (T2,
Especie T1 T2 T3 T4Arothron meleagris 2Balistes polylepis 1Bodianus diplotaenia 1Caranx caballus 10Cirrhitus rivulatus 1 1Chaetodon humeralis 1 1Halichoeres notospilus 1 1Holacanthus passer 1 1Jhonrandallia nigrirostris 1 1Kiphosus elegans 1Lutjanus argentiventris 1 1Microspathodon dorsalis 4 12 4Ophioblennius steindachneri 2Oxycirrhites typus 1Stegastes acapulcoensis 13 11 10Stegastes flavilatus 1Sufflamen verres 1 1Thalassoma lucasanum 10 40 35 70NUMERO TOTAL (S) 13 76 64 89RIQUEZA DE ESPECIES 4 12 9 7ÍNDICE DE MARGALEF 0.811 1.761 1.333 0.927SIMPSON 0.577 0.319 0.355 0.013H'max 2.000 3.585 3.170 2.807H'n 1.145 2.245 1.928 1.169J' 0.573 0.626 0.608 0.417Densidad 0.217 1.267 1.067 1.483

27
T3 y T4) En el caso del transecto 1 ubicado en el ambiente rocoso, este presenta una riqueza de especies más baja ya que en este lugar las condiciones imperantes de oleaje y corrientes son más fuertes, además de que la disponibilidad de hábitats no es tan grande. Respecto al transecto 5 de la zona arenosa, este no presenta valor de riqueza debido a la ausencia de individuos. De acuerdo a los índices de Margalef y Shannon la mayor riqueza corresponde a la zona de coral en el transecto 2 (figs. 9e, 10e, 9g y 10g). En el caso del transecto 3 este presenta una menor riqueza ya que se encuentra más protegido de la acción del oleaje, por lo que especies más territoriales están en una cantidad más elevada. El transecto 4 presentó la menor riqueza de todos los transectos de la zona arrecifal debido a que aquí la acción del oleaje y la incidencia de la fuerza hidráulica es mayor en comparación a los demás. En lo referente al transecto número 1 de la zona rocosa este presentó la menor riqueza según el índice de Margalef y Shannon, ya que aquí las condiciones de oleaje son mayores en relación a las que se presentan en el arrecife y la disponibilidad de hábitats es poca. El transecto 5 de la zona arenosa no presentó individuos de ninguna especie. Dominancia. El valor más alto de dominancia esta en el transecto número 1 de la zona rocosa (Tabla 4, Figuras 9d y 10d), debido al alto número de individuos de Thalassoma lucasanum encontrados en el lugar así como también por la baja abundancia de los organismos de otras especies. Los transectos 2 y 3 correspondientes a la zona arrecifal son los que presentan una dominancia moderada ya que a pesar de que poseen una riqueza de especies alta, muchas de estas solo están representadas por un solo individuo de ahí que la dominancia sea mayor, como es el caso del número 3 en donde se presenta una de las más altas abundancias de Microspathodon dorsalis. En el caso del transecto número 4, este presenta la menor dominancia ya que a pesar de que posee una riqueza de especies no muy alta, muchas de estas se encuentran representadas por más de un individuo, lo cual aumenta la equitatividad entre estas es por esto que aquí se encuentra a un menor número de Microspathodon dorsalis y Stegastes acapulcoensis en relación a otros transectos. No obstante el número de Thalassoma lucasanum es más alto. El valor más alto de equidad lo presenta el transecto número 2 ya que muchas de sus especies están representadas por un solo individuo (Tabla 4, Figuras 9h y 10h), de igual manera el transecto 3 es el que le sigue en valor por que también muchas las especies están representadas por un individuo, a excepción de las que poseen más de un representante de su especie en donde se encuentran principalmente aquellas de carácter territorial y algunas que forman cardúmenes. El tercer transecto en valor de equidad es el número 1, de manera semejante a los anteriores muchas de sus especies están representadas por un solo individuo. Finalmente la menor equidad se presenta en el transecto número 4 de la zona de coral ya que aquí la variación numérica entre los individuos de cada especie es mayor.
28
Figura 9. Abundancia y diversidad de la ictiofauna por transecto en julio 2004.
Riqueza de especies
(b) Número de especies
T1
T2
T3
T4
T5
0 2 4 6 8 10 12 14
Número de individuos
(c) Individuos
T1
T2
T3
T4
T5
0 20 40 60 80 100
Simpson
(d) Dominancia S
T1
T2
T3
T4
0 0.1 0.2 0.3 0.4 0.5 0.6
Indice de Margalef
(e) Diversidad D
T1
T2
T3
T4
0 0.5 1 1.5 2
Diversidad Máxima
(f) H´max
T1
T2
T3
T4
0 1 2 3 4
Indice de Shannon
(g) Diversidad H´n
T1
T2
T3
T4
0 0.5 1 1.5 2 2.5
Pielou
(h) Equidad J
T1
T2
T3
T4
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7
Densidad
(a) Número de individuos
T1
T2
T3
T4
T5
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6

29
Figura 10. Abundancia y diversidad de la ictiofauna en julio de 2004.
(a)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.21
1.261.06
1.48
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Densidad g/m2
(b)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
4
129
7
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Riqueza de especies
(c)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
13
7664
89
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Número de individuos
(d)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.57
0.310.35
0.01
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Dominancia (S)
(e)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.81
1.761.33
0.92
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Margalef
(f)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
2
3.583.16
2.80
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad Máxima (H’max)
(g)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
1.14
2.241.92
1.16
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Diversidad de Shannon
(h)
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
0.57
0.620.60
0.41
0
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
Equidad (J’)

30
El transecto número 5 perteneciente a la zona de arena no presentó índice de equidad debido a la ausencia de especies en éste. Riqueza de especies por tipo de ambiente (Figura 11). La mayor riqueza de especies se presenta en el ambiente arrecifal debido a que en éste se presentan las características más optimas para la supervivencia de las especies, ya que les brinda zonas de resguardo, protección y alimentación lo cual esta básicamente determinado por la irregularidad del sistema y el tipo de substrato (Alevizon et al., 1985). Por su parte el ambiente rocoso presenta una menor riqueza debido a que este ambiente se encuentra más influenciado por la acción del oleaje y las corrientes, así como también por su poca extensión en área, les es más difícil a las especies lograr establecerse en el lugar por lo cual básicamente se encuentran aquí organismos más adaptados a este tipo de ambiente y algunos cuantos visitantes ocasionales. Finalmente el ambiente arenoso no presenta riqueza de especies debido a que aquí es donde se presenta con mayor intensidad la acción del oleaje pues este no se encuentra protegido por alguna barrera física lo que limita el crecimiento de algas y otros organismos que sirva de alimento, por esta razón es que en este ambiente no hay la presencia de especies y las pocas que puedan llegar a encontrarse es muy factible que solo estén de paso en busca de alimento y protección.
Figura 11. Riqueza de especies por ambiente en julio de 2004 correspondiente a la época de lluvias.
0
2
4
6
8
10
Riq
ueza
Roca Arrecife decoral
Arena
Tipo de ambiente
Riqueza de especies por ambiente
Roca
Arrecife de coral
Arena

