Embed Size (px)
Citation preview

Lezione 3
I geni nelle popolazioni (2)

Cambiamenti casuali delle frequenze alleliche dovuti alle dimensioni finite delle popolazioni
Questo tipo di evoluzione dovuto alla deriva non si può predire; è causale o stocastico
L’effetto della deriva dipende dalla dimensione della popolaizone ed è più forte in piccole popolazioni
Una forza evolutiva di grande efficacia
Deriva genetica

Deriva genetica
Un approccio stocastico
Probabilità: breve ripasso
Genetica ed evoluzione hanno una natura stocastica
I possibili risultati di un processo stocastico si chiamano eventi. (Un processo deterministico ha solo un possibile risultato)
La probabilità di un evento è la frazione di risultati (tutti i casi possibili) in cui si verifica tale evento (casi favorevoli). La probabilità di un evento A viene indicata con P(A).
0 < P < 1
La somma delle probabilità di tutti gli eventi mutuamente esclusivi è 1
P(i) = 1i=1
n∑
Deriva genetica
Un approccio stocastico
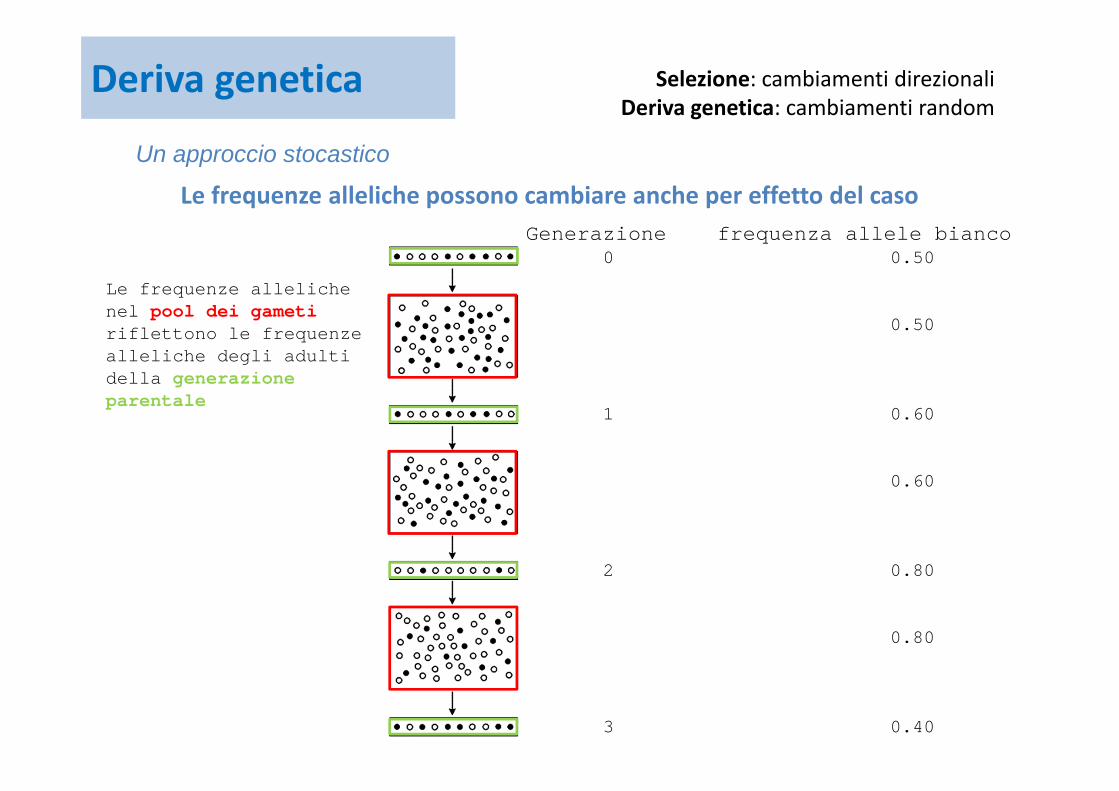
