Embed Size (px)
Citation preview

Depuis les trois dernières décennies,de nombreux travaux ont été effectuéspour comprendre les mécanismes d’ac-tion des molécules phytochimiques,aussi bien dans les pays utilisant cou-ramment les plantes comme sources demédicaments que dans les pays indus-trialisés. Des bases de données informa-tiques se développent pour rassemblerles très nombreuses données ainsi obte-nues pour les rendre plus accessibles etdonc mieux exploitables (Scalbert et al2011).
Cependant, concernant les travauxréalisés chez les animaux d’élevage,en particulier les volailles, le nombred’études est comparativement faible parrapport aux travaux effectués en méde-cine, et actuellement, aucune base dedonnées ne regroupe l’ensemble de cestravaux. Dans cet article, les différenteshypothèses émises actuellement sur lesmodes d’action des principes actifs desphytomolécules chez les volailles ontété rassemblées (Lee et al 2004, Bakkaliet al 2008, Brenes et Roura 2010, Franzet al 2010, Wallace et al 2010, Abbas etal 2012). Ces hypothèses ont été renfor-cées par l’utilisation d’autres donnéesmoins couramment évoquées, issues enparticulier de travaux obtenus chez d’au-tres espèces animales lorsque ces donnéesn’existent pas chez les volailles. Lesexemples de mécanismes qui pourraientêtre impliqués ont été choisis principa-lement parmi les travaux concernant lestrois sources d’Huiles Essentielles (HE)les plus décrites dans la littérature pourleur utilisation en alimentation avicole et
présentées dans l’article précédent (Alle-man et al 2013), c’est-à-dire l’HE dethym, d’origan et de romarin, ainsi queparmi les travaux portant sur leurs com-posants majeurs.
La difficulté de compréhension deces mécanismes a plusieurs origines : lacomplexité de la composition chimi-que des HE (Alleman et al 2013), laprésence de composants mineurs pouvantagir comme modulateurs, la présence decomposés encore non identifiés et lesactions de synergie et/ou d’antagonismeentre ces différents composants. Ainsi,les HE utilisées telles quelles, sont plusefficaces que le mélange de leurs com-posants majeurs.
Des relations entre la structure chi-mique et les activités biologiques de cesmolécules sont proposées, sachant quedes différences d’activité peuvent êtreobservées entre isomères, ainsi qu’entrestéréo-isomères. Par ailleurs, les effetsbiologiques dépendent des doses utili-sées et des ratios entre les différentscomposants atteignant la cible biolo-gique, qui ne sont pas forcément lesmêmes que ceux présents dans l’HEajoutée à l’aliment. De plus, de nom-breux autres facteurs liés à l’animal ouà ses conditions d’élevage (Alleman etal 2013) peuvent également intervenir.L’âge de l’animal chez qui l’HE est uti-lisée a aussi son effet. Dans cet article,les différents mécanismes proposésactuellement pour expliquer les effetsobservés chez les animaux d’élevageseront présentés. Après avoir rappelé les
propriétés biologiques des HE, le deve-nir de leurs principes actifs après leurconsommation par l’animal est détaillé,puis leurs différentes modalités d’actionchez l’animal.
1 / Propriétés biologiques deshuiles essentielles
1.1 / Effets sur les microorga-nismes
a) Activités in vitro De nombreuses HE exercent, in vitro,
des effets négatifs sur la croissance desbactéries de l’appareil digestif, qui sontrapportés comme étant plus importantssur les bactéries Gram positif que surles bactéries Gram négatif (Dorman etDeans 2000, Burt 2004, Brenes et Roura2010, Ouwehand et al 2010). En effet, cesdernières sont protégées par une mem-brane externe supplémentaire au niveaude leur paroi à la différence des bacté-ries Gram positif dont la paroi n’estcomposée que d’un peptidoglycane, bienque plus épais que celui présent chez lesbactéries Gram négatif.
L’activité des composants des HEdépend de leur structure chimique. Lesterpènes oxygénés (carvacrol, thujonepar exemple) présentent une activitéantimicrobienne plus élevée que lesterpènes non oxygénés (α- et β-pinène,limonène par exemple) (Dorman etDeans 2000). De plus, le caractère lipo-
INRA Productions Animales, 2013, numéro 1
INRA Prod. Anim.,2013, 26 (1), 13-24
I. GABRIEL1, F. ALLEMAN2, V. DUFOURCQ2, F. PERRIN2, J.-F. GABARROU2
1 INRA, UR83 Recherches Avicoles, F-37380 Nouzilly, France2 Ecole d’Ingénieurs de Purpan, 75 Voie du TOEC, F-31076 Toulouse, France
Courriel : [email protected]
Utilisation des huiles essentiellesen alimentation des volailles.2. Hypothèses sur les modes d’actionimpliqués dans les effets observés
De nombreux produits contenant des huiles essentielles sont proposés pour améliorer lesperformances de croissance des animaux d’élevage, dont les volailles. Comme cela a été décritdans l’article précédent (Alleman et al 2013), des effets variables sont rapportés dans lalittérature. Une meilleure compréhension des modes d’action de ces composants devraitpermettre d’optimiser leur utilisation.

phile de leur squelette hydrocarbonéainsi que le caractère hydrophile deleurs groupes fonctionnels sont détermi-nants vis-à-vis de leur activité anti-microbienne. Ainsi, l’ordre d’activitéantimicrobienne de ces composés est lesuivant : phénols > aldéhydes > cétones> alcools > éthers > hydrocarbures. Parailleurs, d’autres caractéristiques molécu-laires interviennent : pour les phénols laprésence de groupe hydroxyle, pour lescomposés non phénoliques le type degroupe fonctionnel (alcényle > alkyle), etpour les hydrocarbures la structure cyclique.
Cependant des bactéries Gram positifpeuvent être résistantes à certaines HE,l’effet dépendant des souches bactérien-nes. De leurs côtés, des bactéries Gramnégatif peuvent être sensibles à l’actionde certaines HE. De plus, la sensibilitéde ces bactéries aux HE peut être aug-mentée par l’ajout de substances affec-tant l’intégrité de la membrane externe(Griffin et al 2001). Par ailleurs, il estimportant de noter qu’il existe des diffé-rences notables de résultats selon lesméthodes in vitro utilisées.
Des effets stimulants d’HE ou deleur composants sur le développementdes bactéries ont aussi été observés(Broudiscou et al 2007). L’inhibitiondu développement de certaines bacté-ries pourrait être bénéfique au dévelop-pement d’autres types de bactéries.Cependant, une stimulation directe dela croissance de bactéries a été rappor-tée comme dans le cas de trois souchesde différentes espèces bactériennes,Clostridium perfringens, Bifido-bacteriumbreve et Lactobacillus fermentum (Ouwe-hand et al 2010).
In vitro, certaines HE exercent aussiun effet délétère sur des parasites diges-tifs comme les oocystes d’Eimeria, res-ponsables de la coccidiose, et Histomo-nas meleagridis, le protozoaire respon-sable de l’histomonose chez la dinde(Zenner et al 2003, Remmal et al 2011).L’effet des composants phénoliquesd’HE sur les oocystes d’Eimeria pour-rait être lié à leur groupement phénol(Williams 1997).
Des HE comme celle de thym et sescomposants peuvent inhiber des cham-pignons producteurs de mycotoxinestoxiques pour l’animal (Mukherjee etNandi 1994, Soliman et Badeaa 2002).Les HE peuvent enfin avoir des actionscytotoxiques sur certains virus (Bakkaliet al 2008).
Cas des HE de thym, d’origan et deromarin et de leurs composants
Les HE de thym, d’origan et de roma-rin inhibent le développement de nom-breuses bactéries pathogènes comme
des streptocoques, des salmonelles, descoliformes ou C. perfringens associéaux entérites nécrotiques, mais inhi-bent peu celui de bactéries bénéfiquescomme des bifidobactéries ou des lacto-bacilles qu’elles peuvent même stimuler(Ouwehand et al 2010). Un effet négatifsur H. meleagridis a été observé avecun mélange d’HE de thym et de romarinainsi qu’avec du carvacrol (Grabenstei-ner et al 2007).
Les effets inhibiteurs de ces HE peu-vent s’expliquer en partie par l’actionde leurs composants majoritaires. Ainsi,le carvacrol et le thymol font partie desterpènes les plus efficaces contre lesbactéries, surtout les bactéries patho-gènes (Lee et al 2004, Broudiscou etal 2007, Ouwehand et al 2010). Cepen-dant, bien que les bactéries bénéfiquescomme les lactobacilles et les bifido-bactéries soient généralement résistantesà ces terpènes, certaines peuvent y êtresensibles. Ces terpènes peuvent aussiexercer un effet stimulateur de la crois-sance de certaines bactéries comme desbifidobactéries et des lactobacilles, maisaussi des salmonelles et C. perfringens(Ouwehand et al 2010). Le camphènequant à lui, possède un effet inhibi-teur important, alors que l’α-pinèneexerce une forte stimulation (Broudiscouet al 2007).
b) Mécanismes d’action Du fait du très grand nombre de com-
posants présents dans les HE avec desstructures chimiques très variées, cesmolécules peuvent avoir des cibles trèsdifférentes au niveau cellulaire (Dormanet Deans 2000, Burt 2004, Bakkali et al2008, Brenes et Roura 2010).
La majorité des études se sont con-centrées sur les effets au niveau desmembranes cellulaires et différenteshypothèses ont été émises. Ces molé-cules semblent entraîner des modifica-tions de la surface des bactéries simi-laires à celles observées avec d’autresagents antibactériens (La Storia et al2011). Elles pourraient traverser la paroides bactéries aussi bien Gram positifque Gram négatif et/ou entraîner desmodifications de cette paroi, variablesselon sa composition et sa structure(Sikkema et al 1995, Faleiro 2011, LaStoria et al 2011). Du fait de leurhydrophobicité, les terpénoïdes pour-raient alors agir sur la bicouche phos-pholipidique de la membrane cytoplas-mique bactérienne en s’immisçant àl’intérieur (Sikkema et al 1995). Ceciconduit alors à une modification desinteractions entre les composants de lamembrane, et des perturbations de sesfonctions accentuées dans le cas desterpènoïdes ayant des groupes fonction-nels comme des alcools et des aldéhy-
des phénoliques, avec un systèmed’électrons délocalisés (Ultee et al2002, Ben Arfa et al 2006). Ceci a denombreuses conséquences : une aug-mentation de la perméabilité mem-branaire, une fuite d’ions potassium etde protons avec une acidification dumilieu intracellulaire, un épuisementde la réserve intracellulaire d’ATP, unemodification de l’espace périplasmiquedes bactéries Gram négatif, une pertede contenu cellulaire, une coagulationdu contenu cytoplasmique, et une lysecellulaire (Faleiro 2011).
Par ailleurs, les composants des HEpeuvent posséder différents sites d’ac-tion au niveau intracellulaire en fonc-tion de leur structure chimique. Ils peu-vent agir sur le métabolisme bactérien, parexemple sur la synthèse de différentesprotéines (protéines de choc thermique,flagellines, protéines de la membraneexterne de bactérie Gram négatif, pro-téines impliquées dans le métabolismeou la division cellulaire), inhiber l’acti-vité d’enzymes en se complexant àelles (Burt 2004, Di Pasqua et al 2010,Faleiro 2011).
