Embed Size (px)
Citation preview

1
Centro Universitario de Matanzas
Estación Experimental de Pastos y Forrajes "Indio Hatuey"
Instituto de Ciencia Animal
Estudio regional y clasificación de las leguminosas forrajeras autóctonas y/o naturalizadas en Cuba
Autor: Ing. Juvenal Menéndez Villaurrutia Tutor: Dr. Gustavo Febles Pérez C. Dr. Sc.
Tesis presentada en opción al grado de Candidato a Doctor en Ciencias
1982

i
La producción agropecuaria se desarrollará, principalmente, por el método intensivo- incrementando la productividad del trabajo, el rendimiento- de los cultivos y del ganado y disminuyendo los costos y aprovechando las posibilidades que aún ofrece el método extensivo, incorporando más superficie cultivable al proceso productivo.
Plataforma Programática del PCC
Nuestra producción láctea tendríamos que buscarla fundamentalmente en los pastos de gramíneas y leguminosas en la medida
en que podamos ir introduciendo estas últimas.
Fidel

Dedicatoria
ii
Dedico esta tesis al XXIV Aniversario de la Revolución Cubana y a su Partido
Comunista, que posibilitaron la instauración del primer estado socialista en el
hemisferio occidental, que con su ejemplo y posición ha hecho posible que todos los
pueblos del Tercer Mundo sean desde ese momento más libres y no vean muy lejano
el día en que también serán dueños de sus riquezas, de su destino, sean dueños de sí.

Agradecimientos
iii
AGRADECIMIENTOS
Al Estado Socialista de Cuba que con su política científica ha hecho posible la materialización de este
trabajo. A la Estación Experimental de Pastos y Forrajes "Indio Hatuey", por el apoyo brindado para la
ejecución de las investigaciones. A mi tutor C.Dr.C. Gustavo Febles por sus magníficas sugerencias y
dedicación en la elaboración de la tesis.
A todos los compañeros técnicos e investigadores de la Estación y muy especialmente a los del
Departamento de Introducción y Mejoramiento que de una forma u otra colaboraron activamente en la
materialización de este trabajo, particularmente al Ing. Rey Machado, Juan F. Martínez e Iván Yepes, así
como a los choferes que nos acompañaron en las excursiones a los trabajadores agropecuarios de las
diferentes provincias quienes también brindaron su aporte.
A Blanca R. Montano y Alida Trujillo por la mecanografía, a Andrés Suárez, Rafael Campbell y el grupo de
estudiantes de quinto año de la especialidad de Pastos y Forrajes por la confección y tirada de las tablas y
gráficos, así como al Departamentos de Fotografía del ICA y del CENIT por las vistas tiradas.
A mi esposa Acela Tamayo y a mi hijita Janet por el tiempo que no dediqué a ellas entregado a este trabajo.

Índice
iv
ÍNDICE
INTRODUCCIÓN......................................................................................................................................................1
CAPÍTULO I. REVISIÓN DE LA LITERATURA .......................................................................................................2 Principales centro de origen de las leguminosas.................................................................................................2 Importancia de la prospección .............................................................................................................................2 Necesidad del estudio de las leguminosas..........................................................................................................3 Principales características de las leguminosas más importantes en Cuba .........................................................3 Factores que intervienen en la explotación de las leguminosas..........................................................................5
CAPÍTULO II. METODOLOGÍA EXPERIMENTAL...................................................................................................7 Ubicación y extensión del archipiélago cubano ...................................................................................................7 Principales regiones físico-geográficas o naturales de Cuba..............................................................................7
Región Occidental............................................................................................................................................7 Región Central .................................................................................................................................................7 Región Camagüey-Maniabón ..........................................................................................................................7 Región Oriental ................................................................................................................................................7 Región de Isla de Pinos ...................................................................................................................................8
Ecología................................................................................................................................................................8 Sistema de muestreo ...........................................................................................................................................8 Clasificación de la flora empleada en las regiones físico-geográfica ................................................................10
CAPÍTULO III. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN DE OCCIDENTE Y DE ISLA DE PINOS............................................................................................................................................12
Materiales y métodos .........................................................................................................................................12 Resultados .........................................................................................................................................................12
Generalidades................................................................................................................................................12 Descripción y distribución de los géneros principales ...................................................................................15
Discusión............................................................................................................................................................18
CAPÍTULO IV. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN CENTRAL Y PARTE DE LA DE CAMAGÜEY-MANIABÓN DE CUBA.............................................................................................21
Materiales y métodos .........................................................................................................................................21 Resultados .........................................................................................................................................................21
Generalidades................................................................................................................................................21 Descripción de los géneros principales .........................................................................................................24
Discusión............................................................................................................................................................28
CAPÍTULO V. LEGUMINOSAS FORRAJERAS SILVESTRES DEL NORESTE DE LA REGIÓN CAMAGÜEY-MANIABÓN Y SUROESTE DE LA ORIENTAL..............................................................................30
Materiales y métodos .........................................................................................................................................30 Resultados .........................................................................................................................................................30
Generalidades................................................................................................................................................30 Descripción de los géneros de mayor importancia........................................................................................31 Géneros de menos importancia.....................................................................................................................34
Discusión ............................................................................................................................................................36
CAPÍTULO VI. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN ORIENTAL DE CUBA .............38 Materiales y métodos .........................................................................................................................................38 Resultados .........................................................................................................................................................38
Generalidades................................................................................................................................................38 Descripción de géneros principales...............................................................................................................38
Discusión ............................................................................................................................................................43
CONCLUSIONES...................................................................................................................................................44 RECOMENDACIONES...........................................................................................................................................46 REFERENCIAS BIBLIOGRÁFICAS.......................................................................................................................47

Índice de tablas y figuras
v
ÍNDICE DE TABLAS
Tabla 1. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en la región de Occidente e Isla de Pinos. ........................................................................................................................13
Tabla 2. Especies menos importantes, frecuencia y suelo donde se encuentran en la región de Occidente e Isla de Pinos. ...................................................................................................................................................14
Tabla 3. Vegetación que acompaña a los géneros principales en la región de Occidente e Isla de Pinos...........14
Tabla 4. Vegetación que acompaña a los géneros menos importantes en la región de Occidente e Isla de Pinos...................................................................................................................................................................15
Tabla 5. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en la región Central y Camagüey-Maniabón................................................................................................................................22
Tabla 6. Especies menos importantes, frecuencia y suelo donde se encuentran en la región Central y Camagüey-Maniabón................................................................................................................................23
Tabla 7. Vegetación asociada a los géneros más importantes en la región Central y Camagüey-Maniabón.......24
Tabla 8. Vegetación asociada a los géneros menos importantes en la región Central y Camagüey-Maniabón...28
Tabla 9. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en noreste de la región Camagüey-Maniabón y suroeste de la Oriental. ......................................................................................34
Tabla 10. Vegetación asociada a los géneros más importantes en la Camagüey-Maniabón y parte de la región Oriental. ..................................................................................................................................................34
Tabla 11. Especies menos importantes, frecuencia y suelo de Camagüey-Maniabón y parte de la región Oriental. ..................................................................................................................................................35
Tabla 12. Vegetación asociada a los géneros menos importantes en Camagüey-Maniabón y parte de la región Oriental. ..................................................................................................................................................36
Tabla 13. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en el este de la región Oriental. ..................................................................................................................................................41
Tabla 14. Vegetación asociada a los géneros más importantes en el este de la región Oriental. ........................41
Tabla 15. Especies menos importantes, frecuencia y suelo en el este de la región Oriental................................42
Tabla 16. Vegetación asociada a los géneros menos importantes en el este de la región Oriental. ....................42 ÍNDICE DE FIGURAS
Fig. 1. Principales regiones físico-geográficas.........................................................................................................8
Fig. 2. Precipitaciones en el período lluvioso ..........................................................................................................9
Fig. 3. Precipitaciones en el período seco................................................................................................................9
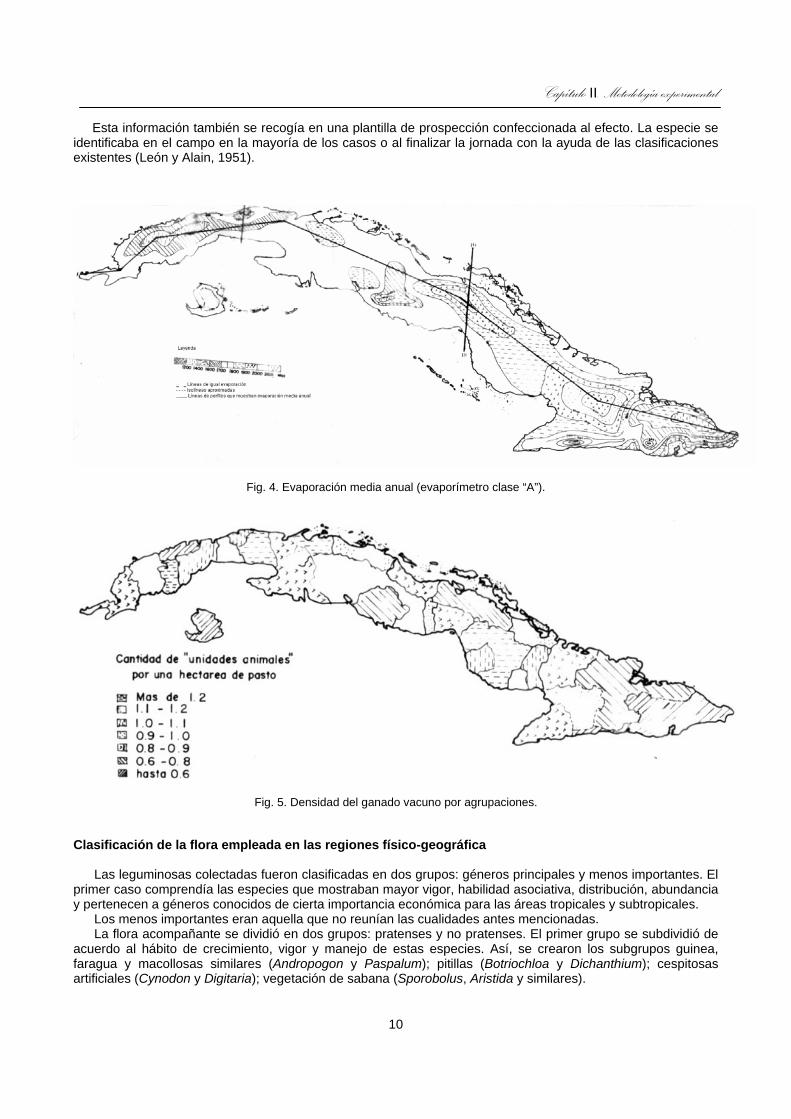
Fig. 4. Evaporación media anual (evaporímetro clase “A”). ...................................................................................10
Fig. 5. Densidad del ganado vacuno por agrupaciones. ........................................................................................10
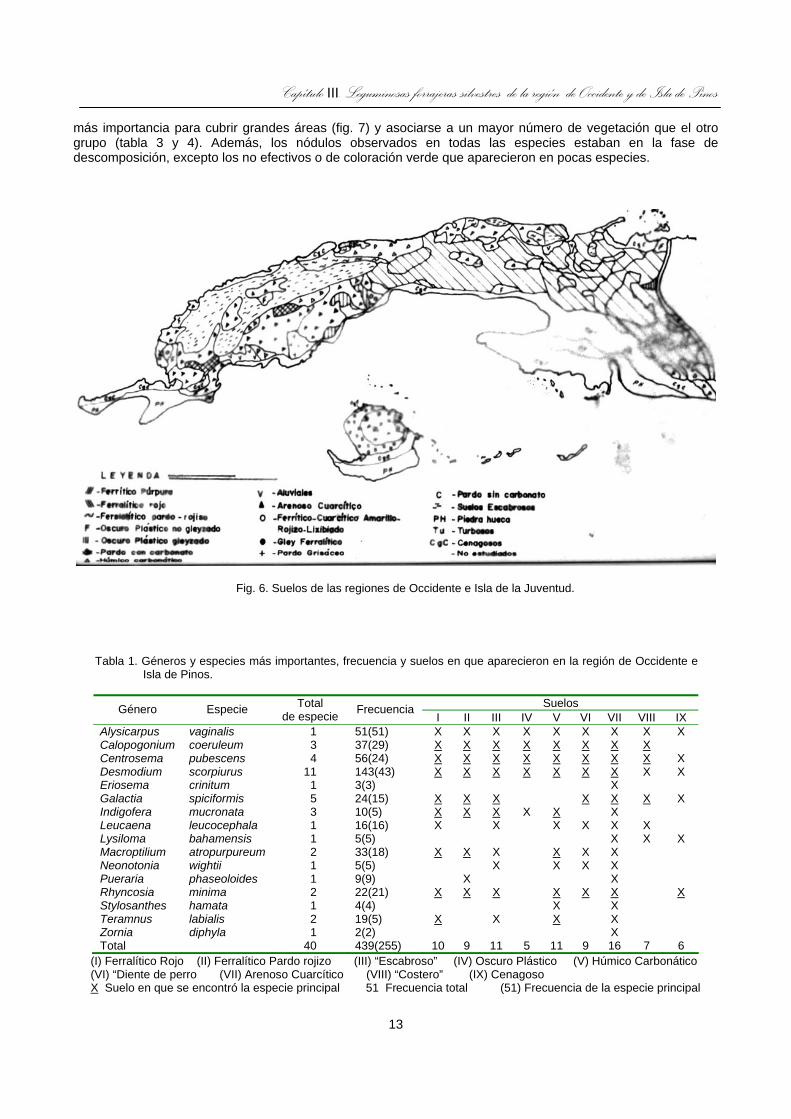
Fig. 6. Suelos de las regiones de Occidente e Isla de la Juventud........................................................................13
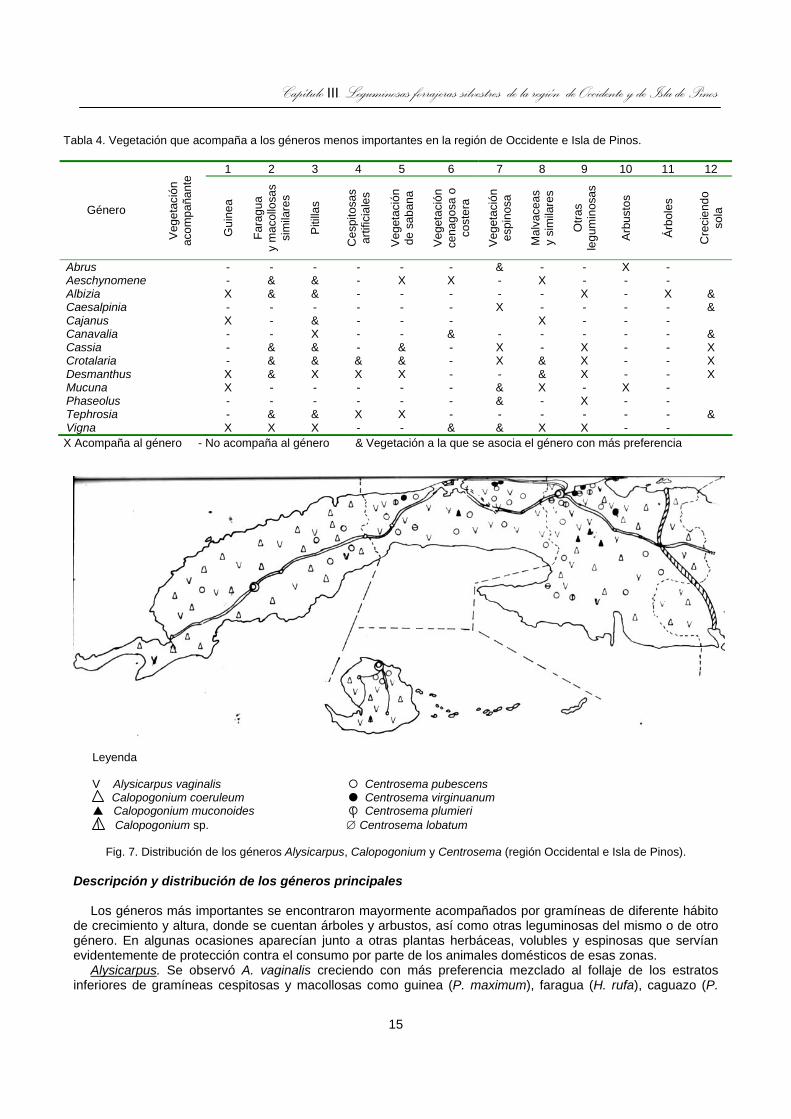
Fig. 7. Distribución de los géneros Alysicarpus, Calopogonium y Centrosema (región Occidental e Isla de Pinos). ....................................................................................................................................15
Fig. 8. Distribución de los géneros Desmodium, Eriosema, Galactia, Indigofera, Leucaena y Lysiloma (Occidente e Isla de Pinos).............................................................................................................................................17
Fig. 9. Distribución de los géneros Macroptilium, Neonotonia, Pueraria, Rhyncosia, Stylosanthes, Teramnus y Zornia (Occidente e Isla de Pinos). ..............................................................................................................19
Fig. 10. Suelos de la región Central .......................................................................................................................22

Índice de tablas y figuras
vi
Fig. 11. Suelos de la región Camagüey-Maniabón. ...............................................................................................23
Fig. 12. Distribución de los géneros Alysicarpus, Calopogonium y Centrosema (región Camagüey-Maniabón)..25
Fig. 13. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Lysiloma (región Camagüey-Maniabón). ................................................................................................................................................26
Fig. 14. Distribución de los géneros Macroptilium, Rhynchosia, Teramnus y Zornia (región Camagüey-Maniabón). ................................................................................................................................................27
Fig. 15. Suelos de la región Oriental. .....................................................................................................................30
Fig. 16. Distribución de los géneros Alysicarpus, Calopogonium, Centrosema, Clitoria y Desmanthus (región Oriental). ....................................................................................................................................................32
Fig. 17. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Macroptilium (región Oriental). ....................................................................................................................................................33
Fig. 18. Distribución de los géneros Rhynchosia, Stylosanthes y Teramnus (región Oriental). ............................35
Fig. 19. Distribución de los géneros Alysicarpus, Calopogonium, Centrosema y Desmanthus (este Oriental). ...39
Fig. 20. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Macroptilium (este Oriental)......................................................................................................................................................40
Fig. 21. Distribución de los géneros Pueraria, Rhynchosia, Stylosanthes, Tephrosia y Teramnus (este Oriental)......................................................................................................................................................42

Introducción
1
INTRODUCCIÓN
Cuba heredó del capitalismo una estructura económica deformada, de base agropecuaria atrasada, donde la
baja calidad y producción de los pastizales naturales y artificiales (cuando estos no disponen de las cantidades
suficiente de fertilizantes) provoca una baja productividad de la masa ganadera y de las tierras dedicadas a su
explotación.
Es por eso que en los principios programáticos del Primer Congreso del Partido Comunista de Cuba se
expresa que entre sus objetivos: “El desarrollo de la ganadería exigirá el mejoramiento de la base técnica y del
manejo de la masa ganadera, el incremento y la atención constante de la correspondiente y decisiva base
alimentaria...”.
Así la introducción y la búsqueda de nuevas especies es una de las vías para mejorar la alimentación de la
masa ganadera de nuestro país.
En la actualidad Cuba cuenta con valiosas introducciones de gramíneas forrajeras que superan en calidad y
producción a las que se explotaban comercialmente aunque estos nuevos cultivares requieren, en su mayoría,
de una aplicación eficiente de fertilización nitrogenada, el riego y un adecuado manejo, que sólo puede ser
practicado en reducidas áreas del país, debido a que éste no cuenta con suficientes recursos para extender
masivamente estos sistemas.
Sin embargo, el uso adecuado de las leguminosas pudiera ser una de las soluciones para esta problemática,
ya que las evidencias experimentales han puesto de manifiesto la posibilidad de su empleo en el trópico y
particularmente en Cuba, donde se han obtenido resultados alentadores como los de Funes, Yepes y
Hernández (1971); Salinas y Milera (1972); Echevarría y Rodríguez (1977); Monzote y Hernández (1977);
Rodríguez (1977); Ruiz y Ayala (1977); Menéndez y Martínez (1980) y Menéndez, Tang, Aleaga y Polunin
(1980), entre otros.
Además, el origen y distribución actual de las leguminosas es tropical (Bogdan, 1966; Williams, 1967).
Mehra y Magoon (1974) especifican que México y los países de América Central son centros ricos en
diversificación de especies de leguminosas tropicales y subtropicales de importancia económica y Yepes
(1971a) planteó que Cuba es el país más rico en leguminosas forrajeras en el área del Caribe.
Las razones anteriores justifican la prospección mundial de leguminosas y particularmente en Cuba, que es
un país en vías de desarrollo, donde la puesta en explotación de áreas vírgenes amenaza con provocar la
erosión genética de tan rico germoplasma de leguminosas autóctonas forrajeras.
Estos argumentos fundamentan nuestros objetivos de trabajo, los cuales en esencia apoyan los principios
de la Plataforma Programática, cuando plantean que “...los trabajos se dedicarán a la investigación de los
recursos naturales del país, su adecuada protección y sus posibilidades de aprovechamiento en la economía y
otras esferas de la vida nacional”.
Estos objetivos de trabajo son:
1. Identificar las leguminosas con características forrajeras autóctonas y/o naturalizadas existentes en la flora de Cuba.
2. Establecer la distribución, diversificación y la regionalización de las leguminosas forrajeras cubana. 3. Brindar una información básica con el fin de facilitar los trabajos futuros de explotación de este grupo de
plantas. 4. Evitar la posible erosión genética e incrementar el germoplasma de este taxón en el país.

Capítulo I. Revisión bibliográfica
2
CAPÍTULO I. REVISIÓN DE LA LITERATURA Principales centro de origen de las leguminosas
Según Hartley (1950) y Bogdan (1966) de la región de América Central y América son importantes para la búsqueda de las leguminosas. Vavilov (1950) basado en los resultados de las expediciones a través del mundo pudo descubrir una rica zona de material vegetal y comprobó que las variaciones dentro de cada planta era mucho más rica de lo que hasta entonces se imaginaba. Esto permitió a N.I. Vavilov llegar a la conclusión de determinar 8 antiguos cuatros principales de los cuales nació la agricultura mundial.
Los géneros principales de leguminosas se distribuyen en los centros de origen de la forma siguiente: Centro Chino. Lo describió como el más antiguo y más vasto de los centros independientes de la agricultura
y de origen de las plantas cultivadas, está formado por las regiones montañosas del centro y occidente de China, junto con las tierras bajas antiguas. El Centro Chino se distingue de todos los demás por su riqueza de especies endémicas y por la magnitud del potencial de las especies cultivadas. Las especies principales de leguminosas son Glycine hispida, Phaseolus angularis, Ph. vulgaris, Vigna sineusis y Stizolobium hassjoo.
Centro Indio. Este centro de origen sigue al chino en importancia y comprende a Birmania y Assam, pero no la parte noreste de la India ni Pakistán occidental, donde se incluyeron las especies Cicer arietinum, Cajanus cajan, Phaseolus acontifolius, Ph. mungo, Ph. aureus, Ph. calcaratus, Dolichos biflorus, D. niger, Vigna sinensis, Triconella foenum, Canavalia gladiata, Psophocorpus tetragonolobus y Cyamopsis psoraloides.
Centro Indomalayo. Este centro, auxiliar del anterior abarca todo el archipiélago malayo, las islas de Java, Borneo, Sumatra, Filipinas e Indochina, de donde es originario Stizolobium deeringianum.
Centro de Asia Central. Es el centro de menor superficie y comprende el noroeste de la India, Cachemira, Pakistán; Afganistán y en la URSS, Tadjikistán, Uzbekistán y Tiam-Sham occidental, en el mismo se ubican Cicer arietinum, Lens esculenta, Phaseolus aureus, Ph. mungo, Pisum sativum y Vicia faba.
Centro de Cercano Oriente. Comprende el interior del Asia Menor, toda la Transcaucacia, Irán y las tierras del Turkmenistán, a este centro corresponden las especies Cicer arietum (subespecie pisiforme), Lathyrus cicera, Lens esculenta, L. albus, L. lenticula, L. nigricans, L. kotschyana, L. orientalis, Lupinus pilosus, L. angustifolius, Medicago sativa, Onobrycyis altisima, O. transcaucasica, Pisum sativum, P. pelatius, P. humile, P. fulvum, Trifolium resupinatum, Triconella foenum, Vicia ervilia, V. pannonica, V. sativa y V. villosa.
Centro Mediterráneo. Se caracteriza por sus plantas cultivadas de importancia más limitada que las del centro anterior e incluye Cicer arietum, Ervum monathos, Hedysarum coronarium, Lathyrus corgoni, L. ochrus, L. cicera, L. satirus, Lens esculenta, Lupinus albus, L. termis, L. luteus, L. angustifolius, Ornithopus sativus, Trifolium repens, T. incarnatum, T. alexandrinum, Ulex europeus, Vicia sativa, V. ervilia y V. faba.
Centro Abisinio. Comprende Etiopía, la que tiene una peculiar flora cultivada; además, las montañas de Eritrea, de donde son originarias Cicer arietum, Dolichos niger, Lathyrus sativus, Lens esculenta, Lupinus termis, Pisum sativum, Trigonella foenum, Vicia faba y Vigna sinensis.
Centro Mexicano Meridional y Centroamericano. Está delimitado por el sur de México y América Central, cuyas floras las componen plantas endémicas cultivadas que se diferencian de forma notable del vasto continente de América del Norte, donde la agricultura ha tenido como base las plantas cultivadas introducidas de otras partes del mundo. Se considera de este centro Canavalia ensiformis, Phaseolus acutifolium, Ph. multiflorus, Ph. lunatus y Ph. vulgaris.
Centro Sudamericano. Lo constituyen Perú, Ecuador y Bolivia, siendo éste el que contiene las principales especies de alta montaña, la puna y la sierra. Las especies son Lupinus mutabilis, Phaseolus lunatus y Ph. vulgaris.
Además se clasifican dos subcentros del anterior, el de Chile que carece de leguminosas y el del Brasil y Paraguay donde se ubican Arachis hypogea y Phaseolus caracalla (Whyte et al., 1955).
Sin embargo, recientemente Mehra y Magoon (1974) han descrito cuatro centros de genes principales de leguminosas que son: Nuevo Mundo, Africano, Indio y Este de África. Todos estos autores han demostrado que en esas regiones es donde ocurre la variabilidad de las especies.
Del total de especies de leguminosas en el mundo el 50% son tropicales, pero solo 10 ó 12 son usadas como pastos a escala relativamente amplia en los trópicos (Funes, 1979).
Importancia de la prospección
El trabajo de prospección con el objetivo de recolectar especies autóctonas y/o naturalizadas ha brindado grandes aportes en especies y han brindado además datos de considerable valor para sustentar las teorías actuales sobre la evolución y distribución de las especies. Particularmente, podemos citar entre otros, los trabajos de W. Hartley y J.L. Stephens en 1947-1948 (Hartley, 1950) con cuyos resultados este autor pudo preparar los mapas de distribución para cada una de las tribus de gramíneas (Whyte, 1958).