Le frequenze alleliche possono cambiare anche per effetto del casoGenerazione frequenza allele bianco
0 0.50
0.50
1 0.60
0.60
2 0.80
0.80
3 0.40
Selezione: cambiamenti direzionaliDeriva genetica: cambiamenti random
Le frequenze alleliche nel pool dei gameti riflettono le frequenze alleliche degli adulti della generazione parentale
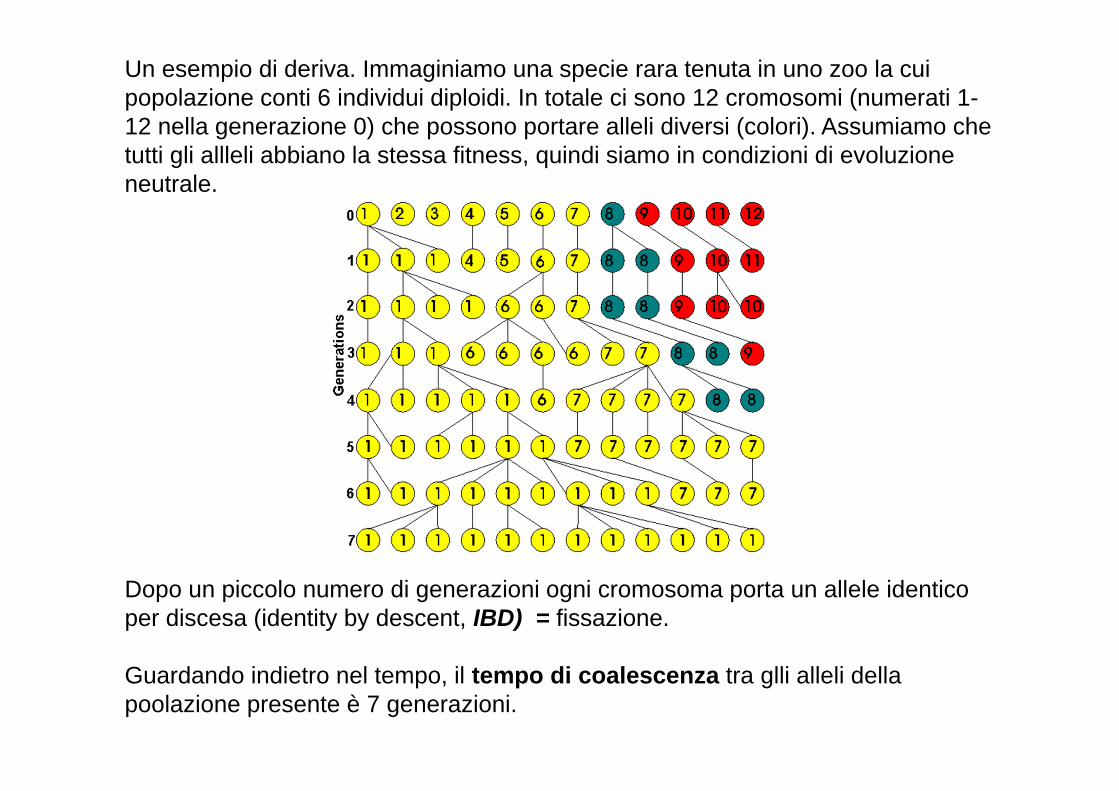
Un esempio di deriva. Immaginiamo una specie rara tenuta in uno zoo la cui popolazione conti 6 individui diploidi. In totale ci sono 12 cromosomi (numerati 1-12 nella generazione 0) che possono portare alleli diversi (colori). Assumiamo che tutti gli allleli abbiano la stessa fitness, quindi siamo in condizioni di evoluzione neutrale.
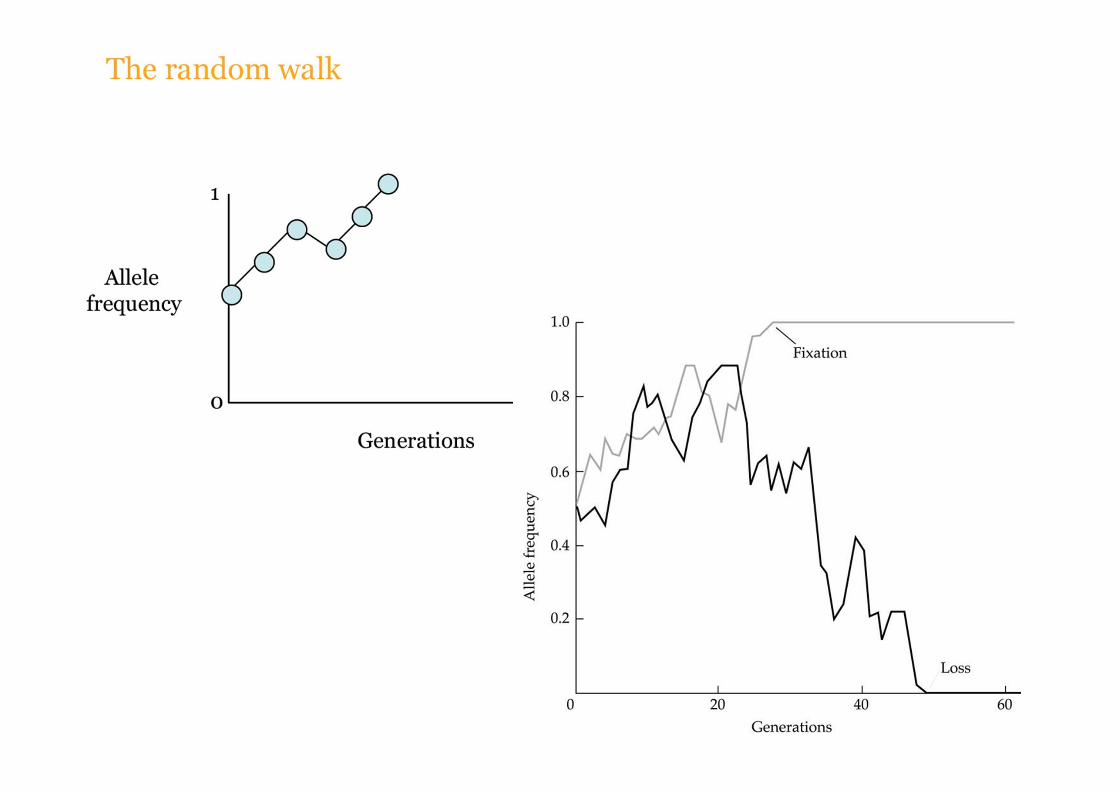
Dopo un piccolo numero di generazioni ogni cromosoma porta un allele identico per discesa (identity by descent, IBD) = fissazione.
Guardando indietro nel tempo, il tempo di coalescenza tra glli alleli della poolazione presente è 7 generazioni.

