Embed Size (px)
Citation preview

Neurochemical Research, Vol. 22, No. 8, 1997, pp. 957-966
Basal Forebrain Cholinergic Immunolesion by 192IgG-Saporin: Evidence for a Presynaptic Location ofSubpopulations of a2-and p-Adrenergic as Well as 5-HT2A
Receptors on Cortical Cholinergic Terminals*
Mechthild Heider,1 Reinhard Schliebs,1,2 Steffen RoBner,1 and Volker Bigl1
(Accepted January 7, 1997)
To study whether the changes in cortical noradrenergic and serotonergic mechanisms observed inpatients with Alzheimer's disease are the consequence of reduced cortical cholinergic activity, anovel cholinergic immunotoxin (conjugate of the monoclonal antibody 192IgG against the low-affinity nerve growth factor receptor with the cytotoxic protein saporin, 1921gG-saporin) was usedto produce a specific and selective loss of cholinergic cells in rat basal forebrain nuclei. To correlatethe responses to cholinergic immunolesion in cholinoceptive cortical target regions with cholinergichypoactivity, quantitative receptor autoradiography to measure adrenoceptors and 5-hydroxytryp-tamine (5-HT) receptor subtypes, and histochemistry to estimate acetylcholinesterase activity, wereperformed in adjacent brain sections. a1-adrenoceptor and 5-HT1A receptor binding were not af-fected by cholinergic immunolesion in any of the cortical and hippocampal regions studied. How-ever, cholinergic immunolesion resulted in significantly reduced a2-and B-adrenoceptor as well as5-HT2A receptor binding in a number cortical and hippocampal regions displaying a reduced activityof acetylcholinesterase, already detectable seven days after a single injection of 192IgG-saporinand persisting up to three months post lesion without any significant recovery. The data suggestthat at least a subpopulation of a2-and (B-adrenoceptor as well 5-HT2A receptor subtype is presenton cortical and hippocampal cholinergic terminals originating in the basal forebrain. The lesion-induced receptor changes suggest that the alterations in cortical 5-HT2 receptor binding observedin patients with Alzheimer's disease might be secondary to the cholinergic deficits.
KEY WORDS: Alpha-adrenoceptor; beta-adrenoceptor; 5-hydroxytryptamine receptor; autoradiography; ace-tylcholinesterase; cerebral cortex; image analysis.
INTRODUCTION
Alzheimer's disease is known to be associated witha very early and progressive loss of cholinergic cells in
1 Paul Flechsig Institute for Brain Research, University Leipzig, Jahn-allee 59, D-04109 Leipzig, Germany.
2 Address reprint requests to: Dr. Reinhard Schliebs, University ofLeipzig, Paul Flechsig Institute for Brain Research, Department ofNeurochemistry, Jahnallee 59, D-04109 Leipzig, Germany. Tel.:+49-341-97-25766; fax +49-341-211 44 92.
* Special issue dedicated to Dr. Annica Dahlstrohm.
the nucleus basalis of Meynert. This cortical cholinergicdysfunction has been implicated for the severe cognitivedeficits in these patients. However, the changes in thecholinergic transmission are accompanied by a numberof alterations in other transmitter systems including glu-tamate, GABA, noradrenaline and serotonin (see e.g. ref.1), suggesting the importance of other neurotransmittersystems in cognition. Among these systems noradrena-line and serotonin are thought to be special significance.This is emphasized by a number of studies demonstrat-ing impairments in memory and cognitive behaviour fol-
9570364-3190/97/0800-0957$12.50/0 C 1997 Plenum Publishing Corporation

958 Heider, Schliebs, Refiner, and Bigl
lowing experimentally-induced alterations in the centralnoradrenergic and serotonergic transmission (2-7). Al-though the administration of both serotonergic and no-radrenergic drugs to animals can improve performancein various learning tasks (5,8,9), the role of the seroto-nergic system in learning and memory processes is stillcontroversial (for review, see ref. 7). It is also possiblethat serotonin affects cognitive processes by modulationof the cholinergic system and vice-versa. Affecting onetransmitter system, e.g. by drugs or by degenerative pro-cesses in the diseased brain, may alter the efficacy ofanother one. Thus, there is a big demand to examine theinteraction of transmitter systems which may play a rolein controlling cognitive function (7). This might contrib-ute to derive more validated rationales for pharmacolog-ical interventions in this process and to find newtherapeutical strategies to treat Alzheimer's disease. Toelucidate the exact nature of the interaction of acetyl-choline with other transmitter systems, adequate animalmodels are required to produce specific cholinergic def-icits in vivo. Cholinergic lesion paradims have been usedto characterize the role of the cholinergic system in cor-tical information processing, learning and memory andin cognitive behaviour. In the past, a number of studieshave applied excitotoxins or cholinotoxins to producecortical cholinergic deficits. However, the cytotoxinsused are far from being selective to cholinergic cells (seee.g. ref. 10). A recently introduced new approach byWiley et al. (11) which uses a monoclonal antibody(192IgG) to the low affinity NGF receptor p75NGFR totarget a ribosome-inactivating cytotoxic protein (saporin)selectively to neurotrophin receptor bearing cholinergicneurons, circumvents these limitations. Intraventricularadministration of this 192IgG-saporin conjugate resultedin a complete disappearance of choline acetyltransferase(ChAT) and NGF receptor immunoreactive neuronsthroughout the basal forebrain, but spared other neuronalsystems in the basal forebrain nuclei (12-18). The spe-cific loss of basal forebrain cholinergic cells led to astrong activation of microglia in all magnocellular basalforebrain nuclei corresponding to the topographic local-ization of degenerating cholinergic cells (14). Substantialimmunotoxin-induced reductions in both ChAT-immu-noreactivity and acetylcholinesterase (AChE) staining aswell as high-affinity choline uptake sites in a number ofcortical regions including hippocampus and olfactorybulb, but not in the striatum and cerebellum, were de-tectable already seven days after lesion (13,19). Thesehistochemical changes are complemented by specific al-terations in cortical cholinergic neurochemical param-eters including muscarinic acetylcholine receptorsubtypes, choline uptake, acetylcholine release and mus-
carinic receptor-mediated production of inositol phos-phates (20,21). Therefore, 192IgG-saporin was used toselectively lesion basal forebrain cholinergic cells and tostudy the impact of cortical cholinergic hypoactivity onnoradrenergic and serotonergic mechanisms in cholino-ceptive cortical target regions. Acetylcholine may influ-ence the neuronal activity of other transmitter systemsby acting through different postsynaptic transmitter re-ceptors in the cerebral cortex. Neurotransmitter receptorsare one of the decisive links in the chain of synapticinformation processing. They can markedly respond toalterations in neuronal activity by adaptive mechanismslike sub/supersensitivity or down-regulation (22), whichmakes them an appropriate tool to monitor for interactivechanges in the neuronal activity of a particular transmittersystem. Therefore, the effect of cholinergic immunolesionfor various periods of time on cortical adrenoceptor (a1-,, a2-, and (B-subtype) and 5-hydroxytrvptamine (5-HT)receptor subtypes (5-HT1A, 5-HT2A) was investigated byquantitative receptor autoradiography, which allows for ascreening of receptor changes through the whole brain.To correlate the responses to cholinergic immunolesionin cholinoceptive cortical target regions with cholinergichypoactivity, acetylcholinesterase (AChE) histochemistryin adjacent brain sections was performed.
EXPERIMENTAL PROCEDURE
Materials. [7-methoxy-3H]Prazosin (2886 GBq/mmol), [methyl-3H]Rauwolscine (2738 GBq/mmol), [ring, propyl-3H(N)]L-Dihydroal-prenolol (DMA, 3922 GBq/mmol), [propyl-2,3-ring-l,2,3-3H]8-Hydroxy-DPAT (4791 GBq/mmol), and [ethylene-3H]Ketanserin(3149 GBq/mmol) were purchased from New England Nuclear,DuPont, Germany.