Capítulo I. Revisión bibliográfica
3
Los trabajos desarrollados por N.I. Vavilov durante el período comprendido entre la primera y segunda guerra mundial, a los que hemos hecho referencia anteriormente (Vavilov, 1950), J.F. Miles quien colectó especies de pastos en África en 1952 (Willians, 1964); R.L. Buró que colectó principalmente leguminosas en América Central y del Sur en 1971; R. Reid que trabajó en igual sentido en Cuba y América del Sur. Estos especialistas y otros han contribuido al descubrimiento de materiales de gran valor que actualmente forman parte de las variedades promisorias tanto en programas de investigación como de desarrollo y comercial.
La búsqueda de especies nativas o autóctonas y las naturalizadas (que se han extendido y adaptado) que en muchos casos han derivado en formas evolucionadas (Funes, 1979) pueden ser empleados frecuentemente como un material valioso en los programas de mejora genética; no obstante, en los países subdesarrollados una de las mejores opciones sería evaluar el potencial productivo de estas especies e introducirlas en la práctica agrícola.
En Cuba existe, después del triunfo de la Revolución, un programa de investigación estructurado para el desarrollo agrícola, donde la prospección ocupa un lugar importante. El desarrollo técnico alcanzado en nuestro país y su riqueza de leguminosas (Yepes, 1971; Menéndez y Machado, 1978) obliga a la búsqueda, estudio y recolección de nuestra flora autóctona, a lo que se añade la importancia de conservar el germoplasma original de estas especies. El alcance de este trabajo puede ser de relevante interés para muchas regiones de América Latina y del Caribe que son homoclimas (Meigs, 1951).
Necesidad del estudio de las leguminosas
Las leguminosas se conocen desde hace más de 6 000 años y durante ese período se ha reconocido su importancia para la agricultura en el mundo debido a su alto valor biológico y a la facultad de fijar el nitrógeno atmosférico, característica que las hacen independientes de los abonos nitrogenados.
Las leguminosas mejoran la estructura del suelo, controlan la erosión y sirven de materia orgánica en el suelo, además sirven de alimento a los animales y al hombre.
Las leguminosas tropicales pueden fijar entre 100 y 280 kg de N/ha/año. El Primer Secretario del PCC planteó en la Plenaria de Ganadería de 1974: “Nuestra producción láctea tendríamos que buscarla fundamentalmente en los pastos de gramíneas y leguminosas en la medida en que podamos ir introduciendo estas últimas”.
Este planteamiento tiene una vigencia actual, ya que la utilización de las leguminosas como un componente del pasto merece consideración especial en aquellos países en desarrollo donde la proteína es frecuentemente un factor limitante para la producción animal, el fertilizante dedicado a los pastos es escaso y de elevados precios y existe una prolongada estación seca que disminuye el crecimiento en las gramíneas (Pérez Infante, 1970).
Según los datos del Banco Mundial, los precios de los fertilizantes nitrogenados mantendrán una tendencia a subir anualmente entre un 8 y 15% (FAO, 1975), más específicamente desde 1978-1985 la tendencia alcista variará entre el 7 y el 36% (Quimimport, 1978, datos inéditos).
En conexión con esto, tenemos que más del 50% de los suelos dedicados a la ganadería en Cuba son de baja fertilidad y la carga animal promedio/ha en vacas lecheras era en 1976 1,7 aproximadamente, mientras que en vacas y novillas fue alrededor de 1,2 cabezas/ha en el mismo período, según indican los datos de la Reunión Nacional de Pastos (1976). En este caso, se puso de manifiesto que las provincias de mayores cargas fueron Pinar del Río, La Habana y Las Villas, produciéndose un descenso progresivo hacia las provincias orientales.
Sin embargo, en otros países donde se han desarrollado programas de investigación para estudiar el uso de las leguminosas (Skerman, 1977; Jones y Jones, 1978) y en Cuba (Quesada y Oliva, 1971; Febles y Padilla, 1972; Funes y Pérez, 1976; Rodríguez, 1977; Monzote, 1977; Menéndez y Martínez, 1978) se ha mostrado que este tipo de cultivo puede soportar hasta 2 cabezas/ha.
Estas evidencias ponen de manifiesto la posibilidad de elevar la carga-animal en grandes áreas dedicadas a la explotación ganadera, solamente con el empleo de las leguminosas.
No obstante, estos resultados experimentales no se corresponden con los prácticos en áreas ganaderas cubanas, argumentándose que una de las causas fundamentales son las dificultades con el manejo de los pastizales de leguminosas, pudiéndose citar los fracasos con el kudzú (Pueraria phaseoloides) a mediados de la década del 60; de la alfalfa (Medicago sativa) y el gandul (Cajanus cajan) a finales de este mismo período en las provincias occidentales principalmente; así como con la glycine (Neonotonia wightii), que fue sembrada en grandes áreas, fundamentalmente en La Habana. Estos fracasos disminuyeron el interés de los ganaderos cubanos por la utilización de las leguminosas.
Principales características de las leguminosas más importantes en Cuba
La familia de las leguminosas (Leguminosae) es una de las más grandes dentro de Spermatophytae con un estimado de 700 géneros y 14 000 especies (Kretschmer, 1974), existiendo en Cuba 125 especies nativas (Yepes, 1971b).

Capítulo I. Revisión bibliográfica
4
Las características comunes de este grupo de plantas son un elevado valor biológico, su beneficio al suelo debido al aporte de nitrógeno, materia orgánica y al mejoramiento de su estructura. Existen formas arbustivas y herbáceas, siendo estas últimas las más importantes para ser empleadas como forrajeras.
Las leguminosas más importantes para Cuba pertenecen en su mayoría a las subfamilias Phaseoloides y Vicioideae (Yepes, S., 1974, datos inéditos) que se caracterizan por agrupar a plantas herbáceas, con flores completas, conspicuas y forma mariposada, normalmente entomófilas. El fruto es una legumbre dehiscente o indehiscente que presenta hojas compuestas (generalmente trifoliadas) y alternas, además posee nódulos radicales y viven en simbiosis con las bacterias del género Rhizobium.
Whyte, Nilsson-Leisner y Trumble (1955) plantean que las leguminosas son sensibles a los cambios climáticos, debido a que la fijación simbiótica y su efecto en el desarrollo de la planta dependen, entre otros factores, del coeficiente de asimilación del carbono, el cual a su vez, se rige por la intensidad y duración de la luz. Además, el carácter mesofítico de las hojas de la mayoría de los ejemplares de este taxón, así como la gran superficie foliar y su fisiologismo, sugieren en general una reacción más intensa entre los cambios climáticos y edáficos, muy superiores a los que suelen tener la mayoría de las demás plantas forrajeras.
En este sentido debemos considerar lo planteado por Clark, Leslie y Hemken (1966) cuando informan que una mezcla de composición botánica adecuada es difícil de mantener durante largos períodos, debido a que las gramíneas y las leguminosas responden de manera diferente a los sistemas de manejo, fertilización y condiciones ambientales.
Sin embargo, Whyte et al. (1955) sugieren que la asociación de las leguminosas con las gramíneas es la forma más sencilla y conveniente de asociación de pastos.
Entre los géneros más importantes forrajeros para Cuba se destacan: Centrosema. Es un género que agrupa alrededor de 30 especies en los trópicos y subtrópicos, son plantas
rastreras, perennes y volubles. Se destaca C. pubescens, que muestra una tendencia trepadora cuando se asocia a diferentes gramíneas. Las variedades foráneas estudiadas en Cuba no han mostrado buen comportamiento en asociaciones o en cultivo puro, bajo sistemas de corte y pastoreo (Menéndez, J, datos inéditos).
Desmodium. El género comprende cerca de 200 especies perennes o anuales que se producen en su mayoría en las regiones templadas y tropicales del hemisferio occidental, África tropical y Australia. Las especies D. intortum y D. uncinatum son los más ampliamente difundidos en la explotación ganadera. Toleran suelos ligeramente ácidos (Yepes, 1974) y con baja fertilidad. D. intortum no se encuentra en tierras bajas tropicales, sino en las de mediana altura, mientras que D. uncinatum aparece en altitudes inferiores y mayores que la especie anterior desde en Veracruz en México hasta el norte de Argentina. En Cuba se desarrollan 37 especies (León y Alain, 1951), pero ninguna se cultiva comercialmente.
Leucaena. Comprende unas 50 especies de adaptación especial, originaria de Centroamérica con especies en las islas del Pacífico. Las especies más conocidas son L. leucocephala, L. retusa y L. pulverulenta (Yepes, 1974).
Macroptilium. Agrupa unas 10 especies de plantas erguidas y trepadoras, generalmente trifoliadas, identificándose 4 especies en la flora cubana (León y Alain, 1951 citados por Menéndez y Pereira, 1979). La especie más importante es M. atropurpureum, cuyo centro de distribución se encuentra en las regiones tropicales de América, de crecimiento más vigoroso en la época de lluvias (Menéndez, 1978), mientras que su crecimiento por debajo de 14°C se detiene (Whiteman y Lulham, 1970). El siratro presenta plántulas vigorosas que le permiten un buen establecimiento y posee una buena habilidad competitiva que le permite establecerse con facilidad mediante el empleo de un cultivo mínimo y entre pasto natural (Monzote, 1977).
Medicago. Este género tiene más de 50 especies que se adaptan a períodos de seca y a todos los suelos que no sean deficitarios de cal o demasiado húmedos. M. sativa es una de las especies que más ampliamente se usa mundialmente. En Cuba, las plantaciones deben renovarse cada año en occidente debido a una elevada humedad relativa y precipitaciones en estas zonas (Zambrana, 1971). Sin embargo, bajo las condiciones climáticas de Guantánamo, los resultados de rendimiento y persistencia en producción corroboran los obtenidos experimentalmente para esa zona (Menéndez et al., 1980).
Neonotonia (antes Glycine). Es una leguminosa originaria de África, aunque se han encontrado algunos tipos en el sur y sureste de Asia. Se ha diseminado por el Caribe, norte, centro y sur de América y Australia. En Cuba se reconocen 3 variedades: Tinaroo, Cooper y Clarence. Neonotonia es conocida en Cuba y en otros países como glycine, kudzoide o soya perenne. Según Bogdan (1966) es una especie perenne y postrada, pero con hábitos de trepar a las gramíneas u otras plantas en busca de luz. Las hojas son trifoliadas. La especie N. wightii tiene gran adaptación a diferentes condiciones climáticas y muestra un buen comportamiento en áreas donde la precipitación anual fluctúan entre 27-33/22-28°C aproximadamente (día/noche). Aunque sus requerimientos de suelos son considerados específicos, prosperan bien suelos Ferralíticos Rojos y los arcillosos fértiles, así como en Aluviales (Thomas y Whiteman, 1971).

Capítulo I. Revisión bibliográfica
5
Pueraria. Es un género tropical originario del Asia Oriental y las islas del Pacífico. El más difundido es el kudzú (P. phaseoloides) que crece en los suelos arenosos. En la actualidad se ha introducido en casi todos los países tropicales, pues se adapta a suelos pobres y se establece con lentitud en suelos arcillosos (Whyte et al., 1955).
Stylosanthes. Posee 30 especies tropicales, especialmente de África y Brasil. En Cuba hay 3 especies: S. hamata y S. viscosa, que también se extienden por Jamaica y América del Sur y S. tuberculata, que crece en Camagüey, Cayo Romano y Las Bahamas (Yepes, 1974). Muchas de las especies son utilizadas en la ganadería, combinan bien con ciertas gramíneas y son muy resistentes a la sequía (Whyte et al., 1955). Posee especies perennes y su desarrollo es mejor en climas cálidos. La especie S. guianensis se deriva de un material introducido en Australia desde Brasil, es muy tolerante a condiciones de baja fertilidad y puede crecer en suelos ácidos, así como ha demostrado alguna tolerancia a condiciones de drenaje deficiente (Humphreys, 1967).
Teramnus. Algunas especies de este género han demostrado posibilidades como planta para el ganado, siendo T. labialis el más promisorio. Es una especie perenne, estolonífera, de tallos finos y de gran habilidad asociativa (Menéndez, J.; Araujo, Sol y Sardiñas, Nancy, 1980, datos inéditos). Yepes (1974) describe 2 variedades encontradas en Cuba a) variedad mayor, de entrenudos y hojas mayores y semillas clara y b) variedad menor, de entrenudos y hojas menores y semillas negras.
Factores que intervienen en la explotación de las leguminosas
Entre los factores más importantes para la explotación exitosa de los pastizales de leguminosas tenemos el establecimiento, la nutrición y el manejo, considerado este último como el fundamental debido al fracaso con este grupo de plantas en nuestro país.
Establecimiento. El establecimiento es la etapa más crítica en el desarrollo de un pastizal, ya que para que ocurra con éxito deben ser favorables las condiciones y preparación del suelo, las labores aerotécnicas antes y después de la siembra, la fecha, distancia y densidad de siembra, así como con el momento de iniciar la explotación del mismo y en leguminosas, particularmente, el empleo de un inóculo efectivo y eficiente y el conocimiento de las exigencias edáficas del cultivo.
El desarrollo inicial de las leguminosas tropicales está afectado por numerosos factores, entre los que merecen citarse, la nodulación tardía, la cual se ha relacionado en Glycine con su lento establecimiento (Sistachs y López, 1979), que junto al tamaño de las semillas dan lugar a plántulas pequeñas que son dominadas con facilidad por las malezas (Yepes, 1973, comunicación personal). Muy relacionados con estos dos factores están la preparación del suelo y el empleo de una cepa efectiva y eficiente de Rhizobium (Sistachs y López, 1979).
La densidad de siembra es otro factor, ya que se ha comprobado que la rapidez con que se cubre el área de siembra está directamente relacionada con el peso de semilla viable sembrada (Bogdan, 1966; Humphreys, 1974). La fecha de siembra también tiene gran influencia en el establecimiento de leguminosas particularmente en regiones como las de Cuba en que existe una prolongada estación seca, que se relaciona con el tipo de suelo, la temperatura, las características de la especie y la incidencia de malezas (Jones y Jones, 1971; Jones, 1975).
Nutrición. Los estudios han demostrado que aunque las condiciones del suelo permitan una adecuada formación de los nódulos, el sistema simbiótico puede fracasar si no hay suficiente fijación de nitrógeno, que afecta el desarrollo normal de la planta hospedera por la interferencia que se crea en el metabolismo del nitrógeno (Sistachs y López, 1979).
Una deficiente nutrición de la planta de azufre, calcio, cobre y fósforo pueden, bajo ciertas circunstancias, disminuir el porcentaje de nitrógeno.
El molibdeno, desempeña un importante papel en la fijación simbiótica del nitrógeno como componente de la enzima nitrogenasa y el boro lo es para el desarrollo de los meristemos y la conformación de los tejidos vasculares (Hudder, 1948; Norris, 1964).
Sin embargo, son pocos los estudios que se han llevado a cabo relacionados en la nutrición en el trópico y particularmente aquellos que se refieren a la fijación simbiótica, que adquiere singular importancia por su necesidad para lograr un buen establecimiento mediante una apropiada nodulación, así como desde el punto de vista económico como ahorro de nitrógeno. Así Sistachs y López (1979), informan que una fijación simbiótica de 100 kg de N/ha equivale a una aplicación de 500 kg de sulfato de amonio. Sin embargo, según estos mismos autores estos beneficios son obtenidos solamente cuando el Rhizobium apropiado está presente en el suelo para tomar los nódulos radicales y hacer efectiva la fijación.
Además, el nitrógeno fijado simbióticamente se encuentra en forma orgánica, la que está menos sometida a los factores que pueden determinar la pérdida de este elemento.
En la práctica, muchos suelos, específicamente en los que se han introducido nuevas especies de la subfamilia Vicioedeas, ocurre que frecuentemente la cepa de Rhizobium adecuada no se encuentra, o es

Capítulo I. Revisión bibliográfica
6
escasa e inefectiva, que es lo que le ha ocurrido con frecuencia en las siembras masivas de alfalfa efectuadas en el país.
Manejo. El manejo del pastizal tanto para el comienzo como para su explotación permanente ha sido determinante en la persistencia de pastizales de leguminosas.
Los trabajos experimentales han puesto de manifiesto que el manejo ha sido determinante en el éxito o fracaso de los pastizales y áreas forrajeras de leguminosas, así Valdés y Pérez (1971) y Febles y Padilla (1972), cuando sometieron asociaciones de gramíneas y leguminosas a pastoreo rotacional con elevadas cargas y un corto período de establecimiento o inicio de explotación, observaron una paulatina desaparición de las leguminosas en la flora del pastizal.
Otros autores como Jones (1967) y Whiteman (1979) que han trabajado con siratro, plantean que cuando se incrementó la frecuencia de corte se redujo la produjo de éste en praderas mixtas, debido a la pequeña regeneración del índice de área foliar (IAF) después del corte. Jones (1971) indica que en verano un IAF de 4 sólo es posible después de 8-12 semanas con este mismo cultivar, mientras que Jones (1974) planteó que el rebrote después del corte tiene una relación lineal con el IAF remanente. Winter (1976) asoció 12 gramíneas con 5 leguminosas en suelo Amarillo poco fértil que al ser sometidas a cortes desaparecieron 4 especies de leguminosas como resultado de los rendimientos más altos en las gramíneas, perdurando solamente Stylosanthes guianensis cv. Shofield.
Con el empleo del sistema de pastoreo diferido Salinas y Milera (1972) han obtenido producciones de hasta 13,1 L de leche/vaca con alta persistencia en el pastizal; Echevarría (1977) también informó alta producción de leche y persistencia en la leguminosa empleando un sistema de manejo similar y la misma leguminosa (glycine). Rodríguez Femenías (1977) señala buen comportamiento en asociaciones de gramíneas y leguminosas, pastoreadas por vacas lecheras de mediano potencial durante más de 5 años en sistema rotacional, pero con intervalos de reposo más cortos y más baja carga que los empleados por Valdés y Pérez (1971) y Febles y Padilla (1972).
Valdés, Montoya y Batista (1977) plantean ganancias de peso de 108 g/animal en seca con animales para producción de carne en la época seca cuando éstos consumían leguminosas, mientras que los que pastoreaban en pasto natural tuvieron pérdida de peso.
Todos estos trabajos nos indican la importancia de las leguminosas en la agricultura. Funes y Pérez (1976) sometieron a corte 3 cultivares de glycine y Teramnus labialis y obtuvieron
rendimientos alrededor de 12 t de MS/ha/año, sin que se apreciaran pérdidas notables en la población. Estos resultados coinciden con los alcanzados por Guzmán (1967); Boado (1969); Funes, Yepes y Hernández (1971) y otros.
Valor nutritivo. Las leguminosas son un alimento de mayor calidad comparado con las gramíneas, lo que está relacionado con la importancia conferida por Hensell (1969) y Hutton (1970) a este grupo de plantas para la alimentación bovina.
Febles y Padilla (1972) en Cuba, obtuvieron rendimientos de 3 t/ha de proteína. Funes et al. (1971) obtuvieron tenores de 16 a 18 y hasta 20% de proteína en Desmodium intortum y Teramnus labialis respectivamente y según Salette (1971) leguminosas con establecimiento satisfactorio pueden contener entre 14,4 y 18% de proteína y alrededor de 10% en malas condiciones.
El nivel de producción animal a partir de los pastos está directamente relacionado con el valor nutritivo de éstos, ya que la producción animal depende fundamentalmente del consumo voluntario, la energía y proteína digestible y el contenido mineral (Milford y Minson, 1966). Además la digestibilidad se ve afectada por la edad del pasto y está altamente correlacionada con el contenido de proteína (Milford y Haydeck, 1965; Henzell, 1969; Hutton, 1970).
Las proteínas son muy ricas en proteínas de las llamadas “completas”, principalmente en lisina, aminoácidos de gran valor en la síntesis de los constituyentes de la leche, especialmente la caseína, tienen buena provisión de vitaminas A, B, C y D, son ricas en calcio y su contenido en fósforo mayor que el de las gramíneas.
Show (1961) informa que la inclusión de la leguminosa Stylosanthes humilis en un pastizal aumentó la capacidad de carga en 3 veces, la ganancia de peso vivo en 5 veces y se disminuyó el tiempo de ceba en 1 año.
Henzell y Norris (1962) y Henzell (1968) indican que las leguminosas pueden elevar el contenido de nitrógeno de la gramínea asociada de 1,8 a 2,4% siendo también mostrado por Moore (1962). Hutton y Banner (1960) señalan que los rendimientos anuales de nitrógeno en las partes aéreas de Leucaena leucocephala sometida a corte son tan altos como 580 kg/ha de este elemento.
Smith (1963); Horrell (1964); Boncearelli y Convarelli (1965); Horrell y Court (1965); Febles y Padilla (1972) sugieren que en general las leguminosas influyen en la transferencia de nitrógeno a la gramínea asociada y en rendimiento de MS y composición botánica, especificando que una asociación de gramíneas-leguminosas, generalmente rendía más cantidad de forraje que cualquier gramínea no fertilizada o leguminosa por sí sola.

Capítulo II. Metodología experimental
7
CAPÍTULO II. METODOLOGÍA EXPERIMENTAL Ubicación y extensión del archipiélago cubano
El archipiélago cubano pertenece a la región del Caribe o Antillas Mayores, limita al norte con el Estrecho de la Florida y la península del mismo nombre; al noreste con las Islas Bahamas y el Océano Atlántico; al noroeste con el Golfo de México; mientras que al oeste limita con el Canal de Yucatán, al este con el Paso de los Vientos y la isla que componen Haití y República Dominicana y al sur con el Mar Caribe.
La integran la isla de Cuba y de la Juventud y un gran número de cayos adyacentes al norte y al sur de Cuba. Su superficie total es de 110 922 km2, y de ellos, Cuba ocupa 105 007 km2, Isla de la Juventud 2 200 km2 y el resto (3 715 km2) a los cayos que rodean la isla mayor (Instituto Cubano de Geodesia y Cartografía, 1978).
El sistema de producción del estado Cubano es socialista y el peso fundamental de la economía está sustentado por la agricultura.
Principales regiones físico-geográficas o naturales de Cuba
El archipiélago cubano está dividido en 5 regiones principales (fig. 1) con las siguientes características. Región Occidental. Es la región más intensa de Cuba, con aproximadamente 535 km de longitud, que se
caracteriza por la gran variedad de las formas del relieve y de sus paisajes naturales y culturales. Una de sus principales subregiones es la Sierra de los Órganos, formada por cadenas montañosas paralelas
que encierran extraordinarios valles intramontanos que presentan numerosos mogotes. En la parte sur de la región occidental se extiende una gran llanura, que es aluvial al oeste y al este presenta características cársicas y suelos rojos.
Posee una red hidrográfica poco desarrollada, pero tiene grandes recursos de agua subterránea. Las ciénagas litorales ocupan extensas porciones de la parte meridional. En la actualidad se caracteriza, por
el más alto grado de aprovechamiento agrícola de la tierra y la que posee la mayor densidad de población del país, principalmente en las cercanías de la capital (Ciudad de La Habana).
La diversidad de las condiciones naturales de la región ha determinado una especialización agropecuaria multifacética.
Región Central. El elemento más importante de esta división es el macizo montañoso de Guamuhaya o alturas de Trinidad-Sancti Spiritus (Escambray) ubicado al sur de la región. Tiene una longitud aproximada de 80 km y se haya dividido de norte a sur en dos subgrupos, por el río Alabama el subgrupo del este se denomina de Sancti Spiritus y el del oeste de Trinidad, en las que se levantan alturas superiores a 1 500 m. Presenta una red hidrográfica relativamente densa y presenta ciénagas litorales en el norte y sur de la región.
En las áreas montañosas queda algo de vegetación boscosa, mientras que en las llanuras hay grandes extensiones cultivadas.
Región Camagüey-Maniabón. Es la región más llana de Cuba, la constituye un gran peniplano que se extiende desde los montes de Guamuhaya hasta las cuencas de Banes y Nipe. Entre la zona coralina del norte y peniplano está la Sierra de Cubitas y entre la ciénaga del sur y el propio peniplano se levanta la Sierra de Najasa, ambas de escasa elevación. Al este del peniplano Florida-Camagüey-Las Tunas y cerca de la costa septentrional, hay una zona bastante diferente (el grupo de Maniabón) donde se encuentran cerros más o menos cónicos, carsificados.
Aunque es una de las regiones menos poblada, fue víctima de la deforestación que realizaron los latifundistas ganaderos y azucareros en etapas anteriores al triunfo de la Revolución y es la zona cañera y azucarera más importante del país.
Región Oriental. Es el territorio de más altas montañas y de los ríos más caudalosos de Cuba. El accidente geográfico más notable de la región es la Sierra Maestra que tiene una longitud de 340 km y 30
km en su mayor ancho, extendiéndose junto a la costa sur desde Cabo Cruz hasta la cuenca de Guantánamo. Está formada por varias cadenas paralelas, con numerosos picos que sobresalen los 1 000 m, siendo su punto culminante el Pico Turquino (1 974 m).
La cuenca de Santiago de Cuba es de origen tectónico y divide a la Serra Maestra en dos partes, la del oeste o Sierra del Turquino y la del este o Sierra de la Gran Piedra. Al norte de la Sierra Maestra se abre el gran valle del Cauto, al este del cual se eleva el domo de la Sierra de Nipe y al este de la misma, se extiende el extremo más oriental de Cuba, la subregión montañosa Sagua-Baracoa, donde también se alturas mayores de 1 000 m.
Esta región se distingue por la variedad de las condiciones climáticas. En ella se encuentran desde las zonas de más elevada precipitación hasta las que poseen rasgos de aridez, como en la costa sur cerca de Guantánamo.