Deriva genetica
Un approccio stocastico
Modello di Wright‐Fisher (popolazione ideale)
La dimensione della popolazione rimane costante
Le generazioni non sono sovrapposte
Tutti gli alleli hanno la stessa probabilità di essere campionati
Sex ratio = 1
Se non c’è selezione e la dimensione della popolazione è infinita le frequenze alleliche non cambiano da una generazione alla successiva (HWE), ma se le dimensioni sono finite le frequenze alleliche fluttuano per via del campionamento
Selezione: cambiamenti direzionaliDeriva genetica: cambiamenti random

Quando 2N gameti vengono campionati da un pool di gameti infinito la probabilità Pi che quel campione contenga esattamente i alleli di tipo A1è data dalla funzione di probabilità binomiale
Pochè Pi è sempre > 0 per popolazioni in cui i due alleli coesistono (0 < p < 1), le frequenze alleliche possono cambiare di generazione in generazione senza l’intervento della selezione
Deriva genetica P(k) =nk
⎛
⎝ ⎜
⎞
⎠ ⎟ pk 1− p( )n − k Distribuzione binomiale
La distribuzione binomiale dà la probabilità di k successi in nprove
Deriva genetica
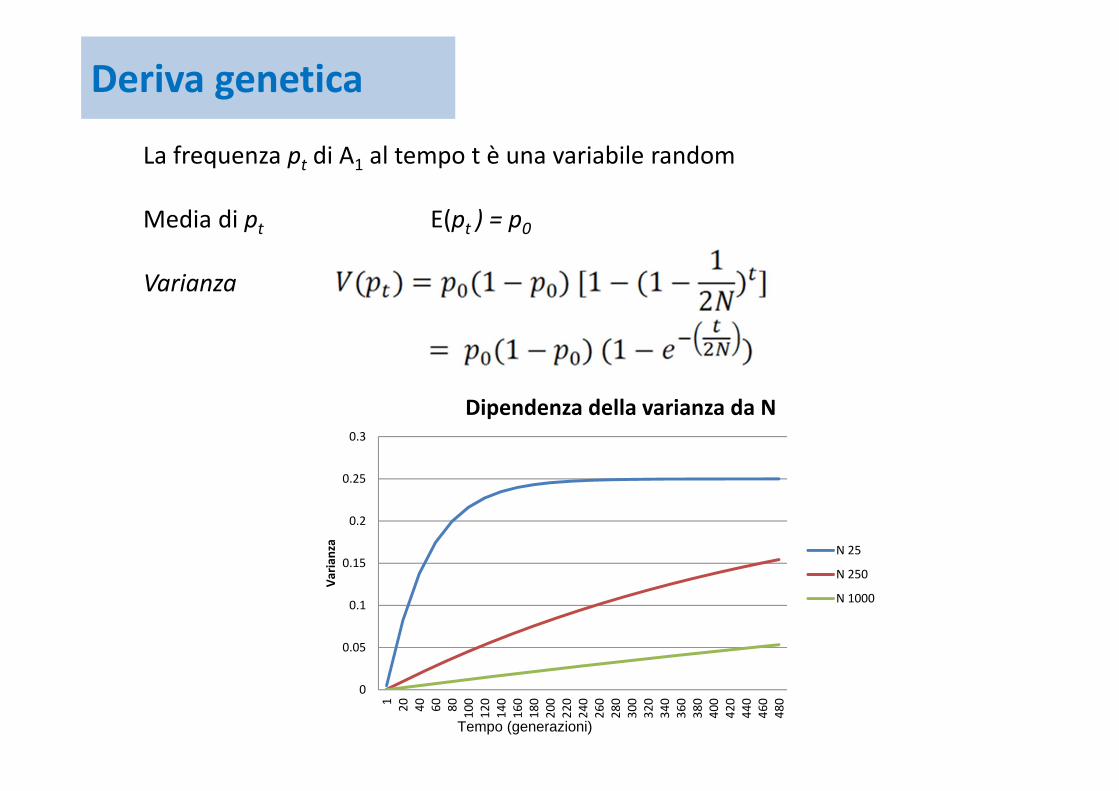
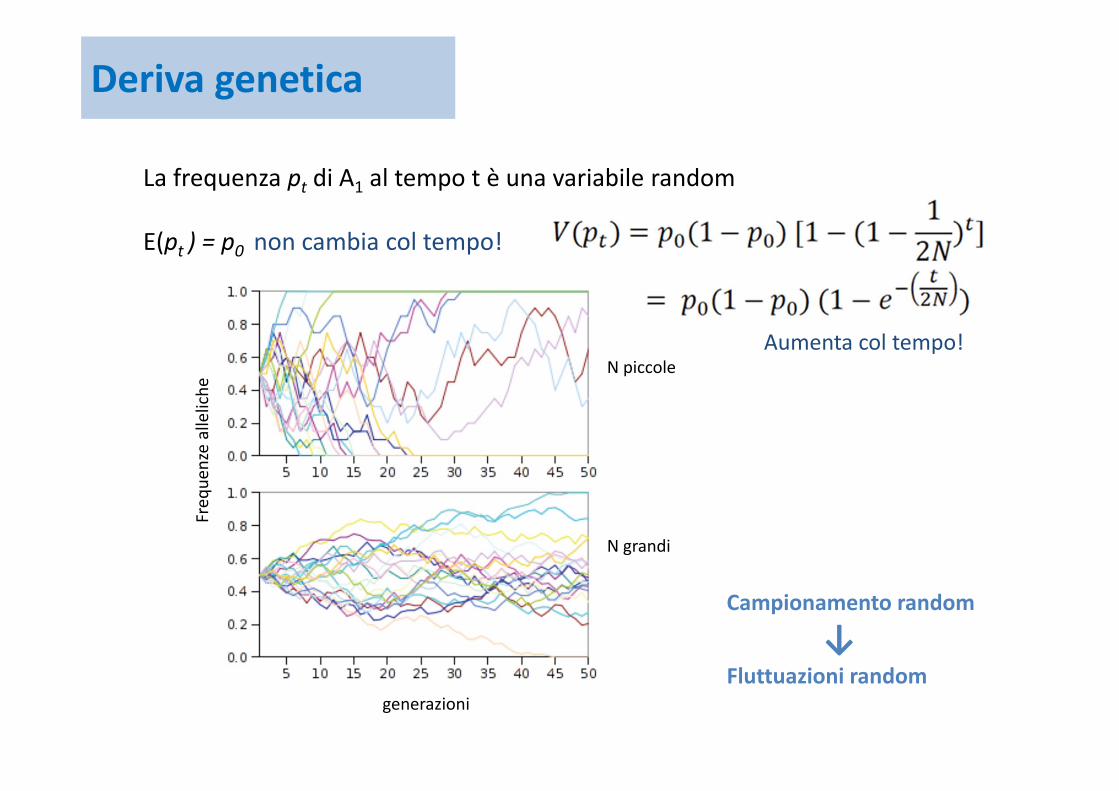
La frequenza pt di A1 al tempo t è una variabile random
Media di pt E(pt ) = p0
Varianza
0
0.05
0.1
0.15
0.2
0.25
0.3
1 20 40 60 80 100
120
140
160
180
200
220
240
260
280
300
320
340
360
380
400
420
440
460
480
Varia
nza
Dipendenza della varianza da N
N 25
N 250
N 1000
Tempo (generazioni)
Deriva genetica
Freq
uenze alleliche
generazioni
N piccole
N grandi
La frequenza pt di A1 al tempo t è una variabile random
E(pt ) = p0 non cambia col tempo!
Campionamento random
↓Fluttuazioni random
Aumenta col tempo!

Deriva genetica
Modello di Wright‐Fisher non realistico:
Sewall Wright (1931) introdusse il concetto di dimensione effettiva (Ne) delle popolazioni che definì come la dimensione di una popolazione ideale che avrebbe lo stesso effetto di campionamento casuale delle frequenze alleliche della popolazione reale

Consideriamo una popolazione con dimensione reali (census size) = Nin cui la frequenza dell’allele A1 alla generazione t = p.
Se il numero di individui che si riproduce = N, allora la varianza della frequenza dell’allele A1 nella generazione successiva, pt+1, si può ottenere dall’equazione della varianza
ponendo t = 1
Deriva genetica

Dato che non tutti gli individui nella popolazione prendono parte al processo riproduttivo la varianza è più grande di quella predetta da:
La dimensione effettiva della popolazione è il valore che sostituisce N per soddisfare l’equazione:
V( pt+1) =pt(1− pt )
2N
V( pt+1) =pt(1− pt )
2Ne
Deriva genetica

Deriva geneticaModello di Wright Fisher: non realistico
Esempi:
•le generazioni si sovrappongono → alcuni individui al tempo t non contribuiscono al pool genico della generazione successiva perché non fertili (non maturi, oltre il periodo fertile) DEVELPEMENTAL STRATIFICATION
Nei and Imazumi (1966): uomo Ne ≈ N/3
•Il numero di maschi e femmine che si riproducono non è uguale (es. specie poligame)
Behavioural ecology 2004

Deriva geneticaModello di Wright Fisher: non realistico
•Il numero di maschi e femmine che si riproducono non è uguale (es. specie poligame)
Se una popolazione ha Nm maschi e Nf femmine, Ne è data da:
SOLO se la sex ratio è di 1:1 (sex ratio “fisheriana” ) allora N = Ne
In ogni altro caso N > Ne

Qual è la dimesione effettiva di un alveare?
N ≈ 100000Nf = 1Nm >> 1Ne = 4
Deriva genetica