The cholinergic immunotoxin 192IgG-saporin was obtained fromChemicon International Inc., Temecula, USA. Phentolamin (SIGMA),L-isoproterenol (SIGMA), mianserin (SIGMA), and serotonin(SIGMA) were used for assessing nonspecific binding, all other chem-icals used were commercial products of highest purity available.
Treatment of Animals and Tissue Preparation. Immunolesion ofthe forebrain cholinergic system was performed as described previ-ously (19). Briefly, adult male Wistar rats with initial weight of 235-250 g were anesthetized with 60 mg ketamine i.m. and 40 mgpentobarbital i.p./kg body weight. The rat's head was placed in a ster-eotaxic frame and the skull was opened at bregma 1.2 mm lateral tothe longitudinal suture by placing a small hole. Stereotaxic injectionof approximately 4 mg (in 10 ml of phosphate buffered saline) of192IgG-saporin was made into the left lateral ventricle (coordinates:posterior 0.8 mm and lateral 1.2 mm from bregma) using a 10 mlsyringe with a 31-gauge needle (Hamilton). The needle was insertedto a depth of 3.4 mm below the cortical surface and the injectionperformed at the rate of 1 m1 per minute. After injection the syringewas kept for additional 5 min at the injection site to allow for a com-plete diffusion. An equal number of animals received the vehicle aloneand were considered as controls. Seven, 15, 30, and 90 days following

Aminergic Receptor Binding after Cholinergic Immunolesion 959
injection the animals were sacrificed; the brains were frozen andtwelve mm thick serial coronal sections were cut with a cryostat mi-crotome (Miles, USA) and thaw-mounted on gelatin coated slides, andstored at — 20°C until use.
The animal experiments performed in this study have been ap-proved by the independent Ethical Committee for Animal Experimentsof the Regierungsprasidium Leipzig, licence no. TW 10/92.
Acetylcholinesterase Histochemistry. Histochemical staining foracetylcholinesterase (AChE) was performed in adjacent sections ineach cortical level according to the method of Andra and Lojda (23).Briefly, air dried cryocut sections were preincubated for 30 min at37°C in 0.1 M Tris-maleate buffer (pH 5.0) containing 30 mM iso-OMPA to inhibit nonacetylcholinesterases. After preincubation sec-tions were incubated in a solution consisting of 1.7 mM acetylthioch-oline iodide, 40 mM sodium citrate, 12 mM cupric sulfate, 8 mMpotassium ferrycyanide, 30 mM iso-OMPA and 75 mM Tris-maleatebuffer (pH 5.0) for 60 min at 37°C. The reaction was stopped byrinsing sections in 0.1 M Tris-maleate buffer (pH 5.0) followed by ashort dipping in distilled water. Finally, the sections were dehydratedand coversliped.
Receptor Binding Assays
Adrenoceptor Subtypes. [3H]prazosin binding to thea1-adenoceptor subtype was carried out using a slightlymodified procedure of Rainbow et al. (24): sections wereprewashed for 15 min in 170 mM Tris-HCl buffer (pH7.4) at 4°C and dried under a stream of air at roomtemperature before performing the binding assay. Afterpreincubation, the dried slides were covered with 170mM Tris-HCl (pH 7.4) containing 1 nM [3H]prazosinand incubated in humid chambers for 30 min at roomtemperature. After incubation, sections were rinsed twiceof 10 min each with ice-cold buffer, followed by a quickdipping into ice-cold distilled water, and blown dry.Nonspecific binding was estimated in adjacent sectionsby adding 10 (iM phentolamine to the incubation buffer,and represented <10% of total binding.
For detecting the a2-adrenoceptor subtype accord-ing to a slightly modified method of Noyer et al. (25),[3H]rauwolscine was used as radioligand. As[3H]rauwolscine does not discriminate between a2-ad-renoceptor and 5-HT1A receptors (26), the 5-HT1A recep-tors were masked with unlabeled serotonin. Sectionswere prewashed for 15 min in 50 mM sodium-potassiumphosphate buffer (pH 7.7) at 4°C and blown dry undera stream of warm air prior to the binding assay. Afterpreincubation, the dried slides were flooded with 50 mMsodium-potassium phosphate buffer (pH 7.7) containing0.3 mM serotonin, 10 mM pargyline, 0.01% ascorbic acidand 5 nM [3H]rauwolscine and incubated in humidchambers for 30 min at room temperature. After incu-bation, sections were rinsed twice of 10 min each withice-cold Tris-HCl buffer (pH 7.7), followed by a quickdipping into ice-cold distilled water, and blown dry.
Nonspecific binding was estimated in adjacent sectionsby adding 100 uM phentolamine to the incubation buf-fer, and represented < 10-20% of total binding.
[3H]Dihydroalprenolol (DHA) binding to (3-adren-oceptor sites was performed as recently described (27).Briefly, after preincubation for 15 min, slides were in-cubated for one hour at room temperature with 170 mMTris-HCl containing 10 mM MgSO4 (pH 7.4) and 1 nM[3H]DHA. After incubation, slides were given two rinsesof 5 min each in ice-cold buffer followed by a shortdipping in ice-cold distilled water before drying in alight air stream. Nonspecific binding was estimated inadjacent sections by adding 1 mM isoproterenol to theincubation buffer, and amounted to about 30%-40% oftotal binding.
Serotonergic Receptor Subtypes. [3H]8-OH-DPATbinding to the 5-HT1A receptor subtype was carried outaccording to Dyck et al. (28). Slides were preincubatedfor 30 min in ice-cold 170 mM Tris-HCl buffer (pH7.6)containing 4 mM CaCl2. After preincubation, the driedslides were covered with 170 mM Tris-HCl containing4 mM CaCl2, 10 (xM pargyline, 0.01% ascorbic acid, and2 nM [3H]8-OH-DPAT (pH 7.6) and incubated in humidchambers for 60 min at room temperature. Followingincubation, slides were rinsed twice for 5 min each withice-cold buffer followed by one quick rinse with ice-colddistilled water before drying in a light air stream. Non-specific binding was estimated in adjacent sections byadding 10 mM unlabeled serotonin to the incubation buf-fer, and amounted to about 15% of total binding.
For [3H]ketanserin binding to the 5-HT2A receptorsubtype (29), sections were preincubated for 15 mineach in 170 mM Tris-HCl (pH 7.7). After drying, slideswere immediately flooded with an incubation solutionconsisting of 170 mM Tris-HCl (pH 7.7) and 2 nM[3H]ketanserin. After two hours of incubation at roomtemperature, slides were given two rinses of 10 min eachin ice-cold buffer, followed by one quick rinse with ice-cold distilled water before drying in a light air stream.Nonspecific binding was determined in adjacent sectionsco-incubated with 1 mM mianserin and did normally notexceed 10% of total binding.
Receptor Autoradiography. The labeled and driedtissue sections were apposed to tritium-sensitive film(3H-Hyperfilm, Amersham) at 4 °C together with slidescontaining standards of known radioactivity level (3H-microscale, Amersham). After exposure of three to eightweeks (depending on the radioligand applied), the filmswere developed with a Kodak D19 developer for 5 minat 20 °C, fixed, rinsed and dried.
Evaluation of Autoradiograms. Quantitative analy-sis of the autoradiograms was done on a video camera-
960 Heider, Schliebs, RoBner, and Bigl
corresponding tissue region using a calibration curveplotted from the radioactivity of the tissue standards(kBq/mg tissue equivalent) and the densitometrically de-termined optical density values of the respective auto-radiograms. Receptor densities were expressed as fhiolspecifically bound radioligand per mg tissue (for detailsof quantification, see also ref. 30).
