Embed Size (px)
Citation preview

일반과제(○), 보안과제( ) (과제번호 : PJ007181)
사람치료를 위한 신약 검정용 질환모델 가축개발에 관한
연구
(Development of animal disease models of the new
drug testing for human treatment)
국립축산과학원
농촌진흥청

- 2 -
< 완결과제 최종보고서 >
제 출 문
농촌진흥청 장 귀하
본 보고서를 “사람치료를 위한 신약 검정용 질환모델 가축개발에 관한 연구(PJ007181)”
과제의 결과물로 제출합니다.
연 구 수 행 기 간 : 2009년 1월 1일 ∼ 2011년 12월 31일
제1세부연구과제 : 당뇨질환 모델 돼지의 개발 및 생리학적 검정
제1협동연구과제 : 전사체 및 단백체 발현분석을 통한 질환모델 가축의 검정
제2협동연구과제 : 당뇨병의 기능해석과 활용 연구를 위한 형질전환 모델 생쥐 개발
2012. 2.
주 관 연 구 기 관 명 : 국립축산과학원 (직인)
주 관 연 구 책 임 자 : 양병철
연 구 원 : 이휘철, 김현미, 고응규, 황성수, 오건봉,
우제석, 정학재, 박진기, 장원경
제1협동연구기관명 : 건국대학교
제1협동연구책임자 : 이창수
연 구 원 : 박혜란, 손은숙, 한우리자랑, 권오철, 김남국
제2협동연구기관명 : 경북대학교
제2협동연구책임자 : 류재웅
연 구 원 : 배기범, 여형수, 김나리, 지영래, 이재웅,
이준구, 박서진, 박정묵

- 3 -
요 약 문
Ⅰ. 제 목
사람 치료를 위한 신약 검정용 질환모델 가축 개발
Ⅱ. 연구개발의 목적 및 필요성
1. 연구개발의 필요성
전 세계적으로 수천 종의 유전자 변형동물들이 가능성 있는 질환모델로 보고되어 있으나,
사람의 질환에 가장 유사한 일부의 동물들만이 실제로 전임상단계의 약효검증에 활용되고
있다. 즉, 바이오신약 후보물질의 효능은 현재까지 주로 질환동물 모델로써 설치류를 포함
한 소동물을 이용한 전임상검정을 실시하고 있으나, 당뇨병, 비만 및 동맥경화 등의 생물의
학적 모델로써 인간과 생리학적으로 보다 유사한 돼지가 주목되고 있다 (Turk 등, 2005).
지금까지 인간질환 모델동물로 가장 많이 이용되고 있는 실험동물은 마우스이다. 일반적
으로 마우스는 포유동물이면서 인간과 유전적으로 비슷하고, 면역체계 등 많은 부분에서
인간을 대변하고 있는 동물이다. 하지만, 당뇨병 관련 연구 분야에선 다시 생각해 볼 여지
를 2009년 Science지에 Frances Brodsky 박사에 의해 제기되었다. 요지는 인간에게는 있지
만 마우스에게는 없는 단백질이 혈당조절에 중요한 역할을 하며 당뇨병의 발병에 영향을
미친다는 내용으로 보고하였다. 또 인간의 경우 혈중의 포도당의 제거 즉 이들의 소모는
90%가 골격근에서 이루어지는 반면, 마우스의 경우는 대부분 간에서 일어난다. 이러한 당
을 소모시키는 기작에 있어 큰 차이는 신약개발 및 질병기작의 해석 등에 있어 반드시 고
려되어야 할 사항이다. 이와 같은 관점에 있어 돼지는 인간의 생리학 및 병리생리학적인
측면에 대해서 매우 훌륭한 모델동물이다. 인간과 돼지는 영양 요구조건과 소화계, 혈장지
질, 췌장의 발달 및 형태학적인 측면뿐만 아니라 대사와 내당 (glucose tolerance)에 있어
많은 유사한 점이 발견된다. 그래서 돼지는 당뇨병과 관련된 일반적인 심혈관생리학 및 병
리생리학의 연구에 대한 좋은 모델동물이기에 당뇨질환모델로써의 개발이 반드시 필요하
다.
2. 연구개발의 목적
지금까지 돼지를 이용한 당뇨 질환모델들은 대부분 streptozotosin이나 alloxan 등에 의한
외과적 처리로 췌장세포를 파괴시키는 방법에 의해 당뇨병이 유도되어지거나, 혹은 선택적
인 육종 방법 등에 의해 당뇨병의 표현형을 도출했다(Junod 등, 1969; Lenzen과 Paten,
1988). 최근에 형질전환 기술을 이용한 당뇨질환 모델 돼지의 생산은 일본의 메이지대학
Nagashima 교수팀과 독일의 Ludwig-Maximilians대학의 Kessler 팀에서 진행되고 있고,
국내에선 2009년 8월 황우석 박사팀이 당뇨질환 모델돼지를 개발하겠다고 발표한 것이 유

- 4 -
일한 상황이다. 본 연구의 목적은 당뇨질환에 대한 전임상 모델동물의 개발에 앞서, 당뇨병
유발 예상 유전자인 PEA15가 과발현되는 형질전환 동물을 개발하여 본 유전자와 당뇨병과
의 관계 및 기능해석, 당뇨질환 모델동물로써의 가능성 등 관련 연구 분야에 기초연구 형
질전환 동물을 공급하기 위해서이다. 또한 형질전환 기술을 이용한 질환모델 생쥐를 개발
하여 질환 발병의 유무를 검정하고, 유전자 재조합 세포배양계의 질환모델로서의 이용성을
검정한다. 전사체 및 단백체 발현양상을 통한 질환모델가축의 검정과 2형 당뇨병 모델동물
및 합병증 형질전환 모델생쥐를 이용한 발병원인 규명과 제어기능을 해석한다.
Ⅲ. 연구개발의 내용 및 범위
1. 형질전환 돼지 증식과 생리학적 분석
2. mPea15과발현 재조합세포주 구축과 당질대사시스템 특성
3. 장기 특이적 promoter 탐색과 유전자 발현 벡터 구축 및 당뇨병 형질전환 생쥐 생산
4. 조직학적 특성 확인과 신규발현 벡터 도입
5. 재조합세포주의 지질대사 시스템특성 및 질환모델가축 발현전사체 특성
6. 당뇨병 형질전환 생쥐 생산과 유전자 기능 분석
7. 신규당뇨질환 모델돼지의 개발 및 In vitro 시스템계의 확립
8. 재조합세포주의 이용성 검토 및 질환모델가축 발현전사체 특성
9. 형질전환 모델생쥐을 이용한 발병원인 규명과 제어기능 해석
Ⅳ. 연구개발결과
□ 당뇨질환 모델 돼지의 개발 및 생리학적 검정
기 생산된 Pea15 과발현 형질전환 돼지의 당뇨질환 모델로써의 활용 가능성에 대한 시험
분석을 실시한 결과, In vivo 당 부하 검사에서는 뚜렷한 당뇨질환을 관찰하지 못했지만,
이들 근육세포를 이용한 In vitro에서 당 소모능의 분석은 현저히 감소됨을 확인하였다. 이
는 질환모델의 도구로써의 활용가능성을 제시하였다. In vitro에서의 당 소모능 저하에 대
한 원인을 규명하고자 Glut4 당수송체의 막이동에 대한 분석을 조사한 결과, Pea15 과발현
에 의해 저해됨을 확인하였다.
□ 전사체 및 단백체 발현분석을 통한 질환모델 가축의 검정
Pea15 과발현 형질전환 돼지의 대사시스템 변화 분석 결과, 인슐린 무처리 세포주에서는
검출되지 않았던 pAkt는 인슐린 처리로 재조합유전자의 과발현과는 관계없이 그 발현량이
매우 높게 증가하였고 이는 Pea15 유전자의 과발현으로 인하여 세포내 당수송 능력의 감소
는 인슐린 비의존적인 경로를 경유하여 작용되었을 것으로 생각된다. 인간유래 간세포주에
Pea15 유전자를 도입한 후 24시간 48시간 후에 간에서의 중성지방 합성 정도를 확인한 결
과, 24시간 후 Pea15 과발현 세포주에서의 중성지방 합성율이 40% 증가 하는 것을 확인하
였다. 또한, 형질전환 돼지의 혈장 단백체에서 3개의 차등발현을 보이는 단백질을 확인하였
고 현재 근육조직에서의 대사변화를 규명하기 위하여 DNA chip 이용한 근육조직전체의 유

- 5 -
전자 발현양상을 분석 중에 있다.
□ 당뇨병의 기능해석과 활용 연구를 위한 형질전환 모델 생쥐 개발
Jazf1 형질전환 생쥐의 개발을 통하여 이들의 혈압이 정상에 비하여 높게 유지되었으며,
심초음파 실험 결과 심장박동이 불규칙함을 확인하였다. 심장조직의 모습 또한 정상 마우
스에 비하여 조직이 치밀하지 못하였으며, 심근세포의 미토콘드리아의 형태가 많이 망가져
있음을 확인하였다. 또한, CTSS 과발현 형질전환 생쥐에서는 지방분화에 관여하는 것으로
비만유도에 의한 합병증이 발병할 것으로 예상하였고 그 결과, 아토피가 발병되었다. 개발
된 2종류의 형질전환 생쥐의 돼지로의 개발에 검토 중에 있다.
Ⅴ. 연구개발결과의 활용계획
□ 당뇨질환 모델돼지의 산업화를 위한 지적 소유권 확보
□ 개발된 질환모델 돼지을 통하여 당뇨병 치료제의 후보물질의 효능 검정에 활용
□ 의료 및 제약업계와의 공동연구로 산업화의 조기달성
□ 당뇨질환 재조합 세포배양계를 이용한 당뇨치료 생약 및 화합물도출을 통한 화합물의
가치를 새롭게 창출하는 시스템 확립
□ 질환모델생쥐의 질환관련 연구를 통해 신규 유전자의 발굴과 질환모델 돼지의 개발에
기초자료 제공

- 6 -
S U M M A R Y
○ Title : Development of animal disease models of the new drug
testing for human treatment
The overexpression of Phosphoprotein Enriched in Astrocytes (PEA15) gene is
commonly found in human diabetic patients. The overexpression of this gene in skeletal
muscle and fat tissues have been reported to cause insulin resistance, thereby impairing
insulin stimulated glucose uptake. We introduced a gene of mouse Pea15 (mPEA15) and
enhanced green fluorescent protein (EGFP) into fertilized one cell pig zygotes using
microinjection, and produced a piglet that showed overexpression of mPea15 in the
muscle tissues and expression of EGFP in the ear tissues and hooves. RT-PCR RFLP,
southern blot and FISH analysis showed that the tissues carried the transgene. Real-time
RT-PCR and western blots demonstrated that Pea15 gene was overexpressed in the
various tissues and muscle tissues, respectively. These facts suggest that expression
vector system is normally expressed in the trnasgenic (TG) pigs.
PEA15 is a 15kD-sized intra-cellular signaling protein, highly expressed in cells of
neural system. Recently, PEA15 was found to be highly expressed in peripheral tissues
of type-2 diabetic patients and are suggested to be involved in the symptoms of insulin
resistance. Although diabetes mellitus is one of the most prevalent metabolic diseases,
development of new cure has been unsuccessful due to the ineffectiveness or severe
side effects. One reason explaining these failures clearly is a lack of appropriate model
animal for the pre-clinical trial. To tackle this problem, we produced a transgenic pig,
over-expressing diabetes-related mPEA15 gene. As a model animal, pig has many
advantages. They produce many offsprings, has a short generation time, and
physiologically similar to human. Using the transgenic pig, we carried out a series of
experiments to establish a link between mPEA15 expression and the insulin resistance.
Our results shows that the transgenic pigs express two immuno-detectable proteins. We
believe the one sized smaller is the endogenous PEA15 and another bigger sized protein
is the introduced mouse PEA15. Among diabetes-related endocrine factors, adiponection,
resistin, insulin receptor, and glucose channel (GLUT4) were investigated using several
biochemical tools. In cases of adiponection, insulin receptor, and total GLUT4, we could
not claim any difference against wild type. In contrast, resistin was elevated in the
blood of mPEA15 pigs. In separate experiments, skeletal muscle cells were treated with
insulin and subsequently the GLUT4 distribution and glucose uptake were analyzed to
prove the involvement of mPEA15 on the GLUT4 translocation. It turns out that both

- 7 -
plasma membrane-embeded GLUT4 level and glucose clearing ability were lower in
mPEA15-expressing pigs and were not elevated by insulin treatment. Finally, by using
oral glucose tolerance test (OGTT), we pursued the blood glucose profile in mPEA15
expressing pigs in vivo. It was found that blood glucose level was elevated after oral
adminstration of glucose solution in both wild-type and transgenic pigs. However, the
blood glucose level was maintained significantly higher in transgenic pigs for 120
minutes. When our all results combined, it can be concluded that, mPEA15 expressing
pigs cannot accelerate glucose uptake in response to insulin treatment due to the
blockage of PKCξ/λ-mediated signaling which is necessary for the activation of GLUT4.
The pathogenesis of type 2 diabetes is characterized both by insulin resistance in
muscle, fat and liver and by impaired insulin secretion. The elevated level of fat is
responsible for the impairment of glucose utilization and a decrease of insulin
sensitivity, which both contribute to the establishment of insulin resistance. Because of
the importance of insulin resistance and fatty acid abnormalities in the development of
type 2 diabetes, many investigators have confirmed that dyslipidemia, hyperglycemia,
and insulin resistance can aggravated each other, which means they are involved in a
vicious circle. Overexpression of the ped/pea-15 gene is a common feature of type 2
diabetes. In the present work, ped/pea-15 mRNA was overexpressed by transfection
assay in hepatocyte and skeletal muscle cells, two major sites of insulin resistance in
type 2 diabetes. Transfection of ped/pea-15 in hepatocyte changed lipid metabolism as
part of the underlying mechanism for insulin resistance, especially the accumulation of
triglyceride, and L6 skeletal muscle cells decreased the uptake of [14C]
2-deoxy-D-glucose. Overexpression of the PED/PEA15 gene may effected to insulin
resistance in accumulation of triglyceride and glucose uptake in type 2 diabetes.
Although the human hepatoma cell line HepG2 is widely used as an in vitro model
human hepatic lipid metabolism. This study first evaluated the impact of overexpression
of the ped/pea-15 gene on TAG synthesis by transfection into HepG2 cells for 24 or 48
hour. When the cells were transfected in the presence of ped/pea 15 gene expression
vector with radio-labeled acetate, theses transfected cells showed 40% (for radio-labeled
acetate) increase against cellular TAG synthesis.
The myotubes were then trnsfected with pea/ped 15 gene expression vector for 24
hour. After appropriate incubation, uptake of 2-deoxy-D-glucose (2DG) was determined
during a further 15min incubation period. To determine transfected cells affect glucose
uptake in myotubes, we measured 2-deoxy-D [1-14C]glucose uptake in the different
experimental groups. The transfected cell glucose uptake was decrease in the presence
of insulin that showed 40%.
Transgenic a piglet that showed overexpression of mPea15 in the liver and muscle

- 8 -
tissues. RT-PCR microarray analysis showed that the overexpression of mouse ped/pea
15 gene. Microarray demonstrated that Pea15 gene overexpression transgenic pig was
type 2 diabetes in the various metabolic pathway change.
Proteomic technology might provide the new solutions for solving this problem, which
can identify large set of the proteins in cells or tissues through high-throughput
methods, and provide a globe view of the protein changes associated with
diabetes(Rong-Xia Li, et al. 2008). It is well known that serum severs the optimal
resource for discovery of disease biomarkers. In the present study, we used
Q-TOF-MS/MS to investigate the protein comparison between the wild type pig(WT)
and ped/pea 15 gene overexpression transgenic pig(TG) serum without removing the
high abundant proteins. After analyzing 3 proteins were identified. In this study, many
individual proteins associated with T2DM reported in previous studies were also
identified. In the protein of ficolin-2precursor. On the other hand, we identified down
regulation protein, apolipoprotein A.
We selected candidated genes to make a transgenic model mice of Type 2 diabetes
and complication. We constructed Biliverdin reductase (BVR), Cathepsin S (CTSS) and
JAZF zinc finger 1 (Jazf1)-expression vectors and producted the transgenic model mice
by using confirmed vectors. The confirmed transgenic mice were investigatived
metabolic dysfunction and identified that these model mice is useful as Type 2 diabetes
model animal. The transcription factor Juxtaposed with another zinc finger gene 1
(JAZF1) is a zinc finger protein that binds to the nuclear orphan receptor TR4. Recent
evidence indicates that TR4 receptor functions as both a positive and negative regulator
of transcription, but the role of JAZF1 in transcriptional mechanisms has not been
elucidated. Recently, the incidence rate of congenital heart malformations was reported
to be significantly elevated in patients who had neurofibromatosis 1 (NF1) with
chromosomal microdeletion syndrome. Furthermore, Joined to JAZF1 (SUZ12) is
expressed at high levels in the hearts of adult patients with NF1 microdeletion
syndrome. Therefore, we hypothesized that ectopic expression of JAZF1 may lead to
cardiac malformations that deleteriously affect the survival of neonates and adults. We
sought to elucidate the role of JAZF1 in cardiac development using a
Jazf1-overexpressing (Jazf1-Tg) mouse model. In Jazf1-Tg mice, Jazf1 mRNA expression
was significantly elevated in the heart. Jazf1-Tg mice also showed cardiac defects, such
as high blood pressure, electrocardiogram abnormalities, apoptosis of cardiomyocytes,
ventricular non-compaction, and mitochondrial defects. In addition, we found that the
expression levels of pro-apoptotic genes were elevated in the hearts of Jazf1-Tg mice.
These findings suggest that Jazf1 overexpression may induce heart failure symptoms
through the upregulation of pro-apoptotic genes in cardiomyocytes.

- 9 -
목 차
제 1 장 서 론 ················································································································· 8
제 1 절 연구개발과제의 개요 ······················································································ 8
제 2 절 연구적 배경 ········································································································ 11
제 2 장 국내외 기술 개발의 현황 ··········································································· 14
제 1 절 연구개발대상 기술의 국내·외 현황 ···························································· 14
제 2 절 국내·외의 연구현황 ························································································ 16
제 3 장 연구개발수행 내용 및 결과 ······································································· 17
제 1 절 형질전환 돼지의 검정 ···················································································· 17
1. 목적유전자의 삽입여부의 확인 ··········································································· 17
2. PEA15의 발현 검정 ······························································································ 20
제 2 절 생리학적 검정 (in vivo) ··············································································· 24
1. 당 부하검사 (Glucose tolerance test) ······························································· 24
2. 생화학적 지표 및 혈중 대사성 호르몬 레벨 ··················································· 28
3. 근위성 세포의 분리 및 배양법 확립 ··································································· 31
제 3 절 PEA15 과발현 세포의 In vitro 검정 ······················································· 34
1. PEA15 과발현 근육세포의 특징 ········································································ 34
제 4 절 유전자재조합 세포배양계의 질환모델로서의 이용성 검정 ···················· 49
1. 재조합세포주 구축 및 당뇨질환과의 연관성 검정 ········································· 49
2. 근육세포주를 이용한 재조합세포주 구축 및 당뇨질환과의 연관성 검정 · 52
제 5 절 전사체 및 단백체 발현양상을 통한 질환모델가축의 검정 ·················· 54
1. 유전체 및 단백체의 발현 특성 ··········································································· 54
제 6 절 질환모델가축의 개발을 위한 기초연구 ···················································· 65
1. 연구배경 ··················································································································· 65
2. 돼지근육 미토콘드리아 단백체 발현특성 ························································· 65
3. 당뇨모델 생쥐의 차등발현 단백체 분석 ····························································· 66
제 7 절 질환모델 형질전환 생쥐의 개발 ································································ 72
1. 형질전환 생쥐의 생산 및 특징 ··········································································· 72
제 4 장 연구개발목표 달성도 및 대외 기여도 ······················································ 86
제 5 장 연구개발결과의 활용 계획 ··········································································· 91
제 6 장 중요 변동사항 ·································································································· 93
제 7 장 참고문헌 ············································································································ 94

- 10 -
제1장 서 론
제1절 연구개발과제의 개요
1. 연구개발의 필요성
지난 수 십년간 전 세계적으로 급격히 증가하고 있는 비만 및 대사질환 유병율은 인류의
건강을 위협하는 요인이 되고 있다. 또한 대사질환은 노령화, 식습관의 변화, 환경의 오염
및 유전적 요인에 의해 발병되는 것으로 보고되고 있다. 당뇨병의 90~95%는 비만, 운동부
족 및 과도한 영양섭취에 따른 지속적인 체내 에너지대사의 불균형으로 초래되는 성인형
당뇨병인 제 2형 당뇨병으로써 세계 각국에서는 성인형 당뇨병에 대한 새로운 치료제 개발
및 병인연구에 연구를 집중하고 있는 실정이다 (Kopelman, 2000). 2008년 한국의 사회지표
조사결과에 따르면 우리나라 국민 사망원인 1위는 악성신생물(암)으로 인구 10만 명당
139.5명이 사망하였으며, 다음은 뇌혈관질환(56.5명), 심장질환(43.4명), 당뇨병(20.7명), 고혈
압(9.6명) 순으로 나타남. 이는 1998년도 대비, 암은 23%, 당뇨병은 36% 증가한 수치로, 우
리나라 전체 인구의 30%가 이들 만성질환을 앓고 있는 것으로 나타났다 (통계청). 현재 비
만과 당뇨병 등의 대사질환은 만성질환으로 치료적 접근이 완치보다는 질병관리에 제한되
어 있으며, 전 세계적으로 의료비의 지출을 상승시키고 노동력의 저하를 가져오는 상태로
이를 예방할 수 있는 새로운 접근 방식이 필요하다.
세계적으로 질환치료용 바이오신약 개발에 많은 인력과 연구비를 투자하고 있고, 2015년
이들의 세계시장 규모는 2,112억$, 국내시장 규모는 13,529억원으로 추산되고 있으며 꾸준
히 증가하는 추세이다. 특히 당뇨병 치료제 세계시장 규모는 603억$, 국내는 4,218억원으로
추정된다(생명공학정책연구센터 분석 ‘07년). 전 세계적으로 수천 종의 유전자 변형동물들
이 가능성 있는 질환모델로 보고되어 있으나, 사람의 질환에 가장 유사한 일부의 동물들만
이 실제로 전임상단계의 약효검증에 활용되고 있다. 즉, 바이오신약 후보물질의 효능은 현
재까지 주로 질환동물 모델로써 설치류를 포함한 소동물을 이용한 전임상검정을 실시하고
있으나, 당뇨병, 비만 및 동맥경화 등의 생물의학적 모델로써 인간과 생리학적으로 보다 유
사한 돼지가 주목되고 있다(Turk 등, 2005).
지금까지 인간질환 모델동물로 가장 많이 이용되고 있는 실험동물은 마우스이다. 일반적
으로 마우스는 포유동물이면서 인간과 유전적으로 비슷하고, 면역체계 등 많은 부분에서
인간을 대변하고 있는 동물이다. 하지만, 당뇨병 관련 연구 분야에선 다시 생각해 볼 여지
를 2009년 Science지에 Frances Brodsky 박사에 의해 제기되었다. 요지는 인간에게는 있지
만 마우스에게는 없는 단백질이 혈당조절에 중요한 역할을 하며 당뇨병의 발병에 영향을
미친다는 내용으로 보고하였다. 또 인간의 경우 혈중의 포도당의 제거 즉 이들의 소모는
90%가 골격근에서 이루어지는 반면, 마우스의 경우는 대부분 간에서 일어난다. 이러한 당
을 소모시키는 기작에 있어 큰 차이는 신약개발 및 질병기작의 해석 등에 있어 반드시 고
려되어야 할 사항이다. 이와 같은 관점에 있어 돼지는 인간의 생리학 및 병리생리학적인

- 11 -
측면에 대해서 매우 훌륭한 모델동물이다. 인간과 돼지는 영양 요구조건과 소화계, 혈장지
질, 췌장의 발달 및 형태학적인 측면뿐만 아니라 대사와 내당(glucose tolerance)에 있어 많
은 유사한 점이 발견된다. 특히 돼지에서 자연발생적으로 발생되는 것으로 알려진 아테롬
성 동맥경화증(atherosclerosis: 대동맥 같은 탄력성 동맥의 내층에 지방과 콜레스테롤로 이
루어진 층이 생기는 병태)은 인간에게서 나타나는 현상과 매우 유사하다. 즉, 돼지는 당뇨
병과 관련된 일반적인 심혈관생리학 및 병리생릭학의 연구에 대한 좋은 모델동물이다.
2. 연구개발의 목적
지금까지 돼지를 이용한 당뇨 질환모델들은 대부분 streptozotosin이나 alloxan 등에 의한
외과적 처리로 췌장세포를 파괴시키는 방법에 의해 당뇨병이 유도되어지거나, 혹은 선택적
인 육종 방법 등에 의해 당뇨병의 표현형을 도출했다(Junod 등, 1969; Lenzen과 Paten,
1988). 최근에 형질전환 기술을 이용한 당뇨질환 모델 돼지의 생산은 일본의 메이지대학
Nagashima 교수팀과 독일의 Ludwig-Maximilians대학의 Kessler 팀에서 진행되고 있고,
국내에선 2009년 8월 황우석 박사팀이 당뇨질환 모델돼지를 개발하겠다고 발표한 것이 유
일한 상황이다. 본 연구의 목적은 당뇨질환에 대한 전임상 모델동물의 개발에 앞서, 당뇨병
유발 예상 유전자인 PEA15가 과발현되는 형질전환 동물을 개발하여 본 유전자와 당뇨병과
의 관계 및 기능해석, 당뇨질환 모델동물로써의 가능성 등 관련 연구 분야에 기초연구 형
질전환 동물을 공급하기 위해서이다. 또한 형질전환 기술을 이용한 질환모델 생쥐를 개발
하여 질환 발병의 유무를 검정하고, 유전자 재조합 세포배양계의 질환모델로서의 이용성을
검정한다. 전사체 및 단백체 발현양상을 통한 질환모델가축의 검정과 2형 당뇨병 모델동물
및 합병증 형질전환 모델생쥐를 이용한 발병원인 규명과 제어기능을 해석한다.