“the lemming cycle”Lemmings breed at regular intervals in such vast numbers as to cause mass migration, for instance, in Lapland. In Greenland, population cycle of the collared lemming is particularly regular, varying over 4‐year periods. According to recent research, the cyclic population dynamics in Greenland are explained by predator‐prey interaction, and not the biological attributes of lemmings or the limited food resources, as was previously assumed. (http://www.helsinki.fi/research/news/2003/week46.htm)
Anche variazoni nel lungo periodo possono rendere Ne < N
Esempi:Catastrofi ambientaliModelli di riproduzione ciclicaEventi di estinzione e ricolonizzazione di una regione geografica
Deriva genetica
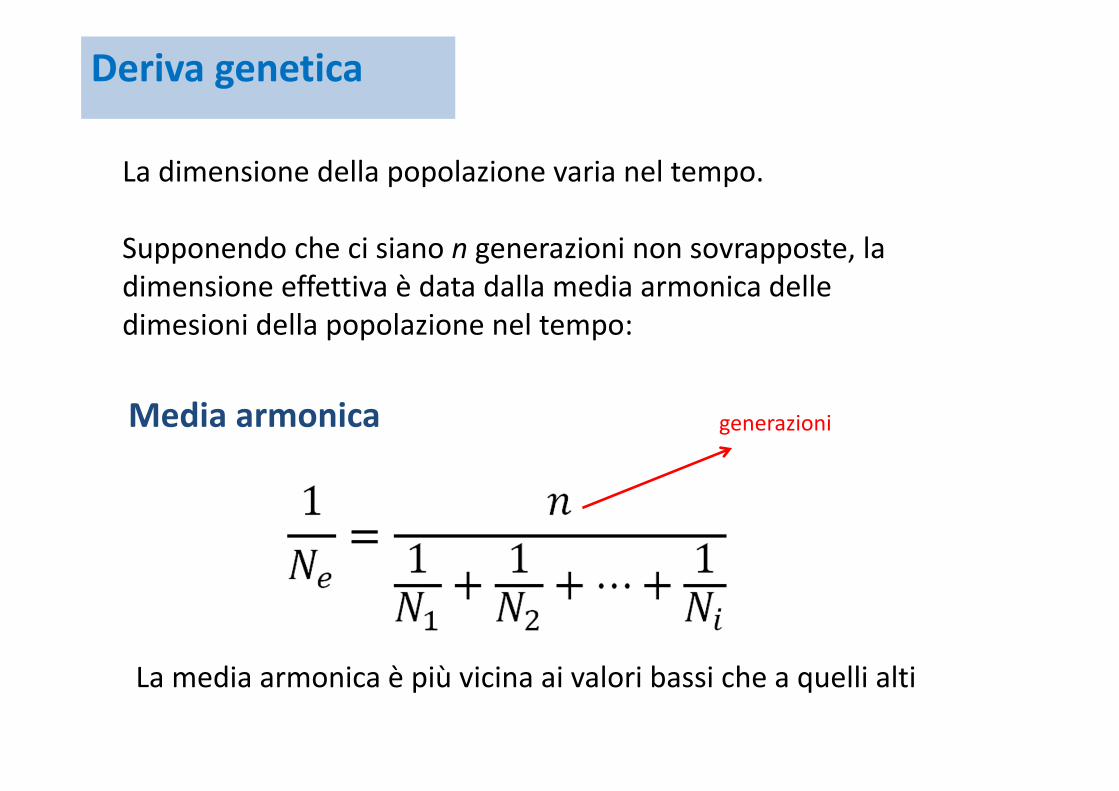
La dimensione della popolazione varia nel tempo.
Supponendo che ci siano n generazioni non sovrapposte, la dimensione effettiva è data dalla media armonica delle dimesioni della popolazione nel tempo:
Deriva genetica
generazioniMedia armonica
La media armonica è più vicina ai valori bassi che a quelli alti
Deriva genetica
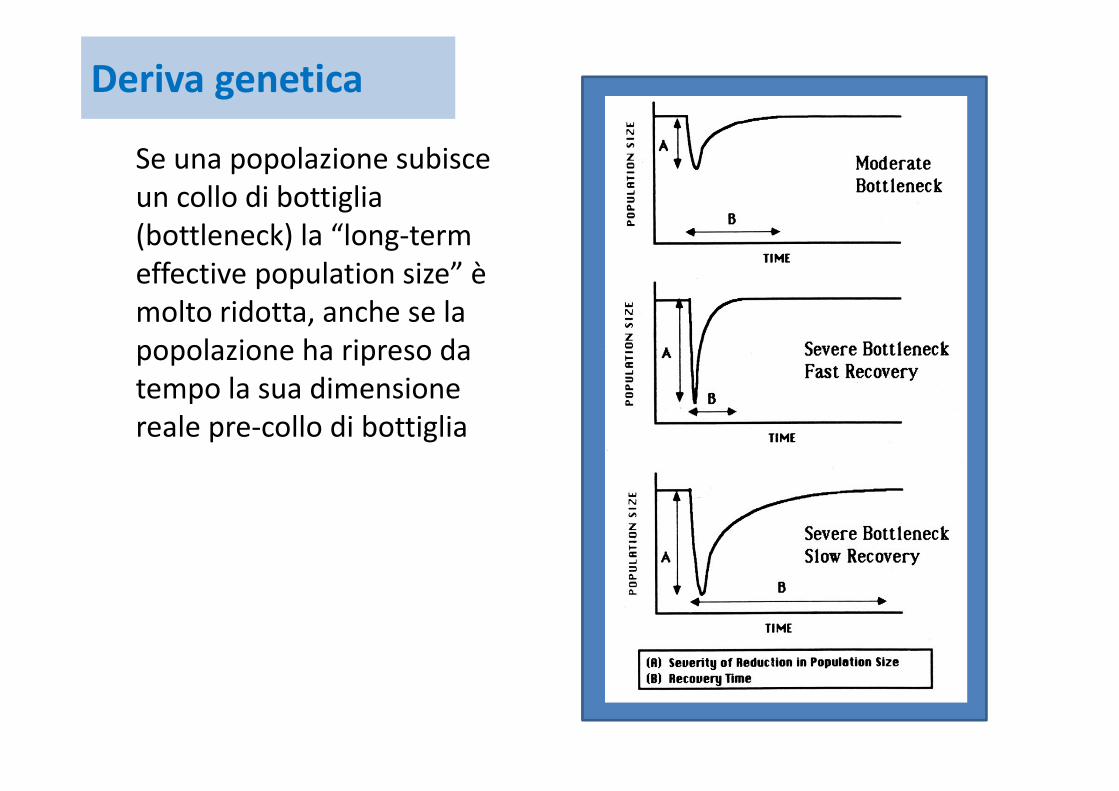
Se una popolazione subisce un collo di bottiglia (bottleneck) la “long‐termeffective population size” è molto ridotta, anche se la popolazione ha ripreso da tempo la sua dimensione reale pre‐collo di bottiglia
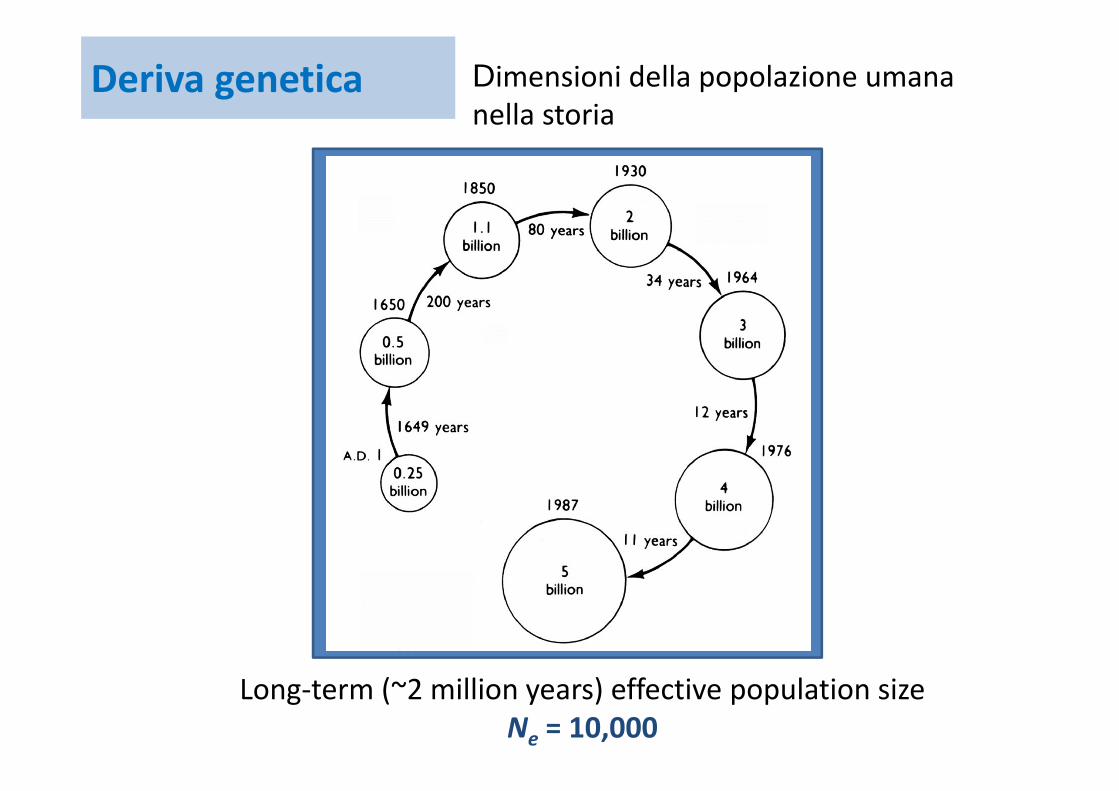
Deriva genetica Dimensioni della popolazione umana nella storia
