Embed Size (px)
Citation preview

1
BIOQUÍMICA HUMANA
REGULACIÓN METABÓLICA
(Clase Nº 20)

2
Nuestro organismo debe satisfacer diversos requerimientos metabólicos esenciales: sintetizar
todos los componentes que las células necesitan, proteger nuestro medio interno de toxinas y
adaptarse a las condiciones cambiantes del medio externo. Para cumplir con estos requisitos,
transformamos los componentes de la dieta mediante el metabolismo oxidativo, el almacenamiento y
movilización de moléculas combustibles, las vías biosintéticas y la detoxificación o eliminación de los
compuestos residuales de las diferentes vías metabólicas.
El organismo debe mantener un balance entre las necesidades de las células y la
disponibilidad de los combustibles, lo que se denomina homeostasis metabólica. La disponibilidad
constante de combustibles en la sangre se denomina homeostasis calórica, mediante la cual el nivel
sanguíneo de combustibles (en equivalentes de ATP) no disminuye por debajo de ciertos límites,
independientemente de si el individuo se encuentra en un estado de buena nutrición o ayuno.
El mantenimiento de la homeostasis metabólica se logra mediante la integración de tres factores
principales:
1) La concentración de nutrientes en la sangre, que afecta la velocidad con la cual éstos son
utilizados y almacenados en los diferentes tejidos,
2) los niveles de hormonas en sangre (primeros mensajeros), que transmiten información a tejidos
específicos sobre el estado del organismo y el aporte o demanda de nutrientes,
3) el sistema nervioso central que por medio de señales neurales, controla el metabolismo
directamente o a través de la liberación de hormonas.
Los nutrientes que utiliza nuestro organismo son los carbohidratos, los lípidos y las proteínas,
aproximadamente en las siguientes cantidades:
Una persona normal que lleva una vida sedentaria consume diariamente alrededor de 200 g de
glúcidos, 70 g de proteína y 60 g de lípidos, lo que le permite afrontar un requerimiento energético de
1600-2400 kcal. Como se observa en la tabla anterior, desde el punto de vista energético, el
principal combustible metabólico son los ácidos grasos.
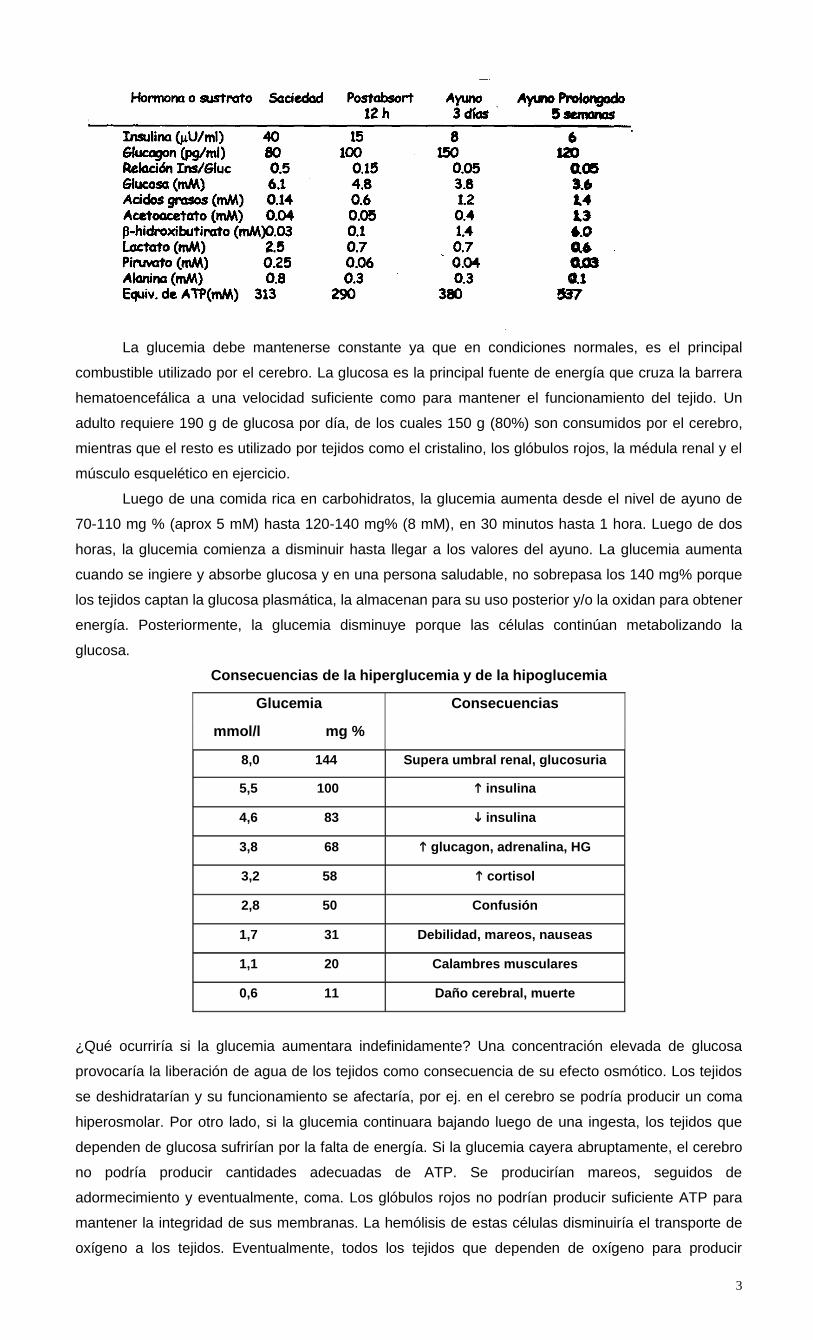
Cuando se estudia la variación en los niveles de nutrientes en las diferentes etapas que van
desde la saciedad hasta el ayuno se observa que mientras que la cetonemia y la concentración de
ácidos grasos en sangre puede variar entre 1 y 2 órdenes de magnitud, la concentración de glucosa
se mantiene dentro de límites muy ajustados.
3
La glucemia debe mantenerse constante ya que en condiciones normales, es el principal
combustible utilizado por el cerebro. La glucosa es la principal fuente de energía que cruza la barrera
hematoencefálica a una velocidad suficiente como para mantener el funcionamiento del tejido. Un
adulto requiere 190 g de glucosa por día, de los cuales 150 g (80%) son consumidos por el cerebro,
mientras que el resto es utilizado por tejidos como el cristalino, los glóbulos rojos, la médula renal y el
músculo esquelético en ejercicio.
Luego de una comida rica en carbohidratos, la glucemia aumenta desde el nivel de ayuno de
70-110 mg % (aprox 5 mM) hasta 120-140 mg% (8 mM), en 30 minutos hasta 1 hora. Luego de dos
horas, la glucemia comienza a disminuir hasta llegar a los valores del ayuno. La glucemia aumenta
cuando se ingiere y absorbe glucosa y en una persona saludable, no sobrepasa los 140 mg% porque
los tejidos captan la glucosa plasmática, la almacenan para su uso posterior y/o la oxidan para obtener
energía. Posteriormente, la glucemia disminuye porque las células continúan metabolizando la
glucosa.
Consecuencias de la hiperglucemia y de la hipoglucemia
Glucemia Consecuencias
mmol/l mg %
8,0 144 Supera umbral renal, glucosuria
5,5 100 insulina
4,6 83 insulina
3,8 68 glucagon, adrenalina, HG
3,2 58 cortisol
2,8 50 Confusión
1,7 31 Debilidad, mareos, nauseas
1,1 20 Calambres musculares
0,6 11 Daño cerebral, muerte
¿Qué ocurriría si la glucemia aumentara indefinidamente? Una concentración elevada de glucosa
provocaría la liberación de agua de los tejidos como consecuencia de su efecto osmótico. Los tejidos
se deshidratarían y su funcionamiento se afectaría, por ej. en el cerebro se podría producir un coma
hiperosmolar. Por otro lado, si la glucemia continuara bajando luego de una ingesta, los tejidos que
dependen de glucosa sufrirían por la falta de energía. Si la glucemia cayera abruptamente, el cerebro
no podría producir cantidades adecuadas de ATP. Se producirían mareos, seguidos de
adormecimiento y eventualmente, coma. Los glóbulos rojos no podrían producir suficiente ATP para
mantener la integridad de sus membranas. La hemólisis de estas células disminuiría el transporte de
oxígeno a los tejidos. Eventualmente, todos los tejidos que dependen de oxígeno para producir
4
energía fallarían. Si el problema fuera suficientemente severo, podría provocar la muerte del individuo.
Estas consecuencias del exceso o deficiencia de glucosa se evitan porque el organismo es
capaz de regular la glucemia. Cuando la concentración de glucosa en sangre se aproxima al valor
normal del ayuno de 70 a 110 mg%, alrededor de 2 horas después de la ingesta, se activa la
glucogenolisis en el hígado. El glucógeno hepático es la fuente principal de glucosa plasmática
durante las primeras horas del ayuno y luego, la gluconeogénesis empieza a ser importante como
fuente de glucosa. Los precursores utilizados en la glucogeneogénesis hepática son aportados por
otros tejidos. El músculo en ejercicio y los glóbulos rojos producen lactato en la glucólisis. La
degradación de proteínas en el músculo provee aminoácidos y el tejido adiposo aporta glicerol cuando
se movilizan los depósitos de triacilglicéridos (TAG). Aún durante el ayuno prolongado, la glucemia no
cae dramáticamente. Luego de 5 a 6 semanas de ayuno, la glucemia aún se encuentra en valores de
alrededor de 65 mg%.
Reservas energéticas en el humano
A pesar de su rol esencial en el metabolismo energético del cerebro y otros tejidos, la
cantidad de glucosa circulante es limitada. Para asegurar su continua provisión, el organismo
almacena combustibles metabólicos para proveer glucosa o energía en caso de necesidad.
Combustible tisular
Reserva en g
Provee energía durante
Ayuno Caminando Maratón
Glucosa
sanguínea
20
40 min
5 min
4 min
Glucógeno hepático 80 3,5 h 70 min 18 min
Glucógeno Muscular 150 14 h 5 h 70 min
Lípidos 9000 – 15000 34 días 11 días 3 días
Proteínas 6000 15 días 5 días 1,3 días
Dentro de los mecanismos homeostáticos que permiten regular la disponibilidad de moléculas
combustibles, el control hormonal es uno de los más importantes. La insulina y el glucagon son las
principales hormonas que regulan el almacenamiento y la utilización de combustibles. La insulina es
una hormona anabólica que promueve el almacenamiento de combustibles, mientras que el glucagon
es la hormona que estimula la movilización de combustibles. Otras hormonas como la adrenalina, son
liberadas como respuesta del SNC a la hipoglucemia, al ejercicio y a otros tipos de estrés fisiológico.
Junto con otras hormonas del estrés (glucocorticoides), la adrenalina aumenta la disponibilidad de
combustibles.
Los principales mecanismos que modifican la velocidad de una vía metabólica a través de la
regulación de la actividad de las enzimas clave de las mismas son:
1. Disponibilidad de sustrato
2. Compartimentación celular
3. Modificación alostérica.
4. Modificación covalente
5. Inducción y represión enzimáticas

