Embed Size (px)
Citation preview

TS. CHU HOÀNG MẬU
CƠ SỞ VÀ PHƯƠNG PHÁP SINH HỌC PHÂN TỬ
NHÀ XUẤT BẢN ĐẠI HỌC SƯ PHẠM

Lời nói đầu.................................................................................................................................4
Chương 1: HỆ GENE..............................................................................................................5
§1. KHÁI NIỆM HỆ GENE (GENOME) ..................................................................5 §2. AXIT NUCLEIC ..................................................................................................9 §3. ADN VÀ TÁI BẢN ADN..................................................................................11 §4. ARN VÀ CƠ CHẾ PHIÊN MÃ .........................................................................19 THẢO LUẬN...........................................................................................................23
Chương 2: ĐẶC ĐIỂM CẤU TRÚC GENE CỦA SINH VẬT PROKARYOT VÀ EUKARYOT ...........................................................................................................25
§1. ĐẶC ĐIỂM CẤU TRÚC GENE Ở PROKARYOT ..........................................25 §2. CẤU TRÚC PHÂN ĐOẠN GENE Ở EUKARYOT .........................................25 §3. MỘT SỐ TRÌNH TỰ ADN................................................................................27 THẢO LUẬN...........................................................................................................32
Chương 3: MỐI LIÊN HỆ GIỮA ADN, ARN, PROTEIN................................................33
§1. THÔNG TIN DI TRUYỀN VÀ MẬT MÃ DI TRUYỀN .................................33 §2. PROTEIN ...........................................................................................................39 §3. QUÁ TRÌNH SINH TỔNG HỢP PROTEIN.....................................................42 §4. ĐIỀU HOÀ BIỂU HIỆN GENE ........................................................................45
THẢO LUẬN......................................................................................................................49
Chương 4: ENZYME SỬ DỤNG TRONG KĨ THUẬT SINH HỌC PHÂN TỬ ............50
§1. ENYME GIỚI HẠN (RESTRICTION ENZYME HOẶC ENZYME CẮT HẠN CHẾ (RESTRICTION ENDONUCLEASE - RE) .............................................50
§2. CÁC NHÓM ENZYME KHÁC.........................................................................57 §3. ENZYME NUCLEASE......................................................................................60
THẢO LUẬN......................................................................................................................61
Chương 5: XÁC ĐỊNH MỐI LIÊN QUAN GIỮA PROTEIN VÀ ĐẶC TÍNH Ở SINH VẬT ...............................................................................................................62
§1. MỘT SỐ LOẠI PROTEIN CHỨC NĂNG........................................................62 §2. QUY TRÌNH XÁC ĐỊNH CHỈ THỊ PHÂN TỬ PROTEIN..............................66 §3. KĨ THUẬT PHÂN TÍCH PROTEIN .................................................................67 §4. KĨ THUẬT PHÂN TÍCH ENZYME..................................................................74 §5. NGHIÊN CỨU RIPS BẰNG WESTERN BLOT ..............................................77
THẢO LUẬN......................................................................................................................78
Chương 6: KỸ THUẬT SINH HỌC PHÂN TỬ TRONG PHÂN TÍCH HỆ GENE SINH VẬT..........................................................................................................................79
§1. PHƯƠNG PHÁP TÁCH CHIẾT ADN..............................................................79 §2. LAI PHÂN TỬ ...................................................................................................83 §3. XÁC ĐỊNH TRÌNH TỰ NUCLEOTIT CỦA ADN ..........................................85 §4. RFLP TECHNOLOGY (Kĩ thuật phân tích hiện tượng đa hình của độ dài các
phân đoạn ADN).................................................................................................87
2

THẢO LUẬN......................................................................................................................91
Chương 7: PHẢN ỨNG CHUỖI POLIMERASE (POLIMERASE CHAIN REACTION - PCR)................................................................................................92
§1. PHẢN ỨNG CHUỖI POLIMERASE (PCR) ....................................................92 §2. PHÂN TÍCH TÍNH ĐA HÌNH CỦA ADN ĐƯỢC NHÂN BẢN NGẪU
NHIÊN (RANDQM AMPLIFIED PQLIMORPHIC DAN - RAPD) ................99 §4. PHÂN TÍCH TÍNH ĐA HÌNH CHIỀU DÀI CÁC PHÂN ĐOẠN ADN ĐƯỢC
NHÂN BẢN CÓ CHỌN LỌC .........................................................................106 THẢO LUẬN.........................................................................................................108
Chương 8: TẾ BÀO CHỦ VÀ VECTOR ..........................................................................109
§1. TẾ BÀO CHỦ ..................................................................................................109 §2. VECTOR ..........................................................................................................111 §3. PHƯƠNG PHÁP CHUYỂN VECTOR TÁI TỔ HỢP VÀO TẾ BÀO CHỦ ..122 THẢO LUẬN.........................................................................................................125
Chương 9: PHÂN LẬP GENE TÁCH DÒNG PHÂN TỬ VÀ BIỂU HIỆN GENE......126
§1. PHÂN LẬP GENE ...........................................................................................126 §2. TÁCH DÒNG GENE .......................................................................................127 §3. BIỂU HIỆN GENE...........................................................................................139 THẢO LUẬN.........................................................................................................151 Tài liệu tham khảo chính .....................................................................................152 những công trình của tác giả cùng các cộng tác viên đã công bố.....................155
CƠ SỞ VÀ PHƯƠNG PHÁP SINH HỌC PHÂN TỬ .....................................................162
3

Lời nói đầu Sinh học phân tử hiện đại đang phát triển mạnh và đã trở thành nòng cốt của Công nghệ sinh học. Ở Việt Nam, thành tựu nghiên cứu Sinh học phân tử và áp dụng kĩ thuật phân tử trong nghiên cứu Khoa học sự sốngvà Công nghệ sinh học đã có nhiều đóng góg trong việc chăm sóc, bảo vệ sức khoẻ con người; trong đánh giá tài nguyên sinh vật; trong chọn giống và sản xuất nông lâm ngư nghiệp. Sinh học phân tử là môn Sinh học hiện đại, được giảng dạy ở nhiều trường đại học và đang có những đóng góp nhất định trong đào tạo lớp người có tri thức về Công nghệ sinh học góp phần vào sự nghiệp Công nghiệp hoá, hiện đại hoá Cơ sở và phương pháp Sinh học phân tử được biên soạn từ nhiều tài liệu, bài giảng và công trình nghiêm cứu mới về Sinh học phân tử hiện đại của các tác giả trong và ngoài nước, nhằm cung cấp những kiến thức cơ bản về nguyên lí và ứng dụng của Sinh học phân tử, làm tài liệu cho nghiên cứu, giảng dạy và học tập môn này ở trường đại học. Cuốn Cơ sở và phương pháp Sinh học phân tử được cấu trúc bởi 9 chương: Chương 1. Hệ gene
Chương 2. Đặc điểm cấu trúc gene của sinh vật Prokaryot và Eukryot Chương 3. Mối liên hệ giữa ADN, ARN, Protein
Chương 4. Enzyme sử dụng trong kĩ thuật sinh học phân tử Chương 5. Xác định mối liên quan giữa protein và đặc tính ở sinh vật Chương 6. Kĩ thuật sinh học phân tử trong tích hệ gene ở sinh vật Chương 7. Phản ứng chuỗi polimerase (PCR) Chương 8. Tế bào chủ và Vector Chương 9. Phân lập gene, tách dòng phân tử và biểu hiện gene.
Tác giả trân trọng cảm ơn PGS.TS. Nguyễn Trọng Lạng, TS. Lương Thị Hồng Vân đã đọc và góp ý cho bản thảo, xin cảm ơn những đóng góp của Hội đồng nghiệm thu đánh giá giáo trình của trường Đại học Sư phạm - Đại học Thái Nguyên và cảm ơn các ý kiến đóng góp quý báu của đông đảo các nhà khoa học. Trong quá trình biên soạn chắc chắn có những sai sót, tác giả rất mong nhận được những góp ý của bạn đọc. Mọi đóng góp xin gửi về Khoa Sinh - Kĩ thuật Nông nghiệp trường Đại học Sư phạm - Đại học Thái Nguyên.
Tác giả
4

Chương 1: HỆ GENE Tóm tắt: Sự ra đời của sinh học phân tử được đánh dấu bằng thời điểm mà Oatsơn và Cric (1953) phát hiện ra cấu trúc ADN. Trải qua hơn 40 năm (1953 - 2000) sinh học phân tử đã đạt được những thành tựu vĩ đại mà đỉnh cao của sự phát triển này là những khám phá bản chất sinh học của sự sống ở cấp độ phân tử và xây dựng các kĩ thuật sinh học phân tử ứng dụng váo thực tiễn. Geneomics và Proteomics là vấn đề đang được đặc biệt quan tâm hiện nay mà cơ sở của các lĩnh vực này là những phát hiện về cấu trúc và chức năng của axit nucleic, về đặc điểm của genome nhân, genome ti thể, genome lạp thể. Những điểm khác nhau về cấu trúc và chức năng của các hệ gene cho phép ứng dụng vào thực tế chọn giống và nghiên cứu ở người. Cùng với cấu trúc ADN và ARN còn có đặc điểm của quá trình tái bản ADN và phiên mã cũng được quan tâm, vì nó là cơ sở của những kĩ thuật sinh học phân tử - các thao tác ở ADN và ARN. Nội dung cơ bản của chương đề cập đến những cơ sở của sinh học phân tử. Nội dung của chương gồm 4 vấn đề cơ bản: (1). Khái niệm hệ gene; (2). Axit nucleic; (3). ADN vá tái bản ADN, (4). ARN và cơ chế phân mã.
§1. KHÁI NIỆM HỆ GENE (GENOME) Quá trình sinh trưởng và phát triển của sinh vật trải qua rất nhiều giai đoạn và tất cả quá trình đó đều phụ thuộc vào sự điều khiển của các gene. Cấu trúc, chức năng của tế bào được quyết định trực tiếp bởi protein, đó là sản phẩm cuối cùng của sự biểu hiện gene. Quá trình thể hiện hoạt động của gene qua protein, bị ảnh hưởng rất lớn bởi các yếu tố ngoại cảnh như ánh sáng, chế độ dinh dưỡng, sự cộng sinh và sự tương tác của các gene trong hệ gene của tế bào. Như vậy, giữa các thành phần của hệ gene trong tế bào sống có sự tương tác với nhau và có mối quan hệ với các yếu tố của môi trường. Trong tế bào, bên cạnh genome trong nhân, hệ thống di truyền còn phân bố trong lục lạp, ti thể và plasmid (vi khuẩn). Các cơ quan tử này cùng tham gia vào quá trình sinh tổng hợp protein của tế bào. Tế bào là đơn vị cấu trúc và chức năng cơ bản của các cơ thể sống, tế bào còn là đơn vị của sự di truyền. Hệ gene là toàn hộ các gene trong tế bào của cơ thể sinh vật, hệ gene hứa toàn bộ thông tin di truyền đặc trưng cho từng loài, cho từng cá thể trong loài. Ở sinh vật Prokaryot hệ gene gồm toàn bộ các gene trong tế bào; còn ở Eukaryot
5

hệ gene gồm toàn bộ các gene trong tế bào đơn bội (n). Tế bào đơn bội có một hệ gene, các sinh vật hoặc tế bào lưỡng bội có hai hệ gene, sinh vật đa bội có nhiều hệ gene. Hệ gene của sinh vật Eukaryot bao gồm hệ gene nhân (genome nhân), hệ gene ti thể (genome ti thể), hệ gene lục lạp (genome lục lạp). Hệ gene ở Prokaryot chỉ có một thành phần gồm các đoạn ADN không giống nhau; còn hệ gene ở Eukaryot có ba thành phần ADN không giống nhau: Các đoạn ADN lặp lại nhiều (chiếm khoảng 25%), các đoạn ADN lặp lại ít (30%) và các đoạn ADN không lặp lại (45%). 1.1. Hệ gene nhân Hệ gene nhân là genome lớn nhất trong tế bào xét về mặt khối lượng cũng như số lượng gene mã hoá. ADN nhân được sắp xếp gọn trong nhiễm sắc thể trong sự liên kết với protein chứa histone và không chứa histone. ADN có vai trò mã hoá thông tin di truyền, còn protein thì bảo vệ và tham gia điều khiển sự sao chép, phiên mã được chính xác. Quá trình cơ bản trong phát triển động, thực vật là sự nhân đôi vật chất di truyền và phân đều cho các tế bào con khi tế bào phân chia. Tế bào thực vật chứa một lượng lớn ADN, khối lượng này thay đổi theo loài. Arabidosis thaliana có lượng ADN nhỏ nhất (0.07 picogram). Allium cepa có lượng ADN nhiều hơn 33,5 pg/ haploid genome (hệ gene đơn bội). 1.2. Hệ gene lục lạp Cho đến năm 1962, người ta mới khám phá ra ADN và ribosom có trong lục lạp. Như vậy, lục lạp hoặc trong lạp thể chứa tất cả bộ máy cần thiết để tự mình biểu hiện hoạt động gene. Hệ gene lục lạp là toàn bộ các gene trong lục lạp hay toàn bộ lượng thông tin di truyền chứa trong ADN của lục lạp. Lượng ADN trong lục lạp rất lớn, chiếm tới 15% tổng lượng ADN cơ thể thực vật, ribosom chiếm 60% lượng ribosom của tế bào, ADN lục lạp được cấu tạo bởi các phân tử có cấu trúc mạch vòng, có trọng lượng phân tử 83 - 128 x 106, trong đó gần 85% phân tử ADN là mạch đơn. ADN lục lạp làm khuôn phiên mã tổng hợp mARN chloroplast. Ở đây các phân tử mARN chỉ có khoảng 20 nucleotit, chúng thực hiện sự tổng hợp protein tại chỗ trong chloroplast. Ribosom trong chloroplast là 70S bao gồm 30S và 50S. Sau khi tổng hợp xong protein được vận chuyển đến nơi mà lục lạp cần thiết. Về cơ bản hệ thống di truyền trong lục lạp tương tự với hệ thống di truyền của sinh vật Prokaryot. Do vậy đã có nhiều cuộc tranh luận, có phải vật liệu di truyền của sinh vật tiền nhân là nguồn gốc của ADN lục lạp? Ngoài ra ADN của lục lạp không chứa một số gene đặc trưng cho những sinh vật có nhân thật. Điều này đã tìm thấy trong ADN ở thuốc lá và ngô. Ribosom của lục lạp cũng giống
6

ribosom của sinh vật Procariot: 70S được cấu tạo bởi 2 tiểu phần 50S và 30S. Ribosom của E. coli và lục lạp thực vật có đặc tính miễn dịch giống nhau. Thông qua việc nghiên cứu sản phẩm biểu hiện gene là protein, người ta đã phát hiện giữa hệ gene lục lạp và hệ gene nhân trong tế bào thực vật có mối quan hệ chặt chẽ. - Genome của lục lạp không có khả năng tổng hợp tất cả các protein có trong chúng. - Phần lớn polipeptid của lục lạp được tổng hợp do genome trong nhân tế bào. - Những polipeptid này được chuyển vào lục lạp theo cơ chế sau dịch mã thực hiện qua vỏ lục lạp. - Hệ thống di truyền lục lạp điều khiển tổng hợp các protein cần cho sự phát triển của chúng với số lượng khoảng 100 polipeptid.
Bảng 1.1. Kích thước một số ADN lục lạp (ct. ADN), ADN nhân và vi khuẩn
AND Kích thước (bp) Kích thước vòng (μ m)
Plasmid 1≈ 200 x 103 - Trực khuẩn E-coli (chromosome) 3,8 x 106 - Trùng roi (Euglena) (ct. ADN) 1,4 x 105 44 - 44 Thuốc lá (Tabaco) (ct. ADN) 1,6 x 105 - Ngô (Maize) (ct. ADN) 1,36 x 105 43 Đậu xanh (mungbean) (ct. ADN) 1,21 x 105 39 - Polipeptid được mã hoá tổng hợp và ARN trong lục lạp đảm nhiệm chức năng của cơ quan tử liên quan đến quá trình quang hợp. Ở lúa người ta đã xác định được vị trí của các gene quan trọng điều khiển quá trình quang hợp như rbcl, atpBE, pSbA, pstA, psbA. Các gene này đóng vai trò quan trọng trong quá trình sinh tổng hợp protein ở hệ thống quang hoá II. 1.3. Hệ gene ti thể (Mitochondria Genome)
Để nghiên cứu hệ gene ti thể điều quan trọng đầu tiên là thu được ADN ti thể. Có thể lấy ví dụ ở cây lúa theo Salech và cộng sự (1989), phương pháp phân tích ADN trong ti thể gồm các bước sau: - Li tâm với tốc độ chậm để loại bỏ nhân, lục lạp và các thành phần khác của tế bào. - Li tâm với tốc độ nhanh để tách ti thể. - Li tâm với tốc độ chậm để lấy nhân, lục lạp và các thành phần khác.
7