Capítulo II. Metodología experimental
8
Fig. 1. Principales regiones físico-geográficas. Región de Isla de Pinos. La compone la Isla de la Juventud y cayos circundantes. En términos generales
geográfica y geológicamente es muy similar que la parte oeste de la región occidental. La parte norte es un peniplano arcilloso y arenoso con drenaje radial y la parte sur es una llanura cársica sin
drenaje superficial. Entre la zona septentrional y la llanura cársica se extiende la ciénaga Lamier
Ecología
El archipiélago cubano está conformado por un mosaico edáfico y vegetal, donde generalmente varía o predomina un tipo y especies de suelo y vegetación respectivamente por cada región, variando también las condiciones climáticas.
Los suelos cubanos (Instituto Cubano de Geodesia y Cartografía, 1978) se clasifican en 13 tipos genéticos más los escabrosos que es la denominación que dan a los montañosos. En la región de Occidente predominan los escabrosos, Arenosos Cuarcíticos y los Ferralíticos Rojos, en la Central los Pardos con y sin carbonato; en Camagüey-Maniabón predominan los Pardos con Carbonato, Ferralítico Rojo y Cuarcítico y Oscuro Plástico; mientras que en la Oriental son mayoritarios los escabrosos y Pardos con Carbonato y en Isla de Pinos los Arenosos Cuarcítico.
El clima del archipiélago es clasificado como tropical de sabana (Köpen, 1936), caracterizado por dos épocas bien definidas, una de lluvia (mayo-octubre) y otra de seca (noviembre-abril) ocurriendo alrededor del 80% de las precipitaciones en el período lluvioso (figs. 2 y 3). La temperatura media anual es de 25,2°C con máxima absoluta de 38,6°C registrada en Guantánamo y mínima absoluta de 1,0°C, registrada en Alacranes, Matanzas. El promedio anual de humedad relativa es de 80%, más alta en que en seca y el promedio anual de precipitación es de 1 375 mm, con menores volúmenes hacia el sur y el este extremo y mayores en las montañas y el norte (figs. 2 y 3). La evaporación media anual es más intensa hacia las provincias orientales (fig. 4).
La flora varía generalmente según el tipo de suelo y manejo, entre ellos, la carga animal es uno de los factores que influyen en la flora y según se aprecia en la figura 5 es variable en todo el territorio e incluso a nivel de provincias.
Sistema de muestreo
Después de definir las diferentes regiones físico-geográficas en que se dividió el archipiélago cubano se procedió a muestrear las mismas. Se empleó el muestreo visual para definir los estratos donde se recolectaron las leguminosas forrajeras en estudio. En este sentido debemos señalar que no se definió a priori un número de muestreos a seleccionar debido a que esto dependía de la topografía aérea. Así, en los estratos seleccionados anteriormente se utilizó un muestreo sistemático definido de la siguiente forma: selección de muestras cada 0,5-10 km de manera que en las áreas llanas se muestreaba cada 10 km y en las occidentales cada 0,5 km.

Capítulo II. Metodología experimental
9
Fig. 2. Precipitaciones en el período lluvioso
Fig. 3. Precipitaciones en el período seco.
Además de las muestras antes mencionadas se seleccionaron muestras representativas en todos aquellos lugares donde crecían leguminosas de interés.
Este sistema permitió tomar muestras al 17% del área total. Para cada muestra tomada a ambos lados y sentido de la ruta trazada se anotó el lugar (con la ayuda del
mapa y/o vecinos de esa zona) con sus características particulares de vegetación existente, cultivo principal, topografía, tipo de suelo, etc.
Se describió la morfología de la leguminosa in situ, su fenofase, uso y abundancia en cada área en particular, así como el vigor, hábito de crecimiento y la presencia o no de nódulos, plagas y enfermedades. Al repetirse dos o más veces la misma especie, solo se tomaban como muestras aquellas que presentaban rasgos morfológicos marcadamente diferentes. Después de haber realizado varios muestreos y determinar la homogeneidad en el material existente y en el hábitat, se continuaba la trayectoria hasta localizar un nuevo cambio y de no haberlo, a los 10 km como máximo, se proseguía el muestreo descrito. Cada muestra así obtenida recibía un número arábigo consecutivo que indicaba la especie y otro romano que representaba el lugar de recolección.
Capítulo II. Metodología experimental
10
Esta información también se recogía en una plantilla de prospección confeccionada al efecto. La especie se identificaba en el campo en la mayoría de los casos o al finalizar la jornada con la ayuda de las clasificaciones existentes (León y Alain, 1951).
Fig. 4. Evaporación media anual (evaporímetro clase “A”).
Fig. 5. Densidad del ganado vacuno por agrupaciones.
Clasificación de la flora empleada en las regiones físico-geográfica
Las leguminosas colectadas fueron clasificadas en dos grupos: géneros principales y menos importantes. El primer caso comprendía las especies que mostraban mayor vigor, habilidad asociativa, distribución, abundancia y pertenecen a géneros conocidos de cierta importancia económica para las áreas tropicales y subtropicales.
Los menos importantes eran aquella que no reunían las cualidades antes mencionadas. La flora acompañante se dividió en dos grupos: pratenses y no pratenses. El primer grupo se subdividió de
acuerdo al hábito de crecimiento, vigor y manejo de estas especies. Así, se crearon los subgrupos guinea, faragua y macollosas similares (Andropogon y Paspalum); pitillas (Botriochloa y Dichanthium); cespitosas artificiales (Cynodon y Digitaria); vegetación de sabana (Sporobolus, Aristida y similares).

Capítulo II. Metodología experimental
11
Las no pratenses se subdividieron en cenagosa o costera (Rhizophora, Cyperus); espinosa (Acacia, Pisonia); Malvaceas y similares (Hibiscus, etc.), arbustos, árboles, otras leguminosas y creciendo solas. Este último caso comprendía especies “más importantes” que aparecían en general sin flora acompañante.
Cuando los géneros de la flora acompañante aparecieron más del 85% de las veces asociada a géneros de leguminosas en las áreas muestreadas se consideraba que era la vegetación la que con preferencia se asociaban las especies de leguminosas.

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
12
CAPÍTULO III. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN DE OCCIDENTE Y DE ISLA DE PINOS
Materiales y métodos
Localización. La región de Occidente está localizada al oeste del archipiélago cubano y comprende las provincias de Pinar del Río, La Habana, Ciudad de La Habana y Matanzas; mientras la de Isla de Pinos comprende la Isla de la Juventud y cayos adyacentes en sendas regiones (fig. 1).
Clima. De acuerdo a la vegetación estas regiones se clasifican de la siguiente forma: a) Zona de bosques tropicales temporalmente húmedos, mayormente caducifolios y acucifolios en algunas
partes pantanosas. La precipitación anual fluctúa entre 1 500 y 2 000 mm, el coeficiente hidrotérmico en el período lluvioso es de 2 a 3 y de 0,7 a 2 en el período de seca, siendo del tipo cálido con un acumulado anual de temperaturas de 9 000 a 9 500°C, esta característica se extiende por toda la zona central de occidente hasta la Ciénaga de Zapata.
b) Zona de bosques no muy húmedos, bosques de sabana con algunos pantanos, con una precipitación anual entre 1 000 y 1 500 mm y coeficiente hidrotérmico 1 a 2 y de 0,5 a 1 en el período de lluvia y de seca respectivamente, con un acumulado o suma de temperatura de 9 000 a 9 500°C. Este tipo climático afecta el norte y el sur de la región de Occidente y la mayor parte de la de Isla de Pinos.
Ambas zonas se caracterizan por presentar, al igual que la media nacional, dos épocas bien definidas de 6 meses cada una, la de lluvia (mayo-octubre) donde caen alrededor del 80% de las precipitaciones y la de seca (noviembre-abril) con el resto de las precipitaciones (fig. 2 y 3), con una evaporación media anual que va de 1 600 a 1 750 con máximas de 1 800 en el norte de La Habana y Matanzas (fig. 4).
Suelos. Occidente es la región más extensa (535 km de longitud aproximadamente) y constituye un verdadero mosaico geológico, caracterizado por las variadas formas del relieve y de sus paisajes naturales y culturales. Entre sus subregiones principales está la Sierra de los Órganos, formada por cadenas paralelas que encierran valles intramontanos que presentan numerosos mogotes y en la parte sur se extiende una gran llanura, que es aluvial al oeste y el este presenta características cársicas y suelos rojos. Tiene la red hidrográfica poco desarrollada pero posee grandes recursos de agua subterránea con ciénagas litorales que ocupan extensas porciones de la parte meridional.
Esta región ha sufrido una gran deforestación provocada por el hombre y en la actualidad posee el más alto grado de explotación de la tierra y diversificación, así como la mayor densidad de población. La región de Isla de Pinos es parecida geográfica y geológicamente a porción sur de Pinar del Río, con la parte norte formada por un peniplano arcillosos y arenoso, con drenaje radial y la parte sur la constituye una llanura cársica sin drenaje superficial, entre esta zona y la septentrional ( se extiende la Ciénaga de Lamier). La isla fue muy deforestada y actualmente se han creado nuevos paisajes culturales y grandes extensiones ocupadas por cítricos y otros frutales, así como por pastos artificiales. La carga media (animales/ha) es de la más alta del país en la región de Occidente (fig. 5).
En Pinar del Río e Isla de la Juventud hay un equilibrio entre los suelos Arenosos Cuarcíticos (entre los que hay Ferralítico Cuarcítico) y escabrosos, mientras que en Matanzas y La Habana predominan los Ferralíticos según se aprecia en la figura 6.
Resultados Generalidades
Fueron efectuadas 423 muestras en 143 paradas, más las observaciones continuas desde el vehículo. Se colectaron 121 muestras de leguminosas que tuvieron una frecuencia total de 566, para promediar 4 leguminosas por parada. Fueron colectadas un total de 29 géneros con características forrajeras entre los que se diferenciaron 61 especies (tablas 1 y 2). Del total de especies 40 (65,5%) pertenecen a géneros de importancia para el trópico. En la tabla 1 se observa que Desmodium fue el género que mayor frecuencia tuvo (143) y el que más especies presentó. Las especies más frecuentes fueron Alysicarpus vaginalis, Desmodium abscendens (45), D. scorpiurus, C. coeruleum y otras cinco que se encontraron 15 ó mas veces (tabla 1). Las especies de menos importancia (tabla 2) tuvieron en total menor frecuencia que el género Desmodium. En general, la abundancia y frecuencia de los géneros más importantes fue casi 3,5 veces mayor que los menos importantes; además los primeros predominaron en más tipos de suelos que los segundos, aunque en los suelos Ferralítico Rojo y Arenoso Cuarcítico fue donde más leguminosas (de ambos grupos) se encontró. Se apreció que los géneros más importantes se mostraron más colonizadores que los demás, ya que 3 aparecieron en todos los suelos y el resto, por lo menos en dos, pero la mayor parte en áreas de explotación; mientras que el otros grupo se ubicó, generalmente, en áreas marginales, lo que evidencia aún más este carácter en los de
Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
13
más importancia para cubrir grandes áreas (fig. 7) y asociarse a un mayor número de vegetación que el otro grupo (tabla 3 y 4). Además, los nódulos observados en todas las especies estaban en la fase de descomposición, excepto los no efectivos o de coloración verde que aparecieron en pocas especies.
Fig. 6. Suelos de las regiones de Occidente e Isla de la Juventud.
Tabla 1. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en la región de Occidente e
Isla de Pinos.
Suelos Género Especie Total de especie Frecuencia
I II III IV V VI VII VIII IX Alysicarpus vaginalis 1 51(51) X X X X X X X X X Calopogonium coeruleum 3 37(29) X X X X X X X X Centrosema pubescens 4 56(24) X X X X X X X X X Desmodium scorpiurus 11 143(43) X X X X X X X X X Eriosema crinitum 1 3(3) X Galactia spiciformis 5 24(15) X X X X X X X Indigofera mucronata 3 10(5) X X X X X X Leucaena leucocephala 1 16(16) X X X X X X Lysiloma bahamensis 1 5(5) X X X Macroptilium atropurpureum 2 33(18) X X X X X X Neonotonia wightii 1 5(5) X X X X Pueraria phaseoloides 1 9(9) X X Rhyncosia minima 2 22(21) X X X X X X X Stylosanthes hamata 1 4(4) X X Teramnus labialis 2 19(5) X X X X Zornia diphyla 1 2(2) X Total 40 439(255) 10 9 11 5 11 9 16 7 6
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) “Escabroso” (IV) Oscuro Plástico (V) Húmico Carbonático (VI) “Diente de perro (VII) Arenoso Cuarcítico (VIII) “Costero” (IX) Cenagoso X Suelo en que se encontró la especie principal 51 Frecuencia total (51) Frecuencia de la especie principal

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
14
Tabla 2. Especies menos importantes, frecuencia y suelo donde se encuentran en la región de
Occidente e Isla de Pinos.
Suelos Género Especie Frecuencia I II III IV V VI VII VIII IX
Abrus precatorius 1 X Aeschynomene americana 3 X X Albizia lebbeck 10 X X X X X Caesalpinia vesicatoria 2 X Cajanus cajan 1 X Canavalia gladiata 1 X Canavalia maritima 8 X X X X X Cassia aeschynomene 1 X Cassia dyphyla 15 X X Cassia savanarum 5 X Crotalaria retusa 13 X X X X X X X X Crotalaria sp. 1 X Desmanthus virgatus 12 X X X X Mucuna pruriens 2 X X Phaseolus adenanthus 1 X Tephrosia cinerea 5 X Tephrosia corallicola 5 X X X Tephrosia senna 7 X X X X X Vigna antillana 4 X X Vigna peduncularis 9 X X X Vigna vexillata 21 X X X X X X X Total 127 14 5 7 2 7 7 12 1 2
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) “Escabroso” (IV) Oscuro Plástico (V) Húmico Carbonático (VI) “Diente de perro (VII) Arenoso Cuarcítico (VIII) “Costero” (IX) Cenagoso
Tabla 3. Vegetación que acompaña a los géneros principales en la región de Occidente e Isla de Pinos.
1 2 3 4 5 6 7 8 9 10 11
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y m
acol
losa
s si
mila
res
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Árb
oles
Cre
cien
do
sola
Alysicarpus X & & & X X X X X - Calopogonium & & & X & & X X & X X Centrosema & & & X & & & & X - X Desmodium X X & & & & X & & & X Eriosema - - - - & - - - - - X Galactia - X X - - & X X X X Indigofera X X & & - - X - - - & Leucaena & & & X X & X X X - X Lysiloma X X X - - X - & & - & Macroptilium & & & & X & X X X X Neonotonia & & X - - & - X X - Pueraria X & X - X - - - - & X Rhyncosia X & & - X & X X - - X Stylosanthes - X & - X - - - - - X Teramnus & & & X - X - X - - Zornia - - - - & - - - - - & X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
15
Tabla 4. Vegetación que acompaña a los géneros menos importantes en la región de Occidente e Isla de Pinos.
1 2 3 4 5 6 7 8 9 10 11 12
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y m
acol
losa
s si
mila
res
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
cena
gosa
o
cost
era
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Árb
oles
Cre
cien
do
sola
Abrus - - - - - - & - - X - Aeschynomene - & & - X X - X - - - Albizia X & & - - - - - X - X & Caesalpinia - - - - - - X - - - - & Cajanus X - & - - - X - - - Canavalia - - X - - & - - - - - & Cassia - & & - & - X - X - - X Crotalaria - & & & & - X & X - - X Desmanthus X & X X X - - & X - - X Mucuna X - - - - - & X - X - Phaseolus - - - - - - & - X - - Tephrosia - & & X X - - - - - - & Vigna X X X - - & & X X - - X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
Leyenda V Alysicarpus vaginalis Centrosema pubescens Calopogonium coeruleum Centrosema virginuanum
Calopogonium muconoides Centrosema plumieri Calopogonium sp. ∅ Centrosema lobatum
Fig. 7. Distribución de los géneros Alysicarpus, Calopogonium y Centrosema (región Occidental e Isla de Pinos).
Descripción y distribución de los géneros principales
Los géneros más importantes se encontraron mayormente acompañados por gramíneas de diferente hábito de crecimiento y altura, donde se cuentan árboles y arbustos, así como otras leguminosas del mismo o de otro género. En algunas ocasiones aparecían junto a otras plantas herbáceas, volubles y espinosas que servían evidentemente de protección contra el consumo por parte de los animales domésticos de esas zonas.
Alysicarpus. Se observó A. vaginalis creciendo con más preferencia mezclado al follaje de los estratos inferiores de gramíneas cespitosas y macollosas como guinea (P. maximum), faragua (H. rufa), caguazo (P.

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
16
virgatum), arbustos y plantas espinosas (tabla 3), donde se observó una mayor altura que cuando crecía entre las gramíneas cespitosas, pero con menor densidad y mayor vigor cuando crecía junto a faragua y otras similares, y no entre las otras macollosas como guinea. No fue vista creciendo sola. También apareció acompañada por otras leguminosas de varios géneros (Desmodium, Centrosema). El estado fenológico que predominó fue el de semilla madura con legumbres secas en sus racimos terminales. Estaba presente en ambos suelos de ambas regiones (tabla 1), pero fue más abundantes en los Ferralíticos (I), Arenosos (VII), Oscuro Plástico (IV) y Húmico Carbonático (V), de toda el área, excepto en La Habana donde su presencia fue escasa en los pastizales en explotación intensiva (fig. 7). En todas las zonas se mostró libre de patógenos y se vio consumido por los animales.
Calopogonium. Fue un género que mostró tres especies (C. coeruleum, C. muconoides y C. sp.), C. coeruleum fue la especie más prometedora por su amplia distribución, vigor y facilidad de asociarse con todo tipo de vegetación por ser voluble, que fue la característica más destacada. Esta especie se caracterizó por cubrir la especie acompañante y sobresalir marcadamente en áreas marginales, mientras que crecía armónicamente entre gramíneas de áreas pecuarias donde era consumido por los animales. El estado fenológico observado fue el de semilla madura, semilla verde, floración plena y vegetativo, los cuales se mantenían en equilibrio mostrándose las plantas vigorosas y libres de plagas y enfermedades. Las dos especies restantes aparecieron equilibradas entre gramíneas, aunque C. muconoides estaba en estado senil y C. sp., que es una especie intermedia entre las anteriores, apareció con semilla verde y madura, ambos son menos vigorosos que C. coeruleum, que se encontró en todos los suelos, excepto en Cenagoso (tabla 1) de todo el área, menos en La Habana (fig. 7), C. muconoides fue encontrado en suelo Ferralítico Rojo de Matanzas y La Habana y C. sp. sólo en el Arenoso mal drenado de Isla de la Juventud.
Centrosema. Este género contribuyó con cuatro especies (C. lobatum, C. plumieri, C. pubescens y C. virginianum), entre los cuales se destacó C. pubescens por mostrar mejores características pratenses, con enraizamiento en los nudos, tallos más finos, mayor ramificación y una distribución más amplia que el resto. Esta especie se observó formando armónica asociación con faragua y otras macollosas de césped abierto, también con pitillas y vegetación de sabana-esportillo (Sporobolus indicus), pajón (Andropogon gayanus) y otras. Además, con Malváceas y similares, así como con otras leguminosas aunque también asociada a guinea, pangola (D. decumbens), bermuda (C. dactylon), entre arbustos y plantas espinosas o creciendo sola, pero con menos vigor que con las primeras (tabla 3).
Sin embargo, en todos los ejemplares de esta especie se observó, en mayor o menor grado, síntomas de virosis o después de concluida la fase juvenil que fueron más marcados en plantas más viejas. C. plumieri se presentó vigorosa y crecía, fundamentalmente, junto a guinea, pitillas y faragua a las que cubría al igual que C. lobatum, que mostró menos vigor, no así C. virginianum, que apareció asociada a plantas erguidas y de poco follaje, como Malváceas, aroma (A. farnesiana) y otros similares, pero sin sobresalir, excepto en La Habana del Este donde apareció cubriendo toda la vegetación acompañante en áreas donde afloran las rocas calizas. Con respecto al estado fenológico C. pubescens mostró todos los estadios, con predominio del de semilla madura y verde, mientras que en C. virginianum predominó la floración plena con semilla verde. En los demás la fase principal fue semilla madura con muchas de las hojas secas en C. plumieri y algunas en C. lobatum, pero libres los tres últimos de plagas y enfermedades.
El género se encontró en todos los suelos de la zona, aunque predominó C. pubescens (tabla, fig. 7); mientras que C. virginianum prefirió los suelos costeros, “diente de perro”, Ferralítico y Arenoso y C. plumieri en los escabrosos, Oscuro Plástico, Húmico Carbonático, Ferralítico y Cenagoso, con mayor exuberancia en las zonas de mayor precipitación. La distribución de estas especies aparece en la figura 7 y todas son volubles. Solamente se observaron consumida por animales C. pubescens y C. virginianum.
Desmodium. Fue el género que más especies presentó (D. adscendens, D. affine, D. axillare, D. barbatum, D. canum, D. distortum, D. obtusum, D. procumbens, D. scorpiurus, D. tortuosum y D. triflorum). De estos los que más sobresalieron fueron D. scorpiurus, que fue uno de los de mayor frecuencia (43) junto a D. triflorum (30) y D. affine (9), que aparecieron entre gramíneas cespitosas, vegetación de sabana media talla y con otras leguminosas sin suponerse a las mismas en áreas de pastoreo y corte, así como en áreas marginales, en la mayoría de los suelos de todo el área (tabla 1). Las demás especies fueron encontradas con menor frecuencia y en lugares muy específicos, excepto D. adscendens que tuvo una frecuencia de 45, pero apareció siempre creciendo bajo arbustos y entre matorrales de suelos escabrosos, Arenosos, Oscuro Plástico y Húmico Carbonático de toda el área (fig. 8). Sin embargo, fue común en todas las especies el vigor y predominó en ellos fase vegetativa, mostrándose libres de plagas y enfermedades. No se observaron nódulos activos.
Todos son consumidos por los animales y ninguno es voluble. Eriosema. Se encontró en tres ocasiones E. crinitum en suelo Arenoso de los menos fértiles en Isla de la
Juventud creciendo junto a espartillo y sola, con semilla madura y pocas hojas (fig. 8). Presentaba pocas hojas, erguido y de menos de 30 cm de altura. Es afectado por pagas en su follaje.

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
17
Galactia. Se encontraron 5 especies (G. earlei, G. galactoide, G. parviflora, G. spiciformis y G. striata), entre las que sobresalió G. spiciformis, ya que se presentó con mayor vigor en la mayoría de los suelos (tabla 1, fig. 8) en áreas marginales de Isla de la Juventud, Pinar del Río y Matanzas. Los restantes fueron encontrados en Isla de la Juventud (norte) G. striata; en Pinar del Río, mientras que en suelo Cenagoso y Húmico Carbonático G. galactoide y G. parviflora y G. earlei se encontraron en suelo Escabroso de Pinar del Río y Matanzas la primera y entre casuarinas de la costa norte en carretera Matanzas-Varadero en el km 16 la segunda, siendo la que mayor vigor mostró comparada con las demás que no llegaban a rebasar la vegetación acompañante, que en su mayoría eran arbustos espinosos. En todas predominó la fase de semilla madura, los animales las consumían y mayormente se presentaron afectados por hongos.
Indigofera. De este género fueron encontradas tres especies: I. hirsuta, I mucronata e I. sumatrana. La más sobresaliente fue I. mucronata de porte decumbente que apareció en cuatro suelos (tabla 1), entre gramíneas cespitosas (tabla 3) sobre las que sobresalía y formaba una adecuada asociación bien consumida por los animales en áreas de pastoreo de la llanura de Matanzas y en suelo Ferralítico Rojo. I. hirsuta se presentó creciendo en población poco densa sin otro tipo de planta acompañante en suelo Arenoso de Cayo Gordo, en Isla de la Juventud, pero los animales no la consumían, ya que todas las plantas se encontraban intactas y en estado senil avanzado. La otra especie fue I. mucronata, de crecimiento erecto, similar a la anterior, pero de mayor altura, apareció con plantas aisladas en suelo Ferralítico, Oscuro Plástico y Arenoso de Matanzas y Pinar del Río, entre gramíneas cespitosas (fig. 8). Todas se observaron en estado senil con gran cantidad de legumbres y hojas secas, pero en I. mucronata predominaban las verdes y mantenían una buena proporción de hojas verdes. Las especies se mostraron libres de plagas y enfermedades y los nódulos se observaron en descomposición.
Leucaena. Aunque solo con L. leucocephala se encontró esta especie en seis suelos (tabla 1) y entre casi toda la vegetación (tabla 3) formando poblaciones compactas, a la orilla de las costas arenosas y rocosas o en otras áreas marginales, pero siempre sobresaliente entre la vegetación acompañante, fundamentalmente, en los suelos alcalinos y bien drenado (fig. 8). No se observaron síntomas de plagas y enfermedades y su estado fenológico fue el de semilla madura con reinicio del estado vegetativo y los animales la ramoneaban.
Lysiloma. Al igual que leucaena se encontró una sola especie (L: bahamensis) distribuida en zonas costeras de la Ciénaga de Zapata hasta los límites con Cienfuegos y en Santa Lucía, en la costa norte de Pinar del Río (fig. 8), crecía fundamentalmente, sola y entre arbustos y con leucaena, con los que formaba poblaciones compactas, aunque entre estas crecían otras leguminosas (tablas 1 y 3). Esta especie se mostró libre de plagas y enfermedades, predominando el estado vegetativo con abundante y vigoroso follaje y pocas legumbres secas y verdes, pero no apareció consumida por animales.
Fig. 8. Distribución de los géneros Desmodium, Eriosema, Galactia, Indigofera, Leucaena y Lysiloma (Occidente e Isla de Pinos).

