Embed Size (px)
DESCRIPTION
bioquimica de nucleotidos
Citation preview

Q.F. Juan Salazar Sánchez

NucleótidosSon ésteres de fosfato de pentosas en las cuales una base nitrogenada está
unida al C1´ de una azúcar.
En los ribonucleótidos la pentosa es la D-ribosa, en los
desoxiribonucleótidos (DNA, desoxinucleótidos), el azúcar es 2´-desoxi-D-
ribosa (los números primos señalan posiciones de átomos del azúcar, los no
primos corresponden a la base nitrogenada).
El fosfato puede estar en posición 3´ó 5´. Si no hay fosfato en la molécula, el
compuesto es un nucleósido.
En general estos compuestos son ácidos moderadamente fuertes.
Las bases nitrogenadas son moléculas planas, aromáticas y heterocíclicas
que son derivados de la purina o pirimidina

Funciones metabólicas de los nucleótidos
Todos los tipos de células contienen una gran variedad de nucleótidos y
derivados. Sus principales funciones son las siguientes:
a) Participación en el metabolismo energético. El ATP (trifosfato de adenosina)
se genera en las células mediante fosforilación, oxidativa y a nivel de sustrato.
Se utiliza para activar las reacciones metabólicas, y está implicado en procesos
tales como: contracción muscular, transporte activo y mantenimiento de la
integridad de las membranas celulares. El ATP es un dador de fosfato para la
generación de otros nucleósidos 5´-trifosfatos .
b) Presencia en unidades monoméricas de ácidos nucleicos. ADN y ARN,
están compuestos por unidades de nucleótidos. En reacciones de síntesis de
ácidos nucleicos, los nucleósidos 5´-trifosfatos son sustratos que se unen al
polímero por enlaces fosfodiéster 3´-5´, liberando pirofosfato

c) Mediadores fisiológicos de procesos metabólicos clave. AMPc (monofosfato
de adenina cíclico) actúa como segundo mensajero en el control de los
procesos glucógenolisis y glucogénesis, mediados por adrenalina y glucagón.
d) Componentes de coenzimas. NAD (dinucléotido de nicotinamida y
adenina), FAD (dinucleótido de adenina y flavina) y CoA (Coenzima A) son
constituyentes metabólicos implicados en gran número de rutas metabólicas.
e) Intermediarios activados, necesarios para diversas reacciones. UDP-
glucosa es un intermediario clave en la síntesis de glucógeno y de
glucoproteínas. GDP-manosa, GDP-fucosa, UDP-galactosa y CMP-ácido
siálico son intermediarios clave de reacciones en las que azúcares son
transferidos para la síntesis de glucoproteínas. CTP se utiliza para generar
CDP-colina, CDP-etanolamina y CDP-diacilgliceroles, compuestos que
intervienen en metabolismo de fosfolípidos.
f) Efectores alostéricos. Las concentraciones intracelulares de nucleótidos
controlan los pasos regulados en las vías metabólicas.
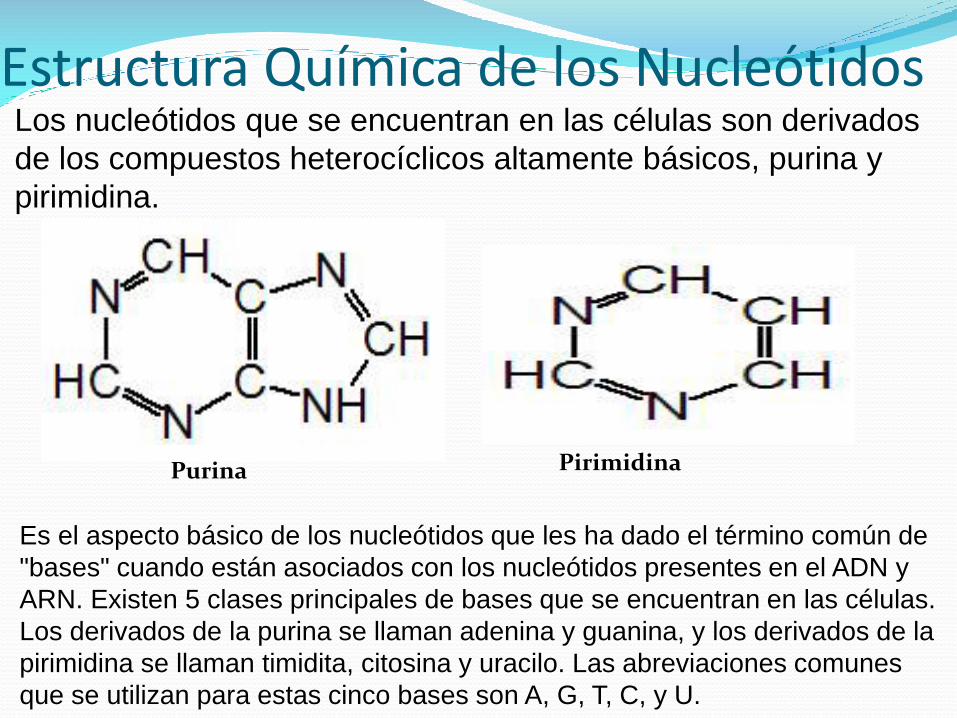
Estructura Química de los NucleótidosLos nucleótidos que se encuentran en las células son derivados
de los compuestos heterocíclicos altamente básicos, purina y
pirimidina.
Es el aspecto básico de los nucleótidos que les ha dado el término común de
"bases" cuando están asociados con los nucleótidos presentes en el ADN y
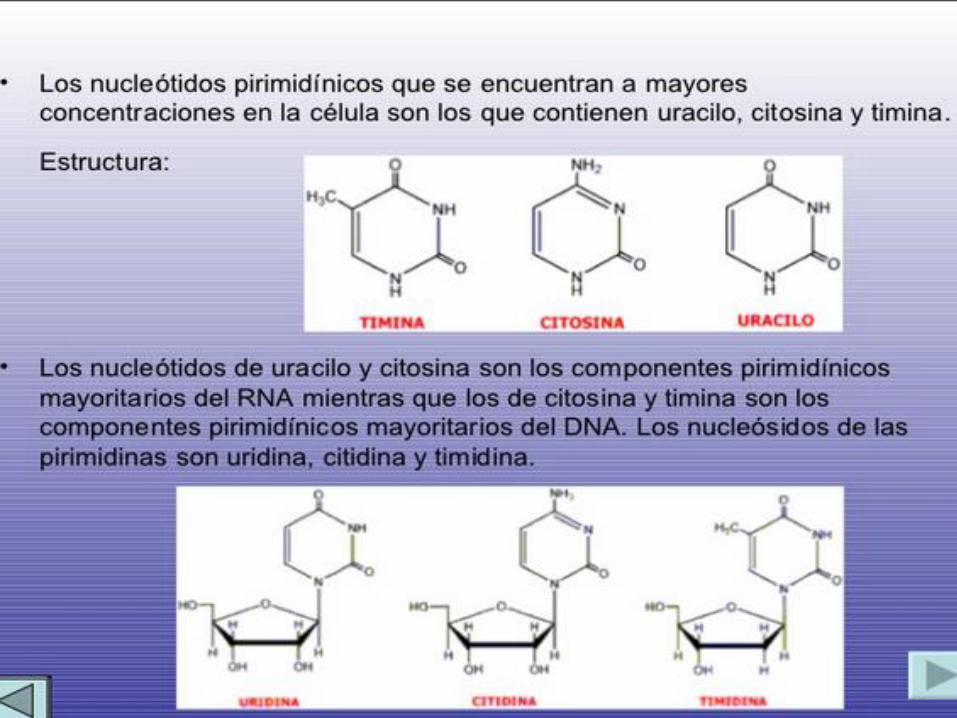
ARN. Existen 5 clases principales de bases que se encuentran en las células.
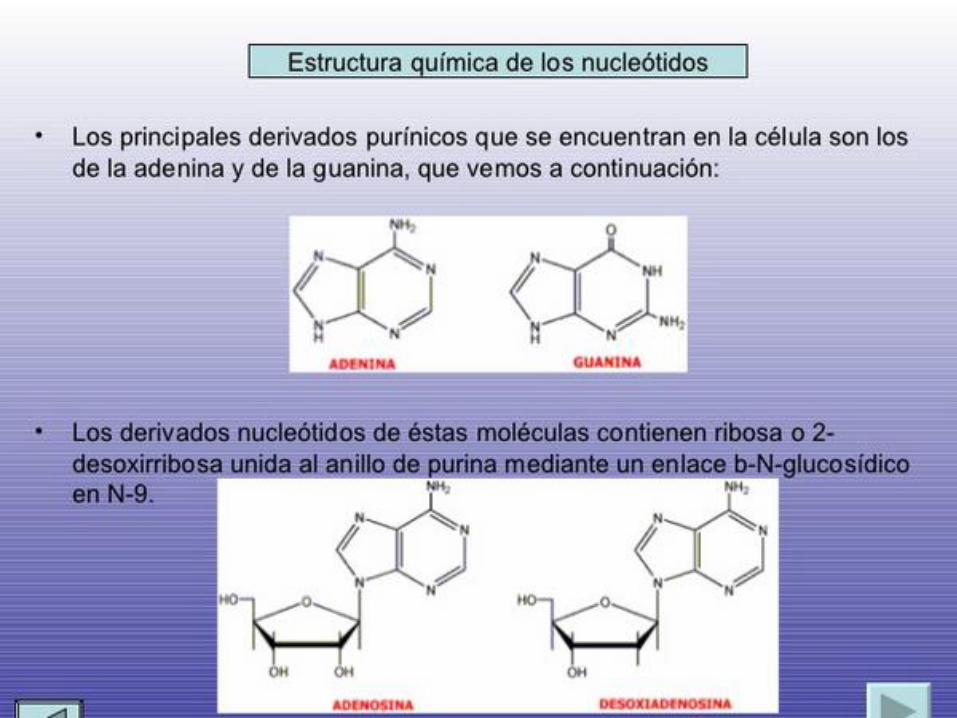
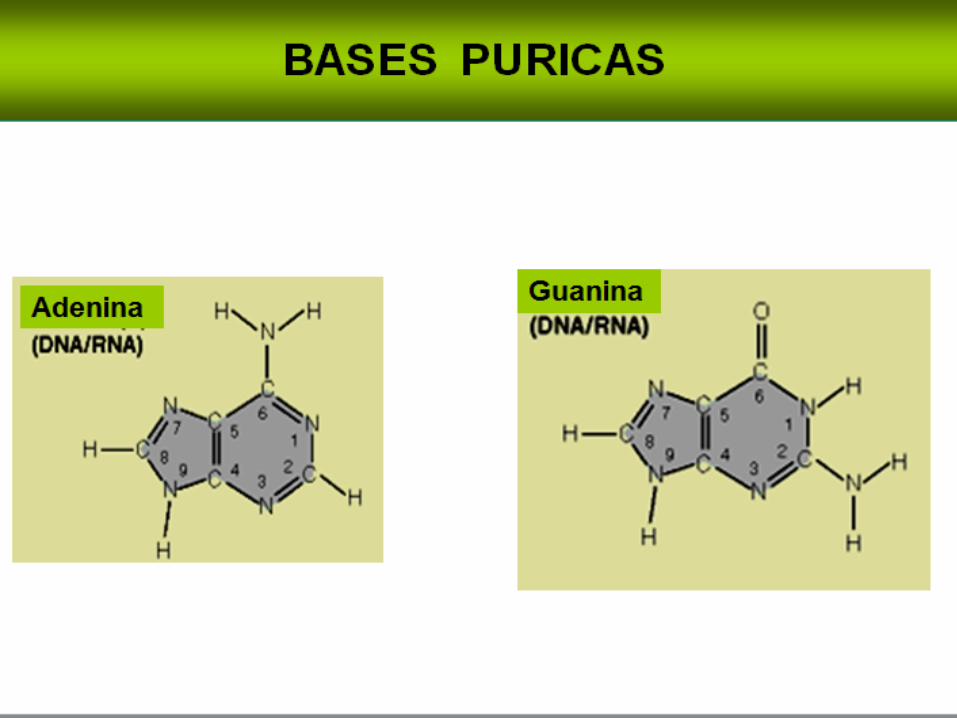
Los derivados de la purina se llaman adenina y guanina, y los derivados de la
pirimidina se llaman timidita, citosina y uracilo. Las abreviaciones comunes
que se utilizan para estas cinco bases son A, G, T, C, y U.
Purina Pirimidina

