Embed Size (px)
Citation preview

INTERRELACIONES METABÓLICAS
Los procesos metabólicos que se llevan a cabo en los diversos tejidos del cuerpo son interdependientes. No todas las rutas metabólicas principales y procesos del cuerpo operan en cada tejido en un momento dado. Según el estado nutricional y hormonal del individuo las rutas metabólicas pueden estar activas o no.
Los principales procesos que se analizarán serán la glucogenogénesis, glucogenolisis, gluconeogénesis, glucólisis, síntesis de ácidos grasos, oxidación de ácidos grasos, actividad del ciclo del ácido cítrico, oxidación de aminoácidos, síntesis de proteínas, proteólisis y síntesis de úrea. Para esto explicaremos qué tejidos son los más activos en diversos procesos, cuándo son estos procesos más y menos activos, y cómo se controlan estos procesos y su coordinación en diferentes estados metabólicos.
Para adquirir conocimiento de las relaciones entre las rutas metabólicas principales, nos familiarizaremos con los cambios que tienen lugar en el metabolismo durante el ciclo ayuno-alimentación.
CICLO AYUNO-ALIMENTACIÓN
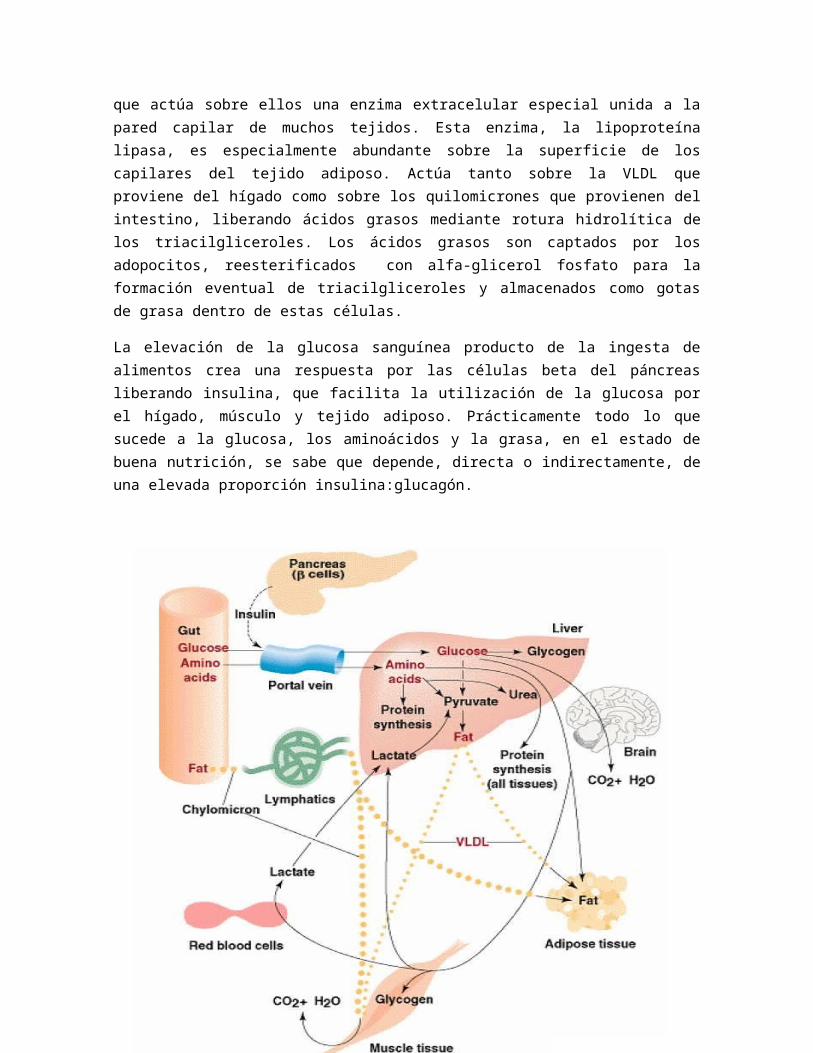
Estado de buena nutrición
En este estado, la glucosa y los aminoácidos pasan directamente a la sangra desde las células epiteliales del intestino, pasando al hígado a través de la vena porta. La grasa, en forma de quilomicrones pasa a la sangre en un sitio de flujo sanguíneo rápido. Esto último permite la rápida distribución de los quilomicrones impidiendo la coalescencia de las partículas de grasa.
