Embed Size (px)
Citation preview

ISTRAŽIVAČKI RAD
ISPITIVANJE UTICAJA ORHIDEKTOMIJE NA STEREOLOŠKE KARAKTERISTIKE GONADOTROPNIH ĆELIJA PACOVA
Autori:
Vojislav Gligorovski1
Božidar Mitrović1
Mentor:
dr Nataša Ristić2
1 Učenik III razreda Matematičke gimnazije u Beogradu (Kraljice Natalije 37)2 Institut za biološka istraživanja Siniša Stanković (Bulevar despota Stefana 142)

2
Sadržaj
1.Rezime 3
2.Uvod 4
2.1.Hormoni 4
2.2.Funkcije hipofize i hipotalamusa 5
2.3.Mehanizam negativne povratne sprege 5
2.4.Struktura LH i FSH hormona 7
2.5.Orhidektromija 7
3.Cilj rada 8
4.Materijal i metode rada 9
4.1.Laboratorijsko održavanje životinja 9
4.2.Dizajn eksperimenta 10
4.3.Priprema tkiva za svetlosnu mikroskopiju i histološka bojenja 10
4.4.Imunocitohemijsko obeležavanje gonadotropnih ćelija 11
4.5.Stereološka merenja 12
4.6.Statistička analiza podataka 13
5.Rezultati 14
5.1.Gonadotropne ćelije hipofize sham operisanih (SO) životinja 14
5.2.Gonadotropne ćelije hipofize orhidektomisanih (Orx) životinja 15
6.Diskusija 19
7.Zaključak 20
8.Zahvalnica 21
9.Literatura 22

3
1.Rezime
U ovom radu ispitan je uticaj orhidektomije odnosno hirurškog uklanjanja testisa na tri
stereološke karakteristike gonadotropnih ćelija hipofize. Stereološke karakteristike koje su
ispitivane su: zapremina jedra, zapremina ćelije i volumenska gustina ćelija. Istraživanje je
vršeno na ukupno četiri mužjaka pacova svrstanih u kontrolnu grupu (n=2) i eksperimentalnu
(orhidektomisanu) grupu (n=2). Kod svake jedinke ispitivane su obe vrste gonadotropnih ćelija
(folikulostimulirajuce i luteinizirajuće) koje su prethodno imunocitohemijski obojene.
Stereološke karakteristike su ispitivane pomoću mikroskopa (uveličanje objektiva 100x) na
osnovu standardizovanog sistema M42. Ispitivanje je pokazalo da se zapremina ćelija i jedara
statistički značajno ne menja nakon orhidektomije dok se volumenska gustina ćelija statistički
značajno povećava: u slučaju folikulostimulirajućih za 136% (p<0.05) i luteinizirajućih ćelija za
90%, (p<0.05). Ovakvo povećanje volumenske gustine gonadotropnih ćelija se može objasniti
diferencijacijom stelatnih ćelija u cilju što veće sinteze folikulostimulirajućeg (FSH) i
luteizirajućeg hormona (LH) koji stimulišu sintezu testosterona i spermatogenezu po principu
negativne povratne sprege.

4
2.Uvod
2.1.Hormoni
Najbitniju ulogu u održavanju homeostaze tela i reagovanju na draži iz spoljašnje sredine
ima endokrini sistem, koji deluje preko hormona. Hormoni su hemijski aktivne supstance koje se
izlučuju direktno u krv i koje putem krvi stižu do mesta delovanja. [1] Hormoni koji određuju
polnu karatkeristiku muškaraca i žena se nazivaju polni hormoni. Oni su ključni za razvoj polnih
organa kao i za polnu diferencijaciju mozga. Endokrine žlezde koje luče polne hormone su
gonade-ovarijumi, kod žena i testisi kod muškaraca. Testisi sekretuju nekoliko muških polnih
hormona koji se zajedno nazivaju androgenima. To su testosteron, dihidrotestoseteron i
androstendion. Testosteron se luči znatno više od ostalih hormona, te je najvažniji hormon
testisa.
Slika 1: Struktura testosterona
Kod muškaraca, pod dejstvom testosterona se razvijaju i održavaju sekundarne seksualne
karakteristike, kao što su: rast, maljavost, uvećanje polnih organa, povcećana produkcija sperme,
povećanje libida i mutiranje glasa. Testosteron se sintetiše u Lejdigovim ćeijama koje se nalaze
u semenim kanalićima testisa. Dnevno čovek izluči 4-9 mg testosterona. Količina testosterona u
telu se reguliše hipotalamo-hipofizno-gonadalnom osovinom (HPG osom), po principu
negativne povratne sprege. [2]

