-
8/9/2019 Hemoglobin Module 6
1/45
Hemoglobin
-
8/9/2019 Hemoglobin Module 6
2/45
Hemoglobin
Human hemoglobin A, present in adults, consists of four
subunits:
two -subunits and two -subunits.
The - and -subunits are homologous and have similar
three-dimensional structures.
The capacity of hemoglobin to bind oxygen depends on
the presence of a bound prosthetic group (heme).
The heme group is responsible for the distinctive red
color of blood.
-
8/9/2019 Hemoglobin Module 6
3/45
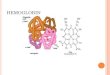
The heme group consists of a protoporphyrin,
and a central iron atom.
Protoporphyrin is made up of four pyrrole rings
linked by methene bridges to form a tetrapyrrole
ring.
Four methyl groups, two vinyl groups, and two
propionate side chains are attached.
-
8/9/2019 Hemoglobin Module 6
4/45
-
8/9/2019 Hemoglobin Module 6
5/45
-
8/9/2019 Hemoglobin Module 6
6/45
-
8/9/2019 Hemoglobin Module 6
7/45
GLOBIN(protein part or apoprotein):
It is a simple protein (histone) which is characterized by
its high content of histidine and lysine.
It is composed of four polypeptide chains 2 and 2
chains.
The -chain contains 141 amino acids and -chain
contains 146 amino acids.
Each -polypeptide chain is folded into 8 right handed -
helices termed A-H starting from NH2-terminal,
-subunit is folded into 7-helices.
The ratio of haem to globin is 4:1. So each haem moietyis linked
to one peptide chain.
-
8/9/2019 Hemoglobin Module 6
8/45
The myoglobin-hemoglobin family of
proteins has produced a way in which Fe++
can be bounded to the proteins so as to
produce an O2 binding site.
-
8/9/2019 Hemoglobin Module 6
9/45
deoxyHbdeoxyMb
Myoglobin and Hemoglobin Structure
F
F
E
E
oxyMb (MbO2)
O2
-
8/9/2019 Hemoglobin Module 6
10/45
The iron atom lies in the center of the protoporphyrin,
bonded to the four pyrrole nitrogen atoms.
Under normal conditions, the iron is in the ferrous (Fe2+)
oxidation state.
The iron ion can form two additional bonds, one on each
side of the heme plane. These binding sites are called
the fifth and sixth coordination sites.
In hemoglobin, the fifth coordination site is occupied by
the imidazole ring of a histidine residue from the protein.
In deoxyhemoglobin, the sixth coordination site remains
unoccupied.
-
8/9/2019 Hemoglobin Module 6
11/45
-
8/9/2019 Hemoglobin Module 6
12/45
Types of normal haemoglobin
I. Adult haemoglobin: There are 2 types HbA1 and HbA2.
a) Majoradult haemoglobin: Hb A1 (2 2)
- Contains 2 alpha chains and 2 beta chains.
This haemoglobin A1 constitutes 95-97% of the
totalhaemoglobin.
b) Minoradult haemoglobin: Hb A2 (2 2)
- HbA2 forms about 2-4% of total haemoglobin.
- Contains 2 -chains and 2 -chains.
- In the -chains there is more than one aminoacid
different than those in -chain e.g. arginine residue at
the position 16 instead of glycine which is normally
present in beta chain
-
8/9/2019 Hemoglobin Module 6
13/45
II. Glycosylated haemoglobin (Hb A1c):
- It is modified form of haemoglobin similar to
haemoglobin A1 but it contains glucose linked to aminogroup
present on lysyl residues and at the NH2 - terminal
ends. The reaction is non enzymatic and its rate
depends on the concentration of glucose .It is present in
normal value 5% of the total haemoglobin.
- This percentage is increased in prediabetic and diabetic
patients up to 8-14%. This glycohaemoglobin gives an
idea about the blood glucose level during the last three
months and is useful in the assessment of diabetic
control
-
8/9/2019 Hemoglobin Module 6
14/45
III. Fetal haemoglobin = HbF (2 2):
It is present normally in newborn and early fetal life and
at age of 7 months 90 % of fetal haemoglobin isreplaced by adult
haemoglobin ( HbA1)
- It consists of 2 alpha chains and 2 gamma chains.
- In gamma chain there is more than one aminoacid
different from those in -chain e.g. His21 residue isser21
- HbF has a great affinity for O2 under physiological
conditions, because -chains do not bind 2,3 BPG well.
BPG is responsible for lowering the O2 affinity of Hb
andallowing Hb to release O2 at the typical PO2 of tissues.