El hígado es el primer tejido que tiene la oportunidad de utilizar la glucosa de la dieta. Después de penetrar la membrana plasmática de este tejido, la glucosa se puede convertir en glucógeno mediante la glucogenogénesis, en piruvato y lactato mediante la glucólisis, o utilizada en la vía de las pentosas fosfato para la generación de NADPH para los procesos sintéticos reductores. El piruvato formado a partir de la glucosa se puede oxidar a acetil CoA, el cual a, a su vez, se puede convertir en grasa por el proceso de la lipogénesis o se puede oxidar a CO2 y agua por el ciclo TCA. Gran parte de la glucosa que tiene del intestino evita el hígado y circula hacia los restantes tejidos corporales. El cerebro es uno de los principales consumidores de la glucosa sanguínea, dependiendo casi exclusivamente del catabolismo de este sustrato a CO2 y agua para la producción de ATP. Otros consumidores importantes de glucosa son los eritrocitos que sólo pueden convertir la glucosa en lactato y piruvato, y el tejido adiposo que la convierte en grasa. El músculo en el estado de buena nutrición tiene la capacidad de utilizar glucosa convirtiéndola en glucógeno o introducirla en las rutas glucolíticas y del ciclo TCA. Diversos tejidos producen, mediante la glucólisis, lactato y piruvato a partir de la glucosa circulante. El piruvato y el lactato circulan por la sangre a los tejidos que son metabolizadores activos de estos sustratos. En estado de buena nutrición, el hígado y el tejido adiposo son consumidores ávidos del piruvato y del lactato para el proceso de la lipogénesis, incluso cuando en el hombre la mayor parte de la grasa se sintetiza en el

hígado y no en el tejido adiposo. En el estado de muy buena nutrición, el hígado utiliza la glucosa y no emplea la gluconeogénesis. Así, el ciclo de Cori, que implica la conversión de glucosa en lactato en los tejidos periféricos, seguida de la reconversión del lactato en glucosa en el hígado, se interrumpe en el estado de buena nutrición.
El hígado es también el primer tejido que tiene la oportunidad de absorber de a sangre los aminoácidos dietarios. Como regla general, el hígado deja pasar a su través la mayoría de aminoácidos, a menos que la concentración de aminoácidos sea excepcionalmente elevada. Esto es de un importancia especial en el caso de los aminoácidos esenciales necesarios a todos los tejidos corporales para la síntesis de proteínas. El hígado puede catabolizar aminoácidos, pero los valores de Km con respecto a los aminoácidos de muchas de las enzimas implicadas son elevados, permitiendo que los aminoácidos se encuentres en exceso antes de que se produzca un catabolismo significativo. Por el contrario, las enzimas cargadores de los tRNA implicados en la síntesis de proteínas tienen unos valores de Km para los aminoácidos mucho más bajos. Esto asegura que en tanto estén presentes todos los aminoácidos, pueda tener lugar la síntesis de proteínas según se necesite para el crecimiento y para la síntesis de reemplazo. Los aminoácidos catabolizados en el hígado se pueden oxidar completamente a CO2 y agua, o bien los intermediarios pueden usarse como sustratos de la lipogénesis, cetogénesis o gluconeogénesis. Así, los aminoácidos en exceso que no son necesarios para la síntesis de proteínas acaban siendo convertidos en cuerpos cetónicos, glucosa o grasa, al tiempo que el nitrógeno amínico se convierte en urea. Los aminoácidos no retenidos por el hígado se pueden utilizar para la síntesis de proteína en otros tejidos. El músculo esquelético y el músculo cardiaco tienen una gran capacidad de transmisión de aminoácidos y de oxidación de los alfa-cetoácidos resultantes dando CO2 y agua. Los aminoácidos ramificados (leucina, isoleucina y valina) son tratados de un modo interesante. El hígado tiene poca capacidad para la transaminación de estos aminoácidos pero tiene una capacidad considerable para la descarboxilación oxidativa de los alfa-cetoácidos derivados de estos aminoácidos. En el músculo esquelético ocurre lo contrario, esto es, este tejido tiene considerable capacidad para la transaminación pero es relativamente deficiente respecto a las enzimas responsables del catabolismo posterior. La consecuencia es que la mayor parte de la transaminación tiene lugar en los tejidos periféricos como el músculo esquelético, los alfa-cetoácidos pasan a la sangre siendo oxidados en el hígado. Los aminoácidos ramificados constituyen una fuente importante de nitrógeno para la producción de alanina a partir de piruvato en el músculo.