For evaluation of cortical regions all over the braincoronal sections at selected distances from the bregma:+2.7, +0.2, -3.3, -5.8 mm (according to ref. 31) wereused. Particularly, optical density readings were per-formed in the following regions: subfields of hippocam-pal formation including dentate gyrus, frontal cortices(Frl, Fr2, Fr3), parietal cortices (Parl, Par2), temporalcortices (Tel/Te2) occipital cortices (OC1, OC2), fore-limb and hindlimb area, piriform, cingulate, entorhinaland retrosplenial cortex as well as corpus striatum. Thedata obtained were corrected for non-specific binding.Measurements were made on three consecutive sectionsfrom each animal. The corresponding data obtained fromfour animals in each experimental group were averaged.
Statistical Analysis. A one-way analysis of variance(ANOVA) was used to examine differences in the neu-rochemical parameters measured between brain regionand coronal level and between brain regions of controland experimental animals, followed by t-tests usingDunnett's correction for multiple simultaneous compar-isons by means of the software package SPSS. Differ-ences between treatments were considered statisticallysignificant when P < 0.05.
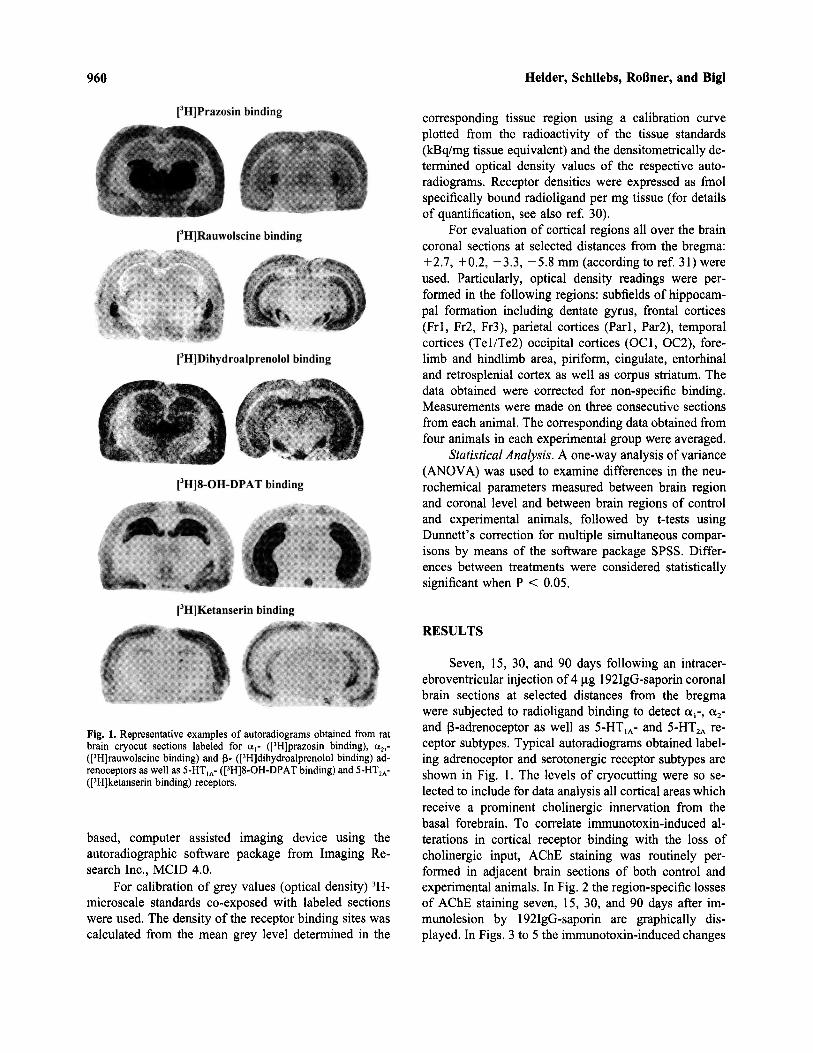
Fig. 1. Representative examples of autoradiograms obtained from ratbrain cryocut sections labeled for a1- ([3H]prazosin binding), a2,-([3H]rauwolscine binding) and (B- ([3H]dihydroalprenolol binding) ad-renoceptors as well as 5-HT1A- ([3H]8-OH-DPAT binding) and 5-HT2A-([3H]ketanserin binding) receptors.
based, computer assisted imaging device using theautoradiographic software package from Imaging Re-search Inc., MCID 4.0.
For calibration of grey values (optical density) 3H-microscale standards co-exposed with labeled sectionswere used. The density of the receptor binding sites wascalculated from the mean grey level determined in the
RESULTS
Seven, 15, 30, and 90 days following an intracer-ebroventricular injection of 4 mg 192IgG-saporin coronalbrain sections at selected distances from the bregmawere subjected to radioligand binding to detect a1-, a2-and 3-adrenoceptor as well as 5-HT1A- and 5-HT2A re-ceptor subtypes. Typical autoradiograms obtained label-ing adrenoceptor and serotonergic receptor subtypes areshown in Fig. 1. The levels of cryocutting were so se-lected to include for data analysis all cortical areas whichreceive a prominent cholinergic innervation from thebasal forebrain. To correlate immunotoxin-induced al-terations in cortical receptor binding with the loss ofcholinergic input, AChE staining was routinely per-formed in adjacent brain sections of both control andexperimental animals. In Fig. 2 the region-specific lossesof AChE staining seven, 15, 30, and 90 days after im-munolesion by 192IgG-saporin are graphically dis-played. In Figs. 3 to 5 the immunotoxin-induced changes
Aminergic Receptor Binding after Cholinergic Immunolesion 961
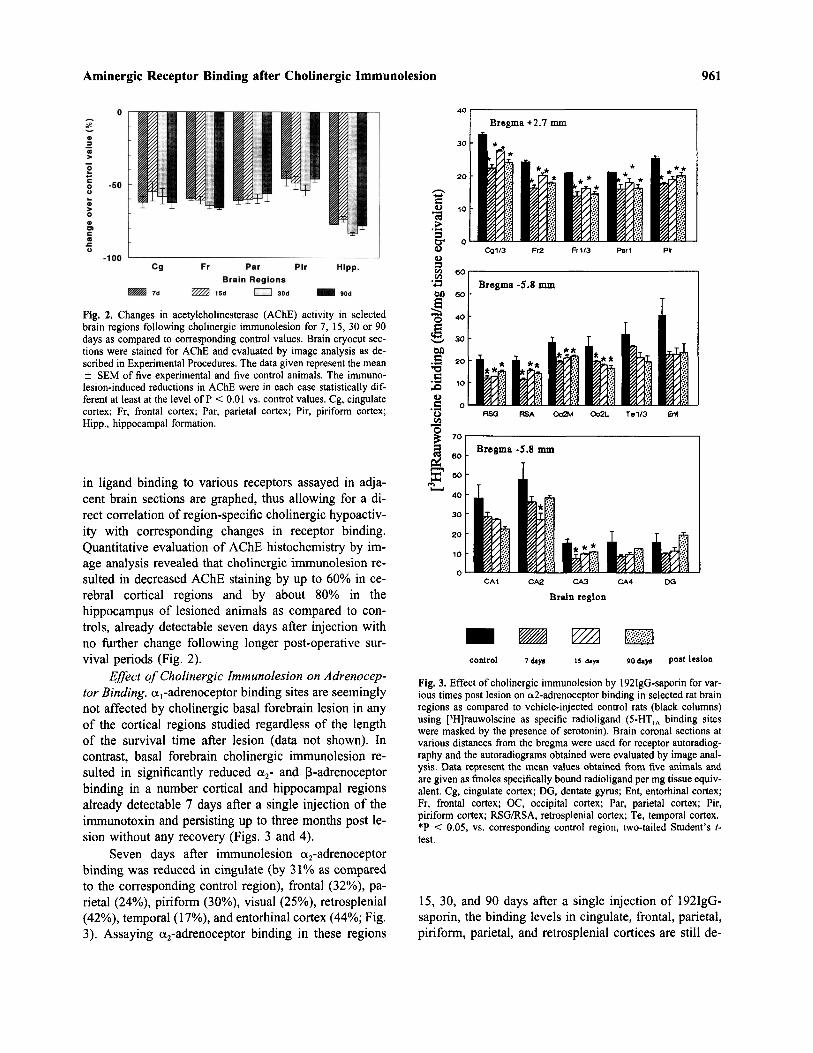
Fig. 2. Changes in acetylcholinesterase (AChE) activity in selectedbrain regions following cholinergic immunolesion for 7, 15, 30 or 90days as compared to corresponding control values. Brain cryocut sec-tions were stained for AChE and evaluated by image analysis as de-scribed in Experimental Procedures. The data given represent the mean± SEM of five experimental and five control animals. The immuno-lesion-induced reductions in AChE were in each case statistically dif-ferent at least at the level of P < 0.01 vs. control values. Cg, cingulatecortex; Fr, frontal cortex; Par, parietal cortex; Pir, piriform cortex;Hipp., hippocampal formation.
