Embed Size (px)
Citation preview

3.00 1
生物の科学・技術(B)
動物生命科学―動物繁殖のバイオテクノロジー
農学生命科学部・園芸農学科
鈴 木 裕 之
はじめに
われわれの生活には,家畜と呼ばれる動物がいろ
いろな形で関わりをもっている。たとえば,直接見
る機会は少なくなったが,食料生産用に飼育されて
いる家畜,研究用の用いられている実験動物,ある
いはもっとも身近な仲間として伴侶(愛玩)動物が
いる。近年,これらの動物の生産にはいろいろな「人
工生殖」技術が利用されている。最近,assisted
reproductive technology(生殖介助術,ART;アー
ト)とも呼ばれている。たとえば,人工授精技術に
よって優秀な遺伝資質をもった雄の子孫を効率よ
く増産することができるし,生殖細胞を凍結保存す
ることによって,貴重な動物の遺伝資源を半永久的
に保存することも可能である。また,一個の胚を人
為的に分離・分割することによって一卵性多数子を
生産することや,核移植によるクローン動物の生産
も可能となっている。この講義では,最近の動物生
産を支える「人工生殖」技術の内容と実際,さらに
は今後に期待されている関連技術を取り上げ紹介
していく。
人工生殖の基本技術は,動物の生殖細胞(卵子や
精子)の人為的操作であり,これらを駆使して自然
に行われるより極めて効率のよい,あるいは自然界
では起こり得ないような動物の生殖システムが実
現している。この講義の前半では,生殖細胞と体細
胞を比較して,生殖にまつわる生物の不思議な現象
を学び,後半では人工生殖という動物バイオテクノ
ロジーの最先端の情報を紹介する。
Ⅰ.生殖にまつわる生物の不思議
1.生命論の変遷
生命の誕生については古代ギリシァ時代から,い
ろいろ論じられている。主に二つの考え方があり,
一つは「男性の分泌物の中に,やがて形成されるは
ずの人間の体の全器官が閉じ込められており,それ
らが成長して生まれてくる」という説(前成説),
他の一つは「母胎内で,胚/胎児が徐々に形態を変
化させながら人間らしい形がつくられていく」とい
う説(後成説)である。前成説によれば,発生過程
の基本は,成体の微小な原型が成長していく変化に
すぎない。古代において科学的な生物学の基礎を作
ったとされるアリストテレスは,独自の観察結果か
ら前成説を否定する見解をもっていた。しかし,ア
リストテレスの説より前成説の方が定着し,影響力
も強いものとなった。この考え方は,その後キリス
ト教的な立場によって支えられたと思われるが,以
来中世以降までの約 2,000 年間に亘って,個体発生
に関する見解には見るべき進展はなかった。
17世紀になって顕微鏡観察がはじまり,イギリス
の物理,生物学者であるロバート・フック Robert
Hooke (1635-1703)によって初めて細胞が発見され
るとともに,動物の解剖,発生の面でも顕微鏡を用
いた研究が盛んとなった。上図はフックが使用した
顕微鏡と彼が観察したコルク片を示す。1672 年,マ
ルピーギ Marcello Malpighi (1628-1694)はニワト
リの胚発生を観察して,それまでは体の一部しか出
現していないと思われていた時期に,胚体の各部分

3.00 2
がすでに形成されている事実を報告した。また,ス
ワンメルダム Jan Swammerdam (1637-1680)は,サ
ナギの中に成体のチョウが折りたたまれて入って
いることを明らかにした(1669)。この時点では,卵
に微小成体が存在すると理解されたので,この様な
前成説は卵原説と呼ばれている。それは,精子の存
在はこれ以降に知られたからである。
ハム Johan Ham が精子の存在を見出し
(1675),レーウェンフック Antony van
Leeuwenhoek(1632-1723)が受精におけ
るその働きを推定(1679)して以来,精子
に成体の原型が存在するという精原説
が現れた。この説によれば,卵子は精子
に栄養を提供する役割をうけもつだけ
のものとされている。右図には,精子内
に膝をかかえた小人が描かれたホムン
クルス(精子微人)を示す。
18 世紀になると精子を綿密に観察しても成体の
原型が見いだされないことが確認され,精子は寄生
微生物とみなされた。そしてボネ Charles Bonnet
(1720-1793)によってアリマキの単為生殖が発見さ
れるに至り(1745),精原説の根拠は完全に失われた。
また,この頃トランブレー Abraham Trembley
(1710-1784)がヒドラの再生現象を実証した(1744)。
前成説が正しければ,失われた部分が再生するはず
はない。これに対して,ボネは従来の卵原説を修正
し,卵には成体そのものではなく,そのもとになる
構造体があり,それが体の各部に分散し,発育状況
に応じて栄養物質を吸着して成体になると考えた。
しかし,再生の発見に続き奇形発生の研究,動物の
発生の比較研究などから,前成説はしだいに不利に
なっていった。科学的な後成説は 19 世紀に,各種
動物の比較研究にもとづきドイツのベーア Karl
Ernst von Baer (1792-1876)によって確立された。
彼は,すべての動物は発生初期には同一の胚葉構造
をもち,発生が進行するとともに各動物の個性が現
れてくると説いた。
19 世紀後半において最も支持を受けたアウグス
ト・ワイスマン August Weismann の生殖質説(1892)
にも前成説の要素がみられるが,同世紀末にはじま
る実験発生学,とくに,ドイツの発生学者ルー
Wilhelm Roux (1850-1924)とハンス・ドリーシュ
Hans Driesch (1867-1941)の論争は注目される。ド