- Xử lí ADN- ase I để phân giải các chất ADN nhân dính trong ti thể. - Làm sạch ti thể. - Tách chiết mtADN từ ti thể (mtADN). Như vậy người ta có thể thu được ADN của mitochondria tinh sạch phục vụ cho các công tác nghiên cứu tiếp theo. mtADN thể hiện một sự khác biệt rất lớn về mặt kích thước và hình dạng. Đối với động vật, mtADN có dạng vòng và kích thước xấp xỉ 15 - 20kb. Ngược lại mtADN của thực vật có nhiều dạng (vòng.,thẳng vaà(cc dạng khác), cũng như có kích thước lớn hơn nhiều: ở cây bắp cải là 200kb, hoặc ở một loại dưa là 2500 kb. Do vậy genome ti thể thực vật tương đối lớn và gồm một vài phân tử ADN. Lượng mtADN trong thực vật chiếm khoảng 0,5 - 1% lượng ADN của tế bào nhưng nó đóng vai trò sống còn cho sự phát triển và sinh sản thực vật. Chức năng của mtADN giống như ctADN. Nó có khả năng mã hoóatổng hợp một số lượng nhỏ pohlieptid nhưng rất quan trọng.,phục vụ cho những hoạt động của chính nó. Những sản phẩm chính mà mtADN điều khiển tổng hợp là 3 tiểu phần của cytochromoxydaza, 2 tiểu phần của phức chất cytochrom bc và một số thành phần khác (Andre, 1991). Ngoài ra mtADN còn đóng vai trò quan trọng trong hiện tượng bất dục đực tế bào chất (CMS). Đây là một đặc tính được khai thác trong sản xuất các dòng lai ở nhiều cây trồng như lúa, ngô... Spruill và cộng sự (1981) đã chỉ ra ở cây ngô có sự khác nhau giữa mtADN trong tế bào cây thường và trong tế bào cây có hiện tượng CMS. Khi phân tích sản phẩm protein thì những mtADN ở cây CMS có khả năng tổng hợp chuỗi peptid 130000 MW, nhưng nó mất khả năng tổng hợp chuỗi polipeptid 21000 MW ở nguyên sinh chất tế bào cây bình thường. Mặt khác người ta còn thấy hiện tượng bất dục tế bào chất còn liên quan đến những phân tử ADN giống như plasmid tìm thấy trong ti thể. Chẳng hạn ở ngô trong tế bào chất của cây CMS có mang hai phân tử AND mạch thẳng với kích thước 6,2 kb và 5,2 kb. Việc sử dụng code mã hóa tổng hợp protein của mtADN có những điểm cần lưu ý: trong hệ thống hoạt động mã di truyền thông thường code UGA cung cấp tín hiệu kết thúc quá trình tổng hợp một chuỗi polipeptid, nhưng ở ADN của ti thể nó lại là code của tryptophan. Ngược lại ở ADN nhân code CGG mã hoá cho tryptophan thì ở ti thể nó lại mã hoá arginin. Ngoài ra người ta đã phát hiện một dạng mới của mARN trong genome ti thể cây trồng và nó được gọi là ARN - Editing. Đây là một vấn đề thể hiện sự tiến hoá của thực vật bậc cao, giúp cho nó có khả năng
8

thích nghi hơn với điều kiện ngoại cảnh.
§2. AXIT NUCLEIC 2.1. Axit nucleic là vật chất di truyền Axit nucleic bao gồm axit deoxyribonucleie (ADN) hoặc axit ribonucleic (ARN) đều đáp ứng tiêu chuẩn của vật chất di truyền. Có rất nhiều bằng chứng đã chứng tỏ ra axit nucleic là một vật chất di truyền ở cấp độ phân tử. Axit nucleic hấp thụ tia tử ngoại cực đại ở bước sóng 260nm và điều này phù hợp một cách chính xác với bước sóng mà tia tử ngoại có thể gây đột biến tối đa ở tế bào, trong khi đó protein ở bước sóng 280nm. - Năm 1928 F. Grifflth phát hiện thấy nòi phế cầu khuẩn S, khuẩn lạc nhẵn do có vỏ bọc polisaccharid (vi khuẩn gây viêm phổi) của vi khuẩn Diplococus pneumoniae làm chuột chết khi tiêm vào cơ thể chuột. Trong khi đó khuẩn lạc R (khuẩn lạc không có vỏ bọc polisaccharid) không gây độc hại gì. Hỗn hợp các phế cầu khuẩn R còn sống và phế cầu khuẩn S đã chết thì làm cho chuột bị viêm phổi, chết và từ máu của chúng đã phân lập được vi khuẩn S sống. Như vậy tác nhân nào đó từ phế cầu khuẩn đã biến nạp và làm cho phế cầu khuẩn R thành S. Người ta đã chứng minh được rằng: ADN tách từ vi khuẩn S nếu được đưa vào vi khuẩn R thì gây nên sự biến đổi kiểu R → S, phát hiện này được khẳng định ADN là vật chất di truyền. Năm 1944 O. T Avery, Mc Leod và Mc Carti đã chứng minh được rằng tác nhân biến nạp là ADN. Phế cầu khuẩn S bị xử lí bằng protease hoặc ARN-aza thì hoạt tính biến nạp vẫn còn: những nếu phế cầu khuẩn S bị xử lí bởi ADN-aza thì hoạt tính biến nạp không còn nữa.
Hình 1.1. Sơ đồ chứng minh vật chất di truyền là ARN
Năm 1957, H. Fraen Kel - Conrat và B. Singer công bố thí nghiệm ở virut đốm thuốc lá có lõi ARN và vỏ protein, có hai dạng a và b. Các thí nghiệm lắp ráp lõi ARN của dạng này với protein của dạng kia và ngược lại đã thành công tạo ra virut có vỏ và lõi thuộc hai dạng khác nhau. Sau đó đem gây nhiễm vào
9

thuốc lá, kết quả các virut con phân lập được từ vết đốm đều mang cả vỏ protein và lõi ARN thuộc cùng một dạng là dạng của lõi ARN mang nhiễm chứ không phải của vỏ protein. Như vậy thông tin di truyền được chứa đựng trong ARN chứ không phải trong protein. 2.2. Axit nucleic ở virut, Prokaryot và Eukaryot Virut là vật chất sống được cấu tạo rất đơn giản, gồm lõi là axit nucleic và vỏ là protein. Vật chất di truyền của virut có hai loại: ADN và ARN. Đa số các loài virut đều sống kí sinh, khi xâm nhập vào tế bào vật chủ (có thì toàn bộ virut hay chỉ A. nucleic). Virut sống kí sinh ở tế bào Prokaryot được gọi là phage hay bacteriophage (thực khuẩn thể). Một số virut có lõi là ARN chủ yếu kí sinh ở thực vật, còn các virut kí sinh ở động vật và tế bào Prokarvot đa số có lõi là ADN. NST phải tạo ra môi trường vật lí để cho phân tử ADN hoạt động, tác động qua lại với các hệ thống trao đổi chất trong tế bào. Hệ thống sắp xếp cách tổ chức, bố trí ADN trong NST ở những tổ chức sinh vật chưa có nhân chính thức và sinh vật nhân chuẩn có sự khác nhau như thế nào? Người ta đã sử dụng ba hướng chính để nghiên cứu tổ chức ADN trong các đối tượng sinh vật khác nhau, đó là việc sử dụng các thiết bị và kĩ thuật như kính hiển vi điện tử, nhiễu xạ tia X, xử lí bởi enzyme biến đổi cấu trúc NST. Đối với sinh vật Prokaryot, tổ chức ADN trong NST ở dạng nucleoid (vùng nhân). Vùng nhân chứa ADN và ADN được gấp và cuộn thành nhiều vòng xoắn: ADN ở dạng siêu xoắn, tính chất siêu xoắn chịu sự kiểm soát của enzyme topoisomerase. Ví dụ ở E. coli ADN có kích thước 300μ m khi co ngắn kích thước dài 25μ m và tiếp tục co ngắn thì kích thước chỉ còn 1,5μ m. Cách sắp xếp này được phát hiện nhờ enzyme ribonuclease và deoxyribonuclease. Prokaryot chứa ADN trần, chuỗi kép, dạng vòng, ngoài ra vật chất di truyền còn ở plasmid (chiếm l-2%). E. coli chỉ có 1 phân tử ADN dạng vòng chứa 3000-4000 gene. Đối với sinh vật Eukaryot, ADN trong tế bào có cả trong nhân, ti thể và lạp thể và chủ yếu ADN nằm trên nhiễm sắc thể trong nhân tế bào. ADN nhân có dạng mạch kép, chủ yếu là B-ADN. ADN ti thể và ADN lục lạp có dạng kép vòng. Tổ chức ADN trong nhiễm sắc thể ở dạng nucleosome (thể nhân). Mỗi nucleosome gồm phân tử ADN và protein (kiềm) được gọi histon. Histon gồm 8 phân tử (octamer). Phân biệt 5 loại histon với tỉ lệ gần như nhau H1, H2A, H2B, H3, H4. Trong đó có 2 phân tử H3, 2 phân tử H4 liên kết với ở vùng trung tâm; 2 phân tử H2A, 2 phân tử H2B liên kết ở vùng ngoài.
10

Các nucleosome nối với nhau bằng một đoạn nucleotit (ADN) dài 15 - 100 cặp nucleotit. NST có cấu trúc xoắn theo nhiều bậc: tập hợp nhiều nucleosome thành sợi cơ bản có cấu trúc xoắn (100Å), sợi cơ bản tiếp tục xoắn thành solenoit (sợi nhiễm sắc) có đường kính là: 250Å: solenoit cuộn xoắn lần nữa tạo nên ống rỗng có đường kính 2000Å; ống rỗng này cuộn xoắn một lần nữa thành sợi chromatit có đường kính bằng 6000Å.
§3. ADN VÀ TÁI BẢN ADN 3.1. Đặc điểm cấu trúc của phân tử ADN Hàm lượng ADN trong nhân tế bào đặc trưng cho từng loài và phụ thuộc và số lượng nhiễm sắc thể trong nhân. ADN là đại phân tử, có khối lượng và chiều dài rất lớn. Ở sinh vật Prokaryot mỗi nhiễm sắc thể có thểcó nhiều phân tử ADN, còn ở sinh vật Eukaryt mỗi nhiễm sắc thể chỉ có một phân tử ADN.
Bảng 1.2. Hàm lượng ADN của một số loài sinh vật Người Ruồi giấm
Tế bào lưỡng bội (2n) 6,6 pg ADN 2 pg ADN
Tế bào đơn bội (n) 3,3 pg ADN. 1 pg ADN.
1 picogram (pg) ADN = 0,965. 109 bp = 6,1.1011 dalton = 29cm Người ta đã phân biệt các dạng ADN là B, A. Z, C, D. Các dạng AND khác nhau bởi chiều xoắn, chiều cao một chu kì xoắn, số cặp nucleotit trong một chu kì.
11
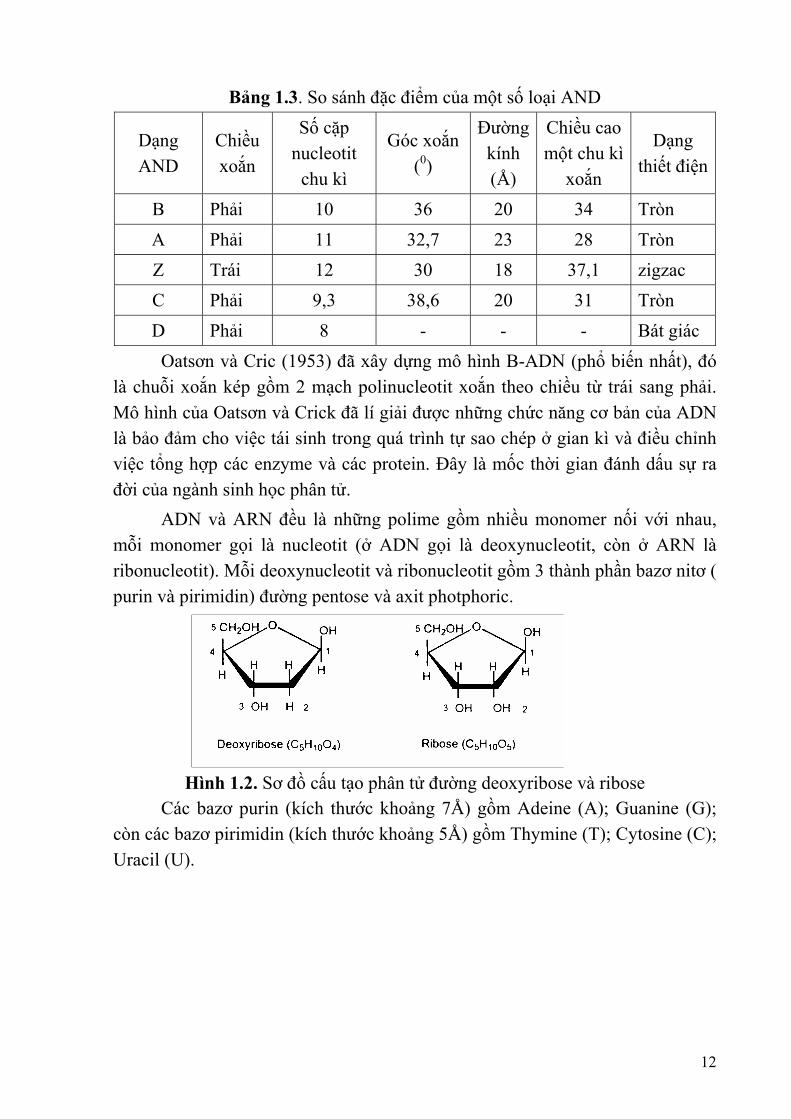
Bảng 1.3. So sánh đặc điểm của một số loại AND
Dạng AND
Chiều xoắn
Số cặp nucleotit
chu kì
Góc xoắn (0)
Đường kính (Å)
Chiều cao một chu kì
xoắn
Dạng thiết điện
B Phải 10 36 20 34 Tròn A Phải 11 32,7 23 28 Tròn Z Trái 12 30 18 37,1 zigzac C Phải 9,3 38,6 20 31 Tròn D Phải 8 - - - Bát giác
Oatsơn và Cric (1953) đã xây dựng mô hình B-ADN (phổ biến nhất), đó là chuỗi xoắn kép gồm 2 mạch polinucleotit xoắn theo chiều từ trái sang phải. Mô hình của Oatsơn và Crick đã lí giải được những chức năng cơ bản của ADN là bảo đảm cho việc tái sinh trong quá trình tự sao chép ở gian kì và điều chỉnh việc tổng hợp các enzyme và các protein. Đây là mốc thời gian đánh dấu sự ra đời của ngành sinh học phân tử. ADN và ARN đều là những polime gồm nhiều monomer nối với nhau, mỗi monomer gọi là nucleotit (ở ADN gọi là deoxynucleotit, còn ở ARN là ribonucleotit). Mỗi deoxynucleotit và ribonucleotit gồm 3 thành phần bazơ nitơ ( purin và pirimidin) đường pentose và axit photphoric.
Hình 1.2. Sơ đồ cấu tạo phân tử đường deoxyribose và ribose
Các bazơ purin (kích thước khoảng 7Å) gồm Adeine (A); Guanine (G); còn các bazơ pirimidin (kích thước khoảng 5Å) gồm Thymine (T); Cytosine (C); Uracil (U).
12
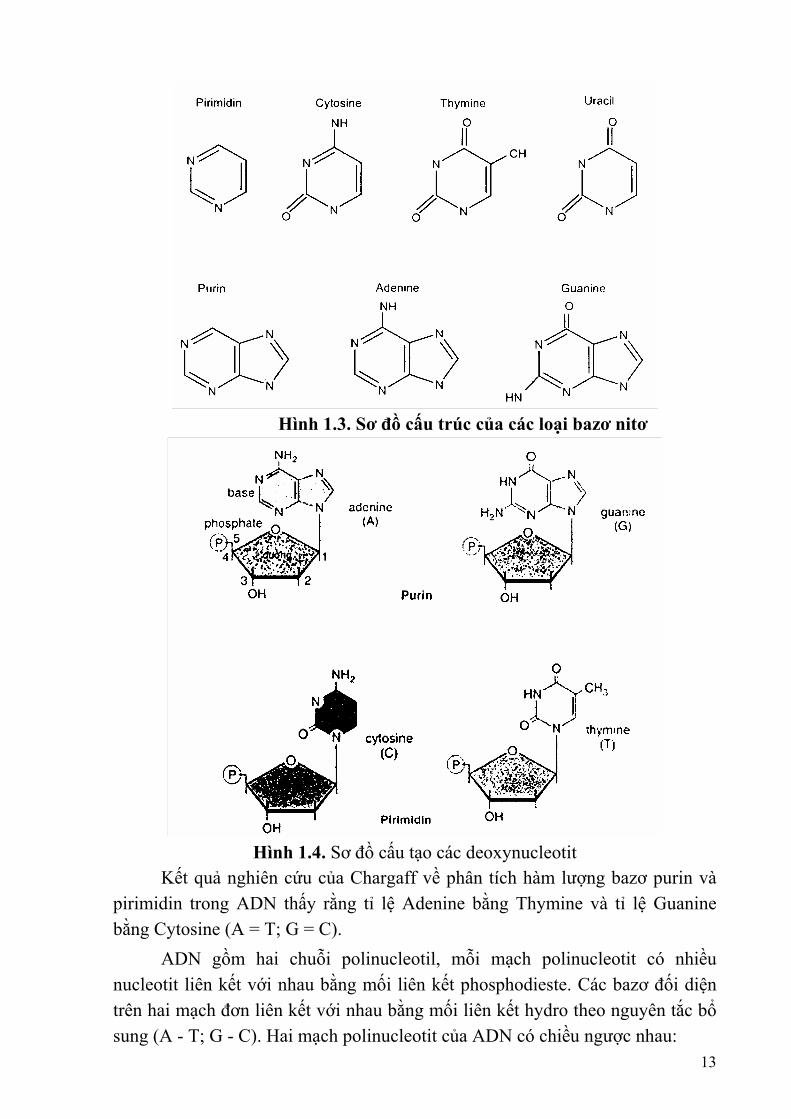
Hình 1.3. Sơ đồ cấu trúc của các loại bazơ nitơ
Hình 1.4. Sơ đồ cấu tạo các deoxynucleotit
Kết quả nghiên cứu của Chargaff về phân tích hàm lượng bazơ purin và pirimidin trong ADN thấy rằng tỉ lệ Adenine bằng Thymine và tỉ lệ Guanine bằng Cytosine (A = T; G = C). ADN gồm hai chuỗi polinucleotil, mỗi mạch polinucleotit có nhiều nucleotit liên kết với nhau bằng mối liên kết phosphodieste. Các bazơ đối diện trên hai mạch đơn liên kết với nhau bằng mối liên kết hydro theo nguyên tắc bổ sung (A - T; G - C). Hai mạch polinucleotit của ADN có chiều ngược nhau: 13

