Embed Size (px)
Citation preview

UNIVERZA V LJUBLJANI
PEDAGOŠKA FAKULTETA
Pouĉevanje - Predmetno pouĉevanje
Biologija in gospodinjstvo
Amela Osmić
VPLIV SELENA IN JODA NA IZBRANE
BIOKEMIJSKE IN MORFOLOŠKE LASTNOSTI
PRI KALICAH KOLERABICE (Brassica oleracea
L. var. gongylodes L.)
Magistrsko delo
Ljubljana, 2017


UNIVERZA V LJUBLJANI
PEDAGOŠKA FAKULTETA
Pouĉevanje - Predmetno pouĉevanje
Biologija in gospodinjstvo
Amela Osmić
VPLIV SELENA IN JODA NA IZBRANE
BIOKEMIJSKE IN MORFOLOŠKE LASTNOSTI
PRI KALICAH KOLERABICE (Brassica oleracea
L. var. gongylodes L.)
Magistrsko delo
Mentorica: izr. prof. dr. Mateja Germ
Ljubljana, 2017


Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
I
Magistrsko delo je zakljuĉek magistrskega študija na Pedagoški fakulteti, smer Biologija in
gospodinjstvo. Delo je bilo opravljeno v prostorih Biotehniške fakultete Univerze v Ljubljani,
na Oddelku za biologijo in Katedre za ekologijo in varstvo okolja.
Študijska komisija Pedagoške fakultete je 12. 9. 2016 odobrila predlagano temo magistrske
naloge z naslovom Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri
kalicah kolerabice (Brassica oleracea L. var. gongylodes L.). Za mentorico magistrskega dela
je imenovala doc. dr. Matejo Germ.
Komisija za oceno in zagovor:
Predsednica: izr. prof. Katarina VOGEL MIKUŠ
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za biologijo
Mentorica: izr. prof. dr. Mateja GERM
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za biologijo
Ĉlanica: prof. dr. Alenka GABERŠĈIK
Univerza v Ljubljani, Biotehniška fakulteta, Oddelek za biologijo
Datum zagovora:
Podpisana izjavljam, da je naloga rezultat lastnega raziskovalnega dela. Izjavljam, da je
elektronski izvod identiĉen tiskanemu. Na univerzo neizkljuĉno, neodplaĉano, ĉasovno in
prostorsko neomejeno prenašam pravici shranitve avtorskega dela v elektronski obliki in
reproduciranja ter pravico omogoĉanja javnega dostopa do avtorskega dela na svetovnem
spletu preko Digitalne knjiţnice Pedagoške fakultete.
Amela Osmić

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
II
ZAHVALA
Zahvaljujem se mentorici izr. prof. dr. Mateji Germ za vso pomoĉ, vodenje in razlage pri
nastajanju mojega magistrskega dela.
Aleksandri Golob, mladi raziskovalki, se iskreno zahvaljujem za pomoĉ pri statistiĉni
obdelavi podatkov.
Hvala tehniĉnemu sodelavcu, Draganu Abramu, za pomoĉ pri eksperimentalnem delu.
Zahvala gre tudi prof. dr. Alenki Gaberšĉik in izr. prof. Katarini Vogel Mikuš za hiter in
temeljit pregled magistrskega dela in tehtne komentarje.
Hvala prof. dr. Nini Kacjan Maršiĉ za pripravo raztopin selena in joda, ki smo jih uporabili pri
poskusu in za strokovne nasvete.
Posebna zahvala gre tudi moji druţini in moţu, ki mi vedno stojijo ob strani in verjamejo
vame.
»Znanje je zaklad, ki venomer spremlja svojega lastnika.« (Kitajski pregovor)

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
III
POVZETEK
Selen (Se) in jod (I) sta esencialna elementa za ljudi in ţivali, medtem ko njuna esencialna
vloga za rastline še ni dokazana. Obstaja tudi zelo malo podatkov o interakciji med Se in I pri
rastlinah, zato je pomembno, da preuĉujemo hkraten vpliv obeh elementov na rastline, ki jih
uporabljamo za prehrano ljudi. Pribliţno 2/3 svetovne populacije ima zdravstvene teţave, ki
so povezane z nezadostnim vnosom Se in I s hrano v telo. Ena izmed najpreprostejših rešitev
za to teţavo je biofortifikacija oz. obogatitev poljšĉin s Se in I, katere namen je poveĉanje
prenosa Se in I v prehransko verigo.
V naši raziskavi so poskusne rastline predstavljale kalice kolerabice (Brassica oleracea L.
var. gongylodes L.). Kolerabica je kapusnica, ki spada v druţino kriţnic in je videti kot
mešanec med zeljem in repo. Raziskavo smo izvajali na Oddelku za biologijo Biotehniške
fakultete, na Katedri za ekologijo in varstvo okolja. Z raziskavo smo ţeleli ugotoviti, ali
razliĉne oblike Se in I posamezno ali v kombinaciji vplivajo na izbrane biokemijske in
morfološke lastnosti pri kalicah kolerabice. Za dosego tega cilja smo si zastavili dve
raziskovalni vprašanji. S prvim raziskovalnim vprašanjem smo poskušali ugotoviti, kako
razliĉne kemijske oblike Se in I ter njune kombinacije vplivajo na izbrane biokemijske in
morfološke lastnosti kalic kolerabice, z drugim raziskovalnim vprašanjem pa smo poskušali
ugotoviti, kako razliĉne kemijske oblike Se in I ter njune kombinacije vplivajo na rast kalic
kolerabice. Raziskava je potekala na sluĉajnostnem vzorcu, ki so ga predstavljala semena
kolerabice. Poskus je bil sestavljen iz devetih obravnavanj. Vsako obravnavanje je obsegalo 4
ponovitve, v eni ponovitvi je vzorec predstavljalo 300 semen kolerabice. Kalice smo vzgojili
iz semen, ki smo jih namoĉili v osem razliĉnih raztopin z razliĉnimi kombinacijami in
oblikami Se in I ter v kontrolno raztopino, ki jo je predstavljala destilirana voda (dH2O).
Ostale raztopine so poleg dH2O vsebovale posamezno dodan selenit (SeO32-
) oz. selenat
(SeO42-
) s koncentracijo 10 mg Se/L, jodid (I-) oz. jodat (IO3
-) s koncentracijo 1.000 mg I/L in
kombinacije razliĉnih oblik Se in I (SeO32-
+ I-, SeO3
2-+ IO3
-, SeO4
2- + I
-, SeO4
2- + IO
3-). Selen
je bil v dH2O dodan v obliki natrijevega selenita (Na2SeO3) oz. natrijevega selenata
(Na2SeO4), I pa v obliki kalijevega jodida (KI) oz. kalijevega jodata (KIO3). Merili smo
koncentracijo klorofila a in b, karotenoidov, antocianov, UV-A absorbirajoĉih snovi, UV-B
absorbirajoĉih snovi ter potencialno fotokemiĉno uĉinkovitost fotosistema II (FS II). Na
koncu poskusa smo izmerili še višino in maso kalic. Zaradi primerljivosti rezultatov smo
celoten poskus po enakem postopku izvedli dvakrat.
Kljuĉne besede: kolerabica, kalice, selen, jod

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
IV

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
V
ABSTRACT
Selenium (Se) and iodine (I) are essential elements for humans and animals, while their
essential role for plants has not yet been established. There is also very little information
about the interaction between selenium and iodine in plants, therefore it is important to study
the combined effect of these two elements on plants that are used for human consumption.
Approximately 2/3 of the world's population has health problems associated with insufficient
intake of Se and I with diet. One of the easiest way to combat this problem is biofortification
or enrichment of crops with Se and I, to increase the transfer of Se and I into the food chain.
In our research we used kohlrabi sprouts (Brassica oleracea L. var. gongylodes L.). Kohlrabi
is a brassica, which belongs to the family of Brassicaceae, and it looks like a hybrid between
cabbage and turnip. The research was conducted at the Department of Biology at the
Biotechnical Faculty in the Chain of Ecology and Environmental Protection. The aim of our
research was to determine the effect of different forms of Se, I and their combinations on
selected biochemical and morphological characteristics of the kohlrabi sprouts. To achieve
this goal we have set ourselves two research questions. With the first research question we
tried to examine how the different chemical forms of Se and I and combinations of both
elements affect selected biochemical and morphological properties of kohlrabi sprouts. With
the second research question we tried to examine how the different chemical forms of Se and
I and combinations of both elements affect growth of kohlrabi sprouts. The study was
conducted on a random sample, which were kohlrabi seeds. The experiment consisted of nine
treatments. There were four replicates for each treatment with 300 seeds per one replicate.
Sprouts were grown from seeds, which were soaked in eight different solutions with different
combinations and forms of Se and I, and in the control solution which was distilled water
(dH2O). Other solutions contained selenite (SeO32-
) or selenate (SeO42-
) with a concentration
of 10 mg Se/L, iodide (I-) or iodate (IO3-) with a concentration of 1000 mg I/L, and their
combinations (SeO32-
+ I-, SeO3
2-+ IO3
-, SeO4
2- + I
-, SeO4
2- + IO
3-). Selenium was applied to
distilled water in the form of sodium selenite (Na2SeO3) and sodium selenate (Na2SeO4),
respectively. Iodine was applied to distilled water in the form of potassium iodide (KI) and
potassium iodate (KIO3), respectively. We measured the content of chlorophyll a and b,
carotenoids, anthocyanins, UV-A and UV-B absorbing substances. We also measured
potential photochemical efficiency of photosystem (PS II). At the end of the experiment we
measured the weight and length of the sprouts. In order to compare the results the entire
experiment was carried out twice.
Keywords: kohlrabi, sprouts, selenium, iodine

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
VI
KAZALO VSEBINE
1 UVOD ..................................................................................................................................... 1
1.1 OPREDELITEV RAZISKOVALNEGA PROBLEMA .................................................. 2
1.2 CILJ RAZISKAVE IN RAZISKOVALNI VPRAŠANJI ................................................ 2
2 PREGLED OBJAV ................................................................................................................. 3
2.1 KOLERABICA ................................................................................................................ 3
2.2 SELEN.............................................................................................................................. 4
2.2.1 LASTNOSTI SELENA ............................................................................................. 4
2.2.2 PRISOTNOST SELENA V TLEH ............................................................................ 4
2.2.3 PRISOTNOST SELENA V VODI IN ZRAKU ........................................................ 5
2.2.4 SELEN V RASTLINAH ........................................................................................... 5
2.2.5 SELEN V LJUDEH IN ŢIVALIH ............................................................................ 8
2.3 JOD ................................................................................................................................... 9
2.3.1 LASTNOSTI JODA .................................................................................................. 9
2.3.2 PRISOTNOST JODA V TLEH ................................................................................. 9
2.3.3 PRISOTNOST JODA V VODI IN ZRAKU ............................................................. 9
2.3.4 JOD V RASTLINAH .............................................................................................. 10
2.3.5 JOD V LJUDEH IN ŢIVALIH ............................................................................... 11
2.4 SELEN IN JOD .............................................................................................................. 12
2.4.1 HKRATNO DELOVANJE SELENA IN JODA ..................................................... 12
3 MATERIALI IN METODE .................................................................................................. 13
3.1 RAZISKOVALNA METODA IN RAZISKOVALNI PRISTOP.................................. 13
3.2 VZOREC ........................................................................................................................ 13
3.3 OPIS POSTOPKA ZBIRANJA PODATKOV .............................................................. 13
3.4 MERITVE ...................................................................................................................... 15
3.4.1 BIOKEMIJSKE IN MORFOLOŠKE MERITVE ................................................... 15
3.5 STATISTIĈNA OBDELAVA PODATKOV ................................................................ 17
4 REZULTATI ......................................................................................................................... 18
4.1 MORFOLOŠKE LASTNOSTI KALIC ......................................................................... 18
4.1.1 Sveţa masa............................................................................................................... 18
4.1.2 Suha masa ................................................................................................................ 19
4.1.3 Višina kalic .............................................................................................................. 20
4.2 BIOKEMIJSKE LASTNOSTI KALIC .......................................................................... 21
4.2.1 Koncentracija klorofila a na enoto suho mase ......................................................... 21

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
VII
4.2.2 Koncentracija klorofila b na enoto suho mase ......................................................... 22
4.2.3 Koncentracija karotenoidov na enoto suho mase ..................................................... 23
4.2.4 Koncentracija antocianov na enoto suho mase ........................................................ 24
4.2.5 Koncentracija UV-B absorbirajoĉih snovi na enoto suho mase ............................... 25
4.2.6 Koncentracija UV-A absorbirajoĉih snovi na enoto suho mase .............................. 26
4.2.7 Potencialna fotokemiĉna uĉinkovitost FS II ............................................................ 27
5 RAZPRAVA .......................................................................................................................... 28
5.1 MASA, VIŠINA IN ŠTEVILO KALIC ......................................................................... 28
5.2 KONCENTRACIJE BARVIL ........................................................................................ 30
5.3 POTENCIALNA FOTOKEMIĈNA UĈINKOVITOST FOTOSISTEMA II ............... 32
6 SKLEPI .................................................................................................................................. 34
7 LITERATURA ...................................................................................................................... 35
PRILOGE ................................................................................................................................. 43
Priloga A: Število kalic ......................................................................................................... 43

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
VIII
KAZALO SLIK
Slika 1: Kolerabica (Brassica oleracea L. var gongylodes L.) Vir: Mek.niif.hu ....................... 3
Slika 2: Poskus, nastavljen 30. 3. 2016. Foto: Amela Osmić. ................................................. 14
Slika 3: 10 dni stare kalice, katerih semena so bila obravnavana z I(V). Foto: Amela Osmić. 14
Slika 4: Sveţa masa kalic ......................................................................................................... 18
Slika 5: Suha masa kalic .......................................................................................................... 19
Slika 6: Višina kalic ................................................................................................................. 20
Slika 7: Koncentracija klorofila a na enoto suho mase ............................................................ 21
Slika 8: Koncentracija klorofila b na enoto suho mase ............................................................ 22
Slika 9: Koncentracija karotenoidov na enoto suho mase ....................................................... 23
Slika 10: Koncentracija antocianov na enoto suho mase ......................................................... 24
Slika 11: Koncentracija UV-B absorbirajoĉih snovi na enoto suho mase ............................... 25
Slika 12: Koncentracija UV-A absorbirajoĉih snovi na enoto suho mase ............................... 26
Slika 13: Potencialna fotokemiĉna uĉinkovitost FS II ............................................................. 27

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
IX
KAZALO PREGLEDNIC
Preglednica 1: Obravnavanja pri 1. in 2. poskusu ter koncentracije razliĉnih oblik Se in I. .... 13

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
X
OKRAJŠAVE IN SIMBOLI
ant
CaCO3
CH3I
dH2O
DMSe
DMDSe
Fm
FS
Fv
H2SeO4
kar
kl a
kl b
n
GSH-Px
I
I2
I- / I(-I)
IO3- / I(V)
K
KI
KIO3
MSeC
Na2SeO3
Na2SeO4
SE
Se
Se2-
SeO32-
/ Se(IV)
SeO42-
/ Se (VI)
SeCys
SeMet
sm
antociani
kalcijev karbonat
metil jodid
destilirana voda
dimetil selenid
dimetil diselenid
maksimalna fluorescenca klorofila a v FS II temotno adaptiranega
vzorca
fotosistem
variabilna fluorescenca temotno adaptiranega vzorca
selenova kislina
karotenoidi
klorofil a
klorofil b
število vzorcev
glutation peroksidaza
jod
elementarni jod
jodid
jodat
kontrolne rastline
kalijev jodid
kalijev jodat
metilselenocistein
natrijev selenit
natrijev selenat
standardna napaka
selen
selenid
selenit
selenat
selenocistein
selenometionin
suha masa vzorca

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
1
1 UVOD
Selen (Se) in jod (I) sta elementa, ki sta potrebna za pravilno delovanje šĉitnice pri ljudeh
(Germ in Stibilj, 2007). Pribliţno 2/3 svetovne populacije trpi zaradi bolezni in zdravstvenih
teţav, ki so povezani z nezadostnim vnosom Se in I s hrano v telo. Eden izmed
najpreprostejših naĉinov za boj proti tej teţavi je biofortifikacija oz. obogatitev poljšĉin s Se
in I, katere namen je poveĉanje prenosa Se in I v prehransko verigo (Smoleń, Kowalska in
Sady, 2014). Številni raziskovalci priporoĉajo, da gojenim rastlinam, ki jih uporabljamo za
hrano ljudi, dodajamo soĉasno oba elementa, saj je Se nujno potreben za metabolizem I v
šĉitnici (Germ idr., 2015).
Dokazov o esencialnosti obeh elementov za rastline še ni. Obstaja zelo malo podatkov o
hkratnem delovanju Se in I pri rastlinah. Oba elementa pozitivno vplivata na rastline v
zmernih koncentracijah, medtem ko lahko visoke koncentracije obeh elementov delujejo
strupeno (Germ in Stibilj, 2007). Selen ima v prehrani ţivali in ljudi dvojno vlogo, saj je pri
manjših koncentracijah esencialen, pri veĉjih pa strupen. Meja med esencialnostjo in
strupenostjo Se je zelo ozka in je med 0,1 in 1 mg Se/kg. Ţivali in ljudje zauţijemo veĉino Se
s hrano v obliki aminokislin (selenocistein (SeCys) in selenometionin (SeMet)). (Ţnidarĉiĉ,
2011). Minimalna hranilna vrednost za ţivali je pribliţno 0,05 do 0,10 mg Se/kg suhe snovi
krme, medtem ko ima lahko izpostavljenost koncentracijam od 2 do 5 mg Se/kg suhe snovi
krme strupen uĉinek (Terry, Zayed, Souza de in Tarun, 2000). Znaki pomanjkanja Se v
ĉloveškem organizmu se pokaţejo pri dnevnem vnosu, manjšem od 19 µg Se/dan pri moških
in 13 µg Se/dan pri ţenskah. Zgornja meja, ki še ne povzroĉa teţav pa je 400 µg Se/dan
(Ţnidarĉiĉ, 2011). Pri rastlinah je meja med esencialnostjo in strupenostjo Se odvisna
predvsem od rastlinske vrste oz. od tega, koliko Se je rastlina sposobna kopiĉiti. Selen
neakumulirajoĉe rastline privzemajo manj kot 25 mg Se/kg suhe snovi, medtem ko selen
akumulirajoĉe rastline privzemajo tudi do 4000 mg Se/kg suhe snovi (Ellis in Salt, 2003).
Vsebnost Se v rastlinah je v veliki meri odvisna od vsebnosti elementa v tleh in sposobnosti
rastlin za njegov privzem (Smrkolj, Pograjc, Hlastan-Ribiĉ in Stibilj, 2005). Znani so
pozitivni vplivi Se na nekatere gojene rastline. Selen zavira procese, povezane s staranjem,
pospešuje antioksidacijsko aktivnost in omili stres, ki ga povzroĉa moĉna svetloba in suša
(Xue, Hartikainen in Piironen, 2001). Pri vplivu I na rastline je kljuĉna njegova koncentracija,
poleg tega pa je vpliv I na rastline odvisen tudi od njegove kemijske oblike. Rastline naj bi
prenesle veĉje koncentracije jodata (IO3-) kot jodida (I
-) (Borst-Pauwels, 1961). Številne
raziskave so pokazale, da lahko nizke koncentracije I (0,02 – 0,2 mg I/kg) v rastnem okolju,
neodvisno od njegove oblike, zelo ugodno vplivajo na številne rastlinske vrste, še posebej na
rastline, ki uspevajo na slanih tleh (Borst Pauwels, 1961; Dai, Zhu, Huang, Zhang in Song,
2006; Zhu, Huang, Hu in Liu, 2003).
Za temo raziskovalne naloge sem se odloĉila, ker obstaja zelo malo podatkov o hkratnem
delovanju Se in I pri rastlinah. S pomoĉjo kavzalne eksperimentalne metode smo v raziskavi,
izvedeni na kalicah kolerabice (Brassica oleracea L. var. gongylodes L.), preverili, kako
razliĉne kemijske oblike Se (selenit (SeO32-
), selenat (SeO42-
)) in I (jodid (I-), jodat (IO3
-))
posamezno ali v kombinaciji vplivajo na koncentracije klorofila a in b, karotenoidov,
antocianov, UV-A absorbirajoĉih snovi, UV-B absorbirajoĉih snovi ter fotokemiĉno
uĉinkovitost fotosistema II (FS II). Preverili smo tudi, kako razliĉne kemijske oblike Se in I
ter njune kombinacije vplivajo na rast in število kalic, ki vzklijejo iz semen. Poskus na kalicah

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
2
smo po enakih korakih izvedli dvakrat in v obliki grafov primerjalno prikazali rezultate 1. in
2. poskusa.
1.1 OPREDELITEV RAZISKOVALNEGA PROBLEMA
Zaradi pomanjkanja podatkov o hkratnem delovanju Se in I pri rastlinah in zaradi ozke meje
med esencialnostjo in strupenostjo Se za ţiva bitja smo z raziskavo ţeleli ugotoviti, kako
razliĉne oblike Se in I ter njune kombinacije vplivajo na izbrane biokemijske in morfološke
lastnosti kalic kolerabice (Brassica oleracea L. var gongylodes L.).
1.2 CILJ RAZISKAVE IN RAZISKOVALNI VPRAŠANJI
Cilj magistrske naloge je ugotoviti, ali razliĉne oblike Se (SeO32-
, SeO42-
) in I (I-, IO3
-)
posamezno ali v kombinaciji vplivajo na izbrane biokemijske in morfološke lastnosti kalic
kolerabice (Brassica oleracea L. var. gongylodes L.).
Za dosego cilja smo si zastavili naslednji raziskovalni vprašanji:
1. Kako bodo razliĉne kemijske oblike Se in I ter njune kombinacije vplivale na rast kalic
kolerabice?
2. Kako bodo razliĉne kemijske oblike Se in I ter njune kombinacije vplivale na izbrane
biokemijske in morfološke lastnosti kalic kolerabice?