in ligand binding to various receptors assayed in adja-cent brain sections are graphed, thus allowing for a di-rect correlation of region-specific cholinergic hypoactiv-ity with corresponding changes in receptor binding.Quantitative evaluation of AChE histochemistry by im-age analysis revealed that cholinergic immunolesion re-sulted in decreased AChE staining by up to 60% in ce-rebral cortical regions and by about 80% in thehippocampus of lesioned animals as compared to con-trols, already detectable seven days after injection withno further change following longer post-operative sur-vival periods (Fig. 2).
Effect of Cholinergic Immunolesion on Adrenocep-tor Binding. a1-adrenoceptor binding sites are seeminglynot affected by cholinergic basal forebrain lesion in anyof the cortical regions studied regardless of the lengthof the survival time after lesion (data not shown). Incontrast, basal forebrain cholinergic immunolesion re-sulted in significantly reduced a2- and 3-adrenoceptorbinding in a number cortical and hippocampal regionsalready detectable 7 days after a single injection of theimmunotoxin and persisting up to three months post le-sion without any recovery (Figs. 3 and 4).
Seven days after immunolesion a2-adrenoceptorbinding was reduced in cingulate (by 31% as comparedto the corresponding control region), frontal (32%), pa-rietal (24%), piriform (30%), visual (25%), retrosplenial(42%), temporal (17%), and entorhinal cortex (44%; Fig.3). Assaying a2-adrenoceptor binding in these regions
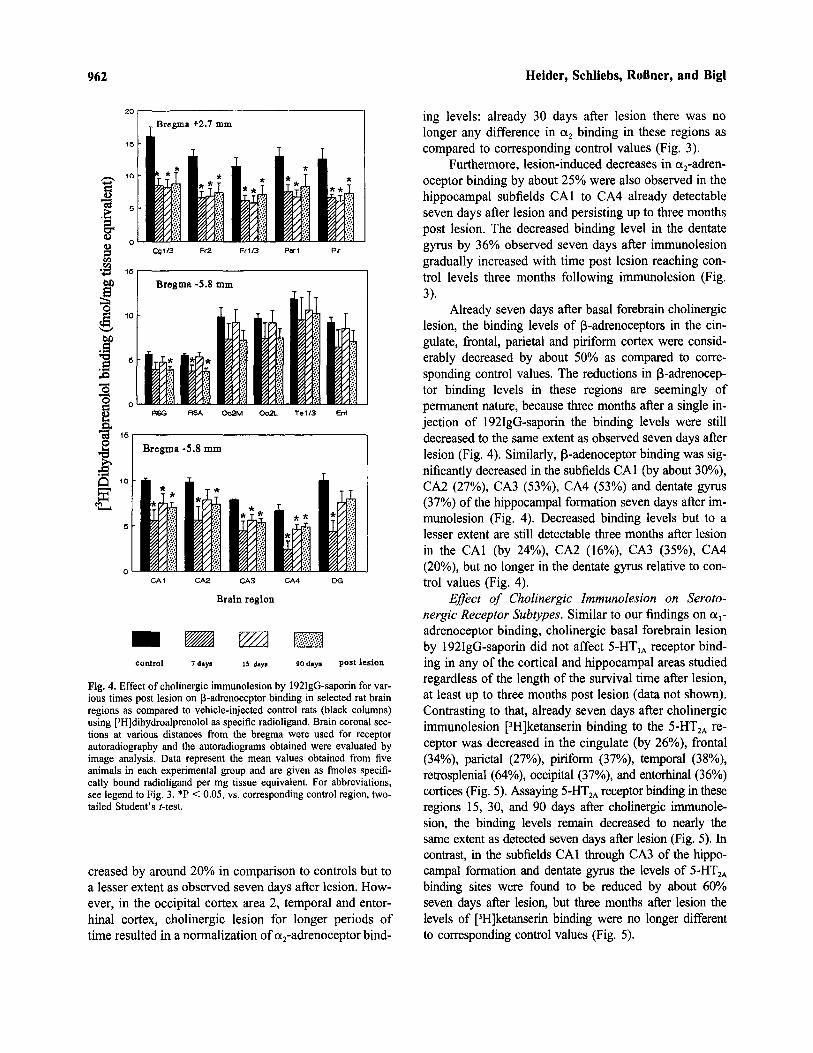
Fig. 3. Effect of cholinergic immunolesion by 192IgQ-saporin for var-ious times post lesion on a2-adrenoceptor binding in selected rat brainregions as compared to vehicle-injected control rats (black columns)using [3H]rauwolscine as specific radioligand (5-HTIA binding siteswere masked by the presence of serotonin). Brain coronal sections atvarious distances from the bregma were used for receptor autoradiog-raphy and the autoradiograms obtained were evaluated by image anal-ysis. Data represent the mean values obtained from five animals andare given as finoles specifically bound radioligand per mg tissue equiv-alent. Cg, cingulate cortex; DG, dentate gyrus; Ent, entorhinal cortex;Fr, frontal cortex; OC, occipital cortex; Par, parietal cortex; Pir,piriform cortex; RSG/RSA, retrosplenial cortex; Te, temporal cortex.*P < 0.05, vs. corresponding control region, two-tailed Student's t-test.
15, 30, and 90 days after a single injection of 192IgG-saporin, the binding levels in cingulate, frontal, parietal,piriform, parietal, and retrosplenial cortices are still de-
962 Heider, Schliebs, RoBner, and Bigl
ing levels: already 30 days after lesion there was nolonger any difference in a2 binding in these regions ascompared to corresponding control values (Fig. 3).
Furthermore, lesion-induced decreases in a2-adren-oceptor binding by about 25% were also observed in thehippocampal subfields CA1 to CA4 already detectableseven days after lesion and persisting up to three monthspost lesion. The decreased binding level in the dentategyrus by 36% observed seven days after immunolesiongradually increased with time post lesion reaching con-trol levels three months following immunolesion (Fig.3).
