Embed Size (px)
Citation preview

Hippocampus og psyken
Udarbejdet af:
Ane Søndergaard Thomsen
Aalborg Universitet
Institut for Kommunikation, Psykologi
Efterår 2005
Publiceret 2006 på hjemmesiden for
Klinisk organisations- og arbejdspsykologi
(KOAP)

Abstract
Rapporten er en analyse af, hvordan hjernestrukturen hippocampus bidrager til
hukommelsen og dermed psyken.
Hippocampus rolle i relation til det samlede hukommelsessystem ridses op for at
placere strukturen i sin psykologiske kontekst, men vægten er lagt på en analyse på
baggrund af neurovidenskabelig forskning.
Hippocampus er primært relateret til eksplicit langtidshukommelse. Strukturen er en
del af den mediale temporallaps hukommelsessystem.
Sanseinformationer omdannes til repræsentationer i neocortex. De sendes videre til
den rhinale cortex (de parahippocampale regioner), der integrerer informationer fra
de forskellige sansesystemer og sender resultatet videre til hippocampus.
Hippocampus er således en ”super konvergenszone”, idet den modtager input fra
flere konvergenszoner i den rhinale cortex. Derfor er den vigtig for den
domæneuafhængige hukommelse pga. sin integrative funktion.
I hippocampus er der et specielt vigtigt trisynaptisk kredsløb, der sender
informationer fra de rhinale områder til inputarealet i hippocampus (dentate gyrus),
derfra via CA3 og CA1 regionerne, sluttende i outputregionen (subiculum), der
sender informationen tilbage til den rhinale cortex. Derved lukkes løkken.
Hippocampus bidrager desuden til at regulere en amygdala-aktiveret stressreaktion.
Ved længerevarende eller intens stress mister hippocampus denne evnen til kontrol.
Forskellig reaktion på stressbelastning internt i hippocampus vidner om, at de
forskellige områder må have forskellig funktion. Atrofi og celledød finder
hovedsageligt sted i CA3 regionen, evnen til neurogenese i den dentate gyrus mistes
og i CA1 regionen vanskeliggøres LTP.
Hukommelsen er central for dannelse og vedligeholdelse af et individs selvbillede.
Uden hukommelse er vi desuden ude af stand til at reflektere, tage beslutninger,
indgå i sociale relationer o.l. Uden at kunne sætte hændelser i sammenhænge er det
vanskeligt at give dem mening. Hukommelsen og dermed hippocampus er således
uundværlig for den menneskelige psyke.

Indholdsfortegnelse
1. Indledning ................................................................................................................ 1
1.2. Problemafgrænsning/problemformulering........................................................ 1
1.3. Disposition og metode....................................................................................... 1
2. Placering af Hippocampus i sin teoriramme ............................................................ 2
2.1. Hukommelse ..................................................................................................... 2
3. Analyse og diskussion af hippocampus’ rolle.......................................................... 5
3.1. Hippocampus og hukommelsen ........................................................................ 5
3.2. Plasticitet er grundlaget for læring og hukommelse ......................................... 8
3.3. Emotionernes indflydelse på hukommelsen.................................................... 11
3.4. Hukommelsesproblemer ................................................................................. 12
3.4.1. Stress ........................................................................................................ 14
3.4.2. Andre almindelige situationer .................................................................. 17
3.5. Hvad betyder det så for psyken? ..................................................................... 18
4. Kritisk reflektion .................................................................................................... 19
4.1. Uklarheder ved teorierne................................................................................. 19
4.2. Forskningens empiriske grundlag ................................................................... 20
4.3. Glemsel ........................................................................................................... 20
4.4. Lokalisering af funktioner............................................................................... 21
5. Konklusion ............................................................................................................. 21
6. Perspektivering....................................................................................................... 22
7. Referenceliste......................................................................................................... 23
7.1. Bøger og redigerede værker ............................................................................ 23
7.2. Forskningsartikler ........................................................................................... 24
7.3. Andet materiale ............................................................................................... 25
8. Procesbeskrivelse ................................................................................................... 26
Appendiks A – grundlæggende neuropsykologi ........................................................ 27
Cellebiologi ............................................................................................................ 27
Molekylærbiologi................................................................................................... 28
Appendiks B – computeranalogi................................................................................ 30

Hippocampus og psyken Ane Søndergaard Thomsen
1
1. Indledning
Læring og hukommelse spiller en stor rolle i at vedligeholde en sammenhængende
personlighed.
De ting, vi er bevidste om vedrørende os selv, udgør de eksplicitte aspekter af selvet.
Resten er implicitte. Begge typer hukommelse er centrale mekanismer i dannelsen og
vedligeholdelsen af selvet. Det er i den forbindelse interessant at gå i dybden med,
hvilken rolle og hvordan hippocampus er involveret i hukommelsen og således er en
væsentlig medspiller i vores psyke.
Hukommelsen er vigtig for alt hvad vi foretager os. Uden den er vi ingenting. Viden
om diverse fakta og erfaringer hjælper os bl.a. definere os selv, tænke og tage
beslutninger.
1.2. Problemafgrænsning/problemformulering
”Hippocampus og psyken” er et bredt emne, hvorfor der foretages en afgrænsning
således, at rapporten behandler hippocampus’ rolle for hukommelsen.
1.3. Disposition og metode
Rapporten er bygget op således, at man bevæger sig fra det generelle mod det mere
specifikke og fra det raske mod dysfunktioner.
I kapitel 2 gennemgås hukommelsen først overordnet for at placere hippocampus i
sin teoretiske kontekst.
Kapitel 3 indeholder en analyse og diskussion af hippocampus og hukommelsen.
Først den overordnede struktur, dernæst et afsnit vedr. plasticitet som grundlag for
læring og hukommelse. Emotionernes indflydelse berøres i det omfang, det har
betydning for hippocampus.
Afsnittet om hukommelsesproblemer uddyber især problematikken omkring stress,
men enkelte andre psykiske tilstande nævnes, idet de kan eksemplificere, hvordan
Hvordan bidrager hippocampus til hukommelsen og dermed psyken?

Hippocampus og psyken Ane Søndergaard Thomsen
2
skader på hippocampus kan have indflydelse på psyken. Kapitlet slutter med at
opsamle hippocampus’ bidrag til psyken.
Kapitel 4 indeholder bl.a. en behandling af et par uklarheder ved især den beskrivelse
LeDoux fremlægger af hukommelsen, men som han deler med de øvrige kilder.
Derudover en diskussion af forskningens empiriske grundlag, glemslens mulige
adaptive funktioner o.lign.
Da den grundlæggende neuropsykologi ikke kan forudsættes bekendt er der lavet et
appendiks. Dette indeholder først et afsnit vedr. cellebiologi, der gennemgår et
neuron, dets forskellige dele og transmission mellem neuroner. Dernæst følger en
gennemgang af de molekylære processer.
Appendiks B indeholder en egenudviklet model for hippocampus’ integrative
funktion.
2. Placering af Hippocampus i sin teoriramme
Indledningsvis introduceres emnet hukommelse for at placere hippocampus i denne
teoriramme.
2.1. Hukommelse
Teorier om hukommelsen beskæftiger sig med såvel hukommelsessystemets struktur,
dvs. organisering af informationerne, som processerne, dvs. aktiviteterne.
Hukommelse involverer flere trin. Først præsentation af diverse indkomne stimuli og
kodning heraf, dernæst lagring af informationerne og endelig genkaldelse af
informationerne (Eysenck, 2000).
Tidligere opererede man med et simplificeret billede af hukommelsen, hvor den
indkomne stimulus først blev lagret i få sekunder i ”sanselagrene”. Disse blev
bombarderet med indtryk. De stimuli, opmærksomheden var rettet mod, kom i
korttidshukommelsen, der med sin begrænsede kapacitet var ustabil; den mindste
distraktion og informationen var væk. Ved repetition kunne det overflyttes til
langtidshukommelsen, der var ubegrænset både med hensyn til tid og plads.
Siden har det vist sig, at informationsstrømmen ikke er så simpel, der er fx adgang til
langtidshukommelsen allerede inden ting kommer i korttidshukommelsen aht.
genkendelse og repetition (ibid.).

Hippocampus og psyken Ane Søndergaard Thomsen
3
At der ér kvalitative forskelle mellem korttids- og langtidshukommelsen, og mellem
forskellige typer erindringer, har forskning vist ved dobbelt dissociation1, men i
stedet for korttidshukommelsen opereres nu med en arbejdshukommelse (ibid.). Den
er involveret i al tænkning og problemløsning.
I den model er der stadig specialiserede systemer involveret i behandling af
specifikke stimuli, der kan holde informationen i få sekunder, dvs. ”sanselagre”.
Selve arbejdshukommelsen består af et generelt system indeholdende et arbejdsareal
og et sæt mentale funktioner. Her integreres informationer på tværs af systemer, så
der kan skabes abstrakte repræsentationer:
Arbejdsarealet indeholder det, vi tænker på og er opmærksomme på lige nu. Det er
derfor helt afhængig af langtidshukommelsen mht. eksisterende viden og erfaringer.
Erindringer er en konstruktion på baggrund af lagret viden.
De mentale funktioner holder bl.a. arbejdsarealet opdateret, udvælger sig de ting
opmærksomheden skal rettes mod og henter relevant viden ind fra
langtidshukommelsen ved at finde specifikke erindringer eller aktivere mentale
skemata2 (LeDoux, 2002).
Sansesystem A
Langtidshukommelsen
(eksplicit) Sansesystem B
Figur 1: Arbejdshukommelsen (LeDoux, 2002, p. 179)
Baddeley opdeler sin beskrivelse af arbejdshukommelsen i tre komponenter og
integrerer derved sanselagrene i arbejdshukommelsen (Baddeley, 1974, 1986, 1990,
if. Eysenck, 2000):
1 Ved hukommelsestab kan nogen typer erindringer rammes uden at andre typer berøres og vice versa. 2 Arbejdshukommelsen er en funktion af kredsløb i frontallabben, specielt den præfrontale cortex. Den
præfrontale cortex er således en konvergenszone, der integrerer input fra mange systemer, herunder
hippocampus og de andre corticale områder involveret i eksplicit langtidshukommelse, se senere.
Perceptuel information om diverse stimuli og situationen integreres med tidligere lagrede
informationer om fakta og erfaringer samt den emotionelle signifikans af de pågældende stimuli. Der
er tilligemed forbindelse til de områder, der styrer motorikken (LeDoux, 2002).
Arbejds-
hukommelsen