- 12 -
제2절 연구적 배경
1. PEA15의 특징
Phosphoprotein Enriched in Astrocytes (PEA15) 유전자는 제 2형 당뇨 환자에게서 높은
수준으로 발현된다. PEA15 유전자는 다양한 조직에서 광범위하게 발현되며, 세포내 중요한
phosphoprotein이다. PEA15는 human chromosome 1q21-22dp에 위치하고 있는 15kDa의
세포질 단백질이다 (Condorelli 등, 1998). 그것은 고도의 조절 단백질이며, 2가지 중요한 인
산화 부위인 Ser104와 Ser116을 포함하고 있다 (Fiory 등, 2009). 특히, Ser104은 protein
kinase C (PKC)에 의해 인산화 되고, Ser116은 cacium-calmodulin kinase Ⅱ (CaMKⅡ)이
나 AKt/PKB에 의해서 인산화 된다 (Kubes 등, 1998; Terencia 등, 2003). 여러 연구 중,
PEA15는 세포 신호경로의 중요한 조절 인자로서 보고되었다. 이런 PEA15는 세포질 골격
단백질이며, 주요 세포 내 여러 signaling에 의해 특정 성분에 결합함으로써 세포 내 다양
한 기능들을 조절하며 (Trencia 등, 2003; Formstecher 등, 2001; Vaidyanathan 등, 2003;
Vigliotta 등, 2004; Zhang 등, 2000), 그리고 PEA-15는 death effector 도메인(DED)을 포함
한다. 이 상호적인 도메인을 통하여, PEA15는 세포사멸 신호(apoptotic signals)와 길항작용
을 하며, 그 신호를 핵에 전달하는 많은 분자를 억제한다 (Trencia 등, 2003; Formstecher
등, 2001; Vaidyanathan 등, 2003). 또한 PEA15는 phospholipase D (PLD)에 결합하고,
PLD의 안정성을 증가 시킨다. PEA15에 안정화된 PLD는 다양한 signaling에 의해 당뇨를
유발시키게 된다 (Zhang 등, 2000). 다른 연구에서는 PEA15가 제 2 당뇨병 유형의 발달에
대하여 중요한 역할을 지지하는 직접적인 증거들을 설명해왔다.
2. PEA15와 인슐린 신호전달
대부분의 제 2 당뇨병 유형의 병리학적 증상은 혈액 내 인슐린이 존재할지라도 말초조직
의 포도당 흡수의 감소를 설명할 수 있다. 일반적으로 인슐린 저항성으로 불리는 병리 생
리학적 현상은 PEA15의 과 발현과 PKCα의 활성, 그리고 포도당 흡수를 위한 인슐린 신호
의 세포 내 중개자인 PKC ξ/λ의 억제와 같은 여러 가지 세포 내 신호체계를 수반한다
(Condorelli 등, 2001).
세포의 인슐린 저항성에 대한 현재 제안된 모델 시스템은 이 세 가지 성분 모두를 포함
하고 있다. PEA15는 아직 정확히 확인되지는 않았지만 과발현이 되었을 때, PKCα을 활성
시키고 이후 활성을 띈 PKCα은 비정형적인 PKC ξ/λ을 억제한다. 이로 인해 말초조직의
포도당 흡수는 매우 감소하였다 (Condorelli 등, 2001). 췌장의 베타세포에 의한 인슐린 분
비는 제 2 당뇨병 유형의 환자에서 부분적으로 손상되었기 때문에, 인슐린 저항은 인슐린
분비의 실패만으로는 설명 되어질 수 없다. 그러나 인슐린 분비 결핍은 당뇨 증상에 크게
기여하기 때문에 극복하기는 어렵지만, 당뇨병의 경우 인슐린 비의존형
(insulin-indepenent)이거나 결핍된 인슐린 의존형 (non-insulin dependent)으로 특징지어져
왔다. 그 중 제 2형 당뇨병은 인슐린 비의존형에 해당된다. PEA15와 PLD의 약리적 억제를
통해 PEA15를 과발현하는 세포 내에서 PKC ξ/λ 기능의 구조(rescue)는 포도당 수송을 평
범한 인슐린 분비로 수복한다 (Vigliotta 등, 2004). 그러므로 배양된 근육과 지방세포 내에

- 13 -
PEA15는 PKC α에 의해 PKC ξ/λ 기능의 일반적인 조절을 손상시킴으로써 포도당 투여
시 췌장 내 인슐린은 분비되나 세포 내에 이러한 기작에 의해 인슐린 저항성이 발생하게
된다. 현재까지 밝혀진 바에 따르면, atypical PKCs ξ과 λ은 골격근과 지방조직 내에 세포
외 인슐린에 반응하여 포도당 수송 기구를 활성 시키는 중요한 하류(main downstream)
effectors라는 것을 확인하였다 (Farese 등, 2002). 인슐린에 의해 발생되는 atypical PKCs
의 활성은 제 2 당뇨 동물 모델과 제 2 당뇨병의 환자에게서는 PKCs의 활성이 억제된다
(Farese 등, 2002; Kanoh 등, 2000; Kanoh 등, 2001). 결정적으로, PEA15의 과발현은 인슐
린 신호를 저해하고 이로 인해 인슐린 저항성이 발생하게 되는 것이다 (Gerolama 등,
1998). PEA15를 제외하고, 당뇨병 환자에서 인슐린 저항에 기여하는 다른 단백질의 발현은
insulin receptor kinase inhibitor 에 관여하게 된다 (Maddux 등, 1995; Myoers 등,1996).
3. PEA15와 당수송체(GLUT4)
더욱이 최근에 들어 근육세포에서의 PEA15 과발현으로 인해 포도당이 glucose
transporter type 4(GLUT4)를 통한 세포막으로 이동이 억제되는 것으로 확인되었다
(Condorelli 등, 1998). 혈액 내에서 포도당은 facilitative transporters의 한 계열인 막을 통과
하는 포도당 채널 단백질인 GLUT를 통하여 이동하였다. 세포 내에서 GLUT는 세포질과
막에 분산되어있다. 그러나 세포막에 위치한 GLUT만이 포도당 채널로서 기능을 한다. 세
포가 인슐린에 의해 자극되면, 세포질 GLUT를 저장하는 소낭이 세포막으로 융합되고 더
많은 GLUT가 세포막에 융합 할 수록 세포 내로 더 많은 포도당을 흡수 할 수 있다.
GLUT는 ATP-비의존적 촉진 확산 기작을 통해 세포막을 가로질러 육탄당 수송을 촉매화
한다 (Hruz와 Mueckler, 2001). GLUT4는 사람의 유전자에서 발현된 13개의 당 운송체 단
백질(GLUT1-GLUT12, and HMIT) 중 하나이다 (Joost와 Thorens, 2001; Wood와
Trayhurn, 2003). 골격근과 지방조직에서 일반적으로 GLUT 계열의 여러 단백질들을 발현
하지만 GLUT4 포도당 수용체의 경우 인슐린에 의해 변형된 주요한 regulatable isoform으
로 여기고 있다. 근육과 지방조직에서 GLUT4의 낮은 수준은 당뇨병에 관련된 인슐린 저항
성과 같은 성향을 수반하고 있다 (Rossetti 등, 1997; Stenbit 등, 1997; Li 등, 2000). 그래서
근육과 지방조직에서 GLUT4는 일반적인 포도당 항상성을 위해 반드시 필요하며, 반면에
이런 근육과 지방조직에서의 인슐린 수용체는 GLUT4에 비해 덜 중요하게 작용된다
(Huang 등, 2007). 포도당 수송율은 세포 내 저장 소낭에서 세포막으로 GLUT4 유닛을 축
적하거나 세포 표면 수송체의 활성을 통해 자극받을 수 있다 (Condorelli 등, 1998).
여러 자료에 따르면, PEA15 과 발현은 제 2 유형의 당뇨병에서 인슐린 저항에 기여 할
수 있을 것이라 제안하였다 (Condorelli 등, 2001; Vigliotta, 등, 2004). PEA15의 발현은 제
1 당뇨병의 섬유아세포로 변환되지 않지만, 제 2 당뇨병 섬유아세포에서 PEA15의 과발현
은 배양된 상태의 최소한 20 세대를 통해 지속된다. 이러한 과정은 환자의 내부환경으로
기인하는 이차 전환이 아니라는 것을 보여주고 있다. 높은 혈당수준과 당뇨병성 망막증, 당
뇨성 신장증 그리고 당뇨성 신경병과 같은 다양한 합병증에 의해 특징을 지닌 진성 당뇨병
은 어느 정도 심각한 만성질병에 따라 현대 사회에서 가장 문제가 많은 질병 중 하나로 간
주되어 진다 (Wild 등, 2004). 자가 면역성 질환인 제 1 유형의 당뇨병(5%)의 소수 경우를

- 14 -
제외하고는, 비만과 인슐린 저항과 연관된 당뇨병의 대다수 경우는 제 2 유형의 당뇨병
(95%)이 이에 속한다 (Rees 등, 2004).
4. 당뇨병관련 유전자
당뇨병과 관련된 호르몬중 하나인 Adiponectin은 에너지 항상성과 포도당 그리고 지질
대사의 조절에 중요한 역할을 한다 (Maeda 등, 1996; Scherer 등, 1995; Hu 등, 1996).
Adiponectin은 지방세포 분화과정을 증가시키는 지방세포의 발현으로부터 분비된 물질이다.
Adiponetin의 감소된 혈청 수준은 당뇨병, 이상 지질증 그리고 고혈압이 일반적인 특징으로
나타난다 (Stefan 등, 2002; Lindsay 등, 2003; Diez 등, 2003). adipokine은 항-염증성, 항-
당뇨병성 그리고 항-아테롬의 특징을 나타낸다고 보고 된 바 있으며 (Ouchi 등, 1999;
Ouchi 등, 2000; Matsuda 등, 2002; Weyer 등, 2001), 근육과 간에 지질과 포도당 대사에
중심역할을 하는 것으로 알려졌다 (Ouchi 등, 2001; Lihn 등, 2005). 또 다른 중요한 호르몬
인 resistin는 지방세포가 분화하는 동안 유도 되어진다. 지방세포에서 발현되고 분비된
resistin 폴리펩티드는 동물의 순환계를 순환하며, resistin의 양은 유전적이거나 식습관으로
인한 비만에서 상당히 높다 (Steppan 등, 2001).
5. 유전자재조합 세포배양계 검정
간 조직은 생체대사를 위한 기본 에너지생산 및 대사가 이루어지는 기관으로 당뇨질환에
있어 간조직의 기능은 중요하게 여겨진다. 특히, 간에서의 지방대사변화는 당뇨질환 발병에
중요한 요인으로 여겨지며, 지방대사의 증가는 간에서의 중성지방생산을 증가시키며(Paul
등, 2005), 중성지방의 증가는 제 2형 당뇨발병의 중요한 원인이면서 병증이기도 하다
(Deborah 등, 1999). 이러한 이유로 PEA/PED15 유전자의 과발현으로 당뇨발병여부를 확인
하기 위해서 우선적으로 간세포주를 이용한 재조합세포주 구축 및 당뇨질환과의 연관성 검
정을 시도 하였다. PEA/PED15 유전자의 과발현으로 제 2형 당뇨가 유발되었다면, 재조합
세포주의 지질 대사시스템 변화로 제2 형 당뇨의 병증인 인슐린 저항성의 가장 큰 원인인
과도하게 생합성된 중성지방을 제공할 것이다. 본 실험에서는 재조합 간세포주에서의 중성
지방 증가된 생합성 변화를 확인하여 재조합세포주의 당뇨질환 모델로서의 가능성을 확인
한다. 제 2형 당뇨에 있어 중요 병증인 간에서의 중성지방 생합성 증가는 인슐린 저항성을
유도 하며, 인슐린 저항성에 의한 병증은 근육조직에서의 당 수송능력 감소로 확인 할 수
있다. 인슐린분비에도 불구하고 혈중의 당을 흡수하지 못하므로 혈중에는 높은수준의 당을
포함하고 있으나 조직 및 세포에는 당의 부족현상이 일어난다. 질병유발 유전자
(PED/PEA15)의 과발현으로 유도된 근육 세포내의 기능적 변화확인을 세포 안으로의 당
수송능력을 glucose uptake 실험을 통해 확인 하였다.

- 15 -
제2장 국내외 기술개발 현황
제1절 연구개발대상 기술의 국내·외 현황
1. 세계적 수준
질환모델 동물은 현재까지 마우스를 대상으로 가장 많이 연구되어지고 있으며, 신경계,
청각계, 시각계, 혈관계, 면역계, 암 및 대사질환 등 1997년까지 110개의 인간질환에 대한
마우스 유전자가 확인되었다 (Bedell MA, ' 97). 그리고 당뇨병 연구의 목적으로 형질전환
기술을 이용하여 특정유전자(Insulin receptor, IRS, Glucokinase, GLUT4 등)의 유전적 조
작을 통해 개발된 형질전환 마우스가 다수 있다. 한편 중추신경계 질환모델로써는 2008년
NIH에서 세계 최초로 Huntington' s disease(중추신경계 질환)의 형질전환 질환모델 원숭이
가 개발되었다. 당뇨유발 후보유전자의 과발현동물 모델을 비롯한 특정 유도물질이나 식이
로서 유도하는 모델들이 개발되어 있고, 질환유발 작용점(당뇨모델 후보유전자의 과발현,
당뇨유발물질처리)의 적합성을 입증하기위해 질환모델 동물을 사용하여 이들의 분자생물학
적, 병리학적인 특성을 분석하여 새로운 모델동물로서의 가능성을 제시하는 연구가 진행
중에 있다.
한편, 미국의 경우 deCODE Genetics, Navegenics 등이 마이크로어레이를 이용하여 대사
질환 및 합병증(고혈압, 당뇨병, 고콜레스테롤혈증, 동맥경화 등)과 같은 특정 질병에 걸릴
가능성을 예측하려는 시도가 있으나 이러한 질병과 유전체 관련 연구는 미흡한 실정이다.
또한, 형질전환 방법을 이용한 대사질환 및 합병증 모델동물의 개발 연구는 인슐린 저항성
을 감소, glucose의 감소, lipid disorder의 정상화 및 lower weight (induced satiety)의 분
자 및 세포 수준의 in vitro 대사질환 및 관련 합병증 연구가 활발히 진행되고 있고, 대사질
환 및 관련 합병증의 발병원인과 치료 기술 개발을 위한 병인 기전의 규명에 관련되는 형
질전환(knock-out, over-expression transgenic animal) 모델 마우스의 개발 연구는 그 이
상의 발전이 이루어지지 못하고 있다.
본 연구와 직접적으로 관련이 있는 유전자인 PED/PEA15 유전자는 제2형 당뇨질환을 갖
는 환자의 혈액과 근육조직, 지방조직에서 정상인보다 2배 이상의 발현증가를 나타나는 현
상을 확인 하였으며, 당뇨질환기작 연구에서 중요한 표지유전자로서 가치가 평가 받고 있
다.
2. 국내수준
국립축산과학원은 형질전환 기술을 이용한 동물생산은 국내에서 독보적인 위치를 차지하
며 세계적인 기술과도 대등한 위치에 있다. 최근엔 일본동경대학 의과학연구소와의 국제공
동연구(‘05~’07)를 통해 제2형 당뇨병 유발 예상유전자 (PEA15)가 삽입된 형질전환돼지를
개발하여 지속적인 연구가 진행되고 있다. 하지만, 질환모델 동물개발은 인간유전체사업을
비롯한 특정연구개발사업에서 일부 추진 중에 있으나 아주 미약한 수준이며 (인간유전체사

- 16 -
업단의 경우, 2단계에서 제작되는 transgenic 및 knockout mice가 수십 종에 불과함) 일본
의 Sankyo제약회사의 1/3수준에 불과하다. 국내에는 KIST, 한국생명공학연구원, 성균관의
대, 이대, 숙대, 포항공대, 경북대, 식약청 등을 중심으로 질환모델생쥐 생산 기술을 확보하
고 있지만, 국내에서 형질전환 기술을 이용한 당뇨질환모델 돼지 개발에 대한 연구는 거의
없는 실정이다.

- 17 -
제2절 국내·외의 연구현황
제2형 당뇨병은 다양한 원인을 동시에 갖는 대사성 질환으로서 인슐린 저항성이 발생하
는 기전은 활발한 연구에도 불구하고 아직까지도 명확히 밝히지 못하고 있는 실정이다. 일
본에서는 2005년 7월 당뇨병 치료를 위해서 인간의 면역유전자 (HLA-G)를 가지는 복제돼
지를 생산하였고, 이 복제돼지의 췌장을 당뇨환자에게 이식함으로써 질병을 치료한다는 목
적을 두고 있다. 그리고 메이지대학교 발생공학과 나가시마교수는 최근 2007년 8월에 제4
세대 복제돼지 생산에 성공하였다고 발표했으며, 이는 세계 최초이며 앞으로 이러한 기술
을 당뇨병 모델 돼지에 응용할 계획이라고 밝혔다. 그리고 Purdue 대학과 Indiana 대학은
공동으로 Ossabaw 섬 미니돼지가 가지고 있는 비만, Type 2 당뇨병을 일으키는 인슐린
저항성, 고혈압 등의 특징을 이용한 공동연구를 추진하였으며 (‘06. 3. 30, Purdue 대학
Newsletter), 네덜란드 Wageningen 대학 연구센터는 제 1형 및 2형 당뇨질환 모델돼지의
개발을 추진 중에 있다. 한편, 독일 뮌헨대학의 Eckhard Wolf 교수팀은 당뇨병 연구를 위
한 lentiviral vector를 이용해 새로운 형질전환 모델 돼지의 개발에 필요한 기술을 확립였
으며, 국내에선 유전자 미세주입법에 의해 생산된 제2형 당뇨병 유발이 예상되는 형질전환
돼지가 F5 세대까지 보유하고 있으며, 이들 개체들의 당뇨유발 유무에 대한 생리학적 검정
을 본 연구에 의해 추진되었다.
유전체 수준에서 유전자 발현을 조사하는 마이크로어레이, 프로테오믹스 등을 사용하여
인슐린 저항성 및 당뇨병 관련 바이오마커의 발굴이 활발하게 진행되고 있으며, 이미 상당
량의 유전자 발현 정보가 NCBI의 GEO, EBI ArrayExpress등에 저장되어 있기에 in silico
에서 인슐린 저항성 및 당뇨병 관련 바이오마커를 발굴하고 생물정보학적 기법을 사용하
여 이의 기능적 특성을 예측하려고 하는 연구도 시도되었다.

- 18 -
제3장 연구개발수행 내용 및 결과
제1절 형질전환 돼지의 검정
1. 목적유전자의 삽입여부의 확인
가. 실험방법
(1) 공시재료
국립축산과학원 동물바이오공학과에서 일본과의 국제공동연구 (‘05~’07)를 통하여 생산된
PEA15 과발현 형질전환 돼지의 후대를 사용하였으며, 랜드레이스 종으로 후대번식은 자연
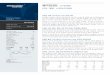
교배를 통하여 증식하였다. 그림1은 최초 F0 생산에 사용된 벡터이며, Pvu I으로 절단한
후 정제하여 유전자 미세주입에 이용했다.
pCAGGS-mPEA15
8559bp
mPEA15
chicken beta-actin promoter
rabbit b-globin polyA
SV40 ori CMV IE enhancer
SV40 promoter
IRES
Amp
SV40 poly A
bovine GH polyA
TK promoter
oriNeo
EGFP
Flag
PvuI (1)
XbaI (3326)
SalI(170
0)
SalI (34
64)
SnaBI (2058)
SnaBI(3830)
ApaI (2561)
ApaI(3967)
ApaI(5654)
BamHI (1694)
BamHI (6778)
BamHI (7115)
HindIII(4074)
HindIII(5421)
HindIII (6593)
StuI(599)
StuI(670)
StuI (6963)
PstI (828)
PstI (1025)
PstI (6588)
EcoRI (3422)
pCAGGS-mPEA15
8559bp
mPEA15
chicken beta-actin promoter
rabbit b-globin polyA
SV40 ori CMV IE enhancer
SV40 promoter
IRES
Amp
SV40 poly A
bovine GH polyA
TK promoter
oriNeo
EGFP
Flag
PvuI (1)
XbaI (3326)
SalI(170
0)
SalI (34
64)
SnaBI (2058)
SnaBI(3830)
ApaI (2561)
ApaI(3967)
ApaI(5654)
BamHI (1694)
BamHI (6778)
BamHI (7115)
HindIII(4074)
HindIII(5421)
HindIII (6593)
StuI(599)
StuI(670)
StuI (6963)
PstI (828)
PstI (1025)
PstI (6588)
EcoRI (3422)
그림1. pCAGGS-mPEA15 벡터의 구조
(2) Southern blotting
10㎍의 genomic DNA를 제한효소 Sph I (Toyobo, Japan) 으로 8시간 반응 시킨 후,
DNA Clean & Concentrator-25 (Zymo Research, USA)로 정제하였다. 그 다음 5㎍의
genomic DNA를 1% 아가로스 겔에 전기영동 (50V, 50mA)하였고, 10분간 Et-Br 염색 후,
순차적으로 탈염색 (15분), denaturation (15분 2회), neutralization (15분 2회)를 수행하였
다. Gel transfer는 삼투압 방식을 이용하여 over night로 nylon membrane (Amersham,
USA)에 흡착 시켰다. 흡착 시킨 membrane은 20X SSC 완충용액으로 세척한 후, UV

- 19 -
croslinker (Bio-RAD, USA)로 고정 시킨 후 probe를 표지하여 hybridization을 수행하였다.
Detection은 NBT/BCIP detection (Roche, USA)으로 발색시켜 필름에 현상하였다. Pea15
Probe DNA는 318bp의 PEA15 PCR 증폭산물 정제한 것을 이용하여 DIG DNA labeling
kit (Roche, USA)로 DIG-11-dUTP를 표지하여 50ng의 probe를 hybridization에 사용하였
다.
(3) Fluorescence In situ Hybridization (FISH)
Metaphase 상태의 백혈구는 헤파린 처리된 전혈에 10㎖의 배양액과 0.4㎖의 PHA-M를
넣고 38.5℃에서 prewarm 시킨 후, CO2 배양기에서 48~72시간 배양시켰다. 이후, 세포분
열을 중지시키기 위해 0.2㎖ colcemid solution 처리를 하고 30분 더 배양시켰다. 500g에서
5분간 원심분리 후 상청액을 제거하고 백혈구만 남을 때까지 0.075M KCl로 반복해서 적혈
구를 융해시킨 후, 차가운 fixative (metanol : acetic acid = 3 : 1)로 혈구를 고정시켜 도말
될 때까지 -20℃에 보관하였다. Probe는 pCAGGS-mPea15 벡터를 heat fragmentation법에
의해 100~600bp로 분절시켜 Direct labeling kit (Kreatech Biotechnology, Netherlands)를
사용하여 547 orang dye (Kreatech Technology, Netherlands)로 labeling하여 24×40 커버글
라스 당 20㎕의 probe mix (4ng/㎕)를 사용하였다.
백혈구 용액 30㎕를 슬라이드에 펼친 후 hot plate 위에서 10초간 가열하고 상온까지 온
도를 내린 후, 펩신처리 과정과 post-fix 과정을 통해서 고정시키고 1X PBS로 세척과
70%, 85%, 100%의 에탄올로 순차적으로 탈수 시킨 후, 건조시켰다. 이후 denaturation 시
킨 probe mix를 커버글라스에 도입하고 슬라이드글라스에 겹친 후 rubber cement로 밀봉
하였다. 이어서 80℃의 hot plate에서 5분간 동시에 denaturation 시키고 위쪽이 바닥으로
향하게 slide holder에 끼워 moist chamber에 넣은 후 42℃에서 12시간 hybridization 하였
다. Hybridization이 끝난 후, 커버글라스를 제거하고 1X Post-wash buffer1 (0.4X
SSC/0.3% NP-40)와 1X Post-wash buffer2 (2X SSC/0.1% NP-40)로 순차적으로 세척하
였다. 마지막으로 DAPI/Antifade Counterstain 용액 (Kreatech Biotechnology,
Netherlands)을 첨가 한 후 매니큐어로 커버글라스 주위를 밀봉하여 BX50 현미경
(OLYMPUS, Japan)으로 검경하였다.
(4) 통계처리
본 연구에 사용된 통계처리는 one-way ANOVA와 Student' s t-test를 이용하여 유의차
를 검증하였다. p<0.05일 때 유의차를 인정하였다.
나. 결과
PEA15 유전자가 삽입된 형질전환 돼지의 증식은 지속적으로 수행하였으며, 본 연구과제
수행에서 F3 및 F4 후대까지 증식하였다 (표1, 2). 이후 당 부하검사에 F4 후대를 사용하
였으며, 근위성 세포의 분리를 위한 근육조직 시료도 본 후대를 활용하였다.

- 20 -
표1. PEA15 형질전환 돼지의 F3 후대증식 현황
모돈수 총 자돈수 형질전환 자돈수 형질전환율
7수컷 암컷 수컷a 암컷b
47.7%40 25 18 (45%) 13 (52%)
a: 폐사돈 2두 포함, b: 폐사돈 1두 포함
표2. PEA15 형질전환 돼지의 F4 후대증식 현황
모돈수 총 자돈수 형질전환 자돈수 형질전환율
4수컷 암컷 수컷 암컷
68.7%21 14 15 (71%) 9 (64%)
유전자미세주입에 의해 생산된 F0의 목적유전자 삽입여부를 Southern blot에 의해 게놈
상의 삽입여부를 확인한 결과, 약 4kb의 위치에 밴드가 검출됨을 확인하였다 (그림2). 또한
그림XX는 mPEA15 유전자가 F1 후대로 전이되는지를 FISH법에 의해 확인한 결과이며,
TG의 염색체 휴지기 (a) 및 염색체의 양이 두배로 증가하는 중기 (b)에서 목적 유전자의
시그널이 검출되었다. 이와 같이 형질전환 개체의 검정은 다양한 방법으로 확인되었으며
후대로 안정적으로 전이됨을 확인하였다 (그림3).
그림2. Southern blot에 의한 mPEA15 유전자의 삽입유무의 확인 . (A) PEA15 과발현
벡터모식도, (B) Southern blotting 결과 4kb에 밴드 검출, (C) 후대개체의 검정결과. M:
size marker, PC: positive control, WT: wild type, TG: transgenic pig

- 21 -
그림3. FISH법에 의한 염색체상의 목적유전자의 존재 유무 확인 . WT: wild type, TG:
transgenic pig, a: probe signals in interphase, b: probe signal in metaphase.
2. PEA15의 발현 검정
가. 실험방법
(1) Real-time RT-PCR
혈액 및 조직에서 total RNA는 ZR Whole-Blood Total RNA Kit (Zymo Research, USA)
및 TRIzol reagent (Invitrogen, USA)를 이용하여 제공된 방법에 의해 각각 추출하였다.
cDNA는 1㎍의 total RNA를 1st Strand cDNA Synthesis Kit (Roche, USA)의 사용법에 따
라 최종 20㎕에 반응시켜 합성했다. RT-PCR은 TaKaRa PCR Thermal Cycler (TaKaRa,
Japan)을 사용하여, 최초 94℃에서 5분간 반응시킨 뒤 94℃에서 30초, 60℃에서 30초, 72℃에서 30초를 총 35회 반복하여 증폭하였으며, 최종 72℃에서 5분간 반응시켰다. PEA15 유전
자 증폭에는 forward primer인 5'-GGACATCCCCAGTGAGAAGA-3'과 reverse primer인
5'-TACGGGTTAGCTTGGTGTCC-3'을 이용하여 최종 50㎕에 반응시켜 318bp의 증폭산물
을 생산했다. mRNA 발현량은 LightCycler FastStart DNA Master SYBR Green I (Roche,
USA)를 이용하여 LightCycler (Roche, USA)로 분석했다. 최초 94℃에서 5분간 반응시킨
뒤 94℃에서 15초, 60℃에서 15초, 72℃에서 15초를 총 30회 반복하여 증폭하였으며, 최종
72℃에서 5분간 더 반응시켰다. PCR 반응 종결 후 melting curve 작성을 통하여 유전자 증
폭의 정확성을 재확인하였다. 유전자 발현량의 내부 보정을 위하여 house keeping 유전자인
돼지의 GAPDH (glyceraldehyde-3-phosphate dehydrogenase)를 이용하였고, 유전자 발현량
은 2-△Ct 방법을 통하여 분석하였다. 여기서 Ct값은 반응을 일으키는 최소 역가
(threshold)의 cycle 수를 의미하며, △Ct 값은 Pea15 유전자의 Ct 값에서 GAPDH 유전자의
Ct값을 뺀 값을 의미한다 (Livak와 Schmittgen, 2001). PEA15의 증폭 primer는 위와 동일하
며, GAPDH는 forward primer인 5'-ATTGCCCTCAACGACCACT-3'과 reverse primer인
5'-GGCCTCTCTCCTCCTCGC-3'을 이용하여 194bp의 PCR 증폭산물을 증폭시켰다.

