Embed Size (px)
Citation preview

Sveučilište u Zagrebu
Prirodoslovno-matematički fakultet
Biološki odsjek
Iva Kleiner
Diferencijalna i citokemijska analiza krvnih stanica
zelene žabe (Pelophylax ridibundus, Pallas, 1771)
Diplomski rad
Zagreb, 2013. godina

Ovaj diplomski rad, izrađen u Zavodu za animalnu fiziologiju Prirodoslovno-matematičkog
fakulteta Sveučilišta u Zagrebu, pod vodstvom doc. dr. sc. Domagoja Đikića, predan je na
ocjenu Biološkom odsjeku Prirodoslovno-matematičkog fakulteta Sveučilišta u Zagrebu radi
stjecanja zvanja magistra molekularne biologije.

Zahvale
Puno hvala tehničarki Mariji Potočić i stručnoj suradnici dr. sc. Anici Horvat-Knežević na
stručno odrađenoj pripremi uzorka i spremnosti da mi uvijek uskoče u pomoć.
Hvala mentoru doc. dr. sc Domagoju Đikiću što mi je pružio potpuno samostalnost i
odgovornost tijekom izrade ovog rada. Smatram to odličnom pripremom za pravi posao.
Hvala cijeloj ekipi Zavoda za animalnu fiziologiju na njihovom pozitivnom stavu! Bilo mi je
zadovoljstvo radit s vama!
Hvala Duji na ustupljenom magistarskom radu koji je pomogao tamo gdje je literatura nije.
Također, hvala kolegici Mateji Jarak na pomoći kod izrade i bojanja uzorka. Bez nje bio ovo
bilo mnogo dugotrajnije i upola manje zabavno.
Za kraj, hvala obitelji i prijateljima na potpori i velika hvala Toniju za pomoć oko obrade
slika.
Hvala vam što ste uz mene!

TEMELJNA DOKUMENTACIJSKA KARTICA
Sveučilište u Zagrebu Prirodoslovno-matematički fakultet Biološki odsjek Diplomski rad
Diferencijalna i citokemijska analiza krvnih stanica zelene žabe (Pelophylax ridibundus, Pallas, 1771)
Iva Kleiner
Rooseveltov trg 6, 10000 Zagreb
U ovom radu koristila se puna krv zelene žabe Pelophylax ridibundus. Izračunala se brojnost eritrocita, leukocita te vrijednost hematokrita. Morfološka i kvantitativna analiza krvnih stanica provedena je bojanjem May-Grünwald Giemsom. Diferencijalnim citokemijskim tehnikama napravljena je citološka karakterizacija različitih tipova leukocita. Glikokonjugati i polisaharidi dokazani su tehnikama alcian blue (AB) i periodic acid Schiff (PAS). Stanični lipidi dokazani su tehnikom sudan black B (SBB). Enzimatsko-citokemijskim tehnikama dokazala se aktivnost alfa-naftil acetat esteraza (ANAE), alfa-naftil butirat esteraza (ANBE) te alkalne fosfataze (ALP). Brojnost i udio leukocita, hematokrit i morfologija stanica slični su ostalim žabama, dok je brojnost eritrocita veća. Bojanje ABom imaju jezgre svih stanica, citoplazma limfocita, monocita, dio neutrofila, trombocita i neke granule eozinofila. PASom se boje limfociti, granule eozinofila, neki monociti, neutrofili i trombociti. SBBom se boje leukociti, trombociti, slabije nezreli i zreli eritrociti. Aktivnost ANAE pokazuju monociti i neki limfociti. Aktivnost ANBE imaju monociti, neki trombociti i limfociti. ALP nije dala pozitivne rezultate.
(63 stranica, 44 slika, 9 tablica, 58 literaturna navoda, jezik izvornika: hrvatski) Rad je pohranjen u Središnjoj biološkoj knjižnici. Ključne riječi: boja, eritrocit, hematologija, leukocit, trombocit, vodozemac Voditelj: Dr. sc. Domagoj Đikić, doc. Ocjenitelji: Dr. sc. Domagoj Đikić, doc.
Dr. sc. Gordana Lacković Venturin, prof. Dr. sc. Renata Šoštarić, doc.
Zamjena: Dr. sc. Davor Zanella, doc. Rad prihvaćen: 23.2.2013.

BASIC DOCUMENTATION CARD
University of Zagreb Faculty of Science Department of Biology Graduation Thesis
Differential and Cytochemical Analysis of Blood Cells in Green Frogs (Pelophylax
ridibundus, Pallas, 1771
Iva Kleiner
Rooseveltov trg 6, 10000 Zagreb, Croatia
This thesis presents results attained from peripheral blood of the frog Pelophylax ridibundus. It contains hematocrit values, erythrocytes and leukocytes counts. The May-Grünwald Giemsa stain was used in morphological and quantitative blood cell analysis. Cytological characterization of leukocytes types used differential cytochemical stains. The alcian blue (AB) dye and the periodic acid Schiff method (PAS) stained glycoconjugates and polysaccharides. The sudan black B (SBB) dye stained cell lipids. Activities of alpha-naphthyl acetate esterase (ANAE), alpha-naphthyl butyrate esterase (ANBE) and alkaline phosphatase (ALP) were proved using enzyme-cytochemical reactions. Leukocyte count and percentage, hematocrit and cell morphology are similar to other frogs, unlike erythrocyte counts which are greater. AB stained nuclei of all cells, cytoplasm of lymphocytes, monocytes, some neutrophils, thrombocytes and some eosinophil granules. PAS stained lymphocytes, eosinophil granules, some monocytes, neutrophils and thrombocytes. All leukocytes, thrombocytes, young and mature erythrocytes stained with SBB. Monocytes and lymphocytes have active ANAE. Monocytes, some lymphocytes and thrombocytes have active ANBE. ALP activity was not recorded.
(63 pages, 44 figures, 9 tables, 58 references, original in: Croatian) Thesis deposited in the Central Biological Library Key words: amphibian, dye, erythrocyte, leukocyte, hematology, stain, thrombocyte Supervisor: Dr. Domagoj Đikić, Asst. Prof. Reviewers: Dr. sc. Domagoj Đikić, doc.
Dr. sc. Gordana Lacković Venturin, prof. Dr. sc. Renata Šoštarić, doc.
Replacement: Dr. sc. Davor Zanella, doc. Thesis accepted: February 23rd, 2013.

POPIS KRATICA:
AB alcijansko modrilo, engl. alcian blue
ACP kisela fofataza, engl. acid phosphatase
ALP alkalna fosfataza, engl. alkaline phosphatase
ANAE alfa-naftil acetat esteraza, engl. α-naphthyl acetate esterase
ANA alfa-naftil acetat
ANBE alfa-naftil butirat esteraza, engl. α-naphthyl butyrate esterase
ANBE alfa-naftil butirat
CFU-E stanice koje formiraju eritrocite, engl. colony-forming unit-erythrocyte
CFU-GM stanice koje formiraju granulocite i monocite, engl. colony-forming unit-
granulocytes,monocytes
CFU-M stanice koje formiraju megakariocite, engl. colony-forming unit-megakaryocytes
dH2O destilirana voda
DKS diferencijalna krvna slika
EG etilen glikol
EGME etilen glikol monometil eter
GAG glikozaminoglikani, engl. glycosaminoglycans
GalNAc N- acetilgalaktozamin
GlcNAc N-acetilglukozamin
MGG May-Grünwald Giemsa
NFR engl. nuclear fast red
N-H Natt- Herrick
NK stanice ubojice, engl. natural killer cells
NSE nespecifična esteraza, engl. non-specific esterase
PAS metoda Schiffove baze i perjodne kiseline, engl. periodic acid Schiff
SBB sudansko crnilo, engl. sudan black B

SADRŽAJ: 1. UVOD ................................................................................................................................................. 1
1.1. Izgled, građa, uloga i razvoj pojedinih krvnih stanica u kralježnjaka .......................................... 1
1.1.1. Eritrociti................................................................................................................................. 3
1.1.2. Trombociti ............................................................................................................................. 5
1.1.3. Leukociti ................................................................................................................................ 7
1.2. Citokemijske tehnike .................................................................................................................. 14
1.3. Citokemijske karakteristike krvnih stanica u kralježnjaka ......................................................... 25
2. CILJ ISTRAŽIVANJA...................................................................................................................... 26
3. MATERIJALI I METODE ............................................................................................................... 27
3.1. Uzorci ......................................................................................................................................... 27
3.2. Određivanje ukupnog broja eritrocita i leukocita ....................................................................... 27
3.3. Određivanje vrijednosti hematokrita .......................................................................................... 28
3.4. Bojanja May-Grünwald Giemsom (MGG) ................................................................................ 28
3.4.1. Morfološke karakteristike krvnih stanica ............................................................................ 28
3.4.2. Računalna obrada podataka ................................................................................................. 28
3.5. Citokemijske analize .................................................................................................................. 28
3.5.1. Alcijansko modrilo (AB) ..................................................................................................... 29
3.5.2. Alfa-naftil acetat esteraza (ANAE) ..................................................................................... 30
3.5.3. Alfa-naftil butirat esteraza (ANBE) .................................................................................... 31
3.5.4. Alkalna fosfataza (ALP) ...................................................................................................... 32
3.5.5. Metoda Schiffove baze i perjodne kiseline (PAS)............................................................... 33
3.5.6. PAS i AB kombinacija ........................................................................................................ 34
3.5.7. Sudansko crnilo B (SBB) .................................................................................................... 34
3.5.8. Morfološke karakteristike krvnih stanica ............................................................................ 35
4. REZULTATI ..................................................................................................................................... 36
4.1. Određivanje ukupnog broja eritrocita i leukocita ....................................................................... 36
4.2. Određivanje vrijednosti hematokrita .......................................................................................... 36
4.3. Diferencijalna analiza pomoću bojanja May-Grünwald Giemsom ............................................ 36
4.3.1 Eritrociti................................................................................................................................ 36
4.3.2. Trombociti ........................................................................................................................... 37
4.3.3. Agranulociti, limfociti ......................................................................................................... 38
4.3.4. Granulociti, neutrofili .......................................................................................................... 38
4.3.5. Granulociti, eozinofili.......................................................................................................... 38
4.3.6. Agranulociti, monociti ......................................................................................................... 38

4.3.7. Granulociti, bazofili ............................................................................................................. 39
4.4. Citokemijske analize .................................................................................................................. 41
4.4.1. Alcijansko modrilo (AB) ..................................................................................................... 41
4.4.2. Alfa-naftil acetat esteraza (ANAE) ..................................................................................... 42
4.4.3. Alfa-naftil butirat esteraza (ANBE) .................................................................................... 43
4.4.4 Alkalna fosfataza (ALP) ....................................................................................................... 43
4.4.5. Metoda Schiffove baze i perjodne kiseline (PAS)............................................................... 43
4.4.6. PAS i AB kombinacija ........................................................................................................ 44
4.4.7. Sudansko crnilo B (SBB) .................................................................................................... 44
4.5. Atipične i neklasificirane stanice................................................................................................ 45
4.5.1. "Raspadnute jezgre". ........................................................................................................... 45
4.5.2. Neobične stanice .................................................................................................................. 46
4.5.4. Azurofili i heterofili ............................................................................................................. 47
4.5.5. Morfološke neobičnosti krvnih stanica ............................................................................... 47
5. RASPRAVA ...................................................................................................................................... 49
6. ZAKLJUČAK ................................................................................................................................... 58
7. LITERATURA .................................................................................................................................. 59

UVOD
1
1. UVOD
Vrlo je malo objavljenih radova o hematologiji vodozemaca. Vrsta koja je najviše
korištena u istraživanjima hematologije vodozemaca je tropska žaba panđašica Xenopus laevis
koja je standardizirana kao laboratorijska životinja i česti je model u laboratorijskom radu i
fiziološkim istraživanjima.
Krvne stanice vodozemaca dijele neke sličnosti (poglavito morfološke) sa ostalim
kralježnjacima, ali u nekim pogledima (npr. hematopoeza u ličinačkim stadijima) sličnije su
nižim kralježnjacima (Claver i Quaglia 2009).
U najčešće rutinske analize krvi spada niz pretraga u kojima se mjeri brojnost leukocita,
eritrocita i trombocita, diferencijalna krvna slika (DKS), koncentracija hemoglobina,
vrijednost hematokrita te indeksi eritrocita (Desai 2006; Dugdale 2011). DKS određuje
relativni broj, odnosno udio svakog tipa leukocita u perifernoj krvi (Dugdale 2011; Piršljin i
Milinković Tur 2009).
U ovom je radu izmjerena brojnost eritrocita i leukocita, dobivena je vrijednost
hematokrita i napravljen je DKS. Također su napravljene citokemijske analize za
karakterizaciju pojedinih tipova i podtipova krvnih stanica zelene žabe Pelophylax
ridibundus.
Podaci o diferencijalnoj citokemijskoj analizi krvnih stanica P. ridibundus ne postoje u
dostupnoj literaturi. Kako ova vrsta, uz još nekoliko drugih vrsta iz roda Pelophylax,
predstavlja eksperimentalnu životinju u različitim fiziološkim, genetičkim i evolucijsko
specijacijskim istraživanjima (Feidantsis i sur. 2012; Fominykh i Liapkov 2011; Marracci i
sur. 2011; Usmanova i sur. 2010), ovaj rad predstavlja temeljno istraživanje sa svrhom
utvrđivanja referentnih podataka o citokemijskim karakteristikama stanica krvi.
1.1. Izgled, građa, uloga i razvoj pojedinih krvnih stanica u kralježnjaka
Kod embrija sisavaca hematopoeza počinje u žumanjčanoj vreći (gdje se stvaraju
eritrociti sa jezgrama), nastavlja se u jetri, zatim u slezeni i limfnim čvorovima i konačno u
koštanoj srži. Sve krvne stanice nastaju iz pluripotentnih hematopoetskih matičnih stanica.
Prilikom njihovih diobi jedna stanica kćer ostaje pluripotentna dok druga ide dalje u
intermedijarnu fazu gdje postaje usmjerena za određenu staničnu liniju. Postoji više različitih
usmjerenih matična stanica (Slika 1.), colony-forming unit-erythrocyte (CFU-E) stanice koje
formiraju eritrocite, colony-forming unit-granulocytes, monocytes (CFU-GM) stanice koje

UVOD
2
stvaraju granulocite i monocite i colony-forming unit-megakaryocytes (CFU-M) stanice iz
kojih nastaju megakariociti (Guyton i Hall 2006).
Kod embrija ptica glavno mjesto hematopoeze također je žumanjčana vreća u kojoj
nastaju samo eritrociti i trombociti. Nakon izlijeganja sve se krvne stanice proizvode u
koštanoj srži. Kod gmazova mjesto hematopoeze je isto kao i u ptica osim što njihova jetra i
slezena sudjeluju u hematopoezi u ranijim razvojnim fazama, slično procesu embrionalne
hematopoeze u sisavaca (Claver i Quaglia 2009).
U embrionalnom razvoju vodozemaca hematopoeza se događa u tkivu analognom
žumančanoj vreći viših kralježnjaka, dok je u ličinačkim stadijima (primjerice stadij
punoglavaca u Anura ili ličinačkom stadiju u Urodela) bubreg centar hematopoeze. Općenito,
kod odraslih žaba koštana srž nema značajnu hematopoetsku ulogu (Claver i Quaglia 2009).
Kod Xenopus laevis glavni organi hematopoeze su jetra, bedrena kost i slezena (Hadji-Azimi i
sur. 1987).
Kako ribe nemaju koštanu srž i limfne čvorove morale su razviti jedinstvene
hemtopoetske organe. Čak se i u embrionalnoj hematopoezi razlikuju od ostalih kralježnjaka.
U većini odraslih koštunjača prozonefros (anteriorni bubreg) glavni je hematopoetski organ.
Neke koštunjače koriste slezenu i srce za hematopoezu, a zabilježeno je da se pojedini tipovi
stanica mogu dijeliti i u cirkulaciji (Claver i Quaglia 2009).

UVOD
3
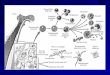
Slika 1. Pojednostavljeni prikaz hematopoeze kod ljudi. Različite krvne stanice nastaju iz pluripotentne
hematopoetske matične stanice. CFU-E (colony forming unit- erythrocytes), CFU-GM (colony forming unit-
granulocytes, monocytes), CFU-M (colony forming unit- megakaryocytes). Obrađeno iz Guyton i Hall
(2006).
1.1.1. Eritrociti
Zreli eritrociti (crvena krvna zrnca) sisavaca (Slika 2.a) nemaju jezgru ni organele čime
gube sposobnost sinteze proteina i nukleinskih kiselina. Veličina im varira između 6 do 9 μm
te izgledaju kao biokonkavni diskovi (Guyton i Hall 2006; Weiss i Wardrop 2010). Deve su
jedina skupina sisavaca koje imaju eliptičan oblik eritrocita. Kod ostalih kralježnjaka eritrociti
su ovalnog ili elipsoidnog oblika sa centralno pozicioniranom jezgrom. Citoplazma je svijetlo
roze boje. Kod ptica (Slika 2.b) čini tanki, bijeli obrub oko jezgre (Arserim i Mermer 2008;
Claver i Quaglia 2009; Weiss i Wardrop 2010). Gmazovi imaju okruglaste jezgre (Slika 2.c)
za razliku od ovalnih kod ptica i vodozemca. Najveće eritrocite imaju upravo vodozemci
(Slika 2.d). Kod salamandera Amphiuma tridactylum iznose 70 x 40 μm što ih čini najvećim
eritrocitima u životinjskom svijetu (Claver i Quaglia 2009). Duljina eritrocita X. laevis varira
od 17 do 19 μm, a širina 9 do 12 μm. Duljina jezgre kreće se od 5 do 9 μm, a širina 4 do 5 μm