Hippocampus og psyken Ane Søndergaard Thomsen
4
• En central udførende enhed.
• En fonologisk løkke, der indeholder information i en form baseret på tale. Dels et
lager forbundet med perception af tale, dels en artikulatorisk kontrolproces
knyttet til produktion af tale.
• En visuel rummelig notesblok.
Langtidshukommelsen kan opdeles på flere måder. Ofte deles den i to: en eksplicit
eller deklarativ hukommelse og en implicit eller non-deklarativ hukommelse.
Den eksplicitte hukommelse består af det, vi er i stand til at genkalde os bevidst og
beskrive verbalt. Det er såvel fakta (semantisk viden) som erfaringer, dvs.
informationer om specifikke begivenheder eller episoder i sin kontekst af tid og rum
(episodisk hukommelse).
Den implicitte viden (procedural hukommelse) reflekteres af måden, vi handler på.
Det er erfaringer, der letter udførelsen af opgaver uden bevidst genkaldelse. ”Øvelse
gør mester”, som man siger, men uden at individet kan forklare hvorfor eller
hvordan. Erindringerne er et middel til at udføre funktionen, fx at kunne holde
balancen på cykel, det er ikke egentlige hukommelsessystemer. Det omfatter
motoriske færdigheder samt læring ved priming og betingning3 (Eysenck, 2000;
LeDoux, 1999, 2002).
Forskellen skyldes, at der er forskellige hjernesystemer involveret i de to former for
hukommelse. PET scanninger har bl.a. vist, at hippocampus, er meget aktiv under
eksplicitte hukommelsestests, mens implicit viden ikke kræver øget aktivitet i
hippocampus (Eysenck, 2000).
Arbejdshukommelsen påvirker den eksplicitte langtidshukommelse, idet
opmærksomheds og perceptuelle processer på lærings tidspunktet bestemmer,
hvilken information, der lagres (LeDoux, 2002). Hvor meget der huskes afhænger
ikke af, hvorvidt det er individets hensigt at skulle lære det eller ej. Jo mere man
analyserer og behandler en given stimulus i dybden4, og jo mere særegen oplevelsen
er5, des mere omfattende og længerevarende hukommelsesspor dannes (Eysenck,
2000).
3 (conditioning) Ved klassisk betingning lærer individet at associere to stimuli. Ved operand eller
instrumentel betingning er det mellem stimulus eller belønning og en adfærd. 4 Der dannes flere associationer og forbindelser (fx ved repetition). 5 Der er færre irrelevante informationer.

Hippocampus og psyken Ane Søndergaard Thomsen
5
Vi lagrer oplysninger om såvel den ting, vi skal huske, og den kontekst, situationen
finder sted i. Hukommelsen er bedst, når der er overensstemmelse mellem
betingelserne under indkodning og genfinding (ibid.).
Der findes ikke ét samlet system eller lager til langtidshukommelse. Som beskrevet
har hippocampus stor indflydelse på den eksplicitte hukommelse, men ikke på den
implicitte hukommelse. Det hænger bl.a. sammen med at priming, betingning og
motoriske færdigheder ikke kræver kontekst information eller bevidst genkaldelse.
Den implicitte viden er derfor som regel tilgængelig under hukommelsestab. De to
former for langtidshukommelse er således forskellige dokumenteret ved dobbelt
dissociation6 (ibid.).
Hippocampus er således primært relateret til eksplicit langtidshukommelse.
3. Analyse og diskussion af hippocampus’ rolle
Efter at have placeret hippocampus inden for teorier om hukommelse analyseres og
diskuteres i dette kapitel hippocampus’ rolle for hukommelsen og psyken.
3.1. Hippocampus og hukommelsen
Psykologiske teorier og praksis bør funderes i neurobiologisk viden, da psykologiske
processer er baseret på hjerneprocesser7, således også hukommelsen. Dette afsnit vil
behandle hippocampus og dennes rolle for hukommelsen nærmere.
Hippocampus sættes primært i forbindelse med lagring af eksplicit og rummelig
viden i langtidshukommelsen samt integration af stimuli i relation til en oplevelse
(Kalat, 2004). Forskning dokumenterer, at blodgennemstrømningen i hippocampus er
høj ved eksplicitte hukommelsesopgaver. Dette indikerer, at hippocampus er meget
aktiv og at hippocampus således spiller en vigtig rolle (Eysenck, 2000).
Navnet hippocampus kommer af det græske ord hipokampus, der betyder søuhyre,
fordi strukturen skulle ligne en søhest (LeDoux, 2002). Der er to, én i hver
hemisfære.
6 J.fr. tidligere. 7 Et emne, der vil blive uddybet under perspektiveringen.

Hippocampus og psyken Ane Søndergaard Thomsen
6
Hippocampus og den rhinale cortex (de parahippocampale regioner) udgør den
mediale temporallaps hukommelsessystem, der er dybt involveret i eksplicit
hukommelse, jfr. figur 2.
Neocortex
Mediale
temporallaps
hukommelsessystem
Figur 2: Den mediale temporallaps hukommelsessystem (LeDoux, 2002, p. 104)8
Informationer om livets erfaringer modtages i hjernen gennem sanserne og
sansesystemerne i neocortex, hvor der skabes domænespecifikke9
sanserepræsentationer, der også trækker på forudgående erindringer.
De sendes herfra til den rhinale cortex (de parahippocampale regioner). Den rhinale
cortex er konvergenszoner, der integrerer informationer fra de forskellige
sansesystemer og skaber en samlet repræsentation uafhængig af de konkrete stimuli,
inden de sendes videre til hippocampus.
I hippocampus er der et specielt vigtigt trisynaptisk kredsløb10, der sender
informationer fra de rhinale områder til inputarealet i hippocampus (dentate gyrus),
8 ”CA” i ”CA1” og ”CA3” står for ”cornu ammonis”. 9 Dedikeret til behandling af specifikke stimuli. 10 Dette kredsløb påvirkes kraftigt under visse psykiske tilstande, hvilket der vil blive fulgt op på
senere.
Rhinale
cortex
Hippocampus
Somatosensoriske
cortex
Auditive cortex Visuelle cortex
Perirhinale
Parahippocampale
Entorhinale
Dentate gyrus
CA3
CA1
Subiculum

Hippocampus og psyken Ane Søndergaard Thomsen
7
derfra via CA3 og CA1 regionerne, sluttende i outputregionen (subiculum), der
sender informationen tilbage til den rhinale cortex. Derved lukkes løkken11.
Noget tyder på, at hvert område i hippocampus’ kredsløb har sin egen funktion.
Forsøg viser fx at skader på CA3-regionen hæmmer tilegnelse og konsolidering af
rummelige erindringer, men ikke genkaldelse.
CA1 aktiveres direkte under genkaldelse. Dog inddrages CA3 i genkaldelsen af
erindringen, hvis den rummelige kontekst er ændret, i forbindelse med reorganisering
af den rummelige repræsentation (Florian et al., 2004).
Regionerne i hippocampus rammes desuden forskelligt, fx under stress, hvilket
uddybes i afsnit 3.4.1.
Hippocampus er en ”super konvergenszone”, idet den modtager input fra flere
konvergenszoner i den rhinale cortex. Derfor er den vigtig for den
domæneuafhængige hukommelse pga. sin integrative funktion.
Scanninger viser således, at der er mere aktivitet i hippocampus ved opgaver, der
kræver integration af informationer; det være sig perceptuelle, semantiske,
tidsmæssige og rummelige (Eysenck, 2000)12.
Hippocampus associerer bl.a. eksterne hukommelsesmarkører (fra sanser) med
interne repræsentationer af kontekst og genopretter kontekst, når der er blevet
uoverensstemmelse (Redish, 2001).
Forbindelsen mellem hippocampus og neocortex går begge veje. Informationerne
kan fra den rhinale cortex sendes tilbage til hippocampus eller neocortex til videre
behandling eller lagring. De corticale områder, der modtog en stimulus, deltager
således i lagringen af samme (LeDoux, 2002).
Som nævnt spiller hippocampus forskellige roller for hukommelsen. Strukturen er
nødvendig for lagring af nye eksplicitte erindringer, men over tid aftager dens rolle i
relation til den enkelte erindring.
Ifølge LeDoux (2002) mener de fleste forskere, at de eksplicitte erindringer gemmes
i neocortex, hvor behandlingen af den indkomne stimulus blev indledt, og at det er
hippocampus, der dirigerer lagringsprocessen:
11 At gemme nye episodiske erindringer afhænger primært af hippocampus, for at kunne sætte
informationen i en rummelig og tidsmæssig kontekst, mens forskning tyder på at semantisk viden
primært afhænger af de corticale dele af den mediale tindingelaps hukommelsessystem. 12 Også den integrerende funktion vendes der tilbage til.