5
2.2.Funkcija hipofize i hipotalamusa
Hipofiza, žlezda smeštena u udubljenju sfenoidne kosti, ima ključnu ulogu u regulaciji
sistema endokrinih žlezda. Sastoji se od neurohipofize (pars neurosa) i adenohipofize (pars
distalis). Ćelije neurohipofize luče dva hormona, dok ćelije adenohipofize luče šest hormona
ključnih za regulaciju funkcije ostalih endokrinih žlezda, to su:
hormon rasta (GH), koji pokreće rast tela delovanjem na deobu i diferencijaciju ćelija adrenokortikotropni hormon (ACTH), koji utiče na metabolizam hranljivih materija tireostimulirajući hormon (TSH), koji kontroliše rad štitne žlezde prolaktin (PRL), koji pokreće razvoj mlečnih žlezda luteinizirajući hormon (LH), i folikulostimulirajući hormon (FSH), čija će funkcija biti detaljno objašnjena u daljem
tekstu.
Hipotalamus je deo mozga, smešten pri dnu treće moždane komore. Hipotalamus je
kontrolni centar za mnoge autonomne funckije perifernog nervnog sistema i njegova
funckcionalna pozicioniranost između nervnog i endokrinog sistema ukazuje da uma važnu
ulogu s jedne strane u održavanju stalnosti unutrašnje sredine (homeostaznu funkciju), a sa druge
strane od njegove aktivacije zavise neki oblici ponašanja koji bi se mogli definisati kao
instinktivno ili emotivno ponašanje. Pri tome treba imati u vidu da su ove dve grupe funckija
hipotalamusa međusobno tesno povezane. Aksonski terminali imaju sposobnost aktivacije
funkcije hipofize.[3]
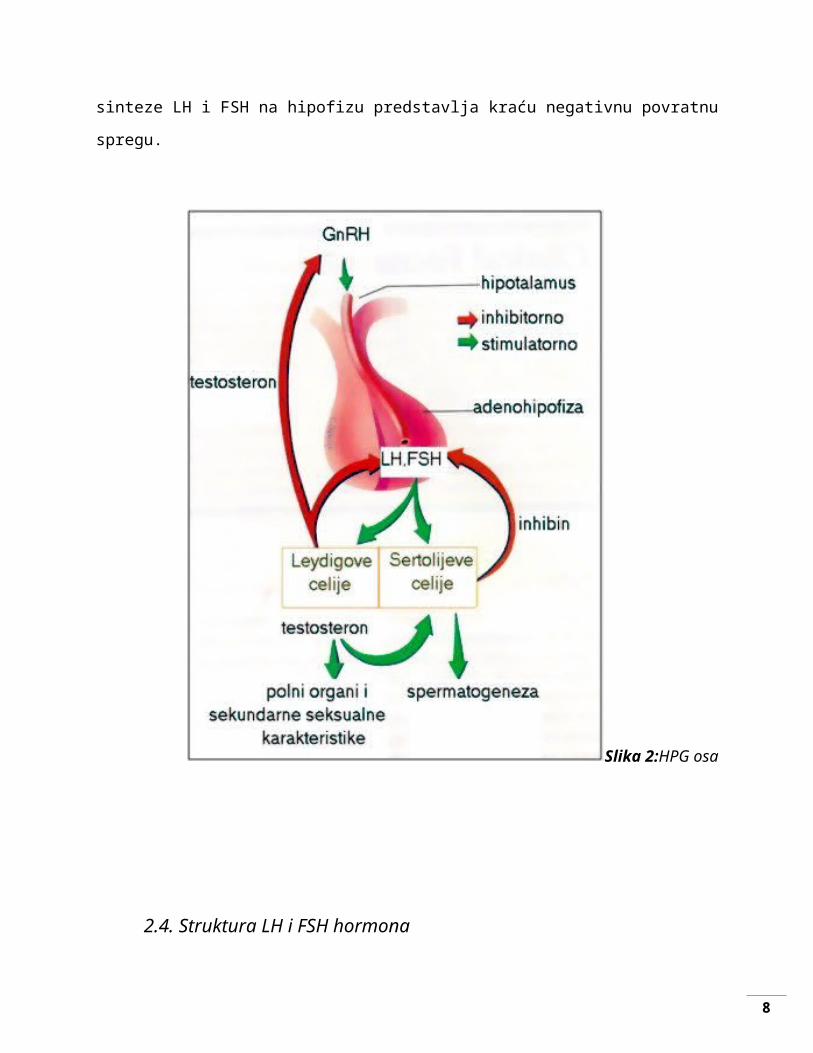
2.3.Mehanizam povratne sprege
Hipotalamus proizvodi gonadotropin-oslobađajući hormon (GnRH) koji dejuje na
hipofizu. [4] Sekretorne ćelije prednjeg dela hipofize pod uticajem GnRH luče gonadotropne
hormone: luteinizirajući hormon (LH) i folikulostimulirajući hormon (FSH). Nakon toga, LH i
FSH putem krvi dolaze do testisa i pokreću proizvodnju testosterona i spermatozoida. LH
stimuliše Lejdigove ćelije da sintetišu testosteron a FSH stimuliše Sertolijeve ćelije koje u
prisustvu testosterona iz Lejdigovih ćelija odvijaju spermatogenezu (proces nastanka i razvoja
spermatozoida). Povećanje nivoa testosterona u krvi negativnom povratnom spregom utiče na
hipotalamus koji smanjuje lučenje GnRH i hipofizu koja smanjuje lučenje FSH i LH.