UVOD
4
(Hadji-Azimi i sur. 1987). Prosječan broj eritrocita ljudi je oko 5 000 000/ml krvi (Guyton i
Hall 2006), a kod X. laevis taj broj iznosi oko 75 000/ ml krvi (Hadji-Azimi i sur. 1987).
Zahvaljujući hemoglobinu, eritrociti vežu, prenose i predaju kisik tkivima, odnosno
uzimaju ugljični dioksid iz njih. Hemoglobin je proteinski kompleks, tetramer sastavljen od
dva αβ dimera. α i β su globinski lanci od kojih svaki nosi hem skupinu (Nelson i Cox 2008).
Hemoglobin se počinje sintetizirati u rastućim eritrocitima, tj. eritroidnim prekursorskim
stanicama.
Prosječni život zrelog eritrocita čovjeka je 120 dana (Guyton i Hall 2006), kod gmazova
600 do 800 dana, a kod vodozemaca 700-1400 dana (Claver i Quaglia 2009).
Kod ljudi nastanak eritrocita počinje iz CFU-E stanica (Slika 3.). Pod određenim
uvjetima prelaze u proeritroblaste, a u idućem koraku postaju bazofilni eritrociti sa vrlo malo
hemoglobina. Kako stanice mijenjaju oblik (polikromatofilni, ortokromatofilni eritrociti) tako
sintetiziraju sve više hemoglobina, jezgra je sve manja i u fazi retikulocita dolazi do
reapsorpcije endoplazmatskog retikuluma i jezgre (koja se može izbaciti izvan stanice).
Retikulociti se sa ostacima mitohondrija i Golgijevog aparata protiskuju kroz koštanu srž u
krvne kapilare procesom dijapedeze. U iduća dva dana sav bazofilni materijal nestane iz
retikulocita i oni postanu zreli eritrociti (Guyton i Hall 2006). X. laevis ima dosta sličan
proces (Slika 4.). Diferencijacija eritrocita se događa na periferiji jetre prilikom čega dolazi do
kondenzacije i smanjenja jezgre, dok veličina stanice ostaje gotovo ista. Iz matične stanice
razvija se proeritroblast, okrugla stanica sa velikim udjelom jezgre. Daljnjom diferencijacijom
nastaju eritroblast I i II gdje dolazi do izduljivanja stanice, povećanja količine hemoglobina te
kondenzacije jezgre. Nezrele eritrocite I i II karakterizira povećanje obujma citoplazme i
dodatno kondenziranje i smanjenje jezgre koja poprimi okrugli oblik. U tom stadiju
citoplazma ima jednolično obojenje zbog visoke količine hemoglobina. Konačno, zreli
eritrocit ovalnog je oblika sa ovalnom, centralno smještenom jezgrom (Hadji-Azimi i sur,
1987). Zanimljivo kako eritrociti gmazova mogu nastati transformacijom trombocita (Pendl
2006).
Membrana eritrocita ima tipični asimetrični fosfolipidni dvosloj u kojem se nalaze
glikolipidi, kolesterol, proteini i glikokonjuganti (Alberts i sur. 1994; Nelson i Cox 2008).
Jedan od bitnih membranskih glikoproteina je glikoforin. Bogat je negativno nabijenom
sijalinskom kiselinom dajući eritrocitu ukupni površinski negativni naboj (Weiss i Wardrop
2010).
Najčešćih poremećaji u broju eritrocita su anemije, poput aplastične i hemolitičke, te
eritrocitoze, odnosno policitemije (Guyton i Hall 2006; Weiss i Wardop 2010). Kod ljudi,

UVOD
5
ptica i gmazova zabilježeno je nekoliko morfoloških poremećaja poput anizocitoze (eritrociti
različitih veličina), poikilocitoze (različiti oblik), hiperkromije (prejaka obojanost) i prisutnost
normoblasta tj. eritroblasta u ljudi, odnosno eritrocita bez jezgre kod gmazova (Pendl 2006;
Piršljin i Milinković Tur 2009).
Slika 2. Eritrocit čovjeka (a), ptice (b), gmaza (c), vodozemca (d) i ribe (e). Proporcije nisu nužno realne.
Preutezo iz Claver i Quaglia (2009) i Reece (2012).
Slika 3.. Nastanak eritrocita u ljudi. Preuzeto iz Guyton i Hall (2006).
Slika 4. Eritropoeza X. laevis. Proeritroblast (a), eritroblast I (b), eritroblast II (c), nezreli eritrocit I (d),
nezreli eritrocit II (e) i zreli eritrocti (f). Obrađeno iz Hadji-Azimi i sur. (1987).
1.1.2. Trombociti
Trombociti ili krvne pločice u sisavaca (Slika 5.) izgledaju kao mali diskovi promjera 1
do 4 μm (Guyton i Hall 2006). Kod ptica, gmazova, vodozemaca i riba najčešće su okrugli ili
ovalni sa okruglom ili izduženom jezgrom. Velika je morfološka sličnost između nezrelih
trombocita i malih limfocita u gmazova i vodozemaca (Claver i Quaglia 2009). Kod ruskih

UVOD
6
kornjača čak su pronađeni pravokutni trombociti (Knotková i sur. 2002) dok je za ribe česti
vretenasti oblik (Claver i Quaglia 2009; Shigdar i sur. 2007). Zreli trombociti X. laevis su
izduženog, ovalnog oblika različith dimenzija. Citoplazma je svijetlo plave boje, oko
centralno smještene izdužene, tamno ljubičaste jezgre. Kod manjih, izuženih trombocita
jezgra je vrlo kondenzirana, za razliku od većih i okruglijih. Njihova se duljina kod kreće od 7
do 16 μm, a širina 3 do 5 μm. Prosječni broj trombocita X. laevis je oko 19 000/ ml krvi
(Hadji-Azimi i sur. 1987), a čovjeka 225 000 000/ ml krvi (Guyton i Hall 2006).
Kod svih kralježnjaka trombociti imaju glavnu ulogu u aktivaciji mehanizma
zgrušavanja krvi (Claver i Quaglia 2009; Guyton i Hall 2006). Kod ptica i riba sposobni su
fagocitirati i u ovih skupina možda sudjeluju u nespecifičnoj imunosti, iako među
komparativnim hematolozima nikada nije postignuto usaglašavanje mišljenja o ovoj temi
(Claver i Quaglia 2009).
Kod sisavaca trombociti nastaju fragmentacijom iz megakariocita koji se sintetiziraju u
koštanoj srži (Guyton i Hall 2006; Weiss i Wardrop 2010). Nastanak trombocita X. laevis
događa se na periferiji jetri (Slika 6.). Umjesto megakariocita postoji protromboblast, velika
jezgrena stanica nepravilnog oblika koja sazrijevanjem smanjuje obujam citoplazme i jezgre.
U idućem stadiju, stadiju tromboblasta, počinje kondenzacija kromatina. Sazrijevanjem u
nezreli trombocit jezgra je potpuno kondenzirana, trombocit ima vrlo malo citoplazme i
poprima ovalni oblik. Konačno, zreli trombocit ima sposobnost rasta i dijeljenja i često se
nalazi u nakupinama s drugim trombocitima (Hadji-Azimi i sur. 1987).
Najčešći poremećaji u broju trombocita sisavaca su trombocitopenija, smanjeni broj
trombocita, i trombocitoza, povećani broj trombocita. Neki poremećaji grušanja krvi (npr.
Von Willebrandova bolest) nastaju zbog abnormalnih trombocita (Weiss i Wardop 2010).
Slika 5. Ljudski trombocit (a), ptičji (b), trombocit gmazova - desni je aktiviran (c), vodozemca (d) i ribe-
skroz desni je aktiviran (e). Proporcije nisu nužno realne. Preutezo iz Claver i Quaglia (2009) i Reece
(2012).

UVOD
7
Slika 6. Trombopoeza X. laevis. Protromboblast (a), tromboblast (b), nezreli trombocit i zreli eritorcit (c) i
nakupina zrelih trombocita (d). Preuzeto iz Hadji-Azimi i sur. (1987).
1.1.3. Leukociti
Leukociti ili bijela krvna zrnca krvne su stanice sa jezgrom, što je potvrđeno kod svih
istraživanih kralježnjaka od riba sve do sisavaca. Glavni su u obrani organizma. Kod sisavaca
postoji pet tipova leukocita: neutrofili, eozinofili, bazofili, monociti i limfociti. Prema
morfološkim svojstvima dijele se na granulocite i agranulocite. Granulociti u svojoj
citoplazmi sadrže specifično obojena zrnca (granule), a prema boji zrnaca razlikuju se
neturofili, eozinofili i bazofili. Mogu imati segmentiranu ili nesegmentiranu jezgru.
Agranulociti nemaju citoplazmatska zrnca i dijele se na limfocite i monocite (Guyton i Hall
2006; Weiss i Wardrop 2010).
Prosječan broj ukupnih leukocita kod ljudi je oko 7000/ ml krvi. Najzastupljeniji su
neutrofili (62 %) i limfociti (30 %) dok monocita (2,3 %), eozinofila (2,3 %) i bazofila (0,4
%) ima najmanje (Guyton i Hall 2006). Kod X. laevis prosječn broj leukocita je 5000/ ml krvi
(Hadji-Azimi i sur. 1987). Limfociti i neutrofili su također najčešći (30 % i 27 %), dok su
eozinofili (1,2 %) i monociti (1,6 %) najrjeđi tipovi. Zanimljivo je kako bazofili čine oko 40
% ukupnih leukocita X. laevis (Hadji-Azimi i sur. 1987). Pojedini tipovi leukociti gmazova
teže se klasificiraju zbog jakih morfoloških varijacija među vrstama (Knotková i sur. 2002).
Prosječan život granulocita čovjeka nakon ispuštanja iz koštane srži u krv je 4-8 sati te 4
do 5 dana u ciljnom tkivu. Prilikom jakih infekcija granulociti žive još kraće. Monociti u krvi
mogu izdržati 10 do 20 sati, a čim uđu u tkivo narastu u makrofage koji mogu živjeti nekoliko
mjeseci (Guyton i Hall 2006). Kod sisavaca granulociti, monociti i manji dio limfocita
proizvode se i čuvaju u koštanoj srži dok se plazma stanice i većina limfocita proizvodi i čuva
u limfogenim tkivima- limfnim čvorovima, slezeni, timusu i mandulama. Po potrebi se

UVOD
8
leukociti ispuštaju u cirkulaciju i prenose u različite dijelove tijela omogućavajući trenutnu
reakciju na infekcije i upale (Guyton i Hall 2006).
Leukociti nastaju iz usmjerenih matičnih stanica (CFU-GM i limfoidne matične stanice)
koje tvore mijelocitne i limfocitne loze. Mijelocitna loza (Slika 7.) počinje mijeloblastom sa
karakterističnim primarnim, azurofilnim granulama. Neutrofili, eozinofili i bazofili sazrijevaju
slično, počevši iz promijelocita kojeg karateriziraju sekundarne bazofilne granule i smanjeni
broj primarnih. Daljnjim sazrijevanjem nastaje usmjereni mijelocit. To je ujedno i zadnja
stanica sposobna za diobu. Prije formiranja zrelih eozinofila i neutrofila, u prijelaznom obliku
zvanom metamijelocit dolazi do kondenzacije i segmentacije jezgre. Monociti nastaju
postepenom diferencijacijom iz CFU-GM stanica u monoblaste, promonocite i konačno
monocite (Venkat 2010; Weiss i Wardrop 2010). Većina granulopoeze X. laevis događa se u
slezeni, a manji dio u jetri. Stadiji i loze su gotovo identični kao u sisavaca (Hadji-Azimi i sur.
1987).
Limfopoeza X. laevis događa se u timusu, gdje se limfocitne populacije zovu timociti, i
u slezeni, gdje se limfocitne populacije zovu limfociti slezene. Limfoidna loza počinje
protimocitom (u timusu), odnosno limfoblastom (u slezeni) koji se postepeno diferencira u
timocite I, II, III i IV, odnosno limfocite I, II, III i IV. Prilikom toga dolazi do značajnog
smanjenja stanice od početnih 13 μm do 6 μm. Kod sisavaca iz limfoidne loze (Slika 8.)
nastaju različiti podtipovi limfocita: B i T stanice, plazma stanice i tzv. stanice ubojice, NK
stanice (engl. natural killer). B limfociti i plazma stanice X. laevis nastaju u slezeni, T
limfociti u timusu, a NK stanice u oba organa (Hadji-Azimi i sur. 1987). Kod ljudi T limfociti
također nastaju u timusu, ali B limfociti sazrijevaju u koštanoj srži. Plazma stanice nastaju iz
aktiviranih B limfocita (Guyton i Hall 2006; Weiss i Wardrop 2010).
Najčešći poremećaji broja leukocita su leukopenije, smanjeni broj leukocita, i
leukemije, povećani broj (Guyton i Hall 2006).

UVOD
9
Slika 7. Shematski prikaz nastanka leukocita iz mijelocitne loze kod ljudi. Preuzeto iz Guyton i Hall
(2006).
Slika 8. Pojednostavljeni shematski prikaz nastanka različitih limfocita. Preuzeto iz Venkat (2010).
Bazofili (Slika 9.) svih kralježnjaka okrugle su stanice sa centralno ili periferno
smještenom jezgrom (Claver i Quaglia 2009; Piršljin i Milinković Tur 2009; Shigdar i sur.

UVOD
10
2007). U citoplazmi se nalaze granule tamno plave do crne boje koje obično ne prekrivaju
jezgru. Sisavci, za razliku od vodozemaca, imaju segmentiranu jezgru bazofila (Piršljin i
Milinković Tur 2009). Kod sisavaca imaju promjer 10 do15 μm i najrjeđe su stanice (Lester i
sur. 2005). U X. laevis dosta su česti leukociti, a promjer im se kreće od 7 do 16 μm (Hadji-
Azimi i sur. 1987). U krvi kornjače također ih ima više nego kod sisavaca, nekada čine 50%
do 60% ukupnih leukocita (Claver i Quaglia 2009, Knotková i sur. 2002). Kod riba imaju
varijabilan izgled, ima ih jako malo i nisu prisutni u svim vrstama (Claver i Quaglia 2009).
Mogu imati segmentiranu jezgru (Shigdar i sur. 2007).
Bazofili ispuštaju u krv heparin, koji sprečava zgrušavanje, te sudjeluju u alergijskim
reakcijama prilikom kojih ispuštaju velike količine histamina, serotonina, anafilaktičkih tvari,
bradikinina i lizosomnih enzima (Guyton i Hall 2006). Žive oko 3 do 10 dana (Reece 2012).
Slika 9. Bazofil čovjeka (a), ptice (b), gmaza (c), vodozemca (d) i ribe (e). Proporcije nisu nužno realne.
Preutezo iz Claver i Quaglia (2009) i Reece (2012).
Eozinofili (Slika 10.) kralježnjaka okruglog su oblika sa blijedo plavom citoplazmom
bogata granulama crvene boje i ljubičastom jezgrom. Sisavci obično imaju dva segmenta
jezgre (Lester i sur. 2005). Ljudski eozinofili imaju promjer od 10 do 15 μm (Piršljin i
Milinković Tur 2009). Citoplazmatske granule okrugle su kod ptica i gmazova, a kod
vodozemca i riba također mogu biti izdužene. Gmazovi i ribe jedini imaju samo okrugle,
nesegmentirane jezgre (Claver i Quaglia 2009). Promjer eozinofila kod X. laevis iznosi od 13
do 17 μm (Hadji-Azimi i sur. 1987).
Granule su vrste modificiranih lizosoma koji sadržavaju razne enzime (npr. hidrolitičke)
i larvacidne polipeptide (Guyton i Hall, 2006). Eozinofili su slabi fagociti koji sudjeluju u
upalnim procesima uzrokovanim parazitima i alergijama. Fagocitiraju i uništavaju parazite te
smanjuju upalne procese (Guyton i Hall 2006). Žive oko 3 mjeseca (Reece 2012).

UVOD
11
Slika 10. Eozinofil čovjeka (a), ptice (b), gmaza (c), vodozemca (d) i ribe (e). Proporcije nisu nužno realne.
Preutezo iz Claver i Quaglia (2009) i Reece (2012).
Limfociti (Slika 11.) su jedni od najčešćih tipova leukocita kralježnjaka. Kod sisavaca
okruglog su oblika sa velikom okruglom jezgrom smještenom periferno. Citoplazma se vidi
kao mali svijetlo plavi prsten. Razlikuju se veliki, promjera 12-15 μm i mali, promjera 10 μm
(Weiss i Wardrop 2010). Limfociti ptica imaju gotovo isti izgled kao kod sisavaca, jedino što
mogu biti nepravilnog oblika. Gmazovi, vodozemci i ribe također imaju slični izgled limfocita
kao sisavci (Claver i Quaglia 2009). Mali limfociti X. laevis promjera su od 5 do 8 μm, a
veliki 10 do 11 μm (Hadji-Azimi i sur. 1987). U riba također spadaju u najčešću vrstu
leukocita (Shigdar i sur. 2007).
T i B limfociti, plazma i NK stanica teško se mogu morfološki razlikovati. NK stanice
dio su urođenog imunološkog odgovora i izgledaju kao veliki granulirani limfociti. T limfociti
(prezentiraju antigene), B limfociti i plazma stanice (proizvode antitijela) dio su stečenog
imunološkog odgovora (Guyton i Hall 2006; Weiss i Wardrop 2010). Ljudski B limfociti žive
4-5 tjedna. T limfociti mogu živjeti od jednog dana do više mjeseci ovisno o njihovoj
aktivnosti (Reece 2012).
Slika 11. Limfocit čovjeka (a), mali i veliki limfociti kod ptice (b), mali limfocit gmaza (c), mali limfocit
vodozemca (d) i limfocit ribe (e). Proporcije nisu nužno realne. Preuzeto iz Claver i Quaglia (2009) i Reece
(2012).
Monociti (Slika 12.) kralježnjaka najveće su krvne stanice, okruglog oblika. Imaju
ljubičastu nesegmentiranu ekscentrično smještenu jezgru, koja zna poprimiti nepravilni,
zaobljeni oblik. Citoplazma im je sivkasto plava, a kromatinska struktura im je rjeđa nego kod
limfocita (Claver i Quaglia 2009; Guyton i Hall 2006; Lester i sur. 2005). Kod ljudi promjer

UVOD
12
im varira od 16 do 22,5 μm (Piršljin i Milinković Tur 2009). Promjer kod X. laevis kreće se
od 11,5 do 15 μm (Hadji-Azimi i sur. 1987).
Za razliku od neutrofila monociti nemaju veliku sposobnost obrane od patogena te
moraju sazreti u makrofage što se događa u tkivima. Kod X. laevis diferencijacija se događa u
jetri (Hadji-Azimi i sur. 1987). Fagocitoza ovisi o više faktora, strukturi tvari, zaštitnim
proteinima (koji nedostaju mrtvim stanicama) i antitijelima (koja se vežu na membrane
patogena označavajući ih metom za fagocitozu). Makrofagi su sposobni fagocitirati stotinjak
bakterija tijekom svog života i probavljati velike čestice poput eritrocita što ih čini jačim od
neutrofila. Zanimljivo je kako i monociti i neutrofili imaju baktericidne agense u slučaju da
probavljanje lizosomima nije efikasno (Guyton i Hall 2006).
Slika 12. Monocit čovjeka (a), ptice (b), gmaza (c), vodozemca (d) i ribe (e). Proporcije nisu nužno realne.
Preutezo iz Claver i Quaglia (2009) i Reece (2012).
Morfologija neutrofila slična je kod svih kralježnjaka (Slika 13.). Ljudski neutrofili, čiji
promjer stanice varira od 10 do 15 μm, imaju fino granuliranu citoplazmu. Jezgra može biti u
komadu ili segmentirana, što je češće kod zrelijih neutrofila gdje se broj segmenta jezgri kreće
od tri od pet, s tim da spojnica među segmentima mora biti manja od 1/3 najdebljeg dijela
(Piršljin i Milinković Tur 2009). Neki autori smatraju kako ptice, gmazovi i ribe nemaju
neutrofile, nego samo heterofile (Claver i Quaglia 2009). Promjer neutrofila X. laevis kreće se
od 11 do 19 μm. Zbog bazofilnih i acidofilnih citoplazmatskih granula poprimaju blijedo rozu
boju dok su im segmenti jezgre tamno ljubičasti. Jedni su od rijetkih leukocita koje nastaju u
koštanoj srži (Hadji-Azimi i sur. 1987).
Uloga neutrofila je napad i uništavanje invadirajućih organizama, poput bakterija i
virusa te uklanjanje nekrotičnog tkiva. Čim uđu u tkiva su sposobni fagocitirati 3 do 20
bakterija prije nego što se inaktiviraju i odumru (Guyton i Hall 2006).