Cuando se considera el envío de grasa hacia los tejidos, se debe diferenciar cuidadosamente entre grasa endógena y grasa exógena. Como se mencionó anteriormente, la glucosa, piruvato y aminoácidos se pueden utilizar para mantener la lipogénesis hepática. La grasa formada a partir de esos sustratos se libera del hígado en forma de VLDL para su transporte principalmente al tejido adiposo. La grasa de la dieta se libera a la sangre en forma de quilomicrones. Tanto los quilomicrones como las VLDL circulan por la sangre hasta que actúa sobre ellos una enzima extracelular especial unida a la pared capilar de muchos tejidos. Esta enzima, la lipoproteína lipasa, es especialmente abundante sobre la superficie de los capilares del tejido adiposo. Actúa tanto

sobre la VLDL que proviene del hígado como sobre los quilomicrones que provienen del intestino, liberando ácidos grasos mediante rotura hidrolítica de los triacilgliceroles. Los ácidos grasos son captados por los adopocitos, reesterificados con alfa-glicerol fosfato para la formación eventual de triacilgliceroles y almacenados como gotas de grasa dentro de estas células.
La elevación de la glucosa sanguínea producto de la ingesta de alimentos crea una respuesta por las células beta del páncreas liberando insulina, que facilita la utilización de la glucosa por el hígado, músculo y tejido adiposo. Prácticamente todo lo que sucede a la glucosa, los aminoácidos y la grasa, en el estado de buena nutrición, se sabe que depende, directa o indirectamente, de una elevada proporción insulina:glucagón.
Disposición de glucosa, amino ácidos, y grasa por varios tejidos en el estado de buena nutrición. Pag. 528. Text Book of Biochemstry with Clinical correlation - Devlin
Estado temprano del ayuno
El principio del ayuno ocurre cuando cesa la captación de combustible por el intestino. Durante este periodo transicional es muy importante la glucogenólisis del glucógeno hepático para el mantenimiento de la glucosa sanguínea.
La lipogénesis está restringida y el lactato, el piruvato y los aminoácidos que se habían utilizado para mantener este proceso se desvían hacia la formación de glucosa. Así, el ciclo de Cori está presente como parte de la producción de glucosa a partir de lactato por el hígado y volviéndose a convertir en lactato mediante la glucólisis en los tejidos periféricos tales como los eritrocitos. El ciclo de la alanina, en el que el carbono vuelve al hígado en forma de alanina en lugar de lactato, también se hace cada vez más importante como mecanismo para el mantenimiento de los niveles de glucosa sanguínea.
Dado que hay menos cantidad proveniente del intestino y que los niveles sanguíneos son generalmente bajos, el catabolismo de los aminoácidos como fuente de energía para los tejidos es muy bajo en la condición de principio del ayuno
Interrelaciones metabólicas de tejidos en el estado de ayuno temprano. Pag. 530. Text Book of Biochemstry with Clinical correlation - Devlin

Estado de ayuno
A medida que un individuo entra en un estado real de ayuno, ocurren modificaciones en el metabolismo para adecuarse a las condiciones actuales. En la situación de ayuno, del intestino no viene absolutamente nada de combustible y no queda glucógeno en el hígado para ayudar a mantener la glucosa sanguínea, la cual ahora depende completamente de la gluconeogénesis hepática, principalmente a partir del lactato, glicerol y alanina.
