-
8/10/2019 Hum. Reprod.-2002-Barnes-88-91.pdf
1/4
Human Reproduction Vol.17, No.1 pp. 8891, 2002
CASE REPORT
The role of LH and FSH in ovarian androgen secretion andovarian
follicular development: Clinical studies in a patientwith isolated
FSH deficiency and multicystic ovaries
Randall B.Barnes1,5, Anne B.Namnoum2, Robert L.Rosenfield3 and
Lawrence C.Layman4
Departments of Obstetrics and Gynecology, 1University of
Chicago, Chicago, IL 60637, 2Emory University, Atlanta, GA
30322,3Department of Pediatrics and Medicine, University of
Chicago, Chicago, IL 60637, 4Department of Obstetrics and
Gynecology,
Medical College of Georgia, Augusta, GA 30912, USA
5To whom correspondence should be addressed at: Dept of
Obstetrics and Gynecology, University of Chicago, 5841 Maryland
Avenue, Chicago, IL 60637, USA. E-mail:
[email protected]
Inactivating mutations have proven to be instructive in
elucidating the role of FSH in human ovarian function. We
performed a detailed reproductive endocrine evaluation of a
patient with inactivating mutations in the FSH
-subunit gene who was hypo-estrogenic and had LH excess. The
patient underwent a pelvic ultrasound andovernight frequent blood
sampling followed by a human chorionic gonadotrophin (HCG)
stimulation test. One
month later she received human recombinant FSH, followed 24 h
later by a second HCG stimulation test. Despite
a mean LH serum concentration and LH pulse characteristics
typical for polycystic ovaries (PCOS), baseline and
dexamethasone-suppressed free testosterone were lownormal. The
administration of HCG led to minimal stimulation
of 17-hydroxyprogesterone and androgens. The patient had
multicystic ovaries containing follicles 35 mm in
diameter and responded to FSH with prompt increases in estradiol
and inhibin B. There were no clinical or
laboratory consequences of LH excess in this FSH-deficient
woman. These findings support the hypothesis that
excessive LH stimulation alone does not cause ovarian
hyperandrogenism. We also found that follicular development
was present in the absence of FSH. These antral follicles had
apparently developed normally, since estradiol and
inhibin B increased promptly after FSH administration.
Key words: androgens/FSH deficiency/LH excess/ovary
Introduction
Inactivating mutations have proven to be instructive in
elucidat-
ing the role of FSH in human ovarian function (Layman and
McDonough, 2000; Themmen and Huhtaniemi, 2000). Our
studies of a patient with compound heterozygous mutations of
the FSH -subunit gene (Layman et al., 1997), undetectableserum
FSH and elevated serum LH have elucidated two aspects
of FSH action which challenge common wisdom. We report
here evidence that FSH is not necessary for the development
of small, healthy antral follicles readily responsive to
FSH,
which contrasts with studies in FSH-knockout mice (Kumar
et al., 1997; Dierich et al., 1998). However, FSH is
necessary
for ovarian theca cell function, contrary to expectations
from
the 2-gonadotrophin, 2-cell model of ovarian steroidogenesis
(Barnes et al., 2000). The latter finding has important
implications for theories about the role of LH excess in the
pathogenesis of polycystic ovary syndrome (PCOS).
88 European Society of Human Reproduction and Embryology
Case report
Subject
The patient presented at 16 years of age with primary
amenorrhoea and absent breast development. Her baseline
and gonadotrophin-releasing hormone (GnRH)-stimulatedFSH
concentrations were undetectable by immunoassay, while
LH concentrations at baseline (30 mIU/ml) and post-GnRH
(150 mIU/ml) were elevated. The patient was subsequently
treated with estrogen and underwent normal breast develop-
ment. Analysis of the FSH -subunit gene demonstrated thatshe was
a compound heterozygote for two different mutations,
a two base-pair deletion in exon three at codon 61 (Val61X)
inherited from her mother, and a missense mutation changing
a cysteine to a glycine at codon 51 (Cys51Gly) inherited
from
her father. When these mutations were stably transfected
into
Chinese hamster ovary cells, they demonstrated no measurable
FSH immuno- or bioactivity (Layman et al., 1997).