UVOD
13
Slika 13. Neutrofil čovjeka (a) i vodozemca (b). Proporcije nisu nužno realne. Preutezo iz Claver i Quaglia
(2009) i Reece (2012).
Heterofili (Slika 14.) vrsta su leukocita prisutnih kod većine kralježnjaka. Sliče
eozinfilima s glavnom razlikom u obliku citoplazmatskih granula izuženog oblika (Claver i
Quaglia 2009; Knotková i sur. 2002). Kod ptica najčešća su vrsta leukocita. Citoplazma im je
bezbojna sa acidofilnim okruglastim granulama. Kod gmazova izgled heterofila varira ovisno
o vrsti, ali većina ima okruglu jezgru. Heterofili vodozemca imaju segmentiranu jezgru i
nekad ih je teško razlučiti od neutrofila i eozinofila. Kod riba imaju varijabilnu morfologiju
granula i jezgre (Claver i Quaglia 2009). U sisavaca su pronađeni kod zečeva i zamorčića gdje
im je promjer 10 do 15 μm (Lester i sur. 2005; Weiss i Wardrop 2010).
Heterofili imaju sličnu ulogu kao neutrofili sisavaca.
Slika 14. Heterofil ptice (a), gmaza (b), vodozemca (c) i ribe (d). Proporcije nisu nužno realne. Preutezo iz
Claver i Quaglia (2009) i Reece (2012).
Azurofili (Slika 15.) su pronađeni kod gmazova i vodozemaca. Morfološki sliče
monocitima, ali sadržavaju azurofilne granule. Neki autori smatraju da potječu iz iste linije
kao monociti (Knotková i sur. 2002) dok ih drugi smatraju vrstom neutrofila ili monocita
(Claver i Quaglia 2009).
Imaju fagocitoznu ulogu.

UVOD
14
Slika 15. Azurofil gmaza (a) i vodozemca (b) Proporcije nisu nužno realne. Preutezo iz Claver i Quaglia
(2009) i Reece (2012).
1.2. Citokemijske tehnike
Za karakterizaciju pojedinih tipova leukocita koriste se posebne boje kojima se
vizualiziraju određene stanične komponente poput lipida, ugljikohidrata i proteina.
Boje imaju dvije funkcionalne skupine, kromofor (nekad zvan i kromogen) za bojanje i
auksokrom za vezanje sa supstratom i pojačanje intenziteta boje. Auksokromi mogu biti
ionizirajuće skupine, mogu se vezati kovalentno sa supstratom ili raditi koordinirane veze.
Mogu biti lužnati zbog amino grupa ili kiseli zbog karboksilnih grupa i fenola (Kiernan
2008). Boju koju vidimo nastaje prolaskom bijelog svjetla koz objekt koji apsorbira određene
valne duljine, nakon čega iz objekta proizlazi boja komplementarna apsorbiranoj. Naime,
bijelo svjetlo nastaje miješanjem komplementarnih boja, a oduzimanjem (apsorbiranjem)
jedne boje (npr. žute), vidimo drugu (plavu). Smatra se kako boja organskih bojila nastaje na
temelju energetskih razlika između elektron donora i elektron akceptora. Obično kromofori
imaju azo, karbonilnu i nitro skupinu te su akceptori, a auksokromi, sa amino i hidroksi
grupom, donori elektrona (Christie 2001). Batokromtski pomak je promjena spektra molekule
prema dužim valnim duljnama, prema crvenom dijelu spektra. Neke grupe jače utječu na
pomak od drugih: aromati sa NH skupinom jače utječu nego NH2 skupina koja je jača od OH
skupine. Suprotna pojava, promjena prema kraćim apsorbiranim valnim duljinama, zove se
hipsokromatski pomak (Christie 2001).
Postoji više interakcija između boja i tkiva. Van der Waalsove sile javljaju se kod
aromatskih boja (azo boje), vodikove veze najviše su prisutne kod boja za vezivna tkiva, a
Coulombove interakcije javljaju se kod kiselih i lužnatih boja. Kovalentne interakcije postoje
kod enizmskih reakcija esteraza te metode Schiffove baze i perjodne kiseline (engl. periodic
acid Schiff, PAS), a hidrofobne veze kod nepolarnih boja (Bancroft i Gamble 2008).
Na selektivnost boja utječe nekoliko faktora: naboj, veličina i polarnost. Kationske
(lužnate boje) pozitivnog su naboja koje interagiraju sa anionskim (negativno nabijenim)
biopolimerima (npr. nukleinskim kiselinama i glikozaminoglikanima). Velike boje (poput

UVOD
15
alcian bluea) sporo difundiraju kroz tkiva i stanice te će lakše prodrijeti u rahle strukture,
naspram kompaktnog kromatina. Za bojanje nepolarnih, hidrofobnih, odnosno lipofilnih
struktura koriste se ne-ionske boje (Kumar i Kiernan 2010).
Postoje dva načina klasificiranja boja- prema namjeni ili prema kemijskoj strukturi.
Prema namjeni razlikujemo lužnate (kationske, pozitivno nabijene) boje koje uz pomoć tzv.
mordanta (pomoćna tvar, najčešće metalni ion, koja učvršćuje vezu molekule boje i tkiva te
pojačava intenzitet boje) vizualiziraju proteine, kisele (anionske, negativno nabijene) boje
koje sadržavaju sulfatne i karboksilne auksokrome, a služe bojanju citoplazmatske i
izvanstanične proteinske strukture te direktne boje, poput azo boja, koje ne trebaju mordant.
Prema kromoforima razlikujemo azo i dušične (engl. azotic) boje, ftalocijanine, karbonilne,
ksantenske, triarilmetane (trifenilmetane) i tiazinske boje te mnoge druge (Kiernan 2008).
1. Azo boje najčešće su korištene komercijalne boje čiji je kromofor azo -N=N-
skupina. Apsorbiraju pri nižim valnim duljima stoga ih ima najviše u žutim, crvenim i
narančastima bojama. Mogu biti hidrofilne poput indikatora metil oranža ili hidrofobne poput
Oil red Oa i različitih sudanskih bojila (Christie 2001).
Za nespecifično dokazivanje nekristaliziranih staničnih lipida koristi se lizokrom
sudansko crnilo B (engl. sudan black B, SBB) (Bayliss High 1981). Lizokromom se smatraju
boje topive u mastima (Stains Files 2012c). Zbog svoje kemijske strukture ne mogu ionizirati,
nego tvore hidrofobne veze otapajući se u mastima. Sudan black B (SBB, Slika 16.) spada u
diazo boje zbog dva azo kromofora (Christie 2001; Kumar i Kiernan 2010). Već se prije znalo
kako je ova tehnika osjetljiva na metode fiksacije te, ovisno o njima, može prikazivati samo
membranske fosfolipide ili lipidne kapljice (Baker 1956).
Slika 16. Sudan black B, azo boja zbog karakterističnih N=N veza. Preuzeto sa www.sigmaaldrich.com
Kompleksni glikokonjugati i polisaharidi mogu se dokazati tehnikama alcian blue i
periodic acid Schiff (PAS).

UVOD
16
2. Alcijansko modrilo (Alcian blue 8 GX, Ingrain Blue 1) se sastoji od kromofora
ftalocijanina (četiri izoindolska prstena spojenih dušikovim atomima) koordinirano vezanog
sa atomom bakra (Slika 17.) te četiri izotiouronska bočna lanca koji mu daju masivnost i
pozitivni naboj čineći ga lužnatom (kationskom) bojom (Christie 2001). Apsorbira svjetlost u
žuto-narančastom dijelu spektra, a reflektira valne duljine koje mi percipiramo kao zelene ili
plave. Topiv je u vodi i polarnim otapalima. Specifično boji heteropolisaharide poput
glikozaminoglikana u hrskavicama, mukopolisaharida na stanicama i sijaliniziranog
glikokaliksa intenzivnom plavom bojom. Glikozaminoglikani (GAG) sastoje su od
ponavljajućih disaharida od kojih je jedan N-acetilglukozamin (GlcNAc) ili N-
acetilgalaktozamin (GalNAc), a drugi uronska kiselina ili iduronska kiselina (Slika 18.) koja
polimeru može dati negativan naboj. Mucini spadaju u glikoproteine (vrsta glikokonjuganta),
oligosaharide kovalentno vezane za proteinsku srž. Mogu se izlučivati ili ostati na membrani
kao dio glikokaliksa. Sadržavaju veliki broj O-vezanih oligosaharidnih lanca povezanih za
protein glikozidnom vezom preko njegove -OH skupine serina ili treonina (Nelson i Cox
2008). Ovisno o sastavu oligosaharida mogu biti neutralni do blago kiseli. Neutralni mucini,
obično prisutni na površinskim i probavnim epitelnim stanicama, imaju visok udio neutralnih
šećera poput manoze i galaktoze. Blago kiseli mucini sadržavaju sijalinske kiseline koje su
monosaharidi od 9 C atoma sa karboksilnom grupom na prvom C atomu. Naziv "sijalinska
kiselina" obično se odnosi na N-acetilneuraminsku kiselinu (Bancroft i Gamble 2008).
Ovisno o pH alcijansko modrilo (AB) radi ionske veze se različitim grupama šećera. U
kiselinama dolazi do ionizacije amino grupa boje. Tkiva sadržavaju anione poput fosfata
nukleinskih kiselina, sulfata ugljikohidrata te karboksilata ugljikohidrata i proteina. Procesom
ionske zamjene pozitivno nabijeni alcian blue (AB) istiskuje male katione (poput H+) iz tkiva
(Kiernan 2008). U rasponima od pH 0,2 do pH 1 AB boji sulfatne polisharide. Pri pH 2,5 do 3
osim njih boji i fosfate DNA i RNA. Od pH 4 do 5 boji sve nabrojano te karboksilne grupe
ugljikohidrata (prisutne na sijalinskim i uronskim kiselinama) i kiselijih proteina. Iznad pH 5
može bojati neutralne proteine, ali pri pH 5,6 postaje nestabilan te se često dodaje magnezijev
klorid kao stabilizator (Green i Pastewka 1974; Kiernan 2008). Iako alcian blue pri
optimalnim uvjetima (pH, prisutnim ionima soli, temperaturi i vremenu reakcije) preferentno
boji glikozaminoglikane, nekad zna bojati nukleinske kiseline. Smatra se da je to zbog
interakcija negativnog naboja fosfatne okosnice i pozitivih bočnih lanaca alciana blue. Zbog
veličine molekule AB teško prodire do jezgrinog kompaktnog kromatina, ali proces je znatno
lakši ukoliko je DNA denaturirana (Kumar i Kiernan 2010).

UVOD
17
Često se kombinira sa metodom Schiffove baze i perjodne kiseline (PAS) upravo zato
što nema odbojavanja (engl. destaining) te ne mijenja boju ovisno o svojoj koncentraciji ili
dodatku drugih supstrata, što je svojstvo ortokromatskih boja (Bancroft i Gamble 2008).
Slika 17. Alcian blue 8GX kationska (lužnata) boja. Kloridni ioni prikazani kao protuioni. Na
ftalocijaninskoj jegri vide se četiri bočna lanca (R). Preuzeto sa www.sigmaaldrich.com
Slika 18. Lijevo N-acetilglukozamin (GlcNAc; GalNAc ima obrnuti položaj OH i H skupine na C-4
atomu), desno iduronska kiselina. Preuzeto iz Nelson i Cox (2008).
3. U skupinu trifenilmetana (triarilmetana) spadaju bazični fuksin (engl. fuchsin basic),
pararozanilin i metil violet (Keusch 2012). Karakteriziraju ih tri fenilna (arilna) prstena
vezana za centralni C atom. Pararozanilin i bazični fuksin imaju amino skupne vezane na
arilnim prstenima (Horobin i Kiernan 2002).
Bazični fuksin (pararozanilin hidroklorid) obično se prodaje kao mješavina rozanilina,
pararozanilina, novog fuksina i Magenta II u omjerima specifičnim o proizvođaču. Za
korištenje u Schiffovom reagensu mora imati visoki udio pararozanilina (Santa Cruz
Biotechnology 2012). Schiffov reagens, koji služi za dokazivanje aldehida, često se koristi u
kombinaciji sa perjodnom kiselinom u tzv. periodic acid Schiff (PAS) metodi. Prvenstveno
služi za dokazivanje glikogena, ali se pomoću nje može prikazati škrob, mucini, bazilarne
membrane i amiloidna tjelešaca. Glikogen je homopolisaharid sastavljen od glukoza

UVOD
18
povezanih (α1→4) glikozidnim vezama koji se prosječno grana na svakih 8 do 12 podjedinica
pomoću (α1→6) veza (Nelson i Cox 2008). Temelj ove metode je oksidacija hidroksilnih
grupa polisaharida pomoću perjodne kiseline (HIO4) u aldehidne grupe koje reagiraju sa
Schiffovim reagensom (Slika 19.). Bazični fuksin, koji stvara kovalentne veze između
reagensa i polisaharida, zaslužan je za crveno obojenje (Kumar i Kiernan 2010). U
Schiffovom reagensu nalaze se sulfitni (HSO3-) ioni koji vezanjem za centralni C atom
bazičnog fuksina njega čine bezbojnim. Vezanjem fuksin sulfita za aldehid nastaje niz
struktura od kojih je zadnja fuksinski kation (Slika 20.) zaslužan za crveno obojenje (Keusch
2012).
Slika 19. Oksidacija glukoze perjodnom kiselnom (HIO4) radi dvije aldehidne skupe i cijepa veze između
C2 i C3 atoma. Preuzeto iz Bancroft i Gamble (2008).
Slika 20. Bazični fuksin, odnosno rozanilin (lijevo) dodatkom sulfita prelazi u bezbojni sulfitni oblik
(sredina) koji vezanjem za aldehid u konačnici daje crveno-ljubičasti kation. Preuzeto iz Keutsch (2012).
Pararozanilin se može zasebno koristiti u Schiffovom reagensu. Radi na istom principu
opisanom kod bazičnog fuksina (Slika 21.; Kumar i Kiernan 2010).

UVOD
19
Slika 21. Pararozanilin (lijevo) dodatkom sumporaste kiseline (H2SO3) prelazi u Schiffov reagent (desno).
Preuzeto iz Kumar i Kiernan (2010).
Metil ili kristal violet (Slika 22.) osim što se često koristi kod bojanja Gram pozitivnih
bakterija, služi za prikazivanje DNA.
Slika 22. Metil violet. Preuzeto sa www.sigmaaldrich.com
Jedna od najčešće korištenih nespecifičnih boja u citologiji je May-Grünwald Giemsa
(MGG). Nastaje kombiniranjem otopina May-Grünwald i Giemse. May-Grünwald sastoji se
od ksantenske boje eozina Y i tiazinske boje metilenskog modrila (Reagena 2003). Giemsa je
mješavina azura B, metilenskog modrila i eozina Y. Ako se radi u lužnatijim uvjetima uzorci
će biti više plavi nego u kiselim uvjetima gdje će biti više crveniji (Kumar i Kiernan 2010;
Reagena 2003).
Kod gmazova i vodozemca za brojanje krvnih stanica često se koristi Natt-Herrickova
otopina. U njoj se nalaze tiazinska boja toluidinsko modrilo i spomenuti metil violet (Lisičić
2005; Vetlab 2013).

UVOD
20
4. Ksantenske boje većinom su flourescentne, a ovisno o naboju mogu biti kationske ili
anionske (Bancroft i Gamble 2008). Eozin Y (eozin G, tetrabromoflourescin; Slika 23.)
anionska je boja koja služi za vizualizaciju DNA dajući joj crveno obojenje. Ukoliko se
kombinira sa kationskim azurom B (vezanim na DNA), doći će do promjene boje DNA iz
plave u ljubičastu. Eozin Y također se veže sa proteinima bogatim lizinima i argininima,
poput hemoglobina u eritrocitima te eozinofilnih granula leukocita (Kumar i Kiernan 2010).
Slika 23. Eozin Y. Preuzeto iz Kumar i Kiernan (2010).
5. Tiazinske boje sadrže šesteročlani prsten sastavljen od 4 atoma ugljika, atoma
sumpora i atoma dušika. Ove većinom kationske boje daju plava i ljubičasta obojenja i topive
su u vodi (Kiernan 2008). Metakromatske su što znači da apsorbiraju svjetlost pri različitim
valnim duljama ovisno o njihovoj koncentraciji i okolini (Bancroft i Gamble 2008).
Metilensko modrilo (engl. methylene blue, Slika 24.a) kationska je boja koja se
specifično veže na DNA. Privuku je fosfatni anioni okosnice, a stabilizira se Van der
Waalsovim silama za regije bogate adeninom i timinom. Ukoliko dođe do oksidacije i
demetilacije metilensko modrilo prelazi u azur B (Kiernan 2008; Kumar i Kiernan 2010)
Azur B (Slika 24.b) također boji DNA isto kao metilensko modrilo. Opaženo je kako
formira dimere i radi hidrofobne interakcije sa purinima i pirimidinima (Kumar i Kiernan
2010).
Toluidinsko modrilo (engl. toluidine blue, Slika 24.c) također je kationska boja koja
boji na istom principu kao alcian blue. Služi za prikazivanje GAGa, s kojima radi ionske
interakcije, ili nukleinskih kiselina, s kojima je povezana Van der Waalsovim silama (Kiernan
2008; Kumar i Kiernan 2010).

UVOD
21
Slika 24. Metilensko modrilo (a) oksidacijom gubi metilnu skupinu i prelazi u azur B (b). Toluidinsko
modrilo (c). Preuzeto iz Kumar i Kiernan (2010).
Kod enzimskih bojanja utvrđuje se aktivnost određenih enzima leukocita poput alkalne
fosfataze i nespecifičnih esteraza. Princip je sljedeći: enzim hidrolizira dodani supstrat koji se
zatim veže sa bojom s kojom tvori obojani talog na mjestu aktivnosti enzima. Enzimske
aktivnosti nisu iste kod svih leukocita, a ponekad su prisutne i u trombocitima.
a) Alfa-naftil butirat esteraza (ANBE) nespecifičan je enzim koji hidrolizira alfa-naftil
butirat (Slika 25.a) u alfa-naftol. Nastali produkt vezan sa pararozanilonom (obogaćenim
nitratnim skupinama) daje crveno obojeni talog (Strober 2001). Aktivnost ovog enzima
pokazuju monociti.
b) Pomoću aktivnost alfa-naftil acetat esterazi (ANAE) mogu se razlikovati stanice
monocitnog i neutrofilnog porijekla. Esteraze hidroliziraju estersku vezi sintetskog supstrata
α-naftil acetata pritom tvoreći α-naftol (Slika 25.b i c) koji se kovalentno veže sa fast blue RR
soli čime radi netopivi tamno plavo obojani talog (Chronolab 2012c; Kiernan 2008).
Slika 25. Esteraze cijepaju α-naftil butirat (a) i α-naftil acetat (b) na α-naftol (c). Obrađeno sa
www.sigmaaldrich.com.