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
3
2 PREGLED OBJAV
2.1 KOLERABICA
Kolerabica je kapusnica, ki spada v druţino Brassicaceae (Ĉerne, 1998; Kalloo in Bergh,
1993; Lim, 2015). Ime izvira iz nemških besed kohl (zelje) in rabi (repa), saj je videti kot
mešanec med zeljem in repo (Lešić idr., 2004). Kolerabica je gojena zelenjavnica, ki se
neredko zaseje zunaj gojitve na ledinah, ob njivah in na podobnih krajih. Njena domovina je
obmorsko skalovje v Evropi (Martinĉiĉ idr., 2007). Kolerabica bolje prenaša vroĉino in sušo
kot veĉina vrst iz druţine kriţnic in zlahka prenese zmrzali (do -12°C) (Lim, 2015). Je
dvoletna rastlina, ki v primeru normalne rasti cvetno odevalo razvije v drugem letu po
jarovizaciji. Ob kalitvi razvije kliĉna lista, hipokotil in glavno korenino s stranskimi
koreninicami. V tkivu nad kliĉnimi listi se steblo odebeli in omeseni. Po jarovizaciji se razvije
golo cvetno steblo, na katerem rastejo vijakasto razporejeni sedeĉi listi. Cvetovi so rumene
barve, ĉašni in venĉni listi pa so razporejeni v obliki kriţa. Cvetovi so dvospolni in zdruţeni v
cvetne grozde. Plod, ki se razvije po oploditvi, imenujemo lusk (Ĉerne, 1998). V dolţino meri
8 do 12 cm, v širino pa 5 mm (Ĉerne, 1998; Lim, 2015). Seme kolerabice je temno rjave
barve, okrogle oblike in v premer meri od 1 do 2 mm. V tehnološki zrelosti rastlina doseţe
višino 40 do 50 cm (Lim, 2015), v fiziološki zrelosti pa lahko zraste tudi do 1 m (Ĉerne,
1998).
Slika 1: Kolerabica (Brassica oleracea L. var gongylodes L.) Vir: Mek.niif.hu

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
4
2.2 SELEN
2.2.1 LASTNOSTI SELENA
Prvi, ki je izoliral in opisal selen (Se), je bil švedski kemik Jons Jacob Berzelius. Odkril ga je
leta 1817, ko je analiziral rdeĉe obloge na stenah svinĉevih komor, ki so se uporabljale pri
proizvodnji ţveplove kisline (Tinggi, 2003; Ţnidarĉiĉ, 2011). Selen ima atomsko število 34,
atomsko maso 78,96 (Kopsell in Kopsell, 2007) in se nahaja v 6. skupini periodnega sistema,
med telurjem (kovina) in ţveplom (nekovina) ter v 4. periodi med arzenom in bromom
(Tinggi, 2003). Spada med polkovine (metaloide), kar pomeni, da ima lastnosti kovin, kot tudi
nekovin (Kopsell in Kopsell, 2007; Reilly, 1998). Nekatere kemijske lastnosti Se kot so
atomska masa, vezavna energija, ionizacijski potencial in elektronska afiniteta, so zelo
podobne kot pri ţveplu (S) (Reilly, 1998; Tinggi, 2003). Naravne Se spojine so analogi in
derivati naravnih S spojin, zaradi ĉesar obstaja povezanost med metabolizmom Se in S.
Namreĉ pri asimilaciji anorganskih Se spojin sodelujejo isti encimi kot pri asimilaciji
anorganskih S spojin, zaradi tega Se in S med seboj tekmujeta za vstop v celico (Dilworth in
Bandurski, 1977; Läuchli, 1993; Terry idr., 2000). Bistvena razlika med Se in S je ta, da je Se
v štirivalenĉni reducirani obliki, S pa v štirivalenĉni oksidirani obliki (Tinggi, 2003).
V naravi najdem 6 naravnih izotopov Se (74
Se, 76
Se, 77
Se, 78
Se, 80
Se, 82
Se) (Brenĉiĉ in
Lazarini, 1995). Selen najdemo tako v organski kot tudi v anorganski obliki. V anorganskih
spojinah ima Se razliĉna oksidacijska števila: -2 (selenid), 0 (elementarni Se), +2 (tioselenat),
+4 (selenit) in +6 (selenat) (Terry idr., 2000). Zaradi razliĉnih oksidacijskih stanj je Se
zmoţen tvoriti organske spojine (dimetilselenid, trimetilselen) in aminokisline
(selenometionin, selenocistein) (Tinggi, 2003). Selen je prisoten tudi v nekaterih hlapnih in
nehlapnih organskih spojinah (Hymer in Caruso, 2006; Läuchli, 1993; Pyrzyńska, 2001;
Uden, Boakye, Kahakachchi in Tyson, 2004). Oblika Se je odvisna od pH in redoks razmer. V
oksiĉnih, nevtralnih do alkalnih razmerah prevladuje selenat (SeO42-
), medtem ko v
anoksiĉnih okoljih prevladujeta selenid (Se2-
) in elementarni Se (Läuchli, 1993; Terry idr.,
2000). V rastlinah lahko pride do pretvorbe anorganskih oblik Se v organske, vendar so te
pretvorbe odvisne od rastlinske vrste (Pedrero in Madrid, 2009). Biokemiĉni in fiziološki
uĉinek Se je odvisen predvsem od oblike in koncentracije Se v organizmu. Elementarni Se je
najmanj biorazpoloţljiv, medtem ko ima SeO42-
niţjo biološko aktivnost in sposobnost
absorpcije kot SeO32-
(Tinggi, 2003). Selen ima pri ţivalskih organizmih tri biološke ravni
aktivnosti (Germ, Stibilj in Kreft, 2007b): (i) koncentracija v sledovih je potrebna za
normalno rast in razvoj; (ii) zmerna koncentracija omogoĉa homeostazo; (iii) medtem ko ima
lahko povišana koncentracija strupene uĉinke.
2.2.2 PRISOTNOST SELENA V TLEH
Selen je eden izmed najbolj razširjenih elementov Zemeljske skorje (Kabata-Pendias in
Pendias, 2001; Kopsell in Kopsell, 2007). Vsebnost Se v tleh je med 0,02 mg Se/kg in 1.000
mg Se/kg in je predvsem odvisna od geološke sestave tal. V tleh, ki so bogata z organsko
snovjo in apnenĉastih alkalnih tleh ga je obiĉajno veĉ kot v granitnih tleh (Kabata-Pendias in
Pendias, 2001). Na porazdelitev Se v tleh vplivajo predvsem klimatske razmere, narava in
izvor tal ter posegi ĉloveka v okolje (Kabata-Pendias in Pendias, 2001). Proces erozije
spodbuja sprošĉanje Se iz kamnin, v okolje se sprošĉa tudi zaradi spiranja tal in biološke
aktivnosti organizmov (Navarro-Alarcon in Cabrera-Vique, 2008). Selen se pojavlja v tleh v

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
5
razliĉnih oblikah: elementarni Se, Se2-
, SeO32-
, SeO42-
in organski Se (Ryser idr., 2006). V
kislih tleh je prevladujoĉa oblika Se SeO32-
, ki je zaradi nizke topnosti manj dostopen za
rastline. V alkalnih tleh pa je Se prisoten v obliki SeO42-
, ki je bolj topen in tako rastlinam
bolje dostopen (Navarro-Alarcon in Cabrera-Vique, 2008). Teţava se pojavi ob obilnih
padavinah, ki povzroĉijo izpiranje Se v topni obliki. Zaradi tega so tla v Novi Zelandiji, na
Finskem, na doloĉenih obmoĉjih Kitajske, v Tasmaniji in Koreji revna s Se (Combs, 2001). V
osrednji Evropi je koncentracija Se v tleh manj kot 1,0 mg Se/kg, na Norveškem okrog 0,25
mg Se/kg in na Finskem 0,04 – 0,07 mg Se/kg (Germ idr., 2007b). V Sloveniji se vsebnost Se
v tleh giblje od 0,1 do 0,7 mg Se/kg (Ţnidarĉiĉ, 2011). Fordyce (2005) navaja, da se vsebnost
Se v tleh po navadi manjša z vsebnostjo gline, ţeleza in organskih snovi v tleh.
2.2.3 PRISOTNOST SELENA V VODI IN ZRAKU
Koncentracija Se v površinskih vodah je le nekaj µg Se/L, kar je zelo malo (Uden, 2005). V
pitni vodi je njegova koncentracija obiĉajno pod 1 µg Se/L, medtem ko je koncentracija Se v
morski vodi še 10-krat manjša (Reilly, 2006). V morski vodi je Se prisoten predvsem v obliki
SeO32-
in SeO42-
. Strokovnjaki predvidevajo, da se prek mikroogranizmov pretvori v
anorganski in elementarni Se, ob prisotnosti planktona pa se pretvori v organske selenove
komponente (Uden, 2005). Selen je prisoten tudi v podzemni vodi. Vsebnost Se v geotermalni
vodi na severovzhodu Slovenije je v mejah od 0,1 do 41 mg Se/L (Ţnidarĉiĉ, 2011). V
atmosferi je Se prisoten predvsem v elementarni obliki in je vezan na prašne delce. Lahko je
antropogenega (seţig fosilnih goriv) ali naravnega izvora. V atmosferi je prisotnega le nekaj
ng Se/m3
(Barceloux, 1999; Uden, 2005).
2.2.4 SELEN V RASTLINAH
Esencialna vloga Se za rastline še ni dokazana, obstajajo pa dokazi, ki kaţejo, da ima Se pri
majhnih koncentracijah spodbujajoĉ vpliv (Germ in Stibilj, 2007; Germ idr., 2007b), medtem
ko lako pri velikih koncentracijah povzroĉi negativne uĉinke na fiziološke lastnosti rastlin
(Hartikainen, Xue in Piironen, 2000). Meje med esencialnostjo in strupenostjo Se so zelo ozke
in so veĉinoma odvisne od koncentracije in kemijske oblike, v kateri se Se pojavlja (Golob
idr., 2016a). Selen pri majhnih koncentracijah vpliva na rastline spodbujajoĉe, saj lahko
izboljša strpnost rastlin na razliĉne abiotske stresorje (Germ idr., 2007b), pospeši rast rastlin,
upoĉasni staranje (Hartikainen idr., 2000; Xue idr., 2001), poleg tega lahko uravnava vodni
status rastlin v obdobju suše (Germ in Stibilj, 2007), tako da poveĉa kapaciteto privzema vode
v koreninskem sistemu (Kuznetsov, Kholodova, Kuznetsov in Yagodin, 2003). Proces
staranja se upoĉasni na raĉun poveĉane antioksidacijske aktivnosti, ki je povezana s
poveĉanjem aktivnosti glutation peroksidaze (GSH-Px) (Germ idr., 2007b). Hartikainen idr.
(2000) navajajo, da je Se pozitivno vplival na rast ljuljke (Lolium perenne). Pri
koncentracijah, veĉjih od 10 mg Se/L, pa so se pokazali negativni uĉinki pri gojenju ljuljke in
solate (Lactuca sativa L.), saj se je biomasa obeh rastlin zmanjšala (Xue idr., 2001). Mechora,
Stibilj, Radešĉek, Gaberšĉik in Germ (2011) so pri raziskavi na rdeĉem zelju (Brassica
oleracea var. capitata L. f. rubra, cv. Erfurtsko rano) ugotovili, da so bile rastline rdeĉega
zelja, izpostavljene SeO42-
, višje kot kontrolne rastline, ki niso bile obravnavane s SeO42-
.
Majhna koncentracija SeO42-
(2 µg Se/L) je pozitivno vplivala na rast rastlin rdeĉega zelja,
medtem ko je rast ostala nespremenjena pri veliki koncentraciji (0,5 mg Se/L). Dodatek
SeO42-
pa ni imel nobenega vpliva na biomaso glav rdeĉega zelja, saj je ostala nespremenjena.

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
6
Na rastlinah ni bilo vidnih znakov strupenosti SeO42-
. Selen zmanjšuje dostopnost teţkih
kovin za rastline in na ta naĉin šĉiti rastline pred poškodbami oksidativnega stresa, ki ga lahko
povzroĉijo teţke kovine (Pedrero, Madrid-Albarrán, Hartikainen in Cámara, 2008; Thangavel,
Sulthana in Subburam, 1999). Germ, Kreft in Osvald (2005) navajajo, da se je respiratorni
potencial rukole (Eruca sativa) poveĉal pri rastlinah, katerih semena so bila izpostavljena Se.
Dokazano je bilo, da Se pozitivno vpliva na akumulacijo ogljikovih hidratov pri krompirju
(Solanum tuberosum) (Turakainen, Hartikainen in Seppänen, 2004). Tudi Pennanen, Xue in
Hartikainen (2002) so v povezavi s tem ugotovili, da se v rastlinah, ki jim je dodajan Se,
poveĉa kopiĉenje škroba v kloroplastih. Na privzem in prenos Se v rastlinah vplivajo
kemijska oblika in koncentracija Se, razliĉni dejavniki tal kot so pH, slanost, koncentracija
drugih ionov v tleh in sposobnost rastlin za privzem Se (Kopsell in Kopsell, 2007). Korenine
rastlin privzemajo Se v obliki SeO32-
, SeO42-
in selenovih organskih spojin kot sta SeCys in
SeMet (Kabata-Pendias in Pendias, 2001). Selenat se v rastlini prenaša laţje in hitreje kot
SeO32-
(Terry idr., 2000). Arvy (1993) je gojil fiţol in v tla dodajal SeO32-
oz. SeO42-
.
Ugotovil je, da se je v manj kot treh urah, 50 % SeO42-
preneslo iz korenin v poganjke. Poskus
je ponovil s SeO32-
in ugotovil, da se je le majhna koliĉina SeO32-
prenesla v poganjke, saj je
veĉina tega ostala v koreninah. Na primeru indijske gorĉice so ugotovili, da se je 10 % SeO32-
preneslo iz korenin v poganjke, medtem ko se je SeO42-
prenašal brez teţav in dvakrat hitreje
kot SeO32-
. Razlog za slabši prenos SeO32-
iz korenin v poganjke je lahko v tem, da se v
koreninah hitro pretvori v selenove organske spojine, ki se v korenini tudi ohranijo, medtem
ko se SeO42-
preko ksilema transportira do poganjka, kjer v kloroplastih poteĉe asimilacija
SeO42-
v selenove organske spojine, te pa se preko floema prerazporedijo po celotni rastlini
(Terry idr., 2000). Vsebnost Se v rastlinah lahko poveĉamo z dodajanjem Se v tla,
namakanjem semen v raztopino Se pred setvijo, z listnim škropljenjem ter z aeroponiĉnim in
hidroponiĉnim gojenjem v hranljivi raztopini, ki vsebuje Se. Kje se bo Se v rastlini kopiĉil in
kakšna bo njegova razporeditev v rastlini, je odvisno od rastlinske vrste, njenega fiziološkega
stanja in faze razvoja ter od koncentracije in oblike Se (Terry idr., 2000). Aktivno rastoĉa
tkiva obiĉajno vsebujejo veĉje koliĉine Se, zato rastline pogosto kopiĉijo veĉ Se v listih in
poganjkih kot v koreninah (Germ in Stibilj, 2007; Smrkolj, Germ, Kreft in Stibilj, 2006),
medtem ko so semena obiĉajno zmerni viri Se (Germ in Stibilj, 2007). Rastline v listih
kopiĉijo znatne koliĉine SeO42-
, veliko manj pa kopiĉijo SeO32-
in SeMet (Terry idr., 2000).
Golob idr. (2016b) navajajo, da listno škropljenje rastlin hibridne in tatarske ajde z raztopino
natrijevega selenata (Na2SeO4) poveĉa kopiĉenje Se v vseh rastlinskih organih, zlasti v listih
in semenih. Pri hibridni ajdi, obravnavani z raztopino Na2SeO4, so bile vrednosti Se v listih
186-krat in v semenih 64-krat veĉje kot pri kontrolni skupini, ki ni vsebovala Na2SeO4. Pri
tatarski ajdi so bile vrednosti Se v listih 83-krat in v semenih 276-krat veĉje kot pri kontrolni
skupini, ki ravno tako ni vsebovala Na2SeO4.
Koliĉina privzetega Se v rastlinskih tkivih je odvisna tudi od rastlinske vrste. Na podlagi teh
sposobnosti loĉimo rastline na (Ellis in Salt, 2003; Terry idr., 2000):
seleno neakumulirajoĉe rastline, ki akumulirajo manj kot 25 mg Se/kg suhe snovi in je
Se veĉinoma prisoten v obliki aminokisline SeMet (v to skupino spada veĉina rastlin);
seleno akumulirajoĉe ali primarne indikatorske rastline, ki akumulirajo do 4000 mg
Se/kg suhe snovi (vrste iz rodu Astragalus, Morinda, Neptunia, Oonopsis, Stanleya,
Xylorhiza). V njih je Se prisoten preteţno v obliki prostih aminokislin, kot so
selenocistationin, metilselenocistein, selenocistin in selenohomocistein;
seleno sekundarno akumulirajoĉe ali indikatorske rastline, ki akumulirajo do 1000 mg
Se/kg suhe snovi (v to skupino spadajo rastline iz rodu Aster, Atriplex, Castillea,