-
8/10/2019 Hum. Reprod.-2002-Barnes-88-91.pdf
2/4
Clinical studies in a patient with FSH deficiency
Figure 1. Outline of clinical research center
protocols.Dexamethasone, 0.5 mg four times daily, was started on
day 1 andcontinued until the blood sample was drawn 24 h after
HCG.
Reproductive endocrine evaluation
The patient underwent a detailed reproductive endocrine
evaluation at 21 years of age in the University of Chicago
General Clinical Research Center (GCRC), after dis-
continuing the oral contraceptive pill for one month and
giving
informed consent. The study protocols were approved by the
University of Chicago Institutional Review Board. Her heightwas
155 cm and her weight was 53.2 kg. There was no
hirsutism (Ferriman and Gallway score of 4; normal 8),
breasts and pubic hair were Tanner stage 5, and the pelvic
examination was normal. An initial blood sample was drawn
for
immunoactive FSH, LH and free testosterone. The patient then
received dexamethasone 0.5 mg four times daily for 4 days
prior to and continuing throughout the endocrine evaluation
to suppress adrenal function. Following admission to the
GCRC, she underwent a transvaginal pelvic ultrasound fol-
lowed by blood sampling every 10 min from 7.00 p.m. to 6.00
a.m. Serum immunoactive FSH and LH were determined for
each sample; inhibin A and B, bioactive FSH and free -
subunit were determined in a pooled sample. The
followingmorning, a human chorionic gonadotrophin (HCG) test
was
performed. A blood sample was drawn for baseline testoster-
one, free testosterone and estradiol. The following steroid
intermediates were also determined: 17-hydroxyprogesterone
(17-Prog), androstenedione, 17-hydroxypregnenolone and
dehydroepiandrosterone. HCG 5000 IU was then administered
i.m. and blood was drawn 24 h later for repeat steroid
measurements.
The patient remained off any sex steroids for another month,
then returned to the GCRC for a second HCG study which
was like the first except she received 300 IU of recombinant
human FSH s.c. in the morning, 24 h prior to the HCG test.
A blood sample was drawn 24 h later for steroids and inhibin
A and B, and HCG 5000 IU was administered. She then
returned 24 h after HCG (48 h after FSH) for a final blood
sample for steroid measurements (Figure 1).
All samples for steroids were frozen at 20C and
measured in the same assay using previously published
methods
(Barnes et al., 1989; Rosenfieldet al., 1994). Serum immuno-
active LH and FSH were measured using a -subunit-specificassay
(Delfia, Wallach, Finland; lower limit of sensitivity
0.15 mIU/ml for LH and 0.2 mIU/ml for FSH). Bioactive
FSH, inhibin A, inhibin B and free -subunit were measuredusing
previously published methods (Christin-Maitre et al.,
89
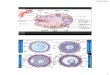
Figure 2. Transvaginal ultrasound of the left ovary before
FSHtreatment. The ovary is fusiform in shape, measuring ~61.5
cm.The stroma is scant but there are multiple follicles throughout
theovary, the largest measuring 5 mm. The right ovary (not
shown)was similar to the left.
1996; Lavoieet al., 1998; Weltet al., 1999). LH pulse
analysis
was performed using the ULTRA program (Van Cauter andCopinschi,
1981).
Results
The ovarian ultrasound prior to FSH treatment is shown in
Figure 2. The ovaries were multicystic containing follicles
35 mm in diameter, but unlike typical polycystic ovaries
(Adams et al., 1986), there was no dense stroma and the
follicles were scattered throughout the ovary rather than
being
predominately subcapsular.