6
Sertolijeve ćelije pri odvijanju spermatogeneze sintetišu i inhibin koji sprečava dalju
sintezu FSH delujući direktno na hipofizu slično kao testosteron.[5]
Delovanje testosterona na hipotalamus u cilju prestanka sinteze GnRH predstavlja dužu
negativnu povratnu spregu, dok zajedničko delovanje testosterona i inhibina u cilju prestanka
sinteze LH i FSH na hipofizu predstavlja kraću negativnu povratnu spregu.
Slika 2:HPG osa

7
2.4. Struktura LH i FSH hormona
Po strukturi gonadotropni hormoni su heterodimerni glukoproteini, tj. sadrže dve
različite subjedinice (Alfa i Beta) koje su sastavljene od ostataka amino-kiselina i ostataka
ugljenih hidrata. Alfa subjedinica je ista za LH i FSH hormon i kod čoveka se sastoji od niza 92
amino-kiseline, a kod ostalih kičmenjaka od niza 96 amino-kiselina na čijem se jednom kraju
nalaze ostaci ugljenih hidrata (fukoze, galaktoze, manoze, galaktozamina i glukozamina). Drugi
kraj alfa subjedinice je povezan sa beta subjedinicom koja je različita za LH i FHS hormon, dok
nevezujući kraj beta subjedinice takođe sadrži ostatke saharida. Ova subjedinica obezbeđuje
biološku aktivnost jer ima sposobnost vezivanja za receptore ćelijske membrane. Kod LH
hormona ona je izgrađena od niza 120 amino-kiselina, dok subjedinica FSH hormona sadrži 111
amino-kiselina. Poluživot (vreme za koje koncentracija hormona nakon lučenja opadne za
polovinu trenutne) LH hormona je 20 minuta dok je poluživot FSH hormona duži i iznosi 3-4
sata. Koncentracija FSH hormona u krvi je niža u odnosu na koncentraciju LH hormona.
2.5.Orhidektomija
Orhidektomija (kastracija) podrazumeva hirurško uklanjanje testisa, pasemenika
(epididymis) i semevoda (ductus deferens) sa okružujućim tkivom. U humanoj medicini,
primenjuje se kod pacijenata sa karcinomom prostate u poodmakloj fazi, kao i kod obolelih od
karcinoma testisa. Zahvaljujući tome pacijenti preživljavaju duži period nakon hirurške
intervencije. Kastracija je u istoriji ljudske vrste zabeležena i kao deo religioznih i socijalnih
običaja u nekim kulturama, vršena je i nad robovima i zarobljenim neprijateljima u toku ratova, a
kastrirani muškarci- evnusi su u nekim društvima činili i posebne socijalne klase. Danas je
ilegalna kastracija prisutna nažalost i u trgovini belim robljem kod oba pola, a u određenim
zemljama i društvima u jako viskom procentu je prisutna ženska kastracija.U mnogim državama
danas “dobrovoljna” hemijska ili hirurška kastracija je zakonom propisana opcija za seksualne
prestupnike, koja im omogućava reintegraciju u društvo, mada su efikasnost i etički aspekt ovog
tretmana predmet žučnih rasprava. Dalekosežne posledice orhidektomije su smanjenje gustine
koštanog tkiva, a time povećanje rizika od frakture kostiju. Njeni prateći efekti su i erektilna
disfunkcija, smanjen libido i impotencija.[6]
8
3.Cilj rada
Cilj ovog istraživačkog rada bio je da se ispitaju stereološke karakteristike luteotropnih i
folikulotropnih ćelija prednjeg režnja hipofize kod orhidektomisanih andropauzalnih pacova, kao
i da se kreira paralela izmedju orhidektomisanih i lažno operisanih pacova (kontrolne grupe).
Hipoteza, koja se postavlja na osnovu činjenica o HPG osi, je da će usled niske
koncentracije slobodnog testosterona u krvi kod orhidektomisanih pacova, doći do povećanja
koncentracije GnRH, a samim tim i LH i FSH hormona u krvi odnosno broja gonadotropnih
ćelija u prednjem režnju hipofize.