UVOD
22
c) Supstrat alkalne fosfataze (ALP) je naftol AS-BI fosfat (Slika 26.) koji se hidrolizira
na fosfat i aril naftolamid. Slično ANAEu, aril naftolamid tvori kovalentne veze sa fast red ili
fast blue soli čime nastaje crveno, odnosno plavo obojani talog. Aktivnost ALP pronađena je
u ljudskim neutrofilima (Chronolab 2012b; Lowe 1998b).
Slika 26. Naftol AS-BI fosfat. Preuzeto sa www.sigmaaldrich.com
6. Dušične boje stabilizirane su diazo i tetraazo soli, obično pripremljenje kao dvostruke
soli cinkovog klorida. Diazo soli (ArN2+) nastaju dodavanjem dušićaste kiseline (HNO2) na
aromatske amine (ArNH2). Izrazito su nestabilne strukture koje za stabiliziaciju trebaju velike
ione suprotnog naboja poput metalnih soli tetraflouroborata, BF4- i tetraklorozinkata, ZnCl4
2-
(Christie 2001).
Fast blue RR salt (Slika 27.) i fast red TR salt spadaju u diazo soli koje se koriste za
prikazivanje aktivnosti hidrolitičkih enzima. Sa enzimskim supstratom rade kovalentne veze
tvoreći netopivi obojani talog (Chronolab 2012b).
Slika 27. Fast blue RR sol. Preuzeto sa www.sigmaaldrich.com.
Uz navedene tehnike često se koriste kontrastne boje za vizualizaciju jezgri. Obično se
uz sudan black B, alkalnu fosfatazu i alcian blue (koji boje crno i plavo) koristi boja nuclear

UVOD
23
fast red (za crveno obojenje), a uz periodic acid Schiff metodu i reakciju alfa-naftil butirat
esteraze (koje boje crveno) hematoksilin (za plavo obojenje jezgri).
7. Karbonilne boje, također često korištene u komercijalne svrhe, dobile su ime po
karbonilnoj C=O grupi. Obično imaju dvije ili više karbonilnih skupina povezanih
aromatskim prstenima.
Najpoznatija grupa karbonilnih boja su antrakvinoni u koje spada nuclear fast red
(NFR). Njihova se struktura sastoji od tri spojena šesteročlana prstena, dva aromatska na
rubovima i kvinona u sredini. Kvinon je struktua od dvije ketonske grupe na sešteročlanom
prstenu sa dvije dvostruke veze (Slika 28.a i b). NFR (Slika 28.d) koristi se za vizualizaciju
jezgre dajući joj crveno obojenje (Christie 2001). Anionska je boja kojoj je potreban mordant,
obično aluminijeva sol. Smatra se kako u interakciji nukleinskih kiselina sa kompleksom
aluminij-NFR sudjeluju ionske i ne-ionske veze (Kiernan 2008). Auksokromi su amino i
hidroksi grupe koje služe kao elektron donori i omogućuju batokromatski pomak, dok je
karbonilna grupa elektron akceptor. Radi se o brzom reverzibilnom redoks sustavu (Slika 29.)
gdje se benzokvinon reducira u hidrokvinon uz prijenos dva vodika (Christie 2001).
Slika 28. Kvinoni, orto- (a) i para-benzokvinon (b). Osnovna struktura antrakvinona (c). Nuclear fast red,
kvinonski dio nalazi se u sredini. Preuzeto iz Christie (2001) i sa www.sigmaaldrich.com.
Slika 29. Kvinon- hidrokvinon redoks sustav, karbonilna grupa kao elektron akceptor. Hidrokvinon skroz
desno. Preuzeto iz Christie (2001).

UVOD
24
8. Hematoksilin monohidrat (Natural Black 1) spada u prirodne boje. Ekstrakt je
grmovitog stabla Haematoxylum campechianum (Bancroft i Gamble 2008; Stains File 2012b).
Sam za sebe nije boja već se mora oksidirati u boju hematein (Slika 30.). Prirodna oksidacija
je dugotrajna i događa se na svjetlu i zraku. Kemijska oksidacija, koja je trenutna i relativno
kratkotrajna, koristi natrijev jodat, NaIO3 (kod Mayerovog, Gillovog i Carazzijevog
hematoksilina) ili živin oksid (Harrisov hematoksilin). Hematein je anionska boja sa slabim
afinitetom za tkiva stoga treba kationski mordant. Najbolje su aluminijeve (Gillov, Mayerov i
Carazzijev hematoksilin), željezove i volframove soli koje kompleksu daju neto pozitivan
naboj (Bancroft i Gamble 2008; IHC World 2011a i 2011b; Stains File 2012a). Ovisno o
mordantu boja se veže za različita tkiva i utječe na krajnje obojenje. Za vezanje na kromatin,
preciznije na nukleoproteinski lizin, koriste se aluminijeve soli. Smatra se kako u toj
interakciji sudjeluju ionske i ne-ionske veze (Kiernan 2008). Pri kiselim pH kompleks
hemateina i aluminija jezgri daje crveno obojenje, ali ispiranjem u slaboj bazičnoj otopini
prelazi u tamno plavu boju. Vodovodna voda je dovoljna, ali nekad treba litijev karbonat za
taj proces zvan "plavljenje" (eng bluing, Bancroft i Gamble 2008; Kiernan 2008) .
Slika 30. Hematoksilin lijevo, hematin desno. Zaokruženo je mjesto oksidacije. Preuzeto sa
www.sigmaaldrich.com
Većini navedenih metoda prethodi fiksacija. Fiksacija prvenstveno služi za očuvanje
izgleda i sastava tkiva, što se postiže narušavanjem aktivnost staničnih enzima (poglavito
autolitičkih), uklanjanjem vode i zamjenom topivih dijelova stanice netopivima te stvaranjem
kovalentnih veza (engl. cross-linking). Fiksirane stanice također su otporne na vanjske faktore
poput patogena i atmosferiliija. Najčešći fiksativi su formaldehid i alkoholi.
Otopljeni formaldehid zove se formalin i stvara različite interakcije sa
makromolekulama. To mogu biti kovalentne veze sa bočnim lancima proteina, modifikacije
nukleotida, stabilizacije nukleoproteinskih veza te vezanje sa nezasićenim lipidima. Međutim,
ne reagira sa ugljikohidratima (Bancroft i Gamble 2008).

UVOD
25
Alkoholi dehidriraju stanice (uklanjaju vodu iz nje), čime narušavaju hidrofobne i
vodikove veze što uzrokuje promjene strukture proteina koji zatim koaguliraju. Time se
učvrste strukturni proteini (netopivi citoskeletni i lipoproteinski), ali se naruši citoplazmatska
struktura gdje proteini međusobno formiraju nakupine (Bancroft i Gamble 2008).
1.3. Citokemijske karakteristike krvnih stanica u kralježnjaka
Diferencijalna citokemijska analiza krvnih stanica otkriva specifične biokemijske
karakteristike pojedinih tipova stanica, posebice među leukocitima. Ovom se analizom mogu
otkriti pojedini podtipovi neutrofila, bazofila, eozinofila i limfocita ovisno o starosti ili stadiju
aktivacije te diferencijalnu pripadnost ovisno o vrsti životinje
U nastavku teksta navedeni su rezultati bojanja alcijanskim modrilom (AB), metodom
Schiffove baze i perjodne kiseline (PAS), sudanskim crnilom B (SBB), alkalnom fosfatazom
(ALP), nespecifičnim esterazama alfa-naftil acetat esterazom (ANAE) i alfa-naftil butirat
esterazom (ANBE) kod krvnih stanica različitih skupina kralježnjaka.
Eritrociti svih kralježnjaka u načelu ne daju pozitivne rezultate na opisane tehnike.
Trombociti kralježnjaka se uglavnom ne boje specifično citokemijskim metodama. Kod
sisavaca zajedno sa megakariocitima pokazuju jaku aktivnost nespecifičnih esteraza (ANAE)
(Chronolab 2012c). Kod X. laevis bojaju se PASom i pokazuju aktivnost nespecifičnih
esteraza (Hadji-Azimi i sur.1987).
Bazofili sisavaca boje se ABom. Ponekad pokazuju aktivnost ALPa. Negativni su na
SBB (Weiss i Wardrop 2010). Kod X. laevis citoplazmatske granule bazofila boje se crveno
metodom PAS (Hadji-Azimi i sur. 1987).
Eozinofili sisavaca pokazuju jaku aktivnost alfa-naftil acetat esteraza (ANAE), alfa-
naftil butirat esteraza (ANBE), alkalne fosfataze (ALP) i bojaju se SBBom (iznimka su
mačke; Weiss i Wardrop 2010).
Limfociti sisavaca rijetko pokazuju reaktivnost na citokemijske metode, npr. na
aktivnost ANAE (Sigma-Aldrich 2006). Aktivnost ALP zabilježena je u B stanicama. NK
stanice pokazuju slabu, fokalnu aktivnost nespecifičnih esteraza i boje se metodom PAS
(Weiss i Wardrop 2010).
Monociti sisavaca pokazuju aktivnost na nespecifične esteraze (ANBE i ANAE) koja se
pojačava sa njihovim sazrijevanjem u makrofage. SBB bojanje daje slabe rezultate (Weiss i
Wardrop 2010). Monociti X. laevis također su pozitivni na aktivnost nespecifičnih esteraza
(Hadji-Azimi i sur. 1987).
Neutrofili sisavaca pokazuju pozitivne rezultate na SBB, ALP i ANBE (Weiss i
Wardrop 2010). Kod X. laevis boje se PAS tehnikom (Hadji-Azimi i sur. 1987).

CILJ ISTRAŽIVANJA
26
2. CILJ ISTRAŽIVANJA
Ustanoviti ukupni broj eritrocita i leukocita, udjele pojedinih tipova leukocita te
vrijednost hematokrita zelene žabe Pelophylax ridibundus.
Utvrditi morfološke karakteristike (izgled i dimenzije) eritrocita, trombocita i različitih
tipova leukocita iz krvnih razmaza.
Diferencijalnim citokemijskim tehnikama utvrditi boje li se pojedini leukociti u
istraživane vrste jednako specifično kao i u ostalih kralježnjaka te postoje li razlike u
biokemijskim osobinama pojedinih stanica. Također, cilj je utvrditi mogu li se temeljem
diferencijalnih bojenja razlikovati pojedini podtipovi leukocita.
Utvrditi referentne podatke o karakteristikama krvnih stanica s obzirom da se rod
Pelophylax koristi u fiziološkim i genetičkim istraživanjima.

MATERIJALI I METODE
27
3. MATERIJALI I METODE
3.1. Uzorci
U ovom radu analizirani su krvni razmazi svježe krvi odraslih zelenih žaba (Pelophylax
ridibundus), oba spola, težine u rasponu od 55 do 80 g. Sve su žabe dekapitirane i špricama
(promjera 0,17mm) im je izvađena krv iz srca. Kroz šprice bio je provučen heparin kako bi se
spriječilo grušanje. Za bojanje May-Grünwald Giemsom (MGG) i citokemijske analize
odmah su napravljeni krvni razmazi, tri do šest stakalca po žabi. Ostavljeni su preko noći da
se osuše. Za određivanje ukupnog broja krvnih stanica krv se odmah razrijedila Natt-
Herrickovom otopinom.
Uzorci sedam žaba žrtvovanih u listopadu 2009. godine koristili su se za kompletnu
krvnu sliku, odnosno bojanje May-Grünwald Giemsom i određivanje ukupnog broja
leukocita. Za citokemijsko bojanje koristili su se uzorci šest žaba žrtvovanih u listopadu 2012.
godine. Svaka se citokemijska tehnika radila minimalno u triplikatu.
3.2. Određivanje ukupnog broja eritrocita i leukocita
Za pripremu Natt- Herrickove (N-H) otopine u 1 l destilirane vode potrebno je otopiti:
3,88 g natrijevog klorida, NaCl (Herbos, Hrvatska)
2,50 g natrijevog sulfata, Na2SO4 (Herbos, Hrvatska)
2,91 g natrijeva-hidrogen fosfat dodekahidrata, Na2HPO4 x 12 H2O (Kemika, Hrvatska)
0,25 g kalijevog dihidrogen fosfata, KH2PO4 (Kemika, Hrvatska)
7,50 ml formalina 37 %-tni (Alkaloid, Makedonija)
0,10 g metil violeta (Herbos, Hrvatska)
0,10 ml toluidinskog modrila, 1% (Herbos, Hrvatska)
Za određivanje ukupnog broja eritrocita nekoagulirana krv razrijedila se N-H otopinom
u omjeru 1:200 u eritrocitnim melanžerim i miješala na rotacijskoj miješalici (Ika) minimalno
15 minuta. Nakon toga kap iz melanžera se kapnula ispod pokrovnice u hemocitometar
(Bürker-Türk komoricu) i nakon 3 minute stajanja započelo se brojanje u 16 polja pod
svjetlosnim mikroskopom (400 x povećanje). Iz dobivenog broja izračunao se broj eritrocita
po ml krvi prema formuli: E = N x 800000 x 103
E = broj eritrocita / ml krvi
N = srednji broj eritrocita po kvadratiću

MATERIJALI I METODE
28
Za brojanje leukocita u leukocitni melanžer se navukla svježa krv do oznake 0,5 i N-H
otopina do oznake 11. Time je krv razrijeđena 20 puta. Na rotacijskoj miješalici uzorak se 3
minute miješao. Jedna kap krvi dodana je na hemocitometar sa 16 polja gdje su se pod
povećanjem 400x brojali leukociti. Broj ukupnih leukocita po ml krvi računao se prema
formuli: L = N x 5000 x 103
L = broj leukocita / ml krvi
N = srednji broj leukocita po kvadratiću
3.3. Određivanje vrijednosti hematokrita
Svježa krv navukla se u heparinizirane kapilare zataljene na jednom kraju, tri kapilare
po svakoj životinji. Centrifugirale su se 5 minuta na 5000 okretaja / min. Nakon toga se na
odgovarajućoj skali očitala vrijednost hematokrita. Prosječna vrijednost hematokrita
izračunala se iz prosjeka grupe koji je izračunat iz prosjeka životinja.
3.4. Bojanja May-Grünwald Giemsom (MGG)
Dobro osušeni krvni razmazi držali su se u May-Grünwald (J.T. Baker, Nizozemska)
otopini 7 minuta, nakon čega su isprani destiliranom vodom (dH2O) i držani u otopini Giemse
12 minuta. Otopina Giemse je napravljena od dva dijela vode i jednog dijela Giemse (J.T.
Baker, Nizozemska). Slijedilo je ispiranje destiliranom vodom i sušenje preko noći.
Takvi su preparati spremni za mikroskopiranje.
3.4.1. Morfološke karakteristike krvnih stanica
Mikroskopirali su se razmazi tretirani MGGom. Slike su dobivene pomoću
Olympusovog mikroskopa BX51 sa Olympusovom kamerom DP70 te pripadajućeg programa
DP Controller. Korištena povećanja bila su 40x bez imerzije te 60x i 100x sa imerzijom.
3.4.2. Računalna obrada podataka
Za izračunavanje udjela pojedinih leukocita iz MGG razmaza ručno su se brojali
bazofili, eozinofili, limfociti, monociti i neutrofili na 100 stanica.
Digitalna mjerenja uslikanih stanica napravljena su pomoću Carl Zeissovog programa
Axio Vision verzija 4.8.2. Statistička obrada podataka učinjena je u Microsoft Excel 2007
programu.
3.5. Citokemijske analize
Radile su se na osušenim krvnim razmazima, jedna tehnika po razmazu (iznimno
kombinacija alciana blue i periodic acid Schiffa). Razmazi su se nakapavali citokemijskim
bojama umjesto uranjanja u posude za bojanje histoloških preparata (engl. coplin jar). Za

MATERIJALI I METODE
29
svako stakalce bilo je potrebno oko 2,5 ml citokemijske boje. Otopine određenog pH
pripremale su se pomoću magnetske miješalici (Yellow line) i pH metra (Schott Instrument).
3.5.1. Alcijansko modrilo (AB)
Prema protokolu (Lowe 1998a) napravljene su otopine alcijanskog modrila (AB) sa
različitim pH.
1. Za pripremu otopine AB pH 3,1 u tikvicu (25ml) trebalo je:
250 mg alciana blue (AppliChem, Njemačka),
0,125 ml 0,5 % ledene octene kiseline (Kemika, Hrvatska)
dH2O.
AB se otopio u octenoj kiselini i dolila se dH2O do 25 ml.
2. Za pripremu ABa pH 2,5 u tikvicu (25ml) trebalo je:
250 mg ABa
0,75 ml ledene octene kiseline
dH2O
AB se otopio u octenoj kiselini i dolila se dH2O do 25 ml.
3. Za pripremu ABa pH 1 trebalo je:
25 ml klorovodične kiseline, HCl (Riedel- de Haën, Njemačka)
250 mg ABa
AB se otopio u HClu.
4. Za pripremu AB pH 0,2 u tikvicu (25ml) se doda
250 mg ABa
2,5 ml 10 %-tne sumporne kiseline, H2SO4 (Carlos Erba Reagents, Italija)
AB se otopio u H2SO4 i dolila se dH2O do 25 ml. Sve pripremljene otopine drže se
dobro zatvorene na 4°C.
Za vizualizaciju jezgri korištena je 0,1 %-tna otopina nuclear fast reda (NFR) prema
IHC World protokolu (2011c), ali bez dodatka timola (konzervans). Za prekrivanje 10
stakalca pripremljeno je 25 ml NFRa iz:
25 ml dH2O
25 mg nuclear fast red (AppliChem, Njemačka)
2,5 g aluminijevog-sulfat oktadekahidrata, Al2(SO4)3 x 18 H2O (Gram-Mol, Hrvatska)
NFR i aluminijev-sulfat otopili su se u vodi. Dobivena otopina kuhala se 5 minuta i ohlađena
filtrirala. Otopina NFRa čuvala se na 4°C.