5
1.- Disponibilidad de sustrato:
La concentración de sustrato puede modificar significativamente la velocidad de una reacción
enzimática. De esa forma, fluctuaciones en la concentración de los sustratos proveen un mecanismo
automático de ajuste de la velocidad de una reacción enzimática a las circunstancias metabólicas
particulares. Un ejemplo de este tipo de regulación es el que ocurre cuando se ingiere una dieta rica
en glúcidos con la actividad de la glucoquinasa hepática. Los cambios en las velocidades de las
reacciones enzimáticas que ocurren en virtud de cambios en la disponibilidad de sustrato son rápidos
ya que la fluctuación en la concentración de sustratos influye directamente en la actividad catalítica de
las enzimas involucradas, sin que medie ningún otro mecanismo adicional.
2.- Compartimentalización celular:
La velocidad del flujo de una vía metabólica también se regula por la accesibilidad de los
sustratos al compartimiento celular en que se encuentran las enzimas de la vía. En este tipo de
regulación es fundamental el mecanismo de transporte de los metabolitos a través de las membranas
celulares y subcelulares. La beta-oxidación de ácidos grasos ocurre en las mitocondrias y por lo tanto
los acidos grasos deben atravesar la membrana mitocondrial interna para ser oxidados. Esta vía
metabólica, por lo tanto, está fuertemente regulada por la velocidad de transporte de los AG a la
mitocondria.
3.- Modificación alostérica:
Se produce cuando se altera la capacidad catalítica de una enzima (generalmente con
estructura cuaternaria) como consecuencia de un cambio en su conformación, inducido por un
metabolito (modificador alostérico). Es un mecanismo rápido de modificación de la actividad
enzimática.
4.- Modificación covalente:
Otro tipo de modificación de la actividad enzimática se produce cuando la enzima une
covalentemente un grupo químico, lo que puede provocar una profunda alteración en su actividad
catalítica. La situación puede revertirse, es decir, la enzima puede perder el grupo unido y retomar la
actividad anterior. La regulación por modificación covalente da como resultado cambios rápidos de la
actividad enzimática, debido fundamentalmente a que el agregado o eliminación del grupo es también
catalizado por enzimas. Este mecanismo de regulación enzimática ocurre en respuesta a una acción
hormonal, por lo tanto responde a una regulación metabólica integral del organismo, es decir,
responde a la necesidad de coordinación de los estados metabólicos de los diferentes tejidos y a la
disponibilidad general de metabolitos, reflejada en los niveles sanguíneos de los mismos.
Entre los grupos que pueden unirse covalentemente se encuentran a los grupos fosfato,
adenilo, uridilo, metilo y ribosil adenil difosfato. La fosforilación es el tipo más común de modificación
covalente. Un tercio de las todas las proteínas regulables en las células eucariotas son capaces de
sufrir este tipo de regulación. Algunas proteínas tienen sólo un sitio de fosforilación, mientras que otras
proteínas poseen numerosos residuos que pueden ser fosforilados. Este modo de modificación
covalente es central en un gran número de pasos regulatorios del metabolismo intermedio.
5.- Inducción o represión enzimática:
Este mecanismo provoca un cambio en la cantidad de enzima presente debido a una
modificación en la expresión génica de la enzima como consecuencia de una cascada de señalización
intracelular generada por un mensajero químico. Es un mecanismo lento y se manifiesta en respuesta

6
a diferentes estímulos (hormonas, cambios en el medio, etc.).
Otros mecanismos de regulación:
Los zimógenos representan un conjunto de enzimas que se activan luego de sufrir la hidrólisis
de un péptido. Esta modificación covalente es irreversible. Generalmente, este mecanismo impide que
la acción de estas enzimas, mayoritariamente enzimas proteolíticas, se manifieste en localizaciones
“inconvenientes” para la célula o el organismo (enzimas digestivas), o permite que se desencadene un
proceso en el momento apropiado (coagulación).
Especialización celular:
Aun cuando todas las células del organismo están dotadas de la información completa
correspondiente al genoma, la diferenciación que conduce a la formación de los distintos tejidos y
órganos es el resultado de la expresión diferencial de la información genética. Es decir, cada grupo de
células especializadas puede expresar diferentes genes y por lo tanto presentar diferentes actividades
enzimáticas.
Mecanismos múltiples:
La superposición de diversos mecanismos de regulación en un paso metabólico no constituye
una redundancia estéril. Cada mecanismo tiene sus particularidades, entre ellas, la velocidad con que
se desencadena y con la que se establece, y la duración de su efecto. Cuando operan varios
mecanismos a la vez, la regulación de la vía es más sensible a cambios metabólicos de diversa
índole. Por otra parte, cuando alguno de los dispositivos de control no funciona correctamente, por
anormalidades del individuo o del ambiente, las consecuencias se atenúan por la existencia de otros
mecanismos de control. Por ello, en la diversidad se encuentra gran parte de la eficiencia y eficacia de
los mecanismos de regulación metabólica, y por consiguiente, no cabe plantearse cuál de ellos es
más importante o desempeña un papel más relevante en el control del metabolismo.
Integración del metabolismo:
Uno de los requisitos para mantener y perpetuar la vida es la conservación de la homeostasis,
es decir, la constancia del medio interno (niveles sanguíneos de iones, lípidos e hidratos de carbono),
dentro de límites muy estrechos. Estas condiciones deben mantenerse tanto aún en situaciones
variadas como el reposo, el ejercicio, la saciedad o el ayuno. ¿Cómo se armoniza nuestro organismo
para sobrevivir en diferentes situaciones metabólicas?
En los mamíferos, la coordinación del metabolismo se logra a través del sistema
neuroendócrino. Las principales hormonas que participan en la regulación del metabolismo intermedio
son: la insulina, el glucagon, las catecolaminas y el cortisol. Por lo tanto, antes de seguir repasemos
sus efectos.
La Insulina
La insulina es una hormona anabólica porque promueve el almacenamiento de nutrientes. En
particular, la insulina promueve
a) el almacenamiento de glucosa como glucógeno en hígado y músculo
b) la conversión de glucosa en TAG en hígado y su almacenamiento en el tejido adiposo
c) el transporte de aminoácidos y la síntesis de proteínas en músculo
d) la síntesis de proteínas en el hígado (por ej. la albúmina)
e) el transporte de glucosa al músculo y al tejido adiposo
La insulina también inhibe la movilización de los combustibles.
¿Qué efectos tiene el glucagon? Como ya dijimos, el glucagon favorece la movilización de

7
combustibles:
1) estimula la liberación de glucosa a partir del glucógeno hepático
2) estimula la gluconeogénesis hepática a partir de lactato, glicerol y aminoácidos
3) moviliza los ácidos grasos de los TAG del tejido adiposo como fuente alternativa de combustible
Otras hormonas cuyos efectos son opuestos a los efectos de la insulina son la adrenalina, la
noradrenalina y el cortisol. Sólo la insulina y el glucagon se liberan como respuesta directa al
cambio de combustibles en la sangre. La liberación de las otras hormonas es mediada por señales
neurales.
Mecanismos homeostáticos en saciedad
La saciedad es la percepción de no tener necesidad inmediata de ingesta de alimentos.
Utilizaremos este término para describir la situación que se observa luego de la ingesta de alimentos.
Luego de la ingesta, el incremento de la glucemia es el estímulo para que las células del
páncreas liberen insulina. Ciertos aminoácidos, particularmente arginina y leucina, también estimulan
la liberación de insulina. Los niveles sanguíneos de glucagon, secretado por las células del
páncreas, pueden aumentar o disminuir dependiendo de la composición de la ingesta. Si la ingesta es
rica en glúcidos, la concentración de glucagon disminuye, pero si es rica en proteínas, los niveles de
glucagon sanguíneo aumentan. Luego de una comida típica mixta con carbohidratos, proteínas y
grasas, los niveles de glucagon permanecen relativamente constantes mientras que los de insulina
aumentan.