9
4.Materijal i metode
4.1.Laboratorijsko održavanje životinja
U eksperimentima su korišćeni andropauzalni pacovi, Wistar soja, starosti 16 meseci i
prosečne telesne mase od 650-700 g, koji su odgajani u odgajalištu Instituta za biološka
istraživanja “Siniša Stanković”. Laboratorijski uslovi ispunjavali su standardne zahteve: životinje
su boravile u plastičnim kavezima sa prostirkom od sterilne strugotine, pri konstantnoj
temperaturi od 20-22ºC i režimom osvetljenja 12 sati svetla, 12 sati mraka, čime je omogućen
dnevno-noćni ritam. Tokom izvođenja eksperimentalne procedure životinjama su bile dostupne
voda i laboratorijska hrana ad libitum. Receptura za pripremu laboratorijske hrane, koja kao
jedini izvor proteina sadrži kazein, preuzeta je od Picherit i sar. Sastav hrane je dat u tabeli 1.
Tabela 1: Sastav laboratorijske hrane
Sastojci %
Proteini
Casein (Alfa Aesar, Johnson Matthey GmbH &
Co.KG, Karlsruhe, Germany)
16.5%
Ugljeni hidrati
Kukuruzno brašno (Mlin produkt, S.Mitrovica)
75.2%
Celuloza
Cellulose, microcrystalline (Alfa Aesar. Johnson
Matthey GmbH & Co.KG, Karlsruhe, Germany)
2.0%
Masti
Suncokretovo ulje (Vital, Zrenjanin, Srbija)
2.0%
Vitaminska mešavina
(INSHRA PKB, Beograd, Srbija)
1.0%
Mineralna mešavina (di-Ca-fosfat, kreda, NaCl)
(INSHRA PKB, Beograd, Srbija)
3.3%
Ukupno 100%

10
4.2. Dizajn eksperimenta (eksperimentalne grupe)
Svi operativni zahvati nad životinjama izvođeni su u ketaminskoj anesteziji (15 mg/kg
t.m. ketamin hidrohlorid, Richter pharma, Wels Austria). Nakon zahvata, životinje su se dve
nedelje oporavljale, a potom su bili podeljene u grupe (n=2) i započet je tretman na sedeći način:
I grupa – SO
Lažno (sham) operisane životinje subkutano su tretirane dozom od 0.5 ml rastvarača
(mešavina 0.17 ml apsolutnog etanola i 0.33 ml sterilnog maslinovog ulja);
II grupa – Orx
Orhidektomisane životinje koje su takođe subkutano tretirane adekvatno zapreminom
rastvarača (0.5 ml mešavine apsolutnog etanola i sterilnog maslinovog ulja).
Eksperimentalni protokoli su odobreni od strane Komiteta za brigu o životinjama u
sklopu Instituta za biološka istraživanja „Siniša Stanković“ (Beograd, Srbija) u saglasnosti sa
preporukama propisanim u European Convention for the Protection of Vertebrate Animals used
for Experimental and Other Scientific Purposes (ETS no. 123, Appendix A).
4.3.Priprema tkiva za svetlosnu mikroskopiju i histološka bojenja
Nakon žrtvovanja, hipofize su fiksirane 48h u Bouin-ovom fiksativu. Dehidratacija tkiva
je obavljena serijom alkohola (30%, 50%, 70%, 96% i 100%), prosvetljavanje ksilolom, a
kalupljenje u parafinu. Ukalupljene hipofize su sečene rotacionim mikrotomom (Leica,
Germany), a debljina preseka je iznosila 5µm. Preseci su posle sušenja pripremljeni za
imunocitohemijsko bojenje. Na presecima hipofiza peroksidaza-antiperoksidaza (PAP;
Sernberger, 1970) metodom su obeležene LH i FSH ćelije.

