
GENC SULCEBE ARBEN HOXHA
I M U N O L O G J I A
TEKST BAZË PËR STUDENTËT E FAKULTETIT TË MJEKSISË
TIRANE, 2001

1
TABELA E LËNDËS KAPITULLI 1. Hyrje në Imunologji. Njohuri mbi imunitetin e lindur (jo specifik per antigjenin) dhe atë të fituar (adoptiv, specifik per antigjenin). Historiku. Imuniteti i lindur dhe i fituar. Imuniteti i lindur: Faktorët që marrin pjesë. Mekanizmat qelizore (me vrasje intraqelizore): Qelizat fagocitare; Makrofagët dhe neutrofilet. Mekanizmat humorale: Rruga alterne e komplementit; Proteinat e fazës akute; Interferonet. Mekanizmat me vrasje jashtëqelizore: Qelizat vrasëse - natyrale (NK); Eozinofilet. KAPITULLI 2. Sistemi imunitar, organizimi i tij: Organet e imunitetit, ndarja dhe përshkrimi i tyre. Organet parësore ose qëndrore. Timusi, përshkrimi, funksionet. Maturimi në timus i limfociteve T. Palca kockore, roli i saj si organ limfoid paresor. Organet dytësore (periferike): Nyjet limfatike; Shpretka; Indi limfoid i lidhur me mukozat (MALT). KAPITULLI 3. Njohuri mbi antigenët. C'jane antigenët dhe haptenet, antigeniciteti, imunogeniciteti. Natyra e antigenëve, llojet e tyre, cil ësite fiziko-kimike të tyre, adjuvantët. Kushtet e antigenicitetit. Përcaktuesit antigenikë (epitopet), natyra dhe llojet e tyre. KAPITULLI 4. Karakteri klonal i përgjigjes imune. Receptorët për antigenin në limfocitet T dhe B. Bazat molekulare të shumëllojshmërise së tyre. Teoria e seleksionit klonal, TCR, BCR, karakteristikat e tyre. KAPITULLI 5. Popullatat limfocitare, shënjuesit dhe funksionet e tyre. Qelizat që marrin pjesë në funksionimin e sistemit imunitar. Nënpopullatat e limfociteve T, B, as T as B, qelizat dentritike, funksionet dhe shënjuesit kryesorë të tyre. Rruga e antigenit kur hyn në organizëm: Në indet (nyjet limfatike), në gjak (shpretka), në mukozat (MALT). Riqarkullimi i limfociteve në organizëm, fenomeni i Homing. KAPITULLI 6. Aktivizimi i limfociteve T dhe B, mekanizmat molekulare. KAPITULLI 7. Antitrupat, struktura dhe funksioni i tyre. C'janë imunoglobulinat, lidhja midis ndërtimit molekular dhe funksionit biologjik të tyre. Ndarja në klasa dhe nënklasa, vetitë biologjike të secilës prej tyre. KAPITULLI 8. Heterogjeneiteti i imunoglobulinave. Bazat molekulare të shumëllojshmerisë së antitrupave. C'janë izotipia, alotipia dhe idiotipia. Antitrupat monoklonalë dhe hibridomat. KAPITULLI 9. Sistemi komplementar, përberësit dhe funksioni. Përbërësit e rrugës klasike dhe asaj alterne të sistemit komplementar, shkaktarët e aktivizimit të tyre, dinamika në formë kaskade e këtij aktivizimi. Pasojat biologjike të aktivizimit të komplementit. Ndryshimet fiziologjike dhe patologjike të tij. KAPITULLI 10. Kompleksi madhor i pajtueshmërise indore (HLA). Genet dhe molekulat e kompleksit HLA, struktura, shprehja, dhe funksionet e tyre në përgjigjen imune. Lidhja e tyre me përgjigjen imune dhe sëmundjet. KAPITULLI 11. Mekanizmat efektorë të imunitetit qelizor. C'janë interleukinat. Ndarja e limfociteve në Th1 dhe Th2. Mekanizmat e veprimit të imunitetit me limfocitet T helper dhe T citotoksikë. KAPITULLI 12. Mekanizmat efektorë të imunitetit humoral. Veprimi i citokinave në prodhimin e antitrupave. Ecuria e përgjigjes antitrupore parësore dhe dytësore. Faktorët që veprojnë dhe vecoritë e secilës prej tyre. Efekti i dozës së antigenit në afinitetin e antitrupave; si realizohet maturimi i afinitetit. Mekanizmat qelizore dhe molekulare të sekretimit të imunoglobulinave serike. Ecuria e imunoglobulinave në lidhje me moshën, seksin dhe situata të tjera. KAPITULLI 13. Njohuri mbi mekanizmat e kontrollit në përgjigjen imune.

2
Roli i antigenit, antitrupave, limfociteve T ndihmës. C'jane limfocitet rregullatore, supresorë llojet e tyre, mekanizmat e veprimit. Hallkat e tjera të imunosupresionit (rrjeta idiotipike, makrofagët etj.). Influenca e faktorëve gjenetikë. Faktorë të tjerë.Toleranca imunitare. KAPITULLI 14. Mekanizmat imunopatollogjikë të inflamacionit. Mediatorët dhe molekulat që marrin pjesë në inflamacion. Ecuria e procesit inflamator. Kontrolli i inflamacionit dhe tërheqja e tij. Inflamacioni kronik. KAPITULLI 15. Mekanizmat imunologjike të mbrojtjes antiinfeksioze I. Mekanizmat imunologjike të mbrojtjes antiinfeksioze ndaj mikroorganizmave ekstraqelizore (bakterie, parazitë etj). Mekanizmat e shpëtimit të mikroorganizmave ndaj reaksioneve imune. KAPITULLI 16. Mekanizmat imunologjike të mbrojtjes antiinfeksioze II. Mekanizmat imunologjike të mbrojtjes antiinfeksioze ndaj mikroorganizmave intraqelizore (bakterie, këpurdha, viruse etj). KAPITULLI 17. Imunologjia e shtatzanisë. Faktorët që pengojnë flakjen e fetusit nga sistemi imunitar i nënes. Bashkëveprimi nëne-fetus dhe mekanizmat e aloimunizimit feto-amnor. KAPITULLI 18. Mangësite imunitare të lindura. Ndarja e mangësive imunitare të lindura sipas mekanizmave imunopatogjenetikë të tyre. Shënjat e tyre kryesore klinike dhe biologjike. KAPITULLI 19. Mangësite imunitare të fituara (me përjashtim të AIDS/SIDA). Ndarja e tyre në vartësi të shkaqeve bazë (rrjedhojë e sëmundjeve të ndryshme bazë). Përshkrimi dhe shënjat kryesore të tyre. KAPITULLI 20. Mangësia imunitare e fituar nga infeksioni me virusin HIV (AIDS-SIDA). Njohuri mbi shkaktarin dhe rrugët e infeksionit. Imunopatologjia e veprimit të virusit. Pasojat e veprimit të tij. KAPITULLI 21. Mekanizmat imunopatologjike I. Klasifikimi sipas Gell dhe Coombs i mekanizmave imunopatologjikë të mbindjeshmerisë të tipit të parë dhe të dytë. Shëmbuj të tyre. KAPITULLI 22. Mekanizmat imunopatologjike II. Klasifikimi sipas Gell dhe Coombs i mekanizmave imunopatologjikë të mbindjeshmërisë të tipit të tretë dhe të katërt. Shëmbuj të tyre. KAPITULLI 23. Autoimuniteti. C'janë reaksionet dhe sëmundjet autoimune. Hipotezat e ndryshme që shpjegojnë cfaqjen e tyre. Karakteristikat e sëmundjeve autoimune. KAPITULLI 24. Sëmundjet autoimune jo organ-specifike. Klasifikimi i tyre, mekanizmat kryesore patogjenetike, shënjat imunologjike. KAPITULLI 25. Sëmundjet autoimune organ-specifike. Mekanizmat imunologjike që marrin pjes ë në cfaqjen e tyre. Karakteristikat e sëmundjeve autoimune organspecifike (S.A.I.O.S.). Gjëndrat endokrine; Diabeti juvenil insulinovartës( DJIV); Tireoiditi i HASHIMOTO, M. BASEDOW-GRAVES, M. ADDISON, STERILITETI; Lëkura; Sistemi Nervor Qëndror dhe NEUROMUSKULAR; Sistemi Gastrointestinal: HEPARI; GJAKU; VESHKA; SYRI; ZEMRA; DIAGNOZA E S.A.I.O.S.). KAPITULLI 26. Sindromet imunoproliferative. Sindromet imunoproliferative monoklonale dhe poliklonale. Klasifikimi i tyre sipas llojit të qelizave. Diferencimi i SIP malinjë nga ato beninje. C'jane krioglobulinemite, llojet e tyre. Përcaktimi i SIP; Karakteri monoklonal; Klasifikimi; SIP i qelizave B; Mieloma multiple; Leucemia mieloide kronike me LB; Sindromi i Valdenstrom; Gamapatite monoklonale beninje; SIP me LT; Sindromi i Sezary si dhe Mycosis fungoide. Leucemia limfoide kronike me LT. Limfoma me qeliza T. SIP me Limfocite të padiferencuara. KAPITULLI 27. Imunologjia e tumoreve. Mekanizmat e bashkëveprimit sistem imunitar-tumor. Faktorët imune që pengojnë rritjet tumorale si dhe ata që favorizojnë rritjen e tyre. Shënjuesit kryesorë tumorale. KAPITULLI 28. Imunologjia e transplantit.

3
Llojet e ndryshme të transplanteve. Mekanizmat e flakjes imune të tyre. Kushtet e mosflakjes së transplanteve, roli i Kompleksit Madhor të Pajtueshmerise Indore (KMPI). KAPITULLI 29. Imunoterapia aktive. Vaksinimet, llojet e tyre. Imunoterapia pasive, llojet. KAPITULLI 30. Imunoterapia pasive, llojet. KAPITULLI 31. Metodologjia imunologjike. Bashkëveprimi antigen-antitrup në diagnostikën klinike. Klasifikimi i metodave dhe përshkrimi i tyre. Karakteristikat e reaksionit antigen-antitrup. Bashkëveprimi antigen-antitrup: specificiteti, komplementariteti, reversibiliteti, forcat që marrin pjesë në bashkëveprim, kushtet e bashkëveprimit. C' ështe afiniteti, aviditeti, konstatja e asocimit dhe disocimit. Klasifikimi i metodave dhe teknikave që studiojnë reaksionet antigen-antitrup. KAPITULLI 32. Reaksionet imunologjike të precipitimit. Principi dhe kushtet e realizimit të tij (Antitrupat dhe Antigenët detyrimisht në fazë të lënget, qënja të pakten bivalente, raporti molar i përshtatshem). Precipitimi cilësor në fazë të lënget: Ring - test-i, aplikimi i tij si metodë e shpejtë cilësore për vënjen në dukje të formimit të antitrupave, shembuj. Precipitimi sasior në fazë të lënget: përshkrimi i tij si një model metode sasiore imunologjike (kurba e precipitimit në fazë të lënget me 3 zonat e saj). Principi i nefelometrise. Precipitimi në xhel: Imunodifuzioni i dyfishte bidimensional në xhel (Ouchterlony) si metodë cilësore dhe gjysëm sasiore p ër studimin e antigenëve dhe antitrupave. Përdorimet e tij për qëllime eksperimentale (reaksionet e identitetit, mosidentitetit, identitetit të pjesshëm) dhe në praktikën diagnostike: Aplikimi në kërkimin e HBsAg apo të antitrupave antiberthamore solubel (anti Sm/RNP etj). Elektrosinereza; principi dhe pë rdorimet e saj. Metodat sasiore të munodifuzionit në xhel (imunodifuzioni i njëfishte bidimensional): Imunodifuzioni radial, rocket-imunoelektroforeza; përshkrimi, shëmbull nga dozimi i imunoglobulinave apo proteinave të tjera plazmatike. Imunoelektroforeza dhe Imunofiksimi : Principi dhe realizimi, aplikimet e tyre në studimin e proteinave serike. KAPITULLI 33. Reaksionet imunologjike të aglutinimit . C' ështe aglutinimi imunologjik. Kushtet e realizimit të tij: Aglutinacioni direkt ose aktiv, shëmbull nga përcaktimi i grupeve të gjakut si dhe në përcaktimin e Antigenëve bakteriale (Reaksioni i Vidal etj.). Aglutinacioni indirek ose pasiv: aplikimi i tij në provën e Coombs (c' është prova e Coombs direkte dhe indirekte, c’vënë ato në dukje, rëndesia diagnostike), përcaktimin e faktorit reumatoid (c' është faktori reumatoid, si dhe ku kërkohët ai me provën e Waaler - Rose dhe Latex) si dhe provën imunologjike të barrës. KAPITULLI 34. Matja e komplementit total dhe fraksioneve të tij. Titrimi i komplementit me metodën e hemolizës CH50, si realizohet dhe interpretimi i rezultatit. Metodat që përdoren në matjen e përbërësve të komplementit, interpretimi i rezultateve. KAPITULLI 35. Reaksionet imunologjike me antigenë ose antitrupa të shënuar. Parimet e përgjithshme të këtyre reaksioneve: dallimi nga metodat ë tjera (saktesia, ndjeshmeria, automatizimi, shpejtësia). Ndarjet e tyre sipas llojeve të ndryshme të shënuesve; fazë e lëngët apo e ngurtë, metodë direkte apo indirekte, sasiore apo cilësore. Reaksionet e imunofluoreshencës: Imunofluoreshenca direkte, përdorimi i saj në biopsitë indore. Imunofluoreshenca indirekte, përdorimi i saj në përcaktimin e antitrupave antibërthamore. Reaksionet imunoenzimologjike: Metoda direkte (në inde), paralelizmi i saj me imunofluoreshencën indirekte. Metoda ELISA indirekte apo sandwich, rënd ësia e sotme e saj në përcaktimin sasior të antitrupave apo antigenëve të ndryshëm, shëmbuj (Antitrupat anti-HIV, IgE totale). Metodat radioimunologjike, principi i tyre: Antigeni në fazë të ngurtë, shëmbull nga dozimi i IgE specifike. Antigeni në fazë të lëngët (në kompeticion me antigenin e shënuar), shëmbull nga dozimi i insulinës dhe hormonëve të tjerë. KAPITULLI 36. Metodat e Imunologjisë qelizore. Numurimi i limfociteve B me imunofluoreshencë direkte si dhe me antitrupa monoklonale. Numurimi i limfociteve T me metodën e rozetave E si dhe me antitrupat monoklonalë. Numurimi i qelizave të tjera limfocitare, metodat bashkëkohore (Fluorocitometria). Studimi i funksionit të qelizave T: me provat kutane (in vivo), dhe provat e stimulimit limfoblastik (in vitro) me mitogenë dhe antigenë. Si realizohen dhe rëndësia diagnostike e tyre. Studimi i citokinave. Studimi i funksionit të limfociteve

4
B: Përgjigja antitrupore in vivo me anë të studimit të përgjigjes antitrupore dhe in vitro me anë të stimulimit të prodhimit të anitrupave in vitro. Studimi i funksionit fagocitar.

5
KAPITULLI 1. IMUNITETI I LINDUR 1.1. Historik i shkurter, disa koncepte baze te imunollogjise 1.2. Barrierat lëkuro-mukoze. 1.3. Mekanizmat qelizore (me vrasje intraqelizore). Qelizat fagocitare; Fagocitoza. 1.4. Mekanizmat humorale: Rruga alterne e komplementit; Proteinat e fazes akute; Interferonet. 1.5. Mekanizmat me vrasje jashtëqelizore: Qelizat vrasëse - natyrale (NK cells); Eozinofilet.
1.1. Historik i shkurter, disa koncepte baze te imunollogjise Imunologjia eshte nje shkence relativisht e re. Fillimet e saj shkencore, megjithese empirike, i
atribuohen mjekut anglez Eduard Jenner i cili zbuloi ne 1796 qe lija e lopeve ose “vaksinia” indukton nje mbrojtje ndaj lijes se vertete (variola vera), semundje kjo ne ate kohe vdekjeprurese. Jenner e quajti kete procedure vaksinim dhe kjo terminologji vazhdon te perdoret akoma sot per te pershkruar imunizimin e individeve te shendoshe me shtame te dobesuar (atenuar) mikroorganizmash patogjene.
Megjithate, edhe per nje shekull me vone akoma asgje nuk do te dihej per shkakun e semundjeve infektive. Vetem ne fund te shekullit XIX mjeku gjerman Robert Koch provoi qe semundjet infektive shkaktohen nga mikroorganizmat patogjene perkates.
Ne te njejten kohe Louis Pasteur zhvilloi nje vaksine ndaj koleres se pulave si dhe ndaj terbimit, ndersa Emil von Behring dhe Shibasaburo Kitasato ne Gjermani zbuluan qe serumi i kafsheve te imunizuara me anatoksine tetanike ose difteritike permban nje “aktivitet antitoksik” (me vone keta u quajten antitrupa), i cili transferon nje imunitet pasiv afat-shkurter ne individet e paimunizuar, duke shenuar keshtu fillimet e imunoterapise (seroterapise) pasive.
Me sistem imunitar quajmë atë tërësi faktorësh indorë, qelizorë apo humoralë me të cilat organizmi mbrohet nga agresionet e huaja.
Imuniteti aktiv është ai që përpunon vetë organizmi pas futjes në të të një lënde të huaj. Imuniteti pasiv kemi kur fusim në organizëm antitrupa të gatshëm (ose dhe qeliza) që
mbrojnë organizmin nga një substance e huaj, të cilën shpesh e quajmë antigjen. Më i qëndrueshëm dhe afatgjatë është imuniteti aktiv qe mund te gjenerohet qoftë nga imunizimet natyrale ashtu dhe nga vaksinimet. Shembull nga imuniteti pasiv është injektimi i gamaglobulinave ose kalimi i imunoglobulinave te klases IgG nga nëna të fetusi gjate shtatzanisë dhe që i japin femijës një mbrojtje pasive derisa ai te aktivizojë mekanizmat aktive te mbrojtjes imunitare specifike.
Imuniteti natyral është krijuar gjatë evolucionit historik të vetë species. P.sh kolera e pulave nuk prek specien humane.
Imuniteti i fituar (adoptiv) ose specifik per antigjenin si psh. prodhimi i antitrupave ndaj nje patogjeni te caktuar quhet i tille sepse ai fitohet gjate jetes se individit si nje adaptim specifik ndaj infeksionit me kete patogjen.
Imuniteti i lindur ose jo specifik per antigjenin quhet i tillë sepse ai nuk kushtëzohet nga një kontakt paraprak me antigenin dhe eshte i gatshem qe ne lindje. Ai përbëhet nga një kompleks faktorësh mbrojtës të organizmit qe përbejne barrierën e parë ndaj një agresioni të jashtëm.
Në filogjenezë (evolucionin e specieve) imuniteti i lindur është i pari që formohet ndërsa imunitetin e fituar (adoptiv) e gjejmë të cfaqet më vonë në vertebrorët. Imuniteti i lindur është i gatshëm, funksional që në lindje në ndryshim nga imuniteti i fituar që aktivizohet vetëm pas një kontakti paraprak me antigjenin specifik. Por nga ana tjetër imuniteti i lindur duke qënë jo specifik për antigjenin ka një gamë relativisht të ngushtë veprimi ndaj agjentëve të jashtëm dhe nuk ka kujtesë imunitare, në ndryshim me imunitetin e fituar i cili duke qenë specifik ka një mundësi pothuajse të pakufizuar stimulimi ndaj cdo lloj antigjeni dhe nga ana tjeter ka nje kujtese imunitare qe bartet nga receptoret specifike per antigjenet ne siperfaqen e limfociteve. (Fig. 1.1.)
Faktorët e imunitetit të lindur:

6
1.2. Barrierat lëkuro-mukoze janë faktorë esencialë. Kur dëmtohet lëkura, për shembull në djegjet, organizmi rrezikohet shumë nga infeksionet. Djersa dhe gjëndrat e yndyrës prodhojnë acid laktik dhe acide yndyrorë që ushtrojnë një veprim inhibitor ndaj rritjes bakteriale si dhe realizojnë një pH të ulët të lëkurës i cili pengon zhvillimin e infeksioneve. Kur prishen këto faktorë mund të kemi zhvillim te infeksioneve në lëkurë dhe më thellë.
Një barrierë të rëndësishme përbën dhe mukusi që largon bakteret dhe pengon aderimin e tyre në mukozat apo ciliet që bëjnë lëvizjen e mukusit dhe agjentëve të huaj. Këtë rol luan dhe kolla apo teshtitja. Faktorë të tjerë mekanikë që ndihmojnë mbrojtjen e sipërfaqeve epiteliale është dhe veprimi shplarës i lotëve, pështymës apo urinës. Urina me pH e saj të ulur ruan rrugët urinare nga infeksionet. Po kështu dhe pH acid i lëngut gastrik. Shumë nga sekrecionet e organizmit përmbajnë përbërës baktericidale si lizozima në lotë, salivë dhe sekrecionet nazale, laktoperoxidaza në qumësht apo spermina dhe zinku në spermë.
Rol të rëndësishëm mbrojtës në mukozat luan flora normale bakteriale saprofite e mukozës intestinale ose flora vaginale. Këto pengojnë rritjen e mikroorganizmave patogjenë duke konkuruar për faktorët ushqyes ose duke prodhuar substanca inhibitore. Psh. flora normale e vaginës nëpërmjet prodhimit të acidit laktik ruan një pH acid që pengon zhvillimin e bakterieve. Kur kjo florë saprofite crregullohet si p.sh. nga përdorimi i antibiotikeve, rritet pH dhe rrezikohet infeksioni nga Candida albicans apo Clostridium difficile. Saprofitet e zorrës prodhojnë gjithashtu kolicina, një klasë baktericidinash.
Neqoftese mikroorganizmat arrijne te depertojne ne organizëm, 2 mekanizma te tjera futen ne loje: veprimi shkatërrues i faktoreve kimike solubël si psh. enzimat baktericidale si dhe mekanizmat e fagocitozës.
1.3. Mekanizmat qelizorë të imunitetit te lindur: Neutrofilet polimorfonukleare dhe Makrofagët (Fig. 1.2.) realizojnë të ashtuquajturën
vrasje intraqelizore: Neutrofilet jane leukocitet më të vegjël, s'kanë aftësi të shumëzohen dhe kanë një jetëgjatësi
të shkurtër. Ata përbëjne shumicën e leukociteve ne gjak dhe kanë nje citoplazëm granulare me 2 lloj granulash: Granulat azurofile primare formohen shpejt dhe përmbajne mieloperoksidazë si dhe shumicen e substancave antimikrobiale jo-oksiduese si katepsinen G, faktorin baktericid të rritjes së permeabilitetit (BPI) dhe difensinat. Granulat specifike sekondare peroksidaze – negative permbajne lizozimë, laktoferrinë, fosfatazë alkaline si dhe citokrom b558.
Funksioni i polimorfonukleareve neutrofile është fagocitoza dhe vrasja e mikroorganizmave me lokalizim jashtëqelizor siç jane psh bakteriet piogene.
Makrofagët janë qeliza me jetëgjatësi të madhe dhe kanë si funksion fagocitozën dhe pastaj vrasjen e mikroorganizmave me lokalizim intraqelizor. Ato jane derivate të monociteve. Monocitet kur lokalizohen në inde shndërrohen në makrofagë. Në mushkëri ato quhen makrofagë alveolare, në hepar qelizat e Kupferit, në shpretkë qelizat splenike, në veshka qelizat mezangiale, në sistemin nervor qelizat mikrogliale, në kockë qelizat osteoklaste.
Makrofagët funksionojnë si fagocite si dhe paraqitës të antigjenëve të përpunuar limfociteve T. Ky funksion i fundit ka të bëjë me imunitetin e fituar. Makrofagët aktivizohen nga limfocitet T CD4(+), produkte bakteriale si lipopolisakaridet (LPS) apo muramil-dipeptidi. Një funksion tjetër është stimulimi i mekanizmave të tjerë të imunitetit të lindur. Këto mekanizma, kryesisht ato humorale janë të rëndësishëm për të favorizuar fagocitozën. Favorizimi i fagocitozës quhet opsonizim. Një pjesë e mikroorganizmave prodhojnë substanca kimiotaktike që i afrojnë fagocitet, pra kanë efekt kimiotaktik, por një numër bakteriesh nuk i prodhojnë këto substanca dhe për të favorizuar kimiotaksis-in organizmi ka zhvilluar një sërë mekanizmash të tjerë.
Fagocitet kane mekanizma qe i mundësojnë ato te njohin përbërësit e vetë organizmit nga perberes te huaj potencialisht te demshem qe mund te jene mikroorganizmat patogjene.
Ne fagocitet janë zhvilluar keshtu nje sistem receptoresh te quajtur receptore te njohjes se llojit (pattern recognition receptors – PRR) te afte te njohin molekula te lidhura me agjente patogjene (pathogen associated molecular patterns – PAMP) te shprehura ne siperfaqen e agjenteve patogjene. PAMP jane kryesisht polisakaride dhe polinukleotide qe ndryshojne pak nga nje patogjen te tjetri por

7
që nuk gjenden në organizmin bujtes. Ndersa PRR ne siperfaqen e fagociteve jane kryesisht molekula te ngjashme me lektinat qe lidhen me shume valenca dhe specificitet te konsiderueshem me polisakaride dhe struktura te tjera PAMP në sipërfaqen e mikroorganizmave.
Ne ndryshim me receptoret specifike per antigjenet ne limfocitet, keta receptore nuk jane specifike per antigjenin dhe si rrjedhoje nuk jane shperndare ne menyre klonale por jane te pranishme ne te gjitha qelizat fagocitare.
Si pasojë e njohjes se PAMP nga ana e PRR kemi gjenerim të një sinjali nëpërmjet faktorit te transkripsionit NFkappaB (nuclear factor kappa B) i cili sinjalizon fagocitin per rrezikun dhe inicion kështu procesin e fagocitozës.
Lektina Lidhese e Manozes (MBL) eshte nje PRR e tille e pranishme si proteine e lire ne plazem dhe qe eshte e afte te inicioje aktivizimin e komplementit.
TLR (toll-like receptors) janë nje familje prej së paku 10 proteinash transmembranore ne siperfaqen e makrofageve dhe qelizave dentritike që njohin molekula mikrobiale te ndryshme. TLR2 njeh peptidoglikanin bakterial të bakterieve Gram – pozitive, TLR4 njeh lipopolisakaridin e bakterieve Gram- Negative (endotoksinën), ndërsa TLR3 dhe TLR5 janë të rëndësishëm në njohjen e ARN virale me varg te dyfishte. Kur aktivizohen TLR ato nxisin nje kaskade biokimike me aktivizim te NFkappaB dhe sintezen e citokinave proinflamatore dhe te peptideve te tjere antimikrobiale qe cojne drejt zhvillimit te imunitetit te fituar specifik per antigjenin.
Fagocitoza kryhet në disa faza (Fig. 1.3.): 1. Faza e kontaktit me agjentët e huaj realizohet nepermjet njohjes se PAMP nga fagocitet. 2. Faza e gëlltitjes dhe formimit të fagozomës shkaktohet nga aktivizimi i nje sistemi kontraktil aktine-miozine qe shkakton formimin e pseudopodeve qe mbeshtjellin pjesezen e fagocituar. 3. Brenda 1 minuti granulat citoplazmike shkrihen me fagozomin dhe shkarkojne permbajtjen e tyre brënda në fagozom duke realizuar keshtu tretjen e mikroorganizmit te fagocituar. 4. Faza e ekskretimit të materialit të fagocituar. Faza e dytë dhe e tretë do pak minuta, kurse e treta pak më tepër. Faza e tretë kryhet me dy mekanizma kryesore:
Mekanizmat oksigjen vartës te vrasjes intraqelizore fillojne me aktivizimin e theksuar te NADPH, kalimin e elektroneve nga kjo e fundit te citokromi cyt b 558 dhe reduktimi i oksigjenit molekular ne anion superoksid. Reaksioni kyc i katalizuar nga NADPH oksidaza qe inicion formimin e ndermjetesve reaktive te oksigjenit (ROI = reactive oxygen internediates) perfundon me formimin e joneve superokside, perokside, hidroksile që janë derivate të oksigjenit dhe që realizojnë vrasjen e mikroorganizmit.
NADPH + O2 oksidaze → NADP+ + O2 - (anion superoksid) Nen veprimin e superoksid dismutazes joni superoksid konvertohet ne peroksid te hidrogjenit
dhe si pasoje ne formimin e radikaleve hidroksile OH, i cili eshte nje nder radikalet e lire reaktive me te njohur. Nga ana tjeter, kombinimi i peroksideve, mieloperoksidazes (MPO) dhe joneve halogjene perben nje sistem te fuqishem halogjenizues i afte te vrase si bakteriet ashtu dhe viruset.
Nje cikel tjeter vrases eshte ai i oksidit nitrik (NO) qe formohet nga nje sintaze NO e
induktuar ne shume qeliza te trupit (iNOS) dhe qe ne neutrofilet dhe makrofaget humane gjeneron nje sistem te fuqishem antimikrobial.
Nderkohe qe NADPH oksidaza vepron kryesisht per vrasjen e mikroorganizmave ekstraqelizore te perfshire me ane te fagocitozes brenda ne fagozom, mekanizmi NO vepron ndaj mikrobeve qe hyjne ne citozol, pra dhe ndaj mikroorganizmave intraqelizore. Per kete aresye aftesia e vrasjes NO – iNOS eshte e pranishme dhe ne shume qeliza jofagocitare qe mund te infektohen nga viruse dhe mikroorganizma te tjere.
Neqoftese mikroorganizmat nuk vriten nga keto sisteme ato mund t’i nenshtrohen me tej nje familje peptidesh te quajtur difensina, dhe qe arrijne nivele shume te larta brenda fagozomit duke vepruar si dezinfektante ndaj nje varieteti te gjere bakteriesh, kepurdhash dhe virusesh. Nje goditje e

8
metejshme i jipet membranave bakteriale nga proteinazat neutrale (katepsina G) si dhe nga faktoret baktericidale dhe bakteriostatike se lizozima dhe laktoferrina. Se fundmi mikroorganizmat e vrare shkaterrohen nga enzimat hidrolitike dhe produktet e degradimit te cliruara jashte fagocitit.
1.4. Mekanizmat humorale. Sistemi komplementar (SK). SK eshte emertimi i nje sistemi kompleks prej 30 proteinash te pranishme ne plazem dhe ne
siperfaqet qelizore dhe i cili bashke me sistemin e koagulimit te gjakut, fibrinolizes dhe te formimit te kininave perbejne nje nga sistemet enzimatike plazmatike me te rendesishme. Te gjithe keta sisteme kane te perbashket gjenerimin e nje pergjigje te shpejte dhe te amplifikuar ndaj nje stimuli nxites dhe qe shoqerohet nga nje fenomen “kaskadë” ku produkti i nje reaksioni sherben si nje enzime katalizatore e reaksionit pasues. Molekulat e aktivizuara ose te copëtuara nga ky reaksion kaskade kane nje shumellojshmeri funksionesh mbrojtese dhe pa dyshim molekulat e komplementit mund te konsiderohen si pjese shume e rendesishme e sistemit imunitar te lindur. Disa nga perberesit e komplementit jane te emertuar nga germa C e ndjekur nga nje numer qe eshte me teper i lidhur me kronologjine e zbulimit te tij se sa me pozicionin ne rrjedhen e reaksionit.
Në imunitetin e lindur rol luan rruga alterne e aktivizimit të komplementit (Fig. 1.4.). Rruga alterne aktivizohet nga kapsulat e bakterieve (lipopolisakaridet bakteriale), endotoksinat bateriale si dhe nga imunoglobulinat A por pa nevojën e pranisë së kompleksit antigen-antitrup sic kërkon rruga klasike e aktivizimit të komplementit.
Molekula kyç e kësaj rrugë është molekula C3 që ndodhet në përqëndrim prej 1,2 mg/ml në plazëm. Ne rrethana normale, nën veprimin e faktorëve serike, sasi te vogla te C3 copetohen vazhdimisht duke dhene produktin C3b ose molekulen funksionalisht te ngjashme me te te quajtur C3bi. Ne praninie e joneve Mg2+ C3b ka aftësi të lidhet me faktorin B dhe formon C3bBb. Ky i fundit aktivizohet nga faktori D dhe gjenerohet keshtu C3bBb. Ky kompleks inhibohet (shkatërrohet) nga faktori H dhe I. Kur C3bBb lokalizohet në polisakaridet e membranave bakteriale ai i fshihet veprimit të faktorëve H dhe I, dhe si rrjedhoje ky kompleks formohet me shumicë dhe vepron si enzime (C3 konvertaze) duke e ndarë vazhdimisht C3 në C3a dhe C3b. Nje proteine tjeter e quajtur properdinë vepron duke e stabilizuar me tej konvertazen C3bBb.
Sistemi komplementar mund te aktivizohet dhe kur karbohidratet ne siperfaqen e bakterieve bashkeveprojne me nje proteine serike te quajtur lektine lidhese e manozes (mannose – binding lectin – MBL). MBL gjendet ne nivele te ulta ne serumin normal dhe lidhet me manozen apo karbohidrate te tjera ne siperfaqet bakteriale. MBL aktivizon komplementin duke ndervepruar me 2 serine proteaza te quajtura MASP1 dhe MASP2. MASP2 copëton dhe aktivizon C4 dhe C2 duke gjeneruar nje C3 konvertaze te quajtur
Kjo seri reaksionesh e provokuar nga vete mikroorganizmat con ne formimin e nje numri te madh molekulash C3b qe fiksohen ne siperfaqen e mikroorganizmit.
Me pas aktivizimi i komplementit vazhdon me bashkimin e nje molekule tjeter C3b me kompleksin
C4b2a.
C3bBb
Vetia tjeter shume e rendesishme e komplementit është ajo opsonizuese (favorizuese e fagocitozës). Ketë veti e ka C3b dhe C3bi. Kjo sepse të gjithë fagocitet kanë receptorë për C3b (CR1) dhe C3bi (CR3) dhe kur C3b mbulon një mikroorganizem atëhere lehtësohet shumë fagocitoza e këtij mikroorganizmi nga fagocitet që përmbajnë receptorë për C3b (CR1 dhe CR3).
duke gjeneruar keshtu nje C5 konvertaze. Kjo e fundit aktivizon C5 nepermjet nje copëtimi proteolitik duke cliruar keshtu nje polipeptid te vogel, C3a dhe duke lene fragmentin me te madh C5b te lidhur jo ngushte me C3b. Lidhja e metejshme e C6 dhe C7 me C5b formon nje kompleks me afinitet per C8 dhe me lidhje membranore tranzitore. C8 vendoset ne membrane dhe drejton ndryshimet konformacionale te C9 qe e transformojne kete te fundit ne nje molekule te afte te krijoje kanale transmembranore ne shtresen bilipidike te membranes te cilet lejojne kalimin e ujit dhe elektroliteve brenda ne qelize dhe si rrjedhim në sipërfaqen e bakterieve krijohen pore që sjellin lizën osmotike të tyre. Ky është veprimi citolitik i aktivizimit të komplementit.
Vetia e tretë është aktivizimi i inflamacionit. Kjo realizohet nga C3a dhe C5a. C5a ka edhe rol kimiotaktik që e dallon nga C3a. Por roli kryesor i C3a dhe C5a është ai anafilatoksinik. Ata veprojnë mbi bazofilet dhe mastocitet dhe realizojnë degranulimin e tyre. Bazofilet dhe mastocitet

9
janë të pasur me substanca inflamatore ku kryesoret janë: histamina, proteoglikanet, ECF (faktori kimiotaktik i eozinofileve), NCF (faktori kimiotaktik i neutrofileve), PAF (faktori aktivizues i trombociteve), interleukinat 3, 4, 5, 6, GM-CSF, TNF, leukotrienet, prostaglandinat dhe tromboksanet. Këta japin një sërë efektesh biologjike si vazodilatacion dhe grumbullim të gjakut në kapilarët, rritje të permebilitetit kapilar, veprojnë si substanca kimiotaktike etj.
C5a vepron direkt ne neutrofilet dhe si C5a ashtu dhe C3a veprojne mbi eozinofilet duke stimuluar shperthimin respirator te shoqeruar me prodhimin e ROI si dhe shprehjen e receptoreve siperfaqesore per C3b. Ne menyre te vecante C5a sherben gjithashtu si nje agjent i rendesishem kimiotaktik per neutrofilet. Si C3a dhe C5a kane nje aftesi te vecante te veprojne direkt ne endoteliumin kapilar duke prodhuar vazodilatacion dhe rritje te permeabilitetit kapilar, efekt ky qe duket se zgjatet nga leukotrieni B4 i cliruar nga mastocitet e aktivizuar, neutrofilet dhe makrofaget.
Kur ngacmohen nga agjentë të huaj, makrofagët sekretojnë citokina si interleukina 1 (IL1), IL6, TNF (faktori i nekrozes humorale) që luajnë rol në stimulimin e reaksioneve imunitare. Rol tjetër i interleukinave 1 dhe 6 është stimulimi i prodhimit të proteinave të fazës akute të inflamacionit (PFAI) ku përfshihen: proteina C reaktive, proteina lidhëse e manozes, a1- glikoproteina acide, përbërësi serik amiloid P, alfa1-anti-kimotripsina, C3, C9, Faktori B, fibrinogjeni, angiotensina, haptoglobina, fibronektina, ceruloplazmina, inhibitorët e alfa-1 proteinazes (Tab. 1.1.). Këta prodhohen kryesisht në melci dhe menjëherë, kur ka inflamacion. Matja e tyre në serum është tregues sasior inflamacioni. Rol të rëndësishëm klinik si tregues kryesor i inflamacionit ka proteina C- reaktive (PCR). Normalisht PCR pothuajse nuk është e pranishme në gjak dhe. në inflamacion mund te rritet deri 400 here. Roli i PCR qe përbëhet nga 5 vargje polipeptidike të njëllojta ështe që të aktivizojë komplementin me rrugën klasike pasi ajo të fiksohet në fosforilkolinat e sipërfaqeve bakteriale. Kjo rezulton ne depozitimin e C3b ne siperfaqen e mikroorganizmit i cili ne kete menyre opsonizohet dhe pergatitet per t’u fagocituar nga fagocitet. Matja e proteines C reaktive eshte nje test laboratorik mjaft i dobishem per te vleresuar shkallen e cdo procesi inflamator.
Ne pergjithesi duket se pergjigja e fazes akute kryen nje efekt te dobishem nepermjet rritjes se rezistences se organizmit, pakesimit te demtimit indor dhe riparimit te lezioneve inflamatore.
Interferonet jane nje familje agjentesh antivirale me spekter te gjere qe induktohen nga viruset dhe veprojne duke kufizuar proliferimin viral si dhe shperndarjen e infeksionit.
Interferonet jane 3 llojesh: Interferoni alfa- prodhohet nga të gjithë leukocitet, ka rreth 14 lloje te tilla, Interferoni beta – prodhohet kryesisht nga fibroblastet megjithese pothuaj te gjitha qelizat me berthame mund t’i prodhojne keto molekula. Interferoni gama - prodhohet nga limfocitet dhe ka kryesisht rolin e nje citokine qe merr pjese ne bashkeveprimin qelizor te pergjigjes imune. Funksioni i Interferoneve a dhe b është kryesisht ai antiviral. Ato sekretohen nga qelizat e infektuara nga viruset. Roli i tyre realizohet në këtë mënyrë: Pasi sekretohen nga qelizat e infektuara ato kapen nga receptorët përkatës të qelizave përreth. Në këto qeliza ato aktivizojnë 2 gene: Njëri gen i aktivizuar është përgjegjës për uljen e sintezës se proteinave. Geni tjetër është përgjegjës për prodhimin e një proteine që shkatërron ADN. Si rrjedhim. rrotull qelizës së infektuar nga virusi krijohet një brez qelizash refraktare ndaj këtij infektimi dhe në këtë mënyrë pengohet përhapja e virusit.
1.5. Vrasja ekstraqelizore. Qelizat e imunitetit të lindur që kryejnë vrasje jashtëqelizore jo specifike për antigenin quhen Qeliza NK. Qelizat NK (natural killer) kanë karakteristikat morfollogjike të qelizave limfoide por s'bëjnë pjesë as në limfocitet T dhe as në ata B dhe nga ana tjetër mund të cfaqin karakteristika nga të dy llojet e qelizave (Fig. 1.5.). Përsa i përket shënjuesve ata nuk kanë shënjuesit e limfociteve T apo B dhe si shënjues karakteristike të tyre kanë CD16 dhe CD56. Quhet ndryshe qelizë e madhe granulare (LGL) sepse duket më e madhe se limfocitet e tjerë dhe ka granula në citoplazëm. Qelizat NK kane nje qelize pararendese te perbashket me limfocitet T dhe shprehin si keto te fundit shenjuesin CD2. Ata kane gjithashtu receptore per interleukinen 2, proliferojne nen ndikimin e kesaj interleukine si dhe prodhojne intereferon gama. Por qelizat NK nuk maturohen ne timus dhe genet TCR te tyre nuk jane rirregulluar. Pikpamja e sotme eshte qe keto qeliza ndahen nga linja qelizore T shume heret ne diferencimin e kesaj te fundit.

10
Funksioni i NK është të vrasë qelizat e infektuara nga viruset si dhe qelizat tumorale por pa një specificitet për një antigjen të caktuar. Qelizat NK quhen ndryshe qeliza as T, as B, ose qeliza nul. Funksionin vrasës ndaj qelizës shënjë këto e realizojnë me ane te receptoreve aktivizues qe qelizat NK kane ne siperfaqen e tyre dhe qe njohin në sipërfaqen e qelizës shenjë glikoproteina me origjinë virale apo tumorale. Ligandet qe njihen nga keta receptore aktivizues mund te ndodhen edhe ne siperfaqen e qelizave normale te painfektuara dhe qelizat NK kane gjithashtu edhe receptore inhibitore qe parandalojne vrasjen e qelizave normale. Keta receptore inhibitore te quajtur KIR (killer immunoglobulin like receptors) njohin molekula ubikuitare (te kudondodhura) sic jane molekulat e Klases I te KMPI qe gjenden normalisht ne te gjitha qelizat me berthame. Duket se KIR mund te ndodhen ne forma te ndryshme molekulare si dhe kane nje llojshmeri alelike qe bartin jo vetem aftesi inhibuese por dhe aktivizuese te qe lizave NK. Sidoqofte qelizat e infektuara nga viruset apo dhe ato tumorale shpesh nuk i shprehin molekulat e klases se pare te pajtueshmerise indore. Pra, ne mungese te molekulave te Klases I te KMPI kemi pengese te stimujve inhibitore dhe si pasoje behet e mundur vrasja e qelizes shenje nga qeliza NK. Kjo njohje nuk është specifike për antigenin viral, por shërben për të kontaktuar qelizën NK me qelizën shënjë. Pas këtij kontakti membranë me membranë, nga qeliza NK dalin shumë substanca qe quhen citolizina ose perforina qe kane ngjashmeri me C9 dhe qe kontribuojne direkt ne vrasjen e qelizes shenje ne menyre te ngjashme me vrasjen me ndermjetesi te komplementit. Pervec perforines, granulat e qelizave NK permbajne limfotoksine alfa dhe nje familje te proteazave te serines qe quhen granzima. Njera nga keto, granzima B, vepron si faktor citotoksik NK duke induktuar apoptozen ne qelizen shenje. Nje sistem tjeter alternativ njohje per vrasjen qelizore me ndermjetesine e qelizave NK perfshin angazhimin e molekulave te aktivizuara te receptorit Fas ne siperfaqen e qelizes shenje nga FasL (Fas-ligand) ne qelizen efektore NK, proces edhe ky qe indukton nje sinjal apoptik ne qelizen shenje. Edhe pjestare te familjes se faktoreve te nekrozes tumorale mund te nderveprojne me receptore ndaj TNF ne qelizat shenje duke ndermjetesuar citotoksicitetin.
Qelizat NK aktivizohen nga interferonet që sic dihet prodhohen nga qelizat e infektuara nga viruset si dhe nga citokina te tjera te prodhuara nga makrofaget. Qelizat NK lidhen me qelizën shënjë edhe me anë të IgG që mund të mbulojnë këto të fundit pasi qelizat NK kanë receptorë për fragmentin Fc të IgG. Ky fenomen quhet dhe ADCC (antibody dependent cellular citotoxicity).
Qelizat NK jo vetem kane aftesine te lizojne qeliza te infektuara me viruse si dhe qeliza tumorale por ato prodhojne gjithashtu pas aktivizimit te tyre nje game te gjere citokinash. Ketu perfshihen citokinat si IL-1 dhe TNF qe luajne nje rol te rendesishem ne inflamacion si dhe faktori granulocitar dhe makrofag stimulues i kolonive (GM-CSF), interferoni gama (IFN gama) si dhe faktori i rritjes dhe transformues beta (TGF beta), te cilet modulojne pergjigjen imunitare te fituar. Qelizat NK shprehin gjithashtu molekula kostimulatore si CD40L (ligandi i CD40) dhe gjithashtu kane demostruar se ne rast aktivizimi te tyre cfaqin nje funksion rregullues te funksionit te limfociteve B.
Qeliza të tjera vrasëse ekstraqelizore jane eozinofilet. Ato janë leukocite të pasura me granula të cilat përmbajnë shumë substanca me efekt vrasës për qelizat shënjë. Këto substanca janë: substanca MBP (proteina madhore bazike), peroxidaza, histaminaza, arilsulfataza B, fosfolipaza D. Funksioni kryesor i eozinofileve është vrasja e parazitëve. Parazitet shume qelizore si psh helmintet nuk mund te fagocitohen dhe vrasja ekstraqelizore me ane te eozinofileve duke se ka evoluar pikerisht per kete qellim. Eozinofilet, pasi prodhohen ne palcen kockore kalojne në gjakun periferik dhe pastaj ne indet periferike si mushkeria apo zorra. Vendosja e tyre ne keto inde tregon se ato luajne nje rol te rendesishem ne mbrojtjen e mukozave. Eozinofilet kanë receptorë për citokinat, kemokinat, molekulat e adezionit, perberesit e komplementit dhe ne aktivizim ata gjenerojne nje shperthim te fuqishem respirator me prodhim te njekohshem te metaboliteve aktive te oksigjenit dhe te citokinave proinflamatore.
Shumica e helminteve provokojne aktivizimin e rruges alterne te komplementit, dhe megjithese ato mund te jene rezistente ndaj sulmit membranor te C9, mbulimi i tyre me C3b lejon aderimin e eozinofileve nepermjet receptoreve C3b te ketyre te fundit. Neqoftese ky kontakt do te coje ne aktivizim, eozinofili fillon sulmin e tij ekstraqelizor qe perfshin clirimin e proteines madhore

11
bazike (MBP) te pranishme ne granulat e eozinofileve dhe e cila demton membranen e parazitit.. Si rrjedhim realizohet vrasja e parazitëve nga eozinofilet nëpërmjet daljes edhe të substancave të tjera citotoksikë që përmbajnë granulat e tyre. KAPITULLI 2. ORGANET E IMUNITETIT 1.1. Organet parësore ose qëndrore : Timusi dhe Palca kockore. 1.2. Organet dytësore (periferike) : Nyjet Limfatike; Shpretka dhe Indi limfoid i lidhur me mukozat (MALT). 1.3. Rruga e antigenit kur hyn në organizëm, qarkullimi i limfociteve.
Përveç imunitetit të lindur, në vertebrorët është zhvilluar edhe imuniteti i fituar që ka këto veçori: 1. Mekanizmat e tij realizojnë një njohje specifike të antigenëve të ndryshëm. 2. Që të realizohet kjo njohje specifike duhet një kontakt paraprak i sistemit imunitar të fituar me antigenin. 3. Gama e veprimit ndaj antigenëve të ndryshëm është shumë më e gjërë për shkak të shumëllojshmërisë shumë më të lartë të specificiteteve antigenike që mund të shohë sistemi i

12
imunitetit të fituar. 4. Imuiteti i fituar zhvillohet pas lindjes si pasojë e kontaktit me agjentët e huaj në ndryshim me imunitetin të lindur që është plotësisht funksional që në lindje. 5. Imuniteti i fituar zoteron kujtese imunitare, pra kontakti i dyte dhe ata te metejshem me nje antigjen specifik gjenerojne nje pergjigje imunitare shume me te shpejte dhe me te fuqishme se kontakti fillestar. Qe te ndodhe nje pergjigje imunitare efikase nevojitet nje bashkepunim qelizor i komplikuar. Antigjeni duhet te bashkeveproje dhe eventualisht te perpunohet nga nje qelize antigjen paraqitese (QAP), e cila nga ana e saj duhet te bashkeveproje me kontakt me limfocitet T dhe B antigjen-specifike. Nder keta te fundit limfocitet T helper duhet te nxisin limfocitet B dhe ato T citotoksike te realizojne funksionet e tyre nepermjet shumefishimit te qelizave efektore potenciale dhe gjenerimit te mediatoreve te imunitetit qelizor dhe humoral. Pervec kesaj nevojitet formimi i limfociteve kujtese, pergjegjes per pergjigjet imunitare dytesore si dhe e gjithe kjo pergjigje imunitare duhet kontrolluar ne menyre qe te jete e pershtatshme me llojin dhe intensitetin e agresionit dhe mos te jete e ekzagjeruar pasi kjo mund te behet e demshme per organizmin. Ky skenar kompleks nderveprimesh qelizore qe perbejne bazen e pergjigjes imunitare realizohet ne indet limfoide te organizuara te quajtura periferike ose sekondare ku perfshihen nyjet limfatike (LN= lymph nodes), shpretka si dhe indi limfoid i painkapsuluar i lidhur me mukozat (MALT). Sistemi imunitar i fituar ka organet, indet dhe qelizat e tij. Keto organe e inde popullohen nga qeliza me origjine retikulare, makrofage dhe limfocite qe rrjedhin nga qelizat baze hematopoietike te palces kockore (HSC = hematopoietic stem cells). Limfocitet T diferencohen ne qeliza imunokompetente gjate nje procesi te komplikuar edukimi ne timus nderkohe qe per limfocitet B ky proces ndodh ne vete palcen kockore. Si permbledhje mund te thuhet qe nyjet limfatike drenojne dhe ne se eshte e nevojshme i pergjigjen molekulave te huaja (te tretshme apo ne perberjen e qelizave) qe vijne nga indet e ndryshme te organizmit, shpretka filtron gjakun, ndersa MALT sherben si barriere mbrojtese e gjithe mukozave te organizmit. Palca kockore kontribuon gjithashtu ne menyre te rendesishme ne prodhimin e antitrupave (2.0.). Komunikimi midis organeve limfoide paresore, dytesore si dhe organeve e indeve te tjera te organizmit realizohet nga nje popullate qarkulluese limfocitesh qe kalojne nga gjaku ne nyjet limfatike, shpretke apo indet e tjera dhe nga keta perseri ne gjak nepermjet eneve kryesore limfatike si psh. duktusi toracik.
Organet dhe indet e sistemit imunitar i ndajme ne ato qendrore ose paresore si dhe ne ata dytesore ose periferike.
1.1.Ne organet qendrore (ose paresore) bejne pjese timusi dhe palca kockore. Ne to ndodh maturimi i qelizave specifike te imunitetit qe jane respektivisht limfocitet T dhe ato B. Keto organe jane te zhvilluar mire qe ne lindje pasi zhvillimi i tyre nuk ka lidhje me stimujt antigjenike sic ndodh me organet limfoide dytesore.
Timusi:. Timusi është organ bilobar i vendosur në mediastinin superior/anterior mbi zemër. Të dy lobet
janë të ndërtuar nga lobule që përbëjnë njësinë funksionale të timusit. Në çdo lobul dallojmë zonën kortikale dhe medulare (Fig. 2.1.). Qelizat epiteliale te vendosura ne te 2 zonat perbejne mikromjedisin qelizor ku diferencohen limfocitet T. Ketu kemi nderveprime komplekse midis proteinave te matriksit ekstraqelizor dhe mjaft integrinave ne popullatat e ndryshme limfocitare, te cilat se bashku me kemokinat dhe me shprehjen e receptoreve ndaj ketyre te fundit luajne rol ne gjetjen e shtepise (fenomeni “homing”) te qelizave pararendese te limfociteve T ne timus dhe ne migrimin e tyre ne kete organ. Qelizat epiteliale prodhojne nje sere hormonesh peptidike sic jane timulina, timozina α 1, faktori timik serik si dhe timopoietina te cilat ndikojne ne cfaqjen e markuesve te diferencimit te limfociteve T. Zona kortikale është e përbërë nga limfocite të dëndur që quhen timocite. Këto ndodhen edhe në medul por më pak të dëndur. Qelizat e specializuara kortikale ne korteksin e jashtem jane te njohura si qeliza edukuese ( nurse cells) sepse ato nderveprojne me nje numer te madh limfocitesh te cilet gjenden ne citoplazmen e ketyre qelizave. Qelizat epiteliale te korteksit me te thelle kane procese dentritike te degezuar dhe te pasur ne

13
molekula te klases se dyte te kompleksit madhor te pajtueshmerise indore (KMPI). Nepermjet desmozomeve te tyre ato nderthuren dhe formojne nje rrjet nepermjet te cilit limfocitet kortikale duhet te kalojne per ne zonen medulare. Limfocitet kortikale jane mjaft me te dendur se sa ata ne zonen medulare dhe shumica e tyre pesojne procesin e apoptozes si rrjedhoje e seleksionit pozitiv dhe negativ. Ne zonen medulare jane te pranishme dhe nje numer qelizash dentritike interdigitale me origjine nga palca kockore dhe qelizat epiteliale te kesaj zone kane zgjatime me te gjera se sa partneret e tyre kortikale si dhe shprehin nivele te larta si te klases se pare dhe asaj te dyte te KMPI. Qelizat epiteliale në pjesën medulare bashkohen edhe në formacione të rrumbullakta që quhen trupëzat e Hassall-it. Përveç kësaj në lobulin timik dallojmë shumë makrofagë. Veçori edhe e ketyre është se shprehin mjaft molekulat e kompleksit madhor të pajtueshmërisë indore (KMPI).
Te njeriu involucioni i timusit fillon qe ne 12 muajte e pare te jetes dhe kjo gjender reduktohet rreth 3% cdo vit deri ne moshat e mesit dhe me pas rreth 1% cdo vit. Madhesia e organit nuk reflekton keto ndryshime pasi kemi zevendesim me ind dhjamor. Timusi diferencon limfocite T jetegjate te cilet i lejojne organizmit te vazhdoje imunokompetencen e tij edhe ne rast heqje te plote te ketij organi ne moshat e rritura ndersa ne rast se heqja e timusit ndodh tek neonati apo ne vitin e pare te jetes kemi efekte te dukshme ne imunokompetencen e femijes.
Diferencimi dhe maturimi i limfociteve T ne timus: Funksioni i timusit është maturimi i limfociteve T (T si timus). Në palcën e kockave kemi qeliza
mëmë (stem cells) që nën ndikimin e qelizave stromale të palcës së kockave diferencohen në të gjithë elementët e figuruar të gjakut. Qelizat meme shprehin receptore te kemokinave te cilat terhiqen ne timus nga bashkeveprimi me kemokinat e prodhuara nga qelizat stromale te timusit.
Pararendesi i limfociteve T eshte pozitiv per shenuesin CD34 si dhe per enzimen terminal deoxynucleotidyl transferaze (TdT) e cila vepron duke rritur diversitetin e receptoreve per antigjenin ne limfocitet T (TCR) (Fig. 2.2.). Nen veprimin e IL-1 dhe te faktorit te nekrozes tumorale (TNF) pararendesi T diferencohet ne protimocit. Ne kete stad keto qeliza shprehin receptorin TCR si dhe shenjuesin CD3 i cili eshte kompleksi invariant i percimit te sinjalit nga TCR dhe keto qeliza behen dyfish pozitive per CD4 dhe CD8. Se fundi, nen drejtimin e kemokinave keto qeliza pershkojne lidhjen kortiko-medulare dhe kalojne ne medule ku ato tashme behen qeliza prekursore imunokompetente T helper njefish pozitiv CD4(+) si dhe T citotoksike CD8(+). Qelizat me TCR gama/delta (λδ) ngelin dyfish negative CD4-CD8- pervec nje popullate te vogel qe shpreh CD8.
Pararendesit T te hershem i kane genet TCR ne konfigurim germinal dhe rirregullimi i pare i tyre perfshin se pari lokuset λ dhe δ. Receptoret α dhe β cfaqen pak dite me pas. Geni Vβ rirregullohet se pari ne qelizat dyfish negative CD4- CD8- dhe asociohet me nje varg invariant pre–α si dhe molekulen CD3 per te formuar nje “pre-TCR”. Shprehja e ketij kompleksi e con qelizen pre-T drejt proliferimit dhe kthimit ne nje qelize dyfish pozitive CD4+CD8+. Diferecimi i metejshem kerkon tashme rirregullimin e e genit Vα duke lejuar formimin e nje receptori TCR αβ te pjekur dhe keshtu keto qeliza jane tashme gati per te pesuar seleksionet pozitive dhe negative si me poshte. Rirregullimi i geneve V β ne kromatidin moter suprimohet me nje proces te quajtur ekskluzion alelik keshtu qe cdo limfocit T shpreh vetem nje varg β te TCR te tij.
Persa i perket shenjuesve te tjere T, timocitet kortikale shprehin menjëherë shënjuesit sipërfaqësore CD2 dhe CD7. Nën influencën e IL-1 si dhe të faktorëve sipërfaqësore të tjerë këtu ndodh një ekspansion i madh i timociteve kortikale. Më pas cfaqen CD1, CD5, CD3 si dhe TCR.
Limfocitet T seleksionohen pozitivisht ne timus te njohin KMPI e vetes: Aftesia e limfociteve T te njohin peptidet antigjenike ne asocim me KMPI e vetes zhvillohet ne
timus. Nje perqindje e vogel e timociteve dyfish pozitive CD4+CD8+ kane TCR qe lidhet me aviditet te ulet me molekulat e KMPI te shprehura ne qelizat epiteliale timike kortikale dhe keshtu ato seleksionohen pozitivisht per te perfunduar progresin e tyre drejt nje qelize T te pjekur. Qelizat e tjera qe kane TCR te paafte te njohe KMPI e vetes eliminohen dhe vdesin brenda 3 ose 4 ditesh ne proces apoptoze. Nje vecori tjeter e ketij seleksioni pozitiv eshte qe qelizat CD4+CD8+ me TCR qe njeh KMPI e vetes ne qelizat epiteliale, selektohen gjithashtu pozitivisht per t’u diferencuar ne qeliza njefish pozitive CD4+CD8- ose CD4-CD8+. Ai timocit qe njeh molekulat e klases se pare te KMPI do te maturohet si qelize CD8+ ndersa nqs. kontakti behet me molekulat e KMPI se klases se dyte qeliza

14
do te diferencohet si CD4+. Ne mangesine imunitare te lindur te rralle qe quhet Sindromi i limfocitit te xhveshur (bare lymphocyte syndrome), molekulat e KMPI nuk cfaqen ne siperfaqen e qelizave. Si pasoje, ne rastet me mungese te KMPI se klases se pare, nuk kemi maturim te Limfociteve T CD8+, ndersa ne rastet me mungese te molekulave te klases se dyte te KMPI nuk kemi Limfocite T CD4+.
Limfocitet T qe njohin autoantigjenet eliminohen ne timus nepermjet seleksionit negativ: Timusi luan një rol të rëndësishëm në eliminimin e limfociteve T autoreaktive (që njohin
antigenët e vehtes : self-antigens). Në këtë mënyrë eliminohet rreziku i autoimunitetit. Shume nga qelizat qe i mbijetojne seleksionit pozitiv kane receptore per antigjenet e vetes
(autoantigjenet) dhe neqoftese ato do te lejoheshin te maturoheshin do te prodhonin nje pergjigje imunitare ndaj autoantigjeneve. Keto limfocite pesojne ne fakt nje seleksion negativ qe eshte i domosdoshem per te ruajtur tolerancen ndaj autoantigjeneve. Ne kete proces autoantigjenet u paraqiten timociteve ne maturim e siper nepermjet qelizave dentritike apo makrofageve dhe cdo timocit qe lidhet me aviditet te larte te TCR se tij per kete autoantigjen do te eliminohet nepermjet apoptozes.
Pra seleksionet pozitive dhe negative që u përshkruan më lart ndodhin në kushtet e afinitetit të ulët (seleksioni pozitiv) dhe atij të lartë (seleksioni negativ) ndërmjet nga njera ane TCR dhe KMPI së vehtes apo TCR dhe KMPI + autoantigen nga ana tjeter. Mundet që qelizat epiteliale kortikale duke qënë paraqitëse relativisht jo efikase të antigenit japin një sinjal të dobët, megjithëse të restriktuar me KMPI e vehtes dhe shkaktojnë kështu një proliferim të timociteve imature. Nga ana tjetër, qelizat dentritike dhe makrofagët i përpunojnë autoantigenët në menyrë mjaft efikase dhe i dërgojnë kështu një sinjal të fortë timociteve autoreaktive (që njohin KMPI të vehtes + autoantigen) duke shkaktuar kështu një seleksion negativ të limfociteve T autoreaktive.
Rezultati i seleksioneve pozitive dhe negative eshte qe te gjithe limfocitet T te pjekur qe gjenden ne medulen e timusit do te njohin peptidet e huaja vetem ne kontekstin e KMPI se vetes dhe nuk do te kene mundesi te zhvillojne pergjigje imunitare ndaj autoantigjeneve dhe ne rastin e limfociteve me TCR α β do te jene njefish pozitive: ose CD4(+) ose CD8(+).

15
Fig. 2.1. Struktura e timusit Palca e kockave: Quhet organ parësor sepse pervec funksioneve te tjera qe ka palca kockore
ajo sherben dhe si organ paresor per maturimin e limfociteve B. Pararendesit e limfociteve B, qelizat pro-B jane te pranishme ne melcine fetale ne javen e 8-9 te barres. Prodhimi i limfociteve B nga hepari gradualisht zhduket dhe ky funksion merret nga palca kockore per gjate gjithe jetes.
Ky maturim bëhet nga kalimi i një qelize meme hematopoietike ne palcen kockore ne qelize pro-B, qelize pre-B, qelize B e papjekur dhe se fundi ne qelize B te pjekur. Qelizat stromale retikulare të palcës kockore shprehin molekulat e adezionit dhe sekretojnë Il-7. Me anë të zgjatimeve dentritike të tyre ato realizojnë një kontakt të ngushtë me pararendësit e limfociteve B që shprehin receptorë për Il-7. Maturimi i limfociteve B në palcën kockore ndjek disa etapa të fitimit të shënuesve diferencues (Fig. 2.3.).

16
Qeliza limfoidememe
Qelize pro-B Qelize pre-B Qelize Be papjekur
Qelize B e pjekur
Rregullim i geneve te Ig.Shprehje e TdT
Shprehje e CD10 "CALLA" Shprehje e CD 19
CD 20CD 21 (CR2)
CD 23 /Fc eps. R IICD 45 R
KMPI KL. IIFc gama R
myM M/D
Fig.2.3. Rirregullimi i geneve te imunoglobulinave gjate maturimit te limfociteve B: Faza 1: Se pari kemi rirregullim te segmenteve D-J ne te dy regjionet koduese te imunoglobulinave
(nje nga cdo prind) (Fig. 2.4.) Faza 2: Rikombinimi V-DJ ndodh ne nje varg te rende. Nqs. Ky rirregullim del jo-produktiv (jo
kodues) ndodh nje rikombinim V-DJ i dyte ne regjionin qe kodon per vargjet e renda te kromatidit tjeter. Nqs edhe ketu nuk kemi rirregullim produktiv qeliza pre-B nuk diferencohet me tej dhe eliminohet me apoptoze.
Faza 3: Nqs. rirregullimi produktiv arrihet, qeliza pre-B mund te sintetizoje tani vargje te renda μ.
Pothuajse ne te njejten kohe, 2 gene, V preB dhe λ5, homologe me segmentet respektive VL dhe CL te vargjeve te lehta λ transkriptohen perkohesisht duke formuar nje “varg pseudo te lehte” qe bashkohet me vargjet μ duke prodhuar nje IgM qe bashke me vargjet Ig-α dhe Ig-β eshte nje imitim receptori ne limfocitet B. Ky receptor “surrogat” ka nje paralelizem ne formimin e tij me receptorin pre-T α/β ne qelizat prekursore pre-T me receptore TCR αβ.
Faza 4: Receptori siperfaqesor sinjalizohet, ndofta nga nje qelize stromale, te suprimoje cdo
rirregullim te metejshem te geneve te vargjeve te renda ne kromatidin tjeter simoter. Ku fenomen quhet ekskluzion alelik.
Faza 5: Mendohet se receptori siperfaqesor fillon tashme nje seri rrirregullimesh gjenike qe ndodhin
ne lokuset e vargut te lehte κ. Ketu perfshihen rekombinimet V-J ne njerin alel κ dhe pastaj ate tjeter derisa arrihet nje rirregullim Vκ-J produktiv. Nqs. Ky deshton iniciohet nje tentative tjeter per te arritur nje rirregullim produktiv te geneve λ. N ekete menyre fillohet sinteza e imunoglobulinave M siperfaqesore (IgMs) qe sherbejne si receptor per antigjenet ne siperfaqen e limfociteve B.