8
¿Qué ocurre con la glucosa que llega al hígado desde el intestino?
El hígado es el primer tejido que tiene la oportunidad de utilizar la glucosa que proviene de la
dieta que le llega por la circulación porta. En saciedad, el hígado oxida glucosa para satisfacer sus
necesidades inmediatas y su exceso se almacena como glucógeno. La glucosa puede convertirse en
glucógeno; en piruvato y lactato (por glucólisis) o puede
utilizarse en la vía de las pentosas. El piruvato puede
oxidarse a acetilCoA, que a su vez se convierte en
ácidos grasos y luego en triacilglicéridos, u oxidarse a
CO2 y agua en el ciclo de Krebs (TCA).
¿Qué mecanismos regulatorios controlan el
almacenamiento de combustibles producidos a
partir de glucosa? Luego de la digestión y absorción de
los glúcidos, la glucosa llega a la vena porta, donde su
concentración puede alcanzar los 180-360 mg% (10-20 mM), y finalmente al hígado. Dado su elevado
S0.5 (Km) para la glucosa, cuando la concentración de glucosa es normal, la actividad de la
glucoquinasa es muy baja. Como consecuencia del incremento de la concentración de glucosa que
llega al hígado por la vena porta, la velocidad de fosforilación de la glucosa por la glucoquinasa
aumenta. Los niveles de la enzima también se INDUCEN con dietas ricas en carbohidratos en
respuesta al incremento en los niveles de insulina. La insulina promueve el almacenamiento de
glucosa como glucógeno. La insulina activa la fosfodiesterasa, disminuyendo los niveles de AMPc, a
las fosfatasas que desfosforilan a la glucógeno sintasa (GS desfosforilada es activa) y a la enzima
clave de la glucogenolisis (glucógeno fosforilasa desfosforilada es inactiva). La insulina también
promueve la síntesis de triacilglicéridos (TAG), que
se liberan del hígado como VLDL.
¿Qué ocurre con la glucosa que no es utilizada
por el hígado?
Parte de la glucosa que proviene del intestino
llega a la circulación general. El cerebro utiliza casi
únicamente glucosa para generar ATP. Otros
grandes consumidores de glucosa son los glóbulos
rojos, que sólo pueden convertir la glucosa en
lactato y piruvato y el tejido adiposo que la
convierte en lípidos. El músculo puede utilizar
glucosa para convertirla en glucógeno o degradarla
por glucólisis. Algunos tejidos producen lactato y

9
piruvato por glucólisis a partir de la glucosa sanguínea. Estos metabolitos son captados por el hígado
y convertidos en lípidos. EN EL ESTADO DE SACIEDAD, EL HÍGADO UTILIZA GLUCOSA Y NO
REALIZA GLUCONEOGÉNESIS. Por lo tanto, el ciclo de Cori (la conversión de glucosa a lactato en
los tejidos periféricos y luego la conversión de lactato en glucosa en el hígado) se interrumpe en
estado de saciedad. En este caso, el lactato que llega al hígado se convierte en piruvato que puede
convertirse en acetilCoA, que a su vez puede oxidarse (ciclo de Krebs) o convertirse en ácidos grasos
y TAG.
Todos los tejidos requieren glucosa para la vía de las pentosas y muchos usan glucosa para la
síntesis de glucoproteínas y otros compuestos que contienen carbohidratos.
La insulina estimula el transporte de glucosa a las células del músculo en reposo y adipocitos
promoviendo el reclutamiento de los transportadores de glucosa de tipo GLUT 4 en la membrana
celular. Otros tejidos, como el hígado, el cerebro y los glóbulos rojos tienen un tipo diferente de
transportador de glucosa que no se afecta significativamente por insulina. Luego de una ingesta, se
sintetiza glucógeno en el músculo por un proceso similar al del
hígado. Existen diferencias metabólicas entre estos tejidos pero
en esencia, la insulina estimula la síntesis de glucógeno en el
músculo en reposo como lo hace en el hígado. Una diferencia
clave entre el metabolismo del glucógeno muscular y hepático, es
que la insulina estimula fuertemente el transporte de glucosa en el
músculo mientras que el transporte de glucosa en las células
hepáticas es independiente de insulina.
En el tejido adiposo, la insulina también estimula el transporte
de glucosa que se oxida para producir energía y también provee
glicerol para la síntesis de TAG. La glucosa también puede
convertirse en ácidos grasos en el tejido adiposo.
¿Qué ocurre luego de una ingesta con los distintos tipos de
nutrientes que la componen?
Los productos finales de la digestión de los alimentos son los
monosacáridos, los ácidos grasos y los aminoácidos que por
diferentes rutas llegan a la circulación sanguínea.
En esta situación, la hormona predominante es la insulina, que favorece la entrada de glucosa
a las células, particularmente a los miocitos y adipocitos. Sumado a la mayor utilización de glucosa en
los hepatocitos, la consecuencia es una disminución de la glucemia. Dos horas después de la ingesta
la glucemia vuelve a los niveles del ayuno, 70 a 110 mg% (aprox. 5 mM).
En un individuo normal, aproximadamente el 50% de la glucosa ingerida se metaboliza por
glucólisis, el 10% se acumula como glucógeno y el 30 - 40% se convierte en lípidos. En términos
generales, en ausencia de insulina, la glucólisis, la glucogénesis y la lipogénesis disminuyen. Sólo el
5% de la glucosa ingerida se transforma en lípidos en un diabético deficiente en insulina.
¿Qué ocurre con las proteínas hidrolizadas en la digestión? Algunos de los aminoácidos de la
dieta son utilizados por las células intestinales para obtener energía, el resto llega al hígado a través
de la vena porta. El intestino metaboliza aspartato, asparagina, glutamato y glutamina y libera alanina
a la sangre portal. En el hígado, sólo se metabolizan aminoácidos cuando su concentración es muy
alta dado que las transaminasas hepáticas tienen un Km alto, lo que permite “priorizar” el uso de
aminoácidos para la síntesis de proteínas. Los esqueletos carbonados de los aminoácidos
degradados en el hígado pueden ser oxidados completamente a CO2 y H2O o bien utilizarse como

10
sustrato para la lipogénesis, y el grupo amino se convierte en urea. En los demás tejidos, los
aminoácidos provenientes de la dieta pueden utilizarse para la síntesis de proteínas o para obtener
energía, lo que dependerá del estado metabólico, es decir, de la disponibilidad de energía. El hígado
tiene baja capacidad para transaminar aminoácidos ramificados que son fácilmente degradados en el
músculo esquelético. De esta forma, el músculo transamina aminoácidos ramificados y libera los
cetoácidos a la sangre, de donde son captados y oxidados en el hígado. En el músculo, los
aminoácidos ramificados se utilizan para sintetizar alanina y glutamina, que se liberan a la sangre.
¿Qué ocurre con los lípidos en la saciedad?
Los lípidos de la dieta llegan a la circulación general contenidos en quilomicrones. En la
circulación también se encuentran las VLDL provenientes del hígado, que contienen los productos de
la lipogénesis a partir de glucosa, lactato, piruvato y aminoácidos. Los Qm y las VLDL son sustrato de
la LIPOPROTEÍN LIPASA, una enzima anclada en las células endoteliales de los capilares, que es
muy abundante en el endotelio del tejido adiposo. Esta enzima cataliza la liberación de ácidos grasos
de los Qm y las VLDL a partir de los TAG. En saciedad, se frena la lipólisis y se produce lipogénesis
en el tejido adiposo y la concentración plasmática de ácidos grasos libres es baja. Los ácidos grasos
captados por los adipocitos son re-esterificados con glicerol 3-fosfato para formar TAG y se
almacenan en forma de gotas de grasa dentro de estas células. El glicerol 3-P se produce en los
adipocitos a través de la glucólisis que genera dihidroxiacetona fosfato (DHAP). La enzima glicerol 3-P
deshidrogenasa reduce la DHAP a glicerol-3P.
Mecanismos homeostáticos en el ayuno
Cambios en los niveles de insulina y glucagon
Cuando la glucemia disminuye como consecuencia de los mecanismos ya descriptos, los niveles
de insulina bajan y las células del páncreas comienzan a liberar glucagon. Estos cambios
hormonales provocan la degradación del glucógeno hepático y la síntesis de glucosa por
gluconeogénesis, lo que permite mantener la glucemia.
Estimulación de la glucogenolisis
El glucagon se une a receptores 7TMS
ubicados en la membrana plasmática y
asociados a la proteína Gs. La activación de
la adenilato ciclasa produce un incremento
en los niveles de AMPc y la activación de la
proteína quinasa A. Como consecuencia, la
glucógeno sintasa es fosforilada e inactivada
y la síntesis de glucógeno disminuye. Al
mismo tiempo, se estimula la degradación de
glucógeno por un mecanismo en dos pasos:
1) la PKA fosforila y activa a la fosforilasa
quinasa. 2) Esta enzima, a su vez, fosforila y
activa a la glucógeno fosforilasa. La
fosforilasa cataliza la glucogenolisis,
produciendo glucosa-1-fosfato, que se
convierte en glucosa-6-fosfato. La
desfosforilación de la glucosa-6-fosfato por la
glucosa 6 fosfatasa produce glucosa libre,

