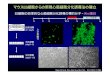
-
ラフト (細胞膜のミクロドメイン)
生体膜の動的な構造からは 膜成分は均一な分布となる。
ミクロドメインの形成
ラフト
コレステロールはミクロドメインを相分離で誘導する (Liquid-ordered phase, Lo)
・コレステロールが豊富に存在するラフトは界面活性剤に不溶な膜画分と
して発見され、スフィンゴ糖脂質とコレステロールが豊富に存在する。
・細胞の情報伝達系に関連する蛋白質が集積している。
細胞には非常に多くの生物機能が作動しているが、ミクロドメインの形成は微小な細の生体膜上胞に多様な機能を存在させるのに有効である。
-
コレステロールとスフィンゴ脂質によるミクロドメイン(液体秩序相・ラフト)形成の原理
DPPC-コレステロール系について、NMRとDSC(―)、ESR(---)、Freeze-fracture EM
(…)などの方法で求めた
相図をスケールを合わせて重ねてプロットした。
[J. H. Ipsen et al. Biochim. Biophys. Acta 905 (1987),
162-172]
液体秩序相
Liquid-ordered phase
ゲル相 液晶相
相転移
コレステロール
コレステロールが相分離を誘起した液体秩序相(Liquid ordered phase)
-
0
5
10
15
20
25
30
35
400 450 500 550 600Wavelength(nm)
10℃
20℃
25℃
30℃
35℃
40℃
45℃
50℃
60℃
65℃
70℃
I440
I490
Em
issi
on i
nte
nsi
ty (
arb.u
nt)
環境感受性蛍光色素 の特性
環境感受性色素Laurdan
GP I440 I490I
440 I
490
膜流動性の指標として GP(Generalized Polarization)を定義
ゲル相 液晶相
スペクトル変化は水の浸入度による
ゲル相
液晶相
DPPC(Tm=41 ℃)中でのLaurdanの発光スペクトル
:Laurdan
[Parassi et al.: Biophys. J. 60 (1991), 179-189]
6-dodecanoyl-2-
dimethylaminonaphthalene
-
10 20 30 40 50
-0.2
0.0
0.2
0.4
0.6
G.P.
Genera
lized P
loariza
tion
Temperature [ C]
DPPC (Tm=41 ℃)のリポソーム中のLaurdan のスペクトルから求めたGeneralized
Polarization (G.P.)の温度変化
ゲル相(Gel phase)
流動相(Fluid phase)
相転移
nmnm
nmnm
II
IIPG
490440
490440..
水の浸入がない。
脂質膜に水が浸入する。
-
膜流動性イメージング顕微鏡システム
Laurdanは発光波長が溶媒緩和によ
り、脂質膜がゲル相で水の浸入がないときの440nmから水が浸入した流動相の490nmにシフトする。
この変化をGP値の画像として定量化する顕微鏡装置を開発した。GP画像を得るために440nmの画像と490
nmの画像ダイクロイックミ
ラーと単色フィルターの組み合わせで分離し、両画像をCCDカメラの受像面を2分割して同時に観察し、
動的アフィン変換処理をパーソナルコンピューターで行いGP画像を
得る。
[Jpn. J. Appl. Phys. 44 (2005), 8733-8738]
-
DMPC/DMPEの2成分巨大リポソームのDSCサーモグラム
DMPC (Tm=23 ℃), DMPE (Tm=49 ℃ )
-
-0.2
-0.1
0
0.1
0.2
0.3
0.4
0 10 20 30 40 50
時間(分)
平均GP
45℃に設定
45℃
43℃に設定
43℃
40℃に設定
35℃に設定
DMPC/DMPE(1:1)巨大リポソームの温度変化の時間経過
リポソームの両脂質成分を混合するために温度を上昇、下降させる(annealing)。
-