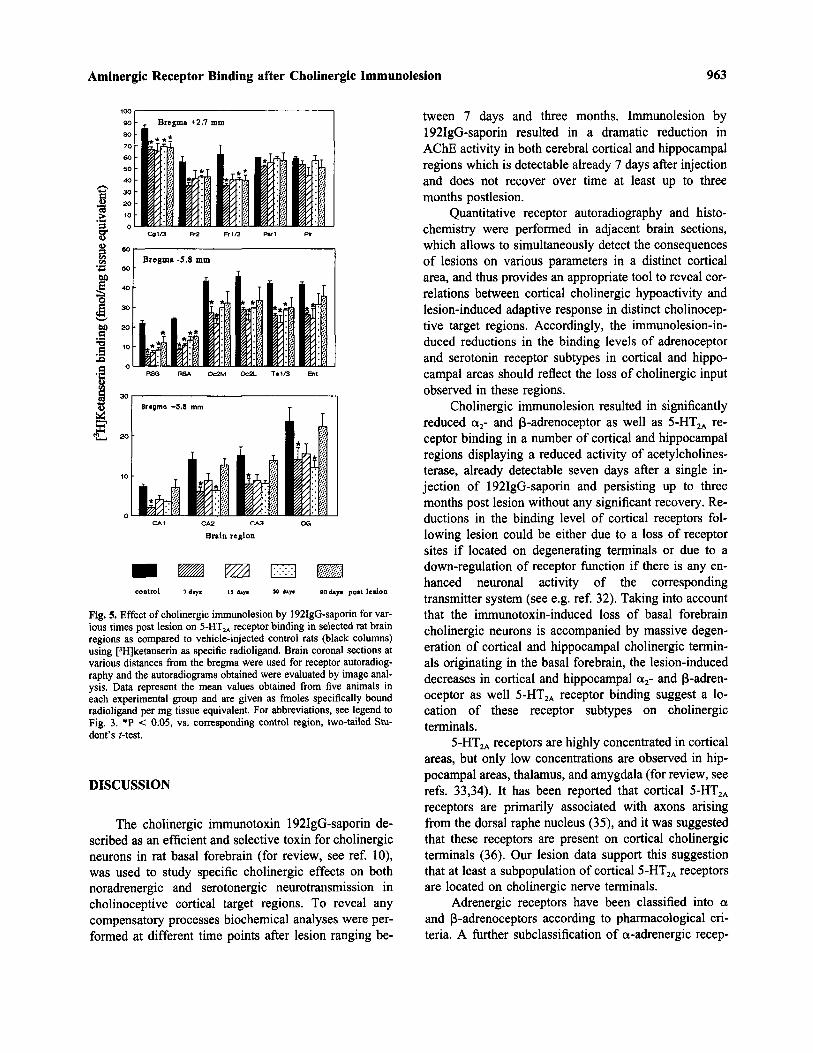
Already seven days after basal forebrain cholinergiclesion, the binding levels of B-adrenoceptors in the cin-gulate, frontal, parietal and piriform cortex were consid-erably decreased by about 50% as compared to corre-sponding control values. The reductions in B-adrenocep-tor binding levels in these regions are seemingly ofpermanent nature, because three months after a single in-jection of 192IgG-saporin the binding levels were stilldecreased to the same extent as observed seven days afterlesion (Fig. 4). Similarly, B-adenoceptor binding was sig-nificantly decreased in the subfields CA1 (by about 30%),CA2 (27%), CA3 (53%), CA4 (53%) and dentate gyrus(37%) of the hippocampal formation seven days after im-munolesion (Fig. 4). Decreased binding levels but to alesser extent are still detectable three months after lesionin the CA1 (by 24%), CA2 (16%), CA3 (35%), CA4(20%), but no longer in the dentate gyrus relative to con-trol values (Fig. 4).
Effect of Cholinergic Immunolesion on Seroto-nergic Receptor Subtypes. Similar to our findings on a,-adrenoceptor binding, cholinergic basal forebrain lesionby 192IgG-saporin did not affect 5-HT1A receptor bind-ing in any of the cortical and hippocampal areas studiedregardless of the length of the survival time after lesion,at least up to three months post lesion (data not shown).Contrasting to that, already seven days after cholinergicimmunolesion [3H]ketanserin binding to the 5-HT2A re-ceptor was decreased in the cingulate (by 26%), frontal(34%), parietal (27%), piriform (37%), temporal (38%),retrosplenial (64%), occipital (37%), and entorhinal (36%)cortices (Fig. 5). Assaying 5-HT2A receptor binding in theseregions 15, 30, and 90 days after cholinergic immunole-sion, the binding levels remain decreased to nearly thesame extent as detected seven days after lesion (Fig. 5). Incontrast, in the subfields CA1 through CA3 of the hippo-campal formation and dentate gyrus the levels of 5-HT2A
binding sites were found to be reduced by about 60%seven days after lesion, but three months after lesion thelevels of [3H]ketanserin binding were no longer differentto corresponding control values (Fig. 5).
Fig. 4. Effect of cholinergic immunolesion by 192IgG-saporin for var-ious times post lesion on B-adrenoceptor binding in selected rat brainregions as compared to vehicle-injected control rats (black columns)using [3H]dihydroalprenolol as specific radioligand. Brain coronal sec-tions at various distances from the bregma were used for receptorautoradiography and the autoradiograms obtained were evaluated byimage analysis. Data represent the mean values obtained from fiveanimals in each experimental group and are given as fmoles specifi-cally bound radioligand per mg tissue equivalent. For abbreviations,see legend to Fig. 3. *P < 0.05, vs. corresponding control region, two-tailed Student's Mest.
creased by around 20% in comparison to controls but toa lesser extent as observed seven days after lesion. How-ever, in the occipital cortex area 2, temporal and entor-hinal cortex, cholinergic lesion for longer periods oftime resulted in a normalization of a2-adrenoceptor bind-
Aminergic Receptor Binding after Cholinergic Immunolesion 963
tween 7 days and three months. Immunolesion by192IgG-saporin resulted in a dramatic reduction inAChE activity in both cerebral cortical and hippocampalregions which is detectable already 7 days after injectionand does not recover over time at least up to threemonths postlesion.
Quantitative receptor autoradiography and histo-chemistry were performed in adjacent brain sections,which allows to simultaneously detect the consequencesof lesions on various parameters in a distinct corticalarea, and thus provides an appropriate tool to reveal cor-relations between cortical cholinergic hypoactivity andlesion-induced adaptive response in distinct cholinocep-tive target regions. Accordingly, the immunolesion-in-duced reductions in the binding levels of adrenoceptorand serotonin receptor subtypes in cortical and hippo-campal areas should reflect the loss of cholinergic inputobserved in these regions.
Cholinergic immunolesion resulted in significantlyreduced a2- and B-adrenoceptor as well as 5-HT2A re-ceptor binding in a number of cortical and hippocampalregions displaying a reduced activity of acetylcholines-terase, already detectable seven days after a single in-jection of 192IgG-saporin and persisting up to threemonths post lesion without any significant recovery. Re-ductions in the binding level of cortical receptors fol-lowing lesion could be either due to a loss of receptorsites if located on degenerating terminals or due to adown-regulation of receptor function if there is any en-hanced neuronal activity of the correspondingtransmitter system (see e.g. ref. 32). Taking into accountthat the immunotoxin-induced loss of basal forebraincholinergic neurons is accompanied by massive degen-eration of cortical and hippocampal cholinergic termin-als originating in the basal forebrain, the lesion-induceddecreases in cortical and hippocampal a2- and p-adren-oceptor as well 5-HT2A receptor binding suggest a lo-cation of these receptor subtypes on cholinergicterminals.
5-HT2A receptors are highly concentrated in corticalareas, but only low concentrations are observed in hip-pocampal areas, thalamus, and amygdala (for review, seerefs. 33,34). It has been reported that cortical 5-HT2A
receptors are primarily associated with axons arisingfrom the dorsal raphe nucleus (35), and it was suggestedthat these receptors are present on cortical cholinergicterminals (36). Our lesion data support this suggestionthat at least a subpopulation of cortical 5-HT2A receptorsare located on cholinergic nerve terminals.