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
18
Macroptilium. Se encontraron M. atropurpureum y M. lathyroides, siendo el primero el de mayor importancia, ya que apareció en equilibrio o predominando entre la vegetación acompañante (tabla 3), aunque prefirió los suelos bien drenados, ya que en Matanzas apareció solo en suelo Ferralítico, en Pinar del Río en el mismo tipo de suelo y en arenoso; mientras que en La Habana cercana a una construcción del central Martínez Prieto en Húmico Carbónatico (fig. 9). En general, se observaron todos los estados fenológicos, pero los ejemplares, aunque vigorosos, mostraron generalmente sus foliolos (en el envés) moteaduras de esporas del hongo Puccinia sp., la otra especie se encontró en plantas aisladas que crecían sin acompañantes de similar altura y/o hábito de crecimiento o entre cespitosas, sobre las que sobresalían por su porte erecto. Su estado fenológico fue el de semilla seca y semilla verde y no se observaron plagas ni enfermedades. Esta especie a diferencia de la anterior no se observó consumida por los animales.
Neonotonia. (antes Glycine). N. wightii fue encontrada en áreas escapadas del cultivo en suelos carbonatados (tabla 1) y vegetación propia de esos lugares (tabla 3, fig. 9). Se mostró vigorosa y voluble en La Habana y Matanzas, predominando el estado vegetativo con muchas semillas verdes, sin tener síntomas patológicos.
Pueraria. P. phaseoloides se presentó en suelos poco fértiles y ácidos de Isla de la Juventud (tabla 1, fig. 9), sobresaliendo entre faragua y especies similares y entre otra vegetación y bajo los pinares (tabla 3), donde se observó vigorosa y en estado reproductivo en equilibrio con el vegetativo. No apreció dañada por plagas y enfermedades, ni consumida por animales y sus nódulos estaban secos.
Rhyncosia. Además de R. minima que fue más abundante se encontró R. reticulata creciendo ambas entre la vegetación de un campo de henequén (Agave fourcroydes) sobre “diente de perro” (tabla 1, fig. 9). Presentaron gran cantidad de legumbres y flores en las fases de desarrollo y pocas hojas, además no llegaban a predominar entre la vegetación acompañante (tabla 3) por sus tallos endebles, además de mostrar generalmente los foliolos moteados con zonas cloróticas similar al ataque viral y no se observaron consumidas por animales.
Stylosanthes. S. hamata fue muy abundante en todo Varadero sobre suelo Arenosos y escaso en Puerto Escondido y playa Santa María al norte de La Habana, siempre entre escasa vegetación (tabla 3). No mostró presencia de plagas y enfermedades y se observó en floración plena con sus características flores amarillas en inflorescencia terminal.
Teramnus. Fueron localizadas T. labialis y T. uncinatus, mayormente en suelos carbonatados (tabla 1) donde abundó más T. uncinatus, que también predominó sobre la vegetación acompañante; mientras que T. labialis lo hacía sin sobresalir (tabla 3). Estas especies no mostraron sintomatología patológica y el estado fenológico que predominó en T. labialis fue el vegetativo, mientras que T. uncinatus se equilibraron semilla madura con semilla verde, siendo su distribución principalmente en Matanzas (fig. 9). Se observó que solo T. labialis era consumido por los animales.
Zornia. Z. diphyla apareció en los suelos ácidos de Isla de la Juventud y Pinar del Río entre vegetación muy pobre o creciendo sola con gran producción de semillas maduras y verdes y pocas hojas sin mostrar plagas y enfermedades, siendo una de las pocas especies del hábitat en esos lugares (fig. 9).
Las especies de menor importancia se encontraron con poca frecuencia y en pocos suelos (tabla 2), más comunes en los suelos Arenosos y Ferralíticos y en general no sobresalieron entre la vegetación acompañante (tabla 4), a excepción de Abrus, Albizia, Mucuna y Tephrosia cinerea que lo hicieron sobre la vegetación abierta o sin acompañante. Ninguna predominó sobre guinea, otras leguminosas, árboles y entre arbustos (tabla 4).
Discusión
Los resultados de la prospección en estas regiones indicaron la presencia de 40 especies y comprueba la presencia de leguminosas forrajeras en grandes extensiones de suelos cubanos, como ha sido planteado por León y Alain (1951) y Yepes (1975). Sin embargo, en amplias zonas de La Habana y Pinar del Río no se observaron géneros importantes o aparecieron muy pocas especies en áreas marginales (figs. 7, 8 y 9), lo que puede indicar una gradual desaparición de las mismas, aunque son especies que pertenecen a la flora de estas regiones (León y Alain, 1951).
Este comportamiento evidencia lo planteado por Anon (1976), Menéndez y Machado (1978, Funes (1979) referente a la necesidad de recolectar el material nativo y a la influencia del manejo de la tierra en la persistencia de las leguminosas, ya que las cargas y niveles de nitrógeno elevados influyen negativamente en la presencia de leguminosas en la flora del pastizal (Bryan y Evans, 1973). Además, Mejías, Hernández, Villamizar y Chaverra (1968) puntualizan que el pastoreo intensivo hace que las leguminosas desaparezcan entre 1 y 2 años. Estas condiciones prevalecen en la mayoría de los pastizales de La Habana (fig. 2), donde Neonotonia; no obstante ser una de las leguminosas que más ampliamente fue sembrada en esta provincia, solo apareció en áreas marginales con poca frecuencia, lo que pone de manifiesto que el manejo para esta especie fue desfavorable. Este aspecto fue planteado por Valdés y Pérez (1971) y Febles y Padilla (1972). Estos resultados

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
19
se reafirman con el hecho de que la mayor distribución de especies en esta región fue en la provincia de Matanzas, donde la carga predominante fue inferior a 0,9 animales/ha aproximadamente (fig. 5).
Fig. 9. Distribución de los géneros Macroptilium, Neonotonia, Pueraria, Rhyncosia, Stylosanthes, Teramnus y Zornia (Occidente e Isla de Pinos).
En La Habana particularmente han desaparecido importantes especies de leguminosas, excepto Alysicarpus,
Desmodium scorpirus y D. triflorum, aunque estuvieron en menor proporción que otras zonas. El hábito de crecimiento rastrero, la morfología y el pequeño tamaño de las especies que prevalecieron favorecieron su permanencia en estas zonas, ya que excepto en casos extremos cuando se produce un sobrepastoreo estas leguminosas son desfoliadas completamente, lo que no ocurre con frecuencia, por lo que en la mayoría de las áreas pratenses el índice de área foliar (IAF) remanente para estas especies es adecuado que constituye uno de los factores importantes para mantener el contenido de leguminosas en el pastizal (Whiteman, 1969; Blazer, 1974).
Un análisis de la frecuencia de distribución muestra que todos los géneros encontrados prevalecieron en suelos Arenoso Cuarcítico, mientras que las especies más importantes aparecieron en los Escabroso, Ferralítico Rojo y Fersialítico Pardo rojizo (tabla 1). Los géneros de mayor frecuencia fueron Alysicarpus, Calopogonium, Centrosema, Desmodium y Macroptilium y las especies predominantes A. vaginalis, D. Scorpiurus y C. coeruleum. Por otro lado, los géneros de menos frecuencia eran Eriosema, Neonotonia, Pueraria, Stylosanthes y Zornia, mientras que E. crinitum, L. bahamensis, N. wightii, S. hamata y E. diphyla fueron las especies menos frecuentes que en su mayoría son de hábito de crecimiento erguido.
Una característica general fue que alrededor del 90% de los géneros de leguminosas encontrados tenían hábito de crecimiento rastrero y voluble y tenían poca altura, además todas fueron observadas asociadas a gramíneas y otras plantas, que se manifestó más marcadamente en los suelos de baja fertilidad, lo que pudiera ser una forma de adaptación de este grupo de plantas para subsistir a las condiciones adversas del medio. Este comportamiento coincide con los resultados de Capote y Shishchenco (1977a, b), quienes trabajando con gramíneas y leguminosas encontraron que al disminuir la nutrición del sustrato, las leguminosas incrementaron la biomasa en sentido inverso a la intensidad luminosa. Estas condiciones son similares a las que ocurren en las asociaciones naturales, donde se observó mayor vigor en las leguminosas asociadas en las que crecían

Capítulo III. Leguminosas forrajeras silvestres de la región de Occidente y de Isla de Pinos
20
como cultivo puro en suelos de baja fertilidad, además eran más abundantes en estos que en los más fértiles, aunque en estos últimos también pudo haber influido el cultivo intenso.
Otras características del suelo también pueden haber alterado la presencia y distribución de este grupo de plantas. Así, Macroptilium atropurpureum apareció, fundamentalmente, en suelo Ferralítico Rojo (asociado a gramíneas cespitosas y macollosas, así como a otras plantas), siendo las propiedades de estos suelos similares a los preferidos por esta especie según t’Mannetje (1967; 1974), aunque también pudo desarrollarse en un amplio rango de suelos desde pH 4,5 (suelos ligeros), alcalinos o en arcillosos en guinea y el noroeste de Australia (Jones y Jones, 1978). Además, esta especie ha tenido buen establecimiento cuando se introduce en pasto natural (Monzote, 1977) debido posiblemente a que posee plántulas vigorosas igual que Calopogonium y logra un establecimiento más rápido que otras leguminosas tropicales (Yepes, Alfonso y Funes, 1971). Sin embargo, C. coeruleum se encontró en todos los tipos de suelos excepto en los Arenosos Cuarcíticos de fertilidad extremadamente baja de Isla de la Juventud y Pinar del Río, que concuerda con las características de suelo señaladas para este género por Whyte et al. (1955).
Centrosema y Galactia fueron encontrados en la mayoría de los suelos, aunque prefirieron los fértiles y se mostraron resistentes a la sequía. Centrosema se presentó en áreas de pastoreo y marginales, mientras que Galactia solo apareció en áreas marginales, donde mostró poco vigor, debido posiblemente, a la susceptibilidad ante el ataque de hongos presentándose también C. pubescens afectado por virus que ha sido el principal causante de la pérdida de vigor y la poca persistencia manifestada en asociaciones y cultivo puro (Menéndez et al., 1980, datos inéditos).
Eriosema, Pueraria, Zornia y Stylosanthes prefirieron condiciones de suelos muy específicos. Los tres primeros se encontraron en los suelos menos fértiles y pH ácido, peor el último prefirió los salinos de pH básico, en todos los casos de textura arenosa y en áreas no pecuarias marginales. Estas especies se incluyen como importantes para las zonas de alto contenido de aluminio y pH ácido en América Latina (CIAT, 1977).
Un análisis aparte merece Leucaena, que apareció fundamentalmente, en zonas costeras de pH básico, aunque Whyte et al. (1955) informa que esta especie se adapta bien a suelos de baja fertilidad, por lo que la no presencia en áreas pecuarias pudiera atribuirse a que la hayan eliminado al confundirla con la aroma y el marabú, ya que es una especie agresiva, según pudo observarse en todas las zonas.
Otras especies de interés como I. mucronata, L. bahamensis, R. minima y T. labialis aparecieron con frecuencia de baja a media, aunque se distribuyeron en general, en más de dos suelos, pero mostraron poco porcentaje en la flora, excepto Lysoloma que fue abundante en suelos mal drenados e Indigofera en suelo Ferralítico Rojo también en abundancia entre gramíneas cespitosas artificiales, mientras que Teramnus se observó poco vigoroso y Rhyncosia apareció atacada por virus, comportamiento que pone de manifiesto la influencia de las condiciones ambientales en la distribución de las especies.
En general, se observó que los géneros de leguminosas prevalecieron en suelos Arenoso Cuarcítico que son particularmente poco fértiles y de pH bajo, mientras que las especies más importantes aparecieron en los suelos escabrosos, Ferralítico Rojo y Fersialítico Pardo rojizo. Sin embargo, las especies menos importantes aparecieron, generalmente, en los suelos Ferralítico Rojo y Arenoso Cuarcítico, pero su frecuencia fue baja y la presencia en esas condiciones fue muy escasa que pone de manifiesto una gradual desaparición de estas especies en la flora autóctona, posiblemente por asociarse a muy pocas plantas y a su estructura en general.
Se observó además que las leguminosas más importantes en general, aparecieron asociadas fundamentalmente a gramíneas cespitosas y macollosas, evidenciándose, la preferencia por este tipo de ecosistema, que pudiera ser una de las formas de mantener el componente de leguminosa en pastizales en explotación, además debe tenerse en cuenta la especificidad por determinado tipo de suelos manifestada por Macroptilium atropurpureum, Eriosema, Pueraria, Stylosanthes y Zornia, así como la mayor adaptación de Centrosema y Calopogonium y aun mayor de Alysicarpus y Desmodium.
Al mismo tiempo se puso de manifiesto la acción modificadora del manejo en la flora de estas regiones y se evidenció la importancia de estudiar tan rico material que, además de servir de germoplasma, especies como Calopogonium, Centrosema, Macroptilium y Leucaena pudieran brindar sus bondades en un futuro no lejano, a grandes áreas ganaderas del país, teniendo en cuenta su distribución natural como una forma de hacer más económica la explotación pecuaria.

Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
21
CAPÍTULO IV. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN CENTRAL Y PARTE DE LA DE CAMAGÜEY-MANIABÓN DE CUBA
Materiales y métodos
Localización. La región Central está ubicada en el centro del archipiélago cubano, limitada al oeste por la región de Occidente y al este con la de Camagüey-Maniabón, extendiéndose en el norte por toda la cayería que comprende el archipiélago de Sabanas y el de Camagüey (fig. 1).
La región Camagüey-Maniabón limita al oeste con la anterior y al este con la Oriental y se prolonga al sur hasta la cayería del archipiélago de los Jardines de la Reina. Este trabajo abarcó en esta región desde el oeste hasta los límites de la provincia de Camagüey y con la de Las Tunas al este (fig. 3).
Clima. La región Central presenta en la zona central del Escambray un clima clasificado como de bosques temporalmente muy húmedos, mixtos temporalmente húmedos, caducifolios-perennifolios con precipitaciones anuales que sobrepasan los 2 000 mm. La suma de temperaturas superiores a 10°C y su coeficiente hidrotérmico es mayor que 1 en período seco y en el lluvioso supera a 3, mientras la zona intermedia del macizo montañoso del Escambray, al suroeste de esta subregión y de la región Central se caracteriza por un clima de bosques tropicales temporalmente húmedos, mayormente caducifolios y acucifolios, en algunas partes pantanosos, con precipitación anual de 1 500 a 2 000 mm y coeficiente hidrotérmico de 2 a 3 y de 0,7 a 2 en el período lluvioso y de seca respectivamente, que incluye el tipo menos cálido con una suma de temperatura menos que 9 000°C y el resto de esta región y la de Camagüey-Maniabón tienen las mismas características climáticas que las descritas para la de Occidente en el acápite b.
En cuanto a la distribución de las lluvias presentan características similares que el resto del país y descritas para la región de Occidente (figs. 2 y 3), pero la evaporación media anual varía desde 1 400 hasta más de 2 000 mm (fig. 4).
Suelos. Esta región se caracteriza por presentar al sur el macizo montañoso de Guamuhaya o alturas de Trinidad-Sancti Spiritus (Escambray) con una altitud superior a 1 000 m, con una subregión llana al norte de Domos al centro y entre estas dos la cordillera de Las Villas, mientras que la región de Camagüey-Maniabón es la más llana de toda Cuba y está constituida por un gran peniplano, con la Sierra de Cubitas y Sierra de Najasa, ambas de baja elevación. Es la más importante zona ganadera y cañera del país.
En ambas regiones hay un predominio de los suelos Ferralítico, Pardo y Oscuro Plástico según puede apreciarse en las figuras 10 y 11.
Resultados Generalidades
Se efectuaron 442 muestreos y las observaciones continuas desde el vehículo. Se colectaron 74 muestras
con una frecuencia promedio de 4 leguminosas observadas por parada, de las que se encontraron 25 géneros que contenían 43 especies. Los géneros y especies principales se muestran en la tabla 5. Se aprecia que Desmodium, Centrosema y Macroptilium tuvieron mayor frecuencia, con los dos primeros presentes en todos los tipos de suelos (Desmodium con 7 especies y Centrosema con 4), pero sin sobresalir de la vegetación acompañante en la mayoría de los casos, Macroptilium con la especie atropurpureum sobresalió entre sus acompañantes, aunque en suelos específicos; otros géneros mostraron baja frecuencia. Sin embargo, Calopogonium con la especie coeruleum tuvo un comportamiento similar a Macroptilium, aunque con una frecuencia discreta, pero mostró gran vigor y agresividad llegando incluso a cubrir árboles medianos y arbustos ocupando grandes áreas y cubriendo toda la vegetación acompañante o formando una mezcla armónica cuando crecía junto a gramíneas, donde se observaba un buen equilibrio en áreas de pastoreo y en general se caracterizaron estas dos especies por abarcar áreas más extensas que otras especies (figs. 12 y 14).
En la tabla 6 aparecen las especies que hemos considerado como las de menos importancia que incluyen forrajeras tropicales, ya que a excepción de Mucuna las restantes presentaron una baja relación hoja/tallo, aparecieron como plantas aisladas y mostraron poco vigor.
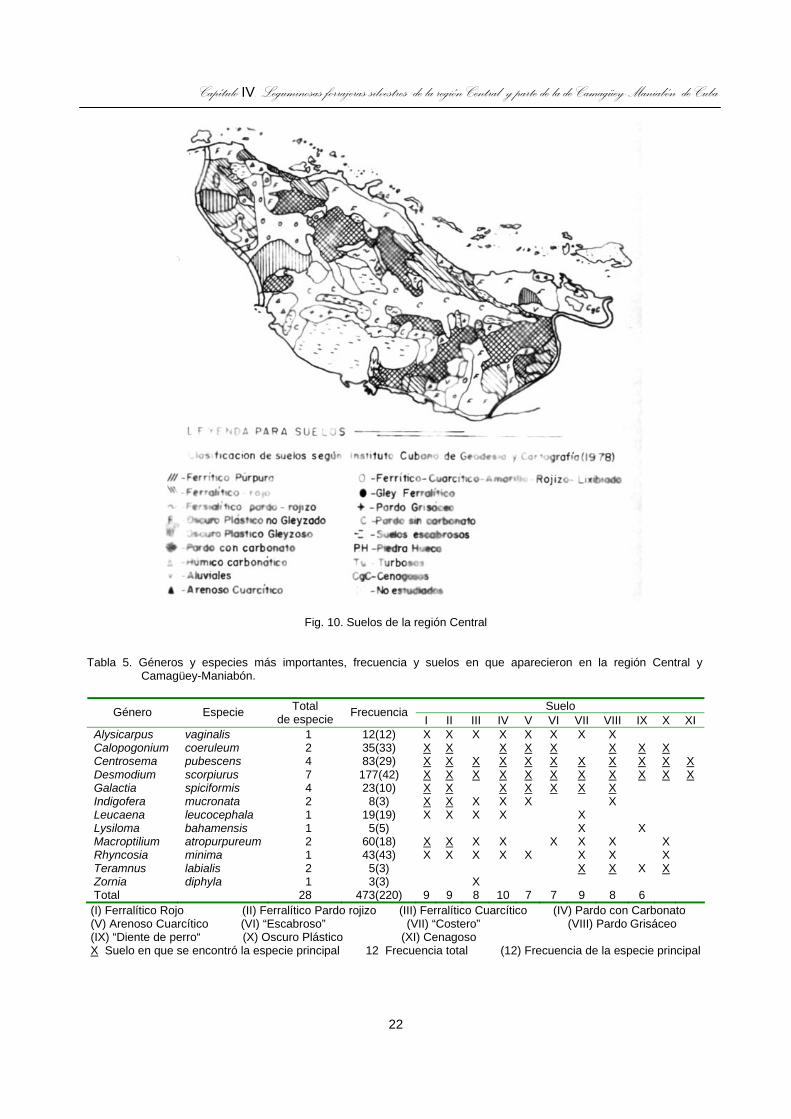
Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
22
Fig. 10. Suelos de la región Central
Tabla 5. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en la región Central y Camagüey-Maniabón.
Suelo Género Especie Total
de especie Frecuencia I II III IV V VI VII VIII IX X XI
Alysicarpus vaginalis 1 12(12) X X X X X X X X Calopogonium coeruleum 2 35(33) X X X X X X X X Centrosema pubescens 4 83(29) X X X X X X X X X X X Desmodium scorpiurus 7 177(42) X X X X X X X X X X X Galactia spiciformis 4 23(10) X X X X X X X Indigofera mucronata 2 8(3) X X X X X X Leucaena leucocephala 1 19(19) X X X X X Lysiloma bahamensis 1 5(5) X X Macroptilium atropurpureum 2 60(18) X X X X X X X X Rhyncosia minima 1 43(43) X X X X X X X X Teramnus labialis 2 5(3) X X X X Zornia diphyla 1 3(3) X Total 28 473(220) 9 9 8 10 7 7 9 8 6
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) Ferralítico Cuarcítico (IV) Pardo con Carbonato (V) Arenoso Cuarcítico (VI) “Escabroso” (VII) “Costero” (VIII) Pardo Grisáceo (IX) “Diente de perro“ (X) Oscuro Plástico (XI) Cenagoso X Suelo en que se encontró la especie principal 12 Frecuencia total (12) Frecuencia de la especie principal
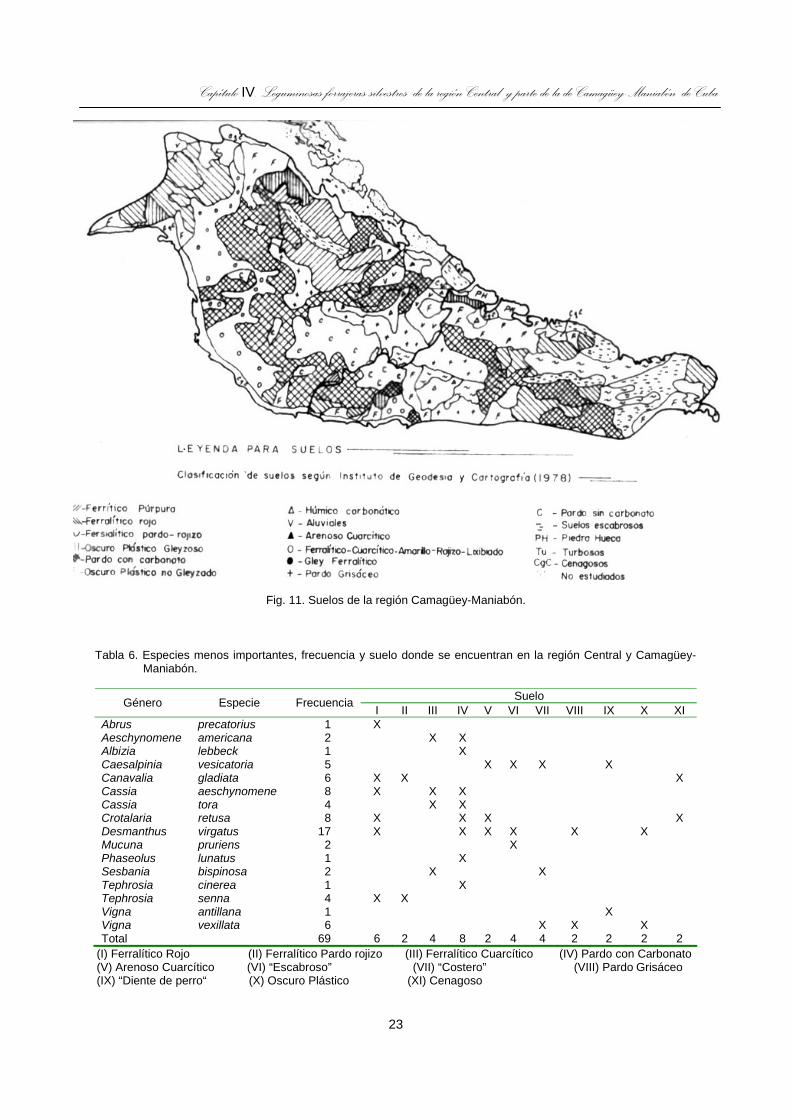
Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
23
Fig. 11. Suelos de la región Camagüey-Maniabón. Tabla 6. Especies menos importantes, frecuencia y suelo donde se encuentran en la región Central y Camagüey-
Maniabón.
Suelo Género Especie Frecuencia I II III IV V VI VII VIII IX X XI Abrus precatorius 1 X Aeschynomene americana 2 X X Albizia lebbeck 1 X Caesalpinia vesicatoria 5 X X X X Canavalia gladiata 6 X X X Cassia aeschynomene 8 X X X Cassia tora 4 X X Crotalaria retusa 8 X X X X Desmanthus virgatus 17 X X X X X X Mucuna pruriens 2 X Phaseolus lunatus 1 X Sesbania bispinosa 2 X X Tephrosia cinerea 1 X Tephrosia senna 4 X X Vigna antillana 1 X Vigna vexillata 6 X X X Total 69 6 2 4 8 2 4 4 2 2 2 2
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) Ferralítico Cuarcítico (IV) Pardo con Carbonato (V) Arenoso Cuarcítico (VI) “Escabroso” (VII) “Costero” (VIII) Pardo Grisáceo (IX) “Diente de perro“ (X) Oscuro Plástico (XI) Cenagoso
Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
24
También se comprobó que en suelos Pardo, Ferralítico, Fersialítico y costeros fue donde apareció mayor número de géneros importantes, mientras que los menos importantes se encontraron con mayor frecuencia en Pardo con Carbonato y Ferralítico Rojo.
La vegetación acompañante de los géneros más importantes aparece en la tabla 7 y estos se mostraron creciendo en comunidad sin sobresalir de la vegetación en el caso de las rastreras (Alysicarpus y Desmodium) y volubles (Galactia) o en equilibrio y/o sobresaliendo sobre gramíneas y otras familias particularmente las volubles (Calopogonium, Centrosema, Macroptilium, Rhyncosia y Teramnus), erectas (Leucaena, Lysiloma y Zornia), así como las decumbentes (Indigofera), Lysiloma y Zornia se encontraron, fundamentalmente, creciendo no acompañados de otros géneros y cuando aparecieron mezclados sobresalían sobre la vegetación existente. Sin embargo, las especies menos importantes se observaron creciendo aisladas y sin formar una población numerosa, exceptuándose Abrus y Mucuna.
Tabla 7. Vegetación asociada a los géneros más importantes en la región Central y Camagüey-Maniabón.
1 2 3 4 5 6 7 8 9 10 11
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y m
acol
losa
s si
mila
res
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Árb
oles
Cre
cien
do
sola
Alysicarpus X & & & & X & & X X Calopogonium & & & X & & & & X X X Centrosema & & & - & & & & X - X Desmodium X & & & & X & & & X Galactia - X X - - & - - X - Indigofera X X & & X X X X X - Leucaena & & & X X & X & & - X Lysiloma X X - - - & X X & - X Macroptilium & & & & X X & & X - & Rhyncosia - & X X - & & & X Teramnus & & & & - X X & - Zornia - X - - & - X - - - &
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
Descripción de los géneros principales Los géneros más importantes abarcaron una amplia zona y aparecieron acompañados por especies del
mismo género o por otros géneros de leguminosas, pero mayormente asociados a gramíneas, entre las que abundaban Andropogon, Bothriochloa, Dichanthium, Panicum y Sporobolus y en menor proporción Paspalum y vegetación de sabana o propia de cada suelo, aunque también se encontraron cubriendo árboles y arbustos.
Alisicarpus. A. vaginalis fue recolectado y observado entre el follaje de pitillas (Dichanthium spp. y Bothriochloa), guinea (P. maximum) y otros, pero donde mayor vigor y población mostró fue entre las cespitosas pangola (D. decumbens), bermuda común y cruzada-1 (C. dactylon) y las pitillas en los suelos Ferralítico, Fersialítico, Ferrítico Cuarcítico (Arenoso Cuarcítico) y Pardo con Carbonato. Apareció con menor frecuencia y poco vigor arenoso, propiamente escabroso y costero (tabla 5 y 7) en Villa Clara, Ciego de Ávila (fig. 12), donde se observó con pocas hojas en las fases de fin de floración y semilla verde madura en equilibrio.
Calopogonium. La especie C. coeruleum fue observado con muy buenas perspectivas asociado a todo tipo de vegetación, pero particularmente a las gramíneas macollosas, las pitillas, a vegetación de sabana, plantas espinosas y arbustos, aunque también se encontró cubriendo árboles (tabla 7). Esta especie se caracterizó por cubrir toda la vegetación de áreas marginales, pero se mantenía en equilibrio en áreas de pastoreo donde se observó un consumo moderado por parte de los animales que pastoreaban. Se distribuyó en la mayoría de los suelos (tabla 5), pero no se encontró en los Ferralíticos Cuarcítico de los límites Villa Clara-Matanzas, en los costeros, ni en los Cenagosos de toda el área explorada (fig. 12). La otra especie encontrada fue C. muconoides (de ciclo anual) que apareció dos veces en suelo Ferralítico Rojo de Ciego de Ávila.
Los estadíos vegetativo y todas las fases reproductivas se observó en equilibrio en C. coeruleum y el reproductivo en su fase final en la otra especie, pero ambos se mostraron vigorosos y libres de plagas y enfermedades.
Centrosema. Este género también fue observado con perspectivas con la especie C. pubescens, que apareció asociada, fundamentalmente, a faragua (Hyparrhenia rufa), pajón (Andropogon gayanus) y pitillas, así

Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
25
como otras leguminosas de su género o de otro y otros componentes vegetales. Sin embargo, no se encontró asociada a cespitosas artificiales (tabla 7), pero estuvo presente en todos los tipos de suelos (tabla 5), excepto en Cienfuegos donde no fue encontrada. Las otras especies fueros C. lobatum, C. plumieri y C. virginianum; las dos primeras aparecieron asociadas entre sí o con C. plumieri y con C. pubescens, junto a gramíneas de diversos hábitos de crecimiento, con los cuales formaban una población armónica.
En ocasiones C. plumieri se asoció a gramíneas macollosas de las que sobresalía, pero siempre en áreas no sometidas a corte y/o pastoreo, mientras que C. virginianum apareció en áreas marginales, asociado a plantas erguidas y de poco follaje en los suelos Ferralíticos y Pardos de Camagüey, Ciego y Villa Clara (fig. 12). En C. lobatum y C. plumieri predominó la fase de semilla madura y vegetativo, mientras que en C. pubescens se armonizaban los estados fenológicos vegetativo, fin de floración, semilla verde y semilla madura, pero predominó floración plena con pocas semillas verdes y maduras en C. virginianum.
C. pubescens se mostró con síntomas de ataque de virus, caracterizado por la presencia de moteaduras cloróticas o deformaciones en los foliolos, aunque las demás especies estaban libres de plagas y enfermedades.
Desmodium. Se determinaron siete especies: D. adscendens, D. axillare, D. barbatum, D. procumbens, D. scorpiurus, D. tortuosum y D. triflorum), de los cuales sobresalieron D. scorpiorus, seguido por D. triflorum caracterizados por asociarse a cespitosas entre sus estratos inferiores (sin sobresalir) en áreas de corte, de pastoreo y marginales en suelos Pardos (con y sin carbonatos), Grisáceo y Ferralítico de la provincia de Camagüey, Ciego de Ávila y Villa Clara. D. adscendens fue otra especie abundante en la flora, pero entre los arbustos y árboles de sabana y arboledas, bajo las cuales crecía junto a alpargata (Paspalum notatum) y otros Paspalum rastreros en suelos escabrosos y Ferralítico en montes del Escambray y de Sierra de Cubitas y alturas de Trinidad y de Villa Clara y al noroeste y noreste de Villa Clara. Las restantes especies aparecieron en diferentes suelos y zonas, pero sin predominar en ninguna excepto D. axillare que apareció con mayor frecuencia en suelos Ferralíticos, debajo de árboles frutales y arboledas en general, a veces asociado a D. adscendens (tabla 5 y 7, fig. 13).
Fig. 12. Distribución de los géneros Alysicarpus, Calopogonium y Centrosema (región Camagüey-Maniabón). Galactia. Aparecieron tres especies: G. revoluta, G. spiciformis y G. striata entre las que se destacó G.
spiciformis, que apareció en numerosas zonas, aunque sin sobresalir en la vegetación, a no ser en áreas marginales, donde se mantenía en equilibrio entre la vegetación erguida constituida, fundamentalmente, por plantas espinosas como zarza (Rubus durus), marabú (Dichrostachys glomerata) y aroma (Acacia farnesiana), bajo las cuales se protegía de los dientes de los animales que pastoreaban estas zona. Las otras dos especies

Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
26
se encontraron con menos frecuencia y se localizaron en suelos Pardos y Ferralíticos de Camagüey, Ciego de Ávila, norte de Villa clara y en el norte y sur de Sancti Spiritus, al igual que G. spiciformis (tabla 5 y 7, fig. 13).
Todas las especies presentaron síntomas de haber sido afectadas por hongos, menos marcados en G. spiciformis y el estado fenológico predominante fue el de semilla madura, con la presencia de pocas hojas verdes. No se encontraron nódulos activos.
Indigofera. Se encontraron dos especies, pero predominó I. mucronata, la que sobresalió entre las cespitosas artificiales (pangola y cruzada-1) y las pitillas (tabla 7) en Villa Clara sobre suelos Pardos, rojos y escabrosos (tabla 5). La otra especie fue I. sumatrana apareció en los suelos escabrosos de Cienfuegos (fig. 13).
Leucaena. Apareció con L. leucocephala, la que se caracterizó por predominar en suelos costeros bien drenados, “diente de perro”, Arenoso, Ferralítico y Fersialítico (tabla 5), pero en estos últimos en menos proporción, con una distribución hacia el norte y noroeste de Camagüey y Ciego de Ávila, respectivamente; al sur de Sancti Spiritus y Cienfuegos y al norte de Villa Clara, siempre bordeando las costas, aunque se observó a orillas de la Carretera Central (fig. 13).
Esta especie se mostró agresiva al parecer asociada y en equilibrio o sobresaliendo con otras plantas vigorosas como Lysiloma, aroma y arbustos, así como con guinea y otras macollosas (tabla 7).
Lysiloma. Se encontró L. bahamensis que es un arbusto y se distribuyó, fundamentalmente, sobre suelo costero de mal drenaje y “diente de perro”, en la costa norte de Camagüey desde la playa Santa Lucía hasta el entronque del poblado de ese mismo nombre y el sur de Sancti Spiritus y Cienfuegos desde Trinidad hasta Boca de Yaguanabo (fig. 13). Esta especie siempre sobresalió entre la vegetación acompañante, aunque mayormente apareció formando poblaciones compactas sin otras especies acompañantes (tabla 5 y 7). En todos los casos se mostró libre de plagas y enfermedades y con mucho follaje y pocas legumbres secas.
Fig. 13. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Lysiloma (región Camagüey-Maniabón).

Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
27
Macroptilium. Aparecieron dos especies (M. atropurpureum y M. lathyroides) con M. lathyroides presente en todos los suelos, pero sin predominar ni formar población en ninguna zona. Sin embargo, M. atropurpureum se mostró prometedora para la explotación pecuaria, ya que se encontró formando verdaderas asociaciones con gramíneas cespitosas y macollosas (tabla 7), aunque apareció sola en suelos Ferralíticos y Ferralíticos Pardo rojizo (tabla 5) en Sancti Spiritus desde el norte y centro has los límites de Villa Clara y en toda esta provincia (fig. 14). La fase fenológica predominante fue semilla madura; no obstante, presentar todas las fases reproductivas y sus hojas (abundantes y vigorosas) aparecieron moteadas en el envés por esporas de hongos de roya (Puccinia sp.) posiblemente; a pesar de lo que esta leguminosa predomina entre la vegetación acompañante con buen vigor.
Rhynchosia. La especie R. minima apareció creciendo poco vigorosa con constitución frágil debido a sus tallos volubles débiles entre la vegetación de áreas marginales no dedicadas a la ganadería, fundamentalmente, en suelos de difícil laboreo y vegetación erguida (tablas 5 y 7). El estado fenológico predominante fue el de floración plena y presentó síntomas de virosis.
Teramnus. Aparecieron dos especies (T. labialis y T. uncinatus), que se mostraron con buen vigor, particularmente T. uncinatus, el que cubría la vegetación acompañante (tabla 7), aunque su distribución fue limitada a dos puntos de la costa norte de Villa Clara (fig. 14).
T. labialis se encontró mezclado con gramíneas y otras plantas (tabla 7) en suelo Pardo de Encrucijada en Villa Clara (fig. 14).
Zornia. Z. diphyla fue localizada formando una población compacta y sin acompañante en suelo Arenoso Cuarcítico al noroeste de Cienfuegos. No se observó otro tipo de vegetación de consideración; mostró buen vigor y estaba libre de plagas y enfermedades, con una proporción balanceada de tallos y hojas y gran cantidad de semillas.
Fig. 14. Distribución de los géneros Macroptilium, Rhynchosia, Teramnus y Zornia (región Camagüey-Maniabón).
Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
28
Las especies de menos importancia aparecieron con poca frecuencia y ocupando pocas áreas y aparecen en las tablas 6 y 8, donde se reflejan los suelos y vegetación acompañante.
Tabla 8. Vegetación asociada a los géneros menos importantes en la región Central y Camagüey-Maniabón.
1 2 3 4 5 6 78 8 9 10 11
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y si
mila
res
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Árb
oles
Cre
cien
do
sola
Abrus - - - - - & - - - - Aeschynomene - X & - & - & X - - X Albizia X & & - - - - X - - X Caesalpinia - - - - - & - - - - Canavalia - & & - X - X - - - & Cassia - & & X X - & X - - X Crotalaria & & & & & - - X - - X Desmanthus - & & & - - X - - - X Mucuna - X - - - & & - - - Phaseolus - - - - - - & - - - Sesbania - - - - - - X - - - & Tephrosia - X & & - - X X - - & Vigna - - - - - & & X - -
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia Estas especies aparecieron, fundamentalmente, en áreas marginales donde su presencia fue escasa,
apareciendo fundamentalmente plantas aisladas, excepto Abrus y Mucuna, la primera formando parte de la vegetación de un monte y la segunda en las márgenes de los montes, además, ninguna fue observada consumida por animales.
Discusión
De los 27 géneros encontrados 14 son principales y de ellos, 8 se ubican entre los 30 informados por Febles (1973) como importantes, otros 3 han dado buenos resultados en el trópico (Funes et al., 1971; Febles, 1973; Menéndez y Machado, 1978) y solo 3 no son recomendados como destacados para condiciones climáticas similares a las nuestras (Galactia, Lysiloma y Rhynchosia). Sin embargo, se incluyen en el grupo de los más importantes por la amplia distribución y el vigor mostrado en algunas zonas (tabla 5).
Desmodium estuvo presente en todos los suelos de esta región con 7 especies, donde se evidenció que además de las condiciones edáficas, algunas características fisiológicas como el hábito de crecimiento y el comportamiento similar al de las plantas esciofilas les permitió distribuirse en diferentes ecosistemas. Así, D. scorpiurus y D. triflorum, ambos de hábito de crecimiento postrado, se encontraron entre gramíneas cespitosas de pastoreo y corte; mientras que D. adscendens de crecimiento más erguido se encontró en áreas abruptas o marginales entre los arbustos, los que prefirieron en el primer caso la sombra parcial y en el segundo la sombra completa. De forma similar se comportó Alysicarpus, aunque no apareció en los suelos de la provincia de Camagüey, posiblemente porque este fue encontrado en otras zonas generalmente entre gramíneas cespitosas que dan sombra parcial y en suelos de mediana a alta fertilidad que son los que mejor de adapta esta especie (Whyte et al., 1955), ya que en Camagüey los suelos de fertilidad más alta están poblados por guinea u otros cultivos que proyectan sombra completa.
Calopogonium, Centrosema y Macroptilium fueron géneros que sobresalieron en la flora de determinadas áreas de la región, posiblemente porque tienen como características comunes, poseer un rápido establecimiento, plántulas vigorosas (Yepes et al., 1971) y la tendencia de cubrir la vegetación acompañante, además sus tallos enraizan cuando hacen contacto con el suelo, que son características fundamentales para que estas especies abunden en zonas pecuarias y en diversas áreas marginales.
Sin embargo, estos géneros se distribuyeron en condiciones edáficas diferentes, correspondiendo a Macroptilium ocupar los suelos Ferralítico y Pardo con y sin Carbonato de la región Central, evidenciándose la preferencia de esta especie por estos tipos de suelos que coincide con las características edáficas que informa t’Mannetje (1974) para esta estirpe. Calopigonium y Centrosema, aunque se distribuyeron por toda la región, el primero no apareció en los suelos menos fértiles y el segundo no estuvo en las zonas montañosas, lo que

Capítulo IV. Leguminosas forrajeras silvestres de la región Central y parte de la de Camagüey-Maniabón de Cuba
29
puede estar relacionado con la preferencia por suelos fértiles del primero y la no admisión de un sombreo completo por el segundo informado por Whyte et al. (1955), además Centrosema, generalmente, prefirió las zonas de precipitaciones más elevadas.
Por otra parte, Leucaena y Lysiloma, aunque ambas se encontraron en áreas costeras, la primera ocupó las zonas bien drenadas de suelos Ferralítico, Fersialítico y Pardo con Carbonato y la segunda se encontró en áreas mal drenadas o ciénagas; no obstante, ser plantas bastante similares. Zornia apareció sin vegetación acompañante en suelo franco de pH ácido y baja fertilidad y una zona de baja precipitación.
Galactia, Indigofera y Teramnus eran las de menor frecuencia en sentido general. Rhychonsia minima tuvo una distribución aceptable, pero presentaba tallos endebles y apareció afectada por virosis que son factores que limitan su empleo.
Las especies menos importantes aparecieron en mayor número en los suelos Ferralítico y Pardo, pero tanto su distribución como la frecuencia resultó muy limitada, pues en general su estructura y poca habilidad asociativa pudieron no favorecer su permanencia en estas condiciones.
La forma en que se encontraron las especies en esta región evidencia la influencia de los factores ambientales en la distribución natural de las plantas, lo que concuerda con lo planteado por Good (1931; 1953); Mason (1936); Gain (1944) y Academia de Ciencias de Cuba–URSS (1970). Whyte et al., (1955) plantean específicamente la influencia de la fertilidad del suelo en el desarrollo de las leguminosas y Wilsie (1970) menciona al pH como factor importante en la distribución de este grupo de plantas.
Así, pudimos comprobar en la práctica comparativamente la región Central es más rica en especies de leguminosas que la parte oeste de Camagüey-Maniabón. Podemos señalar que en la primera abundan los suelos más fértiles y de pH más alto, así como vegetación de gramíneas cespitosas, mientras que en Camagüey-Maniabón predominan los suelos menos fértiles y la vegetación fundamental es de gramíneas macollosas o erectas en áreas pastorales. Teniendo que la condición de suelos poco fértiles no favorece el desarrollo extensivo e intensivo de las leguminosas, provocado, posiblemente, por condiciones de mayor competencia intraespecífica (Sistachs, 1980, comunicación personal).
Sin embargo, el hombre se convierte cada día más en el mayor modificador del hábitat de las plantas y en nuestro caso con las aplicaciones masivas de herbicidas, la labranza excesiva, el manejo animal, etc., modifican el ecosistema, lo que puede contribuir a que un grupo de especies informadas en esta zona no fueran encontradas en ella. Por otra parte, la fisiología y estructura de las leguminosas también pueden haber influido en su permanencia en determinado ecosistema (Jewis, 1972), ya que las especies con yemas o puntos de crecimiento cercanos al suelo, pueden soportar mejor los cortes a baja altura.
Todos estos factores conjugados pueden haber influido en la distribución de las especies, donde las más resistentes se adaptaron a las condiciones actuales, lo que hace posible recomendar como promisorias a Macroptilium atropurpureum para suelos Ferralíticos; Calopogonium para Ferralíticos, Pardos y escabrosos; Centrosema para Pardos y escabrosos y a Leucaena para suelos bien drenados. Estas leguminosas deben ser estudiadas por las perspectivas que ofrecen como constituyentes de los pastizales de esas localidades con pocos recursos, además de tener en cuenta las especies que se desarrollan con exclusividad en condiciones muy específicas.

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
30
CAPÍTULO V. LEGUMINOSAS FORRAJERAS SILVESTRES DEL NORESTE DE LA REGIÓN CAMAGÜEY-MANIABÓN Y SUROESTE DE LA ORIENTAL
Materiales y métodos
Localización. Esta zona comprende el noreste de la región Camagüey-Maniabón y el suroeste de la Oriental (provincias Las Tunas, Granma y el oeste de Holguín (fig. 1).
Clima. La provincia Las Tunas y el oeste de Holguín tienen características climáticas similares a la región Camagüey-Maniabón descritas en el capítulo anterior mientras que en Granma, en la Sierra Maestra, el clima es parecido al de la Sierra Escambray ya descrito. La llanura del Cauto se clasifica como zona de sabana árida, en partes con precipitación anual menor de 1 000 mm y coeficiente hidrotérmico de 0,5 a 1 y de 0,3 a 0,5 para lluvia y seca respectivamente, cálida con un acumulado de temperatura (+10°C) de 9 000 a 9 500°C y en la costa sur de la Sierra Maestra y el noroeste de la región Oriental muy cálida, con suma de temperaturas superior a 9 500°C (figs. 2, 3 y 4).
Suelos. El área presenta un relieve llano al oeste y montañoso al sur y noroeste, estando en el sur las mayores elevaciones del país y está compuesta por suelos escabrosos o de montaña, abundando en el resto de la región los Pardos, Oscuros Plásticos y Aluviales (figs. 11 y 15) soportando los pastos una baja carga animal (fig. 15).
Fig. 15. Suelos de la región Oriental.
Resultados Generalidades
Se efectuaron 690 muestreos, además de las observaciones continuas desde el vehículo para colectar 112 muestras, entre las cuales se determinaron 22 géneros que contenían 52 especies y/o variedades. Los géneros más ricos fueron Centrosema y Desmodium, con 8 y 9 especies respectivamente; mientras que Cassia, Clitoria, Galactia, Indigofera y Tephrosia tuvieron 3 y las restantes solamente dos.
Tanto los géneros más ricos en especies, como los más pobres, se encontraron en una amplia gama de suelos, donde se armonizan la variedad de topografía, altitud y vegetación.