17
Faza 6: Molekula IgMs e formuar ndalon rikombinimet e tjera te geneve te imunoglobulinave me ane
te ekskluzionit alelik. Prodhimi i metejshem i nje imunoglobuline siperfaqesore IgD ne siperfaqen e limfocitit B shenon tashme gatishmerine e ketij limfociti B “naiv” per t’u stimuluar nga antigjeni specifik. Limfocitet B me receptore me afinitet te mesem dhe te larte per autoantigjenet eliminohen nepermjet nje procesi seleksioni negativ te ngjashem me ate qe eliminon limfocitet T reaktive ne timus.
Si pasoje e kontaktit me antigjenin specifik per te, limfociti T naiv transformohet ne plazmocit sekretues te IgM ose peson nje kalim (“switch”) izotipik si pasoje e te cilit IgM dhe IgD membranore zevendesohen me nje klase tjeter te vetme imunoglobulinash si IgG, IGA ose IgE. Ne stadet terminale te jetes se nje plazmociti te maturuar plotesisht prodhohet vetem versioni i sekretuar i kesaj klase imunoglobuline pa prani te atyre siperfaqesore.
Rendesia e ekskluzionit alelik: Meqenese cdo qelize ka dy kromozome prinderore, qeliza B ne diferencim ka 4 komplete
genesh te vargjeve te lehta (2 kapa dhe 2 lambda nga secili prind) dhe 2 te renda (1 nga cdo prind) te gatshem per t’u rekombinuar. Ne rast se ndodh nje rikombinim produktiv VDJ i ADN ne nje komplet genesh te vargjeve te renda, kompleti ne kromozomin tjeter ngel pa rekombinuar si rrjedhoje e ketij procesi te quajtur ekskluzion alelik. Para, qeliza shpreh vetem nje varg te lehte dhe nje te rende. Kjo eshte esenciale per funksionimin e seleksionit klonal sipas te cilit nje limfocit B eshte i programuar te shprehe vetem nje lloj molekule antitrupi dhe pikerisht ate qe perdor si receptor siperfaqesor per njohjen e antigenit.
1.2. Organet dytesore te Sistemit Imunitar. Përveç organeve parësore kemi organet dytesore ose periferike të sistemit imunitar. Dallimi midis tyre qendron ne faktin qe: 1 Ndërsa organet parësore janë të zhvilluar mirë që në lindje të fëmijes, ato periferike
zhvillohen pas lindjes nën veprimin e antigenëve të ndryshëm. 2. Antigenët nuk depërtojnë asnjëherë në organet parësore, ndërsa depërtojnë vazhdimisht në
ato dytësore. 3. Limfocitet që dalin të maturuar nga organet parësore nuk rikthehen me aty ndërsa
limfocitet e organeve periferike qarkullojnë dhe rikthehen vazhdimisht në këta organe. 4. Organet parësore e realizojnë funksionin e tyre maturues pavarësisht nga stimujt
antigenikë, ndërsa organet dytësore nuk e realizojnë funksionin pa stimulim antigenik. Organet sekondare të kapsuluar: Nyjet limfatike janë organe më të vegjël se 1 mm, në formë fasule. Ndodhen në numër rreth
1000 në organizëm. Funksioni i tyre është të drenojnë limfën që vjen nga indet, lëkura dhe mukozat. Me anë të kësaj limfe futen edhe antigenët (me enët limfatike aferente që depërtojnë në kapsulën e nyjeve limfatike). Pra nyjet limfatike mund të konsiderohen si filtër për antigenët qe vijne me rruge limfatike. Në to dallojmë kapsulën ku depërtojnë enët limfatike aferente nga ku limfa kalon në sinusin subkapsular dhe më pas del nëpërmjet enëve limfatike eferente. Ne nyjet limfatike dallojmë zonën kortikale që është e pasur me limfocite B të organizuara në folikuj limfoide të diferencuar si vijon: Folikul parësor ku antigeni nuk është futur asnjëherë dhe folikul dytësor kur antigeni tashmë ka depërtuar në folikul. Karakteristikë e folikulit dytësor është se përmban një qëndër germinative ku ka shumëzim të limfociteve B. (Fig. 2.5) Ne folikujt limfoide ndeshim edhe makrofagë si dhe qeliza dentritike folikulare që realizojnë paraqitjen e antigenit limfociteve B. Folikujt limfoide quhen ndryshe dhe zona timoindipendente (ose B- vartese) e nyjes limfatike. Ajo pjese e limfociteve B qe kalojne ne qeliza B kujtese zejne vend ne zonen mantel dhe jane limfocite te vegjel ne qetesi qe kane si IgM dhe IgD ne siperfaqen e tyre. Zona parakortikale është e pasur me limfocite T dhe qeliza

18
dentritike dhe quhet ndryshe zona timovartese e nyjes limfatike. Zona medulare ka shumë makrofagë si dhe plazmocite që kanë ardhur nga zona kortikale.
Fig. 2.5. Ndërtimi i nyjes limfatike Shpretka: Kjo shërben si filtër i antigenëve që depërtojnë në rrugët e gjakut (Fig. 2.6). Ne te ndodh
shkaterrimi i eritrociteve dhe leukociteve te vjeteruar si dhe pergjigja imunitare ndaj antigjeneve qe hyjne me rruge gjaku sidomos ndaj atyre qelizore.
Shpretka ka 3 zona të ndryshme: a. këllëfi periarteriolar që mbështjell arteriolat ose pulpa e bardhë (ajo ka limfocite T me shumicë prandaj quhet zona timovartëse). b. zona marginale ku ndodhen folikujt limfoidë, përbëhet nga limfocite B dhe quhet zonë B vartëse c. pulpa e kuqe që përbëhet nga sinusoide venoze dhe makrofagë e plazmocite. Ne kete te fundit ndodh shkatërrimi i eritrociteve.
Organet jokapsulare - indi limfoid i lidhur me mukozat (MALT). Ky ndodhet i shpërndarë nën gjithë mukozat e organizmit, p.sh. mukozat digjestive, mukozat
respiratore, urogenitale, gjëndrat e qumështit gjatë laktacionit. Karakteristike e tyre eshte se jane perqendrime subepiteliale te indit limfoid te parrethuara nga nje kapsul e indit lidhor sic ndodh ne limfonodulet. MALT-i në traktin respirator përbëhet nga makrofagë që veshin submukozën. Vende-vende kjo formon grumbullime që quhen nyja limfoide subepiteliale. Ndërsa në traktin tretës dallojmë unazën e Valdeyerit dhe pllakëzat e Peyer-it si dhe indin limfoid të apendiksit. Në MALT antigeni futet nëpërmjet mukozës dhe nxit qelizat limfoide që shndrrohen në limfocite T dhe B efektorë (këta të fundit shndrrohen në plazmocite prodhues kryesisht të imunoglobulinave A sekretore), si dhe në limfocite T dhe B kujtesë që qarkullojnë kryesisht në lokalizime të ndryshme të MALT.
2.3. Rruga që ndjek antigeni kur depërton në organizëm, qarkullimi i limfociteve: Kur një antigen hyn në një ind ai kapet nga disa qeliza të specializuara që quhen qeliza
dentritike dhe transportohet në rrugë limfatike në folikujt limfoide të nyjeve limfatike. Aty ai u paraqitet limfociteve B nga qeliza dentritike folikulare. Kur antigeni kalon në zonën parakortikale ai i paraqitet limfociteve T nga qeliza dentritike interdigitale. Kur antigeni depërton nëpërmjet gjakut në

19
organizëm ai kalon në shpretkë në tre zonat e saj. Sot pranohet se qelizat dentritike realizojnë në përgjithesi paraqitjen e antigenit që futet për herë të parë në organizëm ndërsa makrofagët përpunojnë antigenët që futen për herë të dytë dhe në vazhdimësi.
N.q.s antigeni futet në mukozë pra ai kalon epitelin e mukozave dhe lokalizohet në indin limfoid të submukozës ai stimulon qeliza limfoide të atjeshme. Kur antigeni kalon nëpër organet limfoide nëpëmjet rrugëve limfatike ai stimulon prodhimin e limfociteve T ose B. Këto të stimuluara shumëzohen, kthehen në qeliza efektorë apo kujtesë si dhe qarkullojnë. Nga nyja limfatike ato dalin në enët limfatike eferente dhe derdhen në ductus thoracicus dhe me anë të tij derdhen në vena cava superior. Pastaj kalojnë në qarkullimin arterial, më pas në venulat postkapilare. Aty ka receptorë për këta limfocite që ndodhen në të ashtuquajturin HEV (High Endotelium Venules). Ky qarkullim ka shumë rëndësi se e shpërndan informacionin në të gjithë organizmin (2.7.). N.q.s stimuli ndodh në MALT limfocitet e stimuluar shkojnë në indin limfocitar lokal dhe pastaj kthehen sërish në MALT (p.sh nqs stimulohen në MALT të zorrës ata mund të lokalizohen më tej në indin limfoid të gjëndrës së qumështit dhe nga këtu t’i transmetohet një imunitet pasiv fëmijes). Ky fenomen i rikthimit quhet fenomeni i HOMING (gjetja e shtëpisë).

20
KAPITULLI 3. NJOHURI MBI ANTIGJENËT
C'janë antigjenët dhe haptenët, antigjeniciteti, imunogjeniciteti: natyra e antigjenëve, llojet e tyre. Cilësite fiziko-kimike të tyre, adjuvantët. Rrugët e futjes së antigenit në organizëm. Përcaktimi i antigjenëve nga pikëpamja e sistemit imunitar.
Elementi bazë nxitës i sistemit imunitar është antigjeni. Antigjen quajmë cdo substancë kur ajo shkakton krijimin e antitrupave apo limfociteve T specifikë për të. Antigjeni eshtë elementi fillestar i cili kur futet në organizëm provokon në të një reaksion mbrojtës që ka të bëjë me qelizat imunitare, (ku rolin kryesor e luajnë limfocitet) apo me klasen e proteinave plazmatike ku rolin kryesor e luajnë imunoglobulinat.
Antigjen do të quhet çdo lëndë e huaj që duke u futur në organizëm shkakton një përgjigje imunitare qelizore apo humorale dhe njëkohësisht mund të lidhet me antitrupat specifike apo qelizat të nxitura si rezultat i futjes së tij. Antigjenë të ndryshëm duke u futur në të njëjtat sasi e kushte në organizëm japin përgjigje imunitare të ndryshme. Kjo varet nga imunogjeniciteti i antigjenit. Me këtë kuptojmë cilësinë që ka antigjeni i futur për të bashkëvepruar me organizmin. Imunogjeniciteti ndryshon dhe në vartësi të sasisë së antigjenit që depërton në organizëm. Për çdo lloj antigjeni ekziston një interval sasior i caktuar (dozë e caktuar) që është i ndryshme për antigjenë të ndryshëm dhe në të cilin interval organizmi reagon me një përgjigje imunitare optimale. N.q.s sasia e antigjenit të futur është shumë e vogël apo më e madhe se kjo dozë optimale atëhere përgjigja imunitare nuk shkaktohet.
Tjetër faktor që ndikon në imunogjenicitet është afërsia gjenetike e lëndëve të futura (antigenëve) me indet e vetë organizmit ku futet. Sa më e huaj të jetë një lëndë për organizmin aq më i madh është imunogeniciteti i këtij antigeni.
Rol në imunogjenicitet luan dhe pesha molekulare. Kjo duhet marrë në konsideratë sidomos kur kemi antigjenë proteinikë. Proteinat me peshë më të ulët se 5000-10000 dalton nuk kanë imunogjenicitet. Megjithatë ka dhe përjashtime, p.sh angiotensina ndonëse me peshë 1030 dalton ka imunogjenicitet. Sa më e madhe të jetë pesha molekulare e antigjenit të futur aq më i lartë është dhe imunogeniciteti i tij. Koncepti imunogenicitet dallohet nga antigeniciteti që është cilësia që ka antigeni për t'u lidhur me antitrupa specifikë për të pas futjes së një antigeni në organizëm. P.sh, antigenët e quajtur jo të plotë apo haptenët s'kanë cilësi imunogenike, por kanë antigenicitet.
Ajo pjesë e antigenit që lidhet me antitrupin specifik quhet përcaktues antigenik ose epitop. Pjeset e regjioneve hipervariabel te molekules antitrupore qe lidhet me epitopin quhet ndryshe dhe paratop. Cdo molekule antigjenike permban zakonisht shume epitope ne siperfaqen e saj qe jane te ndryshem dhe nga ana strukturore. Keshtu psh. nje antitrup monoklonal qe bashkevepron me nje percaktues antigjenik zakonisht nuk bashkevepron me epitope te tjere te ndryshem ne te njejten molekule (Fig. 3.1). Nga puikpamja e madhesise, nqs antigjeni eshte nje peptid linear ose karbohidrat epitopi mund te perbehet nga 5-6 aminiacide ose njesi glukoze. Ne proteinat globulare rreth 16 aminoacide mund te bijne ne kontakt me paratopin.

21
Kur në një organizëm futim njëkohësisht 2 antigenë të ndryshëm përgjigja imunitare për secilin prej tyre është më e fuqishme sesa po t'i futim veç e veç. Kjo cilësi përdoret gjatë përgatitjes së polivaksinave. Megjithatë që të realizohet një përgjigje e tillë duhet që të dy antigjenet të futen në sasi të njëjtë dhe të kenë imunogenicitet të njëjtë sepse ndryshe ndodh që njëri të shuajë përgjigjen imunitare të antigjenit tjetër. Po qe se në një ditë të caktuar kemi injektuar një antigen të fuqishëm dhe pas 5-6 ditësh injektojmë një antigen me imunogenicitet më të dobët mund të ndodhë që ndaj të dytit të mos kemi përgjigje imunitare dhe të forcohet përgjigja ndaj antigenit të futur herën e parë. N.q.s antigenë të ndryshëm kanë në përbërje të tyre struktura të ngjashme atëhere antitrupat e prodhuara ndaj një antigeni mund të japin një reaksion pak a shumë të shprehur edhe ndaj antigjenëve të tjerë. Bëhet fjalë në këtë rast për reaksione të kryqëzuara.
Antigjenët i ndajmë në të plotë (1) dhe jo të plotë (2). 1. Kur futet në organizëm antigjeni i plotë e realizon përgjigjen imunitare. 2. Antigjeni jo i
plotë që të realizojë një përgjigje duhet të lidhet me një proteine bartëse. Duke u nisur nga përkufizimi i imunogjenicitetit dhe antigjenicitetit mund të themi se antigeni jo i plotë (hapteni) nuk ka imunogjenicitet por ka antigjenicitet.
Në bazë të afërsisë gjenetike me individin Antigenet ndahen në autoantigjene (1), aloantigjene (2), izoantigjene (3), ksenoantigjene (4):
1. Autoantigjenët janë specifikë për individin. Sëmundje ku perfshihen autoantigjenet quhen sëmundjet autoimune. Për arsye të ndryshme inde të veçantë nuk njihen nga sistemi imunitar, pra sillen si antigjene të huaj dhe ndaj tyre prodhohen (auto)antitrupa të cilat bashkeveprojne me to dhe krijojnë komplekse imune që janë shkaktarë të këtyre patologjive.
2. Aloantigjenët jane antigjene qe i gjejmë në individë te ndryshem të të njëjtës specie, p.sh faktori Rhezus, grupet ABO të gjakut apo antigenët e KMPI (kompleksi madhor i pajtueshmërisë indore).
3. Izoantigjenët janë antigjenë që përmbahen në dy individë të ndryshëm të të njëjtës specie të prodhuar nga e njëjta vezë.
4. Ksenoantigjenët jane antigjenë qe i përkasin specieve të ndryshme. P.sh serumet imune të përgatitura në kafshë. Ato prodhohen kur bejmë transplantin e kornese tek njerezit dhe atij i vihet kornea e demit psh..
Cila është natyra kimike e Ag? Grupin më të madhe të antigenëve e përbëjnë proteinat. Pavarësisht nga struktura e proteinës ajo ka cilësi antigenike. Edhe polisakaridet kanë cilësi antigenike sidomos këto cilësi të tyre përforcohen kur ato janë në trajtë glikoproteinash (si polimere). Polimeret e glukozës përbëjnë mbështjellen e viruseve dhe shkaktojnë patologji të shumta. Cilësi antigenike kanë edhe lipidet vetëm kur janë me proteina si p.sh kardiolipina. Veç këtyre si antigenë mund të sillen edhe lëndë sintetike ose molekula të ndryshme si dinitroklorbenzeni, acidi benzensulfonik etj.
Rrugët e futjes së Ag në organizëm: 1. Me rrugë orale, p.sh kolera, salmonela, virusi i poliomielitit, hepatitit A 2. Me rrugë kutane si piodermitet, stafilokoku 3. Me rrugë gjaku si viruset e hepatitit viral B e C, HIV etj
Për të përforcuar aktivitetin imunogjenik të antigjeneve në procesin e imunizimit përdoren lëndë që quhen adjuvantë. Cilësia e tyre është të provokojnë një reaksion inflamator dhe të ngadalësojnë eliminimin e antigjenit. Reaksioni inflamator favorizon kontaktin e antigjenit me qelizat imunitare, kurse eliminimi i ngadalësuar i antigjenit zgjat kontaktin e tij me qelizat imunitare. Të dy këto faktorë përforcojnë imunogjenicitetin. Adjuvantët që përdoren për injektimin e antigjenit me rrugë kutane përbëhen nga yndyrna të ndryshme si p.sh adjuvanti i Freund-it. Ky përbëhet nga lanolina, vazelina dhe BCG (vaksina e TBC) dhe përdoret në kafshë ndersa te njërëzit përdoren multivaksinat.
Klasifikimi i antigenëve nga pikpamja e sistemit imunitar: Nga pikpamja e sistemit imunitar antigenët mund t'i ndajmë në antigjenë ekzogjene dhe në
antigjenë endogjenë.

22
Antigjene ekzogjenë quhen ato antigjenë që bëjnë pjesë në ato mikroorganizma që vijnë nga jashtë dhe shumëzohen në organizëm në mënyrë të pavarur nga aparati biologjik i qelizave të organizmit. Antigjenet ekzogjene i ndajmë në jashtëqelizore dhe brëndaqelizore. Ata jashtëqelizore shumëzohen jashtë qelizave të organizmit (shëmbull: koket piogjene si streptokoku, pneumokoku, stafilokoku, hemophilus influenzae etj). Mekanizmi mbrojtës ndaj tyre realizohet kryesisht me anë të imunitetit humoral: antitrupat dhe komplementi. Ata intraqelizorë shumëzohen brënda qelizave fagocitare. Shëmbuj: mykobakteriet, kandida, aspergilus, pneumocistis, toxoplazma. Mekanizmi mbrojtës: Limfocitet T helper që aktivizojnë makrofagët të cilët janë qelizat efektorë kryesore.
Antigjenët endogjenë janë ata që përdorin aparatin biologjik të qelizës ku depërtojnë për t'u shumëzuar dhe këtu përfshihen viruset por këtu mund të përfshijmë dhe antigjenët tumoralë. Mekanizmat mbrojtës janë me anë të limfocitëve T citotoksikë, qelizave NK, interferonet.
KAPITULLI 4. KARAKTERI KLONAL I PËRGJIGJES IMUNE
Qelizat bartëse të imunitetit specifik për antigenin jane limfocitet T dhe B. Këto qeliza kanë
receptorë specifike në sipërfaqen e tyre për një antigen të caktuar (përcaktues antigenik), të cilët janë të pranishëm para cdo kontakti me këto antigenë. Futja e një antigeni të huaj për organizmin shkakton ngacmimin e receptorit në sipërfaqen e limfocitit për të cilin ai është specifik dhe si rrjedhojë kemi stimulimin e këtij limfociti.
Cdo limfocit ka vetëm një lloj receptori antigenik specifik për një përcaktues antigenik të caktuar. Stimulimi nga ky përcaktues antigenik shkakton formimin e një kloni limfocitar, e cila përbën një popullatë homogjene e lindur vetëm nga një qelizë mëmë dhe anetarët e së cilës janë identikë me njëri tjetrin, pra kanë dhe të njëjtin receptor për antigenin. Meqenëse shumica e antigenëve që gjënden në natyrë janë kompleksë dhe përbëhen nga shumë përcaktues antigenikë të ndryshëm nga njëri tjetri, përgjigja imunitare ndaj tyre ështe zakonisht poliklonale, pra stimulohen njëkohësisht disa klone limfocitare. Përgjigje imunitare monoklonale kemi zakonisht në 2 situata: 1) Patollogjike, si në rastin e mielomave kur kemi një shumëzim anormal të vetëm një kloni limfocitar 2) Ekspërimentale, si në rastin e prodhimit të antitrupave monoklonalë në laborator me metodën e hibridomave.
Limfocitet B përdorin Imunoglobulinat membranore si receptorë për antigenin: Në një limfocit B që nuk ka rënë në kontakt me antigenin imunoglobulinat membranore janë
të klasës IgM (monomere) dhe IgD. Pas kontaktit me antigenin dhe pas aktivizimit të limfocitit B mund të hasim dhe imunoglobulina të klasës IgG, IgA dhe IgE. Këto imunoglobulina megjithëse mund të ndryshojnë pjesën Fc të tyre mbajnë gjithmonë po të njëjtën pjesë variabël si ato që kishte limfociti i virgjër.
Karakteristikat funksionale të Ig membranore (sipërfaqësore: Igs) : 1. Igs në sipërfaqen e limfocitit B njohin antigenë ekzogjene (të cilët përdorin aparatin
biologjik të tyre për t'u riprodhuar) me lokalizim ekstraqelizor. Këta antigenë janë ata që bëjnë pjësë në mikroorganizma të cilët ciklin jetësor të tyre e realizojnë jashtë qelizave të organizmit (fagociteve e të tjera qeliza). Mbrojtja ndaj tyre realizohet kryesisht me anë të antitrupave dhe të komplementit. Shembuj tipik janë koket piogjene.
2. Igs ka aftësinë të njohë përcaktuesin antigenik (PA) specifik për të në gjëndje native (jo të denatyruar) si dhe në gjëndje të tretshme.

23
3. Igs s'ka nevojë për një qelizë antigen paraqitëse (QAP) apo një molekulë të kompleksit madhor të pajtushmërisë indore (KMPI) për të njohur antigenin.
Limfocitet T përdorin TCR (T-cell receptor) si receptorë sipërfaqësore për antigenin. TCR është një dimer i përbërë nga dy vargje polipeptidike me peshë molekulare 40-50 kilodalton që janë të lidhur me lidhje disulfurore me njëra tjetrën. Cdo varg është i përbërë nga 2 pjesë që kanë homologji me domenet e imunoglobulinave. Domeni proksimal me membranën qelizore është pjesa konstante e vargut (e cila nuk ndryshon nga një klon limfocitar në tjetrin) ndërsa 1/2 distale e vargut përbën pjesën variabël të tij e cila ndryshon nga një limfocit T në tjetrin. Bashkësia e 2 pjesëve të ndryshueshme e të dy vargjeve të TCR përcakton specificitetin antigenik të TCR. Ekzistojnë 4 izotipe vargjesh TCR të cilët përcaktohen nga pjesa konstante e tyre: alfa, beta, gama, lambda. Ato alfa lidhen gjithmonë me vargjet beta dhe përbëjnë receptorët TCR2 ndërsa vargjet gama dhe delta përbëjnë tipin tjetër TCR1. Ky fundit haset në 0,5 - 15% të limfocitëve T në gjakun përiferik të organizmit të rritur, ndërsa në ontogjenezë cfaqet me herët se TCR2. TCR1 ka një variabilitet më të ulët se TCR2 dhe është vëne re se ai është më i shprehur në limfocitët T intraepiteliale të mukozave dhe në lëkurë. Duket se TCR2 luan rol në njohjen e antigenëve mykobakterialë dhe vecanërisht të proteinës të shokut nga nxehtësia (hsp 65). Secili prej dy dimereve është i koduar në një kromozom të vecantë. Pas lidhjes me antigenin nëpërmjet pjesës variabël të TCR sinjali përcohet në brëndësi të limfocitit T me ndërmjetësinë e shënjuesit CD3 i cili përbëhet nga 5 vargje polipeptidike.
Funksioni biologjik i TCR është njohja nga ana e limfociteve T i antigenëve endogjene (viruse apo antigenë tumorale që sintetizohen nga vetë aparati biologjik i qelizës së organizmit) si dhe të antigenëve ekzogjene por me lokalizim brëndaqelizor (qe kanë aftësinë të shumëzohen në brëndësine e qelizave psh makrofagëve sic janë mykobakteriet, myket si Candida albicans, parazitë si Pneumocystis carinii, Toxoplasma Gondii etj.).
Karakteristikat funksionale të TCR: 1. Përcaktuesit antigenikë përbërës të antigenëve të ndryshëm nuk njihen në gjëndje native
por të denatyruar, pra të përpunuar nga qelizat antigen paraqitëse (QAP). Të tillë janë makrofagët, qelizat dendritike interdigitale të nyjeve limfatike, limfocitet B. QAP kanë aftësinë që molekulën antigenike pasi e fusin në citoplazmën e tyre, e përpunojnë dhe e paraqisin në sipërfaqen e tyre së bashku me molekulat e klasës II të KMPI ku njihen nga TCR e limfocive T ndihmëse. Molekulat e klasës II të KMPI kanë aftësinë të lidhin peptidet e denatyruara të antigenit që kanë një madhësi 5-20 aminoacide. TCR njeh peptidin+pjesën e ndryshueshme të molekulës Kl II të KMPI ndërsa molekula CD4 ka afinitet për pjesën jo të ndryshueshme të molekulës së klasës II të KMPI. Pasojë e kësaj njohje është aktivizimi i limfocitit T ndihmës dhe prodhimi me shumicë i interleukinavë nga ky i fundit. 2. Antigenët endogjene nuk paraqiten vetëm nga qeliza të specializuara si ato ekzogjene, por potencialisht nga cdo qelizë e organizmit. Përcaktuesit antigenikë të denatyruar asociohen në këto qeliza me molekulat e klasës I të KMPI dhe të tilla paraqiten në sipërfaqen e këtyre qelizave ku njihen nga limfocitet T citotoksikë CD8 (+). Molekula CD8 njeh në këtë rast pjesën e pandryshueshme të molekulës KMPI kl I ndërsa TCR njeh peptidin e huaj+molekulën kl I KMPI. Pasojë e kësaj njohjeje ështe vrasja e qelizës shenjë (që paraqiste antigenin e huaj).
Mekanizmat e gjenerimit të shumëllojshmërisë së receptorëve për antigenin në limfocitet T dhe B.
Sikur cdo receptor për antigenin të kodohej nga një gen i vetëm në ADN, atëhere, duke patur parasysh shumëllojshmërinë e madhe të antigenëve në natyre nuk do të mjaftonte ADN e gjithë genomës për të koduar këta receptorë. Për këtë arësye natyra ka zhvilluar një mekanizëm të tillë gjenerimi të shumëllojshmërisë së receptorëve për antigenin në mënyrë që duke u nisur nga disa qindra gene të krijohet një numër prej miliarda receptorësh si në limfocitet B ashtu dhe në ato T. Mekanizmat janë të ngjashme si për vargjet e lehta dhe të rënda të imunoglobulinave ashtu dhe për vargjet e TCR.
Nqs shohim si kodohen vargjet e rënda ose të lehta të imunoglobulinve në ADN germinale (ADN germinale quhet genoma e qelizës së padiferencuar, ku ADN s'ka pësuar ende asnjë rirregullim) atëhere vihet re se cdo varg i lehtë kodohet në një kromozom të vecantë ndërsa vargjet e

24
rënde kodohen në një kromozom tjetër. Një vecori tjetër e kodimit gjenetik të cdo vargu receptori për antigenin është se pjesa variabël e vargut kodohet nga kombinimi i rastësishëm me njëri tjetrin i 2 ose 3 ekzonëve secili nga të cilët bën pjesë në një familje ekzonësh anëtarët e së cilës ndryshojnë nga njëri tjetri dhe janë të ndarë nga introne nga njëri tjetri. Cdo familje është ndarë nga një intron më i gjatë nga familja tjetër e ekzonëve variabël.
Ëkzoni i parë 5' është ai L (leader) i cili kodon për një pëptid, që luan rol në transportimin e vargut nëpër qelizë dhe që shkëputet kur vargu del nga qeliza në sipërfaqe të saj ose sekretohet. Familja V eshtë ajo më e rëndësishmja dhe përbëhet nga disa qindra kopje ekzonesh të cilët kodojnë pjesën më të madhe të pjesës variabël. Familja J (junction) përbëhet nga më pak ekzone (rreth 5) i cili shton variabilitetin në pjesën fundore të pjesës variabël. Vargjet e rënda të Ig si dhe vargjet e TCR kanë dhe një familje të tretë ekzonësh variabël që janë ekzonët D(diversity) që ndodhen në 2-3 kopje dhe që janë të vendosur midis ekzonëve V dhe J duke kontribuar kështu në rritjen e shumëllojshmërisë.
Ekzoni që kodon për pjesën konstante të vargut (i cili ndodhet vetëm në një kopje në genomë pra nuk ka shumëllojshmëri idiotipike) ndodhet mjaft me 3', i ndarë nga një intron mjaft i madh nga ekzonët e pjesës variabël.
Kur një limfocit maturohet në timus ose në palcën kockore në një limfocit T ose B të maturuar atëhere mikromjedisi i këtyre indeve dërgon sinjale në qelizë që veprojnë duke rirregulluar genomën e limfocitit. Nëpërmjet aktivizimit të enzimave të ndryshme që veprojnë në ADN kemi kombinimin e ekzonit L me një nga ekzonët V dhe J (ose dhe D për ato vargje që thamë më sipër) ndërsa këputet pjesa e ADN të ndërmjetme. Në këtë mënyrë formohet geni kodues VJ ose VDJ për pjesën variabël të vargut. Ky gen bashkohët më pas në ARN mesagjere me gjenin kodues të pjesës konstante pasi këputet i gjithë introni që ndodhej midis VDJ dhe C. Kjo ARN mesagjere shërben për formimin e vargut polipeptidik. Të dy vargjet që kodojnë për receptorin bashkohën pastaj në sipërfaqen e limfocitit duke krijuar receptorin e plotë membranor për antigenin (Igm ose TCR).
L V1 Vn D1 Dn J1 Jn C
ADN germinale
L V D J
ADN e rirregulluar
C
L V D J C
ARN mesagjere
5' 3'
5' 3'
5' 3'
Nqs për vargun alfa të TCR kemi VxJ kombinime atëhere 200(numri i përafërt i ekzonëve V)x 4 (numri i përafërt i ekzonëve J)=kemi 800 pjesë të ndryshme të ndryshueshme.
Për vargun beta kombinimet jane VxDxJ. Atëhere 200x4x6=5000 jane kombinimet e ndryshme për pjesët variabël të vargjeve beta. Duke patur parasysh se vargjet alfa kombinohen me ato beta në formimin e TCR2 atëhere numri i mundshëm i pjesëve variabël të TCR2 është 800x5000=4x106
Përsa i përket imunoglobulinave shumëllojshmëria e tyre shtohet nga bashkimi i vargjeve të lehta me ato të rënda.
Për vargjet e rënda H kemi kombinimet VxDxJ= 200x2x5=2000 kombinime x 10 (variabiliteti i kombinimeve V me D që s'përputhen asnjëhere cep me cep por mbivendosen lehtësisht)=20000 Vargjet e lehta L formohen nga kombinimi VxJ= 200x5=1000x10 ( variabiliteti i kombinimeve V me J që s'përputhen asnjëherë cep me cep por mbivendosen lehtësisht)= 1000

25
Nga kombinimi i H me L do të kemi pra 2000 x 10000= 2x107 kombinime. Duke patur parasysh se në formimim e Ig kemi dhe ndryshime somatike të nukleotideve (që nuk ndodh me vargjet TCR) shumëllojshmëria e pjesëve variabël rritet me një faktor tjetër 10 pra kemi rreth 2x108 pjesë të ndryshueshme imunoglobulinash që kanë aftësine të lidhin të njëjtin numër përcaktuesish antigenike.
TCR1 TCR2 Ig gama delta
alfa beta H L
kapa lambda ekz. V 8 3 50 57 100 70 25 ekz. D - 3 - 1,1 4 - - ekz. J 3,2 3 70 6,7 6 5 8 Combin. VxJ VxDxJ VxJ VxDxJ VxDxJ VxJ VxJ 8x5 3x3x3 50x70 57(13+7) 100x4x6 70x5 25x8 Total 40 x 27 3500 x 1140 2400 x 350 2400 x 200 103 4 x 106 9 x 106 5 x 106 Mekanizma të tjerë: diversiteti i lidhjeve V-D-Jetj.
106 4 x 109 9 x 109 5 x 109
Mutacionet somatike
- - +++ +++

26
KAPITULLI 5. QELIZAT E IMUNITETIT SPECIFIK
LIMFOCITET. Limfocitet bartin funksionin e imunitetit të fituar. Ky imunitet ndryshon nga imuniteti i
lindur n ë 2 aspekte kryesore: 1. Imuniteti i fituar është specifik për një antigen të caktuar ndërsa imuniteti i lindur nuk
është i tillë. 2. Zhvillimi i imunitetit të fituar ndaj një antigeni të caktuar kërkon një kontakt paraprak me
këtë antigen. Kontakti i antigenit realizohet me qelizat e imunitetit të fituar, pra limfocitet. Karakteristikë
kryesore e këtyre qelizave është prania në sipërfaqen e tyre e receptorëve specifikë për antigenin. Këta receptorë janë të shpërndarë në mënyrë klonale d.m.th. cdo limfocit i diferencuar dhe i pjekur por që s'ka rënë akoma në kontakt me antigenin bart në membranën e tij vetëm një lloj receptori specifik për një përcaktues antigenik të caktuar. Rënja në kontakt e këtij limfociti me përcaktuesin antigenik specifik për të sjell stimulimin dhe shumëzimin e këtij limfociti dhe pra krijimin e një "kloni limfocitar" përbërësit e së cilës janë identikë midis tyre pasi kanë rrjedhur nga limfociti i stimuluar fillimisht.
Limfocitet në mikroskopi optike nuk kanë dallime midis tyre. Dallimi midis tyre është në nivelin molekular dhe kryesisht nga prania në sipërfaqen e tyre të molekulave në formën e glikoproteinave. LIMFOCITET T B % në gjakun përiferik 65 - 80 8 - 15 Ig sipërfaqesore - ++ TCR (CD3) ++ - CD2 ++ - recptore për Fc gama +- ++ receptorë për C3b (CR1,CR2) +- ++ receptorë për virusin e fruthit ++ - receptorë EBV (=CR2) - ++ shënues të tjerë CD 4/5/8/45 CD 5/19/23/40/45 KMPI KL I ++ ++ KMPI KLII (+ në aktivizim) ++

27
AKTIVIZIM POLIKLONAL
anti- CD3 fitohemaglutinina (PHA) pokeweed mitogen (PWM) Concanavalin A supërantigen (p.sh. enterotoksina)
anti- Ig Staph. A(Cowan 1) Pokeweed Mitogen (PWM) EBV
KRAHASIMI I QELIZAVE B ME ATO T:
Ato molekula të cilat karakterizojnë një popullatë ose nënpopullatë limfocitare dhe e dallojnë
atë nga një tjetër quhen shënjues. Shumica e këtyre shënjuesve vihen sot në dukje me anë të antitrupave monoklonale. Antitrupa monoklonale quhen ato antitrupa homogjene (molekula identike me njëra tjetren) që kanë një specificitet të ngushtë, vetëm për një përcaktues antigenik të caktuar i cili mund të jetë një shënjues limfocitar.
Limfocitet T, aktivizimi i tyre: Ka një madhësi prej 8-15 mikron. Bërthama, e rrumbullakët, ze pothuajse të gjithë
citoplazmën, e cila është e varfër në organela citoplazmike. Ndër shënjuesit e tyre kryesorë, që përdoren për numërimin e tyre janë CD (cluster of differentiation) 2 dhe CD3. Limfocitet T zenë rreth 65-80% të te gjithë limfociteve të gjakut përiferik. Limfocitet T i ndajmë në 3 nënpopullata kryesore që dallojnë nga njëra tjetra si nga shënjuesit ashtu dhe funksionet. Popullata më e shumta në numër është ajo e limfociteve T helper (ndihmës). Shënjuesi karakteristik i tyre është molekula CD4. Funksioni i tyre kryesor është që pas kontaktit me antigenin dhe nëpëmjet sekretimit të limfokinave të stimulojnë qelizat e tjera të sistemit imunitar sic janë limfocitet B, limfocitet T supresorë, ata citotoksikë apo dhe makrofagët. Ata luajnë rolin me të rëndësishëm në fenomenet e hipërsensibilitetit të vonuar (tipi i katërt i mbindjeshmërisë).
Limfocitet T citotoksikë (vrasëse qelizore) kanë shënuesin CD8. Funksoni u tyre është vrasja e qelizave të huaja për organizmin, të cilat zakonisht janë qeliza të vetë organizmit të infektuara me një virus ose një qelizë tumorale (e transformuar). Këtë funksion ata e realizojnë me anë të kontaktit membranë-membranë me qelizën shënjë (target) dhe me anë të kalimit të disa substancave vrasëse (citolizina) nga qeliza citotoksike në qelizën shënjë.
Limfocitet T supresorë (frenuese) efektorë kanë edhe ato shënjuesin CD8. Funksioni i tyre është të frenojnë reaksionet imune, pra të mos lejojnë përpjestime të ekzagjeruara të tyre të rrezikshme për organizmin.
Një grup i rëndësishëm limfocitesh T janë limfocitet T kujtesë që kanë rënë një herë në kontakt me antigenin dhe kanë një jetëgjatësi të madhe që shkon nga muaj në mjaft vite. Ata qarkullojnë vazhdimisht në organizëm nga sistemi limfoid dytësor në qarkullimin e gjakut dhe anasjelltas. Ata sillen si qeliza roje qarkulluese të organizmit që bartin kujtesën imunitare ndaj një antigeni të caktuar. Futja e këtij antigeni në një moment të dytë sjell mobilizimin e shpejtë të limfociteve kujtesë dhe realizimin në një nivel mjaft më të amplifikuar të përgjigjes imunitare specifike në krahasim me kontaktin e parë që gjeneroi këto qeliza kujtesë.
Limfocitet T maturohen në timus dhe kjo ndodh gjatë kalimit të tyre nga korteksi në pjesën medulare të tij. Gjatë maturimit ata fitojnë dhe shënjuesit membranorë kryesorë të tyre. Në fillim fitohet CD2 pastaj CD3 dhe njëkohësisht me këtë dhe receptorët specifike për antigenin të limfocitet T (TCR). Me pas fitohen njëkohësisht CD4 dhe CD8. Kur maturohet plotësisht limfociti T humb ose CD4 ose CD8 dhe mbetet një limfocit T i pjekur (CD2+;CD3+;TCR+; CD4+ ose CD8+). Shumica e limfociteve T kanë TCR2 (alfa, beta), pakica TCR1 (gama, delta). Këta të fundit janë CD4 dhe CD8 (-). Në timus ndodhin 2 lloj seleksionesh: seleksioni pozitiv gjatë të cilit eliminohen ato LT që nuk kanë afinitet për KMPI të vetvehtes dhe seleksioni negativ: eliminohen ato LT që njohin KMPI e vehtes + autoAntigenët (LT autoreaktive). TCR përcakton specificitetin e limfocitit T për antigenin dhe është një dimer i përbërë nga 2 vargje polipeptidike.

28
L
s-sC
V
CD3LIMFOCITI T
TCR ka këto vecori në njohjen e antigenit: 1. TCR e njeh antigenin vetëm kur ky i paraqitet nga një qelizë antigen paraqitëse (QAP) e
cila pasi e ka përpunuar antigenin e huaj e paraqet në formën e një polipeptidi. QAP kryesore janë a) makrofagët b) limfocitet B c)qelizat dendritike interdigitale që ndodhen në zonën parakortikale të nyjeve limfatike.
2. TCR e njeh antigenin vetëm kur ky paraqitet bashkë me një molekulë të kompleksit madhor të pajtueshmerise indore (KMPI) të vetë individit (njohje e të huajes -"nonself"- vetëm kur kjo shoqërohet nga një molekulë e vetes-" self-").
3. Antigenin e njeh të denatyruar, në formën e polipeptideve lineare me 10-20 aminoacide. Aktivizimi i limfociteve T. Stimulimi dhe aktivizimi i limfociteve T kërkon përvec njohjes se antigenit dhe ndërhyrjen e
një sërë molekulash dhe faktorësh të tjerë. Kompleksi CD3 është i nevojshëm për të përcuar sinjalin nga TCR në brëndësi të limfocitit T. Molekula CD4 bashkëvepron me pjesën konstante të molekulës KMPI Kl II, ndërsa ajo CD8 me pjesën konstante të KMPI Kl I. Molekula CD2 bashkëvepron me molekulën LFA-3 që gjëndet në sipërfaqen e QAP ndërsa LFA-1 e limfocitit T me ICAM-1 në sipërfaqen e QAP. Edhe molekula CD45 shërbën si kapëse e limfocitit T me QAP. Por dhe këto bashkëveprime molekulare s'jane të mjaftueshme për të nxitur stimulimin e limfocitit T. Sinjali tjetër i nevojshëm është prodhimi i interleukinës 1 nga QAP e cila bashkëvepron me stimulin antigenik për stimulimin e limfocitit T.
Një sinjal tjetër i nevojshëmi vënë rekohëte fundit për aktivizimin e limfociteve T në qetësi është bashkëveprimi midis molekulës B7 në qelizën antigen paraqitëse me molekulën CD 28 në limfocitin T ndihmës ose me molekulën ekuivalente CD 28 në molekulën T citotoksike.
Stimulimi dhe aktivizimi i limfocitit T sjell shumimin e tij dhe formimin e klonit limfocitar si dhe sekretimin e një sërë interleukinash si Il 2,3,4,5,6, Interferoni gama, TNF(tumor necrosis factor), faktori GM-CSF. Nëpërmjet këtyre mediatorëve si dhe mjaft të tjerëve limfocitet T të aktivizuara kryejnr funksionet e tyre efektorë sic janë stimulimi i limfociteve B, citotoksikë, supresorë apo fenomenet e hipërsensibilitetit të vonuar.
Limfociti
TQAP
antigjeni
KMPI kl I/II
LFA-3
ICAM-1LFA-1
CD3
TCR
CD-2CD4/8
Molekulat bashkevepruese te limfocitit T me qelizen antigen paraqitese

29
Limfocitet B. aktivizimi i tyre:. Është një popullatë tjetër limfocitare që ze 8-15% të limfociteve të gjakut përiferik. Quhen
kështu pasi maturohen në palcën kockore (angl: bone marrow). Këtu ato nën influencën e mikromjedisit transformohen në fillim në qeliza pre-B (qe kanë vetëm varg të rëndë të Ig në citoplazëm), pastaj në qeliza B të papjekura dhe së fundi në L.B i pjekur.
Funksioni kryesor i limfociteve B është prodhimi i imunoglobulinave (antitrupave) pas stimulimit nga antigeni dhe transformimi në plazmocite. Shënjuesi kryesor i tyre janë imunoglobulinat membranore (ose sipërfaqesore) Igs që shërbejnë si receptorë për antigenin në limfocitet B. në limfocitet B që s'kane rënë akoma në kontakt me antigenin Igs janë të klasës IgM(monomer) dhe IgD. në limfocitet B që kanë rënë në kontakt me antigenin gjënden dhe Igs të klasave të tjera (IgG, IgA, IgE). në një klon limfocitesh B pjesa variabël e Igs është gjithmone e njëjta ndërsa ndryshimet e pjesës konstante përcaktojnë klasat e ndryshme të Igs.
Igs kanë këto karakteristika kryesore: 1. Igs e njeh antigenin në formë native (jo të denatyruar) si dhe jo detyrimisht (si TCR) në
sipërfaqen e një QAP, por edhe në gjëndje të tretshme. 2. Antigeni njihet pa qënë e nevojshme të paraqitet nga një QAP. 3. S'është e nevojshme paraqitja e antigenit të shoqëruar nga një molekulë e KMPI. Pra, limfocitet B në një molekulë antigenike njohin përcaktues të tjerë antigenike nga ata që
njohin limfocitet T. Përcaktuesit antigenike që njihen nga limfocitet B quhen haptenike ndërsa ata që njihen nga limfocitet T quhen bartës (carrier).
Antigenët që stimulojnë limfocitet B i ndajmë në 3 lloje kryesore: 1. Antigenë timoindipendentë të tipit të parë: Kryesisht këta janë lipopolisakaride (LPS) me origjinë bakteriale që stimulojnë limfocitet B në formë poliklonale, pra jo vetëm një klon të caktuar limfocitesh B, por shumë prej tyre. Këta antigenë nuk ndërveprojnë me pjesën e ndryshueshme të Igs si Antigenët e tjerë por por me molekula të tjera në sipërfaqen e limfociteve B që janë të përbashkëta për të gjitha këto qeliza.
2. Antigenë timoindipendentë të tipit të dytë: Këta janë zakonisht molekula të gjata polisakaridesh me struktura të përsëritura (monotone) të cilat arrijnë të lidhin me njëra tjetren Igs specifike për to në sipërfaqen e limfociteve B (duke krijuar një ure midis tyre) dhe duke i stimuluar këta direkt pa ndihmën e limfociteve T. Këta lloj antigenësh kanë vecori të stimulojnë një prodhim antitrupash me predominim të imunoglobulinave të klasës IgM. Gjithashtu përgjigja antitrupore ndaj tyre s'ka kujtesë imunitare si ajo ndaj antigenëve timovartës.
Antigenët timoindipendente të klasës 2 nuk përpunohen dot nga QAP sic ndodh me Antigenët timovartës. Vetëm disa makrofagë specialë që gjënden në zonën subkapsulare të nyjeve limfatike si dhe në zonën marginale të shpretkes arrijnë t'ua paraqesin këta antigenë limfociteve B.
3. Antigenët timovartës: përbëjne pjesën me të madhe të Antigenëve natyrore dhe kryesisht janë Antigenë proteinike. Quhen të tillë se ndihma e limfociteve T është e domosdoshme për të realizuar përgjigjen antitrupore nga ana e limfociteve B ndaj këtyre antigenëve.
Limfociti B njeh përcaktuesin haptenik në antigenin timovartës, por kjo njohje s'është e mjaftueshme për të stimuluar limfocitin B. Ky i fundit shërben në këtë rast si QAP, duke e përpunuar antigenin dhe duke paraqitur përcaktuesit antigenike "bartës" të tij, bashkë me molekulat e klasës se II të KMPI limfociteve T ndihmës CD4(+). Ky stimul i nevojitet limfociteve T (bashkë me Il 1 të prodhuar nga QAP) për të prodhuar interleukinat e nevojshme (Il 2; 3; 4; 5; 6), të cilat në distancë të afërt shërbejnë si stimuli i dytë (bashkë me përcaktuesit antigenike haptenike të antigenit në fjalë) për diferencimin e mëtejshëm të limfocitit B në qelizë prodhuese të antitrupave ndaj këtij antigeni. Në këtë stimulim luan rol ndihmës dhe molekula CD40 në limfocitin B që bashkëvepron me molekulën p 39 në limfocitin T helper të aktivizuar.

30
Limf B
Limf. T QAP (makrofagu)
Percaktues antigjenike
haptenik bartes
TCR
TCR
IL1
IL2;3;4;5;6
Bashkeveprimi i limfocitit T me ate B ne pergjigjen imunitare.
Limfocitet B CD5(+) dhe CD 5(-) paraqisin dy popullata të ndryshme nga njëra tjetra: Nënpopullata e limfociteve B që shpreh edhe shënjuesin e limfociteve T CD5 bashkë me IgM
sipërfaqesore është ajo popullatë që shprehet më shpejt gjatë ontogjenezes dhe që predominon përsa i përket kontributit në formimin e rrjetës idiotipike, në prodhimin e autoantitrupave IgM multispecifike me afinitet të ulët si dhe të ashtuquajturit antitrupat "natyrale" ndaj polisakarideve bakteriale, të cilët cfaqen në përiudhen neonatale pa qënë nevoja e ekspozimit të dukshëm ndaj Antigenëve të zakonshëm. Mendohet se këto limfocite B paraqesin një popullatë krejt të vecantë nga shumica e limfociteve B që janë CD5 (-). në mënyrë konvencionale L.B. CD5(+) janë quajtur B1 dhe ata CD5 (-) B2. Limfocitet B1 shprehin si CD5 shtu dhe ligandin e saj CD72, gje që nxit prodhimin e vazhdueshem të kësaj linje qelizore, e cila kështu ka aftësine të vetëmbahet (pa qënë nevoja për qeliza pararëndëse nga palca kockore). Por për këtë qellim një rol të rëndësishëm luan edhe prodhimi i Il-10 nga këto qeliza. Predispozimi i kësaj linje për autorigjenerim fsheh edhe rrezikun e transformimit leucemik të saj.
Qelizat B1 kanë tendencë t'u përgjigjen Antigenëve timojovartes të tipit të dytë. Në ndryshim nga limfocitet B2 ato B1 nuk bashkëveprojnë me antigenët timovartës si dhe nuk pësojnë ndryshime somatike me rritje të afinitetit për antigenin. Me sa duket funksioni i tyre është gjenerimi i rrjetës idiotipike me rëndësi kjo në self-tolerancë, përgjigja imunitare ndaj antigenëve mikrobiale të ruajtur (gjatë evolucionit) si dhe probabilisht prodhimi i "antitrupave natyrale" që përbëjnë vijën e parë të mbrojtjes pre-ekzistuese me IgM ndaj mikroorganizmave më të zakonshëm.

31
KAPITULLI 6. IMUNOGLOBULINAT, STRUKTURA DHE FUNKSIONI I
TYRE. Imunoglobulinat janë proteina të glukozuara, pra janë glukoproteina që ndodhen në serum dhe lëngje të tjera biologjike të organizmit. Kryejnë rol mbrojtës. Deri sot njihen 5 klasa imunoglobulinash që emertohen imunoglobulina G,A,M,D,E.
Imunoglobulinat e klasave të ndryshme dallohen nga njëra-tjetra nga pesha molekulare, nga ngarkesa elektrike, përbërja e aminoacideve dhe vetitë biologjike të tyre.
Ontogjeneza e Imunoglobulinave: Tek fëmija i porsalindur mund të vërehen vetëm imunoglobulina të klasave M e D mbi sipërfaqen e limfociteve B. Imunoglobulinat G dhe A dhe E nuk gjënden pothuajse fare.
Të gjitha antitrupat janë imunoglobulina. Për këtë arsye sa më shumë rritet kontakti i sistemit imunitar të organizmit me antigenët e jashtëm aq me shumë rritet llojshmëria e antitrupave ndaj këtyre antigenëve.
Fëmija normalizon së pari nivelin e imunoglobulinave M që arrin atë të te rriturit në rreth 9 muaj. Me pas, rreth 2 vjec, normalizohet niveli i imunoglobulinave G. I fundit normalizohet niveli i i imunoglobulinave A: rreth 12-14 vjec.
Imunoglobulina G (IgG) luan rol kryesor në mbrojtjen imunitare. Në muajt e parë pas lindjes kur sistemi imunitar i fëmijes nuk është maturuar plotësisht për të kryer funksionin e vet mbrojtës, këtë rol e kryejnë antitrupat e klasës IgG të nënes që fëmija i merr me rrugë transplacentare. Pas lindjes, niveli i IgG të nënes bie dhe kjo bëhet e dukshme rreth moshes 4 muajsh të fëmijes kur katabolizimi i IgG të nënes është më i shprehur sesa fillimi i sintezës se IgG të fëmijes.
Struktura e Imunoglobulinave: Një monomer imunoglobuline përbëhet nga 2 vargje të rëndë dhe 2 të lehtë. Zbulimi i
strukturës ndihmon në studimin e funksionit të tyre. Sic thame, ka 4 vargje të lidhura me ura disulfure.

32
s s
ss
ss
FcFab
(Fab')2VL CL
VH CH1 CH2 CH3
Struktura e nje molekule imunoglobuline G. Vargjet e lehta janë të lidhur me ato të rëndë dhe të dy të rëndët midis tyre. Struktura
molekulare e një molekulë IgG paraqet strukturën tipike të një monomeri imunoglobuline. Studimi i strukturës është bërë duke e zbërthyer atë me anë të veprimit të fermenteve
proteolitike. Nën veprimin e papaines (qe vepron majtas lidhjeve disulfurore midis vargjeve të rënda) molekula e Ig ndahet në 2 pjesë Fab dhe 1 Fc. Nën veprimin e pepsines (qe vepron djathtas) molekula e Ig ndahet në një pjesë që quhet F(ab`)2 dhe pjesa Fc copëzohet.
Fab përbëhet nga 1/2 e vargut të rëndë dhe një varg i lehtë. Këtoo përbëjne pjesën aminofundore të Ig. Kurse pjesa Fc përbëhet vetëm nga gjysma e vargjeve të rëndë dhe përbën pjesën korboksifundore të Ig. VH quhet pjesa variabël e vargut të rëndë ndërsa VL e vargut të lehtë. Këtu ndodh lidhja Antitrup - Antigen. Specifiteti i një At për një Ag të caktuar përcaktohet nga llojet e aminoacideve në pjesën VH+VL si dhe nga mënyrë e rënditjes se tyre. Në këtë domen VH, në këtë lak ka 3 zona shumë të ndryshueshme të përbëra nga 5-15 aminoacide secila, që përcaktojnë përgjithësisht lidhjen me Ag. specifik. Pjesa Fc përbëhet nga 3 domene pra 3 laqe që emertohen CH1, CH2 e CH3. Këto ndodhen vetëm tek IgG, D, A, kurse për IgM e IgE kemi edhe një lak tjetër CH4. për këtë arsye Fc për IgM dhe E përmban rreth 300 e ca aminoacide. Ndërmjet CH1 dhe CH2 ekziston një pjesë lidhëse që quhet pjesa menteshe me rreth 15 aminoacide. Kjo i jep molekulave të Ig fleksibilitetin e saj prandaj monomeri Ig mund të marrë formën e germes T apo asaj Y.
Vargjet e rënda i ndajmë në vargje të rënda të shkurtër me rreth 450 aminoacide që i gjejmë tek IgG, D,A dhe vargje të rënda të gjata me rreth 570 aminoacide që i gjejmë tek IgE e M. Gjatësia e një vargu të shkurtër është 100 Angstrom, e një vargu të gjatë 130 Angstrom.
Përsa i përket vargjeve të lehta kemi : -vargje të lehta kapa K dhe vargje të lehta lambda Kappa i gjejmë në 2\3 e molekulave të Ig Lambda i gjejmë në 1\3 e molekulave të Ig Një molekulë imunoglobuline s`mund të këtë 1 varg të lehtë Kapa dhe një verg të lehtë
lambda, por ose të dyja Kappa, ose të dyja lambda. Vargjet e lehta kanë gjatësi 60 Angstrom. Pesha është 20-22 mije dalton. Ato përbëhen nga 2 laqe, pra 2 domene që emërtohen VL e CL.
Klasat e Ig i ndajmë në 5 klasa : G, A, M, D, E. Cilesite biologjike të përgjithshme të Ig: Me anë të fragmentit Fab ato fiksohen në
antigenin. Si pasojë e këtij fiksimi cfaqen vetitë efektorë të imunoglobulinave që barten nga Fragmenti
Fc i tyre. Këto jane: 1- Fiksimi i përbërësit C1q të komplementit dhe stimulimi i aktivizimit të këtij të fundit. Këtë
veti e ka IgG (duhen 2 molekula për këtë qellim) dhe IgM (mjafton një molekulë e kësaj). 2- Favorizimi i fagocitozës si pasojë e fiksimit të fragmentit Fc të IgG në receptorët përkates
të qelizave fagocitare (aftësia opsonizuese). këtë veti e ka vetëm IgG. 3- Kalimi i placentës (vetëm nga IgG). 4- Gjysëm-jeta e imunoglobulinave përcaktohet nga fragmenti Fc i tyre.

33
Nëpërmjet këtyre funksioneve kryesore cfaqet veprimi i tyre: 1) Eliminimi i mikrobeve të ndryshme: këtë e realizojnë me anë të prodhimit të At që i
eleminojnë mikrobet me reaksionin e lizës bakteriale, të aglutinimit, neutralizimit etj. 2) kanë funksion imunorregullues sidomos në gjëndjet alergjike. Krijojnë At bllokuese të
nënklasës IgG4 që neutralizojnë Ag alergjizues që futen në organizëm. 3) Eliminimi i Ag. Bëhet fjalë kryesisht për autoantigenët dhe kjo realizohet me ndihmën e
limfociteve N.K. Imunoglobulina G : Struktura e IgG është e ndërtuar nga 2 vargje të lehtë e dy vargje të
rëndë. Pesha është 150-180 mije dalton. Konstantja e ultracentrifuguar është 75 (unitet Swedberg). E kalon placentën, e fikson komplementin në përgjithësi, 1\2 jeta e Ig është 21 ditë, molekulat e IgG përbëjne70-80% të te gjithë antitrupave. Përqindja në serum është 10-12g\l. Ndahet në 4 nënklasa: IgG1,G2,G3,G4. Më me shumicë janë IgG1 e më me pakice IgG4. Përqendrimet e përafërta në serum janë si vijon: IgG1=8g/L; IgG2=4g/L; IgG3=1g/L; IgG4=0,4 g/L të 4 nënklasat e kalojnë placentën, vetëm IgG4 nuk e fikson komplementin. IgG është molekulë bivalente në lidhje me bashkëveprimin me antigenin. Ajo ka efekt të rëndësishëm opsonizues (favorizues të fagocitozës) pasi të gjitha qelizat fagocitare kanë receptorë për fragmentin Fc të IgG. IgG prodhohet pas IgM në përgjigjen antitrupore parësore, por ajo është klasa predominuese në atë dytësore. Afiniteti i IgG për antigenin rritet vazhdimisht gjatë përgjigjes imunitare në ndryshim me IgM e cila nuk pëson një evolucion të tillë.
Imunoglobulina M: Ka një peshe 920 mije - 1milion. Konstantja e ultracentrifugimit 19 S. Gjysëmjeta është 5-6ditë. Antitrupat e klasës IgM i gjejmë në aglutininat e grupit të gjakut të sistemit AB0.
Struktura e IgM është e përbërë nga 5 struktura bazë të ngjashme me monomerin IgG dhe të lidhura midis tyre në trajte të një ylli nga pjesët konstante, pra pjesët Fc. IgM është penta ose dekavalente në lidhje me antigenin dhe në vartësi të kushteve ekspërimentale.
Pjesët Fc bashkohën me ura disulfure, në vendin e bashkimit të të njesi 5 strukturave ka një pjesëz J.
Funksioni i IgM : realizon përgjigjen primare pas futjes se Ag. Pra At e para janë të klasës IgM. Përqindja në gjak është 0,8-1,2 g\l. E fikson mirë komplementin, s'ka funksion opsonizues, nuk e kalon placentën, gjysëm - jetën e ka rreth 5-6 ditë.
Imuoglobulina A: Gjëndet në forma të monomereve ose polimereve. Pesha molekulare është 150 mije dalton,
konstantja e ultracenrifugimit 7 njësi Svedberg. Vargu i rëndë është alfa. Ka 2 nënklasa A1 dhe A2 ku secila prej tyre ka vargun e rëndë alfa1 dhe alfa2.
IgA ndahet edhe në IgA serike dhe sekretore. IgA serike në 90% të saj përbëhet nga IgA1. IgA sekretore në 60% të saj përbëhet nga IgA2 dhe përmban pjesëzen sekretore që e mbron molekulën e IgA nga veprimi i fermenteve proteolitike të aparatit tretes. Ky polipeptid formohet nga qelizat epiteliale të zorreve të holla . Molekulat e IgA sekretore kanë peshe molekulare 400 mije dalton dhe janë në formën e një dimeri (2 (H2L2) ) tetravalent. Konstantja e cetrifugimit është 11 Svedberg. Përmbajtja në serum e saj është 1-4 g\l. Nuk e fikson komplementin (rrugën klasike të tij) dhe as ka funksion opsonizues si dhe nuk e kalon barrierën placentare. Gjysëm jeta është 5-7 ditë. në serum IgA është në formën e monomerit ( H2L2). Përqendrimi në serum i IgA1 është rreth 3,5 g/L, i IgA2 rreth 0,4 g/L.
Roli kryesor i IgA është mbrojtja e mukozave nga agjentët infeksioze. IgA sekretore përbëjne rreth 90% të imunoglobulinave të sekrecioneve digjestive, respiratore, qumështin e nënes, lotët, peshtyme.
Imunoglobulina D: Kjo klasë imunoglobuline është e pranishme në membranën e një pjesë të limfociteve B dhe shërben si receptor për antigenin në këto qeliza duke vepruar në kontrollin e aktivizimit dhe të supresionit limfocitar. Ka një gjysëm-jetë të shkurtër prej 2,8 ditë. Përqëndrimi në gjak është i ulët (0-0,4 g/l).
Imunoglobulina E: Në gjak përqendrimi i kësaj imunoglobuline është shumë i ulët (17-450mikrogram/liter). Me
anë të pjesës Fc të tyre IgE lidhen ngushtë me receptorët përkates në mastocitet. Kontakti me

34
antigenin con në degranulimin e mastociteve dhe clirimin e aminave vazoaktive dhe të citokinave si dhe në sintezën e një sërë varieteteve të mediatorëve inflamatore derivate të acidit arakidonik. Këto procese janë përgjegjës të simptomave të rrufës alergjike, të astmës bronkiale dhe fenomeneve të tjera alergjike.
Roli kryesor fiziologjik i IgE duket se është mbrojtja indore, kryesisht e mukozave dhe e lëkurës nga agresionet fiziko-kimike apo mikrobiale nëpërmjet provokimit të një reaksioni akut inflamator si pasojë e mobilizimit të faktorëve plazmatike dhe të qelizave efektorë. Agjentet infeksioze pasi depërtojnë barrierën e IgA kombinojnë me IgE specifike mbi mastocitet dhe stimulojnë clirimin e agjenteve vazoaktive dhe të faktorëve kimiotaktike për granulocitet duke shkaktuar kështu një influks të IgG plazmatike, komplementit, polimorfonukleareve dhe të eozinofileve. IgE është divalente në lidhje me antigenin. Kjo do të thotë që një molekulë antigeni duhet të lidhet me 2 molekula IgE në sipërfaqen ë mastocitit apo bazofilit për të degranuluar këto të fundit. KAPITULLI 7. BAZAT GJENETIKE TË SHUMËLLOJSHMËRISË FUNKSIONALE TË ANTITRUPAVE, IZOTIPIA, ALOTIPIA, IDIOTIPIA, HIBRIDOMAT
Bazat strukturale të funksionit të imunoglobulinave: Një monomer imunoglobuline përbëhet nga 2 vargje të lehta, të dy ose kappa ose lambda si
dhe 2 vargje të rënda: my, gama, alfa, delta, ose epsilon. Vargjet e rënda quhen kështu se janë 2 herë me të gjatë se vargjet e lehtë. Vargu i lehtë ka një
gjysëm të ndryshueshme dhe një gjysëm konstante ndërsa vargu i rëndë ka dhe ai një pjesë të ndryshueshme me të njëjtën madhësi me vargun e lehtë ndërsa pjesa konstante është 3 herë me e gjatë se ajo e pjesës konstante të vargut të lehtë.
Ne një monomer Ig dallojme fragmentin Fc të përbërë nga pjesët konstante të vargjeve të rëndë dhe 2 fragmentet Fab të përbërë nga 2 vargjet e lehta dhe 1/2 proksimale e vargjeve të rëndë.

35
sss
ss
ss Fc
Fab
Fab
(Fab') 2 2
Struktura e nje monomeri te IgG:Kur prerja e molekules behet majtas lidhjes bisulfurore midis 2 vargjeve te renda kemi formimime nje fragmenti Fc dhe te dy fragmenteve Fab.Ndersa kur veprimi i enzimes proteolitike ndodh djathtas kesaj lidhjeje bisulfurore formohet fragm(Fab')2 i perbere nga 2 fragmente Fab, ndersa fragmentiFc copezohet ne fragmente shume te vegjel(veprimi i pepsines).
(veprimi i papaines)ente
Fragmentet Fc dhe Fab kanë 2 funksione të ndryshme: Fab ka funksion të lidhet me përcaktuesin antigenik specifik për te. Ky specificitet përcaktohet nga bashkësia e 2 pjesëve variabël të vargjeve të lehta me ato të rënda. Fragmenti Fc përcakton klasën dhe nënklasën e Ig dhe bart funksionet efektorë të imunoglobulinave. Ndër me kryesoret nga këto funksione përmendim: 1- Gjysëm jeta e Imunoglobulinave (21 ditë për IgG, 5-7 ditë për klasat e tjera të IgG). 2 -Fiksimi i komplementit 3- Fiksimi i Fc me receptorët përkates, të pranishem në një sërë qelizash si makrofagët dhe neutrofilet (IgG), bazofilet dhe mastocitet (IgE) etj. Ky funksion është i rëndësishëm për fagocitozën ,për shpërthimin e reaksioneve inflamatore me ndërmjetësine e IgE, si dhe për rregullimin e përgjigjes imunitare. 4- Aftësia për të përshkuar barrierën placentare (kete veti e ka vetëm IgG).
Dualiteti funksional i imunoglobulinave rrjedh nga menyra e kodimit gjenetik të vargjeve të Ig.
Idiotipet: Shumëllojshmeria e pjesëve të ndryshueshme përcakton dhe shumëllojshmerine e fragmenteve Fab. Nqs këto fragmente i konsiderojme si Antigenë dhe i injëktojme, brënda të njëjtes specie, nga një individ që mbart një Fab të caktuar në një tjetër që nuk e ka këtë Fab (psh sepse nuk është imunizuar nga një antigen i caktuar si individi i parë) atëherë në individin 2 prodhohen antitrupa që njohin përcaktues antigenike që ndodhen në fragmentet Fab dhe kryesisht në pjesët variabël të tyre. Këta përcaktues antigenike quhen idiotipe ndërsa antitrupat specifike ndaj tyre quhen antiidiotipe.
Pra antitrupat antiidiotipike lidhen me pjesën e ndryshueshme të imunoglobulinave në mënyrë analoge me antigenin specifik ndaj kësaj pjesë. Ndërsa pjesët Fab kanë një shumëllojshmeri që korespondon me shumëllojshmerine e Antigenëve në natyre fragmentet Fc kanë një

36
shumëllojshmeri shumë me të kufizuar e cila është në dy nivele: shumëllojshmeria izotipike dhe shumëllojshmeria alotipike.
Izotipet: Kur një molekulë Ig e një specie psh. e njëriut e injëktojme në një individ të një tjetër specie psh tek lepuri atëherë krijohen antitrupa që njohin përcaktues antigenike që ndodhen në të gjithë individët e species se parë. Këto përcaktues antigenike ndodhen kryesisht në regjionet konstante të vargjeve të Ig dhe quhen përcaktues izotipike. Përcaktuesit izotipike përcaktojnë nga ana e tyre izotipine nocion ky që është i lidhur me klasat dhe nënklasat e ndryshme të imunoglobulinave. Klasat dhe nënklasat e ndryshme të imunoglobulinave pra izotipet e tyre përcaktohen nga genet koduese C që kodojnë për 1/2 distale të vargjeve të lehta dhe 3/4 distale të vargjeve të rënda. Ndërsa ekzistojnë 2 izotipe për vargjet e lehta (njëri kappa dhe tjetri lambda), për vargjet e rënda kemi 9 izotipe të ndryshme që kodohen nga 9 gjene C, të cilët ndodhen njëri pas tjetrit në regjionin kromozomik që kodon për pjesën konstante të vargjeve të rënda. Secili nga këta izotipe përfaqeson një klasë ose nënklasë imunoglobuline, të cilët përcaktohen pikërisht nga pjesët konstante të vargjeve të rënda.
Pra izotipet janë përcaktues antigenike që gjënden në pjesën konstante të vargjeve të rënda ose të lehta dhe që përcaktojnë tipin e vargut të lehtë (kappa ose lambda) apo klasën dhe nënklasën e imunoglobulines. Ata janë të pranishem në të gjithë individët e të njëjtes specie.
V1 Vn D1 Dn J1 Jn delta my gama3 1 2 4 alfa 1 2 epsilon
genet koduese te pjeseve variabel te vargjeve te renda
genet koduere te pjeseve konstante te vargjeve te renda (izotipet).
5' 3'Kromozomi
s
Alotipet: janë përcaktues antigenike të imunoglobulinave që janë të pranishem në grupe të ndryshëm individësh brënda të njëjtes specie, pra jo në të gjithë individët e një specie si izotipet.
Nga ana gjenetike, ndërsa izotipet janë të koduar nga gene të pranishem në të gjithë individët e një specie, alotipet janë variante alelike të këtyre gjeneve që janë të shpërndare në grupe të ndryshme individësh brënda species. Pra alotipet janë variante alelike të geneve koduese të izotipeve. Antitrupat antialotipike krijohen duke injëktuar alotipin e pranishem në një grup të caktuar popullatë në një grup tjetër në të cilin ky alotip nuk është i pranishem. të njëriu njihen rreth 20 variante alelike (alotipe) të vargjeve të rënda gama (Gm) dhe 3 variante alelike të vargjeve të lehta lambda (Km).
Antitrupat monoklonale dhe hibridomat: Zbulimi i menyres se prodhimit të antitrupave monoklonale me teknikën e hibridomave
përbën një revolucion në biologjine moderne. Antitrupat monoklonale haseshin deri atëhere vetëm në patologji, në mielomat. Studimi i tyre u tregua shumë i vlefshem për sqarimin e strukturës dhe funksionit të imunoglobulinave. Prodhimi i imunoglobulinave monoklonale in vitro me teknikën e hibridomave bazohet me principin e formimit të një plazmociti hibrid, i cili nga një prind trashëgon aftësine të shumëzohet në mënyrë të pakufizuar dhe nga prindi tjetër aftësine të prodhoje imunoglobulina me një specificitet antitrupor të vetëm ndaj një përcaktuesi antigenik të caktuar. për këtë qellim përdoren qeliza të një tumori që provokohët në mijtë dhe që quhet plazmocitomë në ndryshim nga limfocitet B ose plazmocitet normale që kanë një jetëgjatësi të kufizuar, qelizat e plazmocitomes për arësye të transformimit malinj të tyre kanë aftësi të shumëzohen në kultura in vitro në mënyrë të pakufizuar, mjafton që të rriten në terrene kulturore të përshtatshem. Këto plazmocitoma zgjidhen si jo prodhuese imunoglobulinash. Partneri tjetër në formimin e hibridomes janë limfocitet e shpretkës se miut i cili është i imunizuar ndaj një antigeni të caktuar. Ndër këta limfocite gjënden dhe limfocitet B që prodhojnë antitrupin me specificitetin antigenik që deshirojmë. Formimi i hibridomës bëhet duke fuzionuar (me anë të një agjenti fuzionues sic është polietilenglikoli) qelizat e plazmocitomës me limfocitet splenike. Faza ë dytë është seleksionimi i

37
hibrideve nga qelizat e plazmocitomes të pafuzionuar, e cila realizohet me terrene kulture të posacme, ndërsa limfocitet normale eliminohen vetë në kulture për arësye të jetëgjatësise se tyre të kufizuar. Faza e trete është seleksionimi i hibrideve që sekretojnë imunoglobulina. Faza tjetër është klonimi i hibridomave prodhuese të antitrupave deri sa në cdo kulture të kemi deri në qelizë hibridome në shumëzim. Me tej seleksionohen ato hibride që sekretojnë antitrupin me specificitetin që na intereson. Klonimi i mëtejshëm i këtyre hibrideve (ne cdo kulture jo me shumë se 1qelizë prodhuese antitrupash) garanton sigurine që antitrupat e prodhuara nga këta antitrupa të jenë me të vërtete monoklonale, pra me origjinë nga një qelizë e vetme prodhuese antitrupash, e cila në shumëzim të vazhdueshem prodhon një klon homogjen qelizash të cilat janë një burim i pashtershem antitrupash me një specificitet të vetëm të drejtuae ndaj një specificiteti antigenik të caktuar. Këta antitrupa monoklonale të prodhuara në këtë mënyrë in vitro kanë gjetur një fushe shumë të gjërë përdorimi si në aspektin praktik diagnostik apo terapeutik ashtu dhe në aspektin kërkimor. 1. Me anë të antitrupave monoklonalë është bërë e mundur identifikimi i nënpopullatave të ndryshme limfocitare (CD3; CD4; CD8; etj) ashtu si dhe i qelizave të llojeve të tjera, duke përcaktuar shënjuesit membranore të tyre. 2. Me anë të At. mk. është bërë e mundur izolimi i popullatave limfocitare nga njëra tjetra për qëllime të ndryshme. 3. Me anë të tyre është bërë i mundur tipizimi me i sakte i llojeve të ndryshme të leucemive apo dhe i proliferimeve të tjera malinjë duke patur si qellim një trajtim me të përshtatshëm të tyre. 4. Duke e lidhur At. mk. me një substance antitumorale bëhet e mundur që këtë substance ta përdorim për eliminimin e qelizave tumorale pa dëmtuar dhe qelizat e shëndosha të organizmit si me kimioterapinë klasike. 5. Përdorimi i At. mk. si psh CD4 po merr një përdorim gjithnjë e me të madh në modulimin e përgjigjes imunitare në mjaft sëmundje autoimune apo dhe për flakjen e transplanteve.
KAPITULLI 8. SISTEMI KOMPLEMENTAR (S.K.)
Të dhëna të përgjithshme: Sistemi komplementar (S.K.) është një sistem proteinash plazmatike që aktivizohen në
formën e një kaskade enzimatike d.m.th. cdo përbërës i këtij sistemi kur aktuvuzohet nga përbërësi paraardhes shërbën si enzime aktivizuese e përbërësit pasardhes. Proteinat e S.K. janë rreth 20 dhe ato gjënden të paaktivizuara në serum ku përbëjne rreth 5 % të proteinave plazmatike me rreth 300 mg%. Aktivizimi i sistemit komplementar ngjan në shumë aspekte me sisteme të tjerë enzimatike të plazmes si sistemi i koagulimit, i fibrinolizës apo i kininave plazmatike. një nga efektet kryesore biologjike të komplementit është eliminimi i antigenëve të huaj me natyre ekzogjene dhe lokalizim ekstraqelizor. Në këtë grup antigenësh përfshihen shumica e bakterieve me cikël jetësor jashtëqelizor si dhe viruset apo mikroorganizmat e tjera ndaj të cilëve komplementi vepron në momentin e ciklit jashtëqelizor të tyre.