Una vez más, los ciclos de Cori y de la alanina juegan papeles importantes en el suministro de glucosa a los tejidos que dependen de este sustrato para la producción de ATP. No obstante, debe observarse que el Ciclo de Cori y el Ciclo de la alanina no proporcionan carbono para la síntesis neta de glucosa. En estos ciclos, la glucosa formada por el hígado tan solo reemplaza la que se ha convertido en lactato en los tejidos periféricos. No hay síntesis neta de glucosa. El cerebro oxida completamente la glucosa a CO2 y a agua y, probablemente, no participa de modo significativo en ninguno de estos dos ciclos. De ahí que sea absolutamente necesaria la síntesis neta de glucosa a partir de alguna fuente de carbono en esta situación. Los ácidos grasos no se pueden utilizar para la síntesis neta de glucosa ya que no hay ninguna ruta en los animales para tal conversión. De modo específico, el acetil CoA obtenido del catabolismo de los ácidos grasos no se puede convertir en intermediario tricarbonado de la gluconeogénesis. El glicerol, obtenido de la lipólisis de la grasa en el tejido adiposo, funciona como un sustrato importante para la síntesis de glucosa en el estado de ayuno.
Metabolismo del Glicerol. Química Biológica. A. Blanco

La mayor parte del carbono necesario para la síntesis de glucosa en estas condiciones proviene de la proteína de los diversos tejidos corporales, especialmente el músculo esquelético. La secuencia de acontecimientos se puede describir de la siguiente manera. Las proteínas son hidrolizadas dentro de las células musculares (proteólisis) produciendo aminoácidos. Una parte de estos aminoácidos se libera a la sangre y circula hasta los otros tejidos. De manera algo sorprendente la mayor parte de los aminoácidos no se liberan sino que son metabolizados parcialmente dentro de las células musculares en las que tuvo lugar la proteólisis. Sólo se liberan tres aminoácidos en grandes cantidades: alanina, glutamina y glicina. Parece que lo que sucede es que los otros aminoácidos se metabolizan mediante sus diversas rutas catabólicas para dar intermediarios (piruvato y alfa-cetoglutarato) que pueden producir alanina y glutamina. A continuación estos aminoácidos se liberan a la sangre de donde pueden ser extraídos por el hígado para la formación neta de glucosa.
Hay pruebas de que gran parte de la glutamina se convierte en alanina en las células del riñón del epitelio intestinal. La glutamina se oxida parcialmente en estas células proporcionando energía para satisfacer parte de la demanda metabólica de estos tejidos y el carbono y los grupos amino no empleados vuelven a liberarse a la sangre, en parte, en forma de alanina y NH4+. De ahí que la alanina sea, cuantitativamente, el aminoácido gluconeogénico más importante que alcanza el hígado en el estado de ayuno.
Ciclo glucosa-alanina. Química biológica. A. Blanco

La glicina liberada por el músculo se transforma, en parte, en serina en los riñones. La serina liberada por los riñones a la sangre se convierte a continuación en glucosa por el hígado. El tejido adiposo queda también muy involucrado en el estado de ayuno. Debido a la baja proporción insulina:glucagón que hay en esta condición, la lipólisis en este tejido está muy activada. Esto da lugar a niveles elevados de ácidos grasos en sangre que pueden utilizarse como combustibles alternativos a la glucosa por muchos tejidos. De hecho, el corazón y el músculo esquelético prefieren utilizar ácidos grasos antes que glucosa, quedando inhibida la glucólisis por la oxidación de los ácidos grasos. Por otro lado, el cerebro no utiliza ácidos grasos como sustrato ya que estos compuestos penetran muy poco la barrera hematoencefálica.