DMPC/DMPE (1:1)の2成分で調製した巨大リポソームの相分離
[Tm=23 ℃(DMPC/49 ℃(DMPE]
Time; min:sec.00
・GPイメージング顕微鏡システムの性能評価のために DMPC /DMPE (1:1)の2成分巨大リ
ポソームの相分離をビテオレートで観察した。
・膜流動性の異なる部位が屈曲部位(DMPC-rich)と直線部位(DEPC-rich)として相分離して
いる。
-
脂質の形状からのラフト形成のモデル
円筒形のリン脂質 コレステロール スフィンゴミエリン
-
CHO細胞をメチルーβーシクロデキストリン(MβCDで処理してコレス
テロールを細胞膜から除去する。
スフィンゴミエリナーゼ(SMase)処理でスフインゴミエリン(SphM)をセラミド(Cer)に変換する。
Sphingomyelinase(SMase)
この処理による膜流動性 値 の変化を顕微鏡下にイメージングして解析する。
生きているCHO細胞に以下の処理を施し、コレステロールの膜流動性低下効果に注目して、細胞の膜流動性イメージング法により調べる。
Methyl-β-cyclodextrin(MβCD)
生きている細胞(CHO細胞)でのコレステロールとスフィンゴミエリンの相互作用からラフトの存在を確認する。
Phosphocholine
-
Sphingomyelinase
Laurdan 染色 (40 μM, 37 ℃, 30 min)
GP imaging
Methyl-β-cyclodextrin
Laurdan 染色 (40 μM, 37 ℃, 30 min)
GP imaging
スフィンゴミエリンのセラミドへの変換およびコレステロール除去による G.P.値の変化
スフィンゴミエリンのセラミドへの変換は膜流動性を変化させなかった。
コレステロールの除去で膜流動性が上昇した。
G.P. 値の低下は膜流動性
の上昇を示す。
1
1 mM の methyl-β-cyclodextrinはほとんど G.P. 値を低下させない.。
-
0.1 0.4
スフインゴミエリナーゼとメチル-β-シクロデキストリンの逐次処理による膜流動性の変化
最初のスフィンゴミエリナーゼ処理の酵素濃度(37 ℃, 10 min)
0U/ml 0.05U/ml 0.075U/ml 0.1U/ml 0.125U/ml
1mM 1mM 1mM 1mM 1mM
次のメチルβシクロデキストリン処理の濃度(37 ℃, 30 min)
前のスライドで1 mM のメチル-β-
デキストリン処理では膜流動性はほとんど低下しなかった。
この結果はメチル-β-デキストリンによる
コレステロールの除去がスフィンゴミエリンによる前処理の濃度に依存していることを示しており、スフィンゴミエリンとコレステロールの相互作用の存在を強く示唆している。
■: Sphingomyelinase treatment (37 ℃, 10 min)
●:Additional Mβcyclodextrin treatment (37 ℃, 30 min)
-
Liquid-ordered
Liquid-disordered
1 4 7 10 13 16 19 22 25 28
31 34 37 40 43 46 49 52 55 58
61 64 67 70 73 76 79 82 85 88
Bar indicate 10 μm
蛍光脂質の存在する明るい領域は液晶非秩序 ドメインを暗い領域は 液体秩序 ドメインを示している。
巨大リポソーム(GUV)の最初の脂質構成はDOPC/ C16:0-スフィンゴミエリン/コレステロール=1:1:1
(モル比)+0.1 0.1 mol% テキサスレッド-DPPE
巨大リポソームをスフィンゴミエリナーゼ(Streptomyces sp.を用いて室温(20-22 ℃)で処理した。
ラフト・モデルにおけるミクロドメインをスフィンゴミエリナーゼ処理(SphM →Cer)した際
の制御過程の観察
-
DOPC/C16スフインゴミエリン/コレステロールの3成分系にLaurdanを導入して巨大リポ
ソームを調製し膜流動性イメージング顕微鏡でGeneralized Polarization (G.P.)