Hippocampus og psyken Ane Søndergaard Thomsen
8
Først gemmes erindringen i hippocampus13. Når dele af erindringen gentager sig,
deltager hippocampus i genindsættelsen af det mønster af cortical aktivering, som
fandt sted første gang. Hver gang ændres synapserne i cortex en smule. Langsomt
opbygges erindringen som en repræsentation i cortex og til sidst bliver den
selvtilstrækkelig og uafhængig af hippocampus14.
Hukommelseskonsolidering sker bl.a. når vi sover. Forskning tyder på, at de neurale
mønstre gentages som ved playback i hippocampus under søvn (LeDoux, 2002).
Hippocampus og dele af cortex (bl.a. den visuelle cortex) er lige så aktive under
REM søvn som i vågen tilstand, i modsætning til den præfrontale cortex involveret i
arbejdshukommelsen. REM søvn er således også overrepræsenteret tidligt i livet,
hvor vi udvikler os hurtigst (Hobson, 1999).
Fordi hver genindsættelse af erindringen afhænger af hippocampus påvirker en
bilateral skade på hippocampus evnen til efterfølgende at danne nye erindringer. Til
gengæld påvirkes gamle erindringer, der blev konsolideret i cortex før skaden opstod,
ikke. Gamle erindringer er resultatet af akkumuleringen af synaptiske ændringer i
cortex som et resultat af mange genindsættelser af erindringen.
Konsolideringen er en gradvis proces. Det tager dage eller uger (Kalat, 2004).
3.2. Plasticitet er grundlaget for læring og hukommelse
For at komme nærmere, hvordan erindringer lagres, som grundlag for en diskussion
af skader på hippocampus, vil følgende afsnit gå mere i detaljer med det
neurobiologiske.
De fleste systemer i hjernen er plastiske, dvs. kan modificeres med erfaring; de
involverede synapser ændres15. Plasticitet er vigtig for hippocampus’ funktion.
Essensen af, hvem vi er, reflekterer mønstre af neurale netværk i vores hjerner.
Hjernen fungerer ved synaptisk transmission mellem neuroner og ved at trække på
information lagret ved tidligere synaptiske transmissioner (LeDoux, 2002).
13 Dette vendes der tilbage til i den senere diskussion af teorien i afsnit 4.1.
14 Hvis erindringerne gemmes i cortex, hvor den oprindelige stimulus blev behandlet, mangler den
integrerende enhed, hvis hippocampus skades. Nogle forskere mener, evnen til at samle
repræsentationerne læres af neocortex langsomt, hvorfor det ikke er strukturens normale rolle, men
den kan overtage funktionen ved skade på hippocampus. 15 Jfr. Appendiks A – grundlæggende neuropsykologi.

Hippocampus og psyken Ane Søndergaard Thomsen
9
Vores evne til at huske, begære lykke mm. involverer stimulerende synaptisk
transmission af glutamat reguleret af hæmmende GABA og moduleret af peptider,
aminer og hormoner, jfr. Appendiks A - grundlæggende neuropsykologi.
Når glutamat frigives fra den præsynaptiske terminal til synapsen binder den sig på
forskellige receptorer på den postsynaptiske dendrit. Det er AMPA-receptoren, der er
involveret i den almindelige synaptiske transmission og således åbner for
indstrømning af positive ioner i det postsynaptiske neuron, der i sidste ende kan
betyde aktivering af et aktionspotentiale, der sender impulsen videre.
Ud over således at lede en impulsen videre skal de neurale netværk også kunne lære
og huske nyt.
Hebb mente, at hvis to neuroner er aktive samtidigt, den ene præsynaptisk den anden
postsynaptisk, ville forbindelsen mellem dem styrkes. Hans ”fire and wire” teori
forklarer læring, hukommelse, udvikling af synapser (Hebb, 1949, if. LeDoux, 2002).
Det frigivne glutamat fra de præsynaptiske terminaler binder sig til flere slags
postsynaptiske receptorer. Den postsynaptiske celle skaber et link hjulpet af
NMDA16 receptoren.
NMDA receptoren er i cellens hviletilstand blokeret af magnesium, Mg++. Når det
postsynaptiske neuron er aktiveret af almindelig synaptisk transmission og har
initieret et aktionspotentiale, depolariseres cellen og denne Mg frigives. Glutamat
kan da også binde sig til NMDA receptoren og åbner derved for indsivning af
calcium, Ca++17
, til den postsynaptiske celle. Det resulterer i LTP18:
Ca aktiverer en reaktion med proteinkinaser. Disse aktiverer proteiner.
I tidlig LTP (i de tidlige faser) aktiverer kinaserne eksisterende proteiner i cellen,
bl.a. AMPA receptor proteiner. Det bevirker at den samme mængde glutamat frigivet
fra den præsynaptiske terminal næste gang vil have flere receptorer i den
postsynaptiske celle at binde sig til, hvorfor det vil resultere i en større postsynaptisk
reaktion (LeDoux, 2002).
16 N-methyl-D-aspartate (Garcia, 2001).
17 Neurotransmittere som glutamat er førstehånds budbringere; de signaler mellem neuroner.
Andenhånds budbringere, som Ca, initierer kemiske reaktioner inde i cellen på baggrund af
information modtaget udefra under transmission af førstehåndsbudbringerne (LeDoux, 2002). 18 LTP står for ”Long-term potentiation”. Ved kortvarigt at stimulere nervefibre elektrisk ses at den
postsynaptiske reaktion i hippocampus bliver større og virkningen holder i flere timer. Man kan
således simulere læring og plasticitet.

Hippocampus og psyken Ane Søndergaard Thomsen
10
I ”sen LTP”, hvor erindringerne gøres langtidsholdbare, aktiverer Ca en række
kinaser19, der igen aktiverer molekylære processer i cellekernen, herunder aktivering
af proteinet CREB20. CREB aktiverer gener ansvarlig for proteinsyntese. Proteinerne
sendes ud i cellen, hvor de bruges af de aktive synapser. Denne proteinsyntese er en
forudsætning for at det lærte overføres til langtidshukommelse21, idet den styrker de
synaptiske forbindelser. Ligeledes når erindringerne skal konsolideres i forbindelse
med, at vi finder dem frem og gemmer dem igen. Derved opdateres repræsentationen
og tilpasses de neurale netværk, som de ser ud på det tidspunkt, dvs. der tages højde
for viden og erfaringer, der er kommet til efterfølgende (LeDoux, 1999, 2002;
McGaugh, 2000).
Alle den postsynaptiske celles NMDA receptorer depolariseres, dvs. alle aktive
synapser forstærkes på den beskrevne måde, også de der selv var for svage til at
aktivere et aktionspotentiale, hvilket muliggør associativ læring. Forskning viser, at
LTP involverer associative interaktioner mellem postsynaptiske neuroner og
specifikke præsynaptiske input. Dvs. den samme postsynaptiske celle kan deltage i
lagring af informationer vedrørende forskellige erfaringer sålænge det er forskellige
synapser, der er involveret (LeDoux, 2002).
Når aktionspotentialet aktiveres i den postsynaptiske celle frigives neurotrophiner
tilbage til synapsen. Disse optages af presynaptiske terminaler, der netop er aktive,
idet de i forvejen genoptager frigivne molekyler fra synapsen. Neurotrophiner
fremmer overlevelse og vækst af neuroner22. Der sker således en forgrening af det
presynaptiske axon og dannelse af nye synapser. Men kun de aktive terminaler får de
livgivende neurotrophiner. Aktivitet fører til øget synaptisk kompleksitet og celledød
forhindres.
Aktive forbindelser forstærkes således, mens passive går til grunde. Synaptogenese,
eller skabelsen af nye synapser mellem eksisterende neuroner, sker kontinuerligt livet
igennem (LeDoux, 2002).
Plasticitet eller LTP er meget vigtig, hvilket vil fremgå i forbindelse med senere
problematikker i relation til hukommelsen.
19 Cyklisk AMP (cAMP) –> proteinkinase A (PKA), CaMK og MAPK.
20 cAMP response element-binding protein.
21 Blokeres der for NDMA synapserne, blokeres dannelsen af LTP og dermed konsolidering af
hukommelsen. 22 BDNF er et eksempel. Det sænker aktiviteten af Bad (jfr. beskrivelsen af atrofi under stress) og
stimulerer produktion af Bcl-2, hvilket stimulerer mitochondriernes membran.

Hippocampus og psyken Ane Søndergaard Thomsen
11
3.3. Emotionernes indflydelse på hukommelsen
Det mest undersøgte emotionelle23 system er frygt.
Amygdala og hippocampus regulerer frygt på baggrund af vurderingen af
situationen. Amygdala24 er meget involveret i den direkte frygtbetingning, dvs.
reagerer på stimuli direkte fra thalamus (the low road)25 og sætter kroppen i
alarmberedskab endnu inden cortex har registret faren (the high road) (LeDoux,
1999, 2002). Hippocampus bidrager især ved den kontekstuelle betingning. Det
mediale temporallaps hukommelsessystem informerer amygdala om den kontekst,
den emotionelle læring finder sted i. Der er forbindelser fra såvel den rhinale cortex
som subiculum (LeDoux, 2002).
Amygdala er fuldt funktionsdygtig fra fødslen, så pga. hippocampus’ senere
modning mangler den hæmmende indflydelse, hvorfor frygt (og andre emotioner) må
opleves overvældende i den tidlige barndom (Cozolino, 2002).
Frygtbetingning er som nævnt implicit læring, men alle oplevelser og følelser, vi er
bevidste om, involverer arbejdshukommelsen og hvis oplevelsen er signifikant, vil
den blive lagret i det eksplicitte hukommelsessystem.
Følelsen integreres med informationer om den aktuelle stimulus,
langtidshukommelsens erindring om den pågældende stimulus fra hippocampus og
det emotionelle bidrag fra amygdala (LeDoux, 1999). Amygdala transformerer
således en kognitiv perception til en emotionel oplevelse. Amygdala kan sende
informationer til alle stadier i behandlingen af sanseindtryk, også den tidlige
behandling i neocortex.
Som beskrevet i afsnittet om hippocampus og hukommelsen er neocortex afgørende
involveret i aktiveringen af det mediale temporallaps hukommelsessystem. Ved at
påvirke neocortex påvirker amygdala således langtidshukommelsen26.
Eksplicitte erindringer dannet i emotionelle situationer huskes ofte meget tydeligt.
De kaldes ”flashbulb”27 erindringer (LeDoux, 2002; Sapolsky, 2003).
23 En emotion fører til fysiologiske ændringer. Emotioner er ubevidste processer, der kan, men ikke
nødvendigvis, fører til bevidste oplevelser. 24 Amygdala handler ikke bare om frygt.
25 Se senere vedr. stress.
26 Amygdala kan også påvirke den rhinale cortex direkte (LeDoux, 2002).
27 Det betyder blitzpære på dansk, men det engelske udtryk anvendes.