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
31
Desmodium con sus nueve especies mostró una gran diversidad al ser encontrado en todos los suelos y altitudes de la zona, y además formaron comunidades con todos los componentes vegetales del ecosistema. En este último aspecto Centrosema, Macroptilium y Leucaena menos marcadamente presentaron gran similitud con la anterior.
Por otra parte, Desmanthus con una sola especie manifestó gran plasticidad, encontrándose en la mayoría de los suelos y en todas las altitudes, aunque sin sobresalir marcadamente entre la vegetación del hábitat. Teramnus y Leucaena tuvieron una amplia distribución y sobresalieron sobre la vegetación acompañante o estaban en equilibrio cuando crecía junto a gramíneas cespitosas en áreas de pastoreo como en el caso de Teramnus labialis.
Entre los géneros de limitado rango de adaptación se encuentran Sesbania, recolectado exclusivamente en la Mensura en los Pinares de Mayarí, ocurriendo en forma similar con Harpalice. Canavalia se presentó solo en suelos costeros con una sola especie y Clitoria, que aunque muy abundante, se encontró específicamente al noroeste de la ensenada de Mora, formando poblaciones considerables en suelo Pardo con Carbonato, mientras que en otros lugares se encontraban algunos ejemplares completamente aislados.
Descripción de los géneros de mayor importancia
Alysicarpus. Se localizó A. vaginalis formando asociaciones múltiples con Desmodium triflorum y D. Scorpiurus y con las gramíneas cruzada-1, pangola y/o pitillas, creciendo junto a la vegetación de sabana y semisabana en suelos Pardos carbonatados y sin carbonato y Oscuro Plástico en altitudes de 0 a 200 msnm.
En estos lugares apareció en proporciones abundantes y asociada en pastoreo de pangola y pitillas, así como en áreas destinadas a cortes de cruzada-1. Esta leguminosa de hábito rastrero cubría todos los espacios vacíos y se entremezclaba con las otras especies de la comunidad, pero nunca su altura sobrepasó la de las especies acompañantes. Su estado fenológico predominante fue el de semilla madura, con una proporción adecuada de hojas y no presentó síntomas de estar afectado por plagas ni enfermedades.
Calopogonium. C. coeruleum fue encontrada a ambos lodos de la carretera Manzanillo-Las Mercedes en las montañas de La Maestra hasta San Lorenzo y en las estribaciones del Turquino. En general, se localizó en suelos y se evidenció una mejor adaptación a las condiciones montañosas (fig. 16), donde cubría toda la vegetación acompañante, incluso árboles. Sin embargo, cuando crecía sobre cocó (en la carretera) todos sus órganos se mostraron reducidos, aunque sin cambiar de forma. Se evidenció variabilidad en esta especie, y se observó libre de plagas y enfermedades con las fases vegetativas y reproductivas en equilibrio.
Centrosema. Se encontraron seis especies: C. lobatum, C. plumieri, C. pubescens, C. virginianum, dos sin identificar y dos formas de C. plumieri que son volubles con excepción de una de las especies no identificadas. De todos estos tipos la de mayor interés fue C. pubescens que se encontró en áreas marginales y de pastoreo con una población aceptable, asociado a la mayoría de los componentes vegetales y en la mayoría de los suelos (tablas 9 y 10). Centrosema sp. que fue especie rastrera encontrada en los Pinares de Mayarí en suelo Fersialítico Pardo rojizo a 1 000 msnm, creciendo entre vegetación muy rala. Las otras especies se encontraron en una gran extensión de suelo (tabla 9) en altitudes desde 0-200 m, pero sin cubrir áreas compactas, con mayor frecuencia en áreas marginales que en las de explotación y fueron las menos abundantes, C. plumieri y C. lobatum. Las especies volubles se observaron en general, muy agresivas y tendieron a cubrir la vegetación acompañante, excepto C. virginianum que permaneció por debajo de la misma, la cual consistió en plantas erguidas con poco follaje, pero todas las áreas marginales no sometidas a corte o pastoreo. La especie que presentó variabilidad fue C. plumieri que mostró un tipo de semilla redondo y otro de semilla jaspeadas, ambos de ciclo más corto que C. plumieri y se localizaron entre Las Tunas y Jobabo (fig. 16).
Clitoria. La especie C. ternatea fue encontrada mostrando gran variabilidad, ya que se observaron ejemplares con flores azules desde el claro a oscuro y blancas en gran abundancia entre los campos de caña y terrenos aledaños del central “Luis E. Cariacedo” (fig. 16) predominaba en suelo Aluvial, mientras que en áreas del Caney de Las Mercedes se encontraron algunos ejemplares aislados, pero sin mostrar la variabilidad manifestada en el lugar anterior. En ambos lugares no hubo presencia de plagas ni enfermedades y dominaba la fase vegetativa con algunas flores y legumbres verdes y secas.
Desmanthus. Se encontró la especie D. virgatus con hierbas de hábito procumbente o ascendente y tallos leñosos, con pequeñas pinnas, esta especie aunque muy difundida se presentó en plantas aisladas que no eran de consideración en la composición botánica del hábitat (tabla 9).
Desmodium. Es uno de los géneros más difundidos de los encontrados y el más numeroso en especies: D. adscendes, D. axillare, D. barbatum, D. distortum, D. procumbens, D. scorpiurus, D. tortuosum, D. triflorum y D. uncinatus, entre los que se destacaron D. scorpiurus y D. triflorum que se mostraron formando asociaciones múltiples junto a Alysicarpus y gramíneas cespitosas como pangola, cruzada-1 y pitillas en áreas de pastoreo y de corte D. adscendens y D. axillare sobresalieron también, pero en áreas marginales y entre arbustos y matorrales y el resto se distribuyó con menor frecuencia y en menos suelos que los demás. El género se

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
32
encontró en altitudes desde 0 hasta 1 000 m (tabla 9 y 10, fig. 17) y solo D. tortuosom sobrepasó en altura a la vegetación acompañante.
Fig. 16. Distribución de los géneros Alysicarpus, Calopogonium, Centrosema, Clitoria y Desmanthus (región Oriental). Galactia. Se encontraron tres especies (G. monophyla, G. spiciformis y G. striata) entre las que sobresalió G.
spiciformis, que aparecía entre arbustos y plantas espinosas en suroeste de Las Tunas y más cercano a la costa sur de esta misma zona se encontraron las demás. Todas mostraron síntomas de haber sido dañadas por hongos y su estado fenológico predominante fue el de semilla madura con pocas hojas verdes.
Indigofera. Fueron detectadas las especies I. mucronata, I. sumatrana e I. tictoria de las que resultó más interesante I. mucronata (de hábito decumbente) por asociarse con gramíneas cespitosas, donde se incluye la pangola. Esta especie se observó en cantidades considerables en pastizales de Birán, donde era consumida por los animales, predominando la fase vegetativa con semillas verdes y maduras, más abundantes las primeras, así como se mostró libre de plagas y enfermedades.
De las otras especies se destacó I. tictoria por aparecer en zonas desérticas en el sur y en las costas al norte, siempre sin vegetación acompañante, libre de plagas y enfermedades y con todos los estadios en equilibrio, mientras que I. sumatrana se encontró también creciendo sola entre Guisa y Pino de Agua (fig. 17) con las legumbres dañadas por un barrenador y predominando la fase reproductiva.
Leucaena. L. leucocephala apareció en suelos Pardos carbonáticos y no carbonáticos y Oscuros Plásticos más abundante en las zonas costeras y en altitudes desde 0 hasta 200 m, entre gramíneas de diferente hábito de crecimiento sin formar una población densa, además hubo plantas aisladas a orillas de las carreteras y cercados entre Holguín y Cueto (fig. 17). En áreas de pastoreo su altura no rebasaba 1 m; sin embargo, en zonas marginales esta era superior a los 3 m. Todos los ejemplares observados se mostraron libres de plagas y enfermedades y en la fase fenológica de semilla madura con una gran parte de sus legumbres vacías por haber esparcido sus simientes.
Macroptilium. En este género se pudieron definir que fueron M. atropurpureum y M. lathyroides. M. lathyroides se distribuyó en una amplia gama de suelos, topografía y altitudes (tabla 9), aunque sin formar una población numerosa, crecía entre distintas especies aisladamente, excepto en Las Mercedes, en la Ciudad Escolar Camilo Cienfuegos, donde aparece formando verdaderas poblaciones. Esta especie es la que excepcionalmente se desarrollaba entre la bahía (Paspalum notatum) (tabla 10). M. atropurpureum solo se encontró en Bayamo, con poco desarrollo en suelo Oscuro Plástico y en Mayarí, donde se mostró exuberante, pero en un área muy limitada entre los pinares a los cuales subía hasta las ramas más altas. Ambas especies aparecieron libres de plagas y enfermedades con hojas lozanas y abundantes que estaban en equilibrio con el estado reproductivo con todas sus fases manifestadas.
Neonotonia (antes Glycine). N. wightii se encontró en suelo Húmico carbonático de la Ciudad Escolar Camilo Cienfuegos, escapada del cultivo y asociada con gramíneas macollosas y erectas y a arbustos, libre de plagas y enfermedades. Presentó abundante follaje y muchas flores y frutos en diferentes fases de desarrollo.
Rhynchosia. R. minima está muy difundida, fundamentalmente, en los suelos de difícil laboreo en áreas marginales hasta los 200 m de altitud (tabla 9). Apareció asociada a hierbas erectas y decumbentes, pero poco

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
33
vigoroso, a las que cubría por su carácter voluble. Presentó sus foliolos moteados con aparente sintomatología de virosis y no se encontró en zonas donde se practicaba la siega o se pastoreaba.
También se comprobó que en suelos Pardo, Ferralítico, Fersialítico y costeros fue donde apareció mayor número de géneros importantes, mientras que los menos importantes se encontraron con mayor frecuencia en Pardo con Carbonato y Ferralítico Rojo.
La vegetación acompañante de los géneros más importantes aparece en la tabla 7 y estos se mostraron creciendo en comunidad sin sobresalir de la vegetación en el caso de las rastreras (Alysicarpus y Desmodium) y volubles (Galactia) o en equilibrio y/o sobresaliendo sobre gramíneas y otras familias particularmente las volubles (Calopogonium, Centrosema, Macroptilium, Rhyncosia y Teramnus), erectas (Leucaena, Lysiloma y Zornia), así como las decumbentes (Indigofera), Lysiloma y Zornia se encontraron, fundamentalmente, creciendo no acompañados de otros géneros y cuando aparecieron mezclados sobresalían sobre la vegetación existente. Sin embargo, las especies menos importantes se observaron creciendo aisladas y sin formar una población numerosa, exceptuándose Abrus y Mucuna.
Teramnus. Se encontraron dos especies (T. labialis y T. volubilis) que forman verdaderos pastizales mixtos al asociarse a gramíneas cespitosas a las que como ninguna otras se entremezclan estrechamente y se comportaron persistentes ante el pastoreo. Se extienden por amplias áreas en Birán y Holguín. Es apetecido por el ganado bovino y conocido por “chonchohue” por los campesinos de la zona. En ambas especies no se observó daños da plagas ni de enfermedades y el estado fenológico predominante fue el vegetativo con legumbres secas y verdes y pocas flores debajo del follaje.
Fig. 17. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Macroptilium (región Oriental). Stylosanthes. Se encontró S. hamata en suelo Pardo con Carbonato entre Bayamo-Holguín entre pitillas sin
sobresalir y en estado vegetativo, ya que era consumido por el ganado y estaba libre de plagas y enfermedades.
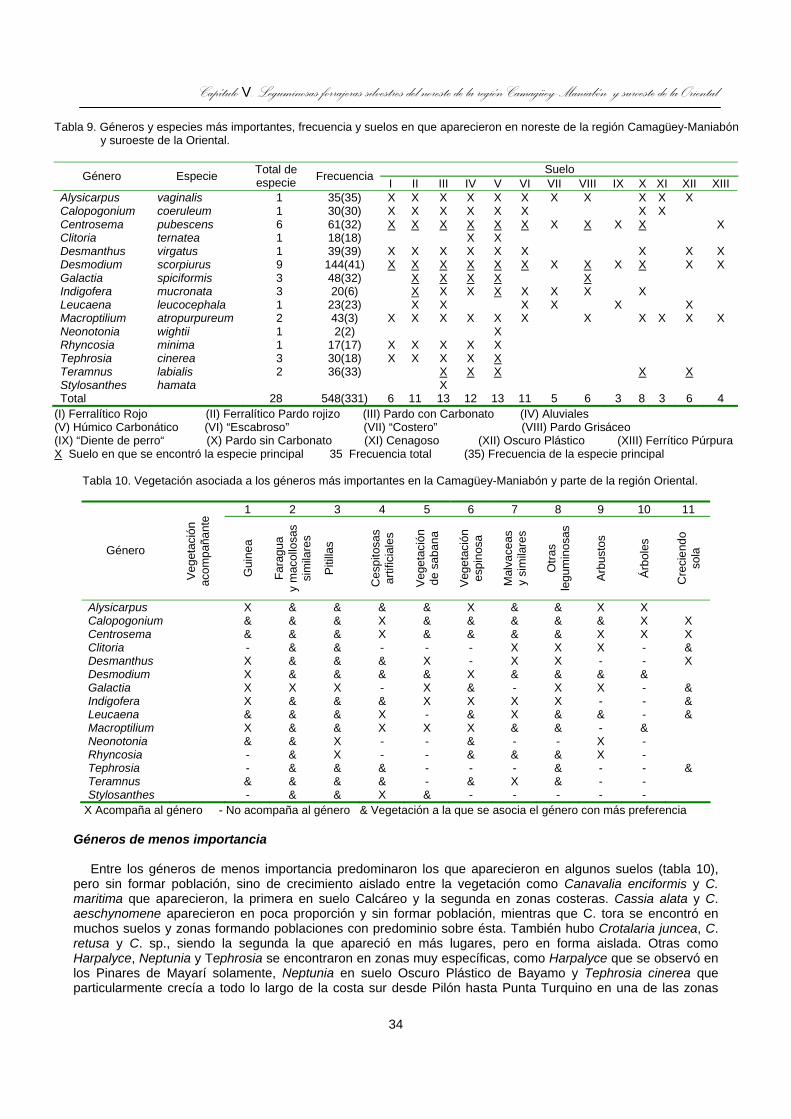
Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
34
Tabla 9. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en noreste de la región Camagüey-Maniabón y suroeste de la Oriental.
Suelo Género Especie Total de
especie Frecuencia I II III IV V VI VII VIII IX X XI XII XIII Alysicarpus vaginalis 1 35(35) X X X X X X X X X X X Calopogonium coeruleum 1 30(30) X X X X X X X X Centrosema pubescens 6 61(32) X X X X X X X X X X X Clitoria ternatea 1 18(18) X X Desmanthus virgatus 1 39(39) X X X X X X X X X Desmodium scorpiurus 9 144(41) X X X X X X X X X X X X Galactia spiciformis 3 48(32) X X X X X Indigofera mucronata 3 20(6) X X X X X X X X Leucaena leucocephala 1 23(23) X X X X X X Macroptilium atropurpureum 2 43(3) X X X X X X X X X X X Neonotonia wightii 1 2(2) X Rhyncosia minima 1 17(17) X X X X X Tephrosia cinerea 3 30(18) X X X X X Teramnus labialis 2 36(33) X X X X X Stylosanthes hamata X Total 28 548(331) 6 11 13 12 13 11 5 6 3 8 3 6 4
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) Pardo con Carbonato (IV) Aluviales (V) Húmico Carbonático (VI) “Escabroso” (VII) “Costero” (VIII) Pardo Grisáceo (IX) “Diente de perro“ (X) Pardo sin Carbonato (XI) Cenagoso (XII) Oscuro Plástico (XIII) Ferrítico Púrpura X Suelo en que se encontró la especie principal 35 Frecuencia total (35) Frecuencia de la especie principal
Tabla 10. Vegetación asociada a los géneros más importantes en la Camagüey-Maniabón y parte de la región Oriental.
1 2 3 4 5 6 7 8 9 10 11
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y m
acol
losa
s si
mila
res
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Árb
oles
Cre
cien
do
sola
Alysicarpus X & & & & X & & X X Calopogonium & & & X & & & & & X X Centrosema & & & X & & & & X X X Clitoria - & & - - - X X X - & Desmanthus X & & & X - X X - - X Desmodium X & & & & X & & & & Galactia X X X - X & - X X - & Indigofera X & & & X X X X - - & Leucaena & & & X - & X & & - & Macroptilium X & & X X X & & - & Neonotonia & & X - - & - - X - Rhyncosia - & X - - & & & X - Tephrosia - & & & - - - & - - & Teramnus & & & & - & X & - - Stylosanthes - & & X & - - - - -
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
Géneros de menos importancia
Entre los géneros de menos importancia predominaron los que aparecieron en algunos suelos (tabla 10), pero sin formar población, sino de crecimiento aislado entre la vegetación como Canavalia enciformis y C. maritima que aparecieron, la primera en suelo Calcáreo y la segunda en zonas costeras. Cassia alata y C. aeschynomene aparecieron en poca proporción y sin formar población, mientras que C. tora se encontró en muchos suelos y zonas formando poblaciones con predominio sobre ésta. También hubo Crotalaria juncea, C. retusa y C. sp., siendo la segunda la que apareció en más lugares, pero en forma aislada. Otras como Harpalyce, Neptunia y Tephrosia se encontraron en zonas muy específicas, como Harpalyce que se observó en los Pinares de Mayarí solamente, Neptunia en suelo Oscuro Plástico de Bayamo y Tephrosia cinerea que particularmente crecía a todo lo largo de la costa sur desde Pilón hasta Punta Turquino en una de las zonas

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
35
más desérticas, donde esta especie, en la mayoría de los casos, era el único constituyente de la flora y se observó ramoneada, aunque en forma ligera por los animales. Las otras dos especies se distribuyeron con más amplitud por la mayoría del área (tablas 11 y 12).
Fig. 18. Distribución de los géneros Rhynchosia, Stylosanthes y Teramnus (región Oriental).
Tabla 11. Especies menos importantes, frecuencia y suelo de Camagüey-Maniabón y parte de la región Oriental.
Suelo Género Especie Frecuencia I II III IV V VI VII VIII IX X Canavalia ensiformis 2 X Canavalia maritima 10 X Cassia alata 2 X Cassia tora 15 X X X X X X Cassia aeschynomene 8 X X X X Crotalaria juncea 3 X X Crotalaria retusa 15 X X X X X X Crotalaria sp 2 X Harpalyce angustifloria 2 X Neptunia plena 2 X Sesbania emerus 1 X Total 62 2 3 3 3 2 3 3 2 3 1
(I) Ferralítico Rojo (II) Ferralítico Pardo rojizo (III) Ferrítico Púrpura (IV) Pardo con Carbonato (V) Húmico Carbonático (VI) “Escabroso” (VII) Pardo Grisáceo (VIII) Pardo sin Carbonato (IX) Oscuro Plástico (X) “Costero”

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
36
Tabla 12. Vegetación asociada a los géneros menos importantes en Camagüey-Maniabón y parte de la región Oriental.
1 2 3 4 5 6 78 8 9 10
Género V
eget
ació
n ac
ompa
ñant
e
Gui
nea
Fara
gua
y
sim
ilare
s
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arb
usto
s
Cre
cien
do
sola
Canavalia & X & Cassia & & X & & X & Crotalaria X & & X & X X & Harpalyce & & & Neptunia & & Sesbania X &
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
Discusión
Los resultados indican que la leguminosas rastreras son las que se encontraron en mayor proporción en las comunidades vegetales de esta región, al representar el 67% del total de especies.
Entre las más abundantes estuvieron Teramnus labialis que se asoció con habilidad a las gramíneas cespitosas y persiste en los pastizales de esa zona. Este comportamiento coincide con los resultados de Menéndez et al. (1980, datos inéditos) y contrastan con los de Funes et al. (1971), quienes al someter a corte esta leguminosa comparada con Glycine en cultivo puro observaron una despoblación parcial en teramnus y buena población en Glycine seleccionada y en pastoreo tampoco teramnus persistió (en cultivo puro) por su alto consumo (Funes y Pérez, 1976). Sin embargo, se observó una buena proporción de esta leguminosa entre pangola en pastizales de más de 5 años en Birán y noreste de Holguín, donde se emplean cargas moderadas y en ocasiones ha dominado la pangola (Puig, 0., 1976, comunicación personal). Este comportamiento puede deberse a la forma en que los tallos volubles de teramnus se entretejen a los de las gramíneas, características que no permiten una defoliación a fondo (Menéndez et al., 1980, datos inéditos) y le permiten un adecuado índice de área foliar con lo que se obtiene un buen rebrote (Whiteman, 1969; Blazer, 1974). Otra condición que pudiera haber favorecido el alto porcentaje de leguminosas en la flora de esta zona es la fertilidad y pH del suelo, ya que esta especie apareció en zonas donde estas características son de media a alta, que es otra premisa para el buen desarrollo de este género (Whyte et al., 1955).
De acuerdo a los datos de León y Alain (1951) de las 150 especies de Desmodium en el mundo 15 pertenecen a nuestra flora, de las cuales fueron encontradas 9 en esta región. Estas especies según se ha observado, tienen en general características esciósfilas (Wilsie, 1970) y se presentan mayormente asociadas a gramíneas cespitosas en todos los suelos y es un género que aunque no se conocen especies de uso agrícola en Cuba, existen algunas con características muy promisorias en muchas partes de África, Hawaii y el Caribe. Este generó predominó en esta región y se encontró en todos los suelos estudiados.
Alysicarpus fue un género que apareció con una sola especie, pero tuvo un comportamiento similar que Desmodium, ya que predominó en la mayor parte de los suelos de la región y mostró un amplio rango de tolerancia que se ha evidenciado en otras especies vegetales (Cain, 1944; Mason, 1936). Esta especie también pide incluirse como esciósfila (Wilsie, 1970), ya que es incapaz de establecerse cuando es sembrada como cultivo puro (Yepes, 1973, comunicación personal).
Calopogonium coeruleum y Centrosema pubescens son especies que también mostraron un gran rango de adaptación al suelo y vegetación acompañante, aunque ambas prefirieron suelos de fertilidad de media a alta. El primero fue más abundante en las zonas montañosas que en las llanuras, mientras que el segundo fue más abundante en las zonas de mayores precipitaciones. Es característica común, en ambos, cubrir la vegetación acompañante por su buena habilidad asociativa y tendencia a cubrir la vegetación acompañante.
Otras especies coma las pertenecientes a los géneros Clitoria, Stylosanthes, Tephrosia y Neonotonia tuvieron un rango de tolerancia estrecho (Cain, 1944; Manson, 1936) al parecer en condiciones muy específicas. Clitoria estaba presente en suelo Aluvial fértil, condiciones similares en la que se ha informado creciendo naturalmente en Oriente, Las Villas y La Habana (León y Alain, 1951). Ha sido estudiada en cultivo puro y asociada con otras leguminosas y gramíneas cespitosas y macollosas y se han encontrado algunas variedades con alguna superioridad Funes et al. (1971); Valdés y Pérez (1971); fue recomendada para asociar con gramíneas forrajeras de corte como mejoradotas de su valor nutritivo (Menéndez y Martínez, 1980).

Capítulo V. Leguminosas forrajeras silvestres del noreste de la región Camagüey-Maniabón y suroeste de la Oriental
37
Stylosanthes apareció en un solo lugar y poco vigoroso entre gramíneas cespitosas en suelo poco fértil, lo que pudiera ser la causa principal por la que esta especie ha estado ausente en la mayoría de los suelos de la región, ya que ha sido encontrada únicamente en suelo franco. Además Torcell, Ive y Cunningham (1976) informaron que se podía esperar dominancia en asociaciones de esta leguminosa con pangola en suelos de alta fertilidad, mientras los fértiles son arcillosos o pesados. El caso de Tephrosia cinerea resultó interesante porque en la zona donde apareció fue prácticamente la única especie de interés encontrada y mostró adaptarse mejor que otras a las condiciones casi desérticas de esa franja costera, donde abundaba el cactus y no crecía el buffel que es una especie que se adapta a condiciones de precipitación relativamente baja (Humphreys, 1967), por lo que no se descarta la influencia de otro factor edáfico, climático o fisiológico que gobierne la distribución de esta especie leguminosa. Neonotonia apareció solamente en suelo Húmico Carbonático que son los más fuertes de esta región. Esto coincide con los criterios de Whyte et al. (1955) y McIlroy (1976) acerca de la necesidad de suelos suficientemente fértiles para esta especie.
Leucaena mostró en esta región una gran plasticidad al ser encontrada tanto en suelos francos como pesados, fértiles y poco fértiles en los cuales de mostró ser agresiva, características que concuerdan con Eavis, Cumberbath y Medford (1974) y Cooksley (1974), quienes plantean que su capacidad de adaptación le permite crecer y producir en las más diversas condiciones edafoclimáticas. Partridge y Ranacov (1974) informan que es la única leguminosa tropical capaz de soportar largos períodos de encharcamiento. Sin embargo, no fue encontrada en altitudes mayores de 200 m que coincide con lo informado por Whyte et al. (1955) sobre esta especie.
Estas características la ubican como una planta de perspectivas para esta zona porque además posee un alto valor biológico (Upadilla, Rekiby y Pathak, 1974) con altos porcentajes de digestibilidad y contenido de proteína bruta en sus hojas que oscilan entre 18,7 a 30% (Falvey, 1976) y se ha recomendado para la producción de carne en el trópico (Falvey, 1976; Blunt, 1976).
Galactia spiciformis e Indigofera mucronata aparecieron en suelos Fersialítico Pardo rojizo, Pardo con Carbonato, Aluviales y Húmico carbonático que son suelos fértiles e indica la preferencia de estas especies por estas condiciones edáficas. Sin embargo, Galactia aunque tuvo una frecuencia aceptable apareció poco vigoroso debido al ataque de hongos en todos los ambientes en que estuvo presente, lo que puede limitar su uso a gran escala. Esto no ocurrió con Indigofera que se mostró vigorosa y libre de plagas y enfermedades, aunque su frecuencia fue generalmente baja.
Macroptilium apareció en un gran número de suelos, pero la especie de mayor importancia que es M. atropurpureum fue específica de los suelos Fersialítico Púrpura de los Pinares de Mayarí, donde se presentó muy vigoroso, incluso cubriendo pinos. Este hallazgo está de acuerdo con lo informado por Whyte et al. (1955) y Humphreys (1974) con relación a la especificidad de esta especie a este tipo de suelo.
Contrariamente a las especies anteriores las no importantes aparecieron con una frecuencia de distribución muy baja además de estar presente en muy pocos suelos y entre muy poca vegetación (tablas 11 y 12) que indica su poca adaptación al medio, ya que ninguna se observó consumida por animales y presentan, en general, una estructura desfavorable para subsistir en áreas pratenses.
Los resultados de este trabajo evidencian la posibilidad que tienen leguminosas como Teramnus labialis en suelos Pardos, Calopogonium y Centrosema en los Pardos y escabrosos de fertilidad voluble y leucaena en diversas condiciones.
Además, otras que aunque han aparecido muy localizadas deben ser protegidas, aún más si tenemos en cuenta que en esta zona existen las más bajas cargas de animales por área y hay poca disponibilidad de fertilizantes nitrogenados para elevarlas. De aquí que se abre un amplio campo para el estudio y explotación de las especies de leguminosas, donde debe tenerse muy en cuenta la influencia de los factores edafoclimáticos en la distribución de este grupo de plantas para hacer más económica la empresa.

Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
38
CAPÍTULO VI. LEGUMINOSAS FORRAJERAS SILVESTRES DE LA REGIÓN ORIENTAL DE CUBA Materiales y métodos
Localización. Esta región se ubica en el extremo noroeste de Cuba, pero el trabajo conducido por nosotros
abarcó el este de Holguín y las provincias de Santiago de Cuba y Guantánamo (fig. 1). Clima. Las características climáticas de esta región son muy contrastantes, ya que tiene las mayores
precipitaciones hacia el norte, noreste y suroeste en las mayores elevaciones, mientras que en la costa sur y en las cuencas de Santiago y Guantánamo y en Punta de Maisí ocurren las precipitaciones más bajas (figs. 2 y 3) al igual que los demás elementos climáticos (fig. 4). La distribución de las lluvias (que difiere de las del resto del país en la costa norte). Presenta un acumulado de temperatura (+10°C) superior a los 9 500°C por el norte desde Punta de Maisí hasta Puerto Padre y entre 9 000 y 9 500°C en el resto de la región.
Suelos. Esta región posee los suelos de más difícil laboreo con predominio de los escabrosos y pocas áreas de Pardos con Carbonato y Oscuros Plásticos (fig. 15), los suelos son en general de topografía alomada y muy propensos a la erosión.
Resultados Generalidades
Fueron encontrados 19 géneros en los que se diferenciaron 42 especies, producto de 53 muestras colectadas de 685 muestreos. Más del 50% del total de los géneros corresponden los más importantes para el trópico. Dentro de estos se observaron como promisorios Teramnus con 3 especies con T. labialis como más destacado; Centrosema con 5 especies sobresaliendo C. pubescens entre los demás y Calopogonium, con 2 especies donde C. coeruleum se destacó sobre la otra, ambas por su amplia distribución, vigor y habilidad con que aparecieron asociadas a las gramíneas macollosas y cespitosas y otras plantas de la comunidad.
Sin embargo, los géneros que consideramos menos importantes para el trópico, se encontraron muy aislados con pocos ejemplares que no sobresalían entre la vegetación acompañante y aparecieron en muy pocos suelos, fundamentalmente, en los calcáreos. Esto contrasta con el comportamiento de los géneros más importantes que con la excepción de Stylosanthes se mostraron en la mayoría de las condiciones edáficas.
Descripción de géneros principales
Todos estos géneros se observaron creciendo asociados con especies del mismo género o por otros de
leguminosas, pero principalmente junto a gramíneas, entre las que abundaban Dichanthium spp.; Bothriocloa pertusa; Cenchrus ciliaris y en menor proporción Paspalum notatum y Panicum maximum, así como arbustos y la vegetación propia de cada suelo.
Alysicarpus. A. vaginalis se recolectó y observó dentro del follaje de paraná (Brachiaria mutica), pitilla y W. americana alrededor de los 70 km de Santiago de Cuba (oeste); Rhychonsia minima, Desmodium spp., guinea y faragua en los suelos de Mayarí arriba y en la costa sur de Santiago de Cuba junto a pitillas. Su estado fenológico fue el de semilla madura, con muchas legumbres secas y pocas hojas y se mostró libre de plagas y enfermedades.
Calopogonium. Se encontraron las especies C. coeruleum y C. muconoides y se destacó particularmente C. coeruleum, el cual sobresalió de la vegetación acompañante, entre pitillas, alpargatas, guinea, arbustos y matorrales desde Songo hasta Guantánamo, alrededor de 10km al oeste de Santiago de Cuba y alrededor de los 18 km al norte Guantánamo, en suelos Pardos y escabrosos, así como en las montañas Moa-Baracoa. C. muconoides apareció a 40 km al norte de Guantánamo sobre el mismo tipo de suelo y similar vegetación. El estado fenológico predominante en la primera floración fue el de floración plena con semillas verdes y maduras con presencia de muchas hojas verdes, mientras que el segundo se observó en estado senil avanzado. Ambas especies se mostraron libres de plagas y enfermedades.
Centrosema. Se encontraron cinco especies: C. lobatum, C. plumieri, C. pubescens, C. sp. y C. virginianum. Se destacó particularmente C. pubescens que apareció asociada, además de las especies comunes (vegetación de sabana y pitillas) a pangola, caña de azúcar (S. Officinarum), entre cafeto (C. arabica) y los arbustos de los bosquecillos y potreros, en otras áreas de cultivo y cercado de los mismos en las zonas de Guantánamo, Santiago y en la costa sur hasta 90 km al oeste, pero predominó entre los 30 y 50 km al noroeste de Guantánamo y a 10 km al oeste. Las otras especies se encontraron en áreas marginales no sometidas a corte ni a pastoreo, por ejemplo, C. plumieri apareció junto a C. pubescens, entre guinea y otras gramíneas al noroeste de Guantánamo, C. lobatum se encontró entre los cañaverales y en las guardarrayas o calles de los campos cañeros en esta misma zona. C. virginianum solo apareció en las maniguas y zonas abruptas y
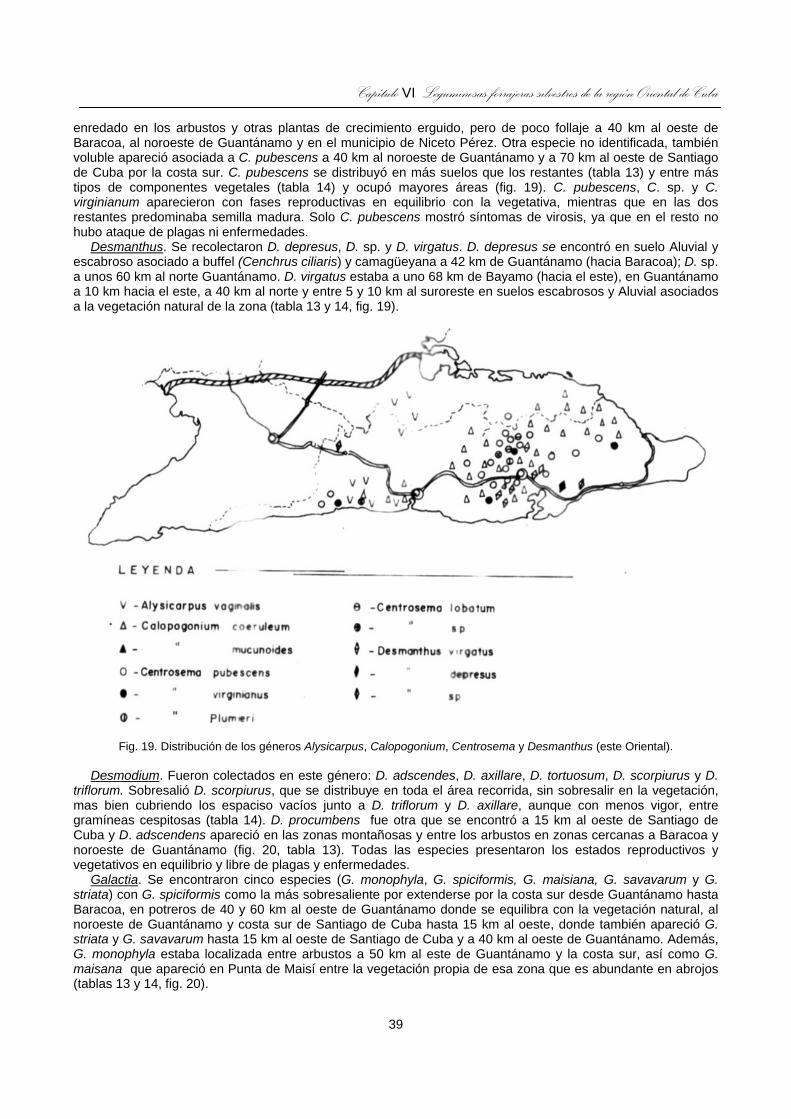
Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
39
enredado en los arbustos y otras plantas de crecimiento erguido, pero de poco follaje a 40 km al oeste de Baracoa, al noroeste de Guantánamo y en el municipio de Niceto Pérez. Otra especie no identificada, también voluble apareció asociada a C. pubescens a 40 km al noroeste de Guantánamo y a 70 km al oeste de Santiago de Cuba por la costa sur. C. pubescens se distribuyó en más suelos que los restantes (tabla 13) y entre más tipos de componentes vegetales (tabla 14) y ocupó mayores áreas (fig. 19). C. pubescens, C. sp. y C. virginianum aparecieron con fases reproductivas en equilibrio con la vegetativa, mientras que en las dos restantes predominaba semilla madura. Solo C. pubescens mostró síntomas de virosis, ya que en el resto no hubo ataque de plagas ni enfermedades.
Desmanthus. Se recolectaron D. depresus, D. sp. y D. virgatus. D. depresus se encontró en suelo Aluvial y escabroso asociado a buffel (Cenchrus ciliaris) y camagüeyana a 42 km de Guantánamo (hacia Baracoa); D. sp. a unos 60 km al norte Guantánamo. D. virgatus estaba a uno 68 km de Bayamo (hacia el este), en Guantánamo a 10 km hacia el este, a 40 km al norte y entre 5 y 10 km al suroreste en suelos escabrosos y Aluvial asociados a la vegetación natural de la zona (tabla 13 y 14, fig. 19).
Fig. 19. Distribución de los géneros Alysicarpus, Calopogonium, Centrosema y Desmanthus (este Oriental). Desmodium. Fueron colectados en este género: D. adscendes, D. axillare, D. tortuosum, D. scorpiurus y D.
triflorum. Sobresalió D. scorpiurus, que se distribuye en toda el área recorrida, sin sobresalir en la vegetación, mas bien cubriendo los espaciso vacíos junto a D. triflorum y D. axillare, aunque con menos vigor, entre gramíneas cespitosas (tabla 14). D. procumbens fue otra que se encontró a 15 km al oeste de Santiago de Cuba y D. adscendens apareció en las zonas montañosas y entre los arbustos en zonas cercanas a Baracoa y noroeste de Guantánamo (fig. 20, tabla 13). Todas las especies presentaron los estados reproductivos y vegetativos en equilibrio y libre de plagas y enfermedades.
Galactia. Se encontraron cinco especies (G. monophyla, G. spiciformis, G. maisiana, G. savavarum y G. striata) con G. spiciformis como la más sobresaliente por extenderse por la costa sur desde Guantánamo hasta Baracoa, en potreros de 40 y 60 km al oeste de Guantánamo donde se equilibra con la vegetación natural, al noroeste de Guantánamo y costa sur de Santiago de Cuba hasta 15 km al oeste, donde también apareció G. striata y G. savavarum hasta 15 km al oeste de Santiago de Cuba y a 40 km al oeste de Guantánamo. Además, G. monophyla estaba localizada entre arbustos a 50 km al este de Guantánamo y la costa sur, así como G. maisana que apareció en Punta de Maisí entre la vegetación propia de esa zona que es abundante en abrojos (tablas 13 y 14, fig. 20).

Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
40
Indigofera. Fueron halladas I. mucronata, I. sumatrana e I. tictoria. La primera abundante en los alrededores de Guantánamo hacia el norte y oeste, fundamentalmente, a orillas de las cercas. I. tictoria se encontró abundante a todo lo largo de la costa sur, desde Punta Turquino hasta Santiago de Cuba, pero sin formar una población compacta y sin acompañante, mientras que I. sumatrana se localizó a 11 km de Contramaestre, creciendo entre todo tipo de vegetación y formando población en esta zona. En todas las especies se armonizaban las fases reproductivas con la vegetativa, apareciendo las legumbres de I. sumatrana perforadas por insecto y las demás libres de plagas y enfermedades.
Leucaena. Se encontró L. leucocephala en suelo costero de Santiago de Cuba hacia el este y a 10 y 15 km de Guantánamo y también al este, creciendo en todo tipo de vegetación (tabla 13 y 14, fig. 20).
Esta especie se observó libre de plagas y enfermedades y todas las muestras tenían pocas hojas y las legumbres secas.
Macroptilium. Fueron encontradas las especies M. lathyroides y posiblemente M. longipedunculatum, ya que durante tres años no pudo observarse la floración de esta que apareció en zonas de suelo rojo (Ferralítico) desde Mar Verde hasta Chivirico en la costa sur de Santiago de Cuba entre pitillas y malváceas (fig. 20). La otra especie estaba presente en diferentes zonas y suelos, pero sin formar población ni sobresalir entre la vegetación. Ambas especies no mostraron daños de plagas ni enfermedades y se observó a M. lathyroides con las fases reproductiva y vegetativa en equilibrio, mientras que la otra especie estaba en estado vegetativo.
Fig. 20. Distribución de los géneros Desmodium, Galactia, Indigofera, Leucaena y Macroptilium (este Oriental). Pueraria. P. phaseoloides se presentó en suelo arenoso y entre pitillas y buffel en Punta de Maisí en
equilibrio estas, en estado vegetativo y algunas legumbres secas y sin síntomas de haber sido dañada por plagas o enfermedades.
Rhynchosia. Se encontró R. minima, asociada a pangola en un área forrajera de Mayarí Arriba, con mucho vigor y abundante en suelo Pardo (tablas 13 y 14), a 30 km al noroeste de Guantánamo en el mismo tipo de suelo y asociada a pitillas y W. americana donde presentó aspecto clorótico y poco vigor. Apareció también y con el mismo aspecto en Punta Turquino asociado con camagüeyana (fig. 21). Su estado fenológico fue el de fructificación.
Stylosanthes. S. hamata apareció formando poblaciones en equilibrio con camagüeyana y buffel desde Tortuguilla hasta Cajobabo (costa sur al este de Guantánamo). La vegetación principal de esta zona es casi desértica, además se halló en la costa sur de Santiago de Cuba hacia el este a 10 km asociado a guinea y otras no gramíneas (tabla 14), formando una población poco densa y en Punta de Maisí junto a camagüeyana y buffel sin predominar. En todos los lugares se presentó en estado vegetativo y floración plena con algunas legumbres maduras y sin presentar sintomatología patológica.
Tephrosia. Aparecieron dos especies: T. cinerea, que se encontró en las mismas zonas que I. tictoria, vigorosa y creciendo sin acompañante o entre I. tictoria, pero sin formar una población densa. En algunos lugares es la única vegetación existente, y era ramoneada por el ganado vacuno. La otra especie fue T. senna encontrada cerca de Cajobabo, creciendo junto a buffel a una altura no mayor de 15 cm sin sobresalir por
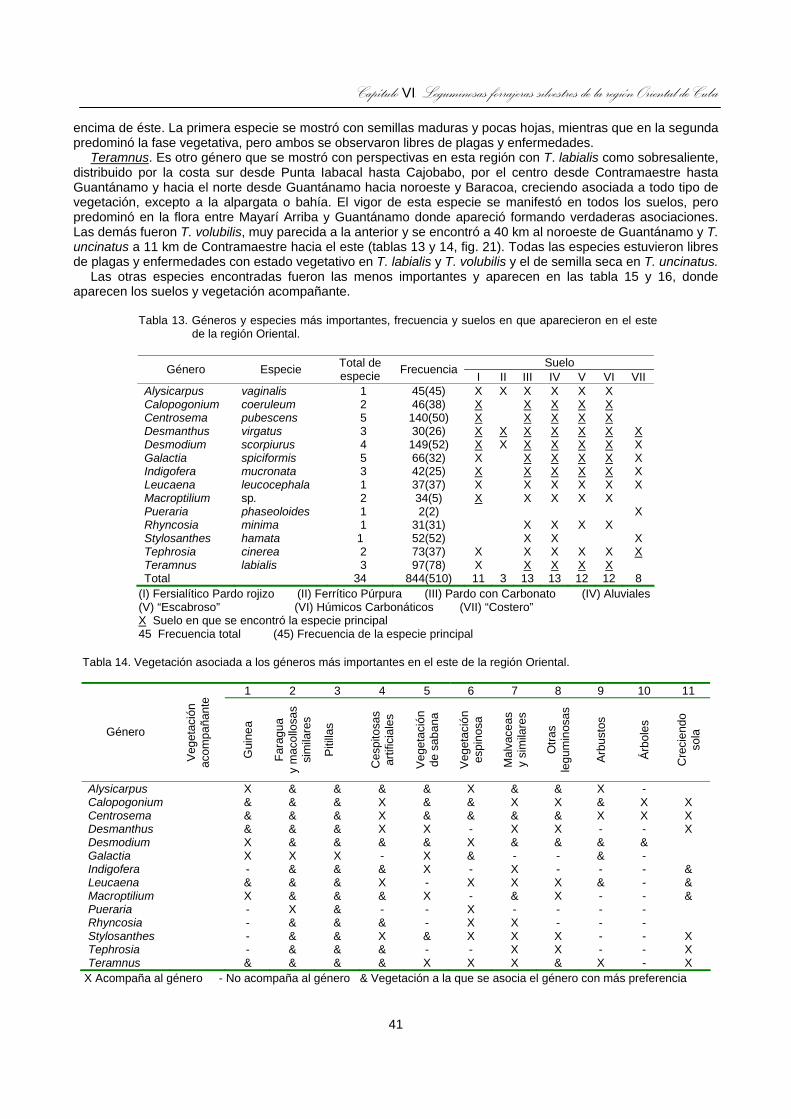
Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
41
encima de éste. La primera especie se mostró con semillas maduras y pocas hojas, mientras que en la segunda predominó la fase vegetativa, pero ambos se observaron libres de plagas y enfermedades.
Teramnus. Es otro género que se mostró con perspectivas en esta región con T. labialis como sobresaliente, distribuido por la costa sur desde Punta Iabacal hasta Cajobabo, por el centro desde Contramaestre hasta Guantánamo y hacia el norte desde Guantánamo hacia noroeste y Baracoa, creciendo asociada a todo tipo de vegetación, excepto a la alpargata o bahía. El vigor de esta especie se manifestó en todos los suelos, pero predominó en la flora entre Mayarí Arriba y Guantánamo donde apareció formando verdaderas asociaciones. Las demás fueron T. volubilis, muy parecida a la anterior y se encontró a 40 km al noroeste de Guantánamo y T. uncinatus a 11 km de Contramaestre hacia el este (tablas 13 y 14, fig. 21). Todas las especies estuvieron libres de plagas y enfermedades con estado vegetativo en T. labialis y T. volubilis y el de semilla seca en T. uncinatus.
Las otras especies encontradas fueron las menos importantes y aparecen en las tabla 15 y 16, donde aparecen los suelos y vegetación acompañante.
Tabla 13. Géneros y especies más importantes, frecuencia y suelos en que aparecieron en el este
de la región Oriental.
Suelo Género Especie Total de especie Frecuencia I II III IV V VI VII
Alysicarpus vaginalis 1 45(45) X X X X X X Calopogonium coeruleum 2 46(38) X X X X X Centrosema pubescens 5 140(50) X X X X X Desmanthus virgatus 3 30(26) X X X X X X X Desmodium scorpiurus 4 149(52) X X X X X X X Galactia spiciformis 5 66(32) X X X X X X Indigofera mucronata 3 42(25) X X X X X X Leucaena leucocephala 1 37(37) X X X X X X Macroptilium sp. 2 34(5) X X X X X Pueraria phaseoloides 1 2(2) X Rhyncosia minima 1 31(31) X X X X Stylosanthes hamata 1 52(52) X X X Tephrosia cinerea 2 73(37) X X X X X X Teramnus labialis 3 97(78) X X X X X Total 34 844(510) 11 3 13 13 12 12 8
(I) Fersialítico Pardo rojizo (II) Ferrítico Púrpura (III) Pardo con Carbonato (IV) Aluviales (V) “Escabroso” (VI) Húmicos Carbonáticos (VII) “Costero” X Suelo en que se encontró la especie principal 45 Frecuencia total (45) Frecuencia de la especie principal
Tabla 14. Vegetación asociada a los géneros más importantes en el este de la región Oriental.
1 2 3 4 5 6 7 8 9 10 11
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y
mac
ollo
sas
sim
ilare
s
Pitil
las
Ces
pito
sas
artif
icia
les
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Arbu
stos
Árb
oles
Cre
cien
do
sola
Alysicarpus X & & & & X & & X - Calopogonium & & & X & & X X & X X Centrosema & & & X & & & & X X X Desmanthus & & & X X - X X - - X Desmodium X & & & & X & & & & Galactia X X X - X & - - & - Indigofera - & & & X - X - - - & Leucaena & & & X - X X X & - & Macroptilium X & & & X - & X - - & Pueraria - X & - - X - - - - Rhyncosia - & & & - X X - - - Stylosanthes - & & X & X X X - - X Tephrosia - & & & - - X X - - X Teramnus & & & & X X X & X - X
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia
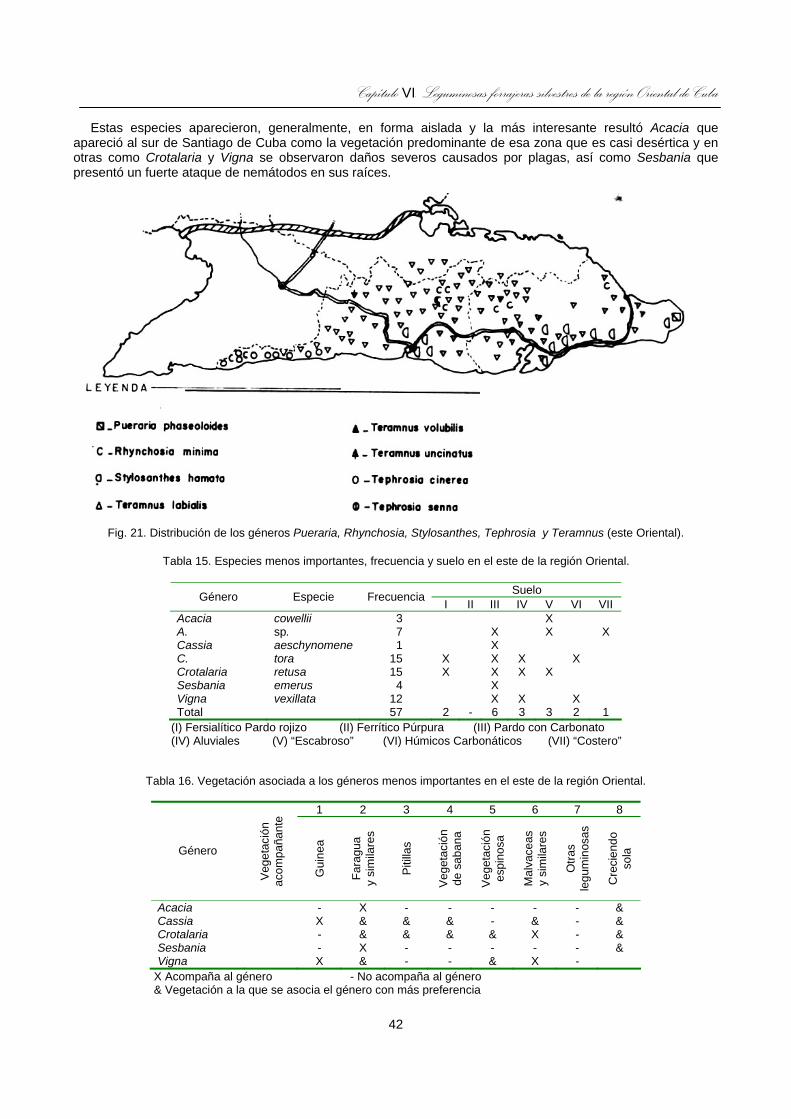
Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
42
Estas especies aparecieron, generalmente, en forma aislada y la más interesante resultó Acacia que apareció al sur de Santiago de Cuba como la vegetación predominante de esa zona que es casi desértica y en otras como Crotalaria y Vigna se observaron daños severos causados por plagas, así como Sesbania que presentó un fuerte ataque de nemátodos en sus raíces.
Fig. 21. Distribución de los géneros Pueraria, Rhynchosia, Stylosanthes, Tephrosia y Teramnus (este Oriental).
Tabla 15. Especies menos importantes, frecuencia y suelo en el este de la región Oriental.
Suelo Género Especie Frecuencia I II III IV V VI VII Acacia cowellii 3 X A. sp. 7 X X X Cassia aeschynomene 1 X C. tora 15 X X X X Crotalaria retusa 15 X X X X Sesbania emerus 4 X Vigna vexillata 12 X X X Total 57 2 - 6 3 3 2 1
(I) Fersialítico Pardo rojizo (II) Ferrítico Púrpura (III) Pardo con Carbonato (IV) Aluviales (V) “Escabroso” (VI) Húmicos Carbonáticos (VII) “Costero”
Tabla 16. Vegetación asociada a los géneros menos importantes en el este de la región Oriental.
1 2 3 4 5 6 7 8
Género
Veg
etac
ión
acom
paña
nte
Gui
nea
Fara
gua
y si
mila
res
Pitil
las
Veg
etac
ión
de s
aban
a
Veg
etac
ión
espi
nosa
Mal
vace
as
y si
mila
res
Otra
s le
gum
inos
as
Cre
cien
do
sola
Acacia - X - - - - - & Cassia X & & & - & - & Crotalaria - & & & & X - & Sesbania - X - - - - - & Vigna X & - - & X -
X Acompaña al género - No acompaña al género & Vegetación a la que se asocia el género con más preferencia

Capítulo VI. Leguminosas forrajeras silvestres de la región Oriental de Cuba
43
Discusión
Con los resultados de este trabajo se completan en lo fundamental, el estudio de la distribución y ubicación de las leguminosas forrajeras cubanas. En toda esta región se puso de manifestó la mayor proporción (63%) de especies de hábito de crecimiento postrado y voluble que fueron las más abundantes y predominaron entre la flora de la región. Se destacaron Teramnus labialis, Desmodium sp., Calopogonium coeruleum y Centrosema pubescens, aunque sobresalieron también en determinadas zonas, las de crecimiento erecto y decumbente, Leucaena, Indigofera, Stylosanthes y Tephrosia, lo que evidencia la adaptación de estas leguminosas a condiciones determinadas que han sido asociadas por Mason (1936) y Cain (1944). Uno de los géneros de mayor interés fue Teramnus con la especie Teramnus labialis, que apareció muy abundante en los suelos Pardos con Carbonato y escabrosos de Guantánamo y Santiago de Cuba y una parte de Holguín, fundamentalmente, entre gramíneas cespitosas, pero no apareció entre los árboles de las zonas montañosas ni en lugares de precipitaciones extremadamente bajas como Punta de Maisí y suroeste de Santiago de Cuba. Sin embargo, no se manifestó afectada por la altitud, ya que se distribuyó hasta 800 m, pero no se encontró en los suelos Fersialíticos de Holguín que se conocen por su baja fertilidad, aunque tampoco aparecieron otras leguminosas de interés en esos suelos (figs. 19 y 21), lo que evidencia que las características edáficas pueden limitar en alto grado la presencia de esta familia en condiciones naturales.
En zonas montañosas predominó Calopogonium, mientras que Centrosema apareció, generalmente, en las de mayor precipitación, lo que confirma lo observado en las demás regiones y lo planteado para estas especies por Whyte et al. (1955) y Humphreys (1974) referente a la necesidad de mediana a alta fertilidad para ambas especies, aunque este último autor informó que Centrosema se puede adaptar también a suelos ácidos, pero que responden bien a las aplicaciones de fósforo.
Desmodium, aunque apareció con un menor número de especies que en las demás regiones, se distribuyó en todos los suelos y altitudes, mayormente asociado a gramíneas cespitosas. Este comportamiento pone de manifiesto lo planteado por Good (1931; 1953). Sin embargo, Alysicarpus apareció solo en suelos fértiles situados a altitudes inferiores a los 100 m, lo que pudiera ser un factor que influya en la distribución de esta especie.
También puede ser una evidencia de la influencia del hombre en la modificación de la flora, ya que en Punta de Maisí existe una vegetación que no parece haber sido alterada en muchos años y aparecieron leguminosas como Galactia maisana y Stylosanthes hamata que son unas de las pocas leguminosas informadas en la flora de esta localidad (León y Alain, 1951). Se evidenció nuevamente la preferencia de algunas especies como Stylosanthes hamata, Tephrosia cinerea e Indigofera tictoria que aparecieron en suelos francos y salinos, el primero desde Santiago de Cuba hasta Cajobabo en la costa sur, lugar en que mostró su habilidad para asociarse a buffel, desde la costa hasta los pastoreos adyacentes, lo que sitúa a esta especie con perspectivas para esa zona, a la vez que confirma la resistencia a la sequía informada para este género (Humphreys, 1974). Tephrosia e Indigofera se encontraron en la costa sur al oeste de Santiago de Cuba en una de las zonas más desérticas y fue común verlas reemplazándose una a otra, pues cuando estaban juntas fue en las zonas de transición donde una especie sucedía a la otra. Sin embargo, otras especies de ambos géneros (T. senna e I. mucronata) se comportaron como mesofitas, ya que aparecieron en suelos de mediana fertilidad y con régimen de lluvia medio. Leucaena se distribuyó, fundamentalmente, en zonas costeras con suelos francos, donde manifestó ser una planta agresiva al sobresalir entre la vegetación acompañante, aunque no se encontró entre los potreros (muy pobres en vegetación) posiblemente porque no soportaba el consumo animal o por haber sido eliminada por el hombre.
Otros géneros como Galactia y Rhyncosia corren el riesgo de desaparecer por la forma en que encontraron, ya que Galactia apareció afectada por ataques severos de hongos y Rhyncosia por ataque viral; además, ninguna de estas especies presentan tallos que enraizan en contacto con el suelo y se observaron generalmente poco vigorosas.
Se observó que en esta zona las leguminosas protegen el suelo contra la erosión y la pérdida de humedad que son dos factores importantes a tener en cuenta por la topografía accidentada y las zonas de bajo régimen de lluvia en esta región.
Jewis (1972) plantea que las leguminosas poseen estas características porque presentan un follaje más compacto que regula la temperatura del suelo.
Las características antes señaladas no fueron observadas en las especies menos importantes, ya que su distribución fue escasa, y generalmente, poseen una estructura desfavorable para persistir en áreas de cultivo.
Hay evidencias para recomendar para esta región como posibles forrajeras a Teramnus labialis, Calopogonium coeruleum, Centrosema pubescens y Leucaena leucocephala, leguminosas que deben ser protegidas y estudiadas comparativamente con cultivares promisorios afines. Si esta idea se aplica a la práctica no pasará mucho tiempo sin conocer los beneficios que este grupo de plantas pueden brindar a la ganadería cubana en esta región.

