Embed Size (px)
Citation preview

Міжклітинна передача електричного збудження
Андрій Чернінськийк.б.н., н.с. відділу фізіології мозку та психофізіології
ННЦ "Інститут біології"
Київського національного університету імені Тараса Шевченка
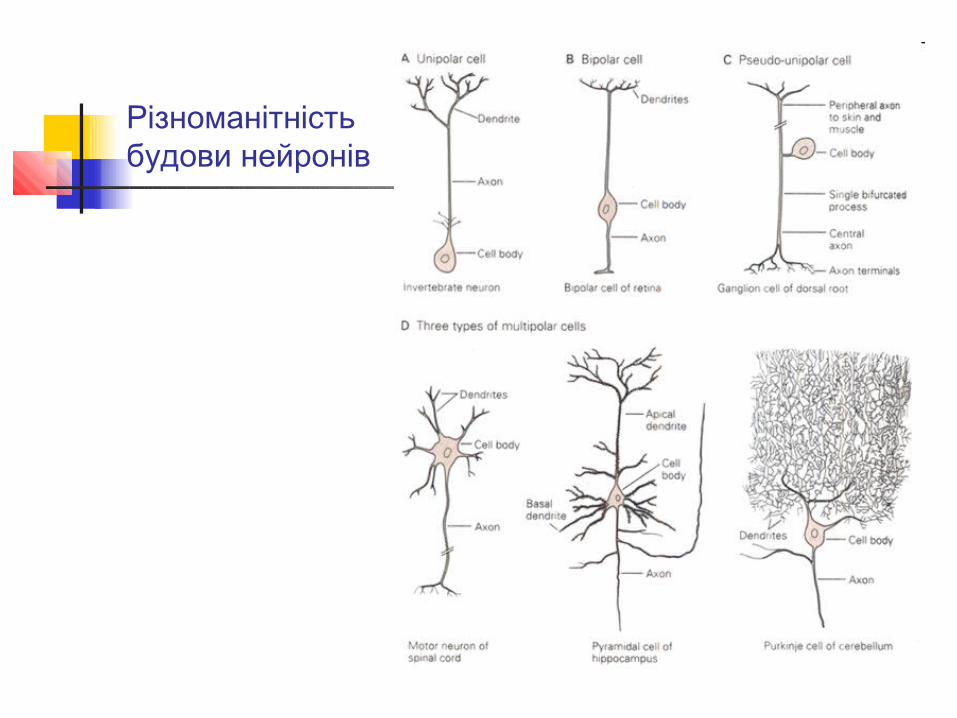
Різноманітність будови нейронів

Контакт двох нейронів: Синапс
Синапс (грец. σύναψις -обіймати, охоплювати) – місце контакту між двома нейронами.
Чарльз Шерінгтон, 1897 р.
В головному мозку людини налічується близько 10^15 синапсів
1 000 000 000 000 000

Будова хімічного синапсапресинаптичне закінчення
синаптичні пухирці
постсинаптична мембрана медіатор
рецептори до медіаторасинаптична щілина