-
8/9/2019 Hemoglobin Module 6
15/45
Hemoglobin Genes and Gene Products
-
8/9/2019 Hemoglobin Module 6
16/45
Haem biosynthesis
Haem is the iron protoporphyrin, synthesized mostly in
the bone marrow( 85%) for incorporation into
haemoglobin and in liverfor synthesis ofcytochromes.
The initial and last three enzymatic steps are catalyzedby
enzymes that are present in mitochondria whereas
the intermediate steps are taking place in cytoplasm.
-
8/9/2019 Hemoglobin Module 6
17/45
Heme synthesis begins with condensation of glycine &
succinyl-CoA, with decarboxylation, to form H-
aminolevulinic acid (ALA). Pyridoxal phosphate (PLP)
serves as coenzyme forH-Aminolevulinate Synthase (ALA
Synthase
ALA synthase
OOC CH2 CH2 C S-CoA
O
+ OOC CH2 NH3+
OOC CH2 CH2 C
O
CH2 NH3+
CO2CoA-SH
H+succinyl-CoA glycine
H-aminolevulinate (ALA)
H-AminolevulinicAcid Synthase
1
1
-
8/9/2019 Hemoglobin Module 6
18/45
-
8/9/2019 Hemoglobin Module 6
19/45
-
8/9/2019 Hemoglobin Module 6
20/45
-
8/9/2019 Hemoglobin Module 6
21/45
-
8/9/2019 Hemoglobin Module 6
22/45
-
8/9/2019 Hemoglobin Module 6
23/45
-
8/9/2019 Hemoglobin Module 6
24/45
-
8/9/2019 Hemoglobin Module 6
25/45
2 ALA dehydratase
3
4
5
6
Uroporphyrinogen-I
Synthase + III cosynthase
Uroporphyrinogen-IIIdecarboxylase
Coproporphyrinogen-III
oxidase
Protooporphyrinogen
-III oxidase
-
8/9/2019 Hemoglobin Module 6
26/45
Heme synthase7
-
8/9/2019 Hemoglobin Module 6
27/45
Summary of heme synthesis
-
8/9/2019 Hemoglobin Module 6
28/45
Regulation of Haem Biosynthesis
Regulation of transcription or post-translationalprocessing
ofenzymes of the heme synthesispathways differs between erythrocyte
forming cells &other tissues.
In erythrocyte-forming cells there is steadyproduction of
pathway enzymes, limited only by ironavailability.
In other tissues expression of pathway enzymes is
more variable & subject to feedback inhibition byheme.
-
8/9/2019 Hemoglobin Module 6
29/45
Allosteric Regulation
ALA synthase enzyme controls the rate-limiting stepof Haem
synthesis.
Haem and also haematin act as a repressor of thesynthesis of ALA
synthase and act as feed backinhibitor at this step.
The block in haem biosynthesis in pantothenic acidor vitamin B6
deficiency occurs at very early step inhaem synthesis (ALA
synthase).
ALA dehydratase is sulfhydryl enzyme and is very
sensitive to inhibition by heavy metals as mercuryor lead.
-
8/9/2019 Hemoglobin Module 6
30/45
Porphyrias
Porphyrias are genetic diseases in which activity of one
of the enzymes involved in heme synthesis is decreased
(e.g., PBG Synthase, Porphobilinogen Deaminase, etc).
Symptoms vary depending on the enzyme
the severity of the deficiency
whether heme synthesis is affected primarily in liveror in
developing erythrocytes
-
8/9/2019 Hemoglobin Module 6
31/45
Sickle Cell Disease
Hemoglobin S (HbS) 50% Hb present.
Homozygotic HbSS (sickle cell anemia) - HbS = 100% Hb
present,Giving Sickle cell disease
HbSA disease - Double heterozygote for HbS and HbA, with
intermediate clinical severity. It is called Sickle cell
trait
-
8/9/2019 Hemoglobin Module 6
32/45
Basic abnormality - glutamic acid is replaced by
valine at the sixth position of the F-globin chain.
2 normal E-globin and 2 abnormal F-globin
chains forms HbS.
HbS carries O2 normally but begins to form
semisolid aggregate structures once O2 is
unloaded to the tissues. These HbS aggregates
distort RBCs and cause them to lose their
normal elasticity.
Molecularand cellularchanges of
hemoglobin S
-
8/9/2019 Hemoglobin Module 6
33/45
-
8/9/2019 Hemoglobin Module 6
34/45
Thalassaemias:
The name is derived from the Greek word
thalassa,which means sea.Greek identified this
disease present around Mediterranean sea.
They are hereditary hemolytic diseases in whichthe synthesis of
either - or - globin chain is
defective.