Hippocampus og psyken Ane Søndergaard Thomsen
12
I emotionelle situationer aktiverer amygdala frigivelsen af hormoner, fx cortisol fra
binyrerne, der returnerer til hjernen med blodbanen. Gennem forbindelsen til
hippocampus og andre regioner involveret i det eksplicitte hukommelsessystem,
styrker amygdala konsolideringen af erindringernes dannelse. De genkaldes derved
lettere og klarere. Emotionelle ting huskes bedre end neutrale (Kuhlmann et al.,
2005; McGaugh, 2000; Roozendaal, 2002).
Som beskrevet under LTP er ny proteinsyntese en forudsætning for gøre en erindring
langtidsholdbar. Det blev initieret af CREB, jfr. ovenfor. Der findes også en gen-
repressor, CREB-2. I nogle situationer er vi så emotionelt tændte, at vi kan huske alt
fra det øjeblik. Det skyldes bl.a. at gen-repressoren fjernes og proteinsyntesen lettere
går i gang (LeDoux, 1999).
At emotionel ophidselse forstærker erindringen er godt i nogle situationer, mindre
heldigt i andre, idet traumatiske oplevelser er svære at slippe igen.
Ved genkaldelsen samarbejder hippocampus og amygdala således, at emotionelle
erindringer (formidlet af amygdala) og erindringer om emotioner (formidlet af
hippocampus) integreres i én bevidst oplevelse (LeDoux, 1999).
Erindringer genkaldes lettere, hvis den emotionelle tilstand på
genkaldelsestidspunktet svarer til tilstanden, da erindringerne blev dannet; muligvis
fordi amygdala ved genkaldelsen forsøger at genskabe den oprindelige emotionelle
tilstand (LeDoux, 2002).
Til gengæld husker man dårligere under overproduktion af cortisol (Kuhlmann et al.,
2005; Roozendaal, 2002), hvilket uddybes i næste afsnit.
3.4. Hukommelsesproblemer
Der kan være mange årsager til hukommelsesproblemer.
Ingen har eksplicitte erindringer fra de første leveår. Det kognitive selv dannes først
fra omkring 2-års alderen og selvgenkendelsen bliver et centrum, hvorom erindringer
kan organiseres (Eysenck, 2000). Det hænger sammen med at hippocampus og
cortex modnes sent og de neurale netværk forbundet med eksplicit hukommelse
udvikles gradvist (Cozolino, 2002).
Rapporten har allerede været inde på, at kontekst betyder noget for genkaldelsen.
Man har lettere ved at huske ting, der er indlært under ca. samme forhold og under
samme følelsesmæssige tilstand som på genkaldelsestidspunktet. Tid ændrer

Hippocampus og psyken Ane Søndergaard Thomsen
13
kontekst, hvorfor det bliver sværere at huske efterhånden som tiden går. Også stress’
styrkelse af amygdala ændrer konteksten, formodentlig også ved genfremkaldte
erindringer, hvilket vil blive uddybet nedenfor. Konteksten er vigtig, da den adskiller
situationer, der ligner hinanden.
En erindring består af såvel videnselementer som markører, der kan aktiveres,
hvorved erindringen rekonstrueres28. De konstrueres af fakta og erfaringer på
baggrund af de informationer, der er lagret i hukommelsen. De reproduceres ikke.
Vores selvskemaer påvirker fx både hvordan vi opfatter og husker personlig
information. Autobiografiske narrativer bevarer begivenheder som de er oplevet,
men er ikke faktuelle rapporter. Erindringer påvirkes også af efterfølgende hændelser
og erfaringer, hvorfor en erindring ikke er en perfekt gengivelse af originalen
(Eysenck, 2000; LeDoux, 2002).
Med hensyn til egentlige tab af erindringer eller viden (hukommelsestab29), kan der
være sket to ting:
Informationen kan være slettet af hukommelsen og derfor ikke længere tilgængelig
eller informationen er fortsat i hukommelsen, men kan ikke findes frem. Det kan
skyldes, at erindringsmarkøren (cue) peger forkert30 (giver falske erindringer) eller er
væk (giver amnesi). Det kan fx være markører som navne og tid eller
stemningsmarkører.
Dette er illustreret i Appendiks B – computeranalogi.
Hukommelsesproblemer er en central ingrediens i flere sygdomme og dysfunktioner.
Det kan der være flere forklaringer på.
For det første kan der være tale om, at problemer med viden skaber afledte problemer
og diagnoser, dvs. at skade på hippocampus forårsager specifikke lidelser.
For det andet, at fejlfunktion i nogle processer medfører lidelser i hippocampus og
dermed skaber hukommelsesproblemer. Det kan dreje sig om problemer med
homeostasis, nogle reguleringsmekanismer eller reparationsmekanismer.
Endelig, som en tredje mulighed, kan der være tale om, at nogle diagnoser rammer
bredt og at hippocampus blot er et af mange berørte organer.
28 Jfr. Appendiks B – computeranalogi.
29 Det er den eksplicitte hukommelse, der er skadet ved amnesi.
30 Det svarer til, at indholdet i en tyk bog er intakt, men alle sidehenvisninger er rodet rundt og peger i
tilfældig retning. Da kan det være svært at finde en specifik oplysning.

Hippocampus og psyken Ane Søndergaard Thomsen
14
Stress er en af de belastninger eller psykiske tilstande, der har forbindelse til
hippocampus og som vil blive uddybet som eksempel.
3.4.1. Stress
Hvis den emotionelle tilstand er meget ophidset, fx under stress31, rammes
hukommelsen ofte. Stress forringer den eksplicitte hukommelse ved at ændre
funktionen i hippocampus. Forskning viser også, at stress kan give falske erindringer.
Hippocampus’ evne til at integrere informationerne i den kontekstuelle situation
forringes (Payne et al., 2002).
Den første af de logiske forklaringsmodeller omhandlede, at problemer med viden i
sig selv kan resultere i sygdomme eller dysfunktioner. Stress kan give en
overrepræsentation af selvnegative tanker, der svækker selvtillid og selvværd.
Manglende selvverifikation kan, ifølge Burke, resultere i depression (Thomsen,
2005).
Hippocampus deltager også i en reguleringsmekanisme, den anden af de tre
forklaringer ovenfor.
Under stress stiger koncentrationen af cortisol. Det system, der kontrollerer
frigivelsen af stresshormonet, kaldes HPA aksen32:
Processen under truende omstændigheder er vel undersøgt33. Amygdala aktiverer
neuroner i hypothalamus, der frigiver CRF/CRH34 fra deres axon terminaler i
hypofysen. CRF/CRH stimulerer hypofysen til at danne ACTH35 til blodbanen,
hvorigennem det påvirker binyrerne til at producere cortisol. Cortisol transporteres
rundt i kroppen til diverse forskellige organer, herunder hjernen. Det har flere
funktioner, bl.a. bevirker det at mængden af blodsukker forhøjes, så der er mere
energi til rådighed.
Cortisol binder sig til receptorer i hippocampus og når tilstrækkeligt mange
receptorer er optaget, sendes der signal til hypothalamus om at stoppe frigivelsen af
31 Selye beskrev ”the general adaptation syndrome” (GAS) med sine tre faser: alarmfasen,
modstandsfasen og udmattelsesfasen. Først mobiliserer det autonome nervesystem kroppen. Så
forbereder HPA aksen vedligeholdelsen af reaktionen (McEwen, 1999). 32 HPA for hypothalamic-pituitary-adrenal axis, idet den involverer hypothalamus, hypofysen og
binyrerne. 33 Jfr. afsnit 3.3.
34 LeDoux skriver, der er tale om et peptid ved navnet corticotropin-releasing factor (CRF), McEwen
kalder det et hormon ved navnet corticotropin releasing hormone (CRH). 35 Adrenocorticotrofin hormon.