Типи синапсів
Акс
о-со
мат
ичн
ий
Акс
о-д
енд
рити
чни
й
Акс
о-ак
сона
льн
ий
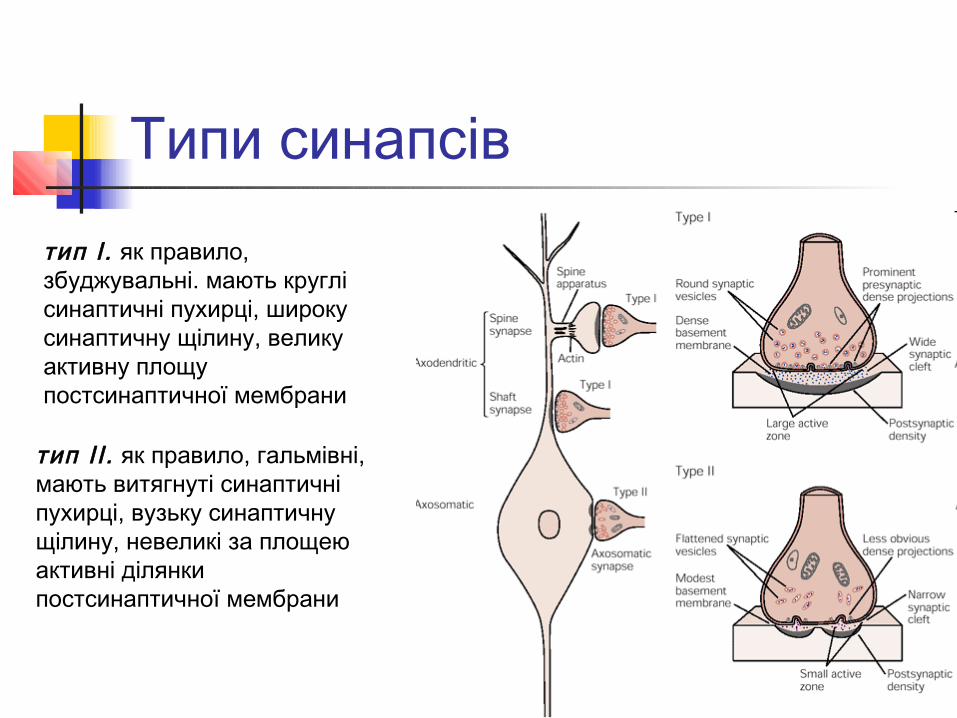
Типи синапсів
тип І. як правило, збуджувальні. мають круглі синаптичні пухирці, широку синаптичну щілину, велику активну площу постсинаптичної мембрани
тип ІІ . як правило, гальмівні, мають витягнуті синаптичні пухирці, вузьку синаптичну щілину, невеликі за площею активні ділянки постсинаптичної мембрани

Типи синапсів
"прохідний" "термінальний"
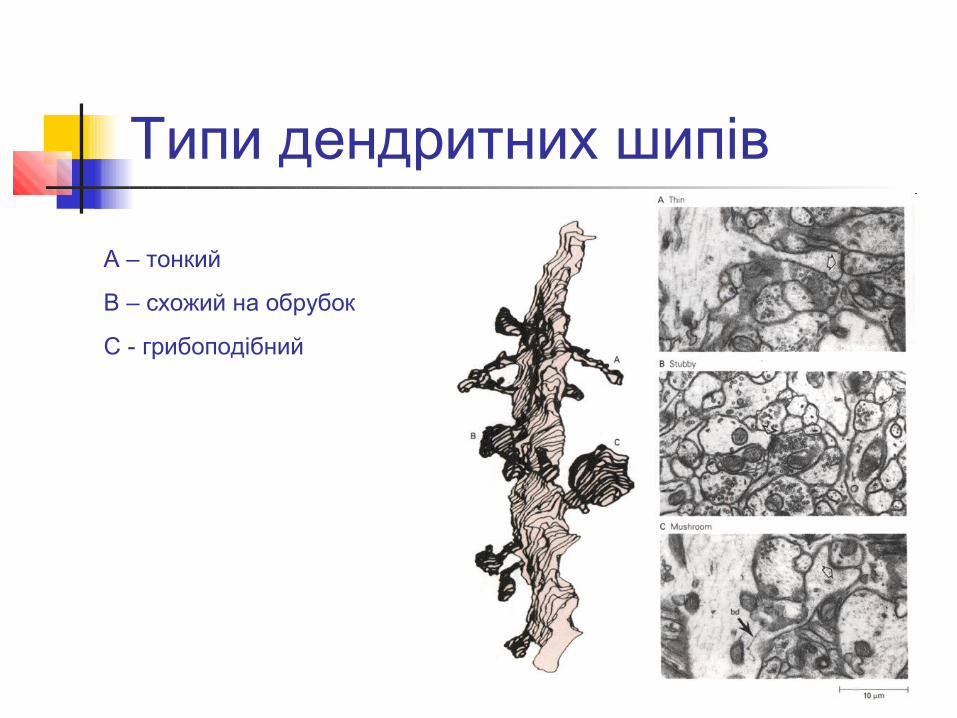
Типи дендритних шипів
А – тонкий
B – схожий на обрубок
C - грибоподібний

На одному нейроні ЦНС можуть закінчуватися сотні і тисячі інших нейронів
зелений колір відповідає певному синапсо-специфічному білку

Ріст дендритних шипів
на момент народження дитини її головний мозок мість набагато більше нейронів, ніж мозок дорослої людини
натомість, число існуючих синапсів на порядок менше
у процесі росту і навчання нефункціональні нейрони відмирають, а ті, що залишися формують нові синаптичні закінчення у відповідності до їх функціонального навантаження