11
que se libera a la sangre.
Estimulación de la gluconeogénesis
Alrededor de 4 horas luego de una comida, el hígado produce glucosa que libera a la sangre no
sólo por glucogenolisis, sino también por gluconeogénesis. Los cambios hormonales activan la
liberación de precursores para la gluconeogénesis desde los tejidos periféricos, específicamente
lactato, aminoácidos y glicerol. Por diversos mecanismos se promueve la conversión de los
precursores gluconeogénicos a glucosa y se evita la producción de ciclos fútiles. En particular, en el
hígado, durante el ayuno se inactivan las enzimas glucolíticas, piruvato quinasa, fosfofructoquinasa 1
(FFQ1) y glucoquinasa promoviendo el flujo de carbonos por la vía gluconeogénica. Estos
mecanismos operan en los tres pasos en los que difieren la glucólisis y la gluconeogénesis:
1. Conversión de piruvato a Fosfoenolpiruvato (PEP). El piruvato (derivado de lactato y alanina) se
convierte por la vía gluconeogenética a PEP. Este no se reconvierte a piruvato (ciclo fútil) porque el
glucagon promueve la fosforilación e inhibición de la piruvato quinasa. El PEP se transforma en
fructosa 1,6 difosfato (reversión de las reacciones de la glucólisis).
2. Conversión de fructosa 1,6 bifosfato en fructosa 6 fosfato. Dados los bajos niveles del regulador
alostérico fructosa 2,6 bifosfato, la enzima glicolítica FFQ1 es relativamente inactiva. Por lo tanto, la
fructosa 6 fosfato no se convierte en fructosa 1,6 bifosfato y se evita así un segundo ciclo fútil. La
fructosa 6 fosfato se convierte en glucosa 6 fosfato.
3. Conversión de glucosa 6 fosfato en glucosa por la glucosa-6 fosfatasa. Dado que la
glucoquinasa tiene un alto S0.5 (Km) para la glucosa y las concentraciones de glucosa son
relativamente bajas en el hepatocito durante el ayuno, la glucosa se libera a la sangre. Por lo tanto el
tercer ciclo fútil potencial no ocurre.
Las enzimas que participan en la gluconeogénesis, pero no en la glucólisis, están activas
durante el ayuno. La piruvato carboxilasa se activa por el acetilCoA que proviene de la oxidación de
ácidos grasos. La fosfoenolpiruvato carboxiquinasa (PEPCK), la fructosa 1,6 bifosfatasa y la glucosa 6
fosfatasa se inducen (aumentan los niveles proteicos de las enzimas) por efectos del glucagon y los
glucocorticoides. Cuando los niveles de fructosa 2, 6 difosfato son bajos, la fructosa 1,6 bifosfatasa es
activa. En esta situación metabólica, el ciclo de Cori contribuye a mantener la glucemia. El hígado
sintetiza glucosa a partir de lactato y luego la glucosa se convierte de nuevo en lactato por glucólisis

12
en los tejidos periféricos como los glóbulos rojos. El ciclo de la alanina, mediante el cual vuelven los
carbonos al hígado como alanina en vez de lactato también contribuye a la gluconeogénesis.
Luego de varias horas de la ingesta, ya no ingresa combustible desde el intestino y queda poco
glucógeno en el hígado. Los tejidos que usan glucosa dependen de la gluconeogénesis hepática,
principalmente a partir de lactato, glicerol y alanina.
En los ciclos de Cori y de la alanina NO SE PRODUCE SÍNTESIS NETA DE GLUCOSA,
sólo se reemplaza la utilizada por los tejidos periféricos. Sin embargo, dado que otros tejidos como el
cerebro oxidan completamente glucosa, es obligatoria su síntesis durante el ayuno.
El glicerol, producto de la lipólisis en el tejido adiposo es un sustrato para la síntesis de glucosa
en el ayuno, en cambio los ácidos grasos no pueden utilizarse para la síntesis de glucosa porque el
acetilCoA no puede convertirse en los intermediarios de tres carbonos de la gluconeogénesis.
Son las proteínas, especialmente las del tejido muscular, las que aportan la mayor parte del
carbono necesario para la gluconeogénesis. Las proteínas musculares se hidrolizan en el músculo
produciendo aminoácidos que en su mayoría, se metabolizan en el músculo. Los principales
aminoácidos que se liberan del músculo son alanina y glutamina. El metabolismo de los demás
aminoácidos produce intermediarios (piruvato, -cetoglutarato) que pueden producir alanina y
glutamina. Estos aminoácidos se liberan a la sangre, de la que pueden ser captados en el hígado o el
riñón y convertidos en glucosa. Cuantitativamente la alanina es el sustrato gluconeogenético más
importante.
El músculo también puede liberar los cetoácidos de aminoácidos ramificados que pueden ser
transformados en el hígado en glucosa o cuerpos cetónicos (dependiendo de su condición de
aminoácidos cetogénicos o glucogénicos). A partir del cetoácido de la valina se puede sintetizar
glucosa, a partir de cetoácido de la leucina se puede sintetizar cuerpos cetónicos y ambos tipos de
compuestos se pueden sintetizar a partir del cetoácido de isoleucina Gran parte de la glutamina
liberada por el músculo es convertida en alanina en el epitelio intestinal. En el intestino, la glutamina
puede ser parcialmente oxidada para producir energía y en parte el esqueleto carbonado y el grupo
amino se liberan a la sangre como alanina y amonio. Esta vía probablemente involucra la formación
de oxaloacetato a partir de glutamina por el TCA y la conversión a PEP y a piruvato que se transamina
a alanina.
La síntesis de glucosa en el hígado durante el ayuno está muy relacionada con la síntesis de
urea. La mayoría de los aminoácidos ceden su grupo amino por transaminación con -cetoglutarato
formando glutamato y un nuevo cetoácido que puede utilizarse para la gluconeogénesis (para un
aminoácido glucogénico). El glutamato (por desaminación oxidativa catalizada por la glutamato
deshidrogenasa) provee el amonio para la síntesis de urea. Una fuente adicional de amonio es la
mucosa intestinal que convierte glutamina en alanina y amonio. Es importante considerar que cuanto
mayor sea la actividad gluconeogénica, mayor será la producción de urea.
¿Qué limita la capacidad del hígado de convertir aminoácidos en glucosa?
La conversión de aminoácidos en glucosa involucra diversos procesos metabólicos:
transaminación, desaminación oxidativa, conversión de NH4+ en urea y finalmente gluconeogénesis a
partir del esqueleto carbonado del aminoácido. La limitación fisiológica para formar glucosa a partir de
aminoácidos es la gran cantidad de energía requerida para este proceso. Se requieren cuatro
moléculas de ATP para convertir amoníaco en urea y seis más para transformar el esqueleto
carbonado de los aminoácidos en glucosa.
En todas las células los niveles de ATP se mantienen relativamente constantes. Por lo tanto,
para la gluconeogénesis, el hígado debe realizar metabolismo aeróbico para producir ATP en
reemplazo del ATP consumido.