3'OH ⎯→⎯ 5'P
5'P ⎯→⎯ 3'OH
Tính đặc trưng của phân tử ADN phụ thuộc vào 3 yếu tố: số lượng, thành phần và trình tự sắp xếp các nucleotit trong ADN, trong đó phụ thuộc nghiêm ngặt vào trình tự các nucleotit. Kích thước của phân tử ADN được tính bằng bp (base pair- cặp bazơ) hoặc kb (kilobase - kb = 1000 bp), vì tính đặc hiệu của nucleotit là do bazơ, cho nên khi nói đến axit nucleic thì bazơ thường được dùng thay cho nucleotit.
Hình 1.5. Sơ đồ chuỗi polinucleotit của B-ADN 3.2. Tái bản ADN 3.2.1. Các kiểu tái bản ADN Người ta đưa ra 3 giả thuyết về các kiểu tái bản ADN: Kiểu bảo toàn: Chuỗi ADN mẹ giữ nguyên, ADN mới được tổng hợp từ nguyên liệu (chưa có bằng chứng trong tự nhiên). Kiểu phân tán: ADN ban đầu đứt ra từng đoạn nhỏ, mỗi đoạn nhỏ làm khuôn để tổng hợp các đoạn nhỏ khác. Các đoạn nhỏ nối lại với nhau thành ADN. Kiểu bán bảo tồn: Meselson và Stahl chứng minh 1958 ở E. coli sử dụng đồng vị phóng xạ N15. Mathew Meselson và Franklin Stahl (Mỹ - California) tiến hành thí nghiệm nuôi E. Coli với nguồn N15, đã chứng minh được cơ chế tái bản ADN. 3.2.2. Hệ enzyme tái bản ADN
14

• Ba loại enzyme ADN-polimerase ở E. Coli - ADN-poIimerase I: giữ vai trò trong tái bản ADN, có thể dễ sửa chữa. - ADN-poIimerase II: xác định sự bắt đầu tổng hợp ADN. - ADN-poIimerase III: điều khiển tổng hợp ADN, thực hiện nhiệm vụ chủ yếu gia tăng chiều dài của một sợi mới.
• Ba loại enzyme ADN - polimerase ở Eukaryot - ADN-polimerase (120.000 - 300.000 dalton) có chức năng tái bản ADN của nhân.
α
- ADN-polimerase β (30.000 - 50.000 dalton) sửa đổi ADN.
- ADN-polimerase γ (150.000 - 300.000 dalton) tái bản hệ gene ti thể. 3.2.3. Cơ chế tái bản ADN theo Okazaki -Tái bản nửa gián đoạn (Semidiscontinous replication ) 3.2.3.1. Hiện tượng duỗi xoắn
Hiện tượng trên có sự tham gia của một loại protein là SSB (Single Strand Binding) bám vào sợi đơn ADN luôn ở trạng thái mở xoắn, mỗi SSB bám vào 8 bazơ và mỗi chạc tái bản có 250 SSB. 3.2.3.2. Khởi đầu tái bản bằng ARN mồi (primer) ARN polimerase hoạt động tổng hợp đoạn ARN mồi ngắn tạo ra đầu 3'OH tự do, sau đó ADN polimerase III hoạt động tái bản. Đối với E. coli primer được tạo ra bởi enzyme primase. 3.2.3.3. Kéo dài, loại bỏ mồi và hình thành các phân đoạn Okazaki Sau khi ARN mồi được tổng hợp, thì ADN-polimerase I hoạt động loại bỏ mồi nhờ cắt từ (5' - 3') và thay vào đó là một đoạn ADN mới (hình thành phân đoạn Okazaki). Mỗi phân đoạn Okazaki có 1000 - 2500 bazơ. Các phân đoạn Okazaki nối với nhau bằng ADN-ligase. Sợi ADN mới được tổng hợp bằng cách nối các phân đoạn okazaki được gọi là sợi ra chậm (lagging strand) là sợi được tổng hợp không liên tục (gián đoạn); sợi kia được tổng hợp liên tục trên khuôn của sợi ADN có chiều 3' - 5'. Cơ chế tái bản ADN có một sợi được tổng hợp liên tục và một sợi được tổng hợp gián đoạn được gọi là tái bản nửa gián đoạn. - Mạch ADN mới được tổng hợp theo chiều 5' - 3', một mạch polinucleotit được tổng hợp liên tục và một mạch được tổng hợp gián đoạn- Mỗi bước của 15
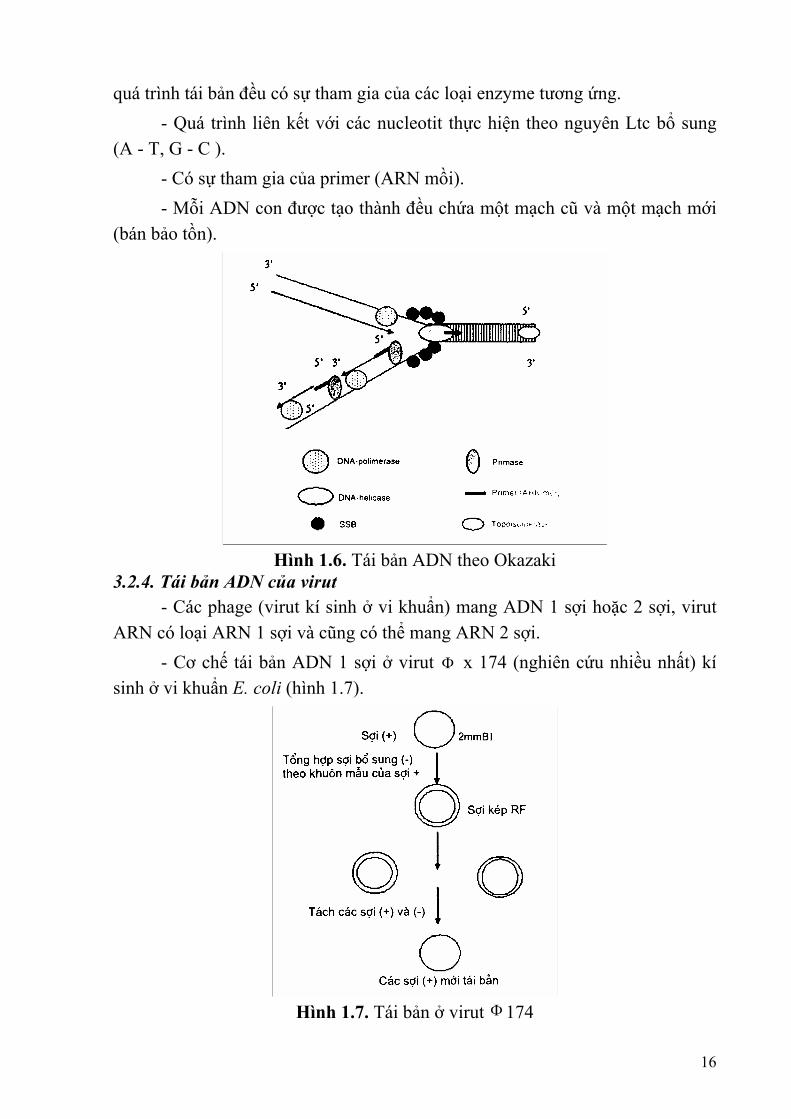
quá trình tái bản đều có sự tham gia của các loại enzyme tương ứng. - Quá trình liên kết với các nucleotit thực hiện theo nguyên Ltc bổ sung (A - T, G - C ). - Có sự tham gia của primer (ARN mồi). - Mỗi ADN con được tạo thành đều chứa một mạch cũ và một mạch mới (bán bảo tồn).
Hình 1.6. Tái bản ADN theo Okazaki
3.2.4. Tái bản ADN của virut - Các phage (virut kí sinh ở vi khuẩn) mang ADN 1 sợi hoặc 2 sợi, virut ARN có loại ARN 1 sợi và cũng có thể mang ARN 2 sợi. - Cơ chế tái bản ADN 1 sợi ở virut Φ x 174 (nghiên cứu nhiều nhất) kí sinh ở vi khuẩn E. coli (hình 1.7).
Hình 1.7. Tái bản ở virut Φ174
16

Chỉ có sợi (+) được tái bản và chỉ có 1 trong 2 sợi trong RF làm khuôn tổng hợp ra sợi (+). 3.2.5. Tái bản ADN ở tế bào của sinh vật Prokaryot 3.2.5.1. Tái bản ADN vòng ở Prokaryot Ở E. coli, quá trình tái bản xuất phát từ một điểm ori (origine) được gọi là điểm xuất phát và triển khai ra cả hai phía. Ở E. coli chỉ có một điểm xuất phát sao chép ori nên cả ADN thành một đơn vị sao chép (replicon). Prokaryot thường chỉ có một replicon (hình 1.8).
Hình 1.8. Đơn vị tái bản (replicon) ở Prokaryot
3.2.5.2. Tái bản kiểu lăn đai thùng (Rolling circle ) - E. coli F+ và F- tiếp hợp. Yếu tố F được truyền từ F+ sang F- và F- biến thành F+, ADN vòng plasmid được tái bản theo kiểu lăn đai thùng tạo thành ADN vòng mới ở tế bào nhận F-.
Hình 1.9. Tái bản kiểu lăn đai thùng (rolling circle)
3.2.6. Tái bản ở tế bào của sinh vật Eukaryot Ở sinh vật Eukaryot sự sinh sản của tế bào là một quá trình sinh trưởng phân nhân và phân bào mang tính chu kì, quá trình đó được gọi là chu kì tế bào (cell cycle). Chu kì tế bào gồm 4 pha: Pha G1 (Gap 1) tiếp sau pha phân chia, nhiễm sắc thể tháo xoắn trở về
17

dạng sợi mánh và xảy ra quá trình tổng hợp các chất cho sự nhân đôi ADN và diễn ra hàng loạt các biến đổi dẫn đến khởi đầu cho sự sao chép ADN. Pha S (Synthesis): vật chất di truyền của chất nhiễm sắc được nhân đôi, đó là quá trình tái bản ADN, các bằng chứng đã chứng tỏ rằng sao chép ADN của sinh vật nhân chuẩn cũng diễn ra theo kiểu nửa gián đoạn như ở sinh vật nhân sơ. Các enzyme cần thiết cho sự sao chép ADN của sinh vật nhân chuẩn bao gồm nhóm ADN-polimerase có ADN polimerase α , ADN-polimerase và ADN-polimerase . Theo quan niệm hiện nay, ở các sinh vật nhân chuẩn bậc cao nói chung - polimerase được sử dụng sao chép ADN của nhân, β -polimerase hoạt động như enzyme sửa chữa,
β
γ
α
γ -polimerase chuyên trách sao chép ADN ti thể.
Hình 1.10. Các sự kiện diễn ra ở mức phân tử trong chu kì tế bào
M. Pha phân chia G1: Pha sau phân chia, chuẩn bị tổng hợp các chất, chuẩn bị tổng hợp ADN S: Pha tổng hợp trong đó diễn ra quá trình tái bản ADN G2: Pha trước khi phân chia tế bào, trong đó diễn ra tổng hợp tubulin và cubuin 1. Bắt đầu phân chia; 2. Kết thúc phân chia;3. Xuất hiện nhân tố cảm ứng tổng hợp ADN
Ở sinh vật nhân chuẩn, mỗi NST có nhiều đơn vị sao chép (Replication Unit) còn gọi là replicon, mỗi replicon có một điểm khởi đầu và hai điểm kết thúc quá trình sao chép. Các NST của sinh vật nhân chuẩn bao gồm ADN và histon cùng nằm trong thể nhân. Những bằng chứng cho thấy các gene mã hoá histon sao mã tạo ra các bản sao để tổng hợp số lượng histon cần thiết cho sự hình thành các thể nhân mới trong pha S.
18

Pha G2 (Gap 2): các nhiễm sắc thể chuẩn bị cho nguyên phân. Một sự kiện quan trọng trong G2 là nơi tổng hợp protein tubulin, cubulin được trùng hợp hoá để tạo ra các vi ống của bộ máy thoi sắc để phân li các NST trong nguyên phân. Pha M (Mitosis): Diễn ra quá trình nguyên phân của tế bào, đặc điểm cơ bản của nguyên phân là có những biến đổi lớn về chức năng và cấu trúc của tế bào. Ở Eukaryot có số lượng ADN lớn, tái bản ADN phức tạp hơn và tốc độ chậm (50 nucleotit/giây). Tái bản của Eukaryot có nhiều replicon và nhiều điểm ori. 3.2.7. Sửa sai trong tái bản Hướng sao chép ADN thực hiện theo chiều từ 5'-3' và hướng sửa chữa sai sót trong tái bản lại diễn ra theo chiều từ 3'-5'. Enzyme ADN-polimerase I, III có hoạt tính của exonuclease (enzyme có hoạt tính cắt nucleotit ở đầu tự do của ADN), nếu có nucleotit lắp sai thì ADN-polimerase sẽ cắt bỏ nucleotit sai đó theo hướng 3'-5'.
Hình 1.11. Tái bản ở Eukaryot (nhiều replicon)
Trong trường hợp ADN ở trạng thái không tái bản thì có xấp xỉ 20 loại enzyme rà soát dọc các NST để tìm ADN có biến đổi cấu trúc, 5 enzyme kiểm soát sự sai sót của liên kết hoá trị, một số enzyme khác phát hiện sai sót bắt cặp giữa các bazơ không bổ sung. Tổng số có xấp xỉ 50 enzyme chuyên phát hiện và sửa các sai sót trong ADN.
§4. ARN VÀ CƠ CHẾ PHIÊN MÃ 4.1. Cấu trúc và chức năng của ARN 4.1.1. Các loại ARN ARN thông tin (iARN; mARN) được cấu tạo từ một mạch poliribonucleotit phổ biến có từ khoảng 600-1500 ribonucleotit. Phân tử mARN có một bộ ba mở đầu (AUG), sau đó đến bộ ba quy định axit amin thứ nhất, thứ hai... cuối cùng là bộ ba kết thúc (UAA, UGA, UAG). Phân tử mARN có chức năng truyền đạt thông tin di truyền từ ADN trong nhân tế bào đến tế bào chất và trực tiếp tham gia tổng hợp protein. Hàm lượng mARN rất ít, chỉ chiếm vài %
19