Будова мембрани нейрона основою є подвійний
фосфоліпідний шар ліпідний шар є
непроникним для заряджених частинок – іонів
іони можуть проникати через мембрану за допомогою білкових молекул – іонних каналів

Іонні канали
Білкові молекули, які дозволяють проникати через мембрану зарядженим частинкам – іонам.
Властивості:
селективність
“все або нічого”
чутливість до певних чинників

Іонні канали (А) хемочутливі
(хімічні чинники) (С) потенціал-
залежні (В) залежні від
фосфорилювання (D) механочутливі

у стані спокою нейрон заряджений всередині негативно – мембранний потенціал спокою
збудження нейрона пов’язане із стрімкою зміною цього потенціалу, яка називається потенціал дії
виникши у одній клітині, потенціал дії повинен перейти до іншої
для цього існують спеціальні механізми
Концептуальні тези

У мембрані існують іонні канали, які здатні пропускати Натрій, Калій, Хлор
Ці канали завжди відкриті
Кількість Калієвих каналів є найбільшою:
nK:nNa:nCl = 1:0.04:0.45
Мембрана неоднаково проникна до різних іонів

Збудження нейрона: потенціал дії
потенціал спокою
деполяризація
гіперполяризація
потенціал дії
поріг збудження

Загальні поняття збудження передається з мембрани
на мембрану мембрана першої клітини
називається пресинаптичною мембрана другої клітини називається
постсинаптичною між двома мембранами є синаптична
щілина

Перехід збудження з однієї клітини на іншу
в межах однієї клітини деполяризація поширюється від збудженої ділянки до незбудженої, спричиняючи розвиток нового збудження
найбільш простий механізм – пряма передача збудження з мембрани на мембрану

Щілинний контакт мембран двох клітин
у мембранах обох клітин є особливі канальні білки – конексини
вони утворюють конексони – молекулярні пори, які дозволяють іонним струмам перетікати з однієї клітини до іншої

Морський молюск аплізія
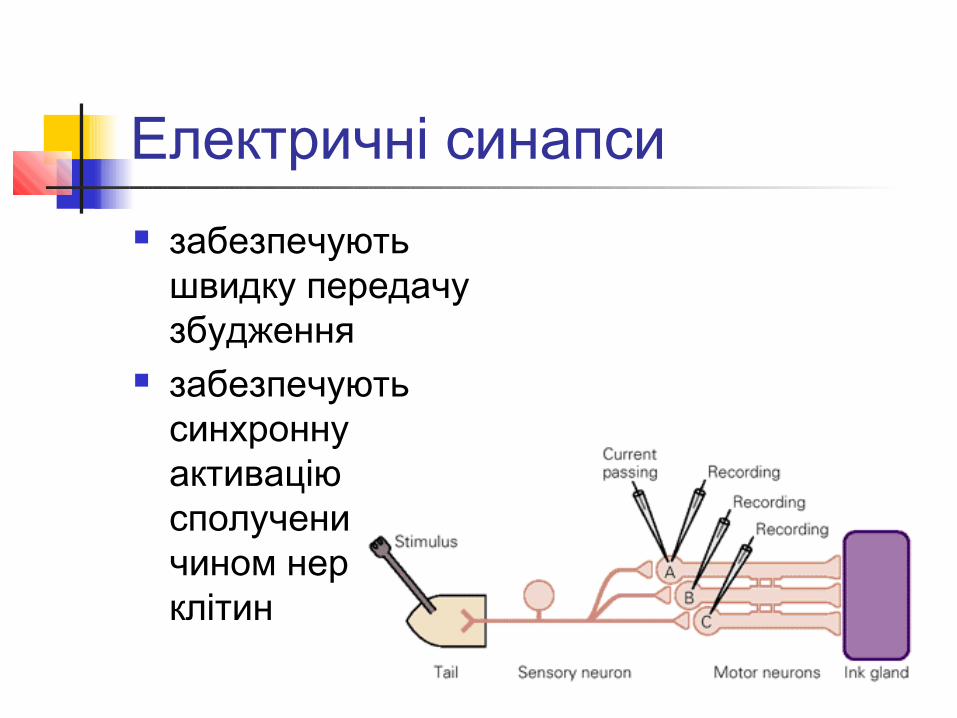
Електричні синапси забезпечують
швидку передачу збудження
забезпечують синхронну активацію сполучених таким чином нервових клітин