Adrenergic receptors have been classified into aand p-adrenoceptors according to pharmacological cri-teria. A further subclassification of a-adrenergic recep-
Fig. 5. Effect of cholinergic immunolesion by 192IgG-saporin for var-ious times post lesion on 5-HT2A receptor binding in selected rat brainregions as compared to vehicle-injected control rats (black columns)using [3H]ketanserin as specific radioligand. Brain coronal sections atvarious distances from the bregma were used for receptor autoradiog-raphy and the autoradiograms obtained were evaluated by image anal-ysis. Data represent the mean values obtained from five animals ineach experimental group and are given as fmoles specifically boundradioligand per mg tissue equivalent. For abbreviations, see legend toFig. 3. *P < 0.05, vs. corresponding control region, two-tailed Stu-dent's Mest.
DISCUSSION
The cholinergic immunotoxin 192IgG-saporin de-scribed as an efficient and selective toxin for cholinergicneurons in rat basal forebrain (for review, see ref. 10),was used to study specific cholinergic effects on bothnoradrenergic and serotonergic neurotransmission incholinoceptive cortical target regions. To reveal anycompensatory processes biochemical analyses were per-formed at different time points after lesion ranging be-

964 Heider, Schliebs, RoBner, and Bigl
tors was first made on the basis of the anatomic location:a1-adrenoceptors were assumed to occur postsynaptically,while a2-adrenoceptors were postulated to exist presy-naptically and to regulate the release of noradrenaline (seee.g. ref. 37). However, noradrenergic lesion experimentshave revealed that a2-adrenoceptors located on noradre-nergic nerve terminals may represent only a minor portionof the total a2-adrenoceptors in rat cerebral cortex (38).Our lesion data provide strong evidence that at least asubpopulation of a2-adrenoceptors may act also as het-eroreceptor on cortical cholinergic nerve terminals a2-ad-renoceptors have been further classified into a2A, a2B, anda2C subtypes (for review, see ref. 39). Recent noradre-nergic lesion experiments provided evidence that a2C ad-renoceptors occur postsynaptically to noradrenergicneurons, while a2A receptors may exist also on non-no-radrenergic terminals in the cortex both pre- and post-synaptically (40). The radioligand used in this studydoes not differentiate between a2-adrenoceptor subtypes.But based on the fact that 90% of the a2-adrenoceptorsin rat cerebral cortex are of the a2A subtype, and theremaining 10% are most likely of the postsynaptic a2C
subtype (40), the lesion-induced decrease in a2-adreno-ceptor binding observed here should mainly reflect theloss of a2A-adrenoceptor subtype.
[3H]Dihydroalprenolol used in this study detectsboth B1- and B2-adrenoceptors. p1-adrenoceptors pre-dominate in forebrain structures such as cerebral cortex,striatum and hippocampus, while (32-adrenoceptors aremore prominent in the cerebellum (39). Therefore, thelesion-induced decrease in [3H]dihydroalprenolol bind-ing in a number of cortical and hippocampal regionsshould mainly reflect changes in the fi1-adrenoceptorsubtype. Noradrenergic denervation studies revealed thatP-adrenoceptors are preferentially located postsynapti-cally with respect to noradrenergic terminals (see e.g.ref. 41). However, cortical P-adrenoceptors are not onlypresent on neurons but were detected also on astrocytes(42). The data presented in this study provide evidencethat a subpopulation of B-adrenoceptors may also existon cortical cholinergic terminals originating in the basalforebrain.
The lesion-induced decrease in a2-, B-, and 5-HT2A
receptor binding could also be considered as a receptordown-regulation and/or subsensitivity. Down-regulationof postsynaptic receptors is assumed to compensate forenhanced presynaptic input. Basal forebrain cholinergiccell loss may affect interactions of the cholinergic sys-tem with the serotonergic and noradrenergic neurotrans-mission which might result in a dysbalance andoveractivation of the non-lesioned systems. Possiblesites for such interactive processes could occur at the
level of convergent projection areas of the two transmit-ter systems, or by direct contacts between cholinergic basalforebrain and serotonergic raphe nucleus and noradrenergiclocus coeruleus neurons. It is well established that the basalforebrain cholinergic nuclei receive projections from themedial and dorsal raphe nuclei (43,44). The loss of basalforebrain cholinergic neurons that are innervated by raphenucleus serotonergic cells could result in an imbalance be-tween cholinergic and serotonergic system leading to anenhancement of cortical serotonergic activity, which mightexplain a down-regulation or subsensitivity of postsynapticserotonergic receptors.
The cerebral cortex receives a prominent noradre-nergic input from cells located within the locus coeruleus.Basal forebrain structures including the nucleus basalis donot provide a cholinergic innervation to the locus coeru-leus (45,46). Therefore, degeneration of cholinergic basalforebrain neurons does not provide any contribution toinfluence noradrenergic activity in the cerebral cortexthrough the locus coeruleus. However, there is a norad-renergic modulation of cholinergic nucleus basalis neu-rons (47), presumably mediated by some locus coeruleusneurons which send collaterals to basal forebrain nuclei(48). Noradrenergic cells could thereby influence corticalactivity by acting upon the cortically projecting cholin-ergic basal forebrain neurons. Whether the immunotoxin-induced loss of cholinergic cells innervated by locuscoeruleus neurons leads to any noradrenergic hyperactiv-ity in the cerebral cortex and thereby to a down-regulationof cortical postsynaptic a2- and p-adrenoceptors mustremain a matter of speculation at this stage of investiga-tion. But interactive processes may occur also at the re-ceptor level, e.g. the modulation of p-adrenoceptor bind-ing characteristics by activation of a2-adrenoceptors in thecerebral cortex as recently demonstrated (49). However,cholinergic immunolesion by 192IgG-saporin did not af-fect the tissue levels of noradrenaline and serotonin in anyof the cortical or hippocampal regions studied at least upto one month post lesion (50), a finding which does notfavour the assumption of any enhanced cortical seroto-nergic and/or noradrenergic activity due to cholinergiccell loss. This further supports our suggestion that thelesion-induced reductions in a2-, P-, and 5-HT2A receptorbinding represent the loss of receptors located on cholin-ergic terminals, rather than being due to receptor subsen-sitivity/down-regulation.
Postsynaptic a1-adrenoceptors seem to be relativelyinsensitive to noradrenaline depletion by specific neu-rotoxins and by specific noradrenergic denervation (51-53) which may explain why cortical a1-adrenoceptorsare unaffected by basal forebrain cholinergic lesion.Cholinergic immunolesion also did not alter cortical 5-

Aminergic Receptor Binding after Cholinergic Immunolesion 965
HT1A receptor binding although they are highly concen-trated in limbic areas such as neocortex and hippocam-pus (33). 5-HT1A receptors are supposed to act asautoreceptors in the raphe nuclei. In the cerebral cortexthey are assumed to be localized postsynaptically withrespect to serotonergic terminals, but may also be situ-ated on presynaptic elements of other neuronal catego-ries (33). Our lesion study provides strong evidence thatboth a1-adrenergic and 5-HT1A receptors are not locatedon cortical cholinergic nerve terminals that originate inthe basal forebrain.
In this study receptor autoradiographies were per-formed at single ligand concentrations only, so changesin binding sites may, therefore, be due to alterations inboth maximum receptor number and/or in receptor affin-ity. In a current neurochemical study we could show thatthe 192IgG-saporin-induced changes in cortical musca-rinic acetylcholine receptor binding are mainly due toalterations in receptor number (21). Whether this is alsotrue for the lesion-induced changes in adrenoceptor andserotonergic receptor subtypes cannot be concluded fromthis study. However, regardless of possible interpreta-tions the immunotoxin-induced changes in adrenoceptorand 5-HT receptor subtypes in cortical regions display-ing reduced cholinergic activity clearly demonstrate on-going interactive processes at the level of convergentprojection areas of the two transmitter systems with thecholinergic one. Moreover, it is interesting to note thatthe same sort of alterations in a2- and 5-HT2A receptorsubtypes observed in rat cortex following basal forebraincholinergic imrmmolesion have been detected in corticalbrain areas from patients with Alzheimer's disease(1,54), suggesting that the reduction in cortical a2- and5-HT2A receptor binding observed in these patients aresecondary to the cholinergic deficits. The data availableon B-adrenoceptors in cortical areas from Alzheimerbrains are controversial demonstrating both increase anddecrease in cortical and hippocampal B-receptor bindingas compared to age-matched controls (55,56, for review,see ref. 1). However, our data also support an aminergicstrategy which might have therapeutic potential in treat-ing Alzheimer's disease, and demonstrate that choliner-gic immunolesion by 192IgG-saporin exhibits a valuabletool to produce specific cholinergic deficits in rats as amodel to study the effect of treatment with variousdrugs.