- 22 -
(2) W estern blotting
-80℃에서 보관 중인 조직을 protease inhibitor cocktail (Roche, USA)이 첨가된 M-PER
단백질 추출 용액 (Thermo Scientific, USA)에서 homogenizer를 이용 분쇄하였다. 얼음에
서 1시간 동안 방치하여 추가로 조직을 용해하였다. 원심분리기를 이용 조직 단백질만을
회수하여 분석 전까지 -80℃에서 보관하였다. 단백질의 농도는 Bradford assay 용액
(Bio-RAD, USA)을 이용하여 결정하였다. 40㎍의 단백질을 SDS-PAGE 겔에서 분리시킨
후 PVDF membrane으로 전이시켰다. Western blot 분석을 위해 blocking buffer는 5%
non-fat dried milk와 0.05% Tween-20이 첨가된 PBS 용액을 사용하였다. 1차 항체는
anti-rabbit polycolonal Pea15 (Cell Signaling Technology, USA)를 사용하였고, 2차 항체
는 anti-rabbit IgG-HRP (Santa Cruz, USA)를 사용하였다.
(3) Immunohistochemical analysis
PEA-15를 Immunohistochemical 분석하기 전에, 준비된 paraffin block 조직을
3-aminopropyl triethoxysilane (TESPA, Sigma)-coated glass slides에 5 mm정도로
paraffin section 한 후에 37°C에서 건조 시켰다. Immunohistochemistry는 EXPOSE Mouse
and Rabbit Specific HRP/DAB Detection IHC kit (abcam, England)의 방법에 따라
paragffin 조직을 면역 염색하였다. 즉, ethanol로 탈수시킨 파라핀 조직을Hydrogen
peroxide을 10분간 처리하였고, 5% non-fat dry milk에 0.05% Tween-20가 첨가된
blocking buffer를 통해 90분간 blocking 하였다. 그 후 1차 항체 PEA15 (1:100, Cell
Signaling Biotech. Inc.)와 flag (1:200, sigma) 항체를 1:200 비율로 사용하여 4℃
overnight 하였다. 2차 항체인 Goat polyclonal antibody against Rabbit IgG는 1:400 비율
로 1시간 동안 실온에서 정치하였다. PEA15를 확인하기 위해 50 mM Tris-HCl (pH 7.6)
가 첨가된 diaminobenzidine (DAB; Sigma)를 사용하여 발색시킨 후에 hematoxylin을 통해
조직의 핵을 염색하였다. 마지막으로 DAPI/Antifade Counterstain (Kreatech
Biotechnology, Netherlands) 용액을 첨가한 후 커버글라스로 밀봉하여 현미경을 통해 관찰
하였다.
나. 결과
본 벡터 시스템에 의해 mPEA15 유전자가 F1 후대 개체의 전 조직에서 과발현되는 것을
Real-time PCR을 통하여 확인하였다 (그림4). 그 발현의 차이는 조직에 따라 혹은 개체에
따라 차이는 보였지만, 정상군에 비해 최대 4×106배 까지 차이가 났다. 또한, 근육조직에서
의 endogenous pig Pea15 단백질은 예상대로 양쪽 모두에서 15kDa 부위에서 발현되었지
만, TG 돼지에서 약 7배정도 높게 발현되었고 (그림5), transgene인 mouse Pea15 단백질
의 발현은 TG 돼지에서만 15kDa 위쪽에서 강하게 발현되었다. 이것은 그림1의 벡터 구조
에서 알 수 있듯이 Pea15 cDNA 앞쪽에 Flag tagging에 의한 단백질이 융합되어 있기 때
문이다. 이러한 이유가 Real-time PCR에 의한 mRNA 발현 차이가 극단적으로 나타날 수
있음을 뒷받침한다. 또한 수술용 칼을 이용하여 약간의 귀 조직을 자른 후, crude한 상태의
조직을 형광현미경으로 관찰하였다. 그 결과 귀 조직에서 강하게 GFP의 발현이 관찰되었

- 23 -
을 뿐만 아니라, 발굽에서도 강하게 발현됨을 확인하였다 (그림6). 비록 본 연구에서는 이
외의 조직에서 GFP의 발현을 관찰하지 않았지만, Naruse 등 (2005)은 돼지의 발굽에서 선
명한 EGFP의 발현과 검시하는 동안 혈액과 간 등의 다른 기관에서도 희미한 발현을 관찰
하였다.
그림4. 다양한 조직에서의 PEA15 mRNA의 발현양상 . (A) WT에 대한 상대적인 양을
표시한 것임, (B) 본 분석에 있어 PEA15 및 GAPDH의 해리곡선을 표시함.
그림5. 형질전환 돼지의 근육조직에 있어 PEA15 단백질의 발현

- 24 -
그림6. 형질전환 돼지의 귀조직 및 발굽에서의 GFP 발현 확인 . A, B: Image under
bright field, C, D: Image under fluorescent, E, F: Image of photograph.
Immunohistochemistry 를 통해 muscle 과 liver, pancreas tissue에서 PEA15 유전자의 발
현을 확인해본 결과이다. 우선 실험에 앞서 H&E를 통해 조직의 형태를 관찰하였다 (그림
7A). 그리고 IHC를 통해 anti-PEA15 antibody를 사용하여 확인해 본 결과, Muscle liver,
pancreas tissue의 형질전환 돼지에서 일반 돼지에 비해 PEA15 유전자가 조직의 전체적인
부분에서 더 강하게 되는 것을 확인하였다. 또한 flag 유전자의 발현을 확인해 본 결과 형
질전환 돼지에서만 발현되고 있음을 확인할 수 있었다(그림7B).
그림7. H&E 염색에 의한 조직형태분석과 면역조직염색에 의한 PEA15 발현 확인. (A)
조직별 H&E 염색, (B) PEA15 및 flag 항체로 IHC 염색. 스케일바 : 200㎛
위 사실을 바탕으로 본 연구에 사용된 벡터 시스템 즉, chicken β-actin promoter와

- 25 -
CMV-IE enhancer로 구성된 CAG promoter는 mPea15를 형질전환 돼지의 전 조직에서 과
발현시킴을 확인하였다. 이러한 사실은 Pea15 유전자의 과발현에 의해 유도되는 인슐린 저
항성 (Vigliotta 등, 2004)이 특히, 당의 분해를 담당하는 근육조직에서 발생할 경우, 제2형
당뇨병을 유발 시킬 수 있다는 사실을 시사한다. 다음 절에서는 형질전환 돼지에 있어 당
뇨에 대한 생리학적 연구에 대한 결과와 당 부하 검사 (glucose tolerance test)등을 포함한
다양한 측면에서 분석한 결과를 바탕으로 당뇨질환 모델 돼지로써의 가능성에 대해서 서술
할 예정이다.
제2절 생리학적검정 (In vivo)
1. 당 부하검사 (Glucose tolerance test)
가. 실험방법
(1) Glucose tolerance test (당 부하검사)
Intravenous glucose tolerance test (IVGTT) 및 Oral glucose tolerance test
(OGTT)
당 부하검사는 두 가지 방법에 의해 실시하였다. 첫 번째는 경정맥에 카테터를 삽입하여
외부로 관을 노출시켜 채혈을 용이하게 하는 수술을 실시하여 IVGTT를 하였다. PEA15
과발현 형질전환 돼지군 (TG)과 일반돼지군 (WT)을 전날 절식을 시켜 다음날 0.5g/체중
(kg)의 포도당을 카테터를 통하여 정맥주사를 하였다. 주사 후, 시간대 별로 헤파린이 첨가
된 채혈 튜브에 채혈을 하였으며, 즉시 혈장을 분리하여 분석에 이용하였다. 또한 인슐린,
중성지방 및 유리지방산의 분석도 함께 하였으며, 모든 분석은 (재)서울의과학연구소에 의
뢰하였다.
두번째는 OGTT에 의한 이전에 분석 확인된 방법을 이용하여 실시하였다 (Kulkarni et
al. 1999; Bruning, et al. 1998). 실험 수행에 앞서 하루 전 돼지를 절식시켰고, glucose
solution (200g)을 형질전환 돼지와 일반돼지에게 경구주입기를 사용하여 직접 입에 주입하
였다. 그 후 혈액 sample은 glucose 투여 후 0, 5, 10, 30, 60, 120, 180, 그리고 240분 후에
채취하였고, 이 혈액 sample은 간이 혈당측정기 (Accu-Check, Roche)를 사용하여 분석하
였다. 또한 당뇨병 유발을 촉진하기 위해서 High-Fat diet를 하였고 IVGTT로 분석하였다.
(2) 통계처리
본 연구에 사용된 통계처리는 one-way ANOVA와 Student' s t-test를 이용하여 유의차
를 검증하였다. p<0.05일 때 유의차를 인정하였다.
나. 결과

- 26 -
(1) IVGTT
그림8은 형질전환 돼지 5두 및 일반돼지 2두를 이용하여 당 부하검사를 실시한 결과이다.
포도당의 정맥투여 하기 전 (0분), -30분과 5, 30, 60, 90, 120 및 180분에 각각 정맥카테터
를 통하여 채혈하였다. TG군에서 30분 이후 지속적으로 일반돼지보다 혈장 글루코스 레벨
이 높은 경향을 보였지만 유의한 차는 보이지 않았다.
그림8. PEA15 과발현 형질전환 돼지의 당 부하검사
(2) IVGTT 및 High-Fat diet
mPEA15 유전자가 과발현된 형질전환 돼지에서 당뇨병 발병의 유무를 검증하기 위해서
당부하검사를 실시하였다. 당 부하검사는 표준식이를 한 WT 및 TG군과 고지방 식이를 한
WT 및 TG군으로 나누어 실시하였다. 그림9 및 10은 개체별로 그 결과를 정리하였다. 표
준식이에 의한 혈당변화는 TG 및 WT군에서 거의 180분 시점에서 떨어졌지만, 그렇지 않
은 개체도 관찰되었다. 한편, 고지방식이에 의한 혈당변화는 대체적으로 180분 시점에서 높
게 유지되는 경향을 보였다. 하지만 개체간 변이차가 심하고 재현성을 확인하기 어려워 그
림 11과 12에서 평균값을 도출하여 그 경향성을 살펴보았고, 당 투여후 30분 및 180분 시
점에서 경향을 해석하였다. 그 결과, 혈당은 표준식이 보다 고지방식이에서 당 투여 후 180
분 시점에서 높았으며, 인슐린의 분비도 고지방 식이에서 낮은 경향을 보였다.

- 27 -
그림9. 표준식이에 의한 W T 및 TG군에 있어 개체별 당 부하검사
그림10. 고지방식이에 의한 W T 및 TG군에 있어 개체별 당 부하검사

- 28 -
그림11. 표준 및 고지방식이에 의한 W T 및 TG군의 당 부하검사 평균 결과
그림12. 표준 및 고지방식이에 의한 W T 및 TG군의 당 부하에 의한 인슐린 레벨
변화
(3) OGTT
형질전환 돼지 (n=3)와 일반돼지 (n=4)의 Glucose의 항상성을 확인해 보기 위해, 일반돼
지와 형질전환 돼지의혈액 내 glucose 수준을 glucose tolerance test 통해 확인해 보았다.
Oral Glucose Tolerance Test (OGTT)를 통한 혈당수준의 변화추이를 확인해 본 결과
glucose loading 전, 형질전환돼지와 일반돼지의 혈당 수준은 일반돼지에 비해 형질전환 돼
지가 약간 낮게 측정되었다 (wild-type : 78±4 mg/dI VS transgenic : 69±5 mg/dI).
Glucose solution loading 후 0, 5, 10, 30, 60, 120 그리고 240분에 혈액 내 혈당 수준을 측
정하였다. 일반돼지에서는 Glucose loading 후 5분에서 증가된 후 5분이 지나자 다시
glucose 투여 전의 수준으로 다시 감소 되었으며, TG의 경우 glucose 투여 후 일반돼지에
비해 천천히 증가되어 10분에서 높은 수준으로 나타났다. (5 min: WT 96±17 mg/dI VS
TG 86±18 mg/dI ; 10 min: WT 79±4 mg/dI VS TG 121±15 mg/dI). 그러나 형질전환 돼
지의 경우, glucose level이 10분에서 120분 동안 높은 수준으로 유지되었으며, 30분대에서
혈액 내 glucose 의 수준이 가장 높게 측정되었다. OGTT 결과 전반적으로 형질전환 돼지
에서 일반돼지에 비해 더 높게 나타나는 것으로 확인되었다 (그림13).

- 29 -
그림13. 경구투여에 의한 당 부하검사 . WT; n=4, TG; n=3, * P<0.05
2. 생화학적 지표 및 혈중 대사성 호르몬 레벨
가. 실험방법
(1) 생화학적 지표분석
혈중 비만 및 당뇨의 생화학적 지표가 되는 인자의 분석은 채취된 혈장에서 혈청을 분리
한 후 시료의 분석은 (재)서울의과학연구소에 의뢰하였다.
(2) ELISA 분석
일반돼지와 형질전환 돼지의 혈장에서 혈청을 회수한 후에, Resistin ELISA Kit
(EIAabTM,China)과 Porcine adiponectin, ADP ELISA Kit (EIAabTM,China)의 방법에 따
라 ELISA 분석하였다. 혈청 protein 농도는 Bradford assay 에 따라 측정 하였다.
Standard와 혈청sample을 37 ℃에서 resistin과 adiponectin 분석 방법에 따라 각각 60분 45
분간 정치하였다. substrate solution 90 ul을 각각의 sample과 standard에 첨가한 후에 37
℃에서 15~30분간 정치하였다. 마지막으로 stop solution을 첨가한 후 450 nM 흡광도에서
측정하였다 (Bio-Rad, model 550).
나. 결과
(1) 혈중 대사성 질환 표지 인자 레벨
혈장 글르코스 레벨은 overnight-fasting 군의 TG에서 다소 높았고, random-feeding 군
에서는 2차 분석 시 유의적으로 TG에서 높았다. 혈장 인슐린 레벨은 random-feeding 군의
2차 분석 시 낮은 경향을 보였다. 중성지방 및 유리지방산 (FFA)은 유의한 차이는 없었지

- 30 -
만 TG에서 모두 높은 경향을 보였다 (그림14).
그림14. PEA15 형질전환 돼지의 혈장내 글루코스 및 인슐린 레벨 (A)과 중성지방 및 유리지방산의 레벨 (B)
고지방 식이는 자유급식 및 절식시킨 군에서 모두 혈당이 높은 경향을 보였지만, WT과
TG군 간에서 차이가 없었다(그림15A). 인슐린 레벨은 자유급식군의 고지방 식이군에서 높
은 경향을 보였고, 절식군에서는 낮은 경향을 보였다(그림15B).
그림15. 자유급식 및 절식에 의한 글루코스 및 인슐린 레벨의 변화
Triglyceride의 레벨은 WT 및 TG간에는 차이가 없었지만, 자유급식군이 높은 경향을 보
였다(그림16A). 마찬가지로 Free fatty acid의 레벨 비슷한 경향이지만 유의적으로 높았다.

- 31 -
그림16. 자유급식 및 절식에 의한 중성지방 및 유리지방산의 변화
(2) Adipinectin 및 Resistin 혈중 레벨
ELISA를 통해 혈중 Adiponectin과 Resistin의 발현량을 확인해본 결과이다. Adiponectin
의 혈중 농도를 일반돼지와 형질전환 돼지에서 비교해 보았을 때 두 개체 집단에서는 약
1.5ng/ml 정도로 거의 차이가 없었다. Adiponectin은 #9-385 (transgenic pig, 7.09 ng/ml)
개체에서 혈중 농도가 가장 높게 발현되었고, #9-115 (wild-type pig, 2 ng/ml) 개체에서
혈중 농도가 가장 낮게 발현되었다 (그림17A). Resistin은 #9-280 (transgenic pig, 2887
pg/ml) 개체에서 혈중 농도가 가장 높게 발현되었고, 반면에 #9-115 (wild-type pig, 19.96
pg/ml) 개체에서는 가장 낮게 발현 되었다. 두 개체 집단간의 차이는 그림17B를 통해 확인
해본 결과, 일반돼지와 비교해 보았을 때 형질전환 돼지에서 약 3배정도 높게 발현 되었다.

- 32 -
그림17. 혈청에서 ELISA를 이용하여 Adiponectin과 Resistin 호르몬의 발현량 분석 .
(A)adiponectin (B) Resistin의 호르몬 발현량 분석. WT : Wild type (n=3 9-115, 9-144,
9-145) TG : transgenic (n=5, 9-385, 9-276, 9-280, 9-282, 9-289). * P<0.05
3. 근위성 세포의 분리 및 배양법 확립
가. 실험방법
(1) 근위성 세포의 분리 및 배양
근육세포의 분리 배양을 위해 이전에 보고된 방법을 이용하여 수행하였다 (Ueyama et
al., 1999). 근육 조직 sample을 trimming 한 후 조직을 가위로 충분히 잘게 자른 후에
pronase (protease; Sigma, MO) 1.25 mg/ml를 첨가하여 37°C에서 한 시간 동안 배양하였
다. 배양된 세포를 충분히 섞어 준 뒤 3000 rpm 4분간 원심분리 후 상층액을 제거하였다.
PBS 40ml을 넣고 20초간 vortex로 섞어 준 뒤 1000rpm 10분간 원심분리 후 새로운 tube
로 상층액을 옮겼다①. 남아 있는 펠렛에 PBS 40ml을 첨가하여 1000rpm 8분간 원심분리
후 새로운 tube로 상층액을 옮겼다② 이와 같은 방법을 한번 더 반복하여 상층액을 얻었다
③. ①, ②, ③ tube의 상층액을 3000rpm 4분간 원심분리 후 상층액을 제거한 후에 DMEM
으로 부유시켜 70um 필터를 통해 하나의 새로운 tube로 옮겼다. poly-L-lysine- and
fibronectin-coated 된 8-well culture plates에 세포를 분주한 후 Dulbeccos’ modified eagle
medium (DMEM-high glucose) 에 10% fetal bovine serum (FBS), 50 Unit/ml penicillin,
50 µg/ml streptomycin and 50 µg/ml gentamicin 을 첨가하여 만든 배양액을 넣어 37°C,

- 33 -
5% CO2에 배양하였다.
(2) 통계처리
본 연구에 사용된 통계처리는 one-way ANOVA와 Student' s t-test를 이용하여 유의차
를 검증하였다. p<0.05일 때 유의차를 인정하였다.
나. 결과
(1) 쥐의 근위성세포의 분리 및 배양
본 실험은 PEA15 과발현 형질전환 돼지의 당뇨병 및 인슐린 저항성과 관련된 신호전달
체계 등 근육에서의 대사과정을 분석하기 위해서 근위성 세포의 분리 및 배양에 대한 기술
을 확립하기 위한 예비실험 결과이다. 그림18은 근위성 세포분리 후, 분화과정에 따른 특정
마커의 발현 시기를 모식화 하였다. 초기에 Pax7이 발현되고 myoblast에서 이들이 융합되
는 시기에 MyoD, myogenin 및 desmin이 발현되고, 다핵성 myotube에서는 desmin 및
MHC가 발현된다.
Proliferating
myoblast
Differentiating
to fuse
Multinucleated
myotube
Satellite cell Myoblast Myotube
Pax7
MyoD
Myogenin
Desmin
MHC
Day 0 Day 1~2 Day 5Day 3~4
Proliferating
myoblast
Differentiating
to fuse
Multinucleated
myotube
Satellite cell Myoblast Myotube
Pax7
MyoD
Myogenin
Desmin
MHC
Day 0 Day 1~2 Day 5Day 3~4
그림18. 근위성 세포의 분화 과정 및 발현 마커
그림19는 근위성 세포 배양 후 2일째의 특정 마커들의 발현 양상을 확인한 결과이고, 그
림20은 배양 5일째의 결과이다. 근위성 세포 배양 후 2일째(Day 2) Hoechst 염색으로 세포
의 핵을 염색하였고, 근위성 세포의 특정 마커인 Pax7, MyoD, Myogenin, 및 Desmin의 항
체를 붙이고, 488nm에서 검출되는 2nd 항체를 사용하여 형광현미경으로 확인하였다. 세포
의 수가 증가 되었고 형태도 방추사 모양으로 변한 것이 많았지만 세포간에 융합은 아직
일어나지 않았다. 또한, myoblast에서 발현이 많은MyoD의 발현이 관찰되었, Desmin의 발
현이 Day 1 보다 많았다 (그림19).

- 34 -
그림19. 근위성 세포 배양 2일째의 세포마커들의 발현 양상
근위성 세포 배양 후 5일째(Day 5) Hoechst 염색으로 세포의 핵을 염색하였고, 근위성
세포의 특정 마커인 Desmin 및 MHC의 항체를 붙이고, 488nm에서 검출되는 2nd 항체를
사용하여 형광현미경으로 확인하였다. 다핵의 myotube가 형성됨을 확인하였고, 세포질에서
Desmin 및 MHC가 염색됨을 관찰하였다. 하지만, 많은 수의 myoblast에서는 염색되지 않
음을 알 수 있었다 (그림20).
그림20. 근위성 세포 배양 5일째의 세포마커들의 발현 양상

- 35 -
제3절 PEA15 과발현 세포의 In vitro 검정
1. PEA15 과발현 근육세포의 특징
가. 실험방법
(1) 근위성 세포의 분리 및 배양
제2절에 설명된 방법과 동일한 방법으로 일반돼지(WT) 3두, 형질전환 돼지(TG) 5두에서
각각 근위성 세포를 분리하여 실험조건에 맞게 배양하였다.
(2) Real Time RT-PCR
Real-Time PCR 반응은 RotorGene 6000 (Corbett) Real-Time PCR기계를 사용하여 분석
하였다. cDNA의 taget gene과 GAPDH은 표3에서 표시된 특정 primer를 사용하여 발현을
확인하였고 GAPDH 발현은 normalized하는데 사용되었다. PCR증폭은 다음과 같은 단계로
진행되었다. denaturation (95°C for 10 min), amplification (95°C for 10 sec, 60°C for 30
sec, 72°C for 30 sec) – 45 cycle, cooling (4°C). 형광 흡광 데이터는 반응 종결 후에 얻
을 수 있었다. PCR 반응 종결 후 melting cure작성을 통하여 유전자 증폭의 정확성을 재확
인 하였다. melting curve는 특정 염기서열마다 다르게 나타나는데, PCR 증폭산물에서 특
이현상을 구별하는데 사용된다. 유전자의 발현의 상대적인 양은 2-ddCt방법을 사용하여 분
석하였다 (Livak and chmittgen, 2001). 그 후, PCR 증폭산물을 2% agarose gel에 전기영
동하여 확인하였다.

- 36 -
표3. 각 유전자의 RT-PCR용 프라이머 염기서열
Gene Primer sequence (5'-3' )Product size
(bp)
Adiponectin F-TGCAAGCTTTCCTCACACAC
335bp R-CACCCCTAACCTCAGTGGAA
Adiponectin-2 F-ATGATGTCACCACTGGCAAA
235bp R-TTCTCTTCATCCCCGTATGC
Resistin F-CTGTGTCCCGTGGATGAAG
301bp R-GATGCGCAGATGCAAACTTA
GLUT4 F-CCTGCAGTTTGGCTACAACA
328bp R-GAGGAACCGTCCAAGAATGA
GLUT4-2 F-CCTGCAGTTTGGCTACAACA
235bp R-CAGCATTGCCTTCTTCCTTC
IR F-CCTGCAGTTTGGCTACAACA
342bp R-TGAGAAACTCGATCCGCTCT
IR-2 F-AGAGCGGATCGAGTTTCTCA
245bp R-CCATCCCATCAGCAATCTCT
PEA15-318bp F-GGACATCCCCAGTGAGAAGA
318bp R-TACGGGTTAGCTTGGTGTCC
PEA15-393bp F-GCAACGTGCTGGTTATG
393bp R-TACGGGTTAGCTTGGTGTCC
GAPDH F-ACCCAGAAGACTGTGGATGG
346bp R-CCCCAGCATCAAAGGTAGAA
GAPDH-2 F-TCGGAGTGAACGGATTTG
240bp R-CCTGGAAGATGGTGATGG
(3) W estern blotting
일반돼지와 형질전환 돼지의 근육, 지방, 췌장, 간 조직 그리고 근육세포로부터 protein을
추출하였다. Total proteins 의 추출은 Mammalian Protein Extraction Reagent (M-PER)
buffer (PIERCE Inc.), protease inhibitor cocktail (Santa Cruz Biotech. Inc.)을 통해 추출
하였다. Plasma membrane protein추출은 plasma membrane protein extraction Kit
(Abcam)를 사용하여 추출하였다. 추출된 proteins sample을 Bradford assay 용액
(Bio-RAD, USA)을 이용하여 측정하였고, 4X Laemmli' s sample buffer로 정량한 후에 5
분간 끓여 주었다. Proteins을 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) gel에서 분리시킨 후 nitrocellulose membranes (Amersham

- 37 -
Biosciences)으로 250 mA 에서 90분 동안 전이 시켰다. 전이된 membrane을 90분간 0.05%
Tween-20이 첨가된 5% non-fat milk를 이용하여 blocking 하였고, 1차 항체는 PEA15
(1:1000, Cell Signaling Tech.), GLUT4 (1:2000, Santa Curz Biotech. Inc.) 그리고 flag
(1:3000, Sigma-Aldrich)를 1:1000비율로 사용하여 4℃ 에서 overnight 하였다. TBS-T 로
10분씩 3회 washing 후 2차 항체 - Goat polyclonal anti-Rabbit IgG 를 1:3000 비율로 1
시간 실온에서 천천히shaking하였다. 그 후에 protein band를 최종적으로 확인하기 위해
enhanced chemiluminescence (ECL)를 사용하여 X-ray film (Model JP-33, JPI, Korea).에
현상하였다.
(4) Glucose uptake 분석
돼지 근위성 세포들은 Dulbecco s̀ modified Eagle s̀ medium (DMEM; 4mM
L-glutamine, 1.5g/L sodium bicarbonate, 4.5g/L glucose)에 FBS 10%를 포함하는 배양액
에 배양한다. 당 수송능력 변화 실험을 위해서 세포는 80% confluent 상태에서 PBS 완충
액으로 세포를 세척한 후 0.5% FBS DMEM(4mM L-glutamine, 1.5g/L sodium
bicarbonate, 1g/L glucose) 배지로 교환하여 24시간 배양하였다. 형질전환 및 일반돼지의
배양된 근위성 세포에 인슐린 호르몬을 일정시간 처리한 후 Krebs-Ringer Buffer (KRB)에
서 30분 배양한다. KRB 완충액에서 배양된 세포에 0.1mM 2-deoxy-D-glucose 와
2-deoxy-D-[14C]glucose(0.2uCi/ml)를 처리한 후 10분 동안 반응시킨다. 반응이 끝난 세포
에서 완충용액을 제거하고 세포를 세척한 후 1M NaOH로 세포를 용해한다. 용해된 세포는
scintillation counter로 측정되었다.
(5) 당수송체 (Glucose transporter) 이동
세포막으로 Glucose 이동의 분석은 이전에 보고된 방법을 이용하여 확인하였다 (Caruso
M. et al. 1997). 간략하게 말하자면, 근육세포에 DMEM (low glucose) 에 2% FBS를 첨가
하여 24시간 배양시킨 후, 세포 내 100 nM insulin 을 처리하여 2, 5, 10, 30, 60 분 동안
배양 하였다. 처리시간에 따라 배양시킨 세포를 plasma membrane protein extraction Kit
(Abcam, England)를 사용하여 plasma membrane protein을 추출 하였고, Mammalian
Protein Extraction Reagent (M-PER) buffer (PIERCE Inc.)를 사용하여 total cellular
proteins을 추출하였다. total protein과 plasma membrane protein로 추출한 sample을 10%
SDS-PAGE gel로 분리한 뒤 nitrocellulose membrane으로 전이 시켰다. 이것을 1차 항체인
polyclonal anti-GLUT4 antibody (Cell Signaling Inc.) 를 사용하여 GLUT4의 발현량을 확
인하였다.
(2) 통계처리
본 연구에 사용된 통계처리는 one-way ANOVA와 Student' s t-test를 이용하여 유의차
를 검증하였다. p<0.05일 때 유의차를 인정하였다.
나. 결과

- 38 -
(1) 세포내 PEA15 발현
형질전환 돼지의 다양한 조직과 근육세포에서 mPEA15가 발현되는지 확인해 보기 위해
Western blot 분석을 통해 확인해 보았다. 확인해본 결과 일반돼지와 형질전환 돼지 모두
endogenous pig PEA15 protein은 15kDa 부위에서 발현되고 있었다. 하지만, exogenous
mouse PEA15 protein의 발현은 형질전환 돼지에서만 15kda 위쪽에 발현되고 있음을 확인
하였다. 이것은 그림1의 벡터구조에서 보여주듯 mPEA15 유전자의 N-terminal 부근에서
flag tagging에 의해 단백질이 융합되었기 때문이다. 따라서 통해 western blotting을 통해
anti-flag를 분석해 본 결과 형질전환 돼지에서만 발현되는 것을 확인해 볼 수 있었다 (그
림21A, 22A).
그림21. 형질전환 돼지와 일반돼지의 근육세포에서 PEA15 발현 확인 . (A) Western
blots결과, WT: wild-type pig ( n=3, W1: 9-115, W2: 9-144, W3: 9-145), TG :
Transgenic pig ( n=5, T1: 9-385, T2: 9-276, T3: 9-280, T4: 9-282, T5: 9-289) (b)
Densitometer를 이용한 형질전환 돼지와 일반돼지의 근육세포에서 mPEA15 단백질 발현을
정량. ** P<0.01

- 39 -
그림22. 형질전환 돼지와 일반돼지의 근육조직에서 PEA15 발현 확인. (A) Western blots
결과, WT: wild-type pig ( n=3, W1: 9-115, W2: 9-144, W3: 9-145), TG : Transgenic
pig ( n=5, T1: 9-385, T2: 9-276, T3: 9-280, T4: 9-282, T5: 9-289) (b) Densitometer를
이용한 형질전환 돼지와 일반돼지의 근육세포에서 mPEA15 단백질 발현을 정량. **
P<0.001
exogenous mPEA15의 발현량을 분석하기 위해 mRNA을 추출한 후 Real-Time
RT-PCR을 통해 확인해 보았다 그림23A와 23B의 결과를 통해 확인해 본 결과, PEA15
318bp의 증폭산물 내부에서는 형질전환 돼지의 근육세포에서 일반돼지에 비해 약 80~100정
도 더 강하게 발현되는 것을 확인해 볼 수 있었고(그림23A), Table 1에서 보는 것과 같이
PEA15 393bp의 프라이머는 mPEA15내에 Flag가 포함된 구조이며, 이 프라이머를 통해
RT-PCR을 수행한 결과 Flag의 염기서열이 포함된 PEA15 393bp의 증폭산물 내부에서는
WT은 전혀 발현되지 않고 TG에서만 발현되고 있었다(그림23B). Real-Time RT PCR을
사용하여 형질전환돼지와 일반돼지에서의 다양한 조직의 발현량을 분석하였다. 형질전환
돼지의 근육 조직에서 PEA15 mRNA 발현량이 일반돼지에 비해 약 80-100배 더 강하게
발현되었다. 또한, 그밖에 다른 조직에서도 조직이나 개체에 따라 차이는 보였지만 일반돼
지에 비해 형질전환 돼지에서 PEA15 유전자가 더 강하게 발현되었다 (그림23C).