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
7
Comandra, Grayia, Grindelia, Gutierrezia, Machaeranthera in vrsti indijska ogršĉica
(Brassica juncea) ter navadna ogršĉica (Brassica napus).
V seleno akumulirajoĉih rastlinah se v zgodnji vegetativni fazi rasti veĉina Se kopiĉi v mladih
listih, medtem ko se v reproduktivni fazi Se kopiĉi predvsem v semenih, veliko manj ga je
takrat v listih (Terry idr., 2000). Velike koncentracije Se imajo lahko negativne uĉinke na
rastline, saj lahko povzroĉijo fiziološke motnje, ki se kaţejo kot zmanjšana izgradnja
beljakovin, slabša rast, listi zaĉnejo rumeneti in se sušiti (Kabata-Pendias in Pendias, 2001;
Kopsell in Kopsell, 2007; Terry idr., 2000), pojavijo se lahko ĉrne pike na listih in rdeĉkaste
pike na koreninah, lahko pa pride tudi do predĉasnega odmiranja rastlin (Kabata-Pendias in
Pendias, 2001; Terry idr., 2000). Velike koncentracije Se imajo lahko negativen vpliv na
izgradnjo encima porfobilinogen sintaze, ki je potreben za biosintezo klorofila, kar poslediĉno
pripelje do kloroz. Selenat lahko negativno vpliva na izgradnjo glutationa. Terry idr. (2000)
navajajo, da je dodatek SeO42-
negativno vplival na kopiĉenje glutationa v špinaĉnih listih.
Tudi smrekove iglice so izpostavili SeO42-
, kar je privedlo do znatnega zmanjšanja vsebnosti
glutationa v iglicah. Motnje v izgradnji glutationa lahko poslabšajo obrambo rastlin pred
hidroksilnimi radikali in oksidativnim stresom. Prisotnost sulfatnih ionov in starost rastlin
vplivajo na raven Se, ki je za rastline strupena. Mlajše rastline koruze in pšenice so bile, za
razliko od starejših rastlin, bolj obĉutljive na velike koncentracije Se, zaradi ĉesar je bilo
zaviranje rasti pri njih veĉje kot pri starejših rastlinah. Najpogostejši strupeni obliki Se za
rastline sta SeO32-
in SeO42-
, saj ju rastline zelo hitro absorbirajo in pretvorijo v organske
selenove spojine (Terry idr., 2000). Strupenost SeO32-
in SeO42-
za semenke je mogoĉe
pripisati kombinaciji treh dejavnikov. Prvi dejavnik je hitra absorpcija SeO32-
in SeO42-
iz tal s
pomoĉjo korenin in hitra translokacija do drugih delov rastline. Drugi dejavnik je hitra
presnova obeh anionov v organske oblike Se znotraj celic. Tretji dejavnik pa predstavljajo
organski selenovi metaboliti, ki delujejo kot analogi esencialnih ţveplovih spojin in motijo
biokemijske reakcije v celicah (Brown in Shrift, 1982).
Rastline so razvile razliĉne mehanizme, ki jim omogoĉajo strpnost do negativnega delovanja
Se. Nekateri obrambni mehanizmi so (Terry idr., 2000):
izkljuĉitev SeCys iz procesa vkljuĉevanja v beljakovine;
kopiĉenje Se v vakuolah (v obliki nebeljakovinskih aminokislin in/ali SeO42-
);
izgradnja nebeljakovinskih aminokislin;
fitovolatizacija oz. pretvorba Se v hlapne spojine, kot sta dimetilselenid (DMSe) in
dimetildiselenid (DMDSe).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
8
2.2.5 SELEN V LJUDEH IN ŢIVALIH
Selen je esencialen mikroelement za ljudi, ţivali in mikroorganizme (Germ idr., 2007b).
Ţivali in ljudje zauţijemo veĉino Se s hrano v obliki Se aminokislin (SeCys in SeMet). V
telesu ga je najveĉ v jetrih, ledvicah in skeletnih mišicah (Foster in Sumar, 1997). Leta 1957
so ugotovili, da dodajanje natrijevega selenita (Na2SeO3) prepreĉuje nekrozo jeter pri
podganah, ki so trpele za pomanjkanjem vitamina E (Schwartz in Foltz, 1957). V majhnih
koncentracijah ima Se pozitivne uĉinke na zdravje ljudi in ţivali, saj deluje kot antioksidant in
zaradi tega prepreĉuje nastanek razliĉnih vrst raka in ostalih bolezni (npr. artritisa,
ateroskleroze in spremenjenih imunoloških funkcij) (Germ in Stibilj, 2007; Pedrero in
Madrid, 2009). Številne raziskave so usmerjene v raziskovanje varovalnih uĉinkov Se v zvezi
z rakom, ki so povezani s prisotnostjo Se v GSH-Px, ki šĉitijo DNK in celiĉne komponente
pred poškodbami s kisikovimi radikali (Letavayova, Vlĉkova in Brozmanova, 2006;
Schrauzer, 2001). V dolgoroĉnih raziskavah je dodajanje Se zmanjšalo nastanek pljuĉnega
raka, raka prostate in debelega ĉrevesa. Metilselenocistein (MSeC) je ena najbolj uĉinkovitih
Se spojin, ki zavirajo nastanek raka (Ellis in Salt, 2003). Ugotovili so, da je umrljivost zaradi
raka manjša na obmoĉjih, kjer je koncentracija Se v tleh veĉja (Shamberger, 1981). Selen je
pomemben tudi za normalno delovanje šĉitnice, saj je sestavni del druţine encimov
jodotironin dejodinaz, ki sodelujejo pri aktivaciji ali deaktivaciji šĉitniĉnih hormonov
(Smoleń idr., 2014).
Preskrbljenost ljudi s Se je v številnih evropskih drţavah manjša od priporoĉenega dnevnega
vnosa Se. Zaradi tega narašĉa zanimanje za rastline, ki naravno vsebujejo veĉje koliĉine Se
(Finley, 2007). Na obmoĉjih, kjer so tla revna s Se, se pojavljata dve bolezni, ki sta povezani s
pomanjkanjem Se v tleh in poslediĉno tudi v prehrani ljudi. Prva je Kashin-Beckova bolezen,
ki prizadene predvsem mišice in sklepe. Posledice bolezni so poveĉani sklepi, šibkost udov,
otekanje, okornost in boleĉine v prstnih ĉlenkih ter atrofija nekaterih progastih mišic. Druga
bolezen je Keshanova bolezen, za katero pa so znaĉilne vrtoglavica, izguba apetita, slabost in
teţave s srcem (srĉne aritmije, poveĉanje srca in srĉno popušĉanje). Bolezen prizadane
predvsem otroke in noseĉnice (Letavayova idr., 2006). Pomanjkanje Se pri ţivalih lahko
povzroĉi mišiĉno distrofijo (Germ idr., 2007b). S premajhno preskrbljenostjo s Se so
povezane še številne druge bolezni kot so: anemija, neplodnost, povišan krvni tlak in multipla
skleroza (Peters idr., 2007). Znaki pomanjkanja Se v ĉloveškem organizmu se pokaţejo pri
dnevnem vnosu, manjšem od 13 µg Se/dan pri ţenskah in 19 µg Se/dan pri moških. Zgornja
meja, ki še ne povzroĉa teţav pa je 400 µg Se/dan (Ţnidarĉiĉ, 2011). Preveliki odmerki Se
lahko povzroĉijo zastrupitev (humana selenoza), zaradi katere pride do poškodb DNK.
Najbolj tipiĉen znak prevelikih zauţitih koliĉin Se je zadah po ĉesnu, ki se pojavi kot
posledica izloĉanja hlapnih Se spojin, še posebno DMSe. Drugi znaki, ki se kaţejo pri humani
selenozi so: izguba las, morfološke spremembe nohtov, pri hudi zastrupitvi pa lahko pride
tudi do degeneracije jeter in ledvic (Letavayova idr., 2006).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
9
2.3 JOD
2.3.1 LASTNOSTI JODA
Jod (I) je leta 1811 po nakljuĉju odkril francoski kemik Bernard Courtois. Kmalu po tem sta
Humphrey Davy in Louis-Joseph Gay-Lussac I opredelila kot nov element in leta 1813 ga je
Gay-Lussac tudi poimenoval. Ime izhaja iz grške besede ioeides, kar pomeni vijoliĉno ali
vijoliĉna (Guthrie, 1863). Namreĉ I je pri sobni temperaturi modro-ĉrna, blešĉeĉa trdnina, ki
lahko sublimira v vijoliĉen plin z draţljivim vonjem. Jod se nahaja v sedmi skupini
periodnega sistema z atomskim številom 53. Elemente, ki se nahajajo v sedmi skupini
periodnega sistema, imenujemo halogeni in so nekovine. V skupini halogenov je I najmanj
reaktiven. Jod je v elementarnem stanju dvoatomna molekula (I2) in je slabo topen v vodi
(Brenĉiĉ in Lazarini, 1995). V naravi lahko obstaja kot monovalentni anion ali pa se pojavlja
v molekularnih spojinah. Jod se pojavlja v razliĉnih oksidacijskih stanjih: -1, 0, +1, +3, +5 in
+7. Kemijske in fizikalne lastnosti I se spreminjajo z elementno ali molekularno obliko
elementa (Kabata-Pendias in Pendias, 2001).
2.3.2 PRISOTNOST JODA V TLEH
Jod se v litosferi nahaja izkljuĉno v sledovih in zaradi preperevanja matiĉne kamnine prehaja
v tla (Fuge, 2005). Sedimentne kamnine lahko vsebujejo bistveno veĉje koncentracije I kot
magmatske kamnine. Razlog za to je lahko kopiĉenje I med procesom sedimentacije in
biološka aktivnost v tleh. Na vsebnost I v tleh vpliva pH, tekstura tal, vsebnost gline v tleh
(Zia, Watts, Gardner in Chenery, 2014), ĉlovekove dejavnosti, vulkanske dejavnosti,
preperevanje kamnin, padavine in razgradnja organskih snovi (Whitehead, 1984).
Koncentracija I v tleh je od 0,1 do 150 mg I/kg tal. Obiĉajno je vsebnost I v tleh veĉja kot v
matiĉni kamnini, iz katere tla nastanejo. Veĉina I v tleh izhaja iz atmosfere in morskega
okolja, zaradi ĉesar se koncentracija I v tleh z oddaljenostjo od morja navadno manjša.
Povpreĉna koncentracija I v litosferi znaša 5 mg I/kg in se nahaja predvsem v anorganskih
oblikah (IO3-, I
-, I2). Iz tal pa se lahko sprošĉa v plinasti obliki kot metil jodid (CH3I) (Fuge,
2005).
2.3.3 PRISOTNOST JODA V VODI IN ZRAKU
Whitehead (1984) navaja, da koncentracije I v rekah in jezerih znašajo okrog 0,5 – 20 µg I/L.
Na splošno so te koncentracije manjše od tistih v morski vodi, ki so med 45 – 50 µg I/L
(Fuge, 2005). V morski vodi se I nahaja tako v organski obliki (npr. CH3I), kot tudi v
anorganski obliki (I-, IO3
-) (Fuge, 2005). Vsebnost I
- je v zgornjih plasteh vodnega stolpca
poveĉana, kar lahko pripišemo delovanju nekaterih ţivih organizmov, ki so s pomoĉjo
encimov sposobni reducirati IO3- v I
- (Tsunogai in Sase, 1969). Morski organizmi imajo torej
pomembno vlogo pri pretvorbi I v razliĉne kemijske oblike in pri njegovem prenosu iz vode v
atmosfero (Fuge, 2005).
Koncentracija I v atmosferi je 10 – 20 ng I/m3
(Whitehead, 1984). V atmosferi se I nahaja
veĉinoma v obliki CH3I (Rasmussen, Khalil, Gunawardena in Hoyt, 1982), nekaj pa ga je tudi
v aerosolih (Fuge, 2005). Lahko je antropogenega (izgorevanje fosilnih goriv) ali naravnega
izvora (Chameides in Davis, 1980).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
10
2.3.4 JOD V RASTLINAH
Številne študije so pokazale, da imajo majhne koncentracije I spodbujajoĉ vpliv na rastline.
(Borst-Pauwels, 1961; Landini, Gonzali in Perata, 2011; Lehr, Wybenga in Rosanow, 1958;
Smoleń, Rożek, Ledwożyw-Smoleń in Strzetelski, 2011; Zhu idr., 2003). Fiziološka,
biokemijska in molekularna narava tega vpliva še ni dokazana, zaradi ĉesar tudi esencialna
vloga I za rastline še ni bila potrjena oz. dokazana (Borst-Pauwels, 1961; Landini, Gonzali in
Perata, 2011; Lehr, Wybenga in Rosanow, 1958; Smoleń, Rożek, Ledwożyw-Smoleń in
Strzetelski, 2011; Zhu idr., 2003). Pri vplivu I na rastline je kljuĉna njegova koncentracija
(Blasco idr., 2011; Borst Pauwels, 1961; Lehr idr., 1958), poleg tega pa je vpliv I na rastline
odvisen tudi od rastlinske vrste in od kemijske oblike I. Rastline naj bi prenesle veĉje
koncentracije IO3- kot I
- (Borst-Pauwels, 1961). Namreĉ pri podobnih koncentracijah je za
rastline I- bolj strupen kot IO3
- (Kato idr., 2013; Mackowiak, Grossl in Cook, 2005). Predvsem
v zaĉetni fazi razvoja naj bi imel IO3- bolj ugoden uĉinek na rast rastlin ko I
- (Borst Pauwels,
1961).
Majhne koncentracije I (0,02 – 0,2 mg I/kg) v rastnem okolju, neodvisno od njegove oblike,
ugodno vplivajo na številne rastlinske vrste, še posebej na tiste, ki uspevajo na slanih tleh
(Borst Pauwels, 1961; Dai idr., 2006; Zhu idr., 2003). Prevelike koncentracije I imajo
negativen vpliv na rastline in lahko delujejo strupeno, kar lahko pripelje do zavrtja rasti,
poškodb listov ali pa celo do smrti rastline (Lehr idr., 1958).
Privzem I iz tal v rastlino je odvisen od adsorpcijsko-desorpcijskih procesov v tleh. Listnata
zelenjava ima veĉjo absorpcijsko sposobnost kot plodovna zelenjava (Weng idr., 2013).
Rastline privzemajo I iz tal preko koreninskega sistema in prednostno privzemajo I v obliki I-
(Fuge, 2005; Smoleń idr., 2011). Jodid ima torej veĉjo biorazpoloţljivost za rastline kot IO3-
(Umaly in Poel, 1971; Whitehead, 1973). Rastline lahko, skozi kutikulo in lepljive delce na
dlaĉicah listov, absorbirajo I direktno iz atmosfere (Fuge, 2005). Poskus na rastlinah riţa je
pokazal, da se je I kopiĉil v koreninah, listih in steblih, medtem ko je bila koncentracija I v
zrnih izredno majhna, kar kaţe na nizek floemski transport. Namreĉ prenos I iz vegetativnih
organov v semena zahteva aktivni floemski transportni sistem za I (Blasco idr., 2011; Zhu
idr., 2003). Koncentracija I se torej zmanjšuje od korenin, listov, stebla do plodov (Weng idr.,
2013). Zia idr. (2014) navajajo, da je koncentracija I v mlajših listih veĉja kot v starejših.
Študije, ki so jih številni avtorji opravili kaţejo, da je proces prerazporeditve I po rastlini, s
pomoĉjo floemskega transporta omejen (Smoleń idr., 2014). Hurtevent idr. (2013) so delali
raziskavo na pšenici. Rastline pšenic so listno škropili z vodno raztopino I. Mobilnost I od
listov do zrna (t.i. translokacijski faktor) je bila zelo nizka (0,2 – 1,1%), vendar ta vrednost še
vseeno kaţe na to, da se je I, ob vsakem listnem škropljenju, prenašal od listov do zrna. Tudi
Landini idr. (2011) so prišli do podobnih zakljuĉkov, saj so s svojo raziskavo na paradiţniku,
ki so ga listno škropili z vodno raztopino I-, ugotovili, da se je I prenašal od listov do plodov.
Dai idr. (2006) so ugotovili, da I- in IO3
-, ki so ga dodali v tla, nista bistveno vplivala na
biomaso rastlin špinaĉe (Spinacia oleracea L.). Tudi Zhu idr. (2003) so preuĉevali vpliv I na
špinaĉo in ugotovili, da I ni koristil rasti špinaĉe, saj je koncentracija I- nad 10 µM I/L,
zmanjšala donos špinaĉe, medtem ko je imel IO3- majhen uĉinek na biomaso. Razlog
negativnemu vplivu I- na rast rastlin naj bi bilo prekomerno kopiĉenje I
- v rastlinskih tkivih.
Blasco idr. (2011) so ugotovili, da je previsok odmerek I- (80 µM I/L), v primerjavi s
kontrolnimi rastlinami, ki niso vsebovale I-, znatno zmanjšal biomaso rastlin vrtne solate
(Lactuca sativa L. var. longifolia). V nasprotju s tem pa je obravnava rastlin z IO3- (20 µM
I/L) poveĉala biomaso rastlin, transpiracijo, prevodnost listnih reţ in stopnjo fotosinteze, v

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
11
primerjavi s kontrolnimi rastlinami, ki niso bile obravnavane z IO3-. Tudi Leyva idr. (2011) so
ugotovili, da je dodatek IO3- poveĉal biomaso rastlin vrtne solate in izboljšal njihov
antioksidacijski odziv. Voogt, Holwerda in Khodabaks (2010) so ugotovili, da obravnava
vrtne solate z I- in IO3
- ni pokazala bistvenega vpliva na njeno biomaso, medtem pa je bilo
kopiĉenje I- uĉinkovitejše od IO3
-. Jod se je veĉinoma porazdelil v zunanjih listih rastline.
Landini idr. (2011) so obravnavali paradiţnik (Solanum lycopersicium L.) z I-, ki je bil v
hranilno raztopino dodan v obliki kalijevega jodida (KI). Ugotovili so, da so rastline
paradiţnika strpne do relativno velikih koncentracij I- (20 mM), ki rastlinam niso povzroĉile
hujših poškodb. Rastline, ki so bile obravnavane z najmanjšo koncentracijo I- (5 mM), so
imele najbolj pigmentirane liste, ki so bili izrazito temno zelene barve, kar kaţe na
proizvodnjo antioksidativnih spojin kot so antociani.
2.3.5 JOD V LJUDEH IN ŢIVALIH
Jod je esencialen element za ljudi in ţivali (Blasco idr., 2011; Germ idr., 2015; Landini idr.,
2011; Zhu, Huang, Hu, Liu in Christie, 2004), saj je potreben za pravilno delovanje šĉitnice
(Smoleń idr., 2015). Namreĉ I je sestavni del šĉitniĉnih hormonov (Fuge, 2005; Kato idr.,
2013), (tri-jodotironina (T3) in tiroksina/tetrajodotironina (T4)), ki uravnavajo stopnjo
metabolizma, izgradnjo proteinov, rast in razvoj veĉine organov, termoregulacijo ter
pomagajo pri ohranitvi centralnega ţivĉnega sistema (Weng idr., 2013). Šĉitniĉna hormona se
nahajata v šĉitniĉnih mešiĉkih, kjer sta vezana na glikoprotein tiroglobulin. Za sintezo
šĉitniĉnih hormonov je potreben I. Jod vstopa v šĉitniĉno celico v obliki I-. Pri vstopu ima
pomembno vlogo beljakovina, imenovana NIS (natrijev / jodidni simporter), ki sodeluje pri
aktivnem transportu I v šĉitnici. Jod kopiĉijo folikelske celice, kjer poteka sinteza šćitniĉnih
hormonov. Ti se potem sprošĉajo v kri (Farhana in Shaiq, 2010). Jod, ki ga zauţijemo v obliki
I-, se hitro in v pribliţno 90 % absorbira v kri. Na podroĉjih, kjer je preskrba z I ustrezna, se
ga do 10% nakopiĉi v šĉitnici, pribliţno 90 % pa se ga izloĉi z urinom. Jod nastopa v hrani v
mineralni obliki I- in IO3
-. Biorazpoloţljivost oz. absorpcija I je veĉja pri I
- kot pri IO3
-,
vendar se obe obliki s hrano dobro absorbirata (Zimmermann, 2009).
Motnje, ki so povezane s pomanjkanjem I, so ene izmed najpogostejših zdravstvenih teţav, ki
jih ima svetovna populacija (Welch in Graham, 1999). Ocenjeno je bilo, da okoli 225
milijonov ljudi po vsem svetu trpi za pomanjkanjem I oz. golšavostjo, medtem ko 1 bilijon
ljudi lahko utrpi pomanjkanje I (Zhu idr., 2004). Pomanjkanje I je povezano s številnimi
motnjami kot so golšavost, mentalna zaostalost, poškodbe moţganov in reproduktivni
neuspeh (Blasco idr., 2011). Vzrok za endemiĉno pomanjkanje I v ĉloveški in ţivalskih
populacijah je skromen prenos I iz tal v rastline, ki so prvi ĉlen v prehranjevalni verigi.
Gnojenje tal z I je najenostavnejši naĉin za poveĉanje vsebnosti I v rastlinah. Ta proces je
opredeljen kot agrotehniĉna metoda, katere namen je obogatiti rastline z mineralnimi
komponentami (White in Broadley, 2009). Eden izmed naĉinov za dopolnitev I v ĉloveški
prehrani pa je tudi jodiranje soli (Zhu idr., 2004). Leta 1953 je bil v Sloveniji uveden obvezen
ukrep jodiranja soli z 10 mg KI/kg soli. Rezultati številnih raziskav so pokazali, da je bil
takrat vnos I premajhen, zato so se predpisi spremenili in se je koncentracija I v kuhinjski soli
poveĉala na 25 mg KI/kg soli (ali 32 mg KIO3/kg soli, kar je enako kot 19 mg I/kg soli). Leta
2003 je stopil v veljavo nov pravilnik o kakovosti soli, po katerem morata biti jodirani le
evaporirana in kamena sol, morska pa ne (Štimec idr., 2007).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
12
2.4 SELEN IN JOD
2.4.1 HKRATNO DELOVANJE SELENA IN JODA
Zelo malo podatkov obstaja o hkratnem delovanju Se in I pri rastlinah. Zaradi njune
esencialnosti za ljudi in ţivali so mnogi raziskovalci mnenja, da bi morali gojenim rastlinam
soĉasno dodajati oba elementa, saj ima Se pomembno vlogo pri metabolizmu I v šĉitnici
(Germ idr., 2015). Ĉeprav je splošno znano, da sta Se in I biološko tesno povezana v ţivalih
(Arthur, Nicol in Beckett, 1992), so interakcije med Se in I v rastlinah le redko preuĉevane. S
tem namenom so Zhu idr. (2004) naredili raziskavo na špinaĉi (Spinacia oleracea L.), katere
namen je bil raziskati hkratno delovanje Se (SeO42-
) in I (IO3-) pri špinaĉi. Ugotovili so, da sta
se Se in I kopiĉila predvsem v listih. Kopiĉenje Se in I v uţitnih delih rastline je zelo
pomembno za izboljšanje in dopolnitev ĉloveške prehrane. Študija je pokazala, da je
koncentracija Se v rastlinskih tkivih ostala nespremenjena z dodatkom I v hranilno raztopino.
Zhu idr. (2004) so v hranilno raztopino, v katero so dodali IO3- pri koncentraciji 6,25 mg I/L,
dodali še Se in ugotovili, da je dodan Se rahlo zniţal zmoţnost rastline za privzem I, saj je
bila koncentracija nakopiĉenega I v poganjkih manjša kot v poganjkih, kjer niso dodali Se. V
koreninah niso opazili nobenih sprememb. Rezultati študije so jasno pokazali, da ni bilo
moĉne interakcije med Se in I v rastlinskih procesih in metabolizmu.
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
13
3 MATERIALI IN METODE
3.1 RAZISKOVALNA METODA IN RAZISKOVALNI PRISTOP
V raziskavi smo uporabili kavzalno eksperimentalno metodo pedagoškega raziskovanja in
kvantitativni raziskovalni pristop.
3.2 VZOREC
Raziskava je potekala na sluĉajnostnem vzorcu, ki so ga predstavljala semena kolerabice.
Poskus je bil sestavljen iz devetih obravnavanj. Vsako obravnavanje je obsegalo 4 ponovitve,
v eni ponovitvi je vzorec predstavljalo 300 semen kolerabice.
3.3 OPIS POSTOPKA ZBIRANJA PODATKOV
Poskusne rastline so predstavljale kalice kolerabice (Brassica oleracea L. var. gongylodes L.).
Poskus je bil sestavljen iz devetih obravnavanj, vsako od obravnavanj je obsegalo 4
ponovitve. Pripravili smo 8 raztopin z razliĉnimi kombinacijami in oblikami Se in I ter
kontrolno raztopino, ki jo je predstavljala destilirana voda (dH2O). Ostale raztopine so poleg
dH2O vsebovale posamezno dodan SeO32-
oz. SeO42-
s koncentracijo 10 mg Se/L, I- oz. IO3
- s
koncentracijo 1.000 mg I/L in kombinacije razliĉnih oblik Se in I. Se je bil v dH2O dodan v
obliki natrijevega selenita (Na2SeO3) oz. natrijevega selenata (Na2SeO4), I pa v obliki
kaljevega jodida (KI) oz. kalijevega jodata (KIO3). Semena smo pred namakanjem stehtali. Po
24 urah namakanja smo semena zopet stehtali, nato pa smo jih dali posamezno za vsako
obravnavanje kaliti na navlaţene papirnate brisaĉke na plastiĉnem podstavku. Pri razporejanju
semen po podstavkih smo za vsako obravnavanje uporabili nove rokavice. Papirnate brisaĉe
smo moĉili ves ĉas trajanja poskusa. Podstavke smo postavili na police v rastni komori.
Kalice smo vzgajali pri konstantni temperaturi 19,3°C, RZV cca 60 % in pod hladno belo
fluorescentno svetlobo z jakostjo 160 µM m-2
s-1
PAR. Dnevno-noĉni cikel je bil v razmerju
16 h : 8 h.
Preglednica 1: Obravnavanja pri 1. in 2. poskusu ter koncentracije različnih oblik Se in I.
Obravnavanja Koncentracija
Se(IV) (mg/L)
Koncentracija
Se(VI) (mg/L)
Koncentracija
I(-I) (mg/L)
Koncentracija
I(V) (mg/L)
K (dH2O) / / / /
Se (VI) / 10 / /
I (-I) / / 1000 /
Se (VI) + I (V) / 10 / 1000
Se (IV) + I (-I) 10 / 1000 /
Se (IV) 10 / / /
Se (VI) + I (-I) / 10 1000 /
Se (IV) + I (V) 10 / / 1000
I (V) / / / 1000

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
14
Semena smo 29. 3. 2016 namoĉili v raztopine. Dne 12. 4. 2016 smo zaĉeli z laboratorijskimi
analizami. 15. 4. 2016 smo kalice pobrali in opravili še rastno analizo. Poskus smo še enkrat
ponovili dne 18. 4. 2016, ko smo semena zopet namoĉili v posamezne raztopine. 3. 5. 2016
smo zaĉeli z laboratorijskimi analizami. Dne 6. 5. 2016 smo opravili rastno analizo. V
laboratoriju smo na listih kalic merili koncentracijo klorofila (kl) a in b, karotenoidov (kar),
antocianov (ant), UV-A in UV-B absorbirajoĉih snovi. Izmerili smo tudi fotokemiĉno
uĉinkovitost FS II kalic ter stehtali sveţo in suho maso kalic. Na koncu smo opravili še rastno
analizo, tako da smo izmerili višine in mase nakljuĉno izbranih kalic iz vsakega obravnavanja
ter prešteli vse kalice, ki so pri posameznemu obravnavanju vzklile iz semen.
Slika 2: Poskus, nastavljen 30. 3. 2016. Foto: Amela Osmić.
Slika 3: 10 dni stare kalice, katerih semena so bila obravnavana z I(V). Foto: Amela Osmić.