リーシュは,本来幼生の一部を構成するはずの割球
を他の部分から分離すると,それだけで完全な幼生
に発生することを示し(1892),後成説の主張に有利
な根拠をあたえた。1902 年,ハンス・シュペーマン
Hans Spemannがついにサンショウウオの胚を二つに
分離させることに成功して,ドリューシュの説は完
全に正当化された。
顕微鏡という革命的な機器が現れたものの,17世
紀から 18 世紀にかけての科学者は前成説から抜け
きれなかった。宗教的な背景から語り継がれた概念
が,いかに大きな壁であったかを物語っている。
【1回目はここまで】
2.生殖
生殖とは,生物が自分と同じもしくは共通する遺
伝子組成をもつ個体を新たにつくることをいう。生
殖には無性生殖と有性生殖の二つの様式がある。有
性生殖は,無性生殖とは対照的に,遺伝子の新しい
組合せを生ずるので,子孫に新しい形質が出現する
可能性が高い。したがって,生物は有性生殖を介し
て環境への適応の多様性を増すことができる。次ペ
ージに無性生殖と有性生殖を比較した表を示した。
3.生殖細胞と体細胞
受精卵は一定のパターンで細胞分裂を繰り返し,
多種類の細胞を生み出していくが,特定の器官を構
成する細胞がどの細胞の子孫であるのかを表す細
胞の家系図を,細胞系譜 cell lineageとよんでいる。
多細胞生物には,大きく分けて2つの細胞系譜が存
在する。ひとつは,卵子や精子である生殖細胞に分
化する生殖系列 germ line,もうひとつは生物個体
を構成する体細胞系列 somatic lineである。体細胞
はいずれ個体の死に伴って死ぬ運命にあるが,生殖
細胞(卵子や精子)は親から子へ伝えられ生き続け
る細胞である。

3.00 3
ふえ方 特徴
無性生殖
体細胞分裂などにより,新しい個体をつくる
分裂の例 出芽の例 栄養生殖の例
雄と雌(性別)によらない
もとの個体と同じ遺伝子をもつため,親とまったく同じ形質を示す
同一な子ができる↓
環境が変化すると全滅する可能性がある
有性生殖
雄と雌による受精によって新しい個体をつくる
動物…精子と卵子の核が合体して受精植物…精細胞と卵細胞の核が合体して受精受精卵が育ち,新しい個体ができる
母親と父親の遺伝子を受けつぐため,種としての形質以外は両親と異なる形質を示す
多様な子ができる↓
環境が変化しても生き残る可能性がある
3-1.卵の成熟と受精の重要性
両親から受け継いだそれぞれの遺伝子を一度分
離して受精による新しい組み合わせを可能にし,ま
た染色体間組換えを起こして同一染色体上の遺伝
子の組み合わせをも変化させて,次代における多様
な遺伝子の組み合わせをつくる。
減数分裂は配偶子形成において遺伝的な多様性
を生じさせ,環境変化への対応や進化に貢献してき
たと考えられている。例えば 2 組 4 本の染色体を持
つ生物では,22=4 通りの組み合わせをもった配偶
子が作られ,ここから得られる次世代は 42=16 通
りである。ヒトの場合では 23 組の相同染色体,計
46 本の染色体を持つため,223=8,388,608 通りの配
偶子,8,388,608 2=70,368,744,177,664 通りの次世
代が生じる可能性をもっている。さらに遺伝の多様
性を生み出す仕組みには,染色体の一部が入れ替わ
る乗換え(交叉)がある。乗り換えは減数第一分裂に
行われる。分子機構としては DNA の二重鎖切断が
起きて,DNA 修復によってつなぎ直される際に起
こる。
3-2.細胞の寿命
すべての動物は,それぞれの種に特有の(最大限
の長さの)寿命をもっている。 スタンフォード大
学のヘイフリックHayflickが明らかにしたように,
その寿命の長さは,その種における胚の細胞の受精
後からの分裂回数の限度に比例している。分裂可能
な体細胞を何度も植え継いで培養を続けると,細胞
には寿命があること,また寿命の長い個体から取っ
た細胞は寿命が短い個体から取った細胞よりもそ
の寿命が長いことが明らかになっている。正常の2
倍体細胞は有限寿命であり,その例外としていつま
でも生き続ける細胞がガン細胞であり,次世代へ受
け継がれる生殖細胞である。
では,正常な2倍体細胞にはどうして寿命がある
のだろうか。そのカギを握るのはテロメアという,
染色体末端に存在する特殊な構造である。テロメア
という言葉は,ギリシャ語で「末端」を意味する telos
と,「部分」を意味する meros を組み合わせて名付
けられた。テロメアはヒトでは TTAGGG という塩
基配列の繰り返しになっている。このテロメアが,
DNA の複製の際に短くなる運命にあることが分か
っている。

3.00 4
上のグラフは,ヒトの繊維芽細胞のテロメア長と
加齢との関係を示す。一回の細胞分裂で,テロメア
は 100~150 塩基対短くなる。ヒト細胞のテロメア
は約 15,000 塩基対からなるので,ヒトの細胞は 100
~150 回分裂することができる計算になるが,実際
には 60 回程度しか分裂できない。その理由は,テ
ロメアの長さが 5,000 塩基対に近づくと,細胞分裂
が止まるからである。
では,生殖細胞はどうして限りなく増殖できるの
だろうか。それはテロメアの合成に必要なテロメラ
ーゼという酵素の活性が高く,テロメアの短縮化が
起こらないためである。大部分のがん細胞(85%)
も高いテロメラーゼ活性を持っている(残り 15%の
がん細胞にはテロメラーゼ活性はないが別な方法
でテロメアの長さを維持している)。これに対し,
正常な体細胞でテロメラーゼはほとんど検出され
ない。
このように,細胞の寿命に限りがあるという現象
は,テロメアの短縮という現象で説明できる。しか