- 40 -
그림23. 형질전환 돼지와 일반돼지의 여러 부위 조직에서 PEA15 유전자의 mRNA 발현량 확
인. 근육세포와 여러 부위의 조직에서 PEA15의 mRNA 발현량을 Real Time PCR을 이용
하여 분석. 같은 양의 RNA로 정량하기 위해 GAPDH를 이용하여 Loading. (A) PEA15
318 bp 유전자를 일반돼지와 형질전환 돼지의 근육세포에서 비교 분석. (B) WT과 TG 그
룹에서 PEA15 318 bp와 PEA15 393 bp를 이용하여 Real Time PCR을 통해 PCR 최종산
물을 얻었고 이것을 아가로스 젤 전기영동을 통해 확인 (C) 형질전환 돼지와 일반돼지의
여러 부위 조직에서 PEA15 318bp 유전자를 확인. *** P<0.001
(2) 당뇨병 관련 유전자 발현
당뇨병 관련 유전자의 발현을 Real Time RT-PCR을 통해 mRNA의 발현량을 확인하였
다. 우리는 먼저 형질전환 돼지와 일반돼지에서의 근육세포를 분리 배양한 후 근육 세포
에서의 당뇨관련 유전자의 mRNA 발현량을 확인하였다. PEA15 유전자는 일반돼지에 비해
형질전환 돼지에서 약 6배정도 더 강하게 발현 된다는 점을 확인하였다 (그림24A).
Adiponectin은 지방세포에서 분비되는 물질로서 지방세포 분화과정에서 발현량이 증가된다.
또한 당뇨, 인슐린 저항이 있는 환자에서 Adiponectin의 mRNA가 낮게 발현된다. 그러나
근육세포에서는 mRNA 발현량이 크게 차이가 없음을 확인할 수 있었다 (그림24B).
Resistin은 비만에서 과발현 되어 진다고 알려진 유전자로써 각 개체별 확인해 본 결과 형
질전환 돼지에서 2배 정도 좀더 높게 발현되는 것으로 확인되었다 (그림24C). GLUT4와
IR 경우 일반돼지와 형질전환 돼지에서 발현량이 크게 차이가 없었다 (그림24D,E).

- 41 -
그림24. 근육세포에서 Real Time PCR을 이용하여 당뇨관련 유전자의 mRNA 발현량 확
인. (A) PEA15, (B) Adiponectin, (C) Resistin, (D) GLUT4, (E) Insulin Receptor (IR) 유
전자의 mRNA 발현량 분석. ** P<0.01
근육세포에서의 당뇨관련 유전자를 분석했던 것과 같이 형질전환돼지와 일반돼지의 다양
한 조직-근육 지방 간 조직에서의 Real Time RT-PCR을 통해 당뇨관련 유전자의 mRNA
의 발현량을 분석하였다. PEA15 유전자의 경우 부위마다 조금씩 차이가 있었지만 형질전
환 돼지의 근육조직에서 일반돼지에 비해 약 70배 이상 더 강하게 발현되는 것을 확인하였
다. Adiponectin의 경우 근육과 지방 조직에서는 크게 차이는 없었지만, 일반돼지의 간 조
직에서 약 2배정도 더 높게 발현되었다. Resistin은 형질전환 돼지의 근육조직에서 약 2배
정도 더 높게 발현되었고, 간, 지방 조직에서는 크게 차이가 없었다. GLUT4는 일반돼지에
서 약 2배정도 더 높게 발현되었으나 다른 부위에서는 크게 차이가 없었다. 마지막으로 IR
은 부위별 근소한 차이는 있었지만 발현량은 크게 차이가 없었다 (그림25).

- 42 -
그림25. 여러 부위의 조직에서 Real Time PCR을 이용하여 당뇨관련 유전자의 mRNA 발현량 확
인. (A)PEA15, (B)Adiponectin, (C)Resistin, (D)GLUT4, (E) Insulin Receptor(IR) 유전자의
mRNA 발현량 분석. * P<0.05, ** P<0.01
(3) 당수송체의 세포막 이동 양상
PEA15가 과발현이 되면 인슐린의 시그널을 저해하고 활성을 억제시켜 GLUT4가 세포막
으로의 이동이 저해 될 것 (Tomas et al., 2002) 이기 때문에, 우리는 다양한 조직에서
GLUT4 발현량을 분석하였다. 일반돼지와 형질전환 돼지의 근육 지방 간 조직의 Total
protein levels에서 Western blot을 통해 GLUT4 발현량을 확인하였다. 근육조직의 경우에
는 일반돼지의 #9-145 개체에서 가장 높은 수준으로 발현되었고, 형질전환돼지의 #9-289
개체에서 가장 낮게 발현되었다. 이와는 대조적으로 일반돼지와 형질전환돼지의 지방조직
에서의 GLUT4 발현은 일반 돼지에서 형질전환돼지에 비해 높게 발현되고 있음을 확인할
수 있었다(P<0.05). 간 조직에서는, 일반돼지의 #9-115 개체에서 가장 낮게 발현되었으나,
다른 개체에서는 거의 차이가 없었다 (그림26).

- 43 -
그림26. 근육 , 지방 , 간 조직에서의 GLUT4 단백질 발현량 분석 . (A) 조직에서 Western
blotting을 이용하여 단백질 분석. WT: wild-type pig ( n=3, W1: 9-115, W2: 9-144, W3:
9-145), TG : Transgenic pig ( n=5, T1: 9-385, T2: 9-276, T3: 9-280, T4: 9-282, T5:
9-289). (B) 근육, (C) 지방 (D) 간 조직 (E) 여러 부위의 조직에서 GLUT4 발현량 분석. *
P<0.05
근육세포를 배양한 후 0, 2, 5, 10, 30, 60분에 걸쳐 인슐린을 처리하였고, 인슐린 처리한
근육세포를 Total protein과 Plasma membrane protein을 추출하여 GLUT4 유전자의 이동
양상을 western blot을 통해 확인해본 결과 이다. 첫번째 밴드 사진은 시간대 별로 인슐린
처리 후 total protein에서 GLUT4 유전자의 발현량을 확인한 결과이다. Total protein에서
는 시간대별 비슷하게 발현되고 있는 것을 확인해 볼 수 있었다. 두번째 밴드 사진은 시간
대 별 인슐린 처리후 plasma membrane에서 GLUT4 유전자의 발현량을 확인한 결과이다.
시간이 지남에 따라 glut4가 세포막 쪽으로 이동하고 있음을 확인해 볼 수 있었다. 반면에
TG의 경우 시간이 지남에 따라 이동하기는 하나 WT 비해 이동량이 현저히 낮은 것으로
확인 되었다. 각 개체 별 western blot을 통해 확인한 후 densitometry를 통해 보다 정확한
수치로 측정한 결과 개체마다 조금씩 차이는 있었지만 대체적으로 Total protein 에서는 시
간대별 변화량이 크게 차이가 나지 않았고, PM의 경우 시간이 지남에 따라 glut4가 세포막
쪽으로 이동하는 것으로 확인 되었다. WT의 경우 5분 정도 지났을 때 glut4 유전자가 세
포막으로 가장 많이 이동하는 것으로 나타났으며 10분 후부터는 서서히 감소하는 것으로
나타났다. 그러나 TG의 경우 WT에 비해 세포막으로의 이동량이 현저히 낮은 것으로 확인

- 44 -
되었다. 따라서 PEA15 과발현으로 인해 세포수준에서 GLUT4 유전자의 세포막으로의 이
동이 저해된다는 사실을 확인해 볼 수 있었다 (그림27B).
Glucose는 인슐린 분비 후 세포막으로 이동하는 것으로 확인되었다 (Tomas et al.,
2002). 따라서 GLUT4의 세포막으로의 이동을 확인해 보기 위해 Western blotting을 통해
일반돼지와 형질전환돼지의 근육세포에서 확인하였다. 근육세포를 배양한 후, PEA15가 과
발현 되면 인슐린 저항성이 발생하여 세포표면으로의 glucose의 이동이 저하된다
(Condorelli et al., 1998). 따라서 우리는 두개의 그룹에서 근육세포를 배양한 후 시간대 별
로 걸쳐 인슐린을 처리하였다. (0, 2, 5, 10, 30, 60분) 인슐린 처리한 근육세포를 Total
protein과 Plasma membrane protein을 추출하여 GLUT4 유전자의 이동 양상을 western
blot을 통해 확인하였다. Total protein과 plasma membrane에서 GLUT4 유전자의 발현양
상을 그림27A를 통해 확인해 볼 수 있다. Total protein 수준에서는 일반돼지와 형질전환
돼지의 근육 세포에서 발현량의 차이가 없음을 확인하였다. 그러나 plasma membrane에서
는 GLUT4 protein 양이 일반돼지에서 높은 수준으로 발현되는 것을 확인하였다 (그림27).
이것은 GLUT4 protein이 인슐린을 처리하였을 때 시간이 지남에 따라 세포막으로 이동하
는 것을 의미한다. 그러나 형질전환 돼지에서는 세포막으로의 이동이 저해되는 것을 확인
해 볼 수 있다. 그림27A에서는 형질전환 돼지와 비교해 보았을 때 일반 돼지에서 GLUT4
protein 수준이 더 많이 발현 되는 것을 확인해 볼 수 있다. 형질전환 돼지에서 GLUT4의
세포막으로의 이동은 인슐린을 100nM 처리하였을 때 개체마다 차이는 있었지만 10분에서
가장 많이 발현하는 것으로 확인 되었다. 반면에, 일반돼지의 경우 인슐린 처리 후 5분에서
발현량이 가장 강한 것으로 확인되었다. 이 결과를 바탕으로 densitometry를 통해 정확한
수치로 측정한 결과 개체마다 조금씩 차이는 있었지만 대체적으로 Total protein 에서는 시
간대별 변화량이 크게 차이가 나지 않았고, PM의 경우 시간이 지남에 따라 glut4가 세포막
쪽으로 이동하는 것으로 확인 되었다 (그림27B).

- 45 -
그림27. 근육세포의 총 단백질과 막 단백질에서 GLUT4 유전자의 이동 양상을
W estern blotting을 통해 확인 . 근육세포 배양 후 인슐린 100 nM을 0분에서 60분 정도 처
리 한 후 GLUT4 분석. (A) 근육세포에서 Western blots을 통해 단백질 발현량을 확인. 왼
쪽 그림은 일반돼지, 오른쪽 그림은 형질전환 돼지. TP : total protein, PM : plasma
membrane protein. (B) 근육세포에서 GLUT4 발현량 정량분석. 왼쪽 그래프는 총 단백질 오
른쪽 그래프는 막단백질. WT: Wild-type (n=3), TG: Transgenic pig (n=5). * P<0.05, ***
P<0.001
PEA15가 과발현이 되면 인슐린의 시그널을 저해하고 활성을 억제시켜 glucose가 세포막
으로의 이동이 저해됨을 확인해 보기 위해, 방사선 동위원소를 통해 glucose uptake를 확인
해 보았다. 근육세포 배양후 방사선 동위원소를 transfection 하였고, 그 후에 인슐린을 24
시간, 15분 동안 처리한 후 분석한 결과이다. 인슐린 처리 24시간 후 각 개체 별
2-deoxy-D-glucose (2-DG) uptake 를 측정한 결과, 형질전환 돼지에 비해 일반돼지가 약
2배정도 더 높게 확인 되었다 (그림28A). 이 때, Glucose uptake level은 일반돼지의 근육세
포 #9-115 개체에서 가장 높게 측정되었고 (109 nmol/mg, +insulin), 형질전환 돼지의
#9-276 개체에서 가장 낮게 측정되었다 (33 nmol/mg, +insulin). 인슐린 처리 15분 후
Glucose uptake 측정 결과 24시간 후의 값과 비슷한 수준으로 측정되는 것으로 확인되었
다. 이를 통해 형질전환 돼지가 일반돼지에 비해 Glucose 의 세포막으로의 이동이 현저하
게 저해되고 있다는 사실을 확인하였다 (그림28B).

- 46 -
그림28. 형질전환 돼지와 일반돼지의 근육세포에서 Glucose uptake 양상 확인 . 근육세
포에서 glucose uptake를 확인하기 위해 2-deoxy-D-glucose (2-DG)를 이용하여
transfection 하였다. 그 후, 근육세포에서 인슐린 100 mM 처리하여 배양 시켰고, 재료 및
방법에 나온 설명에 따라 2-deoxy-D-glucose (2-DG) uptake 팻턴을 확인. 인슐린 처리
(A) 24 시간, (B) 15 분 후에 glucose uptake를 확인하였다. * P<0.05, *** P<0.001
(4) 고찰
PEA15는 multifunctional protein으로 세포 내 여러 기능과 증식, apotosis를 조절하고 인
슐린 조절에 따른 glucose uptake를 조절한다고 연구되어져 왔다 (Brunet et al.. 1999;
Condorelli et al., 1998; Condorelli et al., 1999; Estelles et al.. 1999; Formstecher et al.,
2001; Kitsberg et al., 1999; Trencia et al., 2004; Vaidyanathan et al.. 2003). 또한 제 2형
당뇨환자의 다양한 조직과 세포에서, PEA15 유전자의 발현이 증가되었다 (Condorelli,
1998). 이전 연구에서의 제 2 형 당뇨병에 대한 PEA15 overexpression의 관련성을 확인해
보기 위해, 우리는 mouse PEA15 유전자가 과발현된 형질전환 돼지를 생산했다 (Lee et
al., 2011). 이러한 형질전환 돼지에서, mPEA15의 발현은 형질전환 돼지의 모든 부위에서
발현하며, 근육 조직에서는 50배 이상 발현하고, 다른 조직에서는 근육조직보다는 약하게
발현하지만 일반돼지에 비해 과발현되는 것으로 확인되었고 Western blot 분석 결과
PEA15 일반돼지와 형질전환 돼지 모두 endogenous pig PEA15 protein은 15kDa 부위에서
발현되고 있었고, exogenous mouse PEA15 protein의 발현은 형질전환 돼지에서만 15kda
위쪽에 발현되고 있음을 확인하였다. 이는 mPEA15 유전자의 N-terminal 부근에서 flag
tagging에 의해 단백질이 융합되었기 때문이다.
이번 연구를 통해, 우리는 당뇨 관련 유전자 (Adiponectin, Resistin, GLUT4, IR)의 발현

- 47 -
량을 Real Time PCR을 통해 분석해 보았다. 우리는 형질전환 돼지와 일반돼지의 근육세포
로부터 mRNA 발현량을 확인해 보았다. Adiponectin은 에너지 항상성이나 포도당 지질 대
사에 매우 중요한 역할을 하는 호르몬이다 (Maeda et al., 1996; Scherer et al., 1995; Hu
et al., 1996). Adiponectins은 지방세포에서 분비되는 물질로서 지방세포 분화과정에서 그
발현이 증가되며 지방세포에 특이적으로 발현된다. 흥미롭게도, 혈중 adiponectin의 수준이
증가하면 체중이 감소하며, 이와 관련하여, 일반적으로 비만이나 당뇨, 인슐린 저항이 있는
환자에서 adiponectin의 mRNA의 발현이 급격히 감소하여 adiponectin의 혈중 농도가 낮다
는 연구 결과가 있다 (Xiaobo 등, 2007). Resistin은 adipocytes가 adipogenic으로 발달하는
동안 분비되는 호르몬이다. Resistin 유전자는 당뇨환자에 당뇨병치료제 TZD와 같은 물질
에 의해 현저하게 감소되는 것을 확인해 볼 수 있다. 또한, 후천적으로 식이에 의해 유도된
비만뿐 아니라 유전적으로 발생된 비만과 인슐린 저항성이 있는 환자에서 Resistin의 수준
이 증가된다 (Steppan 등, 2001). 따라서 이를 토대로 Adiponectin의 발현량을 Real Time
PCR을 통해 근육세포와 다른 다양한 조직에서 mRNA의 수준을 확인하였고 형질전환 돼지
와 일반돼지에서는 크게 차이가 없었다. 반면에 Resistin의 경우 일반돼지에 비해 형질전환
돼지에서 약 2~6배정도 더 강하게 발현되었다.
하지만 임상연구에서는, 당뇨병환자의 PEA15 발현은 낮은 adiponectins의 수준과 높은
Resistins의 수준이 확인되었다. 우리는 mPEA15 형질전환 돼지에서 낮은 adiponectins의
수준과 높은 Resistins의 수준이 나타나지는 않았다. 이 돼지의 경우 혈액 내 Resistin의 수
준이 확연하게 증가되었지만 혈액내 adiponectin 수준은 큰 변화가 나타나지 않았다. 이 시
점에서 우리는 명확하게 이 현상을 설명할 수는 없다. 그러나 한가지 가능성은 PEA15 발
현과 같은 강한 signal로 인해, 다른 무언가의 작용으로 adiponectin의 발현에 영향을 줄 것
이라는 점이다.
당뇨병 환자의 경우 PEA15 발현으로 인해 GLUT4의 세포막으로의 이동이 저해되어 체
내 glucose의 함량이 높아짐에 따라 조직의 손상을 가져오게 된다. 따라서 PEA15 유전자
로 인한 GLUT4 활성 여부와 두 유전자의 관계에 대한 존재를 추측하기 위해 GLUT4 유
전자를 분석하였다. 이 실험에서 우리는 주로 근육세포를 이용하여 분석하였는데, 근육의
경우 혈액 내 포도당을 흡수하기 위한 가장 중요한 부분이고 인슐린 uptake를 조절할 수
있다. 이러한 이유로 근육 세포는 세포 내 존재하는 다양한 signaling과 관련하여 GLUT4
traffic을 연구하기 위한 model system이 되었다 (Ueyama et al., 1999; Wang et al., 1999;
Khayat et al., 2000; Tong et al., 2001; JeBailey et al., 2004, 2007; Thong et al., 2007;
Ishikura and Klip, 2008). 근육세포는 PEA15 signaling의 하위 구조를 정의하는데 좋은 툴
을 가지고 있기 때문에 (Chiu et al., 2010),우리는 근육세포를 이용하여 PEA15 과발현으로
인해 세포 내 GLUT4의 세포막으로의 이동에 관련된 signaling을 작용하여 이동을 억제 시
키는지에 관한 메커니즘을 분석하고 정의하였다.
첫째로, 우리는 다양한 조직과 근육세포에서 mRNA 발현량을 확인해 보았다. GLUT4는
조절 메카니즘으로써, 우리는 일반돼지에서의 GLUT4가 높게 발현되는 것을 확인해 보았
다. 그러나 우리의 결과에서는 일반돼지와 형질전환돼지의 근육세포를 비교해 보았을 때
거의 차이가 나지 않았다. 근육조직에서는 일반돼지에서 약 2배정도 높게 발현되었으나, 다

- 48 -
른 부위에서는 차이가 나지 않았다.
두번째 단계로 PEA15 발현과 GLUT4 활성과의 관계를, 일반돼지와 형질전환 돼지의 근
육세포와 세개의 조직(근육, 지방, 간)에서의 GLUT4의 발현량을 Western blottine을 통해
분석하였다. GLUT4의 발현은 대부분의 sample에서 관찰하였으나, 일반돼지와 형질전환 돼
지에서의 GLUT4는 크게 차이가 나지 않았다.
세번째 단계로써, 우리는 PEA15가 과발현된 형질전환 돼지의 세포막에 glucose channel
에 영향을 미치는지 분석하였다. 이것은 형질전환 돼지와 비교하였을 때 일반돼지의 세포
막에 GLUT4가 더 높게 발현되는 것으로 확인 되었다. 즉, mPEA15의 발현은 GLUT4의
이동을 저해시켜 인슐린 저항성이 나타나게 되는데, 이를 확인해 보기 위해 배양된 근육세
포에 인슐린을 처리한 후 시간이 지남에 따라 GLUT4 발현량의 변화를 분석하였다 (Yea
et al., 2009; Zisman et al., 2000). 일반돼지의 경우, 세포막에서의 GLUT4 발현량은 5분에
서 가장 강하게 나타났다 이와 대조적으로 형질전환 돼지의 근육세포에서는, 세포 내 인슐
린 처리 후 10분에서 GLUT4의 발현량이 증가되는 것을 볼 수 있으나 GLUT4의 발현량은
일반돼지에 비해 낮은 수준으로 발현되는 것을 확인해 볼 수 있었다.
네번째 단계에서는 mPEA15의 발현에 따른 근육세포에서의 glucose uptake를 분석해 보
았다. 근육세포 배양후 방사선 동위원소를 transfection 하였고, 인슐린을 24시간, 15분 동안
처리한 후 분석한 결과이다. 세포 막으로의 glucose의 이동량을 확인하기 위해,
radio-labeled glucose, 2-deoxy-[3H]-D-glucose(2-DG)을 배양액에 첨가하였다. 그 후 정
해진 시간 동안 배양 한 후 세포 내 흡수된 ladio-labeled glucose를 scintillation counter를
사용하여 glucose의 이동량을 확인하였다. 그 결과 일반돼지가 형질전환 돼지에 비해 세포
내 2-deoxy-D-glucose (2-DG) uptake가 24시간과 15분에서 더 높게 나타났다. 이 결과는
이전에 세포막에서의 GLUT4의 발현량을 분석한 데이터와 관련하여 근육 세포 내 glucose
uptake도 mPEA15의 발현에 의해 GLUT4가 조절되는 것으로 확인되었다. 즉, 형질전환된
세포 내 glucose의 흡수는 PEA15 과발현에 의해 GLUT4가 세포표면으로의 이동을 억제시
키는 것이며 (Gerolama et al., 1998), 세포막의 GLUT4의 감소는 다른 동물모델에서도
PEA15 발현으로 인해 glucose 의 수송과 GLUT4의 이동이 억제된다고 보고되었다 (Czech
et al., 1992; Marshall et al., 1993).
마지막으로, mPEA15가 발현된 형질전환 돼지에서 Glucose의 항상성은 glucose solution
(200g)을 직접 입에 주입한 후에 정해진 시간에 Oral Glucose Tolerance Test (OGTT)를
통한 혈당수준의 변화추이를 확인해 보았다. 형질전환 돼지와 일반돼지에서의 혈액 내
glucose의 함량을 확인 하였는데 일반돼지에서는 Glucose 투여후 5분에서 증가하였고 10분
후에는 천천히 감소 되었다. 전반적으로 일반돼지에서는 측정 시간 내내 형질전환 돼지에
비해 상대적으로 낮은 수준으로 유지되었다. 그러나 형질전환 돼지의 경우, Glucose 투여
후 30분대에서 혈액 내 glucose 의 수준이 가장 높게 측정되었다. 이 결과와 같이, 혈중
glucose tolerance test 통해 PEA15의 메커니즘의 조절에 의해 나타나는 혈액 내 높은
glucose의 함량은 당뇨병에서 흔히 보이는 증후로 보이며, 따라서 mPEA15가 발현되는 형
질전환 돼지에서 혈액 내 높은 glucose의 함량을 보이고 있었다.
전반적으로, 우리의 결과를 다음과 같이 요약할 수 있다.; (1) mPEA15가 발현된 돼지에

- 49 -
서 당뇨관련 유전자의 발현; Resistin을 제외하고는 다른 유전자들은 우리가 기대했던 결과
를 얻진 못했다. (2) GLUT4는 근육세포에 존재하고 있고, 일반돼지에 인슐린을 처리하였을
때 세포막 내 GLUT4의 발현량이 증가되었음을 보여주었다. 형질전환 돼지의 경우, mRNA
의 발현과 세포 내 total protein의 발현량은 일반돼지와 비교하였을 때 차이가 거의 없었
다. 그러나 형질전환 돼지의 세포에서 인슐린 처리시 세포막에 존재하고 있는 GLUT4는 약
하게 발현되었고 이로 인해 세포막으로의 이동이 저해되는 것을 확인해 볼 수 있다. (3) 마
지막으로 glucose tolerance test에서 형질전환 돼지에서 높은 수준의 혈중 glucose 수준을
보여주고 있다.
결론적으로, 돼지에게 glucose를 투여 한 후, glucose는 인슐린 분비에 의해 혈액 내
glucose의함량은 시간이 지남에 따라 감소되어야 하지만 mPEA15가 발현되고 있는 형질전
환 돼지의 혈액 내 glucose의 수준은 glucose glucose administration 이후 증가되었다.
GLUT4의 경우 형질전환 돼지의 조직에서 일반적인 수준으로 나타났다. 그러나 인슐린 처
리 후 세포막에서의 GLUT4는 약하게 발현되었다. 이것은 아마도 인슐린 분비 시 작용되는
세포 내 여러 signaling이 mPEA15에 의해 특정한 커텍터로 작용하여 PKC ξ / λ를 억제하
였기 때문에 인슐린 저항성이 발생되었을 것이다.
본 연구에서는, 당뇨병 연구를 위한 질환모델 동물로써 PEA15가 발현되고 있는 돼지가
질환모델 동물로써 가능한지 입증하기 위해 연구하였다. 이미 당뇨병은 다양한 유전자가
관여하고 있고, 호르몬과 signaling을 조절하는 복잡한 질환이라고 알려져 있다. 따라서 단
일 유전자의 과발현으로 완벽한 모델동물을 만들 수는 없다. 그럼에도 불구하고, 적절한 모
델 동물이 제공될 수 있다면 신약 훨씬 빠르고 쉽게 개발될 수 있을 것이 분명하다.
mPEA15 돼지는 완벽한 모델 동물의 제작의 한 단계일 것이다.

- 50 -
제4절 유전자재조합 세포배양계의 질환모델로서의 이용성 검정
1. 재조합세포주 구축 및 당뇨질환과의 연관성 검정
가. 실험방법
(1) 실험개요
형질전환 돼지에 사용된 당뇨관련 유전자(PED/PEA15)의 과발현이 미치는 효과를 세포수
준에서 검토 하고자 간 및 근육 세포주를 이용한 과발현재조합세포주를 구축하였다. 재조
합세포주의 대사시스템 변화 분석을 통하여 질병유발 유전자(PED/PEA15)의 과 발현이 당
뇨를 유발하였는지 검정한다. 제2 형 당뇨의 병증인 인슐린 저항성의 가장 큰 원인은 각
조직과 혈중의 중성지방의 수치의 증가이다. 그럼으로 재조합세포주에서의 중성지방 생합
성 증가는 당뇨 유발 유무를 확인하기 위한 중요한 요인이며, 재조합 간세포주에서의 중성
지방 생합성 변화를 확인하여 재조합세포주의 당뇨질환 모델로서의 가능성을 확인한다. 재
조합세포주에서의 재조합유전자가 정상적으로 과발현되는지를 단백질 수준에서 확인 하였
다. 제 2형 당뇨는 근육조직 및 세포에서의 당수송능력 감소로 그 병증을 나타내다. 본 연
구에서는 질병유발 유전자(PED/PEA15)의 과발현으로 유도된 세포내의 기능적 변화확인을
세포 안으로의 당 수송능력을 glucose uptake 실험을 통해 확인 하였다.
과발현벡터는 간세포에 사용한것과 동일한 유전자를 사용하였으며, 세포주는 랫트 근육세
포주( L6 cell)를 사용하였다. 과발현 유무는 PEA15 항체를 이용하여 웨스턴블롯팅으로 확
인하였고, 형질전환 세포의 당 수송능력의 변화 실험은 2-deoxy-D-[14C]glucose을 이용하
여 분석하였다.
(2) 실험재료
PED/PEA15 유전자를 과발현벡터는 국립축산과학원으로부터 제공받아 간 및 근육세포
형질전환에 이용하였다 (그림29). 세포주는 인간 간세포주(HepG 2 cell)를 사용하였으며, 형
질전환 세포의 지방대사활성 변화 확인은 [14C]acetate를 이용하여 분석하였다.