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
15
3.4 MERITVE
3.4.1 BIOKEMIJSKE IN MORFOLOŠKE MERITVE
Biokemijske in morfološke meritve smo opravljali v laboratoriju Biotehniške fakultete na
Katedri za ekologijo in varstvo okolja.
3.4.1.1 Klorofili in karotenoidi
Koncentracijo klorofila a in b ter karotenoidov smo doloĉali spektrofotometriĉno po metodi
Lichtenthalerja in Buschmanna (2001a, 2001b). Vzorce smo strli v terilnici, ekstrahirali v 5
mL 100 % acetona ter centrifugirali (4000 rpm, 4°C, 4 min) v centrifugirkah za klorofil. Po
konĉanem centrifugiranju smo odĉitali prostornine ekstraktov, nato smo izmerili ekstinkcije z
VIS spektrofotometrom pri valovnih dolţinah 470, 645 in 662 nm. Koncentracije klorofilov
(Kl a, b) in karotenoidov (Kar) smo izrazili na suho maso vzorca po naslednjih formulah:
Kl a (mg/g sm) = ca * V / sm / 1000 = (11,24 * E662 – 2,04 * E645) * V / sm / 1000 … (1)
Kl b (mg/g sm) = cb * V / sm / 1000 = (20,13 * E645 – 4,19 * E662) * V / sm / 1000 … (2)
Kar (mg/g sm) = (1000 * E470 – 1,9 * ca – 63,14 * cb) * V / sm / 214 / 1000 … (3)
ca, cb – koncentracija klorofila a, oz. klorofila b [mg/L]
V – volumen ekstrakta [mL]
sm – suha masa vzorca [g]
E – ekstinkcija pri izbrani valovni dolţini
3.4.1.2 Antociani
Koncentracijo antocianov v vzorcih smo doloĉali po metodi Drumm-a in Mohr-a (1978).
Vzorce smo strli v terilnici, ekstrahirali v 5 mL ekstrakcijskega medija (metanol : HCl (37 %)
= 99 : 1 (v/v)) ter centrifugirali (4000 rpm, 4°C, 4 min). Po konĉanem centrifugiranju smo
odĉitali prostornine ekstraktov in jih za 24 ur postavili v temo pri temperaturi 3 – 5°C.
Ekstinkcije smo izmerili z VIS spektrofotometrom pri valovni dolţini 530 nm. Koncentracije
antocianov (Ant) smo izrazili v relativnih enotah na suho maso vzorca po naslednji formuli:
Ant (relativna enota) = E530 * V * sm-1
… (4)
E530 – ekstinkcija pri valovni dolţini 530 nm
V – prostornina ekstrakta [mL]
sm – suha masa [g]

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
16
3.4.1.3 UV-absorbirajoče snovi
Koncentracijo absorbirajoĉih snovi UV-B IN UV-A (UV-B 280 – 320 nm, UV-A 320 – 400
nm) smo doloĉili po Caldwellovi metodi (1968). Vzorce smo strli v terilnici, ekstrahirali v 5
mL ekstrakcijskega medija (metanol : destilirana voda : HCl (37 %) = 79 : 20 : 1 (v/v)), 20
minut smo vzorce inkubirali. Nato smo jih centrifugirali (4000 rpm, 10°C, 10 min) ter odĉitali
prostornine ekstraktov. S pomoĉjo spektrofotometra UV/VIS in kvarĉnih kivet smo izmerili
ekstinkcije pri valovnih dolţinah od 280 do 400 nm. Koncentracije UV-absorbirajoĉih snovi
smo izraĉunali kot integral ekstinkcijskih vrednosti od 280 do 319 nm ter od 320 do 400 nm.
Koncentracije UV-absorbirajoĉih snovi smo izrazili v relativnih enotah na suho maso vzorca
po naslednji formuli:
UV abs (relativna enota) = I * V * sm
-1 … (5)
I - vsota absorpcijskih vrednosti v intervalu 280 – 319 nm (UV-B abs) ter 320 – 400 nm
(UV-A abs)
V - prostornina ekstrakta [mL]
sm - suha masa vzorca [g]
3.4.1.4 Fotokemična učinkovitost FS II
Fotokemiĉno uĉinkovitost rastline doloĉimo s pomoĉjo klorofilne fluorescence klorofila a v
FS II. Merili smo jo s pomoĉjo fluorometra (PAM 2100 Chlorophyll Fluorometer, Heinz
Walz GmbH, Nemĉija) v popoldanskem ĉasu med 14:00 h in 15:00 h pri sobni temperaturi.
Merili smo samo potencialno fotokemiĉno uĉinkovitost, ki se meri na zatemnjenih listih. Iz
vsake ponovitve posameznega obravnavanja smo vzeli po dve kalici, kar pomeni da smo pri
posameznem obravnavanju izbrali po 8 kalic. Na kliĉne liste kalic smo pred merjenjem
potencialne fotokemiĉne uĉinkovitosti pritrdili šĉipalke za temotno adaptacijo, zato da so se
sprejemniki elektronov oksidirali in vsi reakcijski centri v FS II odprli. Po 20 minutah
adaptacije smo list osvetlili s saturacijskim pulzom bele svetlobe (PPFD = 8000 µmol m-2
s-1
),
ki je trajal 0,8 s. Nato smo na napravi odĉitali potencialno fotokemiĉno uĉinkovitost, ki jo
izraţamo s parametrom Fv/Fm (Schreiber, Bilger in Neubauer, 1994).
3.4.1.5 Masa, višina kalic in število kalic
Kalice iz vsake ponovitve posameznega obravnavanja smo po konĉanih laboratorijskih
meritvah porezali tik nad koreniĉico in jih prešteli. Iz vsake ponovitve smo nakljuĉno izbrali
po deset kalic in izmerili višino vsake kalice posebej, nato pa smo vseh deset kalic skupaj tudi
stehtali. Zapakirali smo jih v aluminijasto folijo (po deset kalic iz vsake ponovitve
posameznega obravnavanja), na katero smo napisali iz katere ponovitve in obravnavanja so
kalice ter jih dali za 24 ur sušiti v sušilnik pri temperaturi 105° C. Po 24 urah sušenja smo
zopet stehtali njihovo maso. Vse ostale kalice smo dali v kuverte, oznaĉili ponovitev in
obravnavanje ter vse skupaj pospravili v zamrzovalnik, saj lahko na ta naĉin sluţijo za
nadaljnje analize.