Інший тип передачі: хімічні синапси
мають більшого розміру синаптичну щілину: струми затухають до постсинаптичної мембрани

Принцип роботи хімічного синапса
збудження пресинаптичного закінчення вивільнення з нього у синаптичну щілину
хімічної речовини - медіатора (нейротрансміттера)
вплив медіатора на постсинаптичну мембрану відкриття іонних каналів зміна мембранного потенціала прибирання медіатора з синаптичної щілини

Вивільнення медіатора: потенціал-залежні кальцієві канали
деполяризація пресинаптичної мембрани призводить до відкриття потенціал-залежних кальцієвих (Ca2+) каналів
через них кальцій входить у клітину
зростання концентрації кальцію стимулює екзоцитоз – вивільнення в синаптичну щілину вмісту пресинаптичних пухирців
медіатор опиняється у синаптичній щілині

Екзоцитоз

Потенціал-залежні кальцієві канали обумовлюють у нейроні електро-хімічне спряження
зміна мембранного потенціалу → → зміна концентрації кальцію →
→ екзоцитоз → → вивільнення хімічного посередника
(медіатора)

Виділення медіатора відбувається порціями – квантами
квант медіатора – вміст одного пухирця

Вплив медіатора медіатор дифундує до постсинаптичної
мембрани
він зв’язується з рецепторними молекулами, розміщеними у ній
таке зв’язування призводить до змін структури рецепторної молекули
зміна структури рецептора прямо або опосередковано призводить до відкриття певних іонних каналів

Прямий вплив медіатора: ліганд-залежні іонні канали
медіатор зв’язується з рецепторною молекулою, яка одночасно є іонним каналом (іонотропні рецептори)
зв’язування з медіатором змінює структуру молекули так, що канал відкривається
через канал починає текти відповідний іонний струм

Непрямий вплив медіатора: вторинні посередники
зв’язування медіатора з рецепторною молекулою змінює її структуру(метаботропні рецептори)
ця зміна призводить до активації зв’язаного з рецептором G-білка
активація G-білка призводить до синтезу спеціальної речовини – вторинного посередника (наприклад, цАМФ)
цАМФ впливає на внутрішньоклітинні ферменти, які активують (або інактивують) іонні канали

Вторинні посередники невелика кількість медіатору може призводити до
синтезу значно більшої кількості вторинних посередників
більша кількість вторинних посередників активує більше іонних каналів, ніж міг би активувати/інактивувати медіатор безпосередньо
таким чином, система вторинних посередників дозволяє підсилювати сигнал у синапсі
ефективність системи вторинних посередників може змінюватися у часі – передумови навчання

Постсинаптичні іонні струми іонні канали, на функціонування яких можуть
впливати медіатори, селективні до основних іонів, розглянутих раніше – Na, K, Cl
прямий чи опосередкований вплив медіаторів може привести до активації (відкриття) або деактивації (закриття) іонних каналів
активація іонних каналів призводить до збільшення відповідного іонного струму, деактивація – навпаки
як буде мінятися мембранний потенціал при збільшенні або зменшенні різних іонних струмів?

Постсинаптичні іонні струми і потенціали
деполяризація
гіперполяризація
гіперполяризація
деполяризація
гіперполяризація
деполяризаціяхлор (Cl -)
калій (K+)
натрій (Na+)
зменшеннязбільшенняіон

Іонні канали постсинаптичної мембрани
здійснюють електрохімічне спряження, але у протилежному напрямку: перетворюють хімічний сигнал
(безпосередньо медіатор або вторинний посередник)
на електричні потенціали

Постсинаптичні потенціали
потенціал спокою
деполяризація
гіперполяризація
потенціал дії
поріг збудження
збуджувальні постсинаптичні потенціали
(ЗПСП)
гальмівні постсинаптичні потенціали (ГПСП)

Медіатори у відповідності до того, які зміни
мембранного потенціалу вони викликають, поділяються на збуджувальні і гальмівні
збуджувальні медіатори викликають деполяризацію - ЗПСП
гальмівні медіатори викликають гіперполяризацію - ГПСП