11
4.4.Imunocitohemijsko obeležavanje gonadotropnih ćelija hipofize
Gonadotropne (FSH i LH) ćelije hipofiza mužjaka pacova kontrolnih i ekperimentalnih
grupa obeležavane su imunocitohemijskom metodom peroksidaza-antiperoksidaza (PAP;
Sternberger, 1970). Imunocitohemijske metode, t.j. specifična reakcija između primarnog antitela
i antigena omogućava diferencijalno bojenje ćelija koje nose specifični antigen, odnosno
hormon. Radi izbegavanja nespecifične imunoreakcije, na deparafinisanim i rehidriranim
presecima hipofiza aktivnost endogene peroksidaze je inhibirana inkubacijom u 0.3% vodonik
peroksidu u metanolu. Nakon toga preseci su ispirani u PBS-u (pH 7.4) dva puta po 5 minuta.
Zatim je sledilo inkubiranje u normalnom svinjskom serumu (DAKO A/S, Glostrup, Danska) 1h
na sobnoj temperaturi u koncentraciji 1:10 u PBS-u (pH 7.4), radi sprečavanja nespecifičnog
bojenja. Za detekciju gonadotropnih ćelija korišćena su primarna anti-FSH (1:300) i anti-LH
(1:500) anti-pacovska antitela (National Institut of Health, Bethesda, Md., USA) rastvorena u
PBS-u. Nakon reakcije antigen - primarno antitelo, svi preseci su prošli istu proceduru: ispiranje
u PBS-u, inkubiranje sa sekundarnim antitelima (svinja-antizec rastvorena u PBS-u–biotinylated
link, LSAB System-HRP, DAKO A/S, Glostrup, Danska) 10 minuta na sobnoj temperaturi,
zatim inkubiranje u Streptavidin-HRP kompleksu (Streptavidin-HRP, LSAB System-HRP,
DAKO A/S, Glostrup, Danska) 10 minuta na sobnoj temperaturi. Potom sledi ispiranje preseka
dva puta po 5 minuta u PBS-u (pH 7.4). Vizuelizacija kompleksa antigen antitelo obavljena je uz
pomoć hromogenskog supstrata, 3.3 diaminobenzidin-tetrahlorida (DAB, DAKO A/S, Glostrup,
Danska). Nakon procedure imunocitohemijskog bojenja, preseci su kontrastirani u Mayer-ovom
hematoksilinu (Merck-Alkaloid, Alkaloid, Skoplje, BJRM), a dehidrirani serijom etanola rastuće
koncentracije. Montiranje je obavljeno DPX-om (Mountant for Histology, Sigma-Aldrich USA).
Specifičnost antitela je potvrđena izostavljanjem primarnog antitela tokom
imunocitohemijske procedure, što je rezultiralo izostankom imunoreakcije.

12
4.5.Stereološla merenja Stereologija (od grčkih reč stereos – solidan, trodimenzionalan i logos – nauka) je nauka
o trodimenzionalnom tumačenju ravnih površina pomoću kriterijuma geometrijske verovatnoće.
Stereologija obuhvata multidisciplinaran pristup kojim se iz dvodimenzionalnih preseka nekog
tela (organa) kvantifikovano, trodimenzionalno procenjuje njegova unutrašnja građa,
korišćenjem testnih sistema kojima se pokriva referentni prostor u čijim okvirima se ispituje
određena struktura. M42 mnogonamenska testna mrežica se postavi na okular Zeiss svetlosnog
mikroskopa (Jena, Germany), a zatim nasumično pozicionira na hipofizni isečak na početku
brojanja. Brojanje se sprovodi na 50 testnih polja po isečku, pri uvećanju x 100 na mikroskopu
(Weibel, 1979).
Slika 3:Testni sistem M42
Koristeći stereološke metode određeni su sledeći stereološki parametri: prosečan
volumen LH i FSH ćelija hipofize (Vć, μm3), kao i njihovih jedara (Vj, μm3) i relativna
volumenska gustina ovih ćelija (Vvć, %).
Sve stereološke karakteristike su određene pomoću tri parametra:
Pj-broj tačaka koje su pale na jedro obojene ćelije
Ptć-broj tačaka koje su pale na telo obojene ćelije
Nj-ukupan broj obojenih jedara koji se nalazi unutar mrežice.
Na presecima hipofize jednog pacova izvršeno je ukupno 150 merenja mrežicom,
odnosno izvršeno je ukupno 1200 merenja. Za obradu podataka uzimane su srednje vrednosti
pomenutih parametara.

13
Volumenska gustina (Vvć) je određivana prema sledećoj formuli:
V v ć=Pt ć42
∗100 [%] (1)
Zapremina jedra odnosno zapremina ćelija (Vć,j) je određivana prema sledećim formulama:
V ć=V g∗β∗√Pt ć 3∗Vg [μm3] (2)
V j= Nj42
∗β∗√ Pj3∗Nj42
[μm3], (3)
gde je β koeficijent čija vrednost zavisi od oblika posmatranog objekta i on iznosi 1.382 za lopte
i 1.500 za elipse.
4.6.Statistička analiza podataka
Analizirani su srednji preseci hipofize kontrolnih i orhidektomisanih mužjaka na kojima
su imunocitohemijski obelezene gonadotropne ćelije. Statistička obrada podataka
imunocitohemijski obeleženih LH i FSH ćelija hipofiza pacova obuhvatala je analizu po dva
preseka iz tri različita nivoa hipofize, od dve životinje po grupi. Dobijeni podaci statistički su
obrađeni pomoću kopjuterskog programa Microsoft Office Excel 2007. Statistički značajnom se
smatra razlika između sređnih vrednosti parametara grupa ukoliko je verovatnoća ove razlike
p<0.05. Rezulati su prikazani kao srednja vrednost ± standardna devijacija.