MATERIJALI I METODE
30
Na suhe razmaze nakapala se destilirana voda i držala dvije minute. Nakon što se voda
istresla, uzorci su se nakapali različitim pH vrijednostima alciana blue (1, 2, 3 i 4). Uzorci su
se inkubirali 3, 4 i 5 minuta za sve pH vrijednosti. Slijedilo je ispiranje destiliranom vodom i
sušenje preko noći. Na suhe, obojane razmaze nakapala se otopina NFRa i nakon tri minute
isprala destiliranom vodom. Preparate se ostavilo sušiti preko noći.
3.5.2. Alfa-naftil acetat esteraza (ANAE)
Korišten je prilagođeni protokol (Sigma Aldrich, 2006) gdje se umjesto fast blue RR
salt kapsula (nedefinirane mase) koristio fast blue RR salt prah (Santa Cruz Biotechnology,
Amerika).
Fiskativ citrat-aceton-metanol se uvijek svježe pripremao od:
3,6 ml razrijeđene citratne otopine od tri-natrij-citrat-2-hidrata (Kemika, Hrvatska)
5,4 ml acetona (T.T.T., Hrvatska)
1 ml metanola (T.T.T., Hrvatska)
Za razrijeđenu citratnu otopinu napravila se prezasićena otopina natrijevog citrata
dodavanjem tri-natrij-citrat-2-hidrata u 10 ml dH2O sve dok se mogao otopiti. Otopina se
filtrirala, izmjeren joj je obujam i ulivena je 9 puta veća količina dH2O kako bi se dobila
razrijeđena otopina natrijevog citrata sa pH 4,5. Čuvao se na sobnoj temperaturi dobro
zatvoreno.
Prije pokusa trebalo je napraviti 50 ml razrijeđenog Trizma pufer. Prezasićena otopina
dobivena je otapanjem otprilike 1 žlice Trizma Base-a (Sigma, Njemačka) u 5 ml dH2O sve
dok se nije moglo više otopiti. Otopina je profiltrirana u tikvicu (50ml), pH je namješten na
7,6 i dodano je dH2O do 50 ml. Čuva se na sobnoj temperaturi dobro zatvoreno.
Uzorci su se fiksirali pripremljenim fiksativom minutu, isprali destiliranom vodom i
sušili 20-30 minuta. U međuvremenu se pripremila svježa otopina α-naftil acetata (ANA):
2 mg ANA (AppliChem, Njemačka)
200 μl etilen glikol monometil etera (EGME, Mreck, Njemačka).
ANA se otopio u EGMEu i morao odmah iskoristiti.
Za dva uzorka otopljeno je 5 mg fast blue RR soli u 5 ml Trizma pufera. Unutra se ulilo 200
μl otopljenog ANAa, promiješalo i odmah nakapalo na osušene fiksirane uzorke. Uzorci su se
zaštitili od svjetla i inkubirali 30 minuta. Nakon toga su se isprali destiliranom vodom i
ostavili sušiti preko noći. Nije bilo kontrasnog obojenja.

MATERIJALI I METODE
31
3.5.3. Alfa-naftil butirat esteraza (ANBE)
Za pripremu 25 ml puferiziranog formalin-aceton fiksativa pH 6,6 prema Stroberovom
protokolu (2001) trebalo je:
9,375 ml dH2O
6,25 mg bezvodnog natrijevog-hidrogen fosfata, Na2HPO4 (Kemika, Hrvatska)
31,25 mg kalijevog dihidrogen fosfata, KH2PO4
14,06 ml acetona
7,81 ml formalina
U vodi se otopio natrijev hidrogen fosfat i kalijev dihidrogen fosfat. Na kraju se
umiješao aceton i formalin te je pH namješten na 6,6. Fiksativ se čuvao dobro zatvoren na
4°C. Ova je količina bila dostatna za 10 uzoraka.
Prema Stroberu (2001) unaprijed je pripremljeno 100 ml fosfatnog pufera pri pH 6,3 (I),
2,5 ml pararozanilin-HCla (II) i 2,5 ml 4%-tne otopine natrijevog nitrata (III).
(I) Za pripremu 100 ml fosfatnog pufera pH 6,3 trebalo je:
100 ml dH2O
236,75 mg Na2HPO4
1,53 g KH2PO4
Obje soli su se otopile u vodi i pH im je namješten na 6,3.
(II) Za pripemu 2,5 ml pararozanilina-HCl potrebno je:
2 ml dH2O
0,5 ml koncentrirane HCl
0,1 g pararozanilin basea (Sigma, Njemačka)
HCl se umiješala u vodu nakon čega se dodao i otopio pararozanilin.
(III) Za 4 %-tnu otopine natrijevog nitrata(NaNO2) trebalo je:
2,5 ml dH2O
100 mg NaNO2 (Gram-Mol, Hrvatska)
NaNO2 se otopio u vodi.
Uzorci su bili fiksirani ohlađenim fiskativom 30 sekundi. Bez ispiranja vodom
ostavljeni su sušiti se 20ak minuta. Za pripremu inkubacijske otopine trebalo je svježe
umiješati:
a) 25 μl pararozanilina obogaćenog nitratnim skupinama (engl. hexazotied
pararosanilin) dobivenog miješanjem:

MATERIJALI I METODE
32
12,5 μl 4 %-tnog NaNO2 (III)
12,5 μl pararozanilin-HCl (II)
b) 250 μl otopine α-naftil butirata (ANB) dobivene otapanjem:
250 μl EGME
5 μl ANB
c) 4,75 ml pripremljenog fosfatnog pufera pH 6,3
Prvo su se umiješale otopine ANB i pararozanilina obogaćenog nitratnim skupinama,
zatim je dodan fosfatni pufer, otopina se filtrirala i nakapala na dva uzorka. Nakon 45 minuta
uzorci su se isprali destiliranom vodom i ostavili sušiti.
Nije napravljeno kontrasno obojenje.
3.5.4. Alkalna fosfataza (ALP)
Ovo je prilagođeni protokol (Lowe 1998b) gdje se fast blue RR salt koristio umjesto fast
red TR salt te nuclear fast red (NFR) umjesto hematoksilina.
Za pripremu 100 ml puferske otopine 0,2M Tris HCl u tikvici (150ml) trebalo je:
2,42 g Trisma base (Sigma Aldrich, Njemačka)
0,1M HCl
dH2O
U 40 ml dH2O otopljena je Trisma te je dodavan HCl dok nije postignut pH 8,3. Na
kraju se ulila dH2O do 100 ml. Otopina Tris-HCl može stajati na sobnoj temperaturi, a služi
za pripremu naftol AS-BI matične otopine.
Za pripremu 100 ml naftol AS-BI matične otopine trebalo je:
5 mg naftol AS-BI fosfata (AppliChem, Njemačka)
2 ml N,N-dimetilformamida (Gram-Mol, Hrvatska)
2 ml dH2O
natrijevog karbonata, Na2CO3 (Kemika, Hrvatska)
36 ml pripremljenog Tris-HCl pufera
60 ml dH2O
U N,N-dimetilformamidu otopio se naftol AS-BI fosfat, dodana je dH2O i natrijevim
karbonatom pH je podešen na 8,0. Zatim je dodan Tris-HCl pufer i dH2O.
Inkubirajuća otopina (za 10 uzoraka) svježe je napravljena od 25 ml matične otopine
naftol AS-BI fosfata i 25 mg fast blue RR soli, promješana, filtrirana i odmah nanešena na

MATERIJALI I METODE
33
suhe, nefiksirane razmaze. Vremena inkubacije su 8, 10, 13 i 15 minuta. Za vizualizaciju
jezgri koristio se NFR na isti način kao kod alcian blue tehnike.
3.5.5. Metoda Schiffove baze i perjodne kiseline (PAS)
Za pripremu Schiffova reagensa (Lowe 1998c) bilo je potrebno:
200 ml kipuće destilirane vode
1 g bazičnog fuksina (AppliChem, Njemačka).
2 g kalijevog disulfita (AppliChem, Njemačka)
2 ml koncentrirane klorovodične kiseline (HCl)
Žlica (cca 1 g) aktivnog ugljena (Kemika, Hrvatska).
U kipućoj vodi otopio se bazični fuksin. Kada se otopina ohladila na 50 °C umiješao se
kalijev disulfit i ohladilo na sobnu temperaturu. Zatim se ulila klorovodična kiselina i
ostavljeno je da stoji u mraku preko noći. Idući dan dodan je aktivni ugljen, smjesa se
protresla i filtrirala. Dobivena otopina prozirne je boje i čuvala se na 4 °C dobro zatvorena.
Za provjeru kvalitete Schiffova reagensa u 10 ml 35 % formalina (Alkaloid,
Makedonija) doda se par kapi reagensa. Ovo nije potrebno raditi, ali je korisno.
Za vizualizaciju jezgri koristio se Gillov hematoksilin (IHC World 2011a). Za pripremu
100 ml Gillovog hematoksilina potrebno je:
25 ml etilen glikola (AppliChem, Njemačka)
50 ml dH2O
200 mg hematoksilin monohidrata (AppliChem, Njemačka)
20 mg kalijevog jodida, KI (Kemika, Hrvatska),
2 g aluminijevog-sulfat oktadekahidrata, Al2(SO4)3 x18H2O (Gram-mol, Hrvatska)
2 ml ledene octene kiseline
dH2O
Etilen glikol se dodao u dH2O te se dodao hematoksilin, KI, alumijiev sulfat i ledena
ocetna kiselina. Natočila se dH2O do 100 ml. Otopina se miješala sat vremena na magnetskoj
miješalici. Gillov hematoksilin čuvao se dobro zatvoren na sobnoj temperaturi zaštićen od
svjetla.
Krvni razmazi nisu bili fiksirani. Na njih se nakapala dH2O i držala dvije minute. Višak
vode se izlio i odmah se tretiralo 10 minuta sa 1 %-tnom perjodnom kiselinom (Merck,
Njemačka). Slijedilo je ispranje vodom i tretman Schiffovim reagensom u vremenskim
intervalima 10, 13, 15 i 20 minuta. Na kraju su se razmazi isprali destiliranom vodom i

MATERIJALI I METODE
34
ostavili sušiti. Za vizualizaciju jezgri uzorci su nakapani Gillovim hematoksilinom. Nakon 3
minute isprani su dH2Oom i tretirani 1%-tnom otopinom litijevog karbonata 2 minute kako bi
se obojili plavo.
3.5.6. PAS i AB kombinacija
Prvi dio identičan je tretmanu ABom: nakapavanje nefiksiranih uzorka destiliranom
vodom u trajanju od dvije minute, izlijevanje viška vode, tretman ABom (dužina tretmana
ovisi o pHu: 4 min za pH 0,2 i 1, 3 min za pH 2,5 i 3,1) i ispiranje dH2Oom. Nije potrebno
sušenje.
Tretman PASom odmah počine 10 minutnom inkubacijom 1 %-tne perjodne kiseline.
Slijedi ispiranje destiliranom vodom i tretman Schiffovim reagensom (iste minutaže opisanom
u PAS metodi). Na kraju su uzorci isprani vodom.
Za vizualizaciju se koristio Gillov hematoksilin opisan kod PAS metode.
3.5.7. Sudansko crnilo B (SBB)
Prilagođeni protokol (WebPath 2012) koristi više metoda fiksacija te otopinu sudanskog
crnila B (SBB) čuva dobro zatvorenu na sobnoj temperaturi umjesto 60 °C.
Za pripremu otopine SBB potrebno je:
100 ml etilen glikola (AppliChem, Njemačka)
0,7 g SBBa (Sigma-Aldrich, Njemačka)
Dodani SBB u etilen glikolu se miješao 4 min u kupelj na 95 °C. Otopina se vruća
filtrirala i čim se ohladila na sobnu temperaturu, filtrirala se drugi put.
Koristili su se nefiksirani uzorci, uzorci fiksirani 10 %-tnim formalinom pH 7,4
(Alkaloid, Makedonija) dvije minute, uzorci fiksirani dvije minute mješavinom 96 %-tnog
etanola (Badel, Hrvatska) i 5 %-tne ocetene kiseline u omjerima 3:1, uzorci fiksirani
ohlađenim formalinom i acetonom 30 sekundi (kao u ANBE) i uzorci fiksirani jednu minutu
puferiziranom otopinom citrata, acetona i metanola (kao u ANAE). Za negativnu kontrolu
uzorci su tretirani acetonom (T.T.T., Hrvatska) tri minute kako bi se degradirale masti.
Ocijeđeni preparati uronili su se u etilen glikol (EG) pet miunta, kratko su osušeni i
ponovno vraćeni na pet minuta u EG. Nakon toga uzorci su ocijeđeni, nakapani otopinom
SBBa i inkubirani 5, 6, 7 i 8 minuta. Zatim su se istresli od viška boje i nakapali 85 %-tnim
etilen glikolom na pet minuta. Zadnji korak bilo je ispranje destiliranom vodom i sušenje par
sati. Za vizualizaciju jezrgi korištena je otopina nuclear fast red (NFR) opisana kod alcian
blue metode.

MATERIJALI I METODE
35
3.5.8. Morfološke karakteristike krvnih stanica
Mikroskopirali su se razmazi tretirani svim citokemijskim metodama. Slike su dobivene
pomoću Olympusovog mikroskopa BX51 sa Olympusovom kamerom DP70 te pripadajućeg
programa DP Controller. Koristila su se povećanja od 40x bez imerzije te 60x i 100x sa
imerzijom.

REZULTATI
36
4. REZULTATI
4.1. Određivanje ukupnog broja eritrocita i leukocita
Nakon potrebnih izračuna dobivene su slijedeće vrijednosti za eritrocite i leukocite
(Tablica 1.).
Tablica 1. Ukupni broj leukocita i eritrocita
Srednja vrijednost Standardna devijacija Median Min Max
Eritrociti x 106/ml 0,43 0,13 0,45 0,11 0,68
Leukociti x 103/ml 7,50 6,89 7,19 6,89 7,50
Ručnim brojanjem stanica najviše je izbrojano eritrocita. Od leukocita najčešći su bili
limfociti (veliki i mali), zatim neutrofili, eozinofili i monociti. Bazofila nije pronađeno na 100
stanica, ali se strpljenjem mogu pronaći na uzorcima. Ukupan broj pojedinih tipova leukocita
prikazan je u Tablici 7. Grafički prikaz udjela pojedinih tipova leukocita nalazi se na Slici 33.
4.2. Određivanje vrijednosti hematokrita
Nakon statističke obrade dobivene su slijedeće vrijednosti za hematokrit (Tablica 2.).
Tablica 2. Vrijednosti hematokrita
Srednja
vrijednost
Standardna
devijacija Median Min Max
0,43 0,13 0,45 0,11 0,68
4.3. Diferencijalna analiza pomoću bojanja May-Grünwald Giemsom
4.3.1 Eritrociti
Svi eritrociti, neovisno o starosti imaju ljubičasto obojanu jezgru i blijedu citoplazmu
(Slika 31.). Kod zelene žabe pronađeno je više stadija zrelosti eritrocita. Nezreli eritociti
okruglog su oblika i okrugle jezgre, intenzivnije obojane citoplazme. Prijelazni oblik iz
nezrelog u zreli ima okruglu jezgru, ali izduženiju stanicu nego nezreli. Zreli eritrociti
ovalnog su oblika i ovalne jezgre, smještene u centru. Većih su dimenzija nego nezreli i

REZULTATI
37
jezgra im zauzima manji dio citoplazme. Stari eritrociti imaju bijelu citoplazmu te ovalnu
ili okruglu jezgru svjetlije obojanu od zrelih i nezrelih eritrocita. U usporedbi sa zrelim i
nezrelim eritrocitima najvećih su dimenzija (visina i širina), uključujući i jezgru (Tablice 3.
i 4.).
Broj eritrocita po ml krvi prikazan je u Tablici 7.
Tablica 3. Prosječne dimenzije stanica nezrelih, zrelih i starih eritrocita izražene u μm sa
standardnom devijacijom te najmanjim i najvećim vrijednostima.
Širina Min Max Duljina Min Max
Nezreli 13,547 ± 1,264 10,41 15,14 17,446 ± 1,605 10,41 19,34
Zreli 15,114 ± 0,909 13,44 17,06 22,685 ± 1,595 19,47 25,68
Stari 18,489 ± 1,674 15,27 20,87 24,94 ± 2,169 21,83 28,36
Tablica 4. Prosječne dimenzije jezgri nezrelih, zrelih i starih eritrocita izražene u μm sa
standardnom devijacijom te najmanjim i najvećim vrijednostima.
Širina Min Max Duljina Min Max
Nezreli 6,192 ± 0,749 5,26 7,68 8,264 ± 1,051 7,36 10,97
Zreli 4,752 ± 0,577 3,65 6,29 8,699 ± 0,819 6,54 10,24
Stari 10,213 ± 3,454 5,7 16,89 13,272 ± 3,066 9,48 17,26
4.3.2. Trombociti
Ovisno o starosti mijenjaju oblik (Slika 31.). Nezreli trombociti sferičnog su oblika,
tamno ljubičaste okrugle jezgre sa vrlo malo rozo obojane citoplazme. Zreli trombociti su
ovalnog oblika, imaju ovalne tamno ljubičaste jezgre centralno smještene. Znatno su veći od
nezrelih (Tablice 5. i 6.). Ponekad se na razmazima trombociti mogu naći grupirani.
Tablica 5. Prosječne dimenzije stanica nezrelih, zrelih i starih trombocita izražene u μm sa
standardnom devijacijom te najmanjim i najvećim vrijednostima.
Širina Min Max Duljina Min Max
Nezreli 8,428 ± 0,657 6,33 9,69 10,591 ± 1,034 8,78 12,49
Zreli 7,564 ± 0,925 5,12 17,41 12,863 ± 1,523 10,41 17,41

REZULTATI
38
Tablica 6. Prosječne dimenzije jezgri nezrelih, zrelih i starih trombocita izražene u μm sa
standardnom devijacijom te najmanjim i najvećim vrijednostima.
Širina Min Max Duljina Min Max
Nezreli 6,484 ± 0,759 5,23 8,98 8,094 ± 0,984 6,37 10,58
Zreli 5,448 ± 0,789 3,25 6,62 9,83 ± 1,245 7,82 13,05
4.3.3. Agranulociti, limfociti
Najčešća su vrsta leukocita kod P. ridibundus. Jezgra tipičnog limfocita centralno je
pozicionirana i zauzima veliki udio stanice, dok se citoplazma vidi kao mali plavkasti prsten
oko tamno ljubičaste jezgre (Slika 32.). Prema veličini razlikuju se veliki i mali limfociti.
Prosječni promjer malih limfocita je 9,909 ± 0,869 μm (min 8,45 μm , max 11,92 μm) μm, a
velikih 11,591 ± 0,945 μm (min 10,07 μm, max 13,81 μm).
Ukupan broj malih i velikih limfocita prikazan je u Tablici 7.
4.3.4. Granulociti, neutrofili
Dosta su česti leukociti. Imaju bijelu citoplazmu u kojoj je segmentirana ljubičasta
jezgra. Broj segmenta varira od dva do četiri, najčešće ih je tri. Različitog su oblika, nekad su
smješteni na periferiji stanice, nekada u sredini (Slika 32.). Prosječni promjer neutrofila je
14,086 ± 1,558 μm (min 11,88 μm i max 17,78 μm).
Ukupan broj neutrofila prikazan je u Tablici 7.
4.3.5. Granulociti, eozinofili
Imaju narančasto obojane okrugle granule i ljubičasto obojanu jezgru centralno ili
ekscentrično smještenu (Slika 32.). Citoplazma je bijele boje. Broj jezgrinih segmenta varira
od jednog do četiri. Najčešće ih je dva. Prosječni promjer eozinofila je 12,707 ± 1,446 μm
(min 9,45 μm i max 16,68 μm).
Ukupan broj eozinofila prikazan je u Tablici 7.
4.3.6. Agranulociti, monociti
Sferičnog su oblika sa perifernom ljubičastom jezgrom i blijedo plavom citoplazmom
(Slika 32.). Jezgra može biti okrugla ili nepravilnog oblika, ali nije segmentirana. Spadaju u
jedne od najvećih krvnih stanica. Prosječni promjer je 14,364 ± 1,886 μm (min 11,6 μm i max
18,08 μm).
Ukupan broj monocita prikazan je u Tablici 7.