- 51 -
그림29. 유전자 과발현용 재조합벡터
(3) 방법
(가) 간세포주 배양
인간 간세포인 HepG2 세포는 ATCC (American Type Culture Collection, USA)에서 분
양 받았으며, 세포는 L-glutamine, non-essential amino acids, sodium bicarbonate, 10 %
(v/v) fetal bovine serum (FBS), 1 % antibiotics, 1 mM sodium pyruvate가 첨가된 MEM
를 배양 배지로 37℃, 5% CO2 조건 하에서 배양하였다. Pea15 유전자 발현벡터를 간세포
에 형질전환 후에 12 or 24-well 배양용기에서 1-2 일 배양 후 실험에 사용하였다.
(나) 간세포에서의 중성지방 생합성
형질전환한 HepG2 cell (5×105 cells/well)을 24 well plate에 분주하고 1-2일 동안 배양한
후 0.25 mCi [14C]acetate를 첨가하고 24간 동안 반응시켰다. 반응 후, 세포는 추출용매
(Hexane : 2-propanol = 3 : 2, v/v) 0.5 ml을 첨가하여 중성지방을 포함하는 총 지방을 추
출하였으며, 추출용매를 제거한 전체 지방은 TLC (silica gel 60F254, 0.5 mm, Merck)에
점적하여 전개용매 (Hexane : Diethyl ether : Acetic acid = 80 : 20 : 1, v/v)상에서 전개
하였다. TLC상에서 중성지방을 분리한 후 TLC는 방사선원소의 에너지를 감광 할 수 있는
필름에 감광한 후 이미지 분석(BAS-1500, Fuji Photo Film Co. Ltd.)을 통해 중성지방의
방사선량을 측정하였다. 추출 후 세포의 단백질 농도를 측정하였다. 중성지방의 방사능 양
을 측정하여 단백질 농도로 나눈 값을 실험값으로 하여 각 실험군사이의 실험적 오차를 보
정하였다.
(다) 근육세포주 배양
재조합세포주 L6 세포는 Dulbecco s̀ modified Eagle s̀ medium (DMEM; 4mM

- 52 -
L-glutamine, 1.5g/L sodium bicarbonate, 4.5g/L glucose)에 FBS 10%를 포함하는 배양액
에 배양한다. 당 수송능력 변화 실험을 위해서 세포는 80% confluent 상태에서 PBS 완충
액으로 세포를 세척한 후 0.5% FBS DMEM(4mM L-glutamine, 1.5g/L sodium
bicarbonate, 1g/L glucose) 배지로 교환하여 24시간 배양하여 실시되었다 (Antti 등, 2001).
(라) 근육세포에서의 Glucose uptake 분석
재조합세포주 L6 세포에 인슐린 호르몬을 일정시간 처리한 후 Krebs-Ringer Buffer
(KRB)에서 30분 배양한다. KRB 완충액에서 배양된 세포에 0.1mM 2-deoxy-D-glucose 와
2-deoxy-D-[14C]glucose (0.2uCi/ml)를 처리한 후 10분 동안 반응시킨다. 반응이 끝난 세
포에서 완충용액을 제거하고 세포를 세척한 후 1M NaOH로 세포를 용해한다. 용해된 세포
는 scintillation counter로 측정되었다 (Aas 등, 2005).
나. 결과
1. 간세포주를 이용한 재조합세포주 구축 및 당뇨질환과의 연관성 검정
간세포에서의 PED/PEA 15 유전자 과발현 재조합세포주의 지질대사 분석을 통하여 질병
유발 유전자(PED/PEA15)의 과발현에 의한 당뇨유발 가능성을 확인하였다. 제2형 당뇨의
병증인 인슐린 저항성은 각 조직과 혈중의 중성지방 수치의 증가가 원인으로 보고되고 있
다. 따라서 재조합 간세포주에서의 중성지방 생합성 증가는 당뇨유발 유무를 확인하기 위
한 중요한 요인이다. 인간유래 간세포주에 PED/PEA15 유전자를 도입한 후 24시간 혹은
48시간 후에 간에서의 중성지방 합성정도를 확인하였다. 방사선 동위원소로 표지된 중성지
방 합성에 이용되는 기질을 세포에 처리 후 6시간 반응하였다. 반응 후 세포내 합성된 중
성지방량은 방사선량을 측정함으로서 확인하였으며, 유전자 도입 후 24시간 후
PED/PEA15 과발현 세포주에서의 중성지방 합성율이 40% 증가하는 것을 확인하였다 (그
림30).
그림30. PED/PEA 15 과발현 간세포에서의 중성지방 생합성 변화 . PED/PEA 15 과발현
벡터를 간세포에 처리하여 24시간(□) 동안 발현유도 한 후 중성지방 생합성활성을 측정한 결과
일반세포보다 중성지방합성율이 40%증가 하였으며 48시간(□) 후에는 큰 차이를 보이지 않았다.

- 53 -
2. 근육세포주를 이용한 재조합세포주 구축 및 당뇨질환과의 연관성 검정
(가) PED/PEA15 과발현세포주의 구축
PED/PEA15의 과발현 재조합유전자를 형질전환시켜 재조합세포주를 제작하였다. 구축된
재조합세포주의 유전자 과발현 유무는 세포에서 단백질을 추출한 후 PED/PEA15 항체를 이
용하여 분석되었다. PED/PEA15 유전자를 형질전환시킨 재조합세포주에서는 그렇지 않은
무처리 세포주와 비교해서 단백질 발현량이 약 2.5배 증가되는 것을 확인하였다 (그림31).
그림31. PED/PEA15 재조합세포주 구축 및 인슐린의 신호전달체계 변화 분석 . 그림29
의 마우스의 PEA/ PED 15유전자 발현벡터를 근육세포에 형질전환 한 후 PEA/PED 15 항체를
이용하여 세포내의 PEA/PED15 단백질의 발현을 확인 하였다. 또한, 인슐린 처리에 의한 세포신호
전달변화를 확인 하기 위해 인슐린에 의해 활성화 되는 AKT단백질의 인산화를 확인 한 결과 형질전환
유무와 상관없이 인산화가 일어났다.
(나) 재조합세포주의 당 수송능력 검토
구축된 과발현 재조합세포주에서의 당 수송능력은 [14C]2-deoxy-D-glucose가 과발현 세
포내로 이동되는 정도를 세포내의 방사활성 변화를 통하여 확인하였다. PED/PEA15 유전
자의 재조합세포주에서는 형질전환시키지 않은 세포주에 비해 당 수송능력이 감소함을 확
인하였다. 감소 정도는 형질전환에 사용된 재조합 DNA의 처리량과 비례관례를 나타냈었
다. 이러한 당 수송능력의 감소는 재조합세포주에 인슐린 호르몬의 처리유무에 관계없이
양쪽 모두의 세포주에서 유사한 발현양상이 관찰되었다 (그림32). 결론적으로 PEA/PED 15
유전자의 근육내 과발현은 세포의 기능변화를 야기 시켰으며, 이와 같은 변화는 제 2형 당뇨병
증을 갖은 환자에게서 확인 할 수 있는 세포외에 있는 당의 세포안으로의 수송능력 감소 병증
으로 보인다.

- 54 -
그림32. 재조합세포주의 당수송 능력 변화 분석 . 근육세포에 처리된 PEA/PED 15 발현벡
터 DNA농도 의존적으로 당 수송능력의 감소를 확인 할 수 있었다. 당 수송능력의변화는 인슐
린을 처리하지 않은 세포(■)보다, 인슐린처리 세포(□)에서 2배 이상증가 하였으며, 발현벡터
처리 후 에는 인슐린에 의한 당 수송능력변화가 감소되는 것을 확인 할 수있다.
(다) 당 수송과 인슐린 관련성 검토
과발현 재조합세포주에서 관찰되는 당 수송능력의 감소가 인슐린 호르몬과 연관성이 있
는지를 확인하기 위하여 인슐린 신호전달 과정에 관여하는 신호전달인자(Akt)의 단백질 활
성(인산화 정도)의 변화를 항원-항체 반응 실험을 통해 분석되었다. 인슐린의 신호전달인자
인 불활성형 Akt는 호르몬 처리에 관계없이 재조합세포주와 무처리 세포주 모두에서 그
발현량이 높게 검출되었으나, 인슐린을 처리하지 않았을 경우 활성형 pAkt(인산화 형태)는
관찰되지 않았다. 인슐린을 처리하지 않은 세포주에서는 검출되지 않았던 활성형 pAkt는
인슐린 처리로 재조합유전자의 과발현과는 관계없이 그 발현량이 매우 높게 증가 되었다.
즉, PED/ PEA15 유전자의 과발현으로 인하여 세포내 당 수송능력의 감소는 인슐린 비의
존적인 경로를 경유하여 작용되었을 것으로 판단되어진다 (그림31).

- 55 -
제5절 전사체 및 단백체 발현양상을 통한 질환모델가축의 검정
1. 유전체 및 단백체의 발현 특성
가. 실험방법
(1) 실험개요
PEA/PDE 15 과발현으로 유도된 당뇨모델가축의 유전체 발현 특성을 일반돼지와 형질전환
돼지의 조직별 유전체의 발현변화를 통하여 과발현 유무와 형질변화를 확인 하고자 하였다. 또
한, DNA chip은 질환이 유도된 개체의 전체적인 전사체 분석을 가능하게 하며, 그 결과를 이
용한 당뇨관련 신규 표지분자의 발굴 및 이용성 검토를 가능하게 한다. PEA/PED15 dbwjs자
에 의한 당뇨유발 질환모델가축의 유전자군의 발현변화 분석 결과는 당뇨질환발병 검정 뿐 아
니라 유전자에 의한 간접적인 생화학적 대사변화 분석도 가능하게 하리라 사료된다. 질환모델
에 연구에 있어서 유전자 발현변화양상을 분석 하는 것은 과발현을 유도한 유전자의 발현여부
등을 확인 할 수는 있지만 실질적인 기능변화나 대사변화를 유추하기는 어렵다. 이러한 이유로
유전체 뿐만 아니라 최종적으로 유전자의 기능을 나타내는 단백체 발현 여부가 질환모델을 검
정하는데 있어 중요하게 여겨진다. 특히, 단백체중에 혈장에 존재하는 단백질은 간등의 대사산
물의 분비에 의해 변화하는 단백질이다 (Dominiczak 등, 2011). 이는 질환의 발병이나 대사변
화를 확인 할 때 중요하게 여겨지는 단백체군으로 여겨지며, 특히 당뇨와 같은 대사성질환에
있어 중요한 연구대상이다. 당뇨는 혈중의 당농도의 변화뿐 아니라, 중성지방 합성 및 에너지
대사등 여러 대사이상을 유발한다. 이러한 대사이상에 의한 병증은 특히, 혈액의 단백질의 발
현변화를 야기시키며, 혈장내 단백질 변화를 통하여 발병 유무를 확인할 수 있다. PEA/PDE
15 과발현 당뇨모델가축의 혈장 단백질 변화분석은 도축을 하지 아니하더라도 발병을 확인 할
수 있을 뿐 아니라, 진단 단백질 개발로 인해 당뇨치료제 투여를 통한 치료효과를 모니터링 하
는데 이용 가능 할 것이다. 유전체와 단백체의 변화를 확인 하였다 하더라도 단백질 기능 변화
에 의한 활성이 검정되지 않는다면 질환모델의 당뇨 발병을 확증하기는 어렵다. 이러한 기능적
변화를 확인하기 위해서는 당뇨병증 유발에 있어 가장 확실한 변화인 조직에서 혈중의 당을
흡수하는 능력의 감소를 측정하는 것이 필요하다 . 혈중의 당은 세포내의 에너지 고갈이나 혈
중의 높은 농도의 당존재에 의해 분비되는 인슐린에 의해서 세포내로 흡수 되지만 (Antti
Virkama ̈ ki, et al., 2001), 당뇨가 발병되면 이러한 생체내 신호에도 불구하고 조직에서 당을
흡수하는 능력이 감소 하게된다. 특히, 당뇨발병에 의하여 이당의 흡수능력이 민감하게 영향을
받는 조직은 근육조직으로서, 당뇨발병 후 근육조직의 당 수송능력의 변화 분석은 질환모델의
발병 유무를 확인하는데 가장 중요한 부분일 것이다.
(가) PEA/PDE 15 과발현 당뇨모델가축의 유전체 발현 특성
일반돼지와 형질전환 돼지의 조직별 유전체의 발현변화와 특성분석을 통하여 과발현 유
무와 형질변화를 확인 하고자 하였다. 또한, DNA chip은 질환이 유도된 개체의 전체적인
전사체 분석을 가능하게 하며, 그 결과를 이용한 당뇨관련 신규 표지분자의 발굴 및 이용

- 56 -
성 검토를 가능하게 한다.
(나) 당뇨모델가축의 단백체 발현 특성
당뇨는 혈중의 당농도의 변화뿐 아니라, 중성지방 합성 및 에너지 대사등 여러 대사이상
을 유발한다. 이러한 대사이상에 의한 병증은 특히, 혈액의 단백질의 발현변화를 야기시키
며, 혈장내 단백질 변화를 통하여 발병 유무를 확인할 수 있다. PEA/PDE 15 과발현 당뇨
모델가축의 혈장 단백질 변화분석은 도축을 하지 아니하더라도 발병을 확인 할 수 있을 뿐
아니라, 진단 단백질 개발로 인해 당뇨치료제 투여를 통한 치료효과를 모니터링 하는데 이
용 가능 할 것이다.
(다) 당뇨모델가축의 근육조직세포의 당 수송능력 변화분석
당뇨병증 유발에 있어 가장 확실한 변화는 조직에서 혈중의 당을 흡수하는 능력의 감소
이다. 혈중의 당은 세포내의 에너지 고갈이나 혈중의 높은 농도의 당존재에 의해 분비되는
인슐린에 의해서 세포내로 흡수 된다. 그러나 당뇨가 발병되면 이러한 생체내 신호에도 불
구하고 조직에서 당을 흡수하는 능력이 감소 하게된다. 특히, 당뇨발병에 의하여 이당의 흡
수능력이 민감하게 영향을 받는 조직은 근육조직으로서, 당뇨발병 후 근육조직의 당 수송
능력의 변화 분석은 질환모델의 발병 유무를 확인하는데 가장 중요한 부분일 것이다.
(2) 실험재료
(가) PEA15 형질전환 돼지 유전체 발현특성
일반돼지와 형질전환 돼지의 간, 근육, 지방 조직을 취하여 유전체의 발현변화와 특성분
석을 하였다. 위의 시료들은 국립 축산과학원에서 제공받아 실험에 사용하였다. 또한, DNA
chip은 Agilent 사에서 제공하는 whole Porcine genome chip을 사용하여 전사체 분석을 하
였다.
(나) PEA15 형질전환 돼지 단백체 발현특성
국립 축산과학원에서 제공받은 일반돼지와 형질전환 돼지의 혈액으로 부터 혈장내에 존
재하는 단백질을 분석 하고자 하였다. 2DE를 통하여 차등발현되는 단백질을 선별한 후
Q-TOF 분석기기를 이용하여 단백질을 동정하고자 하였다.
(다) 형질전환 가축의 생화학적 대사변화 분석
일반돼지와 형질전환 돼지의 근육조직으로부터 조직세포를 분리하여 그 기능을 검정 하
고자 하였다. 당뇨의 발병은 근육조직의 당 수송능력의 감소로 확인 할 수 있다. 형질전환
돼지가 PEA/PED15유전자의 과발현에의해 당뇨가 유발되었다면 정상돼지의 당 수송능력보
다 감소함을 확인 할 수 있을 것이다. 본 실험에서는 위와 같은 사실을 확인하고자, 분리된
세포에 2-deoxy-D-[14C]glucose를 처리하여 2-deoxy-D-[14C]glucose 동위원소가 세포내
로 수송되는 정도를 측정 하였다.

- 57 -
(3) 방법
(가) 조직으로부터 total RNA 분리 및 RT-PCR
확보된 일반돼지와 형질전환돼지의 조직으로부터의 total RNA 추출은 RNeasy plus mini
kit(QIAGEN)를 사용하여 분리하였다. 시료에 RLT plus buffer를 첨가해 조직을 변성시킨
후 gDNA Eliminator spin column에 옮겨 원심 분리하여 DNA를 제거하였다. gDNA
Eliminator spin column을 통과한 용액에 100% 에탄올을 혼합하여 RNeasy mini spin
column에 옮긴 후 원심 분리하여 column에 RNA를 결합시켰고, RPE buffer로 column을
세척하였다. 이 후 50㎕의 RNase-free한 증류수를 사용하여 RNA를 column에서 분리하였
다. 그 중 5㎕를 20배 희석하여 spectrophotometer를 이용 O.D. 260/280nm에서 RNA의 농
도 및 순도를 측정하였다. 추출한 total RNA를 template로 사용하여 oligo dT primer와
reverse transcriptase를 통해 first-strand cDNA를 합성하였다.
(나) 돼지 DNA chip을 이용한 근육의 전체 유전자 발현 분석
추출한 total RNA는 Agilent’s 2100 Bioanalyzer System을 이용하여 total RNA의
quality를 측정하였으며, chip 분석을 위한 전처리는 Agilent’s Low RNA Input Linear
Amplification kit PLUS를 이용하여 증폭 및 라벨링 과정을 수행하였다. Microarray
hybridization은 Agilent’s Gene Expression Hybridization Kit를 이용하여 수행하였으며,
Agilent whole Porcine genome 4 X 44K chip을 이용하여 전체 유전자 발현분석을 진행하
였다. Scan 및 이미지 분석은 Agilent’s DNA microarray scanner 및 Feature Extraction
Software를 이용하여 수행하였으며, 이미지 분석 결과는 Agilent' s GeneSpring Software를
이용하여 normalization 및 clustering 등을 수행하여 분석하였다. 유전자 선택은
normalized ratio 값이 2배 이상 차이를 보이는 probe와 반복실험을 통하여 통계적으로 유
의성 있는 유전자 그룹을 Single t-test로 분석하여 선별 하였다.
(다) 혈장으로부터 단백질 분리
일반돼지와 형질전환돼지의 혈액시료로부터 혈장을 분리하여 단백체 분석 시료로 사용하
였다. 혈액시료는 300 X g에서 20분간 원심분리한 후 상등액을 취하여 실험에 사용하였다.
혈액으로부터 분리된 혈장에서 일반돼지와 형질전환돼지에서 차등 발현되는 단백질을 분석
하기 위하여 다량으로 존재하는 알부민과, 면역글로불린을 제거하였다. 알부민과, 면역글로
불린을 제거한 혈장은 단백질 용해액(7M urea, 2M thiourea, 4% CHAPS, 40mM Tri, 1%
DTT, protease inhibitor cocktail)에 녹였으며, 단백질의 농도는 Bradford assay 용액을 이
용하여 분석되었다. 혈장 단백질(400ug)은 단백질의 등전점에 따른 분리가 가능한 IPG
strip을 이용하여 1차 전기영동을 하였다. 1차 전기영동은 pH 4-7 범위에서 단백질의 분리
가 가능한 IPG strip을 이용하였으며, 2차 전기영동은 12.5% SDS-PAGE를 이용하여 단백
질의 분자량에 따른 분리를 수행하였다. 미토콘드리아 단백질의 동정은 SDS-gel상에서 단
백질만을 추출하여 Q-TOF를 이용하여 펩타이드 서열을 분석 하였다.

- 58 -
(라) 세포배양
형질전환 돼지근육 조직세포는 Dulbecco s̀ modified Eagle s̀ medium (DMEM; 4mM
L-glutamine, 1.5g/L sodium bicarbonate, 4.5g/L glucose)에 FBS 10%를 포함하는 배양액
에 배양한다. 당 수송능력 변화 실험을 위해서 세포는 80% confluent 상태에서 PBS 완충
액으로 세포를 세척한 후 0.5% FBS DMEM(4mM L-glutamine, 1.5g/L sodium
bicarbonate, 1g/L glucose) 배지로 교환하여 24시간 배양하여 실시되었다.
(마) Glucose uptake 분석
형질전환 돼지근육 조직세포에 인슐린 호르몬을 일정시간 처리한 후 Krebs-Ringer
Buffer (KRB)에서 30분 배양한다. KRB 완충액에서 배양된 세포에 0.1mM 2-deoxy-D-
glucose 와 2-deoxy-D-[14C]glucose(0.2uCi/ml)를 처리한 후 10분 동안 반응시킨다. 반응이
끝난 세포에서 완충용액을 제거하고 세포를 세척한 후 1M NaOH로 세포를 용해한다. 용해
된 세포는 scintillation counter로 측정되었다.
나. 결과
(1) 조직으로부터 total RNA 분리 및 RT-PCR
형질전환으로 인한 PEA/PED 15 유전자의 과발현를 PCR분석을 통하여 확인하였다. 일반
돼지 5두와 형질전환 돼지 4두의 조직으로부터 전체 RNA를 정제하여 발현분석을 위한 시
료로 사용하였다. 근육과 간에서는 PEA/PED15 유전자의 과발현을 확연히 확인 할 수 있
었으나, 지방조직에서는 일반돼지에서도 PEA/PED15 유전자의 발현이 높은 수준으로 관찰
되었다 (그림33).
그림33. 일반돼지 및 형질전환돼지 조직별 PEA/PED 15 유전자 발현분석 . WT; wild
type, TG; transgenic

- 59 -
(2) 돼지 DNA chip을 이용한 근육의 전체 유전자 발현 분석
일반돼지 2두와 형질전환 돼지 4두의 조직으로부터 전체 RNA를 정제하여 microarray 분
석을 위한 시료로 사용하였다 (그림34). 형질전환 돼지근육조직과 일반돼지의 근육조직에서
차등 발현되는 유전자 선별은 일차적으로 전체 array결과를 분석하여 통계적으로 유의성있는
유전자군을 선별하였다(그림35). 일차적으로 선별된 유전자군을 바탕으로 발현이 증가하는 유
전자와 감소하는 유전자를 선별 하였으며, 선별기준은 t-test를 통해 p값이 0.05이하의 유의성
을 갖는 유전자를 선택하였다(그림36). Array결과 일반돼지보다 형질전환 돼지에서 2배 이상의
발현증가 양상을 보이는 유전자는 전체 44,000개 유전자중에 264개가 선별되었으며, 선별된 유
전자중에 대사기능이 보고된 기능이 알려진 유전자는 175개였다. 반면에 발현이 감소되는 유전
자는 386개가 선별되었고, 이들 유전자 중에서 기능이 알려진 유전자는 90개로 확인 되었다.
그림34. Agilent’s 2100 Bioanalyzer System을 이용한 total RNA의 quality를 측정
그림35. 전체 array에서 통계적으로 유의성 있는 유전자 그룹 선별

- 60 -
그림36. 통계적으로 유의성 있는 유전자 그룹에서 2배 이상의 차이를 보이는 유전자 선별
대사기능이 보고된 유전자를 바탕으로 생화학적 대사와의 연관성을 분석하기위해 KEGG
(www.kegg.jp) 웹상의 구축된 돼지(Sus scrofa; pig)의 대사맵 분석툴을 사용하였다 (표4). 분
석결과 선별된 유전자군이 포함되고, 관여하는 대사경로를 확인 할 수 있었으며, 전체 생화학
적 대사에 관련되는 유전자는 선별된 유전자중 22개가 포함되었다. 특히, 형질전환돼지에서 차
등 발현되는 유전자들 중 일부는 당뇨질환과 직접적으로 연관되어진 해당과정(관련유전자 6개,
그림37)과 인슐린 신호전달(그림38) 및 제 2형 당뇨에 관여하는 대사(그림39)에 관여하는 유전
자(3개)로 확인되었다. 뿐만 아니라 지방대사의 증가 및 지방의 축적을 질병의 원인 및 복합병
증으로 갖고 있는 제 2형 당뇨의 병증을 확인 할 수 있는 또 다른 결과도 확인 되었다. 이와
같은 사실은 아디포사이토카인 분비 및 신호전달 경로와 지방대사관련 유전자의 발현 변화로
알 수 있었다. 위의 결과로 PEA/PED15 유전자 과발현 형질전환돼지는 당뇨병증과 연관되어
있는 생화학적 대사에 관여하는 유전자의 발현에 영향을 미치는 것으로 확인 되었다.

- 61 -
그림37. Glycolysis/Gluconeogenesis. Susscrofa (pig) 일반돼지(WT) 및 형질전환돼지
(TG)의 근육조직에서 차등발현되는 Glycolysis / Gluconeogenesis 대사에 관련된 유전자군
으로 대사지도상에서 붉은색으로 표시된 유전자는 TG에서 높게 발현되는 유전자이다.

- 62 -
그림38. Insulin signal pathway. Susscrofa (pig) 일반돼지(WT) 및 형질전환돼지(TG)의 근육
조직에서 차등발현되는 Insulin 신호전달 대사에 관련된 유전자군으로 대사 지도상에서 붉은색으로
표시된 유전자는 TG에서 높게 발현되는 유전자이며 푸른색으로 표시된 유전자는 발현이 감소돼는
유전자이다.
그림39. Type II diabetes mellitus. Susscrofa (pig) 일반돼지(WT) 및 형질전환돼지(TG)의 근육조
직에서 차등발현되는 Type II diabetes mellitus 대사에 관련된 유전자군으로 대사 지도상에서 붉은색으로 표
시된 유전자는 TG에서 높게 발현되는 유전자이며 푸른색으로 표시된 유전자는 발현이 감소돼는 유전자이다.

- 63 -
표4. 돼지 근육의 미토콘드리아 단백질의 동정
PathwaysRelated
gene
ssc01100 Metabolic pathways - Susscrofa (pig) 22
ssc00010 Glycolysis / Gluconeogenesis- Susscrofa (pig) 6
ssc04010 MAPK signaling pathway - Susscrofa (pig) 5
ssc04920 Adipocytokine signaling pathway - Susscrofa (pig) 4
ssc04930 Type II diabetes mellitus - Susscrofa (pig) 3
형질전환 동물모델에서의 당뇨 표지유전자와 단백질의 발현량과 활성변화를 연구하여 질
환모델로서의 가치 평가와 질환발병의 원인기작 연구를 수행하였다. 형질전환 돼지
(PEA/PED15)의 혈장의 단백질은 2DE 프로테옴 실험기법을 통하여 분석하였고 (그림40),
형질전환 돼지에서 차등발현되는 단백질을 탐색하였다. PEA/PED15 과발현 당뇨모델 돼지
에서 차등 발현 되는 단백질은 Q-TOF 분석을 통하여 peptide sequence를 확인 하였다. 단
백질분리 및 동정이 비교적 용이한 MALDI-TOF 분석을 시도 했으나 돼지의 혈장에 분비된
단백질과 관련된 유전자 및 단백질의 database는 아직 많이 부족하여 동정된다 해도 일치되는
단백질을 확인 할 수가 없었다. 이러한 이유로 알려지지 않거나 database가 미흡한 단백질 동
정에 이용되는 Q-TOF MS/MS를 분석기법을 이용하였다. 이로서 차등 발현되는 3개의 단백
질은 apolipoproteinA-IV, ficolin-2 precursor로 확인 되었다 (그림41, 표5). Apolipoprotein
A는 HDL에 포함되어 있는 지단백으로서 HDL은 당뇨 및 비만이 유도되었을 때 감소하며, 대
사성질환의 분자마커로 이용된다. 또한 ficolin-2는 당뇨환자에서 혈장중에 높게 분비된다는 보
고가 있다. 본 실험에 이어 항체를 이용한 단백질의 발현 분석 및 유전자 발현분석 등 추가적
인 연구가 진행되어야 하겠지만, 결론적으로 질병의 발병을 진단하는 가장 중요한 혈액내의 단
백체 변화는 형질전환돼지의 당뇨질환 유발의 가능성을 확인 할 수 있게 하였다. 특히 형질전
환 돼지의 혈장단백질에서 당뇨질환을 진단하는 대표적인 단백질 apolipoproteinA의 분비 감소
및 당뇨병증을 갖는 환자에게서 높게 발견되는 혈장 ficolin-2 precursor단백질의 검출은
PEA/PED15 유전자의 과발현이 당뇨질환 유발을 가능하게 함을 검정 할 수 있는 중요한 결과
라고 사료된다.