Immunoactive LH was 46 mIU/ml and FSH was undetectable
(0.2 mIU/ml) in the single sample drawn before dexametha-
sone. There was no immunoactive FSH detected in the
samplescollected overnight (Table I), and no bioactive FSH was
detected in the pooled sample. The mean immunoactive
LH in the overnight samples was 46 mIU/ml; mean pulse
amplitude was 15.8 mIU/ml and nine pulses were detected in
11 h (Figure 3). This LH profile is similar to that reported
for
PCOS (Waldstreicher et al., 1988). Free -subunit measuredin the
pooled 11 h sample was in the postmenopausal range
(3501740 pg/ml; Table I).
Although LH concentrations were elevated, the patient had
no laboratory evidence of ovarian hyperandrogenism. The
patients baseline free testosterone was 6 pg/ml (normal
range 310 pg/ml; ovarian hyperandrogenism 10 pg/ml)
(Rosenfield et al., 1994). After 4 days of dexamethasone
suppression the free testosterone fell to 2 pg/ml (normal
range
8 pg/ml; ovarian hyperandrogenism 8 pg/ml; Rosenfield
et al., 1994). The 17-Prog concentration 24 h after HCG was
40 ng/dl which was 2.7 standard deviations (SD) below the
mean in women with ovarian hyperandrogenism (288 ng/dl)
(Levrantet al., 1997) and 1.5 SD below the mean of normal
follicular phase women (139 ng/dl: Levrant et al., 1997).
One month later, during the second GCRC study,
estradiol increased substantially 24 h after FSH
administration
(2880 pg/mL). Inhibin B was near the lower limit of the
normal range at baseline and rose to above the range for
-
8/10/2019 Hum. Reprod.-2002-Barnes-88-91.pdf
3/4
R.B.Barnes et al.
Table I. Gonadotrophin and inhibin concentrations in an
FSH-deficient patient
FAS LH FSH Bioactive FSH Inhibin A Inhibin B(pg/ml) (mIU/ml)
(mIU/ml) (mIU/ml) (pg/ml) (pg/ml)
Baselinea 1247 46 0.2 4 0.6 25.324 h after FSH 8.4 0.8 283Normal
women 103257 015 320 440 0.680 15220
aBaseline is mean of every 10 min samples over 11 h for LH and
FSH. For all others, baseline is a value ofthe pooled 11 h sample.
Normal values are from Christein-Maitre et al., 1996; Lavoie et
al., 1998; Weltet al., 1999.FAS free alpha subunit.
Figure 3. LH pulse analysis. LH was measured at 10 min
intervalsover 11 h. Significant pulses are indicated by an
asterisk.
menstruating women 24 h after FSH injection (Table I). In
contrast, inhibin A was present at low concentrations.
The response of testosterone and the steroid intermediates
to the two HCG tests have been previously reported (Barnes
et al., 2000). In brief, all steroid concentrations 24 h after
the
second HCG injection (48 h after FSH) were greater than at
24 h after the first HCG injection (HCG alone). The
difference
between the two HCG tests was most dramatic for
testosterone,
which was unaffected by the first HCG test (12 ng/dl before
and after HCG), but increased from 16 to 41 ng/dl after the
second, FSH-primed HCG test.
Discussion
This patient, with well characterized inactivating mutations
in
the FSH -subunit gene, was hypo-estrogenic with secondaryLH
excess. Two features were noteworthy: antral follicular
development was present in spite of the lack of FSH, and
there were no clinical or laboratory consequences of the
LH excess.
The point at which FSH is necessary for follicular develop-
ment in the human ovary is debated (Gougeon, 1996; McGee
and Hsueh, 2000). Despite having no detectable FSH, our
patient had multicystic ovaries with follicles up to 5 mm in
diameter. Similar sized follicles have been reported in some
women with FSH receptor mutations (Aittomaki et al., 1996;
Beau et al., 1998); however, those FSH receptor mutations
90
may not be completely inactivating. These findings are in
contrast to FSH- and FSH receptor-knockout mice, in whichantral
follicles are not maintained (Kumar et al., 1997; Dierich
et al., 1998). In studies of human ovarian xenografts trans-
planted into the kidney capsule of immunodeficient and hypo-
gonadotrophic mice, FSH was required for the growth of
follicles beyond the two-layer granulosa cell stage (Oktay
et al., 1998), which is about the point at which the FSHreceptor
gene isfirst expressed in human follicles (Oktay et al.,
1997). Although the xenograft data are in contrast to our
findings, it is likely that the endocrine and growth factor
milieu
of the in-situ human ovary is very different than that in
the
immunodeficient, hypogonadotrophic mouse.