De plus, certaines HE apparaissentcomme étant de puissants inhibiteursdu système de communication entre lesbactéries jouant un rôle essentiel dansleur développement, ou dans la viabilitédes biofilms bactériens, un des méca-nismes impliqués dans la pathogénicité deces microorganismes, ou encore dans lasuppression de l’expression de leurs tox-ines (Faleiro 2011, Solorzano-Santos etMiranda-Novales 2012).
c) Réaction de résistance des bac-téries aux molécules à activité anti-biotique
Sachant que la recrudescence de larésistance des bactéries aux antibio-tiques est à l’origine de l’interdictionde l’emploi des Antibiotiques commeFacteurs de Croissance (AFC), et parconséquent de la nécessité de la re-cherche d’alternatives à leur utilisation,il est indispensable de connaître lesconséquences de ces alternatives faceà ces problèmes de santé animale ethumaine.
Comme pour les antibiotiques de syn-thèse, les molécules à activité antibio-tique issues de plantes peuvent conduireà la sélection de bactéries résistantes(Bakkali et al 2008). Ainsi B. cereuss’adapte au carvacrol lorsqu’il est pré-sent à des concentrations non létales. Ilen est de même pour des mélanges sim-ples composés de deux ou trois phyto-molécules. Par ailleurs, ces moléculesou des mélanges de quelques compo-sants peuvent augmenter la sensibilitéou la résistance de certaines bactéries
14 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
INRA Productions Animales, 2013, numéro 1

Utilisation des huiles essentielles en alimentation des volailles… / 15
à des antibiotiques de synthèse. Dansle cas de mélanges complexes, il estgénéralement considéré comme peuprobable que les bactéries puissentdévelopper une parade contre toutes lesmolécules. Cependant, certaines bacté-ries sont capables de surmonter l’effetinhibiteur des HE. Cette capacité derésistance doit donc être prise en comp-te lors de l’utilisation d’additifs à based’HE.
1.2 / Propriétés antioxydantes De nombreuses HE présentent une
propriété antioxydante (Chemat et al2007, encadré 1). En particulier, lesHE de thym et d’origan, et dans unemoindre mesure l’HE de romarin, présen-tent les activités antioxydantes les plusimportantes parmi les plantes aromati-ques du pourtour méditerranéen (Viuda-Martos et al 2010).
L’activité antioxydante des HE dépendde plusieurs caractéristiques structu-rales des molécules en relation avec leurpropriété d’oxydo-réduction, et estattribuée essentiellement à la forte réac-tivité des substituants des groupeshydroxyles (Ciz et al 2010). Les com-posés phénoliques, comme le thymol,le carvacrol, et l’eugénol font partiedes molécules des HE présentant lesplus fortes activités antioxydantes. Lethymol, avec un plus grand encombre-ment stérique du groupe phénol parrapport au carvacrol, possède une plusgrande activité. Les hydrocarburesmonocycliques, tels que le terpinolène,
l’α-terpinène, et le γ- terpinène, sontaussi des antioxydants fortement actifsdes HE. Les monoterpènes alcools,cétones, aldéhydes, hydrocarbures etéthers contribuent aussi à cette acti-vité. Ces composants d’HE agiraientcomme des éboueurs de radicaux libreset influenceraient les systèmes de défensein vivo antioxydants (Knas-müller et al2008).
Cependant, certaines HE peuvent, àforte concentration, perméabiliser lesmembranes des mitochondries libérantleurs espèces oxygénées réactives les-quelles peuvent convertir certains com-posés antioxydants d’HE en composéspro-oxydants affectant ainsi les celluleseucaryotes (Bakkali et al 2008).
1.3 / Détection par des récep-teurs spécifiques
Dans l’épithélium nasal des oiseauxcomme dans celui des mammifères,les molécules odorantes sont reconnuespar des récepteurs présents au niveaudes neurones olfactifs aboutissant aubulbe olfactif, en relation avec différen-tes structures du cerveau impliquéesdans le comportement (Gomez et Celii2008).
Au niveau de la cavité buccale, desrécepteurs gustatifs peuvent détecter lesmolécules composant les HE, bien qu’enquantité inférieure chez les oiseaux parrapport aux mammifères (Brenes etRoura 2010).
Au niveau de l’épithélium digestif,principalement au niveau de l’intestingrêle, les cellules entérochromaffinespossèdent des récepteurs olfactifs dontla stimulation par des molécules odo-rantes comme le thymol, entraînent lalibération de sérotonine, qui exerce dif-férentes actions sur le tractus digestif(Kidd et al 2008). Ces molécules odo-rantes peuvent aussi être détectées pardes récepteurs au niveau des neuronesentériques.
2 / Devenir des huiles essen-tielles dans le corps des ani-maux
Pour comprendre les mécanismes àl’origine des effets des HE observéschez les animaux, il est nécessaire deconnaître le devenir de leur constituantsdans le corps de l’animal lors de leurconsommation. Ces molécules subis-sent différents processus rassembléssous le terme de pharmacocinétique(cf. encadré 2).
Alors qu’il existe plusieurs études depharmacocinétique de composantsvégétaux effectuées chez des rongeursou chez l’Homme, elles sont très rareschez les animaux d’élevage, commeles volailles. A l’aide de ces différentesétudes et en tenant compte des particu-larités physiologiques des oiseaux, deshypothèses de devenir de ces moléculespeuvent être proposées (figure 1).
2.1 / Interactions des huiles essen-tielles avec les contenus, le micro-biote et le tube digestif
Après ingestion d’un aliment conte-nant des HE, leurs molécules constituti-ves non absorbées par l’intestin sonttransportés avec l’aliment, du jabot à lafin de l’intestin grêle, puis dans les caecapour la fraction liquide des contenus.
Les concentrations des HE dans letractus digestif sont probablementhétérogènes à l’intérieur même d’unsegment digestif du fait de la com-
INRA Productions Animales, 2013, numéro 1
Encadré 1. Radicaux libres et antioxydants.
Les radicaux libres et autres groupes chimiques oxygénés réactifs entraînent l’oxydationdes biomolécules comme les protéines, les acides aminés, les lipides insaturés et l’ADN,conduisant à des altérations moléculaires potentiellement néfastes. Or, du fait des processus métaboliques cellulaires des tissus animaux, des quantités impor-tantes de radicaux libres sont formées en permanence. Par ailleurs, le microbiote digestifcontribue à cette production de radicaux libres. L’emploi de rations enrichies en lipides pouren augmenter la teneur en énergie et fournir des teneurs importantes en acides gras polyin-saturés (en particulier ceux de la série n-3) dans les produits animaux dont la compositionen acides gras correspond mieux aux recommandations nutritionnelles pour l’Homme,augmente ce risque de stress oxydatif entraînant le dépôt de peroxydes dans le corps del’animal. De plus, dans le cas des animaux d’élevage, différentes sources de stress sontresponsables d’une augmentation de cette production de peroxydes, tels que les tempé-ratures élevées et les fortes densités d’élevage. Par ailleurs, dans le cas des volailles, lasélection génétique pour une croissance rapide, ainsi que le faible ratio entre le volumepulmonaire des animaux et leur poids vif, contribuent à ce stress oxydatif. L’organisme animal est équipé d’un système de défense qui peut éliminer les radicauxlibres présents dans presque toutes les cellules, bien qu’une grande différence de capacitéantioxydante existe selon les tissus. Cependant, un déséquilibre entre la production de radi-caux libres et leur élimination par les systèmes antioxydants peut avoir lieu et conduire à unphénomène appelé stress oxydatif. Ce stress oxydatif a des conséquences néfastes sur laphysiologie de l’animal, avec notamment des effets sur sa croissance, ainsi que sur sonimmunité. Dans ce cas, un apport alimentaire d’antioxydants permet d’en limiter les effets. Le rôle desantioxydants alimentaires dans la protection contre le stress oxydatif n’est cependant pasencore très bien compris. Différentes théories contradictoires sont avancées pour expliquerà la fois l’augmentation et la diminution des activités enzymatiques induites par les antioxy-dants naturels. De plus, différents marqueurs sont utilisés pour mesurer les réponses quipeuvent être très variables selon ces marqueurs.
Encadré 2. Pharmacocinétique.
C’est l’étude du devenir d’une substance active dans l’organisme. Elle comprend quatrephases, se déroulant successivement : l’Absorption, la Distribution, le Métabolisme tissulai-re, et l’Excrétion du principe actif et de ses métabolites. De ce fait, elle est désignée sousle nom de phase ADME.

16 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
plexité de son contenu qui est unmilieu hétérogène où interagissent denombreux composants (mélange dedifférents types de protéines, gluci-des, lipides, vitamines et minéraux)présentant des propriétés physico-chimiques différentes.
Les composants des HE peuvent agirsur le microbiote digestif (cf. encadré3), mais aussi être métabolisés parcelui-ci, et ce, dès le jabot pour lesoiseaux. Dans des simulations in vitrode contenus digestifs d’intestin grêle etde caecum de porc, des terpènes phéno-liques comme le thymol et le carvacrolpeuvent être dégradés alors que d’autresrestent stables. Cela dépend des milieuxdigestifs, la dégradation étant plusintense en présence de contenus de finde tube digestif probablement du faitde la plus forte activité microbienneet de la concentration décroissante deces molécules, les fortes concentrationsconduisant à des dégradations moinsimportantes (Michiels et al 2008). Lessensibilités variables à cette dégradation
pourraient être liées à leur structuremoléculaire (présence d’un cycle aro-matique, présence d’oxygène, localisa-tion des doubles liaisons) (Broudiscouet al 2007).
Les composants des HE peuvent aussisubir des modifications purement chi-miques liées aux conditions physico-chimiques rencontrées dans les milieuxdigestifs (Michiels et al 2008). Ainsi, auniveau des contenus de l’intestin grêleterminal possédant un milieu réducteur,les terpènes subissent probablementdes saturations des doubles liaisons,l’élimination d’oxygène et des réactionsd’ouverture des cycles aromatiques(Malecki 2008). Au contraire, au niveaudes muqueuses, en contact avec desmilieux plus oxydants, des réactionsd’oxydation pourraient avoir lieu.
2.2 / AbsorptionComme indiqué précédemment, du fait
de leur volatilité, certains composantsd’HE peuvent passer directement par
les voies nasales, puis, via les poumons,atteindre ainsi la circulation sanguine.
Pour la partie ingérée, contrairementaux molécules d’origine végétale autresque les composants d’HE, dont le ni-veau d’absorption peut être très diffé-rent même pour des structures proches,les composants d’HE sont considéréscomme étant généralement rapidementabsorbés du fait de leur petite taille etde leur caractère lipophile. Chez lesmammifères, leur absorption a lieu dèsla partie supérieure du tube digestif, auniveau de l’estomac et du duodénum(Michiels et al 2008). Chez les oiseaux,on peut supposer l’existence d’un pro-cessus d’absorption de ces moléculeslipophiles dès le jabot, du fait du tempsde rétention prolongé du bol alimentairedans cet organe (en moyenne de 2 à 4 h,voire plus), et de son épithélium struc-turellement similaire à celui du rumendes ruminants dont la perméabilité auxmolécules des HE a été montrée(Campbell et al 2010), bien qu’actuelle-ment à notre connaissance, il n’existepas de données à ce sujet.