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
17
3.5 STATISTIČNA OBDELAVA PODATKOV
Vse podatke meritev smo sproti vnašali v program Microsoft Office Excel 2007, s pomoĉjo
katerega smo izraĉunali povpreĉne vrednosti posameznih parametrov. Za statistiĉno obdelavo
podatkov smo uporabili raĉunalniški program SPSS Statistics, razliĉica 20.0 (IBM). S
pomoĉjo omenjenega raĉunalniškega programa smo izraĉunali standardno napako (SE). Zelo
pomemben predpogoj za nadaljne statistiĉne obdelave predstavlja informacija o normalni
porazdelitvi pridobljenih podatkov. Normalno porazdelitev podatkov smo testirali s Shapiro-
Wilkovim testom. Statistiĉno znaĉilne razlike med posameznimi obravnavanji pri
posameznem parametru smo preverili z enosmernim ANOVA testom in Duncanovim post-
hoc primerjalnim testom. Niĉelno hipotezo smo zavrnili, ko je bil p ≤ 0,05 (statistiĉno
znaĉilna razlika). Pri statistiĉni obdelavi podatkov smo med seboj primerjali rezultate, ki smo
jih dobili pri prvem poskusu in posebej rezultate, ki smo jih dobili po meritvah pri drugem
poskusu. Potem smo rezultate meritev prvega in drugega poskusa, za posamezen parameter,
predstavili v skupnem grafu.
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
18
4 REZULTATI
4.1 MORFOLOŠKE LASTNOSTI KALIC
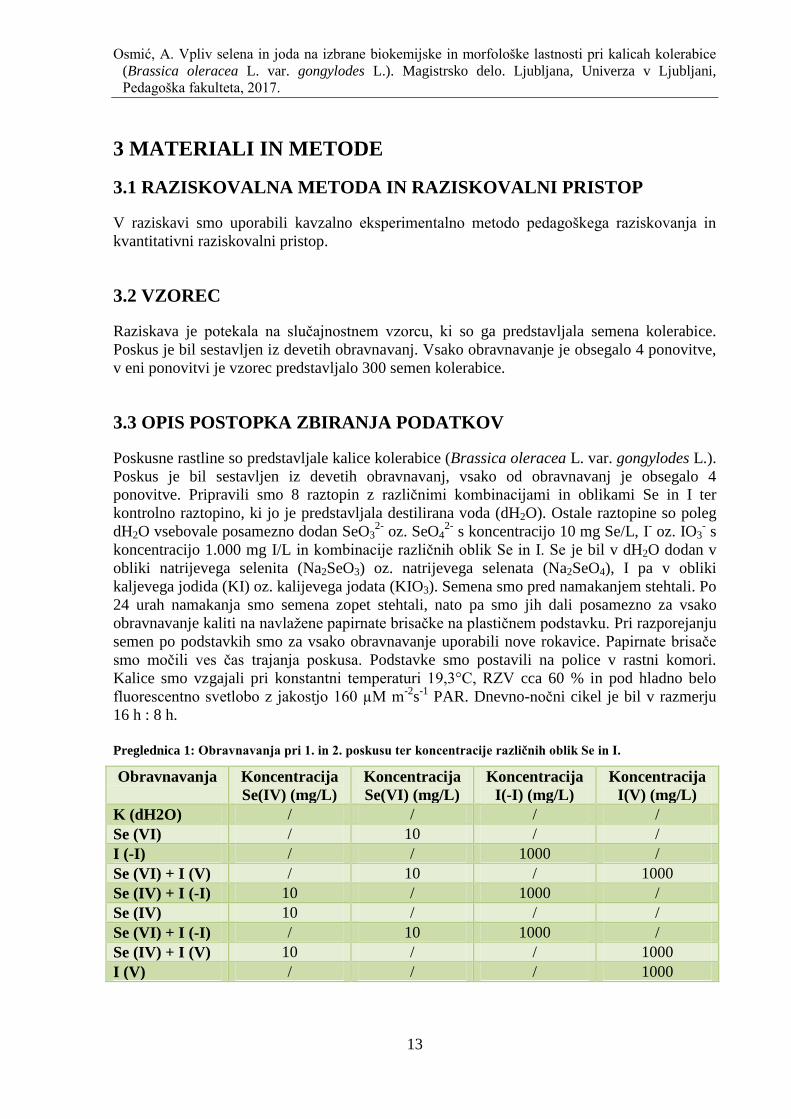
4.1.1 Sveţa masa
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno veĉjo sveţo maso od kalic,
obravnavanih s Se(VI)+I(V) in manjšo sveţo maso od kalic, obravnavanih z I(-I). Kalice,
obravnavane s Se(VI), so imele statistiĉno znaĉilno veĉjo sveţo maso od kalic, obravnavanih
s Se(IV). Prav tako so imele statistiĉno znaĉilno veĉjo sveţo maso od kalic, obravnavanih s
Se(VI)+I(V), Se(VI)+I(-I) in Se(IV)+I(V). Kalice, obravnavane z I(-I), so imele statistiĉno
znaĉilo veĉjo sveţo maso od kalic, obravnavanih z I(V). Imele so tudi statistiĉno znaĉilno
veĉjo sveţo maso od kalic, obravnavanih s Se(VI)+I(V), Se(IV)+I(-I), Se(IV), Se(VI)+I(-I) in
Se(IV)+I(V). Kalice, obravnavane s Se(IV)+I(-I) in I(V), so imele statistiĉno znaĉilno veĉjo
sveţo maso od kalic, obravnavanih s Se(VI)+I(V). Med ostalimi obravnavanji ni bilo
statistiĉno znaĉilnih razlik (Slika 4).
Pri 2. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo sveţo maso od kalic,
obravnavanih s Se(VI), Se(VI)+I(V), Se(IV)+I(-I), Se(IV) in Se(IV)+I(V). Kalice,
obravnavane s Se(VI)+I(-I) in I(V), so imele statistiĉno znaĉilno manjšo sveţo maso od kalic,
obravnavanih s Se(IV)+ I(-I). Med ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik
(Slika 4).
Slika 4: Sveţa masa kalic
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na sveţo maso kalic kolerabice. Rezultati so
predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi
ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z
razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri drugem poskusu (p < 0,05).
ac
cdd
b
ac
abab
abac
C
AB
ABCAB B AB
AC
ABAC
0,00
0,05
0,10
0,15
0,20
0,25
0,30
Sv
eţa
ma
sa k
ali
c (g
)
Obravnavanje
1. poskus
2. poskus
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
19
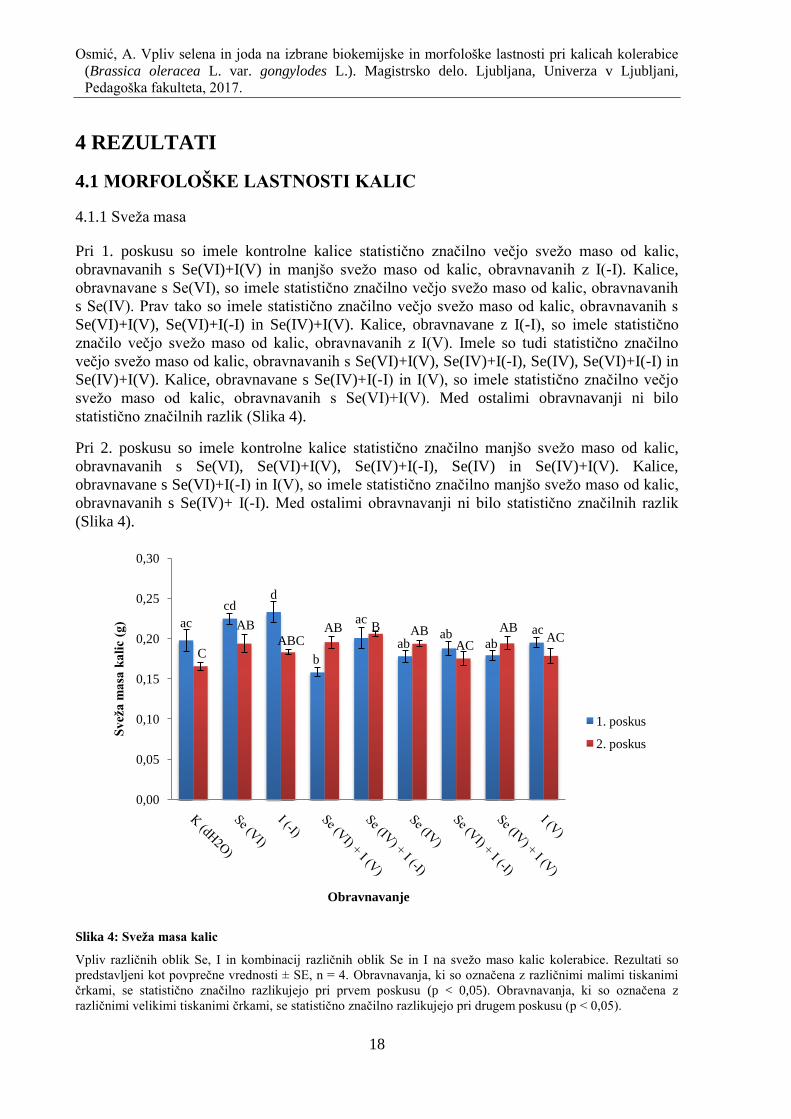
4.1.2 Suha masa
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo suho maso od kalic,
obravnavanih s Se(VI), I(-I), Se(IV)+I(-I) in Se(VI)+I(-I). Kalice, obravnavane s Se(IV)+I(-I),
so imele statistiĉno znaĉilno veĉjo suho maso od kalic, obravnavanih s Se(VI), Se(VI)+I(V),
Se(IV), Se(VI)+I(-I), Se(IV)+I(V) in I(V). Kalice, obravnavane z I(-I), so imele statistiĉno
znaĉilno veĉjo suho maso od kalic, obravnavanih s Se(VI)+I(V) in Se(IV)+I(V). Med ostalimi
obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 5).
Pri 2. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo suho maso od kalic,
obravnavanih s Se(VI)+I(V), Se(IV)+I(-I), Se(IV) in Se(IV)+I(V). Kalice, obravnavane s
Se(VI)+I(V) in Se(IV)+I(-1) so imele statistiĉno znaĉilno veĉjo suho maso od kalic,
obravnavanih s Se(VI), I(-I), Se(VI)+I(-I) in I(V). Kalice, obravnavane s Se(IV) in
Se(IV)+I(V), so imele statistiĉno znaĉilno veĉjo suho maso od kalic, obravnavanih s
Se(VI)+I(-I) in z I(V). Med ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 5).
Slika 5: Suha masa kalic
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na suho maso kalic kolerabice. Rezultati so
predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi
ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z
razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri drugem poskusu (p < 0,05).
b
accd
ab
d
abcac
ab abcAAB AB
CC
BC
A
BC
A
0,00
0,01
0,02
0,03
0,04
0,05
0,06
Su
ha
ma
sa k
ali
c (g
)
Obravnavanje
1. poskus
2. poskus
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
20
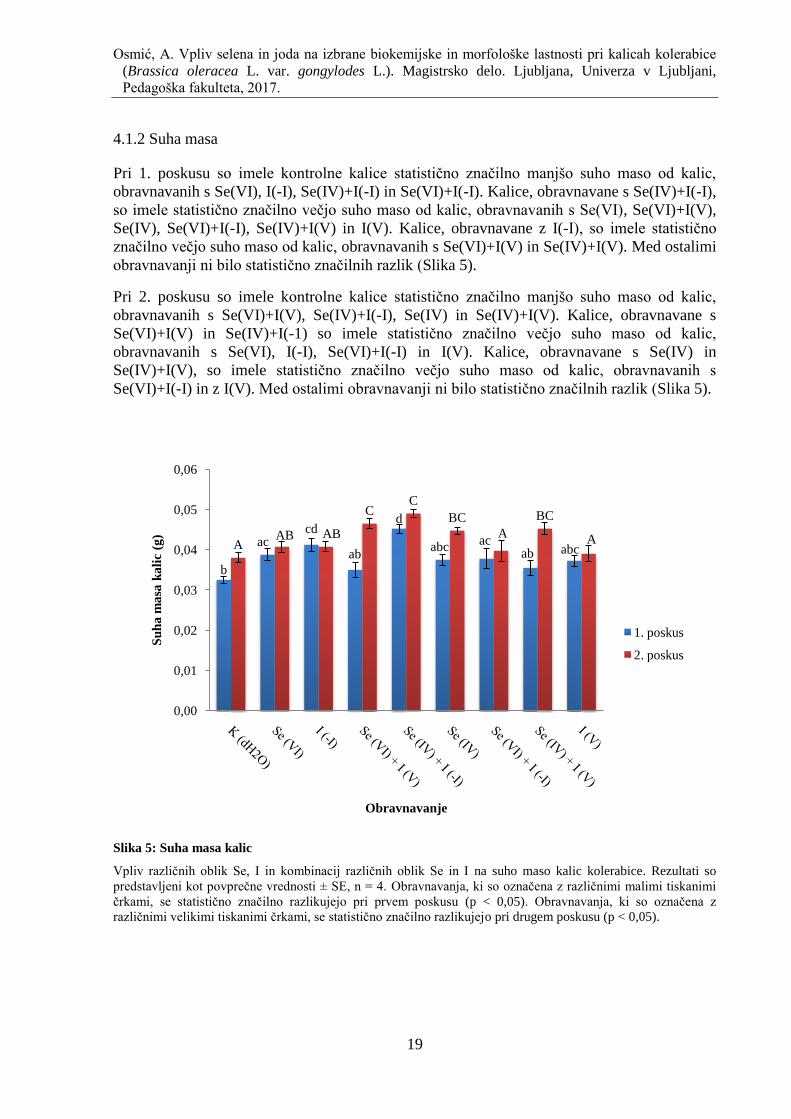
4.1.3 Višina kalic
Pri 1. poskusu so bile kalice, obravnavane s Se(VI), statistiĉno znaĉilno višje od kalic,
obravnavanih s Se(VI)+I(V). Kalice, obravnavane z I(-I), so bile statistiĉno znaĉilno višje od
kalic, obravnavanih s Se(VI)+I(V) in Se(IV). Med ostalimi obravnavanji ni bilo statistiĉno
znaĉilnih razlik (Slika 6).
Pri 2. poskusu so bile kalice, obravnavane s Se(IV)+I(-I), statistiĉno znaĉilno višje od kalic,
obravnavanih z I(V). Med ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 6).
Slika 6: Višina kalic
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na višino kalic kolerabice. Rezultati so
predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi
ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z
razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri drugem poskusu (p < 0,05).
abcbc
c
aabc
ab abc abc abcAB AB AB AB B
AB AB AB A
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
Viš
ina
ka
lic
(cm
)
Obravnavanje
1. poskus
2. poskus
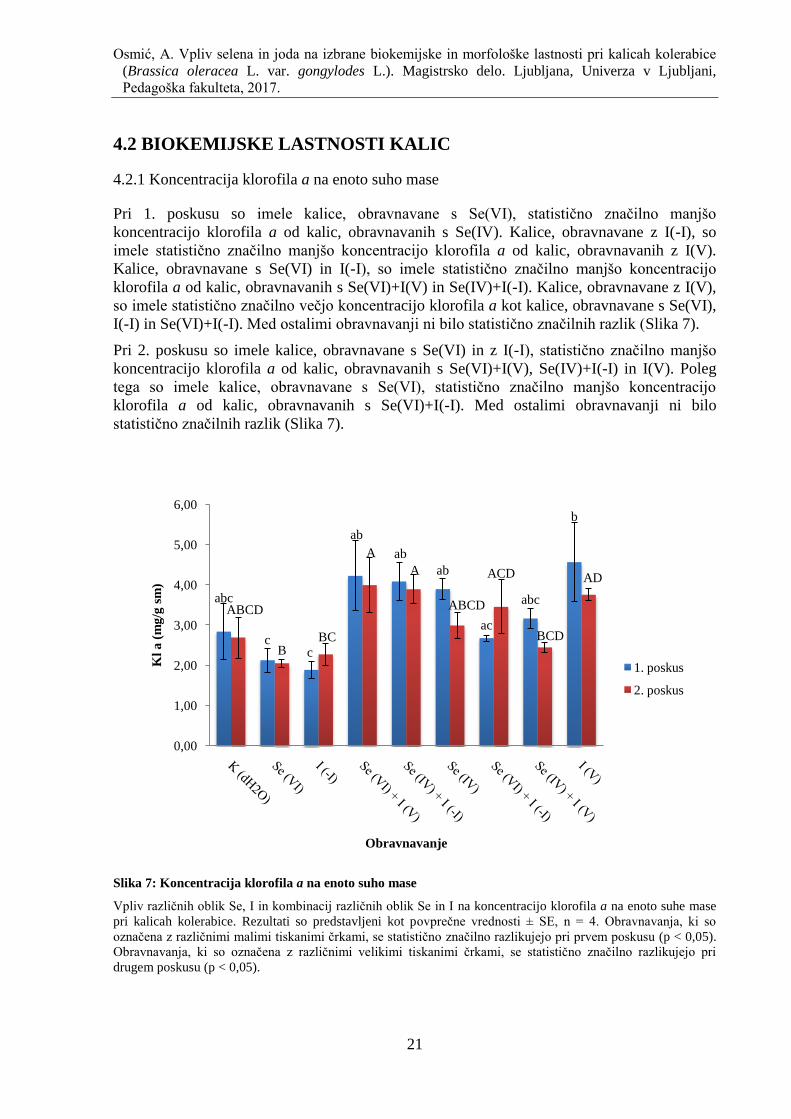
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
21
4.2 BIOKEMIJSKE LASTNOSTI KALIC
4.2.1 Koncentracija klorofila a na enoto suho mase
Pri 1. poskusu so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno manjšo
koncentracijo klorofila a od kalic, obravnavanih s Se(IV). Kalice, obravnavane z I(-I), so
imele statistiĉno znaĉilno manjšo koncentracijo klorofila a od kalic, obravnavanih z I(V).
Kalice, obravnavane s Se(VI) in I(-I), so imele statistiĉno znaĉilno manjšo koncentracijo
klorofila a od kalic, obravnavanih s Se(VI)+I(V) in Se(IV)+I(-I). Kalice, obravnavane z I(V),
so imele statistiĉno znaĉilno veĉjo koncentracijo klorofila a kot kalice, obravnavane s Se(VI),
I(-I) in Se(VI)+I(-I). Med ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 7).
Pri 2. poskusu so imele kalice, obravnavane s Se(VI) in z I(-I), statistiĉno znaĉilno manjšo
koncentracijo klorofila a od kalic, obravnavanih s Se(VI)+I(V), Se(IV)+I(-I) in I(V). Poleg
tega so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno manjšo koncentracijo
klorofila a od kalic, obravnavanih s Se(VI)+I(-I). Med ostalimi obravnavanji ni bilo
statistiĉno znaĉilnih razlik (Slika 7).
Slika 7: Koncentracija klorofila a na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo klorofila a na enoto suhe mase
pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so
oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05).
Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri
drugem poskusu (p < 0,05).
abc
cc
ab
ab
ab
ac
abc
b
ABCD
BBC
A
A
ABCD
ACD
BCD
AD
0,00
1,00
2,00
3,00
4,00
5,00
6,00
Kl
a (
mg
/g s
m)
Obravnavanje
1. poskus
2. poskus
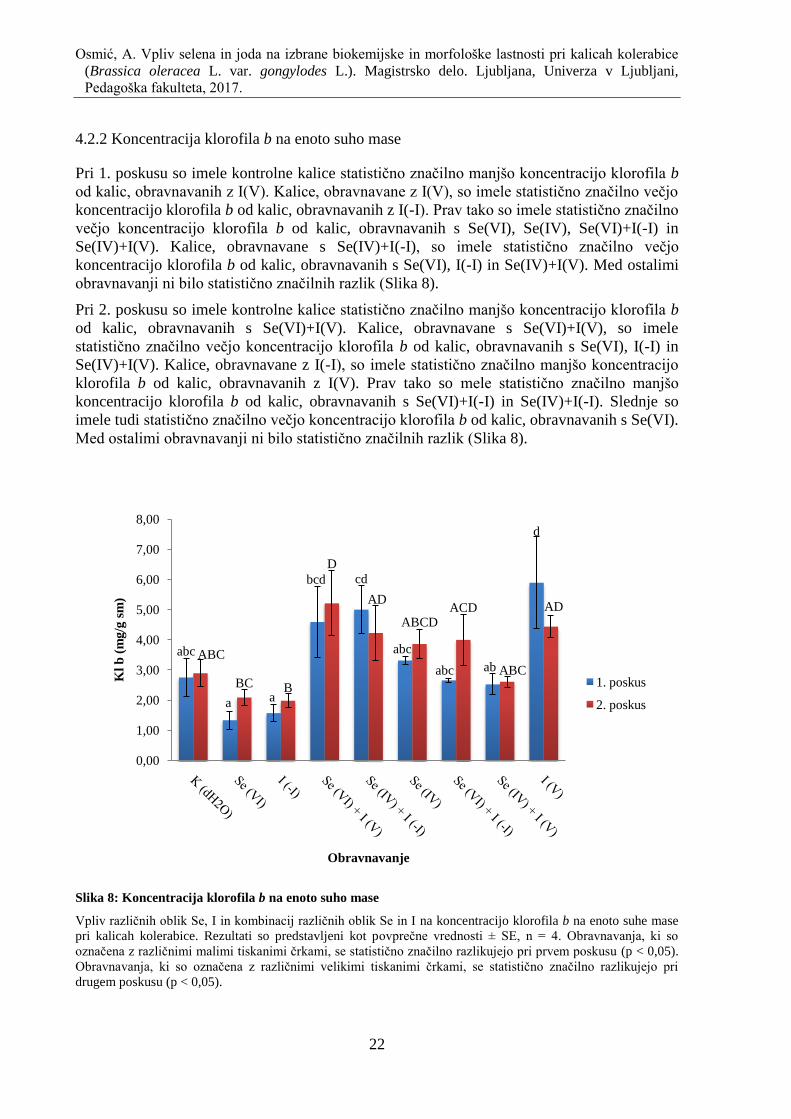
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
22
4.2.2 Koncentracija klorofila b na enoto suho mase
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo koncentracijo klorofila b
od kalic, obravnavanih z I(V). Kalice, obravnavane z I(V), so imele statistiĉno znaĉilno veĉjo
koncentracijo klorofila b od kalic, obravnavanih z I(-I). Prav tako so imele statistiĉno znaĉilno
veĉjo koncentracijo klorofila b od kalic, obravnavanih s Se(VI), Se(IV), Se(VI)+I(-I) in
Se(IV)+I(V). Kalice, obravnavane s Se(IV)+I(-I), so imele statistiĉno znaĉilno veĉjo
koncentracijo klorofila b od kalic, obravnavanih s Se(VI), I(-I) in Se(IV)+I(V). Med ostalimi
obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 8).
Pri 2. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo koncentracijo klorofila b
od kalic, obravnavanih s Se(VI)+I(V). Kalice, obravnavane s Se(VI)+I(V), so imele
statistiĉno znaĉilno veĉjo koncentracijo klorofila b od kalic, obravnavanih s Se(VI), I(-I) in
Se(IV)+I(V). Kalice, obravnavane z I(-I), so imele statistiĉno znaĉilno manjšo koncentracijo
klorofila b od kalic, obravnavanih z I(V). Prav tako so mele statistiĉno znaĉilno manjšo
koncentracijo klorofila b od kalic, obravnavanih s Se(VI)+I(-I) in Se(IV)+I(-I). Slednje so
imele tudi statistiĉno znaĉilno veĉjo koncentracijo klorofila b od kalic, obravnavanih s Se(VI).
Med ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 8).
Slika 8: Koncentracija klorofila b na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo klorofila b na enoto suhe mase
pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so
oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05).
Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri
drugem poskusu (p < 0,05).
abc
a a
bcd cd
abc
abc ab
d
ABC
BC B
D
AD
ABCDACD
ABC
AD
0,00
1,00
2,00
3,00
4,00
5,00
6,00
7,00
8,00
Kl
b (
mg
/g s
m)
Obravnavanje
1. poskus
2. poskus
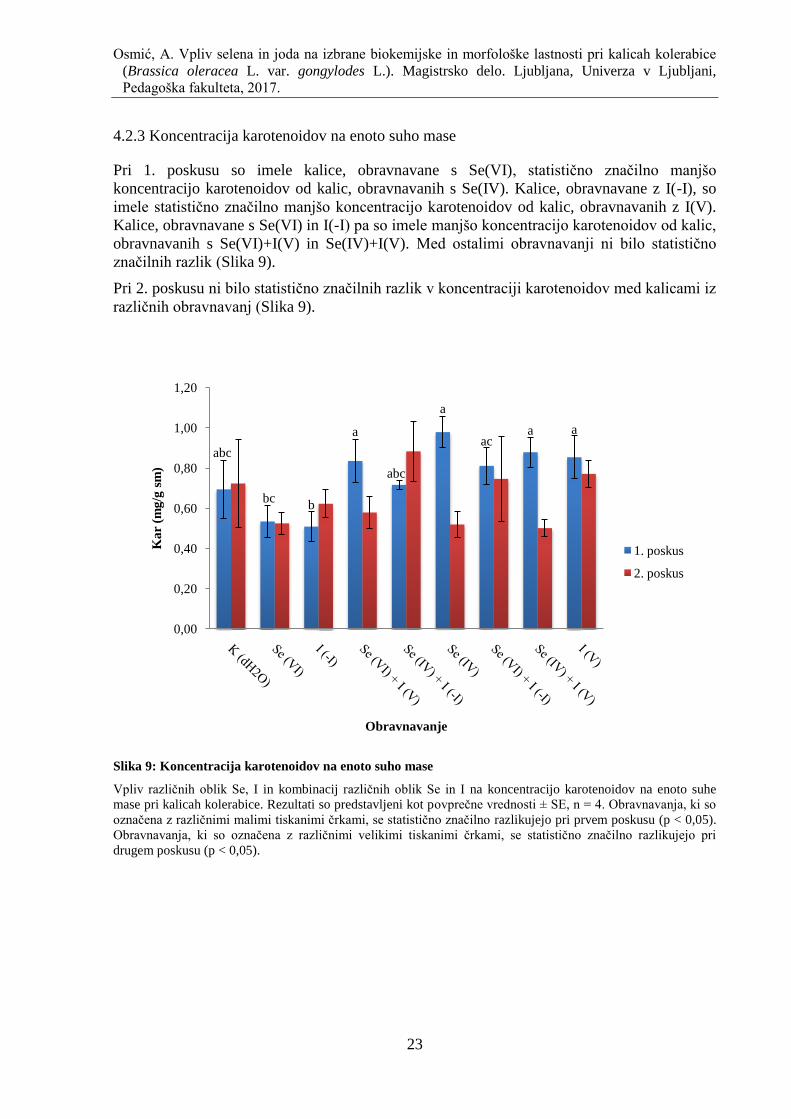
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
23
4.2.3 Koncentracija karotenoidov na enoto suho mase
Pri 1. poskusu so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno manjšo
koncentracijo karotenoidov od kalic, obravnavanih s Se(IV). Kalice, obravnavane z I(-I), so
imele statistiĉno znaĉilno manjšo koncentracijo karotenoidov od kalic, obravnavanih z I(V).
Kalice, obravnavane s Se(VI) in I(-I) pa so imele manjšo koncentracijo karotenoidov od kalic,
obravnavanih s Se(VI)+I(V) in Se(IV)+I(V). Med ostalimi obravnavanji ni bilo statistiĉno
znaĉilnih razlik (Slika 9).
Pri 2. poskusu ni bilo statistiĉno znaĉilnih razlik v koncentraciji karotenoidov med kalicami iz
razliĉnih obravnavanj (Slika 9).
Slika 9: Koncentracija karotenoidov na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo karotenoidov na enoto suhe
mase pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so
oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05).
Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri
drugem poskusu (p < 0,05).
abc
bcb
a
abc
a
aca a
0,00
0,20
0,40
0,60
0,80
1,00
1,20
Ka
r (m
g/g
sm
)
Obravnavanje
1. poskus
2. poskus
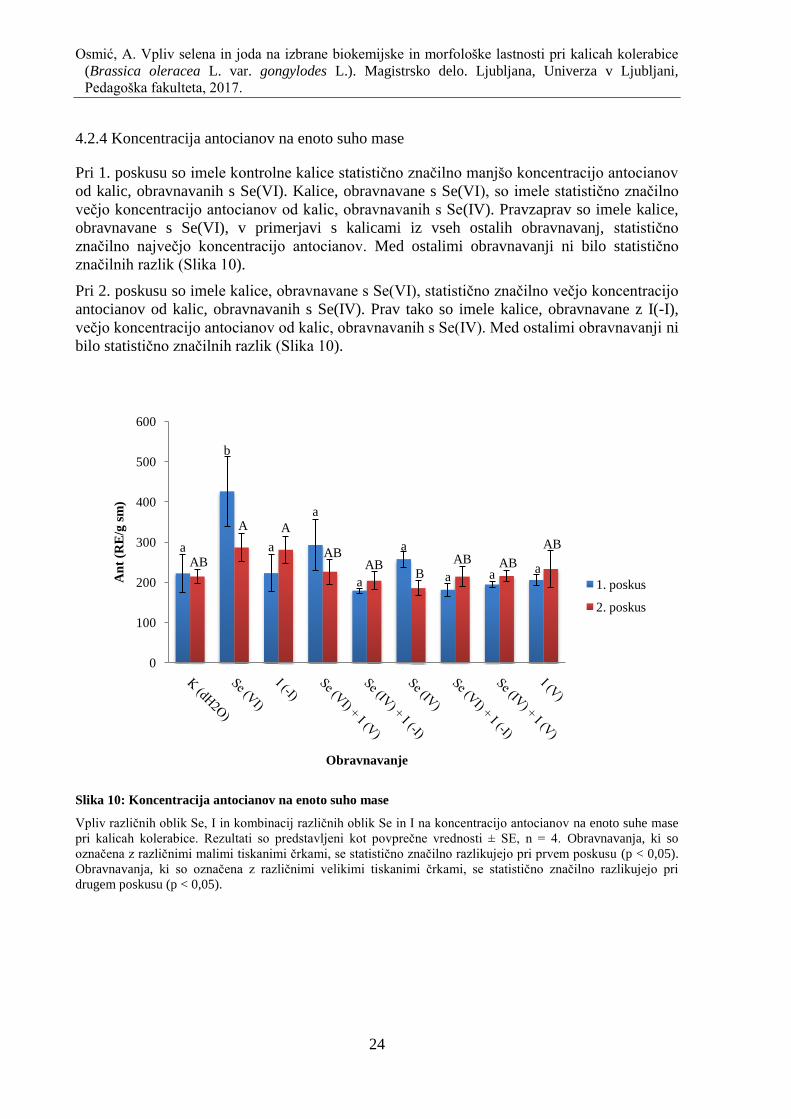
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
24
4.2.4 Koncentracija antocianov na enoto suho mase
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo koncentracijo antocianov
od kalic, obravnavanih s Se(VI). Kalice, obravnavane s Se(VI), so imele statistiĉno znaĉilno
veĉjo koncentracijo antocianov od kalic, obravnavanih s Se(IV). Pravzaprav so imele kalice,
obravnavane s Se(VI), v primerjavi s kalicami iz vseh ostalih obravnavanj, statistiĉno
znaĉilno najveĉjo koncentracijo antocianov. Med ostalimi obravnavanji ni bilo statistiĉno
znaĉilnih razlik (Slika 10).
Pri 2. poskusu so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno veĉjo koncentracijo
antocianov od kalic, obravnavanih s Se(IV). Prav tako so imele kalice, obravnavane z I(-I),
veĉjo koncentracijo antocianov od kalic, obravnavanih s Se(IV). Med ostalimi obravnavanji ni
bilo statistiĉno znaĉilnih razlik (Slika 10).
Slika 10: Koncentracija antocianov na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo antocianov na enoto suhe mase
pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so
oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05).
Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri
drugem poskusu (p < 0,05).
a
b
a
a
a
a
a a aAB
A A
ABAB
BAB AB
AB
0
100
200
300
400
500
600
An
t (R
E/g
sm
)
Obravnavanje
1. poskus
2. poskus
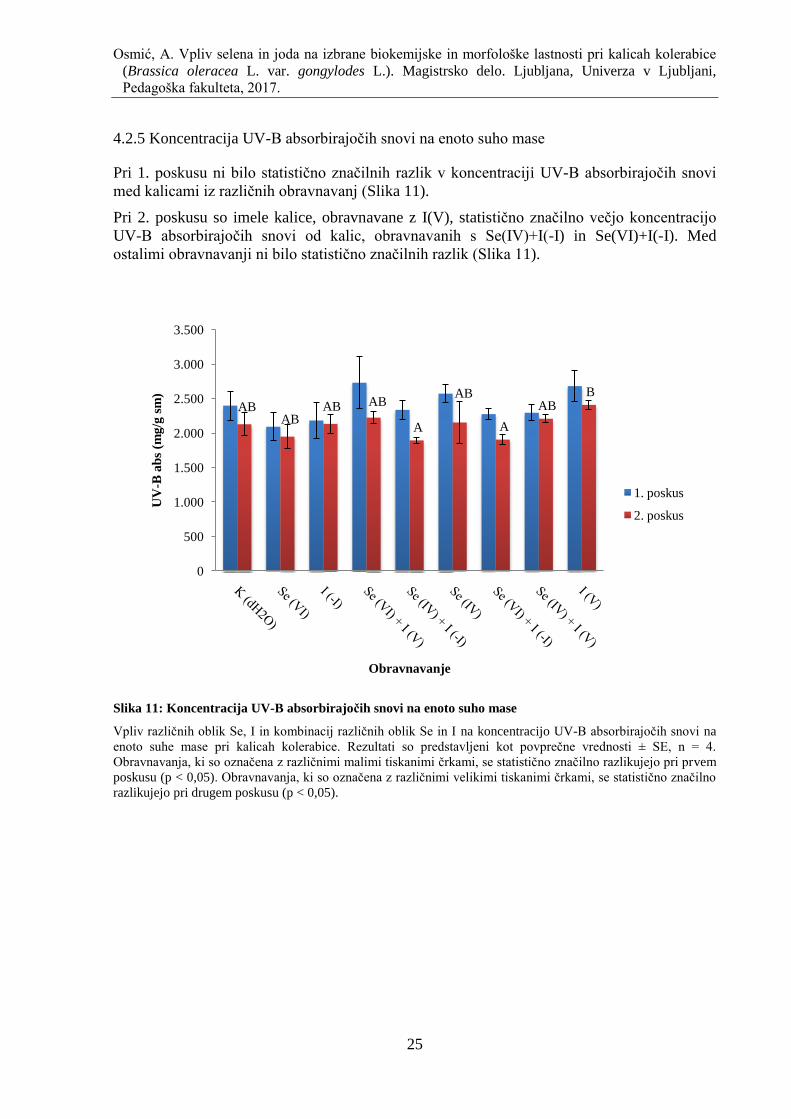
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
25
4.2.5 Koncentracija UV-B absorbirajoĉih snovi na enoto suho mase
Pri 1. poskusu ni bilo statistiĉno znaĉilnih razlik v koncentraciji UV-B absorbirajoĉih snovi
med kalicami iz razliĉnih obravnavanj (Slika 11).
Pri 2. poskusu so imele kalice, obravnavane z I(V), statistiĉno znaĉilno veĉjo koncentracijo
UV-B absorbirajoĉih snovi od kalic, obravnavanih s Se(IV)+I(-I) in Se(VI)+I(-I). Med
ostalimi obravnavanji ni bilo statistiĉno znaĉilnih razlik (Slika 11).
Slika 11: Koncentracija UV-B absorbirajočih snovi na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo UV-B absorbirajoĉih snovi na
enoto suhe mase pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4.
Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem
poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno
razlikujejo pri drugem poskusu (p < 0,05).
ABAB
AB AB
A
AB
A
ABB
0
500
1.000
1.500
2.000
2.500
3.000
3.500
UV
-B a
bs
(mg
/g s
m)
Obravnavanje
1. poskus
2. poskus
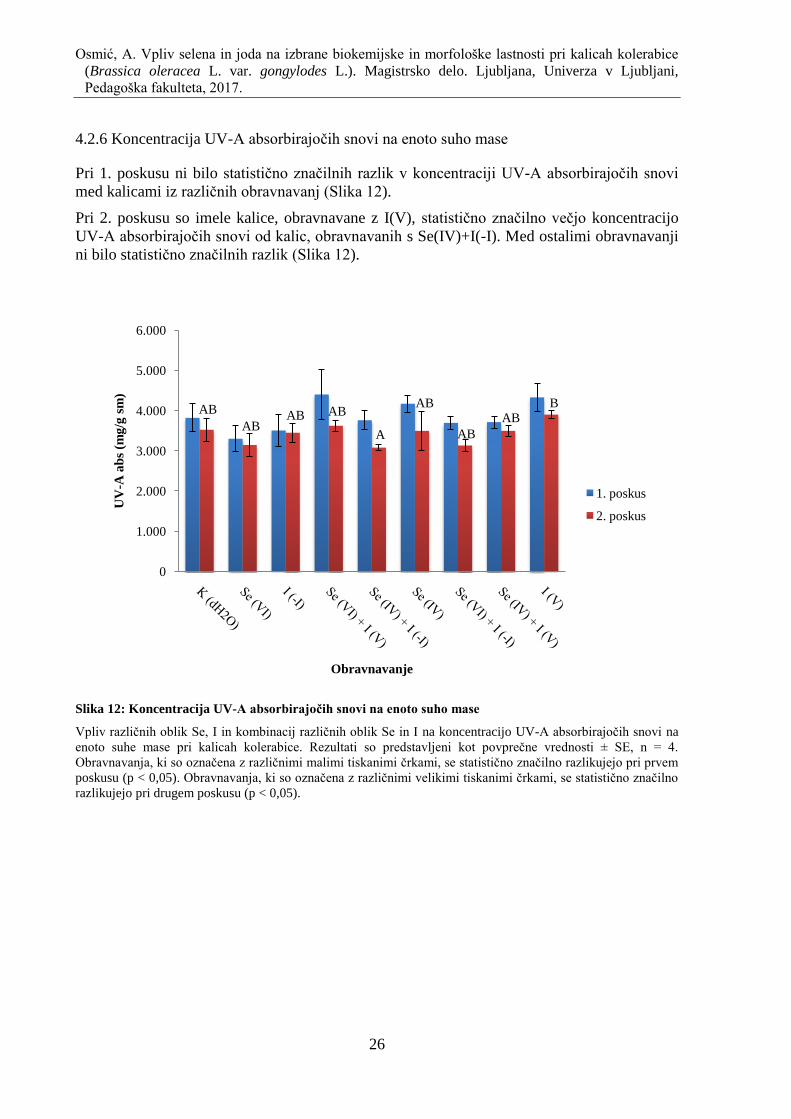
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
26
4.2.6 Koncentracija UV-A absorbirajoĉih snovi na enoto suho mase
Pri 1. poskusu ni bilo statistiĉno znaĉilnih razlik v koncentraciji UV-A absorbirajoĉih snovi
med kalicami iz razliĉnih obravnavanj (Slika 12).
Pri 2. poskusu so imele kalice, obravnavane z I(V), statistiĉno znaĉilno veĉjo koncentracijo
UV-A absorbirajoĉih snovi od kalic, obravnavanih s Se(IV)+I(-I). Med ostalimi obravnavanji
ni bilo statistiĉno znaĉilnih razlik (Slika 12).
Slika 12: Koncentracija UV-A absorbirajočih snovi na enoto suho mase
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na koncentracijo UV-A absorbirajoĉih snovi na
enoto suhe mase pri kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4.
Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem
poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno
razlikujejo pri drugem poskusu (p < 0,05).
AB
ABAB AB
A
AB
AB
ABB
0
1.000
2.000
3.000
4.000
5.000
6.000
UV
-A a
bs
(mg
/g s
m)
Obravnavanje
1. poskus
2. poskus
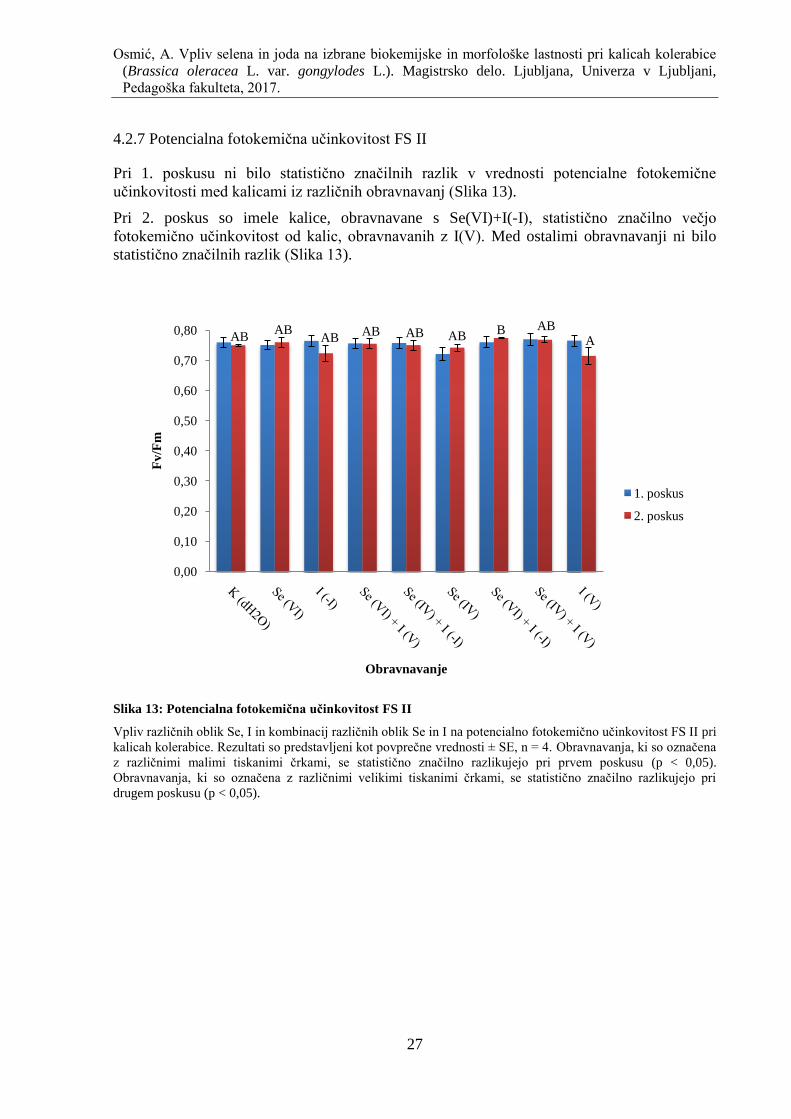
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
27
4.2.7 Potencialna fotokemiĉna uĉinkovitost FS II
Pri 1. poskusu ni bilo statistiĉno znaĉilnih razlik v vrednosti potencialne fotokemiĉne
uĉinkovitosti med kalicami iz razliĉnih obravnavanj (Slika 13).
Pri 2. poskus so imele kalice, obravnavane s Se(VI)+I(-I), statistiĉno znaĉilno veĉjo
fotokemiĉno uĉinkovitost od kalic, obravnavanih z I(V). Med ostalimi obravnavanji ni bilo
statistiĉno znaĉilnih razlik (Slika 13).
Slika 13: Potencialna fotokemična učinkovitost FS II
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na potencialno fotokemiĉno uĉinkovitost FS II pri
kalicah kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4. Obravnavanja, ki so oznaĉena
z razliĉnimi malimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri prvem poskusu (p < 0,05).
Obravnavanja, ki so oznaĉena z razliĉnimi velikimi tiskanimi ĉrkami, se statistiĉno znaĉilno razlikujejo pri
drugem poskusu (p < 0,05).
ABAB
ABAB AB AB
B ABA
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
0,80
Fv
/Fm
Obravnavanje
1. poskus
2. poskus