The prompt estradiol and inhibin B responses 24 h after
exogenous FSH suggest that our patients antral follicles
contained granulosa cells that had developed normally in the
absence of FSH. Our findings are similar to those in two
other
patients with isolated FSH deficiency who ovulated and had a
successful pregnancy after ~14 days of menotrophin therapy
(Rabinowitz et al., 1979; Matthews et al., 1993). Ovulation
after a short exposure to menotrophins implies that somehealthy
follicles had reached the point of recruitability without
FSH exposure, since ~14 days are required for an ovulatory
follicle to develop from follicles a few millimeters in
diameter,
in contrast to the 3 months required to develop from the
pre-
antral stage (Gougeon, 1996). These earlier reports and our
findings suggest that in the complete absence of FSH
stimula-
tion, human ovarian antral follicles can develop up to 5 mm
in diameter.
Our patient had no clinical or laboratory evidence of
ovarian
hyperandrogenism despite a mean LH concentration, LH pulse
characteristics and ovarian follicular sizes typical for
PCOS.
On ultrasound her multicystic ovaries lacked the excess
stroma
of classic polycystic ovaries. Indeed, her ovaries produced
little, if any, androgen. Her baseline and dexamethasone-
suppressed free testosterone were lownormal. The administra-
tion of HCG led to minimal stimulation of 17-Prog or other
thecal cell steroids. However, we have previously shown that
exogenously administered FSH augmented her LH- and HCG-
stimulated production of testosterone and all steroids of
thecal
cell origin (Barneset al., 2000). This suggests that FSH
action,
probably via granulosa cell-produced paracrine
intermediates,
is necessary for thecal cells to respond to LH. One of many
such paracrine factors is inhibin B, which increased
markedly
with FSH administration in this patient (Barnes 1998).
-
8/10/2019 Hum. Reprod.-2002-Barnes-88-91.pdf
4/4
Clinical studies in a patient with FSH deficiency
These findings are relevant to the role of LH excess in the
pathogenesis of the ovarian hyperandrogenism of PCOS. It
has been reported that 75% of women with clinical evidence
of PCOS have an elevated LH concentration and 94% have
an increased LH/FSH ratio (Taylor et al., 1997). These
gonadotrophin secretory abnormalities have been thought to
play an important role in the development of the ovarian
hyperandrogenism characteristic of PCOS (Hall, 1993). In
arelated study, we found that a woman with a constitutively
activating mutation of the LH receptor, identified because
she
was the mother of two sons with gonadotrophin-independent
precocious puberty, had no clinical or laboratory evidence
of
ovarian hyperandrogenism (Rosenthal et al., 1996). Thus,
increased LH stimulation, even in the presence of FSH,
appears
to be insufficient to induce the hyperandrogenism and
stromal
hyperplasia of PCOS. The findings in the previous, as well
as
the current, case report support the hypothesis that thecal
cell
androgen secretion in response to excessive LH stimulation
is
strictly limited by intra-ovarian factors. Ovarian
hyperandro-
genism is more likely a result of escape from
down-regulation
by these intra-ovarian factors than a result of elevated
LHconcentrations (Ehrmann et al., 1995). Taken together, our
studies support the hypotheses that normal ovarian androgen
production depends on both FSH and LH and that excessive
ovarian androgen production is a result of abnormal intra-
ovarian regulation, and not of excessive LH stimulation.