ARN tổng số trong tế bào. ARN vận chuyển - ARN hoà tan (tARN; sARN): Cấu tạo từ một mạch poliribonucleotit, có khoảng 75-100 ribonucleotit, khối lượng phân tử 26000 dalton. Hàm lượng tARN chiếm khoảng 10-20% ARN tổng số. Phân tử tARN có cấu trúc xoắn tạo ra các thùy tròn, một thùy mang bộ ba đối mã, một thùy nhận biết enzyme gắn với axit amin ở một đầu tARN. Một đầu mút mang axit amin kết thúc là ACC, còn đầu mút kia là GGG. Chức năng của tARN là kết hợp với axit amin để tổng hợp protein. Phân tử tARN có 2 chức năng trong sự tham gia tổng hợp protein là tiếp nhận và liên kết.
Hình 1.12. Mô hình cấu trúc không gian và cấu trúc phân tử của tARN
ARN ribosom chiếm phần lớn trong tế bào, khoảng 80% hàm lượng ARN tổng số. Các phân tử rARN cùng với protein cấu tạo nên các ribosome. Bản chất hoá học của ribosome là nucleoprotein, gần 36% protein và 64% rARN. 4.1.2. Cấu trúc của ARN Các loại ARN đều chỉ có một chuỗi poliribonucleotit, mỗi ribonucleotit gồm 3 thành phần (H3PO4; C5H10O5; bazơ nitơ: A, U, G, C). Các ribonucleotit liên kết với nhau bằng mối liên kết phosphodieste, để tạo thành chuỗi poliribonucleotit, có chiều 5' 3'. →
Tất cả các loại ARN đều được tổng hợp từ khuôn mẫu của ADN và chỉ có cấu tạo sợi đơn. Phân tử ARN của virut là genome của chúng, có chức năng duy trì và truyền đạt thông tin di truyền. Riêng các retrovirut mang ARN sợi kép. 4.2. Cơ chế phiên mã 4.2.1. Phiên mã ở sinh vật nhân sơ 4.2.1.1. ARN-polimerase của sinh vật nhân sơ Ở E. coli, ARN-polimerase có hệ số lắng 11S - 13S và khối lượng phân tử 500.000 dalton; từ enzyme ARN polimerase nhân tố sigma ( ) có thể kết hợp σ
20

với enzyme lõi khác.
Hình 1.13. Enzyme ARN -polimerase
Nhân tố σ đóng vai trò quan trọng trong việc tổng hợp ARN, nó giúp cho enzyme lõi có thể khởi đầu tổng hợp ARN tại những điểm đặc thù - gọi là điểm khởi đầu (promotor), nếu thiếu nhân tố ơ thì quá trình tổng hợp ARN sẽ xảy ra ở điểm ngẫu nhiên trên phân tử ADN. Đoạn nhận biết (recognition sequence) gồm 35 cặp bazơ nằm trước điểm khởi đầu promotor. Đoạn để ARN-polimerase nhận biết là đoạn gồm 7 nucleotit được gọi prinow, nằm cách điểm phiên mã 5 - 6 bazơ. Hộp prinow trên sợi đối khuôn (antitemplate) có công thức chung là: 5' TAT phun AT purin 3'. 4.2.1.2. Các giai đoạn của quá trình phiên mã Quá trình phiên mã của Prokaryot bao gồm 3 bước: khởi đầu, kéo dài và kết thúc.
1) ARN-polimerase lõi cùng nhân tố ơ nhận biết và bám vào trình tự khởi động (promotor). ADN polimerase trượt dọc gene (3' - 5') xúc tác biến tính cục bộ 2 sợi ADN lộ ra sợi khuôn tổng hợp ARN.
2) Quá trình tổng hợp ARN bắt đầu và sợi ARN được sinh ra theo chiều 5' - 3'.
3) Đoạn kết thúc ra tín hiệu cho ARN-polimerase dừng phiên mã. Đoạn kết thúc gồm 2 phần: phần có tỉ lệ GC cao và một loại bazơ T trên sợi antitemplate.
4.2.1.3. Đặc điểm Sản phẩm của quá trình phiên mã là sợi đơn ARN. ADN dãn xoắn cục bộ, chỉ có 1 sợi làm khuôn tổng hợp ARN và được gọi là sợi có ý nghĩa (strand sense). Nguyên liệu là các ribonucleotit triphotphat : ATP, GTP, CTP, UTP gọi chung là NTP, enzyme xúc tác là ARN-polimerase. ARN được tổng hợp theo chiều 5' - 3', enzyme ARN-polimerase di chuyển theo chiều 3' - 5' trên sợi strand sense. 4.2.2. Phiên mã ở sinh vật Eukaryot 21

4.2.2.1. ARN polimerase • ARN-poIimerase I: enzyme không bị ức chế bởi a-amanitin (chất ức chế
tổng hợp ARN), chuyên trách việc phiên mã các gene để tổng hợp rARN (28S, 58S và 18S).
• ARN-polimerase II: enzyme bị ức chế bởi α - amanitin, nó cần cho tổng hợp mARN.
• ARN-polimerase III: enzyme bị ức chế bởi α - amanitin ở nồng độ cao, phiên mã tổng hợp các tARN 4S, rARN 5S.
4.2.2.2. Các đoạn kiểm soát phiên mã Hộp TATA hoặc hộp Goldberg - Hogness ở vị trí cách cặp bazơ đầu tiên được phiên mã khoảng 30 cặp bazơ về phía trước được coi là đoạn khởi đầu. Đoạn kết thúc còn nghiên cứu ít, đoạn kết thúc không dài hơn 21 bazơ, ở nấm men có trình tự là: TTTTATA. 4.2.2.3. Các giai đoạn của quá trình phiên mã ở Eukaryot
• Giai đoạn khởi động: chịu sự kiểm tra của một trình tự đặc biệt đó là "hộp TATA" (TATA box) nằm trước vị trí bắt đầu phiên mã khoảng 25 - 35 nucleotit. ARN-polimerase II hoạt động phiên mã nhờ nhiều nhân tố TF có bản chất là protein (trascription factor) cho phép khởi động sự phiên mã.
• Giai đoạn kéo dài: phân tử ARN được tổng hợp từ mạch khuôn ADN. Quá trình này nhờ nhân tố TF IIS.
• Giai đoạn kết thúc: phiên mã kết thúc trước điểm gắn đuôi poli A rất xa. Kết thúc phiên mã có liên quan đến cấu trúc kẹp tóc tiếp ngay sau trình tự giàu GC.
4.2.2.4. Diễn biến quá trình phiên mã 1) Gắn chóp: khi pre mARN đang tạo ra dài 20 - 30 bazơ thì ở đầu 5'P nối
thêm 7-methylguanosine liên kết 5'P - 5'P. Bản phiên mã đầu tiên là tiền mARN gồm cả exon và intron.
2) Thêm đuôi poli A: một đoạn ngắn của mARN bị cắt ra và các bazơ adenine nối vào thành đuôi poli A.
3) Splicing: cắt rời các intron và nối các exon nhờ phức hợp ribonucleoprotein (SnRNP) của nhân, tạo cấu trúc không gian thuận tiện cho các đầu exon gần nhau và xúc tác phản ứng cắt nối hình thành ARN trưởng thành.
22

Hình 1.14. Sơ đồ các giai đoạn của quá trình phiên mã ở Eukayot 4.2.2.5. Đặc điểm của sự phiên mã ở Eukaryot
• ARN-polimerase II chịu trách nhiệm tổng hợp mARN, còn 2 loại enzyme khác tổng hợp rARN và tARN.
• mARN, rARN và tARN chứa thông tin ở dạng bản sao của một gene.
• Quá trình phiên mã phức tạp, ở đầu 5' của mARN có gắn thêm 1 "chóp" (cap: chóp, chụp đèn) là 7-methylguanosine, còn cuối mARN có "đuôi" poliadenine dài 100 - 200 A. Chóp - Methylguanosine và đuôi poliA có tác dụng bảo vệ ARN, ngoài ra đuôi poliA còn là tín hiệu nhận biết mã kết thúc trong quá trình dịch mã.
Bản phiên mã đầu tiên là tiền mARN (premessager ARN-Pre-ARN) bao gồm cả các đoạn intron và exon, sau đó sau đó các intron bị cắt bỏ tạo thành mARN trưởng thành.
THẢO LUẬN 1. Phân biệt các khái niệm: genome, genome nhân, genome ti thể, genome
lạp thể. 2. Đặc điểm về vật chất di truyền của Virut, Prokaryot, Eukaryot. 3. Vấn đề cấu trúc của hệ gene ti thể. Những ứng dụng nghiên cứu hệ gene ti
thể ở cây trồng và ở người. 4. Đặc điểm cấu trúc và chức năng của ADN và ARN. 5. Cơ chế tái bản của ADN. Tái bản ADN ở Virut, Prokaryot, Eukaryot. Tái
bản ADN và tại sao trong tái bản ADN lại cần phải có primer? Tại sao primer là một trình tự ARN?
6. Đặc điểm của cơ chế phiên mã ở Prokaryot, Eukaryot. Vấn đề phiên mã 23

ngược và ứng dụng trong thực tiễn.
24

Chương 2: ĐẶC ĐIỂM CẤU TRÚC GENE CỦA SINH VẬT PROKARYOT VÀ EUKARYOT
Tóm tắt: Những thao tác trên gene được dựa trên cơ sở những hiểu biết về đặc điểm cấu trúc của gene. Gene ở nhóm sinh vật Prokaryot và nhóm Eukaryot có những đặc điểm cấu trúc đặc trưng. Gene của đa số sinh vật Prokaryot có cấu trúc liên tục, bao gồm các trình tự mã hoá axit amin; gene của sinh vật Eukaryot có cấu trúc gián đoạn (gene phân mảnh) gồm các intron và exon xen kẽ nhau. Đặc điểm cấu trúc này đã quyết định sự khác nhau giữa quá trình phiên mã ở Prokaryot và Eukaryot. Các trình tự như promotor, operator, enhancer, yếu tố di truyền vận động (TGE), đoạn xen (IS), trình tự lặp lại CEN, TEL có cấu trúc và chức năng xác định trong hệ thống di truyền của tế bào. Hiện tượng biến tính, hồi tính của ADN, nhiệt độ chảy Tm cũng là cơ sở của một số kĩ thuật di truyền như lai phân tử, PCR, và một số kĩ thuật khác. Nội dung của chương gồm 3 vấn đề cơ bản : (1). Đặc điểm cấu trúc gene của sinh vật Prokaryot; (2). Cấu trúc phân đoạn gene của sinh vật Eukaryot; (3). Một số trình tự ADN.
§1. ĐẶC ĐIỂM CẤU TRÚC GENE Ở PROKARYOT Gene của đa số sinh vật Prokaryol gồm nhiều cistron (đa cistron), trình tự promotor, operator. Gene cấu trúc của Prokaryot có cấu trúc liên tục, bao gồm những đoạn mã hoá axit amin. Promotor là trình tự trên phân tử ADN có ái lực với enzyme ARN-polimerase, có chức năng khởi động quá trình phiên mã. Operator là trình tự có ái lực với protein ức chế, nếu operator gắn với protein ức chế sẽ ngăn cản hoạt động của ARN-polimerase và ức chế quá trình phiên mã. Mỗi cistron chứa thông tin của một loại chuỗi polipeptit và cũng là nhân tố điều khiển quá trình tổng hợp chuỗi polipeptit thông qua quá trình phiên mã và dịch mã.
§2. CẤU TRÚC PHÂN ĐOẠN GENE Ở EUKARYOT 2.1. Cấu trúc gene phân đoạn (gene phân mảnh) Một gene ở Eukaryot có cấu trúc bao gồm đoạn tăng cường (enhancer) đoạn khởi động (promotor), intron và kẽ nhau, cuối cùng là đoạn kết thúc. hoá (Noncoding Sequences) được gọi là intron hay gọi là đoạn xen (Intervening 25

Sequences) (IS) xen vào và làm gián đoạn các đoạn mã hoá (Coding Sequences) được gọi là exon.
Hình 2.1. Cấu trúc gene phân đoạn ở Eukaryot Những gene có cấu trúc gồm exo được gọi là gene khảm (mosaic gene). Hiện nay, chưa rõ chức năng của intron, tuy nhiên có giả thuyết rằng, intron có vai trò như là các khoảng trống xen giữa các exon tạo thuận lợi cho sự tái tổ hợp giữa các exon. Thí nghiệm của Pierre Chambon (Pháp) phát hiện cấu trúc của exon - intron của gene ovalbumine gà. Ovalbumine là protein lòng trắng trứng. gồm một chuỗi polipeptit có 386 axit amin (tập hợp trong ống dẫn trứng ở thời kì gà mái đẻ trứng). Ngoài việc tách mARN của ovalbumine, sử dụng enzyme sao mã ngược tạo ra các bản sao của mARN sợi đơn được gọi là cADN và tạo thành cADN sợi kép. So sánh ADN hệ gene nhân với cADN khi giải trình tự cho phép phát hiện ra intron và exon. Kết quả ovalbumine trứng gà có 7 intron, 8 exon. 2.2. Phương pháp phát hiện các đoạn intron và exon 2.2.1. Phương pháp so sánh trình tự gene và trình tự cADN
Bước 1: 1. Tách chiết ADN hệ gene. 2. Phân lập gene bằng kĩ thuật PCR hay enzyme cắt hạn chế. 3. Tạo dòng ADN tái tổ hợp, nhân dòng và tách dòng gene. 4. Giải trình tự gene. Bước 2: 1. Tách mARN trong tế bào chất (hàm lượng mARN rất lớn, mARN của ovalbumine chiếm tới 50% ARN trong các tế bào này). 2. Sử dụng enzyme sao mã ngược tổng hợp cARN. 3. Giải trình tự cARN. Bước 3: So sánh trình tự ADN hệ gene với trình tự cADN xác định được các đoạn intron và exon.
2.2.2. Phương pháp lai phân tử Phát hiện intron và exon còn được tiến hành theo phương pháp lai phân tử
26

và kiểm tra bằng kính hiển vi điện tử. 1. Tách ADN hệ gene và mARN tế bào chất. 2. Phân lập gene. 3. Lai phân tử giữa ADN và mARN sẽ xác định được intron và exon.
Vấn đề đặt ra là khi phiên mã tổng hợp mARN thì các đoạn không mã hoá intron có được sao không ? Người ta thấy bản sao của ADN có cả exon và intron đó chính là tiền mARN (Pre-mARN), còn gọi là ARN không đồng nhất trong nhân (hn-ARN: heteroge nevusnucliar ARN). Loại hn-ARN chỉ được phát hiện ở sinh vật nhân chuẩn.
§3. MỘT SỐ TRÌNH TỰ ADN 3.1. Promotor Thực chất của khởi đầu phiên mã là quan hệ tương tác giữa ARN-polimerase với trình tự promotor. Khi ARN-polimease gắn vào promotor thì quá trình phiên mã được thực hiện và kết quả sẽ tạo phân tử mARN.
• Phần lớn các promotor ở E. coli về căn bản có cùng một cấu trúc theo sơ đồ sau:
ADN
sao mã hn-ARN tiền mARN
processing
cắt nối mARN
-35 bp -10 bp +1
TTGACA mARN TATAAT
Nếu bazơ đầu tiên được phiên mã ra mARN (thường là A) được đánh số +1 và tất cả các bazơ phía 5' hay phía trước nó không được phiên mã được ghi là (-). Ngay trước +l có 6 bazơ thường là trình tự TATAAT ở xung quanh -10, gọi là trình tự nhất trí (consensus sequense), có thể gọi pribnow box. Xung quanh trình tự -35 có trình tự nhất trí TTGACA. Cả hai trình tự phối hợp với nhau cho phép ARN-polimerase gắn vào promotor và quá trình phiên mã và dịch mã bắt đầu. Promotor ở Eukaryot nằm phía trước điểm xuất phát của mARN. Hộp TATA, định hướng cho ARN-polimerase bắt đầu phiên mã, nằm khoảng dưới
27