31
Distribución de la ictiofauna en cada uno de los am bientes de la bahía Cacaluta.
Tabla 5. Distribución de la ictiofauna en cada uno de los ambientes en la Bahía Cacaluta, en donde P = presente en el ambiente, R = residente, C = visitante cíclico, O = visitante ocasional, B = asociado a bordes y H = asociado a oquedades. La tabla 5 muestra la composición ictiofaunistica presente en los tres ambientes analizados. El ambiente que más especies presenta es el de arrecife de coral teniendo a 27 del total de 29 especies registradas. El ambiente que le sigue en riqueza de especies es el rocoso, el cual presenta a 14 especies de las 29 especies registradas. Finalmente se encuentra el ambiente arenoso el cual solo presenta a una sola especie, por lo que el ambiente más diverso es el de arrecife de coral ya que este ambiente posee las condiciones más favorables para el establecimiento y desarrollo de las especies pues se encuentra protegido por la isla Cacaluta contra el oleaje incidente, tiene una producción
Especie Roca Coral ArenaAbudefduf troschelii PRArothron meleagris PRBalistes polylepis PCBodianus diplotaenia PR PRBCaranx caballus PCBCirrhitus rivulatus PRChaetodon humeralis PR PCRBChromis atrilobata PCR PCRBDiodon holocanthus PC PCB PCEpinephelus labriformis PRHaemulon sexfasciatum PRHalichoeres notospilus PR PRBHemirhamphus saltator PC PCBHolacanthus passer PRHJhonrandallia nigrirostris PR PRBHKyphosus elegans PRBLutjanus argentiventris PCBMicrospathodon bairdii PRBHMicrospathodon dorsalis PR PRBHMugil curema PCBNovaculichthys taeniourus PROphioblennius steindachneri PR PROstracion meleagris PCBOxycirrhites typus PRPrionurus punctatus PRStegastes acapulcoensis PR PRStegastes flavilatus PR PRBSufflamen verres PBThalassoma lucasanum PR PR

32
mayor, disponibilidad de hábitats por la irregularidad y morfología que este presenta y debido a que el oleaje es más tranquilo en esta zona muchos organismos logran tener un establecimiento más exitoso, de ahí que esta zona sea también una de las que presenta más abundancia de individuos. Por su parte en lo que respecta al ambiente rocoso este presenta una menor riqueza por la fuerte incidencia del oleaje, a lo cual se aúna que esta región no es muy extensa en el área que presenta, limitando la disponibilidad de hábitats por lo que la densidad de individuos no es tan alta en comparación a la de la zona de coral y muchas de sus especies están representadas por pocos individuos, siendo aquellas con un número mayor principalmente las que son características de este ambiente por ejemplo Microspathodon dorsalis, así como algunos grupos grandes de otras especies asociadas al hábitat arrecifal como Thalassoma lucasanum y Chromis atrilobata. Finalmente el ambiente arenoso posee la menor diversidad con la presencia de un solo individuo atribuido a que en este ambiente se da la mayor incidencia del oleaje y dado que la irregularidad de su substrato es prácticamente nula, no ofrece hábitats para las especies por eso no hay establecimiento de estas en la zona y probablemente las que logren presentarse, se encuentren de paso. Análisis de las asociaciones presentes entre los tr ansectos y entre las especies para la bahía Cacaluta. El análisis de las asociaciones por especie en Febrero 2004 figura 12a. Muestra la separación de Thalassoma lucasanum del resto de los grupos debido a su alto número de individuos (395 representantes), esta es la especie limpiadora ya que son organismos de talla pequeña que forman agregaciones y se desplazan a través de la columna de agua en busca de alimento, así como por no ser territoriales estos se mueven de un sitio a otro dependiendo de las condiciones que más les favorezcan (Warner y Hoffman, 1980). El segundo grupo que se observa es el conformado por las especies Abudefduf troschelii, Microspathodon dorsalis, Stegastes acapulcoensis, Mugil curema y Chromis atrilobata; donde la asociación más estrecha se da entre las dos primeras especies debido al número de individuos que presentan cada una con 37 y 48 individuos respectivamente a la cual se unen las otras especies Este grupo esta conformado por especies dominantes en donde algunas son altamente territoriales como S. acapulcoensis y M. dorsalis las cuales se encuentran presentes en los ambientes rocoso y arrecifal (Ramos-Santiago, 2004), siendo este ultimo donde son mayoritariamente más abundantes principalmente S. acapulcoensis, también presenta ligeramente un número un poco más alto en la zona arrecifal. M. curema, principalmente se encuentra en los fondos fangosos y arenosos de las lagunas costeras donde penetran para alimentarse del detritus proporcionado por el suelo arenoso (Robins y Ray, 1986), por lo que muy probablemente estaban de paso en el arrecife o en busca de alimento. Este grupo no se une con T. lucasanum debido a que la proporción de individuos que presenta este último es mayor sobre todo en juveniles, en tanto que los censados de las otras especies como M. dorsalis y S. acapulcoensis son adultos. Finalmente se forma un tercer grupo conformado por las especies que poseen muy pocos representantes y que son menos abundantes y por ende mucho más ocasionales y solitarios aún cuando

33
Figura 12. A
sociaciones presentes por especie y por transecto obtenidas mediante el m
étodo de W
ard para cada mes de m
uestreo en la bahía Cacaluta durante el año 2004.
Febrero, 2004
(a)
Especies
Distancia de acoplamiento
050
100150200250300350400
T. lucasanum
S. flavilatus
P. punctatus
H. saltator
D. holocanthus
B. diplotaenia
O. meleagris
M. bairdii
H. notospilus
H. sexfasciatum
O. steindachneri
E. labriformis
A. meleagris
C. atrilobata
M. curema
S. acapulcoensis
M. dorsalis
A. troschelii
Febrero, 2004
(b)
Transectos
Distancia de acoplamiento
0
50
100
150
200
250
300
T4
T3
T2
T5
T1
Abril, 2004
(c)
Especies
Distancia de acoplamiento
050
100150200250300350400
T. lucasanum
C. atrilobata
O. steindachneri
N. taeniourus
H. notospilus
M. curema
H. passer
H. saltator
D. holocanthus
S. verres
B. diplotaenia
S. acapulcoensis
M. dorsalis
A. troschelii
Abril, 2004
(d)
Transectos
Distancia de acoplamiento
0
50
100
150
200
250
T3
T2
T4
T5
T1
Julio, 2004
(e)
Especies
Distancia de acoplamiento
020 40 60 80
100120140160
T. lucasanum
S. acapulcoensis
M. dorsalis
C. caballus
S. flavilatus
O. typus
J. nigrirostris
L. argentiventris
H. passer
S. verres
O. steindachneri
C. rivulatus
B. diplotaenia
H. notospilus
C. humeralis
K. elegans
B. polylepis
A. meleagris
Julio, 2004
(f)
Transectos
Distancia de acoplamiento
0
10 20 30 40 50 60 70 80 90
T4
T3
T2
T5
T1

34
Figura 13.Representación de la agrupaciones obtenidas por el análisis ”cluster” por transectos para los tres censos en la bahía Cacaluta.
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
T1
T2
T3
T4
T5
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
(a) Febrero 2004
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
T1
T2
T3
T4
T5
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
(b) Abril 2004
Playa Cacaluta
Playa Arroyo
Isla Cacaluta
T1
T2
T3
T4
T5
96°09’59’’LW 96°09’11’’LW
15°43’20.5’’LN
15°43’09’’LN
(c) Julio 2004