画像を観察した。
G.P.値の高い(青い領域)がliquid-ordered phaseに対応する。
ラフト・モデルをスフィンゴミエリナーゼ処理(SphM
→Cer)した際の制御過程のLaurdanの観察(テキサスレッド-DPPEをLaurdanに置換)
)
最初は膜流動性が異なる領域が分離している状態であるがスフィンゴミエリナーゼ処理により、スフィンゴミエリンがセラミドに変換すると相分離が解消して、全体が均一化することが示された。
0 min
2
3
5
0.2 0.3 0.4 0.5 0.6 0.7
G P
0 min
1
2
3
4
5
6
7
8
9
1
4
6
9
50pixels
-
未分化のPC12D細胞をLaurdan で染色し、500 mg/ml NGF in BSS (Balanced Salt
Solution)に入れると突起の部分に膜流動性が低い(GP値が高い)部分が観察
された。 A:位相差顕微鏡像 B:蛍光画像 C:GP画像 Barは2μm
PC12D細胞に神経成長因子(NGF)を作用させた際の膜流動性の変化と突起伸展との関連について調べる。
PC12 細胞はラットの副腎髄質の褐色
細胞腫から株化された細胞で、神経成長因子(NGF)で神経細胞に分化する。
-
細胞周辺で盛んに突起を伸ばしている部位はGP値が高い。
細胞周辺のGP値が高い領域が移動して
他のGP値が高い領域に合体していく。
対照実験:NGFで刺激しないとき
はGP値が高い領域の移動は観察されない。(16分間の観察中)
PC12D細胞膜上の高GP値領域の神経成長因子(NGF)刺激に伴う変化
-
突起のない場所からはGP値の高い領
域が移動して、その部位から突起が伸展する。メチル-β-シクロデキスト
リンでコレステロールを除去するとGP値の高い領域は消失する。
NGF刺激によるPC12D細胞からの神経突起伸展とラフトとの関係
PC12D細胞を1日、NGFで処理し、Laurdanとラフトのマーカーであるコレラ
トキシンB(Alexa555蛍光標識)で染色して37℃で観察した。
Aは位相差顕微鏡像であり、CはLaurdan によるGP画像である。
Bは蛍光標識コレラトキシン
による蛍光画像である。
PC12D細胞でNGFによ
り誘導される神経突起はラフト領域から伸展することが示された。
-
Less
functional Functional Less
functional [Mouritsen,O.G. & Bloom, M. Biophys. J. 46
(1984), 141-153]
マットレスモデル:膜蛋白質の正常な機能発現には疎水性領域のマッチング(整合)が必要である。
膜脂質と膜蛋白質の疎水性領域の整合
整合した構造
ラフトへの特定の膜タンパク質の集積機構に疎水性領域の整合性が関与しているかを調べる。
-
NH2-GLY-ILE-GLY-ALA-VAL-LEU-LYS-VAL-LEU-THR-THR-GLY-LEU-PRO-
ALA-LEU-ILE-SER-TRP-ILE-LYS-ARG-LYS-ARG-GLN-GLN-CONH2
メリチン (melittin)
蜂毒のメリチンは26残基のアミノ酸からなるペプチドて゜疎水性アミノ酸領域と塩基性アミノ酸領域をもち、水中では左図のような4量体構造を形成する。
[Jung-Hsin Lin and A. Baumgaertner:Biophys.J 78 (2000),
1714–1724] Top view
Side view
分子動力学法で求めたメリチンの脂質2重層膜内での構造モデル:4分子のメリチンが穴を形成し、脂質の極性頭部がその内面に配置している。静電的な反発でねじれが生じて、拡張していく。
-
0 10 20 30 40 50 60 70
Temperature [ C]
Dimyristoylphosphatidylcholine
(DMPC)とDistearoylphosphatidylcholine
(DSPC)の1対1(モル比)2成分系のDSCサーモグラフ
対照
(メリチンなし)
5 mol% メリチン
DMPC Tm=23 ℃
DSPC Tm=55 ℃
対照ではDMPC-richとDSPC-richの2つのピーク
が観察されたが、メリチンの添加により、DMPC-richのピークが選択的に
減少した。
-
0 10 20 30 40 50 60 70
Temperature [ C]