Cependant, l’absorption des moléculesd’HE dépend également de la composi-tion chimique du bol alimentaire. Ainsi,une corrélation négative est observéeentre le degré d’absorption et la teneuren matière sèche du bol alimentaire(Michiels 2009). Or, celle-ci dépend elle-
INRA Productions Animales, 2013, numéro 1
Fraction liquide
Contenu digestif de l’intestin grêle
Phytomolécule
Caeca
Excréta
Contenu
Jabot
Proventricule
Gésier
Intestin grêle
Foie CœurVentricule
droit
CœurVentricule
gauchePoumonsSacs aériens
Reins
Autres tissus
Urine
Figure 1. Devenir potentiel des composants d’huiles essentielles utilisés dans l’alimentation chez l’oiseau.
Encadré 3. Le microbiote digestif.
Il s’agit de l’ensemble des microorganismes de l’appareil digestif (anciennement appelémicroflore digestive), présent dans les contenus digestifs et au niveau des muqueuses. Ilest modifiable par les conditions du tube digestif (microbiome) liées à la génétique de l’ani-mal et à son environnement dont son alimentation, et exerce de nombreux effets sur sonhôte, aussi bien au niveau digestif qu’au niveau global de l’animal.
Utilisation des huiles essentielles en alimentation des volailles… / 17
INRA Productions Animales, 2013, numéro 1
même des types d’alimentation et del’animal. Un contenu gastrique plusfluide pourrait augmenter les niveauxde solubilisation et d’émulsification desHE et expliquerait ainsi l’augmentationde leur absorption et de la vidange gas-trique de la phase liquide. En revanche,l’absorption des composants des HEpourrait être limitée par leur adsorptionsur la matière organique par des inter-actions hydrophobes ou par formationd’adduits entre les fonctions aldéhydesde certains terpènes et les protéines.
Par ailleurs, l’absorption des HEdépend de leur quantité dans le régime,une faible dose d’HE conduisant à uneabsorption non totale (Kohlert et al2002). Or, les doses utilisées en pratiquesont souvent plus faibles (50-100 ppmpour les phytomolécules les plus concen-trées) que celles utilisées dans les étudesexpérimentales (500 à 1 000 ppm).
De plus, d’autres composants d’origi-ne végétale associés aux HE peuventavoir une influence sur l’absorptioncomme, par exemple, la naringine et lapipérine qui stimulent l’absorption decertains xénobiotiques issus de plantes(Kang et al 2009). En revanche, l’ab-sorption des HE peut être différée versles parties postérieures du tractus diges-tif par certains procédés d’encapsula-tion des HE.
2.3 / Modifications des moléculesissues de plantes par le métabo-lisme de l’hôte
Comme tout xénobiotique, aprèsabsorption digestive, les phytomolécu-les sont métabolisées (par oxydation,hydrolyse, sulfatation, glycosylation…)dans le but d’être détoxifiées, au niveaude la muqueuse gastro-intestinale, puis,après passage dans le système porte, auniveau du foie. Ceci conduit à la forma-
tion de molécules plus solubles et doncplus facilement éliminables. Ces molé-cules peuvent exercer des activités bio-logiques différentes, bénéfiques parl’acquisition d’une plus grande effica-cité biologique, ou néfastes par l’acqui-sition d’un pouvoir de toxicité.
2.4 / Action au niveau des orga-nes cibles
Une fois transférées dans la circula-tion générale, ces molécules peuventatteindre et agir sur différents organesou tissus cibles comme l’appareil diges-tif, le système immunitaire, les muscles,le système nerveux, etc. Au niveau deces organes ou tissus, elles peuvent ànouveau être métabolisées.
2.5 / Elimination Comme pour tous les xénobiotiques,
différentes voies d’élimination sontpossibles via leur incorporation dansla bile, l’urine et l’air expiré après leurpassage dans le foie, les reins et lespoumons respectivement.
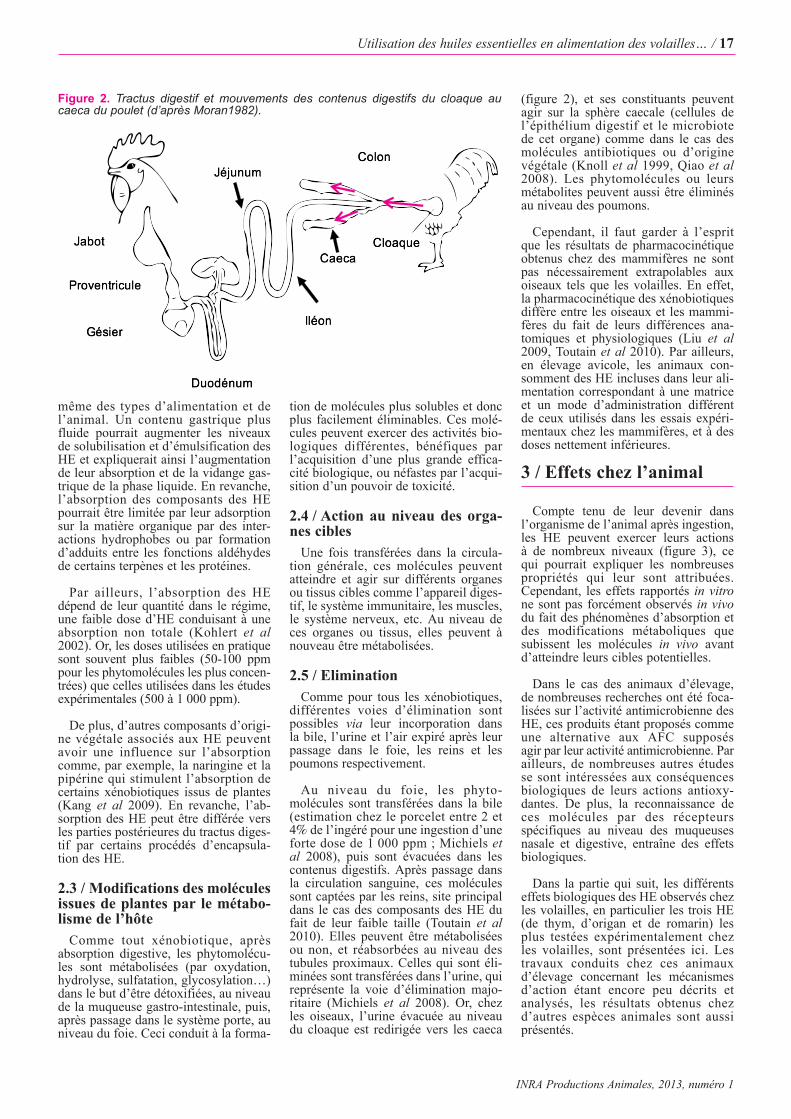
Au niveau du foie, les phyto-molécules sont transférées dans la bile(estimation chez le porcelet entre 2 et4% de l’ingéré pour une ingestion d’uneforte dose de 1 000 ppm ; Michiels etal 2008), puis sont évacuées dans lescontenus digestifs. Après passage dansla circulation sanguine, ces moléculessont captées par les reins, site principaldans le cas des composants des HE dufait de leur faible taille (Toutain et al2010). Elles peuvent être métaboliséesou non, et réabsorbées au niveau destubules proximaux. Celles qui sont éli-minées sont transférées dans l’urine, quireprésente la voie d’élimination majo-ritaire (Michiels et al 2008). Or, chezles oiseaux, l’urine évacuée au niveaudu cloaque est redirigée vers les caeca
(figure 2), et ses constituants peuventagir sur la sphère caecale (cellules del’épithélium digestif et le microbiotede cet organe) comme dans le cas desmolécules antibiotiques ou d’originevégétale (Knoll et al 1999, Qiao et al2008). Les phytomolécules ou leursmétabolites peuvent aussi être éliminésau niveau des poumons.
Cependant, il faut garder à l’espritque les résultats de pharmacocinétiqueobtenus chez des mammifères ne sontpas nécessairement extrapolables auxoiseaux tels que les volailles. En effet,la pharmacocinétique des xénobiotiquesdiffère entre les oiseaux et les mammi-fères du fait de leurs différences ana-tomiques et physiologiques (Liu et al2009, Toutain et al 2010). Par ailleurs,en élevage avicole, les animaux con-somment des HE incluses dans leur ali-mentation correspondant à une matriceet un mode d’administration différentde ceux utilisés dans les essais expéri-mentaux chez les mammifères, et à desdoses nettement inférieures.
3 / Effets chez l’animal
Compte tenu de leur devenir dansl’organisme de l’animal après ingestion,les HE peuvent exercer leurs actionsà de nombreux niveaux (figure 3), cequi pourrait expliquer les nombreusespropriétés qui leur sont attribuées.Cependant, les effets rapportés in vitrone sont pas forcément observés in vivodu fait des phénomènes d’absorption etdes modifications métaboliques quesubissent les molécules in vivo avantd’atteindre leurs cibles potentielles.
Dans le cas des animaux d’élevage,de nombreuses recherches ont été foca-lisées sur l’activité antimicrobienne desHE, ces produits étant proposés commeune alternative aux AFC supposésagir par leur activité antimicrobienne. Parailleurs, de nombreuses autres étudesse sont intéressées aux conséquencesbiologiques de leurs actions antioxy-dantes. De plus, la reconnaissance deces molécules par des récepteursspécifiques au niveau des muqueusesnasale et digestive, entraîne des effetsbiologiques.
Dans la partie qui suit, les différentseffets biologiques des HE observés chezles volailles, en particulier les trois HE(de thym, d’origan et de romarin) lesplus testées expérimentalement chezles volailles, sont présentées ici. Lestravaux conduits chez ces animauxd’élevage concernant les mécanismesd’action étant encore peu décrits etanalysés, les résultats obtenus chezd’autres espèces animales sont aussiprésentés.
Jabot
Proventricule
Gésier
Duodénum
Jéjunum
Iléon
Caeca
Colon
Cloaque
lCJ
elucirtnee
t
vorP
oobaJ C
oloC
aceaC
élI
munujé
euqaolC
no
r
douD
eisééGnoélI
munéd
Figure 2. Tractus digestif et mouvements des contenus digestifs du cloaque aucaeca du poulet (d’après Moran1982).

3.1 / Effets sur le microbiotedigestif
a) BactériesLes molécules des HE sont très sou-
vent considérées comme ayant uneaction directe sur le microbiote bacté-rien intestinal du fait de leur effet anti-bactérien observé in vitro. Cependant,non seulement les effets de ces molécu-les sont variables selon les méthodes invitro utilisées, mais les effets observésen milieu de culture peuvent aussi êtredifférents de ceux observés dans desmatrices complexes tels les contenusdigestifs. Ainsi, les concentrations d’a-gents antibactériens nécessaires pourexercer un effet dans les contenus diges-tifs (dont la teneur en matière organiqueest élevée) sont plus élevées que cellesnécessaires dans un milieu de culturepauvre en matière organique. Par exem-ple, dans le cas du carvacrol, une dosenettement supérieure de ce terpène estnécessaire pour obtenir un effet antibac-térien dans une matrice alimentaire parrapport à un milieu de culture, soit 1,5%et 0,1% respectivement (Kim et al1995).