し,卵母細胞や精母細胞のような生殖細胞でも,体
内の生殖巣に留まっていれば,間もなく死に至る。
これらの細胞は受精して次の世代の胚・個体となっ
て,細胞分裂を続けることができるのである。生殖
細胞になるときに細胞寿命がリセットされるしく
みが備わっている。
下図には生殖細胞の連続性を模式的に示した。
【2回目はここまで】
4.哺乳動物の生殖機構
哺乳動物という高度に進化した生物には,複雑に
して絶妙なる生殖のメカニズムが備わっている。そ
の中には,配偶子形成,発情周期,受精,胚発生,
着床,胎盤形成,分娩,哺育というさまざまな過程
が含まれている。たとえば,動物の性行動の巧みさ
の一つとして,季節繁殖という仕組みがある。これ
は,ある種の動物が一年のうちの決まった季節にの
み繁殖を行う現象のことで,代表的な例としてウマ,
ヒツジやヤギが挙げられる。ウマは春に繁殖(交尾)
期を迎え,翌年の春に子ウマを出産する。また,ヒ
ツジやヤギは秋に交尾をして,やはり翌春に子を産
む。つまり,草食動物であるウマやヤギ・ヒツジが,
若草の萌える春に合わせて出産時期を迎えること
で,新生子の哺育を有利にしているのである。ウマ
の妊娠期間は約 10 ヵ月,ヒツジ・ヤギでは約 5 ヵ
月なので,春に出産時期を迎えるためには,逆算し
てウマは前年の春から初夏に,またヒツジとヤギは
秋から冬にかけて交尾をすればよいことになる。こ
れがかれらの繁殖期となっている。これに対して,
低緯度の熱帯地方のような一年を通じて餌に恵ま
れた地域に住む動物では,季節に関係なく繁殖する
ものが多く,これらは周年繁殖動物と呼ばれている。
本講義では,これらの繁殖周期のうち配偶子形成
から受精までを中心に紹介する。
4-1.精子形成
精子の構造は次図に示すように頭部と尾部の二
つの部分からなっている。頭部には,遺伝情報を含
む細胞核と受精時に働く特異的な酵素が含まれて
いる。尾部は雌の生殖器内を遊泳するための運動装
置である。尾部の付け根のやや太い部位は中片部と
呼ばれ,ここには尾部を動かすためのモータの役割
をするミトコンドリアが存在する。それぞれの動物
種の精子は,頭部の形状や尾部の長さなどに特徴的
な形態を示す。ハムスターやラットの精子などは頭
部が鎌型状を示し,また体が小さいハムスターの精
子の全長が,ウシなどの家畜の精子の約 3 倍もある
というのも興味深い(次ページ各動物の精子の模式
図参照)。

3.00 5
4-2.卵子形成
卵子形成は,動物の種々の繁殖様式に対応して,精
子形成よりはるかに多様である。水中で体外受精を行う
動物では,1回の産卵数は多く,その数は数百から数十
万個である。体内受精を行うものでは,各生殖期に 1
個から十数個の卵子が成熟する。排卵数だけでなく,
卵子の大きさも,母胎外で発生するものと母胎内で発
生するものとでは非常に異なっている。胎生である哺乳
動物の卵子は,卵黄などの栄養物質を多量に蓄積して
おく必要はないので,比較的小さい。それに対して,卵
生である動物の卵子は卵黄を多量に含み,大きいのが
普通である。同じ卵生でも,一般的に水生動物の卵子
は陸生爬虫類や鳥類に比べずっと小さい。動物による
発生環境の違いは,卵子を被う卵膜の多様化にもつな
がっている。卵子形成のもう 1 つの重要な問題は,受精
後の発生に備えた準備である。
4-3.精子形成と卵子形成の相違
精子形成と卵子形成の過程は,遺伝学的には同じ
ように進行するが,形態学的には著しく異なってい
る。減数分裂の結果,精祖細胞が 4 個の精子になる
のに対して,卵祖細胞からは 1 個の卵子しかできな
い。卵母細胞 oocyte は,第 1 減数分裂の結果,細
胞質の大部分を含む大きな細胞とごくわずかな細
胞質しか含まない第 1 極体 first polar body という
細胞になる。大きい卵母細胞は第 2 成熟分裂期にも
著しい不等分裂を行い,卵子と第 2 極体 second
polar body になる(次ページヒト精子形成と卵子形
成の模式図参照)。
女性では,減数分裂の開始は胚子の時期にさかの
ぼるが,分裂は第 1 分裂前期で停止し,その完了は,
最も早い卵母細胞でも思春期になってから,逆に最
も遅いものでは 40-50 年経っても完了しない。一方,
精子形成は思春期になって始まり,生涯継続する。
精子形成過程には,卵子形成時のような長い分裂停
止期は起こらないので,減数分裂の各時期に留まる
時間の割合は,卵子形成の場合とは大きく異なり,
全過程が 2 ヵ月程で完了する。
減数分裂の一時的停止は卵子形成の特徴であ
る。哺乳動物では分裂停止期が 2 回あり,最初の分
裂停止は第 1 減数分裂前期の複糸期に起こる。
この分裂停止は,卵黄の蓄積や受精後に必ず始ま
る合成反応との準備期間として必要であると考え
られている。この分裂停止はホルモンの作用で解除
され,分裂が再開されるが,すぐに2度目の停止が
起こる(脊椎動物では多くの場合,第2減数分裂中
期で停止する)。この2回目の分裂停止は,受精や
卵子の人為的活性化により解除される。
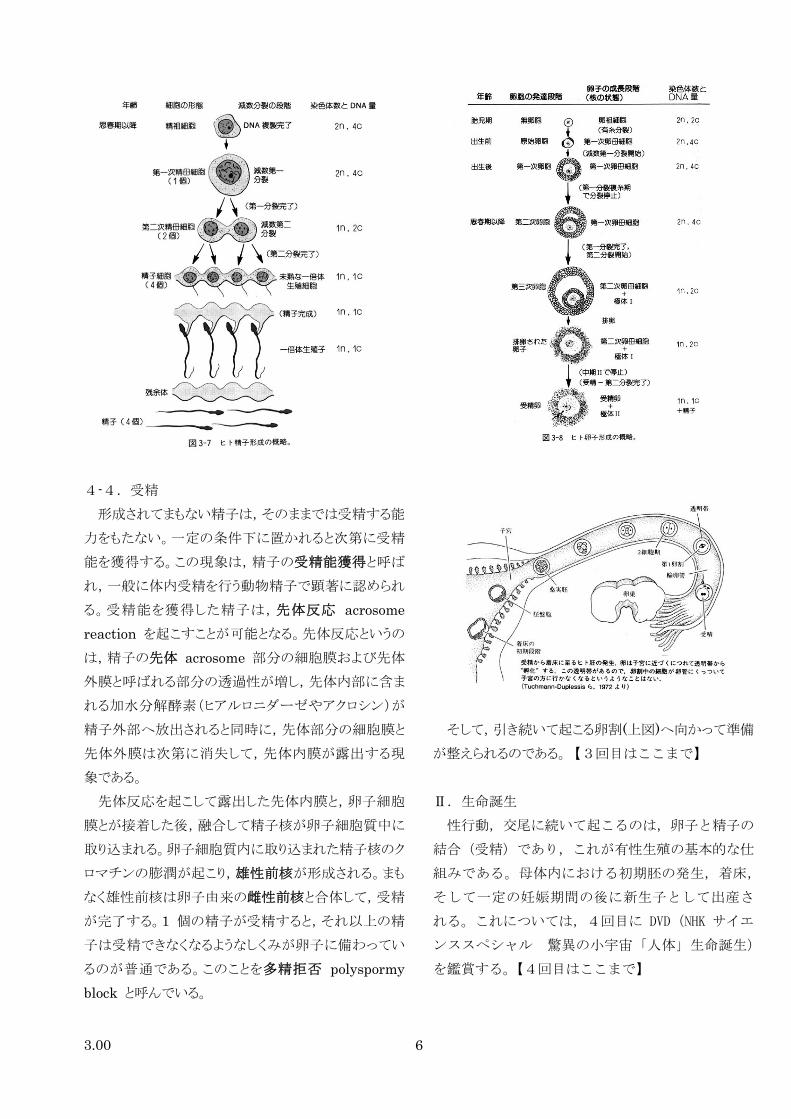
3.00 6
4-4.受精
形成されてまもない精子は,そのままでは受精する能
力をもたない。一定の条件下に置かれると次第に受精
能を獲得する。この現象は,精子の受精能獲得と呼ば
れ,一般に体内受精を行う動物精子で顕著に認められ
る。受精能を獲得した精子は,先体反応 acrosome
reaction を起こすことが可能となる。先体反応というの
は,精子の先体 acrosome 部分の細胞膜および先体
外膜と呼ばれる部分の透過性が増し,先体内部に含ま
れる加水分解酵素(ヒアルロニダーゼやアクロシン)が
精子外部へ放出されると同時に,先体部分の細胞膜と