13
Glucemia durante el ayuno prolongado
Durante el ayuno prolongado, se produce un cambio en la utilización de los combustibles. Los
tejidos usan menos glucosa que durante un ayuno corto y utilizan predominantemente combustibles
derivados de la metabolización de los TAG del tejido adiposo (es decir, ácidos grasos y cuerpos
cetónicos). En consecuencia, la glucemia no cae drásticamente. De hecho, aún después de 5 a 6
semanas de ayuno, la glucemia es de alrededor de 65 mg%.
El principal cambio que ocurre en el ayuno prolongado es un incremento significativo en los niveles
sanguíneos de cuerpos cetónicos luego de 3 a 5 días de ayuno. En estos niveles, el cerebro y otros
tejidos nerviosos comienzan a utilizarlos y consecuentemente oxidan menos glucosa, requiriendo
alrededor de la tercera parte de la glucosa (40 g/día) que requieren en condiciones de alimentación
normal. Como resultado de esta reducción en la utilización de glucosa, la velocidad de la
gluconeogénesis en el hígado disminuye y también lo hace la producción de urea. Por lo tanto, no se
degradan las proteínas del músculo y otros tejidos, dado que hay una menor necesidad de
aminoácidos para la gluconeogénesis.
La proteína corporal, particularmente la muscular, no es fundamentalmente una forma de
almacenaje de combustible, como lo son el glucógeno o los TAG, sino que las proteínas tienen
funciones muy importantes tanto estructurales como funcionales. Por lo tanto, si la degradación de
proteínas aumenta excesivamente, la función corporal puede comprometerse seriamente. Si continúa
el ayuno y no hay otros problemas, como infecciones, el individuo muere, generalmente porque la
pérdida severa de proteínas produce el mal funcionamiento de los órganos principales, como el
corazón. Por lo tanto, el aumento en los cuerpos cetónicos que disminuye la degradación de
proteínas, permite la supervivencia por largos períodos sin ingerir alimentos.
Estimulación de la lipólisis
Los cambios hormonales (baja relación insulina/glucagon) que ocurren durante el ayuno
estimulan la degradación de TAG del tejido adiposo. Consecuentemente, se liberan a la circulación
ácidos grasos y glicerol. Los ácidos grasos (AG) son utilizados como combustible preferentemente a la
glucosa por muchos tejidos. En el corazón y el músculo, la oxidación de los AG inhibe la glucólisis. En
el cerebro, los AG no se oxidan porque no atraviesan la barrera hematoencefálica. La -oxidación de
ácidos grasos en el hígado, permite generar el ATP necesario para la gluconeogénesis y la síntesis de
acetilCoA. En este caso, sin embargo, la mayor parte del Acetil CoA no entra al TAC, sino que se
convierte en cuerpos cetónicos (CC, acetoacetato y -hidroxibutirato), que se transportan a la sangre y
sirven como combustible adicional para los tejidos extrahepáticos. Esto ocurre porque en esas
condiciones los niveles de oxaloacetato hepáticos son muy bajos dado que el oxaloacetato se utiliza
para la síntesis de glucosa. Como en el caso de los AG, los CC son utilizados preferentemente a la
glucosa en muchos tejidos. Pueden atravesar la barrera hematoencefálica y cuando la cetonemia es
suficientemente alta, los CC son un combustible alternativo para el cerebro (Aclaración: esto ocurre
cuando se está en un estado de inanición: es decir con más de 3 a 5 días de ayuno) aunque son
incapaces de reemplazar completamente la necesidad de glucosa del cerebro. Los CC también
inhiben la proteólisis del músculo esquelético y por lo tanto disminuyen la destrucción muscular
durante el ayuno. Mientras la cetonemia esté elevada se necesitará menos glucosa, menos
aminoácidos gluconeogénicos y menor necesidad de usar proteínas musculares. La glucemia tiende a
disminuir en el ayuno por lo que se reduce la secreción de insulina y se favorece la de glucagon.
Además el ayuno reduce la formación de triiodotironina, la forma activa de la hormona tiroidea a partir
de tiroxina lo que puede reducir el requerimiento basal de energía hasta un 25%.

14
¿Cómo se movilizan los aminoácidos durante el ayuno?
Durante el ayuno nocturno, la síntesis de proteínas en el hígado y otros tejidos continua, pero a
una velocidad menor comparada con la del estado postprandial. Hay una degradación neta de
proteínas tanto en el músculo esquelético (que contiene la masa proteica más importante del
organismo) como en otros tejidos. El músculo esquelético puede oxidar aminoácidos ramificados para
producir energía y liberar alanina y glutamina. La liberación de aminoácidos musculares durante el
ayuno nocturno es estimulada por la disminución de los niveles de insulina y el aumento de los niveles
de glucocorticoides, que provocan un aumento en la proteólisis y liberación de aminoácidos a la
sangre. El hígado es el principal órgano que capta la alanina circulante. Además, el hígado también
capta otros aminoácidos libres, alfa-ceto ácidos y algo de glutamina de la sangre. La alanina y otros
aminoácidos son degradados y el amoníaco formado puede ser convertido en urea, mientras que el
esqueleto carbonado es convertido principalmente en glucosa. El glucagon y los glucocorticoides
estimulan la captación hepática de aminoácidos, la gluconeogénesis y ureagénesis. La inducción de
las enzimas gluconeogénicas por glucagon y glucocorticoides durante el ayuno nocturno se
correlaciona con la inducción de las enzimas de la degradación de aminoácidos (ej., tirosina
aminotransferasa) y la inducción de las enzimas del ciclo de la urea. El ciclo de la urea también se
incrementa por el mayor aporte de amonio proveniente de la degradación hepática de los
aminoácidos.
¿Qué ocurre con el metabolismo de los aminoácidos en otros tejidos?
La glutamina es generada en el músculo esquelético por la oxidación de los aminoácidos
ramificados, y en los pulmones y cerebro para la eliminación del amonio formado localmente a partir
del metabolismo de aminoácidos El riñón, el intestino y las células con un alto recambio celular
representan los sitios de mayor captación de glutamina, que actúa como donor de nitrógeno para la
síntesis de bases nitrogenadas y en particular en el riñón, como mecanismo de regulación del
equilibrio ácido-base en situaciones de acidez metabólica.
¿Cuál es la clave de la cooperación metabólica entre los distintos tejidos?
La cooperación metabólica entre células y entre diferentes tejidos es fundamental para la
supervivencia de un individuo. Nuestro organismo está organizado para mantener estable el medio
interno utilizando los mecanismos de regulación enzimática que hemos detallado previamente. Pero,
¿cuál es la “llave maestra” de esta integración?
Se ha descripto que la quinasa activada por AMP (AMPK) es el principal regulador del
metabolismo: La AMPK induce una cascada de eventos dentro de las células en respuesta a

15
cambios en la carga energética de la célula. Su rol como reguladora de la carga energética ubica a
esta enzima en un punto de control central en el mantenimiento de la homeostasis energética. Por
otra parte, recientemente, se ha descripto que la actividad de AMPK también puede ser regulada por
estímulos fisiológico como hormonas y nutrientes.
Cuando se activa la AMPK las células cambian de un estado de consumo activo de ATP (como en la
síntesis de ácidos grasos) a una
producción activa de ATP (como en la
oxidación de glucosa y ácidos grasos).
La activación de la AMPK también ejerce
efectos a largo plazo a nivel de la
expresión génica y la síntesis de
proteínas. Otra actividad importante
atribuida a la AMPK es la regulación de
la síntesis y secreción de insulina a nivel
pancreático y la modulación de la función
hipotalámica involucrada en la regulación
de la saciedad.
El ATP es la fuente principal de energía
bioquímica. Sin embargo los niveles
endógenos de AMP se modifican más significativamente que los de ADP o ATP. Pequeños cambios
en la concentración de ADP o ATP conducen a importantes modificaciones en los niveles de AMP, de
acuerdo a la siguiente reacción:
2 ADP ATP + AMP Adenilato Quinasa
Keq = [ATP] X [AMP] y por lo tanto [AMP] = Keq [ADP]2/ [ATP]
[ADP]2
La unión del AMP (regulador alostérico) a la AMPK aumenta su actividad y favorece su activación
haciendo a la enzima un mejor sustrato para las quinasas que la fosforilan (AMPKK) y peor sustrato
para las fosfatasas que la desfosforilan. Debido a que la AMPK es altamente sensible a las
variaciones en los niveles de AMP, esta enzima modifica su actividad frente a pequeños cambios en la
relación [ATP]/ [ADP]. La regulación alostérica negativa de la AMPK es ejercida por la fosfocreatina.
La AMPK aumenta el metabolismo energético incrementando la captación de glucosa en el
músculo en ejercicio y activando el metabolismo de los ácidos grasos. Simultáneamente, la AMPK
inhibe la síntesis de ácidos grasos, la transferencia del grupo fosfato de alta energía de la
fosfocreatina, la síntesis de colesterol, la transcripción del ADN y la apoptosis.
El balance entre los nucleótidos de adenina está regulado por la actividad de la adenilato
quinasa y a su vez depende fuertemente de la producción mitocondrial de ATP y del metabolismo
anaeróbico de los hidratos de carbono. El ATP consumido se resintetiza mediante las actividades de
la adenilato quinasa, la creatina fosfoquinasa y el metabolismo anaeróbico. Cuando el consumo de
energía es menor y sostenido en el tiempo, se utiliza la oxidación mitocondrial de ácidos grasos para
generar ATP.
La AMPK tiene un dominio que puede unirse al glucógeno. En el músculo, un contenido alto de
glucógeno reprime la actividad de la AMPK. También se ha demostrado que aumenta la actividad la
enzima en respuesta al estímulo de receptores acoplados a fosfolipasa C y a hormonas secretadas
por el tejido adiposo (adipoquinas) tales como leptina y adiponectina.
Quinasa activada por AMP
AMPK
“Coordinación del metabolismo”
Dieta Estres Ejercicio
5´ AMP
AMPK
AMPKK PPasa
-oxidacion
Síntesis de colesterol
Transcripción
ApoptosisCreatina fosfoquinasa
Sintesis de ácidos grasos
Transporte de glucosa