- 64 -
그림40. 일반돼지 및 형질전환 돼지의 차등발현 혈장단백질
그림41. 차등발현 혈장단백질 분석을 위한 단백질 동정 및 Q-TOF 결과

- 65 -
표5. 돼지혈장 단백질의 동정
Spot
NO.
Peptidesequence
ResultsofQ-TOF
proteinID
(mass/pI)matched protein
1
NLEASVAPYADEFK
/ALVQQVEDLR
apolipoproteinA-IV
precursor
(43268/5.68)
1 MFLKAVVLSL ALVAVTGARA EVNADQVATV MWDYFSQLGS NAKKAVEHLQ
51 KSELTQQLNT LFQDKLGEVN TYTEDLQKKL VPFATELHER LTKDSEKLKE
101 EIRRELEELR ARLLPHATEV SQKIGDNVRE LQQRLGPFTG GLRTQVNTQV
151 QQLQRQLKPY AERMESVLRQ NIRNLEASVA PYADEFKAKI DQNVEELKGS
201 LTPYAEELKA KIDQNVEELR RSLAPYAQDV QEKLNHQLEG LAFQMKKQAE
251 ELKAKISANA DELRQKLVPV AENVHGHLKG NTEGLQKSLL ELRSHLDQQV
301 EEFRLKVEPY GETFNKALVQ QVEDLRQKLG PLAGDVEGHL SFLEKDLRDK
351 VNTFFSTLKE EASQGQSQAL PAQEKAQAPL EG
2 VVGLEGSDKLSILR
ficolin-2precursor
(35173/5.63)
1 MDTRGVAAAM RPLVLLVAFL CTAAPALDTC PEVKVVGLE GSDKLSILRGC
51 PGLPGAAGPK GEAGASGPKG GQGPPGAPGE PGPPGPKGDR GEKGEPGPKG
101 ESWETEQCLT GPRTCKELLT RGHILSGWHT IYLPDCQPLT VLCDMDTDGG
151 GWTVFQRRSD GSVDFYRDWA AYKRGFGSQL GEFWLGNDHI HALTAQGTNE
201 LRVDLVDFEG NHQFAKYRSF QVADEAEKYM LVLGAFVEGN AGDSLTSHNN
251 SLFTTKDQDN DQYASNCAVL YQGAWWYNSC HVSNLNGRYL GGSHGSFANG
301 VNWSSGKGYN YSYKVSEMKF RAT
3 VDLVDFEGNHQFAK
ficolin-2precursor
(35173/5.63)
1 MDTRGVAAAM RPLVLLVAFL CTAAPALDTC PEVKVVGLEG SDKLSILRGC
51 PGLPGAAGPK GEAGASGPKG GQGPPGAPGE PGPPGPKGDR GEKGEPGPKG
101 ESWETEQCLT GPRTCKELLT RGHILSGWHT IYLPDCQPLT VLCDMDTDGG
151 GWTVFQRRSD GSVDFYRDWA AYKRGFGSQL GEFWLGNDHI HALTAQGTNE
201 LRVDLVDFEG NHQFAKYRSF QVADEAEKYM LVLGAFVEGN AGDSLTSHNN
251 SLFTTKDQDN DQYASNCAVL YQGAWWYNSC HVSNLNGRYL GGSHGSFANG
301 VNWSSGKGYN YSYKVSEMKF RAT
형질전환 가축의 생화학적 대사변화를 확인하기 위하여, 당뇨유발 유전자의 과발현으로
재조합세포주 및 형질전환 돼지조직 세포의 기능을 비교분석하였다. 과발현시킨 재조합세포
주는 물론 형질전환 근육조직 세포에서도 당뇨의 병증인 당 수송능력의 감소를 확인하였다.
또한 인슐린에 의한 반응성도 세포주 실험계와 동물 실험계 모두에서 감소하였다 (그림42).
결론적으로 당뇨유발 유전자의 과발현으로 재조합세포주와 형질전환 근육조직 세포에서 당
수송능력과 인슐린 반응성이 감소하였고, 이 결과로서 당뇨 병증을 확인할 수 있었다.
그림42. 돼지 근육조직세포 및 형질전환 세포주에서의 당수송능력 분석

- 66 -
제6절 질환모델가축의 개발을 위한 기초연구
1. 연구배경
질병의 연구나 신약개발을 위한 전임상 연구에서 많이 이용되는 설치류를 통한 연구는 결
과를 도출하는데 용이 하기는 하나 생리학적으로 인간과 많은 차이를 갖고 있어 질병이나
신약을 연구하는데 있어 많은 문제점이 야기되는 실정이다. 또한, 이러한 문제의 해결을 위
하여 영장류를 이용한 실험이 제안되고 있으나 경제적 부담과 시설의 열약함 때문에 연구를
함에 있어 제한적이다. 반면에 돼지는 해부 생리학적으로 인간과 가장 유사한 특성을 가지
고 있으며, 영장류 보다 경제적인 연구가 가능하며, 연구시설 확보다 용이하여 이러한 이유
로 질병모델로서의 가치가 높게 평가되고 있다(Gianfranco BRAMBILLA, et al., 2004). 돼지
를 이용한 질환모델 개발이나 연구는 아직은 여러 문제점을 갖고 있다. 질환에 대한 연구를
위해서는 유전체나 단백체, 대사체와 같은 연구기반이 필수적이지만 전체적인 정보를 확인
할 수 있는 DB가 부족한 실정이다. 특히, 질환모델 개발에 있어 발병이나 치료에 따른 생화
학적 변화를 확인하기 위해서는 단백질의 변화와 기능분석이 필수적 이지만 돼지의 단백체
연구는 미진한 상태이다. 특히, 근육조직의 미토콘드리아 단백질은 당뇨질환 발병에 주요 요
인이 되는 당대사나, 지방대사, 에너지대사와 밀접한 관계가 있는 조직이다. 당뇨질환 유발
후 당뇨병의 발병의 유무를 생화학적으로 검정 하기위해서는 돼지근육 조직의 단백질에 대
한 정보가 필수적 이라고 생각된다. 본 연구에서는 돼지를 이용한 당뇨질환 모델의 개발을
위한 기초 연구로서 돼지근육조직의 미토콘드리아 단백질지도를 만들어 질환모델가축의 검
정에 그 정보를 이용하고자 하였다.
본 과제에서는 당뇨질환이 유발되었을 때 간에서 중성지방의 합성이 증가하고, 근육에서
는 당의 수송능력이 감소하는 병증을 기본으로 PEA/PEA15 유전자 과발현 형질전환돼지의
검정을 시도 하였다. 또한, 당뇨관련 신규 표지분자의 발굴을 위해 PEA/PEA15유전자 외에
중성지방의 합성과 당의 수송능력을 감소시켜 당뇨를 유발할 수 있는 유전자를 선별하였다.
위에 설명된 병증의 개선을 가능하게 하는 약물의 검정용 당뇨질환 동물모델로서 선별된 유
전자는 중성지방생합성에서 첫 번째 단계를 촉매하는 단백질로서 glycerol 3-phosphate
acyltransferase로 명명하는 효소이다(Shi, Y et al., 2004). 이 유전자를 이용하여 당뇨모델을
개발하였을 때의 신약검정용으로서의 가치를 평가하기위해 억제제를 처리하여 중성지방의
생합성감소와 당 수송능력변화를 확인 하고자 하였다.
2. 돼지근육 미토콘드리아 단백체 발현특성
가. 실험방법
(1) 재료 및 개요
본 연구에서는 렌드레이스를 이용하여 당뇨질환가축을 생산하고자 하였다. 돼지근육의 미
토콘드리아 단백체지도를 만들기 위해 일반돼지(렌드레이스)의 근육조직을 채취하여 연구
를 수행하였다.

- 67 -
(2) 돼지 근육조직으로부터 미토콘드리아 분리
돼지의 근육은 도축 후 액체질소에 급냉 후 마쇄하여 실험에 이용하였다. 완충용액A
(120mM KCl, 20mM HEPES, pH 7.4, 2mM MgCl2, 1mM EGTA) 10ml에 돼지 근육 1g
첨가 후 균질기를 이용하여 혼합하였다. 균질화된 용액은 600g에서 10분간 원심분리하여
상층액은 확보하였다. 확보된 상층액은 17,000g에서 10분간 원심분리하여 상층액은 제거하
고 펠렛만을 얻어 완충용액B(300mM sucrose, 2mM HEPES, pH 7.4, 0.1mM EGTA)에 균
질화하여 3,500g에서 10분간 원심분리해서 미토콘드리아를 얻었다. 미토콘드리아는 단백질
용해액(7M urea, 2M thiourea, 4% CHAPS, 40mM Tri, 1% DTT, protease inhibitor
cocktail)에 녹였으며, 단백질의 농도는 Bradford assay 용액을 이용하여 분석되었다.
(3) 2차 전기영동법과 질량분석기를 이용한 단백질 동정
미토콘드리아 단백질(200ug)은 단백질의 등전점에 따른 분리가 가능한 IPG strip을 이용
하여 1차 전기영동을 하였다. 1차 전기영동은 pH 3-10 범위에서 단백질의 분리가 가능한
IPG strip을 이용하였으며, 2차 전기영동은 12.5% SDS-PAGE를 이용하여 단백질의 분자량
에 따른 분리를 수행하였다. 미토콘드리아 단백질의 동정은 SDS-gel상에서 단백질만을 추
출하여 MALDI-TOF/TOF mass spectrometer를 이용하였다.
3. 당뇨모델 생쥐의 차등발현 단백체 분석
가. 실험방법
(1) 재료 및 개요
제 2협동과제에서 생산한 Jazf 1 과발현 당뇨질환 생쥐의 간조직 단백체 분석을 수행하
였다. Jazf 1 과발현된 생쥐와 일반생쥐의 간에서 차등 발현하는 단백질을 확인하고자 하였
다. 생쥐로부터 적출한 간조직은 액체질소에 급속동결 후 단백질을 정제하여 시료로 사용
하였다.
(2) 조직으로부터 단백질 분리 및 2DE
Jazf 1 과발현 당뇨모델 동물의 단백체 발현 특성은 2차 전기영동법을 이용한 형질전환
생쥐의 간조직 단백체 분석을 하고자 하였다. 간조직에서 분리된 전체 단백질은 등전점에
따른 분리가 가능한 IPG strip을 이용하여 1차 전기영동을 하였다. 1차 전기영동은 pH
3-10, 5-8 범위에서 단백질의 분리가 가능한 IPG strip을 이용하였으며, 2차 전기영동은
SDS-PAGE를 이용하여 단백질의 분자량에 따른 분리를 수행하였다.
4. 당뇨관련 신규 표지분자의 발굴 및 이용성 검토
가. 실험방법

- 68 -
(1) 재료 및 개요
중성지방의 합성과 당 수송능력의 변화를 확인 할 수 있는 간세포(HepG2)와 근육세포
(L6)를 이용하였다. GPAT의 활성을 억제하는 화합물을 처리하여 당뇨병증의 완화효과를
확인 하고자 하였다. 본 연구의 결과는 GPAT 유전자를 이용하여 당뇨질환 모델을 개발하
였을 때 당뇨질환 개선용 신약의 검정에 효과적으로 사용할 수 있음을 시사 할 수 있다.
(2) 간세포주 배양
인간 간세포인 HepG2 세포는 ATCC (American Type Culture Collection, USA)에서 분
양 받았으며, 세포는 L-glutamine, non-essential amino acids, sodium bicarbonate, 10 %
(v/v) fetal bovine serum (FBS), 1 % antibiotics, 1 mM sodium pyruvate가 첨가된 MEM
를 배양 배지로 37℃, 5% CO2 조건 하에서 배양하였다. Pea15 유전자 발현벡터를 간세포
에 형질전환 후에 12 or 24-well 배양용기에서 1-2 일 배양 후 실험에 사용하였다.
(3) 간세포에서의 중성지방 생합성
HepG2 cell (5×105 cells/well)을 24 well plate에 분주하고 1-2일 동안 배양한 후 0.25
mCi [14C]acetate 또는 [14C]glycerol 를 첨가하고 24간 동안 반응시켰다. 반응 후, 세포는
추출용매 (Hexane : 2-propanol = 3 : 2, v/v) 0.5 ml을 첨가하여 중성지방을 포함하는 총
지방을 추출하였으며, 추출용매를 제거한 전체 지방은 TLC (silica gel 60F254, 0.5 mm,
Merck)에 점적하여 전개용매 (Hexane : Diethyl ether : Acetic acid = 80 : 20 : 1, v/v)상
에서 전개하였다. TLC상에서 중성지방을 분리한 후 TLC는 방사선원소의 에너지를 감광
할 수 있는 필름에 감광한 후 이미지 분석(BAS-1500, Fuji Photo Film Co. Ltd.)을 통해
중성지방의 방사선량을 측정하였다. 추출 후 세포의 단백질 농도를 측정하였다. 중성지방의
방사능 양을 측정하여 단백질 농도로 나눈 값을 실험값으로 하여 각 실험군사이의 실험적
오차를 보정하였다.
(4) 근육세포주 배양
L6 세포는 Dulbecco s̀ modified Eagle s̀ medium (DMEM; 4mM L-glutamine, 1.5g/L
sodium bicarbonate, 4.5g/L glucose)에 FBS 10%를 포함하는 배양액에 배양한다. 당 수송
능력 변화 실험을 위해서 세포는 80% confluent 상태에서 PBS 완충액으로 세포를 세척한
후 0.5% FBS DMEM(4mM L-glutamine, 1.5g/L sodium bicarbonate, 1g/L glucose) 배지
로 교환하여 24시간 배양하여 실시되었다.
(5) 근육세포에서의 Glucose uptake 분석
L6 세포에 인슐린 호르몬을 일정시간 처리한 후 Krebs-Ringer Buffer (KRB)에서 30분
배양한다. KRB 완충액에서 배양된 세포에 0.1mM 2-deoxy-D-glucose 와
2-deoxy-D-[14C]glucose (0.2uCi/ml)를 처리한 후 10분 동안 반응시킨다. 반응이 끝난 세
포에서 완충용액을 제거하고 세포를 세척한 후 1M NaOH로 세포를 용해한다. 용해된 세포

- 69 -
는 scintillation counter로 측정되었다.
나. 결과
(1) 돼지근육 미토콘드리아의 단백체 발현 특성
돼지 근육의 미토콘드리아로부터 단백체 분석은 2차원 전기영동법과 질량분석기를 이용
하여 실시되었다. 단백체의 검출양상은 매우 선명하게 관찰되었으며, 대부분의 spot들이 동
정되었다 (그림43). 동정된 단백질 144개의 목록은 별첨하며 일부 단백질의 종류와 기능은
표5에 실려 있다. 동정된 단백질의 약60%는 대사에 관련된 효소들이며, 그 밖에 세포구조
성분, 물질운반체, 신호전달, heat shock protein 및 chaperone에 관련된 단백질로 구성되
어 있음을 알 수 있었다 (그림44). 돼지의 발현 단백체 중에서 특히 근육 미토콘드리아의
단백체에 관한 정보는 상당부분 보고가 되어있지 않다. 이와 같은 사실은 질환모델 가축의
분자생화학적 발병기작을 연구하는데 많은 어려움을 예상케 한다. 그럼으로, 본 실험에서
확보된 돼지 근육의 미토콘드리아 단백체 정보는 질환모델의 발병 및 대사이상에 관한 기
작을 연구하는데 중요한 정보로서 이용이 가능하리라 사료된다.
그림43. 돼지 근육의 미토콘드리아 단백체 지도
그림44. 돼지 근육의 미토콘드리아 단백체의 기능

- 70 -
표6. 돼지 근육의 미토콘드리아 단백질의 동정
Spot
No.Protein Name
Accession
No.(1)
Mr pI Function
51Voltage-dependent anion-selective
channel protein 1 (VDAC-1) P45879 30675.63 8.84 Calcium signaling pathway
75Probable mitochondrial import
receptor subunit TOM40 homologO96008 37869.14 6.79 Receptor of protein precursors
137Succinate dehydrogenase
[ubiqu -inone] flavoprotein subunitP31039 72897.18 7.55
Carbohydrate metabolism, oxidative
phosphorylation
142 Glycerol-3-phosphate dehydro genase P43304 80763.53 6.98 Glycerol metabolism
55Voltage-dependent anion-selective
channel protein 2 P68003 31560.56 7.49 Calcium signaling pathway
135 Heat shock cognate 71 kDa protein P11142 70854.22 5.37 Chaperone; endogenous inhibitory
regulator of HSC70
140NADH-ubiquinone oxidoreductase 75
kDa subunitQ91VD9 79697.54 5.51
Oxidative phosphorylation, ubiquinone
biosynthesis
138Stress-70 protein
(75 kDa glucose regulated protein)P38647 73482.73 5.91
Chaperone, cell proliferation and
cellular aging
41Thioredoxin-dependent peroxide
reductaseQ91VD9 28177.42 7.15 Redox regulation of the cell
9 Cytochrome c oxidase polypeptide Va P11240 16119.31 6.08 Metabolism; oxidative phosphorylation
61Voltage-dependent anion-selective
channel protein 1Q60932 32331.38 8.55 Calcium signaling pathway
30 Heat-shock protein beta-1 (HSP 27) P42929 22924.66 6.23 Stress resistance and actin
organization
(1) Accession number is SWISS-PROT and NCBI database number.
(2) 당뇨모델 생쥐의 차등발현 단백체 분석
형질전환 동물모델에서의 당뇨 표지유전자와 단백질의 발현량과 활성변화를 연구하여 질
환모델로서의 가치 평가와 질환발병의 원인기작 연구를 수행하였다. 형질전환 동물 (Jazf 1
과발현 당뇨모델)의 간 조직의 단백질은 2DE 프로테옴 실험기법을 통하여 분석하였고, 형
질전환 동물에서 차등발현되는 단백질을 탐색하였다. Jazf 1 과발현 당뇨모델 마우스에서는
새롭게 검출되는 단백질 1종과 높게 발현되는 단백질 1종을 발굴하였다 (그림45).

- 71 -
그림45. Jazf 1 과발현 생쥐 간조직의 차등발현 단백질
(3) 당뇨관련 신규 표지분자의 발굴 및 이용성 검토
중성지방 합성효소로써 기능연구가 시작된 지 약 20년 정도이면, 비만/당뇨치료를 위한
저해제 개발이 활발히 이루어지고 있다. 이것은 GPAT의 분자타겟으로써의 유용성을 암시
하고 있으며 현재 캐나다 National Research Council, Xenon Tegenetics 등에서 연구가 수
행 중에 있다. 따라서 향후 전 세계적으로 개발될 것으로 사료되는 본 저해제의 효능 검증
을 위한 적절한 모델 동물이 필요할 것으로 생각된다. 아래 조사한 결과는 천연물에서 분
리된 GPAT 저해제에 의한 근육세포에서의 당 수송능이 증가함을 보여준 결과이다. 즉
GPAT을 과발현시키면 이러한 저해제는 간세포에의 중성지방 생합성을 감소시킴을 확인하
였고 (그림46), 또한, 당 수송능이 떨어질 것으로 예상되는 결과이다 (그림47).
그림46. 간세포에서의 GPAT 저해제의 중성지방 생합성의 감소 효과

- 72 -
그림47. 근육세포에서 GPAT 저해제(Compound)에 의한 당 수송능력

- 73 -
제7절 질환모델 형질전환 생쥐의 개발
1. 형질전환 생쥐의 생산 및 특징
가. 실험방법
(1) 유전자의 선별
비만 및 합병증 관여할 것으로 예상되는 후보 마커 유전자를 선별하였다. 논문 검색을 통
해서 혹은 타 세부과제 연구결과를 바탕으로 비만이 유도되었을 때 발현에 큰 유의차를 보
이는 유전자를 중심으로 범위를 줄여나갔다. 선별 과정에서 현재까지 형질전환 동물모델로
확립되어 연구되지 않은 유전자를 중심으로, 형질전환 동물을 만들었을 때 적절한 이상여
부를 보일 것으로 예상되는 유전자를 중심으로, 최종적으로 비만과 그에 대한 합병증 유도
에 관여할 것으로 예상되는 유전자 Biliverdin reductase (BVR), Cathepsin S (CTSS) 및
JAZF zinc finger 1 (Jazf1)를 선별하였다.
CTSS는 cysteine protease로서 지방분화과정에서 초기에 분비되어 미성숙지방세포 세포
막의 구성 성분 또는 세포외기질을 분해하여 성숙한 지방세포가 형성되도록 도와주는 역할
을 한다고 보고된 바 있다. 따라서, CTSS 유전자를 과할현 하는 형질전환 모델동물을 확
립하여 고지방식이를 유도하면 정상 마우스에 비하여 미성숙지방세포가 쉽게 성숙한 지방
세포로 분화되어 정상에 비하여 쉽게 비만이 유도 될 것으로 예상된다.
(2) 발현벡터의 구축
CTSS 유전자의 ORF를 클로닝하기 위하여 사람의 태반 조직에서 cDNA를 확보하였다.
태반 조직의 cDNA를 사용하여 선택한 유전자에 특이적인 프라이머를 제작하였고, PCR 방
법을 통해 유전자의 ORF를 클로닝하였다. 사용한 프라이머의 서열은 CTSS ORF-F :
5'-agc aac tgc tgg ttc tat cac-3' , CTSS ORF-R : 5'-agc aac tgc tgg ttc tat cac-3. 클
로닝한 후, pEGFP.N1 발현 벡터와 연결하여 벡터를 구축하고 연결된 여부를 제한효소를
통하여 확인한 뒤, 최종적으로 유전자 서열 분석을 통하여 적합성 여부를 판단하였다.
(3) 타겟 유전자의 과발현 형질전환 모델동물의 생산
형질전환동물의 생산은 통상의 방법에 준하여 실시하였다. 간단히 약술하면, 2IU의
PMSG와 hCG을 이용하여 과배란된 생쥐로부터 1 세포기 수정란을 회수한 후, effendorf사
의 manuplation을 이용하여 웅성전핵에 약 2pl정도의 유전자를 도입한 후, 정관 결찰된 숫
컷과 교배한 가임신 생쥐의 난관에 이 수정란을 이식함으로서 생산하였다.
(4) 형질전환 생쥐의 확인 및 육종
태어난 마우스 가운데에서 형질 전환된 생쥐를 선별하기 위해, 마우스의 꼬리에서
genomic DNA를 추출하였다. 미세주입한 벡터 시퀀스에 특이적인 프라이머를 제작하여 추
출한 genomic DNA를 template로 PCR을 수행하였다. 프라이머의 서열은 pEGFP-N1-F ;

- 74 -
5'-cgt gta cgg tgg gag gtc ta-3' , pEGFP-N1-R ; 5'-tgg tgc aga tga act tca gg-3' 이
며, PCR은 Takara Thermal Cycler를 사용하여 94°C 30 sec, 57°C 30 sec, 72°C 40 sec의
조건으로 30 싸이클 돌려 확인하였다. 전기영동을 하여 밴드의 유무를 positive control 의
밴드와 비교 확인한 뒤, 형질전환 생쥐를 구분하여 사육하였다.
(5) Reverse transcriptase polymerase chain reaction (RT-PCR) 분석
H9C2 세포 및 마우스 심장 조직에서 cDNA를 합성하여 이를 template로 apoptosis 관련 유
전자 (caspase-3, -8, -9, -12, Bax, Bim, Apaf1, Aifm1)의 발현 패턴을 비교 분석 해 보았다.
PCR 한 샘플을 젤 로딩 하여 밴드의 정량적인 밝기 비교를 위하여 Image J 프로그램을 사용
하였다.
(6) 세포배양 및 transfection
마우스 배아 심장 유래의 세포인 H9C2 세포주 (한국세포주은행)를 이용하여 Jazf1 과발현
벡터를 transfection 하였다. H9C2 세포는 DMEM 배지에 10% FBS, 100 units/ml penicillin
과 100 ug/ml streptomycin을 첨가한 세포배양액을 사용하여 37℃, 5% CO2 incubator에서 배
양하였다. Back-bone 벡터인 pEGFP.N1 과 Jazf1 과발현 벡터를 lipofectamine 2000을 사용하
여 transfection 하였다.
(7) Quantitative real-time PCR 분석
정상과 Jazf1 형질전환 마우스의 심장조직에서 RNA를 추출한 후 cDNA를 합성하였다.
Apoptosis 관련 유전자의 발현 패턴을 분석하기 위하여 RT2 ProfilerTM PCR Array
System을 사용하였다. 확인된 유전자의 발현은 GAPDH 유전자의 발현을 기준으로 정량하
였다.
(8) Jazf1 과발현 형질전환 마우스 심장의 기능 분석
당뇨와 심장질환은 매우 밀접한 연관이 있다고 알려져 있기 때문에, 확립된 Jazf1 형질전환
생쥐 심장에서의 기능에 있어서 이상 여부를 확인하였다.
(가) 심전도 측정
실험에 사용된 생쥐들을 마취시킨 후 전극을 연결하여 심전도를 측정해 보았다.
(나) 심박수 측정
(다) 운동능력의 측정
간접적인 심장기능의 확인을 위해 트레드밀을 통한 생쥐의 운동능력을 측정해 보았다.
(라) 혈압측정
심전도를 통한 심장박동의 이상과, 트레드밀을 통한 운동능력의 저하를 확인하면서, 형질

- 75 -
전환 동물과 정상 생쥐의 혈압의 변화를 측정해 보았다.
(9) 13.5일 배아의 심장 발달 비교
심장 발달 초기에 Jazf1 유전자의 영향을 알아보기 위하여, 13.5일째 배아의 심장 모습을
조직학적 분석 해보았다
(10) 성체 생쥐의 심장 발달 비교
심장 발달 초기에 있어서의 영향이 성체 조직에 어떠한 영향을 주었는지를 확인하기 위하
여, 4개월 된 형질전환 생쥐와 동일한 주령의 정상 생쥐의 심장조직을 비교 분석 해 보았다.
심장조직을 적출한 후 파라핀 섹션을 통하여 Hematoxylin and Eosin 염색을 수행하였다.
(11) 심근조직의 세포사멸
심근조직의 치밀하지 못함과 심실벽이 얇아지는 등의 병변의 원인을 확인하기 위하여, 심근
조직의 세포사멸을 Terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling
(TUNEL) 염색을 통하여 확인해 보았다.
(12) 미토콘드리아의 병변 확인
심근조직의 세포사멸로 인해 심근세포의 미토콘드리아의 이상 여부를 전자현미경을 통하여
확인해 보았다.
(13) Reverse transcriptase polymerase chain reaction (RT-PCR) 분석
형질전환 시킨 유전자의 발현 패턴을 분석하기 위하여 형질전환 생쥐와 littermate 정상
생쥐를 희생시켜 다양한 조직들을 떼어내었다. 비교 분석을 위해 피부, 심장, 간, 폐, 신장,
뇌, 고환, 백색지방, 갈색지방의 조직에서 cDNA를 합성하였다. 여러 장기 조직의 cDNA를
template로 사용하여 Cathepsin S의 발현 정도를 비교 분석하기 위하여 미세주입 한 벡터
에 특이적인 프라이머를 제작하였다. 사용한 프라이머 서열은 Cathepsin S-F ; 5’-tgg atc
acc act ggc atc tct-3’, Cathepsin S-R ; 5’-tga aca tgt ggc acg agc at-3’. PCR 방법을
통하여 94°C for 30 sec, 59°C for 30 sec, 72°C for 30 sec 조건으로 30 사이클 증폭하였
다. 정량적인 분석을 위하여 GAPDH (GAPDH-F: 5’-aat gca tcc tgc acc acc aa-3’,
GAPDH-R: 5’-gta gcc ata ttc att gtc ata-3’) 유전자를 함께 PCR 하여, Cathepsin S 유전
자와 정량적인 비교분석을 수행하였다. PCR 한 샘플을 젤 로딩 하여 밴드의 정량적인 밝
기 비교를 위하여 Image J 프로그램을 사용하였다.
(14) RT-PCR 분석을 통한 피부조직에서의 cytokine 발현 비교
16주령의 형질전환 생쥐와 주령이 같은 정상 생쥐의 등 피부 조직을 떼어낸 후 cDNA를
합성하였다. 이것을 template로 사용하여 IL-4, IL-6, IL-10, TNFα. IFN-γ, IL-2, IL-1β의
다양한 cytokine mRNA 발현과 MCP-1 그리고 GM-CSF인 chemokine의 발현을 확인하였
다. 정량적인 분석을 위하여 GAPDH 유전자를 함께 PCR하여 비교분석을 수행하였다. PCR