REZULTATI
39
4.3.7. Granulociti, bazofili
Jedine stanice nepravilnog sferičnog oblika, tamno ljubičaste jezgre i tamno ljubičasto-
plavih (skoro crnih) citoplazmatskih granula (Slika 32.). Najrjeđe su krvne stanice. Prosječni
promjer bazofila je 13,775 ± 1,099 μm (min 12,19 μm i max 15,03 μm).
Ukupan broj bazofila prikazan je u Tablici 7.
Slika 31. Rezultati bojanja eritrocita i trombocita May-Grünwald Giemsom (MGG).

REZULTATI
40
Slika 32. Rezultati bojanja leukocita May-Grünwald Giemsom (MGG).
Tablica 7. Ukupan broj velikih i mali limfocita, neutrofila, eozinofila, monocita i bazofila. Svaki tip
leukocita x 103 / ml krvi.
Srednja
vrijednost
Standardna
devijacija Median min max
Limfociti mali 1,08 0,97 0,9 0,18 2,86
Limfociti veliki 1,91 3,22 0,72 0,29 8,47
Neutrofili 0,96 0,73 0,82 0,03 1,94
Eozinofili 0,41 0,56 0,2 0,04 1,53
Monociti 0,71 0,75 0,6 0,06 2,06
Bazofili 0 0 0 0 0

REZULTATI
41
Slika 33. Udjeli pojedinih leukocita. Brojevi označavaju postotke.
4.4. Citokemijske analize
4.4.1. Alcijansko modrilo (AB)
Najbolji rezultati bojanja alcijanskim modrilom (AB) dobiveni su trominutnim
tretmanom pri pH 2,5 i 3,1 te četverominutnim pri pH 1 i pH 0,2 (Tablica 8.). Što je vrijeme
tretmana bilo dulje, to su se stanice više smežurale, neovisno o pH.
Tablica 8. Optimalne minutaže tretmana ABom ovisno o pH
min
pH
0,2 1 2,5 3,1
3 nema obojenja nema obojenja dobro obojano dobro obojano
4 svijetlo, ali dobro dobro obojano dobro obojano prejako obojano
5 dobro obojano nije dobro prejako obojano prejako obojano
Pri svim navedenim pH jezgre svih stanica obojane su plavo. Zreli eritrociti (Slika 34.a)
imaju neobojanu citoplazmu i oko jezgre daju pozitivnu reakciju na AB. Stari eritrociti (Slika
34.b) imaju vrlo blijedo obojanu jezgru i neobojanu citoplazmu. Pri svim pH pozitivna
reakcija na alcian blue (Tablica 9.) primjećena je kod limfocita (Slika 34.c i d) i monocita
(Slika 34.h) te nešto slabije kod neutrofila (Slika 34.e) i trombocita (Slika 34.g). Eozinofili
37,67
21,30
18,93
14,00
8,09
0,00
Veliki limfociti
Mali limfociti
Neutrofili
Eozinofili
Monociti
Bazofili

REZULTATI
42
(Slika 34.f) imaju obojane i neobojane granule koje se jako dobro vide. Bazofili nisu
pronađeni.
Boja nuclear fast red (NFR) daje dobre rezultate, čak pomaže u razlučivanju
unutarstaničnih struktura. Ljubičasto obojenje jezgre nastaje zbog preklapanja boja AB i NFR
(Slike 34.c, f, g i.h).
Slika 34. Rezultati bojanja alcijanskim modrilom (AB) i nuclear fast redom (NFR). Sve krvne stanice
imaju obojanu jezgru. Nezreli eritrociti pri pH 0,2 (a) i stari eritrociti pri pH 0,2 (b) su bez
citoplazmatskih obojenja. Monociti pri pH 2,5 (h) te limfociti pri pH2,5 sa NFR (c) i pH 3,1 (d) imaju
obojanu citoplazmu, kao i neki neutrofili pri pH 0,2 (e), poneki zreli trombociti pri pH 2,5 sa NFR (g) te
neke granule eozinofila pri pH 2,5 sa NFR (f). Mjerilo 20 μm.
4.4.2. Alfa-naftil acetat esteraza (ANAE)
Fiksacija krvnih razmaza je bila uspješna. Pozitivna je reakcija (Tablica 9.) pronađena
kod monocita (Slika 35.b) i nekih limfocita (crna boja). Ostale su stanice neobojane (Slika
35.a). Bazofili nisu pronađeni.
Slika 35. Rezultati reakcije alfa-naftil acetat esteraze (ANAE). Negativna reakcija kod zrelog trombocita
(a) i pozitivna kod monocita (b). Mjerilo 20 μm.

REZULTATI
43
4.4.3. Alfa-naftil butirat esteraza (ANBE)
Fiksacija je bila uspješna. Pozivitni rezultati (Tablica 9.) pronađeni su kod monocita te
ponekih zrelih trombocita i limfocita (Slika 36. ). Bazofili nisu pronađeni.
Slika 36. Rezultati reakcije alfa-naftil butirat esteraze (ANBE). Pozitivne reakcije kod nekih zrelih
trombocita (a), svih monocita (b) i pojedinih limfocita (d). Negativna reakcija u eozinofilu (c). Mjerilo 20
μm.
4.4.4 Alkalna fosfataza (ALP)
Nije bilo očekivanih pozitivnih rezultata, neovisno o vremenu inkubacije. Negativni
rezultati zapaženi su kod svih pronađenih stanica. Nuclear fast red dobro je obojao jezgre.
4.4.5. Metoda Schiffove baze i perjodne kiseline (PAS)
Provjera kvalitete Schiffova reagensa dala je neobičan rezultat, trenutnu promjenu boje
u tamno ljubičastu.
Pri inkubacijama od 10 i 13 minuta nije primjećeno pozitivno rozo obojenje. Pri 15 i 20
minuta dobivena su dobra obojenja sa normalnim oblikom stanica. Pozitivne reakcije (Tablica
9.) pronađene su kod limfocita (Slika 37.e), granula eozinofila (Slika 37.g) te u citoplazmi
nekih monocita (Slika 37.a), neutrofila (Slika 37.b) i nekih trombocita. Bazofili nisu opaženi.
Bojanje jezgri Gillovim hematoksilinom nije uspjelo.
Slika 37. Rezultati metode Shiffove baze i perjodne kiseline (PAS). Pozitivna reakcija kod monocita (a),
neutrofila pod PH2 filterom (b), negativna reakcija kod zrelih eritrociti pod PH2 filterom (c), nezrelog
trombocita pod PH2 filterom (d), pozitivna reakcija kod limfocita (e), neutrofila (f) i granula eozinofila
(g). Mjerilo 20 μm.

REZULTATI
44
4.4.6. PAS i AB kombinacija
Najbolji rezultati bojom alcianom blue (AB) dobiveni su trominutnom inkubacijom
ABa pri pH 2,5 i 3,1 te četverominutnom pri pH 1 i pH 0,2. Najbolji rezultati PAS metodom
dobiveni su 20 minutnom inkubacijom Schiffovim reagensom (Slika 38.). Za sve navedene
pH jezgre limfocita, trombocita, monocita i neutrofila obojane su plavo (zbog ABa), a
citoplazma istih crveno (zbog PASa).
Bojanje Schiffovim reagensom za sve pH u trajanju od 13 minuta ispalo je presvijetlo ili
je ostalo neobojano. Za 15 minuta najbolji rezultati postignuti su kod AB pH 0,2 i 2,5.
Inkubacija Schiffom 20 minuta dobro je obojala uzorke sa AB pH 1 i 3,1.
Boja nuclear fast red (NFR) dala je ljubičasto obojenje gdje se preklopila sa bojom AB
(najviše u jezgrama svih stanica).
Slika 38. Rezultati kombinirane metode alcijanskog modrila (AB) i periodic acid Schiff metode (PAS).
Pozitivne reakcije u zrelom trombocitu gore i limfocitu dolje AB pH1 (a), limfocitu AB pH1 (b), neutrofilu
AB pH1 (c), eozinofilu AB pH 1 (d), monocitu AB pH 2,5 (e) i bazofilu AB pH 2,5 (f). Mjerilo 20 μm.
4.4.7. Sudansko crnilo B (SBB)
Fiksacije formalinom, etanol octenom kiselinom, puferiziranim aceton-metanolom i
ohlađenim formalin-acetonom bile su uspješne kod svih uzoraka. Nefiksirani uzorci dali su
najbolje rezultate sedmominutnim bojanjem sudan blackom. Formalinom fiksirani uzorci ne
daju zadovoljavajuće rezultate pri ispitivanim minutažama. Kontrolni uzorci fiksirani ostalim
meotdama daju dobre rezultate nakon 5 i 6 minuta bojanja sudanom.
Pozitivna reakcija primjećena je u citoplazmi limfocita, neutrofila, monocita i
trombocita te vrlo slabo kod nezrelih i zrelih eritrocita (Tablica 9., Slika 39.). Stari eritrociti

REZULTATI
45
imaju potpuno neobojanu citoplazmu. Eozinofili imaju izrazito neobojane granule omeđene
crnim rubom (Slika 39.c). Bazofili imaju crne granule (Slika 39.f).
NFR daje odlične rezultate.
Slika 39. Rezultati bojanja tehnikom sudan black B (SBB) i nuclear fast red (NFR). Jezgre svih stanica
crvene su boje. Slaba reakcija u zrelom eritrocitu (a), negativna reakcija u starom eritrocitu (b) i
granulama eozinofila (c). Pozitivne reakcije u zrelom trombocitu (d), limfocitu (e), granulama bazofila (f),
neutrofilu (g) i monocitu (h). Mjerilo 20 μm.
Tablica 9. Rezultati svih citokemijskih analiza. + je pozitivan rezultat, - negativan, / znači da stanica nije
pronađena.
Ostale stanice Leukociti
Eritrociti Trombociti Limfociti Neutrofili Monociti Bazofili Eozinofili
AB - + - + + - + / + -
ALP - - - - - - -
ANAE - - + - - + / -
ANBE - + - + - - + - -
PAS - + - + + - + - / +
SBB + - + + + + + +
4.5. Atipične i neklasificirane stanice
4.5.1. "Raspadnute jezgre".
Nisu bile brojane, ali su dosta česte strukture (Slika 40.). Boja i morfologija odgovara
jezrgama starog eritrocita koje izgledaju manje kompaktno i "napuhano". Proizvoljno su
nazvane "raspadnute jezgre". Nisu uključene u analizu.

REZULTATI
46
Slika 40. Stari eritorcit (1, 5), zreli eritrocit (2, 3) i "raspadnuta jezgra" (4). Primjetiti sličnost oblika
jezgre starih eritrocita sa "raspadnutom". Obojano May-Grünwald Giemsom.
4.5.2. Neobične stanice
Pregledavanjem uzorka relativno često su se pojavljivale stanice proizvoljno nazvane
"ljubičaste jezgre" i nepoznate stanice slične monocitima. Nisu uključene u analize.
"Ljubičaste jezgre" (Slika 41.a) veličine su malih limfocita. Izrazito su tamne i kompaktne.
Ponekad imaju svijetlo roze "mjehuriće" u sredini (Slika 41.a). Stanice slične monocitima
imaju svijetlo ljubičastu jezgru koja izgleda vrlo rahlo i razasuta je po cijeloj citoplazmi.
Imaju citoplazmu blijedo plave do bijele boje. Veličina su kao monocit i veliki limfocit (Slika
41.b i c).
Slika 41. "Ljubičasta jezgra" (a). Na istom uzorku nepoznata stanica (b) i monocit (c). Obojano May-
Grünwald Giemsom.

REZULTATI
47
4.5.4. Azurofili i heterofili
Azurofili (Slika 42.a i b) su veličine monocita sa ekscentrično smještenom ljubičastom
jezgrom. U plavoj citoplazmi se vide plavkasto-ljubičaste granulice. U ovom radu ubrajali su
se u monocite.
Heterofili (Slika 42.c) su dimenzija kao neutrofil. Imaju segmentiranu jezgru ljubičaste
boje. Citoplazma im sadržava crvenkaste granulice. Nisu česte stanice, ima ih dosta manje
nego azurofila. U ovom radu ubrajali su se u neutrofile.
Slika 42. Azurofili (a i b), heterofil (c). Obojano May-Grünwald Giemsom.
4.5.5. Morfološke neobičnosti krvnih stanica
Nađeno je par atipično obojanih eritrocita, bilo u jezgri (Slika 43.a) ili citoplazmi (Slika
43. c). Nađeni su eritrociti čija jezgra izgleda fragmentirano (Slika 43.b). Također, pronađen
je eritrocit oblika suze (Slika 43.d) i eritrocit sa ekscentričnom jezgrom (Slika 43.f). Čak je
pronađen eritrocit u diobi (Slika 43.e).
Nađeno je više "bubrežastih" limfocita (Slika 44.a) te zreli trombocit vretenastog oblika
(Slika 44. b).

REZULTATI
48
Slika 43. Neobični eritrociti. Neobično obojana jezgra (a), fragment jezgre (b), neobično obojana
citoplazma (mogući prelazak u starog eritrocita) (c), oblik suze (d), eritrociti u diobi (e) i ekscentrična
jezgra (f). Obojano May-Grünwald Giemsom osim slike 43.f gdje je korištena boja sudan black B (SBB) i
kontrasno obojenje nuclear fast red (NFR).
Slika 44. "Bubrežasti" limfocit (a) i vretenasti trombocit (b). Obojano May-Grünwald Giemsom.

RASPRAVA
49
5. RASPRAVA
Eritrociti Pelophylax ridibundus najbrojnije su krvne stanice kao i u ostalih kralježnjaka
osim sisavaca. U usporedbi s ljudima (5 000 000 / ml krvi) P. ridibundus (430 000 / ml) ima
oko 10x manje eritrocita po ml krvi (Guyton i Hall 2006). Broj eritrocita žaba varira ovisno o
rodu. Rod Rana ih ima najviše, preko 500 000 / ml. Čini se kako P. ridibundus (prije zvana
Rana ridibundus) spada u ovu skupinu. U usporedbi sa Bombina bombina (250 000 / ml krvi)
i Xenopus laevis (75 000 / ml krvi) P. ridibundus ima daleko veću brojnost eritrocita (Arikan i
sur. 2010; Arserim i Mermer 2008; Hadji-Azimi i sur. 1987). Dimenzije zrelih eritrocita P.
ridibundus (cca 15 x 23 μm) i njihovih jezgri (cca 5 x 9 μm) otprilike su iste kao kod B.
bombina, ali su nešto veće od onih u X. laevis (Arikan i sur. 2010; Hadji-Azimi i sur. 1987).
Eritrociti Rana macrocnemis varijabilnijih su veličina od P. ridibundus (širirne 12 do 18 μm,
dužine 17 do 29 μm), ali prosječno sličnih dimenzija (Arserim i Mermer 2008). Gmazovi
imaju prosječno manje eritrocite i njihove jezgre. Kod poskoka otprilike iznose 13 x 18 μm sa
jezgrom 5 x 8 μm, a kod ruskih kornjača 10 x 20 μm sa jezgrom 4 x 6 μm (Lisičić 2005;
Knotková i sur. 2002). Eritrociti ovdje opisani kao nezreli vjerojatno su slični nezrelim
eritrocitima II opisanim kod X. laevis (Hadji-Azimi i sur. 1987).
Morfologija zrelih eritrocita P. ridibundus gotovo je identična u spomenutim žabljim
vrstama. Za razliku od gmazova (poskok i kornjača) i riba imaju malo izduženiju jezgru.
Vrijednost hematokrita P. ridibundus (43 %), R. macrocnemis (34 %) i X. laevis (37 %)
nalazi se unutar ljudskih granica- od 30 do 45 % (Arikan i sur. 2010; Arserim i Mermer 2008;
Guyton i Hall 2006; Wilson i sur. 2011). Zanimljivo kako kod poskoka vrijednost hematokrita
iznosi 73 % (Lisičić 2005). Vrijednost hematokrita riba varira ovisno o vrsti (Shigdar i sur.
2007).
Trombociti P. ridibundus i ostalih kralježnjaka nemogu se morfološki uspoređivati sa
ljudskima. Prosječni broj trombocita ptica je 25 000 000 / ml krvi, a riba 65 000 000 / ml krvi
što je deseterostruko manje nego u ljudi (Weiss i Wardrop 2010). To se može objasniti malom
veličinom ljudskih trombocita u usporedbi s ostalim kralježnjacima. Prosječni broj trombocita
spomenutih vrsti žaba mnogo je manji. Kod R. macrocnemis iznosi oko 31 000/ ml krvi
(Arserim i Mermer 2008), što je više nego kod X. laevis (19 000/ ml krvi) i B. bombina (13
000 / ml krvi; Arikan 2010). U ovom radu nije mjerena brojnost trombocita P. ridibundus s

RASPRAVA
50
obzirom da je dio njih morfološki jako sličan malim limfocitima, ali očekivano bi imali manju
vrijednost od R. macrocnemis i veću od B. bombina.
Morfologija trombocita također je očuvana u kralježnjaka osim u sisavaca. Kod
spomenutih gmazova Lisičić (2005) i Knotková i sur. (2002) nisu opisali vretanaste oblike, za
razliku od Shigdar i sur. (2007) koji su pronašli te oblike u ribi Maccullochella peelii peelii.
Jedino kod M. peelii peelii trombociti imaju segmentiranu jezgru (Shigdar i sur 2007).
Veličina trombocita i njegove jezgre u P. ridibundus (7,5 x 13 μm) prosječno je manja od
onih u B. bombina (cca 8,5 x 17 μm), ali jako slična poskokovim (8 x 13 μm) opisanih kod
Lisičića (2005).
Ukupan broj leukocita P. ridibundus kreće se od 5070 do 7500 / ml krvi što ga
smještava u ljudski raspon i čini skoro dvostruko većim od onog u R. macrocnemis (3500
leukocita / ml krvi) i X. laevis (Arserim i Mermer 2008, Hadji-Azimi i sur. 1987). Raspon
ukupnog broja leukocita P. ridibundus opisanog kod Gül i sur. (2011) kreće se od 4500 do
8000 / ml krvi, što se podudara sa dobivenim rezultatima.
Udio pojedinog tipa leukocita donekle se poklapa kod ljudi i P. ridibundus. Oboje
imaju najviše neutrofila i limfocita, s tim da ljudi imaju najviše neutrofila (40-75 %), a P.
ridibundus limfocita (prosječno 59 %). Udio monocita sličan je kod obje vrste (<10 %).
Eozinofili su češći u P. ridibundus (14 %) nego kod čovjeka (1-5 %). Bazofili su izrazito
rijetki (<1 %) u obje vrste (Reagena 2003). Uspoređujući P. ridibundus sa drugim vrstama
žaba, R. macrocnemis i X. laevis, također dobivamo slične omjere. P. ridibundus i X. laevis
imaju najviše limfocita, s tim da ih X. laevis ima skoro dvostruko manje (30 %). Udio
neutrofila im je otprilike isti, ali ih X. laevis ima malo više (27 %). X. laevis ima mnogo manji
udio eozinofila (1,2 %) i monocita (<2 %), ali zato skoro 40 % ukupnih leukocita otpada na
bazofile (Hadji-Azimi i sur. 1987). Davis (2011) je opisao udjele kod još 19 vrsti žaba i dosta
su slični ovim rezultatima. Najzastupljeniji su limfociti (prosječno 63 %), zatim neutrofili
(prosječno 19 %), bazofili (9 %) i eozinofili (6 %), a najmanje ima monocita (3 %). Kornjače
opisane kod Knotková i sur. (2002) također imaju najviše limfocita (46 %) i heterofila (37 %),
najmanje imaju azurofila, eozinofila i bazofila (svaki 5 %), dok monocita imaju nešto više od
P. ridibundus (12 %). Limfociti su najčešći i kod ribe M. peelii peelii opisane od Shigdara i
sur. (2007), čineći 61 % ukupnih leukocita, gotovo isto kao i P. ridibundus. Udio heterofila u
iste ribe skoro je trostruko manji (24 %), dok monociti i bazofili zajedno čine oko 10 %
leukocita.