30 bp (động vật có vú) và 6-120 bp (nấm men). Trước hộp TATA có hai trình tự tương ứng phía trước khoảng 40 bp là CCAAT và 110 bp là trình tự giàu GC.
-110 bp -40 bp -30 bp +1
GGGCGGG CCAAT TATA mARN
Sự thay đổi hộp TATA làm giảm tốc độ phiên mã. Hiệu quả của tốt độ phiên mã đo bằng sự thay đổi từng bazơ trong promotor. Hộp TATA và các trình tự phía trước phải được nhận biết bởi các protein điều hoà, chính các protein này gắn với các điểm nhất định trên chúng và hoạt hoá sụ phiên mã. 3.2. Enhancer (trình tự tăng cường) Enhancer là trình tự có tác động cis (đều phía) có chức năng làm tăng tốc độ phiên mã, enhancer có hiệu quả ngay cả khi cách xa vài nghìn cặp bazơ và chúng hoạt động bất kì ở hướng nào, dù là ở trước hay sau promotor. Trình tự tham gia điều hoà - cis có cấu trúc gồm 2 phần đối xứng nhau:
AGGTCA TGACCT TCCAGT ACTGGA
Enhancer được tổ chức gồm một dãy các trình tự có tác động cis để nhận biến các nhân tố trans - nhân tốprotein tham gia điều hoà biểu hiện gene. Nhân tố trans gồm 2 vùng cấu trúc chức năng: vùng gắn trans với ADN và vùng tác động lên phiên mã. Các nhân tố trans có 4 kiểu tác động như:
"Ngón tay kẽm" (zine-finger) "Xoắn-vòng-xoắn" (hehx-turn-hehx) "Xoắn-nút-xoắn" (hehx-loop-helix) "Dây kéo leucine" (leucine-zipper)
Giữa trình tự cis và nhân tố trans thường có các tương tác đặc hiệu do những liên kết yếu hình thành giữa các phân tử của nhân tố trans với các bazơ của trình tự cis. Các trình tự cis thường tiếp nhận protein điều hoà (nhân tố trans) gồm 2 tiểu đơn vị (dimer). Nhân tố trans có thể có 4 nhóm cấu trúc, chúng luôn gắn lên cis. Điều hoà bằng cách biến đổi nhiễm sắc chất hay ADN. Nhiễm sắc thể tồn tại những vùng "nhạy cảm" tương ứng với các gene hoạt động. Các vùng nhạy cảm này dễ tiếp xúc với enzyme sao mã. Tóm lại, các gene của Eukaryot được hoạt hoá bởi hai trình tự ADN có tác động cis là promotor và enhancer. Chúng được nhận biết các nhân tố protein 28

có Lác động trans. Các nhân tố trans này cho phép ARN-polimerase khởi động phiên mã và đạt tốc độ phiên mã tối đa. 3.3. Yếu tố di truyền vận động (Transposable Genetic Element-TGE) Gene nhảy (Jumping genes -JG) Gene nhảy (Jumping genes) còn dược gọi là yếu tố di truyền vận động (TGE) lần đầu tiên được phát hiện bởi Barbara Mc Clintock khi nghiên cứu ở ngô vào những năm 40 của thế kỉ XX và bà đã được nhận giải thưởng Nobel 1983. TGE là những đoạn ADN đặc biệt xen vào một hoặc một số vị trí trong hệ gene, tạo nên những biến đổi di truyền. Loại TGE đơn giản nhất là các đoạn xen (Insertion Sequence - IS ). IS cần thiết cho quá trình xen ADN vào NST, cho quá trình chuyển TGE từ vị trí này sang vị trí khác trong hệ gene. Cấu trúc của IS giống nhau ở những sinh vật khác nhau. Người ta đã phát hiện được ở E. coli một đoạn xen IS1 chứa 720 bp (base pair), nằm giữa đoạn đảo ngược IR (24 bp)
Hình 2.2. Đoạn xen IS1 ở E. coli
Ở E. coli còn phát hiện ra gene nhảy mang tên Tn 1681 (Transposase) gồm 2 đoạn xen IS1 ở hai đầu và 1 gene khác dài 552 bp quy định độc tố chịu nhiệt gây ỉa chảy.
Hình 2.3. Gene nhảy Tn 1681 ở E. coli
Gene nhảy Tn ở E. coli có thể chuyển vị trí và xen vào một nhiễm sắc thể khác theo cơ chế được biểu diễn bằng mô hình ở hình 2.4. 3.4. Biến tính và hồi tính Hai mạch ADN liên kết với nhau bằng những liên kết hyđro. Khi nhiệt độ lên tới 80-950C thì hai mạch ADN tách nhau, được gọi là hiện tượng biến tính của ADN.
29
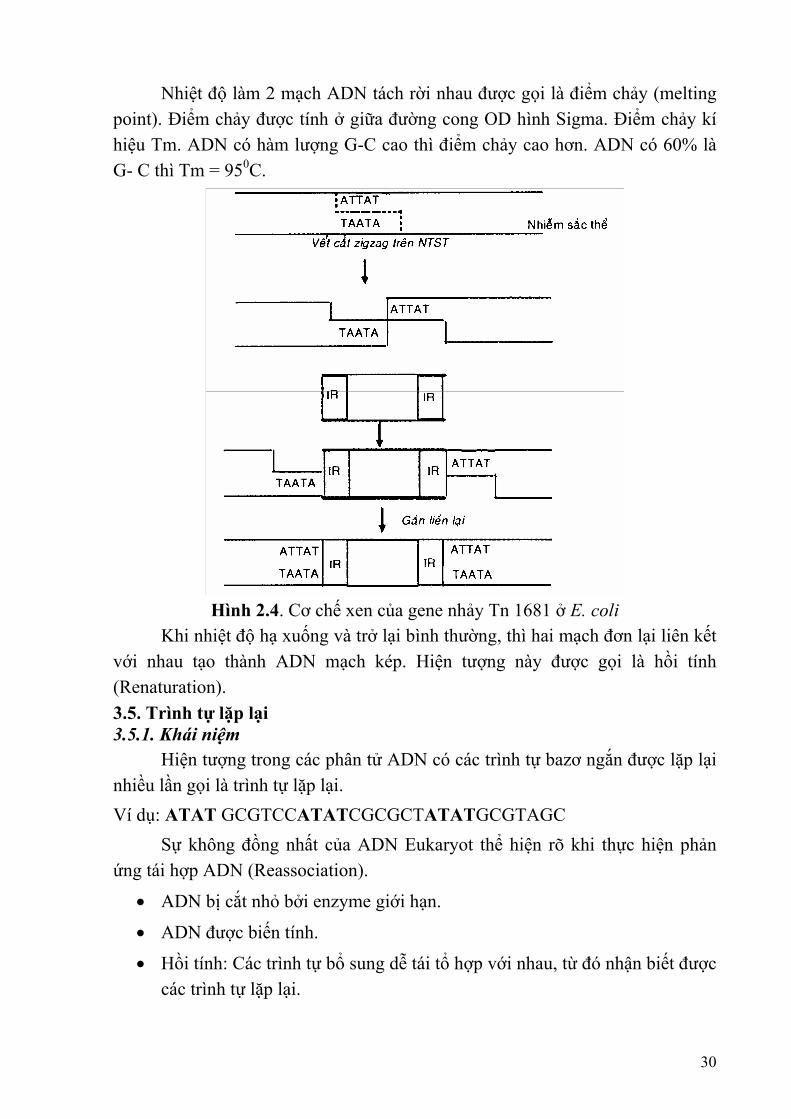
Nhiệt độ làm 2 mạch ADN tách rời nhau được gọi là điểm chảy (melting point). Điểm chảy được tính ở giữa đường cong OD hình Sigma. Điểm chảy kí hiệu Tm. ADN có hàm lượng G-C cao thì điểm chảy cao hơn. ADN có 60% là G- C thì Tm = 950C.
Hình 2.4. Cơ chế xen của gene nhảy Tn 1681 ở E. coli
Khi nhiệt độ hạ xuống và trở lại bình thường, thì hai mạch đơn lại liên kết với nhau tạo thành ADN mạch kép. Hiện tượng này được gọi là hồi tính (Renaturation). 3.5. Trình tự lặp lại 3.5.1. Khái niệm Hiện tượng trong các phân tử ADN có các trình tự bazơ ngắn được lặp lại nhiều lần gọi là trình tự lặp lại. Ví dụ: ATAT GCGTCCATATCGCGCTATATGCGTAGC Sự không đồng nhất của ADN Eukaryot thể hiện rõ khi thực hiện phản ứng tái hợp ADN (Reassociation).
• ADN bị cắt nhỏ bởi enzyme giới hạn.
• ADN được biến tính.
• Hồi tính: Các trình tự bổ sung dễ tái tổ hợp với nhau, từ đó nhận biết được các trình tự lặp lại.
30
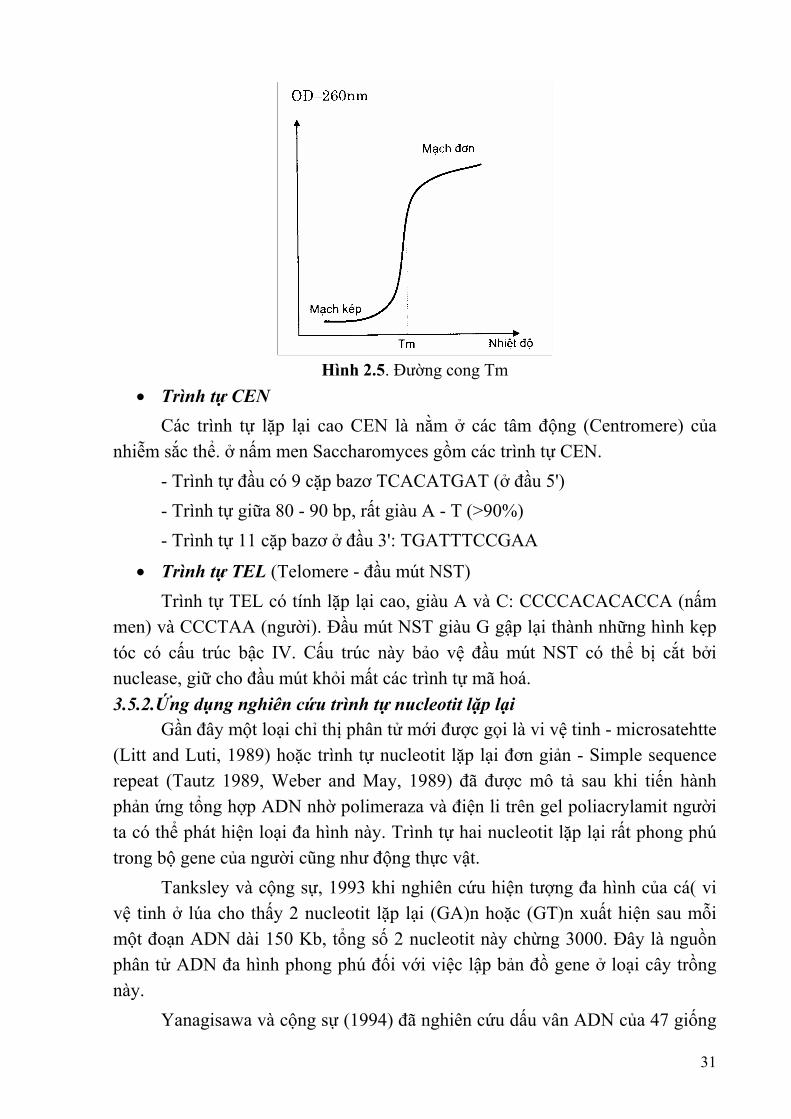
Hình 2.5. Đường cong Tm
• Trình tự CEN Các trình tự lặp lại cao CEN là nằm ở các tâm động (Centromere) của nhiễm sắc thể. ở nấm men Saccharomyces gồm các trình tự CEN. - Trình tự đầu có 9 cặp bazơ TCACATGAT (ở đầu 5') - Trình tự giữa 80 - 90 bp, rất giàu A - T (>90%) - Trình tự 11 cặp bazơ ở đầu 3': TGATTTCCGAA
• Trình tự TEL (Telomere - đầu mút NST) Trình tự TEL có tính lặp lại cao, giàu A và C: CCCCACACACCA (nấm men) và CCCTAA (người). Đầu mút NST giàu G gập lại thành những hình kẹp tóc có cấu trúc bậc IV. Cấu trúc này bảo vệ đầu mút NST có thể bị cắt bởi nuclease, giữ cho đầu mút khỏi mất các trình tự mã hoá. 3.5.2.Ứng dụng nghiên cứu trình tự nucleotit lặp lại Gần đây một loại chỉ thị phân tử mới được gọi là vi vệ tinh - microsatehtte (Litt and Luti, 1989) hoặc trình tự nucleotit lặp lại đơn giản - Simple sequence repeat (Tautz 1989, Weber and May, 1989) đã được mô tả sau khi tiến hành phản ứng tổng hợp ADN nhờ polimeraza và điện li trên gel poliacrylamit người ta có thể phát hiện loại đa hình này. Trình tự hai nucleotit lặp lại rất phong phú trong bộ gene của người cũng như động thực vật. Tanksley và cộng sự, 1993 khi nghiên cứu hiện tượng đa hình của cá( vi vệ tinh ở lúa cho thấy 2 nucleotit lặp lại (GA)n hoặc (GT)n xuất hiện sau mỗi một đoạn ADN dài 150 Kb, tổng số 2 nucleotit này chừng 3000. Đây là nguồn phân tử ADN đa hình phong phú đối với việc lập bản đồ gene ở loại cây trồng này. Yanagisawa và cộng sự (1994) đã nghiên cứu dấu vân ADN của 47 giống
31

đậu tương thông qua chất dò Oligonucleotit nhằm phát hiện trình tự ADN lặp lại đơn giản và cho biết hiện trạng đa hình thể hiện rõ nhất là khi dùng Oligolucleotit (AAT)6 làm đoạn dò ADN tất cả giống đậu tương thử nghiệm đều phân biệt với nhau bởi đoạn dò đó. Khi sử dụng ba đoạn dò ADN khác nhau như (CT)8, (GAA)5 và (AAGG)4 thì không phát hiện được các băng ADN đa hình từ các giống đậu tương thuộc Subgeneus Soja (Glicine max và Glicine Soja), trái lại các băng ADN đa hình xuất hiện trong số các giống thuộc Subgeneus, Glicine. Kết quả nghiên cứu đã chứng tỏ G. max và G. Soja gần gũi nhau về cấu trúc bộ gene.
THẢO LUẬN 1. Phân biệt cấu trúc của gene ở sinh vật Prokaryot và Eukaryot. Vấn đề về
đặc điểm của gene ở sinh vật Eukaryot. 2. Phương pháp xác định các đoạn intron và exon trong gene của sinh vật
Eukaryot. Cho ví dụ. 3. Phân biệt khái niệm về gene nhảy và đoạn xen. Cơ chế xen của gene nhảy. 4. Trình bày các khái niệm: trình tự lặp lại, biến tính và hồi tính.
Ứng dụng của các kĩ thuật này trong phân tích sinh học phân tử.
32