Long‐term (~2 million years) effective population sizeNe = 10,000
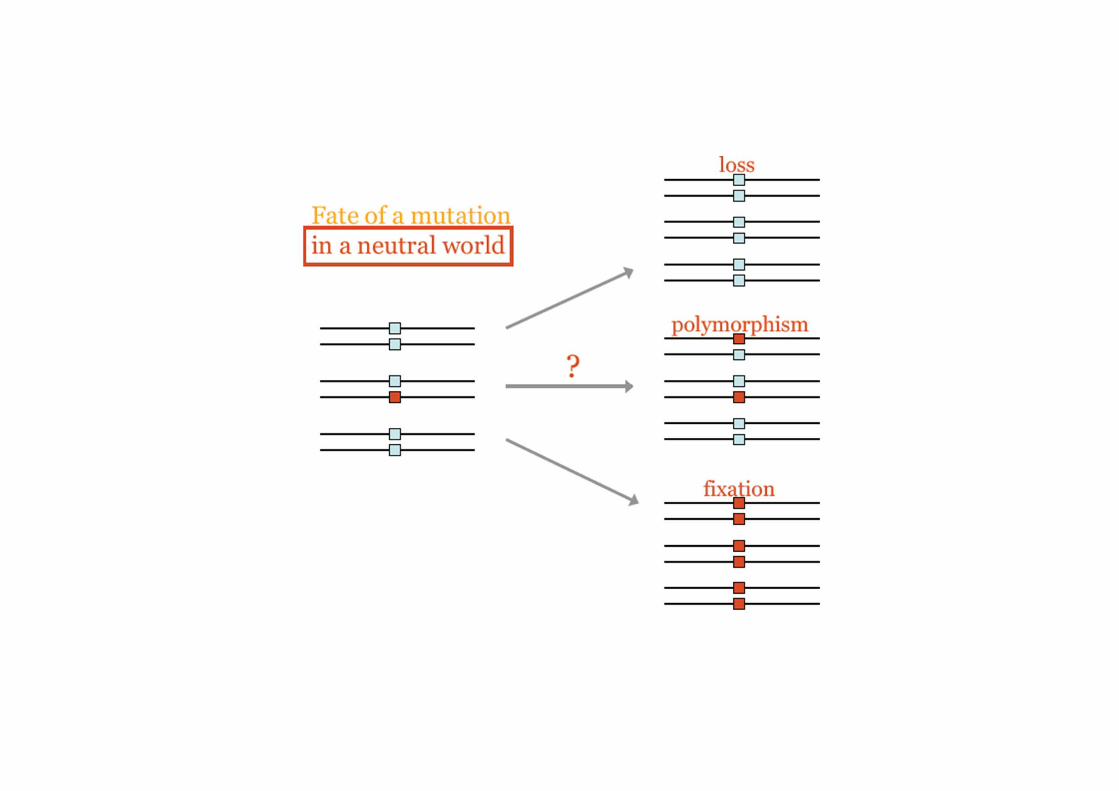
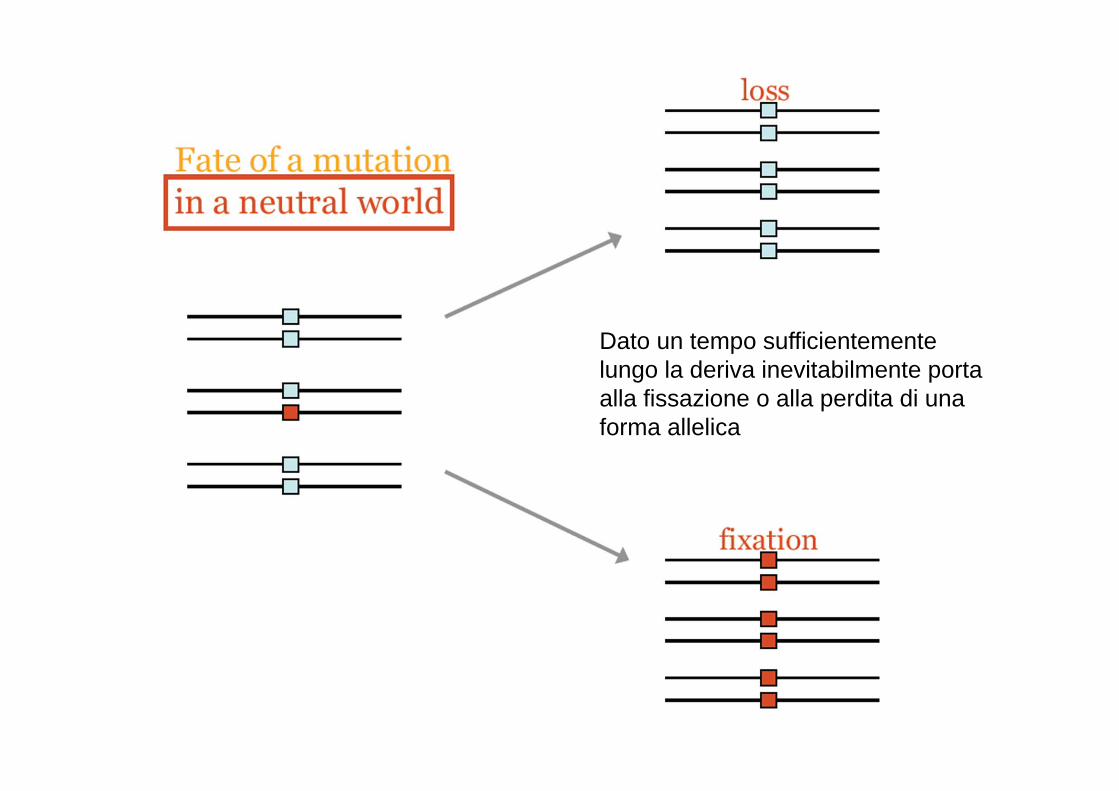
Dato un tempo sufficientemente lungo la deriva inevitabilmente porta alla fissazione o alla perdita di una forma allelica
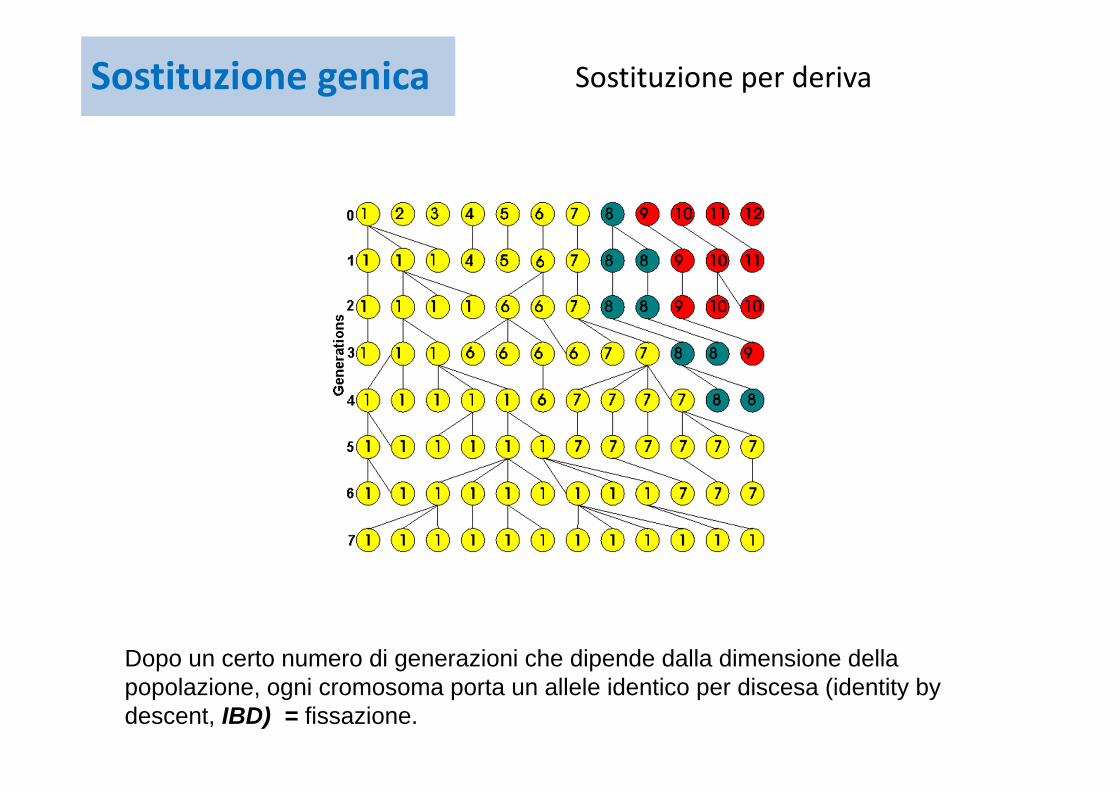
Dopo un certo numero di generazioni che dipende dalla dimensione della popolazione, ogni cromosoma porta un allele identico per discesa (identity by descent, IBD) = fissazione.
Sostituzione genica Sostituzione per deriva
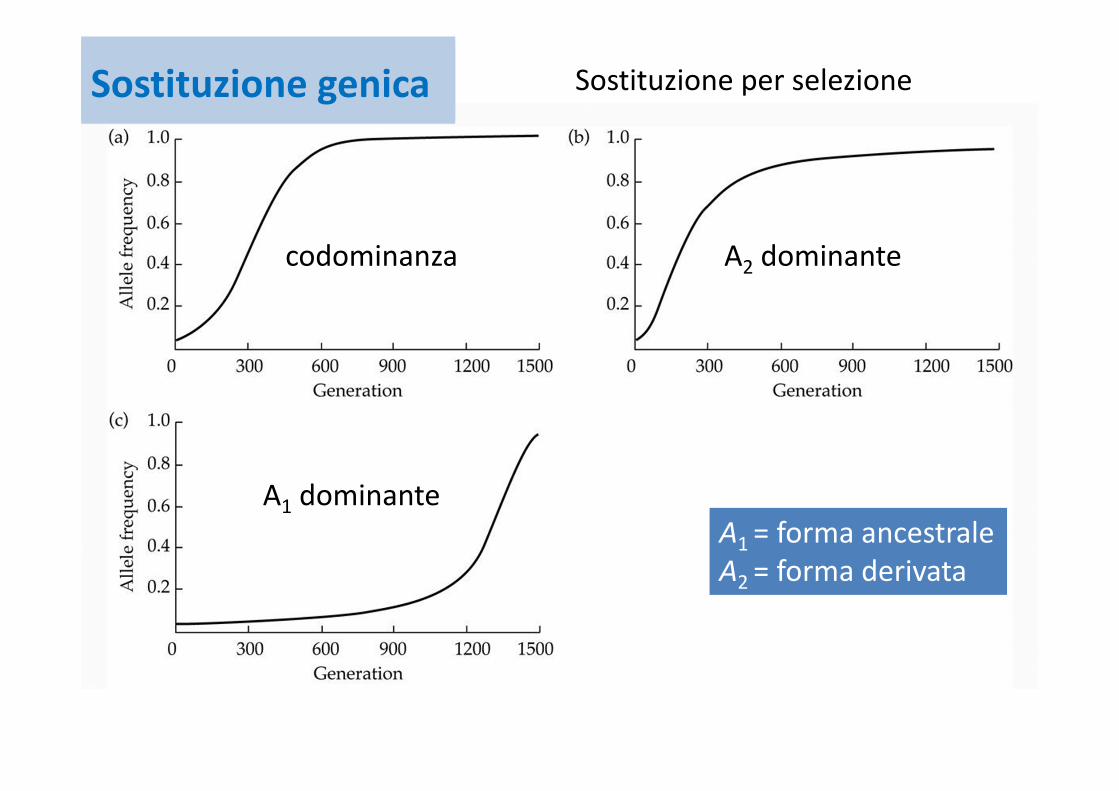
Sostituzione genica
A1 = forma ancestraleA2 = forma derivata
codominanza A2 dominante
A1 dominante
Sostituzione per selezioneSostituzione genica