ACKNOWLEDGMENT
The authors gratefully acknowledge the expert technical assis-tance of Mrs. Renate Jendrek. This work was supported by a grant
from the Bundesministerium fur Forschung und Technik, no. FKZ 01ZZ 9130/2.8 to R.S.
REFERENCES
1. Norberg, A. 1992. Neuroreceptor changes in Alzheimer disease.Cerebrovasc. Brain Met. Rev. 4:303-328.
2. Airman, H. J., Normile, H. J., Galloway, M. P., Ramirez, A., andAzmitia, E. C. 1990. Enhanced spatial discrimination learning inrats following 5,7-DHT induced serotonergic deafferentation ofthe hippocampus. Brain Res. 518:61-66.
3. Bickford, P., Heron, C., Young, D. A., Gerhardt, G. A., and Dela Garza, R. 1992. Impaired acquisition of novel locomotor tasksin aged and norepinephrine-depleted F344 rats. Neurobiol. Aging13:475-481.
4. De Noble, V. J., Schrack, L. M., Reigel, A. L., and De Noble, K.F. 1991. Visual recognition memory in squirrel monkeys: Effectsof serotonin antagonists on baseline and hypoxia-induced per-formance deficits. Pharmacol. Biochem. Behav. 39:991-996.
5. Jackson, W. J., and Buccafusco, J. J. 1991. Clonidine enhancesdelayed matching-to-sample performance by young and agedmonkeys. Pharmacol. Biochem. Behav. 39:79-84.
6. McEntee, W. J., and Crook. T. H. 1991. Serotonin, memory, andthe aging brain. Psychopharmacology (Berl.) 103:143-149.
7. Sleekier, T., and Sahgal, A. 1995. The role of serotonergic-cho-linergic interactions in the mediation of cognitive behaviour. Be-hav. Brain Res. 67:165-199.
8. Arnstein A. F. T., Cai J. X., and Goldman-Rakic P.S. 1988. Thealpha 2 adrenergic agonist guanfacine improves memory in agedmonkeys without sedative or hypotensive side effects: Evidencefor alpha-2 receptor subtypes. J. Neurosci. 8, 4287-2498.
9. Strek, K. F., Spencer, K. R., and De Noble, V. J. 1989. Manip-ulation of serotonin protects against hypoxia-induced deficits of apassive avoidance response in rats. Pharmacol. Biochem. Behav.33:241-244.
10. Schliebs, R., and Bigl, V. 1996. Animal models to produce corticalcholinergic dysfunction. Pages 275-289. in Perez-Polo, R.J. (ed.),Methods in Neurosciences, vol. 30, Academic Press, New York.
11. Wiley, R. G., Oeltmann, T. N., and Lappi, D. A. 1991. Immu-nolesioning: selective destruction of neurons using immunotoxinto rat NGF receptor. Brain Res. 562:149-153.
12. Book, A. A., Wiley, R. G., and Schweitzer, J. B. 1992. Specificityof 192 IgG-saporin for NGF receptor-positive cholinergic basalforebrain neurons in the rat. Brain Res. 590:350-355.
13. Heckers, S., Ohtake, T., Wiley, R. G., Lappi, D. A., Geula, C.,and Mesulam, M.M. 1994. Complete and selective denervation ofrat neocortex and hippocampus but not amygdala by an immu-notoxin against the p75 NGF receptor. J. Neurosci. 14:1271-1289.
14. Roflner, S., Hartig, W., Schliebs, R., Bruckner, G., Brauer, K.,Perez-Polo, J. R., Wiley, R. G., and Bigl, V. 1995. 192IgG-saporinimmunotoxin-induced loss of cholinergic cells differentiallyactivates microglia in rat basal forebrain nuclei. J. Neurosci. Res.41:335-346.
15. RoBner, S., Yu, J., Pizzo, D., Werrbach-Perez, K., Schliebs, R., Bigl,V., and Perez-Polo, J.R. 1996. Effects of intraventricular transplan-tation of NGF-secreting cells on cholinergic basal forebrain neuronsafter partial immunolesion. J. Neurosci Res. 45:40-56.
16. Lee, M. G., Chrobak, J. J., Sik, A., Wiley, R. G., and Buzsaki,G. 1994. Hippocampal theta activity following selective lesion ofthe septal cholinergic system. Neuroscience 62:1033-1047.
17. Torres, E. M., Perry, T. A., Blokland, A., Wilkinson, L. S., Wiley,R. G., Lappi, D. A., and Dunnett, S. B. 1994. Behavioural, his-tochemical and biochemical consequences of selective immuno-lesions in discrete regions of the basal forebrain cholinergicsystem. Neuroscience 63:95-122.
18. Wenk, G. L., Stoehr, J. D., Quintana, G., Mobley, S., and Wiley,R. G. 1994. Behavioral, biochemical, histological, and electrophy-

966 Heider, Schliebs, RoBner, and Bigl
siological effects of 192 IgG-saporin injections into the basal fore-brain of rats. J. Neurosci. 14:5986-5995.
19. RoBner, S., Perez-Polo, J. R., Wiley, R. G., Schliebs, R., and Bigl,V. 1994. Differential expression of immediate early genes in dis-tinct layers of rat cerebral cortex after selective immunolesion ofthe forebrain cholinergic system. J. Neurosci. Res. 38:282-293.
20. RoBner, S., Schliebs, R., Perez-Polo, J. R., Wiley, R. G., and Bigl,V. 1995. Differential changes in cholinergic markers from selectedbrain regions after specific immunolesion of rat cholinergic basalforebrain system. J. Neurosci. Res. 40:31-43.
21. RoBner, S., Schliebs, R., Hartig, W., and Bigl, V. 1995. 192IgG-saporin-induced selective lesion of cholinergic basal forebrain sys-tem: neurochemical effects on cholinergic neurotransmission in ratcerebral cortex and hippocampus. Brain Res. Bull. 38:371-381.
22. Schwartz, J. C., Cortes, C. L., Rose, C., Quach, T. T., and Pollard,H. 1983. Adaptive changes of neurotransmitter receptor mechanismsin the central nervous system. Progr. Brain Res. 58:117-129.
23. Andra, J., and Lojda, Z. 1986. A histochemical method for thedemonstration of acetylcholinesterase activity using semipermea-ble membranes. Histochemistry 84, 575-579.
24. Rainbow, T. C., and Biegon, A. 1983. Quantitative autoradiog-raphy of [3H]prazosin binding sites in rat forebrain. Neurosci. Lett.40:221-226.
25. Noyer, M., de Laveleye, F., Vauquelin, G., Gobert, J., and Wiil-fert, E. 1994. Mivarazol, a novel compound with high spezifityfor a2-adrenergic receptors: binding studies on different humanand rat membrane preparations. Neurochem. Int. 24:221-229.
26. De Vos, H., Convents, A., De Keyser, J., De Backer, J. -P., VanMegen, I. J. B., Ebinger, G., and Vauquelin, G. 1991. Autoradi-ographic distribution of a2-adrenoceptors, NAIBS, and 5-HT1A re-ceptors in human brain using [3H]idazoxan and [3H]rauwolscine.Brain Res. 566:13-20.