16
¿Qué consecuencias tiene la activación de la AMPK?
La cascada de señalización iniciada por la activación de la AMPK tiene efectos sobre el
metabolismo de glucosa y lípidos, la expresión de genes y la síntesis de proteínas. Estos efectos son
importantes en la regulación del metabolismo del hígado, músculo esquelético, corazón, tejido adiposo
y páncreas.
Esquema que representa el rol central de AMPK in la regulación del metabolismo en respuesta a eventos tales como estrés inducido por ejercicio o nutrientes. Las flechas indican efectos positivos de la AMPK, mientras que las líneas T indican efectos inhibitorios
Metabolismo intermedio durante el ejercicio
Regulación de la glucemia durante el ejercicio
Durante el ejercicio operan mecanismos similares a los que ocurren durante el ayuno, lo que
permite mantener la glucemia en valores normales a pesar del aumento en el consumo de glucosa
que ocurre en esta situación. En una etapa inicial, las células musculares obtienen ATP a partir de la
creatina fosfato. Sin embargo, la cantidad de creatina fosfato en el músculo puede sostener el ejercicio
sólo duante períodos muy breves. Por lo tanto, luego de esta etapa inicial comienzan a degradarse los
depósitos de glucógeno muscular, y la glucosa obtenida es oxidada en el músculo para aportar ATP.
La disponibilidad de ATP en el músculo durante el ejercicio se mantiene por los siguientes procesos:
a) Adenilato quinasa
Cuando el ATP se convierte en ADP (durante la contracción muscular), la adenilato quinasa
regenera el ATP produciendo AMP en el proceso, en la siguiente reacción:
adenilato quinasa
2 ADP ATP + AMP
El aumento en los niveles de AMP activa la glucólisis estimulando a la FFQ1 y también a la
fosforilasa b. La fosforilasa b también puede activarse por fosforilación convirtiéndose en fosforilasa a.
La fosforilación de la fosforilasa b también se estimula por un aumento en los niveles de calcio, que

17
proviene del retículo sarcoplásmico. El Ca2+ se une a calmodulina y este complejo activa a la
fosforilasa quinasa que fosforila y activa a la fosforilasa.
2 ADP ATP + AMP
ADP Pi
2 ADP ATP + AMP
Músculo en reposo: 1,0 mM 5mM 0,2mM
Músculo en ejercicio: 1,5mM 4 mM 0,8mM
Con la actividad muscular, los cambios en las concentraciones de ATP y ADP son relativamente
pequeños. Sin embargo, a través de la adenilato quinasa, los niveles de AMP aumentan
marcadamente durante el ejercicio (como vemos en el ejemplo, hay 4 veces de aumento en los
niveles de AMP en el ejercicio respecto del reposo). De esta forma, los niveles de AMP representan
una señal intracelular muy importante en la regulación del metabolismo muscular (por ej. recordar que
es activador de la movilización de glucógeno)
Fuentes de combustible para la contracción muscular
Fuente
Velocidad
max. de prod.
(mmol/seg)
~ P total disp
(mmol)
ATP muscular 223
Creatina-P 73,3 446
Glucógeno muscular Lactato 39,1 6700
Gucógeno muscular CO2 16,7 84000
Glucógeno hepático CO2 6,2 19000
Ácidos grasos (adiposo) CO2 6,7 4000000
b) Creatina Fosfoquinasa/ Fosfocreatina
La fosfocreatina es una reserva de enlaces fosfato de alta energía.
Fosfocreatina + ADP ATP + creatina
Aunque la reacción de la fosfocreatina es la más rápida para producir ATP, la cantidad de ATP
producido en esta reacción es considerablemente pequeña. En un ejercicio intenso, las reservas de
fosfocreatina se agotan aproximadamente a los 30 - 40 segundos. Los corredores olímpicos son
capaces de correr 100 m “casi sin respirar” y aproximadamente la mitad de la energía que usan en la
corrida proviene de los enlaces de alta energía acumulados como creatina fosfato. Mientras que la
fosfocreatina es importante para un máximo rendimiento, otras fuentes de producción de energía
deben ponerse en juego después de los primeros 30 segundos de una corrida rápida y corta. Después
de haber agotado las reservas de fosfocreatina el músculo puede obtener ATP a partir de la
metabolización de glucosa a piruvato o lactato (glucólisis anaeróbica). Por esta vía, sólo se obtienen 2
ATP por cada mol de glucosa o bien 3 ATP por cada resto glicosilo obtenido a partir de la
Piruvato
Adenilato
quinasa Glucógeno
Glucosa-1-P
Glucosa-6-P
+ fosforilasa b
+ FFQ-1

18
glucogenolisis. La glucólisis anaeróbica es un proceso rápido, pero produce ácido láctico que puede
acumularse en el músculo en ejercicio, y producir acidosis. Por otra parte, el lactato puede transferirse
al hígado donde es transformado en glucosa por gluconeogénesis (ciclo de Cori), lo que traslada parte
de la carga metabólica del músculo al hígado. Además, en condiciones anaeróbicas, los ácidos grasos
NO pueden usarse como sustratos. Por lo tanto, si sólo se produce metabolismo anaeróbico en el
músculo en ejercicio, las reservas de glucógeno muscular se agotarán rápidamente y se iniciará la
captación de glucosa sanguínea, lo que puede provocar hipoglucemia y un malfuncionamiento del
SNC.
Los depósitos de glucógeno en el músculo son suficientes para realizar ejercicios como
levantar pesas o empujar un objeto, pero sólo por períodos cortos (aproximadamente 2 min). Durante
el ejercicio intenso, se libera a la sangre la hormona adrenalina que se une a receptores de membrana
en las células musculares, y activa a la adenilato ciclasa. Cuando los niveles de AMPc aumentan, la
PKA se activa y cataliza la fosforilación de la fosforilasa quinasa, que se activa y fosforila a la
glucógeno fosforilasa. La fosforilasa a cataliza la conversión de glucógeno a glucosa 1 fosfato, la que
por la fosfoglucomutasa se convierte en glucosa 6 fosfato que entra a la glucólisis. En los músculos
que contienen muchas fibras glucolíticas (como los pectorales), se produce ATP principalmente por
glucólisis, siendo lactato el producto principal.
A diferencia del ejercicio intenso, el ejercicio moderado puede sostenerse por períodos largos.
Un individuo entrenado, por ejemplo, puede correr durante muchas horas. Los músculos de la pierna
contienen gran cantidad de fibras de contracción lenta (rojas) que son capaces de oxidar combustibles
a CO2 y H2O, dado que contienen más mitocondrias que los músculos compuestos
predominantemente por fibras glicolíticas de contracción rápida (blancas). Los músculos utilizados en
ejercicios violentos y de corta duración (carera de 100 m) tienen predominio de éstas últimas fibras.
Cuando el flujo sanguíneo del músculo en ejercicio aumenta, un proceso que requiere de 5 a
10 minutos, aumenta la llegada de combustibles sanguíneos y de oxígeno al músculo. En estas
condiciones, el músculo puede captar combustibles, principalmente glucosa y ácidos grasos y los
oxida para obtener ATP.
Por lo tanto, en estas condiciones el aporte de glucosa de la sangre debe ser restaurado. El hígado
produce glucosa por degradación de sus depósitos de glucógeno y por gluconeogénesis. La fuente
principal de carbonos para la gluconeogénesis durante el ejercicio es el lactato producido por el
músculo en ejercicio, pero también se utilizan aminoácidos y glicerol. La adrenalina liberada durante el
ejercicio estimula la glucogenolisis y la gluconeogénesis hepáticas, a través de un aumento en los
niveles de AMPc. En estas condiciones, el músculo puede oxidar ácidos grasos y una pequeña
proporción de cuerpos cetónicos provenientes de la circulación, que se producen como consecuencia
de la lipólisis de los TAG del tejido adiposo. Durante el ejercicio prolongado, los ácidos grasos son el
combustible principal del músculo en ejercicio, lo que permite que otros tejidos con dependencia
absoluta por glucosa no se vean afectados. La utilización de ácidos grasos en lugar de glucosa como
combustible en el músculo esquelético depende de diversos factores, como la disponibilidad de ácidos
grasos en el torrente sanguíneo, que a su vez, depende de su liberación por el tejido adiposo por un
proceso regulado por la lipasa hormona-sensible. Durante un ejercicio prolongado, se observa una