Acknowledgements
We are greatly indebted to Patrick Sluss,Reproductive Endocrine
Unit,The Massachusetts General Hospital, Boston, MA, who
performedthe FSH bioassay, FAS, inhibin A, and inhibin B assays; to
ZubieSheikh, Department of Obstetrics and Gynecology, University
ofChicago, for performing the transvaginal ultrasound; and to
Jennifer
M.Cunningham, Department of Medicine, University of Chicago,
forperforming the LH pulse analysis. We also appreciate the
helpfuldiscussions of J.Larry Jameson from the Center for
Endocrinology,Metabolism, and Molecular Medicine, Northwestern
University,Chicago, IL and William F.Crowly Jr from the
ReproductiveEndocrine Unit and the National Center for Infertility
Research, TheMassachusetts General Hospital, Boston, MA.
L.C.L. was supported by NIH grant support PHS NICHD
HD33004;R.L.R. was supported by NIH grant support RR-00055
(CRC).
References
Adams, J., Polson, D.W. and Franks, S. (1986) Prevalence of
polycysticovaries in women with anovulation and idiopathic
hirsutism. Br. Med. J.,293
, 355359.Aittomaki, K., Herva, R., Stenman, U-H. et al. (1996)
Clinical features ofprimary ovarian failure caused by a point
mutation in the follicle-stimulating hormone receptor gene. J.
Clin. Endocrinol. Metab., 81,37223726.
Barnes, R.B. (1998) The pathogenesis of polycystic ovary
syndrome: lessonsfrom ovarian stimulation studies. J. Endocrinol.
Invest., 21 , 567579.
Barnes, R.B., Rosenfield, R.L., Burstein, S. et al. (1989)
Pituitaryovarianresponses to nafarelin testing in the polycystic
ovary syndrome. N. Engl.
J. Med., 320 , 559565.
Barnes, R.B., Namnoum, A., Rosenfield, R.L. et al. (2000) Effect
of follicle-stimulating hormone on ovarian androgen production in a
woman withisolated follicle-stimulating hormone deficiency. N.
Engl. J. Med., 343,11971198.
91
Beau, I., Touraine, P., Meduri, G. et al. (1998) Novel phenotype
related topartial loss of function mutations of the follicle
stimulating hormonereceptor. J. Clin. Invest., 102 , 13521359.
Christin-Maitre, S., Taylor, A.E., Khoury, R.H. et al. (1996)
Homologousin vitro bioassay for follicle-stimulating hormone (FSH)
reveals increasedFSH biological signal during the mid- to late
luteal phase of the humanmenstrual cycle. J. Clin. Endocrinol.
Metab., 81, 20802088.
Dierich, A., Sairam, M.R., Monaco, L. et al. (1998) Impairing
follicle-stimulating hormone (FSH) signaling in vivo: targeted
disruption of theFSH receptor leads to aberrant gametogenesis and
hormonal imbalance.
Proc. Natl Acad. Sci. USA, 95, 1361213617.Ehrmann, D.A., Barnes,
R.B. and Rosenfield, R.L. (1995) Polycystic ovary
syndrome as a form of functional ovarian hyperandrogenism due
todysregulation of androgen secretion. Endocr. Rev., 16 ,
322353.
Gougeon, A. (1996) Regulation of ovarian follicular development
in primates:facts and hypotheses. Endocr. Rev., 17, 121155.
Hall, J.E. (1993) Polycystic ovarian disease as a neuroendocrine
disorder ofthe female reproductive axis.Endocrinol. Metab. Clin.
North Am., 22, 7592.
Kumar, T.R., Wang, Y., Lu., N. et al. (1997)
Follicle-stimulating hormone isrequired for ovarian follicle
maturation but not male fertility.Nature Genet.,15, 201204.
Lavoie, H.B., Martin, K.A., Taylor, E., et al. (1998)
Exaggerated free alpha-subunit levels during pulsatile
gonadotropin-releasing hormone replacementin women with idiopathic
hypogonadotropic hypogonadism. J. Clin.