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
28
5 RAZPRAVA
Na kolerabici je bilo narejeno malo raziskav o vplivu Se in I, zato smo naše rezultate
primerjali z rezultati raziskav, ki so bile opravljene na drugih rastlinah. Vpliv preuĉevanih
dejavnikov je bil podoben v 1. in 2. poskusu. Odziv kalic na razliĉne kemijske oblike Se in I
ter njune kombinacije je bil razliĉen in se je razlikoval med posameznimi merjenimi
parametri. Uporabljene koncentracije obeh elementov ter njunih kombinacij veĉinoma niso
delovale negativno na izbrane biokemijske in morfološke lastnosti ter na rast kalic kolerabice.
Torej ne moremo izpostaviti doloĉene kemijske oblike Se in I oz. njune kombinacije, ki bi
delovale strupeno na naše poskusne rastline. Prav tako smo pri naši raziskavi dobili zelo malo
razlik med razliĉnimi obravnavanji, pri doloĉenih merjenih parametrih, na kar bi lahko
vplivala koncentracija dodanega Se in I. Mogoĉe bi veĉje koncentracije preuĉevanih
elementov imele veĉji vpliv na kolerabico.
5.1 MASA, VIŠINA IN ŠTEVILO KALIC
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo sveţo maso od kalic,
obravnavanih z I(-I) in veĉjo sveţo maso od kalic, obravnavanih s Se(VI)+I(V). To pomeni,
da je obravnavanje z I(-I) pozitivno vplivalo na sveţo maso kalic, medtem ko je obravnavanje
s Se(VI)+I(V) imelo negativen vpliv. Kalice, obravnavane s Se(VI), so imele statistiĉno
znaĉilno veĉjo sveţo maso od kalic, obravnavanih s Se(IV). Kalice, katerih semena so bila
obravnavana z I(-I), so imele statistiĉno znaĉilno veĉjo sveţo maso od kalic, obravnavanih z
I(V) (Slika 4).
Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo suho maso od kalic,
obravnavanih s Se(VI), I(-I), Se(IV)+I(-I) in Se(VI)+I(-I), kar pomeni, da so naštetata
obravnavanja pozitivno vplivala na suho maso kalic kolerabice. Tudi pri 2. poskusu je
obravnavanje s Se(IV)+I(-I) pozitivno vplivalo na suho maso kalic, ki je bila veĉja od suhe
mase kontrolnih kalic. Kontrolne kalice so imele prav tako statistiĉno znaĉilno manjšo suho
maso od kalic, obravnavanih s Se(VI)+I(V), Se(IV) in Se(IV)+I(V), kar pomeni da so ta
obravnavanja prav tako pozitivno vplivala na suho maso kalic kolerabice (Slika 5).
Pri 1. poskusu so bile kalice, katerih semena so bila obravnavana s Se(VI), statistiĉno
znaĉilno višje od kalic, obravnavanih s Se(VI)+I(V). Kalice, obravnavane z I(-I) so bile
statistiĉno znaĉilno višje od kalic, obravnavanih s Se(IV) in Se(VI)+I(V). Pri 2. poskusu so
bile kalice, obravnavane z I(V), statistiĉno znaĉilno niţje od kalic, obravnavanih s Se(IV)+I(-
I) (Slika 6).
Pri številu kalic ni bilo statistiĉno znaĉilnih razlik med obravnavanji pri nobenem od
poskusov. Procent kaljivosti pri posameznih obravnavanjih je bil 67 – 68% pri kontroli, 71 –
73% pri Se(VI), 70 – 72% pri I(-I), 68 – 72% pri Se(VI)+I(V), 74 – 75% pri Se(IV)+I(-I), 71
– 73% pri Se(IV), 68 – 69% pri Se(VI)+I(-I), 72 – 73% pri Se(IV)+I(V) in 70 – 73% pri I(V)
(Priloga A).
Hajiboland in Keivanfar (2012) sta raziskovala vpliv Se(VI) na oljno repico (Brassica napus).
Rastline oljne repice sta 19 tednov listno škropila s Se(VI) pri koncentracijah 0 (kontrola),
0,01 in 0,02 mg Se/L. Ţelela sta raziskati njegov vpliv na rast rastlin. Ugotovila sta, da je bila
suha masa strokov in semen bistveno veĉja pri rastlinah, obravnavanih s Se(VI), kot pri
kontrolnih rastlinah. V nasprotju z drugimi parametri je obravnavanje s Se(VI) vplivalo na

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
29
zmanjšanje števila strokov, vendar ta uĉinek ni bil statistiĉno znaĉilen. Število listov pri
rastlinah oljne repice je bilo nekoliko veĉje pri koncentraciji 0,01 mg Se/L, medtem ko se
število listov pri rastlinah, ki so bile obravnavane s Se(VI), pri koncentraciji 0,02 mg Se/L, ni
razlikovalo od števila listov pri kontrolnih rastlinah. Nasprotno je bila dolţina socvetja v
reproduktivni fazi veĉja pri rastlinah, ki so bile obravnavane s Se(VI) pri koncentraciji 0,02
mg Se/L, kot pri rastlinah, ki so bile obravnavane s Se(VI) pri koncentraciji 0,01 mg Se/L.
Dodatek Se(VI) je pri obeh koncentracijah (0,01 in 0,02 mg Se/L) pospešil rast rastlin oljne
repice. Akbulut in Cakir (2010) sta raziskovala vpliv Se(VI) na velikost kalic jeĉmena.
Ugotovila sta, da so bile kalice, obravnavane s Se(VI), pri koncentracijah 2, 8 in 16 mg Se/L,
niţje od kontrolnih kalic. Pri našem poskusu so bile kalice, obravnavane s Se(VI), statistiĉno
znaĉilno višje od kalic, obravnavanih s Se(VI)+I(V) (Slika 6). Listno škropljenje s Se(VI) ni
vplivalo na suho maso in višino nadzemnega dela ter na suho maso korenin in semen pri dveh
sortah navadne ajde (Tadina idr., 2007). Nasprotno je listno škropljenje krompirja s Se(VI) pri
koncentraciji 10 mg Se/L vplivalo na zmanjšanje mase gomoljev (Germ, Kreft, Stibilj in
Urbanc-Berĉiĉ, 2007a). Uporaba Se ni statistiĉno znaĉilno vplivala na suho maso rastlin
rdeĉega zelja, ki so bile obravnavane vsak drugi dan, 2 meseca, z vodno raztopino Se(VI), pri
koncentraciji 0,002 mg Se/L in 0,50 mg Se/L, s katero so pognojili tla dvakrat v testnem
obdobju (Mechora idr., 2011). Tudi pri rdeĉem radiĉu, ki so ga listno škropili z vodno
raztopino Na2SeO4, pri koncentraciji 1 mg Se/L, raziskovalci niso opazili pomembnega vpliva
na suho maso rastlin (Germ idr., 2007c). Carvalho, Gallardo-Williams, Benson in Martin
(2003) so v svojo raziskavo vkljuĉili rastline paradiţnika, jagod, redkve in solate. Pri
morfoloških lastnostih so se osredotoĉili predvsem na razlike v številu plodov ter teţi rastlin,
ki so jih primerjali s kontrolnimi rastlinami. V tla, kjer so rastline uspevale, so dodali razliĉni
obliki (organski in anorganski) in koncentraciji (1,5 in 40 mg Se/kg) Se. Rezultati raziskave
so pokazali statistiĉno znaĉilno razliko v številu plodov med kontrolnimi rastlinami
paradiţnika in rastlinami, ki so bile obravnavane z 40 mg Se/kg v organski obliki. Število
plodov paradiţnika se je, v primerjavi s kontrolnimi rastlinami, zmanjšalo pri rastlinah
paradiţnika, ki so bile obravnavane s Se v organski obliki. Med ostalimi obravnavanji ni bilo
statistiĉno znaĉilnih razlik. V naši raziskavi smo uporabili veĉjo koncentracijo Se(VI) (10 mg
Se/L), ki ni imela statistiĉno pomembnega vpliva na konĉno število kalic, ki so vzklile iz
semen (Priloga A). Carvalho idr. (2003) so ugotovili, da tudi pri jagodah ni bilo statistiĉno
znaĉilnih razlik v številu plodov med kontrolnimi in s Se obravnavanimi rastlinami. Tako pri
paradiţniku kot tudi pri jagodah ni bilo statistiĉno znaĉilnih razlik v sveţi masi plodov med
posameznimi obravnavanji, medtem ko se je sveţa masa rastlin radiĉa, katerih semena so bila
obravnavana s 40 mg Se/kg v organski obliki in 1,5 mg Se/kg v anorganski obliki, statistiĉno
znaĉilno razlikovala od kontrolnih rastlin. Prav tako se je sveţa masa rastlin vrtne solate,
katere semena so bila obravnavana z 1,5 mg Se/kg v organski obliki, statistiĉno znaĉilno
razlikovala od kontrolnih rastlin. Pri naši raziskavi so imele kalice, obravnavane s Se(VI),
statistiĉno znaĉilno veĉjo sveţo maso od kalic, obravnavanih s Se(IV). Prav tako so imele
veĉjo sveţo maso od kalic, obravnavanih s Se(VI)+I(V) in Se(VI)+I(-I) (Slika 4).
Obravnavanje s Se(VI) je pozitivno vplivalo na suho maso kalic pri 1. poskusu, saj je bila
statistiĉno znaĉilno veĉja od suhe mase kontrolnih kalic (Slika 5).
Dodatek I(-I) v obliki KI in pri koncentraciji 10 mg I/L, je znatno zmanjšal biomaso rastlin
vrtne solate v primerjavi s kontrolnimi rastlinami, medtem ko je dodatek I(V), v obliki KIO3,
poveĉal biomaso rastlin, ki je dosegla najveĉjo vrednost pri odmerku 2,5 mg I/L (Blasco idr.,
2011). Obravnavanje z I(V) ni pokazalo statistiĉno znaĉilnega vpliva na biomaso poganjkov
vrtne solate (Blasco idr., 2008). Listno škropljenje rastlin redkvice (Raphanus sativus) z I(-I)
oz. I(V) ni vplivalo na suho maso listov in korenin (Strzetelski idr., 2010). Pri naši raziskavi

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
30
so imele kontrolne kalice statistiĉno znaĉilno manjšo sveţo maso od kalic, obravnavanih z I(-
I), kar pomeni, da je obravnavanje z I(-I) pozitivno vplivalo na sveţo maso kalic. Prav tako so
imele kalice, obravnavane z I(-I), veĉjo sveţo maso od kalic, katerih semena so bila
obravnavana z I(V) (Slika 4). Blasco idr. (2008) so ugotovili, da je imelo obravnavanje rastlin
vrtne solate z I(-I) (≥ 10 mg I/L) strupen uĉinek na rast rastlin zaradi prekomernega kopiĉenja
tega elementa v rastlinskem tkivu. Rezultati naše raziskave so pokazali, da so bile kalice,
obravnavane z I(V), statistiĉno znaĉilno niţje od kalic, obravnavanih s Se(IV)+I(-I) (Slika 6).
Smoleń idr. (2014) so ugotovili, da obravnavanje rastlin vrtne solate s Se(VI)+I(V) ni imelo
uĉinka na povpreĉno maso glav solate. Oba elementa so dodali v hranilno raztopino, kjer so
rastline uspevale. Se(VI) je bil dodan v hranilno raztopino v obliki Na2SeO4, pri
koncentracijah 0,5 in 1,5 mg Se/L, I(V) pa v obliki KIO3 in koncentraciji 1 mg I/L. Tudi Zhu
idr. (2004) so dodali Se(VI) in I(V) v hranilno raztopino, kjer so uspevale rastline špinaĉe.
Se(VI) je bil dodan v koncentracijah 0,8, 1,6 in 4 mg Se/L, medtem ko je bil I(V) dodan v
koncentracijah 1,25, 2,5 in 6,25 mg I/L. Ugotovili so, da je dodatek Se(VI) pri koncentraciji
0,8 in 1,6 mg Se/L ter I(V) pri koncentraciji 1,25 oz. 6,25 mg I/L zmanjšal biomaso korenin
ter da je biomasa poganjkov špinaĉe teţila k zmanjšanju ob dodatku obeh elementov.
Rezultati naše raziskave so pri 1. poskusu pokazali negativen vpliv dodatka Se(VI)+I(V) na
sveţo maso rastlin, saj je bila ta statistiĉno znaĉilno manjša od sveţe mase kontrolnih kalic.
Pri 2. poskusu so bili rezultati ravno obratni in je bila sveţa masa kalic, obravnavanih s
Se(VI)+I(V), statistiĉno znaĉilno veĉja od sveţe mase kontrolnih kalic (Slika 4). Suha masa
kalic, obravnavanih s Se(VI)+I(V), pri 1. poskusu, je bila statistiĉno znaĉilno manjša od kalic,
obravnavanih s Se(IV)+I(-I) in z I(-I) in veĉja od kalic, obravnavanih (Slika 5). Kalice,
obravnavane s Se(VI)+I(V) so bile statistiĉno znaĉilno niţje od kalic, obravnavanih s Se(VI)
in z I(-I) (Slika 6).
5.2 KONCENTRACIJE BARVIL
Rastline se na spremembe v okolju prilagajajo na razliĉne naĉine, med katere sodijo tudi
spremembe v koncentraciji barvil (Tausz idr., 1999). V naši raziskavi so razliĉne kemijske
oblike Se in I ter njune kombinacije razliĉno vplivale na koncentracijo klorofila a, b in
karotenoidov, medtem ko na koncentracijo antocianov ter UV-A in UV-B absorbirajoĉih
snovi niso statistiĉno znaĉilno vplivale.
Pri naši raziskavi so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno manjšo
koncentracijo klorofila a (Slika 7) in karotenoidov (Slika 9) od kalic, katerih semena so bila
obravnavana s Se(IV). Obratno so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno
veĉjo koncentracijo antocianov od kalic, obravnavanih s Se(IV) (Slika 10). Kalice,
obravnavane z I(-I), so imele statistiĉno znaĉilno manjšo koncentracijo klorofila a (Slika 7),
klorofila b (Slika 8) in karotenoidov (Slika 9) od kalic, obravnavanih z I(V). Pri 1. poskusu so
imele kontrolne kalice statistiĉno znaĉilno manjšo koncentracijo klorofila b od kalic,
obravnavanih z I(V), medtem ko so pri 2. poskusu imele manjšo koncentracijo klorofila b od
kalic, obravnavanih s Se(VI)+I(V) (Slika 8). To pomeni, da sta oba obravnavanja pozitivno
vplivala na koncentracijo klorofila b pri kalicah. Pri. 1 poskusu je na koncentracijo antocianov
pozitivno vplivalo obravnavanje s Se(VI), saj so te kalice imele statistiĉno znaĉilno veĉjo
vsebnost antocianov od kontrolnih kalic (Slika 10). Pri 2. poskusu so imele kalice,
obravnavane z I(V), statistiĉno znaĉilno veĉjo koncentracijo UV-B (Slika 11) in UV-A (Slika
12) absorbirajoĉih snovi od kalic, obravnavanih s Se(IV)+I(-I).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
31
Germ idr. (2015) so naredili raziskavo na kalicah navadne ajde. Pri raziskavi so uporabili
enake koncentracije in oblike Se in I ter njune kombinacije, kot smo jih mi uporabili v naši
raziskavi. Ugotovili so, da so imele kontrolne kalice in kalice, katerih semena so bila
namoĉena v Se(VI) in Se(VI)+I(-I), najmanjšo ter podobno koncentracijo klorofila a in
karotenoidov. Tudi Mechora in Vrhovšek (2012) sta prouĉevala vpliv Se(VI), pri
koncentraciji 10 mg Se/L, na koncentracijo karotenoidov pri dveh vrstah makrofitov (klasasti
rmanec in navadni rogolist) in opazila porast v koncentraciji karotenoidov, vendar brez
statistiĉno znaĉilnih razlik. Mechora idr. (2011) so prouĉevali vpliv Se(VI) na rastline rdeĉega
zelja (Brassica oleracea var. capitata L. f. rubra, cv. Erfurtsko rano). Rastline so posejali in
jih 3 tedne zalivali z raztopino Se(VI). Poskus je bil izveden na dveh skupinah rastlin rdeĉega
zelja. Prvo skupino rastlin so pognojili s 33 ml vodne raztopine, ki je vsebovala Na2SeO4 s
koncentracijo 0,002 mg Se/L, 2 meseca vsak drugi dan. Drugo skupino pa so v 2 mesecih
pognojili samo dvakrat z 0,5 mg Se/L. Ugotovili so, da dodatek Se(VI) ni vplival na
koncentracijo klorofila a, b in antocianov pri rastlinah rdeĉega zelja tako iz 1. kot tudi iz 2.
skupine. Breznik, Germ, Gaberšĉik in Kreft (2005) so listno škropili tatarsko in navadno ajdo
z dH2O, ki je vsebovala Se(VI) v koncentraciji 1 mg Se/L in ugotovili, da ni bilo vpliva na
koncentracijo klorofilov pri nobeni vrsti ajde. Malorgio, Diaz, Ferrante, Mensuali-Sodi in
Pezzarossa (2009) so hidroponiĉno vzgojili zeleno solato (Lactuca sativa L. var. acephala) in
radiĉ (Chicorium intybus L.). V hranilno raztopino so dodali 0,5 in 1,0 mg Se/L v obliki
Na2SeO4. Prvi poskus je bil izveden jeseni, drugi pozimi in tretji spomladi. Po ţetvi so izmerili
koncentracijo klorofilov v listih obeh rastlin ter ugotovili, da dodan Se ni vplival na
koncentracijo klorofilov pri rastlinah solate in radiĉa. V naši raziskavi smo uporabili bistveno
veĉjo koncentracijo Se(VI) (10 mg Se/L). Kalice, obravnavane s Se(VI), so imele statistiĉno
znaĉilno manjšo koncentracijo klorofila a (Slika 7), karotenoidov (Slika 9) od kalic,
obravnavanih s Se(IV), medtem ko so imele veĉjo koncentracijo antocianov od kalic,
obravnavanih s Se(IV) (Slika 10). Hawrylak-Nowak (2008) so rastline koruze (Zea mays var.
„saccharata Kcke‟) hidroponiĉno gojili v hranilni raztopini pri razliĉnih pH. K raztopini so
dodali Na2SeO4 v koncentracijah 0,4, 2, 4 in 8 mg Se/L. Ugotovili so, da je pri rastlinah, ki so
bile obravnavane s Se(VI) pri koncentracijah 2, 4 in 8 mg Se/L, prišlo do zmanjšanja
koncentracije klorofila. Zmanjšane koncentracije klorofila lahko pomenijo to, da rastlina raste
v stresnih razmerah. Koncentracije 0,4 – 4 mg Se(VI)/L niso vplivale na koncentracijo
antocianov pri koruzi, medtem ko je pri koncentraciji 8 mg Se(VI)/L prišlo do poveĉanja
koncentracije antocianov. Razliĉni stresni dejavniki v okolju, v katerem rastlina uspeva, lahko
zmanjšajo koncentracijo klorofila pri rastlinah in pospešijo izgradnjo antocianov. Pri našem
poskusu so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno veĉjo koncentracijo
antocianov od kontrolnih kalic (Slika 10). Hartikainen idr. (2000) so ugotovili, da Se(VI) pri
koncentraciji manjši od 10 mg Se/kg, ki so jo dodali v tla, ni vplivala na koncentracijo
klorofilov v rastlinah ljuljke (Lolium perenne). V tla so dodali tudi Se(VI) pri koncentraciji ≥
10 mg Se/kg in ugotovili, da je zmanjšal koncentracijo klorofilov v mladih listih ljuljke,
medtem ko se je koncentracijo klorofilov v starejših listih poveĉala. Xue idr. (2001) so izvedli
poskus na zeleni solati in ugotovili, da je obravnavanje rastlin zelene solate s Se(VI) v obliki
H2SeO4 in s koncentracijama 0,1 ter 1,0 mg Se/kg tal, poveĉalo koncentracijo klorofilov pri
mladih rastlinah zelene solate. Pri starejših rastlinah zelene solate pa je na poveĉanje
koncentracije klorofilov vplivala le veĉja koncentracija dodanega Se(VI).
Listno škropljenje redkvice (Raphanus sativus) z raztopino, v katero je bil dodan I(-I) in I(V),
v obliki KI oz. KIO3, z 0,2 % koncentracijo ĉistega elementa, ni vplivalo na koncentracijo
fotosinteznih barvil (Strzetelski, Smolén, Roţek in Sady, 2010). Blasco idr. (2011) so v svoji