35
muchas de estas son de tipo arrecifal, lo cual se atribuye a que estas se ven influenciadas por factores físicos y biológicos tanto externos como internos, posiblemente por el tipo de alimentación, por la velocidad de su taza de reproducción y fertilidad, por su cantidad de depredadores y por la irregularidad del tipo de substrato, lo cual puede influir en su biología (Graus y Macintyre, 1989). El análisis de las asociaciones por transecto de Febrero 2004 figuras 12b y 13a. Muestra una separación entre los transectos correspondientes a la zona de coral T4, T3 y T2 que forman el grupo 1 de los pertenecientes a la zona rocosa y arenosa T1 y T5 grupo 2, determinado por la cantidad de individuos por transecto, donde los transectos de la zona arrecifal poseen los valores más altos. Mientras que el transecto 1 de la zona rocosa se encuentra más relacionado con el transecto 5 de la zona arenosa debido a que tienen valores más bajos. Por lo tanto las características del grupo 1 son más parecidas entre sí mismas que a las del grupo 2, es decir, en la zona arrecifal posiblemente se esten cubriendo mejor las necesidades de hábitat que requieren las especies y por eso se tiene mayor abundancia de individuos ya que las especies pueden establecerse exitosamente por el refugio y protección que éste les brinda y dependiendo de su grado de dominancia conformaran la alta o baja abundancia de individuos presentes en el lugar. Mientras que los transectos 1 y 5 se separan de los otros por las diferencias del tipo de ambiente, diferencias que se reflejan en el número de individuos el cual está en función de las necesidades de hábitat que requieren las especies para sobrevivir. El análisis de las asociaciones por especie de Abril 2004 figura 12c. Muestra la formación de 3 grupos. El primero integrado por T. lucasanum y C. atrilobata que se distancian del resto por que presentan él más alto número de individuos lo cual se atribuye a sus hábitos y necesidades las cuales entre más sean cubiertas permiten su permanencia, así como también su talla y forma les dan la oportunidad de ser nadadores más ágiles favoreciendo no ser una presa fácil y el hecho de que su tipo de alimentación sea variada les da la oportunidad de tener una mayor subsistencia. El segundo grupo se encuentra integrado por S. acapulcoensis, M. dorsalis y A. troschelii, donde la relación más estrecha se da entre las dos primeras especies debido a que poseen el número más alto de individuos de este grupo y también son muy territoriales su talla es mediana a grande por lo que su velocidad de reclutamiento es mucho menor a otras especies (Thomsom et al., 1979), de ahí que también la gran mayoría de los individuos censados de estas fueron adultos. Debido a su alta territorialidad es que su proporción es mayor en la zona arrecifal ya que dependen de sitios estables para lograr un exitoso establecimiento, pues es aquí donde llevaran a cabo su reproducción y desarrollo (Ramos-Santiago, 2004). Sin embargo debido a que M. dorsalis se alimenta de algas es por lo que se encuentra en los bordes del arrecife, en algunos recovecos y en las partes rocosas que se mezclan con éste, de ahí que también por eso posiblemente su población no es muy grande.

36
A. troschelii cual no es una especie territorial también se integra a este grupo ya que por su talla pequeña forma agrupaciones numerosas para desplazarse en busca de alimento y protección (Thomson, 1987). Finalmente se forma un tercer grupo integrado por las especies que poseen muy pocos representantes principalmente por ser de hábitos solitarios. El análisis de las asociaciones por transecto de Abril 2004 figuras 12d y 13b. Muestra la separación en dos grupos, el primero conformado por los transectos 3 y 2 pertenecientes a la zona de coral, ya que son los que presentan el más alto número de individuos. El segundo grupo se encuentra conformado por los transectos T4 de la zona de coral, T1 de la zona rocosa y T5 de la zona arenosa, donde la relación más fuerte se da entre el transecto 5 y 1 debido a la baja cantidad de individuos que poseen y el transecto 4 se une debido a que su valor del número total de individuos no es muy alto, siendo este de 62 individuos, mientras que T1 posee 13 individuos y T5 solo un individuo. Los transectos pertenecientes a la zona de coral son más abundantes en su número de individuos debido a que aquí es donde logran algunas especies encontrar las condiciones más optimas para establecerse dando como resultado el incremento en su número de individuos volviendose dominantes Por su parte los transectos 1 y 5 poseen una baja abundancia de individuos debido al tipo de características que posee cada ambiente en su substrato y morfología por eso se separan de la zona de coral pues son diferentes. El transecto 4 de la zona de coral se asocia a estos otros ambientes por la baja abundancia de individuos que presenta debido a las influencias del oleaje, más no porque posea las mismas características de los ambientes rocoso y arenoso. El análisis de las asociaciones por especie de julio 2004 figura 12e. Muestra la separación de las especies en tres grandes grupos, el primero conformado por Thalassoma lucasanum cual no se integra al resto y presenta el mayor número de individuos atribuido principalmente a que esta especie no es territorial y su tipo de alimentación es relativamente variada (Thomson et al., 1979; Warner y Hoffman, 1980). El segundo grupo se encuentra conformado por S. acapulcoensis, M. dorsalis y C. caballus donde la mayor relación se presenta entre las dos primeras especies debido a que son altamente territoriales con una talla mediana a grande dentro de los peces de arrecife de coral y con una alimentación más variada principalmente S. acapulcoensis ya que es omnívoro el cual forrajea pequeños invertebrados y algas bentónicas, en tanto que M. dorsalis se alimenta de algas (Ramos-Santiago, 2004). C. caballus se integra a este grupo debido a que su número de individuos es alto, no obstante esta especie solo se presento en uno de los transectos de la zona arrecifal y no en el resto por lo que no se le considera como especie dominante. No es de hábitos territoriales pero forma agregaciones para desplazarse en busca de peces, cefalópodos, crustáceos y otros invertebrados (Froese y Pauly, 2004). Finalmente el tercer grupo esta conformado por especies que poseen hábitos solitarios.