Післядія: прибирання медіатора дія медіатора не може тривати вічно, інакше
втрачається зміст передачі інформації
через деякий час медіатор прибирається з синаптичної щілини
шляхи елімінації медіатору: > дифузія, > розщеплення спеціальними ферментами,> всмоктування у пресинаптичне закінчення
у пресинаптичному закінченні відбувається синтез або ресинтез медіатору

Порушення елімінації медіатора: тривале продовження його дії
отрути нервово-паралітичної дії (зарин, зоман) блокують фермент ацетилхолінестеразу, яка в нормі швидко розщеплює ацетилхолін
АХ є основним медіатором у нервово-м’язових закінченнях
подовження збудження м’язів спричиняє судоми і параліч

Порівняння синапсів: електричних і хімічних
синаптична щілина мала (3-4 нм)
струми безпосередньо переходять з пресинаптичної мембрани на постсинаптичну
синаптична затримка відсутня
проведення двобічне переважно збуджуючі імпульс передається за
законом “все або нічого”, головне – швидкість реакції
синаптична щілина значних розмірів (20-40 нм)
необхідний хімічний посередник (медіатор)
синаптична затримка суттєва (1-5 мс)
проведення однобічне збуджуючі і гальмівні можуть змінювати свою
роботу (пластичність), що має вирішальне значення для навчання

Найбільш поширені медіатори
ацетилхолін глутамат норадреналін дофамін серотонін (адреналін)
гама-аміномасляна кислота (ГАМК)
гліцин (ацетилхолін)
збуджувальні гальмівні

Основний збуджувальний медіатор в ЦНС: глутамат
рецептори до глутамату можуть бути як іонотропнітак і метаботропні
активовані іонні канали селективні до натрію і калію одночасно
сумарний ефект є збуджуючим (деполяризація)

NMDA-рецептор
Рецептор-канальний комплекс складається з 77 білків
Мутації в генах 49 з цих білків пов'язані з розумовими порушеннями

Найбільш поширені гальмівні медіатори: ГАМК і гліцин
рецептори ГАМК можуть бути іонотропними (активують хлорні канали) або метаботропними (активують калієві канали)
рецептори до гліцина є іонотропними (також активують хлорні канали)

Гальмівний ефект ГАМК

Адренергічна система: блакитне місце
регуляція циклу сон-неспання
REM-сон

Дофамін
регуляція моторики
система позитивного підкріплення

Серотонін
регуляція психоемоційного стану людини, система позитивного підкріплення

Ацетилхолін

Наркотичні речовиниБлокатор зворотного захвату (reuptake) дофаміну
кокаїн – блокатор реабсорбції дофаміну;
подовжує вплив дофаміну, збільшуючи активацію системи позитивного підкріплення
проте! чим більше активуються дофамінові рецептори, тим більше потрібно надалі активатора – природного дофаміну або його аналога

Наркотичні речовини
Опіатні рецептори – протибольова система; регуляція соціальної поведінки
Ендогенні опіати, морфін, героїн

Ліки
Впливаючи на рецептори агоністами (активаторами) або антагоністами (блокаторами) можна досягти лікувального ефекту.

Ліки
На прикладі серотонінергічної системи показано місця впливу ліків, що чинять антидепресантний або ж протилежний ефекти.

Інститут фізіології НАНУПід керівництвом директора інституту – академіка НАНУ Олега Олександровича Кришталя – було відкрито два принципово нові рецептори, що беруть участь у медіації больового відчуття. Це рецептори до АТФ та до протонів.
Зараз йдуть дослідження шляхів блокування больового відчуття, які б не впливали на інші функціональні системи головного мозку.

Myotonia congenita спадкове захворювання, причиною якого є
порушення у гені CLCN1 цей ген кодує білок, який входить до
складу хлорного каналу постсинаптичної мембрани
дефект у цьому гені призводить до порушення хлорної провідності
як при цьому зміняться постсинаптичні струми?