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
32
raziskavi na vrtni solati prouĉevali vpliv razliĉnih koncentracij in oblik I na koncentracijo
fotosinteznih barvil. Semena vrtne solate so obravnavali z I(-I), ki je bil v hranilno raztopino
dodan v obliki KI, pri koncentracijah 2,5, 5, 10 mg I/L ter z enakimi koncentracijami I(V), ki
je bil dodan v obliki KIO3. Med kontrolnimi rastlinami in rastlinami, ki so bile obravnavane z
I(-I), ni bilo razlik v koncentraciji klorofila a, vendar se je koncentracija bistveno zmanjšala v
rastlinah, katerih semena so bila obravnavana z I(V) pri koncentraciji 10 mg I/L. V naši
raziskavi smo uporabili bistveno veĉjo koncentracijo I(-I) oz. I(V) (1000 mg I/L).
Obravnavanje z I(V) je pozitivno vplivalo na vsebnost koncentracije klorofila b, ki je bila
statistiĉno znaĉilno veĉja kot pri kontrolnih kalicah (Slika 8). Kalice, obravnavane z I(V), so
imele statistiĉno znaĉilno veĉjo koncentracijo klorofila a (Slika 7), klorofila b (Slika 8) in
karotenoidov (Slika 9) od kalic, obravnavanih z I(-I). Na koncentracijo UV-B in UV-A
absorbirajoĉih snovi je vplivalo obravnavanje z I(V), saj so imele te kalice statistiĉno znaĉilno
veĉjo koncentracijo UV-B (Slika 11) in UV-A (Slika 12) absorbirajoĉih snovi od kalic,
obravnavanih s Se(IV)+I(-I).
5.3 POTENCIALNA FOTOKEMIČNA UČINKOVITOST
FOTOSISTEMA II
Vrednost potencialne fotokemiĉne uĉinkovitosti nam pove, ali je rastlina zaradi razliĉnih
okoljskih dejavnikov pod stresom. Pri vitalnih rastlinah, ki uspevajo v ugodnih razmerah in
tako niso izpostavljene stresu, dosega razmerje Fv/Fm temotno adaptiranega lista optimalno
vrednost 0,8 – 0,83, kar je teoretiĉni maksimum pri kopenskih rastlinah (Schreiber, Bilger,
Hormann in Neubauer, 1998). Rezultati naše raziskave so pokazali, da so imele kalice,
obravnavane z I(V), statistiĉno znaĉilno manjšo potencialno fotokemiĉno uĉinkovitost od
kalic, obravnavanih s Se(VI)+I(-I) (Slika 13). Vrednosti potencialne fotokemiĉne
uĉinkovitosti so se pri obeh poskusih gibale med 0,7 in 0,8, kar je blizu teoretiĉnega
maksimuma in kaţe na to, da razliĉne oblike Se in I ter njune kombinacije niso povzroĉile
poškodb fotosinteznega aparata in da so kalice uspevale v ugodnih razmerah, zaradi ĉesar niso
bile v stresu (Tadina, Germ, Kreft, Breznik in Gaberšĉik, 2007). Podobne rezultate kot v
našem poskusu so dobili pri raziskavi na rastlinah navadne ajde (Breznik idr., 2005; Tadina
idr. 2007) in tatarske ajde (Breznik idr., 2005), ki so ju listno škropili s Se(VI) (1 mg Se/L).
Golob idr. (2016b) so listno škropili tatarsko (Fagopyrum tataricum) in hibridno ajdo
(Fagopyrum hybridum) z vodno raztopino Na2SeO4 (20 mg Se/L). Ugotovili so, da so imele
kontrolne rastline hibridne ajde bistveno veĉjo potencialno fotokemiĉno uĉinkovitost FS II
kot kontrolne rastline tatarske ajde. Enako velja za rastline, ki so bile obravnavane s Se.
Dodatek Se je statistiĉno znaĉilno poveĉal potencialno fotokemiĉno uĉinkovitost FS II pri
obeh vrstah ajde. Listno škropljenje rastlin radiĉa z vodno raztopino, ki je vsebovala 1 mg
Se/L v obliki Na2SeO4, prav tako ni imelo vpliva na potencialno fotokemiĉno uĉinkovitost FS
II (Germ, Stibilj, Osvald in Kreft, 2007c). Tudi listno škropljenje buĉ z 1,5 mg Se/L v obliki
Na2SeO4, ni vplivalo na potencialno fotokemiĉno uĉinkovitost FS II (Germ idr., 2005).
Pozitiven vpliv Se na potencialno fotokemiĉno uĉinkovitost FS II so opazili pri jagodah
(Fragaria x ananassa), gojenih v tleh, ki so bila obogatena s Se (0,1 mg Se/kg), vendar enako
obravnavanje ni imelo pozitivnega uĉinka pri jeĉmenu (Hordeum vulgare) (Valkama,
Kivimäenpää, Hartikainen in Wulff, 2003). Mechora, Stibilj in Germ (2015) so rastline vodne
leĉe (Lemna minor L.) gojili v nadzorovanih razmerah, ki so jih izpostavili razliĉnim
koncentracijam raztopine Na2SeO3 (0,5, 1, 2, 5 in 10 mg Se/L). Celoten poskus je trajal 7
tednov, vendar so fiziološke in biokemijske parametre merili vsak teden tekom celotnega
poskusa. Obravnavanje rastlin s Se(IV) pri koncentraciji 1 mg Se/L ni vplivalo na potencialno

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
33
fotokemiĉno uĉinkovitost skozi celoten poskus, medtem ko sta obravnavanja z 2 in 5 mg Se/L
negativno vplivala na potencialno fotokemiĉno uĉinkovitost FS II v 3. in 4. tednu. Rastline, ki
so bile izpostavljene najveĉji koncentraciji Se(IV), je bilo treba tekom poskusa dvakrat
zamenjati, ker so propadale. To se je odrazilo tudi na vrednosti potencialne fotokemiĉne
uĉinkovitosti, ki je bila majhna. Tudi v naši raziskavi so raztopine poleg dH2O vsebovale
posamezno dodan Se(IV), s koncentracijo 10 mg Se/L, ki pa pri nobenem obravnavanju ni
vplival na potencialno fotokemiĉno uĉinkovitost FS II kalic kolerabice.

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
34
6 SKLEPI
V magistrski nalogi nas je zanimal vpliv razliĉnih kemijskih oblik Se in I ter njunih
kombinacij na izbrane biokemijske in morfološke lastnosti kalic kolerabice. V ta namen smo
si zadali 2 raziskovalni vprašanji, na kateri smo poskušali odgovoriti s pomoĉjo statistiĉnih
analiz pridobljenih podatkov.
1. raziskovalno vprašanje: Kako bodo različne kemijske oblike selena in joda ter njune
kombinacije vplivale na rast kalic kolerabice?
Razliĉne kemijske oblike Se in I ter njune kombinacije niso statistiĉno znaĉilno vplivale na
število kalic, ki so vzklile iz semen, iz razliĉnih obravnavanj pri 1. in 2. poskusu. Procent
kalitve je bil pri obeh poskusih 67 - 75 %. Pri 1. poskusu so bile kalice, obravnavane s Se(VI),
statistiĉno znaĉilno višje od kalic, obravnavanih s Se(VI)+I(V). Kalice, obravnavane z I(-I) so
bile statistiĉno znaĉilno višje od kalic, obravnavanih s Se(VI)+I(V), medtem ko so bile kalice,
obravnavane z I(V), statistiĉno znaĉilno niţje od kalic, obravnavanih s Se(IV)+I(-I). Višina
kalic se je gibala med 0,70 in 0,80 cm pri obeh poskusih.
2. raziskovalno vprašanje: Kako bodo različne kemijske oblike selena in joda ter njune
kombinacije vplivale na izbrane biokemijske in morfološke lastnosti kalic kolerabice?
Razliĉna obravnavanja s Se in I ter njune kombinacije so razliĉno vplivala na sveţo in suho
maso kalic. Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo sveţo maso
od kalic, obravnavanih z I(-I) in veĉjo sveţo maso od kalic, obravnavanih s Se(VI)+I(V). Pri
2. poskusu so imele kontrolne kalice statistiĉno znaĉilno manjšo sveţo maso od kalic,
obravnavanih s Se(VI), Se(IV), Se(VI)+I(V), Se(IV)+I(-I) in Se(IV)+I(V). Obravnavanje s
Se(VI)+I(V) je torej negativno vplivalo na sveţo maso kalic pri 1. poskusu, medtem ko je pri
2. poskusu vplivalo pozitivno. Vsa ostala omenjena obravnavanja pri 1. in 2. poskusu so imela
pozitiven vpliv na sveţo maso kalic. Na suho maso je pri obeh poskusih pozitivno vplivalo
obravnavanje s Se(IV)+I(-I), saj so imele kalice iz tega obravnavanja statistiĉno znaĉilno
veĉjo suho maso od kontrolnih kalic. V primerjavi s kontrolnimi kalicami so, pri 1. poskusu,
pozitivno vplivala na suho maso kalic obravnavanja s Se(VI), I(-I) in Se(VI)+I(-I), pri 2.
poskusu pa obravnavanja s Se(IV), Se(IV)+I(V) in Se(VI)+I(V).
Razliĉne kemijske oblike Se in I ter njune kombinacije so razliĉno vplivale na koncentracijo
barvil pri kalicah kolerabice. Pri 1. poskusu so imele kontrolne kalice statistiĉno znaĉilno
manjšo koncentracijo klorofila b od kalic, obravnavanih z I(V), medtem ko so pri 2. poskusu
imele manjšo koncentracijo klorofila b od kalic, obravnavanih s Se(VI)+I(V). Obe
obravnavanji sta torej pozitivno vplivali na koncentracijo klorofila b v kalicah. Na
koncentracijo antocianov je, v primerjavi s kontrolnimi kalicami, pozitivno vplivalo
obravnavanje s Se(VI). Kalice, obravnavane s Se(VI), so imele, pri 1. poskusu, statistiĉno
znaĉilno manjšo koncentracijo klorofila a in karotenoidov od kalic, obravnavanih s Se(IV).
Obratno so imele kalice, obravnavane s Se(VI), statistiĉno znaĉilno veĉjo koncentracijo
antocianov od kalic, obravnavanih s Se(IV). Pri 2. poskusu so imele kalice, obravnavane z
I(V), statistiĉno znaĉilno veĉjo koncentracijo UV-B in UV-A absorbirajoĉih snovi od kalic,
obravnavanih s Se(IV)+I(-I).
Razliĉna obravnavanja s Se in I ter njune kombinacije so razliĉno vplivala na potencialno
fotokemiĉno uĉinkovitost FS II. Pri 2. poskusu so imele kalice, obravnavane s Se(VI)+I(-I),
statistiĉno znaĉilno veĉjo fotokemiĉno uĉinkovitost od kalic, obravnavanih z I(V).

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
35
7 LITERATURA
Akbulut, M. in Cakir, S. (2010). The effects of Se phytotoxicity on the antioxidant systems of
leaf tissues in barley (Hordeum vulgare L.) seedlings. Plant Physiology and Biochemistry,
48(2-3), 160-166. doi:10.1016/j.plaphy.2009.11.001
Arthur, J. R., Nicol, F. in Beckett, G. J. (1992). The role of selenium in thyroid hormone
metabolism and effects of selenium deficiency on thyroid hormone and iodine metabolism.
Biological Trace Element Research, 34(3), 321–325. doi:10.1007/BF02783686
Arvy, M. P. (1993). Selenate and selenite uptake remove selenium form soils. Calif. Agric.,
43, 19-20.
Barceloux, G. D. (1999). Selenium. Clinical Toxicology, 37(2): 145-172.
Blasco, B., Ríos, J. J., Cervilla, L. M., Sánchez-Rodríguez, E., Ruiz, J. M. in Romero, L.
(2008). Iodine biofortification and antioxidant capacity of lettuce: potential benefits for
cultivation and human health. Annals of Applied Biology, 152(3), 289–299.
doi:10.1111/j.1744-7348.2008.00217.x
Blasco, B., Ríos, J. J., Leyva, R., Melgarejo, R., Constán-Aguilar, C., Sánchez-Rodríguez, E.,
... Ruiz, M. J. (2011). Photosynthesis and metabolism of sugars from lettuce plants
(Lactuca sativa L. var. longifolia) subjected to biofortification with iodine. Plant Growth
Regulation, 65(1), 137-143. doi:10.1007/s10725-011-9583-0
Borst-Pauwels, G. W. F. H. (1961). Iodine as a micronutrient for plants. Plant and Soil, 14(4),
377-392. doi:10.1007/BF01666295
Brenĉiĉ, J. in Lazarini, F. (1995). Splošna in anorganska kemija. Ljubljana: DZS.
Breznik, B., Germ, M., Gaberšĉik, A. in Kreft, I. (2005). Combined effects of elevated UV-B
radiation and addition of selenium on common (Fagopyrum esculentum Moench) and
tartary (Fagopyrum tataricum (L.) Gaertn.) buckwheat. Photosynthetica, 43(4), 583-589.
doi:10.1007/s11099-005-0091-1
Brown, A. T. in Shrift, A. (1982). Selenium: toxicity and tolerance in higher plants.
Biological Reviews., 57(1), 59-84. doi:10.1111/j.1469-185X.1982.tb00364.x
Carvalho, K. M., Gallardo-Williams, M. T, Benson, R. F. in Martin, D. F. (2003). Effects of
Selenium Supplementation on Four Agricultural Crops. Journal of Agricultural and Food
Chemistry, 51(3), 704-709. doi:10.1021/jf0258555
Chameides, W. L. in Davis, D. D. (1980). Iodine, it's possible role in tropospheric
photochemistry. Journal of Geophysical Research, 85(NC12), 7383-7398.
doi:10.1029/JC085iC12p07383
Combs, G. F. (2001). Selenium in global food system. British Journal of Nutrition, 85(5),
517-547. doi:10.1079/BJN2000280

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
36
Ĉerne, M. (1998). Kapusnice. Ljubljana: Kmeĉki glas.
Dai, L. J., Zhu, G. Y., Huang, Z. Y., Zhang, M, in Song, L. J. (2006). Availability of iodide
and iodate to spinach (Spinacia oleracea L.) in relation to total iodine in soil solution.
Plant and Soil, 289(1), 301-308. doi:10.1007/s11104-006-9139-7
Dilworth, G. L. in Bandurski, R. S. (1977). Activation of selenate by adenosine 5'-
triphosphate sulphurylase from Saccharomyces cerevisiae. Biochemical Journal, 163(3),
521–529.
Ellis, R. D. in Salt, E. D. (2003). Plants, selenium and human health. Current Opinion in
Plant Biology, 6(3), 273-279. doi:10.1016/S1369-5266(03)00030-X
Farhana, A in Shaiq, A. G. (2010). Iodine, Iodine metabolism and Iodine deficiency disorders
revisited. Indian Journal of Endocrinology and Metabolism, 14(1), 13-17.
Fuge, R. (2005). Soils and iodine deficiency. V O. Selinus, B. Alloway, J. A. Centeno, R. B.
Finkleman, R. Fuge, U. Lindh in P. Smedley (ur.), Essentials of Medical Geology (str. 417-
433). London: Elsevier Academic Press.
Finley, W. J. (2007). Increased intakes of selenium-enriched foods may benefit human health.
Journal of the Science of Food and Agriculture, 87(9), 1620–1629. doi:10.1002/jsfa.2943
Fordyce, F. M. (2005). Selenium deficiency and toxicity in the environment. V O. Selinus, B.
Alloway, J. A. Centeno, R. B. Finkleman, R. Fuge, U. Lindh in P. Smedley (ur.),
Essentials of Medical Geology (str. 373-415). London: Elsevier Academic Press.
Foster, L. H. in Sumar, S. (1997). Selenium in health and disease, a review. Critical Reviews
in Food Science and Nutrition, 37(3), 211–228. doi:10.1080/10408399709527773
Germ, M., Kreft, I. in Osvald, J. (2005). Influence of UV-B exclusion and selenium treatment
on photochemical efficiency of photosystem II, yield and respiratory potential in pumpkins
(Cucurbita pepo L.). Plant physiology and biochemistry, 43(5), 445-448.
doi:10.1016/j.plaphy.2005.03.004
Germ, M., Kreft, I., Stibilj, V. in Urbanc-Berĉiĉ, O. (2007a). Combined effects of selenium
and drought on photosynthesis and mitochondrial respiration in potato. Plant Physiology
and Biochemistry, 45(2), 162-167. doi:10.1016/j.plaphy.2007.01.009
Germ, M., Stibilj, V. in Kreft, I. (2007b). Metabolic importance of selenium for plants. The
European Journal of Plant Science and Biotechnology, 1(1), 91-97.
Germ, M., Stibilj, V., Osvald, J. in Kreft, I. (2007c). Effect of selenium foliar application on
chicory (Cichorium intybus L.). Journal of Agricultural and Food Chemistry, 55(3), 795-
798. doi:10.1021/jf0629888
Germ, M. in Stibilj, V. (2007). Selenium and plants. Acta agriculturae Slovenica, 89(1), 65-
71. doi:10.2478/v10014-007-0008-8