37
El análisis de las asociaciones por transecto de Julio 2004 figuras 12f y 13c. Muestra la formación de dos grupos, el primero integrado por los transectos T4, T3 y T2 correspondientes a la zona de coral y el segundo integrado por los transectos T5 de la zona arenosa y T1 de la zona rocosa. La primera integración se debe a que son los que presentan el número más alto de individuos. Estos transectos son mucho más abundantes debido a que la zona arrecifal cubre las necesidades de hábitat de una forma más óptima para ciertas especies tales como refugio, protección y alimento El transecto 4 se ve más influenciado por la acción del oleaje debido a esto su mayor número de individuos esta determinado por Thalassoma lucasanum por esto es que también en este transecto las especies territoriales son menos abundantes, en tanto que otras se hacen presentes, por ejemplo O. steindachneri. El grupo número 2 sus transectos se encuentran más relacionados debido a que presentan menor cantidad de individuos y se separan del resto debido a que las características morfológicas que posee el fondo rocoso y el arenoso son muy diferentes a las presentes en el coral, además de presentar diferencias en el oleaje incidente por la ausencia de una barrera física que lo limite. Diversidad, dominancia, equidad, número total de in dividuos para los tres censos realizados En la figura 14a en el primer mes de muestreo de la época de secas los valores del número total de individuos fluctúan entre 0 y 254; comparando estos valores con los del segundo mes de muestreo también de la época de secas, se tiene que el número es menor ya que este fluctúa entre 1 y 239, sin embargo esta diferencia no es muy marcada. El tercer muestreo de la época de lluvias presentó valores del número total de individuos con una fluctuación menor, siendo esta de 0 a 89. Atribuido a que durante secas el agua es fría, hay surgencias que acarrean nutrientes del fondo y hay mayor circulación en la columna de agua por la ausencia de termoclinas, que en consecuencia genera un incremento en la cantidad de fito y zooplancton, el cual atrae a especies planctófagas en cantidad y por ende hay baja diversidad, de ahí que hay una gran cantidad de individuos Thalassoma lucasanum y Chromis atrilobata. En el último muestreo el cual corresponde a la época de lluvias hay menor cantidad de individuos debido a que en esta temporada el agua es cálida, se establece una termoclina impidiendo la recirculación de la columna de agua y por ende la resuspensión de nutrientes, entonces el fitoplancton y zooplancton es poco y las especies planctófagas se encuentran en baja cantidad o desaparecen. Como Chromis atrilobata cual es estrictamente planctófaga y posiblemente se desplazo hacia zonas con mayor abundancia de este recurso. Por su parte el número de Thalassoma lucasanum es menor en este muestreo pero no desaparece porque sus hábitos alimenticios son más variados. El resto de las especies presentan un comportamiento semejante Esto genera un incremento en la diversidad y por ende una disminución en la dominancia, a lo cual también se puede aunar que en esta época se presenta un incremento en los deslaves, aumento de la sedimentación y aumento en la turbidez del agua en la mayoría de los casos.
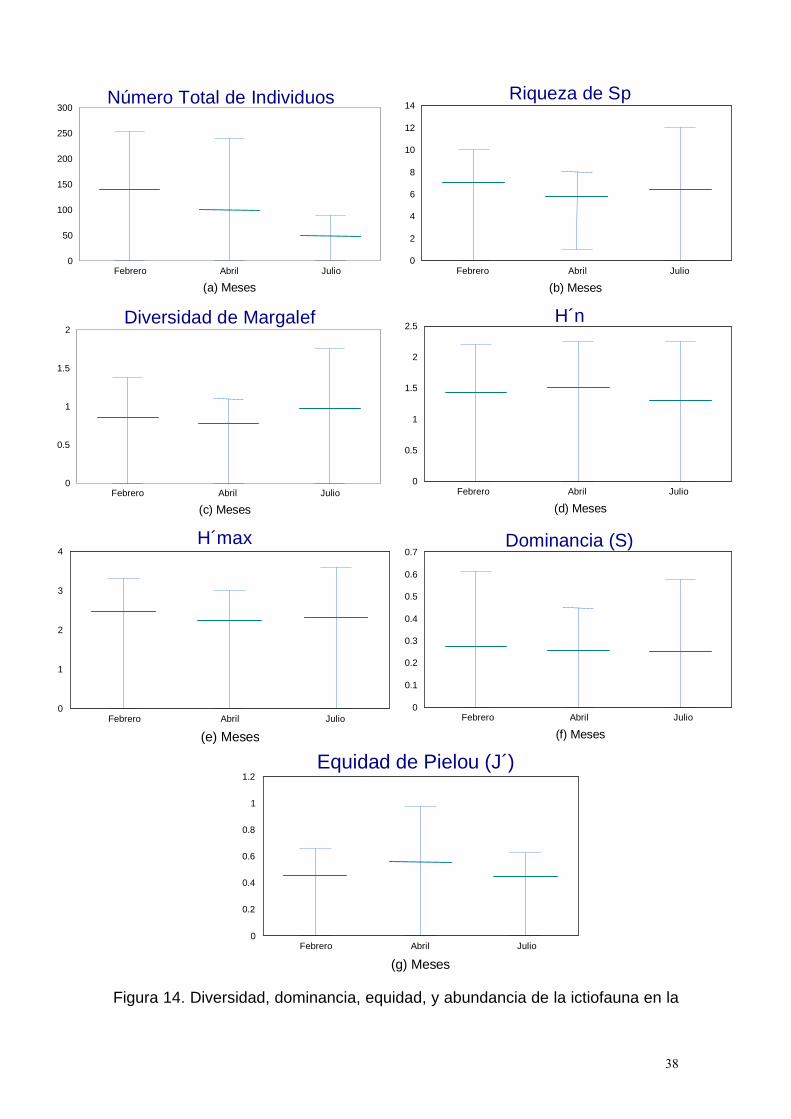
38
Figura 14. Diversidad, dominancia, equidad, y abundancia de la ictiofauna en la
Número Total de Individuos
Febrero Abril Julio
(a) Meses
0
50
100
150
200
250
300Riqueza de Sp
Febrero Abril Julio
(b) Meses
0
2
4
6
8
10
12
14
Diversidad de Margalef
Febrero Abril Julio
(c) Meses
0
0.5
1
1.5
2H´n
Febrero Abril Julio
(d) Meses
0
0.5
1
1.5
2
2.5
H´max
Febrero Abril Julio
(e) Meses
0
1
2
3
4
Equidad de Pielou (J´)
Febrero Abril Julio
(g) Meses
0
0.2
0.4
0.6
0.8
1
1.2
Dominancia (S)
Febrero Abril Julio
(f) Meses
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7

39
bahía de Cacaluta. En la figura 14b la riqueza de especies del primer mes de muestreo correspondiente a la época de secas fluctúa entre 0 y 10, el segundo mes también de la temporada de secas fluctua entre 1 y 8 siendo más bajo que el primer censo. los valores del tercer mes correspondiente a lluvias presenta mayor variación en la riqueza de especies, siendo esta de 0 a 12 y son los valores más altos registrados. Esto es debido a que en la época de secas hay mayor cantidad de nutrientes favoreciendo la presencia de un gran número de individuos de especies planctófagas y de algunas otras cuales poseen cierto grado de territorialidad y dominancia. La figura 14c muestra la diversidad de Margalef o riqueza de especies (D). En el primer mes los datos fluctúan entre 0 y 1.37, mientras que en el segundo mes los valores son más bajos y fluctúan entre 0 y 1.09. Ambos muestreos son de la época de secas. El tercer muestreo realizado (temporada de lluvias) presentó la mayor diversidad de todos los censos con valores que fluctúan entre 0 y 1.76 lo cual se atribuye a que hay un aminoramiento en la cantidad de individuos de especies planctófagas determinado por la variación estacional ya antes explicada. La figura 14d muestra el índice de diversidad de Shannon (H´n) el primer mes de muestreo de la época de secas fluctúa entre 0 y 2.19, mientras que el segundo mes fluctuá entre 0 y 2.25 incrementandose ligaramente. El tercer muestreo realizado en la temporada de lluvias no posee una variación tan marcada ya que sus valores fluctúan entre 0 y 2.24, e inclusive es muy parecido al obtenido en el segundo mes, no obstante esto no fue semejante en el análisis de riqueza de especies del índice de Margalef (Figura 14c) ya que este no toma en cuenta la equitatividad entre las especies para determinar la diversidad de ahí que la temporada de lluvias sea la más diversa según este índice. Entonces lo que nos muestra la figura 14d es que durante los tres muestreos la equitatividad entre las especies se mantuvo relativamente estable. Ahora bien si se toma en cuenta el índice de diversidad máxima (H´max) figura 14e que son los valores esperados, se tiene que el primer mes de muestreo presenta valores que fluctúan entre 0 y 3.32, mientras que en el segundo mes los valores están entre 0 y 3, en tanto que en el tercer mes los valores se encuentran entre 0 y 3.58 siendo esta la más alta diversidad de todos los muestreos. Esto muestra que la temporada potencialmente más diversa es la de lluvias, lo cual coincide con los análisis de riqueza de especies y con la diversidad de Margalef, ya que posee la mayor riqueza de especies y la mayor diversidad. La figura 14f muestra los valores de dominancia según el índice de Simpson (S), para cada uno de los meses de muestreo realizados en las temporadas de secas y lluvias se aprecia que en el primer mes los valores de dominancia

40
fluctúan entre 0 y 0.61, mientras que en el segundo mes los valores están entre 0 y 0.44, siendo estos más bajos en comparación al primero, para el tercer mes los valores fluctúan entre 0 y 0.57, siendo intermedios entre los dos muestreos de la época de secas. Por lo tanto la mayor dominancia se presentó en el primer mes de muestreo por la presencia de una gran cantidad de individuos de especies planctívoras La figura 14g presenta la equidad de Pielou (J´) donde se aprecia que en el primer mes de muestreo los valores fluctúan entre 0 y 0.66, mientras que en el segundo mes los valores se encuentran entre 0 y 0.97 siendo estos más altos en comparación al primer muestreo. Respecto al tercer mes de muestreo los valores fluctúan entre 0 y 0.62 siendo estos los más bajos de todos los muestreos, lo cual se debe a que en la temporada de lluvias hay menor cantidad de plancton por lo que las especies que se alimentan de este recurso reducen su número o se desplazan hacia otras zonas dejando hábitats disponibles para ser ocupados por otras especies las cuales no presentan el mismo o semejante número de individuos entre ellas de forma que la equidad disminuye. Nota: La variación numérica tan amplia entre el valor mínimo y máximo para cada uno de los índices es debida a que en el transecto arenoso solamente una ocasión se registró un individuo y en el resto de los muestreos no se encontró nada, siendo este ambiente el que determino para cada censo los valores mínimos en el número total de individuos cuales fueron de cero, sin embargo estos no se omitieron debido a que el censo si se realizó. Determinación de poblaciones dominantes Utilizando la frecuencia de porcentaje (%) y la abundancia (densidad) para cada época de censo se pudieron determinar cuales son las especies dominantes en todos los transectos durante los periodos de secas y lluvias, siendo tres las especies dominantes encontradas cuales son Thalassoma lucasanum, Stegastes acapulcoensis y Microspathodon dorsalis.
Tabla 6. Especies dominantes en febrero del 2004, época de secas. La tabla 6, figura 15a de la época de secas se aprecian las especies con mayor porcentaje de frecuencia estas son Thalassoma lucasanum (la cual es la que posee la cantidad más alta de individuos de este censo) estando en dos de los tres ambientes (roca y coral), seguida de Chromis atrilobata presente también en los dos mismos ambientes, pero solo en uno de los transectos de la zona de coral; Stegastes acapulcoensis también presente en los ambientes rocoso y coralino; y finalmente Microspathodon dorsalis presente en estos dos
Especie T1 Roca T2 Coral T3 Coral T4 CoralThalassoma lucasanum 4.5 150 125 115Chromis atrilobata 50 30Stegastes acapulcoensis 3.5 8.5 18 35Microspathodon dorsalis 10 9.5 15.5 12.5