Myotonia congenita

Tackling the synaptopathy
challenges - synaptic systems biology from
molecules to networks

Сучасна нейрогенетика

C57BL/A Inbr(A) ?+142. Origin. Little to A c1932. Maint. by A.C57BL/An Little to Andervont 1932. Differs from B6 and B10 at the Ce1 locus.C57BL/GrFa. Origin: Little to Gruneberg 1932, to Falconer 1947. Most British substrains derived from this stock, though 6 and 10 substrains have been imported more recently. This substrain seems to resemble the 6 rather than the 10 substrain.C57BL/KaLwN. To N 1965 from Lw at F35.C57BL/KsC57BL/6 Inbr (J) 150. Origin: substrains 6 and 10 were separated prior to 1937. This substrain is now probably the most widely used of al l inbred strains. Substrain 6 and 10 dif fer at the H9, Igh2 and Lv loci. Maint. by J,N, Ola.C57BL/10 Inbr (J) 158. Origin: see C57BL/6.C57BL/10ScSn. Inbr (J) ? +136. Little to W.L.Russell to J.P.Scott at F26 as a separate substrain. To Snell at F35-36. Behaviour differs from C57BL/10J.C57BL/10Cr Carries spontaneous lipopolysaccharide mutation lps which appears to resemble that found in C3H/HeJ.C57BL/Ola Carries a spontaneous mutation, Wlds, causing a marked slowing of axonal degeneration during Wallerian degeneration
C57BL/6 – найбільш популярна лінія


Для вивчення обрано ключові гени, продукти яких задіяні у розвитку таких захволювань як шизофренія, аутизм, депресія, СДУГ, епілепсія та ін.
1. економічний ефект2. успадкованість3. функціональна роль зазначених генів4. експериментальні напрацювання

В роботі синапса задіяно близько 2000 білків, з яких 1500 у постсинапсі.
Активація одного рецептора може викликати фосфорилювання /дефосфорилювання близько 200 сайтів 130 різних білків.
Мутації у 200 генах пов'язані із розвитком 133 захворювань людини.

Структура захворювань людини, які мають в своїй основі синаптопатології

Структурна організація взаємодії клітинних рецепторів (R), системи вторинних посередників (S) та внутрішньоклітинних ферментів (E).

Схематичне зображення взаємодій різних білкових комплексів, дефекти яких призводять до різних захворювань. Точки уражень, що пов'язані з різними хворобами, перекриваються.
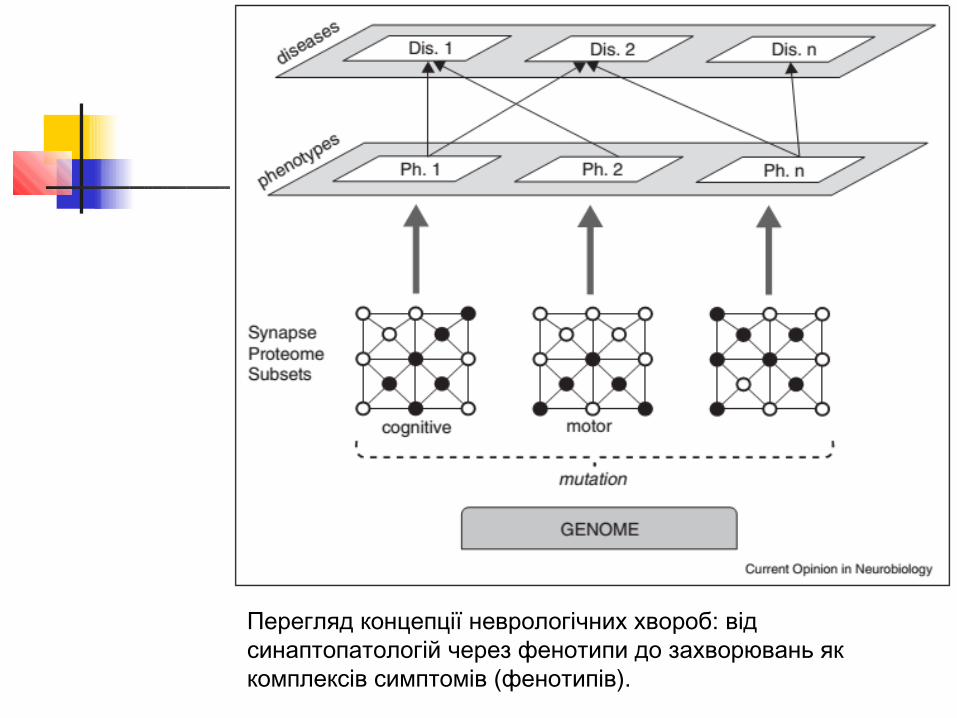
Перегляд концепції неврологічних хвороб: від синаптопатологій через фенотипи до захворювань як комплексів симптомів (фенотипів).

Бажаю усім тривалого і міцного розумового здоров'я!