Dimyristoylphosphatidylcholine(DMPC)とDipalmitoylphosphatidylcholine
(DPPC)の1対1(モル比)2成分系のDSCサーモグラフ
5 mol% メリチン
対照
(メリチンなし)
DMPC Tm=23 ℃
DPPC Tm=41 ℃
対照で1つのピークが観察されたがメリチンの添加により、2つのピークに分離した。
-
UDP-グルコース:セラミド グルコース転移酵素
UDP-glucose:ceramide glucosyl transferase
UDP-グルコース:セラミド グルコース転移酵素
・ゴルジ体外膜表面に局在
・基質は小胞体表面で合成されたセラミド
・UDP-グルコースからグルコースを転移
・グルコシルセラミド(Glucosyl ceramide)を合成
・グルコシルセラミドは内腔側へフリップ・フロップ
・その後、スフィンゴ糖脂質合成の基質に
・膜酵素、脂質要求性
-
UDPグルコース:セラミド
グルコース転移酵素を脂肪酸側鎖の長さの異なる各種のホスファチジルコリンに再構成して、酵素活性を測定する。
酵素活性は側鎖の長さに依存し適正な長さが存在する。
膜蛋白質の活性と脂質膜の厚さ
-
X線断層撮影 (X-ray Computed Tomography)
被験者の周囲を線源と検出器が回転し、被験者は全方位からX線受ける。照射されたX線は検査対象を通過し、対象に一部吸収されて減衰線源の反対側のX線検出装置で記録される。被験者に360度の方向から照射したX線の各方向での吸収度がCTのデータとなる。データからの画像の再構成では1断面を格子状に分割し、各部位の吸収率を未知数として、その合計が
実際の吸収量と等しくなるような連立方程式の解を求めることになる。膨大な行列演算であり、1断面を1024ピクセル四方に分割して処理する空間解像度の高い機種が開発されている。
検出器をらせん状に回転させるヘリカルCTや多数の検出器を並列に配置した多列検出器CTも開発されている。
-
X線CTにおける画像の再構成
X線管からは扇形にX線が照射され検出器に入る。全体はスリップ・リングで回転する。
左はX線の平面像、中央は左図の線で示された部分の湾曲像、右は全体を再構成した像である。
-
The first clinical results were presented at the Annual Meeting
of the British
Institute of Radiology in 1972 by Dr Jamie Ambrose, who showed
images from the
prototype of the very first EMI brain scanner.
X線CT (X-ray computed tomography)
X線CTの技術は英国のEMI社が医療用の画像診断装置として実用化した。
最新装置の螺旋式走査法 左は第1世代の画像、右は第4世代の画像
1972年英国のHounsfield がX線CTスキャン(Computed Tomography)を発表した。
-
X線断層撮影画像の例
健常者の頭部(脳) 癌の転移が見られる胸部(肺)
-
X線断層撮影画像の例
ヒト脳内の血管像
ヒト頭部をスライスした画像
-
陽電子放出断層撮影 Positron emission tomography
核医学領域の生体イメージング法であり、陽電子放出核種をもつ分子を生体内に注入し、陽電子崩壊を利用してその局在を3次元で観察する装置
癌細胞は代謝が活発で多くのエネルギーを必要とするため、そこにはブドウ糖(glucose)が
集積する。
体内で放射性薬剤が放出する陽電子は、周囲の電子と衝突して運動エネルギーをて失うが、ついには周囲の電子と結合して対消滅を生じる。そのとき、全質量エネルギーに等しい消滅光子を180度の反対方向に2個放出する。PETではこの消滅光子を対向する検出器対で同時計数測定し、検出器対を結ぶ線上で電子-陽電子消滅が生じたことを検出する。そして、薬剤の分布位置のデータを取得している。
-
粒子と反粒子が衝突して、エネルギーまたは別の粒子に変換される現象で、対生成の逆の現象である。
電子と陽電子が衝突すると、それぞれの静止エネルギー(511keV/C2)とそれらがもつ運動エネルギーの和に等しいエネルギーをもつ光子が放出されて、γ線として観測される。対消滅では運動量が保存されるので、運動エネルギーをもたない電子と陽電子が対消滅して二つの光子を作り出す場合、エネルギーは均等に分配され、反対方向に放射線が飛翔する。