De plus, comme décrit précédem-ment, les composants des HE étantabsorbés rapidement au niveau du tubedigestif, les concentrations dans lescontenus digestifs sont beaucoup plus
faibles que celles dans l’aliment. Ainsi,les teneurs en composants d’HE pré-sents in vivo dans un segment digestifcomme l’intestin grêle, ne correspon-dent pas à celles nécessaires pour inhi-ber les bactéries digestives. Par ailleurs,chez le poulet, le temps de transit dansce segment digestif étant relativementfaible (environ 1 heure), le temps decontact du microbiote et des HE estpeu important. Ceci conduit à penserque l’effet antibactérien direct n’est pasle phénomène principal expliquantl’action des HE au niveau de l’intestingrêle, sauf si l’absorption des HE estretardée par un procédé d’encapsulationapproprié. En revanche, les HE peuventexercer une action indirecte sur lemicrobiote par l’intermédiaire de mo-difications de la physiologie digestive,dont la digestion, comme on le verra plusloin, entraînant une modification desbiotopes digestifs comme les substratsdisponibles pour le microbiote cor-respondant aux composants non digé-rés par l’animal (glucides dont les fib-res, protéines, lipides…).
En revanche, au niveau des caeca, lasituation pourrait être différente pourdeux raisons, et permettre aux compo-sants d’HE d’exercer un effet anti-microbien. D’une part, comme indiquéprécédemment, les phytomolécules etleurs métabolites pourraient être plusconcentrés du fait du reflux urinaire
dans ces segments digestifs. D’autrepart, le temps de contact entre ces molé-cules et les bactéries est potentiellementplus long au niveau des caeca qu’auniveau de l’intestin grêle (environ 1heure) du fait d’une faible fréquence derenouvellement du contenu (1 à 2 foispar jour).
Ainsi, des modifications de micro-biote ont été observées dans l’intestingrêle avec des mélanges de produitsvégétaux contenant des composantsd’HE (Jamroz et al 2005, Tekeli et al2006), ainsi qu’une réduction des lésionsdues à C. perfringens (Timbermont et al2010). Mais il a été observé que lesmodifications sont plus importantesdans les caeca que dans l’iléon avec unmélange de thymol et de cinnamaldé-hyde (Tiihonen et al 2010) ou avec unmélange composé principalement d’HE(Guardia 2011).
Cependant, avec un mélange d’ex-traits végétaux contenant des compo-sants d’HE, une baisse de l’abondancede C. perfringens a été observée à lafois aux niveaux de l’intestin grêle etdes caeca (Brenes et Roura 2010). Avecce même type de mélange, des modifi-cations de profil bactérien dans ces deuxsegments digestifs ont été rapportées(Hume et al 2011). Par ailleurs, avec unautre mélange contenant de l’HE d’ori-gan ainsi que des extraits de curcuma et
18 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
INRA Productions Animales, 2013, numéro 1
Inhalation
Huile Essentielle
MoléculesDose
Matrice alimentaire
Ingestion
Récepteurs olfactifs (cavité nasale)
Système Nerveux Central
Système Nerveux Entérique
MicrobiotedigestifComportement
Bien-être Animal Santé animaleFonctionsdigestives
Sang Récepteurs olfactifs (TD)
Système immunitaire
Métabolisme
Appareil digestif
Volatilité
Figure 3. Les différentes possibilités d’action des phytomolécules chez l’animal.

de piment de Cayenne, une modifi-cation de la fonctionnalité bactériennemesurée par une augmentation d’uneactivité enzymatique (α-glucosidase) aété observée au niveau de l’iléon, maissans modification des activités bacté-riennes au niveau caecal (Mikulski et al2008).
Les différences d’effets selon les seg-ments digestifs pourraient aussi êtredues à la composition du microbiotedifférente selon ces biotopes (Gabrielet al 2005), avec des microorganismesayant des sensibilités différentes aux HE.
Cas des HE de thym, d’origan et deromarin et de leurs composants
Dans le cas de l’HE de thym utiliséechez le poulet, à une dose de 120 ppm,Tekeli et al (2006) n’ont pas observéd’effet sur le nombre de bactéries aéro-bies au niveau du jéjunum, alors qu’avecune dose de 1 000 ppm, Cross et al(2003, 2011) ont rapporté une réductiondu nombre de coliformes et une modi-fication des produits de fermentationdans les caeca, c'est-à-dire une baissede la proportion d’acides gras volatilsminoritaires (acides isobutyrique et iso-valérique). Avec de l’HE d’origan, uti-lisée à une dose de 120 ppm, Tekeli et al(2006) ont observé une augmentationdu nombre des coliformes dans l’intes-tin grêle, alors que Kirkpinar et al (2011),avec une dose de 300 ppm, n’ont pasobservé de modification du nombre delactobacilles, de coliformes et de strep-tocoques, mais une baisse significativedu nombre de C. perfringens. Avec unmélange d’HE d’origan et de thym, unebaisse de colonisation par des salmo-nelles dans le jabot et les caeca a été ob-servée (Koscova et al 2006). Par ailleurs,avec de la poudre de thym, une baissedu nombre de bactéries totales aérobiesainsi que de coliformes a été rapportéeau niveau de l’intestin grêle, ainsiqu’une baisse de C. perfringens dansles caeca. Cependant, ces effets n’ontpas été confirmés, de même qu’avec duromarin ou de l’origan (Abou-Sekken etal 2007, Brenes et Roura 2010).
Chez le porc, Michiels et al (2010b)n’ont pas observé d’effet significatif ducarvacrol à la dose de 500 ou de 2 000ppm sur le nombre de bactéries dansl’estomac et l’intestin grêle, mais ontdécrit un effet avec du thymol. Ainsi,avec 500 ppm de ce terpène, le pH descontenus de la partie terminale de l’in-testin grêle est diminué traduisant uneaugmentation des fermentations bacté-riennes. Avec 2 000 ppm, dans ce mêmesegment intestinal, ces auteurs ontobservé une augmentation du nombrede streptocoques et une modificationd’activité bactérienne traduite par uneaugmentation de la concentration en
propionate, ces effets dépendant du typede support utilisé pour ce composant(célite ou cellulose).
Les études des effets de régimes en-richis en HE sur le microbiote digestifdu poulet ont, dans la grande majoritédes cas, été conduites en ciblant destypes bactériens cultivables (principale-ment aérobies) alors que la majorité desbactéries du tube digestif des oiseauxn’est pas cultivable (Gabriel et al 2005).Ces travaux ne permettent donc pasd’obtenir une image de l’effet réel glo-bal des HE sur le microbiote. Cependant,l’utilisation d’approches indépendantesde la culture (approches moléculaires)pour étudier les modifications du micro-biote en fonction de facteurs commel’utilisation d’additifs alimentaires com-mence à se développer chez le pouletdepuis une dizaine d’années. Lestravaux concernant les effets des HE surle microbiote digestif sont encore peunombreux (Tiihonen et al 2010, Guardia2011, Hume et al 2011). De plus, ilsnécessitent encore des évolutions tellesque la prise en compte de la fortevariabilité inter-individuelle, ainsi quedes développements méthodologiques.Cette modulation du microbiote bac-térien digestif pourrait être responsabled’un effet indirect des composants desHE sur le tractus digestif de l’animalet son système immunitaire, du fait del’action des bactéries digestives sur letube digestif (Gabriel et al 2005).
b) Parasites digestifsDans le cas de la coccidiose du poulet,plusieurs travaux ont montré un effetbénéfique de produits végétaux liés àleur activité antioxydante au niveau destissus lésés par les parasites respon-sables d’un stress oxydatif conduisantau phénomène de lipoperoxydation, maistrès peu d’études ont porté sur l’effetpropre des HE (Allen et al 1998,Crévieu-Gabriel et Naciri 2001, Abbaset al 2012).
Cas des HE de thym, d’origan et deromarin et de leurs composants
Un effet anticoccidien de l’HE d’ori-gan a été montré contre l’espèceEimeria tenella ciblant les caeca (DaSilva et al 2009, Brenes et Roura 2010).
L’association de camphre et du 1,8-cinéol (appelé aussi eucalyptol) exerceglobalement des effets bénéfiques surune espèce courante d’Eimeria ci-blant le duodénum (E. acervulina) et surE. tenella alors que le cinéol apporté seulentraîne une augmentation des lésionsdues à E. acervulina (Allen et al 1998).Avec un mélange d’HE contenant prin-cipalement du carvacrol, Lillehoj et al(2011) ont observé une réduction de la
perte de poids de l’animal consécutiveà une infection par E. acervulina, ainsiqu’une réduction de la quantitéd’oocystes excrétés. L’effet bénéfiquedes HE serait lié en partie à leur effetdélétère sur ces parasites (comme indi-qué précédemment) ainsi qu’à un effetsur l’immunité. Cet effet bénéfique per-met la limitation du développement decette pathologie et de ce fait celle liéeà C. perfringens, cette bactérie se multi-pliant en situation de coccidiose.
Dans le cas de l’histomonose (infec-tion des caeca par le protozoaire Histo-manas meleagridis), Hafez et Hauck(2006) ont rapporté qu’un mélanged’HE de romarin, de cannelle, d’ail etde citron protège en partie les dindes.Ceci pourrait être lié en partie à l’effetlétal de ces HE sur ces parasites, ainsique sur certaines bactéries digestivesintervenant dans le développement dece parasite dans les caeca.
3.2 / Effets sur les tissus de l’ani-mal
Dans un objectif de santé humaine,les effets de nombreuses phytomo-lécules ont été étudiés, aussi bien auniveau global qu’aux niveaux cellulaireet moléculaire, en mesurant les modifi-cations de l’expression des gènes et dustatut des protéines et des métabolitesproduits par approches transcripto-mique, protéomique et métabolomique(Ovesná et al 2008). Ces travaux surles mécanismes d’action des phyto-molécules à l’origine des effets sur lesfonctions physiologiques sont en pleindéveloppement sur rongeurs commemodèles animaux pour l’Homme.Cependant, concernant les animauxd’élevage, il existe actuellement trèspeu d’études de ce type bien que lespremières apparaissent (Lillehoj et al2011). Dans la très grande majoritédes études disponibles actuellementchez les animaux d’élevage dont lesvolailles, seul un nombre limité demarqueurs biologiques ont été pris encompte, comme nous allons le voir dansla suite de ce texte.
a) Effets sur la sphère oro-nasaleComme les mammifères, les volailles
sont sensibles aux goûts et aux odeurs.Cependant, il semble que, contraire-ment aux mammifères qui peuvent ré-pondre aux HE par le rejet, les oiseauxsoient plus tolérants aux effets sen-soriels des HE (Gomez et Celii 2008,Brenes et Roura 2010). Ceci provientprobablement du fait qu’au niveau de lasphère oro-nasale, le nombre et le typede récepteurs olfactifs et gustatifs sontdifférents entre les mammifères et lesoiseaux.