19
pequeña disminución en los niveles de insulina, y un aumento de los niveles de glucagon,
catecolaminas, cortisol y posiblemente hormona de crecimiento que activan la lipólisis en el adipocito.
Asimismo, productos de la oxidación de los ácidos grasos inhiben la glucólisis muscular, dado que la
actividad de la piruvato deshidrogenada es inhibida por AcetilCoA, NADH y ATP, que aumentan
cuando se produce beta-oxidación. En estas condiciones, la concentración de AMP baja, la de ATP
sube y la actividad de la fosfofructoquinasa 1 (PFK-1) disminuye. El transporte de glucosa hacia el
músculo esquelético está mediado por el transportador GLUT4 que es activado por insulina o el
ejercicio. La oxidación de los cuerpos cetónicos también aumenta durante el ejercicio. Su utilización
como combustible depende de su velocidad de producción hepática. Sin embargo, los cuerpos
cetónicos nunca son el principal combustible para el músculo (el músculo siempre “prefiere” ácidos
grasos libres). Además, para la degradación de ácidos grasos como combustible, la acetil CoA
carboxilasa debe estar inactivada, es decir deben ser bajos los niveles de malonilCoA que inhiben la
entrada de los acilCoA a la mitocondria. Este proceso ocurre porque se activa la AMP-PK que
catalizar (e inactiva) a la acetil-CoA carboxilasa.
El control metabólico es específico de cada órgano
Los mecanismos de integración metabólica pueden diferir en los diferentes tejidos. Si bien
todas las células del organismo están equipadas con la misma información genética, los diferentes
tipos celulares expresan o suprimen diferentes genes, en forma específica. Por lo tanto, los tejidos
difieren en su expresión enzimática, en su respuesta a hormonas y en el transporte de diversas
sustancias.
Hígado
El hígado esta “estratégicamente” ubicado entre la circulación general y el tracto digestivo.
Como ya se mencionó, una de las funciones centrales del hígado es mantener la glucemia dentro de
límites estrictos. Sin embargo, en el hígado se produce también un sinnúmero de reacciones que
permiten monitorear, reciclar, modificar y distribuir todos los metabolitos absorbidos en el tracto
digestivo. El hígado recibe todos los nutrientes que entran a la sangre proveniente del tracto digestivo
por la circulación enterohepática a través del sistema porta. Esto le permite cumplir con algunas de
sus funciones específicas (como por ejemplo la síntesis de las proteínas de la coagulación) y
metabolizar y eliminar compuestos tóxicos ingeridos (por ej. alcohol) o productos metabólicos tóxicos
(como el amoníaco). Además del aporte sanguíneo de la vena porta, el hígado recibe a través de la
arteria hepática, sangre rica en oxígeno y en metabolitos provenientes de tejidos periféricos, que
fueron secretados a la circulación (por ej. glucosa, aminoácidos, ciertas proteínas complejo hierro-
transferrina, LDL remanentes, quilomicrones remanentes y también metabolitos de desecho
producidos en el metabolismo de distintos tejidos). Por otro lado, el hígado cumple una importante
función secretora de moléculas sintetizadas localmente (por ej. VLDL y proteínas séricas). La
secreción de VLDL no sólo distribuye el exceso de calorías hacia el tejido adiposo para su
almacenamiento, sino también fosfolípidos y colesterol hacia otros tejidos. Además, el hígado puede
convertir aminoácidos en glucosa, ácidos grasos y cuerpos cetónicos.
Músculo esquelético
El músculo esquelético representa casi la mitad del peso corporal. A pesar de usar una gran
parte de la glucosa que consumimos diariamente, el músculo no realiza gluconeogénesis, no puede
desfosforilar a la glucosa 6-P y por lo tanto no puede generar glucosa libre y contribuir al
mantenimiento de la glucemia. El músculo carece de receptores para glucagon y por lo tanto, no
responde a las variaciones en los niveles sanguíneos de esta hormona. Las grandes reservas de

20
glucógeno muscular no pueden ser movilizadas para mantener la glucemia, pero son importantes para
el metabolismo energético local, a través de la activación del sistema adrenérgico. Si se realiza
metabolismo anaeróbico, se incrementa la formación de lactato que puede ser transportado a otros
tejidos. El corazón usa grandes cantidades de lactato producido por otros tejidos. A diferencia del
hígado, el músculo esquelético carece de actividad de ácido grasa sintetasa y por lo tanto no puede
sintetizar ácidos grasos ni triglicéridos. Sin embargo, el paso inicial en la síntesis de ácidos grasos (la
síntesis de malonilCoA) está activo y sujeto al control de la AMP Kinasa. Como ya mencionamos, el
malonilCoA regula el transporte de ácidos grasos en la membrana mitocondrial interna y por lo tanto la
velocidad de oxidación de ácidos grasos en el músculo esquelético.
Corazón
El metabolismo del músculo cardíaco difiere del metabolismo del músculo esquelético en
diferentes aspectos. El metabolismo cardíaco en condiciones normales es completamente aeróbico,
mientras que el músculo esquelético puede funcionar anaeróbicamente por períodos cortos. El
corazón no contiene reservas apreciables de combustibles metabólicos, ya sea glucógeno o lípidos,
aunque contiene una pequeña cantidad de fosfocreatina. Por lo tanto, el aporte de oxígeno y
nutrientes debe ser continuo. El corazón puede utilizar ácidos grasos, cuerpos cetónicos, glucosa y
lactato. Luego de una ingesta, predomina la utilización de lactato y glucosa, pero en el ayuno o
durante el ejercicio predomina la de ácidos grasos. Recién en condiciones extremas, como carreras
de 100 m o cuando la irrigación disminuye (como en la ateroesclerosis o infarto de miocardio) puede
realizar glucólisis anaeróbica.
Cerebro
El cerebro debe generar grandes cantidades de ATP para mantener el potencial de membrana,
lo que resulta esencial para la transmisión de los impulsos nerviosos. En condiciones normales, el
cerebro sólo usa glucosa como combustible, oxidándola a través de la glucólisis aeróbica. No utiliza
ácidos grasos, pues éstos no atraviesan la barrera hematoencefálica. De hecho, el 60% del total de
glucosa consumida por el organismo es utilizado por el cerebro. El metabolismo del cerebro es
totalmente aeróbico, consume el 20% del total del oxígeno consumido por el organismo. No posee
reservas apreciables de glucógeno u otros combustibles, por lo que requiere del aporte constante de
oxígeno y glucosa que atraviesan la barrera hematoencefálica con facilidad. Después de 5-10 días de
ayuno, el cerebro comienza a utilizar cuerpos cetónicos además de glucosa, reduciendo notoriamente
el consumo de glucosa.
¿Por qué los cuerpos cetónicos reemplazan sólo parcialmente los requerimientos de glucosa del cerebro? Las células gliales se alinean con los vasos sanguíneos que irrigan el cerebro y forman una barrera que los ácidos grasos no pueden cruzar. Las células gliales captan glucosa y la metabolizan por glucólisis anaeróbica a lactato, que es exportado hacia las regiones más internas del cerebro. Allí, el lactato sirve como sustrato para el metabolismo aeróbico. Por lo tanto, las células de la glía parecen ser parcialmente dependientes del metabolismo anaeróbico y los cuerpos cetónicos no son sustratos de esta vía metabólica. Esto bien puede explicar la dependencia del SNC por glucosa, además de los cuerpos cetónicos, aun en estado de inanición.
Tejido adiposo
La función principal del tejido adiposo es almacenar combustibles lipídicos en forma de TAG.
Como ya se ha descripto, el transporte de glucosa a los adipocitos es un mecanismo que depende de
insulina. Los ácidos grasos se sintetizan a partir del acetilCoA que proviene del piruvato y de NADPH
(procedente de la vía de las pentosas). El glicerol 3 fosfato necesario para la síntesis de TAG proviene
de la reducción de un intermediario de la glucólisis, la dihidroxiacetona fosfato, ya que los adipocitos