- 76 -
한 샘플을 젤 로딩 하여 밴드의 정량적인 밝기 비교를 위하여 Image J 프로그램을 사용하
였다.
(15) Quantitative real-time PCR (Real-time PCR) 분석
아토피 병증의 경도에 따라 Cathepsin S의 발현 강도 차이를 분석하기 위하여 Real-time
PCR을 수행하였다. 정상 생쥐와 littermate 형질전환 생쥐를 희생시켜 등 피부의 조직을 떼
어냈으며, 형질전환 생쥐 경우 아토피 병증의 정도가 약한 쥐와 강한 쥐를 구분하여 조직
을 떼어냈다. 이 분석은 Step One Plus PCR system을 이용하였으며, SYBR green을 함께
사용하였다. 그리고 Cathepsin S의 발현을 살펴보기 위하여 사용된 프라이머 서열은
Cathepsin S-F ; 5’-cgg ctg gtt tgt gtg ctc tt-3' , Cathepsin S-R ; 5’-aga gat gcc agt
ggt gat cca-3' 이다. 먼저 cDNA를 denaturation 시키기 위하여 95°C for 30 sec, 이어서
95°C for 5 sec, 60°C for 30 sec, 72°C for 10 sec 조건으로 50 사이클 증폭하였음. 정량적
인 분석을 위하여 GAPDH 유전자를 함께 분석하였다. 각 샘플의 Cathepsin S 그리고
GAPDH의 발현을 비교하기 위하여 Step One software Version 2.1을 사용하였다.
(16) Cathepsin S 과발현 형질전환 생쥐의 피부의 조직학적 분석
형질전환 생쥐의 귀와 등 피부조직에서의 이상여부를 확인하기 위하여 조직학적 분석을 수행하였
음. 16주령의 형질전환 생쥐와 주령이 같은 정상 생쥐의 조직을 떼어낸 후 하루 동안 Bouin 용액으
로 고정시켰음. 고정된 조직을 파라핀 왁스에 담근 후 5μm 얇기로 조직을 섹션하였다. 섹션한 조직
은 Hematoxylin and eosin 염색을 통하여 조직의 형태적인 면을 확인하였으며, Toluidine blue 염색
을 통하여 조직 내의 비만 세포가 유입된 정도를 파악하고 이를 카운팅하여 도표화하였다.
(17) Cathepsin S 과발현 형질전환 생쥐의 피부의 면역학적 분석
형질전환 생쥐의 등 피부조직에서 Cathepsin S, PAR2, MHC II class, CD11c, CD4,
CD68 유전자의 발현을 살펴보기 위하여 면역학적 분석을 수행하였다. 16주령의 형질전환
생쥐와 주령이 같은 정상 생쥐의 등 피부 조직을 떼어낸 후 하루 동안 Bouin 용액으로 고
정시켰다. 고정된 조직을 파라핀 왁스에 담근 후 5μm 얇기로 조직을 섹션하였다. 섹션한
조직은 0.01M의 citrate 용액(pH6.0)에서 3분 동안 cooking 후, phosphate-buffered
saline(PBS) 용액으로 씻어냈다. 다음으로 언급한 유전자의 primary antibody와 이에 알맞
은 secondary antibody를 사용하여 면역염색을 수행하였다.
(18) 조직 내의 cytokine 분석
16주령의 형질전환 생쥐와 주령이 같은 정상 생쥐의 등 피부 조직을 떼어낸 후 단백질을
추출하였다. Sandwich 방식의 ELISA를 통하여 IL-4, IL-6, IL-10, TNFα. IFN-γ, IL-2,
IL-1β를 측정하였다. 먼저 96well plate에 capture antibody를 코팅하여 하루 정도 붙여두
었다. 세척과 blocking 단계를 거친 후 추출한 단백질과 standard sample을 2시간 동안
plate에 코팅 시켰다. 다음으로 detection antibody를 코팅시킨 후 일반적인 ELISA 방법과
동일하게 450nm 550nm에서 측정하였다.

- 77 -
(19) 혈청에서의 IgE 분석
16주령의 형질전환 생쥐와 주령이 같은 정상 생쥐의 혈액을 채취한 후 3000rpm에서 15분
동안 원심분리를 하여 얻은 혈청을 추출하였음. Sandwich 방식의 ELISA를 통하여 IgE 발
현을 측정하였다. 먼저 96well plate에 capture antibody를 코팅하여 하루 정도 붙여두었다.
세척과 blocking 단계를 거친 후 추출한 단백질과 standard sample을 2시간 동안 plate에
코팅 시켰다. 다음으로 detection antibody를 코팅시킨 후 일반적인 ELISA 방법과 동일하
게 450nm 550nm에서 측정하였다.
나. 결과
(1) 벡터구조 및 형질전환 검정
CMV 프로모터를 사용하여 발현 벡터를 구축하고, 미세주입 방법을 통하여 형질전환 생
쥐를 생산하였다. 생쥐의 꼬리에서 DNA를 뽑아 형질전환 여부를 확인하였다. 최종적으로
9마리의 BVR 형질전환 생쥐를 확립하였다 (그림48).
그림48. Biliverdin reductase (BVR) 발현벡터 구조 및 형질전환 생쥐 생산
CMV 프로모터를 사용하여 발현 벡터를 구축하고, 미세주입 방법을 통하여 형질전환 생
쥐를 생산하였다. 생쥐의 꼬리에서 DNA를 뽑아 형질전환 여부를 확인하였다. 최종적으로
5마리의 CTSS 형질전환 생쥐를 확립하였다 (그림49).

- 78 -
그림49. Cathepsin S (CTSS) 발현벡터 구조 및 형질전환 생쥐 생산
CMV 프로모터를 사용하여 발현 벡터를 구축하고, 미세주입 방법을 통하여 형질전환 생
쥐를 생산하였다. 생쥐의 꼬리에서 DNA를 뽑아 형질전환 여부를 확인하였다. 최종적으로
2마리의 Jazf1 형질전환 생쥐를 확립하였다 (그림50). 당뇨와 심장질환은 매우 밀접한 연관
이 있다고 알려져 있기 때문에, 확립된 Jazf1 형질전환 생쥐 심장에서의 이상 여부를 확인
하였다. 그 결과 아래와 같이 혈압이 높았으며, 심장 박동수가 정상 생쥐에 비해 빠름을 확
인하였다 (그림51).
그림50. JAZF zinc finger 1 (Jazf1) 발현벡터 구조 및 형질전환 생쥐 생산
그림51. Jazf1 형질전환 생쥐의 혈압 및 심박수
(1) Jazf1 과발현 형질전환 마우스의 발현 패턴 분석
형질전환 시킨 Jazf1 유전자의 발현 패턴을 분석하기 위하여 형질전환 생쥐와 littermate

- 79 -
정상 생쥐를 희생시켜 다양한 조직들을 떼어내었다. 비교 분석을 위해 뇌, 심장, 간, 폐, 고
환, 비장, 신장, 췌장 등의 조직에서 과발현 벡터가 제대로 작동하였는지, 발현정도를
GAPDH 유전자와 비교하여 정량적인 분석을 해보았다. Jazf1 유전자는 정상인 생쥐의 11.5
일째 배아에서부터 발현을 시작하였으며, 이후로의 발달단계에서의 발현은 대체적으로 변
함없이 일정하였다. 확립한 형질 전환 생쥐와 정상생쥐에서의 Jazf1 유전자의 발현을 비교
분석한 결과, 확립된 형질전환 생쥐에서 Jazf1 유전자의 발현이 조직 전반적으로 고르게 제
대로 과발현 됨을 확인하였다 (그림52).
그림52. Jazf1 과발현 형질전환 마우스의 발현 패턴 분석
(2) Jazf1 과발현 형질전환 마우스 심장의 기능 분석
당뇨와 심장질환은 매우 밀접한 연관이 있다고 알려져 있기 때문에, 확립된 Jazf1 형질전
환 생쥐 심장에서의 기능에 있어서 이상 여부를 확인하였다. 살아있는 상태에서의 심장의
기능적인 면을 측정하기 위하여 심전도를 측정하여 심박수 등의 심장의 기능적인 측정값들
을 확인하였으며, 운동능력의 측정을 통해 지구력을 알아보았다. 또한, 형질전환 마우스의
운동능력이 정상 생쥐에 비하여 떨어짐을 확인하여, 혈압의 차이도 확인해 보았다.
(가) 심전도 측정
실험에 사용된 생쥐들을 마취시킨 후 전극을 연결하여 심전도를 측정해 보았다. 그 결과,
Jazf1이 과발현 된 형질전환 생쥐의 심장 박동이 정상 생쥐에 비하여 매우 빨랐으며, 불안
정한 박동 모습을 확인 할 수 있었다 (그림53A). 심전도 측정을 통하여 형질전환 생쥐의
심장 박동은 정상에 비하여 적은 양의 혈액을 내 보내기 위해 심장은 오히려 더욱 빨리 뛰
고 있다는 것을 알 수 있었으며, 피크의 높이가 일정하지 않음을 통해 심장의 수축과 이완
기능에 문제가 있음을 확인하였다.

- 80 -
(나) 심박수 측정
심전도 측정과 더불어 심박수 역시 Jazf1 유전자가 과발현 된 생쥐에서는 정상 생쥐에 비
하여 심박수가 유의차 있게 증가해 있음을 확인할 수 있었으며, R 값이 감소됨을 통하여
수축할때의 혈액의 양은 많지 않다는 것을 알 수 있었다 (그림53B).
(다) 운동능력의 측정
간접적인 심장기능의 확인을 위해 트레드밀을 통한 생쥐의 운동능력을 측정해 보았다. 오
랫동안 트레드밀 바퀴 위를 뛸 수 있는 능력을 측정 해 봄으로써 동물의 지구력을 측정해
볼 수 있다. 그 결과, Jazf1 유전자가 과발현 된 형질전환 모델 동물의 경우 정상 생쥐에
비하여 지구력이 유의차 있게 감소함을 확인하였다 (그림53C).
(라) 혈압측정
심전도를 통한 심장박동의 이상과, 트레드밀을 통한 운동능력의 저하를 확인하면서, 형질
전환 동물과 정상 생쥐의 혈압의 변화를 측정해 보았다. 그 결과, Jazf1 유전자가 과발현
된 형질전환 생쥐의 혈압이 정상 생쥐에 비해 수축기, 이완기 혈압 모두에서 높게 측정 되
었다 (그림53C). 이러한 결과는 심장박동의 이상으로 인해 수축과 이완이 불안정 하여, 혈
액의 공급이 원활하지 못하여 혈압이 낮아지는 현상을 동반 한 것으로 예상된다.
그림53. Jazf1 과발현 형질전환 마우스 심장의 기능 분석
(3) Jazf1 과발현 형질전환 마우스 심장의 조직학적 분석
(가) 13.5일 배아의 심장 발달 비교
심장 발달 초기에 Jazf1 유전자의 영향을 알아보기 위하여, 13.5일째 배아의 심장 모습을

- 81 -
조직학적 분석 해보았다. Jazf1 유전자가 과발현 된 형질전환 생쥐의 경우 정상 생쥐에 비
하여 심장 발달이 늦게 진행되고 있음을 확인하였다. 정상 생쥐의 배아 심장의 경우 크기
도 컸으며, 심방과 심실의 모습이 어느정도 구분되어 있음을 확인 할 수 있으나, 형질전환
생쥐의 경우에는 심장의 발달 속도도 느렸으며, 심방과 심실의 구분도 되지 않고 있음을
확인하였다 (그림54A). 이러한 결과를 보았을 때 Jazf1 유전자의 과발현은 생쥐 심장의 발
달 초기에서부터 영향을 주고 있음을 알 수 있었다.
(나) 성체 생쥐의 심장 발달 비교
심장 발달 초기에 있어서의 영향이 성체 조직에 어떠한 영향을 주었는지를 확인하기 위
하여, 4개월 된 형질전환 생쥐와 동일한 주령의 정상 생쥐의 심장조직을 비교 분석 해 보
았다. 심장조직을 적출한 후 파라핀 섹션을 통하여 Hematoxylin and Eosin 염색을 수행하
였다. 그 결과, 전반적인 심장의 크기의 차이는 보이지 않았으나, 형질전환 생쥐의 심장의
심실벽이 정상 생쥐의 심실벽에 비하여 크게 얇아져 있음을 확인하였다. 또한, 형질전환 생
쥐의 심근조직이 치밀하지 못함을 확인하였다 (그림54B). 이러한 심장의 모습은 전형적인
심부전의 모습을 보이고 있었다. 이러한 결과는 Jazf1 유전자의 발현은 초기 심장 발달에
크게 영향을 주었으며, 성체가 되었을 때 심장의 발달에 까지 영향을 주었음을 확인할 수
있었다. 하지만, 이러한 Jazf1 유전자의 과발현에 대한 영향이 심장이 발달하지 못할 정도
의 조절은 하지 않는다는 것을 알 수 있었다.
그림54. Jazf1 과발현 형질전환 마우스 심장의 조직학적 분석
(4) 심근조직의 세포사멸의 확인 및 미토콘드리아의 병변 확인
(가) 심근조직의 세포사멸

- 82 -
심근조직의 치밀하지 못함과 심실벽이 얇아지는 등의 병변의 원인을 확인하기 위하여, 심
근조직의 세포사멸을 Terminal deoxynucleotidyl transferase-mediated dUTP nick
end-labeling (TUNEL) 염색을 통하여 확인해 보았다. 정상 생쥐의 심근조직을 확인하면,
심근조직이 치밀하며 심실벽도 두껍게 형성 되어 있음을 확인 할 수 있으며, 또한, TUNEL
염색 결과 세포사멸의 모습도 보이지 않음을 알 수 있다. 하지만, Jazf1 유전자의 과발현으
로 인한 형질전환 생쥐의 심근조직의 경우 조직이 치밀하지 않았으며, 심실벽이 매우 얇아
져 있음을 알 수 있다. TUNEL 염색 결과 형질전환 생쥐의 경우 검정색 반점으로 표현되
는 세포사멸의 모습을 확인 하였다 (그림55A).
(나) 미토콘드리아의 병변 확인
심근조직의 세포사멸로 인해 심근세포의 미토콘드리아의 이상 여부를 전자현미경을 통하
여 확인해 보았다. 정상 생쥐의 심근세포 미토콘드리아의 모습은 근육세포와 나란하게 일
정한 모습의 배열을 보였으나, Jazf1 유전자가 과발현 된 형질전환 생쥐의 경우, 근육세포
의 배열과는 불규칙하게 무작위하게 배열되어 있었으며, 미토콘드리아의 모습 역시 많이
부풀어져 있음을 확인 하였다 (그림55B). 이러한 미토콘드리아의 병변으로 인하여 심근세
포로의 에너지 공급에 차질이 생기게 되어 심장의 수축과 이완에 문제가 있게 됨을 예상해
볼 수 있었다.
그림55. 심근조직의 세포사멸의 확인 및 미토콘드리아의 병변 확인
(5) CTSS 과발현 형질전환 마우스의 발현 패턴 분석
형질전환 시킨 CTSS 유전자의 발현 패턴을 분석하기 위하여 형질전환 생쥐와 littermate
정상 생쥐를 희생시켜 다양한 조직들을 떼어내었다. 비교 분석을 위해 피부,뇌, 심장, 간,
폐, 고환, 신장, 백색지방, 갈색지방 등의 조직에서 과발현 벡터가 제대로 작동하였는지, 발
현정도를 GAPDH 유전자와 비교하여 정량적인 분석을 해보았다. 확립한 형질 전환 생쥐와
정상생쥐에서의 CTSS 유전자의 발현을 비교 분석한 결과, 확립된 형질전환 생쥐에서
CTSS 유전자의 발현이 조직 전반적으로 고르게 제대로 과발현됨을 확인 하였다 (그림56).

- 83 -
그림56. CTSS 형질전환 마우스의 발현 패턴 분석
(6) CTSS 형질전환 마우스의 피부병 발병확인
유전자의 기능 분석을 위하여 20% 고지방식이를 통한 비만을 유도하였다. 그 과정에서
형질전환 마우스에 특이적으로 피부병의 병변이 발견되었다. 전염성은 없었으며, 피부가 심
하게 짓무르면서 가려움증을 유발하는 것으로 확인하였다 (그림57).
그림57. CTSS 형질전환 마우스의 피부병 발병
(7) CTSS 형질전환 마우스의 피부와 귀 조직의 조직학적 분석
정상과 형질전환 마우스의 피부와 귀 조직을 적출하여 조직학적 분석을 수행하였다.
H&E 염색 결과 형질전환 마우스의 진피와 내피가 많이 두꺼워 져 있었다. 특히, 내피조직
의 보라색으로 염색된 점들은 면역반응으로 인한 면역세포들의 유입으로 보이며, 조직손상
이 심하게 확인되었다 (그림58).
그림58. 피부와 귀 조직의 조직학적 분석
피부 손상으로 염증성 세포들의 유입이 심하게 나타난 것으로 생각하여, mast cell 의 유

- 84 -
입정도를 toluidine blue 염색을 통하여 확인해 보았다. 그 결과 형질전환 마우스의 피부 및
귀 조직에 파란색 반점들이 확연하게 증가함을 확인 하였으며, mast cell 의 개수를 비교해
본 결과 형질전환 마우스의 조직에서 유의차 있는 증가를 확인 하였다 (그림59).
그림59. 피부 및 귀 조직에서의 mast cell 확인
(8) Jazf1 형질전환 마우스 고지방식이를 통한 식이성비만 유도
체중 및 혈당 측정 ; 5주령 생쥐로부터 고지방식이(20% fat)를 통한 식이성 비만을 유도.
정상에 비하여 형질전환 생쥐의 경우 체중증가량이 적음을 확인하였다 (그림60왼쪽). 공복
시 혈당측정 결과 정상에 비하여 형질전환 생쥐의 경우 전반적으로 낮은 혈당수치를 보였
다 (그림60오른쪽).
그림60. Jazf1 형질전환 생쥐의 체중증가 및 혈당변화
생쥐의 간조직 에서 유전자 발현을 비교하였다. Jazf1 형질전환 생쥐의 혈당 감소를 확인
하기 위하여 당신생 관련 유전자인 PEPCK의 발현을 확인하였다. 정상에 비하여 형질전환
생쥐에서 PEPCK의 발현이 낮게 발현됨. TR4의 발현에는 차이가 없었다 (그림61).

- 85 -
그림61. Jazf1 형질전환 생쥐의 간조직에 있어 TR4 및 PEPCK의 발현양상
(9) CTSS 형질전환 마우스
확립한 CTSS 형질전환 생쥐의 경우 식이를 통한 비만 유도 과정 중 피부병변이 확인되
었으며, 대사질환과 관련된 여러 가지 사이토카인의 발현을 확인하였다. 정상에 비하여 형
질전환 생쥐의 경우 관련 사이토카인의 발현이 높게 나타났다 (그림62).
그림62. CTSS 형질전환 생쥐의 다양한 사이토카인의 발현양상
생쥐의 serum에서의 사이토카인 발현은 형질전환 생쥐에서 관련 사이토카인의 발현이 높
게 확인되었다 (그림63).

- 86 -
그림63. CTSS 형질전환 생쥐의 혈청내 사이토카인의 발현 양상

- 87 -
제4장 연구개발목표 달성도 및 대외기여도
제1절 목표대비 달성도
구분 세부연구개발 목표 평가의 착안점 및 기준 달성도
1차년도(2009)
◦형질전환 돼지 증식◦당뇨의 생리학적 분석
◦개체 증식 유무◦인슐린 저항성 확인 유무
100%
◦과발현 재조합세포주 구축◦재조합세포주 당질대사시스템 특성
◦재조합세포주 구축◦재조합세포주의 당질대사분석◦연관성 해석
100%
◦장기 특이적 promoter 탐색 및 유전자 발현 벡터 구축
◦당뇨병 형질전환 생쥐 생산
◦발현 벡터 구축 여부◦형질전환 생쥐 생산 여부
100%
2차년도(2010)
◦조직학적 특성 확인◦신규발현 벡터 도입
◦당뇨의 조직면역학적 분석◦신규발현 벡터의 유용성
100%
◦재조합세포주의 지질대사시스템특성◦질환모델가축 발현전사체 특성
◦재조합세포주의 지질대사변화분석◦연관성 해석◦발현전사체 분석
100%
◦당뇨병 형질전환 생쥐 생산◦당뇨병 형질전환 생쥐의 유전자 기능 분석
◦형질전환 생쥐 생산 여부◦유전자 기능 분석 여부
100%
3차년도(2011)
◦타 질환모델과의 비교검토◦신규발현 벡터의 미세주입
◦당뇨 질환발병의 우수성◦미세주입 두수
미달성(연구내용변경)
◦재조합세포주의 이용성 검토◦질환모델가축 발현전사체 특성
◦당뇨치료물질의 효능 평가◦발현단백체 분석
100%
◦형질전환 모델생쥐을 이용한 발병원인 규명과 제어기능 해석
◦발병원인 규명과 제어 기능 해석 여부 100%
최종 평가
◦당뇨질환 모델 돼지의 개발◦신규 질환모델 돼지의 개발
◦당뇨질환 모델돼지의 유용성◦유전자 미세주입 여부
100%
(신규는 진행 중)
◦과발현 세포주 개발◦당뇨발현 신규 표지분자 발굴
◦과발현 세포주 개발 여부◦당뇨발현 신규 표지분자 발굴여부
100%
◦당뇨병 형질전환 생쥐 생산◦당뇨병 발병원인 규명 및 제어 기능 해석
◦형질전환 생쥐 생산 여부◦발병원인 규명과 제어 기능 해석 여부
100%
연도별 연구목표는 대부분 달성하였으며, 3차년도 타 질환모델과의 비교검토는 연
구내용의 변경으로 수행하지 않았다. 그리고 신규 질환모델 돼지의 개발은 2012년

- 88 -
경상과제를 통하여 지속적으로 과제를 수행 중에 있다. 또한 본 연구는 돼지를 이
용하여 당뇨질환 모델동물을 개발하는 것으로 국내에서 아직 그 연구가 미비한 실
정이다. 물론, 형질전환 생쥐를 이용한 질환모델의 개발은 많은 대학 및 연구소에서
연구되어지고 있지만 돼지는 거의 없다. 돼지가 생리학적측면에서 사람과 매우 유
사한 성질을 가진 특성 때문에 향후엔 전 임상시험에 많이 활용될 것이다. 이에 본
연구는 돼지를 이용한 질환모델 동물의 개발에 대한 다양한 기술을 습득하였고, 이
과정에서 발생된 시행착오로 관련 연구 분야에 다양한 기초정보의 제공이 가능할
것으로 사료된다.

- 89 -
세부(협동)과제명과 제
책임자
성과물
유 형성 과 물 명
성과물
주담당자
적용
년월
승인
여부
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅논문게제
(SCI)
The effects of various antioxidants on
the development of parthenogenetic
porcine embryos
여형수 '09.11 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수논문게제
(SCI)
Comparative studies of skeletal
muscle proteome and transcriptome
profilings between pig breeds
김남국 '10.6 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅논문게제
(SCI)
Overexpression of Jazf1 induces
cardiac malformation through the
upregulation of pro-apoptotic genes
in mice
배기범 '11.10 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅논문게제
(SCI)
Overexpression of Cathepsin S
Induces Chronic Atopic Dermatitis
In Mice
김나리 11,12 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
논문게제
(비SCI)
Production of a Transgenic Pig
Overexpressing Phosphoprotein
Enriched in Astrocytes 15 (PEA 15)
이휘철 ‘11.9 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철특허출원
생쥐 피이에이15 유전자가 과발현된
형질전환 돼지 및 그의 생산방법이휘철 ‘10.4 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수특허출원
영지버섯 추출물 또는 에고스테롤 퍼옥사
이드를 포함하는 또는 가나도마난디올을
포함하는 대사성 질환의 예방 또는 치료
용 성분 조성물
이창수 ‘10.11 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
생물자원
등록·기탁특허미생물 수탁 pCAGGS-mPea15 이휘철 ‘10.3 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
학술
발표
Confirmation of Target Gene Expression
in Transgenic Pig Expressing Mouse
Phosphoprotein Enriched in Astrocytes (mPea15)
and Enhanced Green Fluorescent Protein
이휘철 ‘09.6 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
학술
발표
Phosphoprotein Enriched in Astrocytes
(Pea15) 유전자 과발현 형질전환 돼
지의 당뇨질환 모델로써의 연구
이휘철 ‘09.6 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수학술
발표
Anti-diabetic Activity and the mechanisms
of action of Ganoderma lucidum extracts박혜란 ‘09.12 승인
제2절 정량적 성과

- 90 -
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수학술
발표
Anti-diabetic Activities of Ganoderma
lucidum extracts: Assoiated with
Activation of the AMPK
박혜란 ‘09.12 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수학술
발표
PED/PEA15 gene overexpression
effected to insulin resistance박혜란 ‘09.5 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수학술
발표
Comparative studies of skeletal muscle
proteome and transcriptome profilings
between pig breeds
김남국 ‘09.3 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Effects of hCG and eCG on In Vitro
Parthenogenetic Stimulation of Canine Oocytes김봉수 ‘09.04 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
The Different Rescue Levels in the Circling
Mouse with Transmembrane Inner Ear(tmie)
Gene Are Dependent on the Expression
Patterns of Transgene in the Cochlea
박서진 ‘09.06 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Parthenogenetkc Activation of Canine
Oocytes by Electrical Stimulation and
Chemical Stimulation
이준구 ‘09.06 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
The Effects of Various Antioxidants on
the Development of Parthenogenetic
Porcine Embryos
여형수 ‘09.08 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
RGS19inhibitcardiacmuscledifferentiationt
horoughattenuateWnt/β-cateninsignal 지영래 ‘09.08 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Overexpression of Jazf1 induces heart
failure symptoms through the
upregulation of proapototic genes in mice
배기범 ‘09.10 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
The Entire Structures of Tmie and
Rn49018 are Deleted in the Circling Mice김혜정 ‘10.02 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Transfer of EC-SOD Transgenic Mouse
Embryonic Fibroblasts has Potential
Therapeutic Effect in Inflammatory Arthritis
여형수 ‘10.02 승인
전사체 및 단백체 발현
분석을 통한 질환모델
가축의 검정
이창수학술
발표
Inhibitory effects of GPAT activity and
increase of glucose uptake by
Ganoderma lucidum extracts
손은숙 ‘10.11 승인

- 91 -
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Effect of Jazf1 Overexpression on
Mouse Cardiac Function배기범 ‘10.10 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Overexpression of Jazf1 induces
mouse heart failure배기범 ‘09.3 승인
당뇨병의 기능해석과
활용 연구를 위한 형질
전환 모델 생쥐 개발
류재웅학술
발표
Cathepsin S Induces Chronic Atopic
Dermatitis In Mice김나리 ‘11.9 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
학술
발표
Mouse Pea15 유전자 과발현 형질전환
돼지에 있어 Oral Glucose Tolerance
Test 및 당뇨병 관련 유전자의 발현양
상
김현미 ‘11.6 승인
당뇨질환 모델 돼지의
개발 및 생리학적 검정이휘철
학술
발표
High-fatdiet가 mouse Pea15 과발현 형
질전환돼지에있어 Oral Glucose
Tolerance Test에 미치는 영향
이휘철 ‘11.6 승인

- 92 -
제5장 연구개발결과의 활용계획
제1절 추가연구의 필요성새로운 신약이 개발되기까지는 엄청난 연구비와 시간이 투자되어야 한다. 특히 임상시험
단계에서의 긍정적인 결과를 도출하기 위해선 전 임상시험단계는 무엇보다 중요하다. 한편,
전 임상시험동물로 돼지는 설치류를 포함한 실험동물들에 비해 생리학, 병리생리학 및 이
들의 반응에 대해서 인간과 비슷한 점이 많기 때문에 최근 주목되고 있다. 이에 본 연구과
제는 당뇨병 치료제의 효능 검정을 위한 당뇨질환모델 돼지를 개발하는 것이다. 현재 국내
에서는 돼지를 이용한 질환모델동물의 개발은 거의 없는 실정이다. 특히, 형질전환 기술을
이용해서 당뇨질환모델 돼지 개발에 관한 연구는 본 연구과제가 거의 국내에선 처음 시도
되는 것으로 앞으로의 시행착오가 많다.
본 연구에서 기존에 알려진 유전자 PEA15를 과발현 시킨 형질전환 돼지를 생산하였고,
여러 측면에서 검정을 시도하였다. 그 결과 표현형에 있어서는 당뇨병에 대한 뚜렷한 증상
을 관찰하지 못했지만, 세포수준에서는 당 소모능의 저하현상을 관찰하였다. 이는 생체에서
도 잠재적으로 당뇨병 유발에 대한 가능성을 암시하는 것으로 추가적으로 향후 지속적인
연구가 요구된다. 게다가 본 과제에서 도출된 형질전환 질환모델 생쥐, 특히 당뇨병과 관련
된 Jazf1 형질전환 생쥐에서 당신생과 관련된 문제점을 확인하였으며, 본 유전자에 대한 돼
지로의 도입도 필요한 것으로 사료된다. 또한 중성지방 생합성과 관련된 유전자인 GPAT
유전자가 도입된 형질전환 돼지의 개발도 향후 필요할 것으로 생각된다. 이러한 유전자는
본 연구과제 수행동안 기초적인 내용은 확인이 되었고, 특히 Jazf1 유전자는 형질전환 생쥐
를 개발하여 검정하였다.