Metabolismo de los Nucleótidos

Biosíntesis y Degradación de Nucleótidos
Los nucleótidos son substituyentes ubicuos en la naturaleza que participan
casi en todos los procesos bioquímicos.
1.- Forman las unidades monoméricas de los ácidos nucleicos, que son
sintetizados directamente de nucleósidos trifosfatados, la forma activada
de los nucleótidos.
2.- De los nucleósidos trifosfatados el que participa como donador de
energía química en un mayor número de procesos, es el adenosín
trifosfato (ATP) que además es el producto final de la gran mayoría de los
procesos metabólicos. Muchos intermediarios activados como la UDP-
glucosa en la síntesis de glucógeno, contienen nucleótidos.

3.- Muchas vías metabólicas están reguladas al menos en parte por los
niveles de ATP, ADP o AMP.
De manera semejante, muchas señales hormonales como aquellas que
controlan al metabolismo del glucógeno, son mediadas intracelularmente
por las moléculas cíclicas del AMP o GMP (cAMP o cGMP).
4.- Los nucleótidos de adenina son componentes de las coenzimas
NAD+, NADP+, FMN, FAD y CoA.
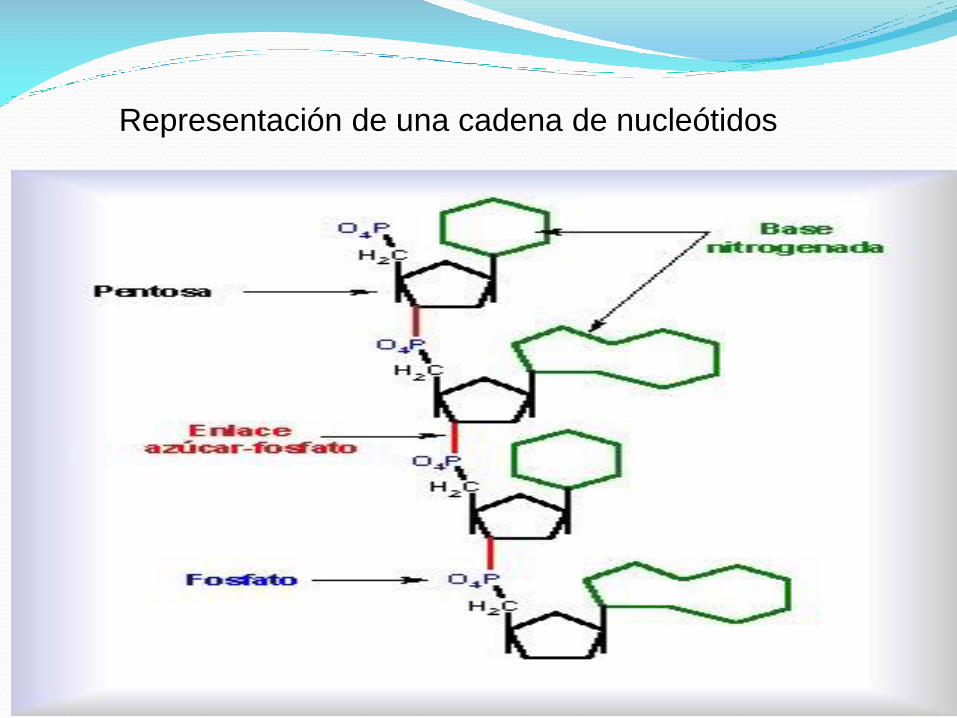
Representación de una cadena de nucleótidos
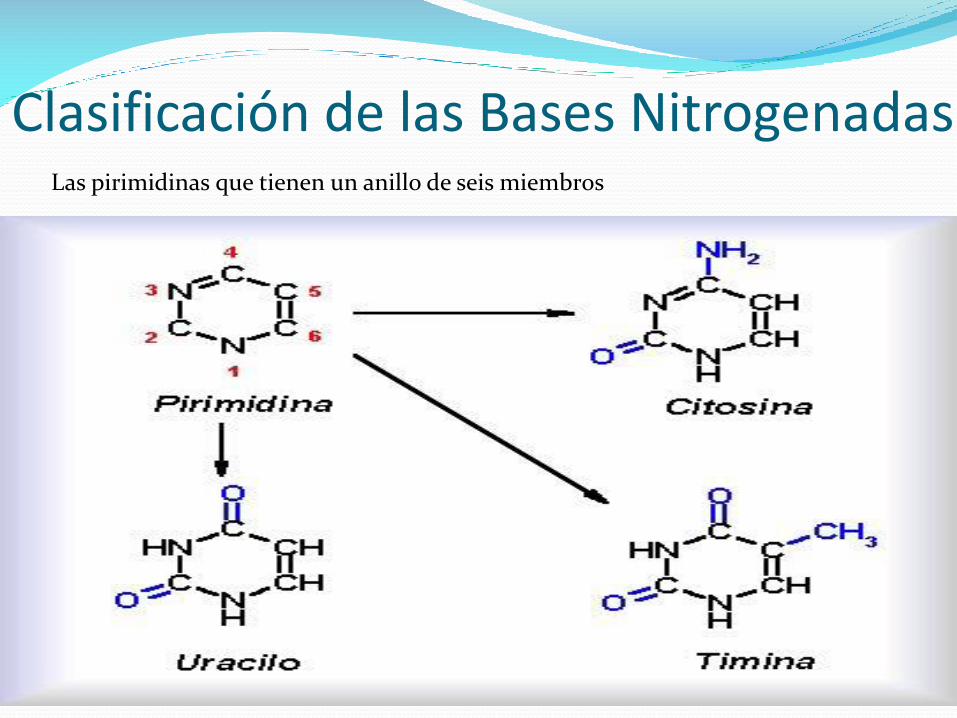
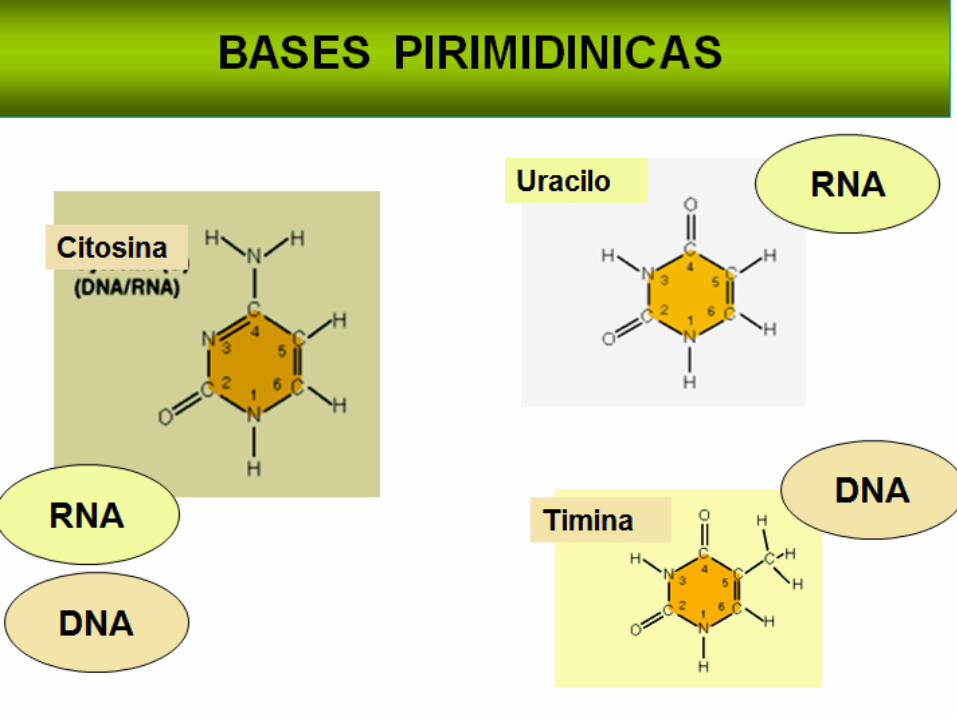
Clasificación de las Bases NitrogenadasLas pirimidinas que tienen un anillo de seis miembros
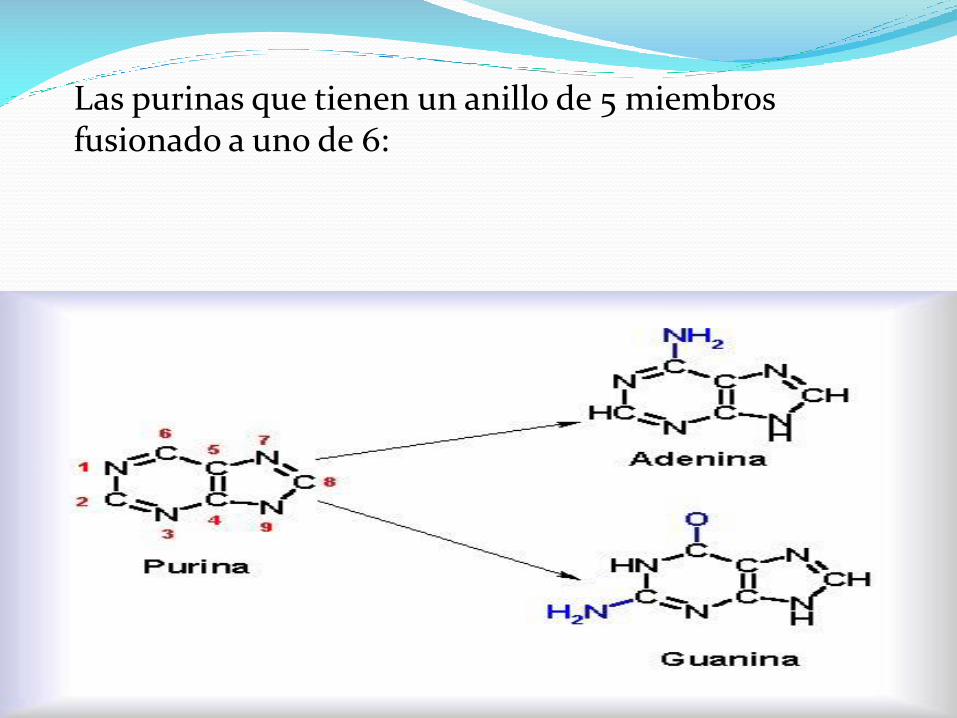
Las purinas que tienen un anillo de 5 miembros fusionado a uno de 6:

Cada ácido nucleico contiene 4 tipos de bases.
Las mismas dos purinas, adenina (A) y guanina (G) están
presentes en el ADN y en el ARN.
Las dos pirimidinas, en el ADN son citosina (C) y timina (T), en
el ARN, la timina se cambia por el uracilo (U).
La única diferencia entre el uracilo y la timina es la presencia
de un substituyente metil en la posición C5.
De tal manera que en el ADN solo existen A,G,C y T, mientras
que en el ARN contiene A,G,C y U.
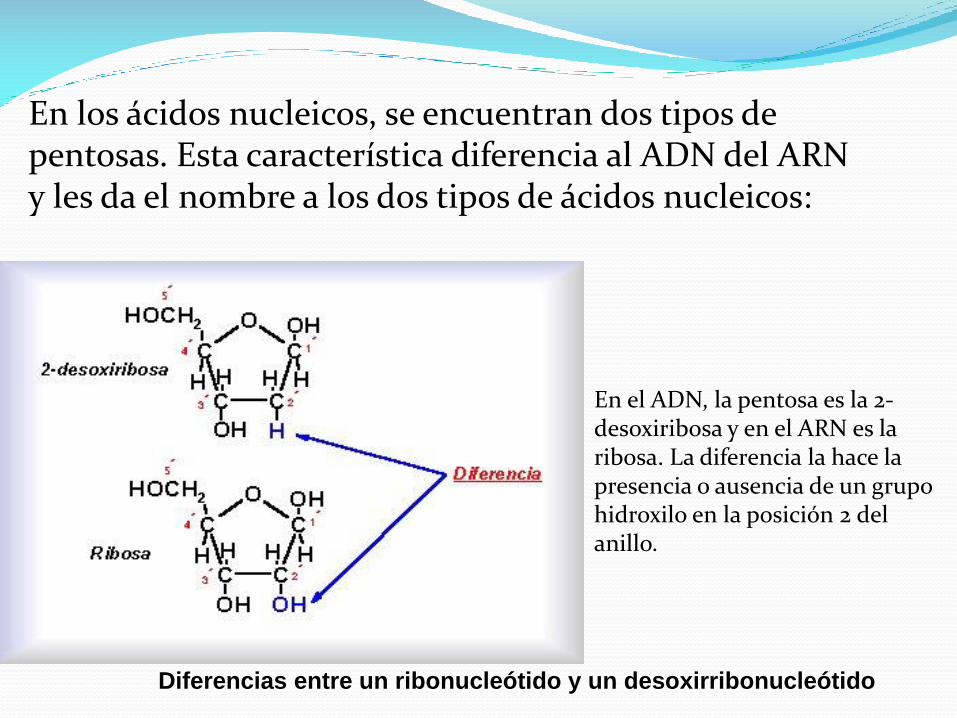
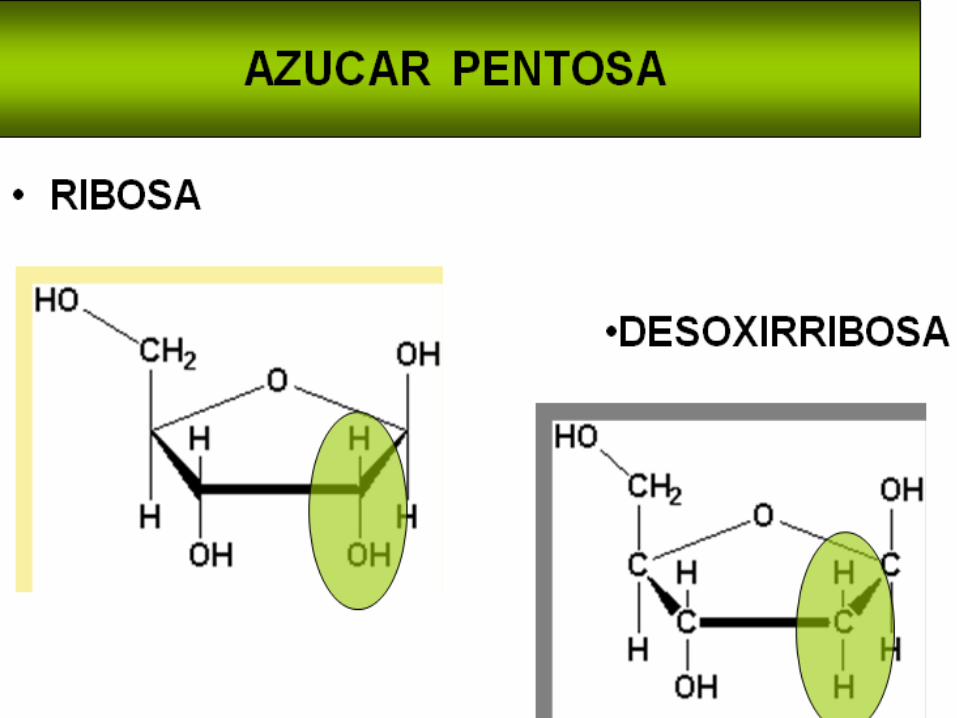
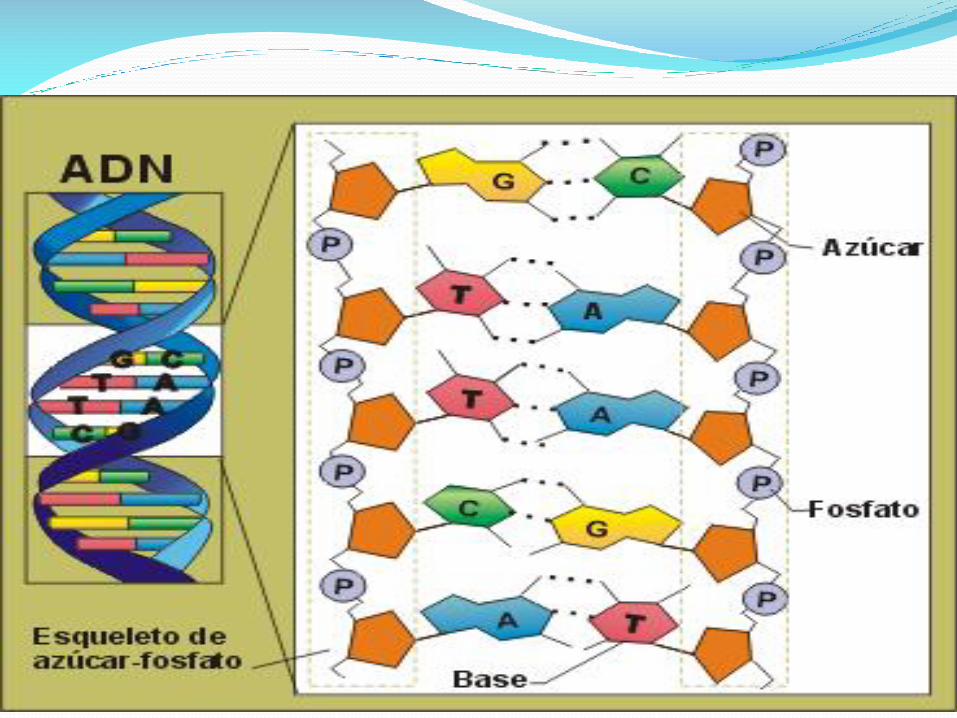
En los ácidos nucleicos, se encuentran dos tipos de pentosas. Esta característica diferencia al ADN del ARN y les da el nombre a los dos tipos de ácidos nucleicos:
Diferencias entre un ribonucleótido y un desoxirribonucleótido
En el ADN, la pentosa es la 2-desoxiribosa y en el ARN es la ribosa. La diferencia la hace la presencia o ausencia de un grupo hidroxilo en la posición 2 del anillo.

La base nitrogenada esta unida a la posición 1 del anillo de
la pentosa por medio de un enlace glucosídico a la posición
N1 de las pirimidinas o a la N9 de las purinas.
Para evitar ambigüedades entre la numeración de los
heterociclos y el anillo del azúcar, las posiciones de la
pentosa se numeran como primos (´).
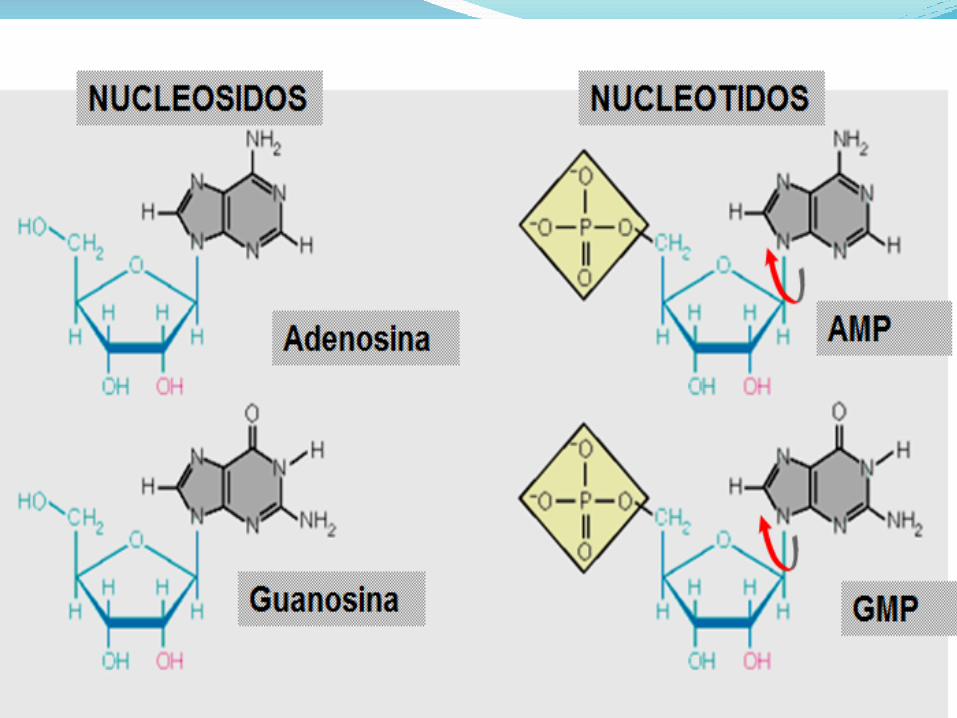
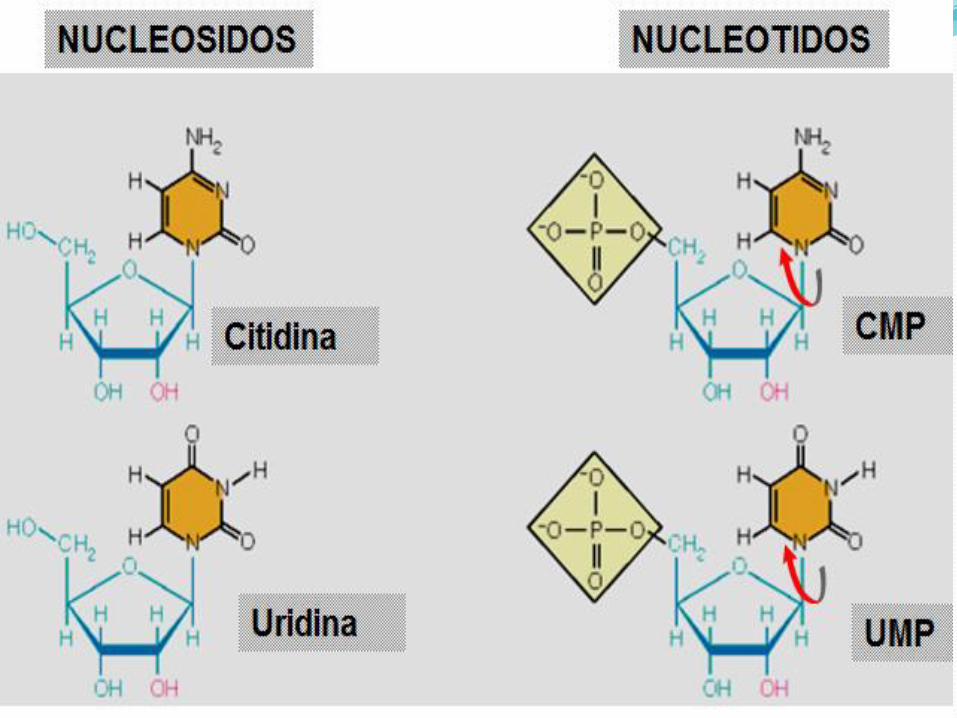
Una base unida a una azúcar se denomina nucleósido,
cuando se une un fosfato, la base-azúcar-fosfato se
denomina nucleotido:

Los diferentes arreglos de los nucleótidos.