14
5.Rezultati
5.1.Gonadotropne ćelije hipofize sham operisanih (SO) životinja
LH i FSH ćelije su kod SO životinja raspoređene pretežno ravnomerno, bilateralno i
rostrokaudalno, u prednjem režnju (parsdistalis), dok u potpunosti odsustvuju u srednjem režnju
(parsintermedia) adenohipofize. Mogu se videti ili kao pojedinačne ili u vidu grupa, posebno duž
sinusoidnih kapilara. Vrlo su krupne, a oblik im varira od ovalnog do poligonalnog. U citoplazmi
se uočava najčešće ekscentrično postavljeno, krupno, ovalno jedro sa jasno izdiferenciranim
jedarcem. Specifične granule su tamnije obojene i raspoređene po čitavoj citoplazmi.
Slika 4:Imunocitohemijski obeležene LH ćelije sham operisanih (SO) pacova (PAP metoda,
uvelićanje: x63)

15
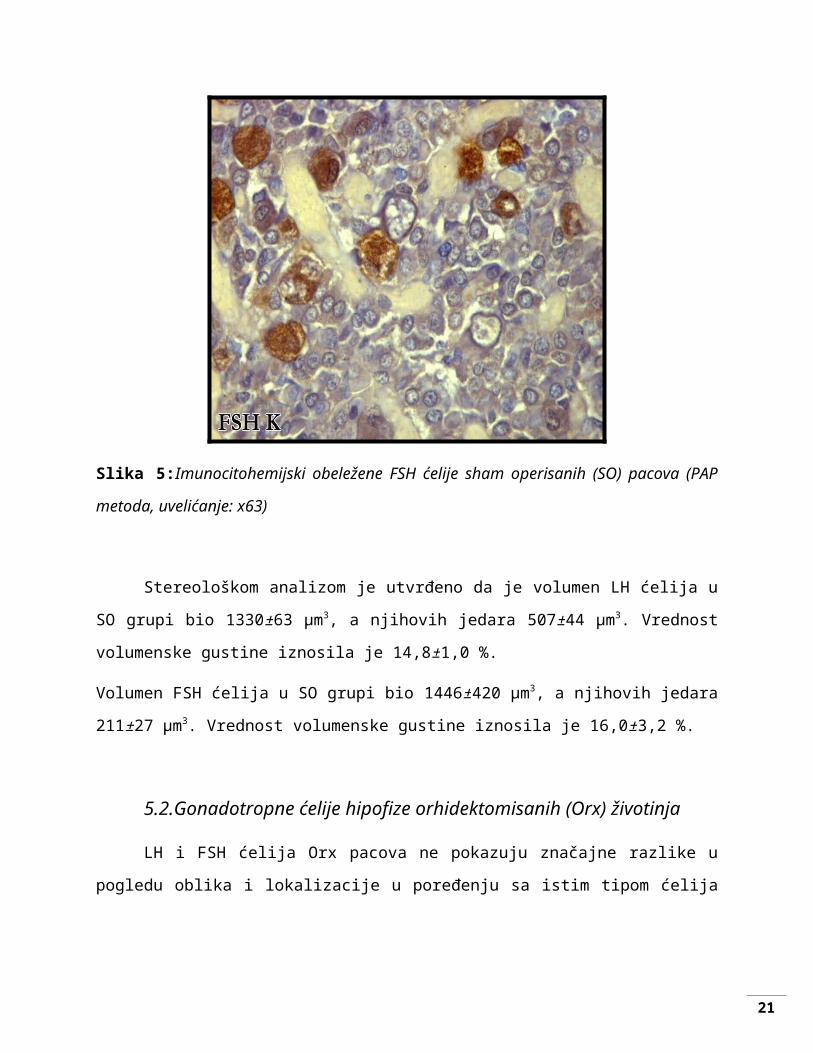
Slika 5:Imunocitohemijski obeležene FSH ćelije sham operisanih (SO) pacova (PAP metoda,
uvelićanje: x63)
Stereološkom analizom je utvrđeno da je volumen LH ćelija u SO grupi bio 1330±63
μm3, a njihovih jedara 507±44 μm3. Vrednost volumenske gustine iznosila je 14,8±1,0 %.
Volumen FSH ćelija u SO grupi bio 1446±420 μm3, a njihovih jedara 211±27 μm3. Vrednost
volumenske gustine iznosila je 16,0±3,2 %.
5.2.Gonadotropne ćelije hipofize orhidektomisanih (Orx) životinja
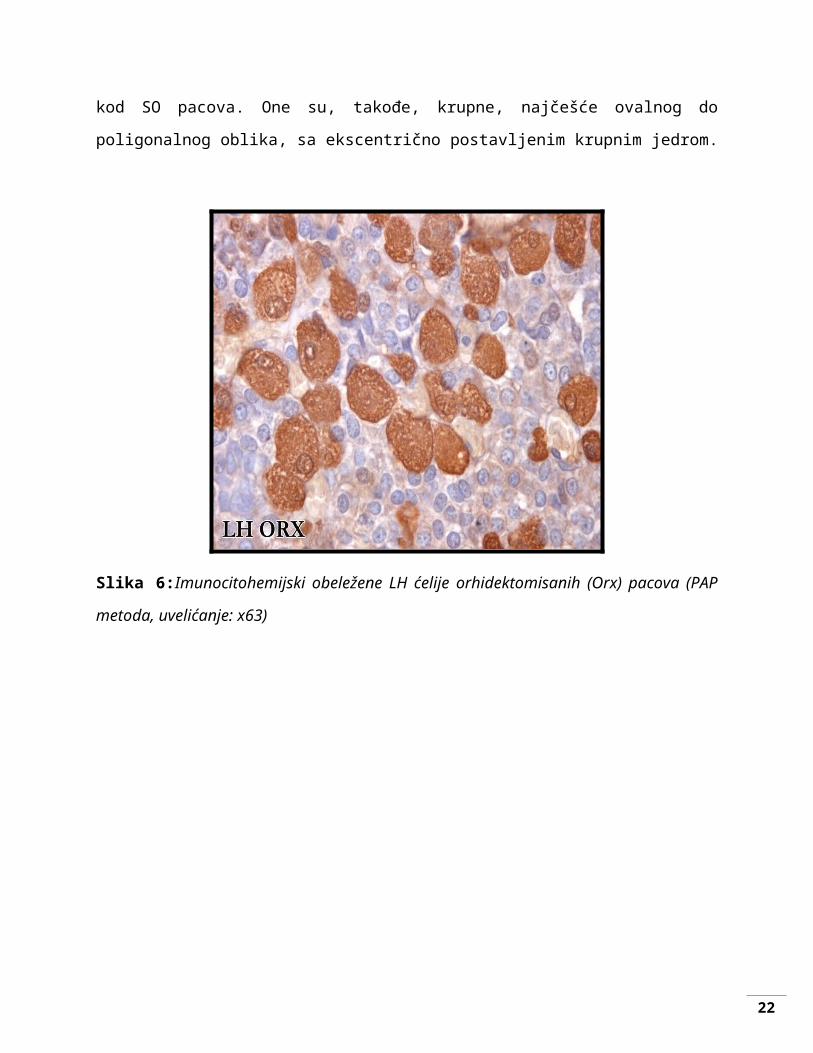
LH i FSH ćelija Orx pacova ne pokazuju značajne razlike u pogledu oblika i lokalizacije
u poređenju sa istim tipom ćelija kod SO pacova. One su, takođe, krupne, najčešće ovalnog do
poligonalnog oblika, sa ekscentrično postavljenim krupnim jedrom.