先体外膜は次第に消失して,先体内膜が露出する現
象である。
先体反応を起こして露出した先体内膜と,卵子細胞
膜とが接着した後,融合して精子核が卵子細胞質中に
取り込まれる。卵子細胞質内に取り込まれた精子核のク
ロマチンの膨潤が起こり,雄性前核が形成される。まも
なく雄性前核は卵子由来の雌性前核と合体して,受精
が完了する。1 個の精子が受精すると,それ以上の精
子は受精できなくなるようなしくみが卵子に備わってい
るのが普通である。このことを多精拒否 polyspormy
block と呼んでいる。
そして,引き続いて起こる卵割(上図)へ向かって準備
が整えられるのである。【3回目はここまで】
Ⅱ.生命誕生
性行動,交尾に続いて起こるのは,卵子と精子の
結合(受精)であり,これが有性生殖の基本的な仕
組みである。母体内における初期胚の発生,着床,
そして一定の妊娠期間の後に新生子として出産さ
れる。これについては,4回目に DVD(NHK サイエ
ンススペシャル 驚異の小宇宙「人体」生命誕生)
を鑑賞する。【4回目はここまで】

3.00 7
Ⅲ.人工生殖技術
哺乳動物の生殖過程の各段階は,極めて精密に調
節されて成り立っている。その生殖過程のある段階
を人工の環境や器具・器械などで置き換え,あるい
は生殖の仕組みを人間の目的に合わせて修飾する
などの技術を「人工生殖技術」という。具体的な例
としては,まず動物の交尾を人工的手技に置き換え
る技術として開発された「人工授精」を挙げること
ができる。また,本来は雌の卵管内で起こる受精を
体外で行う「体外受精」もその一つである。受精に
引き続いて卵管・子宮内で進行する胚の発生を,体
外の人工環境で行う技術も開発されており,「胚の
体外培養」という。さらに,胚を雌の体内から取り
出し,他の借り腹雌(代理母)の子宮内へ移植して
受胎させる技術を「胚移植」と呼んでいる。その他,
最先端の人工生殖技術には,「一卵性多数子生産」,
「キメラ動物」,「核移植」,「雌雄判別」,「遺伝子組
換え動物」の作出など,近年発展が目覚しい発生工
学研究を基礎として開発されたものが多い。
1.人工授精
哺乳動物の生殖の第一段階は,雄と雌との交尾で
あり,この交尾に替わる人工生殖技術が人工授精で
ある。人工授精には,通常,種雄畜から精液が採取
されて用いられる。採取された精液は,まず特殊な
液で希釈される。それにはブドウ糖,クエン酸ナト
リウム,卵黄などを含む液が使用されている。
精液を希釈する意味は,次のようにまとめられる。
1)精子の消耗を防ぎ受精能力を良好に保つこと
2)精液の量を増やすこと
たとえば,ウシの精液量は 3~10 ml/1 回の射精,
その中に 30 億~140 億の精子を含む。これを希釈す
ることによって数百回分もの人工授精用精液を作
ることができる。このことは,優秀な雄ウシの精子
を繰り返し利用して家畜の改良を進めるという畜
産の目的に重要な意味をもつ。
人工授精技術の普及のためには,精液凍結技術の
確立が不可欠であった。畜産における人工授精の利
点の一つは,ある優秀な種雄畜の精子を用いて,そ
の雄を移動しなくても遠隔地の雌畜を受胎させる
ことができる点にある。しかし,射出された精子が
常温で受精能力を保ちうる期間はせいぜい数日間
であるため,精子の保存技術なしには人工授精技術
も十分に効果を発揮することはできないのである。
家畜改良への貢献という点では,凍結精液を用いた
人工授精が極めて大きな役割を果たしている。現在
では,ウシをはじめ種々の家畜の精液の凍結保存が
可能であり,必要に応じて融解され,利用されてい
る。保存温度をマイナス 196℃といった超低温に設
定すれば,半永久的保存も可能である。
ウシを例として,精液の注入操作を下図に示す。
ウシの場合,人工授精用の精液は容量 0.5 ml のプ
ラスチックストローに詰めて用いられる。このスト
ローが装着できるようになっている金属製あるい
はプラスチック製の注入器(AIガンという)によっ
て,内部の精液は‘水鉄砲の原理’で雌の子宮に注
入される。
自然の交尾に比べ,人工授精では子宮内部に注入
される。これによって,通常の精子数に比べてはる
かに少ない量で受胎が成立する。また,受胎成立の
ためには,雌の性周期を把握して,排卵との時間を
考慮して人工授精を行うことが重要である。
人工授精技術の確立によって,家畜の生産におい
ては,能力の確かな雄だけが繁殖に供用されるよう
になり,家畜改良は飛躍的な進歩を遂げることにな
った。また,自然交配による生殖器の接触により感
染する伝染病の蔓延を防止されるようになったの
も人工授精の功績の一つである。パンダなどの野生
動物を対象とする人工授精も,原理的には家畜の技
術と同様である。
人工授精技術の利点を挙げると,次のようにまと
3.00 8
められる。
1)優秀な種雄畜の利用効率の増大(遺伝的改良,致
死遺伝子等の望ましくない遺伝子の排除など)
2)遺伝能力の早期判定
3)生殖器伝染病の蔓延防止
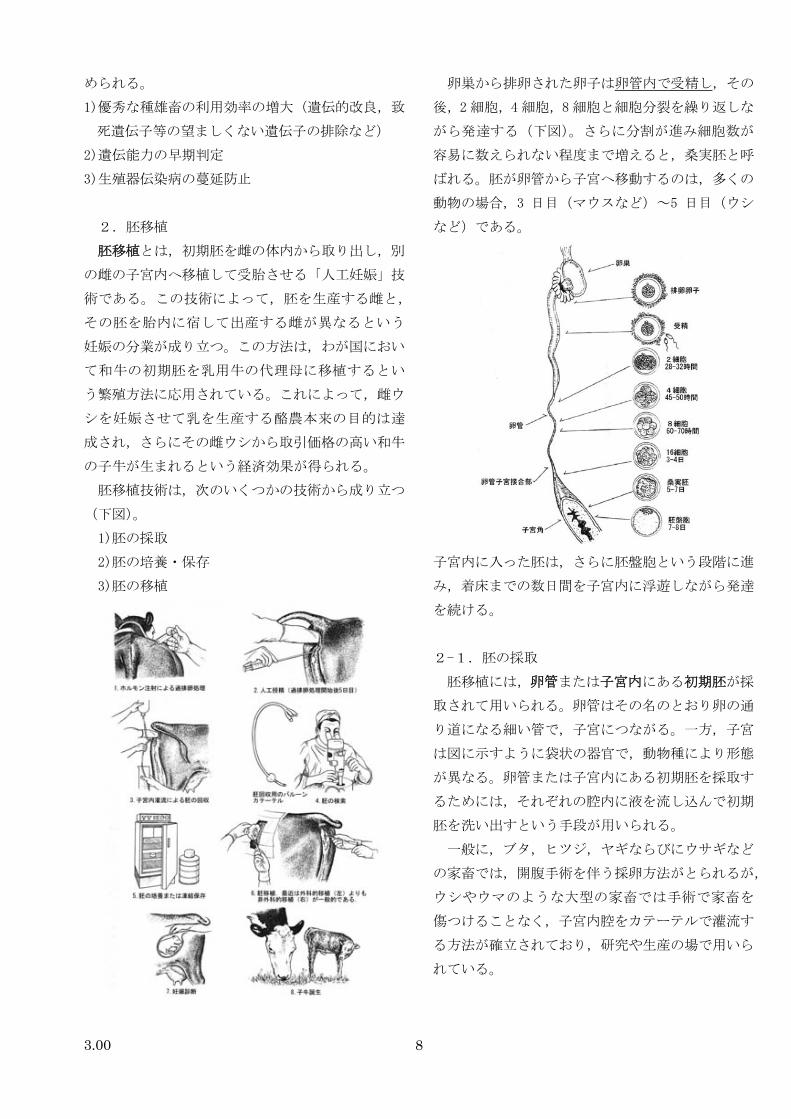
2.胚移植
胚移植とは,初期胚を雌の体内から取り出し,別
の雌の子宮内へ移植して受胎させる「人工妊娠」技
術である。この技術によって,胚を生産する雌と,
その胚を胎内に宿して出産する雌が異なるという
妊娠の分業が成り立つ。この方法は,わが国におい
て和牛の初期胚を乳用牛の代理母に移植するとい
う繁殖方法に応用されている。これによって,雌ウ
シを妊娠させて乳を生産する酪農本来の目的は達
成され,さらにその雌ウシから取引価格の高い和牛
の子牛が生まれるという経済効果が得られる。
胚移植技術は,次のいくつかの技術から成り立つ
(下図)。
1)胚の採取