Los nucleótidos son los bloques de construcción a partir de los cuales se
construyen los ácidos nucleicos.
Los nucleótidos están unidos en una cadena polinucleotídica con un
esqueleto que consiste de series alternadas de residuos de azúcar y fosfato.
La posición 5´ de un anillo de pentosa está conectada a la posición 3´ de la
siguiente pentosa vía un grupo fosfato . Por lo tanto, se dice que el esqueleto
de azúcar-fosfato consiste de un enlace fosfodiester en posición 5´-3´. Las
bases nitrogenadas están por fuera del esqueleto.
El nucleótido terminal en uno de los extremos de la cadena tiene el grupo 5´
libre; y al otro lado de la cadena tiene un grupo 3´ libre.
Es una convención escribir las secuencias de ácidos nucleicos en la
dirección 5´-3´, de tal forma que el extremo 5´ está a la izquierda y el 3´ a la
derecha.
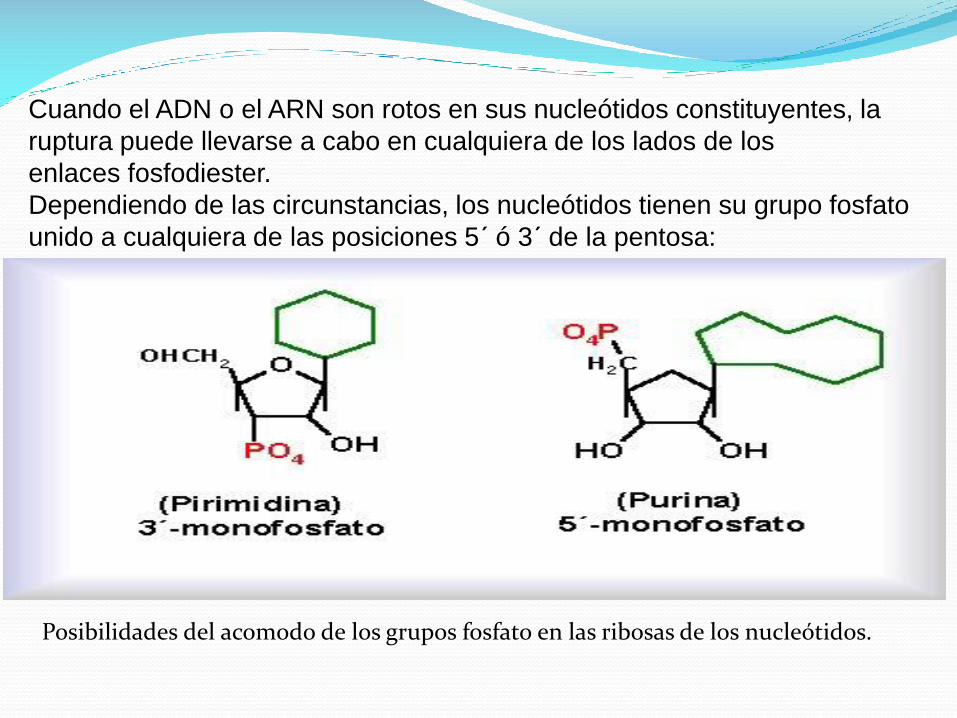
Cuando el ADN o el ARN son rotos en sus nucleótidos constituyentes, la
ruptura puede llevarse a cabo en cualquiera de los lados de los
enlaces fosfodiester.
Dependiendo de las circunstancias, los nucleótidos tienen su grupo fosfato
unido a cualquiera de las posiciones 5´ ó 3´ de la pentosa:
Posibilidades del acomodo de los grupos fosfato en las ribosas de los nucleótidos.
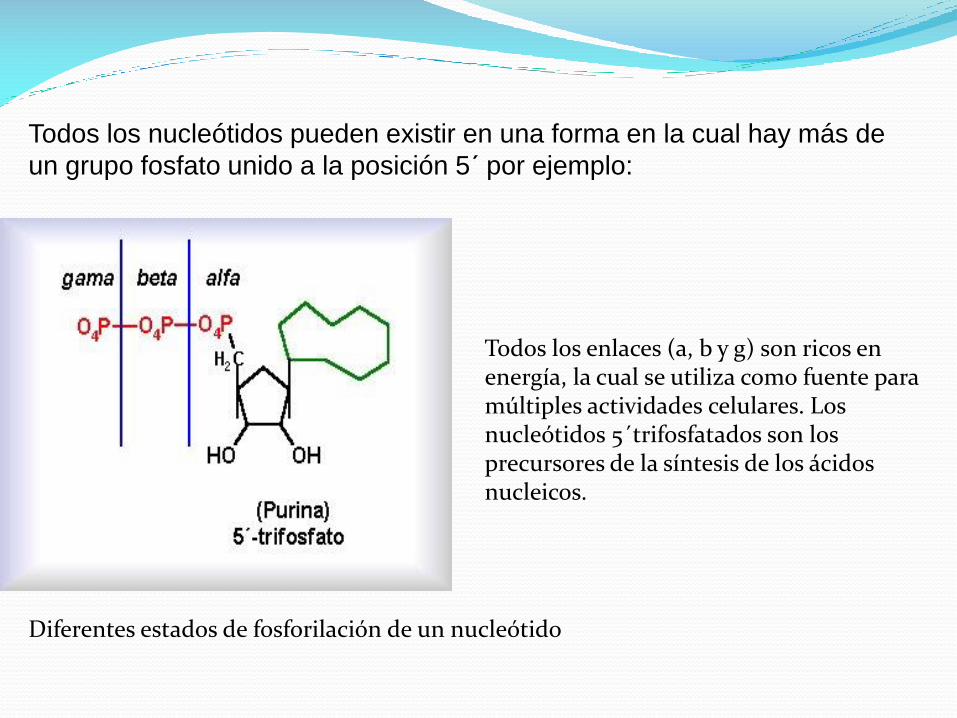
Todos los nucleótidos pueden existir en una forma en la cual hay más de
un grupo fosfato unido a la posición 5´ por ejemplo:
Diferentes estados de fosforilación de un nucleótido
Todos los enlaces (a, b y g) son ricos en energía, la cual se utiliza como fuente para múltiples actividades celulares. Los nucleótidos 5´trifosfatados son los precursores de la síntesis de los ácidos nucleicos.