41
ambientes. Ninguna de estas especies ni de las otras se registro en el ambiente arenoso.
Tabla 7. Especies dominantes en abril del 2004, época de secas. La tabla 7, figura 15b de la época de secas presenta las especies con mayor porcentaje de frecuencia, estas son Thalassoma lucasanum cual posee el número más alto de individuos de todo el censo, pero solo se presenta en el ambiente arrecifal; Chromis atrilobata la cual solo se presenta en la zona arrecifal; Stegastes acapulcoensis cual se presenta en todos los transectos del ambiente rocoso y coralino; y Microspathodon dorsalis que se presentó en todos los transectos de la zona rocosa y de coral.
Tabla 8. Especies dominantes en julio del 2004, época de lluvias. Finalmente la tabla 8, figura 15c de la época de lluvias muestra las especies con los valores más altos de porcentaje de frecuencia y abundancia cuales son Thalassoma lucasanum que presentó el número más alto de individuos de todo el censo y se encontró en todos los transectos de la zona rocosa y de coral; Stegastes acapulcoensis es la que le sigue, sin embargo esta solo se presentó en los transectos de la zona de coral, por último esta Microspathodon dorsalis cual se registró en todos los transectos de la zona de coral. Es obvia la ausencia de la especie Chromis atrilobata durante este muestreo, así como también es notorio que los valores en el número total de individuos de estas especies han sido los más bajos (en 50% a los primeros registrados) y estos han ido disminuyendo paulatinamente desde el primer censo posiblemente debido a la variación climática que se da entre la época de secas y lluvias, así como la influencia que ejercen sobre los flujos de energía presentes en la bahía Cacaluta. Las especies dominantes para las tres épocas de muestreo son Thalassoma lucasanum, Stegastes acapulcoensis y Microspathodon dorsalis. Ya que presentaron los valores más altos en porcentaje de frecuencia, abundancia y número de individuos, así como estuvieron en todos los censos realizados en casi todos los transectos de las zonas rocosa y coralina, a excepción del ambiente arenoso. Chromis atrilobata a pesar de poseer un mayor porcentaje de frecuencia, abundancia y número de individuos que Stegastes acapulcoensis y Microspathodon dorsalis no se puede considerar como dominante ya que una especie dominante es aquella que posee alta frecuencia
Especie T1 Roca T2 Coral T3 Coral T4 CoralThalassoma lucasanum 90 85 36.5Chromis atrilobata 90 90 4Stegastes acapulcoensis 4 22 3.5 9.5Microspathodon dorsalis 2 18 1 5.5
Especie T1 Roca T2 Coral T3 Coral T4 CoralThalassoma lucasanum 10 40 35 70Stegastes acapulcoensis 13 11 10Microspathodon dorsalis 4 12 4Caranx caballus 10
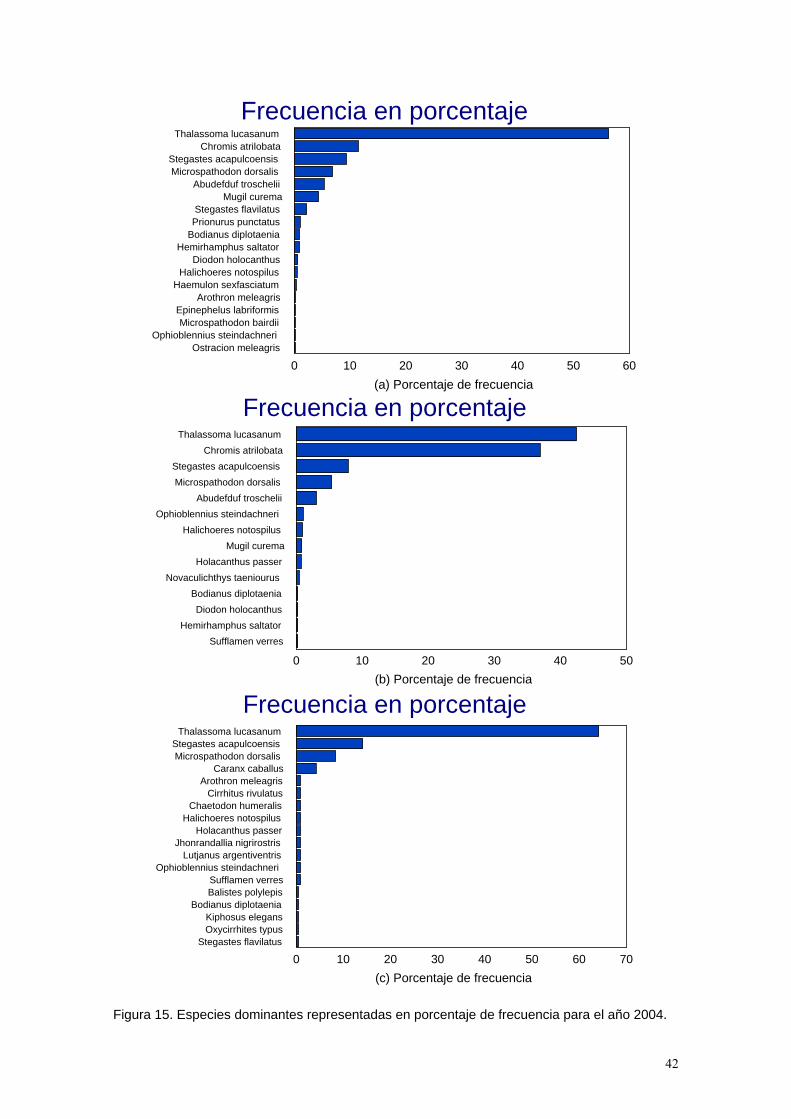
42
Figura 15. Especies dominantes representadas en porcentaje de frecuencia para el año 2004.
Frecuencia en porcentajeThalassoma lucasanum
Chromis atrilobataStegastes acapulcoensisMicrospathodon dorsalis
Abudefduf troscheliiMugil curema
Stegastes flavilatusPrionurus punctatus
Bodianus diplotaeniaHemirhamphus saltator
Diodon holocanthusHalichoeres notospilus
Haemulon sexfasciatumArothron meleagris
Epinephelus labriformisMicrospathodon bairdii
Ophioblennius steindachneriOstracion meleagris
0 10 20 30 40 50 60
(a) Porcentaje de frecuencia
Frecuencia en porcentajeThalassoma lucasanum
Chromis atrilobata
Stegastes acapulcoensis
Microspathodon dorsalis
Abudefduf troschelii
Ophioblennius steindachneri
Halichoeres notospilus
Mugil curema
Holacanthus passer
Novaculichthys taeniourus
Bodianus diplotaenia
Diodon holocanthus
Hemirhamphus saltator
Sufflamen verres
0 10 20 30 40 50
(b) Porcentaje de frecuencia
Frecuencia en porcentajeThalassoma lucasanum
Stegastes acapulcoensisMicrospathodon dorsalis
Caranx caballusArothron meleagris
Cirrhitus rivulatusChaetodon humeralis
Halichoeres notospilusHolacanthus passer
Jhonrandallia nigrirostrisLutjanus argentiventris
Ophioblennius steindachneriSufflamen verresBalistes polylepis
Bodianus diplotaeniaKiphosus elegansOxycirrhites typus
Stegastes flavilatus
0 10 20 30 40 50 60 70
(c) Porcentaje de frecuencia