38
Kur mikroorganizmi depërton në brëndësi të qelizës se organizmit komplementi nuk mund të veproje me. Veprimin e tij S.K. e realizon me anë të aktivizimit të tij. këtë aktivizim mund ta ndajmë në 2 etapa që vijne njëra pas tjetres: 1. Etapa e njohjes se antigenit të huaj 2. Etapa efektore ose vepruese e komplementit. Në etapen e parë dallojme 2 rrugë kryesore të aktivizimit të komplementit: a) Rruga klasike dhe b) Rruga alterne. Rruga alterne është me e vjetra filogjenetikisht dhe haset që në speciet jo vertebrore, pra kjo rrugë ka të bëjë me imunitetin e lindur. Rruga klasike është filogjenetikisht me e re, pasi haset vetëm në vertebroret. Ajo aktivizohet si pasojë e bashkëveprimit të antitrupit me antigenin, pra ka nevoje për imunitetin e fituar që të realizohet.
Aktivizimi i rrugës alterne: Kryhet me ndërmjetësinë e polisakarideve bakteriale, të cilët mund të jenë të pranishëm në kapsulat bakteriale ose në formën e endotoksinave. Përbërësit e kësaj rrugë janë C3, B, D, P, H dhe I. Proteina kryesore e SK është C3 rrotull së cilës aktivizohet SK me të dy rrugët e tij. C3 native ka një PM prej 195 kD dhe është përbërësi i komplementit me përqendrimin me të lartë në serum. Në kondita normale C3 aktivizohet në sasira shumë të pakta në serum. Si pasojë formohet vazhdimisht një sasi e pakët C3b që bashkohët me përbërësin D dhe formohet C3bBb. Ky kompleks quhet ndryshe C3 konvertaze sepse vepron mbi C3 dhe e ndan atë në C3a dhe C3b. Ky është një proces autokatalitik sepse aktivizimi i C3 shkakton formimin e një enzime që nga ana e saj shërbën përsëri për aktivizimin e C3. Në formimin e kësaj konvertaze ndërhyn dhe faktori D dhe P(propërdina) që e stabilizojnë C3 konvertazen. Normalisht aktivizimi i C3 inhibohet nga veprimi i proteinave H dhe I. Të dyja këto pengojnë formomin e C3 konvertazës sepse nuk lejojnë që në mungesë të stimulit antigenik të kemi aktivizim të panevojshëm të C3. Rruga alterne aktivizohet kur një antigen i huaj, në formën e një polisakaridi bakterial psh depërton në organizëm. Atëhere C3 konvertaza i "fshihet" veprimit inhibues të H dhe I dhe ngelet fushë e lirë për aktivizimin e mëtejshëm të C3 nën veprimin e faktorëve D dhe P. Si pasojë formohet një sasi e madhe produktesh aktive te C3 sic janë C3b dhe C3a. Aktivizimi i C3 është pika ku bashkohët rruga alterne me rrugën klasike të aktivizimit të sistemit komplementar. Rruga efektore që pason është e njëjte për të dy format e aktivizimit të komplementit.
Rruga klasike: Kërkon ndërmjetësinë e antitrupave të klasës IgM dhe IgG (vetëm nënklasa IgG4 nuk e aktivizon komplementin). Këto imunoglobulina e aktivizojnë komplementin vetëm kur ato bashkëveprojnë me antigenin e huaj. Si pasojë e këtij bashkëveprimi ndodh një ndryshim hapesinor në domenin CH2 të vargut të rëndë gama ose my. Ky ndryshim bën të mundur fiksimin e C1q. Kjo është një proteine termolabile me peshe të madhe molekulare, rreth 400000 dalton, e cila nevojit fiksimin në 2 molekula të afërta IgG për t'u aktivizuar ose në vetëm një molekulë IgM. Aktivizimi i C1q sjell si pasojë fiksimin në të te C1r dhe C1s. Kompleksi trimolekular i formuar dhe i përbërë nga C1q, C1r, C1s vepron mbi C2 duke e aktivizuar në C2a dhe C2b. Kompleksi tetramolekular C1q,r,s,C2a vepron tani mbi C4 duke e aktivizuar në C4a dhe C4b. Të gjithë këta reaksione të mesipërme kërkojnë praninë e kalciumit dhe të magneziumit. C4b është fragmenti aktiv i C4 dhe se bashku me C2a formon C2aC4b që është bhe C3 konvertaze, pra ekuivalenti i C3bBb i rrugës alterne. Kjo konvertaze e rrugës klasike aktivizon C3 duke e ndarë në C3a dhe C3b.
Me aktivizimin e C3 fillon etapa efektorë e aktivizimit të komplementit e cila është e përbashkët si për rrugën klasike ashtu dhe për atë alterne. Kjo vazhdon me aktivizimin e përbërësve nga C5 tek C9. Edhe C5 ndahet në C5a dhe C5b dhe si konkluzion kemi aktivizimin e kompleksit C6C7C8C9 rezultat i të cilit është formimi i poreve në sipërfaqen e qelizës shënjë (ne sipërfaqen e të ciles realizohet ky aktivizim) dhe për pasojë ndodh liza osmotike e qelizës (funksioni citolitik i komplementit).
Veprimet efektorë të sistemit komplementar: 1. Efekti citolitik i mesipërm shërben për të realizuar lizën e qelizave të huaja për organizmin
(psh bakterieve apo dhe viruseve). 2. Efekti favorizues i fagocitozës (opsonizimi) realizohet kryesisht me anë të fragmentit
C3b. Të gjitha qelizat fagocitare kanë receptorë për C3b në sipërfaqen e tyre. Kur gjenerohet formimi i C3b në sipërfaqen e një qelizë shënjë ose mbi një molekulë antigenike atëherë krijohet mundesia që

39
kjo pjesë e C3b të lidhet me receptorin përkates në sipërfaqen e fagocitit dhe kështu favorizohet fuqishëm fagocitoza e qelizës apo molekulës shënjë.
3. Efekti favorizues i inflamacionit. Inflamacioni është një mekanizem mbrojtës i organizmit me qellim eliminimin e antigenit të huaj. Dy molekula janë favorizuese të fuqishme të inflamacionit: C3a dhe C5a. Këto 2 molekula kanë këto veti proinflamatore: a) Veti kimiotaktike ka kryesisht C5a. Me këtë veti ne kuptojme aftësine e C5a për të tërhequr qelizat fagocitare kryesisht neutrofilet. b) Ato veprojnë si anafilatoksina duke realizuar degranulimin e bazofileve dhe të mastociteve. Nga granulat e këtyre clirohen shumë mediatorë të inflamacionit si histamina etj. veprimi i të cilave është kryesisht vazodilatator dhe rritja e përmeabilitetit kapilar.
Përbërësit e komplementit sintetizohen në qeliza të ndryshme si në hepar, makrofagë, limfocite, apo qelizat epiteliale të intestinit.
Ndryshimet fiziollogjike apo patollogjike të komplementit: Ulja e nivelit të komplementit në plazem (hipokomplementemia) është e 2 llojshme: e lindur
apo e fituar. Të lindur apo hipokomplementemi parësore kemi mangësite e lindura të përbërësve të ndryshëm të tij: relativisht me e shpeshta është mangësia e lindur e C2 që shoqërohet shpesh me lupus eritematoz sistemik, pra kemi mangësi të eliminimit të antigenëve me rrugën klasike të aktivizimit të komplementit. Mangësite e përbërësve të tjerë janë akoma me të rralla. Mangësite e C3 deri C9 shoqërohen nga infeksione të rënda bakteriale me karakter jashtëqelizor si koket piogjene (pneumokoku, streptokoku, meningokoku) apo bakterie të tjera.
Një sëmundje tjetër e lidhur me një mangësi të lindur të një përbërësi të komplementit është angioedema hereditare. Kjo shkaktohet nga mungesa e lindur e inhibitorit të C1(C1INH). Në kushte fiziollogjike ky inhibitor ka për funksion të pengojë aktivizimin e panevojshëm të C1q. Mungesa ë tij sjell në këta të sëmurë aktivizimin e komplementit pas cdo stimuli apo traume si psh ndërhyrjet dentare etj. Ky aktivizim shoqërohet me edema të padhimbshme por që mund të shtrihen mjaft dhe të rrezikojnë deri edemë laringeale.
Hipokomplementemite e fituara ose dytësore janë shumë me të shpeshta dhe i hasim në sëmundje autoimune si psh në lupusin eritematoz sistemik (LES), glomerulonefritet autoimune apo fazen akute të angioedemes hereditare.
Rritje të komplementit serik (hiperkomplementemi) hasim në sëmundje me inflamacion të zgjatur dhe të theksuar si psh në artritin reumatoid (AR) në të cilin nga ana tjetër kemi ulje të nivelit të komplementit në lëngun sinovial të artikulacionit të inflamuar.
Ndryshimet fiziologjike të komplementit serik janë pak të rëndësishme. Norma e tij lëkundet nga 80-120 njësi CH50. Në të porsalindurit niveli është disi me i rritur dhe ulët me pas në normat e mesipërme. KAPITULLI 9. KOMPLEKSI MADHOR I PAJTUESHMERISE INDORE: KODIMI GJENETIK, STRUKTURA MOLEKULARE, FUNKSIONET BIOLOGJIKE, LIDHJA E TIJ ME SËMUNDJET.
Përkufizimet: Kompleksi madhor i pajtueshmerise indore (KMPI) është një kompleks genesh që kodohen
në krahun e shkurtër të kromozomit të gjashtë tek njeriu (ku njihet dhe me emërtimin HLA - human leucocyte antigen-). Karakteristikë kryesore e këtyre geneve është polimorfizmi alelik i tyre, pra brënda të njëjtes specie grupe të ndryshme individësh bartin alele të ndryshme të te njëjteve lokuse. Funksioni i këtij kompleksi është kodimi për molekula membranore në sipërfaqen e mjaft qelizave të organizmit dhe që luajnë një rol të rëndësishëm në përgjigjen imunitare.

40
KMPI e ndajmë në 3 regjione kryesore gjenetike. Sipa vendosjes 5' - 3' në genome dhe pra duke u nisur nga afersia me centromerin këta regjione jane: 1. Regj. të klasës II 2. Regj. të klasës III 3. Regj i klasës I.
Klasat KMPI I III IIHLA DP DQ DR C4 C2 FB etj. B C A G
5' 3'
Regjionet koduese te sist. HLA te krahu i shkurter Kromoz. 6.
Në klasën I të KMPI kemi 3 lokuse kryesore: B,C, A. Secili prej tyre karakterizohet nga
shumëllojshmeria alelike që është me e shprehur për lokusin B pastaj A dhe me pak për lokusin C. Këto gjene kodojnë për një varg polipeptidik që quhet vargu alfa (P.M. rreth 43000 dalton) dhe që është i shprehur në sipërfaqen e të gjitha qelizave me bërthame të organizmit. Ky varg përbëhet nga 3 domene ekstraqelizore (homologe me domenet e imunoglobulinave) që duke filluar nga domeni me distal quhen alfa 1, alfa2, alfa3. në nivelin e domenit alfa 3, këtij vargu i është asociuar me lidhje jo kovalente por hapesinore një varg me P.M. rreth 12000 dalton që quhet beta-2-mikroglobulinë dhe që është i koduar në një kromozom tjetër nga kromozomi 6. Pra molekula e klasës I të KMPI përbëhet nga vargu alfa+beta2mikroglobulina. Shumëllojshmeria alotipike e vargut alfa bartet nga ana molekulare nga domeni alfa1 dhe alfa2 (pra, ndryshimet nga një alotip në tjetrin të KMPI kushtezohen nga ndryshimet në rënditjen e aminoacideve në domenet alfa1 dhe alfa2), ndërsa beta2mikroglobulina dhe domeni alfa3 s'kane një shumëllojshmeri të tillë. Karakteristike e të tre domeneve të vargut alfa si dhe të beta2mikroglobulines është homologjia e tyre me domenet e vargjeve të rënda dhe të lehta të Ig si dhe me ata të vargjeve të TCR. të gjthe këta domene kanë rreth 110 aminoacide si dhe një lidhje kovalente bisulfurore që është e vendosur brënda domenit.
Te gjitha qelizat me bërthame të organizmit bartin në sipërfaqen e tyre molekulat e klasës se I. Me tepër ato janë shprehur me qelizat limfoide, me pak në ato hepatike, mushkeri dhe veshka dhe mjaft rralle në tru dhe muskujt skeletike. Tek njëriu, sipërfaqja e trofoblastit viloz nuk shpreh Antigenët HLA-A, B ose C; në vend të tyre ai shpreh HLA-G i cili nuk shprehet në asnjë lloj indi tjetër. Antigenët me shumë të shprehur të njëriu janë HLA-A dhe HLA-B.
Klasa e II: Molekulat e klasës II janë gjithashtu glikoproteina transmembranore të përbëra nga një një
varg alfa dhe një beta secili me një peshe molekulare përkatesisht prej 34 dhe 28 kD. Ekziston një homologji në sekuencat e këtyre vargjeve me klasën I dhe domenet me afer membranës qelizore, ato alfa2 dhe beta 2, kanë palosjen karakteristike të Ig ndërsa ato alfa1 dhe beta 1i ngjajnë domeneve alfa1 dhe alfa2 të Kl I duke formuar një "gropëz" ku vendoset polipeptidi antigenik. Molekulat e Klasës II janë të shprehura në limfocitet B, qelizat dentritike, dhe makrofagët; sidoqofte dhe qeliza të tjera si endoteliumi kapilar, dhe shumë qeliza epiteliale kur aktivizohen nga gama-interferoni psh. mund të shprehin molekulat e klasës II ashtu sic bëjnë dhe limfocitet T të aktivizuara.
Klasa e III: Ne këtë regjion kodohen një sërë molekulash të komplementit kryesisht si dhe të TNF,
molekula këto që nuk janë në sipërfaqet qelizore, por në serum ose sekretohen nga qeliza të ndryshme.
Polimorfizmi i gjeneve HLA. Ndryshe nga imunoglobulinat të cilat kanë 3 nivele variabiliteti: idiotipik, izotipik dhe alotipik, sistemi HLA ka vetëm 2 nivelet e fundit (izotipik dhe alotipik). Ai izotipik shprehet në lokuset e ndryshme të sistemit HLA (A, B,C, DR, DQ, DR etj). Karakteristika kryesore e HLA është polimorfizmi alelik (ne individë të ndryshëm, alele të ndryshme të te njëjtit lokus). HLA-A dhe B janë shumë polimorfike po kështu dhe peptidet e klasës II DQ alfa, DQ beta dhe DR beta, me pak DP beta ndërsa DR alfa dhe beta-2-mikroglobulina (e cila nuk kodohet në kromozomin 6) janë molekula invariante. Ndryshimet e aminoacideve përgjegjës për këtë polimorfizem ndodhin në domenet alfa1 dhe alfa2 të klasës I si dhe në ato alfa1 dhe beta1 të klasës II. Eshtë shumë sinjifikant fakti që këto

41
ndryshime ndodhin në aminoacidet e domeneve të mesipërme të lokalizuara rreth gropëzes ku vendoset polipeptidi antigenik, pra ato përcaktojnë dhe variabilitetin e lidhjes me antigenin nga alele të ndryshme të sistemit HLA.
Gjenet e sistemit HLA kanë ekspresion kodominant dhe segregojnë në mënyrë mendeliane (si grupet e gjakut p.sh.). Tërësia e geneve KMPI që ndodhen në një nga kromozomet cift formjne haplotipin. Pra cdo individ ka 2 haplotipe dhe fëmija trashegon një haplotip nga cdo prind.
Geni HLA
Prod. gen.
B C E A G F
HLA-B HLA-C HLA-E HLA-A HLA-G HLA-F Gjenet e Klasës I të HLA Gjenet eKlasës II të HLA GeniHLADPB DPA LMP2 TAP2 LMP7 TAP1DQB DQA DRB DRA
Produktigjenik
beta alfaHLA-DP
Kompleksiproteazom
Transporteripeptidik HLA-DQ HLA-DR
DQbeta DQalfa DRbeta DRalfa
MEMBRANA
alfa1
alfa 2
alfa 3beta2mikroglobulina
hapesira e vendosjes seantigjenit
Klasa I
alfa1
alfa2
beta 1
beta2
Klasa II
Funksionet e molekulave të KMPI Kl I. Megjithëse fillimisht të zbuluar nëpërmjet reaksioneve të flakjes se transplantit, molekulat e
HLA kanë funksione jetësore biologjike për organizmin. Ato funksionojnë si shënues qelizorë që i mundësojnë qelizat e infektuara të organizmit të njihen nga limfocitet T helper dhe citotoksikë. Polimorfizmi i pasur alelik i HLA paraqet një përgjigje të species për të amplifikuar mbrojtjen ndaj mikroorganizmave të ndryshëm.
Funksioni kryesor i molekulave të Kl I është restriksioni alotipik i funksionit të limfociteve T citotoksikë. Ky funksion ka të bëjë me faktin që limfocitet T citotoksikë të një organizmi mund të realizojnë vrasjen e një qelizë që bart një antigen të huaj vetëm nqs ky antigen është i shoqëruar me molekulat e KMPI të klasës I të alotipit të këtij organizmi. Pra limfocitet T citotoksikë të një

42
organizmi e kryejnë funksionin e tyre citolitik ndaj qelizave të infektuara me një virus vetëm nqs këto qeliza bartin të njëjtin alotip të KMPI të atij organizmi ku janë prodhuar dhe edukuar këto limfocite. Pra këta qeliza T citotoksikë s'do të jenë në gjëndje të kryejnë efektin citotoksik mbi qelizat e një organizmi tjetër (qe bart alotipe të tjera të KMPI) të infektuara me të njëjtin virus.
Fenomeni i restriksionit alotipik realizohet me anë të aftësise se molekulave të klasës I të KMPI për t'u lidhur me antigenë endogjene. Me antigenë endogjene quajme antigenët virale, autoantigenët ose antigenët tumorale, që kanë karakteristikë të përbashkët faktin që ata përdorin aparatin biologjik të vetë qelizës për t'u sintetizuar. Antigenët endogjene sintetizohen në citoplazmën e qelizës dhe aty ata lidhen me molekulat KMPI të Kl I. Antigenët endogjene paraqiten në formën e polipeptideve prej 9-20 aminoacideve dhe kuy polipeptid vendoset pikërisht në hapësirën që formohet midis domenit alfa1 dhe alfa2. Me pas molekulat e Kl I bashkë me polipeptidin e huaj do të shprehen në sipërfaqen e qelizës dhe këtu ndodh njohja e këtij asociacioni nga limfociti T cititoksik. Receptori TCR i këtij limfociti njëh kryesisht polipeptidin e huaj+domenet alfa1 dhe alfa2 të KMPI të KL I të vetë organizmit.
Funksionet e Klasës II të KMPI: Funksioni kryesor imunologjik i këtyre molekulave është paraqitja e Antigenëve ekzogjene
limfociteve T helper, të cilët njohin antigenin + KMPI të klasës II. Kjo paraqitje bëhet nga qelizat antigen-paraqitëse si limfocitet B, makrofagët apo qelizat dendritike.
Megjithëse fillimisht të zbuluar nëpërmjet transplanteve, molekulat HLA përdoren për funksione të rëndësishme vitale. Roli i tyre në përgjigjen imune është shumë i rëndësishëm dhe në këtë aspekt polimorfizmi i pasur i KMPI paraqet një përgjigje të species për të rritur variabilitetin e përgjigjes imune ndaj shumëllojshmerise se mikroorganizmave.
Përdorimi klinik i tipizimit të KMPI të njeriu (HLA): 1. Përdorimi kryesor është pa dyshim përcaktimi i përshtatshmerise midis dhuruesit dhe
marrësit të transplantit. Kjo bëhet me metodat klasike të limfocitotoksicitetit ose ato me bashkëkohore të biologjise molekulare. në rastin e parë limfocitet e individit për t'u tipizuar vihen në fillim në kontakt me serume me specifitet të njohur për alelet e ndryshme HLA. në një etape tëdytë shtohet komplementi i cili realizon lizën e limfocitit nqs. ky individ përmban alotipin HLA të antiserumit.
2. në mjeksine ligjore tipizimi i HLA të fëmija dhe prindet është metoda me e efektshme për përjashtimin e atesise dhe e konfirmon atë der në 99%.
3. në gjenetiken e popullatave studimi i aleleve të HLA është metoda me e sakte për ndjekjen e përzjerjes se popullatave të ndryshme dhe origjinës se tyre.
4. Disa alele të HLA janë të lidhura me sëmundje të ndryshme kryesisht autoimune: kështu HLA B-27 haset në 95% të te sëmurëve me Spondiloartrit ankilozant. Po kështu HLA-DR3 dhe 4 hasen në një frekuence mjaft të rritur në të sëmurët me diabet juvenil insulinovartes. Mendohet se agjentë infeksioze (Antigenë të tyre) paraqiten në mënyrë të vecantë nga sistemi imunitar i individëve me këto alotipe dhe si pasojë reaksioni imun ndaj antigenit të huaj bëhet i demshem edhe ndaj autoAntigenëve për arësye edhe të një reaksioni të kryqezuar midis tyre.
KAPITULLI 10. MEKANIZMAT VEPRUESE (EFEKTORË) TË IMUNITETIT QELIZOR
CITOKINAT SHËRBEJNË SI MEDIATORË NDËRQELIZORË. Ndërsa aktivizimi fillestar i limfociteve T dhe limfociteve B T-vartese kërkon një kontakt të
ngushtë me qelizat antigen - paraqitëse, proliferimi dhe maturimi i mëtejshëm i tyre drejtohet nga një kaskade citokinash që prodhohen nga limfocitet T dhe që ndërmjetësojnë informacionin midis qelizave. Nën termin citokina përfshijme ato molekula që ndryshe emertohen dhe limfokina, monokina, interleukina dhe interferone. Këto janë proteina me peshe të vogel molekulare, 15-25 kDa, që ndërmjetësojnë rritjen qelizore, inflamacionin, përgjigjen imunitare, diferencimin qelizor dhe

43
cikatrizimin indor. Meqënëse rregullojnë amplituden dhe kohëzgjatjen e përgjigjeve imune dhe inflamatore, citokinat kanë një gjysëmjetë të shkurtër, por kanë efekt të fuqishem pasi veprojnë në përqendrime femtomolare (10-15 M). Ndryshe nga hormonet endokrine shumica e citokinave veprojnë në afersi, në mënyrë parakrine.. Disa citokina si psh. IL-2 kanë veprim autokrin, d.m.th. ato veprojnë duke stimuluar po ato qeliza që i kanë prodhuar. Disa prej tyre kanë veprim endokrin (ne distancë si psh. IL-1). Limfokinat, pra citokinat e qelizave limfoide, hasen rralle në gjak, ndërsa qelizat jo limfoide si makrofagët p.sh. mund të stimulohen nga Antigenë bakteriale për të prodhuar citokina që dalin në qarkullim të gjakut me efekte sistemike (endokrine). Nga ana tjetër mendohet se disa citokina si IL-1 dhe TNF mund të ekzistojnë në formë membranore dhe realizojnë pra veprimin e tyre stimulator pa qënë nevoja të jenë detyrimisht në formë solubël.
Një veti tjetër e citokinave është efekti pleiotropik i tyre: dmth. ato kanë veprime të shumëfishta e të njëkohshme mbi rritjen dhe diferencimin e një sërë qelizash të ndryshme dhe për prodhimin e një sërë proteinash të ndryshme: përmendim rolin e tyre në gjenerimin e qelizave efektorë T dhe B, rregullimin e reaksioneve inflamatore kronike, kontrollin e hematopoiezes, veprimi mbi imunitetin e lindur (jo antigen specifik). Kështu, diferencimi i qelizave mëmë të palcës kockore drejt elementeve qelizore përbërës të gjakut nxitet nëpërmjet prodhimit të citokinave nga qelizat stromale të palcës kockore. Këtu përfshihet GM-CSF (granulocyte-macrophage- colony stimulating factor), G-CSF (granulocyte-colony stimulating factor), M-CSF (macriphage -colony stimulating factor), IL-6, IL-7 dhe LIF(leukemia inhibitory factor) si dhe faktorë të tjerë që prodhohen nga limfocitet T dhe makrofagët. në rastet e inflamacioneve kronike citokinat e prodhuara stimulojnë vazhdimisht prodhimin e prekursoreve të rinj qelizore në palcën kocore. IL-3 vecanerisht duhet përmendur për aftësine e saj të stimuloje qelizat prekursore hematopoietike në sinergji dhe me IL-6 dhe G-CSF.
Ndërveprimet midis citokinave: Genet që kodojnë për IL-3, 4, 5 dhe GM-CSF janë prane njëri tjetrit në kromozomon 5 në të
njëjtin regjion që përmban dhe genet kodues për M-CSF dhe receptorin qelizor për të si dhe disa faktorë të tjerë rritje qelizore apo receptorë të tyre. Ndërveprimi i këtyre mediatorëve ndjek një kaskade në të cilen një citokine indukton prodhimin e një citokine tjetër ose nxit shprehjen e një receptori për të si dhe nëpërmjet një veprimi sinergjik apo antagonist midis dy citokinave.
CITOKINA BURIMI VEPRIMI

44
IL-1 (alfa, beta, gama) makrofagët, fibroblastet -Proliferim i L-T dhe B të aktivizuara; -Indukton PGE2 dhe citokinat nga makrofagët; -Indukton IL-6, IFN-beta 1 dhe GM-CSF -Indukton në qelizat endoteliale shprehjen e molekulave të adezionit të neutrofileve dhe limfociteve T. - Nxit tempëraturen, proteinat e fazes akute, rezorbimin kockor nga osteoklastet
IL-2 limfocitet T -Rritje e limfociteve T dhe B të aktivizuar; aktivizim i qelizave NK
IL-3 limfocitet T, mastocitet -Rritje dhe diferencim i prekursoreve hematopoietike - Rritje e mastociteve
IL-4 limfocitet T CD4(+), mastocitet, qelizat stromale të palcës kockore
- Proliferim i limfociteve T dhe B të aktivizuara, i prekursoreve të mastociteve dhe të qelizave hematopoietike -Induktim i KMPI Kl II dhe i receptorëve FceR në LB dhe i p75 IL2R në LT. - Nxit kalimin në izotipet IgG1 dhe IgE. -Nxit paraqitjen e antigenit, funksionin citotoksik dhe aftësine fuzionuese të maktofageve.
IL5 limfocitet CD4, mastocitet -Nxit proliferimin e LB të aktivizuara dhe prodhimin e IgM dhe IgA. -Nxit proliferimin e eozinofileve si dhe shprehjen e p55 IL-2R.
IL6 LT CD4(+), makrofagët, mastocitet, fibroblastet
- Nxit rritjen dhe diferencimin e LT dhe B - Nxit proteinat e fazes akute
CITOKINA PRODHIMI VEPRIMI IL-7 Qelizat stromale të palcës
kockore -Proliferim i limfociteve pre B, limfociteve CD4(-)CD8(-) dhe i limfociteve T mature.

45
IL-8 Monocitet -Kemotaksis dhe aktivizim të neutrofileve. -Kemotaksis i limfociteve T.
IL-9 Limfocitet T -Rritja dhe prolif. i LT. IL-10 LTCD4(+), LB, makrofagët Inhibon sekrecionin eIFNg
-Inhibon inflamacionin me qeliza mononukleare
IL-11 qelizat stromale të palcës kockore
-,Indukton proteinat e fazes akute
IL-12 limfocitet T aktivizon qelizat NK IL-13 limfocitet T inhibon inflamacionin me
fagocite mononukleare FAKTORËT STIMULUES TE KOLONIVE (CSF) GM-CSF L.T., makrofagët,
fibroblastet rritjen e granulociteve dhe makrofagëve
G-CSF fibroblastet, endoteliumi rritjen e granulociteve M-CSF fibrobl., endotel, epitelium rrit kolonite e makrofagëve STEEL FACTOR qel. stromale të P.K. nxit ndarjen e qel. meme FAKTORËT E NEKROZES TUMORALE (TNF) TNF-alfa makrofagët, limfocitetT -Citotoksicitet tumoral'
kaheksi - Nxisin proteinat e fazes akute - Aktivitet antiviral dhe antiparazitar - Aktivizim i qelizave fagocitare - Induktim i IFNgama. TNF alfa, Il-1, GM-CSF, IL-6 - Shok endotoksik
TNF- beta limfocitet T Njëlloj INTERFERONET (IFN) IFN alfa leukocitet anti-viral; nxit shprehjen e
KMPI të KL. I IFN beta fibroblastet njëlloj IFN gama limfocitet T - Nxit shprehjen KMPI KL I
dhe II në makrofagë dhe qeliza të tjera - Nxit diferencimin e L. T citotoksikë - Nxit sintezën e IgG2a nga qelizat B të aktivizuara - Ka veprim antagonist ndaj disa veprimeve te IL-4

46
TGF-beta (transforming growth factor)
limfocitet T, B - Inhibon shprehjen e IL-2R dhe të proliferimit të qelizave T dhe B IL-2 vartes -Inhibim i hematopoiezes se induktuar nga IL-3+CSF - Nxitje e prodhimit të IgA - Riparim i plages (kemotaksine fibroblastike) dhe angiogenezes - Transformim neoplastik i disa qelizave normale
LIF (leukemia inhibitory factor)
limfocitet T -Proliferim i qelizave meme hematopoietike embrionale pa prekje të diferencimit - Kemoatraktim dhe aktivizim të eozinofileve
Citokinat: Prodhimi dhe funksioni
Th1 Th2 B Antitrupi
Il2
IL-4/5/6
IL-10
IFN-gama
Kontrolli i rritjes se limfociteve
ata
(------ = inhibim ; ______ = stimulim)
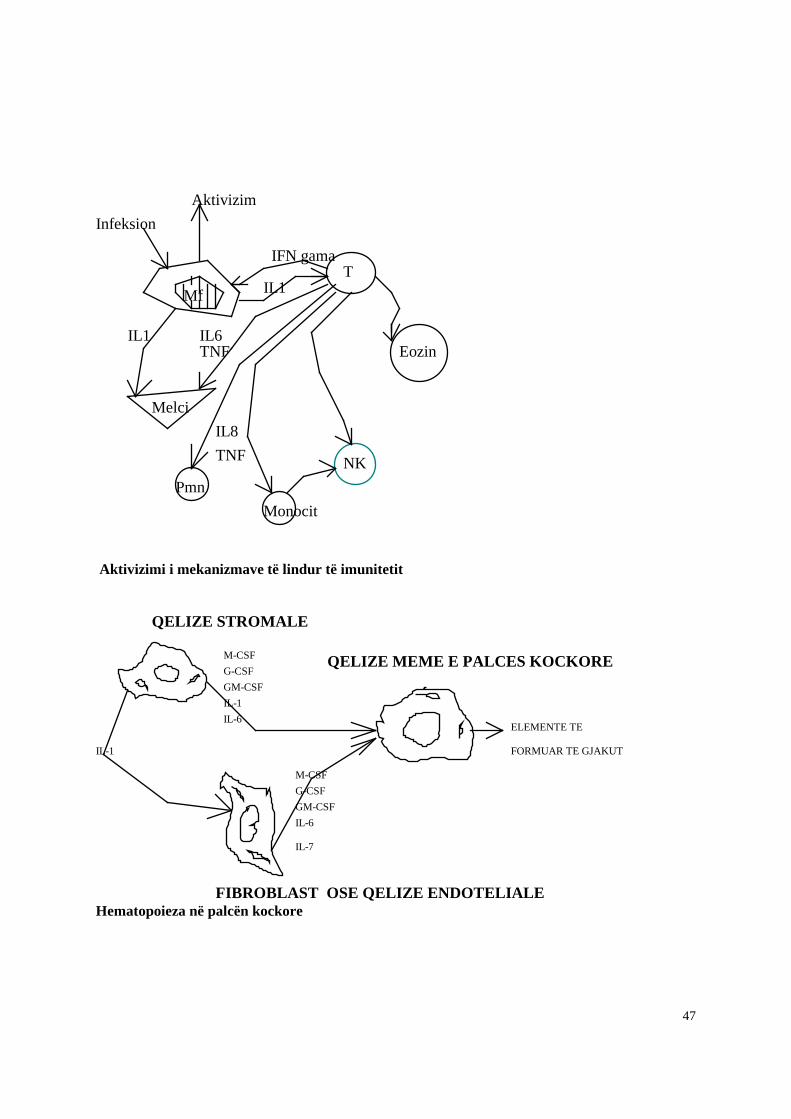
47
Mf
Melci
PmnMonocit
NK
Eozin
T
IL1
InfeksionAktivizim
IFN gama
IL1
TNFIL6
IL8TNF
Aktivizimi i mekanizmave të lindur të imunitetit
QELIZE STROMALE
FIBROBLAST OSE QELIZE ENDOTELIALE
QELIZE MEME E PALCES KOCKOREM-CSFG-CSFGM-CSFIL-1IL-6
IL-1
M-CSFG-CSFGM-CSFIL-6
IL-7
ELEMENTE TE
FORMUAR TE GJAKUT
Hematopoieza në palcën kockore

48
LLOJET E NDRYSHME TE LIMFOCITEVE T CD4 PRODHOJNË PROFILE TË NDRYSHME CITOKINASH:
Studimet e kryera në minjte me anë të kulturave afatgjatë të limfociteve T in vitro kanë vënë në dukje 2 lloje limfocitesh T CD4 (+) që dallohen nga prodhimi i citokinave të ndryshme: TH1 dhe TH2. Kuptimi biologjik i kësaj ndarje është që limfocitet TH1 nëpërmjet prodhimit të citokinave si IFN gama janë me tepër efektive kundër infeksioneve intraqelizore me viruse dhe mikroorganizma që rriten brënda makrofagëve ndërsa Th2 janë ndihmës të mirë për limfocitet B dhe duket se janë përshtatur për mbrojtjen kundër parazitëve që janë të ndjeshëm ndaj prodhimit të IgE që nxiten nga IL-4, eozinofileve që nxiten nga IL-5 dhe mastociteve të stimuluara nga IL-3/4. Duket se makrofagët i paraqesin antigenin limfociteve Th1 ndërsa limfocitet B atyre Th2.
Llojet e citokinave ne klonet e
limfociteve T tek miu
TH 1 TH 2 T citotoksikë IFN gama ++ - ++ IL-2 ++ - - TNF beta ++ - + TNF alfa ++ + + GM-CSF ++ + ++ IL-3 ++ ++ + Met. enkephalin + ++ + IL-4 - ++ - IL-5 - ++ - IL-6 - ++ - IL-10 - ++ _
Kjo ndarje e prerë e profilit të citokinave të prodhuara nuk është vënë re qarte të njeriu. Megjithatë mendohet se limfociti T i hershëm prodhon një përzierje të 2 llojeve të limfokinave dhe me vone diferencimi në Th1 apo Th2 bëhet në vartësi të stimulit antigenik. Këtë e dëshmojnë studime në njerëz të hiperimunizuar. IFN gama luan rol frenues ndaj Th2 ndërsa IL-4 dhe IL-10 frenojnë Limfocitet Th1.
Limfocitet T të aktivizuara proliferojnë nën stimulimin e citokinave. Pas aktivizimit nga antigeni të limfociteve T proliferimi i mëtejshëm i tyre është në vartësi të
IL-2. Kjo limfokine është një peptid me peshe molekulare 15.5 kDa, e cila vepron vetëm mbi qelizat T që shprehin receptorë me afinitet të lartë ndaj IL-2. Këta receptorë nuk janë të pranishem në qelizat T në qetësi, por sintetizohen brënda pak oresh pas aktivizimit. Limfoblastet T (limfocitet T të aktivizuara) prodhojnë gjithashtu një numër të madh citokinash dhe efekti proliferues i IL-6 përforcohet nga veprimi i IL-4 dhe disi të IL-6 të cilët gjithashtu bashkëveprojnë me receptorët përkates të limfociteve T në proliferim e sipër. Edhe ky proliferim ka mekanizmat e tij të kontrollit. Këtu mendohet se ndërhyn TGFbeta e cila bllokon proliferimin e induktuar nga IL-2.
Limfocitet T efektore të imunitetit me ndërmjetësi qelizore: Limfokinat e prodhuara nga limfocitet T janë gjithashtu përgjegjëse për gjenerimin e
reaksioneve inflamatore kronike antigen-specifike të drejtuara ndaj mikroorganizmave brëndaqelizore. Ngacmimi fillestar është probabilisht një përgjigje inflamatore lokale ndaj dëmtimit indor të shkaktuar nga agjenti infeksioz. Kjo përgjigje stimulon sintezën e molekulave të adezionit si VCAM-! dhe ICAM-1 në sipërfaqen e qelizave vaskulare endoteliale të përafërta. Kjo do të lejoje hyrjen e limfociteve T kujtesë në vendin e infektuar nëpërmjet receptorëve të tyre të tipit "homing" VLA-4 dhe LFA-1. Kontakti me Antigenët e përpunuar të parazitit intraqelizor do të aktivizoje limfocitet T specifike dhe do të induktoje clirimin e citokinave të sekretuara. TNF do të stimuloje me tej shprehjen e molekulave të adezionit duke rritur kështu shanset e qelizave të tjera kujtesë qarkulluese të vijne në vatren inflamatore.

49
Nga ana tjetër, limfocitet T të aktivizuar në këtë mënyrë do të sekretojnë citokinat IFN gama, GM-CSF, IL-2 dhe TNF të cilat do të aktivizojnë makrofagët për të kryer veprim vrasës ndaj mikroorganizmave të fagocituar prej tyre. Gjatë këtij procesi disa makrofagë eshe mund të vdesin dhe të clirojnë mikroorganizma të gjalle, por këta do të trajtohen tashmë nga makrofagë të rinj të aktivizuar nga citokinat e prodhuara lokalisht.
Përsa i përket infektimit të qelizave nga viruset strategjia mbrojtëse është e ndryshme dhe përfshin fillimisht ndërhyrjen e imunitetit të lindur me anë të interferonit gama si dhe të TNF alfa dhe beta të cilat mbrojnë qelizat nga infektimi viral. TNF gjithashtu ka aftësine të vrase qeliza të infektuara nga virusi gje që ka efekt të dobishem për pengimin e përhapjes se virusit. Duket se ka një sinergjizem midis IFN gama dhe TNF beta në faktin se IFN gama nxit formimin e receptorëve ndaj TNF në qelizat e infektuara.
Antigjeni
B
AntitrupaIgM,G,A, E
Plazmocit
TMf
IL-1, 6TNFa, IFNaG-CSFGM-CSF
FaktorekemotaktikeIL-8MCP-1
MIP-2 Th1= IL4,5,6,3GM-CSF
2
Th1= IFNg, TNFa/bIL-3, GM-CSF
Inflamacion me ndermjetesine e limfociteve T
IL-8
Citokinat kontrollojnë prodhimin e antitrupave si dhe të inflamacionit me ndërmjetësine e limfociteve T (MCP-1= macrophage chemotactic protein; MIP-2= macrophage inflammatory protein janë të ngjashme me IL-8.
Limfocitet T vrasëse (citotoksikë) (LT ct): Limfocitet T citotoksikë paraqesin anën tjetër madhore të përgjigjes imune me ndërmjetësi
qelizore. Ata janë me rëndësi kryesore për vrasjen e qelizave të infektuara nga viruset dhe mendohet se luajnë rol edhe në mbikqyrjen imune ndaj qelizave kanceroze.
Qeliza T citotoksikë prekursore njëh antigenin e përpunuar viral në sipërfaqen e qelizës të infektuar se bashku me molekulat e klasës I të KMPI. Ashtu si dhe limfociti B edhe LT citotoksik ka nevoje për ndihmën e limfociteve T helper, por me mekanizma të ndryshme. Bashkëpunimi efikas T-B është "i afërt" d.m.th. qelizat bashkëpunuese njohin 2 epitope të cilët janë fizikisht të lidhur menjëri tjetrin (zakonisht në të njëjten molekulë), por të ndryshëm. Kjo për faktin se imunoglobulinat sipërfaqesore në limfocitet B kapin antigenin në gjëndje native ( përcaktuesit antigenike "haptenike" të tij), e përpunojnë atë brënda në qelizë dhe ia paraqesin limfocitit T helper si një peptid (përcaktuesi antigenik "bartës") të lidhur me molekulat e klasës II të KMPI. Limfocitet T citotoksikë nuk njohin dot antigenin në gjëndje native dhe është me e mundëshme që Th dhe Tct prekursore bashkëveprojnë me të njëjten qelizë antigen-paraqitëse e cila ka përpunuar antigenin viral dhe i paraqet peptidet virale të lidhura me Antigenët e klasës II (për limfocitin Th) dhe të klasës I (për qelizën T citotoksikë prekursore). Kjo qelizë mund të jetë vetë qeliza e infektuar nga virusi. Citokinat e cliruara nga limfociti T helper do të clirohen në afersi të limfocitit T citotoksik prekursor i cili ka bashkëvepruar

50
me kompleksin KMPI KL. I+antigen dhe do të stimulohet të proliferoje dhe të diferencohet në limfocit T citotoksik nën influencën e IL-2 dhe IL-6.
Procesi letal: Limfocitet T citotoksikë janë zakonisht të nënpopullates CD8 dhe lidhja e tyre me qelizën shënjë nëpërmjet nëpërmjet njohjes se antigenit +KMPI kl I nga TCR ndihmohet nga bashkëveprimi midis CD8 dhe kl I e KMPI si dhe nga molekula të tjera aksesore si CD2 dhe LFA-1. Si pasojë e aktivizimit të qelizës vrasëse shumica e granulave të citoplazmës lokalizohen shpejt midis bërthames dhe qelizës shënjë se bashku me proteinën skeletale intraqelizore tailin e cila akumulohet nën membranën e qelizës citotoksikë mesa duket duke e mbrojtur atë nga dëmtimi gjatë procesit vrasës. Ka të dhëna se kemi ekzocitoze të përmbajtjes se granulave që përmbajne përforone, granzime dhe TNF të cilat shkaktojnë lezione në membranën e qelizës shënjë dhe vdekje të saj nëpërmjet apoptozes. Pas "puthjes se vdekjes" limfociti T citotoksik clirohet nga qeliza e vrare dhe dh kërkon një viktime të re. Nuk duhet harruar se limfocitet T ct CD8(+) sintetizojnë dhe citokina të tjera si IFN gama, e cila ka gjithashtu një potencial anti-viral.

51
KAPITULLI 11. MEKANIZMAT EFEKTORË TË IMUNITETIT TË FITUAR ME NDËRMJETËSINE E LIMFOCITEVE B.
Proliferimi dhe maturimi i përgjigjes imunitare me limfocite B është i ndërmjetësuar nga citokinat:
Aktivizimi i limfociteve B nga limfocitet T helper nëpërmjet njohjes nga TCR të këtyre të fundit të peptidit antigenik të lidhur me KMPI si dhe ndërveprimi bashkëstimulues p39/CD40, con në shprehjen e receptorit për IL-4 në sipërfaqen e limfociteve B. Clirimi i theksuar i IL-4 nga limfocitet T helper gjeneron një proliferim klonal të fuqishem dhe një ekspansion të limfociteve B të aktivizuara. në këtë proces kontribuon gjithashtu dhe IL-2 .
Nën influencën e vetëm IL-4 klonet e ekspanduara diferencohen dhe maturohen në plazmocite që sintetizijne IgE. TGF beta bashkë me IL-5 stimulon qelizat B të prodhojnë IgA. Plazmocitet që prodhojnë IgM kërkojnë nxitjen e IL-4 + IL-5 ndërsa ata që prodhojnë IgG duan influencën e kombinuar të IL-4 , IL-5 dhe IL-6 me një kontribut të mundshem nga IFN gama.
G 0 THII TCRCD 40 p39
IL-2, 4, 5, 6, IFN g
G1
S S
Aktivizim
Proliferim klonal
Maturim
IL-4 (2)
IL-4TGF beta
IL-5
IgE IgA
IL-4, 5
IL-4, 5, 6, IFN gama
IgMIgG
Përgjigja e L B ndaj një antigeni timo -vartes
Ecuria e përgjigjes imunitare me limfocitet B: Ngacmimi dytësor me antigen ose komplekse imune shkakton zgjerimin e qëndrave
gjerminative të folikujve limfoide dytësore, formimin e qendrave të reja, cfaqjen e limfociteve B kujtesë dhe zhvillimin e plazmociteve që prodhojnë imunoglobuline me afinitet të lartë për antigenin.
Si pasojë e stimulit antigenik dytësor limfocitet B specifike për këtë antigen aktivizohen nga limfocitet T helper parakortikale në bashkëpunim me qelizat dentritike interdigitale ose makrofagët dhe si pasojë migrojnë në qendrat gjerminative të folikujve limfoidedytësore. Aktivizimi mund të ndodhë edhe nga komplekse imune që ndodhen në qelizat dentritike folikulare. Limfocitet B që hyjne në qëndrat germinative bëhen centroblaste me një cikël ndarje cdo 6 oresh. Me pas ato bëhen centrocite që nuk ndahen me dhe që lokalizohen në zonën e qarte të folikulit. Ato që nuk vdesin nga apoptoza shndrrohen në imunoblaste, qeliza prekursore e plazmocitit, sekretues të sasirave të medha të imunoglobulinave. Gjatë gjithë këtij procesi ndodhin ndryshime somatike të geneve që kodojnë për pjesën variabël të Ig që prodhohet. Ato që kanë afinitet me të lartë për antigenin i shpetojnë procesit të apoptozes që realizohet me ndërhyrjen e makrofagëve. Pra kemi një seleksion natyror pozitiv për klonet e limfociteve B me afinitet me të lartë për antigenin. Kjo aq me tepër edhe për faktin se me uljen e përqendrimit të antigenit ai lidhet vetëm me receptorët me afinitet të lartë për te. Kjo rritje e

52
afinitetit të antitrupave për antigenin është mjaft e dibishme me vone kur sasia e antitrupave bie gradualusht.
Pas diferencimit të mëtejshëm qelizat ose migrojnë në mjediset ku përqendrohen plazmocitet (medula e limfonodulave p.sh.), ose transformohen në qeliza B kujtesë, kjo në bazë të sinjalit që merret. CD23 solubel si dhe IL-1alfa që duket se vijne nga qelizat folikulare dentritike stimulojnë formimin e plazmociteve që prodhojnë dhe sekretojnë sasi të medha imunoglobulinash, ndërsa CD40 e sinjalizuar nëpërmjet një limfociti T e drejton qelizën në kompartimentin e kujtesës.
Ecuria e përgjigjes antitrupore: Sinteza e antitrupave sipas klasave dhe nënklasave të ndryshme të imunoglobulinave kryhet
sipas etapave të ndryshme. Pas një përiudhe latence 3-5 ditësh (qe është në funksion të llojit dhe dozes se antigenit ) zakonisht kemi një përgjigje IgM të hershme që ka tendencë të bjere shpejt. Sinteza e IgG e arrin maksimumin e saj për një përiudhe me të gjatë kohë. Këto i përkasin përgjigjes antitrupore primare. Pas një përgjigje antitrupore dytësore (me futjen e dytë të antigenit) përgjigja e IgM në kohë i përngjan asaj primare, por koha e latences në këtë rast është më e shkurtër. Përkundrazi sinteza e IgG zhvillohet më shpejt dhe në nivele më të larta se përgjigja primare. Edhe rënja e nivelit të IgG në përgjigjen sekondare është më e ngadalshme dhe më e paket nga ana sasiore. E njëjta ndodh edhe për IgA dhe janë këto 2 klasa imunoglobulinash që do të garantojnë mbrojtjen humorale të mëtejshme ndaj këtij antigeni. E njëjta qelizë mund të kaloje nga prodhimi i IgM në atë të IgG duke ruajtur gjithmone të njëjtin idiotip (te njëjtet regjione të ndryshueshme).
Për të studiuar në se një infeksion është i hershëm apo i tanishëm ndihmon përcaktimi i klasës së antitrupave ndaj një mikroorganizmi. Kur mbizoterojnë antitrupat e klasës IgM konkludojmë se infeksioni është mjaft i afërt në kohë. Në rast se mbizoterojnë antitrupat e klasës IgG ky është një argument në favor të një kohë më të hershme të infektimit nga ky mikroorganizem. IgG mund të përfaqesojnë në këtë rast një shënjë të një infeksioni të mëparshëm me këtë mikroorganizem. për të gjykuar nëse kemi një infeksion tashmë të kaluar apo në zhvillim e sipër ndihmon përcaktimi i nivelit të antitrupave (pavaresisht nga klasa e tyre) në dinamike. Ngritja sinjifikative (prej të paktën 3 herë) e nivelit (titrit) të antitrupave ndaj një agjenti infeksioz të caktuar në një interval kohë prej 3-4 javesh është një argument i fortë në favor të një infeksioni në zhvillim e sipër dhe jo vetëm anamnestik.
Sinteza e IgG, IgA dhe IgE varet shumë nga ndihma e limfociteve T (Antigenët timovartës) ndërsa ajo e IgM shumë më pak (Antigenët jotimovartes)..
Një qelizë e vetme prodhuese antitrupash (plazmocit) prodhon vetëm një lloj imunoglobuline, të një alotipi dhe një idiotipi të vetëm. Ky quhet fenomeni i ekskluzionit alelik, pra kjo qelizë në një subjekt heterozigot prodhon imunoglobulina me shënues alelike ose maternale ose paternale, por kurre jo të dy bashkë. Idiotipi (regjionet variabël) i një imunoglobuline të prodhuar nga një plazmocit i korrespondon gjithmone idiotipit të imunoglobulines sipërfaqësore të limfocitit B nga i cili derivon ky plazmocit. Pra në një limficit të vetëm B ndërsa idiotipi dhe alotipi i imunoglobulinës së prodhuar nuk ndryshon, izotipi mund t ndryshoje (IgM në IgG ose IgA ose IgE), ndryshim ky që varet nga lloji i antigenit dhe lloji i ndihmës nga limfocitet T me citokinat përkatese.
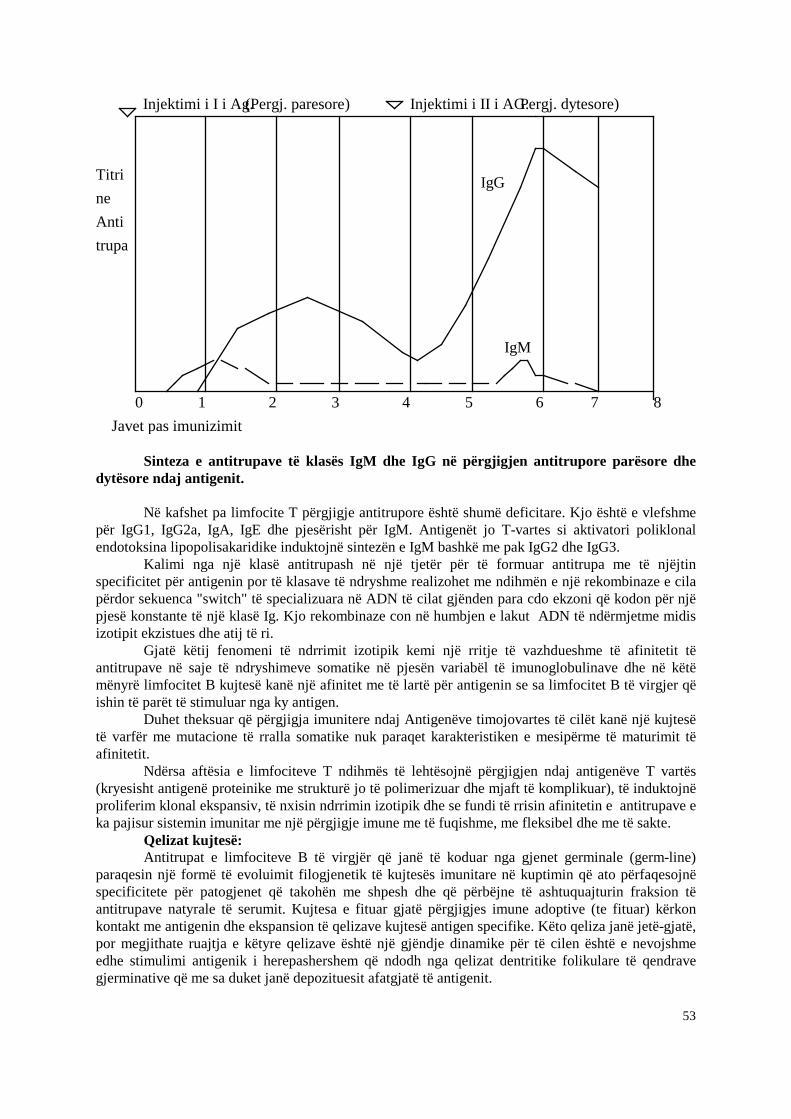
53
0 1 2 3 4 5 6 7 8Javet pas imunizimit
TitrineAntitrupa
Injektimi i I i Ag. Injektimi i II i AG.
IgM
(Pergj. paresore) Pergj. dytesore)
IgG
Sinteza e antitrupave të klasës IgM dhe IgG në përgjigjen antitrupore parësore dhe dytësore ndaj antigenit. Në kafshet pa limfocite T përgjigje antitrupore është shumë deficitare. Kjo është e vlefshme për IgG1, IgG2a, IgA, IgE dhe pjesërisht për IgM. Antigenët jo T-vartes si aktivatori poliklonal endotoksina lipopolisakaridike induktojnë sintezën e IgM bashkë me pak IgG2 dhe IgG3.
Kalimi nga një klasë antitrupash në një tjetër për të formuar antitrupa me të njëjtin specificitet për antigenin por të klasave të ndryshme realizohet me ndihmën e një rekombinaze e cila përdor sekuenca "switch" të specializuara në ADN të cilat gjënden para cdo ekzoni që kodon për një pjesë konstante të një klasë Ig. Kjo rekombinaze con në humbjen e lakut ADN të ndërmjetme midis izotipit ekzistues dhe atij të ri.
Gjatë këtij fenomeni të ndrrimit izotipik kemi një rritje të vazhdueshme të afinitetit të antitrupave në saje të ndryshimeve somatike në pjesën variabël të imunoglobulinave dhe në këtë mënyrë limfocitet B kujtesë kanë një afinitet me të lartë për antigenin se sa limfocitet B të virgjer që ishin të parët të stimuluar nga ky antigen.
Duhet theksuar që përgjigja imunitere ndaj Antigenëve timojovartes të cilët kanë një kujtesë të varfër me mutacione të rralla somatike nuk paraqet karakteristiken e mesipërme të maturimit të afinitetit.
Ndërsa aftësia e limfociteve T ndihmës të lehtësojnë përgjigjen ndaj antigenëve T vartës (kryesisht antigenë proteinike me strukturë jo të polimerizuar dhe mjaft të komplikuar), të induktojnë proliferim klonal ekspansiv, të nxisin ndrrimin izotipik dhe se fundi të rrisin afinitetin e antitrupave e ka pajisur sistemin imunitar me një përgjigje imune me të fuqishme, me fleksibel dhe me të sakte.
Qelizat kujtesë: Antitrupat e limfociteve B të virgjër që janë të koduar nga gjenet germinale (germ-line)
paraqesin një formë të evoluimit filogjenetik të kujtesës imunitare në kuptimin që ato përfaqesojnë specificitete për patogjenet që takohën me shpesh dhe që përbëjne të ashtuquajturin fraksion të antitrupave natyrale të serumit. Kujtesa e fituar gjatë përgjigjes imune adoptive (te fituar) kërkon kontakt me antigenin dhe ekspansion të qelizave kujtesë antigen specifike. Këto qeliza janë jetë-gjatë, por megjithate ruajtja e këtyre qelizave është një gjëndje dinamike për të cilen është e nevojshme edhe stimulimi antigenik i herepashershem që ndodh nga qelizat dentritike folikulare të qendrave gjerminative që me sa duket janë depozituesit afatgjatë të antigenit.

54
Edhe qelizat T kujtesë gjithashtu varen nga stimulimi i herëpashershëm i antigenit dhe nëqoftese pranojmë që antigeni zakonisht persiston si kompleks në qelizat dentritike folikulare, atëhere edhe limfocitet T stimulohen me sa duket nga limfocitet B që përpunojnë antigenin përpara se t'ja paraqesin atyre. Edhe receptorët për antigenin në limfocitet T kujtesë kanë një afinitet me të lartë për antigenin por kjo nuk vjen nga mutacionet somatike në përgjigjen parësore. Me sa duket qelizat T naive me receptorë me afinitet me të lartë proliferojnë në mënyrë selektive si rrjedhojë e lidhjes preferenciale me antigenin. Limfocitet T kujtesë e rrisin gjithashtu aviditëtin e tyre lidhës për qelizën antigen paraqitëse nëpërmjet një rritje të shprehjes se molekulave aksesore të adezionit si CD2, LFA-1, LFA-3 dhe ICAM-1. Meqënëse disa nga këto molekula shërbejnë gjithashtu për të nxitur kalimin e sinjalit brënda në qelizë kjo shpjegon pse limfocitet T kujtesë stimulohen më shpejt se limfocitet T naive.
Një shënues interesant i cili diferencon limfocitet T naive nga ata kujtesë është antigeni i zakonshëm leukocitar CD45R. Forma CD45RA e këtij antigeni përcakton limfocitet T naive ndërsa forma CD45RO identifikon limfocitet T kujtesë të aftë të përgjigjen ndaj stimulimit dytësor antigenik. Gjithashtu limfocitet T CD45RO(+) kanë dhe karakteristika të tjera të qelizave të aktivizuara si p.sh. shprehjen receptorëve të "homing" për endoteliumin e vatrave të inflamacionit e cila u jep atyre një mundesi me të madhe për të takuar Antigenët që depërtojnë në organizëm. Nga ana tjetër ekziston mendimi se limfocitet T kujtesë CD45RO(+) në mungesë të stimulit antigenik mund të kthehen në formën RA dhe të humbin statusin e tyre të aktivizuar për t'u kthyer në limfociteT në gjëndje qetësie. në një depërtim tjetër të antigenit limfociti T kujtesë në gjëndje qetësie mund të transformohet në limfocit T kujtesë CD45RA(+) të aktivizuar dhe me tej në limfocit T efektor.
Limfocitet B të virgjera pas aktivizimit nga antigeni e humbin IgM dhe IgD në sipërfaqen e tyre dhe pësojnë ndrrimin izotipik me shprehje të IgG, A ose E në sipërfaqe, duke ruajtur gjithmone të njëjtin specificitet për antigenin.
Ontogjenia e përgjigjes imunitare: Limfonodulët dhe shpretka janë relativisht të pazhvilluar të neonati vecse nqs. fetusi i është
ekspozuar në uterus infeksioneve kongjenitale si rubeola, citomegalovirusi, toksoplazmoza etj. Aftësia për të flakur transplantet apo për të realizuar një përgjigje antitrupore është relativisht e mirë të neonati megjithëse niveli i imunoglobulinave të ky është mjaft i ulët (sigurisht në mungesë të infeksioneve intrauterine). Këtu bën përjashtim IgG që e kalon placentën nga nëna tek fetusi dhe tek ky i fundit ndodhet në nivelet e nënes(10-12g/L). Ky nivel bie në 3 muajt e parë dhe me pas fillon sinteza e IgG të vetë fëmijes që e arrijnë nivelin e 60-80% të te rriturit rreth moshes 1 vjec. IgM në moshen 1 vjec e kapin në përgjithësi 60-90% të nivelit të te rriturit, ndërsa IgA në këtë moshe janë 30-50% e nivelit të te rriturit. Ato e arrijnë nivelin e të rriturit pas moshes 10 vjec. Po kestu dhe IgE e arrin nivelin ete rriturit pas kësaj moshe me përjashtim të rasteve të atopise kur mund të kemi rritje ter saj edhe në moshat e vogla. Duke patur parasysh nivelet e ndryshme të imunoglobulinave në lidhje me moshen kjo duhet të kihet parasysh në interpretimin e nivelit të tyre.

55
KAPITULLI 12. NJOHURI MBI MEKANIZMAT E KONTROLLIT NË PËRGJIGJEN IMUNE.
Roli i antigenit, antitrupave, limfociteve T ndihmës. C'jane limfocitet supresorë llojet e tyre, mekanizmat e veprimit. Hallkat e tjera të imunosupresionit (rrjeta idiotipike, makrofagët etj.). Influenca e faktorëve gjenetike. Faktorë të tjerë.
Roli i antigenit në kontrollin e përgjigjes imune: Antigeni është një faktor i rëndësishëm rregullues i përgjigjes imune sepse me rënjen e
përqendrimit të antigenit kemi dhe një ulje paralele të përqendrimit të antitrupave ndaj tij për arsyen e thjeshtë se pakësohet stimuli kryesor për prodhimin e tyre, stimuli antigenik. P.sh. eliminimi i shpejtë i antigenëve me anë të injektimit me shumicë të antitrupave gjatë përgjigjes imune con në një ulje të shprehur të prodhimit të antitrupave si dhe të qelizave prodhuese të tyre.
Prania e një antigeni në një përzjerje antigenike mund të ulë në mënyrë drastike përgjigjen ndaj antigenëve të tjerë. Kjo është e vërtete edhe për epitope të ndryshëm brënda të njëjtes molekulë. P.sh. përgjigja imunitare ndaj pjesës Fab të molekulës se imunoglobulines G është më e madhe kur Fab injëktohet e vecuar se sa kur injëktohet molekula e tërë e IgG. Pra në këtë rast kemi efekt inhibues të fragmentit Fc të IgG. Mekanizmi i këtij efekti inhibues qëndron në kompeticionin e peptideve antigenike të procesuara për gropëzen e molekulës se KMPI. Disa përcaktues antigenike (peptide të procesuara) mund të kene një afinitet me të lartë për gropëzen e KMPI se të tjerë.
Antitrupi ushtron një kontroll "feed-back": Kështu p.sh. injektimi i antitrupave anti-D nënave të rrezikuara nga imunizimi anti-Rhesus i
mbron ato nga ky imunizim. Heqja e antitrupave nga qarkullimi i gjakut me anë të plazmaferezës gjatë një përgjigje antitrupore mund të coje në rritjen e sintezës se antitrupave përkates, ndërsa injektimi i IgG të gatshme nga jashtë ul numrin e qelizave formuese të antitrupave. Mendohet se ky rol frenues realizohet nëpërmjet bashkëveprimit të Fc të IgG me receptorin përkates (Fc gama R) në sipërfaqen e limfocitit B.
Në të kundërt me këtë mekanizëm injektimi i antitrupave të klasës IgM e nxit përgjigjen nga ana e limfociteve B, pasi kjo klasë nuk bashkëvepron me receptorin për Fc gama, por bën lidhje të kryqezuar të antigenit të lidhur me IgM sipërfaqesore. Meqënëse antitrupat e klasës IgM formohen në fillim të përgjigjes imune ato luajnë pra një rol nxites të prodhimit të antitrupave, ndërsa IgG që prodhohet me vone luan rol inhibues.
Roli i limfociteve T në rregullimin e përgjigjes imune: Fenomeni i supresionit të përgjigjes imune me anë të limfociteve T u vu në dukje me anë të
fenomenit të quajtur "tolerancë infeksioze": Nqs. te miu induktohet mos përgjigje imunitare ndaj eritrociteve të dashit me anë të injektimit të eritrociteve me doza të larta, atëherë ky supresion mund të transferohet në një mi tjetër me anë të limfociteve T. Eksperimentet e metejshme vune në dukje që supresioni me limfocite T vepron mbi mjaft përgjigje imune humorale apo qelizore. Në këto të fundit përmendim hiperndjeshmerine e tipit të vonuar, limfocitet T citotoksikë dhe proliferimin antigen-specifik të limfociteve T.
Ne mënyrë eksperimentale është vënë gjithashtu në dukje se epitope të ndryshme të një antigeni mund të kene disa funksion supresor dhe të tjerë funksion stimulues të përgjigjes imune. Qelizat T helper dhe ato supresorë mund të bashkëveprojnë me njëra tjetren duke u gjëndur prane njëra tjetres në sipërfaqen e qelizës antigen paraqitëse, e cila paraqet si epitopin helper edhe atë supresor.

56
Antigjeni
Epitopisupresor
Epitopi helper
procesim
Ts
Th
qeliza antigjen paraqitese
KMPI
Mekanizmat imunosupresore Antigen / Ts / Th
Mjaft shembuj eksperimentale kanë vënë në dukje në fillim të mijtë dhe pastaj të njeriu se disa alotipe të KMPI të caktuar japin ndaj një antigeni të dhene me tepër një përgjigje imune supresorë se sa stimuluese. Pra, me sa duket ky alotip ka afinitet të lartë për epitopet imunosupresorë të këtij antigeni.
Qelizat vepruese të supresionit: Ne mjaft sisteme ekspërimentale qelizat që ndërmjetësojnë supresionin janë shumë me të
ndjeshme se limfocitet T helper ndaj timektomise në moshe të rritur (miu), irradimit me rreze X apo ciklofosfamidit. Analiza e përgjigjes imunitare ndaj një sërë antigenësh në popullata humane ka vënë në dukje se përgjigja e ulët varet në mënyrë dominante nga antigenët e klasës II të KMPI. In vitro, deplecioni (varfërimi) i popullatave limfocitare nga qelizat CD8(+) rrit përgjigjen imunitare ndaj antigenit. Pra qelizat efektorë të supresionit specifik për antigenin duket se janë CD8(+) dhe njohin KL II të KMPI. Nga ana tjetër klone limfocitare të tilla prodhojnë citokina të tipit Th2, sidomos Il 4, ndërkohë që limfocitet T citotoksikë, edhe këto CD8(+), prodhojnë kryesisht IFN gama, pra një profil citokinash të ngjashem me tipin Th1. Pra, kjo sugjeron se ndarja Th1 /Th2 është e vlefshme edhe për limfocitet CD8(+).
Menyra e veprimit të limfociteve T supresorë: Faktet e shtruara me sipër vënë në dukje konkluzionin e mundshem se frenimi i L.T. H1
ndodh nëpërmjet L.T. CD8(+) të tipit 2 që prodhojnë IL-4, ndërsa supresioni i L.T. H2 kryhet nga limfocitet T citolitike CD8(+) prodhuese të interferonit gama. Të gjitha këto qeliza ndërveprojnë në sipërfaqen e qelizës antigen-paraqitëse.