- 93 -
제2절 타 연구에의 응용 및 산업화 방안본 연구과제에서 도출된 결과에 대한 활용방안 및 기대성과는 아래와 같다.
□ 당뇨질환 모델돼지의 산업화를 위한 지적 소유권 확보
□ 개발된 질환모델 돼지을 통하여 당뇨병 치료제의 후보물질의 효능 검정에 활용
□ 의료 및 제약업계와의 공동연구로 산업화의 조기달성
□ 당뇨질환 재조합 세포배양계를 이용한 당뇨치료 생약 및 화합물도출을 통한 화합물의
가치를 새롭게 창출하는 시스템 확립
□ 질환모델생쥐의 질환관련 연구를 통해 신규 유전자의 발굴과 질환모델 돼지의 개발에
기초자료 제공
또한, PEA15 과발현 형질전환 돼지에서 분리된 근위성 세포는 일반돼지의 그것 보다 당
소모능이 저하되는 특징이 있다. 이는 In vitro에서 당뇨질환 모델로써의 활용이 가능하며,
당뇨병 치료제의 효능 검정 및 혈당강하 물질의 선별에 일차적으로 활용이 가능하다. 가령,
수많은 천연물 및 생약물질에서 혈당강하에 효능이 있는지를 일차적으로 선별하기 위한 도
구로 간단히 활용할 수 있을 것이다. 물론 다른 종류의 세포들도 있지만, 돼지에서 유래된
세포가 다른 실험동물 유래의 세포보다 더 효율적일 것으로 생각된다. 그리고 기업화 및
산업화를 추진하기 위해선 앞으로 우선적으로 당뇨질환 모델돼지에 대한 지적 소유권의 확
보와 이들의 유용성에 대한 다양한 결과들의 축적 및 확보를 위해서 추가적인 연구가 더
이루어져야 할 것이다.

- 94 -
제6장 중요 변동사항
제1절 과제책임자 변경
1. 변경 내용
주관과제명 세부과제명주관 및 세부과제 책임자
사유변경 전 변경 후
사람치료를 위한 신약 검정용
질환모델 가축 개발
당뇨질환 모델 돼지의 개발 및
생리학적 검정
국립축산과학원
기획조정과
이휘철
국립축산과학원
동물바이오공학과
양병철
인사이동
* 관련 공문 : 생명자원관리과-3867 (2011.10.25.)
제2절 연구내용 변경
1. 변경 내용
세부 및 협동연구과제 문제점/대책 및 주요 변경사항
당뇨질환 모델 돼지의 개발
및 생리학적 검정 (1세부)
○ 문제점 : PEA15 과발현 형질전환 돼지의 in vivo 검정(당부하검사)
에서 뚜렷한 당뇨병의 현상이 관찰되지 않았다.
-대책 : 현재 검토 중인 후보유전자(GPAT, Jazf1등)를 이용한
새로운 질환모델을 개발하고자 한다. 한편, 본 형질전
환 돼지에 있어서 뚜렷한 당뇨병의 현상은 관찰되지
않았지만, PEA15의 과발현은 확인되었고 당소모능을
분석한 세포수준에서는 당소모능이 저하되는 현상을
보였다. 그래서 당뇨병 기전의 연구에 활용 가능토록
세포수준에서 더 연구를 하고 in vitro 시스템을 구축
하는 방향으로 변경하고자 한다.
* 근거 : 2010년 연차실적보고서

- 95 -
제7장 참고문헌
1. Antti Virkama ̈ ki, Elena Korsheninnikova, Anneli Seppa ̈ la -̈Lindroos, Satu
Vehkavaara, Takashi Goto, Juha Halavaara, Anna-Maija Ha k̈kinen, and Hannele
Yki-Ja r̈vinen. 2001. Intramyocellu-lar Lipid Is Associated With Resistance to In
Vivo Insulin Actions on Glucose Uptake, Antili-polysis, and Early Insulin Signaling
Pathways in Human Skeletal Muscle. DIABETES. 50:2337-2343
2. Aas V, Rokling-Andersen M, Wensaas AJ, Thoresen GH, Kase ET and Rustan
AC. 2005. Lipid memtabolism in human skeletal muscle cells: effects of palmitate
and chronic hyperglycemia. Acta physiol Scand 183:31-41.
3. Bedell MA, Largaespada DA, Jenkins NA, Copeland NG. 1997. Mouse models of
human disease. Part II: recent progress and future directions. Genes Dev. 11:11-43.
4. Brüning R, Penzkofer H, Schöpf U, Becker C, Mayer T, Spuler A, Berchtenbreiter
C, Steiger HJ, Reiser M. 1998. Calculation of absolute cerebral blood volume and
cerebral blood flow by means of electron-beam computed tomography (EBT) in
acute ischemia. Radiologe. 38:1054-9.
5. Caruso M, Miele C, Formisano P, Condorelli G, Bifulco G, Oliva A, Auricchio R,
Riccardi G, Capaldo B, Beguinot F. 1997. In skeletal muscle, glucose storage and
oxidation are differentially impaired by the IR1152 mutant receptor. J Biol Chem.
14:272:7290-7.
6. Chiu TT, Patel N, Shaw AE, Bamburg JR, Klip A. 2010. Arp2/3- and
cofilin-coordinated actin dynamics is required for insulin-mediated GLUT4
translocation to the surface of muscle cells.. Mol Biol Cell. 15:21:3529-39.
7. Condorelli G, Vigliotta G, Iavarone C, Caruso M, Tocchetti CG, Andreozzi F, Cafieri
A, Tecce MF, Formisano P, Beguinot L, Beguinot F. 1998. PED/PEA-15 gene
controls glucose transport and is overexpressed in type 2 diabetes mellitus. EMBO
J. 17:3858-3866.
8. Condorelli G, Vigliotta G, Cafieri A, Trencia A, Andalò P, Oriente F, Miele C,
Caruso M, Formisano P, Beguinot F. 1999. PED/PEA-15: an anti-apoptotic molecule
that regulates FAS/TNFR1-induced apoptosis. Oncogene. 18:4409-4415.

- 96 -
9. Condorelli G, Vigliotta G, Trencia A, Maitan MA, Caruso M, Miele C, Oriente F,
Santopietro S, Formisano P, Beguinot F. 2001. Protein kinase C (PKC)-alpha
activation inhibits PKC-zeta and mediates the action of PED/PEA-15 on glucose
transport in the L6 skeletal muscle cells. Diabetes. 50:1244-1252.
10. Czech MP, Clancy BM, Pessino A, Woon CW, Harrison SA. 1992. Complex
regulation of simple sugar transport in insulin-responsive cells. Trends Biochem.
Sci. 17:197-201.
11. Deborah M. MUOIO, Kimberly SEEFELD, Lee A. WITTERS and Rosalind A.
COLEMAN. 1999. AMP-activated kinase reciprocally regulates triacylglycerol
synthesis and fatty acid oxidation in liver and muscle: evidence that
sn-glycerol-3-phosphate acyltransferase is a novel target. Biochem.J. 338:783-791.
12. Diez JJ, Iglesias P. 2003. The role of the novel adipocyte-derived hormone
adiponectin in human disease. Eur J Endocrinol. 148:293-300.
13. Dominiczak MH, Caslake MJ. 2011. Apolipoproteins: metabolic role and clinical
biochemistry applications, Ann Clin Biochem. 48:498-515.
14. Estelles A, Charlton CA, Blau HM. 1999. The phosphoprotein protein PEA-15
inhibits Fas- but increases TNF-R1-mediated caspase-8 activity and apoptosis. Dev
Biol. 216:16-28.
15. Farese, R. V. 2002. Function and dysfunction of aPKC isoforms for glucose
transport in insulin-sensitive and insulin-resistant states. Am. J. Physiol. Endocrinol.
Metab. 283:E1-E11.
16. Fiory F, Formisano P, Perruolo G, Beguinot F. 2009. Frontiers: PED/PEA-15, a
multifunctional protein controlling cell survival and glucose metabolism. Am J
Physiol Endocrinol Metab. 297:E592-601.
17. Formstecher E, Ramos JW, Fauquet M, Calderwood DA, Hsieh JC, Canton B,
Nguyen XT, Barnier JV, Camonis J, Ginsberg MH, Chneiweiss H. 2001. PEA-15
mediates cytoplasmic sequestration of ERK MAP kinase. Dev Cell. 1:239-250.
18. Gerolama C, Giovanni V, Carlo I, Matilde C, Carlo GT, Francesco A, Almerinda C,

- 97 -
Mario F.T, Pietro F, Laura B, Francesco B. 1998. PED/PEA-15 gene controls
glucose transport and is overexpressed in type 2 diabetes mellitus. EMBO j.
15;17:3858-66.
19. Gianfranco BRAMBILLA and Alfredo CANTAFORA. 2004. Metabolic and
cardiovascular disorders in highly inbred lines for intensive pig farming: how animal
welfare evaluation could improve the basic knowledge of human obesity. Ann Ist
Super Sanità 40:241-244.
20. Hruz, P.W., and Mueckler, M.M. 2001. Structural analysis of the GLUT1
facilitative glucose transporter (review). Mol. Membr. Biol. 18, 183-193.
21. Hu E, Liang P, Spiegelman BM. 1996. AdipoQ is a novel adipose-specific gene
dysregulated in obesity. J Biol Chem. 3:271:10697-10703.
22. Huang S, Czech MP. 2007. The GLUT4 glucose transporter. Cell Metab. 5:237-52.
23. Ishikura, S., Bilan, P. J., and Klip, A. 2007. Rabs 8A and 14 are targets of the
insulin-regulated Rab-GAP AS160 regulating GLUT4 traffic in muscle cells.
Biochem. Biophys. Res. Commun. 353:1074-1079.
24. JeBailey L, Wanono O, Niu W, Roessler J, Rudich A, Klip A. 2007. Ceramide- and
oxidant-induced insulin resistance involve loss of insulindependent Rac-activation
and actin remodeling in muscle cells. Diabetes. 56:394-403.
25. Joost HG, Thorens B. 2001. The extended GLUT-family of sugar/polyol transport
facilitators: nomenclature, sequence characteristics, and potential function of its novel
members (review). Mol. Membr. Biol. 18:247-256.
26. Junod A, Lambert AE, Stauffacher W, Renold AE. 1969. Diabetogenic action of
streptozotocin: relationship of dose to metabolic response. J Clin Invest.
48:2129-2139.
27. Kanoh Y, Bandyopadhyay G, Sajan MP, Standaert ML, Farese RV. 2000.
Thiazolidinedione treatment enhances insulin effects on protein kinase C-zeta
/lambda activation and glucose transport in adipocytes of nondiabetic and
Goto-Kakizaki type II diabetic rats. J. Biol. Chem. 275:16690-16696.

- 98 -
28. Kanoh Y, Bandyopadhyay G, Sajan MP, Standaert ML, Farese RV. 2001.
Rosiglitazone, insulin treatment, and fasting correct defective activation of protein
kinase C-zeta/lambda by insulin in vastus lateralis muscles and adipocytes of
diabetic rats. Endocrinology 142:1595-1605.
29. Khayat ZA, Tong P, Yaworsky K, Bloch RJ, Klip A. 2000. Insulin-induced actin
filament remodeling colocalizes actin with phosphatidylinositol 3-kinase and GLUT4
in L6 myotubes. J. Cell Sci. 113:279-290.
30. Kitsberg D, Formstecher E, Fauquet M, Kubes M, Cordier J, Canton B, Pan G,
Rolli M, Glowinski J, 1999. Knock-out of the neural death effector domain protein
PEA-15 demonstrates that its expression protects astrocytes from
TNFalpha-induced apoptosis. J Neurosci. 19:8244-8251.
31. Kopelman, P. G, 2000. Obesity as a medical problem. Nature 404:635-643.
32. Kubes M, Cordier J, Glowinski J, Girault JA, Chneiweiss H. 1998. Endothelin
induces a calcium-dependent phosphorylation of PEA-15 in intact astrocytes:
identification of Ser104 and Ser116 phosphorylated, respectively, by protein kinase C
and calcium/calmodulin kinase II in vitro. J Neurochem. 71:1307-14.
33. Kulkarni RN, Winnay JN, Daniels M, Brüning JC, Flier SN, Hanahan D, Kahn CR.
1999. Altered function of insulin receptor substrate-1-deficient mouse islets and
cultured beta-cell lines, J Clin Invest. 104:R69-75.
34. Lee HC, Kim HM, Lee S, Oh KB, Chung HJ, Yang BC, Kim KW, Lee P, Park
JK, Chang WK. 2011. Production of a Transgenic Pig Overexpressing
Phosphoprotein Enriched in Astrocytes 15 (PEA 15). Reprod Dev Biol. 35:239-245.
35. Lenzen S, Panten U. 1988. Alloxan: history and mechanism of action. Diabetologia.
31:337-42.
36. Lihn AS, Pedersen SB, Richelsen B. 2005. Adiponectin: action, regulation and
association to insulin sensitivity. Obes Rev. 6:13-21.
37. Lindsay RS, Funahashi T, Hanson RL, Matsuzawa Y, Tanaka S, Tataranni PA.
2002. Adiponectin and development of type 2 diabetes in the Pima Indian population.
Lancet 360:57-58.

- 99 -
38. Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using
real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods
25:402-408.
39. Maddux BA, Sbraccia P, Kumakura S, Sasson S, Youngren J, Fisher A, Spencer
S, Grupe A, Henzel W, Stewart TA, et al. 1995. Membrane glycoprotein PC-1 and
insulin resistance in non-insulin-dependent diabetes mellitus. Nature. 1995
373:448-451.
40. Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K.
1996. cDNA cloning and expression of a novel adipose specific collagen-like factor,
apM1 AdiPose Most abundant Gene transcript Biochem Biophys Res Commun.
221:286-9.
41. Marshall BA, Ren JM, Johnson DW, Gibbs EM, Lillquist JS, Soeller WC, Holloszy
JO, Mueckler M. 1993. Germline manipulation of glucose homeostasis via alteration
of glucose transporter levels in skeletal muscle.. J. Biol. Chem. 268:18442-18445.
42. Matsuda M, Shimomura I, Sata M, Arita Y, Nishida M, Maeda N. 2002. Role of
adiponectin in preventing vascular stenosis. The missing link of adipo-vascular axis.
J Biol Chem 277:37487-91.
43. Naruse K, Ishikawa H, Kawano HO, Ueda H, Kurome M, Miyazaki K, Endo M,
Sawasaki T, Nagashima H, Makuuchi M. 2005. Production of a transgenic pig
expressing human albumin and enhanced green fluorescent protein. J Reprod Dev
51:539-46.
44. Ouchi N, Kihara S, Arita Y, Maeda K, Kuriyama H, Okamoto Y. 1999. Novel
modulator for endothelial adhesion molecules: adipocyte-derived plasma protein
adiponectin. Circulation. 100:2473-6.
45. Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y. 2001.
Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and
class A scavenger receptor expression inhuman monocyte-derived macrophages.
Circulation. 103:1057-63.
46. Paul Dowell, Zhiyuan Hu, and M. Daniel Lane. 2005. Monitoring energy balance:

- 100 -
metabolites of fatty acid synthesis as hypothalamic sensors. Annu.Rev.Biochem.74:515-534
47. Rees DA, Alcolado JC. 2005. Animal models of diabetes mellitus. Diabet Med.
22:359-70.
48. Rossetti L. Stenbit AE, Chen W, Hu M. Barzilai, N., Katz, E.B., and Charron,
M.J. 1997. Peripheral but not hepatic insulin resistance in inuclear storage
compartment involves distinct carboxyl-tail motifs. J. Cell Sci. 113:1705- 1715.
49. Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF.A. 1995. novel serum
protein similar to C1q, produced exclusively in adipocytes. J Biol Chem.
270:26746-26749.
50. Shi, Y and Paul, B. 2004. Lipid metabolic enzymes: emergin drug targets for the
treatment of obesity. Nature Reviews Drug Discovery3:695-705.
51. Steppan CM, Bailey ST, Bhat S, Brown EJ, Banerjee RR, Wright CM, Patel HR,
Ahima RS, Lazar MA. 2001. The hormone resistin links obesity to diabetes. Nature.
409:307-12.
52. Stefan N, Bunt JC, Salbe AD, Funahashi T, Matsuzawa Y, Tataranni PA. 2002.
Plasma adiponectin concentrations in children: relationships with obesity and
insulinemia. J Clin Endocrinol Metab. 87:4652– 4656.
53. Thong FS, Bilan PJ, Klip A. 2007. The Rab GTPase-activating protein AS160
integrates Akt, protein kinase C, and AMP-activated protein kinase signals
regulating GLUT4 traffic. Diabetes. 56:414-423.
54. Trencia A, Perfetti A, Cassese A, Vigliotta G, Miele C, Oriente F, Santopietro S,
Giacco F, Condorelli G, Formisano P, Beguinot F. 2003. Protein kinase B/Akt binds
and phosphorylates PED/PEA-15, stabilizing its antiapoptotic action. Mol Cell Biol
23:4511-4521.
55. Trencia A, Fiory F, Maitan MA, Vito P, Barbagallo AP, Perfetti A, Miele C,
Ungaro P, Oriente F, Cilenti L, Zervos AS, Formisano P, Beguinot F. 2004.
Omi/HtrA2 promotes cell death by binding and degrading the anti-apoptotic protein
ped/pea-15. J Biol Chem. 279:46566-46572.

- 101 -
56. Turk JR, Henderson KK, Vanvickle GD, Watkins J, Laughlin MH. 2005. Arterial
endothelial function in a porcine model of early stage atherosclerotic vascular
disease. Int J Exp Pathol. 86:335-345.
57. Ueyama A, Yaworsky KL, Wang Q, Ebina Y, Klip A. 1999. GLUT-4myc ectopic
expression in L6 myoblasts generates a GLUT-4-specific pool conferring insulin
sensitivity. Am J Physiol. 277:E572-E578.
58. Vaidyanathan H, Ramos JW. 2003. RSK2 activity is regulated by its interaction
with PEA-15. J Biol Chem. 278:32367-32372.
59. Vigliotta G, Miele C, Santopietro S, Portella G, Perfetti A, Maitan MA, Cassese A,
Oriente F, Trencia A, Fiory F, Romano C, Tiveron C, Tatangelo L, Troncone G,
Formisano P, Beguinot F. 2004. Overexpression of the PED/PEA-15 gene causes
diabetes by impairing glucosestimulated insulin secretion in addition to insulin
action. Mol Cell Biol 24:5005-5015.
60. Wang Q, Somwar R, Bilan PJ, Liu Z, Jin J, Woodgett JR, Klip A. 1999. Protein
kinase B/Akt participates in GLUT4 translocation by insulin in L6 myoblasts. Mol
Cell Biol. 19:4008-4018.
61. Weyer C, Funahashi T, Tanaka S, Hotta K, Matsuzawa Y, Pratley RE. 2001.
Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin
resistance and hyperinsulinemia. J Clin Endocrinol Metab. 86:1930-5.
62. Wild S, Roglic G, Green A, Sicree R, King H. 2004. Global prevalence of diabetes.
Diabetes Care. 27:1047-1053.
63. Wood, I.S., and Trayhurn, P. 2003. Glucose transporters (GLUT and SGLT):
expanded families of sugar transport proteins. Br. J. Nutr. 89:3-9.
64. Xiaobo HU, Meihua SHE, Hongjie HOU, Qinkai LI, Qingyun SHEN, Yi LUO, and
Weidong YIN. 2007. Adiponectin Decreases Plasma Glucose and Improves Insulin
Sensitivity in Diabetic Swine. Acta Biochimica et Biophysica Sinica. 39: 131-136
65. Yea K, Kim J, Yoon JH, Kwon T, Kim JH, Lee BD, Lee HJ, Lee SJ, Kim JI, Lee
TG, Baek M C, Park HS, Park KS, Ohba M, Suh PG, Ryu SH. 2009.
Lysophosphatidylcholine Activates Adipocyte Glucose Uptake and Lowers Blood

- 102 -
Glucose Levels in Murine Models of Diabetes. J Biol Chem. 284:33833-40.
66. Zhang Y, Redina O, Altshuller YM, Yamazaki M, Ramos J, Chneiweiss H, Kanaho
Y, Frohman MA. 2000. Regulation of expression of phospholipase D1 and D2 by
PEA-15, a novel protein that interacts with them. J Biol Chem 275:35224-35232.
67. Zisman A, Peroni OD, Abel ED, Michael MD, Mauvais-Jarvis F, Lowell BB,
Wojtaszewski JF, Hirshman MF, Virkamaki A, Goodyear LJ, Kahn CR, Kahn BB.
2000. Targeted disruption of the glucose transporter 4 selectively in muscle causes
insulin resistance and glucose intolerance. Nat Med. 6:924-928.

- 103 -
<참고 1> 연차평가 지적사항 및 조치결과
* 연차평가 시 지적사항과 그에 대한 조치결과
지 적 사 항 수 정 보 완 사 항
○ ○
○ ○
○ ○

- 104 -
<참고 2> 주요연구 성과
당뇨병 질환 모델 세포 및 생쥐 개발
□ 과제명 : 사람치료를 위한 신약검정용 질환모델 가축 개발
□ 연구 배경
○ 당뇨질환 인구의 증가와 새로운 치료제의 효능 검증을 위한 전임상 모델 필요
- 인간과 생리학적으로 가까운 전임상 모델로써 돼지의 장점이 부각
□ 주요 연구성과
○ PEA15 과발현 형질전환 돼지 개발
- PEA15 과발현 단백질 확인 및 형광유전자 발현확인
- PEA15 과발현 형질전환 돼지 특허 출원 (‘10년)
○ 돼지 근위성 세포분리 및 배양
- 형질전환 돼지의 근육세포에서 당 소모능 저하 확인
- 당뇨질환 모델 세포로써 활용가능성 확인
○ Jazf1 과발현 형질전환 마우스 개발
- 당신생 저하 및 아토피증상 유발 질환모델 (Transgenic Research ' 11년)
마우스 Pea15 과발현 및 형광유전자(GFP) 발현 형질전환 돼지
형질전환 돼지 근육세포에서의 당 소모능 저하 결과
Jazf1 과발현 형질전환 마우스의 혈당 및 아토피 증상
□ 파급효과
○ 과학기술적 성과
- 국내에서 돼지에 형질전환기술을 적용한 당뇨질환 모델 돼지의 개발은 거의 없
는 실정으로 향후 관련 연구 분야에 많은 기초자료 제공
○ 경제사회적 성과
- 당뇨병 치료제 효능 검증에 시간, 연구비 등의 절감으로 신약 원가 절감 효과 기대

- 105 -
주요 결과 요약서
□ 과제명 : 사람 치료를 위한 신약 검정용 질환모델 가축 개발
▶ 연구목적 : 당뇨병 치료제 효능 검정 및 당뇨병 발병기작 원인 규명에 활용하기
위한 당뇨질환모델 동물의 개발과 이들의 생리학적 분석, 유전자
재조합 세포 배양계 분석, 전사체 및 단백체의 분석을 통한 이들의 특성을
규명한다.
▶ 주요연구내용 :
○ 1세부과제명 : 당뇨질환 모델 돼지의 개발 및 생리학적 검정
- 기 생산된 Pea15 과발현 형질전환 돼지의 당뇨질환 모델로써의 활용 가능성
에 대한 시험분석을 실시한 결과, In vivo 당 부하 검사에서는 뚜렷한 당뇨질
환을 관찰하지 못했지만, 이들 근육세포를 이용한 In vitro에서 당 소모능의
분석은 현저히 감소됨을 확인하였다. 이는 질환모델의 도구로써의 활용가능성
을 제시하였다. In vitro에서의 당 소모능 저하에 대한 원인을 규명하고자
Glut4 당수송체의 막이동에 대한 분석을 조사한 결과, Pea15 과발현에 의해
저해됨을 확인하였다.
○ 2세부과제명 : 전사체 및 단백체 발현분석을 통한 질환모델 가축의 검정
- Pea15 과발현 형질전환 돼지의 대사시스템 변화 분석 결과, 인슐린 무처리
세포주에서는 검출되지 않았던 pAkt는 인슐린 처리로 재조합유전자의 과발
현과는 관계없이 그 발현량이 매우 높게 증가하였고 이는 Pea15 유전자의
과발현으로 인하여 세포내 당수송 능력의 감소는 인슐린 비의존적인 경로를
경유하여 작용되었을 것으로 생각된다. 인간유래 간세포주에 Pea15 유전자를
도입한 후 24시간 48시간 후에 간에서의 중성지방 합성 정도를 확인한 결과,
24시간 후 Pea15 과발현 세포주에서의 중성지방 합성율이 40% 증가 하는
것을 확인하였다. 또한, 형질전환 돼지의 혈장 단백체에서 3개의 차등발현을
보이는 단백질을 확인하였고 현재 근육조직에서의 대사변화를 규명하기 위
하여 DNA chip 이용한 근육조직전체의 유전자 발현양상을 분석 중에 있다.

- 106 -
○ 3세부과제명 : 당뇨병의 기능해석과 활용 연구를 위한 형질전환 모델 생쥐 개발
- Jazf1 형질전환 생쥐의 개발을 통하여 이들의 혈압이 정상에 비하여 높게 유
지되었으며, 심초음파 실험 결과 심장박동이 불규칙함을 확인하였다. 심장조직
의 모습 또한 정상 마우스에 비하여 조직이 치밀하지 못하였으며, 심근세포의
미토콘드리아의 형태가 많이 망가져 있음을 확인하였다. 또한, CTSS 과발현
형질전환 생쥐에서는 지방분화에 관여하는 것으로 비만유도에 의한 합병증이
발병할 것으로 예상하였고 그 결과, 아토피가 발병되었다. 개발된 2종류의 형질
전환 생쥐의 돼지로의 개발에 검토 중에 있다.

- 107 -
주 의
1. 이 보고서는 농촌진흥청에서 시행한 「차세대바이오그린21사업
」의 연구보고서입니다.
2. 이 보고서의 내용을 인용 • 발표할 때는 반드시 농촌진흥청에서
시행한 「차세대바이오그린21사업 」의 결과임을 밝혀야 합니
다.
3. 국가과학기술 기밀유지에 필요한 내용은 대외적으로 발표 또는
공개하여서는 아니 됩니다.