43
reproductiva, alta fecundidad y amplia temporada reproductiva lo que la hace dominante en número y también si esta es capaz de colonizar los diferentes tipos de zonas por su gran capacidad de obtener alimento a diferencia de otras especies. Y el hecho de que no se presente en ninguno de los transectos de los ambientes censados en el último muestreo, tal vez es debido a que su capacidad de obtener alimento no es tan alta o a que su ítem es poco variado, ya que esta especie es planctívora (Froese y Pauly, 2004). Thalassoma lucasanum. Esta es la especie más dominante debido a su alta frecuencia y gran cantidad de individuos; es una especie de hábitos diurnos muy abundante en los arrecifes coralinos del pacífico tropical, su dieta es variada ya que se alimenta de plancton, algas, crustáceos, coral blando e inclusive de excremento de otros peces (Thomsom et al., 1979; Warner y Hoffman, 1980). Así como esta presenta una alta frecuencia reproductiva, alta fecundidad y amplia temporada reproductiva, su mayor abundancia en el arrecife esta ligada a que en éste se da el crecimiento de algas bentónicas y algunos crustáceos los cuales forman parte de su dieta (Williams, 1980), además de brindarle refugio y protección. Stegastes acapulcoensis. Es una especie territorial de hábitos diurnos con alimentación omnívora, la cual forrajea pequeños invertebrados y algas bentónicas, con una reproducción que inicia en los meses de primavera extendiéndose hasta el verano (Ramos-Santiago, 2004). Es mucho más abundante en el hábitat arrecifal aunque también se encuentra distribuida en el ambiente rocoso. Microspathodon dorsalis. Esta especie es altamente territorial defendiendo las zonas de alimentación y reproducción de otros peces, su alimentación es a base de algas. Su reproducción comienza desde el mes de abril, continuando durante el verano en el Golfo de California (Ramos-Santiago, 2004). Habita las zonas rocosas y de arrecife de coral aunque se conoce muy poco sobre los factores que determinan su distribución y abundancia en estas zonas. Chromis atrilobata. Esta especie habita las zonas de arrecife de coral y rocosas, no es territorial ya que se desplaza en busca de plancton el cual esta suspendido en la columna de agua y deriva con la corriente por lo que no puede mantenerse en un solo lugar, además de que por su pequeña talla sería una presa fácil para sus depredadores si se mantuviera estático a diferencia de otros peces damisela, tampoco realiza migraciones. Forma grandes agregaciones en las aguas abiertas cercanas a los arrecifes donde se alimenta exclusivamente de zooplancton (Froese y Pauly, 2004). Por tipo de ambiente en el caso del arrecifal es más dominante T. lucasanum y S. acapulcoensis. La primera realiza acciones de limpieza en otras especies que tienen mayor abundancia en esta zona. En tanto que S. acapulcoensis reside aquí ya que se cubren mejor sus necesidades de hábitat. M. dorsalis esta presente solo en los bordes y oquedades. En el ambiente rocoso es mayormente dominante M. dorsalis ya que sus necesidades son mejor cubiertas, pues este se alimenta de algas las cuales crecen adheridas a las rocas.

44
Conclusión
La ictiofauna de la bahía Cacaluta se encuentra determinada principalmente por la irregularidad del substrato en primera instancia, en segundo grado por la hidrodinámica de las masas de agua (el oleaje y las corrientes) y su variación así como cantidad de individuos esta dada primordialmente por la temporada del año (secas y lluvias) ya que durante estos periodos se da una serie de cambios en la estructura de la comunidad ictiofaunal determinada por la abundancia del plancton presente en cada época. La composición ictiofaunística se encuentra integrada por 29 especies pertenecientes a 27 géneros, 18 familias y a 2 subfamilias, sin embargo no se descarta la presencia de más especies debido a que el censo se realizó durante el día y dado que algunas especies son de hábitos nocturnos es muy alta la probabilidad de que estas no hayan sido registradas. Las especies dominantes para esta bahía durante todo el año son tres las cuales son Thalassoma lucasanum, Stegastes acapulcoensis y Microspathodon dorsalis ya que estas fueron las que presentaron el mayor porcentaje de abundancia de todas las especies debido a su amplia distribución en los ambientes rocoso y coralino así como por estar presentes durante los tres censos realizados. La variación espacio-temporal de la riqueza de especies en el lugar se encuentra determinada principalmente de la siguiente manera. Por zonas la región arrecifal es la que presenta la mayor riqueza de especies debido a la irregularidad de su substrato, ya que este les brinda refugio, protección y alimento teniendo de esta manera las características más óptimas para el establecimiento de las especies, así como también por su gran abundancia de hábitats a lo cual se encuentra aunado la presencia de la isla Cacaluta cual aminora las influencias del fuerte oleaje de ésta bahía. Mientras que la riqueza de especies para los ambientes rocoso y arenoso es baja debido a que estos son menos irregulares, poseen una menor extensión (en el caso del ambiente rocoso) lo que reduce su cantidad de hábitats y están más influenciados por el oleaje dada la ausencia de una barrera física que lo aminore de tal manera que el establecimiento de las especies es más difícil por la ausencia de zonas que brinden protección y refugio a las especies contra sus depredadores y contra los factores ambientales. Por épocas predominantes en la región (secas y lluvias) las cuales tienen gran influencia en esta debido principalmente a la relación que tienen con los nutrientes y la circulación de la columna de agua favoreciendo la abundancia del plancton y otros procesos los cuales promueven el incremento en número de individuos de las especies que se alimentan de este recurso principalmente en la época de secas por lo que la riqueza de especies en esta temporada es baja, mientras que durante lluvias hay una reducción en el número de individuos por especie debido a la ausencia del plancton favoreciendo la disponibilidad de hábitats para ser ocupados por otras especies dando como resultado que la riqueza de especies se incremente en ésta temporada y por lo