Endocrinol. Metab., 83 , 241247.
Layman, L.C. and McDonough, P.G. (2000) Mutations of follicle
stimulating
hormone-beta and its receptor in human and mouse:
genotype/phenotype.Mol. Cell. Endocrinol., 161 , 917.
Layman, L.C., Lee, E.J., Peak, D.B. et al. (1997) Delayed
puberty andhypogonadism caused by mutations in the
follicle-stimulating hormone -subunit gene. N. Engl. J. Med., 337 ,
607611.
Levrant, S.G., Barnes, R.B. and Rosenfield, R.L. (1997) A pilot
study of thehuman chorionic gonadotrophin test for ovarian
hyperandrogenism. Hum.
Reprod., 12, 14161420.
Matthews, C.H., Borgato, S., Beck-Peccoz, P. et al. (1993)
Primaryamenorrhoea and infertility due to a mutation in the
-subunit of follicle-stimulating hormone. Nature Genet., 5 ,
8386.
McGee, E.A. and Hsueh, A.J.W. (2000) Initial and cyclic
recruitment ofovarian follicles. Endocr. Rev., 21 , 200214.
Oktay, O., Briggs D. and Gosden, R.G. (1997) Ontogeny of
follicle-stimulatinghormone receptor gene expression in isolated
human ovarian follicles. J.Clin. Endocrinol. Metab., 82 ,
37483751.
Oktay, O., Newton H., Mullan J. et al. (1998) Development of
humanprimordial follicles to antral stages in SCID/hpg mice
stimulated withfollicle-stimulating hormone.Hum. Reprod., 13 ,
11331138.
Rabinowitz, D., Benveniste, R., Lindner, J. et al. (1979)
Isolated follicle-stimulating hormone deficiency revisted. N. Engl.
J. Med., 300 , 126128.
Rosenfield, R.L., Barnes, R.B. and Ehrmann, D.A. (1994) Studies
of thenature of 17-hydroxyprogesterone hyperresponsiveness to
gonadotropin-releasing hormone agonist challenge in functional
ovarianhyperandrogenism.J. Clin. Endocrinol. Metab., 79,
16861692.
Rosenthal, I.M., Refetoff, S., Rich, B. et al.(1996) Response to
acute challengewith gonadotropin releasing hormone agonist in two
half-brothers and theirmother with a constitutively activating
mutation of the luteinizing hormonereceptor. J. Clin. Endocrinol.
Metab., 81 , 38023806.
Taylor, A.E., McCourt, B., Martin, K.A. et al. (1997)
Determinants ofabnormal gonadotropinsecretion in clinically defined
women with polycysticovary syndrome. J. Clin. Endocrinol. Metab.,
82, 22482256.
Themmen, A.P.N. and Huhtaniemi, I.T. (2000) Mutations of
gonadotropins
and gonadotropin receptors: elucidating the physiology and
pathophysiologyof pituitarygonadal function. Endocr. Rev., 21,
551583.
Van Cauter, E. and Copinschi, G. (eds) (1981) Human
pituitaryhormones: circadian and episodic variations. Martinus
Nijhoff, The Hague,pp. 221235.
Waldstreicher, J., Santoro, N.F., Hall, J.E., et al. (1988)
Hyperfunction of thehypothalamicpituitary axis in women with
polycystic ovarian disease:indirect evidence for partial
gonadotroph desensitization.J. Clin. Endocrinol.
Metab., 66 , 165172.
Welt, C.K., Adams, J.M., Sluss, P.M. et al. (1999) Inhibin A and
inhibinB responses to gonadotropin withdrawal depends on stage of
follicledevelopment.J. Clin. Endocrinol. Metab., 84 , 21632169.
Received on February 28, 2001; accepted on August 21, 2001





















![Sophismata Buridani ([Reprod.])](https://img.pdfslide.tips/doc/110x75/625d10ffa98da525ef7f60fa/sophismata-buridani-reprod.jpg)