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
37
Germ, M., Kacjan Maršić, N., Turk, J., Pirc, M., Golob, A., Jerše, A., … Stibilj, V. (2015).
The effect of different compounds of selenium and iodine on selected biochemical and
physiological characteristics in common buckwheat and pumpkin sprouts. Acta biologica
Slovenica, 58(1), 35-44.
Golob, A., Gadţo, D., Djikić, M., Gavrić, T., Kreft, I. in Germ, M. (2016a). Sulphur interferes
with selenium accumulation in Tartary buckwheat plants. Plant Physiology and
Biochemistry, 108, 32-36. doi:10.1016/j.plaphy.2016.07.001
Golob, A., Germ, M., Kreft, I., Zelnik, I., Kristan, U. in Stibilj, V. (2016b). Selenium uptake
and Se compounds in Se-treated buckwheat. Acta Botanica Croatica, 75(1), 17-24.
doi:10.1515/botcro-2016-0016
Guthrie, F. (1863). “On the iodide of iodammonium”. Journal of the Chemical Society, 16,
239–244.
Hajiboland, R. in Keivanfar, N. (2012). Selenium supplementation stimulates vegetative and
reproductive growth in canola (Brassica napus L.) plants. Acta agriculturae Slovenica,
99(1), 13–19. doi:10.2478/v10014-012-0002-7
Hartikainen, H., Xue, T. in Piironen, V. (2000). Selenium as an anti-oxidant and pro-oxidant
in ryegrass. Plant and Soil, 225(1), 193-200. doi:10.1023/A:1026512921026
Hawrylak-Nowak, B. (2008). Changes in anthocyanin content as indicator of maize sensitivity
to selenium. Journal of Plant Nutrition, 31(7), 1232-1242.
doi:10.1080/01904160802134962
Hurtevent, P., Thiry, Y., Levchuk, S., Yoschenko, V., Henner, P., Madoz-Escande C., ...
Kashparov, V. (2013). Translocation of 125I, 75Se and 36Cl to wheat edible parts
following wet foliar contamination under field conditions. Journal of Environmental
Radioactivity, 121, 43–54. doi:10.1016/j.jenvrad.2012.04.013
Hymer, C. B. in Caruso, J. A. (2006). Selenium speciation analysis using inductively coupled
plasma-mass spectrometry – review. Journal of Chromatography A, 1114(1), 1-20.
doi:10.1016/j.chroma.2006.02.063
Kabata-Pendias, A. in Pendias, H. (2001). Trace Elements in soils and plants. Boca Raton,
Florida: CRC Press.
Kalloo, G. in Bergh, B. O. (ur). (1993). Genetic improvement of vegetable crops. Oxford:
Pergamon press. Pridobljeno s
https://books.google.si/books?hl=sl&lr=&id=oh1S2zLjAGsC&oi=fnd&pg=PP1&dq=kohlr
abi+genetic+improvement+of+vegetable+crops&ots=oT5PBdKXb&sig=1y34F_iLp8H0w
xfYHVYWBwYPh1Y&redir_esc=y#v=onepage&q=kohlrabi%20genetic%20improvement
%20of%20vegetable%20crops&f=false
Kato, S., Wachi, T., Yoshihira, K., Nakagawa, T., Ishikawa, A., Takagi, D., ... Takahashi, M.
(2013). Rice (Oryza sativa L.) roots have iodate reduction activity in response to iodine.
Frontiers in Plant Science, 4(227), 1-11. doi: 10.3389/fpls.2013.00227

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
38
Kopsell, A. D. in Kopsell, E. D. (2007). Selenium. V A. V. Barker in D. J. Pilbeam (ur.),
Handbook of plant nutrition (str. 515-549). Boca Raton, Florida: CRC Press.
Kuznetsov, V. Vas., Kholodova, P. V., Kuznetsov, Vl. V. in Yagodin, A. B. (2003). Selenium
Regulates the Water Status of Plants Exposed to Drought. Doklady Biological Sciences,
390(1-6), 266-268. doi:10.1023/A:1024426104894
Landini, M., Gonzali, S. in Perata, P. (2011). Iodine biofortification in tomato. Journal of
Plant Nutrition and Soil Science, 174(3), 480–486. doi:10.1002/jpln.201000395
Läuchli, A. (1993). Selenium in plants – uptake, functions, and environmental toxicity.
Botanica Acta, 106(6), 455-468. doi:10.1111/j.1438-8677.1993.tb00774.x
Lehr, J. J., Wybenga, M. J. in Rosanow, M. (1958). Iodine as a micronutrient for tomatoes.
Plant Physiology, (33)6, 421-427.
Lešić, R., Borošić, J., Buturac, I., Ćustić, M., Poljak, M. in Romiĉ, D. (2004). Povrčarstvo.
Ĉakovec: Zrinski.
Letavayova, L., Vlĉkova, V. in Brozmanova, J. (2006). Selenium: From cancer prevention to
DNA damage. Review. Toxicology, 227(1-2), 1-14. doi:10.1016/j.tox.2006.07.017
Leyva, R., Sánchez-Rodríguez, E., Ríos, J. J., Rubio-Wilhelmi, M. M., Romero, L., Ruiz, M.
J. in Blasco, B. (2011). Beneficial effects of exogenous iodine in lettuce plants subjected to
salinity stress. Plant Science, 181(2), 195-202. doi:10.1016/j.plantsci.2011.05.007
Lim, T. K. (2015). Edible Medicinal and Non Medicinal Plants. Dordrecht, New York:
Springer. Pridobljeno s
https://books.google.si/books?id=t22vBQAAQBAJ&pg=PA768&dq=brassica+oleracea+l+
var+gongylodes+l&hl=sl&sa=X&ved=0ahUKEwicgZ3Y-
6fMAhXDApoKHeijCkcQ6AEINTAD#v=onepage&q=brassica%20oleracea%20l%20var
%20gongylodes%20l&f=false
Mackowiak, C. L., Grossl, P. R. in Cook, L. K. (2005). Iodine toxicity in a plant-solution
system with and without humic acid. Plant and Soil, 269(1), 141-150.
doi:10.1007/s11104-004-0401-6
Malorgio, F., Diaz, K. E., Ferrante, A., Mensuali-Sodi, A. in Pezzarossa, B. (2009). Effects of
selenium addition on minimally processed leafy vegetables grown in a floating system.
Journal of the Science of Food and Agriculture, 89(13), 2243-2251. doi:10.1002/jsfa.3714
Martinĉiĉ, A., Wraber, T., Jogan, N., Podobnik, A., Turk, B., Vreš, B., ... Surina, B. (2007).
Mala flora Slovenije. Ključ za določanje praprotnic in semenk. Ljubljana: Tehniška
zaloţba Slovenije.
Mechora, Š., Stibilj, V., Radešĉek, T., Gaberšĉik, A. in Germ, M. (2011). Impact of Se (VI)
fertilization on Se concentration in different parts of red cabbage plants. Journal of Food,
Agriculture and Environment, 9(2), 357-361.

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
39
Mechora, Š. in Vrhovšek, M. (2012). Response of two submersed macrophytes
Ceratophyllum demersum and Myriophyllum spicatum to selenium in water. Acta
Biologica Slovenica, 55(1), 9-14.
Mechora, Š., Stibilj, V. in Germ, M. (2015). Response of duckweed to various concentrations
of selenite. Environmental Science and Pollution Research, 22(4), 2416-2422.
doi:10.1007/s11356-014-3270-4
Navarro-Alarcon, M. in Cabrera-Vique, C. (2008). Selenium in food and the human body: A
review. Science of The Total Environment, 400(1-3), 115-141.
doi:10.1016/j.scitotenv.2008.06.024
Pedrero, Z., Madrid-Albarrán, Y., Hartikainen, H. in Cámara, C. (2008). Protective effect of
selenium in Broccoli (Brassica olearacea) plants subjected to cadmium exposure. Journal
of Agriculture and Food, 56(1), 266-271. doi:10.1021/jf072266w
Pedrero, Z. in Madrid, Y. (2009). Novel approaches for selenium speciation in foodstuffs and
biological specimens: A review. Analytica Chimica Acta, 634(2), 135–152.
doi:10.1016/j.aca.2008.12.026
Pennanen, A., Xue, T. in Hartikainen, H. (2002). Protective role of selenium in plant
subjected to severe UV irradiation stress. Journal of Applied Botany, 76(1-2), 66-76.
Peters, U., Foster, C. B., Chatterjee, N., Schatzkin, A., Reding, D., Andriole, G. L., ... Hayes,
R. B. (2007). Serum selenium and risk of prostate cancer – a nested case-control study.
American Journal of Clinical Nutrition, 85(1), 209–217.
Pyrzyńska, K. (2001). Analysis of selenium species by capillary electrophoresis. Talanta,
55(4), 657–667. doi:10.1016/S0039-9140(01)00511-2
Rasmussen, R. A, Khalil, M. A. K, Gunawardena, R. in Hoyt, S. D. (1982). Atmospheric
methyl iodide (CH3I). Journal of Geophysical Research, 87(C4), 3086 - 3090.
doi:10.1029/JC087iC04p03086
Reilly, C. (1998). Selenium: A new entrant into the functional food arena. Trends in Food,
Science and Technology, 9(3), 114-118. doi:10.1016/S0924-2244(98)00027-2
Reilly, C. (2006). Selenium in Food and Health. New York: Springer.
Ryser, A. L., Strawn, D. G., Marcus, M. A., Fakra, S., Johnson-Maynard, J. L. in Moller, G.
(2006). Microscopically focused synchrotron X-ray investigation of selenium speciation in
soils developing on reclaimed mine lands. Environmental Science and Technology, 40(2),
462-467. doi:10.1021/es051674i
Schrauzer, G. N. (2001). Nutritional selenium supplements: product types, quality, and safety.
Journal of the American College of Nutrition, 20(1), 1–4.
doi:10.1080/07315724.2001.10719007

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
40
Schreiber, U., Bilger, W. in Neubauer, C. (1994). Chlorophyll fluorescence as a nonintrusive
Indicator for rapid sssessment of in vivo photosynthesis. V E. D. Schulze in M. M.
Caldwell (ur.), Ecophysiology of Photosynthesis (str. 49-70). Dordrecht, New York:
Springer.
Schreiber, U., Bilger, W., Hormann, H. in Neubauer, C. (1998). Chlorophyll fluorescence as a
diagnostic tool: basics and some aspects of practical relevance. V A. S. Rghavendra (ur.),
Photosynthesis, a Comprehensive Treatise (str. 320-336). Cambridge: University Press
Cambridge.
Schwartz, K. in Foltz, C. M. (1957). Selenium as an integral part of factor 3 against dietary
necrotic liver degeneration. Journal of the American Chemical Society, 79(12), 3292-3293.
doi:10.1021/ja01569a087
Shamberger, J. R. (1981). Selenium in the environment. Science of The Total Environment,
17(1), 59-74. doi:10.1016/0048-9697(81)90108-X
Smoleń, S., Roţek, S., Ledwożyw-Smoleń, I. in Strzetelski, P. (2011). Preliminary evaluation
of the influence of soil fertilization and foliar nutrition with iodine on the efficiency of
iodine biofortification and chemical composition of lettuce. Journal of Elementology,
16(4), 613-622. doi:10.5601/jelem.2011.16.4.10
Smoleń, S., Kowalska, I. in Sady, W. (2014). Assessment of biofortification with iodine and
selenium of lettuce cultivated in the NFT hydroponic system. Scientia Horticulturae, 166,
9-16. doi:10.1016/j.scienta.2013.11.011
Smoleń, S., Skoczylas, L., Rakoczy, R., Ledwożyw-Smoleń, I., Kopeć, A., Piątkowska, E., ...
Sady, W. (2015). Mineral composition of field-grown lettuce (Lactuca sativa L.)
depending on the diversified fertilization with iodine and selenium compounds. Acta
Scientiarum Polonorum Hortorum Cultus, 14(6), 97-114.
Smrkolj, P., Pograjc, L., Hlastan-Ribiĉ, C. in Stibilj, V. (2005). Selenium content in selected
Slovenian foodstuffs and estimated daily intakes of selenium. Food Chemistry, 90, 691-
697. doi:10.1016/j.foodchem.2004.04.028
Smrkolj, P., Germ, M., Kreft, I. in Stibilj, V. (2006). Respiratory potential and Se compounds
in pea (Pisum sativum L.) plants grown from Se-enriched seeds. Journal of Experimental
Botany, 57(14), 3595-3600. doi:10.1093/jxb/erl109
Strzetelski, P., Smoleń, S., Rożek, S. in Sady, W. (2010). The effect of diverse iodine
fertilization on nitrate accumulation and content of selected compounds in radish plants
(Raphanus sativus L.). Acta Scientiarum Polonorum, Hortorum Cultus, 9(2), 65-73.
Štimec, M., Miš, F. N., Smole, K., Širca-Ĉampa, Kotnik, P., Zupanĉiĉ, ... Kriţišnik, C.
(2007). Iodine Intake of Slovenian Adolescents. Annals of Nutrition & Metabolism, 51(5),
439-447. doi:10.1159/000111164

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
41
Tadina, N., Germ, M., Kreft, I., Breznik, B. in Gaberšĉik, A. (2007). Effects of water deficit
and selenium on common buckwheat (Fagopyrum esculentum Moench.) plants.
Photosynthetica, 45(3), 472-476. doi:10.1007/s11099-007-0080-7
Tausz, M., Bytnerowicz, A., Weidner, W., Arbaugh, M. J., Padgett, P. in Grill, D. (1999).
Pigments and photoprotection in needles of Pinus ponderosa trees with and without
symptoms of ozone injury. Phyton, 39(4), 219-224.
Terry, N., Zayed, M. A., Souza de, P. M. in Tarun, S. A. (2000). Selenium in higher plants.
Annual Review of Plant Physiology and Plant Molecular Biology, 51, 401-432.
doi:10.1146/annurev.arplant.51.1.401
Thangavel, P., Sulthana, A. S. in Subburam, V. (1999). Interactive effects of selenium and
mercury on the restoration potential of leaves of the medicinal plant, Portulaca oleracea
Linn. Science of The Total Environment, 243-244, 1-8. doi:10.1016/S0048-
9697(98)00181-8
Tinggi, U. (2003). Essentiality and toxicity of selenium and its status in Australia: a review.
Toxicology letters, 137(1-2), 103-110. doi:10.1016/S0378-4274(02)00384-3
Tsunogai, S. in Sase, T. (1969). Formation of iodide-iodine in me ocean. Deep-Sea Research
and Oceanographic Abstracts. 16(5), 489-486. doi:10.1016/0011-7471(69)90037-0
Turakainen, M., Hartikainen, H. in Seppänen, M. M. (2004). Effects of selenium treatments
on potato (Solanum tuberosum) growth and concentrations of soluble sugars and starch.
Journal of Agricultural and Food Chemistry, 52(17), 5378-5382. doi:10.1021/jf040077x
Uden, P. C., Boakye, H. T., Kahakachchi, C. in Tyson, J. F. (2004). Selective detection and
identification of Se containing compounds – review and recent developments. Journal of
Chromatography A, 1050(1), 85-93. doi:10.1016/j.chroma.2004.05.077
Uden, P. C. (2005). Speciation of Selenium. V R. Cornelis, J. Caruso, H. Crews in K.
Heumann (ur.), Handbook of Elemental Speciation II - Species in the Environment, Food,
Medicine and Occupational Health (str. 346-365). Chichester: John Wiley & Sons Ltd.
Umaly, R. C. in Poel, L. W. (1971). Effects of iodine in various formulations on the growth of
barley and pea plants in nutrient solution culture. Annals of Botany, 35, 127–131.
Valkama, E., Kivimäenpää, H., Hartikainen, H. in Wulff, A. (2003). The combined effects of
enhanced UV-B radiation and selenium on growth, chlorophyll fluorescence and
ultrastructure in strawberry (Fragaria x ananassa) and barley (Hordeum vulgare) treated
in the field. Agricultural and Forest Meteorology, 120, 267–278.
Voogt, W., Holwerda, H. T. in Khodabaks, R. (2010). Biofortification of lettuce (Lactuca
sativa L.) with iodine: the effect of iodine form and concentration in the nutrient solution
on growth, development and iodine uptake of lettuce grown in water culture. Journal of the
Science of Food and Agriculture, 90(5), 906–913. doi:10.1002/jsfa.3902

Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
42
Welch, R. M in Graham, R. D. (1999). A new paradigm for world agriculture: meeting human
needs – productive, sustainable, nutritious. Field Crops Research, 60, 1–10.
Weng, H., Hong, C., Xia, T., Bao, L., Liu, H. in Li, D. (2013). Iodine biofortification of
vegetable plants – an innovative method for iodine supplementation. Chinese Science
Bulletin, 58(17), 2066-2072. doi:10.1007/s11434-013-5709-2
White, P. J., Bowen, H. C., Parmaguru, P., Fritz, M., Spracklen, W. P., Spiby, R. E., ...
Broadley, M. R. (2004). Interactions between selenium and sulphur nutrition in
Arabidopsis thaliana. Journal of Experimental Botany, 55(404), 1927–1937.
doi:10.1093/jxb/erh192
White, P. J. in Broadley, M. R. (2009). Biofortification of crops with seven mineral elements
often lacking in human diets – iron, zinc, copper, calcium, magnesium, selenium and
iodine. New Phytologist, 182(1), 49–84. doi: 10.1111/j.1469-8137.2008.02738.x.
Whitehead, D. C. (1973): Uptake and distribution of iodine in grass and clover plants grown
in solution culture. Journal of the Science of Food and Agriculture, 24(1), 43–50.
Whitehead, D. C. (1984). The distribution and transformations of iodine in the environment.
Environment International, 10(4), 321-339. doi:10.1016/0160-4120(84)90139-9
Xue, T., Hartikainen, H. in Piironen, V. (2001). Antioxidative and growth-promoting effect of
selenium on senescing lettuce. Plant and Soil, 237, 55-61. doi:10.1023/A:1013369804867
Zhu, Y. G., Huang, Y. Z., Hu, Y. in Liu, Y. X. (2003). Iodine uptake by spinach (Spinacia
oleracea L.) plants grown in solution culture: effects of iodine species and solution
concentrations. Environment International, 29(1), 33-37. doi:10.1016/S0160-
4120(02)00129-0
Zhu, Y. G., Huang, Y., Hu, Y., Liu, Y. in Christie, P. (2004). Interactions between selenium
and iodine uptake by spinach (Spinacia oleracea L.) in solution culture. Plant and Soil,
261(1), 99-105. doi:10.1023/B:PLSO.0000035539.58054.e1
Zia, H. M., Watts, J. M., Gardner, A. in Chenery, R. S. (2014). Iodine status of soils, grain
crops, and irrigation waters in Pakistan. Environmental Earth Sciences, 73(12), 7995-8008.
doi:10.1007/s12665-014-3952-8
Zimmermann, M. B. (2009). Iodine deficiency. Endocrine Reviews, 30(4), 376-408.
doi:https://doi.org/10.1210/er.2009-0011
Ţnidarĉiĉ, D. (2011). Selen in njegove zvrsti v okolju. Acta agriculturae Slovenica, 97(1), 73-
83.
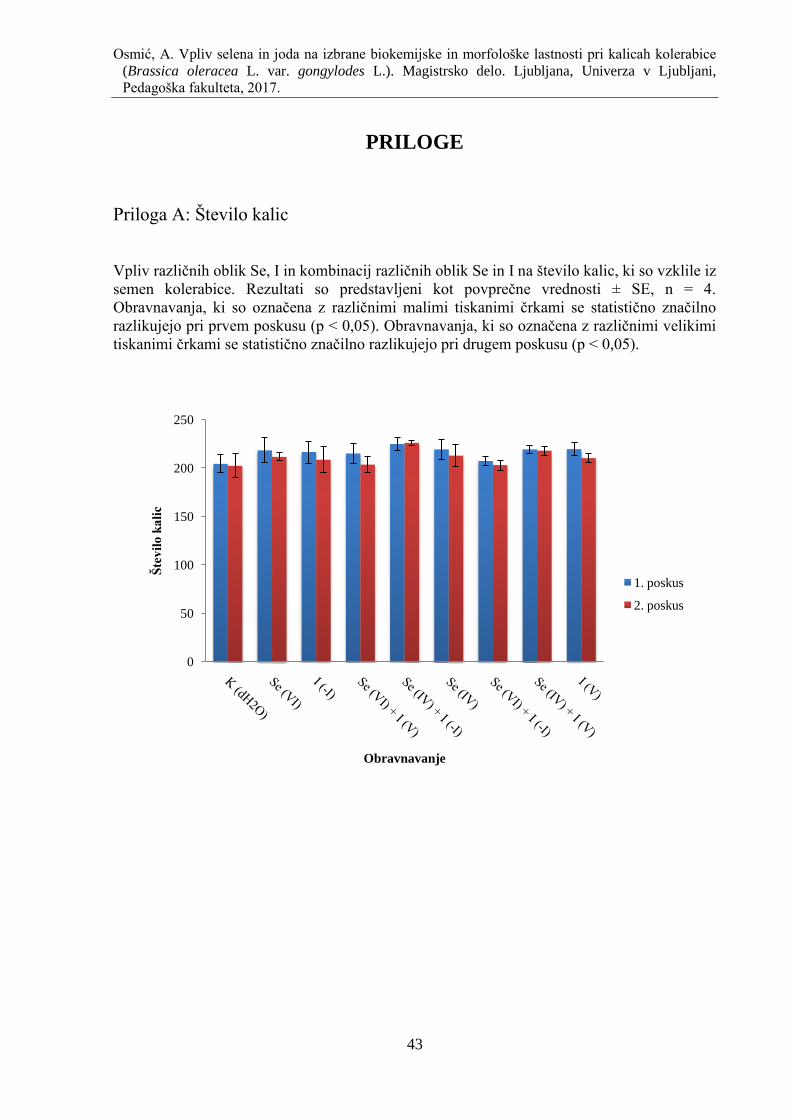
Osmić, A. Vpliv selena in joda na izbrane biokemijske in morfološke lastnosti pri kalicah kolerabice
(Brassica oleracea L. var. gongylodes L.). Magistrsko delo. Ljubljana, Univerza v Ljubljani,
Pedagoška fakulteta, 2017.
43
PRILOGE
Priloga A: Število kalic
Vpliv razliĉnih oblik Se, I in kombinacij razliĉnih oblik Se in I na število kalic, ki so vzklile iz
semen kolerabice. Rezultati so predstavljeni kot povpreĉne vrednosti ± SE, n = 4.
Obravnavanja, ki so oznaĉena z razliĉnimi malimi tiskanimi ĉrkami se statistiĉno znaĉilno
razlikujejo pri prvem poskusu (p < 0,05). Obravnavanja, ki so oznaĉena z razliĉnimi velikimi
tiskanimi ĉrkami se statistiĉno znaĉilno razlikujejo pri drugem poskusu (p < 0,05).
0
50
100
150
200
250
Šte
vil
o k
ali
c
Obravnavanje
1. poskus
2. poskus





