Utilisation des huiles essentielles en alimentation des volailles… / 19
INRA Productions Animales, 2013, numéro 1

b) Effets antioxydants Il existe peu d’études concernant les
effets d’antioxydants systémiques desproduits d’origine végétale comme lesHE chez les volailles, l’essentiel desétudes de ces effets portant sur la qua-lité de la viande. En effet, l’emploi detelles sources d’antioxydants permet destabiliser les viandes de volailles face aurisque de lipoperoxydation, ces viandesétant riches en acides gras insaturés.
Cas des HE de thym, d’origan et deromarin et de leurs composants
Parmi les études menées chez le pouletsur l’effet antioxydant systémique desHE issues de plantes, une augmentationde la concentration en α-tocophérol a étérapportée dans le sérum avec l’utilisationde 100 ppm d’HE d’origan (Brenes etRoura 2010). Chez le rat, l’utilisation d’HEde thym ou de thymol peut entraîner unenrichissement en acides gras polyinsa-turés des phospholipides de différentsorganes (foie, cerveau, rein et cœur)(Lee et al 2004). Avec l’emploi de 500ppm de poudre de romarin, Spernakovaet al (2007) ont observé chez le poulet,une augmentation de la concentrationen α-tocophérol sanguin. Avec une fortedose de poudre d’origan ou de romarin(20 000 ppm) ou avec l’association deces deux plantes, Botsoglou et al (2009)ont rapporté, chez le rat, une diminutionde l’état d’oxydation induit par un stressoxydant expérimental.
Concernant l’impact des produits àbase de plantes sur l’état d’oxydationdes produits animaux, les HE de thym,d’origan ou de romarin exercent chezle poulet des propriétés antioxydantesdans la viande et le gras abdominal(Bolukbasi et al 2006, Brenes et Roura2010). Les composés antioxydants duthym dans les suppléments alimentairespeuvent chez la poule pondeuse aussi setransférer à l’œuf (Lee et al 2004).
c) Effets sur l’immunité Les HE exercent des effets anti-
inflammatoires et stimulateurs del’immunité spécifique. L’action anti-inflammatoire des HE peut s’avérerparticulièrement intéressante chez lesanimaux d’élevage du fait de l’état deperpétuelle inflammation de l’intestinconsécutive à la présence du microbiotedigestif (Gabriel et al 2005), pouvantdiminuer le coût métabolique que repré-sente cette stimulation de l’immunité.L’inflammation est un processus haute-ment complexe. Il s’accompagne, entreautre, d’une augmentation du stressoxydatif (John et al 2011) qui pourraitêtre une des cibles des HE comme indi-qué précédemment. Les HE possède-raient aussi des actions de stimulationdu système immunitaire spécifique.
Cas des HE de thym, d’origan et deromarin et de leurs composants
Chez le poulet de chair recevant desrations enrichies en HE de thym, Najafiet Torki (2010) ont observé une baissedu nombre des hétérophiles et une aug-mentation de celui des lymphocytessanguins. L’HE d’origan possède un effetanti-inflammatoire in vitro (Ocana-Fuentes et al 2010). Par ailleurs, Basma-cioglu et al (2010) ont rapporté que cetteHE conduit à une augmentation du titred’anticorps contre le Virus de Newcastle.L’HE de romarin exerce un effet anti-inflammatoire chez le rat (Takaki et al2008), ce même type d’effet étant observéchez la souris par la combinaison d’HEde thym et d’origan (Bukovska et al 2007).
Le carvacrol et le thymol ont une acti-vité inhibitrice de la cyclo-oxygénase,inhibant ainsi la production de prosta-glandine E2 responsable de la réactioninflammatoire (Marsik et al 2005, Landaet al 2009). Chez le porc, Michiels et al(2010b) ont observé, avec du thymol,une baisse du nombre de lymphocytesintra-épithéliaux de la muqueuse del’intestin grêle. Chez le poulet, unemodification de l’expression de gènesimpliqués dans l’immunité au niveaudes lymphocytes intra-épithéliaux duduodénum a été rapportée avec l’utili-sation de carvacrol (Lillehoj et al 2011).
d) Effet sur l’appareil digestif et lesprocessus de digestion
Au niveau de l’appareil digestif, deseffets des HE ont été rapportés sur latranscription des gènes des tissus intes-tinaux. Ainsi, Kroismayr et al (2008)ont observé chez le porcelet, une baissedu niveau d’expression de marqueursprotéiques de l’apoptose aux niveaux dujéjunum et de l’iléon.
Les effets bénéfiques observés surl’épithélium digestif d’extraits végétauxcomme les HE ayant une action anti-oxydante, pourraient s’expliquer par laréduction du stress oxydatif respon-sable d’une dégradation de la barrièrede l’épithélium gastro-intestinal (Johnet al 2011). Les extraits végétaux peu-vent aussi permettre une réduction deslésions au niveau de l’appareil digestifconsécutives à des entérites bactérien-nes (Acosta et al 2008).
De plus, de nombreux composantsd’HE exercent une action stimulantesur les processus digestifs avec une sti-mulation des sécrétions salivaires, gas-triques, biliaires et pancréatiques, et desactivités des enzymes intestinales, ainsique des effets stimulants de la motricitégastrique et intestinale (Kidd et al 2008,Brenes et Roura 2010). Des améliora-tions de la digestion des aliments ont
ainsi été observées avec un mélangecommercial de composants d’HE (Caoet al 2010, Bravo et al 2011). Les HEpeuvent aussi stimuler la productionintestinale de mucus, limitant l’adhé-sion des microorganismes pathogènes.
Par ailleurs, ces modifications auniveau digestif peuvent entraîner deseffets bénéfiques sur l’état de santé glo-bal des animaux. Ainsi, une diminutionde la quantité d’eau éliminée dans lesexcréta avec l’apport de composantsd’HE (Lee et al 2004) pourrait amélio-rer la qualité des litières et donc la santédes animaux en réduisant les pododer-matites et les troubles respiratoires.
Cas des HE de thym, d’origan et deromarin et de leurs composants
Chez le poulet, avec de l’HE de thym,Demir et al (2005) ont observé unebaisse de la profondeur des cryptes del’iléon. Avec de l’HE d’origan, Tekeli etal (2006) ont rapporté une augmenta-tion du poids du jéjunum et Da Silva etal (2009) une baisse de la profondeurdes cryptes reflétant une diminution durenouvellement cellulaire intestinal,celle-ci entraînant donc un coût méta-bolique moindre pour le tissu intestinal,et sans modification de la hauteur desvillosités. Basmacioglu et al (2010) ontobservé une augmentation de l’activitéde la chymotrypsine et une baisse de laviscosité dans l’iléon, ce qui pourraitcontribuer à l’augmentation de la diges-tibilité fécale des protéines observéechez le poulet. Avec de l’HE de thym,une activité anti-spasmodique a étéobservée au niveau de l’iléon chez le rat(Begrow et al 2010). Avec de l’HE dethym ou de romarin seule incluse à1 000 ppm, Cross et al (2007) n’ont pasobservé de différence de digestibilitéfécale de la matière sèche et de l’énergie,mais avec 5 000 ppm d’un mélange d’unextrait contenant du thym et du romarin,Hernandez et al (2004) ont rapporté uneaugmentation de la digestibilité fécalede la matière sèche et des lipides, ainsique de la digestibilité iléale de la matièresèche et de l’amidon.
Quelques travaux ont été effectuéspour analyser les effets des moléculesconstitutives de ces HE. Avec du cinéol,il a été observé chez le rat des propriétésde protection de la muqueuse gastrique,avec réduction du volume de « jus »gastrique et de la production d’acides(Santos et Rao 2001). Il semblerait queles effets antioxydants et d’inhibitionde la lipoxygénase du cinéol soient àl’origine de son effet gastro protecteur.Dans le cas du thymol, une réduction dela sécrétion d’ion chlorure a été observéeau niveau du tissu intestinal du porcelet,ainsi qu’une réduction de l’absorptionactive de nutriments (Michiels et al
20 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
INRA Productions Animales, 2013, numéro 1

2010a). Avec cette molécule, il n’a pasété observé de modification de l’activitédes enzymes digestives pancréatiques,alors que celle de l’amylase a augmentédans les contenus digestifs avec l’em-ploi d’un mélange contenant du thymolchez le poulet de 21 j, mais pas de 40 j(Lee et al 2004). Le camphre aurait uneffet stimulateur sur la sécrétion bili-aire chez l’Homme (Petracca et Cavaz-zuti 1983). Avec du carvacrol ou du thy-mol, une augmentation du rapportvillosité/crypte a été observée dans l’in-testin grêle distal du porcelet, pouvantconférer de meilleures capacités diges-tives (Michiels et al 2010b). Avec ducinéol, du thymol ou du carvacrol, uneactivité anti-spasmodique a été obser-vée chez des rongeurs (Magalhaes etal 1998, Begrow et al 2010). Avec unmélange d’HE riche en carvacrol, uneaugmentation de la quantité de mucusdans le proventricule et dans le jéjunuma été rapportée chez le poulet, conférantune protection de la muqueuse (Breneset Roura 2010). Ce mélange a modifié lataille des cryptes du jéjunum en fonctiondu type de régime, soit à la baisse avecl’emploi d’un régime maïs, soit au con-traire à la hausse avec l’emploi d’unrégime blé-orge.
e) Effets sur le métabolisme Les HE auraient un effet sur le méta-
bolisme lipidique, avec une régulationdu dépôt des tissus adipeux chez lesmammifères (Brenes et Roura 2010).Chez les volailles, les travaux sont peunombreux.
Une meilleure efficacité protéiquea été rapportée avec de la marjolaineou du basilic (Osman et al 2010), ainsiqu’une amélioration de la rétention azotéeavec un mélange de thymol et de cinna-maldéhyde (Cao et al 2010). Cependantavec un mélange commercial de car-vacrol, de cinnamaldéhyde et d’oléoré-sine de capsicum (constituée majori-tairement de capsaïcine), l’utilisationdes protéines n’est pas apparue modi-fiée de façon significative (Muhl etLiebert 2007).
Cas des HE de thym, d’origan et deromarin et de leurs composants
Avec de l’HE de thym, Bolukbasi etal (2006) ont observé chez le poulet uneaugmentation des niveaux plasmatiquesde triglycérides, de LDL-cholestérol etde HDL-cholestérol, mais ceci n’a pasété confirmé par les travaux de Tekeli etal (2006) et de Najafi et Torki (2010).
Avec de l’HE d’origan, Basmaciogluet al (2010) ont rapporté une augmenta-tion de la concentration plasmatique decholestérol, bien que Tekeli et al (2006)n’aient pas observé de différence.