Hippocampus og psyken Ane Søndergaard Thomsen
15
CRF/CRH. Hippocampus bidrager således til at regulere stressreaktionen aktiveret af
amygdala (LeDoux, 2002; McEwen, 1999).
Stress er en evolutionær tilpasning, der er god i nogle situationer, men dysfunktionel
på andre tidspunkter. Stress mobiliserer kroppens alarmberedskab. Men under
vedvarende eller voldsom stress mister hippocampus evnen til at foretage sin stress
kontrol pga. en reduktion af antallet af glucocorticoid receptorer (Garcia, 2001;
Garves, 2003; Lupien, 2001). Derved kan der ske såvel somatiske som psykiske
skader.
For meget eller for lidt CRF/CRH forbindes med en række mentale tilstande. For lidt,
dvs. lavere niveau af cortisol, findes ved fx sløvhed, udmattelse, kronisk træthed,
SAD36 og atypisk depression. For meget CRF/CRH associeres med melankoli og
depression (Sternberg, 1999).
Stress skader desuden hippocampus ved at føre til atrofi af dendritter og til sidst til
celledød. Cellerne bliver ekstra sensitive for glutamat og de får nærmest en
forgiftningsreaktion. Det sker fordi neuronerne tømmes for deres energikilde,
glukose, hvilket gør det vanskeligt for dem at fungere37. Mængden af neurotrophiner,
der er essentielle for cellevækst og overlevelse38, falder. Neuronerne begynder at
degenerere. Atrofi og celledød finder hovedsageligt sted i CA3 regionen i
hippocampus. Samtidig mister den dentate gyrus sin evne til neurogenese39. Derfor
ses et mindre volumen af hippocampus hos personer med vedvarende forhøjet niveau
af cortisol (LeDoux, 2002; McEwen, 1999). Det bliver vanskeligere både at danne
rummelige, kontekstuelle repræsentationer af omgivelserne (Garcia, 2001; LeDoux,
2002) og at fremkalde hippocampal LTP40 i CA1 regionen
41 (Garcia, 2001; Lupien,
2001; Zheng et al., 2004). Forskning tyder på at ændringerne i hippocampus er
reversible, i al fald i begyndelsen (Akil et al. 2003; McEwen, 1999; Sapolsky, 2003;
Shankaranarayana Rao et al., 2001).
36 Seasonal affective disorder.
37 Overaktivitet af glutamat skader neuroner. Indstrømning af Ca++ til cellelegemet er øget under
stress. Det kommer ind i mitochondrierne, hvor det accelerer en proces styret af Bcl-2-associated
death protein (Bad). Dette protein sammen med andre molekyler åbner en pore i mitochondriernes
membran, hvorved væske flyder ind og mitochondrierne sprænges. Når cortisol binder sig på
glucocorticoid receptoren, påvirker det desuden mængden og effektiviteten af glukose transportere.
Tilsammen reduceres glukose transporten. Cellen ”sulter ihjel”. 38 Jfr. afsnittet om plasticitet.
39 Dentate gyrus er et af de få områder, hvor der foregår produktion af nye nerveceller, neurogenese.
40 Jfr. afsnit om plasticitet.
41 Skader på CA1 producerer kun anterograde amnesia, dvs. hvor patienten ikke kan lære ny
information (ikke retrograde amnesia, hvor man ikke kan huske ting fra før skaden).

Hippocampus og psyken Ane Søndergaard Thomsen
16
Effekten af vedvarende stress på hippocampus er ret dramatisk, hvilket svækker det
mediale temporallaps hukommelsessystems evne til at danne eksplicitte erindringer.
De tre vigtige strukturer i det trisynaptiske kredsløb påvirkes således ved stress, jfr.
figur 2. Reduceret LTP i CA1, atrofi af CA3 og reduktion af neurogenesen i dentate
gyrus kan delvist forklare hukommelsesforandringer i stressrelaterede psykologiske
dysfunktioner som PTSD42 og depression (Akil et al., 2003; LeDoux, 2002 )
43.
Depression fastlåser patienter i en tilstand af psykologisk tilbagetrækning, hvor
hjernens evne til engagement, opmærksomhed og læring er reduceret. SSRI44
produkterne er de nyeste antidepressive midler. Ved at forhindre genoptagelsen af
serotonin i de præsynaptiske terminaler, øges niveauet af serotonin i synapserne.
Derfor genereres en større intracellulær respons og aktivering af andenhånds
meddelere, hvilket giver øget genaktivering og proteinsyntese, jfr. afsnittet om
plasticitet (sen LTP). Den antidepressive behandling skipper læringen og bevirker at
mængden af Ca i cellen øges. Når effekten først opnås efter nogle uger, kan det
skyldes, at det tager tid at ændre de neurale netværk.
Antidepressiv medicin øger således plasticitet. Ved at lette dannelsen af nye
erindringer kan patienten, evt. med hjælp fra en terapeut, lære nye mentale stader og
ny adfærd (LeDoux, 2002)45.
I modsætning til den hæmmende påvirkningen af hippocampus fremmer cortisol den
implicitte hukommelsesfunktion i de emotionelle systemer. Fx amygdalas bidrag til
frygt, hvorved evnen til at lagre implicit viden om de traumatiske situationer øges46.
Da vi imidlertid ikke er bevidste om, hvad det er, vi lærer om i de situationer, kan
senere angstreaktioner være svære at forstå og kontrollere (LeDoux, 2002).
Forskydning mellem hippocampus og amygdala må betyde noget for erindringerne,
ikke mindst selv-relevante erindringer, jfr. afsnittet om emotioners indflydelse.
Under normale omstændigheder hæmmer hippocampus hypofysen, mens amygdala
stimulerer hypofysen. Det øgede niveau af cortisol under stress hæmmer
hippocampus’ evne til at hæmme hypofysen, hvilket resultere i øget aktivitet i
42 Posttraumatisk stress disorder.
43 Nogle forskere påpeger, at det kan være medfødte forskelle, der gør den berørte sårbar for lidelserne
(Akil et al., 2003 ) 44 Selective serotonin reputake inhibitors.
45 En pointe, der er noget omdiskuteret.
46 Jfr. afsnit 3.3.

Hippocampus og psyken Ane Søndergaard Thomsen
17
hypofysen. Samtidig stimulerer cortisol amygdala, hvilket også resulterer i øget
aktivitet i hypofysen. Der er således risiko for at processen løber løbsk (LeDoux,
2002).
Endelig skal det nævnes, at kroppens aktive organer modtager størst blodforsyning,
således også de aktive dele af hjernen. Derfor kan man forestille sig, af fx arbejde
kan påvirke via en særlig aktivering af processer med en afgrænset hjernefysiologi.
Medarbejdere i særligt erindrings- eller videntunge funktioner kan således få lokale
forandringer i bl.a. hippocampus pga. det høje niveau af cortisol netop hér. Det er et
eksempel på den tredje forklaringsmodel for hukommelsesproblemer, nemlig at
binyrerne sender cortisol rundt i hele kroppen og at de tilfældigvis aktive organer
rammes hårdest.
3.4.2. Andre almindelige situationer
Forskning viser, at der er strukturelle forskelle i visse hjernestrukturer, når man
sammenligner patienter med forskellige psykiatriske diagnoser og normale
kontrolgrupper. Der kan fx være tale om atrofi, ændringer i blod flow og/eller neural
aktivitet (LeDoux, 2002).
Hippocampal atrofi findes i en række forskellige situationer foruden stress. Det
forekommer under almen aldring47, demens og ved Alzheimer. Ved Cushing’s
syndrom forårsager tumorer øget cortisol niveau (McEwen, 1999). Alle med samme
resultat for hukommelsen og psyken.
Hippocampus er således også mindre hos patienter med skizofreni. Disse patienter
har desuden et lavere antal GABA celler i hippocampus, hvilket kan resultere i en
glutamat forgiftning (McEwen, 1999). Undersøgelser tyder på, at synaptisk
interaktion mellem den præfrontale cortex, hippocampus og amygdala er ansvarlig
for nogle af sygdommens aspekter (LeDoux, 2002). Hippocampus og amygdala
regulerer den præfrontale aktivering af accumbens gennem deres rolle i henholdsvis
kontekstuel og emotionel behandling. Den præfrontale aktivitet tilpasser sig således
under normale omstændigheder omgivelserne såvel som specifikke emotionelle
stimuli, men pga. ændringer i hippocampus og amygdala hos skizofrene reduceres
deres evne til at reagere på passende vis.
47 Formodentlig pga. hippocampus’ følsomhed for manglende oxygen pga. mindre blodtilførsel
(Cozolino, 2002).

Hippocampus og psyken Ane Søndergaard Thomsen
18
3.5. Hvad betyder det så for psyken?
Kapitel 3 har i detaljer behandlet, hvilken rolle hippocampus har for hukommelsen
og på hvilken måde det sker. Der har ligeledes ind imellem været beskrevet, hvordan
det kan influere psyken. Her til slut opsamles nogle af de væsentlige konsekvenser
for psyken.
Der er enighed om, at hippocampus har stor indflydelse på dannelsen af eksplicit
hukommelse. Da den indeholder alle de erfaringer og erindringer, vi har gjort os
gennem tiden, er hippocampus uundværlig for, hvordan vi tænker og føler.
Det er ikke kun et spørgsmål om at kunne huske fakta nok til at bestå en eksamen.
Selvet udvikles kontinuerligt gennem en historie af nye og forandrede identiteter.
Med beskadiget hippocampus er der store problemer med at opretholde et
selvbillede. Pga. hippocampus’ integrative rolle kan man forestille sig, at problemer
med at holde del-selver adskilt og/eller forbinde forskellige identiteter ud fra
fællestræk bl.a. kunne have sit udspring i lokale forandringer her. Kerneselvet må
indeholde en række principper for forvaltning af selv-relevante erindringer og del-
selver.
Med anterograde amnesi vil man ikke kunne genkende sig selv, efterhånden som
man ændrer sig, eller huske sine oplevelser. Biologisk vil man ældes, men for
bevidstheden er man fastlåst i billedet fra før skaden skete.
På samme måde kan man ikke opretholde sociale relationer, da man ikke vil kunne
etablere eller gemme de episodiske erindringer. Derved kan man ikke huske, hvem
man har talt med eller hvorom, for ikke at tale om at genkende, hvem man taler med.
Gennem evolutionen har mennesket opnået en evne til at danne og vedligeholde
komplekse repræsentationer for gruppesammenhænge. Uden vil vi ikke kunne
indplacere os i og udfylde en social rolle. Eller handle og reagere passende for
situationen.
Selv mindre skader påvirker de intellektuelle evner. Stress bevirker bl.a., at man
genkalder erindringer langsommere og de er mere fejlbehæftede. Man tænker og
lærer langsommere, ligesom kreativiteten daler (det er sværere at forbinde ny viden
til eksisterende eller gammel viden på nye måder)48.
48 Både hippocampus og den præfrontale cortex er ramt.