2)胚の培養・保存
3)胚の移植
卵巣から排卵された卵子は卵管内で受精し,その
後,2細胞,4 細胞,8細胞と細胞分裂を繰り返しな
がら発達する(下図)。さらに分割が進み細胞数が
容易に数えられない程度まで増えると,桑実胚と呼
ばれる。胚が卵管から子宮へ移動するのは,多くの
動物の場合,3 日目(マウスなど)~5 日目(ウシ
など)である。
子宮内に入った胚は,さらに胚盤胞という段階に進
み,着床までの数日間を子宮内に浮遊しながら発達
を続ける。
2-1.胚の採取
胚移植には,卵管または子宮内にある初期胚が採
取されて用いられる。卵管はその名のとおり卵の通
り道になる細い管で,子宮につながる。一方,子宮
は図に示すように袋状の器官で,動物種により形態
が異なる。卵管または子宮内にある初期胚を採取す
るためには,それぞれの腔内に液を流し込んで初期
胚を洗い出すという手段が用いられる。
一般に,ブタ,ヒツジ,ヤギならびにウサギなど
の家畜では,開腹手術を伴う採卵方法がとられるが,
ウシやウマのような大型の家畜では手術で家畜を
傷つけることなく,子宮内腔をカテーテルで灌流す
る方法が確立されており,研究や生産の場で用いら
れている。

3.00 9
2-2.胚の体外培養
体外に取り出された初期胚は,移植までの間「培
養液」という人工の環境下に置かれる。再び子宮に
戻されるまでの間,胚の発生能が損なわれないよう
に保つことが目的である。
培養液には種々の塩類,エネルギー源,タンパク
質源などが含まれ,温度,空気中の炭酸ガス,酸素,
または窒素濃度などがコントロールできる特殊な
培養器内で培養される。
現在,種々の動物の胚を培養によって発育させる
ことが可能であるが,一般的に長時間培養された胚
を移植した場合の受胎率は新鮮胚の場合に比べ低
い。これは,現在の培養液および培養環境がまだ完
全でないことを示すものである。とくに,ゴールデ
ンハムスター胚の体外培養は困難で,培養胚を移植
したあとの産子生産率はきわめて低い。
初期胚の培養技術が部分的にも可能となり,本来
雌動物の胎内で進行する胚の初期発生過程を体外
で観察できるようになった。さらに,体外の人工環
境下で胚を長時間培養できるようになれば,その間
に胚に対してさまざまな処理,操作を加えることも
可能となる。
2-3.卵子,初期胚の凍結保存
精子の場合と同様に,卵の凍結においても細胞内
氷晶形成および溶液中塩類の濃縮による塩害,細胞
収縮などを防止するためには凍害保護物質が用い
られる。さらに,卵の凍結では緩慢な速度で冷却す
る必要がある(胚の凍結の模式図参照)。
一般に,桑実胚や拡張胚盤胞期の胚は凍結後の生
存性が比較的高く,発達初期の胚ほど耐凍性が低い。
とくに,未受精卵や 1細胞期胚の凍結生存性は非常
に低い。この理由として,1~4細胞期胚のような分
割初期の割球は比較的後期の胚の割球に比べては
るかに巨大なため,凍害保護物質の浸透や凍結過程
における脱水がスムーズに起こりにくいことが考
えられている。
2-4.胚の移植
体外に取り出された初期胚を再び代理母に移植
する方法にも,開腹手術を行うものと行わないもの
と二通りある。そのどちらかを用いるかは,胚移植
の目的や動物種によって決められる。中小動物では,
開腹手術を行い細いピペットで胚を少量の培養液
とともに子宮内に注入する。これに対し,大型家畜
では開腹手術をせず,合成樹脂や金属製の注入器を
子宮内に挿入して胚を注入する方法がとられる。
胚移植技術の利点を挙げると,次のようにまとめ
られる。
1)優良雌牛から多くの後代が得られる。
2)卵巣吸引法と体外受精法(IVF)により,多排卵反
応が良くない牛 ( 妊娠中であっても ) からも後
代が得られる。
3)どこでも新たな品種の胚を移植で,他品種の産子
を得ることができる。
4)また,この技術の利用により,今後いろいろな胚
の体外操作や遺伝子工学的手法の導入が可能とな
る。【5回目はここまで】
3.体外受精
ウシの体外受精技術は,家畜の生体または屠体の
卵巣から卵子を採取し,これを人工的環境下で成熟,
受精させ,移植可能なステージまで発育させる技術
である(次ページ図1)。最近では,大量かつ低コ
ストの受精卵の生産技術として,肉用牛の増産に用
いられるだけでなく,超音波ガイド・経膣生体卵子

3.00 10
吸引技術との併用による育種改良への応用や,クロ
ーン研究などの試験研究分野における材料の供給
源として期待されている。
日本国内では 100以上の機関でウシ体外受精技術
が実施されており,実用的な移植頭数も 5,700~
7,700 頭(H11~H14 年)に達し,民間ベースでの体
外受精卵の販売も行われている。下の写真は体外受
精直後のウシ前核期卵を示す。
ウシ体外受精卵(PN:雌雄前核)
4.性判別(雌雄生み分け)
家畜の生産の場においては,人間の目的に合った
一方の性の子畜だけを得たい場合がある。たとえば,
乳用牛では実際に乳を生産する雌が望まれるし,逆
に肉用家畜では成長速度や産肉量の点で雄の方が
都合がよい。また,実験動物や愛玩動物でも,雌雄
のどちらかがとくに必要とされることがある。この
ように,経済上,研究上あるいはそのほかの理由で
動物の雌雄の生み分けが要求されている。
4-1.哺乳動物の性決定のしくみ
哺乳動物では,性染色体の組み合わせによって性
が決定される。すなわち,哺乳動物のすべての卵子
は X 染色体を 1本ずつもち,他方精子は X染色体を
もつものと,Y 染色体をもつものが半数ずつ形成さ
れる。生殖の過程で,X 染色体をもつ精子(X-精子)
が受精すれば,受精卵は XX となり雌が生じ,Y 染色
体をもつ精子(Y-精子)が受精すれば,XYとなり雄
となる。このように,哺乳動物の遺伝的性は受精の