57
Qeliza antigjen-paraqitese
_ _ _ _ _ _
_ _ _ _ _ _
TCR KMPI Epitopi helper Epitopi supresor
CD8T 2
CD8T1
IL-4 etj. IFN- gama
IFN gama
IL-4/10
Limfociti T citotoksikLimfociti T supresor "profesional"?
?
TH1TH2
Antagonizmi i ndërsjellte midis nënpopullatave të limfociteve T të lidhur indirekt nga antigeni i përpunuar nga qeliza antigen - paraqitëse con në menyra të ndryshme supresioni. Nuk është akoma e qarte nqs. midis nënpopullatave CD8 T1 dhe T2 kemi një rregullim të ngjashem si midis popullatave të tjera.
Sipas modelit të mesipërm, kur përgjigja imunitare është orientuar në një drejtim të vecantë p.sh. drejt imunitetit me ndërmjetësi qelizore Th1, tipet e tjera të përgjigjeve si p.sh. bashkëpunimi T-B janë të restriktuara nëpërmjet një efekti inhibues nga citokinat që prodhohen nga limfocitet Th1 si p.sh. IFN gama. Megjithëse këto qeliza kryejnë në këtë mënyrë një efekt T-supresor ato nuk mund të quhen qeliza supresorë "profesionale" sepse aftësia supresorë e tyre është një produkt anesor i funksionit të tyre kryesor që është ai mbrojtës. Ngelet për t'u vërtetuar nese limfocitet CD8(+) të tipit 2 kanë vetëm funksion supresor apo edhe një rol tjetër mbrojtës. Ndofta këto qeliza janë të nevojshme për të parandaluar që qelizat Th1 të dalin jashtë kontrollit me një proliferim të tepërt ashtu sic IgG frenon qelizat B me anë të një kontrolli feed-back.
Ne fakt ka mjaft të dhëna ekspërimentale që vejne në dukje që qelizat supresorë induktohen nga vetë limfocitet Th1.
Rrezatimi me rreze UVB inaktivizon qelizat dentritike dhe stimulon supresionin T-vartes. IL-2 dhe IFN gama frenohen ndërsa rritet prodhimi i IL-4.
Rrjeta idiotipike: Fragmentet e ndryshueshme në molekulën e imunoglobulines, të cilët formojnë vendin kombinues me antigenin mund të provokojnë vetë përgjigje imunitare (përcaktues antigenike idiotipike) dhe të nxisin formimin e antitrupave specifike për to (anti-idiotipe). Kjo mund të ndodhe brënda të njëjtit individ dhe në këtë rast kemi auto-anti-idiotipe. në një individ mund të kemi qindra dhe mijra në mos me tepër idiotipe. në këtë mënyrë kjo shumëllojshmeri idiotipike mund të reflektoje brënda organizmit shumëllojshmerine e antigenëve të jashtëm. Pra, nqs. limfocitet janë të aftë të njohin një diapazon të gjërë përcaktuesish antigenike të huaj ato mund të njohin gjithashtu antiidiotipet në sipërfaqen e limfociteve të tjerë duke krijuar të ashtuquajturen "rrjetë idiotipike". Kjo teori e hartuar nga Jerne është vërtetuar sot. Auto-anti-idiotipi mund të quhet si "imazhi i brëndshem i antigenit". Auto-anti-idiotipet janë vënë re që në jetën fetale, pra për formimin e tyre nuk është e nevojshme stimulimi antigenik. Këto limfocite B që kanë specificitete auto-antiidiotipike janë të shumte në fillim të zhvillimit ontogjenik (pakesohen me pas me rritjen e organizmit) dhe janë kryesisht limfocite B që bartin shënuesin CD5. Me sa duket këto qeliza janë të destinuara të

58
sintetizojnë specificitete antigenike antitrupore të tipit germinal dhe të kontribuojnë në formimin e të ashtuquajtureve antitrupa natyrore të cilët janë të pranishem edhe në kafshe aksenike (te rritur në kushte sterile), pra pa stimulim të jashtëm antigenik.. Kjo rrjetë idiotipike është vënë në dukje edhe për limfocitet T. Mendohet se një numër i mirë i limfociteve T auto-antiidiotipike kanë një funksion supresor "natyror" të përgjigjes imune.
Nga ana tjetër mendohet se këto limfocite B CD5(+) prodhojnë antitrupa natyrore germinale (kryesisht IgM) që kanë specificitet për Antigenët me të shprehur në natyre si p.sh. fosforil-kolinën e polisakarideve bakteriale. Por gjithashtu ata kanë afinitet të ulur edhe ndaj një sërë autoAntigenësh si ADN, IgG, proteinat e shokut të nxehte, apo elemente të citoskeletit.
Mendohet se rrjeta idiotipike jo vetëm ka një rol të rëndësishëm rregullues të përgjigjes imune, por edhe mbajtes të kujtesës imunitare me limfocitet B kujtesë.
Influenca e faktorëve gjenetike në kontrollin e përgjigjes imune: Jane me tepër se 10 gjene të ndryshëm që ndikojnë në përgjigjen e përgjithshme imunitare.
Kjo u vu në dukje në mënyrë ekspërimentale duke seleksionuar një popullatë të përzier minjsh gjatë me tepër se 10 gjenerata në dy grupe kryesore, një me përgjigje të ulët antitrupore dhe tjetri me përgjigje të lartë ndaj një varieteti Antigenësh (Ekspërimenti i Biozzi). Kjo realizohet duke kryqezuar me njëri tjetrin vetëm ata me përgjigje të lartë ndërsa ata me përgjigje të ulët kryqezohen gjithashtu vetëm me njëri tjetrin. Ndërmjet geneve përgjegjës, disa ndikonin në një proliferim dhe diferencim me të theksuar të limfociteve B, ndërsa disa të tjerë ndikonin në aktivitetin e makrofagëve. të dy linjat e mijve ishin të ngjashem në aftësine për të fagocituar pjesëzat e karbonit ose eritrocitet e dashit me anë të fagocitozës, por makrofagët e atyre me përgjigje me të forte e paraqesnin në mënyrë me efikase antigenin. Por nga ana tjetër ata me përgjigje të ulët antitrupore e kalonin me mirë infeksionin me Salmonella typhimurium dhe makrofagët e tyre lejonin një shumëzim shumë me të ngadalshem të Listeria. Këto fakte flasin për një aftësi mikrobicidale me të theksuar të grupit të kafsheve me përgjigje të ulët antitrupore.
Influenca e geneve të imunoglobulinave: Pa dyshim, mekanizmi gjenerues i shumëllojshmerise se përgjigjes antitrupore është aq i
fuqishem sa është mjaft e veshtire të kemi mangësi imunitare si pasojë e një repërtori të manget gjenesh V. Por megjithate janë vënë në dukje edhe mangësi të këtij repërtori si pasojë e mungesës se një geni V: P.sh. mangësia për t'iu përgjegjur sheqerit polimer dekstran alfa1-6 është një vecori e kafsheve pa gen V dex. Nga ana tjetër mijtë me mangësi të genit Valfa2 për receptorin TCR përkates nuk mund të gjenerojnë përgjigje me limfocite T citotoksikë ndaj antigenit mashkullor (H-Y).
Përgjigja imunitare influencohet nga KMPI: Ne mënyrë eksperimentale është arritur të vihet në dukje që përgjigja antitrupore ndaj një
numri substancash të thjeshta me përgjigje timovartëse kodohet nga gjene brënda KMPI. P.sh. mijtë e haplotipit H-2b te KMPI përgjigjen mirë ndaj polipeptidit sintetik (T,G)-A-L, ndërsa mijtë H-2k përgjigjen jo mirë. Ndërsa ndaj antigenit tjetër sintetik (H,G)-A-L ku ndryshon vetëm histidina në vend të tirozines përgjigja është e kundërt, mijtë e haplotipit H-2b përgjigjen jo mirë ndërsa ata H-2k përgjigjen mirë. Këta dy lloj mijsh janë gjenetikisht identike me njëri tjetrin me përjashtim të gjeneve të KMPI. Këta gjene përgjegjës për përgjigjen imunitare u quajten gjene Ir që komandojnë përgjigjen imunitare ndaj një përcaktuesi antigenik (epitopi) të vecantë. Ata janë të vecantë nga genet që kontrollojnë përgjigjen imunitare në përgjithësi dhe që u vune në dukje me ekspërimentet e Biozzi-t. Mendohet se genet Ir të KMPI ndodhen në klasën II të saj dhe komandojnë përgjigjen imunitare me ndërmjetësine e limfociteve T. Kjo mund të ndodhe me anë tre mekanizmave të ndryshëm:
1. Paraqitje defektive: një përcaktues antigenik (epitop) mund të mos ketë afinitetin e nevojshëm për t'u lidhur me molekulat e KMPI të një individi të caktuar dhe si pasojë kemi mungesë të stimulimit të limfociteve T të tij nga ky antigen. 2. Repertor deficitar i limfociteve T: është i pranuar tani fakti që limfocitet T me afinitet të mesem dhe të lartë për molekulat e KMPI të vetës (self) si dhe për komplekset e këtyre me autoAntigenë të procesuar eliminohen në timus duke krijuar një "boshllek" në repërtorin e limfociteve T. Nëqoftëse kemi një reaksion të kryqezuar si rrjedhojë e ngjashmerise strukturale midis një autoantigeni dhe një antigeni të huaj atëherë do të mungojnë në organizmin bujtës limfocitet T me TCR specifike për antigenin e huaj. Si pasojë ky individ do të jetë

59
një "reagues i dobët " ndaj këtij antigeni të huaj. 3. Supresioni me anë të limfociteve T supresorë: Statusi i një "reaguesi të dobët" mund të përcaktohet nga aktiviteti i qelizave T supresorë CD8(+). Përgjigja e ulët në këtë rast është dominante për heterozigotet në klasën II të KMPI, gje që tregon se supresioni mund të veproje ndaj limfociteve Th të restriktuar me cdo lloj molekulë të KMPI kl II. në këtë drejtim ai ndryshon nga modelet 1 dhe 2 ku në një heterozigot është përgjigja e lartë ajo dominante.
MfPerpunim i antigenit
Fuqia mikrobicidale
Genet e mijve"Biozzi"
Genet e lidhura meKMPI
Ig.
T
BProliferimDiferencim
Genet e lidhura me
Kontrolli gjenetik i përgjigjes imune.
Ndikimi i faktorëve neuroendokrine në proceset imune: Qelizat imune kanë receptorë ndaj një game të gjërë hormonesh si kortikosteroidet, insulina,
hormonet e rritjes, estradioli, testosteroni, agjentët beta-adrenergjike, acetilkolina, endorfinat dhe enkefalinat. në përgjithësi androgjenet dhe kortikosteroidet e frenojnë përgjigjet imune ndërsa estrogjenet, hormonet e rritjes, tiroksina dhe insulina bejne të kundërten. Sekrecioni i glukokortikoideve është një përgjigje madhore ndaj streseve të induktuar nga një game e gjërë stimujsh si ndryshimet e medha të tempëratures, frika, uria, dhe dëmtimet fizike. Ato sekretohen gjithashtu edhe si pasojë e përgjigjeve imune dhe i kufizojnë këto të fundit. Ka të dhëna ekspërimentale sipas të cilave glukokortikoidet suprimojnë limfocitet Th1 dhe nxisin ato Th2. Individët me një predispozicion gjenetik ndaj niveleve të larta të kortikoideve të prodhuara nga streset duhet të kene një prekshmeri me të lartë ndaj patogjeneve intraqelizore si psh. Mykobakteriet mbrojtja ndaj të cileve realizohet me anë të limfociteve Th1.
Estrogjenet janë gjithashtu një faktor q influencon në përgjigjen në përgjigjen imune me aktive në femrat se sa në meshkujt. Kështu femrat kanë nivel serik me të lartë të imunoglobulinave apo të IgA sekretore se meshkujt. Ato kanë gjithashtu një përgjigje antitrupore me të lartë ndaj Antigenëve T-indipendente, një rezistence relative ndaj tolerancës me limfocite T si dhe një rezistence me të madhe ndaj infeksioneve. Por femrat jame me të prekshme nga sëmundjet autoimune dhe kontraceptivet orale psh. mund të induktojnë akutizim të një sëmundje autoimune si Lupusi eritematoz sistemik (LES) p.sh.
Edhe sistemi nervor mund të ndikoje mbi sistemin imun: në një ekspëriment që ve në dukje këtë fakt mijtë morren ciklofosfamid si trajtim jokushtezues dhe sakarine si trajtim kushtezues (për të provokuar një reaksion të kushtezuar të tipit Pavlovian). Si pasojë përgjigja e ulët antitrupore u vu re edhe kur kafsha u stimulua me antigen bashkë me stimulin e kushtezuar (sakarinën).
Nga ana tjetër edhe IL-1 është e aftë të stimuloje prodhimin e glukokortikoideve. Gjithashtu duket se ekziston një ndërveprim midis inflamacionit dhe rritjes nervore në vendet e cikatrizimit të plageve. në kulturat e nervit shiatik kemi grumbullim të mastociteve, induktim nga IL-6 të rritjes

60
neurale dhe IL-1 nxit nga ana e saj prodhimin e faktorit të rritjes nervore. IL-1 nxit gjumin me vale të ngadalta dhe bashkë me IFN ka efekt pirogjen duke vepruar në qendrat e kontrollit të temperatures.
Veprimet e dietes, aktivitetit fizik, traumave dhe moshes mbi imunitetin: Prekshmeria e rritur e individëve të kequshqyer ndaj infeksioneve mund t'i atribuohet mjaft
faktorëve: kushtet higjenike të ulta, mbipopullimi në ambiente banimi të kufizuara dhe edukata e papërshtatshme shendetsore. Por kequshquerja proteino-kalorike vepron gjithashtu direkt mbi sistemin imunitar. Atrofia e gjërë e sistemit limfoid si dhe ulja në 50% e limfociteve T CD4 (+) qarkullues shoqërohet me dëmtim serioz të imunitetit me ndërmjetësi qelizore. Përgjigjet antitrupore mund të jenë të paprekura por të një afinitti të ulur. Fagocitoza bakteriale është relativisht normale, por baktericidia intraqelizore është e manget.
Mangësite në piridoksine, acid folik, si dhe vitaminat A, C, E cojnë në përgjithësi në një dëmtim të përgjigjes imune. Vitamina D është një rregullator i rëndësishëm. Ajo prodhohet jo vetëm nga lekura e rrezatuar me rreze ultraviolet, por gjithashtu edhe nga makrofagë të aktivizuar. Kjo vitamine është një inhibitor i fuqishem i proliferimit të limfociteve T dhe i prodhimit të citokinave nga limfocitet Th1. Mangësia në zink mund të shoqërohet me ulje të aktivitetit të hormoneve timike dhe rrjedhimisht edhe në imunitetin me ndërmjetësi qelizore.
Nga ana tjetër njihet që pakesimi i moderuar i marrjes se kalorive në teresi si dhe pakesimi i shprehur i marrjes se yndyrnave përmirëson sëmundjet e lidhura me moshen si p.sh. autoimunitetin. Vajrat me një lidhje n-3 të dyfishte si p.sh. vajrat e peshkut janë gjithashtu mbrojtës ndofta si pasojë e një sintezë të shtuar të prostaglandinave imunosupresorë.
Sforcimet fizike dhe sidomos ato me të rënda induktojnë stres dhe dhe rrisin nivelet plazmatike të kortizolit, katekolaminave, IFN alfa, IL-1, beta-endorfinave dhe metenkefalinave. Kjo mund të coje në ulje të nivelit të IgA, mangësi imunitare dhe prekshmeri të rritur ndaj infeksioneve. Po kështu dhe traumat multiple, kirurgjia dhe djegjet e medha janë gjithashtu imunosupresive dhe kështu mund të kontribuojnë në një rritje të rrezikut nga sepsisi.

61
KAPITULLI 13. TOLERANCA IMUNITARE
C'është toleranca imune:
Ka një vlere të madhe biologjike sepse me anë të asaj parandalohet ose evitohet reaksioni i sistemit imunitar ndaj përbërësve të vetvehtes. Me njohjen me të thelle të mekanizmave të tolerancës imune bëhet e mundur njohja me e mirë e fenomeneve të autoimunitetit dhe në përspektive gjetja e mjetëve me efikase për parandalimin dhe mjekimin e sëmundjeve autoimune. Toleranca imune është një fenomen kompleks që prek limfocitet T dhe ato B. Toleranca imune, si fenomen imun specifik është vënë në dukje që para 40 vjetësh me ekspërimentet e Owen. Ky konstatoi se në vicat binjake biovulare dhe që kishin një qarkullim placentar të përbashkët, qelizat e një binjaku nuk shkaktonin reaksion imunitar kur injëktoheshin tek binjaku tjetër pas lindjes. Injëktimi i qelizave në të njëjten mënyrë tek binjaku biovular por me qarkullim të ndare të placentave rezulton në përgjigje imunitare të rregullt me formim të antitrupave. Ky ekspëriment pra konstatoi se toleranca imune është një fenomen specifik që rezulton nga kontakti i ngushtë dhe i vazhdueshem i sistemit imunitar me Antigenët gjatë gjithë procesit të embriogjenezes. Burnet dhe Fenner nga ekspërimente të tjerë formuan konceptin që kontakti i Antigenëve me qelizat limfoide gjatë fazes se diferencimit të këtyre nga qeliza të papjekura në ato të pjekura, frenon cdo përgjigje të ardhshme ndaj këtyre Antigenëve kur organizmi e ka arritur pjekurine e plotë të sistemit të tij imunitar. Kjo do të ishte një mënyrë me anë të se ciles arrihet mos përgjigja imunitare ndaj përbërësve të vetvehtes dhe që bën të mundur që sistemi limfoid të dalloje vehten nga jo-vetja ("self nga "non-self"). Pra cdo antigen apo qelizë e huaj që futet në organizëm në përiudhen kur sistemi limfoid s'e ka arritur pjekurine e tij (kjo përiudhe intrauterine ndryshon nga lloji në lloj) do të bëjë që antigeni apo qeliza e huaj të konsiderohet si "self" dhe si rrjedhojë s'do të kemi përgjigje imunitare në një futje të tij kur sistemi imunitar është tashmë i pjekur.
Persistenca e antigenit në organizëm është e domosdoshme për ruajtjen e tolerancës për arësye se nqs. ai zhduket nga qarkullimi atëhere qelizat e reja imunokompetente që maturohen nuk janë më tolerante ndaj këtij antigeni dhe do të kemi si rrjedhojë përgjigje imunitare ndaj tij.
Induktimi i self tolerancës (tolerancës ndaj vehtes) në timus nëpërmjet delecionit klonal
intratimik: Eksperimentet e sotme me minj transgjenike kanë vënë në dukje se limfocitet T autoreaktive
eliminohen fizikisht në timus (pësojnë një seleksion negativ në të). Në fakt ky seleksion negativ paraprihet nga një seleksion pozitiv që ndodh nëpërmjet angazhimit të receptorit TCR për diferencimin e mëtejshëm të timociteve dyfish pozitive CD(4+)(8+). Ato klone që kanë afinitet për molekulat e KMPI (self) të shprehura në qelizat epiteliale kortikale të timusit ("nurse cells"= qeliza edukuese) do të kenë mundësi të shumëzohen dhe si rrjedhojë do të kemi seleksion pozitiv për ato klone që njohin KMPI (self). Mendohet se ky seleksion ndodh edhe në nivel të afinitetit të ulët midis TCR dhe KMPI. Si rrjedhojë timociti CD4(+) CD8(+) që ka afinitet për KMPI Kl I do të humbe CD4 dhe ai që ka afinitet për KMPI Kl II do të humbe CD8.
Përsa i përket seleksionit negativ ai ndodh si rrjedhojë e një sinjali të fortë (afiniteti të lartë) midis timociteve që kanë kaluar seleksionin pozitiv në korteks dhe nga ana tjetër makrofagëve kortiko-medulare dhe qelizave dentritike interdigitale që paraqesin KMPI + Autoantigenët. Pra në këtë mënyrë eliminohen ato timocite me TCR specifike (me afinitet të lartë) për KMPI + Antigenët (self).
Toleranca e limfociteve T si pasojë e anergjisë klonale: Një klon T-helper CD4 (+) i përgjigjet stimulimit nga peptidi antigenik respektiv i paraqitur
nga qeliza antigen-paraqitëse(QAP)me proliferim dhe sintezë të citokinave. Nqs. peptidi i shtohet një QAP të fiksuar, kloni helper bëhet jo-përgjigjes: njohja nga ana e tij e peptidit+KMPI nis një sinjal që con në mobilizimin e joneve të Ca por jo në aktivizim të Il-2. Arësyeja është se mungon në këtë rast sinjali I dytë i nevojshëm për stimulimin e limfocitit T helper (sinjali i parë është njohja e TCR

62
me antigenin + KMPI). Ky sinjal I dytë është CD28 (në limf. helper) që duhet të bashkëveproje me molekulën B7 në QAP si dhe cifti HSA + molekulë (akoma e panjohur e QAP). Mungesa e këtij sinjali co-stimulues (sic ndodh psh. në një QAP të fiksuar) do të sjellë në vend të stimulimit një anergji ose paralizë të limfocitit T helper. Kjo tolerancë është periferike dhe ndodh me limfocite T të pjekur. Anergjia përiferike T mund të ndodhe mund të ndodhe në nivele të ndryshme në vartësi të rrethanave të ekspozimit antigenik si dhe mund të paraqesë shkallë të ndryshme të reversibilitetit.
Toleranca imunitare mund të vijë edhe nga mungesa e komunikimit: Nqs. molekula self (autoantigeni) nuk bie në kontakt me TCR respektive atëherë nuk mund të
kemi përgjigje imunitare. Izolimi anatomik i antigenëve si psh. proteinat e kristalinit të syrit apo proteina bazike e mielines në tru pengon cdo përgjigje të mundshme imunitare ndaj këtyre autoantigenëve.
Edhe nqs. një ind është i ekspozuar ndaj limfociteve qarkullues mundet që përqendrimi i peptideve të përpunuara nga QAP (në mungesë në këto të fundit të molekulës ko-stimuluese B7) të jetë i pamjaftueshem për të terhequr vemendjen e një qelizë potencialisht autoreaktive. Kjo është vënë në dukje në mënyrë mjaft elegante në kafshë (minj) që kanë 2 transgjene: një për TCR e një L.T. citotoksik CD4(+) specifik për glikoproteinën virale të virusit LCM dhe tjetri për glikoproteinën virale vetë ( e shprehur kjo në qelizat beta të pankreasit nëpërmjet promotorit të insulines. Rezultati ishte heshtja imunitare e plotë. Limfocitet T as nuk ishin të eliminuara dhe as të anergjizuara. Po kështu dhe qelizat beta të pankreasit që shprehnin glikoproteinën virale të huaj nuk ishin të prekura. Nqs. këto minj infektohen me pas në mënyrë "natyrale" me virusin LCM, limfocitet T transgjenike vihen tashmë në kontakt me doza të larta të antigenit si pasojë kjo dhe e efektit "adjuvant" të infeksionit të vërtete dhe kemi stimulim të tyre. Pasardhesit e këtyre limfociteve të stimuluara kanë tashmë një aviditët të rritur ndaj antigenit dhe janë pra të aftë të njohin edhe përqendrimet e ulta të glikoproteines se përpunuar virale në qelizat beta të pankreasit dhe të sulmojnë këto qeliza (edhe në mungesë të molekulës ko-stimulatore B7 ) duke shkaktuar kështu diabet. Duket se ky mekanizem është shumë i rëndësishëm në gjenerimin e autoimunitetit.
Një rast tjetër të vecantë paraqesin autoantigenët që ndodhen në ato inde që normalisht nuk shprehin molekulat e klasës II të KMPI dhe si rrjedhojë nuk e kanë aftësine të eliminojnë ose të paralizojnë klonet e limfociteve CD4 (+) organ specifike.
Heshtje imunologjike mund të kemi edhe kur një individ nuk ka gene koduese për receptorë limfocitesh të drejtuara ndaj autoAntigenëve të vecantë. Analiza e përgjigjes autoimunitare ndaj citokromit, e induktuar në mënyrë ekspërimentale sugjeron që vetëm ato pjesë të molekulës që kanë variabilitet ndërmjet specieve janë autoantigenike ndërsa pjesët e molekulës se citokromit që janë të ruajtura nga një specie të tjetra nuk kanë reaktivitet antigenik sepse me sa duket limfocitet autoreaktive ndaj tyre janë zhdukur gjatë evolucionit.
Përsa i përket ndërhyrjes se limfociteve T supresorë në self-tolerancë kjo nuk është akoma mjaft e qarte, por duket se nqs. autoimuniteti induktohet në një kafshe normale ose aktivisht me një antigen që paraqet reaksion të kryqezuar me vehten, ose në mënyrë pasive me anë të injëktimit të limfociteve T autoreaktive, klonet autoreaktive zakonisht shtypen nga supresioni me qeliza T supresorë. Shkurt, limfocitet T supresorë probabilisht nuk e parandalojnë autoimunitetin por mund ta smbrapsin atë pasi ai cfaqet.
Induktimi i tolerancës në limfocitet B: Edhe për limfocitet B toleranca mund të realizohet nëpërmjet eleminimit klonal si dhe të
anergjise klonale. Me sa duket kur autoantigeni është i shprehur mjaft në qelizat që janë në kontakt me limfocitet B në maturim, do të kemi "cross-linking"(urezim) të sIg në sipërfaqen e limfociteve B dhe si rrjedhojë do të kemi eliminim të klonit respektiv.
Ne rast se shprehja e antigenit është e dobët dhe univalente nuk realizohet as eliminimi klonal dhe as stimulimi, por do të kemi fenomenin e anergjise klonale.
Toleranca mund të cfaqet edhe nga mungesa e ndihmës për limfocitet B:

63
Te pakten me proteina të tretshme limfocitet T mund të "tolerizohen" më shpejt se sa limfocitet B. në vartësi të përqendrimit të proteines qarkulluese mund të kemi në organizëm një numër limfocitesh B autoreaktive që nuk mund të stimulohen nga këta autoAntigenë timovartës për arësye se limfocitet T të nevojshëm për të realizuar ndihmën T-B respektive janë tashmë tolerante ndaj tyre. Pra, nqs. në kemi parasysh që një determinant i një autoantigeni bashkëvepron si një epitop haptenik me limfocitin B ndërsa një determinant tjetër si epitop bartës me limfocitin T, atëherë nqs. limfociti T është tolerant ndaj epitopit përkates ai nuk mund të realizoje funksionin e tij ndihmës dhe si rrjedhojë do të kemi mungesë përgjigje imunitare ndaj autoantigenit mgjs. limfocitet B specifike ndaj tij janë të pranishme.
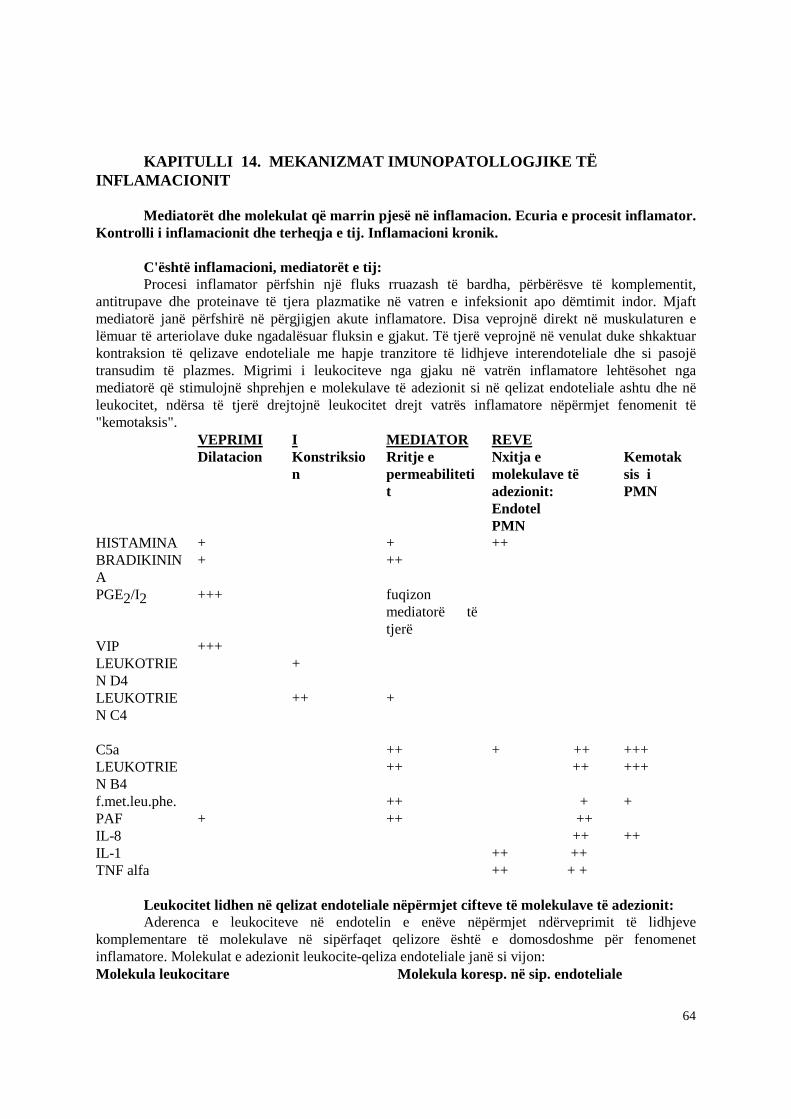
64
KAPITULLI 14. MEKANIZMAT IMUNOPATOLLOGJIKE TË
INFLAMACIONIT
Mediatorët dhe molekulat që marrin pjesë në inflamacion. Ecuria e procesit inflamator. Kontrolli i inflamacionit dhe terheqja e tij. Inflamacioni kronik.
C'është inflamacioni, mediatorët e tij: Procesi inflamator përfshin një fluks rruazash të bardha, përbërësve të komplementit,
antitrupave dhe proteinave të tjera plazmatike në vatren e infeksionit apo dëmtimit indor. Mjaft mediatorë janë përfshirë në përgjigjen akute inflamatore. Disa veprojnë direkt në muskulaturen e lëmuar të arteriolave duke ngadalësuar fluksin e gjakut. Të tjerë veprojnë në venulat duke shkaktuar kontraksion të qelizave endoteliale me hapje tranzitore të lidhjeve interendoteliale dhe si pasojë transudim të plazmes. Migrimi i leukociteve nga gjaku në vatrën inflamatore lehtësohet nga mediatorë që stimulojnë shprehjen e molekulave të adezionit si në qelizat endoteliale ashtu dhe në leukocitet, ndërsa të tjerë drejtojnë leukocitet drejt vatrës inflamatore nëpërmjet fenomenit të "kemotaksis". VEPRIMI I MEDIATOR REVE Dilatacion Konstriksio
n Rritje e permeabilitetit
Nxitja e molekulave të adezionit: Endotel PMN
Kemotaksis i PMN
HISTAMINA + + ++ BRADIKININA
+ ++
PGE2/I2 +++ fuqizon mediatorë të tjerë
VIP +++ LEUKOTRIEN D4
+
LEUKOTRIEN C4
++ +
C5a ++ + ++ +++ LEUKOTRIE N B4
++ ++ +++
f.met.leu.phe. ++ + + PAF + ++ ++ IL-8 ++ ++ IL-1 ++ ++ TNF alfa ++ + +
Leukocitet lidhen në qelizat endoteliale nëpërmjet cifteve të molekulave të adezionit: Aderenca e leukociteve në endotelin e enëve nëpërmjet ndërveprimit të lidhjeve
komplementare të molekulave në sipërfaqet qelizore është e domosdoshme për fenomenet inflamatore. Molekulat e adezionit leukocite-qeliza endoteliale janë si vijon: Molekula leukocitare Molekula koresp. në sip. endoteliale

65
ICAM-1 (CD54) supërfamilja e IgG
LFA-1
LFA-1(CD11a/18) beta2integrinat ICAM-1 P-SELEKTINA (CD62) familja e selektinave
Sialyl Lewis
LYMPH. HOM. MOL. (PROTEOGL. I SIP. QEL.)
Adresinat mukozale
LEUKOSIALINA (CD43)MUCINA E SIP. QEL.
CD62?
Fillimi i përgjigjes inflamatore akute: Një ngjarje e hershme është stimulimi i shprehjes se P-selektines dhe faktorit aktivizues të
trombociteve (PAF) në qelizat endoteliale të venulave nëpërmjet histamines apo trombines se cliruar nga stimuli inflamator fillestar. Mobilizimi i molekulave të adezionit nga vezikulat e ruajtjes intraqelizore shkakton që ato të shfaqen shpejt në sipërfaqen qelizore. Bashkëveprimi i domenit të ngjashem me lektinën në majen e molekulës se gjatë të P-Selektines me karbohidratin Sialyl Lewis x në mucinat e sipërfaqes se polinukleareve (mundesisht leukosialinat) shkakton që neutrofili të "rreshqase" gjatë parëtit endotelial dhe ndihmon që PAF të lidhet me receptorin përkates të tij. Kjo nga ana e saj rrit shprehjen sipërfaqesore të integrinave LFA-1 dhe MAC-1 të cilat e lidhin mjaft mirë neutrofilin me sieprfaqen e endotelit.
Aktivizimi i neutrofileve i bën ato me të ndjeshëm ndaj agjentëve kemotaktike dhe nën influencën e C5a dhe të leukotrienit B4, PMN del nga qarkullimi i gjakut duke levizur në hapesirat midis qelizave endoteliale si dhe membranës bazale (diapedeza) dhe duke ndjekur gradientin kemotaktik përfundon në vatren inflamatore.
Dëmtimi i endotelit vaskular i cili ekspozon membranën bazale si dhe endotoksinat apo lipopolisakaridet bakteriale nxisin sisteme të tjerë komplekse. Aktivizimi i trombociteve nga PAF ose kontakti me kolagjenin e membranës bazale con në formimin e trombeve. Aktivizimi i faktorëve të koagulimit nxit sistemet e kinines dhe të plazmines. Disa nga produktet që formohen influencojnë procesin inflamator duke rritur përmeabilitetin vaskular, duke aktivizuar endotelin, duke amplifikuar në mënyrë autokatalitike prodhimin e faktorit Hageman si dhe duke aktivizuar C3.
Vijimi i procesit inflamator: Ne këtë aspekt ka rëndësi roli i makrofagëve indore, të cilët nën stimulin e infeksionit apo
dëmtimit lokal sekretojnë një numër të konsiderueshem mediatorësh. në vecanti citokinat IL-11 dhe TNF- alfa veprojnë me vone se histamina ose trombina duke stimuluar qelizat endoteliale. një nga produktet e vonshme është E-selektina, një molekulë e adezionit që lidhet dhe aktivizon neutrofilet, dhe tjetra, IL-8, është një kemotaksine efikase për polinuklearet. IL-1 dhe TNF alfa veprojnë gjithashtu në qelizat endoteliale, fibroblastet dhe qelizat epiteliale duke sekretuar një proteine të fuqishme kemotaktike të monociteve, MCP-1, e cila terheq fagocitet mononukleare në vatren inflamatore duke forcuar dhe mbajtur kështu reaksionin mbrojtës ndaj infeksionit. në këto vatra qelizat endoteliale shprehin VCAM-1, e cila vepron si receptor "homing" për limfocitet T kujtesë të aktivizuar dhe që janë VLA-4 pozitiv.
I gjithë ky reaksion i tere shërbën për të fokusuar mbrojtjet imunitare rreth mikroorganizmave invadues. Këta mbulohen me antitrupa, C3b dhe disa proteina të fazes akute dhe pregatiten për fagocitozë nga ana e granulociteve dhe e makrofagëve. Nën influencën e mediatorëve të inflamacionit këta kanë shprehur tashmë receptorët për C3 dhe Ig.si dhe kanë rritur aktivitetin fagocitar dhe aftësine baktericide.
Kontrolli dhe rezolucioni i inflamacionit: Ne nivelin humoral kemi një sërë proteinash rregulluese të komplementit: C1 inhibitori,
proteina lidhëse e C4 (C4bp), proteinat kontrolluese të C3 (faktorët H dhe I), receptori i komplementit CR1, faktori akselerues i zbërthimit (DAF), proteina kofaktorë membranore (MCP),

66
dhe imunokonglutinina si dhe se fundi proteinat që bllokojnë kompleksin e sulmit membranor. Disa nga proteinat e fazes akute me origjinë nga transudati plazmatik veprojnë si inhibitore të proteazave.
Ne nivelin qelizor PGE2, TGFbeta dhe glukokortikoidet janë rregullatore të fuqishem. PGE2 është një inhibitor i fuqishem i proliferimit limfociter si dhe i prodhimit të citokinave nga limfocitet T dhe makrofagët.
TGF beta caktivizon makrofagët duke inhibuar prodhimin e ndërmjetëseve reaktive të oksigjenit dhe duke ulur shprehjen e KMPI të klasës sedytë. Ai gjithashtu frenon aktivitetin citotoksik të makrofagëve si dhe të qelizave NK të aktivizuara nga IFN gama.
Glukokortikoidet endogjene të prodhuara nga aksi hipotalamo-hipofizo-suprarenal e ushtrojnë veprimin e tyre antiinflamator probabilisht ndërmjet kontrollit të lipokortines 1 e cila lidhet në proteinat sipërfaqesore të monociteve dhe neutrofileve. IL-10 inhibon paraqitjen e antigenit, prodhimin e citokinave si dhe vrasjen NO-vartese nga makrofagët.
Një proteine tjetër kontrolli e inflamacionit është antagonisti i receptorit të IL-1 beta. Pas eliminimit të agjentit inflamator, këto procese rregulluese do të normalizojnë vatrën. TGF
beta luan një rol madhor në shërimin e plagës nëpërmjet stimulimit të ndarjes se fibroblasteve dhe rikrijimit të elementeve të reja ekstraqelizore të matriksit.
Inflamacioni kronik: Nëqoftëse agjenti inflamator përsiston për arësye të rezistences se tij ndaj katabolizmit ose
për arësye të pamundesise të sistemit imun të manget për të eliminuar një agjent mikrobial, karakteri i përgjigjes qelizore ndryshon. Vatra dominohet nga makrofagë me morfollogji të ndryshme: shumë kanë një pamje të aktivizuar, disa formojnë gershetime të quajtura qeliza epitelioide dhe të tjerë fuzionojnë me njëri tjetrin duke shkaktuar qeliza gjigante. Nqs. kemi gjithashtu edhe një përgjigje imune të fituar do të cfaqen edhe limfocite të llojeve të ndryshme. Kjo "granulomë" karakteristike do të përpiqet të izoloje agjentin persistues nga pjesa tjetër e trupit.
KAPITULLI 15. MEKANIZMAT IMUNOLOGJIKE TË MBROJTJES ANTIINFEKSIOZE NDAJ BAKTERIEVE EKSTRAQELIZORE.
Ndaj mikroorganizmave ekstraqelizore (me antigenë ekzogjene me lokalizim ekstraqelizor) mbrojtja realizohet kryesisht me anë të imunitetit humoral nëpërmjet ndërhyrjes se komplementit dhe të antitrupave. Bakteriet patogjene gjatë filogjenezes kanë zhvilluar një sërë mekanizmash me anë të të cilëve ato i shpëtojnë këtyre faktorëve:
Evitimi i fagocitozës: Paretet qelizore të bakterieve patogjene janë komplekse dhe në mjaft raste rezistente ndaj
mjaft mekanizmave mikrobicidale. Një faktor i zakonshëm me ndihmën e të cilit format virulente i shpetojnë fagocitozës është sinteza e një kapsule të jashtme e cila pengon aderencen në qelizat fagocitare si dhe mbulon në sipërfaqen bakteriale molekula polisakaridesh të cilat përndryshe do të njiheshin nga receptorët e fagociteve. P.sh. edhe 10 pneumokoke të inkapsuluar mund të vrasin një mi, ndërsa nqs. kapsula hiqet me anë të hialuronidazes atëherë nevojiten 10000 bakterie për këtë qellim. Disa mikroorganizma sekretojnë ekzotoksina që "helmojnë" leukocitet.
Sfidimi i sistemit komplementar: Qelizat normale të gjitareve janë të mbrojtura nga komplementi me anë të proteinave
rregulluese si CR1, MCR, dhe DAF të cilat realizojnë shkaterrim të C3 konvertazes. Bakteriet nuk i kanë këto mekanizma rregullatore dhe si rrjedhim edhe në mungesë të antitrupave shumica e tyre aktivizojnë rrugën alterne të komplementit duke stabilizuar C3bBb konvertazen në sipërfaqet e tyre. Sidoqofte, kapsulat bakteriale në përgjithësi janë aktivizues të varfër të rrugës alterne të komplementit. Nga ana tjetër disa bakterie kanë aftësine të akselerojnë degradimin e C3 në sipërfaqen e tyre. Disa bakterie të tjera sekretojnë enzima që degradojnë peptide si C5a, i cili luan një rol të rëndësishëm në përgjigjen inflamatore akute. Disa specie bakteriale e evitojnë lizën duke devijuar vendin e aktivizimit të komplementit ose në një proteine të sekretuar ose në një pozicion të sipërfaqes bakteriale larg membranës qelizore.

67
Bakteriet Gram-pozitive bartin shtrese të trashe peptidoglikani e cila parandalon vendosjen e kompleksit të sulmit litik C5b-9 në paretin bakterial.
Kundërveprimi i organizmit: Mekanizmat mbrojtëse shfrytëzojnë specificitetin dhe variabilitetin e molekulës antitrupore.
Antitrupat neutralizojnë molekulat antifagocitare si psh ekzotoksinat dhe lidhen në sipërfaqen e mikroorganizmave duke fokusuar kështu fiksimin e komplementit, duke i opsonizuar ato për tretje nga fagocitet si dhe duke i përgatitur për sulmin terminal nga kompleksi i sulmit membranor të komplementit.
Bakterie të inkapsuluara, të cilat i rezistojnë fagocitozës bëhen mjaft tërheqëse ndaj polimorfonukleareve dhe makrofagëve kur mbulohen me antitrupa. Nënklasat e IgG që lidhen me fort me receptorët përkates në sipërfaqen e fagociteve (si IgG1 dhe IgG3 të njëriu) e fiksojnë gjithashtu mirë komplementin. Nga ana tjetër dhe C3b që aktivizohet në sipërfaqen e bakterit ka një rol të rëndësishëm opsonizues. Komplekset imune që përmbajne C3b shprehin një aderence imune në receptorët CR1 të komplementit në eritrocite duke formuar kështu agregate që transportohen me pas në melci për t'u fagocituar. Receptorët CR1 për C3b janë gjithashtu të pranishem në neutrofile, makrofagë, limfocitet B si dhe në qelizat folikulare dentritike në nyjet limfatike. Se bashku me receptorin CR3 ato kanë përgjegjësine kryesore për pastrimin nga komplekset imune që përmbajne C3. Receptorët CR2 për C3d, C3dg dhe iC3b janë të pranishem në limfocitet B si dhe qelizat folikulare dentritike dhe ato mund të përcojnë sinjale aktivizimi për limfocitet B vecanerisht në qendrat germinative. Afiniteti i tyre për virusin EB përbën menyren e futjes se virusit në limfocitin B.
Receptorët CR3 në polimorfonuklearet, makrofagët dhe qelizat NK lidhin formën e inaktivizuar të C3bi. Bashkë me LFA-1 dhe CR4 ata janë anëtarë të familjes së beta2integrinave.
Disa shtame të bakterieve Gram-negative, të cilat kanë një paret të jashtëm lipoproteinash janë të ndjeshme ndaj veprimit baktericidal të serumit të fresket që përmban antitrupa. Antitrupi fillon zhvillimin e një lezioni me ndërmjetësine e komplementit i cili nga ana e tij lejon veprimin e lizozimes serike në paretin e brëndshëm prej peptidoglikani të bakterit duke shkaktuar vdekjen e qelizës. Aktivizimi i komplementit nëpërmjet bashkëveprimit të antitrupit me bakterien do të gjenerojë prodhimin e anafilatoksinave C3a dhe C5a që cojnë në dalje ekstravazale të përbërësve të serumit duke përfshire këtu me tepër antitrupa, dhe duke nxitur migrimin kemotaktik të polimorfonukleareve gje që ndihmon fagocitozën.
Sistemi imun sekretor ndihmon mbrojtjen e mukozave të jashtme: Lidhja e viruseve dhe e bakterieve me qelizat epiteliale të mukozave është esenciale për
realizimin e patogjenicitetit të tyre. Antitrupat e klasës IgA përbëjne mbrojtjen specifike të sekrecioneve mukoze, lotëve, peshtymes, sekrecioneve nazale, zorres, bronkeve duke mbuluar bakteriet dhe viruset dhe duke parandaluar kështu aderencen e tyre në epitelin e mukozave. Gjithashtu, në makrofagët dhe polimorfonuklearet ekzistojnë receptorë me afinitet të lartë për IgA të cilët realizojnë fagocitozën e mikroorganizmave të mbuluar me IgA.
Nqs. një agjent infeksioz arrin të kaloje barrierën e IgA, ai bie në vijen tjetër të mbrojtjes të mukozave që përbëhet nga IgE. Eshtë për t'u shënuar që shumica e IgE serike vjen pikërisht nga plazmocitet e mukozave dhe e limfonodulave që i drenojnë ato. Megjithëse në përqendrime mjaft të ulta IgE është e lidhur fort me receptorët ndaj Fc përkatese në sipërfaqen e mastociteve. Kontakti me antigenin con në clirimin e mediatorëve të cilët mobilizojnë ndërmjetës të tjerë të përgjigjes imune duke gjeneruar si përfundim një reaksion inflamator akut. Kështu, histamina duke rritur përmeabilitetin vaskular shkakton daljen ekstravazale të IgG dhe të komplementit në vatren inflamatore, ndërsa faktorët kemotaktike të neutrofileve dhe eozinofileve terheqin qelizat efektorë të nevojshme për të eliminuar komplekset e formuar nga agjenti infeksioz të mbuluar me IgG dhe C3b. Lidhja e këtyre komplekseve me receptorët e Fc gama dhe C3b të makrofagëve lokale do të coje në sekretimin e peptideve të tjerë që do rrisin me tej përmeabilitetin vaskular dhe mobilizimin kemotaktik qelizor.
Nqs. mikroorganizmi i opsonizuar është tepër i madh për t'u fagocituar, vrasja e tij mund të realizohet nëpërmjet një mekanizmi ekstraqelizor që pason lidhjen e komplekseve imune me receptorët e fagociteve për Fc gama. Ky mekanizem i quajtur dhe citotoksicitet me ndërmjetësi

68
qelizore vartes nga antitrupat (angl. ADCC= antibody dependent cell-mediated cytotoxicity) ndërhyn në infeksionet parazitare.
Nga sa u tha me lart ekziston një paralelizëm i qarte midis rrugëve me të cilat anafilatoksinat me prejardhje komplementare nga njëra ane dhe IgE nga ana tjetër përdorin për të nxitur mastocitet drejt amplifikimit lokal të mbrojtjes imune nëpërmjet inflamacionit.
KAPITULLI 16. MBROJTJA IMUNE NDAJ BAKTERIEVE QE RRITEN NE
MJEDIS BRËNDAQELIZOR SI DHE NDAJ VIRUSEVE DHE PARAZITËVE.
Mbrojtja ndaj bakterieve me lokalizim brëndaqelizor: Disa lloje bakteriesh si psh. Mykobakteriumi i tuberkulozit dhe i lepres, brucela apo listeria
arrijnë t'i shpetojnë sistemit imunitar duke u strehuar pikërisht në makrofagët. Këta fagocite mononukleare janë shumë të përshtatshem për këto lloj mikroorganizmash sepse makrofagët janë mjaft të levizshem dhe me anë të tyre infeksioni mund të përhapet në të gjithë organizmin. Gelltitja e bakterieve të opsonizuara lehtësohet nga mbulimi i tyre me IgG dhe C3 fale receptorëve përkates. Pas futjes se tyre në makrofag ato e sfidojnë këtë duke devijuar aftësine baktericide të makrofagut në disa menyra. Mykobakteri i tuberkulozit p.sh. inhibon fuzionimin e lizozomeve me vakuolen fagocitare që përmban bakterin e fagocituar. Listeria monocitogene përdor një lizine të vecantë për t'i shpetuar fagolizozomit dhe për të qendruar lirisht në citoplazëm të fagocitit. Po kështu veprojnë dhe disa riketcie si dhe protozoari Trypanosoma cruzi. Legionelat nga ana e tyre inhibojnë shpërthimin respirator brëndaqelizor.
Mackaness vuri në dukje rëndësine e imunitetit me ndërmjetësi të qelizave T (INQ) për vrasjen e mikroorganizmave brëndaqelizore: Kafshet e infektuara me doza të moderuara të M. tuberculosis e përballojnë infeksionin dhe bëhen imune ndaj futjeve të mëvonshme të këtij bacili. Ky imunitet mund të transferohet në një marrës normal tjetër të paimunizuar me anë të limfociteve T por jo të makrofagëve apo të serumit të një dhenesi të imunizuar.
Ne formën tuberkuloide të lepres kemi një gjëndje të mirë hipërndjeshmerie me ndërmjetësine e limfociteve T që vihet re me reaksionin pozitiv të vonur ndaj lepromines. Kemi pra një predominim të përgjigjes imunitare me L.T. të tipit TH1 megjithëse këta të fundit nuk arrijnë të crrenjosin sëmundjen. Si rrjedhojë kjo formë e sëmundjes karakterizohet nga lezione me vatra që përmbajne një numër të paket mikroorganizmash të gjalle. Forma lepromatoze karakterizohet nga lezione në të cilat kemi një numër të madh mikroorganizmash të gjalle brënda makrofagëve. në këtë formë kemi një reaktivitet të paket limfocitesh T ndaj këtyre bakterieve dhe si rrjedhojë edhe prova e

69
lepromines është negative. Ajo që mbizoteron në këtë formë është përgjigja me limfocitet T TH2 dhe si rrjedhojë kemi plazmocite të shumta me nivel të lartë antitrupash qarkullues që nuk kanë efekt mbrojtës sepse bakteriet janë brënda makrofagëve. Pra është e qarte që është INQ ai që luan rol mbrojtës ndaj M. Leprae dhe jo imuniteti humoral.
Kur monocitet kalojnë në inde të ndryshme për t'u transformuar në makrofagë "rezidente" ato janë në gjëndje qetësie si përsa i përket shprehjes se receptorëve sipërfaqesore ashtu dhe funksionit të tyre. Makrofagët mund të aktivizohen nga stimuj të ndryshëm. Makrofagët e vatrave inflamatore janë mjaft të rritur në madhësi, në përmbajtje me hidrolaza acide, në sekretim të proteazave neutrale si dhe në funksionin fagocitar.
Aftësia e këtyre makrofagëve inflamatore për të ushtruar vrasje të mikroorganizmave brëndaqelizore arrihet vetëm kur ata stimulohen në një stad tjetër aktivizimi kësaj rradhe nga faktorët e aktivizimit të makrofagëve që prodhohen nga limfocite T specifike të aktivizuar. Ndër këta faktorë dallohet Interferoni gama. Makrofagu i aktivizuar është pa dyshim një nga qelizat me të fuqishme të organizmit, e aftë për të kryer një sërë funksionesh mjaft të rëndësishme në inflamacion si vijon:
Inflamacion dhe tempërature: Prodhon IL-1, TNFalfa, IL-6; IFN beta, Leukotriene, Prostaglandina, Faktorë të komplementit, Faktorë të koagulimit.
Aktivizimi limfocitar: Përpunimi i antigenit, Paraqitja e antigenit, Prodhim i IL-1 Cikatrizimi indor: Faktorë të angiogenezes, Faktorë të fibrogenezes, Elastaze, Kolagjenaze,
Hialuronidaze. Dëmtim indor: Prodhim i H2O2, Hidrolazat acide, C3a. Aktivitet mikrobicidal: O2-vartes, O2- jo-vartes (NO, Lizozime, Hidrolaza acide, Proteinat
kationike, Laktoferrina). Aktivitet tumoricid: Citotoksicitet direkt, H2O2, C3a, Arginaze, Proteazat citolitike, TNF
alfa, NO-vartes. Mekanizmi i imunitetit me ndërmjetësi qelizore është tashmë i qarte. Specificiteti i kësaj
përgjigje qendron në nivel të limfociteve T specifike për antigenin i cili paraqitet i përpunuar në sipërfaqen e makrofagut të infektuar. Si pasojë e njohjes nga ana e kloneve të limfociteve Th1 të përcaktuesve antigenike specifike për to do të kemi clirimin e citokinave nga ana e këtyre limfociteve dhe si rrjedhojë aktivizimin e makrofagëve rajonale që fitojnë kështu aftësine të vrasin mikroorganizmat e fagocituar.
Por shpesh makrofagët e infektuar nuk arrijnë dot të realizojnë vetë vrasjen e mikroorganizmave që kanë fagocituar. në këtë rast ndërhyjne limfocitet citotoksikë CD8(+) dhe CD4(+) të cilët bashkë me qelizat NK (te aktivizuara nga IL2 e prodhuar nga L.T.CD4(+)) realizojnë vrasjen e këtyre makrofagëve "te paaftë". Mikroorganizmat e cliruar me pas nga shkaterrimi i këtyre makrofagëve fagocitohen dhe vriten nga makrofagë të rinj të mobilizuar rishtaz dhe të aktivizuar.
Duke patur parasysh këto mundesi potenciale të Imunitetit me ndërmjetësi qelizore lind pyetja përse disa individë nuk arrijnë të crrenjosin infeksionet me mikroorganizma brëndaqelizore si psh. Mykobakteriet e lepres apo tuberkulozit apo të tjerë megjithëse ato kanë një përgjigje të instaluar Th1. një përgjigje kësaj pyetje vjen nga demonstrimi në mijtë eksperimentale që lloje të ndryshme konsanguine të tyre ndryshojnë në mënyrë të shprehur nga njëri tjetri përsa i përket aftësise të tyre për të luftuar mikroorganizma intraqelizore si mykobakteriet, leishmaniat apo salmonelat. Kontrolli i rezistences apo prekshmerise nga këto infeksione u gjet se kontrollohet nga 2 alele të ndryshme të një geni të vetëm autozomal dominues. Gjëndja e rezistences është e lidhur me një gjëndje të aktivitetit të lartë baktericidik të makrofagut e pavarur kjo nga limfocitet T. Makrofagët e llojeve rezistente kanë një shprehje me të rritur të Antigenëve KMPI të Kl. II, një aktivitet respirator me të fuqishem (O2-vartes), aktivizohen me lehtë nga IFN gama dhe stimulojnë me mirë limfocitet T. Makrofagët e llojeve me të prekshem paraqesin një efekt supresor me të madh ndaj proliferimit të limfociteve T nga antigenët mykobakteriale.
Limfocitet T TCR1 (gama (+); delta (+)) kanë aftësine të stimulohen nga antigenët e mykobakterieve. Këto limfocite gjënden me një shpështësi të lartë në lezionet nga mykobakteriet dhe paraqesin aftësine të lizojnë qelizat shënjë që paraqesin antigent mykobakteriale.

70
Kur organizmi ka veshtiresi të eliminoje në mënyrë efikase mikroorganizma intraqelizore si mykobakteriet p.sh., përgjigja kronike imune me ndërmjetësi qelizore ndaj tyre do të coje në akumulimin e makrofagëve të dendur të cilët clirojnë faktorë angiogjenike dhe fibrogjenike dhe stimulojnë formimin e indit granulomatoz dhe se fundi të fibrozes. Makrofagët e aktivizuar nën stimulin ndofta dhe të IL-4 transformohen në qeliza epiteloide dhe fuzionojnë në qeliza gjigante. Granuloma që rezulton përfaqeson një tentative të organizmit për të izoluar vatren e inflamacionit kronik.
Imuniteti ndaj infeksioneve virale. Një roj dominues në mbrojtjen antivirale luajnë faktorë konstitucionale gjenetike të cilët e
bejne një organizëm (ose disa qeliza të tij) rezistente ndaj viruseve. P.sh. makrofagët mund t'i kapin dhe t'i vrasin viruset në mënyrë jo specifike. Megjithate, në disa raste makrofagu e lejon replikimin viral dhe nëqoftëse virusi është i aftë të shkaktoje efekt citopatik në organe të ndryshme, infeksioni mund të jetë letal. në rast se virusi nuk ka efekt citopatik kemi një infeksion përsistues.
Ndryshimi antigenik i virusit nëpërmjet "drift" dhe "shift": Gjatë dyluftimit të vazhdueshem të tyre me sistemin imun, viruset i ndryshojnë vazhdimisht
Antigenët sipërfaqesore të tyre. P.sh. në rastin e virusit të gripit ndryshimet minore në antigenicitetin e hemaglutinines (proteine sipërfaqesore virale nëpërmjet të ciles virusi kapet në qelizat që i infekton me pas) ndodhin nëpërmjet ndryshimeve pikesore të nukleotideve të genomes virale (drift), ndërsa ndryshime madhore (shift) realizohen nëpërmjet shkembimeve të materialit gjenetik me viruse të tjerë që infektojnë specie të tjera. Ndërsa "drift" nuk pengon imunitetin mbrojtës ekzistues, "shift-i" e ndryshon plotësisht imunogjenicitetin dhe si pasojë e gjithë popullata që ai infekton është e pambrojtur nga ky virus. në këtë mënyrë shpjegohen edhe pandemite e medha nga gripi të shoqëruara me forma të rënda vdekjeprurëse.
Format mutante të viruseve mund të favorizohen edhe nga presioni selektiv i antitrupave. në këtë princip bazohet dhe variacioni karakteristik i rinoviruseve, shkaktare të rrufave sezonale.
Antitrupat qarkullues përbëjne gjithashtu një mjet efikas mbrojtje antivirale. Kjo është me e dukshme sidomos në ato infeksione virale në të cilat koha e inkubimit është relativisht e gjatë si psh në rastin e poliomielitit apo të rabies (terbimit). Antitrupat qarkullues qofte dhe në përqendrim të ulët i neutralizojnë viruset në rrugën e tyre jashtëqelizore me apo pa bashkëpunimin e komplmentit.
Ne disa sëmundje të tjera virale si në grip apo në rrufën e zakonshme koha e inkubacionit është e shkurtër pasi organi shënjë i virusit është i njëjti me porten e hyrjes dhe s'kemi fazen e ndërmjetme të rrugës që përshkon virusi. për këtë arsye nuk ka kohë për një përgjigje antitrupore parësore dhe sipas mjaft gjasave prodhimi i shpejtë i interferonit është mekanizmi me i rëndësishëm i mbrojtjes antivirale. Por megjithate edhe sepse titri i antitrupave serike ndaj këtyre viruseve mund të jetë i ulët, ky nivel mund të jetë i lartë në sekrecionet lokale të mukozave të infektuara si psh. në mukozat bronkiale apo nazale. Kjo i dedikohët indit limfoid të lidhur me mukozat në të cilin prodhohen antitrupat antivirale (kryesisht të klasës IgA ) të cilët mund të jenë në titer të lartë në sekrecionet mukoze dhe që mund të luajnë rol të rëndësishëm sidomos në riinfeksionet e mëpasme. Përsa i përket rrufave të zakonshme, për fat të keq riinfeksionet e mëpasme janë zakonisht nga viruse me antigenicitet të ndryshëm nga ai i pari dhe prandaj dhe imuniteti i përgjithshem ndaj rrufave të zakonshme është vështirë të arrihet.
Ndaj viruseve me lokalizim brëndaqelizor vepron imuniteti me ndërmjetësi qelizore (limfocitet T):
Antitrupat lokale apo sistemike mund të bllokojnë përhapjen e viruseve citolitike të cilët clirohen nga qeliza bujtëse sapo kjo është vrare nga ky virus. Por vetëm antitrupat janë zakonisht të paaftë të mbrojnë nga viruset të cilët dalin në formë sythi nga nga qeliza e infektuar dhe nga këtu menjëhere infektojnë qelizën prane pa qendruar në hapësirën ekstra qelizore. në këtë grup përfshihen : viruset oncorna (viruset onkogjenike me ARN), ortomikso (v. i influencës), paramikso (parotiti, fruthi), toga (dengue), rhabdo (rabies), arena (koriomeningiti limfocitar), adeno, herpes (simplex, varicela-zoster, citomegalo, Epstein-Barr, semundja e Marek), pox (vaccinia), papova (SV40, polioma) dhe virusi i rubeoles. Rëndësia e imunitetit me ndërmjetësi qelizore për t'u sheruar nga infeksioni me këto lloj virusesh nënvizohet nga pamundesia e fëmijve me mangësi primare të

71
limfociteve T për të luftuar këto viruse, ndërsa të sëmurët me mangësi të antitrupave por me imunitet qelizor të paprekur i luftojnë ata me sukses.
Njohja e një qelizë të organizmit të infektuar nga një virus dhe vrasja e saj sa me e hershme janë mjaft dobiprurëse për mbrojtjen nga ky virus. Rëndësia e qelizave NK në këtë aspekt si përbërëse të imunitetit të lindur mbështëtet nga vrojtimet në të sëmurët e rralle me mungesë të plotë të këtyre qelizave, të cilët vuajne nga infeksione virale rekurrente dhe mjaft të rënda si p.sh. nga virusi EBV, virusi i variceles apo i citomegalovirusit. Sipërfaqja e një qelizë të infektuar nga viruse të ndryshme pëson modifikime që e bejne atë atraktive për qelizat NK. Këto të fundit pas kontaktit membranë me membranë me qelizën shënjë realizojnë vrasjen e kësaj të fundit në mënyrë pra jo specifike për antigenin.
Prodhim i interferonit alfa gjatë infeksionit viral mbron lokalisht qelizat nga ky infeksion por aktivizon gjithashtu dhe qelizat NK si dhe nxit shprehjen e molekulave të KMPI në ato qeliza.
Limfocitet T citotoksikë luajnë rol të rëndësishëm në mbrojtjen antivirale duke vrare qelizat e infektuara nga virusi që i sensibilizon ato dhe duke qënë specifike vetëm për ato qeliza që kanë KMPI të Klasës I të këtij individi (fenomeni i restriksionit me KMPI). Veprimi i tyre kërkon sigurisht një sensibilizim paraprak gjatë të cilit gjenerohen L. T. citotoksikë specifike, të cilët në një kontakt tëdytë me të njëjtin virus aktivizohen më shpejtësi duke penguar përhapjen e virusit.
Pas një infeksioni natyral viral kemi përgjigje imune të fituar si me antitrupa ashtu dhe me limfocite T citotoksikë, e cila është mjaft afatgjatë ndërsa imunizimi artificial p.sh. vaksinimi me viruse të vrare të influencës prodhon antitrupa, por jo limfocitet T citotoksikë dhe mbrojtja është afatshkurtër. Fakte të reja kanë vënë në dukje se edhe limfocitet T citotoksikë CD8 (+) mund të prodhojnë citokina si p.sh. IFN gama dhe kemotaksinat e makrofagëve dhe monociteve. Fagocitet mononukleare të terhequra me anë të chemotaxis në vatren inflamatore aktivizohen dhe sekretojnë TNF i cili në sinergji me IFN gama pengon replikimin viral me anë të transferimit interqelizor. Ashtu si IFN alfa dhe IFN gama mund të rrise citotoksicitetin jospecifik të qelizave NK për qelizat e infektuara.
Antitrupat antivirus veprojnë edhe me anë të një mekanizmi tjetër, ADCC ku përseri futet në loje mekanizmi vrasës i qelizave NK. Edhe komplementi mund të marrë pjesë gjithashtu në eliminimin e pjesëzave virale në ciklin e tyre ekstraqelizor.
Përgjigja imunitare ndaj infeksioneve parazitare: Me miliona në bote llogariten përsonat e infektuar nga parazitët e llojeve të ndryshëm. Këtu
futen parazitët protozoare ( plasmodiumi, leishmania, tripanozoma, toksoplazma) si dhe ata shumëqelizore ose helmintet ( shistozomat, trikinela, strongiloides, onkocerka, askaridet, oksiuret, trikocefalus etj). Pasojat nga infeksioni me këto parazitë do të ishin në një ekstrem mungesa e përgjigjes imunitare që con në një supërinfeksion të rrufëshem dhe në ekstremin tjetër një përgjigje imunopatollogjike e ekzagjeruar. të dyja këto pasoja ekstreme janë të demshme për parazitin sepse cojnë në pamundesine e transmetimit të tij nga një individ në tjetrin. Prandaj, për të qënë i suksesshem në mbijetësen e tij parazitë duhet të ruajë një ekuiliber midis këtyre ekstremeve, duke evituar si vdekjen e individit bujtës ashtu dhe eliminimin e vetë parazitit nga sistemi imunitar. Ky ekuilibër është jo i lehtë për t'u mbajtur.
Përgjigja imunitare e bujtësit: Përgjigja imunitare ndaj cdo paraziti është mjaft e gjërë por në përgjithësi pranohet se kur
paraziti invadon gjakun (malaria, tripanozomiaza) përgjigja kryesore imunitare është humorale, ndërsa ndaj parazitëve që rriten brënda indeve (p.sh. Leishmanioza ose Toksoplazmoza) zakonisht imuniteti mbrojtës është i tipit qelizor.
Shpesh, një organizëm i infektuar në mënyrë kronike është rezistent ndaj reinfektimit me mikroorganizma të rinj, një situatë e quajtur imunitet konkomitant si p.sh. në shistozomiaze apo malarie.
Ndaj disa lloj parazitësh me stade shumëzimi jashtëqelizor si psh. Trypanosoma brucei apo stadet sporozoite dhe merozoite të malaries mekanizmat humorale mund të jenë efikase me anë të opsonizimit, fagocitozës dhe lizës me ndërmjetësi nga antitrupat dhe komplementi.

72
Një shënjë karakteristike e reaksionit imun ndaj infeksioneve nga helmintet mund të jetë dhe eozinofilia si dhe rritja e nivelit serik të IgE. Kjo është vënë re në infeksionet nga Trichinella spiralis të njëriu apo Nippostrongylus brasiliensis të mijtë. Duket se kjo lloj përgjigje imune është me vartësi nga Th2 pasi në mënyrë eksperimentale është vënë re që injektimi i anti-IL-4 e ul mjaft sekretimin e IgE ndërsa anti-IL-5 suprimon eozinofilinë. Duket se sekretimi lokal i IgE nga depërtimi i parazitit rrit inflamacionin lokal të nevojshëm për mobilizimin e mëpastajme të faktorëve humorale mbrojtës nga sektori intraqelizor. Nga ana e tyre eozinofilet luajnë një rol mbrojtës nëpërmjet citotoksicitetit në vartësi nga antitrupat: ata kanë receptorë për pjesën Fc të IgG dhe nëpërmjet tyre lidhen me parazitët të mbuluar me IgG. Si pasojë, përmbajtja e granulave të eozinofileve (proteina madhore bazike e tij) kalon në brëndësi të parazitit duke provokuar ngordhjen e këtij të fundit. Të dhëna të tjera kanë vënë në dukje se në citotoksicitetin antitrup-vartes të eozinofileve marrin pjesë dhe IgE dhe IgA.
Nga ana tjetër disa parazitë si psh. Toxoplasma Gondii, Trypanosoma cruzi apo Leishmania kanë lokalizim intraqelizor (brënda makrofagëve) ku arrijnë të evitojnë aftësine baktericide të makrofagut. Kjo aftësi stimulohet nga limfocitet T prodhuese të citokinave që stimulojnë makrofagët sic janë gama-interferoni dhe TNF alfa. Pra limfocitet TH1 luajnë rol kyc në mbrojtjen nga këto lloj parazitësh.
Disa mikroorganizma të tjerë si plasmodiumi i malaries apo riketsiet dhe klamidiet strehohen në qeliza që nuk janë fagocite profesionale. Në këtë rast eliminimi i tyre mund të realizohet me anë të aktivizimit të mbrojtjes intraqelizore nga IFN gama që clirohet nga limfocitet T CD8 (+) ose dhe nëpërmjet citotoksicitetit direkt si psh. është rasti i qelizave hepatike që strehojnë sporozoitet e malaries.
Përsa i përket eliminimit të helminteve nga zorra ky është një opëracion me i ndërlikuar që kërkon kombinimin e mekanizmave qelizore me ata humorale. Pra këtu ndërhyjne si limfocitet T prodhues të citokinave ashtu dhe mastocitet si dhe antitrupat e prodhuara nga limfocitet B. Në fakt strategjia mbrojtëse varet mjaft nga lloji i infeksionit.
Mekanizmat mbrojtës të parazitëve ndaj sistemit imunitar: Disa parazitë me jetëse jashtëqelizore si Tripanozoma apo Skistozoma mbrohen duke
shpërbërë përbërësit e komplementit që aktivizohet në sipërfaqen e tyre. Parazitë brënda qelizore si Toxoplazma Gondii inhibojnë fuzionin fagozom-lizozom ndërsa
Leishmania rrethohet nga një lipofosfoglikan i cili e mbron nga baktericidia O2-vartese duke neutralizuar radikalet e oksigjenit.
Disa parazitë "mimetizojnë" vetvehten duke i ngjarë bujtësit. Kjo arrihet me anë të mimetizimit molekular sic ndodh psh. me Antigenët e Ascaris që i ngjajnë kolagjenit human. një mënyrë tjetër është duke mbuluar sipërfaqen e tyre me proteinat e bujtësit. Psh. format e rritura të shistozomave vishen me glikoproteinat e eritrociteve, molekulat e KMPI si dhe IgG humane dhe kështu qarkullojnë lirisht në vazat mezenterike të bujtësit megjithë praninë e antitrupave qarkullues kundër shistozomave që mbrojnë vetëm nga reinfeksioni.
Një mënyrë tjetër evitimi të sistemit imunitar është variacioni antigenik me anë të të cilit parazitët i shpetojnë veprimit mbrojtës të antitrupave. Psh. tripanozomat edhe pas formimit të antitrupave ndaj tyre vazhdojnë qetësisht jetën e tyre pasi ato ndryshojnë antigenët. Organizmi formon përseri antitrupa ndaj këtyre antigenëve por ndërkohë ato kanë ndryshuar rishtas duke lejuar kështu përsistencën e parazitëve në organizëm.
I njëjti fenomen është vënë re me Plasmodiumin e malaries dhe kjo mund të shpjegojë faktin që në zonat hipërendemike fëmijtë pësojnë atake të përsëritura të malaries në vitet e para të jetës dhe me pas ata janë imune ndaj infeksioneve të mëvonshme. Pra në ta është zhvilluar tashmë imuniteti ndaj të gjithë varianteve të ndryshme të plasmodiumit. Eshtë gjithashtu e njohur që IgG e përftuar nga serumi i individëve me imunitet solid ndaj Plasmodiumit ka efekt mjekues për infeksionin nga malaria tek fëmijtë.
Ne shumë infeksione parazitare është vënë në dukje shtim i imunosupresionit në saje ndofta dhe të ngarkeses se lartë antigenike si dhe të modulimit të nënpopullatave të limfociteve T. Ky imunosupresion i gjeneralizuar rrit prekshmerinë nga infeksionet bakteriale dhe virale.