Les HE peuvent exercer des effetspositifs sur le métabolisme osseuxcomme rapporté avec les HE de romarinet de thym (Putnam et al 2007), cequi pourrait améliorer l’état généraldes volailles à croissance rapide. Parailleurs, les HE et leur composants peu-vent avoir des effets sur le métabolismedes xénobiotiques, comme cela a étéobservé chez la souris avec du thymolou du carvacrol (Sasaki et al 2005).
f) Effets sur le système nerveux et lecomportement
Les composants des HE peuvent agirsur le système nerveux, par l’intermé-diaire de deux voies, la voie sanguine etla voie nerveuse. Ils peuvent atteindre lacirculation sanguine directement aprèsinhalation et passage par la voie pulmo-naire ou indirectement après ingestion,absorption et métabolisation, puis dufait de leur faible taille et de leur lipo-solubilité, traverser la barrière sang/cerveau. Ils peuvent aussi agir aprèsactivation des récepteurs olfactifs auniveau nasal.
Cas des HE de thym, d’origan et deromarin et de leurs composants
Chez les rongeurs, il a été observéque les HE de thym, de romarin et d’ori-gan peuvent bloquer les sensations dedouleur (Takaki et al 2008, Taherian etal 2009, Mikaili et al 2010), ce qui peutêtre favorable au bien-être des animaux.Par ailleurs, des effets analgésiques demolécules composant ces HE, commel’α-pinène, le cinéol, le thymol et le car-vacrol ont aussi été observés chez lesrongeurs (de Sousa 2011). Ces effetspeuvent être liés à la phase inflamma-toire de la douleur ou à l’activation dedifférents récepteurs, comme les récep-teurs opioïdes. De plus, des composéstels que le thymol et le carvacrol possè-dent des propriétés inhibitrices del’acétylcholinestérase (Jukic et al 2007),conduisant à une activation prolongéedes récepteurs cholinergiques.
Les HE et leurs composants exercentaussi des effets sur le comportementanimal. Ainsi, chez la souris, l’ingestionde carvacrol a entraîné un effet anxioly-tique (Melo et al 2010) et l’inhalationd’HE de thym a exercé un effet sédatif(Lim et al 2005). Au contraire, il a étéobservé une augmentation de l’activitélocomotrice lors de l’inhalation ou l’in-gestion d’HE de romarin par la souris(Kovar et al 1987).
Conclusion
Les composants des HE présententdonc plusieurs types d’activités biolo-giques, antimicrobiennes, antioxydantes
et stimulatrices de récepteurs spéci-fiques. L’action de ces molécules chezl’animal dépend à la fois de leur phar-macocinétique et des conditions rencon-trées in vivo. En effet, le devenir de cesmolécules chez l’animal lors de leurconsommation peut être très variableselon la composition de l’aliment, leurmode de présentation (sous forme libreou protégée) et la dose utilisée. Ellespeuvent ainsi être présentes au niveau dela sphère olfactive, de la sphère diges-tive ou dans la circulation sanguine aprèsleur absorption dans l’appareil digestif.Ces molécules ont donc de nombreusescibles potentielles d’action au niveau del’animal, tant au niveau digestif quesystémique. Ainsi, elles peuvent exercerdiverses actions, tant sur le microbiotedigestif, que sur l’animal lui-même àdifférents niveaux (état d’oxydation desdifférents tissus ; immunité et en parti-culier l’inflammation ; fonctionnementdigestif, métabolisme global ; systèmenerveux ; comportement). De plus, lesdifférents composants des HE peuventexercer des actions synergiques permet-tant de réduire leur taux d’incorporationdans les régimes et donc le coût globalde ces additifs. Ces molécules représen-tent donc un grand potentiel pour leurutilisation chez l’animal d’élevage.
Ainsi, de nombreux travaux rappor-tent des effets bénéfiques de mélangesde ces composants chez les oiseauxd’élevage. Cependant ces effets ne sontpas toujours observés, ils dépendent desmélanges utilisés, ainsi que des condi-tions rencontrées dans les milieux bio-logiques de l’animal, qui dépendent del’état physiologique de celui-ci, lui-même dépendant de la génétique et del’âge de l’animal, ainsi que de son envi-ronnement d’élevage dont son alimenta-tion. Par ailleurs, chez les animaux derente, la grande majorité des études effec-tuées jusqu’à présent a concerné un nom-bre limité de marqueurs biologiquespotentiellement modifiés par ces com-posants. Les mécanismes d’action pré-cis au niveau moléculaire ne sont doncpas connus. De plus, dans la grandemajorité de ces études, la compositionexacte des mélanges utilisés n’est pasdéterminée ou diffusée. Il n’est doncpas possible actuellement de définir, àpartir des données disponibles de lalittérature, les différentes conditionsdans lesquelles des mélanges d’HEsont efficaces, ou de déterminer la com-position d’un mélange pour une actionet des conditions d’élevage définies. Enparticulier, dans les cas des oiseaux, ledevenir de ces composants après inges-tion n’ayant pas encore fait l’objet d’étu-des, les cibles potentielles de ces molé-cules ne sont pas réellement connues.
A l’avenir, des travaux sont doncnécessaires pour déterminer la pharma-
Utilisation des huiles essentielles en alimentation des volailles… / 21
INRA Productions Animales, 2013, numéro 1

cocinétique de ces composants, ainsique leurs effets chez l’oiseau, en parti-culier à l’aide d’approches non cibléesaussi bien sur les tissus animaux que surle microbiote digestif. Ceci permettra demettre en évidence des mécanismes
d’action non encore mis à jour. Cesconnaissances participeront à l’optimi-sation de l’utilisation en alimentationanimale de ces composants en les com-binant entre eux, voire à d’autres addi-tifs comme les prébiotiques, les probio-
tiques, les enzymes, les acides orga-niques, les composants d’origine miné-rale (argile, charbon), les fibres non fer-mentescibles... pour tirer profit de leursactions synergiques.
22 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
INRA Productions Animales, 2013, numéro 1
Références
Abbas R.Z., Colwell D.D., Gilleard J., 2012.Botanicals: an alternative approach for thecontrol of avian coccidiosis. World Poult. Sci.J., 68, 203-215.
Abou-Sekken M.S., Moustafa K.E.M.E.,Elalfy T.S., 2007. Effect of fennel, thyme andprobiotic as feed additives on the performanceand the microbial content of the intestine ofMuscovy ducks. Egypt. Poult. Sci. J., 27, 1009-1029.
Acosta Y., Acosta A., Pasteiner S., Rodri-guez B., Mohnl M., 2008. Effect of a probioticand a phytobiotic blend on the performance,health status and carcass yield of broilers.Cuban J. Agric. Sci., 42, 179-184.
Alleman F., Gabriel I., Dufourcq V., PerrinF., Gabarrou J.F., 2013. Utilisation des huilesessentielles en alimentation des volailles. 1.Performances de croissance et réglementation.INRA Prod. Anim., 26, 1, 3-12.
Allen P.C., Danforth H.D., Augustine P.C.,1998. Dietary modulation of avian coccidiosis.Int. J. Parasitol., 28, 1131-1140.
Bakkali F., Averbeck S., Averbeck D.,Idaomar M., 2008. Biological effects of essen-tial oils - A review. Food Chem. Toxicol., 46,446-475.
Basmacioglu Malayoglu H., Baysal S.,Misirlioglu Z., Polat M., Yilmaz H., Turan N.,2010. Effects of oregano essential oil with orwithout feed enzymes on growth performance,digestive enzyme, nutrient digestibility, lipidmetabolism and immune response of broilersfed on wheat-soybean meal diets. Br. Poult.Sci., 51, 67-80.
Begrow F., Engelbertz J., Feistel B.,Lehnfeld R., Bauer K., Verspohl E.J., 2010.Impact of thymol in thyme extracts on theirantispasmodic action and ciliary clearance.Planta Med., 76, 311-318.
Ben Arfa A., Combes S., Preziosi-Belloy L.,Gontard N., Chalier P., 2006. Antimicrobialactivity of carvacrol related to its chemicalstructure. Lett. Appl. Microbiol., 43, 149-154.
Bolukbasi S.C., Erhan M.K., Ozkan A.,2006. Effect of dietary thyme oil and vitamin Eon growth, lipid oxidation, meat fatty acidcomposition and serum lipoproteins of broilers.S. Afr. J. Anim. Sci., 36, 189-196.
Botsoglou N.A., Taitzoglou I.A., BotsoglouE., Zervos I., Kokoli A., Christaki E.,Nikolaidis E., 2009. Effect of long-term dietaryadministration of oregano and rosemary on theantioxidant status of rat serum, liver, kidneyand heart after carbon tetrachloride-inducedoxidative stress. J. Sci. Food Agric., 89, 1397-1406.
Bravo D., Utterback P., Parsons C.M., 2011.Evaluation of a mixture of carvacrol, cinnamal-dehyde, and capsicum oleoresin for improvinggrowth performance and metabolizable energyin broiler chicks fed corn and soybean meal.J. Appl. Poultry Res., 20, 115-120.
Brenes A., Roura E., 2010. Essential oilsin poultry nutrition: main effects and modes ofaction. Anim. Feed Sci. Technol., 158, 1-14.
Broudiscou L.P., Cornu A., Rouzeau A.,2007. In vitro degradation of 10 mono- andsesquiterpenes of plant origin by caprinerumen microorganisms. J. Sci. Food Agric., 87,1653-1658.
Bukovska A., Cikos S., Juhas S., Il'kova G.,Rehak P., Koppel J., 2007. Effects of a combi-nation of thyme and oregano essential oils onTNBS-induced colitis in mice. Mediat.Inflamm., doi : 10.1155/2007/23296., 9p.
Burt S., 2004. Essential oils: their antibacte-rial properties and potential applications infoods - A review. Int. J. Food Microbiol., 94,223-253.
Campbell E.J., Frost R.A., Mosley T.K.,Mosley J.C., Lupton C.J., Taylor C.A., Jr.Walker J.W., Waldron D.F., Musser J., 2010.Pharmacokinetic differences in exposure tocamphor after intraruminal dosing in selec-tively bred lines of goats. J. Anim. Sci., 88,2620-2626.
Cao P.H., Li F.D., Li Y.F., Ru Y.J., Peron A.,Schulze H., Bento H., 2010. Effect of essentialoils and feed enzymes on performance andnutrient utilization in broilers fed a corn/soy-based diet. Int. J. Poult. Sci., 9, 749-755.
Chemat F., Vian M.A., Dangles O., 2007.Essential oils as antioxidants. Int. J. Essent. OilTher., 1, 4-15.
Ciz M., Cizova H., Denev P., KratchanovaM., Slavov A., Lojek A., 2010. Different meth-ods for control and comparison of the anti-oxidant properties of vegetables. Food Control,21, 518-523.
Crévieu-Gabriel I., Naciri M., 2001. Effet del'alimentation sur les coccidioses chez le poulet(Dietary effect on chicken coccidiosis). INRAProd. Anim., 14, 231-246.
Cross D.E., Svoboda K., McDevitt R.M.,Acamovic T., 2003. The performance of chick-ens fed diets with and without thyme oil andenzymes. Br. Poult. Sci., 44, S18-S19.