16
Slika 6:Imunocitohemijski obeležene LH ćelije orhidektomisanih (Orx) pacova (PAP metoda,
uvelićanje: x63)
Slika 7:Imunocitohemijski obeležene FSH ćelije orhidektomisanih (Orx) pacova (PAP metoda,
uvelićanje: x63)

17
Volumen LH ćelija u Orx grupi je bio 1643± 923 μm3, volumen njihovih jedara 620±217
μm3, a volumenska gustina 34,9±3,4%.
Volumen FSH ćelija u Orx grupi je bio 1087±300 μm3, volumen njihovih jedara 230 ± 9 μm3, a
volumenska gustina 30.5±0,3%.
Tabela 2: Stereološke karakteristike gonadotopnih ćelija pacova
Grupa Zapremina jedra [μm3] Zapremina ćelije [μm3] Volumenska
gustina [%]
LH Kontrolna 507±44 1330±63 14,8±1,0LH Orhidektomisana 620±217 1643±923 34,9±3,4FSH Kontrolna 211±27 1446±420 16,0±3,2FSH Orhidektomisana 230±9 1087±300 30.5±0,3
Tabela 3:Relativna promena stereoloških karakteristika gonadotropnih ćelija pacova
Tip
ćelij
a
Relativna promena
zapremine jedra [%]
Relativna promena zapremine
ćelije [%]
Relativna promena
volumenske gustine [%]
LH 22,3 23,6 135,9
FSH 8,7 -24,8 90,0