73
Gjithashtu, persistimi i parazitëve në kontakt me përgjigjen imune mund të rezultojë në reaksione imunopatogjene si sindrom nefrotik nga komplekset imune, granuloma hepatike si dhe lezione autoimune të zemres.
KAPITULLI 17. IMUNOLOGJIA E SHTATZANISE
Faktorët që pengojnë flakjen e fetusit nga sistemi imunitar i nënes. Bashkëveprimi nëne-fetus dhe mekanizmat e aloimunizimit feto-amnor.
Një pasojë e polimorfizmit të një popullatë të përzier është fakti që nëna dhe fetusi kanë
Antigenë të ndryshëm të KMPI. Kjo është dobiprurëse për specien sepse rritet mundesia e mbijetëses se species gjatë evolucionit.
Por ky polimorfizem shtron pyetjen se si është e mundur që nuk kemi flakje imunollogjike të fetusit nga sistemi imunitar i nënes ndërkohë që një haplotip i KMPI se fëmijes (pra 1/2 e trashegimise gjenetike së tij) është e ndryshme nga nëna dhe pra nga ana imunollogjike fetusi duhet konsideruar si një transplant alogjenik 1/2 identik me marrësin. Megjithë këtë ndryshim gjenetik si dhe me praninë e kontaktit fizik në placentën humane të limfociteve imunokompetente të nënes me trofoblastin e fëmijes nuk kemi flakje imunollogjike ndonëse është e pranishme një përgjigje imunitare e nënes ndaj Antigenëve të HLA të babait gje që dëshmohet nga formimi i antitrupave citotoksikë anti HLA të babait. Madje këto antitrupa në mënyrë klasike janë përdorur (dhe vazhdojnë të përdoren) në tipizimin e Antigenëve të HLA. Në fakt, në mënyrë ekspërimentale është vërtetuar se edhe një sensibilizim paraprak i nënes me Antigenë të babait nuk e pengon zhvillimin e barrës. Pra me sa duket qelizat e trofoblastit janë imunollogjikisht të mbrojtura dhe në fakt ato janë rezistente ndaj mjaft mekanizmave citotoksikë megjithëse të ndjeshme ndaj qelizave NK të aktivizuara nga IL-2.
Mekanizmat që mendohet se luajnë rol në mosflakjen imunollogjike të fetusit mund të përmblidhen si me poshte: 1 - Barriera fizike nga vetë uterusi 2 - Prani në placentë të proteinave kontrolluese të komplementit 3 - Prania e HLA-G dhe rezistenca ndaj citotoksicitetit 4 - Faktorë të lidhur me placentën dhe citotrofoblastin 5 - Mungesa në placentë të Antigenëve HLA klasike të Klasës I dhe II 6 - Imunosupresioni lokal jo-specifik psh. TGF beta, IL-10 7 - Absorbimi i antitrupave anti-HLA në qelizat e placentës
Nga faktorët e mesipërm ai me i rëndësishmi është mungesa e shprehjes se Antigenëve HLA të Klasës I dhe II në trofoblastin viloz placentar gje që e mbron fetusin nga flakja alogjenike. Këto ndryshime themelore në rregullimin placentar të geneve të HLA cojnë gjithashtu në shprehjen krejt të vecantë të Antigenëve HLA-G në citotrofoblastin ekstraviloz. Kjo shprehje me sa duket e mbron trofoblastin nga limfocitet NK të nënes.
Antitrupa të klasës IgG të nënes anti-HLA të babait janë të pranishme në 20% të barrave të para dhe në 75-80% të grave multipare. Disa nga këta antitrupa mund të jenë të drejtuar dhe kundër HLA-G por ndjeshmëria e trofoblastit ndaj komplementit është e bllokuar nga prania në sipërfaqen e këtij e proteinave të kontrollit si DAF (decay accelerating factor) apo MCP(membranë cofactor protein) të cilat inaktivizojnë C3 konvertazen.
Edhe citokinat duken gjithashtu se luajnë një rol kompleks në ruajtjen e barrës duke patur patur parasysh prodhimin e faktorëve të rritjes si CSF-1 dhe GM-CSF, të cilët kanë një rol trofik për placentën si dhe të TGF beta e cila frenon aktivizimin e qelizave NK në placentë.

74
KAPITULLI 18. MANGËSITE IMUNITARE TË LINDURA
Ndarja e mangësive imunitare të lindura sipas mekanizmave imunopatogjenetike të
tyre. Shënjat e tyre kryesore klinike dhe biologjike.
Mangësite imunitare janë një grup sëmundjesh në të cilat një nga hallkat e sistemit imunitar është e manget. Meqënëse funksioni kryesor i sistemit imunitar është mbrojtja nga infeksionet, mangësia imunitare do të shprehet klinikisht me infeksione të ndryshme. Lloji i këtyre infeksioneve do të jetë në vartësi të hallkes se demtuar të imunitetit. P.sh. në kemi parë se ndërveprimi i komplementit, antitrupave dhe qelizave fagocitare përbëjne bazën e mbrojtjes ndaj infeksioneve piogjene (formuese të pus-it) me bakterie që kërkojnë një opsonizim paraprak para fagocitozës. Pra, mangësia në një nga tre këta faktorë do të favorizojë cfaqjen e infeksioneve të përsëritura dhe të rënda me bakterie patogjene si streptokok, pneumokok, meningokok, stafilokok, hemofilus influenzae etj.
Ndërsa të sëmurët me mangësi të limfociteve T do të paraqesin një profil të ndryshëm infeksionesh, kryesisht me viruse, këpurdha apo bakterie dhe parazitë që luftohen me anë të imunitetit qelizor.
Nga ana tjetër në të sëmurët me mangësi imunitare është vënë re edhe një shpështësi e rritur e tumoreve dhe fenomeneve autoimune. Këto të dhëna klinike vejne në dukje faktin që sistemi imunitar luan një rol kyc edhe në mbrojtjen anti-tumorale dhe mangësia e hallkave të ndryshme të tij sjell si pasojë fenomene autoimune.
Shumica e mangësive të lindura të imunitetit kanë karakter gjenetik nga ndryshime gjenike. Forma e trashegimise me shpesh është recesive autosomale ose e lidhur me kromozomin X. në raste me të rralla (S. i Di George) kemi të bejme me embriopati.
Mangësite e imunitetit të lindur sipas mekanizmave.

75
Ne Sëmundjen granulomatoze kronike polimorfonuklearet dhe monocitet nuk arrijnë të prodhojnë derivatet reaktive të oksigjenit si pasojë e një difekti në sistemin e citokrom b-245 oksidazes. Ky sistem normalisht aktivizohet nga fagocitoza. Citokromi b-245 përbëhet nga vargjet 92 dhe 22 kDalton dhe në formën e lidhur me kromozomin X të sëmundjes kemi një mutacion në genin që kodon vargun me të madh. Si rrjedhojë citokromi nuk sintetizohet, por ekziston dhe një variant i mutacionit të gp92 që lejon sintezën e niveleve të ulta të kësaj proteine. në këtë rast dhenja e interferonit mund të rrise nivelin e prodhimit të kësaj enzime. Semundja trashegohet me anë të kromozomit X, por në 30% të rasteve me sëmundje granulomatoze kronike kemi një trashegimi të tipit autosomal recesiv. në këtë rast difekti genik qendron në formën difektoze të sintezës se vargut 22 kDa si dhe të përbërësve p47 dhe p67 të sistemit të oksidazes NADPH.
Është interesant fakti që spektri i mikroorganizmave patogjene që prekin këta të sëmurë është relativisht i ngushtë dhe karakteristik: Infeksionet që hasim janë zakonisht nga Staphylococcus aureus si dhe disa bakterie gram-negative, ndërsa ndërmjet fungeve hasen me shpesh Aspërgillus fumigatus si dhe Candida albicans. Kjo se pari vjen nga fakti se shumë bakterie prodhojnë vetë H2O2 që shërbën pra për "vetvrasjen" e tyre. Por nëqoftëse bakteria është katalaze pozitiv (si psh. Staph. aureus) përoksidi shkaterrohet dhe kështu bakteria arrin të mbijetoje brënda fagocitit. Se dyti, mikroorganizmat me virulente janë ata që janë mjaft rezistente ndaj mekanizmave mikrobicidale jo-oksigjen vartese.
Ne sëmundjen Chediak-Higashi lizozomet janë deficitare në elastaze dhe në katepsine G dhe si pasojë të sëmurët vuajne nga infeksione piogjene që mund të jenë fatale.
Mangësia në mieloperoksidaze është një sëmundje tjetër gjenetike me karakter hereditar dhe me prekje të fagocitozës. në të hasim një prekshmeri të lartë ndaj kandidozave sistemike.
Ne Sindromin e leukociteve përtace hasim një përgjigje të manget të polimorfonukleareve ndaj stimujve kemotaktike.
Ne Mangësine e adezionit leukocitar mungesa e nën-njësise beta (CD18) të beta 2 integrinave shoqërohet me një kemotaksis të demtuar të leukociteve dhe infeskione bakteriale rekurrente. Ndërsa migrimi i monociteve, eozinofileve dhe limfociteve është i paprekur sepse këta kanë sistemin alternativ të integrinave VCAM-1/VLA-4.
Agranulocitoza kongenitale shoqërohet me infeksione të rënda nga mungesa e fagocitozës se mikroorganizmave. Injëktimi i përditshem në këta të sëmurë i G-CSF rekombinante është inkurajues pasi ai rrit numrin e granulociteve në shumicën e të sëmurëve.
Mangësite e sistemit komplementar. Difektet në proteinat e kontrollit: Rëndësia e komplementit për mbrojtjen anti-infeksioze u vu në dukje nga shpështësia e
infeksioneve të rënda me bakterie piogjene në një individ të i cili mungonte faktori I (inaktivatori i C3b). për shkak të paaftësise të shkaterroje C3b kemi një aktivizim të vazhdueshem të rrugës alterne që con në nivele të ulta të C3 dhe faktorit B, ndërsa nivelet e C1, C4 dhe C2 janë normale.
Cdo eritrocit bombardohet cdo ditë me rreth 1000 molekula C3b të cilat gjenerohen në mënyrë spontane nga C3 serik nëpërmjet aktivizimit të C3 konvertazes alterne. janë disa molekula që merren me inhibimin e këtij procesi duke disociuar kompleksin C3 konvertaze. Ndër to përmendim faktorin akselerues-zbërthyes (DAF = decay - accelerating factor), receptorët CR1 të komplementit në sipërfaqen e eritrociteve si dhe faktorin H. Me pas vetë C3b copëzohet nga faktori I që vepron bashkë me CR1, proteinën kofaktorë membranore (MCP) ose dhe faktorin H. Ekzistojnë gjithashtu 2 inhibitore të kompleksit të sulmit membranor që janë faktori homolog i restriksionit (HRF) dhe CD59 të cilët duke lidhur C8 parandalojnë aktivizimin e molekulës se parë C9 e nevojshme kjo për inserimin në membranë. DAF, HRF si dhe CD 59 "ankorohen" në membranë nëpërmjet disa molekulave glikozil-fosfatidil-inozitoli. në senundjen e njohur si Hemoglobinuria paroksizmale nokturne (HPN) kemi një difekt në sintetizimin e këtyre "ankorave". Si pasojë e mungesës se inhibitoreve të aktivizimit të komplementit ndodh aktivizimi i këtij të fundit me rrjedhojë lizën eritrocitare.
Ne Angioedemen hereditare, e cila është mangësia me e shpështë e një faktori të komplementit kemi mungesë të lindur të inhibitorit të C1 (C1INH). Kjo con në episode të përsëritura

76
të edemes akute jo-inflamatore dhe të kufizuar, por që në raste të vecanta mund të cfaqet në formë të rëndë si psh me edeme laringeale. Këto atake të edemes, të cilat vijne si pasojë e aktivizimit të pakontrolluar të rrugës klasike të komplementit, shkaktohen nga trauma, ndërhyrje kirurgjikale, dentare etj. Zakonisht të sëmurët janë heterozigote dhe arrijnë të sintetizojnë sasira të pakta të inhibitorit të cilat mund të rriten në nivele të mjaftueshme me anë të dhenjës se medikamentit danazol. një alternative tjetër mjekimi është acidi epsilon-aminokaproik i cili bllokon clirimin nëpërmjet plazmines të përbërësit C2 të komplementit. Ataket e rënda të sëmundjes mund të kërkojnë administrimin e vetë C1 inhibitorit të purifikuar.
Mangësite e përbërësve të komplementit: Në një numër individësh me mangësi të C1q, C1r, C4 dhe C2 (kjo e fundit është nga
mangësite e lindura me të shpeshta të sistemit komplementar) është konstatuar shpeshtësi e rritur e fenomeneve autoimune të ngjashme me lupusin sistemik eritematoz (LES). Si pasojë e mangësise se këtyre faktorëve kemi paaftësi të formimit të C3 konvertazës klasike dhe si rrjedhojë aftësi e ulur e përgjigjes imune ndaj agjentëve infeksioze apo dhe e eliminimit efikas të komplekseve antigen-antitrup. Përsa i përket faktorëve C5, C6, C7, C8, C9, mangësi të lindura tyre janë konstatuar në individë, të cilët përvec një fare tendencë për të bërë me shpesh nfeksione të gjeneralizuara me Neisseria gonorrhoeae dhe N. meningitidis nuk kanë ndonjë prekshmeri me të lartë ndaj infeksioneve në përgjithësi. Me sa duket, këta përbërës të rrugës terminale të aktivizimit komplementar nuk janë esenciale për mbijetësen e species dhe antitrupat opsonizues apo dhe mekanizmat e aderences imune japin një mbrojtje të mjaftueshme imunitare përsa i përket imunitetit humoral.
Mangësite parësore të limfociteve B: Agamaglobulinemia e lindur (M. Bruton) është një nga mangësite e lindura të imunitetit,
geni përgjegjës për të cilët gjëndet në kromozomin X. Pra preken meshkujt dhe sëmundja transmetohet nga femrat. Në meshkujt e prekur nga kjo sëmundje kemi një ulje deri në zero të te gjitha klasave të imunoglobulinave ndërsa në biopsite e nyjeve limfatike janë të pranishem një numër i papërfillshem plazmocitesh dhe folikujsh limfoide.
Difekti është në nivelin e qelizës pre-B dhe kemi një pamundesi rirregullimi të geneve VH me prodhim të pjesëzave të ndërprere D-J-my. Mutacioni genik ndodh në një gen që kodon për tirozine-kinazen në kromozomin X dhe që i ngjan familjes src se proteine-kinazave citoplazmike.
Fëmijet e prekur zakonisht s'kane simptoma deri 5-6 muaj të jetës, kohë gjatë se ciles sigurohet mbrojtja pasive nga imunoglobulinat e marra nga nëna. Pas kësaj moshe fillojnë infeksione të shpeshta, të rënda dhe të përsëritura me mikroorganizma piogjene (ekzogjene ekstraqelizore) si Staphylococcus aureus, Streptococcus pyogenes dhe pneumoniae, Neisseria meningitidis, Haemophilus influenzae si dhe nga viruse me cikel biollogjik të gjatë ekstraqelizor si p.sh. Viruset e hepatitit, ekoviruset etj. Përgjigjet imunitare me ndërmjetësine e limfociteve T janë të paprekura në këta të sëmurë dhe infeksione virale të tilla si psh. fruthi apo lija kalojnë pa probleme. Mjekimi i vetëm për këta të sëmurë është dhenja e imunoglobulinave me rrugë parënterale (me mirë venoze) të pakten cdo muaj (rreth 400 mg/kg/peshe) duke patur si qellim mbajtjen e një niveli të pranueshem serik të IgG (minimumi mbi 2 g/l).
Mangësia e lindur e IgA është forma me e shpeshtë e mangësive të lindura të limfociteve B. Kemi një pamundesi të limfociteve B bartëse të IgA membranore që të diferencohen në plazmocite prodhuese të IgA dhe niveli serik i IgA është zero ose në nivele gjurme. Mjaft nga këta individë janë krejt asimptomatike dhe zbulohen krejt rastesisht në një depistim të rastit si psh. ndër dhuruesit e gjakut. Mendohet se roli i rëndësishëm i IgA sekretore kompensohet nga IgM në sekrecionet mukoze. në një numur individësh të tjerë kemi infeksione të shpeshta në rrugët e frymëmarrjes apo ato digjestive. në mjaft prej tyre është vënë re një mungesë e njëkohshme e një apo dy nënklasave të IgG.
Trajtimi me imunoglobulina nuk rekomandohet në këta të sëmurë sepse mund të kemi imunizim me IgA dhe formim të antitrupave anti-IgA që në një injektim të mëvonshem mund të japin një reaksion anafilaktik.
Mangësia imunitare e quajtur "e zakonshme dhe e ndryshueshme (common variable)":

77
është ajo me e shpeshta dhe mendohet se përbëhet nga disa forma. Shënjat klinike me të shpeshta janë infeksionet piogjene rekurrente, si bronkitet, sinuzitet, otitet etj. si dhe diarrete deri në malabsorbim. Zakonisht sëmundja cfaqet në moshen mbi 15 vjec dhe preken me tepër femrat se meshkujt. Palca kockore e këtyre individëve zakonisht përmban numër normal limfocitesh B imature, por në 1/3 e pacienteve mund të mungojnë limfocitet B qarkullues ndërsa 1/3 tjetër ka një numër të ulët të tyre. Edhe në ata paciente me numër normal limfocitesh B këta shprehin anomali funksionale si paaftësi për t'u diferencuar në plazmocite apo për të sekretuar antitrupa. Edhe limfocitet T mund të jenë të prekur. Ata mund të paraqesin një nivel të ulët të 5-nukleotidazes sipërfaqesore, rreth 30% e tyre kanë një përgjigje të ulët ndaj aktivatorit poliklonal të limfociteve T fitohemaglutinine (PHA) dhe një përqindje e ulët kanë limfocite T të fenotipit CD8(+), KMPI Kl II (+) dhe një aktivitet supresor të theksuar për limfocitet B. Baza gjenetike e kësaj lloj mangësie është akoma e panjohur por një numër i madh të këtyre të sëmurëve kanë të njëjtet haplotipe të HLA si të sëmurët me mangësi të IgA gje që sugjeron një crregullim gjenetik të përbashkët.
Hipogamaglobulinemia tranzitore e latantit karakterizohet nga infeksione respiratore rekurrente dhe shoqërohet me nivele të ulta të IgG që fillojnë dhe normalizohen afer moshes 4 vjec. Ekziston një mangësi në numrin e limfociteve qarkullues si dhe në aftësine e tyre për të gjeneruar ndihmë efektive për prodhimin e imunoglobulinave nga limfocitet B të aktivizuara nga Pokeweed mitogen (PWM). Kjo normalizohet me kalimin e sëmundjes. Por megjithate limfocitet B janë të pranishem dhe jo zero si në M. Bruton.
Një fare shkallë hipogamaglobulinemie mund të cfaqet në mënyrë fiziollogjike në cdo latant me uljen natyrale të nivelit të Ig. të nënes që ndodh rreth moshes 5 muaj. Kjo mund të bëhet problem sidomos në fëmijtë premature.
Mangësite parësore të limfociteve T: Sindromet DiGeorge dhe Nezelof karakterizohen nga mungesa e zhvillimit të timusit nga
xhepat e trete dhe të katërt embrionale gjatë embriogjenezes. në DiGeorge kemi gjithashtu mungesë të paratireoideve dhe anomali kardiovaskulare dhe të fytyrës. Si pasojë e mungesës se timusit qelizat bazë hematopoietike nuk mund të diferencohen në limfocite T dhe si pasojë zonat timovartëse në indet limfoide janë të populluara varfër. Folikujt limfoide janë të pranishem por jo të zhvilluar mirë. Përgjigjet imunitare me ndërmjetësi qelizore s'jane të pranishme dhe, megjithëse fëmijet e prekur me këtë sindrom mund t'i rezistojnë infeksioneve bakteriale të zakonshme, ata kanë pasoja fatale nga fruthi ose vaksinimi me BCG ose lijen. Përgjigja antitrupore serike mund të provokohet, por ajo është mjaft e dobët duke reflektuar në këtë mënyrë mungesën e limfociteve T ndihmës. Trajtimi me transplant të timusit të neonatit con në restaurimin e imunokompetences.
Ne shumicën e rasteve me sindromin e diGeorge nuk kemi mungesë të plotë të timusit por një "diGeorge të pjesshem" në të cilët numri i limfociteve T lëkundet nga 6% në 30% të limfociteve qarkullues nga fundi i vitit të parë të jetës. Përgjigjet antitrupore janë zakonisht të pranueshme.
Mangësi selektive të limfociteve T mund të kemi në Mangësine e Purine-nukleozid-fosforilazes. Si pasojë e mungesës se lindur se kësaj enzime kemi akumulim të metaboliteve toksike si psh. dGTP ndaj të cileve limfocitet T të aktivizuar janë vecanerisht të ndjeshëm. Mungesa e përgjigjeve imunitare T-vartese i bën këta të sëmurë mjaft të prekshem nga infeksionet me viruset e variceles dhe të lijes(vaccinia), por megjithëse ata kanë me pak se 10% limfocite T qarkulluese ata kanë një përgjigje antitrupore normale, gje që sugjeron se bashkëpunimi T-B vepron të njëriu në nivele shumë me të ulta të limfociteve T se sa të miu.
Imuniteti me ndërmjetësi qelizore është i deprimuar në të sëmurët me Ataksi-Teleangiektazi si dhe në ata me trombocitopeni dhe ekzema (Sindromi i Wiskott-Aldrich). është me interes fakti se në të dy këto sëmundje rreth 10% e të sëmurëve të studiuar deri tani kanë vdekur nga sëmundje malinjë të indit limfoid ose tumore epiteliale. Sindromi i Wiskott-Aldrich është i lidhur me kromozomin X dhe në meshkujt e prekur mungon një molekulë e sipërfaqes qelizore, sialoforina (CD43), e cila është ligandi i ICAM-1. Ky sindrom është i shoqëruar me një nivel të ulët të IgM dhe një përgjigje të manget ndaj shumë polisakarideve. Këta të sëmurë kanë pak limfocite B që bartin imunoglobulina sipërfaqesore me idiotipe specifike për Antigenët polisakaridike. Përgjigja imunitare e këtij lloji limfocitesh B, i cili maturohet me vone në zhvillimin e limfociteve B në përgjithësi,

78
kërkon ndihmën e limfociteve T helper, por në mënyrë jo specifike dhe të pavarur nga kompleksi receptor CD3/TCR. Pyetja vjen në mënyrë të natyrshme: Mos valle CD43 është receptori jo-specifik për polisakaridet të cilët stimulojnë kështu ndihmën jo specifike të limfociteve T për limfocitet B përkates?
Ataksia-teleangiektazia është një sëmundje gjenetike me trashegimi autosomale recesive që karakterizohet nga një ataksi cerebelare progresive me degjenerim të qelizave Purkinjë si dhe një hipërndjeshmeri ndaj rrezeve X. Mendohet se difekti gjenetik në këtë sëmundje qendron në mangësine e mekanizmave të riparimit të ADN. Kjo shpjegon përqendrimin e ndërprerjeve kromozomike rrotull geneve të familjes supërgjenike të imunoglobulinave dhe vecanerisht TCR dhe IGH. Si pasojë kemi mangësi si qelizore ashtu dhe të prodhimit të antitrupave. Mangësia e njëkohëshme e IgA dhe e IgE në këta të sëmurë spjegon pjesërisht shpeshtësine më të lartë të infeksioneve të rrugëve të sipërme respiratore në këta të sëmurë në krahasim me ata që kanë mangësi të izoluar të IgA.
Raste të izoluara të mangësise se sistemit T-vartes janë përshkruar në individë në të cilët janë gjetur antitrupa limfocitotoksikë të cilët janë kryesisht të drejtuar kundër limfociteve T.
Limfocitet T të disa të sëmurëve me Kandidoze mukokutane kanë rezultuar të paaftë të prodhojnë citokinën MIF (faktori inhibues i migrimit të makrofagëve) kur ata stimulohen in vitro. Mendohet se ekzistojnë edhe mangësi të tjera të imunitetit me shkak mungesën e sintezës se citokinave.
Gjithashtu janë përshkruar edhe raste të rralla të mangësise se CD3 si pasojë e mutacionit në vargjet gama-theta të këtij receptori.
Ne disa të sëmurë të tjerë me mangësi të imunitetit qelizor është konstatuar një difekt në faktorin e transkriptimit bërthamor NF-AT të limfociteve T të aktivizuar.
Mangësite imunitare të lindura të kombinuara (MILK): Këto janë nga mangësite me të shpeshta të lindura të imunitetit. Në to kemi një difekt
gjenetik në enzimat rekombinaze që janë të domosdoshme për formimin e receptorëve të limfociteve T dhe B. Si pasojë këta limfocite nuk bëhen imunokompetente dhe kjo sjell si pasojë një mangësi të rëndë si të imunitetit qelizor dhe të atij humoral.
Ne fëmijet që kanë këtë mangësi funksionet imune mund të rivendosen me anë të transplantimit të palcës kockore me dhurues HLA të përshtatshem nga vellezërit apo motrat. Qelizat nga dhurues të tjerë edhe në rast se janë të përshtatshem në HLA mund të shpërthejne një reaksion të dhuruesit ndaj bujtësit (graft versus host) i cili mund të jetë vdekjeprurës. Për këtë arësye është e domosdoshme që qelizat imunokompetente të eliminohen nga qelizat e tjera të palcës kockore para se të futen të bujtësi. Disa të sëmurë me MILK kanë mungesë të enzimes adenozine deaminaze (ADA). Si pasojë kemi akumulim të metaboliteve toksike të purines dATP ashtu si mangësia e enzimes tjetër, purine-nukleozid-fosforilazes. Por ndryshe nga kjo e fundit nga metabolitet toksike preken në këtë rast edhe limfocitet B ashtu si ato T. Një pjesë e këtyre të sëmurëve përmirësohen me anë të transfuzionit të eritrociteve normale që përmbajne enzimen që mungon ndërsa të sëmurë të tjerë që vuajne për një kohë të gjatë nga një mangësi të rëndë të kësaj enzime mund të pësojnë dëmtim të epitelit timik dhe si pasojë kanë nevoje për një trajtim me enzime të modifikuar me polietilenglikol, i cili e zgjat gjysëm-jetën e saj. Këta të sëmurë janë kandidatë ideale për terapinë gjenike. Në këtë rast limfocitet T të këtyre të sëmurëve mund të transfektohen me gjenin e ADA të futur në një vektor retroviral dhe më pas t’i riinjektohen të sëmurit. Forma me e rëndë dhe fatale e MILK është e lidhur me mungesën e plotë të prekursoreve mieloide dhe quhet disgeneze retikulare. një tjetër formë e MILK është e lidhur me mungesën e shprehjes se molekulave të HLA në sipërfaqen limfocitare.
Diagnoza e mangësive imunitare: Mangësite në imunoglobulina mund të vleresohen me matje sasiore: niveli prej 2g/L mund të
konsiderohet arbitrarisht si kufiri minimal i normes. Përgjigja imune humorale mund të studiohet me anë të matjes se antitrupave natyrale (izohemaglutininat A dhe B, heteroantitrupat ndaj eritrociteve të dashit, baktericidinat ndaj E. Coli) si dhe me matjen e antitrupave specifike pas imunizimit me vaksinën e difterise apo tetanozit, por jo me vaksina të gjalla.

79
Te sëmurët me mangësi të limfociteve T janë areaktive në provat e lëkurës ndaj antigenëve si tuberkulina, Candida, trikofitina, streptokinaza/streptodornaza apo parotiti. Me ane të dinitroklorobënzenit (DNCB) mund të studiohet in vivo sensibilizimi primar i imunitetit qelizor. Reagimi i limfociteve të gjakut përiferik ndaj stimulimit të tyre in vitro me fitohemaglutininë ose antigenë si ato in vivo është një tregues i ndjeshëm i funksionit të limfociteve T. Për këtë shërbën dhe reaksioni i përzier limfocitar i njëanshem gjatë të cilit limfocitit e të sëmurit që ekzaminohet vihen në kontakt me limfocite të inaktivizuara (me rrezatim p.sh.) të një individi tjetër.
Numërimi i limfociteve T realizohet me mikroskop me fluoreshencë ose me citofluorometri me antitrupa monoklonale ndaj popullatave të ndryshme limfocitare.
Edhe për studimin e funksionit të sistemit komplementar përdoren metoda të ndryshme funksionale apo sasiore. Për aftësine baktericidale apo funksione të tjera të polimorfonukleareve përdoren metoda të ndryshme, ndërsa për studimin e enzimave oksidative të lidhura me aktivitetin e fagocitozës përdoren testi i reduktimit të nitroblutetrazoliumit (NBT) ose stimulimi i prodhimit të supëroksideve.
KAPITULLI 19. MANGËSITE IMUNITARE TË FITUARA
Përgjigja imune mund të dëmtohet në mënyrë jo specifike nga shumë faktorë. Vecanërisht imuniteti qelizor mund të dëmtohet në gjëndjet e kequshqyerjes qoftë kjo e fundit edhe në format jo ekstreme të saj. Mangësia në hekur është mjaft e reëndësishme në këtë drejtim. Infeksionet virale janë shpesh shkak imunosupresioni dhe shembulli më tipik është fruthi, virusi i të cilit ka veprim direkt citotoksik mbi qelizat limfoide. Virusi më i njohur me veprim imunosupresiv është pa dyshim virusi HIV, por ky do të shtjellohet më gjatë në kapitullin tjetër. Në leprën lepromatoze si dhe në malarien kemi prekje të funksionit të përgjigjes imunitare si pasojë e një anomalie të qarkullimit limfocitar apo si në rastin e malaries si pasojë e crregullimit të funksionit të makrofagëve. Crregullimi i ekuilibrit ndërmjet limfociteve Th1 dhe Th2 si pasojë e infeksionit mund të prishë funksionin e asaj popullatë që ka veprim mbrojtës për infeksionin në fjalë.
Shumë agjentë terapeutikë si psh. rrezet X, medikamentet citotoksikë apo dhe kortikosteroidet kanë veprim mbi sistemin imunitar. Sëmundjet limfoproliferative të limfociteve B si leucemia limfoide kronike, mieloma, makroglobulinemia e Waldenstrom janë të lidhura me shkallë të ndryshme të hipogamaglobulinemisë si dhe të dëmtimit të përgjigjes antitrupore. Infeksionet që hasen më shpesh në këta të sëmurë janë nga bakteriet piogjene, ndërsa në sëmundjen e Hodgkin të sëmurët kanë mangësi të imunitetit qelizor dhe si pasojë hasim infeksione nga bacili i tuberkulozit, Brucella, Cryptococcus apo Herpes Zoster virus.

80
KAPITULLI 20. SINDROMI I MANGËSISE IMUNITARE TË FITUAR NGA
INFEKSIONI ME VIRUSIN HIV (SMIF - SIDA/AIDS)
AIDS është një sëmundje e shkaktuar nga infeksioni me virusin HIV dhe që ka marrë përpjestimet e një pandemie botërore. Në disa pjesë të Afrikës ku incidenca e infeksionit është e lartë transmetimi është kryesisht me rrugë heteroseksuale, ndërsa në pjesët e tjera të botës shumica e rasteve janë cfaqur në meshkuj homoseksualë, përdorues të drogave intravenoze, hemofilikë që kanë marrë faktor VIII të nxjerrë nga plazma apo në fëmijë të lindur nga gra të infektuara me virusin HIV. Megjithatë edhe në këto rajone numri i të infektuarve me rrugë heteroseksuale është në rritje të vazhdueshme. Shënjat kryesore klinike janë infeksionet oportuniste nga viruset sic janë Pneumocystis carinii, Cytomegalovirus, Epstein-Barr virus (EBV), Herpes simplex virus, këpurdhat Candida, Aspërgillus, Cryptococcus, parazitët Toxoplasma Gondii, Pneumocystis carinii, bakterie si Mycobacterium tuberculosis etj. Gjithashtu në këta të sëmurë kemi një shpeshtësi të lartë të sarkomës së Kaposi. Sidomos në fazat e para të sëmundjes haset sindromi i quajtur ARC (AIDS related complex) i karakterizuar nga tempëraturë, humbje në peshë dhe limfadenopati.
AIDS shkaktohet nga infeksioni me virusin HIV: Transmetimi I sëmundjes ndodh zakonisht nëpërmjet infeksionit me gjak ose spërmë që
përmban virusin HIV. Ky virus shumëzohet në sistemin limfoid të të infektuarit. Përiudha e latencës pas infektimit mund të zgjasë me vite dhe jo të gjithë të infektuarit zhvillojnë AIDS më tej. HIV është një retrovirus me ARN që bën pjesë në grupin e lentiviruseve, të cilët kanë aftësinë të shkatërrojnë qelizën që infektojnë, shkaktojnë sëmundje me përiudhë të gjatë latence, dhe kanë aftësinë t’I shpëtojnë sistemit imunitar.

81
Glikoproteina gp 120 e mbështjelljes së virusit lidhet me aviditët me molekulën CD4 të limfociteve T dhe nëpëermjet proteinës gp41 fillohet fuzioni i virusit me limfocitin T dhe si pasojë infeksioni i këtij të fundit. Qelizat shënjë janë limfocitet T helper që kanë me shumicë CD4 në sipërfaqen e tyre, por prania e CD4, megjithëse me dendësi të ulët, edhe në makrofagët dhe qelizat mikrogliale I bën këto të fundit të prekshëm nga infeksioni. Qelizat e infektuara lejojnë replikimin viral që mund të infektojë kështu qeliza të tjera. Qelizat folikulare dendritike mund ë infektohen gjithashtu nga HIV nëpërmjet një procesi që nuk përfshin CD4. Kështu qelizat dendritike përbëjnë kështu një rezervuar të rëndësishëm të virusit në organizëm. HIV përdor një transkriptazë riverse për të konvertuar ARN e vet në ADN e cila integrohet në genomën e qelizës bujtëse ku mund të qëndrojë në gjëndje latence për një kohë të gjatë. Stimulimi i limfociteve T dhe makrofagëve të infektuar por në gjëndje latence con në aktivizimin e prodhimit të proteinave virale dhe pra të replikimit viral.
Historia natyrale e infeksionit: Pas infektimit me virusin HIV mund të pasojë një sindrom retroviral akut që shoqërohet me
tempëraturë, mialgji, artralgji, viremi dhe pozitivitet për p24 (nukleokapsid dominant antigenik viral) në gjak. Përgjigja imunitare ndaj virusit, e identifikueshme nga cfaqja e antitrupave qarkullues ndaj p24 dhe ndaj proteinave të mbështjelljes gp120 dhe gp41 si dhe nga gjenerimi i limfociteve T citotoksikë specifike për gp120, ndërpret vireminë dhe con në sekuestrimin e HIV në indin limfoid. Kapja e pjesëzave virale të kompleksuara me antitrupa dhe komplement stimulon hipërplazinë folikulare si dhe infeksionin e qelizave folikulare dendritike. Në fakt folikujt limfoidë bëhen vendi kryesor i replikimit viral si dhe i infektimit të qelizave të tjera të sistemit imunitar. Më pas mund të ndodhë involucioni folikular që con në një degjenerim gradual të qelizave folikulare dendritike me një rritje të ngarkesës virale dhe të replikimit në qelizat mononukleare të gjakut përiferik. Si pasojë numri i limfociteve T CD4 (+) në gjakun përiferik bie progresivisht. Kur ky është nën 50/mm3 fillojë të cfaqen pasojat për imunitetin me ndërmjetësi qelizore si psh. negativizim i provave intrakutane me antigenë të zakonshëm si tuberkulina, kandidina, tetanos apo mosgjenerimi i limfociteve T citotoksikë ndaj viruseve të zakonshëm si psh. citomegalovirusit. Në këtë situatë i sëmuri është tashmë i pambrojtur ndaj infeksioneve të rënda të shkaktuara nga agjentë jopatogjene në kushte normale (oportunistë) sic janë Pneumocystis carinii apo Cytomegalovirus.
Mekanizmat e pakësimit (deplecionit) qelizor: Disa hipoteza përpiqen të shpjegojnë pakësimin e theksuar të limfociteve T CD4 (+) në të
sëmurët me AIDS: 1. Veprimi direkt citopatik i virusit mbi qelizë ose nëpërmjet prodhimit të bashkimeve sinciciale
midis limfociteve T CD4(+). 2. Veprimi i HIV si supërantigen duke ndërvepruar me disa familje TCR dhe eliminim të
limfociteve T që bartin këta receptorë. 3. Në të sëmurët asimptomatikë HIV(+) stimulimi nga virusi HIV i L.T. CD4(+) con në apoptozë të
këtyre të fundit. Në qelizat dendritike të infektuara kemi një replikim viral aktiv si dhe një pakësim i shprehjes së molekulave të KMPI Kl. II si dhe të B7. Si pasojë, molekulat e shumta të gp 120 të pranishme në sipërfaqen qelizore gjatë “celjes” së pjesëzave virale në sipërfaqet qelizore induktojnë apoptozën duke bashkëvepruar me CD4 në sipërfaqen e L.T.
4. Qelizat dendritike dhe makrofagët e humbin aftësinë e tyre për të paraqitur antigenët solubël limfociteve T CD45RO në të sëmurët asimptomatikë HIV(+). Si pasojë e mosmbajtjes të aktivizuar të popullatës së L.T. kujtesë CD45RO(+) do të kemi si pasojë edhe pakësim të popullatës kujtesë së L. T. në gjëndje qetësie (CD45RO+). Këto ndryshime mund të shpjegojnë devijimin e limfociteve T helper drejt nënpopullatës Th2, gjë që vihet re gjatë avancimit të sëmundjes.
5. Induktim i një përgjigje autoimune si pasojë e reaksionit të kryqëzuar midis proteinave të HIV dhe epitopeve kriptikë në limfocitet T të aktivizuar, ose nëpërmjet interferimit me mekanizmat e tolerancës normale intratimike.
6. Humbje qelizore si pasojë e mbulimit të CD4 me komplekse imune me gp120.

82
Të sëmurët me AIDS kanë si pasojë hipërgamaglobulinemi si dhe një numër të lartë të limfociteve B që prodhojnë imunoglobulina in vitro në mënyrë spontane dhe kjo sugjeron faktin që këta limfocite janë të stimuluar në mënyrë poliklonale.
Diagnoza e AIDS: Një individ me infeksione oportuniste, limfopeni, numër të ulë t të L.T. CD4(+), por numër
relativisht të ruajtur të L.T. CD4(+), nivele të larta të IgG dhe IgA si dhe negativitet të provave të lëkures ndaj antigenëve të zakonshëm mund të jetë me AIDS sidomos nqs. ai i takon një grup-popullate në risk. Konfirmimi i diagnozës realizohet me vënjën në dukje të antitrupave antivirale anti-HIV me ELISA si dhe me imunobloting (shih Kap. ). Në sëmundje aktive kemi gjithashtu dhe pozitivitet të antigenit p24. Prova e pakundërshtueshme dhe përfundimtare e infeksionit me virusin HIV është izolimi i virusit ose demonstrimi i genomës së virusit HIV.
Kontrolli i infeksionit nga HIV: Masa më e mirë për kontrollin e këtij infeksioni do të ishte parandalimi i tij nëpërmjet vaksinimit.
Antitrupat e parë që cfaqen pas infeksionit janë të drejtuar ndaj glikoproteinës gp120 të mbështjelljes virale; antitrupat ndaj proteinave të brëndësisë virale p24 dhe p18 mund të cfaqen më vonë. Shumica e antitrupave janë jo-neutralizues, pra ata nuk kanë aftësinë të parandalojnë infeksionin e qelizave in vitro. Deri tani nuk është bërë e mundur të prodhohen antitrupa mbrojtës edhe për faktin se molekula gp120 pëson ndryshime edhe brënda të njëjtit individ.
Terapia antivirale është drejtuar ndaj transkriptazes riverse si psh. AZT(zidovudina). Medikamente të tjerë antiviralë janë antiproteazat, që bashkë me AZT përbëjnë tashmë një mjet të fuqishëm terapeutik për tëe ndaluar përparimin e sëmundjes.
KAPITULLI 21. REAKSIONET E MBINDJESHMËRISË, 4 LLOJET E TYRE
Përgjigjet imunitare të tepruara mund të cojnë në dëmtime indore. Kur një individ ka rënë në kontakt me një antigen të caktuar kontaktet e metejshme me këtë
antigen mund të cojnë në fuqizim të tepruar të përgjigjes imunitare. Ky reaksion mund të jetë i ekzagjeruar dhe të shkaktoje dëmtime indore (mbindjeshmeri) nëqoftëse antigeni është i pranishem në sasi relativisht të larta ose nëqoftëse përgjigja imunitare humorale apo qelizore është e ekzagjeruar.
Duhet të kemi parasysh se mekanizmat që qendrojnë pas këtyre reaksioneve të papërshtatshme janë po ato që përdoren normalisht për luftimin e infeksioneve. Reaksionet e mbindjeshmërisë janë klasifikuar sipas Gell dhe Coombs në 4 tipe, të cileve i është shtuar dhe një tip i peste. Tipet I, II, III, dhe V varen nga ndërveprimi i antigenit me antitrupat humorale dhe quhen në përgjithësi reaksione të tipit të shpejtë, megjithëse disa janë me të shpejtë se të tjerët. Tipi i IV përfshin reaksionet me ndërmjetësine e limfociteve T dhe për arësye të kohë-cfaqjes me të vonshme është quajtur mbindjeshmeri e tipit të vonuar. Por mbindjeshmeria mundte cfaqet edhe nga ndërveprimi direkt i agjentit shkaktar me elemente të sistemit të imunitar të lindur pa ndërhyrjen e përgjigjeve të imunitetit të fituar.
Tipi i I - Mbindjeshmeria e shpejtë (anafilaktike): Te dhënat klinike me të hershme mbi një përgjigje imunitare të tepruar janë mbi anafilaksine.
një injektim i vetëm i 1 mg albumine të vezës në një kavie nuk ka asnjë efekt të dukshëm. Por nqs. injektimi përsëritet 2-3 jave me vonë, kafsha e sensibilizuar reagon tashmë në mënyrë dramatike me shënjat e anafilaksisë së gjeneralizuar: pothuajse menjëhere kafsha fillon të gulcojë dhe pas disa

83
minutash ngordh nga asfiksia. Të dhënat patollogjike tregojnë për konstriksion të bronkeve dhe bronkioleve dhe në përgjithësi kemi kontraksion të muskujve të lemuar dhe dilatacion të kapilareve. Reaksione të tila të rënda janë vënë re të njëriu pas pickimit të grerezes ose bletes si dhe pas injektimit të penicilines në individë të sensibilizuar. në shumë nga këto raste vetëm injëksioni i shpejtë intravenoz i adrenalines, që parandalon kontraktimin e muskujve të lemuar si dhe dilatacionin kapilar mund të parandaloje vdekjen. Këta reaksione shkaktohen nga degranulimi eksploziv i mastociteve dhe bazofileve dhe ky i fundit është përgjegjës për clirimin e histamines dhe të mjaft mediatorëve të tjerë. Degranulimi i bazofileve shkaktohet nga ndërveprimi i alergenit me antitrupat e klasës IgE këto të fundit të fiksuar mbi membranën e mastociteve nëpërmjet receptorëve me afinitet të lartë për fragmentin Fc të IgE (FcεRI) që përmban kjo membranë. Kalimi i limfociteve B në qeliza plazmocitare prodhuese të IgE kontrollohet nga IL-4, e cila vepron me një sinjal bashkëstimulues, i cili është TNF-alfa i lidhur me membranën e limfociteve T. Stimulimi i sintezës se IgE nga IL-4 fuqizohet nga veprimi i IL-5 dhe IL-6 si dhe nga CD23 (forma e tretshme e FcεRII me afinitet të ulët) e cila lidhet me CD21 të limfociteve B (receptori CR2/EBV) dhe që modulohet nga IFN alfa, IFN gama, TGF beta dhe IL-10.
Reaksionet anafilaktike shpërthehen nga bashkëlidhja (cross-linking) e molekulave të IgE në sipërfaqen e mastociteve.
Si mastocitet ashtu dhe bazofilet, pararëndësit qarkullues të te parëve, kanë në sipërfaqen e tyre receptorë me afinitet të lartë për regjionin lidhës Cε2:Cε3 të fragmentit Fc të IgE (FcεRI). Edhe IgG4 lidhet me anë të pjesës Fc në sipërfaqen e mastociteve, por me afinitet me të ulët se IgE. Bashkëlidhja nga antigeni i antitrupave të klasës IgE në sipërfaqen e mastociteve nxit degranulimin e mastociteve dhe bazofileve dhe si pasojë clirimin e mediatorëve. Sa me tepër vende lidhëse me antitrupat të këtë antigeni aq me efikase është nxitja e clirimit të mediatorëve: tetrameret janë me efikase se trimeret dhe këta të fundit me efikase se dimeret. Degranulimi induktohet gjithashtu kur IgE bashkëlidhen nga anti-IgE bivalente, por anti-IgE-te monovalente (Fab) janë inaktive. Urezimi i receptorëve FcεRI (bashkëlidhja e tyre) rezulton në një fosforilim të menjëhershem të vargjeve beta dhe gama, por ky reaksion rikthehet shpejt nga veprimi i fosfatazave pas clirimit të ligandit të lidhur me receptorët. Aktivizimi i receptorëve pasohet shpejt nga zbërthimi i fosfatidilinozitolit në inozitoltrifosfat (IP3), formimi i diacilglicerolit dhe nga një rritje e kalciumit të lire intracitoplazmatik. Kaskada biokimike prodhon “fuzogjene” membranoaktive si acidi lizofosfatidik, i cili favorizon fuzionin e membranave të granulave dhe degranulimin e tyre si dhe serine e metaboliteve të acidit arakidonik që formohen nga rrugët e ciklooksigjenazes dhe lipooksigjenazes. Përfundimisht, mediatorët e paraformuar të cliruar nga granulat përfshijne histaminën, heparinën, faktorët kemotaktike të eozinofileve dhe neutrofileve si dhe faktorin aktivizues të trombociteve, ndërsa leukotrienet LTB4, LTC4 dhe LTD4, PGD2 dhe tromboksanet sintetizohen rishtas. Gjithashtu kemi dhe clirim të IL-4, IL-5, IL-6 si dhe të GM-CSF, një profil citokinash tipik për limfocitet Th2.
Ne rrethana normale, të gjithë këta mediatorë nxisin zhvillimin e një reaksioni akut inflamator mbrojtës. Por në rast se kemi clirim masiv të këtyre mediatorëve në kushte jo normale si psh. në terren atopik, efekti bronkokonstriktor dhe vazodilatator predominon dhe bëhet kercenues.
Përgjigjet klinike ndaj Antigenëve të inhaluar: Pothuaj 10 % e popullates vuan në një masë më të lehtë apo me të rëndë nga alergji që përfshijnë
reaksionet anafilaktike të lokalizuara dhe të shkaktuara nga alergjenë ekstrinseke me ndërmjetësine e IgE. Ndër këta alergjene përmendim polenet, qimet e kafsheve apo ekskrementet e akarieneve (parazitë si Dermatophagoides pteryonissimus) që gjënden me shumicë në pluhurin e shtëpise. Kontakti i alergjenit me IgE të lidhur me membranën mastocitare të mukozave bronkiale, nazale, apo të indeve konjuktivale cliron mediatorët e anafilaksise dhe jep sipas rastit simptomat e astmës, rinitit alergjik apo konjuktivitit. Kjo lloj mbindjeshmerie është e tipit të shpejtë, por shumë të sëmurë atopikë zhvillojnë shënja të zgjatura pas ekspozimit ndaj alergjeneve dhe një sh]mbull tipik i kësaj lloj mbindjeshmërie është astma bronkiale. Të sëmurt me këtë sëmundje ndahen në trë kategori kryesore: 1. Astmë ekstrinseke të lidhur me atopi dmth. predispozicion gjenetik për të sintetizuar nivel të rritur të IgE ndaj alergjeneve të jashtëm. 2. Astmë intrinseke në të sëmurët jo-atopikë. 3.

84
Atmë okupacionale të shkatuar nga proteina specifike ose substanca kimike me peshë ë vogël molekulare.
Sëmundja është e karakterizuar nga ngushtim reversibël i rrugëve të frymëmarrjes i shoqëruar ky nga një ngacmueshmëri bronkiale e lartë ndaj një spektri të gjërë stimujsh irritue apo farmakllogjikë. Në secilën prej grupeve të mësipërme kemi dhe një inflamacion kronik ku përfshihen limfocitet T, makrofagët dhe sidomos eozinofilet.
Akumulimi preferencial i eozinofileve në këtë sëmundje vjen nga veprimi i IL-3, IL-5, dhe GM-CSF. Veçanërisht IL-5 nxit eozinofilet për të pësuar një veprim lokomotor tërheqës që ushtrohet nga PAF dhe LTB4 të mastociteve, rrit adezionin në endotelin vaskular mëpërmjet beta2-integrinave si dhe ul pragun ndaj stimujve që shkaktojnë çlirim të granulave. Proteina madhore bazike e eozinofileve shkakton një hipërreaktivitet të bronkeve dhe është toksike për qelizat epiteliale respiratore. Faktorë të tjerë patogjenikë lanë dhe mediatorët PAF dhe LTC4, që janë bronkokonstriktorë, ndërsa LTC4 është gjithashtu një stimulues i fuqishëm i sekrecioneve bronkiale. IL-5, i cili është një nga faktorët kryesorë të infiltrimit eozinofilik sekretohet pjesërisht nga mastocitet lokale, por në kronicitetin e sëmundjes kontribuon prodhimi i IL-5 dhe IL-4 nga L.T H2.
Predispozicioni atopik mund të çfaqet edhe në formën e dermatitit atopik nga akarienët e pluhurit të shtëpisë ose kafshët shtëpijake si psh. macet etj.
Shumë alergjene të tjerë mund të shkaktojnë alergji ushqimore në subjektë atopikë si psh. e bardha e vezës, qumështi i lopeës apo konservantë ushqimorë si sulfitet. Kontakti i këtyre substancave me IgE specifike të fiksuara në mastocitet e mukozës intestinale mund të shkaktojë një reaksion lokal me të vjella dhe diarre ose mund të lejojë që alergjeni të depërtojë lirisht nëpërmjet zorrës duke ndryshuar përmeabilitetin e mukozës së saj nga veprimi i mediatorëve. Alergjeni mund të kompleksohet kështu me antitrupa dhe mund të shkaktojë lezione në distancë si psh. artikulacionet. Ai mund të difuzojë gjithashtu në regjione të sensibilizuara si psh. lëkurë ose mushkëri ku mund të shkaktojë reaksione të tjera anafilaktike. Psh. ngrënja e manatokave ose e vezës mund të japin përkatësisht urtikarie ose atak astmatik në individë të sensibilizuar.
Faktorët gjenetikë: Ekziston në fakt një predispozicion i fortë familiar në cfaqjen e atopisë alergjike, por megjithëse
kjo është elidhur me një haplotip HLA të caktuar brënda një familje të dhënë asnjë lidhje e përgjithëshme me antigenë HLA të caktuar nuk është vënë në dukje. Një faktor gjenetik i vënë qartë në dukje është aftësia për tëë patur nivele të larta të IgE: Sa më i lartë niveli serik i IgE totale aq më i madh është probabiliteti për të zhvilluar reaksione apo sëmundje alergjike.
Testet klinike të alergjisë: Mbindjeshmëria alergjike vlerësohet zakonisht me anë të provave intrakutane me alergjenet më të
zakonshëm. Çlirimi i histaminës dhe i mediatorëve të tjerë jep shpejt një eritemë dhe papulë që arrijnë maksimumin brënda 30 minutave dhe pastaj tërhiqen. IgE përgjegjëse mund të vihen në dukje nëpërmjet anafilaksisë kutane pasive duke testuar aftësinë e serumit të të sëmurit pë të sensibilizuar në mënyrë pasive lëkurën e individëve normale (reaksioni i Prausnitz-Kustner). Ky sensibilizim pasiv mund të bllokohët duke injëktuar përpara serumit të të sëmurit IgE të gatshme mielomatoze por jo imunoglobulina mielomatoze të klasave të tjera. Kjo ndodh sepse receptorët përkatës në mastocitet e lëkurës saturohen me regjionet Fc të IgE mielomatoze dhe rrjedhimisht IgE e të sëmurit s’kanë se ku të fiksohen.
Reaksioni i menjëhershëm i mësipërm pasohet nga një reaksion i vonshëm që zgjat deri 24 orë. Ai karakterizohet nga një infiltrim i dëndur me eozinofile dhe limfocite T dhe është më edematoz se reaksioni i hershëm. Ngjashmëria me histopatollogjinë e infiltrimit inflamator të astmës kronike është e dukshme dhe këta reaksione të vonshme mund të shihen pas stimulimit të bronkeve dhe të mukozës bronkiale të subjekteve alergjikë.
Përsa i përket provave in vitro, përveç matjes së IgE totale serike një vlerë të rëndësishme diagnostike paraqet edhe matja e IgE specifike për alergjene të caktuar. Ekziston një korrelacion mjaft i mirë midis IgE specifike serike dhe pozitivitetit të provave të lëkurës. Edhe kur këta dy teste diagnostikë janë negativë stimulimi intranazal me alergen mund të rezultojë pozitiv si pasojë e prodhimit kryesisht lokal të IgE specifike.

85
Trajtimi: Pa dyshim trajtimi i parë është eliminimi i alergjenit shkaktar, por jo gjithmonë kjo është e
mundëshme pasi nqs alergjeni është ubikuitar si psh, akarienët e pluhurit të shtëpisë, myket e shtëpisë apo polenet e luleve, evitimi i tyre është shumë i vështirë.
Tentativat për të desensibilizuar të sëmurët nëpërmjet imunizimit me injëktime të përsëritura të alergenëve mund të çojnë në përmirësim të dukshëm, por jo në të gjithë të sëmurët. Mendohet se si rezultat i këtyre injëktimeve kemi sintezën e antitrupave “bllokues” funksioni i të cilëve është të pengojnë kontaktin e alergjenit me IgE specifike të fiksuara në mastocitet. Këta antitrupa bllokues mund të jenë të izotipeve IgG ose IgA dhe kanë funksion mbrojtës.
Gjithashtu duke patur parasysh që bashkëpunimi me limfocitet T është i rëndësishëm për sintezën e IgE dhe për veprimin e eozinofileve, veprimi dobiprurës i injëktimit të alergjeneve mund të induktojë limfocite T supresorë.
Përsa i përket terapisë medikamentoze veprim efikas kanë agjentë të tillë inhalatorë si sodium kromoglikati i cili lidhet në receptorë të veçantë mbi mastocite duke i bërë këta rezistentë ndaj degranulimit. Beta stimuluesit lokale luajnë rol të rëndësishëm bronkodilatator sidomos në acarimet e astmës bronkiale ndërsa kortizoniket lokale luajnë rol parandalues të riakutizimeve të astmës.
Simptomat e alergjisë mund të kontrollohen relativisht mirë (por jo gjithmonë) me anë të antihistaminikëve. Në reaksionet kronike alergjike nevojitet përdorimi i imunomodulatorëve si steroidet, apo së fundi i ciklosporinës A.
Tipi i dytë i reaksioneve të mbindjeshmërisë: Hipersensibiliteti citotoksik në vartësi të
antitrupave: Kur një antigen është i pranishëm në sipërfaqen e një qelizë, lidhja e tij me antitrupin përkatës do
të nxisë eliminimin e kësaj qelizë me mekanizmat si vijon: 1- Ulje e ngarkesës sipërfaqësore të qelizës. 2- Veprimi opsonizues i fragmenteve Fc të Ig nëpërmjet receptorëve përkatës në qelizat e sistemit imunitar. 3- Veprimi opsonizues i i C3 të aktivizuar në sipërfaqen e qelizës shënjë. 4- Vdekja qelizore mund të pasojë gjithashtu aktivizimin e të gjithë sistemit komplementar deri të C8 dhe C9 dhe duke shkaktuar dëmtim direkt membranor (hapje të poreve dhe lizë osmotike. 5- Qelizat shënjë të mbuluara me përqëndrime të ulta të IgG mund të vriten në mënyrë jo-specifike nëpërmjet një mekanizmi jo-fagocitar jashtëqelizor që realizohet nga qeliza limforetikulare jo të sensibilizuara paraprakisht, të cilat lidhen me qelizën shënjë me receptorë specifikë për pjesën Fc të IgG.
Ky citotoksicitet me ndërmjetësi antitrupore (ADCC = antibody dependent cell-mediated cytotoxicity) kryhet si nga qelizat e linjës mieloide (polimorfonuklearët dhe monocitet) ashtu dhe nga limfocitët e mëdhenj granularë (LGL = large granular lymphocytes) të cilët janë quajtur dhe qeliza K (killer), të cilët janë identikë me qelizat NK. Megjithëse ky fenomen deri tani është vënë re vetëm in vitro mendohet se ky fenomen ka vlerë fiziollogjike në rastet kur qelizat shënjë janë mjaft të mëdha për t’u vrarë nga fagocitoza si psh. parazitët e mëdhenj apo tumoret solidë.
Reaksionet e tipit të dytë me karakter aloimun: Ndër përbërësit e shumtë polimorfikë të membranës eritrocitare sistemi ABO i grupeve të gjakut
përbën sistemin dominant. Grupet antigenikë A dhe B e kanë prejardhjen nga substanca H nën veprimin e glycosyl transferazave që kodohen nga genet A dhe B respektivisht. Individët e grupit AB i kanë të dy antigenët në membranën e eritrociteve të tyre ndërsa ata të grupit 0 sintetizojnë vetëm substancën H sepse të ata mungojnë si geni A ashtu dhe ai B. Antitrupat anti-A apo anti-B çfaqen kur antigeni përkatës mungon: kështu një përson i grupit A do të ketë antitrupa anti-B dhe anasjelltës. Këto izohemaglutinina janë kryesisht të klasës IgM dhe i përkasin klasës së antitrupave natyralë: prodhimi i tyre stimulohet nga kontakti me antigenët e florës saprofite intestinale të cilët kanë strukturë të ngjashme me karbohidratet e grupeve të gjakut. Në këtë menyrë antitrupat e formuar kanë një reaksion të kryqëzuar me grupin eritrocitar që mungon tek ky individ. Nqs. një individ është me grup gjaku A, ai do të jetë tolerant ndaj antigenëve të ngjashëm me të ndërsa do të formojë antitrupa të aftë të aglutinojnë eritrocitet e grupit B. Në mënyrë të ngjashme individi i grupit O do të sintetiojë antitrupa si anti-A ashtu dhe anti-B. Në rast transfuzioni të papërshtatshëm erutrocitet e transfuzuara

86
do të mbulohen me antitrupat përkatës dhe do të ndodhë liza e shpejtë e tyre me pasojë reaksione të rënda deri vdekjeprurëse.
Papajtueshmëria në sistemin Rhesus: Grupi i gjakut Rhesus formon sistemin tjetër antigenik madhor në të cilin antigeni RhD është
shkaktari kryesor i reaksioneve izoimune si pasojë e aloimunizimit. Një nënë me grup gjaku RhD- (genotipi dd) mund të sensibilizohet nga eritrocitet e fëmijës kur ky ka antigenin RhD + (genotipi DD ose Dd). Kjo ndodh më shpesh në lindjen e fëmijes së parë kur nga gjakosja e placentës shumë eritrocite të fëmijës kalojnë tek nëna. Në këtë rast antitrupat e formuar janë të klasës IgG që janë të aftë të përshkolnë placentën në rast të një barrë tjetër. Veprimi i tyre me antigenin D të eritrociteve të fëmijës çon në shkatërrimin e këtyre të fundit kryesisht në sajë të aderencës opsonizuese duke shkaktuar në këtë mënyrë sëmundjen hemolitike të neonatit. Këta antitrupa nuk arrijnë të shkaktojnë aglutinimin e eritrociteve RhD+ in vitro (për shkak të densitetit të ulët të përcaktuesve antigenikë në sipërfaqen e eritrociteve) nqs. nuk përdoren albumina si mjedis apo dhe një serum anti-Ig (reagenti i Coombs ky i fundit).
Nqs. një nënë ka izohemaglutinina natyrale që reagojnë me eritrocitet fetale që arrijnë në qarkullimin e saj, sensibilizimi me antigenin D është më pak i mundshëm. Psh. një nënë grup O Rh- që ka një fëmijë grup A Rh+ do të shkatërrojë eritrocitet e fëmijës me anë të antitrupave anti-A të saj përpara se këta eritrocite të mund të provokojnë sintezëen e antitrupave anti-D. Sipas këtij principi nënat Rh+ në momentin r lindjes së një fëmije Rh- trajtohen në mënyrë profilaktike me antitrupa anti-D duke reduktuar kështu në një masë të konsiderueshme rrezikun e sensibilizimit.
Transplantet e organeve: Një organ i transplantuar, i cili i ka rezistuar reaksioneve me ndërmjetësi qelizore të flakjes akute,
mund të provokojë prodhimin e antitrupave tek bujtësi të drejtuar ndaj antigenëve sipërfaqësorë të të transplantimit (HLA) të dhuruesit. Këta antitrupa mund të veprojnë në mënyrë citotoksikë direkte mbi qelizat e organit të transplantuar ose mund të shkaktojnë aderencën e qelizave fagocitare ose sulmin “jospecifik” të qelizave K. Ata mund të skaktojnë gjithashtu aderencën trombocitare kur këta antitrupa ndërveprojnë me antigenë në sipërfaqen e endotelit vaskular. Ndërsa flakja hipërakute ndërmjetësohet nga antitrupa të paraformuar të marrësit.
Reaksionet autoimune të mbindjeshmerisë të tipit të dytë: Autoantitrupa ndaj eritrociteve të vetë pacientit vihen re në anemitë hemolitike autoimune.
Këta antitrupa bashkëveprojnë në 370C me përcaktues antigenikë (epitopë) të antigenëve të kompleksit Rhesus të ndryshëm nga ata që provokojnë reaksionet e transfuzionit. Eritrocitet e mbuluara me këta antitrupa kanë një gjysëm-jetë më të shkurtër kryesisht si rrjedhojë e aderimit të tyre me qelizat fagocitare të shpretkës. Mekanizma të ngjashëm janë përgjegjës edhe për aneminë në të sëmurët me hemaglutinina të ftohta të cilat janë antitrupa monoklonalë anti-I (antigen sipërfaqësor i eritrociteve) që krijohen pas infeksionit me Mycoplasma pneumoniae si dhe në disa raste të hemoglobinurisë paroksizmale nga të ftohtit të lidhur me antitrupat lizues të Donath-Landsteiner që janë specifikë për grupin P të gjakut. Këta antitrupa janë kryesisht të klasës IgM dhe bashkëveprojnë në tempëratura nën 370C.
Serumet e të sëmurëve me tiroidit të Hashimoto përmbajnë antitrupa që në prani të komplementit janë citotoksikë për qelizat tiroidiene. Në sindromin e Goodpasture hasen nga ana tjetër antitrupa ndaj membranës glomerulare të veshkave por dhe të mushkërive. Biopsitë kanë vënë në dukje edhe fiksimin e komplementit bashkë me antitrupat në këtë membranë dhe si rrjedhojë dëmtimin e kësaj të fundit.
Një shëmbull tjetër i kësaj lloj mbindjeshmërie është dëmtimi nga autoantitrupa i receptorëve për acetilkolinën në sëmundjen myasthenia gravis.
Reaksionet medikamentoze me tip të dytë të mbindjeshmërisë: Medikamentet mund të lidhen me përbërës të organizmit dhe kështu nga haptenë të shëndrrohen
në antigenë të plotë të aftë të sensibilizojnë disa individë. Nqs. prodhohen antitrupa të klasës IgE mund të kemi reaksione anafilaktike. Në disa rrethana, sidomos me pomada apo krema me përdorim lokal mund të çfaqet mbindjeshmëri me ndërmjetësi qelizore. Në disa raste të tjera, sidomos kur medikamenti lidhet me proteina serike, mund të kemi reaksione të tipit të tretë të mbindjeshmërisë.