Chương 3: MỐI LIÊN HỆ GIỮA ADN, ARN, PROTEIN Tóm tắt: Thông tin cấu trúc của phân tử protein, tARN, rARN được mã hoá trong gene bằng trình tự các nucleotit. Trình tự nucleotit trong gene mã hoá cho chuỗi polipeptit đã được xác lập bằng thực nghiệm và thể hiện trong bảng mã di truyền. Mã di truyền là mã bộ ba nucleotit (codon), sự thay đổi trong codon có thể dẫn đến sự thay đổi trong chuỗi polipeptit và đột biến ở tính trạng. Những đặc điểm về cấu trúc vá chức năng của protein và mối liên hệ giữa ADN, ARN, protein và sự điều hoà biểu hiện gene cũng là cơ sở quan trọng để xây dựng kĩ thuật biểu hiện gene. Nội dung của chương gồm 4 vấn đề cơ bản: (1). Thông tin sinh học và mật mã di truyền; (2). Protein; (3). Tổng hợp protein; (4). Điều hoà biểu hiện gene.
§1. THÔNG TIN DI TRUYỀN VÀ MẬT MÃ DI TRUYỀN Thông tin quy định cấu trúc một loại protein, một loại tARN, rARN được gọi thông tin di truyền. Thông tin di truyền được chứa đựng và được mã hoá trong phân tử ADN bởi trình tự các nucleotit. Hiện tượng 3 nucleotit kế tiếp nhau trong gene quy định một axit amin trong chuỗi polipeptit được gọi là sự mã hoá bộ ba. Còn một bộ ba cụ thể xác định cho một axit amin cụ thể ( ví dụ: UGU: serine) được gọi là bộ ba mã hoá (codon - code). Phân tử tARN có một bộ ba bổ sung với bộ ba mã sao của mARN được gọi là bộ ba đối mã (anticodon) ADN chứa thông tin di truyền, đó là thông tin quy định cấu trúc một loại protein, rARN và tARN. Trình tự của các bazơ trong ADN quyết định trình tự của axit amin trên protein tương ứng. Phân tử ADN được cấu tạo từ 4 loại nucleotit, và protein lại được cấu tạo từ 20 loại axit amin, nếu, một nucleotit quy định 1 loại axit amin sẽ tạo ra 4 tổ hợp mã; còn hai nucleotit quy định 1 loại axit amin sẽ tạo ra 42 = 16 tổ hợp mã bộ 2, như vậy vẫn còn thiếu 4 loại axit amin chưa được xác định bởi các mã bộ hai. Và ba nucleotit quy định 1 loại axit amin sẽ tạo ra 43 = 64 tổ hợp mã bộ 3. Người ta đã chứng minh bằng thực nghiệm: mã di truyền là mã bộ ba. Đối tượng là gene rII ở phage T4. Trong thí nghiệm người ta tạo đột biến mất (-) và thêm (+) một cặp bazơ trên ADN → biến đổi ADN → thành phần axit amin của protein. Giả sử phân tử mARN chỉ có thành phần CAG và trong phân tử protein chỉ có một loại axit amin:
33

mARN CAG CAG CAG CAG CAG Protein a1 a1 a1 a1 a1 - Nếu đột biến làm mất C (-) ở vị trí mã số 2 trên mARN: mARN CAG AGC AGC AGC AGC protein a1 a2 a2 a2 a2 - Nếu đột biến (+) thêm G vào giữa bộ ba thứ 2: mARN CAG AGC GAG CAG CAG CAG protein a1 a2 a3 a1 a1 a1 Các số liệu thực nghiệm thu được trên thí nghiệm rII ở thực khuẩn T4 đã chứng tỏ lập luận trên là đúng. Vậy mã di truyền là mã bộ ba nucleotit. 1.1. Giải mã di truyền Năm 1961 M. Nirenberg sử dụng hệ thống vô bào để giải mã di truyền. Hệ thống vô bào được chiết từ E. coli có chứa ribosome, ARN, enzyme aminoacylsynthetase, mARN, các axit amin và một số phụ gia khác. Phản ứng diễn ra trong vài phút rồi dừng lại, nếu bổ sung thêm mARN thì việc tổng hợp lại tiếp diễn. Nếu mARN nhân tạo chỉ toàn uraxin thì chuỗi polipeptit được tổng hợp chỉ có loại axit phenylalanine, nếu mARN chỉ toàn adenine thì chuỗi polipeptit được tổng hợp chỉ có loại axit lisine; nếu mARN toàn cytosine thì chuỗi polipeptit được tổng hợp chỉ có loại axit proline. Từ đó có thể suy ra bộ ba UUU quy định axit amin phenylalanine; AAA : lisine; CCC : proline. Bằng phương pháp đó người ta đã tìm ra 61 bộ ba mã hoá. Ba bộ ba còn lại là UAA, UAG, UGA làm nhiệm vụ nhận biết tín hiệu kết thúc quá trình tổng hợp chuỗi polipeptit, còn gọi là bộ ba vô nghĩa (non sense) vì chúng không quy định axit amin. 1.2. Các đặc tính của mã Thông tin được đọc theo từng cụm ba nucleotit một cách liên tục không ngắt quãng.
- Thông tin được đọc theo một chiều, bắt đầu từ một điểm xác định. - Mã di truyền mang tính phổ biến (umversal). - Mã di truyền mang tính thoái hoá (degenerate), trừ 2 ngoại lệ AUG và
UGG. - Mã di truyền mang những bộ ba khởi đầu (AUG) ở đầu 5'. Các bộ ba kết
thúc UAG, UAA, UGA.
34

Bảng mật mã di truyển (mã sao mARN)
U C A G UUU. Phe UCU Ser UUC Phe. UCC Ser UUA Leu UCA Ser
U
UUG Leu UCG Ser
UAU Tir UAC Tir UAA KT UAG KT
UGU Cys UGC Cys UGA KT UGG Trp
U C A G
CUU Leu CCU Pro CUC Leu CCC Pro CUA Leu CCA Pro
C
CUG Leu CCG Pro
CAU His CAC His CAA Gin CAG Gin
CGU Arg CGC Arg CGA Arg CGG Arg
U C A G
AUU Isoleu ACU Thr AUC Isoleu ACC Thr AUA Isoleu ACA Thr
A
AUG Met ACG Thr
AAU Asn AAC Asn AAA Lis AAG Lis
AGU Ser AGC Ser AGA Arg AGG Arg
U C A G
GUU Val GCU Ala GUC Val GCC Ala GUA Val GCA Ala
G
GUG Val GCG Ala
GAU Asp GAC Asp GAA Glu GAG Glu
GGU Gli GGC Gli GGA Gli GGG Gli
U C A G
Mã khởi đầu AUG có 2 chức năng: vừa tạo ra sự bắt đầu dịch mã (mã mở đầu) vừa mã hoá axit amin. (Một vài trường hợp mã mở đầu không phải là AUG mà là UGU điều khiển axit amin formyl methionine đi vào đầu chuỗi polipeptit). Mã kết thúc (codon vô nghĩa - nonsense):
UAA (ochae) Tín hiệu kết thúc UGA (opal) quá trình tổng hợp protein UAG (amber)
- Tính linh hoạt Trong tế bào có những loại tARN có thể đồng thời có một số bộ ba khác nhau. Phân tích trình tự tARN đã cho thấy rằng một số tARN có cấu trúc inosine là một trong các bazơ của cụm mã đối (anticodon). Đặc điểm của inosine được thể hiện ở hình 3.1. Phân tích trình tự cho thấy đầu 5' của codon đối mã (bổ trợ với bazơ thứ ba của cụm mã) khác với 2 bazơ kia, có khả năng kết hợp với một số bazơ trên đầu 3' của cụm mã. Hiện tượng này còn được gọi là tính "linh hoạt" trong kết cặp bazơ và tính linh hoạt là có giới hạn:
35
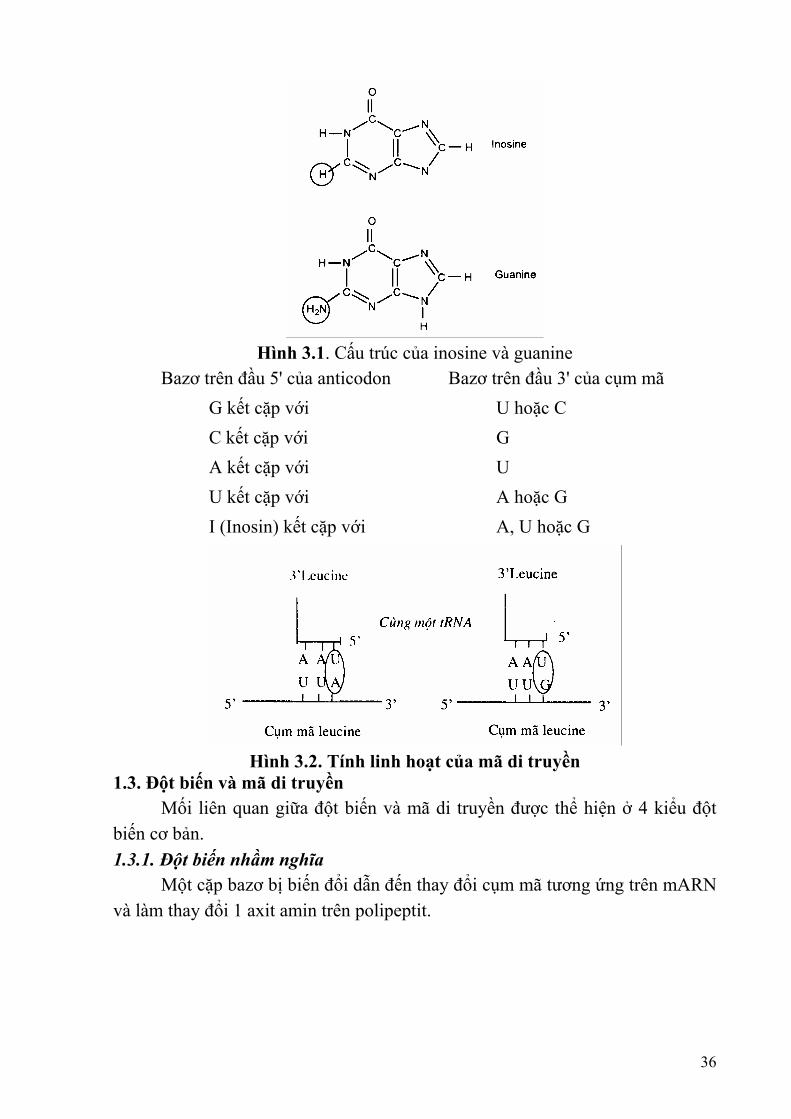
Hình 3.1. Cấu trúc của inosine và guanine
Bazơ trên đầu 5' của anticodon Bazơ trên đầu 3' của cụm mã G kết cặp với U hoặc C C kết cặp với G A kết cặp với U U kết cặp với A hoặc G I (Inosin) kết cặp với A, U hoặc G
Hình 3.2. Tính linh hoạt của mã di truyền
1.3. Đột biến và mã di truyền Mối liên quan giữa đột biến và mã di truyền được thể hiện ở 4 kiểu đột biến cơ bản. 1.3.1. Đột biến nhầm nghĩa Một cặp bazơ bị biến đổi dẫn đến thay đổi cụm mã tương ứng trên mARN và làm thay đổi 1 axit amin trên polipeptit.
36

1.3.2. Đột biến vô nghĩa
3' AGT CAA GGT TGC CAT 5' 1 2 3 4 5 Đột biến thay thế ở bộ ba thứ 4 (G-C bị thay thế bởi A-T) 3' AGT CAA GGT TAC CAT 5' Thay đối ở mARN 5' UCA GUU CCA AUG GUA 3' mARN
Biến đổi một cặp bazơ làm xuất hiện một codon vô nghĩa dẫn đến xuất hiện tín hiệu kết thúc chuỗi polipeptit làm cho chuỗi polipeptit được tổng hợp ngắn (ít axit amin) hơn bình thường.
3' AGT CAA GAT TGC CAT 5' 1 2 3 4 5 Đột biến măt cặp bazơ G-X ở bộ ba thứ 3 3' AGT CAA ATT GCC AT 5' Xuất hiện mã kết thúc UAA ở mARN 5' UCA GUU UAA CGG UA 3' mARN
1.3.3. Đột biến cặp bazơ ở bộ ba quy định axit amin 1àm xuất hiện codon mới (codon thoái hoá) không ảnh hưởng đến chuỗi polipeptit Một loại axit amin có thể được quy định bởi một vài bộ ba: ví dụ GUU, GUC, GUA, GUG cùng quy định axit amin valine. Trường hợp bình thường:
3' AGT CAA GGT TGC CAT 5' 1 2 3 4 5 5' UCA GUU CCA ACG GUA 3' mARN Scrine - Valine - Proline - Threonie - Valine.. Protein
Trường hợp đột biến.
37

1.3.4. Đột biến dịch khung
3' AGT CAA GGT TGC CAT 5' 1 2 3 4 5 Đột biến thay thế ở bộ ba thứ 2 (A-T bị thay thế bởi G-X) 3' AGT CAG GOT TAC CAT 5' Thay đối ở mARN 1àm BD đổi GUU thành GUC 5' UCA GUC CCA AUC GUA 3' mARN ... Serine - Valine - proline - Threonie - Valine..... Protein (GUU và GUC cùng quy định valine)
Hiện tượng mất hoặc thêm một cặp bazơ dẫn đến tổ hợp lại các codon và làm thay đổi thành phần axit amin của chuỗi polipeptit.
• Trường hợp bình thường:
3' AAC ACA CAG GTT CAA AGG AGC 5' Gene Phiên mã 5' UUG UGU GUC CAA GUU UCC UCG 3' mARN Dịch mã - Leucinne - Cysteine-Valine - Glicine - Valine - Tryptophane - Serine Protein
• Trường hợp đột biến dịch khung:
3' AAC ACA CAG GTT CAA AGG AGC 5' Gene Đột biến mất cặp C-G ở bộ ba thứ 3 3' aAC acAn agg ttc aaa Gga gc 5' Gene đột biến Phiên mã 5' UUG UGU UCC AAG UUU CCU CG 3' mARN Dịch mã - Leucinne - Cysteine - Serine - Lisine - Tryptophane - Proline Protein
38

Như vậy chuỗi polipeptide có thành phần axit amin: - Leucinne - Cysteine-Valine - Glicine - Valine - Tryptophane - Serine Biến đổi thành: - Leucinne - Cysteine - Serine - Lisine - Tryptophane - Proline.
§2. PROTEIN 2.1. Chức năng của protein trong tế bào Protein chiếm khối lượng lớn trong tế bào và tham gia vào hầu hết các quá trình sinh học. Thành phần cấu tạo nên các bộ phận của tế bào, quy định những đặc điểm về hình thái và cấu tạo của tế bào và cơ thể. Protein enzyme có khả năng xúc tác lớn và tính đặc hiệu cao. Kháng thể: chống lại các tác nhân xâm nhập vào tế bào và cơ thể. Tác nhân gây miễn dịch (kháng nguyên) xâm nhập vào cơ thể động vật có xương sống sẽ kích thích sự sản sinh ra kháng thể. Protein vận động (myosin), cảm ứng. 2.2. Cấu trúc của protein Axit amin là đơn vị cấu tạo nên protein, có khoảng 20 loại axit amin cấu tạo nên các phân tử protein khác nhau. Axit amin có cấu tạo chung như sau:
Dựa vào diện tích của chúng các aminoacit có thể chia làm 4 nhóm:
• Nhóm I. Các axit amin có tính kiềm: lisine, argimne, histidinc. ở độ pH của tế bào nhóm amine (NH)2 ở nhánh bên bị ion hoá thành NH3, nên chúng mang điện tích dương.
• Nhóm II. Các axit amin mang tính axit, vì nhánh bên bị oxy hoá thành COO-, bao gồm: aspartic và glutamic.
• Nhóm III. Các axit amin trung tính kị nước (không mang điện tích) như: alanine, valine, leucine... có nhánh bên mang các nhóm kị nước.
• Nhóm IV. Các axit amin trung tính phân cực có nhánh bên nhóm OH để tạo các liên kết hyđrô ưa nước.
39

Các axit amin liên kết với nhau bằng mối liên kết peptit. Protein có cấu trúc theo nhiều bậc:
Bậc I: chuỗi polipeptit có dạng mạch thẳng. Bậc II: chuỗi polipeptit có dạng mạch xoắn α . Bậc III: chuỗi polipeptit có dạng mạch hình cầu. Bậc IV: tổ chức protein có nhiều chuỗi polipeptit :
Hình 3.4. Liên kết tạo chuỗi polipeptit
Hình 3.5. Sơ đồ các bậc cấu trúc của protein Một phân tử protein chỉ có một chuỗi polipeptit (cấu trúc bậc I, II, III) hoặc gồm nhiều chuỗi polipeptit (cấu trúc bậc IV).
40