Sostituzione genica
Processo per il quale un allele mutante sostituisce la forma piùcomune o “wild type”
Tempo di fissazioneMutante → Fissazione
Probabilità di fissazione
Tasso di sostituzione= numero di sostituzioni / tempo

Sostituzione genica
Processo per il quale un allele mutante sostituisce la forma piùcomune o “wild type”
Mutante → FissazioneProbabilità di fissazione
Da che cosa dipende?
1. Dalla frequenza iniziale q
2. Dal coefficiente di selezione s (selezione!)
3. Dalla dimensione effettiva Ne (deriva!!)

Sostituzione genica
Processo per il quale un allele mutante sostituisce la forma piùcomune o “wild type”
Mutante → FissazioneProbabilità di fissazione
Kimura (1962): la probabilità di fissazione di A2 è data da
Dimensione effettiva Coefficiente di selezione Frequenza iniziale di A2

Sostituzione genica
Mutante → FissazioneProbabilità di fissazione
quando x è piccolo e –x ≈ 1‐x
Se Nesq è piccolo, cioè quando s→0, l’equazione si riduce a
P ≈ qLa probabilità di fissazione di un allele neutrale è pari alla sua frequenza nella popolazione
Se s tende a 0: assenza di selezione = neutralità

Sostituzione genica
Mutante → FissazioneProbabilità di fissazione
Se N è la dimensione della popolazione, la frequenza iniziale di un nuovo allele sarà
q=1/(2N)
Sostituiamo q con 1/(2N)
Probabilità di fissazione di un nuovo allele quando s ≠ 0
Se s è ≠ 0: presenza di selezione

Sostituzione genica
Mutante → FissazioneProbabilità di fissazione
Se Ne è grande, quindi Ne → N
Probabilità di fissazione di un nuovo allele quando s è piccolo (ricorda: e –x ≈ 1‐x) ed Ne → N
Probabilità di fissazione di un nuovo allele quando s ≠ 0
Per valori positivi e piccoli di s e grandi valori di N

Sostituzione genica
Per valori positivi di s e grandi valori di N
Quindi se una mutazione vantaggiosa insorge in una popolazione grande e ha un vantaggio selettivo inferiore al 5% (s < 0.05, piccolo), la probabilità che si fissi è approssimativamente il doppio del suo vantaggio selettivo
Esempio: la probabilità di fissazione di una nuova mutazione codominante con s = 0.01 è 2%.

Sostituzione genica
Per valori positivi di s e grandi valori di N
Dan Graur
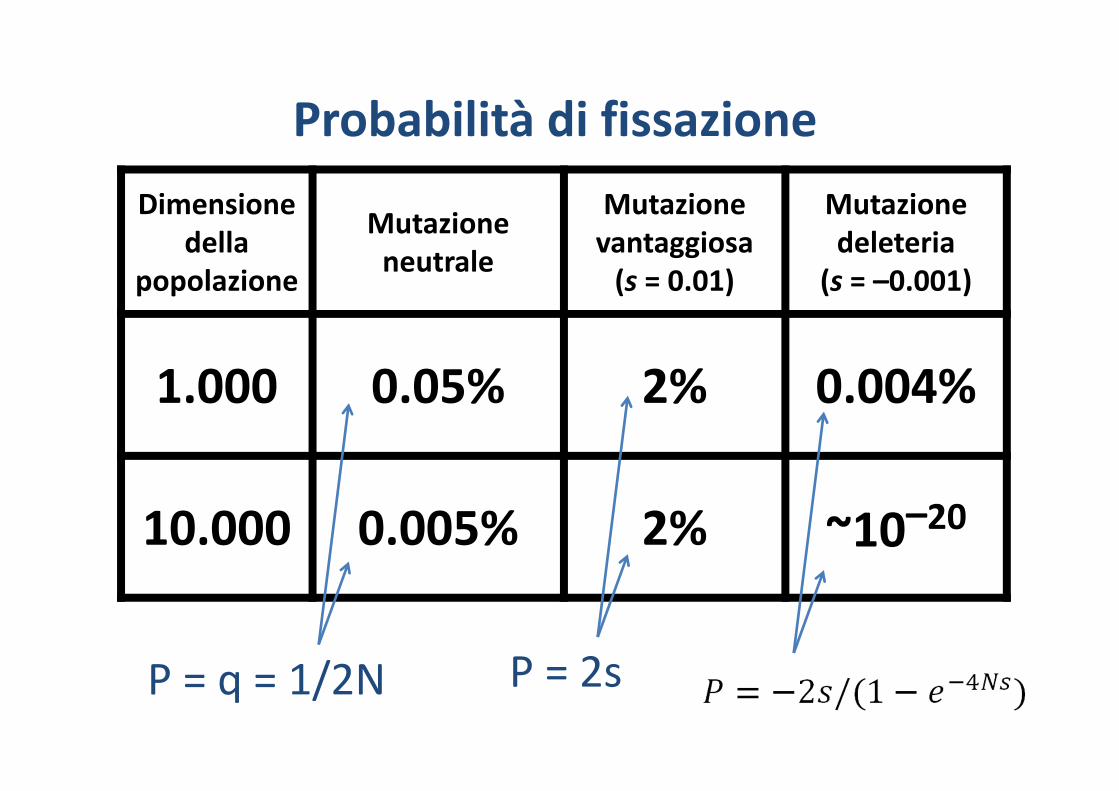
Dimensionedella
popolazione
Mutazioneneutrale
Mutazionevantaggiosa(s = 0.01)
Mutazionedeleteria
(s = –0.001)
1.000 0.05% 2% 0.004%
10.000 0.005% 2% ~10–20
Probabilità di fissazione
P = q = 1/2N P = 2s