Cross D.E., McDevitt R.M., Hillman K.,Acamovic T., 2007. The effect of herbs andtheir associated essential oils on performance,dietary digestibility and gut microflora inchickens from 7 to 28 days of age. Br. Poult.Sci., 48, 496-506.
Cross D.E., McDevitt R.M., Acamovic T.,2011. Herbs, thyme essential oil and condensedtannin extracts as dietary supplements forbroilers, and their effects on performance,digestibility, volatile fatty acids and organo-leptic properties. Br. Poult. Sci., 52, 227-237.
da Silva M.A., Pessotti B.M.D., Zanini S.F.,Colnago G.L., Rodrigues M.R.A., Nunes L.D.,Zanini M.S., Martins I.V.F., 2009. Intestinalmucosa structure of broiler chickens infectedexperimentally with Eimeria tenella and treat-
ed with essential oil of oregano. Cienc. Rural,39, 1471-1477.
de Sousa D.P., 2011. Analgesic-like activityof essential oils constituents. Molecules, 16,2233-2252.
Demir E., Sarica S., Ozcan M.A., SuicmezM., 2005. The use of natural feed additives asalternative to an antibiotic growth promoter inbroiler diets. Arch. Geflügelk., 69, 110-116.
Di Pasqua R., Mamone G., Ferranti P.,Ercolini D., Mauriello G., 2010. Changes in theproteome of Salmonella enterica serovarThompson as stress adaptation to sublethalconcentrations of thymol. Proteomics, 10,1040-1049.
Dorman H.J., Deans S.G., 2000. Anti-micro-bial agents from plants: antibacterial activityof plant volatile oils. J. Appl. Microbiol., 88,308-316.
Faleiro M.L., 2011. The mode of antibacter-ial action of essential oils. In: Science againstmicrobial pathogens: communicating currentresearch and technological advances. Méndez-Vilas A. (Ed). Formatex, Spain, 1143-1156.
Franz C., Baser K.H.C., Windisch W., 2010.Essential oils and aromatic plants in animalfeeding - a European perspective. A review.Flavour Fragr., J. 25, 327-340.
Gabriel I., Mallet S., Sibille P., 2005. Lamicroflore digestive des volailles : facteurs devariation et conséquences pour l'animal. INRAProd. Anim., 18, 309-322.
Gomez G., Celii A., 2008. The peripheralolfactory system of the domestic chicken:Physiology and development. Brain Res. Bull.,76, 208-216.
Grabensteiner E., Arshad N., Hess M.,2007. Differences in the in vitro susceptibilityof mono-eukaryotic cultures of Histomonasmeleagridis, Tetratrichomonas gallinarum andBlastocystis sp. to natural organic compounds.Parasitol. Res., 101, 193-199.
Griffin S.G., Wyllie S.G., Markham J.L.,2001. Role of the outer membrane ofEscherichia coli AG100 and Pseudomonasaeruginosa NCTC 6749 and resistance/suscep-tibility to monoterpenes of similar chemicalstructure. J. Essent. Oil Res., 13, 380-386.
Guardia S., 2011. Effets de phytobiotiquessur les performances de croissance et l'équili-bre du microbiote digestif du poulet de chair.Thèse de doctorat de l’Université FrançoisRabelais, Tours, France, 472p.
Hafez H.M., Hauck R., 2006. Efficacy of aherbal product against Histomonas meleagridisafter experimental infection of turkey poults.Arch. Anim. Nutr., 60, 1-7.
Hernandez F., Madrid J., Garcia V., OrengoJ., Megias M.D., 2004. Influence of two plantextracts on broilers performance, digestibility,and digestive organ size. Poult. Sci., 83, 169-174.

Hume M.E., Barbosa N.A., Dowd S.E.,Sakomura N.K., Nalian A.G., Kley A.M.V.,Oviedo-Rondon E.O., 2011. Use of pyrose-quencing and denaturing gradient gel electro-phoresis to examine the effects of probioticsand essential oil blends on digestive microflorain broilers under mixed Eimeria infection.Foodborne Pathog. Dis., 8, 1159-1167.
Jamroz D., Wiliczkiewicz A., Wertelecki T.,Orda J., Skorupinska J., 2005. Use of activesubstances of plant origin in chicken dietsbased on maize and locally grown cereals. Br.Poult. Sci., 46, 485-493.
John L.J., Fromm M., Schulzke J.D., 2011.Epithelial barriers in intestinal inflammation.Antioxid. Redox Signal., 15, 1255-1270.
Jukic M., Politeo O., Maksimovic M., MilosM., Milos M., 2007. In vitro acetylcholi-nesterase inhibitory properties of thymol,carvacrol and their derivatives thymoquinoneand thymohydroquinone. Phytother. Res., 21,259-261.
Kang M.J., Cho J.Y., Shim B.H., Kim D.K.,Lee J., 2009. Bioavailability enhancing act-ivities of natural compounds from medicinalplants. J. Med. Plant Res., 3, 1204-1211.
Kidd M., Modlin I.M., Gustafsson B.I.,Drozdov I., Hauso O., Pfragner R., 2008.Luminal regulation of normal and neoplastichuman EC cell serotonin release is mediatedby bile salts, amines, tastants, and olfactants.Am. J. Physiol. Gastrointest. Liver Physiol.,295, G260-272.
Kim J.M., Marshall M.R., Cornell J.A.,Preston J.F., Wei C.I., 1995. Antibacterialactivity of carvacrol, citral, and geraniolagainst Salmonella typhimurium in culturemedium and on fish cubes. J. Food Sci., 60,1364-1368.
Kirkpinar F., Unlu H.B., Ozdemir G., 2011.Effects of oregano and garlic essential oils onperformance, carcase, organ and blood charac-teristics and intestinal microflora of broilers.Livest. Sci., 137, 219-225.
Knasmüller S., Nersesyan A., Misík M.,Gerner C., Mikulits W., Ehrlich V., Hoelzl C.,Szakmary A., Wagner K.H., 2008. Use of con-ventional and -omics based methods for healthclaims of dietary antioxidants: a criticaloverview. Br. J. Nutr., 99, ES 3-52.
Knoll U., Glunder G., Kietzmann M., 1999.Comparative study of the plasma pharmaco-kinetics and tissue concentrations of dano-floxacin and enrofloxacin in broiler chickens.J. Vet. Pharmacol. Ther., 22, 239-246.
Kohlert C., Schindler G., Marz R.W., AbelG., Brinkhaus B., Derendorf H., Grafe E.U.,Veit M., 2002. Systemic availability and phar-macokinetics of thymol in humans. J. Clin.Pharmacol., 42, 731-737.
Koscova J., Nemcova R., Gancarcikova S.,Jonecova Z., Scirankova L., Bomba A., BulecaV., 2006. Effect of two plant extracts andLactobacillus fermentum on colonization ofgastrointestinal tract by Salmonella entericavar. Dusseldorf in chicks. Biologia (Bratisl.),61, 775-778.
Kovar K.A., Gropper B., Friess D., AmmonH.P., 1987. Blood levels of 1,8-cineole andlocomotor activity of mice after inhalation andoral administration of rosemary oil. PlantaMed., 53, 315-318.
Kroismayr A., Sehm J., Pfaffl M.W.,Schedle K., Plitzner C., Windisch W., 2008.Effects of avilamycin and essential oils on
mRNA expression of apoptotic and inflamma-tory markers and gut morphology of piglets.Czech J. Anim. Sci., 53, 377-387.
Landa P., Kokoska L., Pribylova M., VanekT., Marsik P., 2009. In vitro anti-inflammatoryactivity of carvacrol: Inhibitory effect on COX-2 catalyzed prostaglandin E(2) biosynthesis.Arch. Pharmacal Res., 32, 75-78.
La Storia A., Ercolini D., Marinello F., DiPasqua R., Villani F., Mauriello G., 2011.Atomic force microscopy analysis shows sur-face structure changes in carvacrol-treatedbacterial cells. Res. Microbiol., 162, 164-172.
Lee K.W., Everts H., Beynen A.C., 2004.Essential oils in broiler nutrition. Int. J. Poult.Sci., 3, 738-752.
Lillehoj H.S., Kim D.K., Bravo D.M., LeeS.H., 2011. Effects of dietary plant-derivedphytonutrients on the genome-wide profilesand coccidiosis resistance in the broiler chick-ens. BMC Proc., 5, 10.1186/1753-6561-1185-S1184-S1134.
Lim W.C., Seo J.M., Lee C.I., Pyo H.B., LeeB.C., 2005. Stimulative and sedative effects ofessential oils upon inhalation in mice. Arch.Pharmacal Res., 28, 770-774.
Liu K., He L., Gao H., Huang X., Jiang Z.,Zeng Z., Liu K.Y., He L.M., Gao H., HuangX.H., Jiang Z.G., Zeng Z.L., 2009. Simul-taneous determination of andrographolide anddehydroandrographolide in chicken plasmafor application to pharmacokinetic studies.Chromatographia, 70, 1441-1445.
Magalhaes P.J.C., Criddle D.N., TavaresR.A., Melo E.M., Mota T.L., Leal-CardosoJ.H., 1998. Intestinal myorelaxant and anti-spasmodic effects of the essential oil of Crotonnepetaefolius and its constituents cineole,methyl-eugenol and terpineol. Phytother. Res.,12, 172-177.
Malecky M., 2008. Métabolisme des terpé-noïdes chez les caprins. Thèse de doctorat del’école doctorale Agric. Biol. Environ. Santé,206p.
Marsik P., Kokoska L., Landa P., NepovimA., Soudek P., Vanek T., 2005. In vitro inhibito-ry effects of thymol and quinones of Nigellasativa seeds on cyclooxygenase-1-and-2-catalyzed prostaglandin E-2 biosyntheses.Planta Med., 71, 739-742.
Melo F.H., Venâncio E.T., de Sousa D.P., deFrança Fonteles M.M., de Vasconcelos S.M.,Viana G.S., de Sousa F.C., 2010. Anxiolytic-like effect of carvacrol (5-isopropyl-2-methylphenol) in mice: involvement withGABAergic transmission. Fundam. Clin.Pharmacol., 24, 437-443.
Michiels J., 2009. Effect of essential oils ongut bacteria and functionality in the pig, GhentUniversity, Belgium, Ghent, Belgium, 282p.
Michiels J., Missotten J., Dierick N.,Fremaut D., Maene P., De Smet S., 2008. Invitro degradation and in vivo passage kineticsof carvacrol, thymol, eugenol and trans-cinnamaldehyde along the gastrointestinal tractof piglets. J. Sci. Food Agric., 88, 2371-2381.
Michiels J., Missotten J., Dierick N.,Fremaut D., De Smet S., 2010a. Thymol andtrans-cinnamaldehyde reduce active nutrientabsorption and chloride secretion in the pigjejunal Ussing chamber model. Livest. Sci.,134, 27-29.
Michiels J., Missotten J., Van Hoorick A.,Ovyn A., Fremaut D., De Smet S., Dierick N.,
2010b. Effects of dose and formulation ofcarvacrol and thymol on bacteria and somefunctional traits of the gut in piglets after wean-ing. Arch. Anim. Nutr., 64, 136-154.