18
0200400600800
10001200140016001800
Vć[μm3]
LH K LH ORX FSH K FSH ORX
0
100
200
300
400
500
600
700
Vj[μm3]
LH K LH ORX FSH K FSH ORX
0.00%5.00%
10.00%15.00%20.00%25.00%30.00%35.00%40.00%
Vvć
LH K LH ORX FSH K FSH ORX
Histogram 1. Promene A) volumena LH i FSH ćelija (μm3); B) volumena njihovih jedara (μm3);
i C) volumenska gustina (%) kod: Sham operisanih pacova (K,SO) i orhidektomisanih pacova
(Orx).
A
B
C
19
6.Diskusija
Statistički značajno povećanje parametra uočava se samo kod volumenske gustine, koja
je kod LH ćelija veća za 136% a kod FSH ćelija za 90% kod orhidektomisanih pacova. Ova
činjenica može se objasniti postojanjem nediferenciranih stelatnih ćelija u adenohipofizi koje
kod kontrolnih jedinki imaju potpornu i nutritivnu ulogu ostalim ćelijama adenohipofize.
Kod orhidektomisanih pacova, koncentracija testosterona u krvi je niska što po principu
povratne sprege izaziva lučenje gonadotropin-oslobađajućeg hormona koji putem aksona stiže do
hipofize gde stimuliše rad luteotropnih i folikulotronih ćelija. Ove ćelije luče hormone kako bi se
sintetisao testosteron u što većoj količini, sve dok se ne dostigne njegova optimalna
koncentracija i ne počne odvijanje spermatogeneze, što kod orhidektromisanih pacova nije
moguće usled nedostatka Lejdigovih i Serotolijevih ćelija odstranjenih hirurškim putem. Dakle,
nepostojanje testosterona i inhibina iz testisa u HPG osi se direktno odražava na stimulaciju LH i
FSH ćelija koje onda teže da luče što više hormona.
Iz tog razloga javlja se potreba i za novim LH i FSH ćelijama. Stelatne ćelije koje su
imale potpornu i nutritivnu ulogu, nakon orhidektomije menjaju svoju funkciju i diferencijacijom
prelaze u gonadotropne ćelije, u cilju što većeg lučenja luteinizirajućeg i folikulostimulirajućeg
hormona.

20
7.Zaključak
Na osnovu rezultata dobijenih ispitivanjem efekata orhidektomije na stereološke
karakteristike LH i FSH ćelija možemo izvesti zaključak da je posledica orhidektomije porast
volumenske gustine ćelija, kao što je i pretpostavljeno. Takođe se zaključuje da orhidektomisani
pacovi nemaju značajno izmenjenu zapreminu gonadotropnih ćelija adenohipofize kao ni
zapreminu njihovih jedara, u odnosu na kontrolnu grupu.
Volumenska gustina LH ćelija orhidektomisanih pacova u odnosu na kontrolnu grupu je
veća za 136%, dok se ova karakteristika poveća 90% kada su u pitanju FSH ćelije.
Zaključuje se da nakon orhidektomije, jedan broj stelatnih ćelija u prednjem režnju
hipofize koje su ranije imale nutritivnu ulogu diferencijacijom prelaze u gonadotropne, odnosno
LH i FSH ćelije kako bi obezbedile što veću količinu luteinizirajućeg i folikulo-stimulirajućeg
hormona u krvi.
21
8.Zahvalnica
Želeli bismo da se zahvalimo našem mentoru, stručnom saradniku Odeljenja za citologiju
IBISS-a, dr. Nataši Ristić koja nam je svojim iskustvom i znanjem pružila neizmernu podršku u
izradi rada, zatim dr. Verici Milošević, naučnom savetniku i rukovodiocu Odeljenja za citologiju
IBISS-a na ideji za projekat i konstruktivnim predlozima pri izradi eksperimentalnog dela rada.
Takođe se zahvaljujemo i svim ostalim stručnim saradnicima Odeljenja za citologiju IBISS-a
koji su nam sugestijama pri osmišljavanju i realizaciji projekta učinili ovaj rad boljim. Veliku
zavhalnost dugujemo i našoj profesorki biologije, Jasmini Stošić na podsticaju za rad na ovom
polju biologije.
22
9.Literatura:
1.Kovačević R., Kostić T., Andrić S., Zorić S.: “Opšta fiziologija životinja, skripta za
studente biologije“
2.Ciccone N., Raiser U.: “The biology of gonadotroph regulation“
3.Petrović V., Cvijić G.: “Endokrinologija opšta i uporedna“
4.Huang B., Leu S., Yang H., Norman R.: “Testosterone effects on luteinizing horome
and follicle-stimulating hormone responses to gonadotropin-releasing hormone in the mouse “
5.Winters S., Moore J.: “Intra-pituitary regulation of gonadotroph in male rodents and
primates“
6.Bijelić D.: “Efekti estradiola i izoflavona soje na somatotropne ćelije hipofize u
animalnom modelu andropauze“
7.Vogel D., Sherins R.: “Orchiectomy in young rats results in differential regulation of
FSH and LH hormone content“





