Hippocampus og psyken Ane Søndergaard Thomsen
19
Bevidst genkaldelse af episodisk erindring afhænger af integrativ behandling og
kontekst.
Mister man fx tidsmarkøren, mister man fornemmelsen for tid. Man vil ikke kunne
vurdere, hvor langt tid, noget plejer at tage og mister derfor evnen til at estimere
opgaver. Det gør det også vanskeligt at huske, hvilken rækkefølge begivenheder har
fundet sted i, hvilket kan påvirke personligheden. Uden at kunne sætte hændelser i
sammenhænge er det vanskeligt give dem mening. Resultatet er manglende tiltro til
egne vurderinger.
Tydeligt ses det i forsøgene med rumsans. Uden hippocampus er man ikke i stand til
at navigere – et af de forsøg, der er lettest at foretage med dyr. Men kontekst er også
vigtig for mennesker. Den er med til at skille forskellige erindringer fra hinanden.
Integration af relevante informationer er vigtig, når erindringer skal genkaldes. Uden
fejlrepræsenteres informationerne. Det kan være afgørende, når man i forbindelse
med nye problemstillinger drager paralleller. Uden kendskab til tidligere erfaringer er
vi ikke i stand til at vurdere situationen og handle adækvat eller tage (bare lidt)
rationelle beslutninger.
Vores autobiografiske hukommelse og identitetsfølelsen afhænger således af, at vi
kan huske vores historie, hvad enten det drejer sig om de specifikke/generelle eller
trivielle/vigtige ting.
4. Kritisk reflektion
Analysen har givet anledning til nogle spørgsmål, der behandles i det følgende.
4.1. Uklarheder ved teorierne
De beskrevne teorier har nogle udestående punkter. Bl.a. tager LeDoux ikke højde
for at viden ofte huskes bedre, hvis den relaterer sig til eksisterende viden.
Han beskriver, hvordan information etableres, lagres og genfindes. Men mangler
modifikation af eksisterende viden som et overordnet emne. Han er inde på temaet i
form af konsolidering af viden og at man, hver gang man genkalder sig gamle
erindringer og lagrer dem igen, tager højde for eventuelle erfaringer, der er kommet i
mellemtiden. Ligeledes i forbindelse med associativ læring. Men i den virkelige
verden er meget af vores nye viden supplementer, der skal knyttes til, eller
modifikationer af eksisterende viden. Det er ikke undtagelsen, der bekræfter reglen.

Hippocampus og psyken Ane Søndergaard Thomsen
20
Når de fleste erindringer er opdateringer af eksisterende viden. Er det så ikke muligt
at en skade på hippocampus kan skade eksisterende erindringer, fx ved at overskrive
markører? Eller ved at genlagre information ad en defekt vej og derved få den
”forlagt” som en bog, der kommer på forkert hylde på biblioteket?
LeDoux beskriver desuden, hvordan erindringer først lagres i hippocampus, hvorfra
de konsolideres og lagres i de corticale regioner. Han antyder således, at der må være
et temporært lager i hippocampus. Hvordan vil han og de øvrige forskere ellers
forklare, at arbejdshukommelsen er meget kort og langtidshukommelsen
konsolideres over dage?
4.2. Forskningens empiriske grundlag
Meget af forskningen er baseret på forsøg med rotter, fx i forbindelse med den
rummelige hukommelse. Rotters og menneskers hjerner er imidlertid forskellige. Der
findes fx to typer corticosteroid receptorer (MR og GR). MR/GR fordelingen i
hjernen er forskellig fra dyreart til dyreart. Det er derfor nødvendigt at medtage typen
og distributionen af receptorer i hjernen, når man studerer effekten af stress på
hippocampus og den præfrontale cortex. Det er en potentiel fejlkilde, når man slutter
ud fra forsøg med rotter (Lupien, 2001).
Det er dog uomtvisteligt, at hippocampus hár betydning for, hvem vi er, vores
kognitive evner og sociale kompetencer.
Endelig kan man i relation til forskning i nogle sygdomme diskutere, om atrofi er
årsag eller resultat af forhøjet cortisolniveau pga. feedback mekanismen på HPA
aksen. Er niveauet for højt pga. manglende hæmning (grundet atrofi), eller er
niveauet årsag til atrofien (ibid.)?
4.3. Glemsel
Mangelfuld hukommelse behandles som et problem. Glemsel kan imidlertid være en
adaptiv mekanisme. Uden at kunne slette noget, ville vi overbelastes af unyttige
informationer.
Der må ligeledes være nogle reparationsmekanismer. Fx en sletningsfunktion for
selvnegative oplevelser, der ellers ville blive overrepræsenteret. Erfaring viser, at der
derimod er en grundlæggende disposition til at overvurdere egne muligheder og
evner. De selvnegative erfaringer har svært ved at fæste rod.

Hippocampus og psyken Ane Søndergaard Thomsen
21
Glemsel er også en forsvarsmekanisme. Der refereres fx af psykoanalytikere til
fortrængning (repression).
4.4. Lokalisering af funktioner
Hukommelsesforskning begyndte med HM’s amnesi, men HM manglede mere end
hippocampus.
Hukommelse er afhængig af flere områder i hjernen. Der er ingen tvivl om, at
hippocampus spiller en væsentlig rolle for langtidshukommelsen, men som beskrevet
deltager andre strukturer i processen. Det er et distribueret netværk af processer og
systemer, der samarbejder.
Man kan forestille sig, at andre strukturer pga. hjernens plasticitet kunne overtage
nogle af funktionerne, hvis der sker en bilateral skade på hippocampus.
Forsøg viser, at dyr med skade på hippocampus under nogle betingelser kunne lære
at navigere. Opgaven løses således af systemer, der involverer mere end
hippocampus, evt. ved hjælp af sanserne. Alternative strategier tages i anvendelse
(Redish, 2001).
Ofte inddrages accumbens i navigationsmodeller.
Endelig kan hukommelse kun studeres gennem genkaldelse af erindringer. Fejl
tilskrives hippocampus, men det kan fx også være arbejdshukommelsen, der fejler i
sit forsøg på at finde tidligere erindringer. Genkaldelse involverer den præfrontale
cortex.
• Der kan bruges en forkert markør, hvorved en forkert erindring hentes.
• Corticosteroid/stress påvirker også andre regioner i hjernen (Lupien, 2001).
• Den præfrontale cortex deltager også i regulering af HPA aktivitet (ibid.) etc.
5. Konklusion
LeDoux slutter således
”A lifetime of learning and memory, encoded in the constantly
renewed connections between and among our brain cells, makes us
who we are.” (LeDoux .1999, p. 178)

Hippocampus og psyken Ane Søndergaard Thomsen
22
Det opsummerer ganske godt emnet Hippocampus og psyken. Hippocampus er
essentiel for eksplicit hukommelse. Uden kan vi ikke lagre ny eksplicit viden og ville
ikke være i stand til at leve i nuet. Vi ville være fanget i en fortid som den så ud, da
skaden på hippocampus fandt sted.
Hippocampus er involveret i flere mentale sygdomme og dysfunktioner, fx
depression, hvor der ligefrem forekommer hippocampal atrofi, der dog kan vise sig
at være reversibel. Emotionelle erindringer involverer netværk, der inkluderer både
amygdala og hippocampus. Disse strukturer er derfor vigtige for vores psyke og
psykopatologi.
6. Perspektivering
Det er bemærkelsesværdigt, hvor lidt psykologiske grundbøger relaterer psyken med
dens grundlæggende biologiske forudsætninger. Hukommelse beskrives i bøger om
kognitiv psykologi ud fra abstrakte begreber om struktur (ex. langtidshukommelse)
og processer, men næsten uden at nævne hjernens strukturer og involvering.
Neuropsykologisk viden er eksploderet gennem de sidste ti år. Der er kommet nye
neurovidenskabelige forskningsmetoder til og der sker i stigende omfang en
integration af de beslægtede videnskabsgrene. Biologisk psykologi eller
neuropsykologi vil således i fremtiden i højere grad kunne danne baggrund for såvel
psykologisk teori som praksis.
Livets erfaringer efterlader varige aftryk ved at blive lagret som erindringer i
synaptiske kredsløb bl.a. vha. hippocampus. På samme måde virker terapi gennem ny
læring, dvs. gennem ændringer i synaptiske forbindelser.
Et behandlingsforløb vil således ofte bestå af en kombination af psykofarmaka, der
påvirker de molekylærbiologiske processer medicinsk, og psykoterapi, der gør det
samme gennem læring.
Hjernens biologi, ikke mindst hukommelsessystemer, og psyken er således tæt
forbundet.

Hippocampus og psyken Ane Søndergaard Thomsen
23
7. Referenceliste
7.1. Bøger og redigerede værker
Cozolino, L. (2002). The neuroscience of psychotherapy: building and rebuilding the
human brain. New York: W. W. Norton & Company, Inc.
Eysenck, M. W. & Keane, M. T. (2000). Cognitive psychology: a student’s
handbook. 4. udgave. Hove, UK: Psychology Press.
Hobson, J. A. (1999). Order from chaos. In Conlan, R. (Ed.), States of mind: new
discoveries about how our brains make us who we are (pp. 179-199). New
York: John Wiley & Sons, Inc.
Kalat, J. W. (2004). Biological psychology. 8. udgave. Wadsworth
Kandel, E. (1999). Of learning, memory, and genetic switches. In Conlan, R. (Ed.),
States of mind: new discoveries about how our brains make us who we are (pp.
151-178). New York: John Wiley & Sons, Inc.
LeDoux, J. (1999). The power of emotions. In Conlan, R. (Ed.), States of mind: new
discoveries about how our brains make us who we are (pp. 123-149). New
York: John Wiley & Sons, Inc.
LeDoux, J. (2002). Synaptic self: how our brains become who we are. New York:
Penguin Books.
McEwen, B. (1999). Stress and the brain. In Conlan, R. (Ed.), States of mind: new
discoveries about how our brains make us who we are (pp. 81-101). New York:
John Wiley & Sons, Inc.
Sternberg, E. (1999). Emotions and disease: a balance af molecules. In Conlan, R.
(Ed.), States of mind: new discoveries about how our brains make us who we
are (pp. 103-121). New York: John Wiley & Sons, Inc.