RASPRAVA
51
Morfološki leukociti kralježnjaka jako su slični, iznimke su heterofili (izgledom sliče
eozinofilima i neutrofilima) i azurofili (podsjećaju na monocite).
Dimenzije velikih i malih limfocita spomenutih žabljih vrsta otprilike su istih promjera.
Najmanjih su dimenzija kod X. laevis, a najvećih kod P. ridibundus (Arserim i Mermer 2008;
Hadji-Azimi i sur. 1987). Obje vrste limfocita B. bombina veći su nego kod spomenutih
(Arikan i sur. 2010). Kod poskoka također su opisani mali i veliki limfociti, gotovo istih
dimenzija kao kod P. ridibundus (Lisičić 2005). Limfociti opisani kod kornjača od Knotková
i sur. (2002) izrazito su mali, promjera 6 μm. Morfološki izgledaju gotovo isto kod ostalih
spomenutih vrsta.
Neutrofili spomenutih žaba prosječno su nešto veći od ljudskih (Piršljin i Milinković
Tur 2009). X. laevis, P. ridibundus i poskok imaju gotovo iste promjere neutrofila, nešto
manje nego R. macrocnemis (Arserim i Mermer 2008; Hadji-Azimi i sur. 1987; Lisičić 2005).
Vodozemci i ljudi, za razliku od gmazova, imaju segmentiranu jezgru neutrofila. Najčešći
broj segmenta P. ridibundus je tri. Jezgra je ponekad smještena acentrično, slično kao kod
gmazova (Lisičić 2005).
Eozinofili P. ridibundus sličnih su dimenzija (promjera 13 μm) kao kod ljudi, X. laevis i
ruskih kornjača (Hadji-Azimi i sur. 1987; Knotková i sur. 2002). Kod poskoka nešto su većeg
promjera i imaju ekscentrično smještenu okruglastu jezgru slično ribi M. peelii peelii (Lisičić
2005; Shigdar i sur. 2007). Ljudi, zajedno sa pticama i vodozemcima imaju segmentiranu
jezgru eozinofila (Claver i Quaglia 2009). Najčešći broj segmenta u P. ridibundus je dva, isto
kao i kod ljudi.
Monociti ljudi nešto su većeg promjera od P. ridibundus (Piršljin i Milinković Tur
2009). Žabe R. macrocnemis i P. ridibundus te riba M. peelii peelii imaju gotovo iste
promjere, oko 14 μm, što ih čini nešto većima nego kod X. laevis, ali manjim nego kod B.
bombina (Arikan i sur. 2010; Arserim i Mermer 2008; Hadji-Azimi i sur. 1987; Shigdar i sur.
2007). Monociti P. ridibundus nešto su manji od onih u ruske kornjače (prosječni promjer 15
μm) i poskoka (prosječni promjer oko 20 μm). Slično gmazovima i ribama imaju ekscentrično
smještenu nesgementiranu jezgru. Kod poskoka i ljudi uglavnom je bubrežasta, kod kornjača
kockasta, dok kod žaba može bit okruglasta ili nepravilnog oblika. Citoplazma im je
uglavnom blijedo obojena (Lisičić 2005; Knotková i sur. 2002; Shigdar i sur. 2007). Kod
većine kralježnjaka monociti su najveće stanice.
Bazofili ljudi i X. laevis otprilike su istih veličina (prosječni promjer 12 μm), manji su
od onih u P. ridibundus koji su manji od onih u B. bombina (Arikan i sur. 2010; Hadji-Azimi
i sur. 1987). Bazofili gmazova manji su od žabljih, ali su ovalnog oblika sa centralnom

RASPRAVA
52
jezgrom. Jezgra nekad može biti skroz zaklonjena zbog citoplazmatskih granula kao što je to
slučaj kod kornjača. Kod poskoka nepravilnog je oblika, kod ljudi i ribe M. peelii peeli može
biti segmentirana, za razliku od okruglaste jezgre P. ridibundus (Lisičić 2005, Knotková i sur.
2002; Shigdar i sur. 2007).
Heterofili namjerno nisu bili opisivani u ovom radu iz više razloga. Glavni je što su se
May-Gründwald Giemsa (MGG) bojanjem jasno mogli razaznati neutrofili istih karakteristika
kao u sisavaca. Heterofili nisu opisani kod spomenutih vrsti žaba. Neki autori smatraju kako
bi trebalo promijeniti terminologiju u smislu da neutrofili budu samo kod sisavaca, a heterofili
kod ostalih kralježnjaka (Claver i Quaglia 2009). No, neki gmazovi i vodozemci imaju dobro
prepoznatljive heterofile i azurofile (Claver i Quaglia 2009; Hadji-Azimi i sur. 1987; Lisičić
2005).
Azurofili također nisu bili opisivani u ovom radu iz sličnih razloga. Kod žaba Rana
catesbeiana i X. laevis jako su rijetke stanice i morfološki se mogu lako zamijeniti sa
monocitima (Bricker i sur. 2012). Češće su opisivani kod gmazova nego kod vodozemaca
(Knotková i sur. 2002).
Neobične stanice pronađene kod P. ridibundus pojavile su se na večini uzorka. Stanice
nazvane "raspadnute jezgre" sliče jezgri starog eritrocita. Homogene su obojane, imaju
mjehuriće što im daje nepravilni oblik i većih su dimenzija od jezgri zrelih eritrocita. Moguće
da su prilikom rađenja razmaza stari eritrociti popucali i ispustili jezgru. Eritrocit čija jezgra
izgleda fragmentirano mogao bi sadržavati Howell Jollyjevo tjelešce koje nastaje nepravilnom
diobom jezgre. Eritrociti čudnog obojenja mogu biti ili prelazni oblici prema starim
eritrocitima ili mogu biti polikromatski. Eritrociti sa tamnim citoplazmatskim točkicama
(bazofilnim tjelešcima) koje nisu sličile jezgri su također pronađeni. Ukoliko gmazovi imaju
preko 40 % (a ptice preko 55 %) eritrocita s abnormalnom morfologijom to ukazuje na
različite poremećaje. Npr. poikilocitoza može nastati uslijed metaboličkih promjena ili
hemolize, bazofilna tjelešca mogu označavati pristunost parazita, hipokromija najčešće
karakterizira manjak željeza ili hemoglobina i tako dalje (Pendl 2006). Stanice nazvane
"ljubičaste jezgre" veličinom i bojom odgovaraju nezrelim limfocitima. Ponekad imaju
mjehuriće što nije karakteristika homogenih, kompaktnih nezrelih limfocita. Nepoznata
stanica sa vrlo rahlim kromatinom mogla bi biti raspadnuti monocit, makar sliči limfoblastu
X. laevis pronađenom u slezeni (Hadji-Azimi i sur. 1987). No, kod P. ridibundus ove "čudne"
stanice nisu bile česte.

RASPRAVA
53
Buduća istraživanja krvnih stanica P. ridibundus mogla bi uključiti heterofile i azurofile
s obzirom da su pronađeni kod vodozemaca i gmazova (Claver i Quaglia 2009; Knotková i
sur. 2002). Također bi se mogla raditi analiza brojnosti krvnih stanica P. ridibundus ovisno o
spolu. Razlike u brojnosti eritrocita između mužjaka i ženki pronađene su kod žabe
Pseudepidalea viridis (Gül i sur. 2011). U R. macrocnemis i Bufo bufo nisu pronađene
značajne statističke razlike, ali je opaženo kako ženke R. macrocnemis imaju nešto više
eritrocita, ali manje ukupnih leukocita (Arserim i Mermer 2008; Dönmez i sur. 2009). Razlika
u sastavu krvi punoglavca i odrasle žabe primjećena je kod R. catesbeiana, jedine istraživane
vrste do sad. Prosječno limfociti punoglavaca čine oko 80 % leukocita, a kod odraslih oko 50
% (Davis 2011).
Elektronska mikroskopija pruža odličan uvid stanične mikrostrukture i mogla bi riješiti
neka morfološka pitanja, poput identifikacije nepoznatih stanica.
Zanimljiva ideja bila bi istražiti postoje li krvne grupe ili analozi krvnim grupama
sisavaca kod P. ridibundus. Najviše istraživanja bilo je na sisavcima. Kod pasa postoji 8
krvnih grupa ovisnih o DEA (engl. dog erythrocyte antigen) antigenima, dok mačke imaju tri
grupe A, B i AB. Konji imaju najviše krvnih grupa, njih 30 od kojih su 8 glavne (Cornell
University College of Veterinary Medicine, 2010). Kod gmazova pronađeno je kako gušteri
imaju krvne grupe sličnije ljudskim Rhesus faktorima nego ABO grupi. Od ispitivanih vrsti
žaba (Rana temporaria) i riba (Gadus merlangus i G. luscus) nisu pronađene krvne grupe
slične ljudskoj (Ashhurst 1956).
Rezultati alcijanskog modrila (AB) pri pH 0,2, 1, 2,5 i 3,5 (Lowe 1998a) nisu se
značajno razlikovali što bi moglo značiti kako su krvne stanice P. ridibundus bogate sulfatnim
polisharidima (Green i Pastewka 1974; Kiernan 2008). Jezgre svih krvnih stanica obojale su
se ABom. To je bilo očekivano pri pH 2,5 i više, ali ne pri nižima (Kiernan 2008). Obično se
to dogodi zbog predugog tretmana, što ovdje nije bio slučaj. Produljene minutaže tretmanom
ABa (neovisno o pHu) rezultiralo je smežuranim stanicama, što je bilo očekivano s obzirom
da nisu bile fiksirane. Trebalo bi isprobati ovu tehniku sa fiksativom. Postoji mogućnost da je
jezgrin kromatin rahliji nego u ostalih kralježnjaka, ali u dostupnoj literaturi jedino je kod
sisavaca bilo bojanja ovom tehnikom (Weiss i Wardrop 2010). Dokaz da se zbilja radi o
jezgrama potvrđen je kontrasnim obojenjem nuclear fast red (NFR). NFR zasebno boji
nukleinske kiseline crveno, a poklapanjem sa AB nastaje ljubičasto obojenje jezgri.
Kod P. ridibundus pozitivnu reakciju imaju limfociti i monociti te nešto slabiju reakciju
neutrofili i trombociti. Eozinofili imaju obojane i neobojane granule. Očekivano bi bilo

RASPRAVA
54
bojanje membrane eritrocita s obzirom da je bogata negativno nabijenom sijalinskom
kiselinom. Kod sisavaca jedino su se granule bazofila bojale ABom (Weiss i Wardrop 2010),
ali oni nisu pronađeni u P. ridibundus.
Prije tretmana periodic acid Schiffom (PAS) preporuča se napraviti kontrolu Schiffova
reagensa. Dobar Schiffov reagens u reakciji sa formaldehidom brzo promjeni boju u crveno-
ljubičasto. Schiff kojem ističe trajnost ima dulju reakciju i daje tamno ljubičasto-plavu boju
(IHC World 2011d). Schiffov reagens u ovom radu odmah je dao tamno ljubičasto obojenje.
Zbog tog razloga produljeni su tretmani na 20 minuta umjesto 15 opisanih u Loweovom
protokolu (1998c).
Kod P. ridibundus pozitivne reakcije na PAS metodu imaju limfociti u citoplazmi i
eozinofili u granulama te neki monociti, neutrofili i trombociti u citoplazmi. Slične rezultate
dobili su Hadji-Azimi i sur. (1987) kod X. laevis gdje pozitivne rezultate daju trombociti i
neutrofili. Kod ribe M. peelii peelii pozitivnu reakciju na PAS daju trombociti, bazofili,
heterofili i dio limfocita. Primijećeno je kako se stare stanice riba intenzivnije boje PASom
(Shigdar i sur. 2009).
Kombinacija AB + PAS dala je očekivane rezultate kod P. ridibundus. Plava boja dolazi
od ABa, crveno-roza od PASa, a ljubičasta je na mjestu preklapanja ovih boja. Sve stanice
koje su se bojale zasebno sa jednom i drugom tehnikom, boje se sa ovom kombinacijom. To
se odnosi na limfocite, monocite, eozinofile, neutrofile, čak i trombocite. Tipične boje
navedenih stanica su plave za jezgru i ljubičaste ili crvenkaste za citoplazmu.
Reakcije nespecifičnih esterazi podudaraju se sa rezultatima opisanim kod riba M. peelii
peelii (Shigdar i sur. 2009), sisavaca (Chronolab 2012c; Weiss i Wardrop 2010), R.
catesbeiana (Bricker i sur. 2012) i X. laevis (Hadji-Azimi i sur. 1987). Kod spomenutih vrsti
samo se monociti boje nespecifičnim esterazama (alfa-naftil acetat esterazom, ANAE, i alfa-
naftil butirat esterazom, ANBE).
Pozitivna reakcija na ANAE kod monocita identična je u spomenutim vrstama. Svi
limfociti sisavaca i ribe M. peelii peeli te neki limfociti P. ridibundus imaju aktivne ANAE.
Ostale stanice P. ridibundus nisu pozitivne na ANAE, a bazofili nisu pronađeni. Bazofili i
heterofili ribe M. peelii peeli, kao i eozinofili, megakariociti te trombociti sisavca i trombociti
X. laevis pokazuju pozitivnu reakciju na ovu estarazu (Shigdar i sur. 2009, Weiss i Wardrop
2010).

RASPRAVA
55
Pozitivnu reakciju na ANBE također pokazuju monociti spomenutih vrsti osim ribe M.
peelii peelii (Shigdar i sur. 2009). Kod P. ridibundus pozitivne rezultate daje samo poneki
zreli trombocit i limfocit. Neutrofili R. catesbeiana imaju aktivnu ANBE kao i neutrofili
sisavaca (Bricker i sur. 2012; Weiss i Wardrop 2010). Eozinofili sisavaca i X. laevis također
imaju aktivne ANBE, čija je aktivnost pronađena u bazofilima X. laevis (Hadji-Azimi i sur.
1987; Weiss i Wardrop 2010). Bazofili P. ridibundus nisu pronađeni, makar bi mogli dati
pozitivni rezultat.
Iz rezultata nespecifičnih esterazi može se vidjeti kako su ANAE i ANBE očuvane u
monocitima skoro svih kralježnjaka. Za ostale stanice ima premalo podatka za iznošenje
zaključka.
Alkalna fosfataza (ALP) P. ridibundus nije dala nikakve pozitivne rezultate, neovisno o
duljini tretmana. Iako je protokol opisan sa fast red TR (Sigma-Aldrich 2006) soli, u ovom
radu korištena je fast blue RR sol iz razloga što oboje spadaju u istu vrstu boja i trebale bi na
isti način tvoriti obojani produkt (Kiernan 2008) te radi uštede s obzirom da se koristila i kod
alfa-naftil acetat esteraze (ANAE). Paralelno su bojani uzorci riba babica (Barbatula
barbatula) i miševa (Mus musculus) koji su pod istim uvjetima dali očekivano tamno plavo do
crno obojenje limfocita. Limfociti se također boje kod sisavaca i djelomično kod ribe M.
peelii peelii. Zanimljivo kako neutrofili sisavaca i heterofili (funkcijski isti neutrofilima) te
ribe pokazuju pozitivnu reakciju na ALP (Shigdar i sur. 2009; Weiss i Wardrop 2010). Kod
sisavaca eozinofili u citoplazmi također pokazuju aktivnost (Weiss i Wardrop 2010). Jedino
su Bricker i sur. (2012) ispitivali ovaj enzim u R. catesbeiana, gdje su samo granule bazofila
dale dobar rezultat, a kod X. laevis gdje nije bilo pozitivnih rezultata.
Ovo bi moglo značiti da limfociti P. ridibundus i X. laevis imaju drukčije ili
modificirane enzime, možda sa promjenjenim aktivnim mjestom. Zanimljivo je kako rod
Rana (sličniji P. ridibundusu) ima aktivnost u bazofilima, a udaljenija X. laevis nigdje.
Bojanje metodom sudan black B (SBB) jako je variralo o metodi fiksacije. Nefiksirani
uzorci dali su odlične, očekivane rezultate, crnu citoplazmu neutrofila, limfocita te uglavnom
crnu staničnu membranu. U ovom radu bilo je dosta eksperimentiranja sa fiksacijom
kontrolnih uzoraka P. ridibundus. Formalin je dao najlošije rezultate jer je otapao lipide
(Bancroft i Gamble 2008). Zato su on i aceton upotrebljeni kao negativne kontrole. P.
ridibundus pokazuje jake pozitivne rezultate u citoplazmi limfocita, neutrofila i monocita.
Slabiju reakciju pokazuje citoplazma trombocita te nezrelih i zrelih eritrocita. Kod starih

RASPRAVA
56
eritrocita, kao i u drugim citokemijskim tehnikama, nema obojenja. Granule eozinofila nisu
obojane nego imaju crni rub, za razliku od bazofilnih granula. To bi značilo kako su granule
eozinofila unutar lipidne membrane, dok su granule bazofila bogate lipidima. Zajedničko
ostalim žabama (X. laevis i R. catesbeiana) i sisavcima su crna citoplazmatska obojenja
neutrofila. R. Catesbeiana, kao i P. ridibundus, pokazuje pozitivne rezultate u citoplazmi
eozinofila. Kako se SBBom boje heterofili ribe M. peelii peelii, to dodatno utvrđuje njihovu
povezanost sa neutrofilima. Pozitivne rezultate također pokazuju neki limfociti (Shigdar i sur.
2009). Eozinofili sisavca također pokazuju pozitivne reakcije na SBB kao i neki monociti
(Weiss i Wardrop 2010).
Postoje i druge citokemijske metode koje nisu testirane u ovom radu, većina zbog
ograničenih financijskih mogućnosti. Osim za identifikaciju tipova leukocita, citokemijske
metode pomažu klasifikaciji različitih akutnih leukemija (Chronolab 2012a, 2012b, 2012c i
2012d).
Peroksidaze se nalaze u fagocitnim vakuolama i služe u obrani od patogena. Što su one
aktivnije, to su leukociti intenzivnije obojani. Ako su aktivne u mijelodinoj lozi onda se
nazivaju mijeloperoksidazama. Obično se rezultati peroksidaza i SBBa poklapaju (Chronolab
2012d; Shigdar i sur. 2009). Najjaču aktivnost u sisavaca imaju eozinofili, monociti i
neutrofili (u primarnim granulama). Štoviše, peroksidaze neutrofila aktivne su kod svih
kralježnjaka, odnosno kod riba aktivne su u njihovim ekvivalentima, heterofilima (Shidgar
2009, Bricker i sur. 2012; Hadji-Azimi i sur. 1987). Eozinofili ptica i gmazova pokazuju
aktivnost ovog enzima, kao i poneki žablji eozinofili, heterofil i trombociti (Bricker i sur.
2012; Claver i Quaglia 2009; Hadji-Azimi i sur. 1987). Zanimljivo je kako kod gmazova i
ptica aktivnost mijeloperoksidaza postoji u neutrofilima, ali ne i u heterofilima (Claver i
Quaglia 2009). To bi moglo značiti kako su ipak neutrofili glavni fagociti za razliku od
heterofila.
Kisela fosfataza (ACP, engl. acid phosphatase) aktivna je u većini hematopoetskih
stanica sisavaca. Može pomoći kod identifikacije razvojnih stadija stanica i vrsta limfocita.
Jaku aktivnost ACP pokazuju makrofagi, dok nešto slabiju plazma stanice, megakariociti i
monociti. Slabe reakcije imaju neutrofili, mijelociti i promijelociti (Chronolab 2012a). ACP je
aktivna kod eozinofila i heterofila ptica (Claver i Quaglia 2009) te u svim leukocitima ribe M.
peelii peelii (Shigdar i sur. 2009). Kod vodozemaca u dostupnoj literaturi nije testirana
aktivnost ovog enzima.