45
tanto esta no se mantenga constante durante las dos épocas sino presenta variación. La variación espacio-temporal de la diversidad se presenta de la siguiente manera. Por ambiente varia dependiendo de la época, durante secas la mayor diversidad la presenta el ambiente rocoso ya que la zona arrecifal a pesar de poseer una mayor irregularidad de su substrato, mayor extensión y estar protegida por la isla; posee un alto número de individuos de especies planctívoras, siendo esta la temporada en la que este recurso es más abundante y dado las condiciones que el arrecife proporciona son más favorables para el establecimiento de una gran cantidad de individuos es que muchos de estos llegan y permanecen aquí lo que incrementa la dominancia en la zona, sin embargo como la zona rocosa no es tan amplia y las condiciones que aquí imperan no favorecen el establecimiento satisfactorio de muchas especies y mucho menos de una gran cantidad de individuos es por lo que a pesar de la gran abundancia del plancton no se encuentran muchos individuos de especies planctívoras. Mientras que durante lluvias la mayor diversidad la presenta el ambiente arrecifal ya que en esta temporada hay una reducción en el número de individuos de las especies planctívoras así como el desplazamiento de algunas de estas debido a la menor abundancia del plancton, por lo que dado a la gran cantidad de hábitats presentes en el lugar los cuales albergan a muchas especies, estas una vez que la dominancia ejercida por otras disminuye hacen que la diversidad en la zona se incremente. La variación espacio-temporal de la dominancia se presenta de la siguiente manera. Por ambiente varia dependiendo de la época, durante secas la mayor dominancia la presenta el ambiente arrecifal debido al gran número de individuos que se alimentan de plancton, siendo esta la temporada en la que este recurso es más abundante, mientras que durante lluvias la mayor dominancia la presenta el ambiente rocoso ya que la cantidad de plancton se reduce y consigo hay la reducción de especies planctívoras en su cantidad y en su número total de individuos, permitiendo de esta manera la entrada y colonización en el arrecife de otras especies por lo que la dominancia se reduce. El ambiente arenoso posee una nula diversidad y dominancia por la ausencia de especies de la ictiofauna en el lugar, debido a que la irregularidad en el substrato es nula con lo cual no hay la disponibilidad de sitios que brinden resguardo y protección a las especies contra sus depredadores; por la fuerte incidencia del oleaje que se presenta en el lugar, siendo esta la zona donde golpea más fuerte por no estar protegida por alguna barrera que lo aminore y por su basta extensión la cual no se encuentra en colindancia con los otros dos ambientes por su ubicación, lo cual la aísla de estos impidiendo que funja como una zona de transición de especies entre ambos ocasionando así la ausencia de individuos de la ictiofauna en el lugar. La bahía Cacaluta posee una gran riqueza específica, amplia diversidad y dominancia cuales actúan de forma periódica por la temporalidad del año y por los procesos biológicos de los organismos del lugar en donde todavía la influencia de la actividad humana es poco marcada dado el difícil acceso por

46
tierra hacia el lugar. El cuidarlo y preservarlo aislado favorecerá la conservación de las especies presentes en el lugar tanto de la ictiofauna, como las de corales, crustáceos, moluscos, equinodermos, etc., así como también de la flora submarina y de la flora y fauna terrestres típicas de la zona.
Agradecimientos A mis padres. Juan Luna Peralta y Mercedes Monsivais Castillo: Por brindarme su apoyo, comprensión y por todos los sacrificios realizados a través de sus esfuerzos para que yo lograra terminar mi carrera profesional siendo ésta para mí la más valiosa herencia que me pudieron otorgar. A mis hermanos.
Cynthia y Edwin: Quienes siempre se preocuparon por mí y me brindaron todo su apoyo, por los esfuerzos y sacrificios que pasaron solo con la finalidad de que yo pudiera concluir mis estudios, así como por estar siempre conmigo tanto en los buenos como en los malos momentos; siempre les estaré agradecido. A mis profesores.
Dr. Margarito Tapia García: Quien me brindó la oportunidad de incorporarme a su grupo de trabajo y por ser mi asesor de servicio social otorgándome sus amplios conocimientos y apoyo para concluir mi informe de servicio social. Maestro Ricardo Campos Verduzco: Por brindarme su apoyo moral y académico para poder desempeñarme en el ámbito profesional. Dr. Antonio Valencia Hernández: Por otorgarme su amistad y amplios conocimientos académicos en el ámbito de estudios de censos visuales con videocámaras. Al Parque Nacional Huatulco.
Por todas las facilidades y el apoyo otorgado en cuanto a las instalaciones, información, embarcaciones y acceso a las bahías del parqué especialmente a la de Cacaluta, por permitir obtener toda la información necesaria para la realización de esta investigación e informe de servicio social.
A mis amigos y compañeros de la licenciatura en hidrobiología.
Anderson Alberto Ramírez Ramírez, Axel Tonathiu Martínez Morgan, Alonso I. Aguirre Peñalosa, Luis Gibran Juárez Hernández y demás compañeros quienes siempre me han brindado su amistad, apoyo y comprensión en los buenos y malos momentos, estaré siempre agradecido con ustedes.
Gracias a todos

47
BIBLIOGRAFÍA
Alevizon, W.S., R. Richardson, P. Pitts y G. Serviss. 1985. Coral zonation and
patterns of community structure in Bahamian reef fishes. Bull. Mar. Sci ., 36:304-318.
Allen, G.R. y D.R. Robertson. 1998. Peces del Pacífico Oriental Tropical .
CONABIO, Agrupación Sierra Madre y CEMEX. 327p. Amezcua-Linares, F. 1996. Peces demersales de la plataforma continental
del Pacífico Central Mexicano . Instituto de Ciencias del Mar y Limnología, UNAM. México, 184p.
Barrientos-Villalobos, J. 2000. Diversidad y abundancia de la ictiofauna de
los arrecifes coralinos del Parque Nacional Huatulc o. Tesis de licenciatura. Escuela de Biología. Universidad Autónoma de puebla, México. 54p.
Birkeland, C. (ed). 1997. Life and death of coral reefs . Chapman & Hall., New
York, 536p. Bohnsack, J.A. y S.P. Bannerot. 1986. A stationary visual census technique for
quantitatively assessing community structure of coral reef fishes. NOAA Tech. Rep., NMFS ., 41:1-15.
Brock, V.E. 1954. A critique of the visual census method for assessing coral
reef fish populations. Bull. Mar. Sci ., 32:269-276. Brock, V.E. 1982. A critique of the visual census method for assessing coral
reef fish population. Bull. Mar. Sci ., 32: 269-276. Buckley, R.M., y G.J. Hueckel. 1989. Analysis of visual transect for fish
assessment on artificial reef. Bull. Mar. Sci ., 44(2): 327-342. Caley, M.J. 1993. Predation, recruitment and the dynamics of communities of
coral reef fishes. Mar. Biol ., 117: 33-43. CONABIO, 2003. www.conabio.gob.mx DeMartini, E.E. y D. Roberts. 1982. An empirical test of biases in the rapid
visual technique for species-time censuses of reef fish assemblages. Mar. Biol ., 70:129-134.
Elorduy-Garay, F.J. y S.V. Jiménez-Gutiérrez. 2000. Metodología para el
estudio de los peces de arrecifes, p. 209-228. In: Aburto-Oropeza y C.A. Sánchez-Ortiz (eds). Recursos arrecifales del Golfo de California . Universidad Autónoma de Baja California Sur. México.

48
Fischer, J., W. Krupp, F. Schneider, W. Soomer, C. Carpenter, K.E. y V.H. Niem. 1995. Guía FAO para la identificación de especies para los fines de la pesca. Pacífico centro-oriental. Roma, FAO ., 2: 647-1200.
Flores-Coto, C y J.C Alvarez.1980. Estudios preliminares de distribución y
abundancia del ictioplancton en la laguna de términos, campeche. Mar. y Limnol. Univ. Nal. Autón. México., 7:1-27.
Fowler, A.J. 1987. The development of sampling strategies for population
studies of coral reef fishes. Coral Reefs ., 6:49-58. Froese, R. Y D. Pauly. Editors. 2004. Fishbase. World Wide web electronic
publication . www.fishbase.org, version (06/2004). Gauch, H.G. 1989. Multivariate análisis in comunity ecology . Cambridge
University Press. Glynn, P.W. y G.E . Leyte Morales. 1997. Coral reefs of Huatulco, West México:
reef development in upwelling Gulf of Tehuantepec. Rev.Biol.Trop ., 45:1033-1047.
Graus, R.R. y I.G. Macintyre. 1989. The zonation patterns of Caribbean coral
reefs as controlled by wave and light energy input, bathymetric setting and reef morphology: computer simulation experiments. Coral Reefs ., 8: 9-18.
Greene, L.E. y W.S. Alevizon. 1989. Comparative accuracies of visual
assessment method for coral reef fishes. Bull. Mar. Sci ., 44: 899-912. Helfman, G.S. 1978. Patterns of community structure in fishes: Summary and
overview. Environ. Biol. Fish ., 3: 129-149. Hobson, E.S. (1974).Diel feeding migration in tropical reef fishes. Helgoländer
Wissensch Meersunters ., 2: 361-370. INVEMAR, 2004. www.invemar.org. co/redcostera1/invemar/noticias. Juárez-Hernández, L. G. 2005. Estructura de la comunidad de peces en la
bahía Maguey, Huatulco, Oaxaca. Informe final de servicio social. Universidad Autónoma Metropolitana-Iztapalapa. México. 61 p.
Leyte-Morales, G. E. 1995. Zonas coralinas de Bahías de Huatulco. Res. III
Congreso Nacional de Zoología. Morelia, Michoacán. 68p. Leyte-Morales, G. E. 1996. Importancia biogeográfica de las comunidades
coralinas de Oaxaca, México. Memorias 1er Encuentro Regional de Investigación y Desarrollo Sustentable: Guerrero, Oaxaca y Chiapas. 63p.