87
Reaksionet e tipit të dytë çfaqen zakonisht kur medikamenti lidhet me sipërfaqen e një qelizë gjaku dhe dhe për rrjedhojë provokon prodhimin e antitrupave që janë citotoksikë për kompleksin qelizë-medikament. Kur medikamenti eliminohet plotësisht zhduket dhe kjo lloj mbindjeshmërie. Shembuj të kësaj mbindjeshmërie janë parë në aneminë hemolitike nga marrja e klorpromazinës apo fenacetinës, në agranulocitozën e lidhur me amidopirinën apo kinidinën apo në purpurën trombocitopenike të shkaktuar nga sedativi Sedormid. Në rastin e fundit serumi i freskët i të sëmurit lizon trombocitet në prani të Sedormid-it, por jo kur ky s’është i pranishëm. Inaktivizimi i komplementit me anë të ngrohjes së serumit në 560C për 30 minuta e pengon këtë efekt gjë që tregon se aktivizimi i komplementit është i domosdoshëm në këtë mekanizëm.
KAPITULLI 22. MEKANIZMAT IMUNOPATOLOGJIKE, TIPI I III DHE IV
Klasifikimi sipas Gell dhe Coombs i mekanizmave imunopatologjike të mbindjeshmerisë të
tipit të tretë dhe të katërt. Shembuj të tyre.
TIPI I TRETE I REAKSIONEVE IMUNOPATOLOGJIKE (MBINDJESHMËRIA E TIPIT TË VONUAR):
Organizmi mund t'i ekspozohet një teprice antigeni gjatë një përiudhe të zgjatur kohë në një numër rrethanash si në infeksione të zgjatura, sëmundje autoimune apo në kontakte të vazhdueshme me agjentë të mjedisit. Komplekset antigen-antitrup të formuar në këtë mënyrë mund të precipitojnë në inde apo organe të caktuara dhe të shkaktojnë reaksione inflamatore. Nëqoftëse kemi dhe fiksim të komplementit do të ndodhë çlirim i anafilatoksinave (C3a dhe C3b) dhe degranulim të mastociteve me vazodilatacion dhe rritje të përmeabilitetit kapilar si pasojë. Faktorë kemotaktikë gjithashtu të prodhuar (si psh. C3a) do të shkaktojnë një tërheqje në vatrën inflamatore të polimorfonuklearëve të cilët do të fillojnë fagocitozën e komplekseve imune. Si pasojë e këtyre fenomeneve do të kemi çlirim ekstraqelizor të përmbajtjes së granulave të neutrofileve. Kur kompleksi imun depozitohet mbi një membranë bazale dhe nuk mund të fagocitohet kemi të ashtuquajturën fagocitozë të frustruar. Në këtë rast do të çlirohen me shumicë enzimat proteolitike (ku përfshihen proteinazat dhe kolagenazat), enzimat formuese të kininës, proteinat polikationike, dhe përbërësit reaktivë të oksigjeni dhe azotit, të cilat do të shkaktojnë dëmtim lokal të indeve dhe intensifikim të reaksioneve inflamatore. Dëmtim i

88
mëtejshëm mund të shkaktohet nga liza reaktive si rrjedhojë e aktivizimit të C5,6,7,8,9. Nqs. krijohen kushtet mund të kemi agregim trombocitar me dy pasoja: 1 - çlirim të mëtejshë të aminave vazoaktive; 2 - Formimi i mikrotrombeve që çon në iskemi lokale. Nga ana tjetër komplekset imune të patretshme të fagocituara nga makrofagët nuk arrijnë të treten dhe përbëjnë një stimul aktivizues përsistent që nxit çlirimin e citokinave IL-1 dhe TNF, të përbërësve reaktivë të oksigjenit si dhe të oksidit nitrik.
Kompleksi antigjen - antitrup
Agregim trombocitar Aktivizim i komplementit Aktivizim i makrofagëve
Mikrotrombe
Çlirim aminashvazoaktive
Çlirim enzimashproteolitike dheproteinashpolikationike
Çlirim i mediatorëvemastocitarë
Çlirim i IL-1, TNF, R.O.INO
Tërheqje e poli-nuklearëve
Anafilatoksinat
Fig. : Tipi i III i mbindjeshmërisë (me ndërmjetësinëe komplekseve imune)
Pasojat e formimit të komplekseve imune in vivo varen jo vetëm nga sasitë absolute të
antigenit dhe antitrupit, të cilat përcaktojnë intensitetin e reaksionit, por gjithashtu dhe nga raportet relative të tyre me njëri tjetrin, të cilat përcaktojnë natyrën e kompleksit dhe rrjedhimisht shpërndarjen dhe fatin e tyre në organizëm. Midis tepricës në antitrupa dhe tepricës së lehtë në antigen, komplekset precipitojnë shpejt dhe kanë tendencë të lokalizohen në vendin e futjes së antigenit, ndërsa në tepricë të moderuar ose të shprehur në antigen formohen komplekse imune të tretshëm.
Fati i këtyre komplekseve është i lidhur ngushtë me aktivizimin e rrugës klasike të komplementit. Fiksimi i komplementit inhibon precipitimin e komplekseve imune nëpërmjet lidhjes kovalente të C3b, i cili parandalon ndërveprimet Fc - Fc të nevojshme për të krijuar komplekse të mëdha të patretshme. Këta komplekse të vegjël që përmbajnë C3b lidhen me aderencë imune në receptorët CR1 të komplementit në eritrocite dhe transportohen në makrofagët e heparit ku ata inaktivizohen. Kur kemi difekte në këtë sistem psh. mangësi në rrugën klasike të komplementit ose në receptorët CR1 të eritrociteve, ose në rast mbingarkese të sistemit, atëhere komplekset imune do të jenë të lirë në serum dhe ata do të depozitohen në veshka, kyçe apo në lëkurë.
Dëmtimet inflamatore si pasojë e komplekseve imune të depozituar lokalisht: Reaksioni i Arthus: Maurice Arthus vuri në dukje që injëktimi intrakutan i një antigeni të tretshëm në lepuj të
hipërimunizuar dhe me nivele të larta antitrupash precipitues do të japë një reaksion edematoz dhe eritematoz që arrin një maksimum pas 3-8 orësh dhe më pas zhduket. Lezioni karakterizohet nga një infiltrim i theksuar me leukocite polimorfonukleare. antigeni shpesh precipiton bashkë me antitrupin përkatës brënda në venulë pa i lënë kohë rrugës klasike të komplementit të aktivizohet dhe të parandalojë precipitimin. Aktivizimi i komplementit bëhet më pas, pas precipitimit dhe do tëkemi prodhim të anafilatoksinave me pasojë degranulimin e mastociteve. Komplekset imune lokale me lokalizim intravaskular do të shkaktojnë gjithashtu agregim trombocitar dhe çlirim të aminave vazoaktive me pasojë rritjen e eritemës dhe të edemës. Formimi i faktorëve kemotaktikë do të çojë në një influks të polimorfonuklearëve. Reaksioni i Arthus mund të bllokohët me anë të deplecionit

89
(eliminimit) të komplementit ose të neutrofileve polimorfonuklearë (me nitrogjen mustard ose serum specifik antipolimorfonuklearë).
Reaksionet ndaj antigenëve të inhaluar: Reaksionet intrapulmonarë të tipit Arthus ndaj antigenëve ekzogjene të inhaluar duket se
janë përgjegjës për një sërë çrregullimesh të hipërsensibilitetit të njëriu. Crregullimet e rënda respiratore të lidhura me "mushkërinë e fermerit" ndodhin brënda 6-8
orësh pas ekspozimit ndaj pluhurit të kashtës së mykur. Të sëmurët janë të sensibilizuar ndaj Actinomycetave termofilike që rriten në kashtën e mykur. Ekstraktet e këtyre mikroorganizmave japin reaksione precipitimi me serumin e të sëmurit si dhe reaksion të tipit Arthus nëqoftëse injëktohet intrakutan. Inhalimi i sporeve bakteriale të pranishëm në pluhurin e kashtës bën që antigeni të depërtojë në mushkëri dhe të ndodhë një reaksion hipërsensibiliteti me ndërmjetësi nga komplekset imune. Situata të ngjashme ndodhin në sëmundjen e rritësve të pëllumbave në të cilën antigeni është një proteinë serike e pranishme në pluhurin e feceve të thara, në rritësit e mijve të sensibilizuar nga proteinat serike të miut të eliminuara në urinë, si dhe në raste të tjera të ngjashme të emërtuara si alveolit alergjik ekstrinsek që shkaktohet nga inhalimi i vazhdueshëm i pjesëzave organike si psh. në sëmundjen e larësve të djathit (sporet e Penicillium casei), në mushkerinë e përgatitësit të gëzofeve (proteinat e gëzofit të dhelprës) etj. Në disa situata patollogjike të tilla ka mundësi që të ndërhyjnë edhe mekanizma të hipërsensibilitetit të tipit të parë që ndërhyjnë në fillimin e reaksionit të Arthus. Kështu, të sëmurët me Aspërgilozë bronkopulmonare alergjike kanë si nivele të larta të IgE ashtu dhe IgG precipituese ndaj species së Aspërgillus. Reaksionet ndaj antigenëve të brëndshëm:
Reaksionet e tipit të tretë provokohën shpesh nga clirimi lokal i antigenit nga mikroorganizma infeksiozë brënda trupit. Psh. krimbat e gjallë të filaria si psh. Wuchereria bancrofti janë relativisht të padëmshëm, por parazitët e ngordhur të pranishëm në enët limfatike shkaktojnë një reaksion inflamator që mendohet se është përgjegjës për obstruktimin e enëve limfatike dhe si pasojë për elefantiazën që mund të pasojë. Antibioterapia mund të shkaktojë clirim të theksuar të të antigenëve bakterialë, të cilët në individët me nivel të lartë të antitrupave mund të shkaktojnë reaksione dramatike inflamatore me ndërmjetësinë e komplekseve imune. Kështu psh. shpjegohet eritema nodoze leproze në lëkurën e të sëmurëve me lepër lepromatoze nën trajtim me dapson.
Një variant interesant i reaksionit Arthus është vënë re në artritin reumatoid në të cilin formohen komplekse imune lokalisht në artikulacione si rrjedhojë e bashkëveprimit IgG - anti-IgG (kjo e fundit prodhohet nga plazmocitet sinoviale).
Sëmundje që shkaktohen nga komplekset imune: Sëmundja e serumit: Injëktimi i dozave të larta të serumit heterolog (psh. serumi anti difteritik apo antitetanik i
kalit) është përdorur gjërësisht për arësye terapeutike. Rreth tetë ditë më pas mund të cfaqet një gjëndje e quajtur si sëmundje e serumit: Kemi rritje të tempëraturës, enjtje të gjëndrave limfatike, eritemë urtikariale të gjeneralizuar si dhe poliartrit të shoqëruar me një ulje të komplementit serik si dhe proteinuri tranzitore. Të gjitha këto shkaktohen nga depozitimi i komplekseve antigen-antitrup që formohen në tepricë të antigenit:
Këta individë fillojnë të sintetizojnë antitrupa ndaj proteinave të huaja, zakonisht globulinave të kalit. Meqenëse antigeni është akoma i pranishëm në tepricë të konsiderueshme në këtë moment, formohen komplekse qarkullues të tretshëm me përbërje Ag2Ab, Ag3Ab2, Ag4Ab3 etj. Që të jenë patogjenikë, komplekset imune duhet të kenë madhësinë e nevojshme: shumë të mëdhej ata gëlltitem kollaj nga makrofagët ndërsa kur janë shumë tëvegjël (<19S) ata nuk janë të aftë të induktojnë një reaksion inflamator. Edhe kur ata kanë madhësinë e nevojshme, ata lokalizohen në parëtet vaskulare vetëm nqs. këta kanë pësuar një ndryshim në përmeabilitetin vaskular. Kjo mund të ndodhë nëpërmjet clirimit të 5-hidroksitriptaminës (5HT) nga trombocitet që bashkëveprojnë me komplekse të mëdhenj ose dhe nëpërmjet formimit të histaminës, leukotrieneve dhe faktorit aktivizues të trombociteve (PAF) nga degranulimi i bazofileve dhe mastociteve me ndërmjetësinë e IgE apo të komplementit. Veprimi në kapilarë shkaktohet nëpërmjet shqitjes së qelizave endoteliale dhe ekspozimit të membranës bazale ku fiksohen komplekse imune me madhësi të përshtatshme. Indet më të ekspozuara

90
për këto efekte janë lëkura, kycet, veshkat dhe zemra. Me rritjen e sintezës së antitrupave kemi eliminimin e antigenit dhe shërimin e të sëmurit.
Glomerulonefritet nga komplekset imune: Depozitimi i komplekseve imune është një proces dinamik dhe sëmundjet kronike vihen re
vetëm kur antigeni përsiston në organizëm si në infeksionet kronike dhe sëmundjet autoimune. Dixon provokoi lezione glomerulare kronike në lepujt nëpërmjet administrimit të përsëritur në to të proteinave të huaja. Jo cdo kafshë cfaqi lezione dhe ndofta vetëm ato që janë gjenetikisht të afta të prodhojnë antitrupa me afinitet të ulët ose antitrupa ndaj një numri të kufizuar përcaktuesish antigenikë. Me sa duket vetëm këta kafshë cfaqën komplekse të tretshëm me madhësinë e nevojshme. Komplekset më të vegjël arrijnë në anën epiteliale të glomerulit, por komplekset progresivisht më të mëdhenj kapen në/ose mbi anën endoteliale të membranës bazale glomerulare. Ata grumbullohen si granula të trasha që, në imunofluoreshencë ngjyrosin pozitivisht pëer antigen, imunoglobulina si dhe komplement. Në mikroskopi elektronike këto depozitime duken si masa të mëdha amorfe.
Shumë raste me glomerulonefrit janë të shoqëruara me komplekse imune qarkulluese dhe në biopsi vihet re një imunofluoreshencë e ngjashme me atë të komplekseve DNA/anti-DNA/komplement në tësëmurët me lupus eritematoz sistemik. I njohur mirë është dhe glomerulonefriti që mund të pasojë infeksionin nga disa shtame streptokokësh të quajtur nefritogjene apo dhe sindromi nefrotik në fëmijë nigerianë të prekur nga malaria kuartane ku janë implikuar komplekse imune me Antigenë të plasmodiumit.
Glomerulonefriti nga komplekset imune mund të cfaqet dhe gjatë një infeksioni viral: psh. mijtë të infektuar nga virusi i koriomeningitit llimfocitar zhvillojnë një glomerulonefrit ku hasen komplekse imune të përbërë nga virusi dhe antitrupi. Ky model mund të haset dhe të njëriu.
Depozitimi i komplekseve imune në inde të tjera: Pleksusi koroid i sistemit nervor është një vend kryesor filtrimi dhe si rrjedhojë favorizohet
depozitimi në të i komplekseve imune në LES, gjë që shpjegon shpeshtësinë e shënjave të prekjes së sistemit nervor qëndror në këtë sëmundje. Të sëmurët me SLE dhe me prekje neurollogjike kanë ulje të C4 në lëngun cerebrospinal (LCS) dhe pas vdekjes në të sëmurët me LES, crregullime neurologjike dhe titër të lartë të anti-ADN janë vënë në dukje depozitime të shpërndara të imunoglobulinave dhe të ADN në pleksusin koroid. Në panëncefalitin subakut sklerozues vihet re një një raport i ngritur LCS/serum i antitrupave anti-fruth, ndërsa në indin nervor janë kapur depozitime imunoglobulinash ndër të cilat hasen antitrupa anti-fruth.
Cfaqjet vaskulitike të lëkurës janë një karakteristikë kryesore e sëmundjes së serumit, por dhe e lupusit eritematoz sistemik dhe atij diskoid. Biopsia e lëkurës vë në dukje në këtë rast depozitime amorfe të të imunoglobulinave si dhe të përbërësit C3 të komplementit në membranën bazale të lidhjes dermo-epidermale.
Nga ana tjetër, arteriti nekrotizues i cfaqur në lepuj me anë të sëmundjes ekspërimentale të serumit i ngjan mjaft histollogjisë së përiarteritit nodoz dhe në lezionet e kësaj sëmundje janë vënë në dukje komplekse imune që përmbajnë antigenin sipërfaqësor të hepatitit B. Në disa raste të tjera, medikamente të tilla si penicilina bëhen antigenike pas lidhjes me proteina të organizmit dhe shkaktojnë formimin e komplekseve imune që ndërmjetësojnë reaksione të hipërsensibilitetit.
TIPI I KATËRT I REAKSIONEVE IMUNOPATOLOGJIKE (MBINDJESHMËRIA E TIPIT TË
VONUAR): Kjo lloj mbindjeshmërie haset në shumë reaksione alergjike ndaj bakterieve, viruseve dhe
këpurdhave, në dermatitet e kontaktit nga sensibilizimi me substanca kimike të thjeshta si dhe në flakjen e indeve të transplantuara. Shëmbulli më i njohur i këtij lloj hipërsensibiliteti është “Reaksioni i Mantoux” që përftohet nga injëktimi i tuberkulinës në lëkurën e një individi në të cilin kontakti i mëparshëm me Mykobakteriumin e tuberkulozit ka krijuar një gjëndje imuniteti me ndërmjetësi qelizore (INQ). Reaksioni karakterizohet nga skuqje dhe indurim i lëkurës në vëndin e injëktimit. Reaksioni cfaqet vetëm pas disa orësh (prandaj dhe termi “i vonuar”) dhe arrin maksimumin pas 24-48 orë duke u pakësuar më pas. Histologjikisht faza e hershme e reaksionit

91
cfaqet si një infiltrat përivaskular me qeliza mononukleare që pasohet nga një përqëndrim i shprehur qelizash mono dhe polimorfonukleare.
Qelizat polimorfonukleare migrojnë shpejt larg vendit të inflamacionit duke lënë atje një infiltrat me qeliza mononukleare ku predominojnë limfocitet dhe qelizat e serisë monocitare-makrofagëale. Kjo është në kontrast me karakterin kryesisht polimorfonuklear të reaksionit të Arthus.
Ndryshe nga format e tjera të mbindjeshmërisë, tipi i katërt nuk mund të transferohet nga një individ i sensibilizuar në një tjetër të pasensibilizuar me anë të antitrupave, pra me anë të serumit por me anë të limfociteve T.

92
KAPITULLI 23. AUTOIMUNITETI
Ç’janë reaksionet dhe sëmundjet autoimune. Hipotezat e ndryshme që shpjegojnë
çfaqjen e tyre. Karakteristikat e sëmundjeve autoimune.
Repertori kolosal i sistemit imunitar adoptiv (të fituar) ka evoluar në mënyrë të tillë që të mund të njohë çdo molekulë të huaj për organizmin e gjallë. Por ky evolucion nuk ka mundur të pengojë plotësisht gjenerimin e limfocitëve që mund të reagojnë edhe ndaj përbërësve të vetë organizmit. Kjo edhe për faktin se megjithëse shumë mekanizma fiziollogjikë parandalojnë reagimin e sistemit imunitar ndaj përbërësve të vetvehtes, ashtu si me çdo mekanizëm të komplikuar ekziston gjithmonë mundësia e mosfunksionimit të rregullt dhe sa më i moshuar individi aq më e madhe kjo mundësi. Pa marrë parasysh praninë pothuajse të rregullt të autoantitrupave të klasës IgM me afinitet të ulët, që prodhohen nga limfocitet B CD5+ si pjesë e antitrupave natyralë, fenomenet autoimune shoqërojnë mjaft sëmundje tek njëriu.
Në mënyrë ideale termi sëmundje autoimune duhej të përdorej për ato sëmundje në të cilat reaksionet autoimune janë shkaku direkt i patogjenezës së tyre, dhe jo për ato situata në të cilat çfaqjet autoimune psh. autoantitrupat shkaktohen si pasojë e dëmtimit indor (si psh. autoantitrupat anti-ind kardiak që çfaqen pas infarktit të miokardit). Por, roli i autoimunitetit në shumë sëmundje është akoma jo i qartësuar mirë, prandaj edhe në këto raste në do të flasim për sëmundje autoimune, përveç se në ato patollogji ku është e qartë që fenomenet autoimune janë thjesht çfaqje dytësore.
Spektri i sëmundjeve autoimune (SA): SA mund të konsiderohen sikur formojnë një spektër (Tabela ): Në një skaj kemi
“Sëmundjet autoimune organ-specifike (SAOS)” në të cilat hasim autoantitrupa organ-specifike. Sëmundja e Hashimoto-s që prek gjëndrën tiroide është një shëmbull: në të vihet re infiltrim me qeliza mononukleare (limfocite, histiocite dhe plazmocite), shkatërrim i qelizave folikulare, formim qëndrash gjerminative dhe prodhim autoantitrupash qarkullues me specificitetë për disa përbërës tiroidienë. Drejt qëndrës së spektrit mund të vendosim ato sëmundje në të cilat lezionet kanë tendencë të lokalizohen në një organ të veçantë por antitrupat nuk janë organ-specifikë. Një shëmbull tipik është cirroza biliare primare në të cilën duktuli biliar është objekti kryesor i infiltrimit qelizor, por antitrupat serikë të pranishëm janë anti-mitokondrialë, pra pa specifitet për një organ të caktuar.
Në skajin tjetër të spektrit janë “Sëmundjet autoimune jo-organ specifike (SAJOS) ose sistemike” ku përfshihen shumë sëmundje reumatollogjike me shëmbull tipik Lupusin eritematoz sistemik (LES). Në këto sëmundje si lezionet ashtu dhe autoantitrupat nuk janë të kufizuar në një organ të caktuar. Ndryshimet patollogjike janë difuze dhe janë kryesisht lezione të indit lidhor të shoqëeuara me nekrozë fibrinoide. Lezionet hasen në lëkurë (eritema lupike në formë fluture është mjaft tipike), veshka, artikulacione, seroza, enë gjaku etj. Preken gjithashtu dhe elementët e formuar të gjakut.
Autoantitrupat që hasen janë të larmishëm dhe bashkëveprojnë me ADN dhe përberës të tjerë bërthamorë (ribonukleoproteinat psh.) apo citoplazmikë të të gjithë qelizave të organizmit. Organ - Specifike - Tiroiditi i Hashimoto - Miksedema primare
-Tirotoksikoza -Anemia përnicioze
-Gastriti autoimun atrofik

93
- Sëmundja e Addison - Menopauza premature (pak raste) - Infertiliteti mashkullor (pak raste) - Miastenia grave - Diabeti juvenil - Sindromi i Goodpasture - Pemfigus vulgaris - Pemfigoidi - Oftalmia simpatike - Uveiti fakogjenik - Skleroza multiple - Anemia hemolitike autoimune - Purpura trombocitopenike idiopatike - Leukopenia idiopatike - Cirroza primare biliare - Hepatiti kronik aktiv HBs negativ - Koliti ulçeroz - Sindromi i Sjogren - Artriti reumatoid - Skleroderma - Granulomatoza e Wegener - Poli/Dermatomioziti - Lupusi eritematoz diskoid - Lupusi eritematoz sistemik (LES)
Jo - specifike për organin (sistemike)
Spektri i sëmundjeve autoimune
Mbivendosja e sëmundjeve autoimune: Njihet tendenca që të i njëjti i sëmurë të haset më tepër se një sëmundje autoimune dhe kjo
ndodh zakonisht me sëmundje që ndodhin në të njëjtin regjion të spektërit autoimun. Kështu, të sëmrët me tiroidit autoimun (Sëmundja e Hashimoto ose miksedema primare) kanë një incidencë më të lartë të anemisë përnicioze se sa mund të pritej nga rastësia e thjeshtë(10% ndaj 0.2%). Gjithashtu, si tiroiditi dhe tirotoksikoza hasen në të sëmurët me anëmi përnicioze me një shpeshtësi relativisht të lartë. Lidhje të tjera janë parë ndërmjet sëmundjes së Addison dhe sëmundjeve autoimune të tiroides si dhe në rastet e rralla të të rinjve me anëmi përnicioze dhe poliendokrinopati ku përfshihen sëmundja e Addison, hipoparatiroidizmi, diabeti dhe tiroiditi.
Ekziston gjithashtu edhe një mbivendosje e gjëre në hasjen e autoantitrupave. 30% e të sëmurëve me sëmundje autoimune të tiroides kanë edhe antitrupa ndaj qelizave parietale të stomakut. Gjithashtu, antitrupat antitiroidienë hasen deri në 50% të të sëmurëve me anëmi përnicioze.
Përsa i përket kahut të sëmundjeve jo-specifike për antigenin, sëmundjet sistemike autoimune si psh. LES (lupusi eritematoz sistemik) shoqërohen shpesh me artritin reumatoid si dhe me sëmundje të tjera autoimune si anemia hemolitike, leukopenia idiopatike, purpura trombocitopenike, dermatomioziti apo sindromi i Sjogren. Një karakteristikë e zakonshme e këtij regjioni të spektrit janë dhe antitrupat antibërthamorë si dhe faktori reumatoid.
Të sëmurët me SAOS kanë një tendencë të zhvillojnë një transformim neoplazik në organin e prekur, ndërsa ata me SAJOS neoplazira të gjeneralizuara limforetikulare.
Faktorët që ndikojnë në çfaqjen e sëmundjeve autoimune: Faktorët gjenetikë në sëmundjet autoimune: Fenomenet autoimune kanë tendencë të çfaqen me frekuencë më të madhe brënda një familje
të caktuar. Psh. të afërmit e gradës së parë (vëllezërit/motrat, prindët dhe fëmijët) e të sëmurëve me

94
sëmundje të Hashimoto kanë një incidencë të lartë të antitrupave antitiroide si dhe të tiroiditit subklinik dhe klinik. Gjithashtu autoantitrupat hasen më shpesh në ato familje në të cilat më shumë se një antar janë të prekur nga sëmundja. Përsa i përket sëmundjeve sistemike, në të afërmit e të sëmurëve me LES kemi një prekshmëri më të lartë nga sëmundje të indit lidhor si dhe çrregullime të sintezës së imunoglobulinava.
Këto lidhje familjare mund t’i dedikohën edhe faktorëve mjedisorë si psh. mikroorganizmave, por ka fakte të forta që flasin për rolin e rëndësishëm të faktorëve gjenetikë. Një lidhje e rëndësishme ekziston dhe midis disa sëmundjeve autoimune dhe antigenëve HLA si psh. DR3 në sëmundjen e Addison apo DR4 në artritin reumatoid.
Në 50% të rasteve diabeti juvenil insulinovartës (DJIV) shfaqet në të dy binjakët homozigotë. Kjo shifër është shumë më e ulët në binjakët dizigotikë. Por, nqs. binjakët monozigotë janë DR3/DR4, DJIV do të shfaqet në të dy fëmijët në 75% të rasteve dhe vetëm në 40% të rasteve nqs nuk kanë haplotipin e mësipërm të HLA. Këto fakte tregojnë rolin e rëndësishëm të faktorit gjenetik dhe sidomos të sistemit HLA. Në sëmundjet sistemike si LES kontributi gjenetik është më i ulët me një përqindje konkordance prej 23% në homozigotët dhe prej vetëm 9% në dizigotikët. Ekzistojnë gjithashtu shumë shëmbuj të afërmish asimptomatikë të të sëmurëve me LES, të cilët kanë një prani më të shpeshtë të antitrupave antibërthamorë nqs kanë jetuar bashkë me të sëmurin në krahasim me ata që kanë jetuar larg nga i sëmuri. Sidoqoftë, në tënjëjtën banesë, bashkëshortja ka më pak gjasi të ketë autoantitrupa në krahasim me të afërmit me lidhje gjaku. Faktet e gjetura në sëmundjet autoimune humane dëshmojnë për faktorë gjenetikë multifaktorialë. Pra janë disa gene që ndikojnë në çfaqjen e sëmundjeve autoimune. Këto gene duket se janë të ndryshë në SAIOS në krahasim me SAIJOS.
Si konkluzion, në disa sëmundje faktorët gjenetikë luajnë rol madhor ndërsa në të tjera janë faktorët mjedisorë që predominojnë.
Ndikimi i seksit në sëmundjet autoimune: Ekziston një tendencë e përgjithshme që sëmundjet autoimune të çfaqen më shpesh në seksin
femër se në atë mashkull. Mendohet se estrogjenet mund të luajnë një rol në këtë drejtim. Barra është shpesh e shoqëruar me një përmirësim të sëmundjes si psh. në artritin reumatoid dhe nganjëherëpas lindjes, kur ndodhin ndryshime drastike hormonale, ndodh një acarim i saj. Duhet shënuar gjithashtu shpeshtësia e çfaqjes së hipotiroidizmit pas lindjes në gratë me sëmundje autoimune preekzistuese të tiroides.
Veprimi i mjedisit: Faktorët jo infeksiozë: Ndër faktorët jo-infeksiozë përmëndim dietën. Vajrat e peshkut me përmbajtje acidesh
yndyrore me vargje të gjatë të pasaturuar të tipit omega-3 janë të njohur si mbrojtës ndaj artritit reumatoid. Rrezatimi diellor është një ngacmues i njohur i dëmtimeve të lëkurës në LES. Gjithashtu njihet fakti që ekspozimi ndaj tretësve organikë mund të predispozojë për dëmtim autoimun të membranave bazale: individët DR2 që punojnë në pastrimet kimike kanë një incidencë të lartë të sindromit të Goodpasture.
Faktorët mikrobialë: Ethja reumatizmale pason një tonsilit me streptokok të grupit A në 2-3% të individëve me
predispozicion gjenetik për këtë sëmundje. Në shumicën e sëmundjeve autoimune kronike ekziston një kohë e gjatë latence midis një infeksioni probabilisht shkaktar të sëmundjes dhe vetë çfaqjes së saj. Nga ana tjetër është shumë e vështirë në mos e pamundur të izolohen mikroorganizma të gjallë nga indet e prekur.
Disa studiues janë përqëndruar në disa lloje mykobakteriesh me rritje shumë të ngadaltë të cilët janë jashtëzakonisht vështërë të kultivohen por duket se janë të lidhur me disa gjëndje mbindjeshmërie. Psh. M. paratuberculosis duket se është i lidhur me sëmundjen e Johne, një infeksion intestinal granulomatoz kronik tek bovinët që duket i ngjashëm me sëmundjen e Crohn. Tek njëriu, me anë të sondave gjenetike janë identifikuar sekuenca të mykobakterieve në sarkoidozë. Artriti granulomatoz i Takayasu shoqërohet shpesh me përgjigje të fuqishme ndaj M. Tuberculosis. Sëmundja e Whipple, e shoqëruar me artralgji dhe ndryshime në lëkurë, evoluon kaq ngadalë sa

95
kalojnë të paktën 10 vjet deri në vendosjen e diagnozës. Megjithëse me anë të mikroskopisë elektronike janë vënë në dukje mikroorganizma në brëndësi të makrofagëve, ka qënë e pamundur që ata të kultivohen. Një zbulim interesant në artritin reaktiv të lidhur me HLA-B27 dhe të provokuar nga infeksioni me klamidia, yersinia ose salmonela, ka qënë fakti i pranisë në këta të sëmurë i përgjigjes imunitare të limfociteve T ndaj fragmenteve bakteriale edhe vite pas infeksionit primar.
Sot tashmë njihet që reaksione të kryqëzuara me përbërës mikrobialë mund të iniciojnë autoimunitetin. Kohët e fundit është vënë në dukje që infeksioni me helmintin Nippostrongilus brasiliensis mund të prishë tolerancën ndaj një supërantigeni stafilokoksik të pa lidhur me të. Ndoshta infeksionet e kësaj natyre mund të stimulojnë një prodhim të pazakontë citokinash, të cilat mund të aaktivizojnë limfocite T potencialisht autoreaktive. Kompleksiteti i gjëndjes shtohet nga njohja që mikrobet mjedisorë mund edhe të mbrojnë ndaj sëmundjeve spontane autoimune. Incidenca e diabetit rezulton e shtuar nqs. mijtë NOD ruhen në kushte sterile, ndërsa virusi Sendai inhibon zhvillimin e artritit në mijtë MRL/lpr. Gjithashtu, variacioni i jashtëzakonshëm në incidencën e diabetit në mijtë NOD në vartësi kjo të kushteve mjedisore ku ata rriten, tregon për ndikimin e rëndësishëm të florës mjedisore në çfaqjen e sëmundjeve autoimune.
Autoreaktiviteti vjen në mënyrë të “natyrshme”: Mekanizmat e tolerancës nuk arrijnë të eliminojnë të gjithë limfocitët autoreaktivë.
Përpunimi i një autoantigeni përfundon që disa peptidë të ashtuquajtur “dominantë“ paraqiten në mënyrë preferenciale nga qelizat antigen-paraqitëse (QAP), ndërsa të tjerë, të ashtuquajtur “kriptikë“, çfaqen në gropëzën e KMPI vetëm në përqëndrime shumë të ulta gjë që nuk lejon gjenerimin e sinjalit për seleksionim negativ në timus të limfociteve T koresponduese. Pra, limfocitet T autoreaktivë me specificitet për “epitopet kriptike” do të jenë të pranishëm në repertorin normal të limfociteve T. Në mënyrë të ngjashme njihet dhe prania e një popullatë limfocitesh B CD5+, të cilët prodhojnë “antitrupa natyralë“, term ky që përfshin ato antitrupa serikë që mendohet se janë të pranishëm para çdo kontakti me antigenë të jashtëm dhe pra ato prodhohen në mënyrë të pavarur nga çdo stimulim antigenik. Këto antitrupa janë të klasës IgM dhe përfshijnë autoantitrupa me afinitet të ulët për specificitete multiple dhe të cilët zotërojnë reaktivitet të kryqëzuar me antigenë bakterialë të zakonshëm, kyesisht të natyrës polisakaridike. Kjo mund të konsiderohet si një strategji e cila bën të mundur që stimulimi paraprak i këtyre lloj limfocitesh B nga autoantigenët (duke përfshirë këtu dhe idiotipat) do të sigurojë një mbrojtje antibakteriale duke patur parasysh dhe faktin që natyra polimerike e antigenëve polisakaridikë bën të mundur që këta antitrupa të klasë IgM, edhe pse me afinitet të ulët, do të lidhen me aviditët të lartë me bakteriet. Këta autoantitrupa të klasës IgM që prodhohen nga limfocite B CD5+, janë të pranishëm në titër të ulët në popullatën e përgjithshme dhe shpeshtësia e hasjes së tyre rritet me moshë deri rreth 6-70 vjeç. Këta autoantitrupa janë të padëmshëm, në kuptimin që ata nuk shkaktojnë reaksione mbindjeshmërie ndaj organizmit, por nuk mund të përjashtohet mundësia që këta limfocite B në rrethana të caktuara mund të stimulohen në mënyrë të tillë që të kalojnë në prodhimin e IgG tashmë të dëmshme për organizmin. E njëjta gjë mund të thuhet dhe për limfocitet T autoreaktivë.
Roli i antigenit në induktimin e autoimunitetit: Dihet se limfocitet mund të stimulohen nga antigenët, nga aktivizues poliklonalë si dhe nga
antiidiotipet. Në rastin e autoantigenëve pyetja shtrohet në se këta veprojnë si autoimunogjene dmth. janë ata që shpërthejnë reaksionin imunitar, apo ata shërbejnë thjesht si autoantigenë dmth vetëm bashkëveprojnë me produktet e reaksionit imunitar, ndërsa ky i fundit induktohet nga faktorë të tjerë. Të dhënat nga sëmundjet organ-specifike:
“Pula obeze” (një lloj i veçantë pule që përdoret për qëllime eksperimentale) zhvillon në mënyrë spontane autoantitrupa precipitues të klasës IgG ndaj tiroglobulinës si dhe një përgjigje inflamatore antitiroide që shkatërron gjëndrën duke shkaktuar hipotiroidizëm. Nqs. me anë të tiroidektomisë neonatale hiqet burimi antigenik s’kemi formim të autoantitrupave. Nqs. në këta shpendë injëktojmë tiroglobulinë normale atëhere do të kemi formim antitrupash. Gjithashtu, tiroidektomia e shpendëve me tiroidit tashmë të shfaqur do të pasohet nga një rënjë e shprehur e nivelit të antitrupave. Pra, konkluzoni është që imuniteti spontan anti-tiroglobulinë iniciohet dhe mbahet nga autoantigenë të gjëndrës tiroide. Meqenëse përgjigja imunitare është plotësisht T-vartëse,

96
mund të përfundojmë që si limfocitet T ashtu dhe ata B janë të stimuluar nga tiroglobulina në këtë model eksperimental.
Te njeriu, përgjigja imunitare është vështirë që të analizohet si të kafshët por bazohemi kryesisht në të dhëna më indirekte. Psh., nga gjëndra tiroide e të sëmurëve me tireotoksikozë janë zhvilluar linja qelizore me limfocite T, të cilat mund të stimulohen direkt nga qelizat e tiroides in vitro. Si një provë e rëndësishme e seleksionit të limfociteve B nga ana e antigenit në një përgjigjen T-vartëse shërbën formimi i autoantitrupave të klasës IgG me afinitet të lartë i shoqëruar ky formim dhe me mutacione somatike.
Të dhëna nga autoimuniteti sistemik (jo-specifik për antigenin): Të dhënat eksperimentale janë këtu më vështirë të gjinden sepse nuk mund të realizojmë
“heqjen e antigenit” si në sëmundjet organ-specifike. Përsa i përket limfociteve B edhe në këto sëmundje kemi formim autoantitrupash IgG me
afinitet të lartë dhe me specificitet për autoantigenë ubikuitarë si ata bërthamorë psh. Por dhe limfocitet T janë të domosdoshëm në këto lloj përgjigjesh gjë që është vënë në dukje
në mijtë ekspërimentalë në të cilët varfërimi në limfocitet T CD4+ sjell dhe mosformimin e autoantitrupave. Problemi është se këtyre limfociteve T nuk u njihet specificiteti antigenik. Një hipoteze është se ata janë të drejtuar ndaj antigenëve të mikroorganizmave që janë vështërë të identifikohën siç u përmënd më lart. Por këtu ngel për t’u shpjeguar formimi i autoantitrupave dhe roli patogjenik i tyre së disa sëmundje sistemike si LES psh. Një pikëpamje tjetër është ajo që limfocitet T nuk duhet të kenë aspak specificitet të veçantë pëer antigenët konvencionalë, si psh. anti-ADN në LES, por ata janë anti-idiotipikë. LES psh. do të ishte një sëmundje tipike “e idiotipeve” e cila shfaqet nga çrregullimi i rrjetës idiotipike.
A është autoantigeni në kontakt me limfocitet? Një pikëpamje e hershme, të paktën për autoantitrupat organ-specifikë, ka qënë ajo që
antigenët janë të sekuestruar brënda një organi të caktuar dhe, si rrjedhojë e mungesës së kontaktit me sistemin limforetikular, nuk arrihet të krijohet ndaj tyre toleranca imunollogjike e zakonshme. Një çrregullim i këtij izolimi që shkakton çlirimin e antigenit jashtë organit do të krijojë mundësitë e krijimit të autoantitrupave. Kjo është e vërtetë për disa organe apo inde të caktuar si psh. lens cristallini, zemrën apo spërmën, lëshimi jashtë organit i autoantigenëve të të cilëve do të provokojë formimin e autoantitrupave. Por, në përgjithësi, punimet ekspërimentale kanë vënë në dukje që injëktimi i ekstrakteve të pamodifikuar të atyre indeve të prekur në sëmundjet autoimune organ-specifike nuk shkakton formim autoantitrupash. Në fakt, studime të hollësishme me autoantigenin e tiroides, tiroglobulinën kanë vënë në dukje që kjo e fundit nuk është plotësisht e sekuestruar brënda gjëndrës, por kalon në lëngun ekstraqelizor nëpërmjet folikujve dhe del nga gjëndra me enët limfatike eferente duke arritur të individi normal një përqëndrim prej rreth 0.01-0.05 mikrogram/ml. Në fakt në shumicëen e sëmundjeve autoimune si në rastin e eritrociteve në aneminë hemolitike autoimune, të ADN në LES apo të receptorëve sipërfaqësorë në shumë raste me sëmundje autoimune organ-specifike, autoantigenët janë aksesibël ndaj limfociteve qarkullues.
Antigenët të pranishëm në përqëndrime adekuate në fluidet ekstraqelizore do të mund të proçesohen normalisht nga qelizat antigen-paraqitëse profesionale, por për autoantigenët e lidhur me qelizat, peptidet përkatëse do të mund të bashkëveprojnë në mënyrë efikase me limfocitet T specifikë vetëm në kushtet e favorshme për këtë si: përqendrim i mjaftueshëm i peptidit të proçesuar dhe, në rastin e limfociteve T në qetësi, nëqoftëse kemi dhe sinjalet ko-stimulatorë të pranishëm.
KONTROLLI NGA ANA E LIMFOCITEVE T ËSHTË KRYESOR: Koncepti i sotëm është që limfocite T autoreaktivë potencialisht aksesibël ndaj
autoantigenëve janë të pranishë në organizëm, por mekanizmat rregulluese janë të tillë që në rrethana normale ata pengojnë çfaqjen e autoimunitetit. Nëpërmjet mekanizmave të delecionit klonal, anergjisë klonale, limfociteve T supresorë, apo paraqitjes së pamjaftueshme të autoantigenëve limfocitet T autoreaktive ngelin në gjëndje qetësie, të paaktivizuar.
Meqenëse vartësia e proceseve autoimunitare nga limfocitet T është thelbësore, problemi është të njihen pra ato procese imunopatollogjike që prishin mekanizmat rregullues, të cilët pengojnë çfaqjen e autoimunitetit.

97
AUTOIMUNTETI MUND TË ÇFAQET NGA “BYPASS”-I I LIMFOCITEVE T HELPER:
Pajisja e autoantigenit me një përcaktues antigenik të ri të tipit bartës mund të bëjë që të prishet toleranca ndaj tij dhe të kemi aktivizim të limfociteve T autoreaktive me stimulim të limfociteve B dhe prodhim të autoantitrupave (modeli i Allison-Weigle). 1. Modifikimi i autoantigenit:
Një përcaktues i ri antigenik i tipit “carrier” mund të çfaqet si pasojë e modifikimit të molekulës antigenike, psh. nga difekte në sintezë ose nga anomali në proçesimin lizozomal me pasojë produkte ndarje që ekspozojnë përcaktues të rinj antigenikë. Kështu psh. është vënë në dukje që në të sëmurët me artrit reumatoid kemi një galaktozilim të ulët të karbohidrateve të Fcgama.
Inkorporimi i mjaft proteinave autologe me adjuvantin e plotë të Freund dhe imunizimi në këtë gjëndje i kafshëve ekspërimentale krijon mundësinë e gjenerimit të reaksionit autoimun ndaj këtyre proteinave. Kjo mund të spjegohet me ndryshimin e konfiguracionit të proteinave në fazën ujë - vaj të emulsionit dhe me krijim kështu të përcaktuesve të rinj antigenikë “bartës” në këto proteina.
Modifikimi mund të arrihet gjithashtu me anë të kombinimit me një medikament. Anemia hemolitike autoimune si pasojë e marrjes së alfa-methyl-dopa (aldomet) mund t’i atribuohet modifikimit të sipërfaqes eritrocitare në mënyrë të tillë që kjo shërbën tashmë si përcaktues i ri antigenik me anë të cilit stimulohen limfocite B që njohin antigenin Rhesus. Ky antigen konsiderohet si një antigen i “dobët” me aftësi më të pakët për të induktuar një tolerancë të limfociteve B se sa antigenët e “fuqishëm” të pranishëm në eritrocit. Izoniacidi gjithashtu mund të provokojë një artrit të shoqëruar me çfaqjen e antitrupave antibërthamorë dhe, ndryshe nga rastet e tjera me autoimunitet të induktuar nga medikamentet, sinteza e këtyre autoantitrupave vazhdon edhe pas ndërprerjes së medikamentit. Gjithashtu, në një numër të konsiderueshëm të sëmurësh në trajtim të vazhdueshëm me prokainamid çfaqen antitrupa antibërthamorë dhe në 40% prej tyre janë të pranishëm shënja klinike të LES. Miastenia grave dhe shënja të pemfigus janë vënë re në disa të sëmurë nën penicilinaminë. Sidoqoftë nuk është e qartë në të gjitha rastet nëse medikamenti jep efektin e tij “bartës“(carrier) nëpërmjet modifikimit direkt të autoantigenit apo të ndonjë molekulë tjetër që ka reaksion të kryqëzuar me autoantigenin e interesuar. 2. Reaksionet e kryqëzuar me epitope ndërveprues me limfocitet B:
Njihen mjaft shëmbuj në të cilët përcaktues potencialisht autoantigenikë janë të pranishëm në një antigen ekzogjen dhe si rrjedhojë kur ky antigen ekzogjen depërton në organizëm, këta përcaktues mund të ushtrojnë efektin e tyre bartës duke provokuar formim të autoantitrupave. Psh. encefaliti post-vaksinal i rabies (sëm. së tërbimit) mendohet se shkaktohet nga një reaksion autoimun ndaj trurit, që iniciohet nga indi trunor heterolog i pranishëm në vaksinë. Disa mikroorganizma bartin përcaktues antigenikë që kanë reaksion të kryqëzuar me njëriun dhe kjo mund të jetë një rrugë e rëndësishme e induktimit të autoimunitetit. Në ethen reumatizmale, antitrupat e prodhuar ndaj streptokokut reagojnë gjithashtu edhe me indin kardiak. Po kështu dhe antitrupat anti-kolon që vihen re në kolitin ulçeroz japin reaksion të kryqëzuar me Escherichia coli 014. 3. Mimetizimi molekular i epitopeve të limfociteve T:
Nqs. kemi parasysh modelin e Allison-Weigle, pas eliminimit nga organizmi të agjentit që kishte reaksion të kryqëzuar me limfocitet T autoreaktive, mbajtja e autoimuniteti realizohet tashmë me anë të limfociteve B të aktivizuara të cilat kapin autoantigenin qarkullues, e përpunojnë atë dhe ia paraqesin limfociteve T helper.
Ndryshe është mekanizmi me antigenët qelizorë. Autoantigeni i paraqitet limfociteve T autoreaktivë zakonisht në formën e epitopeve kriptikë që nuk kanë mundësinë të japin një sinjal aktivizues. Agjenti i huaj me reaksion të kryqëzuar do të rrisë mjaft sasinë e antigenit që i paraqitet qelizave antigen paraqitëse (QAP) dhe këta të fundit do të rrisin shkallën e stimulimit të limfociteve T autoreaktive. Këta të fundit do të rrisin shprehjen e molekulave të adezionit dhe në këtë menyrë ata do të fitojnë aviditetin e mjaftueshëm për të bashkëvepruar dhe t’u stimuluar më pas nga auto-epitopi kriptik. Një numër i konsiderueshëm sekuencash të peptideve mikrobiale me shkallë të ndryshme homologjie me proteinat humane janë identifikuar deri tani, por kjo nuk është një provë e

98
mjaftueshme. Zhvillimi i autoimunitetit do të varet nga mënyra se si këto proteina do të përpunohen nga qelizat antigen-paraqitëse si dhe nga lloji i peptidit që do të paraqitet dhe në çfarë përqëndrimi. 4. Epitopet “bartës” të limfociteve T.
Një përbërës membranor mund të japë ndihmë për përgjigjen imunitare ndaj një përbërësi tjetër (njohja asociative). Në kontekstin e autoimunitetit mund të krijohet një përcaktues helper i ri nëpërmjet modifikimit nga ana e një medikamenti ose nëpërmjet inserimit të një antigeni viral në membranën e një qelizë të infektuar. Në studime ekspërimentale është vënë në dukje që infeksioni i një qelizë tumorale me virusin e influencës nxit rezistencën (gjeneron imunitet edhe ndaj qelizave tumorale të painfektuara. Një shëmbëll i ngjashëm në patologjinë njërëzore është vënë re në formimin e aglutininave të ftohta (të klasës IgM) pas infektimit me Mycoplasma Pneumoniae. 5. Mekanizmat e bypass-it idiotipik.
Rrjeta idiotipike, pra prania e autoantitrupave antiidiotipe që janë të pranishem në mënyrë natyrale në organizëm mund të bëhet shkas për aktivizimin e autoantitrupave që normalisht janë të suprimuar. Kjo mund të shkaktohet nga futja në organizëm e antigenëve të huaj që kur kanë aftësi antigenike të fuqishme mund të aktivizojnë prodhimin e antitrupave autoreaktivë. 6. Aktivizimi poliklonal. Mikroorganizma të ndryshëm cfaqin shpesh aftësi adjuvante pasi ata posedojnë aktivatorë poliklonalë të limfociteve si psh. endotoksinat bakteriale që veprojnë direkt mbi limfocitet B pa patur nevojë për ndihmën e limfociteve T. Psh. në infeksionin nga virusi Epstein Barr ( Mononukleoza infeksioze) hasen autoantitrupa të ndryshëm që mund t’i atribuohen stimulimit poliklonal të limfociteve B (probabilisht të tipit CD5+) nga ky virus. Por këta autoantitrupa, ndryshe nga ata që hasen në sëmundjet autoimune të njëriu, janë të klasës IgM dhe nuk përsistojnë pas eliminimit të mikroorganizmit nga organizmi.
AUTOIMUNITETI MUND TË ÇFAQET NGA “BYPASS”-I I MEKANIZMAVE RREGULLUES:
Duhet theksuar që mekanizmat e by-pass-it të limfociteve T nuk janë të mjaftueshëm për të mbajtur autoimunitetin në mënyrë të zgjatur, por kjo mund të ndodhë kur mekanizmat e kontrollit të përgjigjes imune janë të dëmtuar si psh:
Pamundësia e limfocitit T të tolerizohet ose të bashkëveprojë me limfocite T supresorë. Mangësi në sistemet T supresorë antigen-specifike, idiotip-specifike apo jo-antigen-specifike. Një mundesi tjetër është derepresioni i gjeneve të klasës II që con në shprehjen e
papërshtatshme qelizore të molekulave të klasës së dytë të KMPI duke thyer kështu “heshtjen” e ndërveprimit midis autoantigenëve qelizorë dhe limfociteve T induktorë autoreaktivë. Kjo mund ta kthejë qelizën në një shënjë për limfocitet T të aktivizuar.
Crregullimet e ekuilibrit të citokinave mund të ndikojnë gjithashtu në cfaqjen e autoimunitetit.

99
KAPITULLI 24. SËMUNDJET AUTOIMUNE SISTEMIKE (JO-ORGAN-
SPECIFIKE)
Veprimi patogjenetik i autoantitrupave dhe i komplekseve imune. Mbindjeshmëria me ndërmjetësine e limfociteve T si faktor tjetër patogjenetik në sëmundjet autoimune. Vlera e përcaktimit të autoantitrupave në diagnozen e sëmundjeve autoimune sistemike.
Sëmundjet autoimune jo organspecifike (SAJO) janë ato sëmundje autoimune në të cilat autoantigenët e implikuar nuk janë specifike për një organ të caktuar. Si rrjedhojë procesi autoimun nuk është i lokalizuar në një organ, por mund të preken disa inde apo organe dhe për këtë arsye këto sëmundje quhen ndryshe dhe sëmundje sistemike. Edhe autoantitrupat që hasen në këto sëmundje nuk janë specifike për një ind apo organ, por janë të drejtuar ndaj autoantigenëve që janë të pranishëm në cdo qelizë të organizmit si psh. ADN apo ribonukleoproteinave në Lupusin Eritematoz Sistemik (LES).
Nga ana tjetër në sëmundjet autoimune organspecifike dëmtimi autoimun intereson vetëm një ind apo organ të caktuar dhe autoantigeni është specifik për këtë organ apo ind. Kjo ndarje s’është aspak e prerë sepse ka sëmundje autoimune organspecifike si psh. cirroza biliare primitive ku autoantitrupat antimitokondri që hasen s’jane specifike për heparin. Po kështu në një sëmundje sistemike sic është Sindromi i Sjogren hasen dhe autoantitrupa specifike ndaj një indi të caktuar sic janë autoantitrupat ndaj qelizave të duktuseve të gjëndrave të pështymes.
Veprimi patogjenik i autoantitrupave humorale: Ky efekt është i qarte në Anemite hemolitike autoimune në të cilat eritrocitet e mbuluara me
antitrupa antieritrocitare kanë një gjysëm-jetë të shkurtuar kryesisht si rrjedhojë e aderences se tyre në receptorët Fc gama të qelizave fagocitare të shpretkes.
Po kështu në purpuren trombocitopenike idiopatike kemi prani të antitrupave antitrombocitare që janë shkak i trombocitopenise që haset në këtë sëmundje.
Edhe limfopenia e hasur në SLE është pasojë e antitrupave antilimfocitare që mund të cfaqen në këtë sëmundje sistemike autoimune.
Ne Granulomatozen e Wegener hasen autoantitrupa të drejtuara ndaj një antigeni intracelular sic është proteinaza III. Mendohet se si pasojë e një agresioni infeksioz kemi induktim të TNF i cili aktivizon qelizat endoteliale për të prodhuar IL-1 dhe IL-8 që nga ana e tyre tërheqin neutrofilet të cilët nxiten për të shprehur LFA-1 si dhe vetë proteinazen III në sipërfaqen e tyre. Kështu kemi gjenerim të antitrupave anti proteinaze III (c-ANCA - antitrupa anti-citoplazmike të neutrofileve). Si pasojë kemi dhe gjenerim të radikaleve të lira të oksigjenit nga neutrofilet me dëmtim të qelizave endoteliale.
Sindromi antifosfolipid primar karakterizohet nga fenomene tromboembolike rekurrente venoze dhe arteriale, aborte rekurrente, trombocitopeni dhe prani të antitrupave anti-kardiolipine. Shkaku është bashkëveprimi i autoantitrupave me një kompleks të kardiolipines me beta-2 - glikoproteinën 1, gje që inhibon aktivizimin e kaskades se koagulimit.
Veprimi patogjenetik i komplekseve imune me autoantigenë Kur ndaj autoantigenëve solubel krijohen autoantitrupa dhe komplekset imune të formuar nuk
janë të eliminueshem, atëherë mund të shkaktohen lezione indore të ngjashme me ato të sëmundjes se serumit. Kjo mund të ndodhe psh. në mangësite e përbërësve të rrugës klasike të komplementit si C1q dhe C4 në të cilat nga mungesa e eliminimit të komplekseve imune mund të kemi cfaqje të Lupusit Eritematoz Sistemik (LES) që është semundja tipike autoimune sistemike. Në LES janë identifikuar një varietet i gjërë autoantigenësh shumica e të cileve janë lokalizuar brënda në bërthamë. Ndër këto ADN ka një rol patogjenetik të përcaktuar qartë. Komplekset e ADN dhe antigenëve të tjerë bërthamorë me imunoglobulinat si dhe komplementin janë detektuar me imunofluoreshence në biopsitë e veshkave të të sëmurëve me SLE dhe dëmtime të veshkave nga kjo sëmundje. Gjatë fazes aktive të LES kemi rënje të nivelit të komplementit serik pasi ky konsumohet

100
nga komplekset imune qarkulluese dhe ata të depozituar në veshka apo në organe të tjera. Depozitim të komplekseve imune kemi jo vetëm në veshka por dhe në organe të tjera si në lëkurë, artikulacione, mushkëri, gjak, tru apo zemër.
Artriti reumatoid Ne artritin reumatoid kemi një rritje reaktive të qelizave sinoviale me pasojë dëmtim të
kartilagos si dhe të kockës. Membrana sinoviale, e cila rrethon dhe mbron hapësirën artikulare pëson një proliferim si pasojë e një hiperreaktiviteti imunologjik që dëshmohet nga një numër i madh qelizash të aktivizuara si limfocite T, kryesisht të tipit CD4(+), qeliza dendritike, makrofagë si dhe plazmocite që prodhojnë imunoglobulina
Hallka kryesore imunologjike e sëmundjes është sinteza e autoantitrupave ndaj pjesës Fc të IgG, të njohur si Faktor reumatoid. Ky faktor haset në shumicën e të sëmurëve me artrit reumatoid. Një nga të dhënat biologjike të AR është glikozilimi anormal i imunoglobulinave G të këtyre të sëmurëve. Ky crregullim biologjik që shpesh paraprin cfaqjet klinike të sëmundjes mendohet se është shkaku i një ndryshimi konformacional në pjesën Fc të IgG me pasojë të mundshme rritjen e autoantigenicitetit të IgG apo të bashkëveprimit të shtuar me receptorët Fcgama në qelizat efektorë apo dhe me komplementin. Ne artritin reumatoid nuk kemi ulje të komplementit serik si në LES por shpesh edhe rritje të tij si pasojë e inflamacionit të shtuar.
Sëmundjet e tjera të reumatizmit Përvec LES dhe AR, edhe shumë sëmundje të tjera reumatizmale futen në grupin e Sëmundjeve autoimune jo-organspecifike apo sistemike. Këtu futen Sindromi i Sjogren, Sklerodermia, Dermatomioziti, Polimioziti apo Vaskulitet e tipeve të ndryshëm. të gjitha këto sëmundje kanë të përbashkët me LES dhe AR shumë cfaqje klinike sistemike, praninë e autoantitrupave jo specifike për organin dhe shpesh ato i shoqerojnë që të dy sëmundjet e mesipërme. Megjithate, shumë prej këtyre semudjeve kanë karakteristika klinike të sëmundjeve organ-specifike si psh. prekja e gjëndrave të peshtymes në sindromin e Sjogren, prekja e lëkurës në Sklerodermi apo prekja e muskujve në polimiozit. Megjithate duhet nënvizuar fakti se edhe në këto sëmundje kemi prani të autoantitrupave jo-specifike për organin.
Roli diagnostik i autoantitrupave jospecifike për organin Karakteristike kryesore e Sëmundjeve autoimune joorganspecifike (sistemike) është prania në serumin e këtyre të sëmurëve e autoantitrupave jo specifike për organin. Kështu antitrupat antibërthamore me titër të lartë janë karakteristike për LES pasi hasen në 98% të rasteve me këtë sëmundje. Po kështu Faktori reumatoid është shënjë kryesore imunologjike e Artritit reumatoid ku haset në 70% të rasteve. Megjithatë faktori reumatoid haset edhe në rreth 20% të te sëmurëve me LES si dhe antitrupat antibërthamore në rreth 25% të te sëmurëve me AR. Te dy këto shënues hasen me shpeshtësi të ndryshme edhe në sëmundjet e tjera të reumatizmit. Megjithate, në popullatën e antitrupave antibërthamore kemi mjaft nënlloje të tyre që kanë specificitet për një sëmundje sistemike të caktuar. Psh. antitrupat anti ADN native kanë një specificitet të lartë për LES, ndërsa antitrupat anti SSA dhe SSB për Sindromin e Sjogren. Po kështu antitrupat anti Scl kanë specificitet për Sklerodermine ndërsa ata anti-Jo-1 për Dermatomiozitin. Antitrupat anti citoplazëm të neutrofileve janë shënjues të rëndësishëm diagnostike për disa vaskulite si psh Sindromin e Wegener, atë Churg-Strauss, apo vaskulitin mikroskopik që prek kryesisht veshkat.

101
KAPITULLI 25. SËMUNDJET AUTOIMUNE ORGAN-SPECIFIKE
Karakteristikat e sëmundjeve autoimune organ-specifike: Gjëndrat endokrine; Lekura;
Sistemi nervor qendror dhe neuromuskular; Sistemi gastro-intestinal; Hepari; Gjaku; Veshka; Syri; Zemra. Diagnoza e Sëmundjeve autoimune organ-specifike.
Sëmundjet autoimune organ-specifike (SAIOS) janë patollogji në të cilat dëmtimi autoimun prek një organ ose ind të caktuar. Po kështu mekanizmi autoimun është specifik për indin apo organin e prekur dhe karakteristika patogjenetike kryesore është prania e autoantitrupave specifike për organin apo indin. Sistemi endokrin: Ky sistem është nga më të prekurit nga SAIOS. Shembell tipik është gjëndra tiroide e cila mund të preket në forma dhe cfaqje të ndryshme. Në disa rrethana mund të formohen psh. autoantitrupa ndaj sipërfaqes se një të qelizë dhe që mund luajnë një rol stimulues. Kjo ndodh në Sëmundjen e Tirotoksikozes (Semundja e Graves ose Basedow) në të cilen janë vënë në dukje autoantitrupa të drejtuar ndaj receptorit për TSH në qelizat e gjëndrës tiroide. Këto autoantitrupa veprojnë në mënyrë të ngjashme me TSH duke nxitur prodhimin e hormoneve tiroidiene, por njëkohësisht veprimi i tyre është i pavarur nga veprimi feed-back i aksit pituitaro-hipofizar dhe si rrjedhojë kemi stimulim të pakufizuar të tiroides me shënja të tirotoksikozes dhe zmadhim të gjëndres si pasojë e veprimit nxites të autoantitrupave. Semundja e Graves është shpesh e shoqëruar me ekzoftalmi si pasojë e veprimit të kryqezuar të autoantitrupave anti receptor të TSH me një proteine membranore prej 64 kDa të pranishme si në gjëndren tiroide ashtu dhe në muskujt e syrit.
Përkundrazi, në Miksedemen primare, sëmundje e tiroides në të cilen kemi frenim të funksionit të saj, hasim autoantitrupa që kanë veprim të kundërt me ato të mesipërm pasi ata bllokojnë në vend që të nxisin receptorët për TSH.
Në Tiroiditin autoimun (Semundja Hashimoto) që prek kryesisht gratë dhe është e lidhur me HLA-DR3 mekanizmat janë të ndryshëm. Në këtë sëmundje kemi infiltrim limfocitar dhe mendohet se kjo është një shprehje e një mekanizmi autoimun me ndërmjetësine e limfociteve T.
Është për t’u shenuar fakti se në të gjithë këto sëmundje autoimune të tiroides haset në serum në përqindje të ndryshme prania e antitrupave anti-tiroglobuline dhe të atyre anti- përoksidaze të tiroides. Këta autoantitrupa janë me tepër pasojë e dëmtimit autoimun të gjëndres se sa shkak i këtij dëmtimi. Diabeti melitus insulinovartes (DMIV): Ashtu si dhe në tiroiditin autoimun, në DMIV kemi infiltrim kronik inflamator si dhe destruksion të qelizave beta të ishujve të Langerhansit në pankreas. Edhe në këtë rast mendohet se janë limfocitet T ata që përbëjne hallken patogjenetike të sëmundjes. Kjo sëmundje është e lidhur fort me alelet HLA DR-3 dhe 4 si dhe me disa alele HLA-DQ. Përcaktimi i antitrupave anti qelizat beta të ishujve të Langerhansit me imunofluoreshence indirekte ka vlere diagnostike në këtë sëmundje. Infertiliteti Jane vënë re raste infertiliteti në disa meshkuj me prani të autoantitrupave antispërmatozoite që bejne aglutinimin e këtyre dhe pengojnë kalimin nga ana e tyre të mukusit vaginal. Stomaku: Anemia pernicioze është një sëmundje autoimune e stomakut në të cilen dëmtimi histopatologjik konsiston në një gastrit atrofik në të cilin kemi një invadim limfocitar të mukozes e shoqëruar kjo me degjenerim të gjëndrave sekretore dhe me dëmtim të prodhimit të acidit gastrik. Aklorhidria akselerohet nga veprimi inhibues i antitrupave të drejtuar ndaj pompes protonike gastrike. Nga ana tjetër dëmtimi i gjëndrave sekretore sjell pakësimin e prodhimit të faktorit intrinsek me pasojë mosthithjen e mjaftueshme të vitamines B12 (ciankobalamines) dhe shkaktimin e një anemie megaloblastike. Një pasojë e këtij dëmtimi autoimun është prania e antitrupave anti qeliza parietale të stomakut që shërben dhe si shënjë diagnostike e këtij dëmtimi autoimun.

102
Sistemi nervor: në Myasthenia gravis autoantitrupat anti-receptorë të acetilkolines (Ach-R) janë përgjegjës të dëmtimit autoimun. Si pasojë, numri i receptorëve funksionale ulet vazhdimisht me pasojë dëmtimin e përcimit neuromotor që vihet re në këtë sëmundje. Ndërsa në Sklerozen multiple (ose Skleroza në pllaka) mekanizmi autoimun mendohet se është me ndërmjetësine e limfociteve T. Ky mekanizem con në demielinizim dhe formimin e pllakave karakteristike të kësaj sëmundje. Lekura: Si Pemphigus vulgaris dhe Pemphigus foliaceus janë sëmundje autoimune të lëkurës ku janë identifikuar Antigenët që janë shënjë e autoantitrupave që hasen në këtë sëmundje.
Roli i autoantitrupave: Autoantitrupat në sëmundjet autoimune, qofte me rol patogjenetik në to, qofte thjesht pasojë e dëmtimit autoimun janë një shënjues diagnostik i rëndësishëm në to. Përcaktimi i tyre në serum përbën sot mjetin diagnostik me të rëndësishëm të tyre dhe ai realizohet me metoda imunologjike të ndryshme duke filuar nga ato me klasiket si imunoprecipitimi dhe aglutinimi të ato me të avancuara si imunoenzimologjia (EIA), imunofluoreshenca (IF) apo metodat radioimunologjike.
KAPITULLI 26. SINDROMET IMUNOPROLIFERATIVE
Sindromet imunoproliferative monoklonale dhe poliklonale. Klasifikimi i tyre sipas llojit të qelizave. Diferencimi i SIP malinjë nga ato bëninjë. C'jane krioglobulinemite, llojet e tyre:
Përcaktimi i SIP: Karakteri monoklonal: Klasifikimi: SIP i qelizave B: Mieloma multiple: Leucemia mieloide kronike me LB; Sindromi i Valdenstrom; Gamapatite monoklonale bëninjë; SIP me LT: Sindromi i Sezary si dhe Mycosis fungoide. Leucemia limfoide kronike me LT. Limfoma me qeliza T. SIP me Limfocite të padiferencuara.
Përcaktimi I SIP Sindromat imunoproliferative janë sëmundje malinje të qelizave limfoide që nisen nga një limfocit i vetëm gjë që i jep SIP një karakter monoklonal. Ne disa raste lloji i qelizave që proliferojnë në mënyrë malinje përcaktohet relativisht lehtë si psh. në rastin e proliferimeve plazmocitare (mielomave) ku kemi prodhim të sasive të rëndësishme të imunoglobulinave monoklonale. Në rastet kur nuk kemi prodhim të imunoglobulinave të tilla diagnoza është me e veshtire, pasi ajo duhet të bëhet me anë të studimit të shënjuesve qelizore.

103
Në disa raste natyra e qelizave që proliferojnë në mënyrë malinje është e pamundur të përcaktohet pasi ato mund të jenë shumë të papjekura dhe të mos paraqesin asnjë nga shënjuesit e limfociteve B ose T. Karakteri monoklonal i sindromeve imunoproliferative Ky karakter mund të përcaktohet më lehtë kur SIP prek qelizat limfoplazmocitare dhe kemi sintetizim nga ana e tyre e një imunoglobuline monoklonale. Kjo mund të vihet re në elektroforezë të proteinave serike në formën e një piku anormal në regjionin e gamaglobulinave. Ky pik është simetrik dhe në formë i ngjan atij të albumines pasi të gjitha molekulat e imunoglobulines monoklonale janë identike dhe me të njëjtën ngarkesë elektrike. Në faze të dytë është e nevojshme të kryhet imunoelektroforeza ose imunofiksimi që përcaktojnë dhe izotipin e vargut të rëndë dhe të lehtë të kësaj imunoglobuline monoklonale. Proliferimet monoklonale të limfociteve B Mieloma multiple (Semundja e Kahler) Është forma me e shpeshtë e SIP e karakterizuar nga një proliferim plazmocitar monoklonal me cfaqje të plazmociteve anormale në palcën kockore dhe në përqindje të rritur (mbi 20-30%). Diagnoza mbështetet në vënjen në dukje të një imunoglobuline monoklonale në serum dhe/ose urine. Katër situata të ndryshme mund të vihen në dukje: 1 – Prani e izoluar e një imunoglobuline monoklonale në serum (10% e rasteve). 2 – Prani e një imunoglobuline monoklonale të kombinuar dhe me sekretim të tepërt të vargjeve të lehtë (60% e rasteve). Këta vargje të lehtë të lire mund të eliminohen në urine duke cfaqur në këtë mënyrë proteinurine e Bënce-Jones. një vecori fizike e kësaj proteinurie është precipitimi në 56 grade dhe ritretja e këtij precipitati në 100 grade. Por proteinuria e Bënce-Jones nuk është konstante dhe mungesa e saj nuk lejon të eliminohet diagnoza e Mielomes multiple. 3 – Prani e vetëm vargjeve të lehtë në serum dhe urine (proteinuri e Bence-Jones) në 30% të rasteve. 4 – në raste të rralla (1% e rasteve) mund të kemi mungesë të cdo imunoglobuline monoklonale apo vargu të lehtë të vecuar në serum apo urine. Kemi të bejmë me mielome jo-sekretuese, diagnoza e së cilës mund të realizohet vetëm duke studiuar shënjuesit e plazmociteve në palcën kockore si psh. me anë të përcaktimit të imunoglobulinave intracitoplazmike.
Forma me e shpeshtë e mielomes multiple është ajo që shoqërohet me imunoglobuline monoklonale të klasës IgG me varg të lehtë kapa. Me të rralla janë imunoglobulinat monoklonale të klasës IgA dhe shumë të rralla ato IgM. Prania e Ig monoklonale në serum ka si pasojë rritjen e eritrosedimentacionit, rritjen e viskozitetit të gjakut si dhe sindromë hemorragjike. Prania e vargjeve të lehta me tepricë sjell dëmtim renal ndërsa infiltrimi plazmocitar në palcën kockore mund të jape lakuna (boshlleqe kockore), fraktura kockore, anemi si dhe mangësi imunitaredytësore nga dëmtimi i palcës.
Leucemia limfoide kronike Është një proliferim malinj i një kloni të vetëm limfocitesh B të pjekur që shoqërohet me një
limfocitozë të konsideruar (> 10 000/mm3). Këto limfocite nuk janë sekretues imunoglobulinash pra si rregull nuk kemi prani të imunoglobulinave monoklonale në serum përvec se në 10% të rasteve ku mund të haset një IgM monoklonale me përqendrim të ulët. Diagnoza vendoset duke përcaktuar natyren B të limfociteve të shtuar të gjakut dhe ajo që ka me tepër rëndësi karakterin monoklonal të këtyre (prani e vetëm një lloj imunoglobuline sipërfaqesore në to). Komplikacionet e leucemise limfoide kronike janë si me poshte: -Instalim i një mangësie imunitare që prek progresivisht IgM pastaj IgG dhe IgA duke prekur se fundi dhe imunitetin me ndërmjetësi qelizore. Kjo mangësi është përgjegjëse për një prekshmeri të rritur ndaj infeksioneve në këta paciente.
-Cfaqje e manifestimeve autoimune shumë të shpeshta të tipit anemi hemolotike autoimune ose trombocitopeni autoimune. Semundja e Waldenstrom

104
Është një proliferim limfoplazmocitar monoklonal por heterogjen i karakterizuar nga prania e limfociteve në stade të ndryshme të diferencimit (limfocite B të vegjel, të medhenj si dhe plazmocite) por me origjinë nga i njëjti limfocit B. Ky proliferim shoqërohet me prodhim dhe të IgM monoklonale që vihet në dukje si me elektroforeze (rritje e beta ose me rralle e gamaglobulinave) ashtu dhe me imunoelektroforeze ose imunofiksim. Klinikisht semundja shoqërohet me rritje të viskozitetit të gjakut që mund të jape komplikacione respiratore ose neurologjike, insuficience medulare si pasojë e infiltrimit të palcës kockore apo dhe manifestime autoimune si anemi apo trombocitopeni.
Semundja e vargjeve të rënda Është një proliferim monoklonal i plazmociteve prodhues kryesisht të IgA (format me IgG
dhe Ig M janë shumë me të rralla) të cilat kanë pesuar një keputje të vargut të rëndë alfa në nivelin e bashkimit të fragmentit variabël V alfa me pjesën konstante C alfa. Si pasojë kemi një pamundesi të kombinimit të vargjeve të rënda me ato të lehta dhe vargjet e rënda të vecuara dalin në qarkullim duke dhene një rritje monoklonale në regjionin alfa ose beta të globulinave. Semundja e vargjeve të rënda alfa haset kryesisht në popullatat e rajonit mesdhetar dhe cfaqet zakonisht me prekje digjestive (limfome me plazmocite alfa) në formë malabsorbimi. Evolucioni I sëmundjes është variabël dhe në disa raste është vënë re një përmirësim pas trajtimit me antibiotike.
Prodhim i Imunoglobulines monoklonale pa proliferim plazmocitar malinj Ne disa paciente është detektuar një prodhim anormal i Ig monoklonale e pashoqëruar kjo
nga një proliferim limfoplazmocitar malinj. Karakteri “bëninj” i këtij afeksioni afirmohet nga kriteret që vijojnë:
- Nivel normal i Ig serike - Rritje me e vogel se 10 g/L në formë piku në elektroforeze e imunoglobulines
monoklonale - Niveli i vargjeve të lehta në serum me i vogel se 1 g/L. - Mungesë e proliferimit limfoplazmocitar në gjak dhe palcë.
Ky lloj proliferimi mund të jetë krejt i izoluar ose dytësor si pasojë e disa sëmundjeve si psh. cirrozat, sëmundjet autoimune, infeksionet bakteriale ose virale.
Me shpesh kjo imunoglobuline monoklonale mund të regresoje në mënyrë spontane ose mund të ngele e stabilizuar. Megjithate nevojitet një mbikqyrje e rregullt me mielograma përiodike për disa vite e këtyre të sëmurëve.
Proliferimet monoklonale të limfociteve T Njihen 4 tipe kryesore të tyre: 1- Sindroni i Sezary: Kemi të bejme me një hematodermi të karakterizuar nga një proliferim
limfocitesh T anormale në lëkurë ndonjëhere deri në nyjet limfatike lokale dhe me rralle edhe në gjak (format leucemike).
2- Mycosis fungoide: Edhe këtu kemi infiltrim monoklonal me limfocite T por i kufizuar vetëm në lëkurë.
3- Leucemia limfoide kronike me limfocite T: është një formë e rralle (2% e rasteve) në Europe, por e shpështë në Japoni. Ajo është elidhur me faktorë gjenetike si dhe me me virusin onkogen HTLV.
4- Limfomat me limfocite T jane direkt të lidhur me virusin onkogen HTLV. 5- Proliferimet limfocitare monoklonale të paklasifikuar:
1- Leukoza limfoblastike akute (LLA): në 80% të rasteve limfocitet proliferues janë qeliza shumë të papjekura të xhveshur nga cdo shënues karakteristik T ose B. Ne 20 % të rasteve (forma të rënda të shoqëruara me hipërplazi timike) qelizat malinjë janë të natyres T.
2 – Limfomat dhe limforsarkomat : Këto proliferime (sem. e Hodgkin) janë me natyre zakonisht të paklasifikueshme.