時点で決定され,受精卵は雄と雌にそれぞれ特異的
な性染色体の組み合わせをもつことになる。そこで,
この組み合わせの違いを指標として,初期胚の雌雄
を判別する試みがなされている。
4-2.精子の分別による雌雄生み分け
哺乳動物の性決定が X-精子と Y-精子のどちらが
受精するかによって支配されているので,X-精子ま
たは Y-精子のどちらかで選択的に受精させる方法
も動物の雌雄生み分けにつながる。現在,可能性の
ある方法として注目されているのは,X 染色体と Y
染色体の大きさの差に起因するDNA量の差を元にし
た分別方法である。DNA に蛍光色素を結合させ,レ
ーザー光線を照射し,それによって各精子が発する
蛍光の強度をコンピュータで解析して X-精子と Y-
精子を識別する方法である。ただし,この方法の問
題点は,分離された精子の受精能力が低下すること
である。
4-3.初期胚の性判別による雌雄生み分け
初期胚の段階で雌雄の判別する方法の一つは,切
断二分した胚の一方を性判別の材料に用いる方法
である。たとえば,ウシ桑実胚を切断二分して,一
方の性染色体を調べて性別を知り,その後もう一方
の胚を移植して希望通りの性の子牛を生産するこ
とが可能である。この他に,1 個の胚から性判別の
ための細胞を得る方法として,胚盤胞の栄養膜細胞
層を切り取る方法や,桑実胚から少数の割球を吸い
取る方法などが試みられている。
1)細胞学的方法
動物の染色体は,それぞれの種によって数と形が
決まっている。たとえば,ウシでは全部で 60 本の

3.00 11
染色体中に 2 本の性染色体が含まれる。X 染色体と
Y 染色体が 1 本ずつあれば雄,X 染色体が 2 本あれ
ば雌と判定する。他の動物の染色体数は,ウマ 64
本,ブタ 38 本,イヌ 78 本,ヤギ 60 本,ヒツジ 54
本,ネコ 38 本,マウス 40本,ウサギ 44本である。
理論的には,あらゆる動物において初期胚の染色
体を調べることで,雌雄の判別は可能だが,実際に
は動物種によって染色体標本の作製や染色体像の
分析が困難な場合が多い。
2)DNA 配列による方法
Y 染色体上に存在する DNA 配列を指標として判定
する。初期胚の細胞から抽出した DNA 中にその配列
が見つかれば雄,それがなければ雌とするという方
法である。
5.キメラ動物の作出
キメラとは,ギリシャ神話に登場する怪物に由来
する用語である。神話のキメラは,ライオンの頭,
ヤギの胴体,そしてヘビの尾をもった,いわば合体
動物である。
英国家畜生理学研究所のグループによって作ら
れたヒツジとヤギのキメラは,ヤギの特徴を示す顔
と角をもち,体毛はカールした羊毛の部分と比較的
短くまっすぐなヤギの毛の部分とが,それぞれ斑を
なして入り交じっていた。また,尾はヒツジの形で
あった。このキメラ個体は,ヒツジとヤギの「交雑
種」ではない。体を構成する細胞がヒツジの部分と
ヤギの部分とのモザイク状になっていたのである。
キメラの作出方法
キメラ動物は,二個以上の初期胚を混合して複合
胚を作り,移植することで作出される。この複合胚
はやがて胚盤胞を形成し,それを代理母に移植する
と,材料とした両方の胚の特徴をもったキメラ個体
が誕生する。たとえば,白マウスに黒マウスの胚を
ミックスしてキメラマウスを作れば,それは灰色に
なるのではなく,白黒まだら模様の個体になる。
また,胚盤胞の内細胞塊を他の胚盤胞の胞胚腔中
に注入することによって,キメラ胚を作る方法もあ
る。この方法は,胎子の発生の源である胚盤胞の内
細胞塊を,複数の胚に由来する混成細胞集団にする
ことを意図している。
妊娠の謎とキメラ
ヤギ(染色体数 60 本)とヒツジ(同 54 本)とは,
人工授精によって両者を掛け合わせても雑種を形
成しない。また,ヤギ胚をヒツジの雌に,あるいは

3.00 12
その逆にヒツジ胚をヤギ雌に移植しても,妊娠を継
続させることはできない。受胎は成立しても,着床
期またはそれ以降の段階で妊娠は中断してしまう。
このような異種動物間の交配(または胚移植)に
おける妊娠中断の原因は,雑種胎子あるいは異種胎
子が異種抗原をもった「異物」として,母胎の免疫
反応によって排除されるためと考えられる。すなわ
ち,異種動物間には交雑妊娠の成立を妨げる障壁が
存在するのである。この障壁の生物学的機構の解明
と障壁の突破を目的として行われたのが,先に紹介
したヒツジとヤギのキメラ作出の実験である。
この実験では,ヤギとヒツジの 8細胞期胚を材料
として,1 個のヒツジ胚の周囲に 3 個のヤギ胚を配
して,複合胚が作られた。この胚を発達させたとこ
ろ,胚の外側の栄養膜細胞はヤギ胚由来の細胞で構
成され,内細胞塊はヤギ細胞とヒツジ細胞の複合状
態(キメラ状態)になった胚盤胞が得られた。この
胚盤胞を雌ヤギに移植すると,栄養膜細胞がヤギ胚
由来であるために,キメラ個体のヒツジ由来組織が
もつ異種抗原性が母体に認識されることなく,キメ
ラ胚はあたかもヤギ胚であるかのようにヤギ雌の
子宮内で発育を遂げた。胚盤胞の栄養膜細胞層は,
そもそも母体にとっては異物である胎子(半分は父
親由来であるため異物と認識されるはずである)の
抗原性を覆い隠す役目を果たすとされており,その
機能のおかげで,異種であるヒツジの組織をもった
キメラ胎子がヤギの母体から排除されることなく
発達したと考えられる。
以上のように,ヤギとヒツジのキメラ作出実験は,
免疫学的障壁というヤギ-ヒツジ間の交雑や胚移
植を阻んでいた要因を,キメラ胚の構築でクリアす
る方策を示したものである。
ヒツジがヤギを生む
異種動物間の免疫学的障壁がクリアされれば,ヒ
ツジの母親にキメラではない純粋なヤギの子(ある