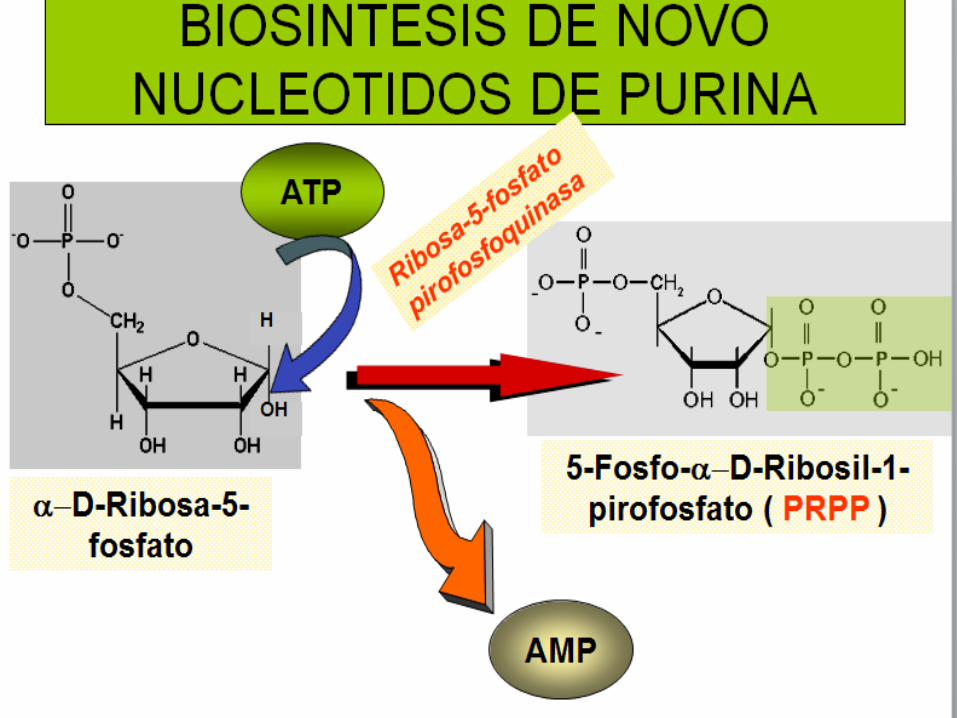
Síntesis de Ribonucleótidos de Purinas
Las primeras evidencias de las síntesis de novo de estos
compuestos se hallaron al alimentar palomas con una variedad
de compuestos isotópicamente marcados.
Se determinó químicamente las posiciones de los átomos
marcados en el producto de excreción, el ácido úrico (una
purina).
Se utilizó aves porque excretan el nitrógeno casi enteramente
como ácido úrico, substancia insoluble, fácilmente de aislar.
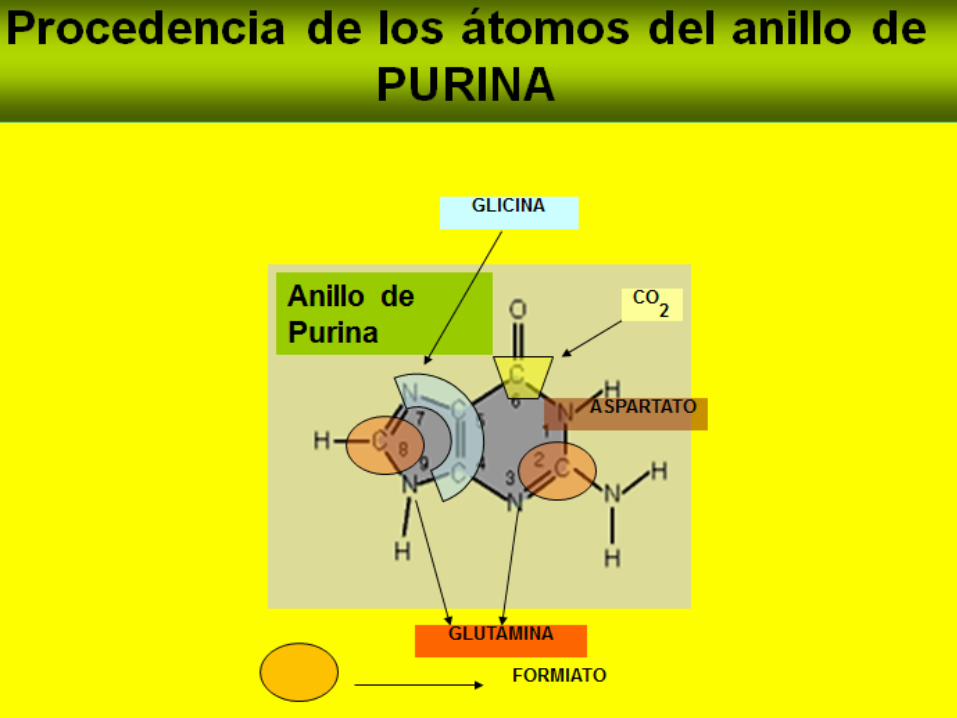
Los C4, C5 y N7 provienen de la glicina, mientras que cada uno
de los otros átomos, derivan de un precursor independiente.
El formato proviene del tetrahidrofolato, en el metabolismo de
unidades de un carbono.
El procedimiento consta de 11 reacciones
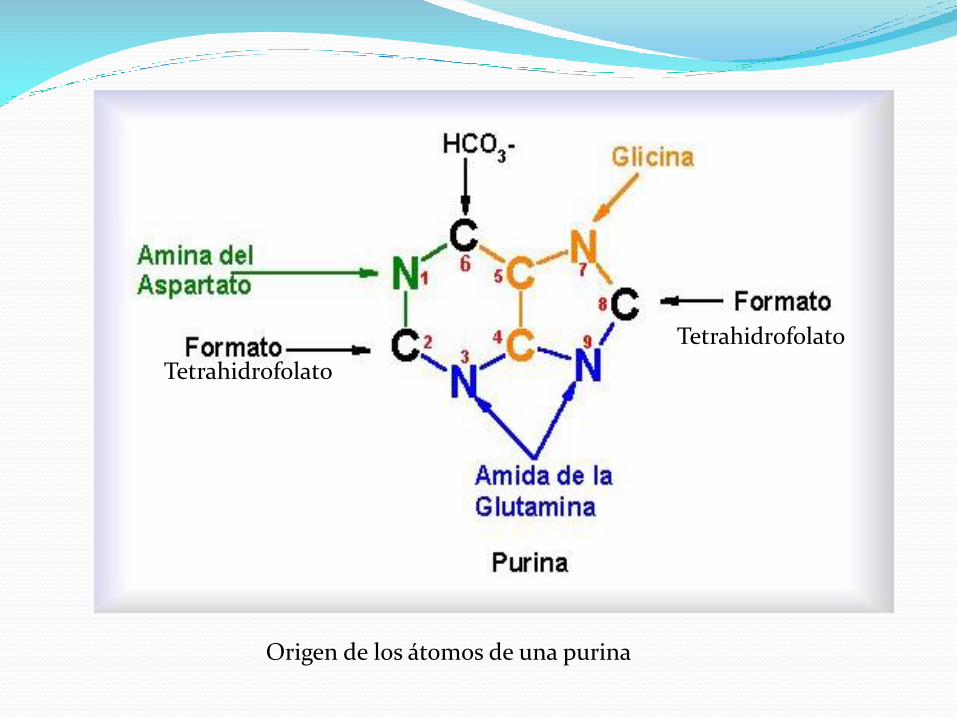
Origen de los átomos de una purina
Tetrahidrofolato
Tetrahidrofolato
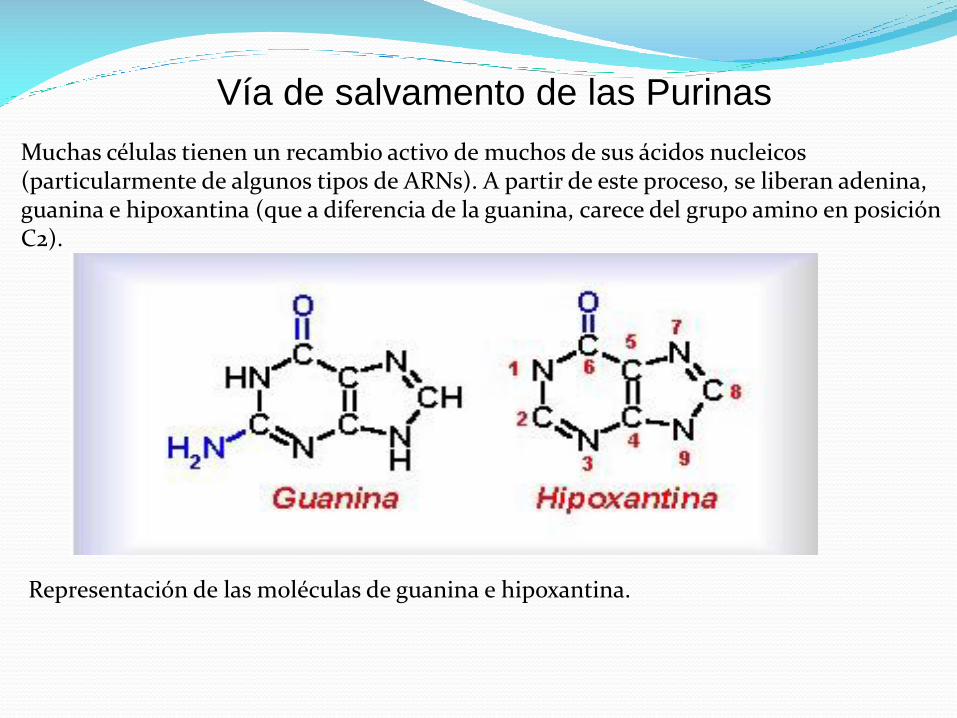
Vía de salvamento de las Purinas
Muchas células tienen un recambio activo de muchos de sus ácidos nucleicos (particularmente de algunos tipos de ARNs). A partir de este proceso, se liberan adenina, guanina e hipoxantina (que a diferencia de la guanina, carece del grupo amino en posición C2).
Representación de las moléculas de guanina e hipoxantina.