Los ácidos grasos juegan un papel importante en el hígado, siendo oxidados a través de la vía de la beta-oxidación para la provisión de la mayor parte del ATP necesario para respaldar las necesidades energéticas de la gluconeogénesis. Muy poco acetil CoA generado por la beta-oxidación del hígado se oxida completamente a CO2 y a agua por el ciclo TCA. Dado que durante la gluconeogénesis hay una tasa pequeña de actividad del ciclo TCA, en estas condiciones, el acetil CoA formado a partir de los ácidos grasos se convierte en cuerpos cetónicos por las mitocondrias hepáticas. Los cuerpos cetónicos se liberan a la sangre para ser usados como fuente de energía por los diversos tejidos. Al igual que con los ácidos grasos, muchos tejidos prefieren los cuerpos cetónicos a la glucosa de modo que la presencia de cuerpos cetónicos y ácidos grasos tiene un efecto de ahorro de la glucosa sanguínea, de modo que puede existir glucosa disponible al cerebro. A diferencia de los ácidos grasos, los cuerpos cetónicos penetran la barrera hematoencefálica. Una vez que su concentración en sangre es suficientemente elevada, los cuerpos cetónicos actúan como buen combustible alternativo para el cerebro. Son incapaces, sin embargo, de reemplazar completamente las necesidades de glucosa que tiene el cerebro. Los cuerpos cetónicos disminuyen la utilización de glucosa por el cerebro y, tal como si se anticipasen a este efecto, también suprimen la proteólisis en el músculo esquelético, disminuyendo hasta cierto punto el despilfarro que, de forma inevitable, tiene lugar durante la inanición. Mientras se mantienen los niveles de cuerpos cetónicos a valores elevados por la beta-oxidación hepática, hay
Catabolismo de glutamina en enterocitos. Text Book of Biochemstry with Clinical correlation - Devlin. Pag. 352

menos necesidad de glucosa, menos necesidad de aminoácidos gluconeogénicos y menos necesidad de utilizar tejido muscular mediante proteólisis.
Existe entonces una relación efectiva entre hígado, músculo y tejido adiposo en orden al suministro de glucosa al cerebro. El hígado funciona sintetizando glucosa, el musculo suministra el sustrato (alanina) y el tejido adiposo suministra el ATP (vía oxidación de ácidos grasos) necesarios para la gluconeogénesis hepática. Esta interacción depende, y es favorecida, por una baja proporción insulina:glucagón opuesta a la que favorece procesos característicos del estado de buena nutrición. Los niveles de glucosa son menores en la condición de ayuno, impidiendo la liberación de insulina y favoreciendo la liberación de glucagón del páncreas para dar una proporción insulina:glucagón mucho más baja que en el estado de buena nutrición.
Interrelaciones metabólicas de tejidos en el estado de ayuno. Pag. 531. Text Book of Biochemstry with Clinical correlation - Devlin

Estado de Re-nutrición
En este estado, la grasa es sintetizada al igual que en el estado de buena nutrición. La glucosa es pobremente extraída por el hígado. Es más, el hígado permanece es un estado gluconeogénico por un par de horas más luego de recibir alimento. En vez de proveer glucosa a la sangre, la gluconeogénesis hepática provee glucosa-6-fosfato para la glucogenogénesis. La glucosa es catabolizada en tejidos periféricos, produciendo lactato como suministro para la gluconeogénesis.
La gluconeogénesis de aminoácidos específicos absorbidos por el intestino también juega un rol importante en el restablecimiento normal de los niveles de glucógeno hepático por vía indirecta.
Al disminuir la tasa de gluconeogénesis, la glucólisis se convierte en el principal mecanismo de disposición de la glucosa, y el glucógeno hepático es mantenido por la vía directa de síntesis a partir de la glucosa en sangre.
Interrelaciones metabólicas de tejidos en el estado de re-nutrición. Pag. 533. Text Book of Biochemstry with Clinical correlation - Devlin





