49
Leyte-Morales, G. E. 1996. The coral communities of Oaxaca, México. VIII International Coral Reef Symposium. Panama, Panama. 117p.
Leyte-Morales, G.E. 1997. La colección de corales de la Universidad del Mar.
Cienc. Mar ., 1: 3-16. Lirman, D., Glynn.P.W., Baker. C y G. E. 2001. Leyte Morales. Combined
effects of three sequential stroms on the Huatulco coral reef tract, México. Bull. Mar. Sci ., 69(1): 267-278.
Ludwing, J.A. y Reynolds, J.E. 1988. Statical Ecology. A primer on methods
and computing . Wiley, New York. 337p. Luna-Vargas, E. L. I. 1993. Impacto de la infraestructura turística en Bahías
de Huatulco, Oaxaca. Tesis profesional de Licenciatura en Geógrafa. Univ. Nal. Autón. Méx . 189p.
Magurran, A.E. 1998. Ecological diversity and its measurement . Princeton
University Press. 179p. Margalef, R. (1958). Perspectives in Ecological Theory . Univ. Press,
Chicago, 240p. Martínez-Morgan, A. T. 2004. Estructura de la comunidad de peces en la
bahía Riscalillo, Huatulco, Oaxaca. Informe final de servicio social. Universidad Autónoma Metropolitana-Iztapalapa. México. 61 p.
McGehee, M.A. 1994. Correspondence between assemblages of coral reef
fishes and gradients of water motion, depth and substrate size off Puerto Rico. Mar. Ecol. Prog. Ser ., 105: 243-255.
Meekan, M.G., A.D.L. Steven y M.J.Fortin. 1995. Spatial patterns in the
distribution of damselfishes on a fringing coral reef. Coral Reefs ., 14:151-161.
Nelson, J.S. 1994. Fishes of the World . 3ra ed. Jhon Wiley & Sons, Inc. 600p. Ogden, J.C. y J.P. Ebersole. (1981). Scale and community structure of coral
reef fishes: A long-term study of a large artifitial reef. Mar. Ecol. Prog., Ser., 4: 97-03.
Oliver, J., M. Noordelous, Yusuf, Y., M. Tan, N. Nayan, C.Fou, and F.
Shahriyah. Reefbase: A Global Information System on Coral Ree fs. Available from: http://www.reefbase.org, version (2004).
Pielou, E.C. 1975. Mathemathical Diversity . 1-165. Wiley. New York. Pielou, E.C. 1977. Mathemathical Ecology . 1-385. Wiley. New York.

50
Pielou, E.C. 1984. The interpretation of ecological data. A primer on classification and ordination . Wiley, New York. 145p.
Ramírez-Ramírez, A. A. 2004. Estructura de la comunidad de peces en la
bahía San Agustín, Huatulco, Oaxaca. Informe final de servicio social. Universidad Autónoma Metropolitana-Iztapalapa. México. 53 p.
Ramos, S.E. ( 2002). Estructura de la Comunidad de peces en la Bahía la
Entrega, Huatulco, Oaxaca . Tesis para obtener el grado de maestría. UAMI.19p.
Ramos-Santiago, E. 2004. Estructura de la comunidad de peces en la Bahía
la Entrega, Huatulco, Oaxaca. Tesis de maestría. División de Ciencias Biológicas y de la Salud. Universidad Autónoma Metropolitana Unidad Iztapalapa, México. 119p.
Robins, C.R y G. C. Ray. 1986. A fiel guide to Atlantic coast fishes of North
America. Houghton Company, Boston, U.S.A. 354p. Sale, F.P. 1980. The ecology of coral reef fishes. Oceanogr. Mar. Biol. Ann.
Rev., 18: 367-421. Sale, F.P. 1991. Introduction: Ecology of Coral Reef Fishes, pp: 3-11 In : The
Ecology of Coral Reef Fishes. P. Sales (ed.) Acedemic Press, Inc., San Diego. C.A.
Sale, F.P., y B.J. Sharp. 1983. Correction for bias in visual transect censuses of
coral reef fish. Coral Reef ., 2: 37-42. Sale, F.P., y W.A. Douglas. 1981. Precision and accuracy of visual census
technique for fish assemblage of coral patch reef. Environ. Biol. Fishes ., 6: 333-339.
Sánchez, A.J. y A. Raz-Guzmán, "Arrecifes y comunidades coralinas"
(mapa), en Atlas Nacional de México, 1990. Secretaría del Medio Ambiente y Recursos Naturales. 2002. Programa de
Manejo del Parque Nacional Huatulco. Oaxaca, México . 164p. Secretaría del Medio Ambiente y Recursos Naturales. 2003. Programa de
Manejo del Parque Nacional Huatulco. Oaxaca. México . 208p. SEDUE, 2001. www.sedue.gob.mx Shannon, C.E., y W. Wiener, 1963. The mathematical Theory of
comunication . University of Illinois, 117p. Sielfeld K., Walter y Vargas. F., Mauricio.1996. Composición y estructura de la
ictiofauna demersal en la zona norte de Chile. Investig. mar ., 24:3-17.

51
Sorokin, I.Y. 1995. Coral Reef Ecology . Springer, New York. 465p. Thomson, D. A. 1987. Reef fishes of the sea of Cortez. The rocky-shore
fishes of the Gulf of California. The University of Arizona Press. Tucson. 310p.
Thomson, D. A., L. T. Findlye y A. N. Kerstitch. 1979. Reef fishes of the sea of
Cortez . University of Arizona Press. Arizona. 302p. Ward, J. 1963. Hierarchichal grouping to optimise an objetive function. Amer.
Statist. Ass ., 58: 236-244. Warner, R. R. y G. S. Hoffman. 1980. Local population size as a determinant of
mating system and sexual composition in two tropical marine fishes (Thalassoma spp.). Evolution ., 34(3):508-518.
Westmacott, S., Teleki, K., Wells, S and West, J. M. 2000. Manejo de arrecifes
de coral blanqueados o severamente dañados. UICN, Gland, Suiza y Cambridge, Reino Unido. 46p.
Williams, D. 1980. Dynamics of the pomacentrid community on small patch
reefs in one Tree Island Lagoon (Great Barrier Reef). Bull. Mar. Sci ., 30: 159-170.
Williams, D. 1986. Temporal variation in the structure of reef slope fish
communities (Central Great Barrier Reef): Short-term effects of Acanthaster planci infestation. Mar. Ecol. Prog. Ser ., 28: 157-164.
Williams, D. Y P. F. Sale. 1981. Spatial and temporal patterns of recruitment of
juvenil coral reef fishes to coral habitats within “One Tree Lagoon” Great Barrier Reef. Mar. Biol ., 65: 245-253.
Zar, J.H. 1988. Bioestadistical analisis . Prentice-Hall, Inc, New Jersey. 718p. Zar, J. H. 1996. Bioestatistical analysis. 3rd edn. Prentice Hall, Princeton, New
Jersey. 863p. FIRMA DEL ALUMNO VISTO B UENO DE LOS ASESORES _____________________ ______________________________ Criterios de evaluación





