Sostituzione genica
Tempo di fissazioneNel caso di un nuovo allele mutante neutrale (q=1/2N) il tempo di fissazione è stato calcolato da Kimura e Ohta (1969)
Se la mutazione ha un vantaggio selettivo di s si può approssimare a
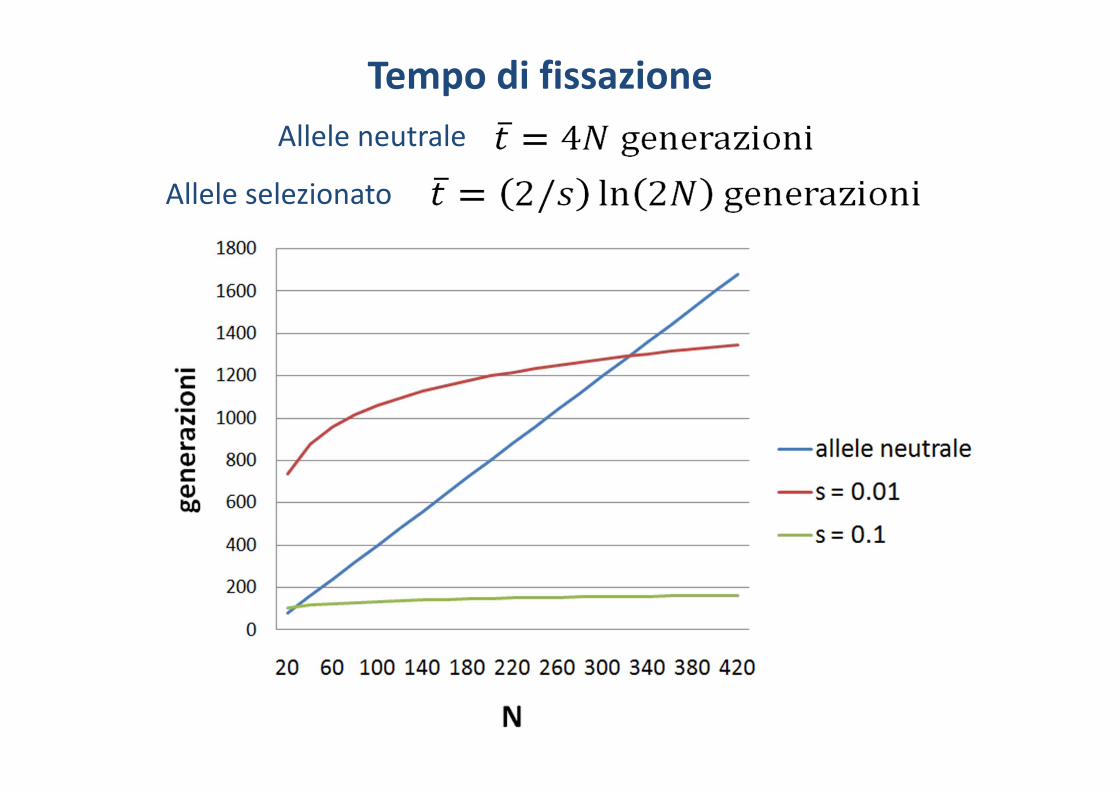
Allele neutrale
Allele selezionato
Tempo di fissazione
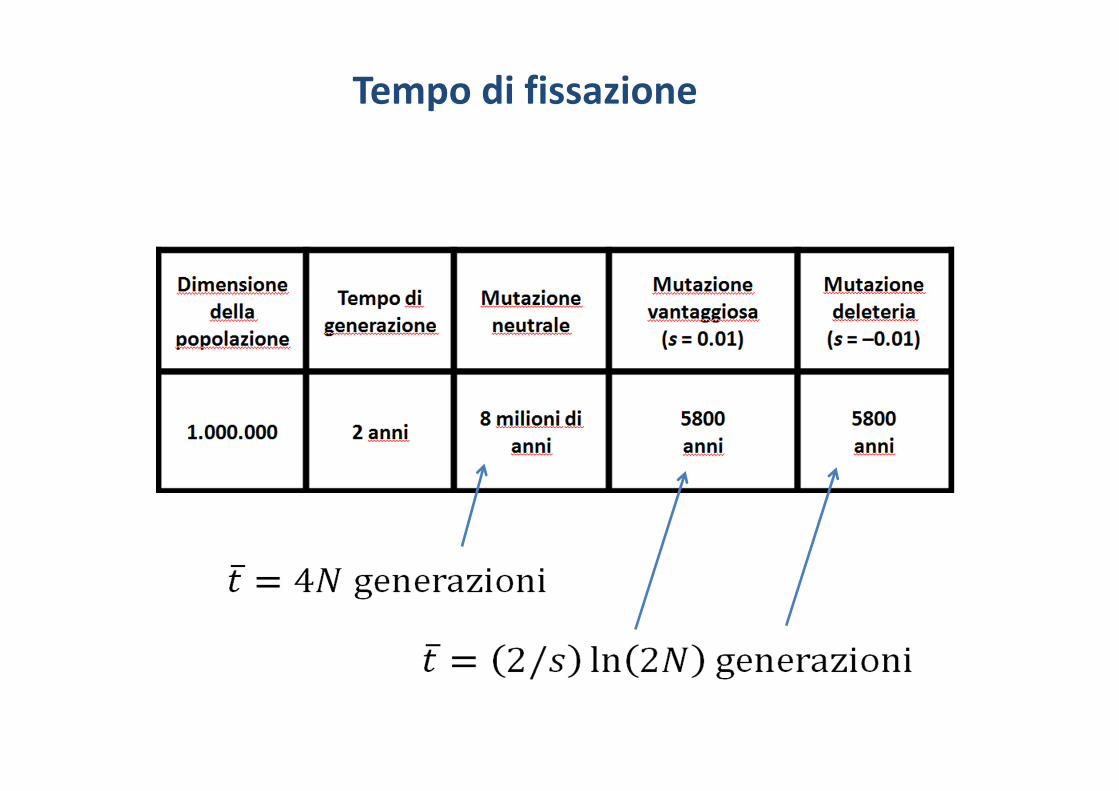
Tempo di fissazione

Sostituzione genica Tasso di sostituzioneNumero di mutanti che raggiungono la fissazione nell’unità di tempo
Mutazione neutraleMutazioni neutrali avvengono con un tasso
µ per gene per generazione
Il numero di mutanti per locus in una popolazione diploide didimensione N sarà
2N µ per generazione
La probabilità di fissazione di queste mutazioni è P= 1/(2N)Il tasso di sostituzione è K = 2N µ P
K = 2N µ (1/2N)= µ
Kimura 1968: Evolutionary rate at the molecular level, Nature

K = µ se la mutazione è neutrale
Sostituzione genica Tasso di sostituzione
N grande = molte mutazioni ma P= 1/(2N) quindi poche si fissano
N piccolo = poche mutazioni ma molte di queste si fissano
Il tasso di sostituzione è INDIPENDENTE da N!

Se la mutazione è sottoposta a selezione positiva il tasso di sostituzione si ottiene moltiplicando il numero di mutazioni per generazione, 2N µ per la probabilità di fissazione P = 2s
Il tasso di sostituzione è DIPENDENTE da N, s e µ
K = 4Nsµ
Sostituzione genica Tasso di sostituzione