This decreased rate of synthesis of the globin
chains is due to mutation affecting the regulatory
gene rather than the structural gene.
-
8/9/2019 Hemoglobin Module 6
35/45
A) - thalassaemias: there are decreased or
absent synthesis of -chains of haemoglobin
with compensatory increase in the synthesis ofother chains.
a.Homozygous -chain thalassaemia
(thalassaemia major):
Incompatible with life, and present as hydrops
foetalis usually die in utero , due to complete
absence of -chains which are required for
synthesis of HbF.
b.Heterozygous -chain thalassaemia
thalassaemia minor,(trait):
-
8/9/2019 Hemoglobin Module 6
36/45
B) - thalassaemias: Synthesis of of -chains is
decreased or absent whereas synthesis of -chains
is normal and will combine with -chains givingexcess of HbA2
(22) or it may combine with -
chains producing excess of HbF (22).
The abnormal haemoglobin do not function as normal
haemoglobin
- Homozygous (Thalassaemia-major = Cooley's
anaemia = Mediterranian sea anaemia):
There is complete absence of -globin chain andthere is marked
increase of HbF.
- Heterozygous thalassaemia (Thalassaemia-
minor):
There is slow rate of synthesis of -globin chain.
-
8/9/2019 Hemoglobin Module 6
37/45
Heterozygous thalassaemia (Thalassaemia-
minor):
There is slow rate of synthesis of -globin chain.
-
8/9/2019 Hemoglobin Module 6
38/45
Methemoglobinemias:
This is oxidized Hb,
The Fe2+ normally present in heme being
replaced by Fe3+,the ability to react as an O2
carrier is lost. The normal erythrocyte contains small amount
of
met Hb, formed by spontaneous oxidation of Hb.
Met Hb is normally reconverted to Hb by
reducing systems in the RBC, the mostimportant of which is
NADH-methemoglobin
reductase.
-
8/9/2019 Hemoglobin Module 6
39/45
Congenital methemoglobinemias
A. Hemoglobin M (Hb-M):
It is a congenital condition due to mutation inglobin
biosynthesis in which distal or proximal
histidine is replaced by tyrosine.
B. Deficiency of NADH cytochrome b5methemoglobin reductase
system
Acquired (toxic) methemoglobinemea
Usually arises following the ingestion of large
amounts of drugs e.g. phenacetin or thesulphonamides, excess
ofnitrites or certain
oxidizing agents present in the diet.
-
8/9/2019 Hemoglobin Module 6
40/45
Glucose Metabolism in RBCs
1- Glycolysis2- Hexose mono-phosphate shunt
(HMPS)
-
8/9/2019 Hemoglobin Module 6
41/45
Glycolysis in RBCs
Glucose
2 NAD
2 NADH+H
2 ATP
2 Lactate
Glucose
1,3bisphosphoglycerate
2,3bisphosphoglycerate
-
8/9/2019 Hemoglobin Module 6
42/45
Importance of glycolysis in red cells:
a) Energy production: it is the only pathway thatsupplies the
red cells with ATP.
haemolytic anaemia may occur due to an inherited
deficiency of glycolytic enzymes mainly
pyruvate kinase deficiency.b) Reduction of methaemoglobin:
glycolysis
provides NADH for reduction of met Hb by
NADH-Cyto.b5 reductase
c) In red cells 1,3 bisphosphoglycerate is
converted to 2,3 bisphosphoglycerate which
binds to oxy Hb and helps release of O2 to
tissues.
-
8/9/2019 Hemoglobin Module 6
43/45
Importance of HMPS in Red cells:
Red cells are liable for oxidative damage byH2O2 due to their
role in O2 transport.
In RBCs, H2O2 can cause both oxidation of iron
in haemoglobin (to form methaemoglobin) andlipid peroxidation
(increases the cell membrane
fragility).
The major role of HMS in red cells is the
production of NADPH, which protect these cellsfrom oxidative
damage by reduction of
glutathionethat helps removal of H2O2.
-
8/9/2019 Hemoglobin Module 6
44/45
Non- Oxidative PhaseOxidative Phase
Reversible
Non-Regulatory
Irreversible
Regulatory
6 moles ofPentose-P
5 moles of G-6-P
6 moles of G-6-P
6 moles ofPentose-P
12 NADP
12 NADPH +12 H
6 H2O
6 CO2
-
8/9/2019 Hemoglobin Module 6
45/45
Role of NADPH+H in reduction of glutathione
G-S-S-G 2-GSH
2 GSH
+H2O2
G-S-S-G
+ 2 H2O
Glutathione
Reductase
Glutathioneperoxidase
NADPH+H NADP