41

Hình 3.7. a, b: Cấu trúc bậc II; c, d, e, g: Cấu trúc bậc III; m. n, p, q: Cấu trúc bậc IV của protein.(theo Phan Văn Chi, 2002)
§3. QUÁ TRÌNH SINH TỔNG HỢP PROTEIN 3.1. Các yếu tố tham gia tổng hợp protein 3.1.1. Các phân tử ARN Mỗi phân tử mARN mang thông tin xác định trình tự của một polipepetit, thông tin này được phiên mã từ ADN. Số lượng loại tARN biến động theo loài từ 30 - 40 loại (Prokaryot), 50 loại (Eukaryot), nhưng cấu trúc của các loại tARN rất giống nhau. Chức năng của tARN được thực hiện nhờ các enzyme đặc biệt là các aminoacyl-tARN-synthetase. Có 20 loại aminoacyl-tARN-synthetase tương ứng với 20 loại axit amin. Quá trình gắn axit amn với tARN với sự tham gia của enzyme này là quá trình tiêu tốn năng lượng và trải qua 2 bước:
• Enzyme nhận biết và gắn với 1 axit amin đặc hiệu
• Enzyme + a.a + ATP → enzyme - aminoacyl - AMP + P.P Axit amin được chuyển từ phức hợp enzyme aminoacyl sang tARN tương ứng. Enzyme - aminoacyl - AMP + tARN → tARN- aminoacyl + AMP + E 3.1.2. Ribosome (RBX)
42

Ribosome (RBX) được cấu tạo từ rARN và protein tạo thành ribonucleoprotein. RBX có 2 tiểu phần (tiểu đơn vị); tiểu đơn vị lớn chứa 1 phân tử rARN lớn và tiểu đơn vị nhỏ chứa 1 rARN nhỏ. RBX ở Prokaryot và Eukaryot cơ bản giống nhau.
Hình 3.8. Ribosome
3.2. Các giai đoạn của quá trình tổng hợp protein Quá trình sinh tổng hợp protein trong tế bào trải qua hai khâu: phiên mã và dịch mã (hình 3.9). Ở đây chỉ trình bày khái quát các giai đoạn của quá trình dịch mã. 3.2.1. Khởi động Giai đoạn có sự tham gia của nhiều nhân tố protein - nhân tố khởi động (IF - Initiation Factor); ở Prokaryot có 3 nhân tố IF, còn ở Eukaryot có 6 nhân tố IF. Dấu hiệu khởi động là codon AUG (mã hoá methyonine). Có 2 loại tARN mang a.a Met có khả năng kết hợp với codon AUG:
(1) tARNmet : kết hợp với Codon AUG ở giữa mARN (2) tARimet : kết hợp với AUG khởi động, gắn a.a methyonine đầu tiên của
chuỗi polipeptit. Các bước của giai đoạn khởi động: Aminoacyl - tARN - synthetase chuyên biệt (methionyl - tARN - synthetase) gắn 1 phân tử met vào một đầu của tARNmet tạo thành Met - tARNimet. Hình thành tiểu đơn vị nhỏ ribosome - met - tARNimet - mARN với sự tham gia của nhân tố khởi động (IF2 - Prokaryot ; IF4 - Eukaryot). 3.2.2. Kéo dài Khi a.a met đặt vào vị trí chuỗi polipeptit bắt dầu được tổng hợp. 43
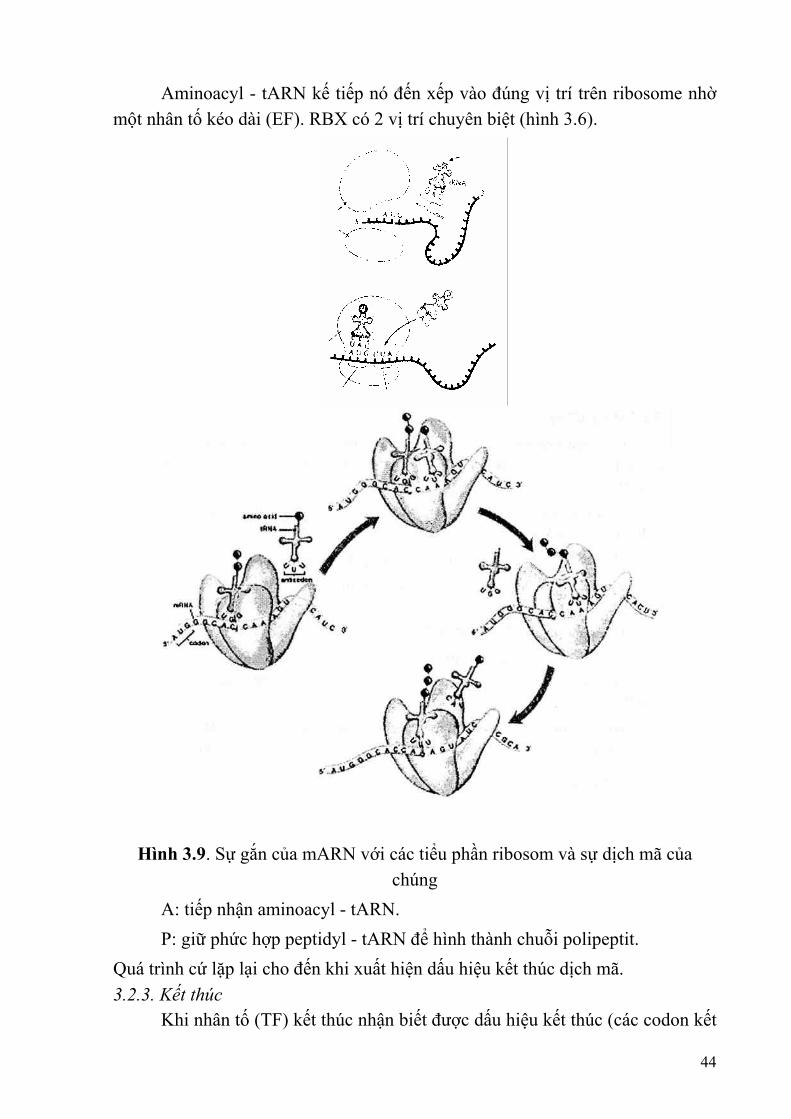
Aminoacyl - tARN kế tiếp nó đến xếp vào đúng vị trí trên ribosome nhờ một nhân tố kéo dài (EF). RBX có 2 vị trí chuyên biệt (hình 3.6).
Hình 3.9. Sự gắn của mARN với các tiểu phần ribosom và sự dịch mã của
chúng A: tiếp nhận aminoacyl - tARN. P: giữ phức hợp peptidyl - tARN để hình thành chuỗi polipeptit. Quá trình cứ lặp lại cho đến khi xuất hiện dấu hiệu kết thúc dịch mã. 3.2.3. Kết thúc Khi nhân tố (TF) kết thúc nhận biết được dấu hiệu kết thúc (các codon kết
44

thúc: UAG, UAA, UGA), phức hợp peptidyl - tARN lập tức tách thành tARN tự do và chuỗi polipepetit. RBX tách ra thành 2 tiểu đơn vị và mARN được giải phóng.
Hình 3.10. Dịch mã tổng hợp protem ở các ribosom trên lưới nội chất
§4. ĐIỀU HOÀ BIỂU HIỆN GENE Ba thành phần cơ bản của sự điều hoà hoạt động gene.
(1). Những tín hiệu làm thay đổi biểu hiện gene. (2). Điều hoà biểu hiện gene thực hiện ở giai đoạn nào từ sao chép đến dịch mã. (3) Cơ chế phân tử của sự điều hoà biểu hiện gene.
4.1. Điều hoà biểu hiện gene ở Prokaryot Các gene được phiên mã tạo ARN, được gọi là gene cấu trúc, protein được dịch mã từ mARN có thể enzyme hoặc không phải là enzyme. Trong số các protein không phải enzyme có protein điều hoà (regulatory prtotenin) chúng tương tác với trình tự ADN đặc hiệu kiểm soát phiên mã của gene cấu trúc. Gene điều khiển tổng hợp protein điều hoà gọi là gene điều hoà (regulatory gene), phía trước nhóm gene cấu trúc có trình tự protnotor, nơi ARN-polimerase nhận biết.
45

Hìnhh 3.11. Mối quan hệ giữa AND, ARN và protein
Thực chất của khởi đầu phiên mã là quan hệ tương tác giữa ARN - polimerase với promotor. Khi ARN-polilmese gắn vào promotor gene sẽ tạo phân tử mARN. 4.1.1. Cấu trúc của operon Sự điều hòa biểu hiện gene ở Prokaryot phần lớn xảy ra ở giai đoạn phiên mã. Cơ chế điều hòa thông qua operon. Mỗi operon bao gồm: nhóm gene cấu trúc (cistron) cạnh nhau và các trình tự AND khác nhau tham gia điều hòa. Operon là đơn vị phiên mã gồm ít nhất 1 promotor (trình tự khởi động) và mARN kế cận mã hóa cho các trình tự của một hay nhiều mạch polipeptit. Promotor có ái lực với ARN-polimerase, một trình tự AND nơi mà protein ức chế (repressor protein) gắn vào được gọi là operator (điểm điều hành), kế tiếp là các gene cấu trúc (cistron). Prokaryot thường tạo ra mARN đa gene (policistronic), nhưng mARN của Eukaryot chỉ có 1 gene. Kiểm soát âm (Negative control): Sự gắn của repressor protein vào operator làm cản trở sự phiên mã của tất cả các gene trong cùng một operon. Kiểm soát dương (Positive control): Các protein cần thiết cho gene biểu hiện gọi là activator (chất hoạt hoá), activator có thể gắn với các điểm khởi động (initiator site) của operon hay gắn với enhancer site (điểm tăng cường) sẽ kích
46

thích sự phiên mã của các gene cấu trúc. Chất giúp cho gene phiên mã được gọi là chất cảm ứng (inducer), chất có tác dụng ngược lại gọi là chất kìm hãm. Các gene cảm ứng (inducible gene) thường thường tham gia vào các phản ứng thoái dưỡng (kiểm soát âm, cảm ứng); còn các gene ức chế (repressible gene) thường tham gia vào các phản ứng biến dưỡng. Khả năng có thể xảy ra 4 kiểu tổ hợp để kiểm soát sự phiên mã:
• Âm, cảm ứng (negative, inducible control);
• Âm, ức chế (negative, repressive control),
• Dương, cảm ứng (positive, inducible control);
• Dương, ức chế (positive, repressive control). 4.1.2. Điều hoả thoái dưỡng: kiểm soát âm, cảm ứng (negarive, inducible control) Các chất dinh dưỡng được phân huỷ để tạo năng lượng hoặc tạo nguyên liệu cần thiết cho quá trình tổng hợp. Cơ chế điều hoà ở đây là sự có mặt của cơ chất, dẫn tới tổng hợp các enzyme phân huỷ. Đó là operon cảm ứng mã hoá cho các enzyme của con đường dị hoá. Điển hình cho kiểu operon cảm ứng là operon lactose. Tín hiệu điều hoà - đường lactose. Không có lactose gene mã hoá các enzyme chuyển hoá lactose không biểu hiện; Khi có lactose gene mã hoá các enzyme chuyển hoá lactose biểu hiện. Sự điều hoà xảy ra hoàn toàn ở mức phiên mã. Cơ chế này dựa vào sự tương tác của protein điều hoà (nhân tố kìm hãm - repressor) với một trình tự ADN, promotor ở đầu 5' không mã hoá của các gene cấu trúc. Repressor liên kết với operator nằm giữa promotor và gene cấu trúc. Có 2 khả năng: (1) Không có lactose: Khi môi trường không có lactose mà chỉ có glucose thì repressor liên kết với operator và các cistron không tổng hợp mARN - lac.
Hình 3.12. Môi trường không có lactose repressor liên kết với operator và các
47

cistron không phiên mã (2) Có lactose:
Hình 3.13. Môi trường có lactose inductor gắn với repressor và các cistron phiên mã
Khi môi trường có lactose, gọi là nhân tố cảm ứng (inductor). Inductor gắn với repressor dẫn đến sự thay đổi cấu trúc không gian của repressor làm cho repressor không còn ái lực với operator dẫn đến quá trình phiên mã của các cistron bắt đầu và các enzyme chuyển hoá lactose được tổng hợp. 4.1.3. Điều hoà biên dưỡng: âm, ức chế (negative, repressive control) Biến dưỡng (anabolism) là quá trình tổng hợp nên các chất cần thiết cho tế bào (như các axit amin), đó là operon kìm hãm liên quan đến con đường đồng hoá - điển hình là operon - triptophan. Quá trình tổng hợp tryptophan bắt đầu từ tiền chất chorismique axit, trải qua 5 giai đoạn kế tiếp do 5 enzyme xúc tác. Hệ thống tổng hợp amino axit tryptophan ở E. coli do operon kiểm soát âm, ức chế. Khi môi trường không có tryptophan tế bào tổng hợp tryptophan nhờ tích cực sản xuất enzyme chịu trách nhiệm tổng hợp tryptophan. Môi trường có tryptophan thì enzyme không được sản xuất nữa. Nhân tố kìm hãm repressor không có ái lực với operator. Axit amin đặc trưng kết hợp với repressor sẽ làm thay đổi cấu hình của repressor.
• Khi môi trường có tryptophan, repressor kết hợp với tryptophan gắn lên operator làm cho các gene cấu trúc của operon không phiên mã.
• Môi trường thiếu triptophan, repressor không gắn được vào operator các gene cấu trúc của operon phiên mã.
4.2. Điều hoà biểu hiện gene ở Eukaryot Bộ gene của Eukaryot có kích thước lớn, phân tử ADN được nén trong nhân tế bào. Cơ chế điều hoà biểu hiện gene có thể xảy ra ở 5-6 mức độ khác 48

nhau và sự điều hoà biểu hiện gene ở Eukaryot có một số đặc điểm cơ bản sau: Ở Prokaryot, các gene điều hoà và promotor nằm gần nhau; còn ở Eukaryot các gene điều hoà ít khi nằm gần các promotor do chúng kiểm soát. Các enhancer là những trình tự cùng nằm trên một phân tử với các promotor, có thể có hàng trăm bp ở phía trước hoặc sau mà chúng kích thích. Trình tự điều hoà 5' ở phía trước promotor của Eukaryot có thể dài tới hàng chục kb. Có nhiều kiểu điều hoà ở dạng các nhân tố tác động trans là các protein. Sự điều hoà hoạt động gene ở Prokaryot phần lớn đáp lại từ những tín hiệu bên ngoài; còn ở Eukaryot chủ yếu là tín hiệu bên trong. Promotor ở Eukaryot nằm phía trước điểm xuất phát của mARN. Hộp TATA, định hướng cho ARN-polimerase bắt đầu phiên mã. Sự thay đổi hộp TATA làm giảm tốc độ phiên mã. Hiệu quả của tốc độ phiên mã đo bằng sự thay đổi từng bazơ trong promotor. Hộp TATA và các trình tự phía trước phải được nhận biết bởi các protein điều hoà, chính các protein này gắn với các điểm nhất định trên chúng và hoạt hoá sự phiên mã. Enhancer là trình tự có tác động cis (đều phía) có chức năng làm tăng tốc độ phiên mã đáng kể từ các promotor, enhancer có hiệu quả ngay cả khi cách xa vài nghìn cặp bazơ và chúng hoạt động bất kì ở hướng nào, dù là ở trước hay sau promotor. Tóm lại, các gene của Eukaryot được hoạt hoá bởi hai trình tự ADN có tác động cis là promotor và enhancer. Chúng được nhận biết các nhân tố protein có tác động trans. Các nhân tố trans này cho phép ARN-polimerase khởi động phiên mã và đạt tốc độ phiên mã tối đa. THẢO LUẬN
1. Trình bày về mật mã di truyền. 2. Đột biến gene và mật mã di truyền. 3. Mối liên quan giữa protein và tính chống chịu. 4. Đặc điểm cấu trúc của protein. Quá trình tổng hợp protein. 5. Phương pháp phân tích cấu trúc protein và đặc điểm cấu trúc của protein. 6. Điều hoà biểu hiện gene và ứng dụng trong kĩ thuật di truyền.
49