RASPRAVA
57
Kloroacetat esteraza specifična je esteraza prisutna u svim leukocitima ribe M. peelii
peelii, osim monocita (Shigdar i sur. 2009). Kod žaba aktivna je u monocitima te u
neutrofilima R. catesbeiana (Bricker i sur. 2012).
Zanimljivo je kako u dostupnoj literaturi nema citokemijskih analiza azurofila.
Što se tiče fiksacije ugljikohidrata, varira ovisno o tome što se želi očuvati. Ako se
proučava glikogen, koji je topiv u vodi, treba se izbjegavati formalin, makar je moguće sa
puferiziranim formalinom. Način fiksacije ovisi koje strukture želimo očuvati. Ako su u
pitanju lipidi, osmijev tetraoksid pokazao se dobrim fiksativom, dok formaldehid (CH2O),
alkohol i aceton nisu dobar izbor jer otapaju lipide. Sa druge strane alkohol i aceton dobri su
izbor fiksacije kod ispitivanja aktivnosti enzima jer im ne narušavaju aktivnost, ali ni ne
fiksiraju proteine. Fiksacija također utječe na promjene pH stanica. Formalin radi lužnatije
uvjete koji mogu smanjiti broj kationa (npr. NH3+ grupa) za razliku od alkohola koji može
smanjiti broj anionskih skupina, poput fosfata u kromatinu (Bancroft i Gamble 2008).
Pozitivne i negativne kontrole nisu bile svugdje moguće, ali nisu ni bile nužno potrebne.
Za pozitivne kontrole koristili su se mišji uzorci, gdje se zna kako stanice izgledaju kod
pojedinih citokemijskih bojanja. Pozitivne kontrole citokemijskih tehnika također se vide u
specifičnom bojanju određenih stanica (npr. nespecifične esteraze koje bojaju monocite crno).
Primjećeni su različiti rezultati kod usporednih tretmana citokemijskim tehnikama
hibernirajućih i kontrolnih uzoraka P. ridibundus. Hibernirajući uzorci (korišteni za
uhodavanja protokola) reagirali su očekivano na fiksaciju formalinom kod sudan black B
tehnike za razliku od kontrolnih. To bi moglo indicirati na drukčiju membransku strukturu.
Kod alcian blue tehnike primjećeno je kako se jezgre hibernirajućih uzoraka slabije boje nego
kod kontrolnih. To bi moglo značiti kako imaju kompaktniju jezgru, moguće zbog smanjenje
transkripcije uslijed stanja mirovanja. Na ovome bi se mogla raditi genomska istraživanja, čak
i epigenetska kako bi se utvrdilo što točno uzrokuje ove promjene. Ostali rezultati nisu se
značajno razlikovali od kontrolnih uzoraka korištenih u ovom radu. Također, bilo bi
informativno imati citokemijske analize ostalih žaba i usporediti ih na evolucijskoj skali, što
bi moglo dati uvid o mogućoj evoluciji i adaptaciji enzima.
Dobiveni rezultati mogli bi se sagledati i sa filogenetskog stajališa koje bi sigurno
rasvjetlilo mnoga pitanja iz ovog rada.

ZAKLJUČAK
58
6. ZAKLJUČAK
Brojnost eritrocita i leukocita Pelophylax ridibundus te vrijednost hematokrita nalazi se
u rasponu ostalih žaba. Najveći udio leukocita redom čine limfociti, neutrofili, eozinofili,
monociti dok je udio baozfila najmanji.
Morfologija i dimenzije eritrocita i leukocita, sa manjim razlikama, odgovaraju onima
opisanim kod ostalih žabljih vrsti. Leukociti s najvećim prosječnim promjerom su monociti,
zatim slijede neutrofili, bazofili, eozinofili te veliki i mali limfociti. Svi neutrofili i eozinofili
imaju segmentiranu jezgru koja može biti centralno i acentrično smještena. Monociti i
leukociti imaju ekscentričnu nesgmentiranu jezgru, koja je ponekad u monocitima
nepravilnog oblika. Bazofili imaju centralnu nesegmentiranu jezgru koja može biti zaklonjena
zbog citoplazmatskih granula.
Citokemijskim analizama pokazanao je kako nije lako identificirati pojedini tip
leukocita. Limfociti se boje alcijanskim modrilom (AB), metodom Schiffove baze i perjodne
kiseline (PAS) i sudanskim crnilom B (SBB) te ponekad pokazuju aktivnost nespecifičnih
esterazi (NSE). Neutrofili se boje jedino SBBom, ponekad ABom i PASom. Monociti uvijek
pokazuju aktivnost nespecifičnih esterazi, boje se ABom, SBBom i ponekad PASom.
Eozinofili nemaju aktivne NSE. Boje se SBBom, PASom i ponekad ABom. Bazofili su
negativni na alfa-naftil acetat esterazu (ANAE), bojaju se SBBom, ali nisu pronađeni kod AB,
alfa-naftil butirat esteraze (ANBE), alkalne fosfataze (ALP) i PAS metode. Jezgra svih
eritrocita boji se ABom. U citoplazmi nezrelih i zrelih eritrocita postoji slabo obojenje
SBBom. Na ostale metode ne reagiraju. Stari eritrociti se ne boje niti jednom metodom.
Trombociti se boje SBBom, slabo ABom i PASom te ponekad pokazuju aktivnost ANAE.
Niti jedna stanica nije pozitivna na ALP.
Azurofili i heterofili nisu opisani što ne isključuje mogućnost njihovog postojanja.

LITERATURA
59
7. LITERATURA
Alberts B., Bray D., Lewis J., Raff M., Roberts K., Watson J.D. (1994): Molecular Biology of
the Cell. 3.izdanje. Garland Publishing.
Arserim S.K., Mermer A. (2008): Hematology of the Uludağ Frog, Rana macrocnemis
Boulenger, 1885 in Uludağ National Park (Bursa, Turkey). E.U. Journal of Fisheries &
Aquatic Sciences 25: 39-46.
Ashhurst D.E. (1956): Red Blood-Cell Antigens in some lower Vertebrates. The Journal of
Experimental Biology 33: 249-255.
Baker J.R. (1956): Improvements in the Sudan Black Technique. Quarterly Journal of
Microscopical Science 97: 621-623.
Bancroft J.D., Gamble M. (2008): Theory and practice of histological techniques. 6. izdanje.
Edinburgh: Churchill Livingstone.
Bayliss High O.B. (1981): The histochemical versatility of Sudan Black B. Acta Histochem
Suppl. 24: 247-55.
Bricker N.K., Raskin R.E., Densmore C.L. (2012): Cytochemical and immunocytochemical
characterization of blood cells and immunohistochemical analysis of spleen cells from 2
species of frog, Rana (Aquarana) catesbeiana and Xenopus laevis. Vet Clin Pathol 41: 356-61.
Christie R.M. (2001): Colour Chemistry. 1.izdanje. The Royal Society of Chemistry.
Chronolab (2012a): Acid phosphatase. http://www.chronolab.com/hematology/5_3_3.htm;
pristupljeno 27.9.2012.
Chronolab (2012b): Alkaline phosphatase.
http://www.chronolab.com/hematology/5_3_1.htm; pristupljeno 27.9.2012.
Chronolab (2012c): Alpha- naphtyl- acetate- esterase.
http://www.chronolab.com/hematology/5_3_2.htm; pristupljeno 27.9.2012.
Chronolab (2012d): Myeloperoxidase. http://www.chronolab.com/hematology/5_3.htm;
pristupljeno 27.9.2012.
Claver i Quaglia J.A., Quaglia A.I.E. (2009): Comparative Morphology, Development, and
Function of Blood Cells in Nonmammalian Vertebrates. Journal of Exotic Pet Medicine 18:
87-97.

LITERATURA
60
Cornell University College of Veterinary Medicine (2010): Blood types.
http://ahdc.vet.cornell.edu/clinpath/modules/coags/bldtype.htm; pristupljeno 3.1.2013.
Davis A.K. (2011): Amphibian Leukocyte Profiles.
http://wildlifehematology.uga.edu/Amphibians/index.htm; pristupljeno 5.10.2012.
Desai S.P. (2006): Laboratorijske pretrage u kliničkoj medicini, praktični pristup. 1.izdanje.
Placebo.
Dönmez F., Tosunoğlu M., Gül C. (2009): Hematological values in hermaphrodite, Bufo bufo
(Linnaeus, 1758). North-Western Journal of Zoology 5: 97-103.
Dugdale D.C. (2011): Blood differential, A.D.A.M. Medical Encyclopedia.
http://www.nlm.nih.gov/medlineplus/ency/article/003657.htm; pristupljeno 8.1.2013.
Feidantsis K., Anestis A., Vasara E., Kyriakopoulou-Sklavounou P., Michaelidis B. (2012):
Seasonal variations of cellular stress response in the heart and gastrocnemius muscle of the
water frog (Pelophylax ridibundus). Comp Biochem Physiol A Mol Integr Physiol 162: 331-
339.
Fominykh A.S., Liapkov S.M. (2011): The formation of new characteristics in life cycle of
the marsh frog (Rana ridibunda) in thermal pond conditions. Zh Obshch Biol 72: 403-421.
Green M.R., Pastewka J.V. (1974): Simultaneous differential staining by a cationic
carbocyanine dye of nucleic acids, proteins and conjugated proteins II Carbohydrate and
sulfated carbohydrate-containing proteins. The Journal of Histochemistry and Cytochemistry
22: 774-781.
Gül C., Tosunoğlu M., Erdoğan D., Özdamar D. (2011): Changes in the blood composition of
some anurans. Acta Herpetologica 6: 137-147.
Guyton A.C., Hall J.E. (2006): Textbook of Medical Physiology. 11. izdanje. Elsevier
Saunders.
Hadji-Azimi I., Coosemans V., Canicatti C. (1987): Atlas of adult Xenopus laevis
hematology. Developmental and comparative immunology 11: 807-874.
Horobin R., Kiernan J. (2002): Conn's Biological Stains: A Handbook of Dyes, Stains and
Fluorochromes for Use in Biology and Medicine. 1. izdanje. Williams & Wilkins.
IHC World (2011a): Gill's Hematoxylin Protocol.
http://www.ihcworld.com/_protocols/counterstain_solutions/gill_hematoxylin.htm;
pristupljeno 10.10.2012.

LITERATURA
61
IHC World (2011b): Mayer's Hematoxylin Protocol.
http://www.ihcworld.com/_protocols/counterstain_solutions/mayer_hematoxylin.htm;
pristuupljeno 10.10.2012.
IHC World (2011c): Nuclear Fast Red Counterstain Protocol.
http://www.ihcworld.com/_protocols/counterstain_solutions/nuclear_fast_red.htm;
pristupljeno 19.9.2012.
IHC World (2011d): PAS (Periodic Acid Schiff) Staining Protocol.
http://www.ihcworld.com/_protocols/special_stains/pas.htm; pristupljeno 20.9.2012.
Keusch P. (2012): Schiff's Reagent, Objective: Test for Aldehydes. http://www.uni-
regensburg.de/Fakultaeten/nat_Fak_IV/Organische_Chemie/Didaktik/Keusch/D-ald_add-
e.htm; pristupljeno 14.1.2013.
Kiernan J.A. (2008): Histological and Histochemical Methods. 4. izdanje. Scion Publishing
Ltd.
Knotková Z., Doubek J., Knotek Z., Hájková P. (2002): Blood Cell Morpholody and Plasma
Biochemistry in Russian Tortoises (Agrionemys horsfieldi). Acta Vet. Brno 71: 191-198.
Kumar G.L., Kiernan J.A (2010): Education Guide Special Stains and H & E. 2. izdanje.
Dako North America.
Lester V. K., Taropley H. L., Latimer K.S. (2005): Small Mammal Hematology: Leukocyte
Identification in Rabbits and Guinea Pigs.
http://www.medirabbit.com/EN/Hematology/Cells/Leukocyte.htm; pristupljeno 12.1.2013.
Lisičić D. (2005): Hematologija poskoka, Vipera ammodytes (Serpentes, Viperidae).
Magistarski rad. Biološki odsjek, Prirodoslovno-matematičkoga fakulteta Sveučilišta u
Zagrebu
Lowe (1998a): Alcian Blue, Histopathology, University of Nottingham.
http://www.nottingham.ac.uk/pathology/protocols/alcblue.html; pristupljeno 26.9.2012
Lowe (1998b): Alkaline Phosphatase, Histopathology, University of Nottingham.
http://www.nottingham.ac.uk/pathology/protocols/alkphos.html; pristupljeno 27.9.2012.
Lowe (1998c): Periodic Acid, Schiffs, University of Nottingham.
http://www.nottingham.ac.uk/pathology/protocols/pas.html; pristupljeno 26.9.2012.

LITERATURA
62
Marracci S., Michelotti V., Guex G.D., Hotz H., Uzzell T., Ragghianti M. (2011): RrS1-like
sequences of water frogs from Central Europe and around the Aegean Sea: chromosomal
organization, evolution, possible function. J Mol Evol. 72 : 368-382.
Nelson, D.L., Cox M.M. (2008): Lehninger Principles of Biochemistry. 5 .izdanje. W.H.
Freeman and Company.
Pendl H. (2006): Morphological changes in red blood cells of birds and reptiles and their
interpretation. Israel journal of veterinary medicine 61: 1-13.
Piršljin J., Milinković Tur S. (2009): Diferencijalna krvna slika.
http://www.vef.unizg.hr/org/physio/files/diferencijalna_krvna_slika.pdf; pristupljeno
14.11.2012.
Reagena (2003): May-Grünwald-Giemsa Staining Solutions.
http://www.reagena.fi/www/en/products/pdf/instructions/MGG_eng.pdf; pristupljeno
19.12.2012.
Reece M. . (2012): Physiology of Self Renewal.
http://www.medicalsciencenavigator.com/physiology-of-self-renewal; pristupljeno 11.1.2013.
Santa Cruz Biotechnology (2012): Product book Basic Fuchsin (CAS 632-99-5).
http://www.scbt.com/datasheet-203731-basic-fuchsin.html; pristupljeno 6.1.2013.
Shigdar S., Cook D., Jonest P., Harford A., Ward A.C. (2007): Blood cells of Murray cod
Maccullochella peelii peelii (Mitchell). Journal of Fish Biology 70: 973–980.
Shigdar S., Harford A., Ward A.C. (2009): Cytochemical characterisation of the leucocytes
and thrombocytes from Murray cod (Maccullochella peelii peelii, Mitchell). Fish & Shellfish
Immunology 26: 731-736.
Sigma- Aldrich (2006): Naphthol AS-D Chloroacetate Esterase and α- Napthyl Acetate
Esterase (Procedure No.90).
http://www.sigmaaldrich.com/etc/medialib/docs/Sigma/General_Information/1/91.Par.0001.F
ile.tmp/90.pdf ; pristupljeno 24.9.2012.
Stains File (2012a): Carazzi's Alum Hematoxylin.
http://stainsfile.info/StainsFile/stain/hematoxylin/aluminum/carazzi.htm; pristupljeno
10.10.2012.
Stains File (2012b): Hematoxylin & Hematein.
http://stainsfile.info/StainsFile/dyes/75290.htm; pristupljeno 8.1.2013.

LITERATURA
63
Stains File (2012c): Lysochromes. http://stainsfile.info/StainsFile/theory/lysochrm.htm;
pristupljeno 17.9.2012.
Strober W. (2001): Wright-Giemsa and nonspecific esterase staining of cells. Curr Protoc
Immunol. App. 3C. doi: 10.1002/0471142735.ima03cs21.
Usmanova N.M., Litvinchuk S.N., Kazakova E.A., Kazakov V.I. (2010): Variation of BM224
microsatellite in green frogs of the genus Rana. Tsitologiia 52: 858-862.
Venkat C. (2010): Myelodysplastic syndrome. http://updates.clltopics.org/2126-
myelodysplastic-syndrome; pristupljeno 12.1.2013.
Vetlab (2013): Natt & Herrick's Stain for Direct counting of Avian and Reptilian Blood Cells.
http://www.vetlab.com/Natt%20&%20Herricks%20Procedure.pdf; pistupljeno 13.1.2013.
WebPath (2012): Anatomy-Histology Tutorials
http://library.med.utah.edu/WebPath/HISTHTML/MANUALS/SUDANF.PDF, pristupljeno
18.9.2012.
Weiss D. J. i Wardrop K. J. (2010): Schalm's Veterinary Hematology. 6. izdanje. Wiley-
Blackwell.
Wilson S., Felt S., Torreilles S., Howard A., Behan C., Moorhead R., Green S. (2011): Serum
Clinical Biochemical and Hematologic Reference Ranges of Laboratory-Reared and Wild-
Caught Xenopus laevis. J Am Assoc Lab Anim Sci (50): 635-640.