105
KAPITULLI 27. IMUNOLOGJIA E TUMOREVE
Mekanizmat e bashkëveprimit sistem imunitar-tumor. Faktorët imune që pengojnë
rritjet tumorale si dhe ata që favorizojnë rritjen e tyre. Shënjuesit kryesore tumorale.

106
Përcaktimi
Proliferimi tumoral (ose kanceroz) është i karakterizuar nga proliferimi i cfrenuar i një ose disa kloneve qelizore që i shpëtojnë (devijojnë) mekanizmave normale të kontrollit të diferencimit dhe të proliferimit qelizor. Si pasojë këto qeliza poliferojnë në mënyrë të corganizuar dhe të pandalshem në gjirin e organizmave shumëqelizore. Si pasojë crregullohet si organi i prekur ashtu dhe organe dhe inde të tjerë nga metastazat në distancë të tumorit. Klasifikimi
Sipas kushteve të cfaqjes se tumorit mund të klasifikohën në tre lloje: 1 – Tumoret “spontane” për të cilët asnjë faktor kancerogen i njohur nuk është vënë në dukje. 2 – Tumoret e “induktuar” që cfaqen në kafshe ose qenjë të gjalla të ekspozuara ndaj kontaktit me faktorë të njohur karcinogene. Në realitet është mjaft e mundshme që shumë tumore megjithëse në dukje “spontane” të jenë në fakt të induktuar nga faktorë akoma të panjohur. 3 – Tumoret e “transmetuar” nga një kafshe në tjetrën ose nga një njëri në tjetrin. Ky transmetim mund të realizohet si horizontalisht (sipas modelit infeksioz) ashtu dhe vertikalisht nga prindi të fëmija (faktorë të trasheguar). Kjo mënyrë transmetimi vihet re shpesh tek kafshet ekspërimentale. Por është e mundur që edhe të njeriu disa tumore në dukje spontane mund të jenë në realitet të transmetuar në mënyrë okulte.
Etapat e karcinogjenezes Ne fillim kemi një crregullim një ose disaqelizor shumë i kufizuar. Ky crregullim që ndodh në genomen qelizore është i pamjaftueshem për të shpjeguar zhvillimin e tumorit. Ky ndodh në përgjithësi si rrjedhojë e crregullimit të mekanizmave rregullues të rritjes qelizore si dhe si crregullim i mekanizmave imunologjike që normalisht duhet të eliminojnë cdo qelizë anormale ose të huaj për organizmin. Këto anomali mund të shkaktohen nga veprimi i faktorëve të ndryshëm që ushtrojnë një efekt “promotor” duke favorizuar kështu zhvillimin e klonit ose kloneve neoplazike. Disa faktorë, të quajtur karcinogene të plotë mund të shkaktojnë njëkohësisht efekte induktore dhe promotore. Njëkohësisht, nën influencën e faktorëve të ndryshëm (mutacione somatike, shkaterrim i qelizave neoplazike me të ndjeshme ndaj faktorëve imuniatare etj.) ndodh një seleksion i qelizave me të ndjeshme tumorale të afta t’I shpetojnë reaksionit të mbrojtjes antitumorale madje dhe trajtimeve antikanceroze.
Faktorët e karcinogjenezes A – Faktorët gjenetike : të njëriu : Kanceri i qafes se mitres është me i shpështë të raca e zeze
ndërsa kanceri i gjirit të raca e bardhe. Disa tumore (retinoblastoma, leukozat, Sem. e Hodgkin, mielomat) janë me të shpeshta në disa familje. Ky predispozicion familjar është vënë re në binjaket monozigote. B – Viruset onkogene: Këto viruse, duke u inkorporuar në ADN qelizore krijojnë mundesine që faktorët onkogene të shpërthejne ato mekanizma që mund të cojnë në proliferim tumoral. Roli kancerogen i viruseve të njëriu është akoma i diskutueshem. Vetëm në disa tumore është provuar ky rol: Ky është rasti psh. rasti I virusit EBV në limfomen e Burkitt dhe në kancerin nazofaringeal, i papiloma virusit në disa tumore të lëkurës, i virusit të HBV në tumoret e heparit apo i HTLV në disa leukoza. C – Faktorët kimike: Rreth 1000 substanca kimike janë tashmë identifikuar si kancerogene, por 20 prej tyre kanë një shtrirje ose përdorim të gjërë si psh. hidrokarburet policiklike dhe derivatet e katranit, disa derivate azotike të përdorur si ngjyrues, uretani i përdorur në industrine plastike etj. Rezultatet e studimeve epidemiologjike të lejojnë të mendosh që përgjegjësia e plotë ose e pjesshme e këtyre faktorëve mund të ndërhyje në rreth 70% të tumoreve të njëriu. D – Agjentet fizike: Tumoret e lëkurës shfaqen me frekuence të rritur në subjektet e ekspozuar rregullisht ndaj rrezeve ultraviolet diellore (UVA dhe UVB). Po kështu rrezatimet jonizuese luajnë rol onkogjen që është verejtur në individët e ekspozuar (punonjësit e radiologjise) apo në shpështësine e rritur të tumoreve dhe leukozave në banoret e Hiroshimes dhe Nagasakit.
E – Faktorët endokrine mund të luajnë rolin e tyre si psh në kanceret e gjëndres mamare

107
apo të uterusit të lidhura me hipërestrogenemi. F - Faktorë të ndryshëm: Shumë studime epidemiologjike kanë vënë në dukje që faktorët ushqimore ndikojnë në shkallë të ndryshme në 5 deri 50 % të tumoreve. Këtu mund të përmendim praninë e disa karcinogeneve kimike në disa ushqime ose rolin favorizues të disa mangësive ushqimore kryesisht të vitaminave (C, E, A) dhe mineraleve (Mg). Nga ana tjetër ushqimet hipërlipidike mund të favorizojnë cfaqjen e tumoreve. Duhani: Efekti kancerogen i duhanit për tumoret e rrugëve të frymemarrjes është tashmë i vërtetuar qarte. Ai vepron nëpërmjet faktorëve mutagene e kancerogene kimike, efektit promotor të irritimit kronik të rrugëve të frymemarrjes si dhe nga prania e faktorëve me vecori imunodepresive të ndryshme si reduktimi I kimiotaktizmit të polinukleareve etj. Një nga karakteristikat kryesore e proliferimeve tumorale është cfaqja e disa Antigenëve tumorale specifike që kanë fituar një vlere të vecantë diagnostike. Kështu përmendim Alfafetoproteinën (AFP) që haset në vlera të rritura në serum në tumoret hepatike, antigenin karcinoembrionar (CEA) që haset në tumoret e traktit intestinal apo Antigenin specifik të prostates (PSA) që haset në tumoret e prostates.
Mekanizmat e mbrojtjes antitumorale Qelizat NK: Këto qeliza janë të implikuara në mjaft reaksione flakjeje të tumoreve sidomos në fazat e hershme të procesit neoplazik (qeliza prekanceroze, qeliza tumorale të reja). Duket në fakt se qelizat NK njohin neoAntigenët e qelizave tumorale që shprehin në sasi të paket molekulat e KMPI. Edhe makrofagët të cilët shpesh infiltrojnë masen tumorale mund të ushtrojnë efekt antitumoral nëpërmjet prodhimit të TNF dhe ROI. Përsa I përket imunitetit specifik roli i limfociteve T citotoksikë është pa dyshim kryesor. Po kështu dhe limfocitet T helper luajnë një rol të rëndësishëm nëpërmjet prodhimit të citokinave ku duhen përmendur në rradhe të parë interferoni. Edhe imuniteti humoral nuk mund të lihet pa përmendur në mbrojtjen antitumorale. Fenomeni ADCC (antibody-dependent cellular citotoxicity) që realizohet nga të njëjtat qeliza NK realizohet nga antitrupa antitumorale të klasës IgG. Por dhe qelizat tumorale arrijnë t’u shpetojnë survejimit imunitar nëpërmjet mjaft mekanizmave. Këtu përmendim ekspresionin e dobët të neoAntigenëve në qelizat tumorale, antigenicitetin e dobët të këtyre Antigenëve, ekspresionin shumë të paket të molekulave të KMPI Kl.I në qelizat tumorale, veshjen me me mucine të këtyre qelizave. Nga ana tjetër edhe sistemi imunitar i dobesuar mund të favorizoje cfaqjen e proliferimeve tumorale. Kjo është vënë re si në mangësite e lindura ashtu dhe në ato të fituara në të cilat shpështësia e rritur e tumoreve është një fakt i dokumentuar qarte.
KAPITULLI 28. IMUNOLOGJIA E TRANSPLANTIT
Llojet e ndryshme të transplanteve. Mekanizmat e flakjes imune të tyre. Kushtet e
mosflakjes se transplanteve, roli i Kompleksit Madhor të Pajtueshmerise indore.
Me transplant quajme zevendesimin e një indi apo organi me një ind apo organ tjetër. Transplantet mund t’i ndajmë në ato indore dhe në transplant organesh. Në rastin e parë (psh. në

108
transplantimin e lëkurës apo të palcës kockore) zakonisht nuk kemi anastomoza të enëve të indit të transplantuar me ato të organizmit ku realizohet transplanti. Ndërsa në rastet e transplanteve të organeve kemi gjithmone lidhje të enëve të organit që transplantohet me ato të organizmit ku realizohet transplanti. Me dhurues kemi gjithmone parasysh individin ku është marrë indi apo organi i transplantuar ndërsa marrësi (apo bujtësi) është individi ku realizohet transplanti.
Sipas specieve ku realizohet transplanti ato i ndajmë në autotransplant, izotransplant, alotransplant, dhe heterotransplant.
Ne autotransplant dhuruesi dhe marrësi janë i njëjti individ. në izotransplant transplanti realizohet midis individëve gjenetikisht identike si psh. binjaket monoovulare. Alotransplantet janë transplantet midis individëve të te njëjtes specie por të ndryshëm gjenetikisht. Ksenotransplantet (ose heterotransplantet) janë transplantet midis individëve të specieve të ndryshme.
Transplanti më i shpeshtë është pa dyshim alotransplanti. Autotransplantet, kryesisht të indeve (lekura, palca kockore) kanë gjithashtu një përdorim të shpështë, ndërsa heterotransplantet po ngjallin një interes gjithmone e me të madh. Alotransplanti me i shpeshtë indor është pa dyshim transfuzioni i gjakut në të cilin njihen mirë pasojat e papajtueshmerise. Transplanti i lëkurës ka qënë gjithmonë një model interesant ekspërimental për studimin e fatit të një transplanti. ë rastin e alo- apo heterotransplantit të lëkurës vihet re midis ditës se 3 dhe të 9 një pakesim i qarkullimit të gjakut në indin e transplantuar si dhe një rritje e infiltrimit të këtij indi me limfocite, plazmocite dhe shumë pak plazmocite. Nekroza fillon të cfaqet makroskopikisht dhe brënda një ose pak ditësh indi i transplantuar nekrotizohet plotësisht (flakja e plotë). Natyra e kësaj flakje është krejtesisht imunologjike. Flakja primare dhe ajo sekondare: Transplanti për herë tëdytë i indit të te njëjtit dhurues të i njëjti marrës shkakton një flakje të përshpejtuar. Vaskularizimi fillestar është i paket ose mungon plotësisht. Ndodh një invadim i shpejtë me leukocite polimorfonukleare si dhe qeliza limfoide përfshi plazmocite. Tromboza dhe shketerrimi akut i indit të transplantuar ndodh shpejt, që në ditën e trete deri të katërt në flakjen sekondare. Flakja sekondare është gjithmone specifike për dhuruesin, pra ajo ndodh nqs. transplanti i dytë ndodh nga i njëjti dhurues. Nqs transplanti idytë ndodh nga një dhurues gjenetikisht i ndryshëm nga i pari atëherë flakja nuk është e tipit sekondar, por primar. Roli i sistemit imunitar: Eksperimentet në kafshe kanë vënë qartë në dukje se përgjegjës të flakjes se transplantit janë limfocitet T. Njëkohësisht kemi dhe një përgjigje imunitare humorale me formim antitrupash ndaj Antigenëve jo të njëjte (alo ose – heteroAntigenë) të indit të transplantuar. Ekspërimentet kanë vënë qarte në dukje se Antigenët kryesore përgjegjës për reaksionet e flakjes se organit apo indit të transplantuar janë Antigenët e Kompleksit Madhor të Pajtueshmerise Indore (KMPI). Në individin marrës proliferojnë limfocite T helper CD4 (+) dhe limfocite T citotoksikë CD (8+) specifike për molekulat e Klasës I të KMPI të dhuruesit. Rolin kryesor e luajnë ato CD (4+) që kanë aktivitet “helper” për limfocitet CD (8+) citotoksikë dhe prodhojnë citokina efektorë për reaksionet e hipërsensibilitetit të vonuar. Mekanizmat e flakjes se transplantit:
Njihen në përgjithësi 3 mekanizma flakje të transplantit: 1. Flakja hipërakute fillon që disa minuta pas realizimit të transplantit dhe ndodh në individë bujtës
që kanë antitrupa humorale preekzistues si pasojë e papajtueshmerise se grupeve të gjakut ose e sensibilizimit paraprak nga molekulat e klasës I të KMPI psh si pasojë e një transfuzioni gjaku të meparshem.
2. Flakja akute e hershme ndodh deri 10 ditë pas kryerjes se transplantit dhe shkaktohet me ndërmjetësine e limfociteve T.
3. Flakja akute e vonshme ndodh pas ditës se 11 pas transplantit në të sëmurët që marrin azatioprine dhe prednizon dhe mendohet se shkaktohet nga antitrupat dhe komplementi që fiksohen në arteriolat si dhe në glomerulat.

109
4. Flakja kronike e vonshme karakterizohet nga një depozitim subëndotelial I imunoglobulinave dhe komplementit në membranën bazale të glomerulave ndoshta si pasojë e sëmundjes bazë ose dhe si pasojë e depozitimit të komplekseve imune me Antigenë solubel të veshkes se transplantuar.
Parandalimi i flakjes se transplantit Meqënëse Antigenët e HLA (KMPI të njëriut) janë përgjegjësit kryesore të flakjes se
transplanteve, përshtatja sa me e mirë e këtyre Antigenëve midis marrësit dhe dhuruesit është rruga kryesore për evitimin e flskjes se transplantit në mënyrë të ngjashme kjo me nevojen e pajtueshmerise se grupeve ABO në transfuzionet e gjakut.
Përcaktimi i Antigenëve HLA realizohet me metoden e limfocitotoksicitetit ose me metodat e biologjise molekulare.
Përmirësimi i teknikave opëratore dhe përdorimi I medikamenteve imunodepresore të reja si psh. ciklosporina kanë pakesuar mjaft efektet e papajtueshmerise se Antigenëve HLA në flakjen e transplantit. Studimet kanë vënë në dukje se përshtatshmeria në alelet DR ka rëndësine me të madhe në parandalimin e flakjes e pasuar kjo nga lokuset B dhe me pas ato A. Transplantimi I palcës kockore kërkon një shkallë sa me të lartë pajtueshmerie dhe saktesia me e madhe e metodave të biologjise molekulare nëpërmjet tipizimit të ADN është një kontribut I rëndësishëm në këtë drejtim.

110
KAPITULLI 29. IMUNOTERAPIA AKTIVE, LLOJET E SAJ
Imunoterapia aktive ka të bëjë me vaksinimet. Ajo konsiston në stimulimin e mekanizmave
specifike të mbrojtjes humorale dhe celulare. Vaksinimet japin pra një mbrojtje jo immediate por të zgjatur (muaj deri në vite).
Imunoterapia pasive ka të bëjë me seroterapine (ose analoget e saj si psh. imunoglobulinat humane). Ajo jep një mbrojtje immediate por të përkohëshme, e lidhur kjo me jetën e antitrupave pasive të transmetuar.
Imunoterapia aktive: Vaksinimet Kjo ka për synim të parandaloje cfaqjen e sëmundjeve infektive në grupe të caktuar popullatë
apo në individë të vecantë. Vaksinat antibakteriale: Antigenët e përdorur mund të përbëhen nga:
1 - bakterie të ngordhura si vaksina antipertusis (kundër kolles se mirë) dhe antitifoide apo dhe: 2 - bakterie të gjalla të dobesuara (jo patogjene) në kultura speciale. Shembull i rastit të fundit është vaksina e tuberkulozit (BCG) që realizohet me bacilin e tuberkulozit bovin të dobesuar në pasazhe suksesive në kulture in vitro. Imuniteti që jep kjo vaksine është e tipit qelizor me ndërmjetësine e limfociteve T. 3 - një lloj tjetër vaksine bakteriale janë anatoksinat bakteriale që përfaqesojnë ekzotoksina të cilat kanë humbur toksicitetin nëpërmjet trajtimit me formol dhe nxehtesi por jo imunogjenicitetin. të tilla vaksina janë anatoksinat tetanike dhe ato difteritike. 4 – Ekstraktet ose fraksionet e bakterieve: të tillë janë përdorur për vaksinat antikolerike, antipneumokoksike, antimeningokoksike dhe antihemofilus. Sot kjo formë po përdoret gjithmone e me tepër duke izoluar fraksione imunogjene të këtyre vaksinave sa me të purifikuar dhe duke I konjuguar me adjuvante imunogjene si psh. vaksina antipërtusike acelulare apo vaksina antihemofilike. Vaksinat antivirale:
Antigenët e përdorur mund të jenë: 1 – Viruse të përafërt me antigenicitet të kryqezuar dhe jo patogjene. Shembulli është vaksina e parë e përdorur që në 1796 nga Jenner që është vaksina e lise. Virusi i përdorur është ai i lise se lopes që s’është patogjen për njëriun. 2 – Viruse të inaktivizuar: të tillë janë vaksina antipoliomielitike injëktabel (Salk), vaksina antirabike si dhe vaksina antigripale. 3 – Viruse të gjalle të dobesuar: Avantazhi i tyre vjen nga fakti që ato përsistojnë duke u replikuar në organizëm pa prodhuar efekte patogjene. Imunogjeniciteti I këtyre vaksinave është shumë I forte dhe jetëgjatë. të tillë janë virusi i poliomielitit i Sabin (oral), i fruthit, rubeoles, shytave (parotitit). 4 – Fragmentet virale: Vaksina e hepatitit B është shembulli i një antigeni sipërfaqesor të purifikuar dhe inaktivizuar.

111
KAPITULLI 30. IMUNOTERAPIA PASIVE
Principi: Ai konsiston në injektimin një individi të antitrupave të sintetizuar në një individ
paraprakisht të imunizuar. Produktet e para kanë qënë serume humane që vine nga të sëmurë konvaleshente ose kafshe të hiperimunizuara paraprakisht. Në të dy rastet kemi të bejme me seroterapi. Më pas këta serume janë zevendesuar gradualisht nga gamaglobulina serike të përftuara me precipitim dhe që janë ose polivalente (globale) me origjinë serike ose placentare, ose specifike me prejardhje nga subjekte të hipërimunizuar të cileve u bëhet plazmafereze.
Përdorimi: Ne përdorimet me karakter terapeutik objektivi kryesor është shpejtësia e veprimit në një
sëmundje infektive në të cilen nuk mund të pritet të bëhet vaksinimi. Kjo psh. është përdorur në fruth apo në tetanos në fillim të infeksionit në individë të pavaksinuar.
Ne infeksionet e rënda kryesisht në të imunodeprimuar, të djegur etj. është shpesh e nevojshme futja e imunoglobulinave apo plazmes për të ngritur aftësite mbrojtëse të imunitetit humoral. Gjithashtu futja e imunoglobulinave intravenoze është një nga mjetët kryesore terapeutike në mangësite imunitare humorale. Se fundi, një efekt të padiskutueshem kanë treguar imunoglobulinat polivalente intravenoze në trombocitopenite idiopatike dhe në sindromin e Kawasaki si dhe efekti i tyre është duke u provuar e shumë sëmundje autoimune.

112
KAPITULLI 31. BASHKËVEPRIMI ANTIGEN - ANTITRUP
Karakteristikat e bashkëveprimit antigen-antitrup. Specificiteti ; Reversibiliteti. Afiniteti, aviditeti, konstantja e asocimit. Metodat dhe teknikat që studiojnë reaksionet antigen – antitrup. Antitrupa të shumtë prodhohen nga organizmi si rezultat i depërtimit të një substance imunogjene. Vënia në dukje in vitro mbështetet në aftësinë e tyre të bashkimit specifik me antigenin nga i cili kanë lindur. Reaksionet e mundshme janë atëhere të shumta: në praktikë zgjedhja e një reaksioni të dhënë varet veçanërisht nga natyra e antigenit, nga ndjeshmëria e kërkuar, nga shpejtësia e pritur për të arritur një rezultat, nga saktësia e dëshiruar.
BASHKËVEPRIMI ANTIGEN-ANTITRUP Sapo antigeni është në prani të antitrupit specifik korrespondues ai bashkohet me këtë
antitrup. Kemi të bëjmë më një fenomen të shpejtë, të padukshëm dhe të përbashkët për të gjithë reaksionet antigen-antitrup. Ai është konstant në kundërshtim me fenomenet e dukshme siç janë precipitimi ose aglutinimi, shfaqja e të cilëve është funksion i një numri të caktuar kushtesh.
Është fjala për një ekuilibër dinamik të formuluar sipas ligjit të veprimit të masës. k1 [Ag]+[At] ⇔ [Ag-At] k2 ku [Ag] i përgjigjet njërit prej përcaktuesve antigenikë të molekulës; [At] tregon një sit korrespondues të lirë të antitrupit; kurse K=k1/k2 përfaqëson konstanten e ekuilibrit ose të afinitetit. Vlera e lartë e kësaj konstante shpjegon ndjeshmërinë e madhe të matjeve në imunologji, e shoqëruar me zbulim të sasive shumë të vogla të antigenëve ose antitrupave.
Forcat e lidhjes Është fjala për lidhje të dobëta të quajtura elektrostatike. Në të vërtetë lidhjet e forta të tipit kovalent nuk marrin pjesë në reaksion imunologjik, sepse këto përdorin sasi të energjisë që janë shumë më të mëdha se ato që vihen në lëvizje gjatë bashkimit antigen-antitrup. Këto lidhje elektrostatike me një energji të dobët janë tri tipe si vijon: lidhje jonike ose elektrovalente që ushtrohen midis radikaleve kimike bartës të ngarkesave elektrike të kundërta (anione dhe katione, për shembull); lidhje hidrogjenore që vihen re kur një atom hidrogjeni është tashmë i bashkuar me një atom elektro-negativ nga një lidhje kovalente. Hidrogjeni është atëhere i aftë të bashkohët me një atom tjetër elektronegativ me një lidhje të quajtur hidrogjenore. forcat e Van Der Waals që vijnë nga tërheqja elektrostatike e ushtruar nga elektronet e një molekulë mbi bërthamën e molekulave fqinjë. Këto forca lidhjeje varen nga afërsia e molekulave; ato shtohen kur distanca molekulare pakësohet. Megjithëkëtë, ato ngelin shumë të dobëta dhe të pamjaftueshme për të siguruar vetë kohëzionin e komplekseve antigen-antitrup. Por fuqia e tyre rritet shumë, sepse ato ushtrohen midis dy fragmente molekulash të cilat kanë përputhshmëri hapësinore specifike. Sa më e madhe përputhshmëria ndërmjet këtyre dy molekulave, aq më e fuqishme është forca e lidhjes antigen -antitrup, siç është rasti për antitrupat që shfaqen në fund të imunizimit dhe që kanë një aviditët shumë më të madh për antigenin.
KINETIKA E BASHKËVEPRIMIT ANTIGEN-ANTITRUP

113
Metoda të ndryshme fiziko-kimike (reaksionet e ekuilibrit të dializës, reaksionet e neutralizimit, metodat spektrofluorometrike, reaksionet që përdorin antigenë të shënuar) kanë bërë të mundur të vihet në dukje shpejtësia shumë e madhe e bashkimit antigen-antitrup pasi ajo është e rëndit të disa sekondave madje dhe në 0°C. Kjo shpejtësi rritet rregullisht me rritjen e tempëraturës, për të rënë shumë brutalisht në tempëratura të larta, pa dyshim për shkak të denatyrimit të antitrupave nga nxehtësia. Pëveç kësaj, kur antigeni është një hapten, shpejtësia e reaksionit është 250 herë më e madhe sesa kur është fjala për një pjesëz të figuruar (qelizë psh.).
KARAKTERISTIKAT E LIDHJES ANTIGEN-ANTITRUP Bashkëveprimi i antitrupit me antigenin e tij realizohet midis përcaktuesit antigenik dhe një
valence antitrupore (ose një TCR) nëpërmjet komplementaritetit hapesinor. Kjo lidhje nuk është kovalente, por realizohet me forca tërheqëse me energji të ulët të cilat veprojnë vetëm në distanca mjaft të vogla. Këto forca janë sic thame me sipër lidhjet jonike (elektrostatike) midis grupeve karboksilike (COO-) dhe grupeve aminike (NH3+), lidhje hidrogjenore, lidhje hidrofobe midis aminoacideve aromatike si dhe forcat e Van der Wals që realizohen me terheqjen e reve elektronike me ngarkesa të kundërta midis 2 molekulave.
Bashkëveprimi antigen-antitrup është një reaksion ekzotermik. Ai varet dhe nga disa kushte si tempëratura (ajo me e përshtatshme është 37 grade Celcius), pH i mjedisit ku kryhet reaksioni dhe që duhet të jetë afer atij fiziologjik (7,2 deri 7,4) si dhe përqendrimi jonik i mjedisit që duhet të jetë sa me fiziologjik. Bashkëveprimi varet sigurisht dhe nga përqendrimet molare reciproke të antigenit dhe antitrupit.
Lidhja antigen-antitrup ka dy karakteristika kryesore: 1. Specificiteti i cili megjithate nuk është absolut. Shembull janë reaksionet e kryqezuar midis Antigenëve me strukturë të ngjashme. 2. Reversibiliteti i cili realizohet me anë të nxehtesise ose pH acid 2,8. në këto kushte kompleksi disociohet dhe antitrupi ndahet nga antigeni përkates. Kjo realizohet në mjaft kushte ekspërimentale si psh në anemite hemolitike autoimune për të studiuar autoantitrupat e fiksuar mbi eritrocitet e të sëmurit.
Ndaj të njëjtit antigen, antiserume specifike ndaj tij por me origjinë të ndryshme paraqesin reaktivitet të ndryshëm që varet nga "cilesia" e antitrupave të prodhuar. në fillim të imunizimit antitrupat janë pak ose mesatarisht reaktive me antigenin përkates. në injëksionet e metejshme të antigenit reaktiviteti (si dhe sasia) e antitrupave rritet. Kjo rrjedh nga seleksioni nga ana e antigenit e kloneve qelizore që janë gjithmone e me reaktive me antigenin. Ky reaktivitet antitrupor është i përcaktuar nga dy parametra: afinitet dhe aviditet.
Afiniteti është intensiteti i lidhjes se antitrupit me antigenin (një përcaktues antigenik të vetëm) e matur kjo nga konstantja e asocimit intrinseke e reaksionit. k1 Ag + At --------- Ag -At + Nxehtesi k2
(Ag).(At)k1 = (Ag-At)k2 (Ag-At) k1 ---------- = ------ = K (Ag).(At) k2 ku K është konstantja intrinseke e asocimit të reaksionit. Kjo vlerë ka për njësi inversin e një përqëndrimi dhe matet me litra për mol.(l. mol-1). Tabela më poshtë jep vlerat e K që i korespondojnë llojeve të ndryshme të lidhjeve ligand-receptor (antitrupa, hormone, transportues). Vlerat e larta të arritura me një reaksion imunologjik shpjegojnë mundësine e arritjes se një ndjeshmerie të lartë në dozimet e substancave të pranishme në mjediset biologjike në sasi të vogla dhe tregojnë supërioritetin e antitrupit si ligand. Antitrupa afine/ Antigen 1010 - 1013 l. mol -1

114
Antitrupa pak afine/ Antigen 107 - 1010 l. mol -1 Receptor qelizor/ Hormon 109 - 1011 l. mol -1 Transportues serik/ Hormon 108 - 1010 l. mol -1 Enzime/ Substrat 107 - 106 l. mol -1 Albumine/ Medikament 104 - 106 l. mol -1 Konstantja e asocimit e sistemeve të ndryshme me substancat lidhëse ose ligandet
Aviditeti. Quhet i tillë intensiteti i lidhjes se një antitrupi me antigenin e tij, e përcaktuar kjo më shpejtësine e realizimit të një reaksioni antigen-antitrup "dytesor": precipitim, aglutinim, neutralizim viral. Aviditeti është pra një nocion praktik i përcaktuar mbi një serum. Ai varet nga koeficienti i asocimit, nga numri i valencave të antitrupave të ndryshme ndaj një antigeni (diferenca midis IgG dhe IgM), nga numri i epitopeve të këtij të fundit si dhe nga karakteristikat e mjedisit rrethues (tempëratura, pH, forca jonike). Psh. afiniteti i një antitrupi të klasës IgG është me i lartë se ai i një antitrupi të klasës IgM ndaj të njëjtit antigen. Ndërsa përkundrazi aviditëti i IgM është me i lartë se ai i IgG sepse kjo e fundit ka vetëm 2 vende lidhëse me antigenin ndërsa IgM ka 10 të tilla.
3. Ekzistojnë klasifikime të ndryshme të reaksioneve antigen-antitrup.Njëri është i tillë: Reaksionet primare: Reaksioni i observuar nuk kërkon vecse lidhjen antigen-antitrup dhe e
ve këtë direkt në dukje kryesisht nepemjet shenimit të antitrupit ose antigenit sic janë reaksionet radioimunologjike, imunoenzimologjike, imunofluoreshente apo imunoluminishente. Këto janë dhe metodat me të shpejta, të ndjeshme, sasiore dhe të automatizueshme, pra me bashkëkohoret. Nga ana tjetër ato kërkojnë Antigenë dhe antitrupa në gjëndje të purifikuar si dhe konjugim të njërit prej tyre me një shënues. Gjithashtu ato kërkojnë aparatura matese të ndjeshme dhe të sofistikuara. për këtë arësye dhe kosto e këtyre metodave është me e lartë.
Reaksionet dytësore: Fenomeni i observuar (precipitimi, aglutinimi, reaksioni i fiksimit të komplementit) është rezultat i lidhjes antigen - antitrup por dhe i kushteve të tjera si përqendrimi respektiv i antigenit dhe antitrupit, aftësia apo jo e antitrupit për të fiksuar komplementin, stabiliteti i pjesëzave antigenike etj. në këtë grup përfshihen reaksionet e precipitimit, të aglutinimit, të lizës imunologjike, reaksionet e fiksimit të komplementit etj. Këta janë reaksione me të thjeshta në realizim, por shpesh kërkojnë disi kohë (përjashtim bën këtu turbidimetria dhe nefelometria) dhe ndjeshmeria është disi me e ulët. Avantazhi i vetëm është kostua me e ulët e realizimit të tyre dhe mungesa e nevojes për aparatura matese të kushtueshme si dhe për reagente të purifikuar dhe konjuguar. Përjashtim këtu bejne turbidimetria dhe nefelometria që kërkojnë aparatura matese dhe që kanë kosto me të lartë se metodat e tjera të precipitimit, aglutinimit apo fiksimit të komplementit.
Reaksionet terciare janë manifestime të cfaqura in vivo. në këtë grup metodash ndërhyjne jo vetëm faktorët e mesipërm por dhe karakteristikat e vecanta të species apo të organizmit. Këtu përmendim shokun anafilaktik, anafilaksine kutane pasive, reaksionin e Arthus, hemolizën in vivo. Mund të përmendim këtu dhe provat në lëkurë me Antigenë ose alergjene të ndryshëm. Këto reaksione kanë dizavantazhin se kanë mjaft rezultate fallco-pozitiv ose fallco negativ dhe janë deri diku invazive për të sëmurin si dhe janë pak sasiore. Rezultati i tyre varet dhe nga reaktiviteti biologjik i organizmit. Avantazhi i tyre në krahasim me testet in vitro është kostoja e tyre relativisht e ulët.
Një klasifikim tjetër i ndan reaksionet antigen-antitrup në vartësi të venjës në dukje të reaksionit antigen-antitrup:
Kështu kemi reaksionet e 1) Precipitimit, 2) Aglutinimit, 3) Fiksimit të Komplementit, 4) Ato që përdorin Antigenë ose antitrupa të shënuar me : lende fluoreshente, lumineshente, lënde radioaktive, ose enzima.
Ne do të studiojme reaksionet antigen-antitrup në bazë të këtij klasifikimi të fundit.

115
KAPITULLI 32. REAKSIONET E PRECIPITIMIT

116
Ato ndodhin me antitrupa të shfaqur nën stimulimin e një antigeni të tipit të tretshëm. Ato mund të vëzhgohen në një mjedis qoftë të lëngshëm, qoftë xhelatinoz (gjysëm të ngurte) dhe mund të jenë si cilësore ashtu edhe sasiore. Këto janë metoda specifike por pak të ndjeshme, dhe në përgjithësi kanë nevojë për sasira të rëndësishme antitrupash. Gjithashtu këto metoda kanë nevoje për kohë të rëndësishme inkubimi për realizimin e tyre (përjashtim bejne këtu turbidimetria dhe nefelometria). Avantazhi i tyre është se kanë kosto të ulët pasi s’kane nevoje në përgjithësi për aparatura matese të sofistikuara dhe antitrupa apo Antigenë të purifikuar. që të realizohet një reaksion imunoprecipitimi duhet të plotësohen disa kushte të përgjithshme:
1. Si antigeni dhe antitrupi duhet të jenë në faze të tretshme (solubel). 2. Qe të dy duhet të jenë të pakte dyvalente. Nqs. edhe vetëm njëri (antitrupi ose antigeni )
është monovalent nuk mud të kemi reaksion precipitimi imunologjik. 3. Qe të dy duhet të jenë në përqendrime të tilla molare që të korespodojnë me zonën e
ekuivalences (shih me poshte). Nqs njëri ga të dy është në teprice përqendrimi atëherë mund të mos kemi fenomen precipitimi.
REAKSIONET NË MJEDIS TË LËNGSHËM Teknika cilësore Metoda e formimit të unazës së precipitimit e quajtur edhe "ring test (prova e unazes)"
përbën teknikën cilësore më të thjeshtë për të ditur në qoftë se një serum përmban ose jo antitrupa. Në një tub prej disa mm/diametër futet serumi që do të studiohet; pastaj depozitohet me kujdes antigeni në sipërfaqen e tij. Në qoftë se serumi përmban antitrupa, në zonën e ndarjes të dy lëngjeve formohet një unazë e bardhë që përfaqëson precipitimin e kompleksit antigen antitrup. Që reaksioni të ndodhë është e domosdoshme të ekzistojë një raport përqendrimi i favorshëm midis antigenit dhe antitrupit. Kjo teknikë është pak e ndjeshme por ajo ka epërsinë e shpejtësisë dhe të thjeshtësisë. Ajo lejon për shembull të diktohet shfaqja e antitrupave tek kafsha gjatë imunizimit, të identifikohën njollat e gjakut, të zbulohen mashtrimet në përpunimin e mishit, të klasifikohët një streptokok në grupet serologjike të Lancefield, të zbulohet antigenët e Bacilit të antrax-it në një ekstrakt indor (reaksioni i Ascol-it).

117
Fig. 32.1 - Metoda e diskut ose e formimit të unazës së precipitimit në proven e unazes.
Teknikat sasiore Matja e sasisë së antitrupave të precipitatit.
Në një seri tubash që përmbajnë sasi fikse të serumit, shtohen doza në rritje të antigenit duke ruajtur konstant vëllimin total. Me qëllim që precipitimi të jetë i plotë, përgjithësisht lihen antigeni dhe antitrupi në kontakt të paktën gjatë 48 orëve në +4°C. Pastaj studiohet supërnatanti (pjesa e lëngshme sipër precipitatit) i çdo tubi i cili ndahet në dy pjesë: në të parën shtohet antigeni kurse në të dytën shtohet antitrupi. Në supërnatantin e tubave të parë vetëm shtimi i antigenit lejon të fitohet një precipitim; pra kanë mbetur antitrupa në solucion: është zona e tepricës së antitrupit. Në supërnatantët e tubave të fundit vetëm shtimi i antitrupave shkakton një precipitim: është zona e tepricës së antigenit. Përsa u përket supërnatantëve të tubave të ndërmjetëm ato nuk përmbajnë as antigen as antitrup pasi i gjithë antitrupi gjëndet në precipitat: është zona e ekuivalencës. Sasia e antitrupave që përmbahet në precipitat mund të përcaktohet atëhere duke u nisur nga dozimi i sasisë së proteinave ose i sasisë së azotit të precipitatit. Që matja të jetë e vlefshme është gjithashtu e domosdoshme nga njëra anë të përdoret një serum i dekomplementuar në mënyrë paraprake sepse komplementi është i përbërë nga fraksione proteinike që fiksohen në disa komplekse antigen-antitrup- si dhe nga ana tjetër të hiqet sasia e azotit që përmbahet në antigen në qoftë se ky ka natyrë proteike. Megjithëkëtë ekziston një shkak për gabim: Kur supërnatanti përmban komplekse antigen-antitrup të tretshëm, një pjesë e antitrupave i shpëton atëhere dozimit. Sidoqoftë proteinat e precipitatit Fig 32.2 - Kurba e precipitimit sasior Ajo fitohet pas reaksionit midis ovalbuminës dhe serumit ndaj antiovalbuminës (At: antritrup në sasi fikse; Ag : antigen në sasi në rritje dozohen ose më metodë kolorimetrike (teknika Folin, biuret, ose me ninhidrinë), ose me spektrofotometri në ultraviolet, ose me turbidimetri, metodë që konsiston në vlerësimin e intensitetit

118
të turbullirës të shkaktuar nga precipitimi; edhe me nefelometri, që ka të bëjë me matjen e sasisë të difraksionit të dritës nga precipitati. Metoda e Heidelberger dhe Kendall, me dozim të azotit me mikrometodën kimike e Kjeldahl përbën metodën më të ndjeshme dhe më të saktë megjithëse më delikate. Pesha e azotit e fituar atëhere mund të shumëzohet me një faktor 6,25 për të fituar rezultatin në peshë të proteinës. Mund të fitohen kështu kurba që shprehin sasinë e antitrupave të precipitatit në funksion të sasisë të antigenit të shtuar. Pamjet janë të ndryshme sipas llojit të kafshës që prodhon antitrupa: përcaktohet kështu një kurbë e tipit lepur dhe një kurbë e tipit kalë. Kurba e quajtur e tipit lepur është ajo që haset më shpesh. - në një zonë të parë ku antitrupi është në tepricë, fitohet një precipitat i dobët sasia e të cilit shtohet me të shpejtë duke shtuar antigenin; - në një zonë të dytë, zona e ekuivalencës, sasia e precipitatit që fitohet është maksimum; Fig. 32.3 - Kurba e precipitimit e quajtur "tip lepuri" Ajo fitohet pas imunizimit me një antigen polisakaridik; poshtë pas imunizimit me një antigen proteik. - në një zonë të tretë më në fund, ku antigeni është gjithnjë në tepricë, vihet re një pakësim pak a shumë i shpejtë i sasisë të precipitatit. Ky pakësim është shumë më i ngadaltë me serumet imune ndaj Antigenëve polisakaridikë sesa me ata që u korrespondojnë Antigenëve proteinikë. Ky pakësim i të precipituarit përfundon në një inhibim të precipitimit në tepricë të antigenit, dhe me formimin e komplekseve antigen-antitrup të tretshëm. Kurba e quajtur e tipit të kalit vihet re në serumet imune të kalit antiproteine por jo me antitrupat antipolisakaridikë ose me antiserumet humanë të drejtuar kundër Antigenëve që kanë pak përcaktues. Ajo karakterizohet nga një mungesë e precipitatit në pjesën fillestare të zonës të tepricës në antitrupa (prè-zonë) si pasojë e formimit të komplekseve antigen-antitrup të tretshëm.

119
Fig. 32.4. Kurba e precipitimit e quajtur e tipit "kali" Pré-zona dhe pas-zona janë të karakterizuara nga formimi i komplekseve antigen-antitrup të tretshëm
Këto reaksione përdoren veçanërisht: - për të përcaktuar sasinë e antitrupave që janë të pranishëm në një serum; -për të matur një antigen në një solucion. Mbi një kurbë standart të realizuar me të njëjtin serum dhe me një tretësirë antigenike me koncentrim të njohur, gjehet sasia e precipitatit të fituar me tretësirën me përqëndrim të panjohur.
Matja e kohës të shfaqjes të precipitimit Precipitimi midis antigenit dhe antitrupit korrespondues është shumë më i shpejtë kur ekziston një raport optimum i përqëndrimit midis dy reaktivëve: këto përqëndrime përcaktojnë zonën e ekuivalencës për sistemin e përdorur. Ky është parimi i metodës të quajtur në mënyrë jo të drejtë "e flokulimit fillestar". Dy teknika janë të mundshme. Në metodën Ramon, në një seri tubash që përmbajnë sasi konstante antigeni shtohet sasi në rritje të antitrupave. Në metodën e Dean dhe Webb përqëndrimi i antitrupave është konstant, është ai i antigenit që ndryshon. Në të dy metodat ka ekuivalencë në tubin ku precipitimi duket së pari; megjithëkëtë në mënyrë paradoksale, të paktën në dukje, ky "flokulim fillestar" nuk shfaqet medoemos me po ato raporte antigen-antitrup sipas metodës të përdorur. Në të vërtetë, këto dy teknika realizohen zakonisht në një vëllim të ndryshueshëm; Raynaud dhe Relyveld gjithashtu kanë kodifikuar këto dy protokolle duke ngulur këmbë mbi nevojën që të mbahet konstant përqëndrimi total i përzierjes antigen-antitrup. Këto metoda kanë qënë përdorur sidomos për përcaktimin e sasive të toksinës, të anatoksinës ose të antitoksinës. Është bërë e mundur të përcaktohet një njësi e flokulimit të toksinës ose njësia Lft, e cila është sasia e toksinës që lejon të fitohet precipitimi më i shpejtë kur ajo i shtohet një sasie të dhënë serumi standart flokulimi.
INTERPRETIMI TEORIK I REAKSIONIT TË PRECIPITIMIT Marrack i pari më 1934, ka formuluar hipotezën që është rimarrë pastaj nga autorë të tjerë:
është ajo e formimit të rrjetës. Sipas kësaj teorie molekula e antigenit duhet të jetë të paktën bivalente për përcaktuesin antigenik që merret në shqyrtim; po kështu edhe molekula e antitrupit. Kur raportet e antigenit dhe antitrupit janë të përshtatshme (zona e ekuivalencës) mund të prodhohet një rrjetë në gjirin e të cilës molekulat e antigenit dhe të antitrupit alternohen në mënyrë të rregullt (fig.). Precipitati shfaqet kur masa e kompleksit të formuar në këtë mënyrë i tejkalon forcat që e mbajnë në tretje. Pengesa e precipitimit nga teprica në antitrupa ose teprica në antigen shkaktohet nga ngopja e të gjitha valencave të molekulave të reaktivit tjetër.

120
Mungesa e precipitimit, e vënë re në disa sisteme antigen-antitrup mund të ketë dhe shumë shpjegime. Kështu, disa antitrupa antiproteinikë të kalit (për shembull) zotërojnë një numër të madh grupesh hidrofile; kur ata janë në prani sasish të pakta antigeni, ata formojnë komplekse antigen-antitrup që kanë akoma grupime hidrofile të mjaftueshëm për të qëndruar në tretje. Paaftësia për precipitim mund akoma të jetë e lidhur me monovalencën e Ag për epitopin e konsideruar ose me antitrupa monovalente të fituar nga ndarja enzimatike e molekulës të imunoglobulinës.
Fig. 32. 5 Interpretimi teorik i reaksionit të precipitimit: teoria e rrjetit
Metodat sasiore të precipitimit sasior në mjedis të lëngshem të përshkruara me sipër kanë qënë historikisht metodat e para sasiore imunologjike. për arësye të kohëzgjatjes, kompleksitetit dhe ndjeshmerise se ulët sot këto metoda nuk kanë me vlere praktike. Por teorikisht ato qendrojnë në bazë të te gjithë reaksioneve sasiore antigen-antitrup. Aplikimi bashkëkohor i këtyre metodave është turbidimetria dhe nefelometria që përdorin kryesisht pjesën ngjitese (me teprice në antitrupa) të kurbes klasike në formë kembane të precipitimit në faze të lënget. në këto raste matet ndryshimi i refraksionit të rrezes se drites që kalon në tretesiren antigen antitrup si pasojë e formimit të komplekseve imune në këtë tretesire pa pritur të formohen precipitate të dukshme të komplekseve antigen-antitrup. Turbidimetria dhe nefelometria përdoren gjërësisht sot për matje sasiore të proteinave serike si imunoglobulinat, fibrinogjeni, përbërësit e komplementit, proteina C reaktive (PCR) etj.
TEKNIKAT E PRECIPITIMIT IMUN ME DIFUZION NË XHEL Në këto teknika antigeni dhe antitrupi difuzojnë njëri drejt tjetrit nëpërmjet xhelit, dhe precipitimi ndodh kur ekziston një raport optimum përqëndrimi midis dy reaktivëve. Në qoftë se përgatitja përmban shumë sisteme antigen antitrup mund të identifikohën vija precipitimi të veçanta. Vecoria e xhelit është se ai ngadaleson levizjen e molekulave dhe precipitimi duket si vije e bardhe në xhelin transparënt. Si xhel përdoret agari apo agaroza që janë të tejdukshëm dhe kanë aftësine të

121
jenë në gjëndje të lëngshme në tempëratura të larta (afer vlimit) dhe në gjëndje xheli (faze gjysëm e lëngshme) në tempërature dhome.
Sipas asaj që difuzioni ka të bëjë me një reaktiv ose me të dy, dhe sipas asaj që ajo bëhet sipas njërit aks ose njërit plan, dallohen: - teknikat e difuzionit të njëfishtë, - teknikat e difuzionit të dyfishtë, - teknikat njëdimensionale, - teknikat bideminsionale. Së fundi teknika të tjera kombinojnë difuzionin në xhelozë me elektroforezën: këto janë teknikat e imunoprecipitimit me elektroforezë (imunoelektroforeza).
Reaksionet e difuzionit të njëfishtë dhe bidimensional Teknika e Mancinit Reaksioni kryhet në një pllakë qelqi ose plastike të mbuluar me një xhelozë me lartësi
konstante mbi të gjithë sipërfaqen e pllakës dhe në të cilën është përzier antiserumi. antigeni është i depozituar në gropëza të vogla dhe duke u nisur prej tyre ai shpërndahet në xhel. Kjo shpërndarje vazhdon disa ditë; formohen si pasojë disqe precipitimi sipërfaqja e të cilëve rritet në mënyrë progresive deri në një maksimum të arritur në 2-10 ditë, dhe qëndron kështu gjatë disa muajve. Diametri i unazes se precipitimit D është proporcionale me sasinë e antigenit që është i pranishëm në gropëz. Imunodifuzioni quhet i njëfishte se difuzon vetëm antigeni dhe bidimensional pasi difuzioni ndodh në 2 drejtime (rrethor). Kjo metodë është kryesisht sasiore; ajo lejon një matje të saktë të antigenit që përmbahet në një tretje; mjafton që të matet D2 e diskut të përftuar me antigenin që studiohet dhe ajo të vendoset në një kurbë standart të përftuar me tretje antigenike me përqëndrime të njohura (fig.). Me këtë metode maten në mënyrë sasiore proteinat serike apo dhe Antigenë të tjerë. Metoda ka një kohëzgjatje të lartë (24 – 72 ore), ndjeshmeri jo të mirë, por ka kosto të ulët se s’kërkon reagente të purifikuar apo aparatura matese.
Reaksionet e difuzionit të dyfishtë dhe bidimensional Teknika e Ouchterlony
Ajo realizohet duke derdhur xhelozën qoftë në fundin e një pjate të Petrit qoftë në një pllakë të sheshtë prej qelqi. Teknika e përdorur zakonisht është ajo e Ouchterlony në të cilën antigeni dhe antitrupi shpërndahen duke u nisur nga gropëzat e hapura në xhelozë dhe që janë në distancë prej rreth 15 mm.. Këto metoda janë para së gjithash cilësore: ato kanë për qëllim kryesor të krahasojnë midis tyre tretje antigenike. Në një vendosje më të thjeshtë serumi imun është i vendosur në qendër dhe solucionet antigenike janë në përiferi. Fitohen kështu linja precipitimi në vendin ku antigeni dhe antitrupi takohën në proporcion optimal; në qoftë se solucioni antigenik përmban shumë Antigenë, vihen re shumë vija precipitimi. Veçanërisht, sipas pamjes të vijës së precipitimit, është e mundur të nxirret konkluzioni në qoftë se të dy agjentët janë identike (vijat e precipitimit bashkohën dhe shkrihen në një vazhdimësi totale) , në qoftë se ata paraqitin midis tyre një reaksion të kryqëzuar (vijat e precipitimit bashkohën por formojnë një zgjatje) ose në qoftë se janë totalisht të ndryshëm (vijat e precipitimit kryqëzohen). Kur të dy antigenët janë identikë është gjithashtu e mundshme të dihet në qoftë se përqëndrimi i tyre ndryshon nga një tretje antigenike në tjetrin: në të vërtetë vija e precipitimit është aq më tepër e afërt me rezervuarin e antigenit sa me pak të jetë e përqëndruar tretja antigenike .

122
Fig. 32.6 - Teknika e Mancini-t: reaksioni i precipitimit me difuzion të njëfishtë dhe bidimensional
Fig. 32.7 - Parimi i dozimit të një antigeni me anën e teknikës së Mançinit Ndërtohet në fillim një kurbë standart (antigenëve 1,2,3, me përqëndrim të njohur i përgjigjen sipërfaqet S1, S2, S3); sipërfaqja Sx e precipitatit që fitohet me antigenin që duhet dozuar lejon të përcaktohet kështu përqëndrimi i tij.

123
.Fig. 32. 8. Reaksioni i Ouchterlony: reaksion i precipitimit me difuzion të dyfishtë dhe bidimensional (AC: antitrupi A1, A2: tretje antigenike). a) Të dy Antigenët A1 dhe A2 janë identikë b) Të dy Antigenët A1 dhe A2 paraqitin midis tyre një reaksion të kryqëzuar c) Të dy antigenët A1 dhe A2 janë totalisht të ndryshëm d) Të dy Antigenët janë identikë, por A1 është në një përqëndrim më të dobët se A2
Teknika e imunoprecipitimit me elektroforezë a) Imunoelektroforeza e Grabar dhe Williams
Kur është fjala për përzierje antigenike komplekse teknikat e përmendura deri tani nuk lejojnë identifikimin e saktë të të gjithave vijave të përcipitimit të formuar. Imunoelektroforeza bën të mundur të përmirësohet kjo mundësi e identifikimit sepse ajo përmban para difuzionit dhe precipitimit në xhelozë, një ndarje paraprake të proteinave me elektroforezë. Ajo realizohet në një pllakë të sheshtë prej qelqi në të cilën është hedhur xheloze. Faza e lëngëshme e këtij xheli është një bufer, karakteristikat e të cilit modifikojnë lëvizshmërinë elektrike të proteinave. Përzierja e Antigenëve që duhet të studiohen është e vendosur në një gropëz. Pllaka e përgatitur në këtë mënyrë i nënshtrohet në fillim gjatë disa orëve veprimit të korentit elektrik. Me t'u mbaruar elektroforeza hapet sipas aksit të madh të pllakës dhe paralelisht me aksin e migrimit elektroforetik, një kanal i vogël në xhelozë. Në këtë kanal futet antiserumi. antigeni dhe antitrupi difuzojnë atëhere njëri drejt tjetrit dhe një vijë precipitimi prodhohet në pikën e ekuivalencës. Kjo vijë ka formën e një harku sepse teorikisht Ag i vendosur në një pikë difuzon në të gjitha drejtimet ndërsa At difuzon sipas një vije ballore (ne front). Difuzioni kërkon të paktën 48 orë. Pastaj duhet tharë preparati, të zbulohen harqet e precipitimit me anën e ngjyrosjes dhe mundësisht të fotografohet dokumenti i fituar. Është fjala pra për një teknikë të gjatë për t'u realizuar, por epërsia e të cilit është që të lejojë analizën e përzierjeve antigenike komplekse. Informacionet e marra janë kryesisht cilësore. Kështu imunoelektroforeza e serumit lejon të identifikohën proteinat plazmatike. Përdoren zakonisht shumë lloje serume imune, disa u përgjigjen fraksioneve të ndryshme proteinike të një serumi human normal, të tjerët janë të drejtuar vetëm kundër disa prej këtyre fraksioneve (serum anti-imunoglobulina për shembull). Arrihet kështu të numërohen në serumin njërëzor rreth 30 përbërës që janë të klasifikuar sipas lëvizjes së tyre elektroforetike. Në të vërtetë, imunoelektroforeza e një serumi bëhet gjithnjë njëkohësisht me atë të serumit normal me qëllim që të krahasohen harqet e fituar. Mundet kështu të zbulohet në serumin e studiuar, prania e një proteine anormale që shprehet me një hark precipitimi të veçantë (nga lëvizshmëria dhe forma e saj ) që nuk ekziston në një serum normal. Mundet gjithashtu të zbulohet rritja sasiore e një fraksioni që shfaqet nëpërmjet një harku më të qartë, më të gjatë, më të shtrirë, ashtu si mund të objektivizohen pakësimi ose zhdukja e një proteine serike.

124
Imunoelektroforeza mund në të vërtetë, të shërbejë për studimin e përbërësve antigenikë të çdo lloj lëngu biologjik, si është lëngu cerebrospinal, urina, lëngu i pleurës ose asciti. në praktiken klinike IEF ka vlere kryesisht për studimin cilesor dhe gjysëmsasior të proteinave serike. Kjo është metoda e zgjedhjes për të karakterizuar praninë e një imunoglobuline monoklonale megjithëse sot metoda e imunofiksimit është ajo me e ndjeshme për këto qellim.
b) Teknika e Laurell Serumi imun është i përzier me xhelozën. Tërësia derdhet pastaj në një pllakë prej qelqi. Në këtë pllakë hapen një seri gropëzash të vendosura paralelisht me aksin e madh të pllakës .Antigeni vendoset në këta rezervuarë; migrimi i tij përshpejtohet nën efektin e një fushe elektrike. Pas 3 orë të elektroforezës e pH 8.6 pllaka lahet thahet e ngjyroset. Reaksioni antigen-antitrup shprehet me një vijë precipitimi në formë të një rakete (së këtejmi emri anglez i rocket-elektrophoresis që i është dhënë kësaj teknike) ose po të doni të një maje , lartësia e të cilës është proporcionale me përqëndrimin e antigenit. Duke i vendosur vlerat e fituara në një kurbë standart të hartuar me tretësira me përqëndrim të njohur ( si në metodën Mançini ) është e mundur të dozohen Antigenët që përmbahen në një tretesirë.
c) Imunoelektroforeza bidimensionale Mbi një pllakë të sheshtë prej qelqi të mbuluar me xhelozë hapet një gropëz në të cilën vendoset tretësira antigenike që do të studiohet dhe realizohet kështu në fillim një elektroforezë e parë e kësaj tretjeje në xhelozë. Pritet pastaj, paralelisht me drejtimin e migracionit, banda e xhelozës që përmban antigenët e ndarë tashmë nga rryma elektrike. Katërkëndëshi vendoset atëhere në një pllakë tjetër qelqi; nga çdo anë derdhet një përzierje xheloze dhe imunserumi. Tërësia i nënshtrohet një elektroforeze të re, migracioni i të cilës kryhet në mënyrë pingule me të parën, ndërsa reaksioni i precipitimit prodhohet në të njëjtën kohë midis antigenit dhe antitrupit. Fitohen vija precipitimi që i përgjigjen secilit prej sistemeve antigen dhe antitrup; këto vija kanë një formë maje, lartësia e të cilës është proporcionale me përqëndrimin e antigenënve që studiohen. Kjo metodë e quajtur gjithashtu metoda e dytë Laurell, është delikate në realizim, por ajo jep rezultate njëkohësisht sasiore dhe cilësore, që bëjnë të mundur të veçohen deri 50 përbërës në serumin njërëzor.
Fig. 32.9 Parimi i imuno-elektroforezës
d) Kundër imunoelektroforeza ose elektro-osmoforeza ose elektrosinereza. Si dhe për teknikat e tjera, në një pllakë të sheshtë prej qelqi derdhet xheloza; antigeni dhe antitrupi depozitohen në gropëza rrethore prej 2 mm diametër, në një distancë prej 10 mm. Gjatë elektroforezës antigeni i ngarkuar negativisht në pH-in që përdoret migron drejt polit pozitiv; ai

125
ndeshet kështu me antitrupin dhe shfaqet një vijë precipitimi. Është fjala për një metodë cilësore, por precipitati përftohet më shpejt dhe me një ndjeshmëri më të madhe sesa në teknikat pa elektroforezë. Ky reaksion mund të përdoret duke vënë në dukje Antigenët ose antitrupat e sistemit HBs, antitrupat anti- ADN ose edhe për diagnostikimin e sëmundjeve parazitare. Fig. 32.10 - Teknika e Laurell Fig. 32.11 - Imunoelektroforeza bidimensionale Fig. 32.12 - Kundër imunoelektroforeza
e) Imunofiksimi Kjo teknikë është e afërt me imunoelektroforezën. Në fillim, përbërësit e një përzierje
antigenike komplekse siç është serumi për shembull ndahen me elektroforezë. Pastaj pista e

126
elektroforezës inkubohet me një serum imunspecifik, nga ku ndodh precipitimi in situ i Antigenëve korrespondues. Molekulat që nuk njihen nga antitrupat eliminohen me anën e larjes. Komplekset antigen-antitrup zbulohen me anën e ngjyrosjes. Kjo teknikë është më e shpejtë dhe veçanërisht më e ndjeshme se imuno-elektroforeza. Ajo bën të mundur të zbulohen shirita shumë të holla të ngushta që u korrespondojnë imunoglobulinave monoklonale dhe që nuk zbulohen medoemos në imunoelektroforezën klasike.
KAPITULLI 33. REAKSIONET E AGLUTINIMIT

127
Pas një imunizimi me Antigenë të grimcuar antitrupat e përftuar bashkëveprojnë kryesisht me strukturat molekulare të vendosura në sipërfaqen e kësaj grimce (kryesisht qeliza por dhe grimca inerte si psh. lateksi). Reaksioni i antitrupave me këto epitope të sipërfaqshëm shkakton pra aglutinimin e këtyre Antigenëve të figuruar. Reaksionet e aglutinimit të këtij tipi janë quajtur aktivë ose të drejpërdrejtë. Ato janë 1000 herë më të ndjeshme se reaksionet e precipitimit. Është e mundur të zbatohet kjo ndjeshmëri me reaksione që u përgjigjen edhe Antigenëve jo të figuruar. Mjafton që të fiksohen këto antigenë të tretshëm në sipërfaqen e pjesëzave të figuruara që janë imunologjikisht inerte; në qoftë se antigeni i fiksuar në këtë mënyrë vihet në prani të antitrupit korrespondues, reaksioni antigen-antitrup do të shkaktoje aglutinimin pasiv të pjesëzave që shërbejnë vetëm si trrgues. Këto janë reaksionet e aglutinimit pasiv ose jo të drejtpërdrejte ose edhe të kushtëzuar. Më në fund në disa reaksione, bashkimi i antitrupit me antigenin pengon që të ndodhë një aglutinim: këto janë reaksionet e inhibimit të aglutinimit.
REAKSIONET E AGLUTINIMIT AKTIV Aspekti cilesor Sipas sistemit antigen-antitrup që studiohet reaksioni mund të kryhet në tuba ose edhe në
pllakë qelqi (ose edhe plastike apo qeramike). Në reaksionin në tuba, pas përzierjes së antiserumit dhe të Antigenëve të figuruara (eritrocite, bakterie), formohet një aglutinat. Pas një farë kohë të inkubimit tundet lehtësisht tubi. Leximi bëhet ose me sy, ose me një pasqyrë zmadhuese. Në qoftë se sasia e sedimentacionit copëzohet në pjesëza shumë të qarta, reaksioni është pozitiv (fig. ). Për të interpretuar në mënyrë të drejtë rezultatet shpesh janë të domosdoshëm tuba kontrolli pozitiv ose negativ. Reaksioni mbi pllaka qelqi kryhet duke depozituar një pikë serumi dhe një pikë suspensioni antigenik. Leximi bëhet me sy ose më mikroskop. Në rast reaksioni pozitiv ekzistojnë grumbuj pak a shumë voluminozë, kurse baza e preparatit sqarohet; në rast reaksioni negativ, përgatitja mbetet homogjene . Një reaksion i këtij tipi përdoret veçanërisht për të përcaktuar grupet e gjakut si dhe për përcaktimin e antitrupave antibakterore si psh. reaksioni i Vidal në përcaktimin e antitrupave atisalmonele apo reaksioni Wright për antitrupat antiBrucela. Fig. 32.13 -Reaksioni i aglutinimit në tuba a) Reaksion negativ. b)Reaksion pozitiv

128
Fig. 32.14 - Reaksioni i aglutinimit në lame a) Reaksion negativ. b) Reaksion pozitiv.
Aspekti sasior Reaksionet e aglutinimit, shumë të ndjeshëm, bëjnë të mundur që të zbulohen sasi shumë të vogla antitrupi. Por ato lejojnë me vështirësi që të bëhen studime sasiore shumë të sakta. Megjithëkëtë dy teknika propozohen: është fjala për metodën e Heidelberger-it dhe për metodën e hollimeve të njëpasnjëshme.
Metoda e Heidelberger-it përcakton sasinë e azotit që përmbahet në aglutinatet; atëhere duhet ekstraktuar azoti antigenik për të llogaritur azotin e antitrupave dhe në këtë mënyrë, sasinë e antitrupave që ka marrë pjesë në reaksion. Ky reaksion është shumë i vështirë të kryhet, sepse sasitë ë azotit të antitrupave janë shumë më të pakta se ato që përmbahen në Antigenët e figuruara.
Metoda e hollimeve të njëpasnjëshme është ajo që realizohet në praktiken laboratorike dhe është në të vërtetë gjysmë-sasiore. Ajo realizohet me ndihmën e një serie tubash në të cilët futen sasira fikse antigeni dhe hollime të njëpasnjëshme të serumit. Është hollimi që përmbahet në tubin e fundit dhe që jep akoma një aglutinim të pastër ai që përcakton titrin e serumit. Për shembull kur një serum i holluar 1/128 është akoma pozitiv, titri është 128. Megjithëse kjo metodë realizohet thjesht dhe shpejt, ajo nuk është e saktë. Në të vërtetë leximi i tubit të fundit pozitiv është delikat. Për më tepër meqënëse hollimet bëhen në seri, kemi rritje të mundësisë së gabimit. Kështu në serologji, në mënyrë që një variacion sasior i nivelit të antitrupave gjatë dy marrjeve të njëpasnjëshme të gjakut të jetë sinjifikative, duhet që ajo të jetë të paktën me dy hollime.
Masat teknike Reaksionet e aglutinimit kërkojnë njëfarë numri kushtesh për t'u krijuar në mënyrë korrekte. Tempëratura optimale e reaksionit është zakonisht 37°C (antitrupa "të ngrohtë"). Por disa antitrupa nuk reagojnë veçse në tempëraturën e laboratorit (20°C); kurse të tjerë nuk zbulohen veçse në +4°C (antitrupa "të ftohtë"). Veçanërisht aglutinimi mund të pengohet nga përqëndrime shumë të lartë të serumit; ai rishfaqet më vonë me serume më të holluar. Ky fenomen i pre-zonës (i quajtur edhe fenomen i zonës) vihet re me disa serume, antibrucelë për shembull; ai zhduket në prani të përqëndrimeve kripore të larta ose pas shtesës të disa proteinave siçështë albumina.
Zbatime Këto reaksione kanë zbatime të shumta: - për diagnostikimin e disa sëmundjeve bakteriale, duke u bërë reaksioni qoftë me vetë mikrobin patogjen (serodiagnostikimi i Widal për përcaktimin e antitrupave antisalmonela apo

129
reaksioni i Wright për përcaktimin e antitrupave anti-Brucela), qoftë me një bakterie të ndryshme por të afërt antigenikisht (serodiagnoza i Weil dhe Feliks e cila vë në reaksion serumin e të sëmurëve që dyshohen për tifo ekzantemike me një proteus të shtamit OX19); - për të zbuluar antitrupa antileukocitare, për shembull ata që vëzhgohen gjatë disa neutropenive medikamentoze.
- për të kërkuar aglutinina antieritocitare (grupet e gjakut). Antitrupat që vihen në dukje në reaksionet e aglutinimit direkt janë IgG por kryesisht IgM që kanë një aviditët me të lartë dhe një fuqi aglutinuese me të madhe. një dizavantazh i kësaj metode përvec karakterit gjysëmsasior është mungesa e dallimit të antitrupave IgG nga ato IgM. AGLUTINIMI INDIREKT OSE PASIV Ne këtë lloj aglutinimi antigeni nuk është pjesë përbërëse e një qelizë por mbivendoset në mënyrë pasive (me metoda kimike të ndryshme si fiksim me glutaraldehid, acid tanik etj.) në sipërfaqen e një qelizë si psh. eritrocit apo grimce pasive si psh. lateksi. Eritrociti apo grimca e lateksit shërbejnë në këtë rast si mbartës pasive të antigenit dhe aglutinimi i tyre shërbën si tregues pozitiv i pranise se antitrupave ndaj antigenit specifik të fiksuar në eritrocit apo lateks. Testi i Coombs
Antitrupat antieritrocitarë mund të vihen në dukje në sajë të provës së Coombs (sipas emrit të autorit që e realizoi i pari këtë teknike në 1941). për realizimin e kësaj metode përdoret reagenti i COOMBS i cili është një antiserum antiimunoglobuline humane i përgatitur në kafshe. Antitrupat anti-imunoglobuline humane që ndodhen në këtë antiserum bashkëveprojnë me antigenin respektiv të tyre që janë antitrupat antieritrocitare në sipërfaqen e eritrociteve duke realizuar aglutinimin e këtyre të fundit.
Prova e COOMBS është direkte dhe indirekte. në proven direkte në përcaktojme praninë e antitrupave antieritrocitare në sipërfaqen e eritrociteve të individit që ekzaminohet. Kjo prove përdoret kryesisht në anemite hemolitike autoimune pikërisht për përcaktimin e këtyre lloj antitrupave që shkaktojnë shkaterrimin e përshpejtuar të eritrociteve.
Ne proven e COOMBS indirekt përcaktohen jo antitrupat antieritrocitare të fiksuar në eritrocite por të lire, qarkullues. Këto lloj antitrupash vihen re në të politransfuzuarit që kanë antitrupa antieritrocitare qarkullues të drejtuar ndaj grupeve eritrocitare të huaj për individin e politransfuzuar.
Kërkimi i një papajtueshmërie fetus-nënë në sistemin Rhezus përbën një shembull klasik të përdorimit të këtyre provave. Dy teknikat janë të mundshme: sipas rastit kur materiali merret nga i porsalinduri ose kur merret nga gruaja shtatzanë Rhezus negativ (fig). Tek një i porsalindur Rhezus pozitiv, antitrupat anti-Rhezus me origjinë amnore rrezikojnë që të fiksohen mbi rruazat e kuqe, në qoftë se nëna është Rhezus negativ (fig.). Këtyre eritrociteve u shtohet një serum antigamaglobulinë humane (reagenti i COOMBS). Ky serum nuk aglutinon eritrocitet normalë por ai shkakton aglutinimin e rruazave të kuqe që kanë në sipërfaqen e tyre antitrupa anti-Rhezus: Ky është testi i Coombs i drejtëpërdrejtë (direkt). Prova e Coombs direkt bëhet gjithashtu për të përcaktuar praninë e antitrupave antieritrocitare në sipërfaqen e eritrociteve të te sëmurëve me anëmi hemolitike. Tek një grua shtatzanë Rhezus negativ antitrupat anti-Rhezus mund të jenë të pranishem në serum. Në fillim i shtohen pra këtij serumi eritrocite grup 0 Rhezus pozitiv mbi të cilat shkojnë dhe fiksohen antitrupat anti-Rhezus. Pastaj shtohet serumi antigamaglobulinë, si në reaksionin e mëparshëm: Ky është testi i Coombs indirekt.