21
carecen de la capacidad de fosforilar al glicerol, por no expresar la enzima glicerol quinasa. Por lo
tanto, la síntesis de TAG depende absolutamente de glucosa. La glucosa funciona como sensor del
metabolismo del tejido, cuando sus niveles son adecuados, se produce glicerol 3 fosfato para la
síntesis de TAG y cuando los niveles son bajos, los ácidos grasos sintetizados se liberan de los
adipocitos y son utilizados por otros tejidos.
Eritrocitos
El eritrocito es una célula especializada en el transporte de oxígeno, dióxido de carbono y
protones. La falta de mitocondrias le impide un metabolismo aeróbico, reduciendo sus posibilidades de
obtención de energía a la glucólisis anaeróbica con formación de lactato, que pasa a sangre y puede
utilizarse como combustible por tejidos como el corazón o como precursor gluconeogenético por el
hígado. La reducción del piruvato a lactato es fundamental para reoxidar el NADH y permitir que la
glucólisis siga ocurriendo. Los requerimientos energéticos del eritrocito se basan en el mantenimiento
de su forma bicóncava (que asegura un transporte adecuado de gases e impide la remoción por las
células del sistema retículo endotelial) y de su ambiente iónico interno. Además de la glucólisis
anaeróbica, el eritrocito utiliza la glucosa para obtener NADPH por la vía de las pentosas. El NADPH
se utiliza para evitar la oxidación del hierro de la hemoglobina, que en esa forma no podría transportar
el oxígeno. Además, el NADPH es necesario para mantener elevados los niveles de glutatión reducido
que se emplea como defensa contra los radicales libres del oxígeno. Finalmente, la glucosa también
es utilizada en el glóbulo rojo en una derivación de la vía glucolítica para sintetizar 2,3 difosfoglicerato
(2,3 DPG) a partir de 1,3 difosfoglicerato, cuya función es disminuir la afinidad de la hemoglobina por
el oxígeno, permitiendo una mayor liberación de oxígeno en los tejidos.
REGULACIÓN HORMONAL DE LA UTILIZACIÓN DEL COMBUSTIBLE METABÓLICO
Las principales hormonas que intervienen en la regulación de la glucemia son la insulina, el
glucagon y las catecolaminas. También son importantes los glucocorticoides y la hormona de
crecimiento.
La concentración de glucosa en sangre es el estímulo primario para la secreción de insulina y
glucagon. La hipoglucemia estimula la secreción de glucagon que favorece la degradación del
glucógeno hepático (pero no del muscular). Una hipoglucemia severa induce la secreción de
adrenalina de la médula adrenal que estimula la degradación de glucógeno muscular. El glucagon y la
adrenalina promueven además la lipólisis en el tejido adiposo, aumentando la concentración
sanguínea de ácidos grasos y aportando sustrato para la cetogénesis hepática.
Por el contrario, la hiperglucemia induce un aumento en la secreción de insulina. Al igual que
los hepatocitos, las células expresan el transportador de glucosa de baja afinidad GLUT2 y la
enzima glucoquinasa de baja afinidad por la glucosa. Por ese motivo, sólo en presencia de altas
concentraciones de glucosa en sangre se produce captación y fosforilación de glucosa en estas
células. La subsecuente metabolización de la glucosa-6-fosfato parece ser el evento que
desencadena la liberación de insulina.
Un balance adecuado de estas dos hormonas antagónicas permite una adecuada homeostasis
de la glucemia, efecto que puede perderse en condiciones de deficiencia de insulina (como la
diabetes), quemaduras graves y shock hemorrágico. Además la concentración sanguínea de glucagon
se reduce por la somatostatina, hormona que también inhibe la secreción de somatotrofina (hormona
de crecimiento).
Insulina
En términos generales la insulina promueve:
1) la captación de sustratos por determinadas células

22
2) el almacenamiento de combustibles como glucógeno y lípidos
3) la biosíntesis de macromoléculas como ácidos nucleicos y proteínas
Más en detalle, la insulina produce:
-Aumento de la permeabilidad a la glucosa en músculo y tejido adiposo, por exposición del
transportador GLUT4 en las membranas plasmáticas.
- Activación de la glucólisis y de la vía de las pentosas en el hígado y tejido adiposo por
inducción de las enzimas clave (fosfofructoquinasa y glucosa-6-fosfato deshidrogenasa,
respectivamente), lo que permite la generación de energía y de precursores para la síntesis de AG
(NADPH y acetilCoA) y de TAG (glicerol-3-fosfato, proveniente de la reducción de la dihidroxiacetona
fosfato).
- Activación de la síntesis de ácidos grasos y triacilglicéridos en hígado y tejido adiposo, al
aumentar el aporte de precursores y por inducción de la enzima acetilCoA carboxilasa. De esta
manera se favorece la formación de la lipoproteína VLDL en el hígado, que por sangre llega a los
tejidos periféricos donde es sustrato de la lipoproteín- lipasa, enzima cuya síntesis también se induce
por insulina.
- Inhibición de la gluconeogénesis en hígado al disminuir las actividades de fructosa- 1,6-
difosfatasa y de fosfoenolpiruvato carboxiquinasa. Este efecto se debe en parte al aumento de los
niveles de fructosa-2,6-difosfato.
- Aumento de la síntesis e inhibición de la degradación de glucógeno en hígado y músculo.
A nivel hepático, se ha descripto recientemente una proteína quinasa B, que se activa por fosforilación
desencadenada por insulina. La proteína quinasa B fosforila a una enzima denominada glucógeno
sintasa quinasa (GSK), inactivándola. De esta forma, no fosforila a la glucógeno sintasa y ésta se
mantiene activa, con la consecuente síntesis de glucógeno. Tanto en el hígado como en el músculo, la
activación de fosfodiesterasas disminuye los niveles de AMPc inactivándose por lo tanto la proteína
quinasa A. Además, la insulina activa fosfatasas con lo cual disminuye el grado de fosforilación de la
fosforilasa quinasa y de la glucógeno fosforilasa inactivando ambas enzimas. En consecuencia
disminuye la glucogenólisis. La desfosforilación de enzimas también contribuye a mantener la
glucógeno sintasa en estado activo.
-Efecto antilipolítico debido a la disminución en los niveles de AMPc intracelular por activación de
fosfodiesterasas. También contribuye la activación de fosfatasas. En consecuencia, se inactiva la
lipasa hormono sensible y se reduce la hidrólisis de TAG en tejido adiposo.
- Aumento de la captación de aminoácidos en músculo, con incremento de la síntesis de
proteína muscular.
Glucagon
El principal órgano blanco del glucagon es el hígado, donde interactúa con receptores
específicos acoplados a la adenilato ciclasa. Su mecanismo de acción involucra el incremento en los
niveles de AMPc. Consecuentemente se activan la glucogenólisis (glucógeno fosforilasa fosforilada y
por lo tanto, activa) y se inhibe la glucogenogénesis (glucógeno sintetasa fosforilada y por lo tanto,
inactiva). Además por fosforilación de la FFQ2 disminuye su actividad de quinasa y aumenta su
actividad de fosfatasa, con lo que se hidroliza la fructosa-2,6-difosfato, se inhibe la glucólisis y se
activa la gluconeogénesis. El glucagon también inhibe a la piruvato quinasa hepática causando una
acumulación de PEP y una disminución en los niveles de piruvato. La acumulación de PEP promueve
la gluconeogénesis, mientras que la inhibición de la piruvato quinasa disminuye la actividad de la vía
glucolítica.
Varias enzimas se inducen cuando los niveles de glucagon son elevados, fundamentalmente
las que catalizan los pasos claves de la gluconeogénesis, la glucosa-6-fosfatasa, la fosfoenolpiruvato

23
carboxiquinasa y la fructosa-1,6-difosfatasa.
El glucagon también aumenta los niveles de AMPc en el tejido adiposo, donde promueve la
movilización de los depósitos de TAG por activación de la lipólisis (activa la lipasa hormono sensible).
Asimismo, al aumentar el nivel de ácidos grasos libres y su captación por el hígado, los acilCoA
inhiben alostéricamente la acetilCoA carboxilasa, que también se inhibe por fosforilación inducida por
glucagon. Como resultado, la síntesis de ácidos grasos en el hígado se inhibe.
Catecolaminas
Las catecolaminas, adrenalina y noradrenalina, funcionan como hormonas al liberarse a la
sangre por la médula adrenal. La hipoglucemia es uno de los estímulos para la liberación de estas
hormonas. En el músculo, sus receptores están acoplados a la adenilato ciclasa y se produce un
incremento en los niveles de AMPc activando la glucogenólisis. En consecuencia, la captación de
glucosa por el músculo disminuye y la glucemia aumenta. La adrenalina también inhibe la secreción
de insulina y estimula la de glucagon. Este efecto aumenta la producción y liberación de glucosa por el
hígado con incremento en la glucemia. Al igual que el glucagon, la adrenalina estimula la lipólisis (a
través de la proteína quinasa A).
Glucocorticoides
Los glucocorticoides tienen acciones diversas que afectan la mayoría de los tejidos del
organismo, alguno de estos efectos parecerían ser contradictorios, pero si se los analiza en conjunto,
se puede ver que promueven la sobrevida en situaciones de estrés. Son moduladores lentos del
metabolismo de los combustibles circulantes. Sus efectos se relacionan con la regulación de la
expresión de enzimas clave de varias vías metabólicas. Inducen la expresión de las enzimas de la
gluconeogénesis: piruvato carboxilasa, fosfoenolpiruvato carboxiquinasa, fructosa-1,6-difosfatasa y
glucosa-6-fosfatasa. Asimismo, por sus efectos catabólicos sobre las proteínas tisulares en
condiciones de estrés, aportan sustratos para la gluconeogénesis. Además, facilitan la lipólisis
inducida por otros agentes como glucagón y catecolaminas.
Hormona de crecimiento
Ejerce un efecto hiperglucemiante y lipolítico, es decir, opuesto al de la insulina. En ese
sentido, disminuye la utilización periférica de glucosa y estimula su producción hepática por
gluconeogénesis. Se ha demostrado también que la hormona de crecimiento acentúa la movilización
de grasas de los depósitos, incrementando los niveles de glicerol y ácidos grasos libres en plasma.