Mikaili P., Nezhady M.A.M., Shayegh J.,Asghari M.H., 2010. Study of antinociceptiveeffect of nepeta meyeri, raphanus sativas andoriganum vulgare extracts in mouse by acutepain assessment method. Int. J. Acad. Res., 2,126-128.
Mikulski D., Zdunczyk Z., Jankowski J.,Juskiewicz J., 2008. Effects of organic acids ornatural plant extracts added to diets for turkeyson growth performance, gastrointestinal tractmetabolism and carcass characteristics. J.Anim. Feed Sci., 17, 233-246.
Moran E.T. 1982. Comparative nutrition offowl and swine : the gastrointestinal systems.University of Guelph, Ontario, Canada.
Muhl A., Liebert F., 2007. Growth, nutrientutilization and threonine requirement of grow-ing chicken fed threonine limiting diets withcommercial blends of phytogenic feed additi-ves. J. Poult. Sci., 44, 297-304.
Mukherjee P.S., Nandi B., 1994. Poultryfeed preservation from fungal infection bycinnamon oil. J. Mycopathol. Res., 32, 1-5.
Najafi P., Torki M., 2010. Performance,blood metabolites and immunocompetaence ofbroiler chicks fed diets included essential oilsof medicinal herbs. J. Anim. Vet. Adv., 9,1164-1168.
Ocana-Fuentes A., Arranz-Gutierrez E.,Senorans F.J., Reglero G., 2010. Supercriticalfluid extraction of oregano (Origanum vulgare)essentials oils: Anti-inflammatory propertiesbased on cytokine response on THP-1macrophages. Food Chem. Toxicol., 48, 1568-1575.
Osman M., Yakout H.M., Motawe H.F., El-Arab W.F.E., 2010. Productive, physiological,immunological and economical effects of sup-plementing natural feed additives to broilerdiets. Egypt. Poult. Sci. J., 30, 25-53.
Ouwehand A.C., Tiihonen K., Kettunen H.,Peuranen S., Schulze H., Rautonen N., 2010.In vitro effects of essential oils on potentialpathogens and beneficial members of the nor-mal microbiota. Vet. Med. (Praha), 55, 71-78.
Ovesná J., Slabý O., Toussaint O., KodícekM., Marsík P., Pouchová V., Vanĕk T., 2008.High throughput 'omics' approaches to assessthe effects of phytochemicals in human healthstudies. Br. J. Nutr., 99, ES 127-134.
Petracca E.G., Cavazzuti C., 1983. Clinicalevaluation of a new choleretic administered innormal and high doses to patients with chronichepatocholecytopathies. Clin. Ter., 107, 109-119.
Putnam S.E., Scutt A.M., Bicknell K.,Priestley C.M., Williamson E.M., 2007.Natural products as alternative treatments formetabolic bone disorders and for maintenanceof bone health. Phytother. Res., 21, 99-102.
Qiao H.Y., Dahiya J.P., Classen H.L., 2008.Nutritional and physiological effects of diet-ary sinapic acid (4-hydroxy-3,5-dimethoxy-cinnamic acid) in broiler chickens and itsmetabolism in the digestive tract. Poult. Sci.,87, 719-726.
Remmal A., Achahbar S., Bouddine L.,Chami N., Chami F., 2011. In vitro destructionof Eimeria oocysts by essential oils. Vet.Parasitol., 182, 121-126.
Utilisation des huiles essentielles en alimentation des volailles… / 23
INRA Productions Animales, 2013, numéro 1

Santos F.A., Rao V.S., 2001. 1,8-cineol, afood flavoring agent, prevents ethanol-inducedgastric injury in rats. Dig. Dis. Sci., 46, 331-337.
Sasaki K., Wada K., Tanaka Y., YoshimuraT., Matuoka K., Anno T., 2005. Thyme(Thymus vulgaris L.) leaves and its constituentsincrease the activities of xenobiotic-metaboli-zing enzymes in mouse liver. J. Med. Food, 8,184-189.
Scalbert A., Andres-Lacueva C., Arita M.,Kroon P., Manach C., Urpi-Sarda M., WishartD., 2011. Databases on food phytochemicalsand their health-promoting effects. J. Agric.Food Chem., 59, 4331-4348.
Sikkema J., De Bont J.A.M., Poolman B.,1995. Mechanisms of membrane toxicity ofhydrocarbons. Microbiol. Rev., 59, 201–222.
Soliman K.M., Badeaa R.I., 2002. Effect ofoil extracted from some medicinal plants ondifferent mycotoxigenic fungi. Food Chem.Toxicol., 40, 1669-1675.
Spernakova D., Mate D., Rozanska H.,Kovac G., 2007. Effects of dietary rosemaryextract and alpha-tocopherol on the per-formance of chickens, meat quality, andlipid oxidation in meat storaged under chil-ling conditions. Bull. Vet. Inst. Pulawy, 51,585-589.
Taherian A.A., Babaei M., Vafaei A.A.,Jarrahi M., Jadidi M., Sadeghi H., 2009.Antinociceptive effects of hydroalcoholicextract of Thymus vulgaris. Pak. J. Pharm. Sci.,22, 83-89.
Takaki I., Bersani-Amado L.E., VendruscoloA., Sartoretto S.M., Diniz S.P., Bersani-AmadoC.A., Cuman R.K.N., 2008. Anti-inflammatoryand antinociceptive effects of Rosmarinusofficinalis L. essential oil in experimental ani-mal models. J. Med. Food, 11, 741-746.
Tekeli A., Celik L., Kutlu H.R., Gorgulu M.,2006. Effect of dietary supplemental plantextracts on performance, carcass characteris-tics, digestive system development, intestinalmicroflora and some blood parameters of broil-er chicks. In 12th Eur. Poultry Conf., Verona,Italy, 6p.
Tiihonen K., Kettunen H., Bento M.H.L.,Saarinen M., Lahtinen S., Ouwehand A.C.,Schulze H., Rautonen N., 2010. The effect offeeding essential oils on broiler performanceand gut microbiota. Br. Poult. Sci., 51, 381-392.
Timbermont L., Lanckriet A., Dewulf J.,Nollet N., Schwarzer K., Haesebrouck F.,Ducatelle R., Van Immerseel F., 2010. Controlof Clostridium perfringens-induced necroticenteritis in broilers by target-released butyricacid, fatty acids and essential oils. AvianPathol., 39, 117-121.
Toutain P.L., Ferran A., Bousquet-Melou A.,2010. Species differences in pharmacokineticsand pharmacodynamics. Handb. Exp. Pharma-col., 19-48.
Ultee A., Bennik M.H., Moezelaar R., 2002.The phenolic hydroxyl group of carvacrol isessential for action against the food-bornepathogen Bacillus cereus. Appl. Environ.Microb., 68, 1561-1568.
Viuda-Martos M., Ruiz Navajas Y., SanchezZapata E., Fernandez-Lopez J., Perez-AlvarezJ.A., 2010. Antioxidant activity of essentialoils of five spice plants widely used in amediterranean diet. Flavour Fragr. J., 25, 13-19.
Wallace R.J., Oleszek W., Franz C., Hahn I.,Baser K.H.C., Mathe A., Teichmann K., 2010.Dietary plant bioactives for poultry health andproductivity. Br. Poult. Sci., 51, 461-487.
Williams R.B., 1997. Laboratory tests ofphenolic disinfectants as oocysticides againstthe chicken coccidium Eimeria tenella. Vet.Rec., 141, 447-448.
Zenner L., Callait M.P., Granier C., ChauveC., 2003. In vitro effect of essential oils fromCinnamomum aromaticum, Citrus limon andAllium sativum on two intestinal flagellates ofpoultry, Tetratrichomonas gallinarum andHistomonas meleagridis. Parasite, 10, 153-157.
INRA Productions Animales, 2013, numéro 1
24 / I. GABRIEL, F. ALLEMAN, V. DUFOURCQ, F. PERRIN, J.-F. GABARROU
Les composants des huiles essentielles peuvent présenter différentes activités biologiques, i.e. antimicrobienne, antioxydante et stimu-latrice de récepteurs spécifiques. Chez l’animal, leur action dépend de leur devenir dans l’organisme lors de leur consommation, puisde leur action au niveau de leurs cibles biologiques. Le site de leur absorption le long du tractus digestif peut être modulé par diffé-rents facteurs, tels que la composition chimique de l’aliment, leur mode de présentation (sous forme libre ou protégée par encapsu-lation) et la dose d’utilisation. Selon leur site d’absorption, ils peuvent agir à différents niveaux : pour les composants absorbés dèsla partie supérieure du tractus digestif, ils peuvent avoir un effet systémique, et pour ceux persistant jusqu’en fin d’intestin grêle, ilspeuvent avoir une action dans les contenus digestifs jusqu’au niveau de ce segment intestinal, puis de façon systémique après leurabsorption. Les produits absorbés après modifications métaboliques au niveau du foie peuvent ensuite exercer une action au niveaudes différents organes de l’animal. Après élimination au niveau des reins, et compte tenu des mouvements de reflux de l’urine versles caeca chez les oiseaux, les composants issus des huiles essentielles pourraient agir au niveau de ces deux diverticules. Ils peuventavoir un effet sur le microbiote digestif (précaecal ou caecal), sur les tissus de l’animal lui-même, c’est-à dire sur leur état d’oxyda-tion, sur l’immunité en particulier l’inflammation, sur l’appareil digestif et ses fonctions digestives, sur le métabolisme animal, surson système nerveux et son comportement. Ces composants ont donc un grand potentiel d’action, qui reste à explorer, pour pouvoiroptimiser leur utilisation.
Résumé
Abstract
Essential oils in poultry feeding. 2. Hypotheses of modes of action implicated
The components of essential oils may have various biological activities, i.e. antimicrobial, antioxidant, and stimulant of specific recep-tors. In animals, their action depends on their fate in the body after feeding, and their biological targets. The absorption site alongthe digestive tract is modulated by various factors such as diet chemical composition, their form of presentation (free or protected byencapsulation) and dose level. Depending on their absorption site, they can act at various levels: for compounds absorbed in the upperpart of the digestive tract, they can act at the systemic level, and for those attaining the end of the small intestine, they can act in diges-tive contents until this intestinal segment, and at the systemic level after their absorption. The adsorbed products, undergo metabolicchanges in the liver, and then act on various organs of animals. After their excretion by kidneys, and due to refluxes from urine to caecain birds, the compounds generated from essential oils may act on these two blind sacs. They can act on digestive microbiota (precaecalor caecal), animal tissues , that is on their oxidative status, immunity and particularly on inflammation, the digestive tract and its diges-tive functions, and on animal metabolism, nervous system and behaviour. These compounds have thus a great potential of action, whichneed to be further studied in the future, in order to optimise their use.
GABRIEL I., ALLEMAN F., DUFOURCQ V., PERRIN F., GABARROU J.-F., 2013. Utilisation des huiles essentielles enalimentation des volailles. 2. Hypothèses sur les modes d’action impliqués dans les effets observés. INRA Prod. Anim., 26, 1, 13-24.





