Hippocampus og psyken Ane Søndergaard Thomsen
24
7.2. Forskningsartikler
Akil, H., Landis, S. C., Gage, F. H. (2003). Stress and the brain. Brain briefings,
nov/dec.
Florian, C. & Roullet, P. (2004). Hippocampal CA3-region is crucial for acquisition
and memory consolidation in Morris water maze task in mice. Behavioural
Brain Research, 154 (pp. 365-374).
Garcia, R. (2001). Stress, hippocampal plasticity, and spatial learning. Synapse, 40
(pp. 180-183).
Kuhlmann, S., Kirschbaum, C. & Wolf, O. T. (2005). Effects of oral cortisol
treatment in healthy young women on memory retrieval of negative and neutral
words. Neurobiology of Learning and Memory, 83 (pp. 158-162).
Lupien, S. J. & Lepage, M. (2001). Stress, memory, and the hippocampus: can’t live
with it, can’t live without it. Behavioural Brain Research, 127 (pp. 137-158).
McGaugh, J. L. (2000). Memory – a century of consolidation. Science, vol 287, 14.
januar (pp. 248-251).
Payne, J. D., Nadel, L., Allen, J. J. B., Thomas, K. G. F., & Jacobs, W. J. (2002). The
effects of experimentally induced stress on false recognition. Memory, 10, 1
(pp. 1-6).
Redish, A. D. (2001). The hippocampal debate: are we asking the right questions?
Behavioural Brain Research, 127 (pp. 81-98).
Roozendaal, B. (2002). Stress and memory: opposing effects of glucocorticoids on
memory consolidation and memory retrieval. Neurobiology of Learning and
Memory, 78 (pp. 578-595).
Sapolsky, R. M. (2003). Stress and plasticity in the limbic system. Neurochemical
Research, vol. 28, 11 (pp. 1735-1742).

Hippocampus og psyken Ane Søndergaard Thomsen
25
Shankaranarayana Rao, B. S., Madhavi, R., Sunanda & Raju, T. R. (2001). Complete
reversal of dendritic atrophy in CA3 neurons of the hippocampus by
rehabilitation in restraint stressed rats. Current science, vol. 80, 5 (pp. 653-
659).
Zheng, H., Yang, Q. & XU, C. (2004). Effects of chronic stress and phenytoin on the
long-term potentiation (LTP) in rat hippocampal CA1 region. Acta Biochimica
et Biophysica Sinica, vol. 36, 5 (pp. 375-378).
7.3. Andet materiale
Garves, C. A. (2003). The effects of the stress mediated release of glucocorticoids on
the hippocampus: an overview of the biological mechanisms of stress.
Præsentationsmateriale fra 12/01/03.
Thomsen, A. S. (2005). Identity Control Theory – en samling af otte artikler af Peter
J. Burke. Ligeledes publiceret 2006, Aalborg Universitet, Institut for
kommunikation, Psykologi, Klinisk organisations- og arbejdspsykologi
(KOAP).

Hippocampus og psyken Ane Søndergaard Thomsen
26
8. Procesbeskrivelse
”Hippocampus og psyken” er et bredt emne der først gav anledning til et utal af ideer
til afgrænsninger. Både overordnede emner og delbestemmelser, der involverer
hippocampus, fx
• Hukommelseskonsolidering.
• Emotionel modulation af deklarativ langtidshukommelse.
• Stress og plasticitet.
• Effekten af stress på hjernen.
Pga. personlig interesse var det først hensigten at skrive om hippocampus og stress,
men for at give mulighed for at gå i dybden med det emne på et senere tidspunkt,
endte det med at opgaven er blevet brugt som en anledning til en mere
tilbundsgående gennemgang af den biologiske psykologi.
I retrospekt burde opgaven nok have begrænset den neurobiologiske gennemgang til
fordel for en diskussion af hippocampus’ rolle i psykologiske dysfunktioner. Fx
interaktionen mellem amygdala og hippocampus og hvor det går galt i konkrete
situationer. I relation til PTSD kunne det dreje sig om de voldsomme,
multisensoriske flashbacks. Hvordan disse er repetative og stereotype pga.
manglende information om kontekst fra hippocampus etc.

Hippocampus og psyken Ane Søndergaard Thomsen
27
Appendiks A – grundlæggende neuropsykologi
For ikke at anse den grundlæggende neurovidenskab for bekendt gennemgås i dette
Appendiks A de mest basale elementer. Teksten er dannet på baggrund af LeDoux
(2002).
Cellebiologi
Den enkelte hjernecelle, dvs. et neuron, omgives af en cellemembran og består af :
• Et cellelegeme indeholdende bl.a. cellekernen, der lagrer det genetiske materiale,
DNA, og står for produktionen af proteiner, samt mitochondrier, cellens
energifabrik.
• Dendritter, der er cellens inputveje. De har små udposninger (spines), der er
vigtige modtageområder. Der er mange dendritter med forskellige receptorer,
hvorfor cellen kan modtage input fra mange celler (konvergens).
• Et axon, som udgør outputkanalen. Et axon kan være såvel kort som lang,
hvorfor cellerne kan kommunikere over lange afstande. Enden af et axon kaldes
en terminal. Da axonet kan forgrene sig meget, kan cellen have mange terminaler
og derfor kommunikere med mange andre celler (divergens).
Cellerne kommunikerer direkte via synapser, små ekstracellulære mellemrum
indeholdende cerebrospinalvæske.
Oftest går kommunikationen mellem den præsynaptiske terminal til en postsynaptisk
dendrit (evt. spine), men den kan også ske til cellelegemet eller andre axoner.
Tilsvarende kan der forekomme kommunikation mellem dendritter (LeDoux, 2002).
Den biologiske impuls spredes fra neuron til neuron via en elektrokemisk reaktion.
I nervecellen sker det via aktionspotentialet. Det aktiveres, hvor axonet i den
stimulerede celle udgår fra cellelegemet, når området er blevet tilstrækkeligt positivt
ladet49. Det bevæger sig som en bølge ned gennem axonet til terminalen ved at den
positive elektriske ladning bevæger sig ned gennem axonet som en dominoeffekt.
Når impulsen når terminalen, frigives neurotransmittere fra små blærer i den
præsynaptiske terminal til synapsen. Her kommer neurotransmitterne i kontakt med
49 I hvile er den elektriske ladning på indersiden af axonets cellemembran negativ, mens der er positivt
på ydersiden.

Hippocampus og psyken Ane Søndergaard Thomsen
28
og binder sig til selektive receptorer på postsynaptiske dendritter, der efterfølgende
aktiverer processen i den modtagene celle.
Ladningen produceret i dendritten føres til cellelegemet og til axonet, hvor den
initierer et nyt aktionspotentiale. Cellen går da fra at være modtager til afsender; den
bliver præsynaptisk i relation til efterfølgende celler.
Enkelte steder er transmissionen mellem neuroner ren elektrisk. Det sker ved at
neuronerne er i kontakt med hinanden via såkaldte ”gap junctions”.
Bl.a. i hippocampus er hæmmende GABA celler50 forbundne med ”gap junctions”.
Stimuleringen spredes således, at de sammenkoblede celler aktiveres samtidig.
Der er to typer neuroner. Projektions neuroner, der er lange og typisk stimulerer en
postsynaptisk celle, og interneuroner, der er korte og virker inden for et lokalt neuralt
netværk. Interneuronerne kan have en stimulerende effekt, der forstærker impulser,
men de har ofte en hæmmende funktion. De fungerer som en modvægt mod den
stimulerende aktivitet hos projektions neuronerne. De hæmmende neuroner er ofte
konstant aktive i et vist omfang og forhindrer derved projektions neuronerne i at
aktiveres i flæng. Hæmningen skal først overvindes. Hvordan det nærmere sker
beskrives i det efterfølgende afsnit om de molekylærbiologiske processer.
Molekylærbiologi
Cerebrospinalvæsken, der omgiver neuronerne, indeholder kemiske molekyler, der
påvirker cellefunktionen. Når neuronerne er i hvile er ladningen inde i cellen mere
negativ end væsken udenfor.
Den vigtigste stimulerende neurotransmitter er glutamat51. Når glutamat bindes til
ydersiden af en postsynaptisk receptor, åbnes der en passage i cellemembranen, der
tillader positivt ladede ioner fra cerebrospinalvæsken, bevæge sig ind i cellen. Det
påvirker den kemiske balance. Hvis tilstrækkelig mange receptorer på den
postsynaptiske celle er optaget samtidigt, bliver indstrømningen stor nok til, at
cellelegemet nær axonet bliver tilstrækkelig positivt ladet til at aktivere
aktionspotentialet.
50 Jfr. næste afsnit for beskrivelsen af GABA celler.
51 Glutamat er en aminosyre, der har flere funktioner i kroppen, bl.a. visse metaboliske processer.