Estas purinas libres, son recuperadas para formar sus nucleótidos
correspondientes a través de vías de salvamento. A diferencia de la síntesis
de novo que es prácticamente idéntica en todas las células, las vías de
salvamento, son diversas. En los mamíferos, las purinas son salvadas
principalmente por medio de dos enzimas, la adenina fosforibosiltransferasa
(APRT), forma AMP a través de la transferencia de adenina al fosforibosil
pirofosfato con la liberación de PPi:
Adenina + PRPP AMP + PPi
Y la hipoxantina-guanina fosforibosiltransferasa (HGPRT), que cataliza una reacción análoga tanto para hipoxantina como para guanina:
Hipoxantina IMP + PPio + PRPP
Guanina GMP + PPi
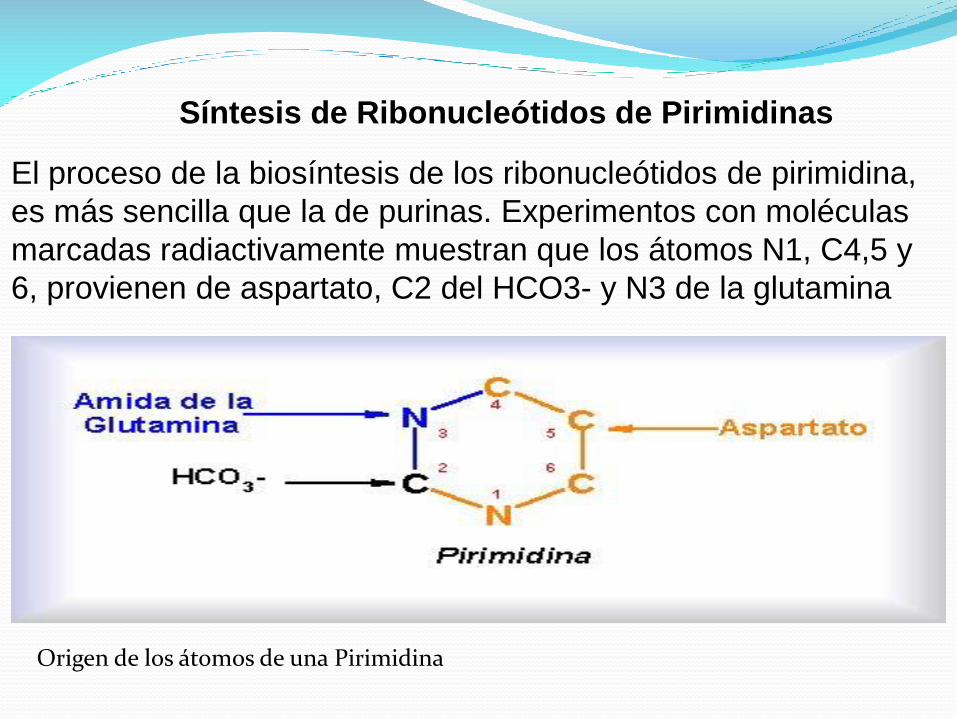
Síntesis de Ribonucleótidos de Pirimidinas
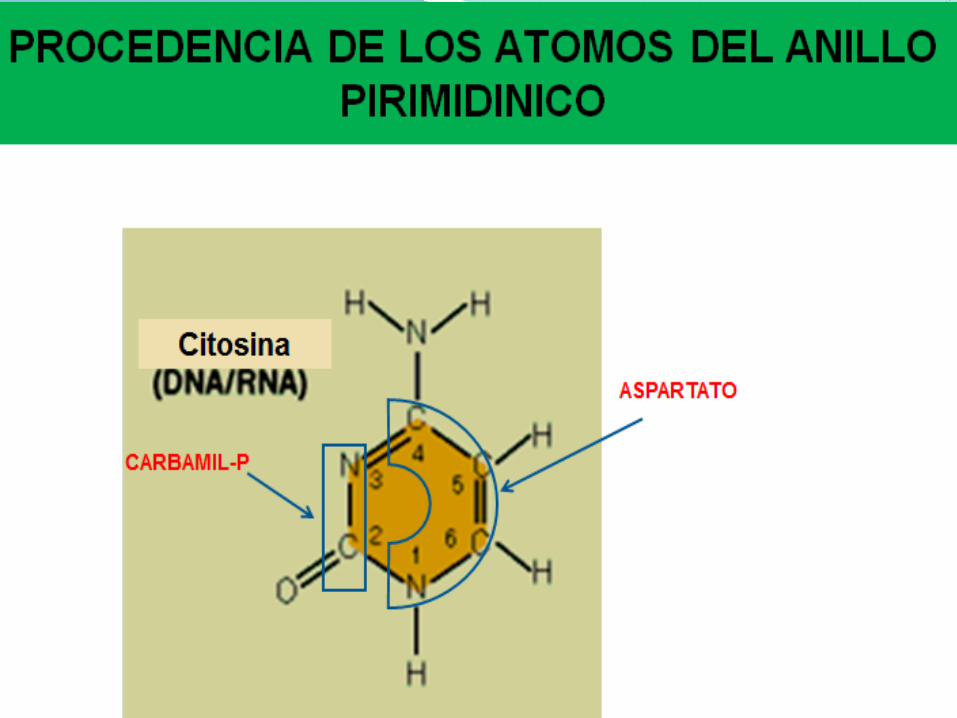
El proceso de la biosíntesis de los ribonucleótidos de pirimidina,
es más sencilla que la de purinas. Experimentos con moléculas
marcadas radiactivamente muestran que los átomos N1, C4,5 y
6, provienen de aspartato, C2 del HCO3- y N3 de la glutamina
Origen de los átomos de una Pirimidina

.
El ADN es una molécula que se ha organizado de
manera eficiente que permite almacenar la
información genética para que se pueda mantener a
través del tiempo y ser traspasada de
generación en generación entre las especies.
Organización genética del ADN

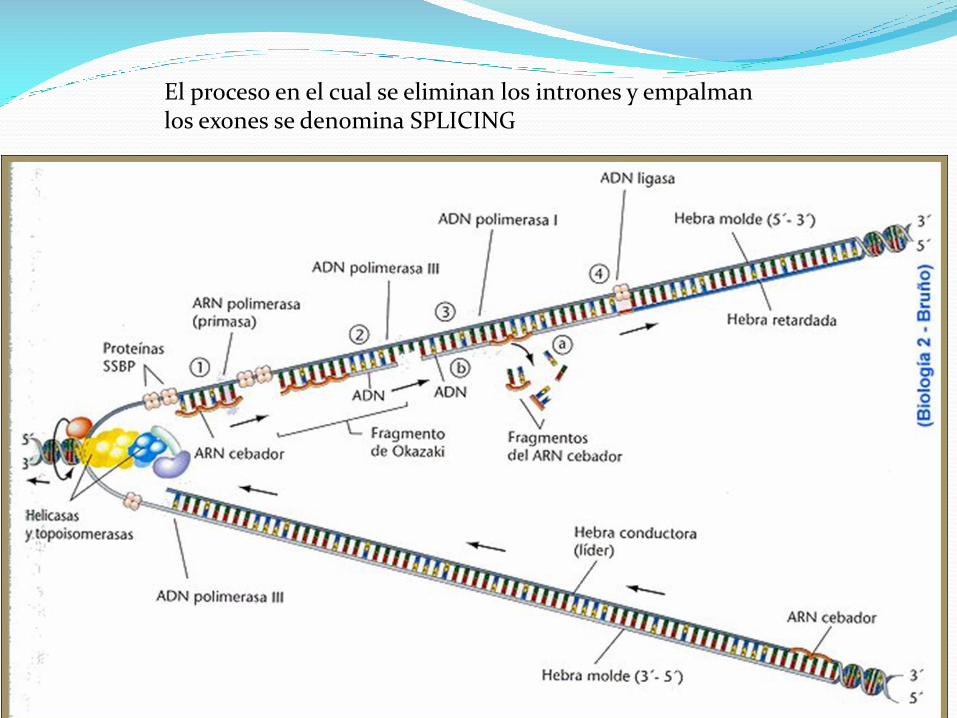
Replicacion del ADN
El proceso de replicación de ADN es el mecanismo que permite
al ADN duplicarse (es decir, sintetizar una copia idéntica).
De esta manera de una molécula de ADN única, se obtienen dos
o más "clones" de la primera.
Esta duplicación del material genético se produce de acuerdo
con un mecanismo semiconservativo, lo que indica que las dos
cadenas complementarias del ADN original, al separarse, sirven
de molde cada una para la síntesis de una nueva cadena
complementaria de la cadena molde, de forma que cada nueva
doble hélice contiene una de las cadenas del ADN original.