いはその逆)を出産させることが可能となる。内細
胞塊がヤギ胚で占められ,栄養膜細胞層をヒツジ細
胞で構成される状態を作れば,ヒツジがヤギを生む
ことが可能である。
このような異種動物間の胚移植は,妊娠期間や着
床様式に類似性の高い動物間には限定されるが,今
後,保護の必要な野生動物を近縁種の家畜を用いて
生産する方法としても応用されるだろう。
【6回目はここまで】
6.クローン動物の作出
クローン clone とは,同一の遺伝資質をもった
個体群のことで,現在では細胞や遺伝子についても
一群のコピーをいう場合に用いられる。元来はギリ
シャ語 klon で〈小枝〉という語から派生している。
1 本の木から出る小枝は,なん本あっても遺伝的に
同じ資質をもっていることから,ウェッバー H. J.
Webber が上のような生物学的意味で初めてこの語
を使った(1903)。
コピーのようにそっくりな個体といえば,まず思
い浮かぶのは一卵性双生子である。一卵性双生子と
はそもそも一個の受精卵に由来する二個体である
から,自然にできたクローンということができる。
6-1.胚の分離による双子作出
一卵性双子を人為的に作出する方法が開発され
ている。それは,一度体外に取り出した初期胚を毛
細ガラス針または微小メスなどで二つに分離し,一
対の「双子胚(1/2 胚)」を作る方法である。この二
分離した胚のペアを代理母に移植することで,これ
までウシ,ウマ,ブタ,ヤギ,ヒツジ,ウサギ,ラ
ット,マウスなどの一卵性双子が作り出されている。
下図に,毛細ガラス針を用いて初期胚を二分離する
顕微操作を模式的に示した。
2 細胞期胚から分離された二個の割球は,それぞ
れが一個体に発達する能力をもつ。この能力を分化
全能性という。それぞれを借り腹雌に移植すること
によって一卵性双子が生まれる。同様に,4 細胞期
胚の割球を 2 個ずつに,あるいは 8細胞期胚を 4個
ずつ分けた「双子胚(1/2 胚)」のペアからも一卵性
双生子が生み出すことができる。
桑実胚を用いる場合には,胚を二つに切断(分割)
する方法がとられる。この場合,切断面の割球は破
壊されるが,その程度が小さければ二分された「双

3.00 13
子胚(1/2 胚)」は十分発達できる。胚盤胞を用いる
場合には,胚を構成する二つの部分(内細胞塊と栄
養膜細胞層)を均等に二分しなければならない。胚
盤胞では,細胞の分化がすでに起きており,内細胞
塊は将来胎子に,また栄養膜細胞層は胎盤・胎膜に
発達するよう運命づけられている。したがって,そ
れぞれの要素をほぼ均等に二分することが必要と
なる。この点は,4 細胞期~桑実胚までの胚では,
切断の方向が全く任意であったのとは異なってい
る。
6-2.一卵性四つ子・・・その限界
胚を二分離して一卵性双生子を作り出すことが
できるのは,初期胚の細胞(割球)に分化全能性が
備わっているからである。
さて,哺乳動物の初期胚を複数の断片に分ける方
法で,一卵性多数子の作出が可能なのであろうか。
胚の四等分によって一卵性四つ子までの生産の
成功例は報告されているが,さらに胚を八等分して
八つ子の作出となると,その成功例がない。ただし,
他の胚から得た細胞(割球)を付与するなどして,
発達を支援した場合には発生する例が報告されて
いる。
胚細胞は受精直後の1細胞期から,2 細胞期,4
細胞期,8 細胞期へと細胞分裂を繰り返すに従い,
娘細胞は 1/2,1/4,1/8 の大きさになってゆく。こ
れは体細胞が細胞分裂を繰り返して自己増殖を続
ける体細胞分裂とは大きく異なる点である。体細胞
分裂では,分裂によって得られた娘細胞は母細胞の
レベルまで成長した後に,分裂が繰り返される。
6-3.核移植-無限のクローンへの挑戦
8 細胞期胚を 8 個の割球に分けても,これらの小
割球は単独では個体へ発生できない。しかし,それ
ぞれの割球の核は分化全能性をもっているので,あ
らかじめ核を抜き取った未受精卵に移植すること
によって,8 個の再構築した胚を新たに作り出すこ
とができる。再構築された 8 個の胚はもともと 1個
の胚に由来する核をもっているので,それぞれの遺
伝子構成は全く等しく,したがってこれらの胚から
生じる個体は胚細胞クローン(受精卵クローン)と
呼ぶことができる。
1997 年に体細胞をドナー細胞としたクローンヒ
ツジ「ドリー」の誕生が報じられた。これを体細胞
クローンという。ドリー誕生後,ウシ,マウス,ブ
タなど多くの動物種で体細胞クローンが作出され
ている。
胚細胞クローンと体細胞クローンの作出の方法
は,ドナー細胞として割球を用いるか,体細胞を用
いるかが異なるだけで,基本的な技術は同様である。
すなわち,除核した未受精卵にドナー細胞の核を導
入して再構築胚を作出し,体外で培養後に代理母へ
胚移植する。このドナー核を導入する際に,除核卵
とドナー細胞を電気的に融合させたり,またはドナ
ー核のみを除核卵に直接移植したりするなどいく
つかの手技が工夫されている。
胚細胞クローンと体細胞クローンの作出技術は
共通する部分が多いが,誕生した個体群がもつ生物
学的な意味は大きく異なる。上述のように,胚細胞

3.00
クローンはもともと 1 個の胚に由来するので,利用
できる割球数には限度があり,また,用いられる胚
は種雄畜と卵を提供する母畜の人工授精または体
外受精により得られるために,両親の遺伝的能力を
どのように受け継いでいるか,成長後に検定しない
と判明しない。そして,この場合生まれた子畜同士
がクローンということになる。これに対して,体細
胞クローンはドナー細胞を提供した親と遺伝的に
は同一なので,たとえば年間乳量
する雌ウシの体細胞由来のクローン子畜は遺伝的
には親と同じ年間乳量 20,000kg
が保証付きであることになる(下図)