Hippocampus og psyken Ane Søndergaard Thomsen
29
De hæmmende neuroner frigiver GABA52. Når GABA bindes til sine receptorer sker
der tilsvarende en indstrømning af negativt ladede ioner til cellen, CL-. Det gør det
vanskeligere for glutamat at opnå den stimulerende virkning, idet negative ioner
neutraliserer positive og det er nettoeffekten, der skal være positiv for at aktivere
aktionspotentialet.
Dette forstærkes af, at receptorerne for glutamat ofte sidder på dendritterne, mens
dem for GABA sidder på cellelegemerne eller dendritterne tæt på cellelegemet.
Stimulationen skal således forbi det hæmmende skjold.
Uden GABA hæmning ville neuronerne aktivere sig selv ihjel. Overaktivitet af
glutamat skader neuroner, hvilket spiller en væsentlig rolle i flere sygdomme.
Flere typer psykofarmaka virker således ved at regulere GABA hæmning. Valium
øger fx GABA’s naturlige evne til at hæmme glutamat i amygdala ved at lette
transmissionen og gør det derved vanskeligere at aktivere en angstreaktion.
Efter neurotransmitterne har haft deres effekt, bliver de som en del af
reguleringsmekanismen suget tilbage i de terminaler, der oprindeligt frigav dem.
Foruden glutamat og GABA er en række mere langsomme, men langtidsvirkende,
modulatorer involveret.
Peptider frigives ofte fra samme terminaler som glutamat og GABA, men binder sig
til specifikke receptorer, og kan derved enten fremme eller hæmme de hurtige
neurotransmitteres stimulerende virkning. Opiater som endorfiner hører til denne
type. De kan ændre smertefølsomhed og humør.
Monoaminer produceres mest i celler i hjernestammen, men neuronernes axoner er
lange og breder sig ud i hele hjernen, hvorfor effekten ikke er ret specifik. De
påvirker mange dele af hjernen og giver generel højt eller lavt niveau af ophidselse.
De virker ligeledes ved enten fremme eller hæmme glutamat eller GABA. Serotonin,
dopamin, epinephrin og norepinephrin hører til den type. Mange psykofarmaka
virker ved at påvirke niveauet af monoaminer, herunder SSRI-præparaterne. Ved at
undgå at serotonin suges tilbage i terminalerne efter brug er der mere serotonin i
synapserne. Mere serotonin i fx amygdala stimulerer GABA celler og fremmer
GABA cellernes hæmning, hvilket giver mindre angst.
Hormoner frigives fra kroppens endokrine organer53 og sendes rundt i blodbanen.
Cortisol fra binyrerne produceres fx under stress og påvirker i hjernen GABA’s evne
til at hæmme glutamat. Det sker fx i amygdala ved at sænke niveauet af serotonin,
jfr. ovenfor, hvilket giver øget angst.
52 GABA er en forkortelse for aminosyren gamma-aminobutyl syre.
53 Fx binyrer, hypofysen og kønskirtlerne.

Hippocampus og psyken Ane Søndergaard Thomsen
30
Appendiks B – computeranalogi
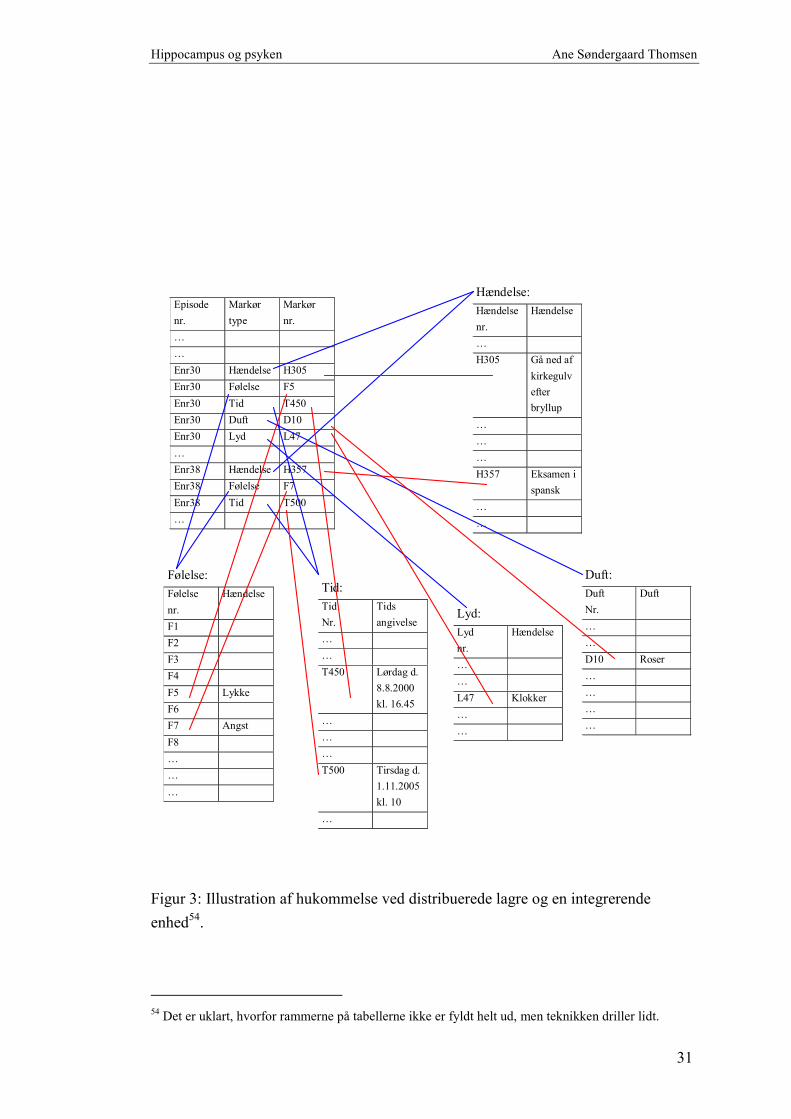
Forestiller man sig hippocampus’ integrative funktion i en computeranalogi er det
bedste sammenligningsgrundlag en relationsdatabase. En sådan indeholder tabeller,
hvor relationer knyttes sammen (hippocampus), og egentlige arkiver bestående af
indekserede datatabeller (neocortex), jfr. figur 3 næste side.
Hippocampus indeholder samlende information om diverse episoder. I den simple
model nedenfor er det skitseret som en tabel med tre kolonner, hvor information om
en given episode er indekseret ved et tilfældigt løbe-/episodenummer (EnrX).
Heri forbindes oplysning om, hvilken type markør der er lagret information om i
relation til episoden. Fx er der lagret information om episode 30 fem forskellige
steder distribueret ud efter type af markør – de blå streger. Disse er igen illustreret
ved hver deres tabel. Endelig indeholder hippocampus i modellen information om,
hvor i de fem ”markør tabeller”, der skal findes yderligere information – de røde
streger.
Episode 30 består således af en:
• hændelse - gå ned af kirkegulvet efter ens eget bryllup
• følelse - lykke
• tid - lørdag den 8.8.2000 kl. 16.45
• duft - roser
• lyd - kirkeklokker.
Episode 38 er en:
• hændelse - eksamen i spansk
• følelse - angst
• tid - tirsdag d. 1.11.2005 kl. 10.
Dette simple eksempel illustrerer, at man kan glemme både bagud og fremad i tid. Fx
eksamen en dag i fremtiden. Det kan ske ved at selve erindringen er væk – det
distribuerede lager i neocortex er slettet. Eller ved at markøren er væk eller peger
forkert. Sidstnævnte kan give falske erindringer. Selv med de meget få forbindelser i
figur 3 er det ved at være uoverskueligt med alle de streger.
Hippocampus og psyken Ane Søndergaard Thomsen
31
Figur 3: Illustration af hukommelse ved distribuerede lagre og en integrerende
enhed54.
54 Det er uklart, hvorfor rammerne på tabellerne ikke er fyldt helt ud, men teknikken driller lidt.
Episode
nr.
Markør
type
Markør
nr.
…
…
Enr30 Hændelse H305
Enr30 Følelse F5
Enr30 Tid T450
Enr30 Duft D10
Enr30 Lyd L47
…
Enr38 Hændelse H357
Enr38 Følelse F7
Enr38 Tid T500
…
Hændelse:
Hændelse
nr.
Hændelse
…
H305 Gå ned af
kirkegulv
efter
bryllup
…
…
…
H357 Eksamen i
spansk
…
…
Følelse:
Følelse
nr.
Hændelse
F1
F2
F3
F4
F5 Lykke
F6
F7 Angst
F8
…
…
…
Tid:
Tid
Nr.
Tids
angivelse
…
…
T450 Lørdag d.
8.8.2000
kl. 16.45
…
…
…
T500 Tirsdag d.
1.11.2005
kl. 10
…
Duft:
Duft
Nr.
Duft
…
…
D10 Roser
…
…
…
…
Lyd:
Lyd
nr.
Hændelse
…
…
L47 Klokker
…
…

Hippocampus og psyken Ane Søndergaard Thomsen
32
Dette er blot et eksempel. Indgangsnøglen er næppe et løbenummer og opbygningen
er væsentlig mere kompleks. Men at der let kan ske sammenblandinger af
informationer, der har indflydelse på psyken, er hævet over enhver tvivl, selv her.
Modellen udelukker ikke, at man kan få adgang til de distribuerede lagre uden om
hippocampus. Man kan i nogle situationer aktivere de distribuerede lagre direkte, de
har hver sin nøgle til indeksering. Selvom relationen går i stykker, kan erindringen
godt aktiveres af andre veje.
Det er endda muligt at fremkalde følelser eller erindringer ved stimulation af hjernen
under en operation.
Det kan forklare, hvordan erindringer kan blive uafhængig af hippocampus efter
konsolidering. Og hvorfor kontekst er en af hippocampus’ væsentlige funktioner – i
det tilfælde skal information findes frem flere steder fra. Her er
relationerne/forbindelserne vigtige.
Modellen illustrerer ligeledes, at forskellige episoder kan pege på den samme
information. Følelsen af angst kan således knyttes til forskellige erindringer og
episoder, uden at den gemmes flere gange i det distribuerede lager. Det kan minimere
redundans. Hjernen, og hele organismen, er evolutionært udviklet til at holde en god
energiøkonomi – hvorfor så ikke også på lagerområdet?





