130
Fig. 32.15 - Testi i Coombs i drejtpërdrejt (direkt) A: Antitrupa anti-Rhezus me origjinë amnore B: Rruaza të kuqe Rhezus pozitiv C: Antitrupa anti-gamaglobulinë

131
.Fig. 32.16 - Testi i Coombs jo i drejtpërdrejt (indirekt) A: Antitrupa anti-Rhezus me origjinë amnore B: Rruaza të kuqe Rhezus pozitiv C: Antitrupa anti-gamaglobulinë
Interpretimi teorik i reaksionit të aglutinimit Si dhe për reaksionet e precipitimit, mund të zbatohet këtu teoria e rrjetit të Marrack. Për më
tepër, studime me mikroskopi elektronike që vëzhgojnë reaksionin e zhvilluar midis disa viruseve dhe antiserumit homolog kanë mundur të vënë në dukje një lloj rrjeti të formuar nga pjesëzat virale të bashkuara midis tyre prej degësh cilindrike që përfaqesojnë molekulat e antitrupave. Megjithëkëtë, për disa sisteme antigen-antitrup mund të ketë një mungesë aglutinimi, që mund të jetë lidhur me lokalizimin e veçantë, të fshehur, kriptoAntigenët; këto epitope të fshehur janë shumë të thellë për t'i lejuar antitrupit që të bënte lidhjen midis dy Antigenëve. Metoda të ndryshme lejojnë megjithëkëtë që të fitohet një aglutinim: disa përdorin veprimin paraprak të enzimeve proteolitike mbi antigenin me qëllim që të çlirohen këto hapësira të fundosura, për shembull nëpërmjet tripsinizimit të eritrociteve; të tjerat kanë të bëjnë me shtimin e antitrupave anti-imunoglobulinë humane që realizojnë atëhere një lidhje midis antitrupave të fiksuara qysh më parë mbi Antigenët: ky është parimi i testeve të Coombs.
REAKSIONET E AGLUTINIMIT PASIV Si mbështetje për antigenët e tretshëm shërbejnë pjesëza të figuruara të tilla si polistereni i latex-it, eritrocite apo dhe kristale të kolesterolit. Këto pjesëza të ndryshme nuk duhet të përdoren pa diferencim për të gjithë Antigenët; për secilin prej tyre është e nevojshme të kërkohën pjesëzat më të përshtatshme.
Reaksionet me lateks Pjesëzat e lateksit kanë epërsi të shumta. Ato janë antigenikisht inerte dhe pak të brishta. Në formë sferike, pjesëzat mund të kenë tri diametra të ndryshme: 1, 17, 0.20 ose 0.81 mikron; është kjo përmasë e fundit që përdoret zakonisht. Për të evituar çdo aglutinim spontan, punohet me një pH fiks prej 8.2. Parimi i reaksionit është i thjeshtë: me anën e një kontakti të thjeshtë, fiksohet një antigen i dhënë mbi pjesëzat e lateksit dhe shtohet pastaj serumi i studiuar. Në qoftëse ai përmban antitrupin korrespondues me antigenin, ndodh aglutinimi i pjesëzave të lateksit (fig. ). Ky reaksion ka zbatime të shumta: në serologjinë reumatizmale për të kërkuar faktorin reumatoid tek të sëmurët që dyshohen për poliatrit reumatoid (faktori reumatoid është një autoantitrup i drejtuar ndaj IgG humane), në bakteriologji ose në parazitologji për të vënë në dukje

132
antitrupat specifikë ndaj shkaktarëve të ndryshëm.. Të gjitha këto reaksione ndaj lateksit janë jo vetëm cilësorë por edhe gjysmësasiore në saje të përdorimit të hollimeve të njëpanjëshme të serumit (hollimi i fundit që jep akoma një aglutinim të pastër përcakton titrin e serumit).
Fig. 32.17 Parimi i testeve me lateks a) Reaksioni pozitiv (aglutinimi). b) Reaksioni negativ (mungesa e aglutinimit)
Reaksione që përdorin eritrocite Këto janë reaksione të hemaglutinimit pasiv. Të përdorura veçanërisht për diagnostikimin serologjik të sifilizit (TPHA) ato kanë si epërsi lehtësinë e tyre të madhe të leximit dhe ndjeshmërinë e tyre të lartë; por nga ana tjetër ato kanë disa dobësi: brishtësia e eritrociteve, (megjithëse përdorimi i eritrociteve të formalinizuara mund të pakësojë këtë vështirësi) dhe veçanërisht ekzistenca e mundshme e pranisë në serumin që studiohet të antitrupave aktivë kundër epitopeve të vetë eritrociteve që nuk përbëjnë me të vërtetë një suport inert. Përballë kësaj vështirësie të fundit, është e domosdoshme ose të përdoren rruaza të kuqe njërëzore të grupit 0, ose të absorbohen në mënyrë paraprake këta antitrupa kur përdoren eritrocite të kafshës (në përgjithësi të dashit). Disa Antigenë fiksohen praktikisht (menjëherë) me lehtësi, pas një inkubacioni të shkurtër mbi eritrocitet. Është rasti i dinitrofenolit, i acidit pikrik, i penicilinës (mundësia për të kërkuar antitrupa antipeniciline me anën e kësaj teknike). Shumica e Antigenëve me natyrë polisakaridike vendosen gjithashtu në mënyrë spontane: është rasti për antigenin polisakaridik i nxjerrë nga bacili i Koch, ose të Antigenëve Vi të Salmonelave ose të Esherichia Coli. Antigenët proteinike nuk fiksohen veçse pas një përgatitje speciale të eritrociteve. Mund të përdoren eritrocite të trajtuara paraprakisht me acid tanik; sidomos, përdoren mjaft substancat që shërbejnë si lidhje midis eritrocitit dhe proteinës: ky është rasti i bënzidinës e bis-diazotuar, i difluorodinitrobënzenit, i karbodiimidit, të disa joneve plurivalente siç është kloruri i kromit. Përsa i përket reaksionit të Waaler-Rose që përdoret për të vënë në dukje edhe ky faktorin reumatoid ai kryhet si vijon: eritrocite të njëriut të grupit zero vihen në kontakt me antitrupa të lepurit anti-eritrocite të njëriut; shtohet pastaj serumi që studiohet. Faktori reumatoid sillet si një anti-gamaglobulinë, prania e tij shprehet me një aglutinim të rruazave të kuqe.

133
Fig. 32.18 - Reaksioni i Waaler-Rose. a) Reaksioni pozitiv (aglutinimi). b) Reaksioni negativ (mungesa e aglutinimit)

134
KAPITULLI 34. REAKSIONET QË PËRDORIN KOMPLEMENTIN
REAKSIONET E FIKSIMIT TË KOMPLEMENTIT
Të quajtura edhe reaksione të devijimit të komplementit ose edhe reaksione të hemolizës, ato janë vënë në jetë që më 1901 nga Bordet dhe Gengou dhe janë adoptuar që më 1906 nga Wasermann për diagnostikimin e sifilizit.
Parimi Në qoftë se një kompleks antigen-antitrup formohet ai fikson komplementin e pranishëm në
mjedisin ku ndodh reaksioni antigen-antitrup. I fiksuar në këtë mënyrë ky komplement nuk është më i pranishëm për të shkaktuar shkatërrimin (lizën) e rruazave të kuqe të sensibilizuara (dmth. të mbuluar me antitrupa antieritrocitare) të shtuara në një kohë të dytë. Përkundrazi, në qoftë se asnjë kompleks imun nuk është formuar, komplementi i papërdorur devijon drejt eritrociteve të sensibilizuara që atëhere lizohen. Megjithëkëtë mund të ndodhë që antitrupat (ngandonjëherë antigenët) të fiksojnë komplementin në mënyrë jo imunologjike. Thuhet atëhere se serumi (ose antigeni) ka një aktivitet antikomplementar. Veçanërisht ky fenomen vihet re me serumet të pasura në antitrupa, me ata që vijnë nga të sëmurë me mielomë IgG ose me serume të vjetër. Ky aktivitet antikomplementar është veçanërisht i lidhur me imunoglobina të tipit IgG që formojnë agregate mbi të cilët fiksohen disa përbërës të komplementit. Në rast serumi antikomplementar interpretimi i reaksionit të fiksimit të komplementit nuk është i mundur.
Masa teknike Meqenëse faktorë të shumtë ndikojnë në fiksimin e komplementit, reaksioni duhet të bëhet
në kushte të përcaktuara. Fiksimi i komplementit varet nga përqendrimet relative të antigenit dhe antitrupit; këta duhen të futen në sasira të përcaktuara sepse ekziston një raport optimal përqëndrimi midis antigenit dhe antitrupit në të cilin fiksimi i komplementit është maksimal. Gjithashtu antigeni duhet të përdoret me hollimin që mund të zbulojë sasi më të vogël të antitrupave në tipin e reaksionit që bëhet fjalë. Gjithashtu, sasitë e eritrociteve janë fikse dhe hemolizina (serumi me antitrupat antieritrocitare) duhet gjithnjë të titrohet paraprakisht. Si pasojë e ligjit të zëvendësimit të Rodet dhe Favre, reaksioni bëhet me tepri hemolizine sepse sipas këtij ligji, për të njëjtën sasi globulash të kuqe që duhen hemolizuar, komplementi dhe antitrupi kompensojnë njëri-tjetrin. Si pasojë, në një reaksion të hemolizës ku hemolizina është me tepri, është një sasi minimum e komplementit që ndërhyn.

135
Sasia e komplementit e futur në reaksion duhet të jetë e njohur në mënyrë shumë të përcaktuar. Meqenëse situata të shumta patologjike mund të bëjnë që të ndryshojë niveli i komplementit serik dhe këto ndryshime mund të futin në rrugë të gabuar serumi i ekzaminuar dekomplementohet pra paraprakisht me anën e ngrohjes në 56° gjatë 30min, dhe komplementi i përdorur për reaksionin është i titruar që më parë. Në reaksionet serologjike futen zakonisht dy njësi CH100 ose 5 njësi CH50. Në përgjithësi përdoret komplementi i kavies që vjen nga një përzierje e shumë serumeve. Por në disa reaksione mund të përdoret komplementi njërëzor që vjen nga përzierja e serumeve të shumë dhuruesve të përzgjedhur paraprakisht. Si burim komplementi përdoret serumi i freskët ose serum i ngrirë dhe i ruajtur në 40°C (afati i konservimit është më shumë se 6 muaj) ose serumi i liofilizuar (konservimi është praktikisht i pakufizuar) Për të zbuluar aktivitetin antikomplementar të një serumi ose të antigenit është e domosdoshme të realizohen njëkohësisht reaksionet e kontrollit negativ ku njëri prej reaktivëve mungon: antigeni në tubin e kontrollit antigen. Është gjithashtu e domosdoshme që të kontrollohet vlera e sistemit hemolitik (kontrolli i sistemit hemolitik) dhe cilësia e mirë e eritrociteve të përdorura (kontrolli - eritrocite)
Zbatimet e këtyre reaksioneve të deviacionit të komplementit janë të shumta. Fillimisht, të përdorura për diagnostikimin e sifilizit, ato lejojnë që të zbulohet praktikisht çdo antitrup, në qoftë se disponohet antigeni korrespondues; përkundrazi, ato mund të venë gjithashtu në dukje një antigen në sajë të antitrupit specifik të njohur. Kështu, ato janë të përdorura për diagnostikimin serologjik të shumë infeksioneve (serodiagnostikimi i mikoplazmave), të adenoviruseve, të mikroviruseve ose të parazitozave. Ato janë 20-100 herë më të ndjeshme se reaksionet e precipitimit; por saktësia e tyre për sa I përket matjes sasiore është e kufizuar.
NDRYSHIMET SASIORE TË KOMPLEMENTIT
TEKNIKAT E MATJES
Dozimi i veprimtarise komplementare totale Në praktikë të gjithë titrimet përdorin fenomenin e hemolizës. Përdoret zakonisht një sasi e caktuar dhe e saktë e rruazave të kuqe të dashit të sensibilizuara nga një tepricë antitrupash dhe në kushte ekspërimentale rigorozisht të fiksuara. Hemoliza vlerësohet në fotometër. Përcaktohet kështu njësia 50% e hemolizës (CH50): është sasia më e vogël e komplementit në gjëndje të shkaktojë 50% të eritrociteve të përdorur. Kjo vlerë është shumë më e saktë se njësia e hemolizës 100% (CH100). Një vlerë normale e nivelit të komplementit serik nuk mund të përcaktohet veçse në funksion të kushteve të matjes: modifikime të kohës ose temperaturës të inkubacionit, ndryshime të vëllimit total të reaksionit ose të përqëndrimit eritrocitar sjellin me vetë ndryshimin e rezultateve. Është pra e domosdoshme të krahasohen titrat e fituara me titrat normalë, të përcaktuar për metodën e përdorur. Më në fund, një ekuacion i quajtur i Von Krogh bën të mundur që të lidhet matematikisht sasia e komplementit dhe e përqindjes të hemolizës. Veprimtaria funksionale e rrugës alterne të komplementit vlerësohet sipas të njëjtit parim si CH50: kjo është VA50. Sistemi zbulues është i përbërë nga eritrocite të lepurit (aktivizues i rrugës alternative).
Dozimi i fraksioneve Me anën e teknikës së hemolizës mund të vlerësohen disa fraksione të komplementit duke i shtuar lëngut biologjik të studiuar reaktivë që përmbajnë të gjithë faktorët me përjashtim të komponëntit që duhet përcaktuar. Në këtë teknikë, meqënëse përbërësit e tjerë futen me teprim, reaksioni është vetëm funksion i sasisë së faktorit të kërkuar. Një dozim sasior i saktë i disa fraksioneve të plotësuesit mund të fitohet me teknikat sasiore të imunodifuzionit radial.

136
Ndryshimet fiziologjike Ato janë mjaft të dobët. Tek fetusi një veprimtari e komplementit ekziston që në moshen 3
muaj, por niveli i tij është i ulët dhe, në lindje, ai është rreth 20 p 100 më i dobët se tek i rrituri; ai shtohet pastaj lehtë në funksion të moshës (më i ngritur pas 35 vjeç), të peshës ose të qëndrimeve në lartësi. Ai shtohet në fund të barrës, por me përjashtim të këtij rasti nuk ka dallim të lidhur me seksin.
Ndryshime patologjike Disa lloje kafshësh kanë gjenetikisht një mangësi në njërin nga komponëntet e plotësuesit.
Kur është fjala C3 kafshët janë shpesh të brishta dhe të vështira për t'i rritur; është rasti për një racë lepujsh (Freiburg 3) dhe për një racë kaviesh të zbuluar në SHBA më 1919 dhe që sot është zhdukur. Kur bëhet fjalë për faktorë që ndërhyjnë shpejt (C4) ose vonë (si C5 ose C6) në rrugën e aktivizimit, kafshët janë me shëndet të mirë të dukshëm dhe dukët se kanë një ekzistencë normale të paktën në kushtet e laboratorit: është rasti i kavieve që nuk kanë C4, të minjve që u mungon C5, të lepujve që kanë një deficit në C6. Në llojin njërëzor shumë familje me një mangësi të trashëguar në C2 janë të njohura: anomalia është transmetuar si një karakter autosomal recesiv. Subjektet homozigote për këtë sëmundje duken se nuk kanë një ndjeshmëri të rritur ndaj infeksionit, ndoshta sepse rruga alterne është e paprekur. Nga ana tjetër, ata janë shpesh të prekur nga sëmundje të indit lidhor (lupus eritematoz sistemik, polimiozit). Një deficit i trashëguar në C3 është gjetur gjithashtu tek disa subjekte që sillen atëhere si subjekte agamaglobulinemike. Deficiti i trashëguar në inhibitor të C1-esterazë, i përmëndur më sipër është në origjinën e edemës angionerurotike të trashëguar, sëmundje kjo e transmetuar me mënyrë autosomale dominante. Një shtim i nivelit të komplementit serik vihet re në shumë gjëndje patologjike ku kemi inflamacion siç janë infeksionet akute, ikteret obstruktivë, sëmundjet malinjë, reumatizmi artikular akut, infarkti i miokardit. Paksimi i komplementit vihet re gjatë cirrozave (mungesë prodhimi) dhe veçanërisht në lupusin
eritematoz sistemik në acarim si dhe gjatë disa glomerulopative, veçanërisht atyre që shoqërohen me dëmtime proliferative të rënda (hipokomplementemitë nga konsumi për shkak të pranise të komplekseve imune).

137
KAPITULLI 35. TEKNIKAT QË PËRDORIN SHËNUES. Ne të gjitha këto teknika studimi i kompleksit antigen-antitrup realizohet me anë të shenimit të antigenit apo antitrupit me një lende fluoreshente, lumineshente, enzime, apo radioizotop. Këto metoda paraqesin supërioritiet ndaj metodave të mesipërme sepse kanë një ndjeshmeri detektimi me të lartë, shpejtësi ekzekutimi dhe automatizim të kenaqshem. Por kostoja e tyre është me e lartë sepse kërkojnë reagente të purifikuar (Antigenë apo antitrupa) dhe të konjuguar si dhe aparatura matese të sofistikuara dhe të automatizuara. Pavaresisht nga lloji i shënjuesit principi i gjithë këtyre metodave është i ngjashem qofte për përcaktimin e antigenit apo dhe të antitrupit. Këto metoda mund t’i klasifikojme sipas disa menyrave:
Një klasifikim i ndan ato: 1 – në faze të ngurte dhe 2 – në faze të lënget. Ne faze të ngurte antigeni apo antitrupi ndodhet në një qelizë, ind apo material tjetër bartës
inert si psh. polistiren etj. në këtë rast komplekset antitrup-antigen formohen dhe studiohen në sipërfaqen e suportit të ngurte. në këto metoda pas cdo inkubimi antigen-antitrup nevojiten shplarje të vazhdueshme për eliminimin e antitrupave apo Antigenëve të pafiksuar. Ne faze të lënget komplekset antigen-antitrup formohen në solucion. për matjen e tyre nevojitet vecimi i tyre nga antitrupat dhe Antigenët në gjëndje të lire. Një klasifikim tjetër i ndan në : 1 – Metoda “sandwich” ; 2 – Me kompeticion Ne rastin e metodes sandwich (këto metoda realizohen në faze të ngurte) teknikat janë të ndryshme në vartësi në se duam të masim/përcaktojme antigenin apo antitrupin si edhe në se kompleksin antigen-antitrup e përcaktojme direkt duke shenuar një nga të dy (metode direkte) apo nëpërmjet një antitrupi tëdytë të shenuar. Përcaktimi/matja e antigenit me metoden “sandwich”:
Ne rast se duam të masim/përcaktojme antigenin me metode indirekte, është antitrupi i njohur ai që fiksohet (ose është i pranishem) në faze të ngurte. Pas inkubimit me lëngun biologjik (serum, lëng cerebrospinal etj.) ku duam të bejme përcaktimin e antigenit, realizojme shplarjen për elimininim e Antigenëve jo të fiksuar. Inkubimi idytë realizohet me një antitrup të njëjte me të parin por kësaj rradhe të shenuar me një nga shënuesit e mesipërm. Bëhen përseri shplarje të përsëritura që largojnë antitrupat jo të fiksuar dhe se fundi bëhet matja e shënjuesit në sipërfaqen e fazes se ngurte.
Ne rastin kur duam të masim/përcaktojme antigenin me metode direkte atëherë ai kerkohët në se është i pranishem në sipërfaqen e fazes se ngurte me anë të antitrupit të shenuar specifik për te. Kjo metode zakonisht përdoret për përcaktimin e pranise apo jo të substancave me interes (psh. depozitimin apo jo të imunoglobulinave apo komplementit) në inde të ndryshëm si veshka, lëkurë etj. Përcaktimi/matja e antitrupit me metoden “sandwich”:
Zakonisht për këtë qellim përdoren metodat indirekte që janë me të ndjeshme dhe me të sakta. është antigeni i njohur që kësaj rradhe është i fiksuar në faze të ngurte. Pasi kjo e fundit vihet në inkubim me lëngun biologjik ku duam të masim antitrupin e panjohur bëhen përseri shplarje të bollshme dhe inkubohet se dyti me një anti-imunoglobuline humane të shenuar që përcakton praninë/apo sasine e antitrupit të kerkuar.
Ne metodat me kompeticion komplekset antigen-antitrup mund të formohen qofte në faze
të ngurte apo të lënget. Ne rast se duam të përcaktojme një antigen të caktuar në faze të ngurte atëherë është
antitrupi specifik që është i fiksuar në faze të ngurte. Kjo faze vihet njëkohësisht në inkubim me një solucion me antigenin e njohur dhe të shenuar në një përqendrim të caktuar si dhe me lëngun biologjik ku duam të masim praninë apo përqendrimin e antigenit që kërkojme. Antigeni i njohur i shënjuar vihet në kompeticion për t’u lidhur me antitrupin në faze të ngurte me antigenin që matet. Sasia e antigenit të njohur dhe të shënjuar që lidhet me antitrupin është në përpjestim të zhdrejte me përqendrimin e antigenit që kerkohët.

138
Ne metodat me kompeticion në faze të lënget komplekset antigen-antitrup formohen në faze të lënget dhe në një faze tëdytë këto komplekse vecohen me metoda fiziko-kimike nga antigeni i shenuar dhe në gjëndje të palidhur. në këto metoda matja e shënuesit mund të bëhet qofte në komplekset antigen-antitrup të vecuar qofte dhe të antigenit të shenuar dhe në gjëndje të palidhur. në këtë rast përqendrimi i antigenit të shenuar dhe në gjëndje të palidhur është në përpjestim të drejte me përqendrimin e antigenit të kerkuar.
IMUNOFLUORESHENCA Coons, përshkroi në 1941 një teknikë që lejon me anë të konjugimit të një ngjyrosësi
fluoreshent në antitrupat të bëjë të dukshëm komplekset imunë në mikroskop me rreze ultraviolet. Në të vërtetë ky fiksim i një molekulë fluoreshente mbi imunoglobulinat nuk prek veçoritë e tyre imunologjike dhe veçanërisht aftësinë e tyre për bashkëveprim specifik me antigenin. Mund të përdoren shumë ngjyrosës fluoreshentë (ose fluorokromë) ; ngjyra që ata lëshojnë duhet të jetë e ndryshme nga fluoreshenca ngjyrë blu që lëshohet në mënyrë natyrore nga indet. Derivatet e fluoreshencës prodhojnë një rrezatim të ngjyrës së gjelbër, por në vend të izocianatit të përdorur fillimisht nga Coons, preferohet izotiocianati. Derivatet e rodaminës (lisaminë dhe rodamina B veçanërisht) japin një fluoreshencë ngjyrë portokalli. Përsa u përket D.A.N.S. (acidi dimetil-amino-naftalen-sulfonik) ai jep një fluoreshencë të gjelbër por është pak i përdorur. Konjugimi me antitrupat duhet të bëhet në prani të një teprice të moderuar fluorokromi. Ai ka nevojë për një kohë kontakti të zgjatur: 18 orë të paktën në +4°C në pH alkalin (9 - 9.4). Teprica e ngjyrosësit të pakonjuguar eliminohet më vonë, për shembull me absorbim në pluhur karboni vegjetal, me dializë të zgjatur, ose me filtrim mbi xhel të sefadeksit. Katër metoda mund të përdoren. Në çdo rast reaktivet e ndryshme lihen në kontakt gjatë 30 min; midis çdo etape larjet e kujdesshme eliminojnë të gjitha substancat e pafiksuara në reaksion imunologjik. Leximi bëhet me ndihmën e një mikroskopi të pajisur për fluoreshencë. Sistemi zotëron një burim rrezesh ultraviolet; kjo dritë kalon nëpërmjet filtrave të quajtur filtra të eksitimit që lënë të depërtojnë vetëm rrezet e gjatësisë së valës të afta të nxisin fluorokromin e përdorur. Rrezatimi që lëshohet nga antitrupi fluoreshent kalon më pas nëpër një filtër i quajtur filtri i ndalesës që ndalon rrezatimet parazitë.
Fig, 32.19 - Parimi i reaksioneve të imunofluoreshencës

139
Metoda direkte
Me këtë metodë antitrupi korrespondues fluoreshent fiksohet drejtpërdrejt mbi antigenin që studiohet. Kjo metode ka nevojë për konjugimin e antiserumit specifik për çdo antigen, gjë që kufizon zbatimet e saj. Megjithëkëtë ajo përdoret shpesh për kërkimin e depozitimeve të imunoglobulinave, të komplementit ose të fibrinës në biopsi indore (lëkure, veshke). Ajo përdoret gjithashtu për tipizimin imunologjik të qelizave (psh. limfociteve) me anë të antitrupave monoklonale të konjuguar me lende fluoreshente.
Metoda e quajtur "sandwich" Ajo shtron të njëjtin problem si metoda e drejtpërdrejtë. Me anën e saj vihen në dukje antitrupa intraindorë. Mbi indin e studiuar vihet në veprim në fillim një tretësirë e antigenit, shtohet pastaj antitrupi specifik i bërë fluoreshent.
Metoda indirekte Ajo përdoret sidomos për kërkimin e antitrupave të ndryshëm në një lëng biologjik si psh serum. Ajo nuk ka mungesat e teknikave të mësipërme si dhe ka një ndjeshmeri me të lartë. Pasi të jetë vënë në bashkëveprim reaktivi që luan rolin e antigenit (prerje indore ose strisho qelizore) dhe serumi që duhet të testohet dhe supozohet se përmban antitrupat korrespondues, vendoset anti-imunoglobulina humane fluoreshente e cila bashkëvepron me antitrupat e fiksuar. Pas cdo faze inkubimi që zgjat secila 30 miniuta beheh shplarje të bollshme për të larguar antitrupat e pa fiksuar. Nëqoftëse rezultati është pozitiv dmth. në serumin e studiuar ishin të pranishem antitrupat e kerkuar do të kemi fluoreshence pozitive indore (ne vend të fushes se erret në mikroskop do të kemi imazh të ndritshem, fluoreshent të strukturave indore të interesuara). Anti-imunoglobulinat vijnë nga një serum i përftuar pas imunizimit në kafshë (zakonisht lepur dele ose dhi) me imunoglobulina me origjinë njërëzore. Pas konjugimit me fluorokrom fitohet një reaktiv që mund të përdoret për reaksione të shumta (për kerkimin e cdo lloj antitrupi human). Reaksioni mund të bëhet gjysmë sasior dhe titri i serumit të testuar shprehet nga emëruesi i hollimit më të madh që jep akoma një fluoreshencë pozitive. Kjo metode ka një përdorim të gjërë për kerkimin e autoantitrupave të ndryshëm si antitrupat antibërthamore (AAB) apo antitrupat anti ADN(AAADN) në lupusin eritematoz, antitrupat anti mitokondri në cirrozen biliare primitive, antitrupat antimuskuj të lemuar në hepatitin autoimun, antitrupat anti qelizat parietale të stomakut në gastritin autoimun etj. për këto qellime si burim autoantigeni përdoren prerje disamikroneshe (4 deri 6 mikron) të organeve përkates që ruhen në gjëndje të ngrire. Psh. për antitrupat antibërthamore përdoren prerjet e melcise se miut apo dhe qelizat Hep-2, për antitrupat antitiroidiene përdoren prerje të tiroides etj. Kjo metode përdoret gjërësisht dhe për zbulimin e antitrupave ndaj shumë agjentëve bakteriale, parazitare apo virale (ne rastin e këtyre të fundit përdoren qekiza humane të infektuara nga virusi pra që shprehin edhe Antigenët virale). Duke përdorur antitrupa fluoreshente specifike për klasa të ndryshme imunoglobulinash (anti-IgG, anti IgM ose anti IgA) kjo metode na lejon edhe përcaktimin se kujt klasë i përkasin antitrupat që na interesojnë. Kjo ka vlere të vecantë diagnostike, psh. prania e antitrupave të klasës IgM flet për një infeksion të fresket, ndërsa vetëm e atyre të klasës IgG për një infeksion me të larget dhe përfaqeson me tepër praninë e një kujtesë imunitare ndaj një infeksioni të caktuar. Ndërsa prania e antitrupave të klasës IgA përfaqeson me tepër një reaksion imunitar (autoimun apo infeksioz) që ndodh në mukozat e ndryshme.
TEKNIKAT RADIOIMUNOLOGJIKE
Ne metoden me dozim me kompeticion në faze të lënget në mënyrë skematike janë të pranishme tre reaktivë:

140
një antigen për shembull hormon i shënuar me një izotop radioaktiv; një antiserum specifik (që përmban pra antitrupa antihormone) i titruar, i cili përgatitet me anën e hipërimunizimit të kafshëve të një specie tjetër; lëngu biologjik në të cilin kërkohët të përcaktohet sasia e antigenit (në shembullin e zgjedhur hormoni jo i shënuar ose "i ftohtë"). Antiserumi mund të kombinohet pa dallim me hormonin radioaktiv ose me atë jo të shënuar që përmbahet në serumin që shqyrtohet. Kur hormoni i shënuar është vetëm i pranishëm përcaktohet në fillim se çfarë sasie kapet nga antiserumi. Më tej matet niveli i kapjes së hormonit radioaktiv nga antiserumi kur ky i fundit vihet në prani të një përzierjeje hormoni të shënuar dhe hormoni të pashënuar. Sa më e madhe të jetë sasia e këtij të fundit aq më pak hormon radioaktiv kapet nga antitrupi. Hartohet atëhere një kurbë standart, në prani të përzierjeve të përbëra nga një sasi konstante të hormonit të shënuar të cilit i shtohen doza të njohura dhe në rritje të hormonit "të ftohtë". Niveli i hormonit të pranishëm në lëngun biologjik përcaktohet duke e kalkuluar rezultatin e fituar mbi kurbën e referencës. Fig. 32.20 - Paraqitje skematike e kurbës së referencës që shërbën për dozimin radioimunologjik të një hormoni.
Variantet teknike Ndërsa parimi bazë mbetet gjithnjë po ai, ekzistojnë disa variante teknike në lidhje me
metodën që u përshkrua për herë të parë për insulinën nga Yalow dhe Berson më 1960. Kjo metodë është realizuar në fazë të lëngshme (dozimi radioimunologjik i mirëfilltë ose radio-imuno-analizë), ndërkohë që shumica e testeve në ditët e sotme bëhën në fazë të ngurtë, duke qënë njëri nga reaktivët i fiksuar mbi një suport (fundi i gropëzave të një pllake mikrotitrimi, grimca lateksi ose disqe të celulozës). Për shembull, radio-imuno-sorbënt-test (RIST) i përdorur për dozimin e IgE (totale) ku janë antitrupat anti IgE që janë fiksuar mbi suportin e ngurtë.
Zbatime Për shkak të ndjeshmërisë së tyre të madhe, meqenëse ato lejojnë të zbulohen sasira të rëndit
të nanogramit për mililitër, këto metoda kanë zbatime të shumta. Ato mund të përdoren për dozimin e

141
çfarëdolloj antigeni: medikamente, vitamina, prostaglandina, leukotriene, IgE totale dhe sidomos hormonet. Megjithëse realizimi i këtyre metodave është delikat ato tani kanë hyrë në praktikën e zakonshme.
IMUNORADIOMETRIA Kjo metodë u realizua më 1968 nga Hales dhe Miles. Ajo përdoret shumë, veçanërisht kur
nuk mund të realizohet shënimi i antigenit. Ajo praktikohët kryesisht në fazë të ngurtë dhe në fakte nuk është gje tjetër vecse një metode “sandwich” tipike për përcaktimin e antigenit. Antitrupi që i përgjigjet antigenit që duhet dozuar vihet në fillim për t'u inkubuar në një gropez polistireni. Pas shplarjes, shtohen antigenët që duhen studiuar të cilët kombinohen me antitrupin. Pas një shplarjeje të re shtohet një tepricë antitrupi specifik i shënuar. Matet pastaj radioaktiviteti: ai është proporcional me përqëndrimin e antigenit. IgE totale, disa shënues tumoralë dhe viralë (HBs, anti HBs, HBe) dhe mjaft Antigenë të tjerë dozohen më këtë teknikë.
TEKNIKA ME SHËNUES FLUORESHENTË
Shumë metoda mund të përdoren. Vetëm dy shembuj janë të përmbledhur këtu: Mundësia e parë përdor ekstinsionin (shuarjen) e fluoreshencës që pason një rregullim të strukturës elektronike molekulare, që vihet re kur një antitrup kombinohet me një antigen të lidhur paraprakisht me lende fluoreshente. Ashtu si në teknikat radioimunologjike, antitrupi mund të lidhet pa dallim me një antigen fluoreshent ose me një antigen jo të shënuar që përmbahet në lëngun që duhet të shqyrtohet. Sa më i lartë të jetë përqëndrimi në antigenin jo fluoreshent aq më pak formohet kompleksi antitrup-antigen fluoreshent dhe aq më shumë pakësohet fluoreshenca. Krijohet kështu një kurbë standart me kontroll pozitiv të njohur dhe nuk mbetet veçse të kalohen mbi të rezultatet e fituara me lëngjet biologjike të panjohur. Mundësia tjetër është quajtur e mbrojtjes të fluoreshencës. Në qoftë se vihen në prani një antigen i shënuar me një substancë fluoreshente dhe një antitrup korrespondues, fitohet një kompleks fluoreshent. Nga ana tjetër kur ky antigen fluoreshent është i përzjerë me një antitrup i drejtuar kundër substancës fluoreshente formon një kompleks jo fluoreshent. Në qoftë se më në fund realizohet një përzierje ku vihen së bashku antigeni jo i shënuar, antigeni fluoreshent dhe antitrupi korrespondues, ndodh një garë midis këtyre dy Antigenëve në mënyrë që sa më i lartë është përqëndrimi i antigenit jo të shënuar, aq më tepër mbetet antigen fluoreshent i lirë, i pakombinuar me antitrupin dhe pra i aftë të bashkohët më vonë me antitrupin e drejtuar kundër substancës fluoreshente që shtohet më vonë.
TEKNIKAT ME SHËNUES ENZIMATIKË
Dozimi i antigenëve Njëra nga më të përdorurat është një metodë imunoenzimatike e drejtpërdrejtë me
kompeticion apo dhe sandwich (Enzyme Linked Immunoabsorbant Assays ose ELISA). Edhe këtu, parimi është po ai si në teknikat radioimunologjike. Antitrupi këtu është i palëvizshëm, i fiksuar në një suport të ngurtë (faqja e epruvetës, grimcat plastike për shembull). Ai do të reagojë me: nga njëra anë një antigen i shënuar me një enzimë (përoxidazë, fosfatazë alkaline, betagalaktosidazë); nga ana tjetër me një antigen jo i shënuar i pranishëm në një lëng biologjik. Sa më e lartë të jetë sasia e këtij reaktivi të fundit aq më pak i rëndësishëm është formimi i kompleksit midis antitrupave dhe Antigenëve të shënuar. Pasi hartohet kurba standart me solucione kontrolli (antigen të njohur), mund të përcaktohet më tej sasia e antigenit (veçanërisht hormone) që përmbahet në lëngun e panjohur. Një teknikë tjetër e zbatueshme ndaj molekulave të vogla (barna, hormone steroid, tyroksina) mbështetet në matjen e ndryshimeve të veprimtarisë enzimatike (glukozë 6-fosfat dehidrogenazë, malikodeshidrogenaze), sipas rastit që antigeni i shënuar me enzimë është i lirë ose i kombinuar me antitrupin e tij. Ky është parimi i reaksioneve të tipit EMIT (Enzyme Multiplied Immunoassay) që realizohet e mjedis të lënget.

142
Metoda të tjera të quajtura "sandwich" nuk përdorin fenomenet e konkurimit. Për shembull, antigeni që duhet të përcaktohet inkubohet në fillim me një sasi të caktuar të antitrupit korrespondues të fiksuar në një suport. Pastaj vihet në veprim një antitrup i shënuar me një enzimë dhe që vjen e fiksohet mbi hapësirat e antigenit të lëna të lira gjatë reaksionit të mëparshëm. Veprimtaria enzimatike e fiksuar në këtë mënyrë mbi kompleksin është drejtpërdrejtë funksion i sasisë së antigenit.
Dozimi i antitrupave Parimi është po ai si dhe për dozimin e antigenëve. Antigeni është i fiksuar mbi suportin e
ngurtë. Shtohet pastaj mjedisi biologjik që përmban antitrupat që duhen dozuar, pastaj një anti-imunoglobulinë e shënuar nga një enzimë (fosfatazë ose përoksidazë) dhe më në fund një reagent kromogjen që do të ndryshojë ngjyrën për të kaluar nga pa ngjyrë në ngjyrën e verdhë me fosfatazën alkaline dhe në ngjyrën blu me përoksidazën. Çdo etapë shoqërohet me një kohë inkubimi dhe mjaft shplarje. Shumica e antitrupave me rëndësi diagnostike në sëmundjet infektive si antitrupat anti-HIV, anti –HbsAg, anti –HCV apo autoantitrupat e sëmundjet autoimune etj. maten sot pikërisht me këto metoda ELISA.
Teknika të tjera Metodat ELIEDA (enzyme-linked immuno-electro-diffusion assay) dhe ELIFA (enzyme-linked immuno-electro-filtration assay) kombinojnë një imunoprecepitim nëpërmjet imunoelektrodifuzionit me zbulimin e komplekseve imune të precipituara në këtë mënyrë me antitrupa anti-imunoglobulinë të shënuara nga një enzimë. Metodat e imunotransferimit ose "western blot" të quajtura edhe imunoprint ose imunoblot kryhen në tri etapa. E para ka të bëjë me një elektroforezë në xhel të poliakrilamidit të përzierjes antigenike. Kjo ndiqet nga një elektro-transferim mbi nitrocelulozë (nën veprimin e një fushe elektrike) dhe një zbulim imunoenzimatik të reaksionit antigen antitrup. Këto metoda të shumta përdoren veçanërisht për vënien në dukje të antitrupave anti-HIV por gjithashtu dhe për karakterizimin e imunoglobulinave monoklonale ose të disa antitrupave antibërthamorë, ose në diagnostikimin e disa sëmundjeve infektive. Përdorimi i shënjuesve të ndryshëm në metodat me shënjues:
- në metodat radioimunologjike (RIA – radioimmuoassay) si shënjues radioaktiv përdoret kryesisht J125 që ka një ndjeshmeri mjaft të mirë por një gjysëm jetë të shkurtër (60 ditë) si dhe përbën rrezik ndotje radioaktive të ambjentit. për këto arësye metodat radioimuologjike (RIA) po zevendesohen gjithnjë e me shumë me metoda të tjera po aq të ndjeshme por me të “pastra” për ambjentin dhe me një qendrueshmeri shumë me të lartë:
-Metodat imunoemzimologjike (EIA). Kur këto metoda përdoren në faze të ngurte quhen dhe ELISA (enzyme-linked-immunosorbënt assay). Enzimat që përdoren me shpesh janë përoxidaza dhe fosfataza alkaline . Pas shtimit të substratit specifik ndodh zhvillimi i një ngjyre që matet me fotometri.
-Shënjimi me lende fluoreshente. Fillimisht ky shënjim u përdor për imunofluoreshence indore, direkte apo direkte me anë të mikroskopit me fluoreshence në substrate biologjike si inde, qeliza, bakterie, parazitë. Shënjuesit me shpesh të përdorur janë FITC (izotiocianati i fluorosceines si dhe rodamina). Sot shënues fluoreshente të tjerë të quajtur fluorofore po përdoren gjithmone e me tepër për teknika të ngjashme nga ndjeshmeria me metodat RIA. Këto metoda quhen edhe FIA (metoda imunofluoreshente). Fluoroforet janë molekula që absorbojnë energji dhe e rrezatojnë rishtas atë në një gjatësi vale me ndryshim 30-50 nm. një shënjues i tillë I përdorur shpesh është 4-Methylumbelliferonphosphat, i cili nëpërmjet fosfatazes alkaline defosforilohet në fluoroforin 4-Methylumbelliferon.
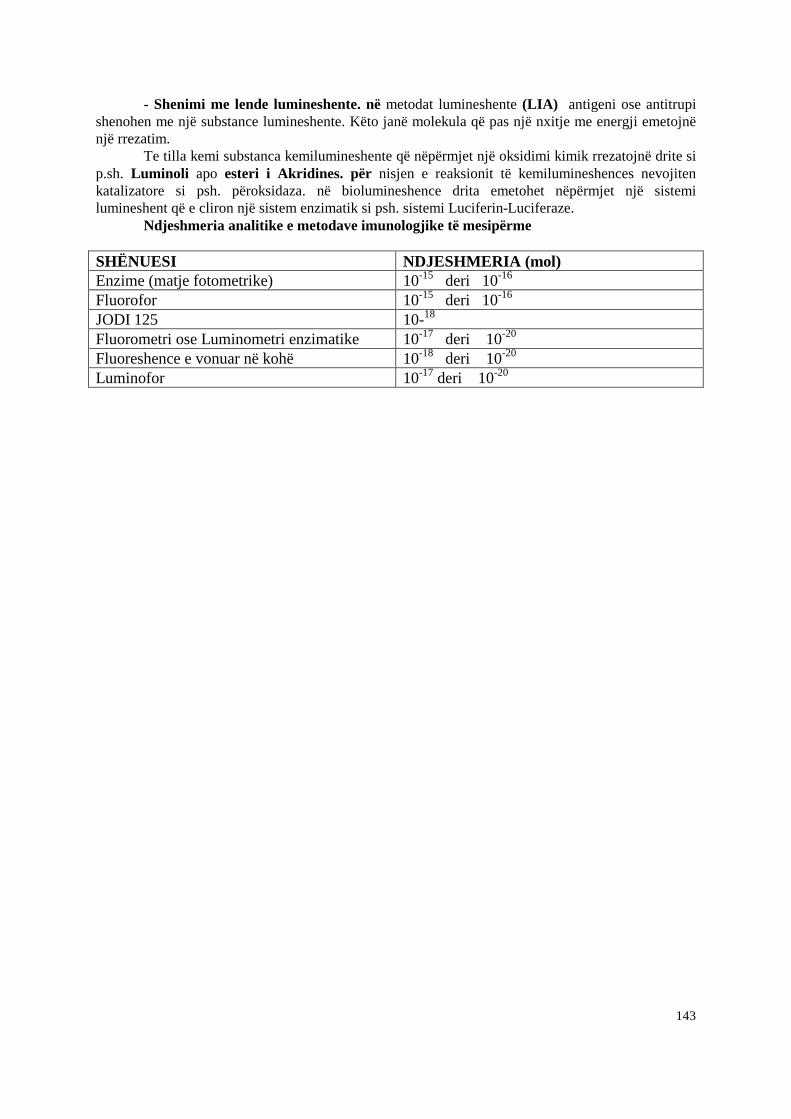
143
- Shenimi me lende lumineshente. në metodat lumineshente (LIA) antigeni ose antitrupi shenohen me një substance lumineshente. Këto janë molekula që pas një nxitje me energji emetojnë një rrezatim.
Te tilla kemi substanca kemilumineshente që nëpërmjet një oksidimi kimik rrezatojnë drite si p.sh. Luminoli apo esteri i Akridines. për nisjen e reaksionit të kemilumineshences nevojiten katalizatore si psh. përoksidaza. në biolumineshence drita emetohet nëpërmjet një sistemi lumineshent që e cliron një sistem enzimatik si psh. sistemi Luciferin-Luciferaze.
Ndjeshmeria analitike e metodave imunologjike të mesipërme
SHËNUESI NDJESHMERIA (mol) Enzime (matje fotometrike) 10-15 deri 10-16 Fluorofor 10-15 deri 10-16 JODI 125 10-18 Fluorometri ose Luminometri enzimatike 10-17 deri 10-20 Fluoreshence e vonuar në kohë 10-18 deri 10-20 Luminofor 10-17 deri 10-20

144
KAPITULLI 36. METODAT E IMUNOLOGJISE QELIZORE Paralelisht reaksioneve imunitare humorale, funksionojnë edhe reaksionet e tipit qelizor. Ekzistenca e tij është vënë në dukje që prej ekspërimenteve të Koch më 1891. Injëktimi brënda lëkurës (intradermë) i bacileve tuberkulare në një kavie shkakton zhvillimin brënda dy ose tri javësh të një plage lokale që nuk ka asnjë tendencë për shërim, kurse kafsha vdes brënda disa muajve me tuberkuloz të gjeneralizuar. Por, në qoftë se në një kavie të tillë të sensibilizuar riinjëktohen bacilet tuberkulare vihet re pas 24 orë deri 48 orë në pikën e injëktimit një reaksion lokal inflamator (fenomen i hipërndjeshmërisë) si dhe një shërim i kësaj vatre të dytë tuberkulare (fenomen i imunitetit të mbi infektimit), ndërsa plaga e parë vazhdon të zhvillohet. Ky eliminim i bakterieve të mbiinfektimit vjen nga makrofagët që shkojnë me shumicë në vendin plagës së dytë. Në kundërshtim me reaksionin me antitrupa, ky imunitet nuk mund t'i transmetohet një kafshe të re me anën e serumit, por mund të transmetohet me anën e injëktimit të limfociteve (të marra në ductus toracicus, në ganglionet limfatike ose në gjakun qarkullues). Është provuar më tej së limfocitet janë në origjinë të këtij reaksioni imunitar. Si pasojë e ngacmimit antigenik zhvillohen limfocite T efektorë të aftë të sigurojnë qoftë drejt për drejt (limfocitet T citotoksikë) qoftë në ndërmjetësinë e mediatorëve (citokina, të cilat veprojnë mbi kategori të tjera qelizore që janë në origjinë të një reaksioni inflamator), eliminimin e antigenit. Është fjala për një proces mbrojtjeje fiziologjik të organizmit, zakonisht e papërceptushme në mënyrë klinike, e cila siguron eliminimin e agjentëve patogjene intraqelizorë si dhe atë të qelizave të organizmit të infektuara nga viruset. Kjo përgjigje imunitare varet nga limfocitet CD4 prodhues të citokinave që veprojnë mbi qelizat fagocitare. Kur në një subjekt të sensibilizuar ky reaksion ka një shfaqje klinike (granulomë me shkatërrim indor) bëhet fjalë për reaksion hipërndjeshmërie të vonuar (maksimum 48 orë pas kontaktit të ri antigenik). Reaksione të tilla, kronike ose të gjeneralizuara, mund të shkaktojnë kaheksi si pasojë e një prodhimi të zgjatur të një citokine siç është TNF. SKEMA E REAKSIONIT IMUNITAR QELIZOR ANTIGENËT E INTERESUAR Shumica e imunogjeneve mund të shkaktojnë një reaksion imunitar qelizor. Është fjala veçanërisht për antigenë proteinike, për përbërës të agjentëve të shumtë infektivë (për shembull tuberkulina ose derivati proteinik i saj i purifikuar (PPD) i konsideruar prej kohësh si një model për studimin e këtij reaksioni), të Antigenëve të përshtatshmërisë indore që shkaktojnë reaksionet e flakjes së shartesës, në të cilat fenomenet e imunitetit qelizor lozin një rol themelor. Disa rrugë të futjes të antigenit janë më të favorshme se disa të tjera për zhvillimin e një reaksioni qelizor: është fjala sidomos për rrugët që sigurojnë një drenim të zgjedhur drejt ganglioneve limfatike

145
veçanërisht të rrugëve brënda lëkurës ose nën lëkurë, ndërsa rrugët intravenoze ose intrapëritoneale shkaktojnë më tepër një përgjigje të tipit humoral. Më në fund, adjuvanti i plotë i Freund favorizon dhe ai zhvillimin e këtij reaksioni të tipit qelizor. PËRGJIGJA PRIMARE Në përgjigjen imunitare me ndërmjetësinë qelizore, ngjarjet gërshetohen si vijon; -Etapa e parë ka të bëjë me paraqitjen e antigenit, gjithnjë me natyrë proteike, i degraduar dhe i shoqëruar me molekulat e kompleksit madhor të përshtatshmërisë indore. Kjo paraqitje sigurohet nga qelizat e quajtura Ag-paraqitëse (monocitet, makrofagët, qelizat dendritike, qeliza të Langerhans, limfocitet B), që lëshojnë një sinjal të përfaqesuar nga prodhimi i një citokine (interleukine 1) të destinuar për imfocitet T. Peptidet imunogjene të shoqëruara me molekulat e kompleksit të histopërshtatshmërisë së të klasës II u paraqiten limfociteve CD4+. Endonatigjenet siç janë proteinat virale të sintetizuara nga qeliza bujtëse e infektuar janë të shoqëruar me molekula të klasës I dhe u paraqiten limfocitet CD8+. -Etapa e dytë ka të bëjë me një aktivizim të limfocitit T. Ajo ndjek njohjen e antigenit nga receptori për antigenin e këtij limfociti T (TCR). Ky aktivizim shoqërohet me një rritje të vëllimit, me një bazofilli të citoplazmës dhe qeliza kalon atëherë në fazën e rritjes. -Etapa e tretë ka të bëjë me një proliferim të qelizës, nga e cila lind pas disa ciklesh të njëpasnjëshme një klon limfocitësh T me origjinë nga kjo qelizë. Kështu shfaqen nga njëra anë qeliza citotoksikë të tipit CD8, nga ana tjetër qeliza kujtesë me jetë të gjatë dhe të afta, gjatë një kontakti të ri me antigenin, për një proliferim të fuqishëm dhe për një prodhim të rëndësishëm citokinash. (Përgjigjadytësore). PËRGJIGJA DYTËSORE Ashtu si për përgjigjen sekondare humorale, përgjigjadytësore qelizore karakterizohet nga zhvillimi i saj i shpejtë dhe nga intensiteti i saj më i madh. Kjo përgjigjedytësore e organizimt që mund të vihet lehtë në dukje kur antigeni është rifutur me rrugë intradermike emërtohet zakonisht me emrin e hiperndjeshmërisë së vonuar. Reaksioni intradermik ndaj turbekulinës është një shembull fare klasik. Ka hipërndjeshmëri sepse reaksioni që vihet re gjatë këtij kontakti të dytë antigenik është më i fuqishëm sesa pas kontaktit të parë; ajo është e vonuar sepse zhvillohet 24-48 orë mesatarisht pas depërtimit të antigenit (në kundërshtim me fenomenet e hipërndjeshmërisë të lidhur me antitrupat që zhvillohen aty për aty, ose të paktën më shpejtësi). Dallohen tri faza të njëpasnjëshme të kësaj përgjigje sekondare. E para, e quajtur e induksionit, zhvillohet brënda disa orëve. Ajo është e karakterizuar nga ardhja me shumicë, si pasojë e takimit me antigenin, e limfociteve T kujtesë, nën ndikimin e citokinave me veprimtari kimiotaktike si interleukinat 1, 6 dhe TNF (shih më poshtë). E dyta, e quajtur e aktivizimit, shoqërohet me ndërhyrjen, gjithnjë nën ndikimin e citokinave, të qelizave fagocitare e të qelizave vrasëse, si dhe me një proliferim limfocitar që zgjeron reaksionin duke rritur prodhimin e citokinave. Këta të fundit aktivizojnë gjithashtu fibroblastet lokale që prodhojë fibrinë dhe kolagjen. Kjo fazë shoqërohet me një shkaterrim të indeve dhe me formimin e një fibroze që realizon kështu një granulomë. E treta merr fund me shërimin, me restitutio ad integrum. IMUNITETI QELIZOR I FETUSIT DHE I TË PORSALINDURIT . Zhvillimi i reaksionit imunitar qelizor të fetusi është akoma jo i njohur mirë. Megjithkëtë është provuar se limfocitet fetale janë të afta të reagojnë ndaj fitohemaglutininës qysh prej muajit të tretë ose të katërt të jetës intrauterine. Një reaksion pozitiv ndaj turbekulinës është e mundur të vihet në dukje të kafshët e porsalindura të ngacmuara in utero me BCG ose me bacile tuberkulare. Po kështu Uhr ka mundur të fitojë reaksione të hipërndjeshmërisë së vonuar tek kaviet e porsalindura të cilat kanë marrë gjatë jetës fetale injëktime antigenike bashkë me adjuvant të Freund.

146
Është e mundur të tregohet gjithashtu se reaksioni imunitar i flakjes së shartesës shfaqet rreth gjysmës së jetës fetale të qingjit. Pranohet se i porsalinduri ka një reaksion imunitar qelizor mjaft normal. Ështe në sajë të kësaj bindjeje që BCG mund të jepet në lindje sepse mbrojtja kundër tuberkulozit ka të bëjë më tepër me një mekanizëm qelizor. Ekzistojnë megjithatë ndryshime individuale sepse orvajtjet e sensibilizimit të të lindurve para kohë ose të të porsalindurve në kohë të bëra nga Uhr dhe bashkëpunëtorët e tij me dinitrofluorobënzen (shih më poshtë) nuk kanë pasur efekt veçse në rreth 1/3 e rasteve. MËNYRAT E STUDIMIT TË REAKSIONIT IMUNITAR QELIZOR MËNYRAT E STUDIMIT IN VIVO Është fjala kryesisht për teste të lëkurës që bëjnë të mundur qoftë të zbulohet ekzistenca e një kontakti antigenik të mëparshëm (reaksioni ndaj tuberkulinës për shembull), qoftë të vlerësohen mundësitë globale të reaktivitetit imunitar qelizor të një organizmi. Reaksioni imunitar që hulumtohet në këtë rast i përket tipit të katërt të hipërsensibilitetit (tipi i vonuar). Këto teste realizohen me një seri antigenësh ndaj të cileve subjekti është zakonisht i sensibilizuar dhe duhet të shfaqë një reaksion pozitiv. Një defiçit reaktiviteti imunitar qelizor dyshohet kur një numër i madh përgjigjesh ndaj këtyre testeve është negativ, sidomos në qoftë se kihet parasysh nocioni i përgjigjes pozitive të mëparshme ( negativizmi i reaksioneve ndaj tuberkulinës për shembull). Antigenët e përdorur Midis antigentëve të shumtë të përdorur, mund të përmënden:
-tuberkulina në formë të purifikuar (PPD) -kandidina -trikofitina -histoplazmina -varidaza (përzierje e streptokinazës dhe streptodornazës)
Përsa i përket dinitroklorobenzenit (DNCB) ose dinitrofluorobenzenit është fjala për substanca me të cilat organizmi nuk është në kontakt në mënyrë natyrore, por ndaj të cilëvë çdo subjekt normal mund të zhvilloje një reaksion imuniteti qelizor. Testet pra bëhen në dy kohë: në fillim sensibilizohet subjekti ndaj këtyre substancave; në kohën e dytë verifikohët fitimi i këtij reaktiviteti të ri. Këto prova lëkure realizohen me injektim në lëkurë (intrakutan) të një prej substancave të mesipërme (zakonisht 0,1 ml). Reaksioni i pritshem vërehet pas 48 oresh në formën e një papule të shoqëruar me eriteme diametri i të ciles në një reaksion pozitiv tejkalon një diameter prej 3-5 mm. Një tjetër substance e përdorur për këtë qellim është mitogjeni Fitohemaglutinine (PHA). Kjo është një lektine që ka efekt nxites jo specifik të limfociteve T. Ana pozitive e përdorimit të kësaj substance është se veprimi mitogjenik, nxites jo-specifik i limfociteve T nuk kërkon sensibilizim paraprak si në rastin e Antigenëve që nxisin limfocitet T në mënyrë specifike. Por nga ana tjetër PHA mund të shkaktoje dhe reaksione jo-specifike që mund të japin reazultat pozitiv fals. MËNYRAT E STUDIMIT IN VITRO Numërimi i limfociteve Ai përbën në fakt provën e parë që kërkohet për vlerësimin in vitro të imunitetit me ndërmjetësi qelizore. Ai realizohet me ndihmën e antitrupave monoklonalë të shenuar me lende fluoreshente të drejtuar kundër shënuesve të diferencimit (CD = cluster of differentiation) që janë

147
molekula specifike në sipërfaqen e limfociteve. Këta antitrupa të shenuar anti CD përdoren për identifikimin e një popullatë limfocitare të caktuar duke përdorur metodat e imunofluoreshences direkte. Kështu ështe e mundur që të njihet si numri absolut i limfociteve T (shënjuesit CD2, CD3) ashtu edhe numri i limfociteve T që i përgjigjet një nënpopullate të dhëne (CD4 për Limfocitet T ndihmës dhe CD8 për Limfocitet T citotoksikë dhe supresorë). Numërimi i limfociteve realizohet me anën e mikroskopit me fluoreshence ose me mirë me anë të imuocitometrise me fluks. Një metode tjetër tashmë e tejkaluar është edhe prova e rozetave E ( E si eritrocit) që përdor vetine që kanë limfocitet T të lidhin eritrocitet e dashit në sipërfaqen e tyre (eritrocitet e dashit kanë receptorë për molekulën CD2 në sipërfaqen e limfociteve T). Për realizimin e gjithë këtyre metodave nevojitet si hap i parë izolimi i limfociteve nga gjaku përiferik ose inde të tjera si psh. indi limfoid. Kjo zakonisht realizohet me anë të gradientit të densitetit me një përzjerje Ficoll-Izopak. Kjo përzjerje e një polisakaridi me peshe të madhe molekulare si Ficoll-I me Izopak- një substace me densitet të lartë që përdoret dhe si substance kontrasti në radiologji – ka aftësine që gjatë centrifugimit të lejoje kalimin dhe sedimentimin në fund të tubit të eritrociteve, trombociteve dhe polimorfoukleareve dhe izolimin në formë unaze të qelizave mononukleare (kryesisht limfocite) në zonën e kontaktit midis gjakut dhe përzjerjes me densitet të lartë. për këtë qellim gjaku me antikoagulant vendoset në një tub mbi përzjerjen e mesipërme Ficoll-Izopak dhe centrifugohet në 2500 rrotullime/min. për 30 minuta. Pas këtij centrifugimi përftohet unaza e bardhe me mononukleare (limfocite+monocite) midis fazes gjak dhe fikol-izopak-ut. Me këto qeliza mund të kryhen studime sasiore (numërim) ose funksionale. Numri I limfociteve T me këto metoda është 60-80 % i gjithë limfociteve. Ndër këto të fundit raporti CD4:CD8 është përafersisht 2:1. Ai konvertohet si pasojë e uljes se numrit të limfociteve CD4 (ne AIDS/SIDA) apo rritjes se limfociteve CD8 si në disa sëmundje virale. Edhe limfocitet B maten me të njëjtat metoda si limfocitet T. Shënjuesit që përdoren për këtë qellim janë imunoglobulinat membranore (Igs apo Igm) apo shënjuesit CD19 dhe CD20 për limfocitet B. Përqindja e limfociteve B në gjakun përiferik është 5 deri 15% e numrit të përgjithshem të limfociteve. Ndërsa shënjuesi CD54 përdoret për identifikimin e qelizave NK që në gjakun përiferik janë 10 deri 25% e numrit të përgjithshem të limfociteve. Provat funksionale: Testi i shndërrimit limfoblastik Parimi Limfociti i vogël ka veçorinë që të shndërrohet në një qelizë limfoblastike kur ai vihet në prani të disa substancave stimuluese. Midis këtyre substancave të quajtura blastogjene disa janë specifike: është fjala atëhere për antigenë ndaj të cilëve limfocitet e vegjel të organizimit janë të sensibilizuara, ndërsa të tjerat jo specifikë (mitogjenet siç është fitohemaglutinina). Teknika Duke u nisur nga gjaku venoz i mbledhur në mënyrë aseptike me heparinë, veçohen qelizat mononukleare d.m.th. kryesisht limfocitet plus monocitet; këto qeliza vendosen pastaj në një mjedis kulture (përqëndrimi final i rëndit 1x106 qeliza për ml). Shtohet agjenti blastogjenik i studiuar (antigeni ose dhe mitogjeni jo specifik). Një vështirësi madhore është në zgjedhjen me saktësi të madhe të hollimit të këtij antigeni: në qoftë se ai është i përqëndruar në mënyrë të pamjaftueshme, aftësia stimuluese e tij nuk mund të vihet në dukje; përkundrazi, përqëndrimet shumë të larta rrezikojnë të shkaktojnë një frenim të limfociteve. Tuba kontrolli pa agjent ngacmues janë të domosdoshëm me qëllim që të përcaktohen niveli i shndërrimeve limfoblastike spontane. Kultura inkubohet në inkubator në 37°C për një kohë minimum prej 3 ditë dhe deri 7 ditë. Në teknikën klasike sedimenti qelizor mblidhet, ndahet dhe ngjyroset (me May-Grunwald-Giemsa). Një numër i caktuar limfocitesh janë shndërruar në qeliza blastike (12-15 mikron, bërthamë voluminoze dhe citoplazmë shumë bazofile). Zakonisht një test quhet pozitiv në qoftë

148
se kultura përmban më tepër se 4% qeliza blastike, me kusht që tubi i kontrollit pa antigen të ketë më pak se 2% limfoblaste. Një teknikë më moderne mat inkorporimin e timidinës radioaktive nga kultura e limfociteve. Kjo timidinë e shtuar në mjedisin e kulturës në të njëjtën kohë me substancën blastogjenike, bashkohët me ADN bërthamore sinteza e së ciles nga limfoblastet është veçanërisht aktive. Koha e inkubimit është po ajo si dhe në teknikën e zakonshme; nxirret më vonë acidi dezoksiribonukleik, dhe përcaktohet radioaktiviteti. Mundet gjithashtu të studiohet edhe masa e sintezës të acidit ribonukleik në saje të uridinës ose adenozinës radiaktive. Substancat mitogjenike (quajtur ndryshe blastogjenike) jo specifike: Midis agjentëve jospecifikë, fitohemaglutinina (P.H.A.) shkakton shndërrimin e 50-70% të limfociteve të një subjekti të rritur normal. Ekziston një ndërlidhje mjaft e ngushtë midis aftësisë së individit për të zhvilluar një reaksion imunitar qëlizor dhe aftësisë së shndërrimit blastik të limfociteve të tij në prani të P.H.A. Kjo përgjigje ndaj P.H.A. konsiderohet si një pasqyrim i imunokompetencës të limfocitit. Ajo është e pakësuar shumë në sëmundjen e Hodgkin në zhvillim, gjatë sarkoidozës, në neoplazi të zhvilluara, kur kemi trajtime me imunosupresorë. Janë studiuar agjentë të tjerë blastogjenikë siç është konkanavalina A (Con A), serumi anti-limfocitar me dozë ngacmuese ose PWM (poke weed mitogen); këto kanë një veprim analog me atë të P.H.A. d.m.th. shkaktojnë induksion të shndërrimit të limfociteve T në limfoblaste. Përsa i përket pokeweed mitogen, ai shkakton shndërrimin njëkohësisht të limfociteve T dhe B. Agjentë mitogjenike Është fjala për Antigenë ndaj të cilëve organizmi ka qënë paraprakisht i sensibilizuar (antigenë bakterialë, medikamentozë, indorë). Fitohet atëhere një nivel shndërrimi që ndryshon nga 5-40 %. Bëhet fjalë për një reaksion besnik, pozitiviteti i të cilit bën të mundur që të afirmohet një sensibilizim ndaj një antigeni të dhënë, veçanërisht medikamentoz. Në dyshimet e autoimunitetit mund të gjëndet një test shndërrimi limfoblastik pozitiv kur limfocitet janë vënë në prani të një ekstrakti të organit që supozohet se është prekur nga një proces autoimunitar. Së fundi, kur përzihen me sasi të barabarta limfocitet që vijnë nga dy subjekte të ndryshëm A dhe B (kulturë mikste limfocitare) fitohet një numër i caktuar qelizash limfoblastike, përqindja e të cilave duket se është e lidhur me dallimet e antigenëve të pajtueshmërise indore që ekzistojnë midis dy individëve që testohen. Nuk vihet re asnjë shndërrim kur limfocitet vijnë nga dy binjakë hemozigotë. Në praktikë, me teknikën me të cilën matet integrimi i timidinës radioaktive bllokohet sinteza proteike e njërit prej llojit të limfociteve (për shembull limfociteve të individit A); kështu mund të matet veprimi ngacmues i këtyre limfociteve A mbi limfocitet e individit B. Vlerësimi i sintezës të interleukinave Me anën e metodave të sotme të ndjeshme imuologjike është bërë e mundur përcaktimi i sakte i të gjitha interleukinave në lëngjet e ndryshme duke përdorur antitrupa monoklonalë specifikë ndaj tyre. Studimi i funksionit të limfociteve B. Edhe funksioni i limfociteve B studiohet in vivo dhe in vitro. In vivo funksioni i tyre hulumtohet nëpërmjet studimit të përgjigjes antitrupore ndaj Antigenëve të ndryshëm si psh. anatoksina tetanike. një rritje sinjifikative e titrit të antitrupave antitetanike pas injëktimit të anatoksines tregon për një funksion normal të limfociteve B. Edhe niveli i përgjithshem i imunoglobulinave serike është tregues i rëndësishëm i funksionit antitrupor të limfociteve B. Këtu duhet të kihen parasysh normat e klasave të ndryshme të imunoglobulinave që ndryshojnë në vartësi të moshes. Kjo është sidomos karakteristike për IgA që ndryshon mjaft në përputhje me moshen dhe arrin nivelin e të rriturit pas adoleshences. Prandaj dhe vleresimi i një hipo-IgA eventuale (qe është mangësia me e shpeshtë e imunitetit humoral) duhet bërë me kujdes në vartësi të normave sipas moshes. Studimi i funksionit fagocitar

149
Duke patur parasysh praninë e mundshme të një mangësie imunitare të lindur në të sëmurë me infeksione të rënda dhe të përsëritura del e nevojshme hulumtimi i funksionit të fagocitozës në këto individë. Prova klasike e përdorur për këtë qellim është ajo e NBT (nitroblutetrazoliumit) që studion fagocitozën oksidative nëpërmjet ndryshimit të ngjyres intraqelizore të substances se përdorur. Një tjetër metode me e sofistikuar për studimin e fagocitozës është “kemilumineshenca”, një metode me e ndjeshme kjo për këtë qellim.
Recommended

