7.DNA 組換え動物
DNA 組換え技術は,1970 年代以降,大腸菌や酵母
のような微生物を対象として急速な進歩を遂げた。
この技術により,体外で作製したクローン化遺伝子
を動物の培養細胞に取り込ませることができるよ
うになった。細胞の DNA 組換えでは,
子のことを「外来遺伝子」,外来遺伝子を入れられ
る細胞を「宿主」,そして外来遺伝子
まれた細胞を「DNA 組換え体」という
マウス受精卵へ外来遺伝子 DNA
示す。受精後数時間経っただけのマウス受精卵には
卵自身の雌性前核と受精した精子由来の雄性前核
が融合する前の状態にある。微細なガラスピペット
を用いて,前核の中へ DNA 分子を
クションする。融合前の前核へ外来遺伝子
入すれば,ほぐれた DNA に外来遺伝子が紛れ込み,
受精卵ゲノムに組み込まれる確率が高まる
14
個の胚に由来するので,利用
できる割球数には限度があり,また,用いられる胚
は種雄畜と卵を提供する母畜の人工授精または体
外受精により得られるために,両親の遺伝的能力を
どのように受け継いでいるか,成長後に検定しない
と判明しない。そして,この場合生まれた子畜同士
がクローンということになる。これに対して,体細
胞クローンはドナー細胞を提供した親と遺伝的に
は同一なので,たとえば年間乳量 20,000kg を生産
する雌ウシの体細胞由来のクローン子畜は遺伝的
20,000kg を生産できる能力
(下図)。
年代以降,大腸菌や酵母
のような微生物を対象として急速な進歩を遂げた。
この技術により,体外で作製したクローン化遺伝子
を動物の培養細胞に取り込ませることができるよ
組換えでは,導入する遺伝
子のことを「外来遺伝子」,外来遺伝子を入れられ
る細胞を「宿主」,そして外来遺伝子 DNA が組み込
組換え体」という。
DNAを注入する方法を
だけのマウス受精卵には
卵自身の雌性前核と受精した精子由来の雄性前核
が融合する前の状態にある。微細なガラスピペット
分子をマイクロインジェ
する。融合前の前核へ外来遺伝子 DNA を注
に外来遺伝子が紛れ込み,
に組み込まれる確率が高まると考え
られる。受精卵の遺伝子セットの中に外来遺伝子が
組み込まれてしまえば,その後受精卵が分割してい
く過程でも,やはりその外来遺伝子は各細胞へ伝え
られ,最終的には体細胞のすべてに外来遺伝子をも
つ DNA 組換え個体が得られることになる。
導入される外来遺伝子には特定の
(調節領域)と連結させたものが用いられる。この
理由は,たとえば特定の重金属の投与によって遺伝
子のスイッチを ONとなるよう誘導するためである。
あるいは特定の器官でのみ外来遺伝子のスイッチ
が ONとなるように誘導するためである。
医薬品生産
ウシやヤギ,ヒツジなどの家畜の泌乳能力に着目
し,これらの家畜の乳汁中に有用タンパク質を生産
させ,それを医薬品として利用しようという試みも
ある。その一つに,ヒトの血液凝固因子(第九因子)
の遺伝子を導入した DNA
されている。ヒトの血液凝固因子は血友病の治療に
用いられるが,いわゆる血液製剤は材料用血液の確
保の困難さや,製剤中への病原ウイルスの混入の問
題を抱えている。これに対して,乳タンパク質であ
るカゼインのプロモーター(調節領域)に連結した
ヒト血液凝固因子遺伝子を,ヒツジやウシの受
に導入して DNA 組換え個体を作れば,目的の凝固因
子タンパク質が乳腺で生産され,乳汁中からそれを
回収,利用することが期待できる。
これまでの DNA 組換え技術による医薬品(生理活
性タンパク質)製造の例には,ヒトのインスリンや
成長ホルモン,インターフェロンなどがある。
【7回目はここまで】
られる。受精卵の遺伝子セットの中に外来遺伝子が
組み込まれてしまえば,その後受精卵が分割してい
く過程でも,やはりその外来遺伝子は各細胞へ伝え
られ,最終的には体細胞のすべてに外来遺伝子をも
組換え個体が得られることになる。
導入される外来遺伝子には特定のプロモーター
(調節領域)と連結させたものが用いられる。この
理由は,たとえば特定の重金属の投与によって遺伝
となるよう誘導するためである。
あるいは特定の器官でのみ外来遺伝子のスイッチ
となるように誘導するためである。
ウシやヤギ,ヒツジなどの家畜の泌乳能力に着目
し,これらの家畜の乳汁中に有用タンパク質を生産
させ,それを医薬品として利用しようという試みも
ある。その一つに,ヒトの血液凝固因子(第九因子)
DNA組換えヒツジの作出が報告
されている。ヒトの血液凝固因子は血友病の治療に
用いられるが,いわゆる血液製剤は材料用血液の確
保の困難さや,製剤中への病原ウイルスの混入の問
題を抱えている。これに対して,乳タンパク質であ
るカゼインのプロモーター(調節領域)に連結した
ヒト血液凝固因子遺伝子を,ヒツジやウシの受精卵
組換え個体を作れば,目的の凝固因
子タンパク質が乳腺で生産され,乳汁中からそれを
回収,利用することが期待できる。
組換え技術による医薬品(生理活
性タンパク質)製造の例には,ヒトのインスリンや
成長ホルモン,インターフェロンなどがある。
はここまで】

![線虫学入門 - nematodenematode.net/Data/Presentations/yukoJAPuse.pdf[Onychophora]、緩歩動物門 [Tardigrada]、類線形動物 [Nematomorpha]、動吻動物 [Kinorhyncha]、胴甲動物](https://img.pdfslide.tips/doc/110x75/60bdce5effbe421b83568329/cee-onychophoracce-tardigradaecc.jpg)



