Conclusiones
44
CONCLUSIONES
Se recolectaron en total 30 géneros que contenían 76 especies y de ellos se destacaron como importantes 18 géneros (Teramnus, Macroptilium, Leucaena, Centrosema, Calopogonium, Neonotonia, Stylosanthes, Alysicarpus, Desmodium, Indigofera, Pueraria, Zornia, Galactia, Lysiloma, Rhynchosia, Eriosema, Desmanthus y Tephrosia) que agruparon a 53 especies; mientras que los menos importantes fueron 12 géneros (Abrus, Aeschynomene, Albizia, Caesalpinia, Cajanus, Canavalia, Crotalaria, Cassia, Mucuna, Phaseolus, Sesbania y Vigna donde se incluyen 23 especies.
De los géneros importantes se observaron como prometedores para la explotación pecuaria Teramnus, Macroptilium, Leucaena, Centrosema y Calopogonium, los cuales estaban presentes en numerosas áreas de pastoreo (excepto la leucaena), se distribuyeron en amplias zonas y aparecieron asociados a un buen número de otras especies y formaron asociaciones equilibradas en áreas de pastoreo, lo que demostró su adaptación a esas condiciones, así como su persistencia ante el pastoreo.
El tipo de suelo y el clima influyeron marcadamente en la distribución de la mayoría de las especies de leguminosas en todo el territorio cubano por lo que muchas leguminosas de difundieron ampliamente en determinada región, mientras que en otras aparecieron pocos ejemplares o no estuvieron presentes.
Fue evidente también la influencia del manejo en la presencia de leguminosas en la flora, ya que géneros valiosos no estuvieron presentes o aparecieron muy pocos ejemplares en amplias zonas de La Habana donde la carga animal es alta y en otras provincias donde el cultivo es intensivo y se hacen aplicaciones periódicas de herbicidas, como son los macizos cañeros de las regiones de Occidente, Central y Oeste de Camagüey-Maniabón, áreas de frutales, viandas y vegetales entre las más notables.
Se evidenció la preferencia de la mayoría de las leguminosas para convivir en asociación con gramíneas, mayormente cespitosas, y a otras especies que además de servirles de tutores las protegen de ser consumidas completamente por los animales, siendo este ecosistema el más común en las zonas de suelo de fertilidad más baja que son en los que se asientan la mayoría de los planes ganaderos del país.
En cuanto a la distribución de los géneros más importantes se comprobó que: Teramnus labialis se distribuyó abundantemente en los suelos escabrosos y Pardos carbonatados de la
región Oriental y la provincia de Holguín en el sureste de Camagüey-Maniabón, donde esta leguminosa apareció asociada, fundamentalmente, a gramíneas cespitosas en áreas de pastoreo, donde persistía con cargas moderadas, así como en otras marginales, mientras que en las demás regiones aunque estuvo presente no se destacó.
Macroptilium atropurpurem se distribuyó y sobresalió en lo suelos Ferralíticos y Pardos Rojizos, fundamentalmente, en la Región Central desde Sancti Spíritus hasta Villa Clara y en la provincia de Matanzas (región Occidental), donde apareció asociado a gramíneas cespitosas y macollosas en áreas de pastoreo y marginales.
Calopogonium coerulem apareció en las 5 regiones del país, no estando presente en los suelos Oscuros Plásticos (arcillosos negros) de ninguna región, ni en los de acidez extrema ni cenagosos. En la región Oriental apareció, fundamentalmente, en las zonas montañosas, mientras que en Camagüey-Maniabón no apareció en los suelos Pardos Grisáceos (de acidez y fertilidad baja) del Este, pero en el área restante de esta región estuvo presente en pastizales de guinea y pasto natural con baja carga pero en las regiones restantes se encontró en áreas marginales constituidas por suelos francos de mediana a baja fertilidad y pH ligeramente ácido.
Centrosema pubescens se distribuyó en todas las regiones donde la precipitación fue superior a la media nacional, independientemente del tipo de suelo y sobresalió más en las áreas marginales que en los pastizales. Leucaena leucocephala apareció en zonas costeras de suelo franco y "Diente de Perro" bien drenados y en otras áreas en suelos alcalinos, pero no apareció en altitudes mayor de 200 m.
Otras especies tuvieron un rango de adaptación más estrecho, porque aparecieron en condiciones muy específicas. Entre éstas se sitúan, Stylosanthes hamata que fue encontrado en suelos franco salino y de pH alto en Guantánamo, Varadero y Santa María del Mar, donde la precipitación es baja.
En condiciones similares en zonas costeras del sur de Santiago de Cuba aparecieron Tephrosia cinerea e Indigofera tictorea y en esta misma región en la ensenada de Mora apareció en suelo Aluvial Clitoria ternatea donde mostró una gran variabilidad con ejemplares con morfología, flores y semillas diferentes. Zornia y Eriosema fueron especies que prefirieron suelos francos con acidez y pH extremadamente bajo y Pueraria que apareció en suelos franco y fertilidad y pH bajo, pero superior que en los que se presentaron las dos anteriores. Lysiloma se encontró en los suelos de mal drenaje de la mayoría de las regiones, mientras que Galactia, Rhynchosia y las restantes especies de Indigofera se encontraron en suelos diversos de fertilidad y pH intermedio.
Sin embargo, las especies que mostraron mayor rango de adaptación fueron Desmodium que apareció en todas las condiciones y Alysicarpus, aunque este último no se encontró en altitudes mayores que 100 m, ni

Conclusiones
45
entre la vegetación que proyectaban mucha sombra, como son los pastizales de guinea y arboledas o bosques, así como tampoco en suelos muy ácidos.
Es indudable que en las zonas donde se desarrollan en forma natural y durante muchos años especies forrajeras de leguminosas hay un equilibrio suelo-planta-bacteria y por tanto es factible explotar en esas condiciones a esas especies o a cultivares afines con un gasto mínimo de recursos.

Recomendaciones
46
RECOMENDACIONES 1. Los resultados de este trabajo deben ser tomados como base para el futuro desarrollo de las leguminosas
forrajeras en las zonas ganaderas del país, para lo cual debe darse continuidad el mismo evaluando a las especies más importantes (Calopogonium coeruleum, Centrosema pubescens, Leucaena leucocephala, Macroptilium atropurpureum y Teramnus labialis) in situ mediante experimentos de regionalización donde se comparen estas especies con cultivares que se recomiendan comercialmente.
2. Dirigir los estudios con cada especie de acuerdo a su predominio porque región, suelo y zona, de modo que quedaría de la forma siguiente:
a) Calopogonium coeruleum, en cualquier región, excepto en los suelos de acidez extrema, fertilidad baja y arcillosos pesados.
b) Centrosema pubescens, en cualquier región donde la precipitación promedio anual sea superior a 1 300 mm.
c) Macroptilium atropurpureum, en las regiones de Occidente, Central e Isla de Pinos con suelos Ferralíticos Cuarcíticos y Ferralítico en todas sus fases.
d) Leucaena leucocephala, en cualquier región de suelos Calcáreos de buen drenaje y que estén situados en altitudes inferiores a los 200 m.
e) Teramnus labialis, en suelos Pardos, Oscuros Plásticos, escabrosos, Húmicos Carbonáticos y Aluviales, cuyo pH sea superior a 6 y de fertilidad media de las regiones Oriental y sureste de Camagüey-Maniabón, aunque pudieran efectuarse trabajos en los suelos Ferralítico Rojo de las demás regiones.
3. Para las restantes especies de rango de adaptación más estrecho y de condiciones extremas debe trazarse un programa donde se incluyan cvs foráneos que son promisorios para esas condiciones y proteger las demás áreas pecuarias evitando el empleo de herbicidas.
4. Concluir los trabajos, fundamentalmente, en asociación en los suelos menos fértiles y sistema de pastoreo en las especies perennes, teniendo en cuenta que la carga no debe ser superior a 2 animales por hectárea.
5. Para facilitar la agilidad de este trabajo pueden recolectarse semillas en cantidades suficientes de las especies recomendadas para estudio en las zonas donde éstas se desarrollan en los meses de febrero y marzo.

Referencias bibliográficas
47
REFERENCIAS BIBLIOGRÁFICAS Academia de Ciencias de Cuba–URSS. 1970. Atlas Nacional de Cuba. Academia de Ciencias de Cuba-URSS.
La Habana
Anon. 1976. I Reunión Nacional de Genética Vegetal. CNCT. La Habana
Anon. 1976. Informe. Reunión Nacional sobre Pastos y Forrajes. La Habana.
Blazer, R.E. 1974. CIDA. Serie Ganadera 1
Blunt, C.G. 1976. Aust. Proc. of the Australian Society of Animal Production. 11
Boado, J. 1969. Pruebas de rendimiento de cinco variedades de alfalfa (Medicago sativa L.). Rev. cubana Cienc. agric. 3:47
Bogdan, A.V. 1966. En: Tropical Pasture. Faber and Faber. London
Boncearelli, F. & Convarelli, G. 1965. First results from trials on the improvement of pastures on the Apenines of umbria. Ann. Fac. Agr. Univ. Perurgia. 18:203
Bryan, W. & Evans, T.R. 1973. Aust. J. Exp. Agric. Anim. Husb. 10:431
Cain, S.A. 1944. Foundation of Plant Geography. Harpers, New York
Capote, S. & Shishchenc, S.V. 1977a. Efecto de diferentes factores ambientales sobre procesos fisiológicos de gramíneas y leguminosas. II. Frijol y maíz en cultivos puros y en asociación. Ciencias Biológicas. 1:57
CIAT. 1977. Resumen analítico sotare pastos tropicales. Cali, Colombia
Clark, N.A.; Leslie, J.J. & Hemken, H. 1966. Comparation of nitrogen fertilized grasses with a grass-legume mixture as pasture for dairy eves. I. Dry mather production, carrying capacity and milk production. Agron. J. 58:280
Cooksley, D.G. 1974. Growing and grazing Leucaena. Qd. Agric. Trin. 51:293
Eavis, B.W.; Cumberbath, E.R.St.J. & Medford, D.L. 1974. Factor influencing regeneration of natural vegetation on reformed Scotland District soil of Barbados. Trop. Agric. Trin. 51:293
Echevarría, N. 1977. Resumen VI Reunión ALPA. La Habana II
Echevarría, N. & Rodríguez, P. 1977. Resumen VI Reunión ALPA. II. La Habana, Cuba
Falvey, L. 1976. Productivity of Leucaena leucocephala in the Daly basin Northern Territory. Trop. Grasslds. 10:177
Febles, G. & Padilla, C. 1972. Evaluación de asociaciones. Rev. cubana Cienc. agric. 6:405
Febles, G. 1973. Efecto del pastoreo y el corte en la composición botánica y el rendimiento de asociaciones de gramíneas y leguminosas tropicales. Rev. cubana Cienc. agric. 7:127
Funes, F. 1979. Introducción de pastos y forrajes. En: Los pastos en Cuba. Tomo I Producción. La Habana, Cuba. p. 125
Funes, F. & Pérez, C. 1976. Comparación de variedades de soya perenne y teramnus sometidas a corte y pastoreo. Rev. cubana Cienc. agric. 10:205
Funes, F.; Yepes, S. & Hernández, D. 1971. Estudios de introducción de pastos de Cuba. II. Leguminosas más productivas. Memoria EEPF "Indio Hatuey". Matanzas, Cuba
Good. R. 1931. A theory of plant gepgraphy. New Phytol. 30:149
Good, R. 1953. Geography of the flowering plants. Longman Green and Company. London
Guzmán, J. 1967. Estudio sobre cuatro leguminosas perennes. Memoria anual. Centro de Investigaciones Agropecuarias. Universidad Central de Las Villas. Cuba. 71
Hartley, W. 1950. Aust. J. Agric. Res. 1:355
Henzell, E & Norris, D. 1962. CAB Bull No. 46
Henzell, E.F. 1968. Crop Soil and Crop Sci. Fla. 27:322

Referencias bibliográficas
48
Henzell, E.F. 1969. Tropical pasture legumes in northern Australia. Proc. Soil and Crop Sci. Fla. 27:322
Horrell, C. 1964. Effect of two legumes on the yield of fertilized pastures at serene. E. Afr. Agric. For. J. 30:94
Horrell, C. & Court, M. 1965. Effect of the legumes Stylosanthes gracilis on pastures yields at serene. Uganda J.B.R. Grass Soc. 20:272
Humphreys, L.R. 1967. En: Pasturas mejoradas para regions tropicales y subtropicales. Un guía. Wright. St. Co. Willmanpg. Co. Pty. Ltd. Victoria, Australia
Humphreys, L.R. 1964. A guide to better pastures for the tropics and subtropics. 3rd edition
Hutton, E.M. 1970. Tropical pastures. Advances in Agronomy. 22:2
Hutton, E.M. & Bonner, I.A. 1960. Dry matter and protein yield in tour strains of Leucaena glauca. J. Aust. Int. agric. Sci. 26:276
Instituto Cubano de Geodesia y Cartografía. 1978. Atlas de Cuba. La Habana, Cuba
Jewis, G.R. Geometría vegetal y fisiología del crecimiento. Conservación de forrajes. Instituto cubano del Libro. La Habana, Cuba. 77
Jones, R.J. 1967 Effects of close cutting and nitrogen fertilizer on growth of Siratro (Macroptilium atropurpureum) pasture at Sanford, south eastern Queensland. Aust. J. Int. Agric. Sci. 35:62
Jones, R.J. 1974. Aust. J. Exp. Agric. Anim. Husb. 14:334
Jones, R.J. & Jones, R.M. 1971. Trop. Grasslds. 5:3
Jones, R.J. & Jones, R.M. 1978. The ecology of Siratro, based pastures. Plan relations in pastures. CSIRO
Jones, R.M. 1975. Management of improved tropical pastures. Refresher Curse. Univ. Queensland, Australia
Köppen, W. 1936. Hand der Klimat. Bd-I, Text C:44
Kretschmer, A.E. 1974. En: Distribution, introduction, evolution of tropical species. Fot Pierce ABC Research Report. Florida, USA
León, Hno. & Alain, Hno. 1951. Flora de Cuba. Cols. II. La Habana
t’Mannetje, L. 1967. Trop. Grasslds. 1:9
t’Mannetje, L. 1974. CSIRO. Tropical Agronomy. Divisional Report
Mason, H.L. 1936. The prienciples of geografic distribution as applied to floral analysis. Madrano. 3:181
McIlroy, R.J. 1976. Introducción al cultivo de los pastos tropicales. Limusa
Mehra, K.L. & Magno, M.L. 1974. Proc. XII Intern. Grassld. Conf. Moscow : 908
Meigs, P. 1951. World distribution of arid and semi-arid homoclimate. Reviews of research on arid zones hidrology. UNESCO. Paris
Mejías, E.; Hernández, G.; Villamizar, F. & Chaverra, H. 1968. Reunión ALPA. Lima, Perú
Menéndez, J. 1978. Botánica de las leguminosas. Conferencia EEPF "Indio Hatuey". CUM. Matanzas, Cuba
Menéndez, J. & Machado, R. 1978. Leguminosas silvestres de Cuba. Oeste de las provincias orientales. Pastos y Forrajes. 1:349
Menéndez, J. & Martínez, J.F. 1980. Comportamiento de leguminosas asociadas al sorgo forrajero. Pastos y Forrajes. 3:83
Menéndez, J. & Pereira, E. 1979. Siratro (Macroptillum atropurpureum) Moc. y Sesse Urb. Pastos y Forrajes. 2:323
Menéndez, J.; Reid, R.; Machado, R. & Martínez, J.F. 1979. Leguminosas silvestres de Cuba. Pastos y Forrajes. 2:377
Menéndez, J.; Tang, M.; Aleaga, V & Polunin, N. 1980. Comparación de cvs de alfalfa (Medicago sativa L.) en Guantánamo. Pastos y-Forrajes. 3:213
Milford, R. & Haydock, K.P. 1965. Aust. J. Exp. Agric. Anim. Husb. 5:13

Referencias bibliográficas
49
Milford, R. y Minson, D.J. 1966. Proc. Aust. Sci. Anim. Prod. Husb. 1:815
Monzote, Martha. 1977. Resumen VI Reunión ALPA. La Habana, Cuba:152
Monzote, Martha & Hernández, T. 1977. Rev. cubana Cienc. Agric. 11:323
Moore, A. 1962. Emp. J. Agric. 30:239
Mulder, E.G. 1948. Pl. Soil. 1:179
Norris, D.O. 1967. The intelligent use of inoculants and line pelleting for tropical legumes. Trop. Grasslands. 1:107
Partridge, I.J. & Ranacou, E. 1974. The effects of supplemental Leucaena leucocephala on steers grazing Dichanthium caricosum in Fiji. Trop. Grasslds. 8:107
Pérez-Infante, F. 1970. Efecto de tres intervalos de corte y tres niveles de nitrógeno en las ocho gramíneas más extendidas en Cuba. Rev. cubana Cienc. agric. 4:145
Pérez-Infante, F. 1977. Posibilidades de los pastos en el trópico. Plenary Paper XIII Int. Grasslds. Congr. Leipzig:1
Quesada, R.R. & Oliva, O. 1971. Estudio sobre distancia de siembra en maíz (Zea mays L.) y frijol terciopelo (Stizolobium deeringianum Bent). Memoria EEPF ”Indio Hatuey”. Matanzas, Cuba. 115 p.
Rodríguez-Femenía, P. 1977. VI. Resúmenes VI Reunión ALPA. I. La Habana, Cuba. 104 p.
Salette, J.E. 1971. Intensifications prospects of forage production in the wet tropics. Colloque sur I’intensification de la production fourragere en milieu tropical humide et son utilization par les ruminants. 47 p.
Salinas, A. & Milera, Milagros. 1972. Pastoreo de pangola (Digitaria decumbens Stent.) comparada con la combinación de pangola y glycine en la producción de leche. Rev. cubana Cienc. Agric. 3:157
Shaw, N.H. 1961. Increase beef production from towsville lucerne (Stylosanthes aundica Taub) in the spear grass pastures of Central costal Queensland. Aust. J. Exp. Agric. Anim. Husb. 1:73
Shishchenco, S.V. & Capote, S. 1977. Efecto de diferentes factores ambientales sobre procesos fisiológicos de gramíneas y leguminosas. III. Soya en cultivo puro y en asociación con maíz. Ciencias Biológicas. 1:91
Sistachs, E. & López, Mirta. 1979. Bacteriología de las leguminosas tropicales. Los pastos en Cuba. Tomo I. p. 167
Skerman, P.J. 1977. Tropical forage legumes. FAO. Roma. 331 p.
Smith, C.A: 1963. Oversowing legumes into the Hyparrhenia grassland of Northern Rhodesia. Nature. 200(408):811
Thomas, D. & Whiteman, P.C. 1971. Aust. J. Exp. Agric. Anim. Husb. 52:513
Torcell, BW.R.; Ive, J.R. & Cunningham, R.B. 1976. Competition and population dynamics in legume grass swards with Stylosanthes hamata L. Taub. (sens. lat.) and Stylosanthes humilis (H.B.K.). Aust. J. Agric. R. 27:71
Upadhayay, V.S.; Rekib, A. & Pathak, P.S. 1974. Nutritive values of Leucaena leucocephala (Lam.) de Wit. Indian Vet. J. 51:534
Valdés, L.R. & Pérez, D. 1971. Observaciones en mezclas de gramíneas/leguminosas en pastoreo. Memoria EEPF "Indio Hatuey". Matanzas, Cuba. p. 38
Valdés, L.R.; Montoya, M. & Batista, J. 1977. Uso de las leguminosas o suplementación para la producción de carne. Resumen VI Reunión ALPA. II. La Habana, Cuba
Vavilov, N.I. 1950. Chron bot. 13:364
Whiteman, P.C. 1969. Aust. J. Exp. Agric. Anim. Husb. 9:287
Whiteman, P.C. & Lulham, A. 1970. Seasonal changes on growth and nodulation of serennial tropical pasture legumes in the field. I. The influence of planting date and grazing and cutting on Desmodium uncinatum and Phaseolus atropurpureum. Aust. J. Agric. Res.. 21:195
Whyte, R.O. 1958. En: Propección recogida e introducción de especies vegetales. 41. FAO. Estad. agrop. Roma

Referencias bibliográficas
50
Whyte, R.O.; Nilsson-Leissner, G. & Trumble, H.C. 1955. Las leguminosas en la agricultura. FAO
Williams, R.J. 1964. In: Some concepts and methods in subtropical pastures research. CSIRO. CAB. Bull 47. England
Wilsie, C.P. 1970. Cultivos: Aclimatación y distribución. Edición Revolucionaria
Williams, W.A. 1967. The role of Leguminosae in pasture and soil improvement in the neotropics. Trop. Agric. Trin. 44:103
Winter, W.H. 1976. Preliminary evaluation of twelve tropical grasses with legumes in northern cape York peninsula. Trop. Grasslds. 10:15
Yepes, S. 1971. Introducción al estudio de las leguminosas. Memoria EEPF "Indio Hatuey". Matanzas, Cuba
Yepes, S. 1971a. En: Observaciones sobre la evolución de leguminosas. Ser. 1. Ing. Agron. 7. Cienc. Agrop. Univ. de La Habana
Yepes, S. 1971b. Cienc. Agrop. Ing. Agron. Univ. de La Habana. 8:25
Yepes, S. 1974. Características botánicas de las principales leguminosas tropicales de pastoreo. Ser. 1. Ing. Agron. 15. Cienc. Agrop. Univ. de La Habana
Yepes, S. 1975. Centro de diversificación de las leguminosas forrajeras en América. Serie Técn. Cient. A-7. EEPF "Indio Hatuey". Matanzas, Cuba
Yeps, S.; Alfonso, F.M. & Funes, F. 1971. En: Introducción al estudio de las plántulas de leguminosas. Serie 1. Ing. Agron. 6. Cienc. Agrop. Univ. de La Habana
Zambrana, T. 1971. Evaluación del potencial agronómico de la alfalfa (Medicago sativa L.) bajo condiciones de Cuba. Tesis M.Cs. Agric. Univ Habana



![Perbandingan Metode Pembelajaran E-learning Dan …repository.gunadarma.ac.id/795/1/Perbandingan Metode Pembelajaran … · Literatul Review Darin E. Hartley [Hartley, 2001] yang](https://img.pdfslide.tips/doc/110x75/5abe64707f8b9aa3088ccfd0/perbandingan-metode-pembelajaran-e-learning-dan-metode-pembelajaran-literatul.jpg)