対消滅(pair annihilation)
PETに用いられる主な陽電子放出核種と生成反応
-
陽電子崩壊と対消滅の過程
陽電子崩壊による消滅放射線の飛翔
陽電子を放出する核種
放射線
(511 keV)
放射線
(511 keV)
180 ゜
飛程
-
PET装置の概略図
-
PET装置の概略図
-
フッ素(F-18)を含むブドウ糖 (fluorodeoxyglucose ,FDG) をヒトに注入し
て得たPET画像
人体のPET画像 PETの臨床利用例(:東北大学CYRIC核医学研究部)
運動(ゴルフ)経験者と初心者の筋活動
ハンディ0 初心者 癌の検出
脳のPET画像
-
2003年のノーベル医学生理学賞:米イリノイ大のポール・ラウターバー(Paul C. Lauterbur,1929
年生)教授と英ノッティンガム大のピーター・マンスフィールド(Sir Peter
Mansfield,1933年生)名誉教授の磁気共鳴断層撮影(MRI)装置開発の基礎を作った業績にに対して。
MRI装置の外観
MRI (Magnetic Resonance Imaging)
-
核磁気共鳴画像法 (magnetic resonance imaging, MRI)
人体の約60%は水であり、その水素原子核は核スピンをもち、外部から(強い)静磁場を作用させると、核スピンの磁化は磁場の向きに量子化され、磁場の向きに巨視的磁化が生じる。この核磁化に特定の周波数のラジオ波を照射すると核磁化は、静磁場方向を軸としてラーモア周波数で歳差運動を行う。ラジオ波のパルスの照射を止めると平衡状態に戻る(緩和現象、relaxation)。これには縦緩和(T1)と横緩和(T2)があり、それぞれの組織によって緩和時間が異なる。核磁気共鳴画像法では、各組織における緩和時間の差をパルスシーケンスのパラメーターを工夫して画像化している。このNMR信号の位置情報を得るためには静磁場に加えて、距離に比例した強度をもつ勾配磁場をかけている。勾配磁場によって原子核の位相や周波数が変化するので、これらの観測信号フーリエ変換して、2次元画像を得、さらにこれを3次元に再構成する。
1
0
T
MMBM
dt
dM zeff
2
,,
T
MBM
dt
dM yxeff
yx
ブロッホ方程式
-
1945年
BlochとPurcellによりNMR現象が発見され、最初はスペクトロスコピーとして化学分析用の装置として用いられた。
1971年 Damadianが腫瘍の良性、悪性の鑑別ができる可能性があることを示唆した。
1973年 Lauterburにより傾斜磁場を用いて二次元画像を得るMRIの基礎が確立した。
1970年代後半 Damadian, Hinshow,Mansfieldが人体の映像化に成功した。
MRIの本格的な応用は1977年頃にイギリスで行われ、その後20年余の変遷を経て現在に至っている。
MRI実用化への歴史
-
生物体はその60 %は水であり、生体内には水を含めて無数の水素原子核(=プロトンproton)が存在する。
個々のプロトン(1H)は核スピンをもち小さな磁石と同じと見なせる。
強い静磁場の中にプロトンをおくと、一斉に縦方向に整列する(Zeeman効果)。
共鳴条件を満たす周波数の電磁波を照射すると、プロトンが横向きに倒れる(磁気共鳴現象)。
外部の電磁波を切ると、電磁波を出しながらプロトンが平衡状態に戻っていく(緩和現象)。
緩和の速度は、組織により、また病変により異なるので、この電磁波を分析すれば緩和の速度を知ることができる。
これを、画像の白黒または色に反映させて診断する。 2次元の情報は磁場の強度を傾斜させ、共鳴点の位置を取得する。
3次元化は2次元のスライス情報を再構成することで行う。
MRIの基本的な原理




![iPS細胞の倫理と国際戦略 - Waseda University · 2016-06-20 · iPS 細胞は無限に ... ・コ ピーであり、劣化するカーボン・コピーでは ない。例えば「ヒーラ[HeLa]細胞」(Skl](https://img.pdfslide.tips/doc/110x75/5cc4072288c993df118e1409/ips-waseda-university-2016-06-20-ips-.jpg)