27. Schliebs, R., Walch, C., and Stewart, M. G. 1989. Laminar patternof cholinergic and adrenergic receptors in rat visual cortex usingquantitative receptor autoradiography. J. Hirnforsch. 30:303-311.
28. Dyck, R. H., and Cynader, M. S. 1993. Autoradiographic locali-zation of serotoninreceptor subtypes in cat visual cortex: transientregional, laminar and columnar distributions during postnatal de-velopment. J. Neurosci. 13:4316—4338.
29. Dewar, D., Graham, D. I., and McCulloch, J. 1990. 5-HT2 recep-tors in dementia of the Alzheimer type: a quantitative autoradi-ographic study of frontal cortex and hippocampus. J. NeuralTransm. [P-D Sect.] 2:129-137.
30. Kumar, A., and Schliebs, R. 1992. Postnatal development of cho-linergic receptors, protein kinase C and dihydropyridine-sensitivecalcium antagonist binding in rat visual cortex. Effect of visualdeprivation. Int. J. Devl. Neurosci. 10:491-504.
31. Zilles, K. 1985. The cortex of the rat. A stereotaxic atlas. Springer,Berlin.
32. Klein, W. L., Sullivan, J., Skorupa, A., and Aguilar, J. S. 1989.Plasticity of neuronal receptors. FASEB J. 3:2132-2140.
33. Radja, F., Laporte, A.-M., Daval, G., Verge, D., Gozlan, H., andHamon, M. 1991. Autoradiography of serotonin receptor subtypesin the central nervous system. Neurochem. Int. 18:1-15.
34. Palacios, J. M., Waeber, C., Mengod, G., and Hoyer, D. 1991.Autoradiography of 5-HT receptors: A critical appraisal. Neuro-chem. Int. 18:17-25.
35. Blue, M. E., Yagaloff, K. A., Mamounas, L. A., Hartig, P. R., andMolliver, M. E. 1988. Correspondence between 5-HT2 receptorand serotonergic axons in rat neocortex. Brain Res. 453:315-328.
36. Quirion, R., Richard, J., and Dam, T. V. 1985. Evidence for theexistence of serotonin type 2 receptors on cholinergic terminals inrat cortex. Brain Res. 333:345-349.
37. Bylund, D. B. 1992. Subtypes of al- and a2-adrenergic receptorsFASEB J. 6:832-839.
38. Grofi, G., Gothert, M., Glapa, U., Engel, G., and Schumann, H.-J. 1985. Lesioning of serotoninergic and noradrenergic fibres of
the rat brain does not decrease binding of 3H-clonidine and 3H-rauwolscine to cortical membranes. Naunyn Schmiedebergs Arch.Pharmacol. 328:229-235.
39. Nicholas, A. P., Hokfelt, T., and Pieribone, V. A. 1996. The dis-tribution and significance of CNS adrenoceptors examined with insitu hybridization. Trends Pharmacol. Sci. 17:245-255.
40. Ordway, G. A. 1995. Effect of noradrenergic lesions on subtypesof ct2-adrenoceptors in rat brain. J. Neurochem. 64:1118-1126.
41. Wolfman, C., Abo, V., Calvo, D., Medina, J., Dajas, F., and Sil-veira, R. 1994. Recovery of central noradrenergic neurons oneyear after the administration of the neurotoxin DSP4. Neurochem.Int. 25:395-400.
42. Stone, E. A., and Jone, S. M. 1991. Further evidence for a gliallocalization of rat cortical fi-adrenoceptors: studies of in vivo cy-clic AMP responses to catecholamines. Brain Res. 549:78-82.
43. Milner, T. A., and Veznedaroglu, E. 1993. Serotonin-containingterminals synapse on septohippocampal neurons in the rat. J. Neu-rosci. Res. 36:260-271.
44. Tork, I. 1985. Raphe nuclei and serotonin containing systems.Pages 43-78, in Paxinos, G. (ed.) The Rat Nervous System. Vol.2: Hindbrain and Spinal Cord, Academic Press, Sydney.
45. Aston-Jones, G., Ennis, M., Pieribone, V. A., Nickel, W. T., andShipley, M. T. 1986. The brain nucleus locus coeruleus: restrictedafferent control of a broad efferent network. Science 234:734-737.
46. Woolf, N. J., and Butcher, L. L. 1989. Cholinergic systems in therat brain. IV. Descending projections of the pontomesencephalictegmentum. Brain Res. Bull. 22:519-540.
47. Fort, P., Khateb, A., Pegna, A., Muhlethaler, M., and Jones, B. E.1995. Noradrenergic modulation of cholinergic nucleus basalisneurons demonstrated by in vitro pharmacological and immunoh-istochemical evidence in the guinea-pig brain. Eur. J. Neurosci. 7:1502-1511.
48. Jones, B. E., and Yang, T. Z. 1985. The efferent projections fromthe reticular formation and the locus coeruleus studied by anter-ograde and retrograde axonal transport in the rat. J. Comp. Neurol.242:56-92.
49. Nakamura, T., Tsujimura, R., and Nomura, J. 1991. Interactionbetween a2- and B-adrenergic receptors in rat cerebral corticalmembranes: clonidine-induced reduction in agonist and antagonistaffinity for P-adrenergic receptors. Brain Res. 542:181-186.
50. Waite, J. J., Wardlow, M. L., Chen, A. C., Lappi, D. A., Wiley,R. G., and Thal, L. J. 1994. Time course of cholinergic and mon-oaminergic changes in rat brain after immunolesioning with192IgG-saporin. Neurosci. Lett. 169:154-158.
51. Dooley, D. J., Bittiger, H., Hauser, K. L., Bischoff, S., and Wald-meier, P. C. 1983. Alteration of central alphal- and beta-adrener-gic receptors in the rat after DSP-4, a selective noradrenergicneurotoxin. Neuroscience 9:889-898.
52. Reader, T. A., and Briere, R. 1983. Selective noradrenergic de-nervation and 3H-prazosin binding sites in rat-neocortex. BrainRes. Bull. 10:155-158.
53. Sharma, V. K., Harik, S. I., Busto, R., and Banerjee, S. P. 1981.Effects of noradrenaline depletion on the cerebral cortex, hippo-campus, and cerebellum. Exp. Neurol. 72:179-194.
54. Kalaria, R. N., and Andorn, A. C. 1991. Adrenergic receptors inaging and Alzheimer's disease: Decreased a2-receptors demon-strated by [3H]p-aminoclonidine binding in prefrontal cortex. Neu-robiol. Aging 12:131-136.
55. Kalaria, R. N., Andorn, A. C., Tabaton, M., Whitehouse, P.J.,Harik, S.I., and Unnerstall, J.R. 1989. Adrenergic receptors in ag-ing and Alzheimer's disease: Increased p52-receptors in prefrontalcortex and hippocampus. J. Neurochem. 53:1772-1781.
56. Shimohama, S., Taniguchi, T., Fujiwara, M., and Kameyama, M.1987. Changes in P-adrenergic receptor subtypes in Alzheimer-type dementia. J. Neurochem. 48:1215-1221.



![05 Cholinergic drugs - Western · 2019-07-31 · Z H d I k N V c ] : 7 ^ X : d ] g L ] k G 4 V c M Z L 4 e V o V U h L 4 e V ] ` L L 4 d [ 4 i \ e ] e T e V I ` K M g e U M ` 4 =](https://img.pdfslide.tips/doc/110x75/5f04878d7e708231d40e6c79/05-cholinergic-drugs-western-2019-07-31-z-h-d-i-k-n-v-c-7-x-d-g-l.jpg)

