Gracias a la complementación entre las bases que forman la secuencia de
cada una de las cadenas, el ADN tiene la importante propiedad de
reproducirse idénticamente, lo que permite que la información genética se
transmita de una célula madre a las células hijas y es la base de la herencia
del material genético.
La molécula de ADN se abre como una cremallera por ruptura de los
puentes de hidrógeno entre las bases complementarias liberándose dos
hebras y la ADN polimerasa sintetiza la mitad complementaria añadiendo
nucleótidos que se encuentran dispersos en el núcleo.
De esta forma, cada nueva molécula es idéntica a la molécula de ADN
inicial.
La replicación empieza en puntos determinados: los orígenes de replicación
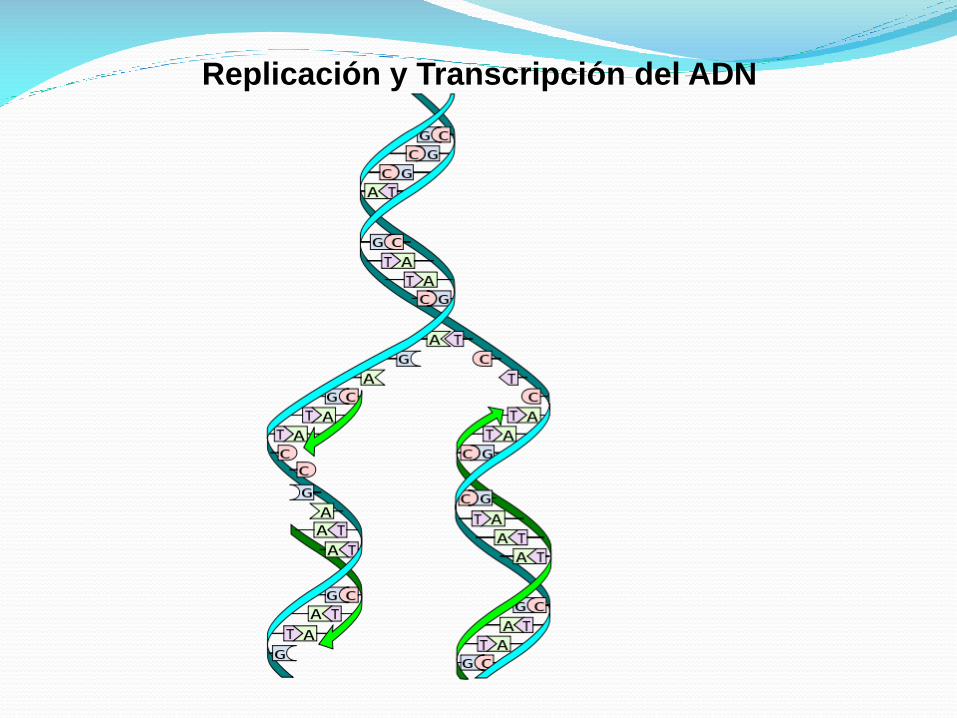
Replicación y Transcripción del ADN

Fragmentos de Okazaki son fragmentos cortos de ADN recién
sintetizadas, que se forman en la cadena molde en retraso
durante la replicación del ADN.
Ellos son complementarios a la cadena molde en retraso,
formando secciones de ADN de doble cadena cortos.
Fragmentos de Okazaki son entre 1.000 y 2.000 nucleótidos
de longitud en Escherichia coli y están entre 100 y 200
nucleótidos de longitud en eucariotas.
Están separados por cebadores de ARN 10-nucleótidos y son
no ligado hasta que se eliminan los cebadores de ARN,
seguido de enzima ligasa conecta los dos fragmentos de
Okazaki en una cadena complementaria recién sintetizada
continua.
Fragmentos de Okasaki
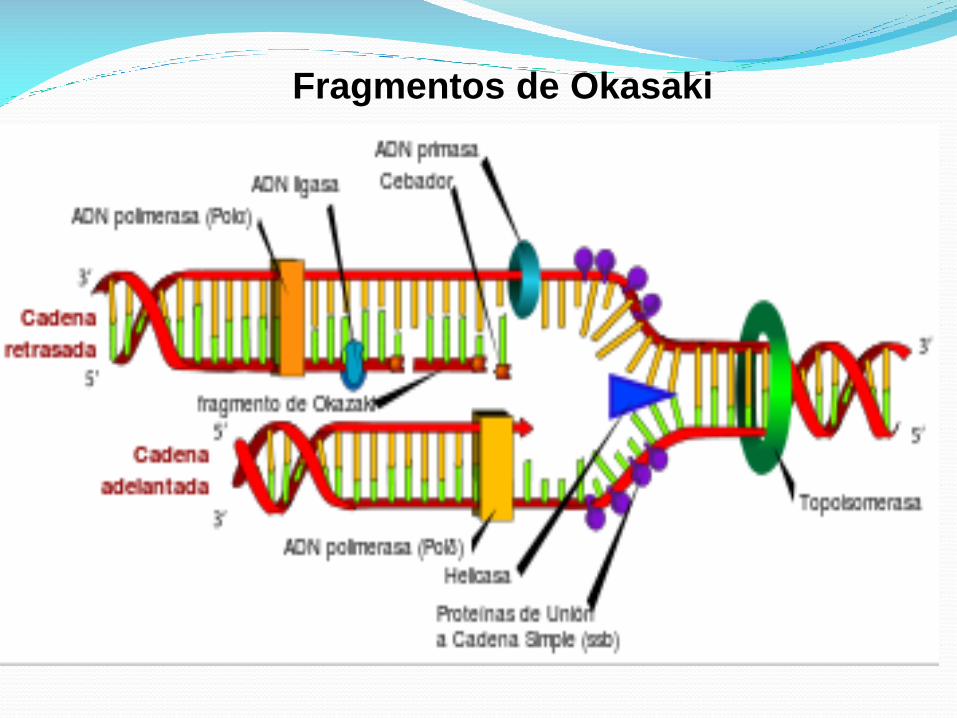
Fragmentos de Okasaki

Proceso de Transcripción del ADN
La transcripción es el proceso por el cual se sintetiza un ARN
usando como molde al ADN.
Muchos tipos de ARN pueden ser sintetizados asì por la
enzima ARN polimerasa, el ARN ribosomal el de transferencia,
los pequeños ARN nucleares o citoplasmáticos y por supuesto
los ARN mensajeros, que serán luego traducidos a una cadena
polipeptídica.
El proceso de la transcripción de los mensajeros es diferente
en procariotas y eucariotas.
Esto es debido a las diferencias propias entre los genes de las
bacterias y los de las células de animales superiores.

Los genes eucariotas son complejos y discontinuos es decir que
poseen regiones codificantes (que formarán parte de la proteína) y
otros que son no codificantes y se remueven rápidamente antes que el
ARN salga al citoplasma a ser traducido. Las regiones codificantes se
llaman EXONES y las no codificantes se llaman INTRONES.
La transcripción comienza en el punto 0 (cero) muy cerca del promotor
y termina en las bacterias en una secuencia llamada terminadora. La
polimerasa al copiar esa región de ADN, se enlentece y se desprende
del molde. En algunos casos hay una proteína que ayuda en ese
proceso denominada Rho.
Un esquema del ARN transcripto de esa región terminadora se pliega
en el espacio formando una horquilla ya que el ARN es de cadena
simple.
El proceso en el cual se eliminan los intrones y empalman los exones se denomina SPLICING

Los genes eucariotas son complejos y discontinuos es decir que poseen
regiones codificantes (que formarán parte de la proteína) y otros que no son
codificantes y se remueven rápidamente antes que el ARN salga al
citoplasma a ser traducido. Las regiones codificantes se llaman EXONES y
las no codificantes se llaman INTRONES.
La transcripción comienza en el punto 0 (cero) muy cerca del promotor y
termina en las bacterias en una secuencia llamada terminadora. La
polimerasa al copiar esa región de ADN, se enlentece y se desprende del
molde. En algunos casos hay una proteína que ayuda en ese proceso
denominada Rho.
Un esquema del ARN trasncriptor de esa región terminadora se pliega en el
espacio formando una horquilla ya que el ARN es de cadena simple.