Chương 4: ENZYME SỬ DỤNG TRONG KĨ THUẬT SINH HỌC PHÂN TỬ Tóm tắt: Enzyme là những protein hoạt tính sinh học vả dựa vào loại phản ứng xúc tác mà trong sinh học phân tử có thể chia enzyme thành 6 nhóm khác nhau. Trong đó các nhóm enzyme giới hạn, enzyme polimerase, enzyme nối, các enzyme nuclease được quan tâm nghiên cứu và ứng dụng trong các lĩnh vực của sinh học phân tử và kĩ thuật di truyền. Enzyme được sử dụng trong các kĩ thuật sinh học phân tử như RFLP, PCR, tách dòng phân tử......... Nội dung của chương gồm 3 vấn đề cơ bản: (1). Enzyme giới hạn, (2). các nhóm enzyme khác; (3). Enzyme nuctease. Hệ thống enzyme có thể chia thành 6 nhóm tuỳ theo loại hình phản ứng mà các enzyme này xúc tác. Nhóm enzyme nuclease có tác dụng cắt và phá huỷ ADN, nhóm ADN-ligase có chức năng nối các trình tự ADN lại. nhóm enzyme ADN-polimerase tham gia tổng hợp chuỗi polinucleotit, nhóm enzyme ADN-polimerase sửa chữa có chức năng sửa chữa những sai sót trong phân tử ADN, nhóm enzyme topoisommerase tác dụng mở xoắn ADN, nhóm enzyme phiên mã ngược reverse-transcriptase xúc tác cho quá trình phiên mã ngược từ ARN thành cADN. Các nhóm enzyme trên có vai trò hết sức quan trọng trong các thao tác với ADN và trong công nghệ ADN tái tổ hợp. Trong số đó có các nhóm enzyme giới hạn, enzyme sửa chữa ADN, ADN-polimease, ADN-ligase....
§1. ENYME GIỚI HẠN (RESTRICTION ENZYME HOẶC ENZYME CẮT HẠN CHẾ (RESTRICTION ENDONUCLEASE - RE) 1.1. Khái niệm Nhóm enzyme nuclease gồm DN-ase và RN-ase có khả năng bẻ gãy liên kết phosphodiester kết quả là phân huỷ phân tử ADN hoặc ARN. Nhóm enzyme ADN-ase gồm 2 loại: Exonuclease (enzyme cắt ADN ngoại bào) có tác dụng cắt từng cặp nucleotit từ hai đầu của đoạn ADN đi dần vào bên trong (hình 4.1). Enzyme exonuclease có thể cắt tỉa từng cặp nucleotit ở cả hai mạch (cắt kép) hoặc cắt từng nucleotit trên một sợi ADN (cắt đơn). Endonuclease (enzyme cắt ADN nội bào) có tác dụng cắt rời ADN tại những vùng xác định gồm vài cặp nucleotit nằm cạnh nhau định vị trong phân tử ADN, tạo thành những phân đoạn ADN nhỏ hơn. Enzyme endonuclease có thể cắt đơn
50

hoặc cắt kép (hình 4.1).
Hình 4.1. Đặc điểm cắt của exonuclease và endonuclease
Một loại enzyme endonuclease có thể nhận biết vị trí nhất định bên trong phân tử ADN và cắt đặc hiệu, cắt một lần tại một điểm đó. Những kết luận trên đã được phát hiện trong thập kỉ 60 của thế kỉ XX. Trong những thí nghiệm ở vi khuẩn người ta đã phát hiện một nhóm các enzyme có thể nhận biết và phân cắt phân tử ADN của phagơ khi ADN ngoại lai này bảo vệ tế bào chủ xâm nhập. Các enzyme này bảo vệ tế bào chủ khỏi bị nhiễm phagơ - đó là các enzyme giới hạn. Enzyme cắt hạn chế (enzyme giới hạn) là loại enzyme nuclease có khả năng nhận biết được điểm cắt và cắt tại điểm xác định. Tuy nhiên vấn đề đặt ra là, liệu enzyme giới hạn cắt ADN của chính tế bào chủ? Những nghiên cứu cho thấy các enzyme giới hạn phân huỷ bất kì ADN ngoại lai nào xâm nhập vào tế bào; trong tế bào hệ thống enzyme sửa đổi (methylase) sửa đổi ADN tế bào chủ bằng cách methyl hoá các bazơ xác định của đoạn trình tự nhận biết, nhờ vậy ngăn cản enzyme giới hạn cắt vào ADN của tế bào chủ. Những enzyme này là đại diện của một trong các nhóm enzyme quan trọng nhất 51

dùng trong thao tác ADN. 1.2. Các nhóm enzyme giới hạn Các enzyme giới hạn có ba nhóm I, II, III và các enzyme giới hạn được dùng phổ biến ngày nay thuộc nhóm II. Enzyme giới hạn nhóm II có cơ chế tác động đơn giản nhất, chúng là các nuclease, và vì chúng cắt tại một vị trí nằm bên trong sợi ADN và không phân huỷ ADN từ hai đầu, nên gọi là endonuclease. Bảng 4.1. Đặc điểm của các loại enzyme giới hạn
Đặc điểm Nhóm I Nhóm II Nhóm III
Điểm cắt Xa điểm nhận biết hơn 1000 bp
Nằm trong điểm nhận biết
Nằm ngoài điểm nhận biết nhận biết
Khả năng methy hóa gốc Adenine
Có Không Có
Điều kiện để cắt ATP,Mg++, Mg++ S-AdoMet
Mg++ hoặc Mn++ Mg++, S-AdoMet
Cấu trúc của enzyme (số chuỗi polipeptide)
Khác nhau Giống nhau Khác nhau
Hình 4.2. Quá trình methyl hoá bởi enzyme methylase
Cách gọi tên các enzyme giới hạn dựa trên các quy ước quốc tế. Tên chi và tên loài của sinh vật, mà ở đó tìm thấy enzyme, được dùng để đặt cho phần
52

đầu của tên enzyme, phần đó gồm chữ thứ nhất của tên chi và hai chữ đầu của tên loài. Chẳng hạn enzyme tách chiết được từ một nòi của Escherichia coli thì đặt tên là Eco, còn enzyme tách từ Bacillus amiloliquefaciens thì gọi là Bam... Hai loại enzyme được sử dụng rộng rãi được tách ra từ các vi khuẩn nói trên là EcoRI và BamHI. Ví dụ: EcoRI: RE tách từ vi khuẩn E. coli, dòng R, nhóm I. HindIII: RE tách từ vi khuẩn Heamophilus influenzae, dòng d, nhóm III. Enzyme giới hạn có tính đặc hiệu, mỗi enzyme cụ thể nhận biết một đoạn trình tự đặc thù các bazơ trên ADN, đoạn nhận biết phổ biến nhất có chiều dài từ 4 - 6 bp. Nếu một trình tự ADN có 4 bp thì cứ 44 = 256 bp lặp lại một lần, trình tự có 5 bp thì cứ 45 = 1024 bp lặp lại, trình tự có 6 bp thì cứ 46 = 4096 bp mới lặp lại. Chẳng hạn, một enzyme nhận biết trình tự có 4 bp sẽ tạo ra đoạn ngắn hơn so với trình tự có 6 bp. Loại hình của các đoạn ADN do một enzyme cụ thể tạo ra tuỳ thuộc vào trình tự nhận biết và vào vị trí của điểm cắt trong trình tự này. Như chúng ta đã thấy, chiều dài của đoạn này phụ thuộc vào tần số lặp lại của trình tự nhận biết. Điểm cắt thực tế của enzyme quyết định hình dạng của đầu bị cắt. Hình dạng này rất quan trọng đối với việc thao tác tiếp ADN. Có hai kiểu hình dạng các đoạn có thể sinh ra: (1) Các đoạn đầu bằng (blunt ends); (2) Các đoạn đầu dính (so le) với 2 kiểu: đoạn có đầu 3' nhô ra; và đoạn có đầu 5' nhô ra.
Hình 4.3. Các kiểu cắt của RE nhóm II
Các enzyme như PstI và EcoRI tạo ra các đoạn ADN có đầu dính, chúng có thể bắt cặp bazơ với trình tự bổ sung cũng do chính enzyme đó tạo ra. Như vậy, bằng cách cắt hai mẫu ADN khác nhau bởi cùng một enzyme và trộn các 53

đoạn đó với nhau có thể tạo ra ADN tái tổ hợp. Bằng cách này các đoạn ADN nguồn khác nhau có thể nối lại với nhau nếu chúng có các đầu dính do enzyme giới hạn sinh ra. Khi các vùng bổ trợ ghép với nhau thì bộ khung photphodiester được gắn lại bằng ADN ligase. Trong thực nghiệm, người ta trộn một lượng thích hợp enzyme với ADN caanf nghiên cứu trong dung dịch đệm, cho phản ứng diễn ra ở 370C. Hoạt tính enzymc được do bằng đơn vị tương đương với lượng enzyme cần thiết để một microgam ADN trong một giờ ở 370C. 1.3. Lập bản đồ cắt giới hạn Phần lớn các đoạn ADN đều có các điểm nhận biết đối với nhiều enzyme giới hạn khác nhauvà nếu biết được vị trí tương đối của một số điểm này thì có thể lập được bản đồ các điểm giới hạn. Trong kĩ thuật này, người ta sử dụng các enzyme giới hạn để cắt một đoạn ADN, sau đó điện di trên agrose, chụp ảnh phân tích được kích thước các đoạn ADN bị cắt. Từ các kết quả phân tích có thể xác định được vị trí tương đối của các điểm được cắt trên phân tử ADN.
Hình 4.4. Kiểu cắt của một số loại enzyme
Giả sử chúng ta muốn lập bản đồ các điểm cắt của các enzyme BamHI, EcoRI và Psd đối với đoạn ADN có chiều dài 15kb, kết quả thu được như sau: Nếu chỉ sử dụng riêng lẻ một loại enzyme thì đoạn ADN dài 15 kb sẽ bị cắt thành 2 đoạn có kích thước khác nhau. BamHI cắt đoạn ADN thành 2 đoạn với kích thước 14 kb và 1 kb EcoRI cắt đoạn ADN thành 2 đoạn với kích thước 12 kb và 3 kb PstI cắt đoạn ADN thành 2 đoạn với kích thước 8 kb và 7 kb
54

Nếu sử dụng phối hợp 2 hoặc 3 loại enzyme thì đoạn ADN bị cắt thành 3 hoặc 4 đoạn nhỏ: BamHI + EcoRI cắt đoạn ADN thành 3 đoạn với kích thước 11 kb, 3 kb và 1 kb. BamHI + PstI cắt đoạn ADN thành 3 đoạn với kích thước 8 kb, 6 kb và 1 kb. EcoRI + PstI cắt đoạn ADN thành 3 đoạn với kích thước 7 kb, 5 kb và 3 kb. BamHI + EcoRI+PstI cắt đoạn ADN thành 4 đoạn với kích thước 6kb, 5kb, 3kb, 1kb.
Hình 4.5. Các enzyme BamHI, EcoRI và PstI cắt trình tự 15kb thành 2 phân
đoạn ADN có kích thước khác nhau Việc phát hiện các enzyme giới hạn là bước đột phá quan trọng trong lĩnh vực sinh học phân tử nói chung cũng như công nghệ tái tổ hợp AND nói riêng. Thực chất những enzyme dùng trong công nghệ ADN tái tổ hợp thuộc enzyme giới hạn nhóm II. Chúng phân cắt khu vực bên trong phân tử ADN tại một trình tự nucleotit nhất định và tạo ra hỗn hợp (quần thể) các đoạn ADN với kích thước khác nhau. Cho đến ngày nay người ta đã tách được hơn 1200 enzyme giới hạn nhóm II từ các cơ thể procariot. Trên 70 enzyme giới hạn hiện đang được bán sẵn trên thị trường. Các RE có thể nhận biết những trình tự ngắn của ADN và cắt đúng vào vị trí đó phân chia phân tử ADN thành những đoạn có độ dài đặc trưng. Các RE khác nhau thì có đoạn nhận biết với trình tự bazơ đặc trưng khác nhau. Sơ đồ mô tả các vị trí cắt khác nhau của một phân tử ADN được gọi là bản đồ cắt hạn chế. Nhiễm sắc thể của Eukaryot chứa phân tử ADN gồm nhiều gene và nó có thể bị cắt bởi enzyme cắt hạn chế thành nhiều đoạn nhỏ, chính vì vậy có thể sử dụng enzyme giới hạn trong ứng dụng lập bản đồ cắt hạn chế.
55

Hình 4.8. Điện di đồ các phân đoạn ADN bị cắt bởi sự phối hợp các enzyme
a. BamHI + EcoRI b. BamHI + PstI c EcoRI + PstI d. BamHI + EcoRI+PstI Quy trình lập bản đồ cắt hạn chế theo quy trình dưới đây:
1. Nghiên cứu đa dạng kiểu hình trong quần thể. 2. Tách chiết phân tử ADN. 3. Sử dụng enzyme RE. 4. Điện di trên gene agarose.
Phân đoạn cắt bởi RE-A ADN chuẩn Phân đoạn cắt bởi RE-B
Hình 4.7. Sơ đồ các bảng điện di của các mẫu thí nghiệm và ADN chuẩn Ngoài ứng dụng thiết lập bản đồ cắt giới hạn người ta còn sử dụng enzyme giới hạn kết hợp với enzyme nối để tạo vector tái tổ hợp (hình 4.8).
56

Hình 4.8. Sử dụng enzyme giới hạn và enzyme nối để tạo ADN tái tổ hợp
§2. CÁC NHÓM ENZYME KHÁC 2.1. Polimerase Các polimerase tham gia vào quá trình tái bản ADN và phiên mã tổng hợp ARN và được sử dụng nhiều trong kĩ thuật di truyền. Các pohmeasase gồm 2 nhóm: (1) ADN-polimerase xúc tác tổng hợp chuỗi polinucleotit theo chiều từ 5' đến 3'. Taq polimerase (enzyme tách từ vi khuẩn suối nước nóng) có khả năng chịu nhiệt, sử dụng trong phản ứng chuỗi PCR. Enzyme phiên mã ngược (Reverse Transcriptase: RT-ase) xúc tác tổng hợp CADN từ khuôn ARN; (2) Các ARN-polimerase tham gia tổng hợp ARN. Ngoài các enzyme trên còn có nhiều enzyme tham gia sửa chữa những sai sót trên phân tử ADN. 2.1.1. Taq polimerase Taq polimerase được tách chiết từ một loại vi khuẩn Thermus aquaticus suối nước nóng. Taq polimerase được sử dụng trong PCR và RAPD tổng hợp đoạn ADN trong điều kiện có dung dịch đệm, Mg+ primers. Hiện nay các nhà khoa học có thể phân lập gene xác định enzyme polimerase, tách dòng và biểu hiện gene này. Chính vì vậy bằng công nghệ ADN tái tổ hợp có thể tạo ra lượng lớn enzyme Taq polimerase sử dụng trong kĩ thuật di truyền. 2.1.2. ADN-polimerase I Enzyme ADN-polimerase I, ngoài chức năng polimerase, còn có hoạt tính
57

của exonuclease 5' → 3' và 3' → 5'. Enzyme này xúc tác phản ứng thay thế sợi, trong phản ứng đó chức năng exonuclease 5' → 3' phân huỷ sợi không làm khuôn trong khi polimerase tổng hợp bản sao mới. Chức năng exonuclease 5'→3' của ADN polimerase I có thể được loại bỏ bằng cách cắt enzyme để tạo ra đoạn Klenow. Đoạn này vẫn duy trì các hoạt tính polimerase và exonuclease 3'→ 5'. Đoạn Klenow được sử dụng khi cần sao chép một phân tử ADN mạch đơn. 2.1.3. Enzyme phiên mã ngược Enzyme phiên mã ngược được phát hiện từ 1970 có chức năng tổng hợp ADN từ ARN. Trong tự nhiên, enzyme này do Retrovirus sản sinh để phiên mã ngược từ hệ gene ARN của chúng để thực hiện quá trình xâm nhập vào hệ gene của tế bào chủ. Reverse Transcriptase được sử dụng trong kĩ thuật tạo cADN bằng phản ứng PCR ngược (RT-PCR). Hiện nay người ta sử dụng các loại enzyme phiên mã ngược như: Avian Myeloblastosis virus reverse transcriptase (AMV-RT) và Moloney Murine Leukemia virus reverse transcriptase (M-MLV-RT), trước đây AMV-RT phải tách chiết từ virus Myeloblastosis gia cầm, còn M-MLV-RT từ virus gây bệnh Leukemia ở chuột. Các enzyme này là sản phẩm của gene pol của virus và hiện nay các enzyme này đã nhận được từ công nghệ ADN tái tổ hợp. Enzyme phiên mã ngược là một ADN-polimerase phụ thuộc ARN, vì vậy nó sản sinh ra sợi ADN từ sợi khuôn ARN. Nó không có hoạt tính exonuclease kèm theo. Enzyme này chủ yếu được sử dụng để sao chép các phân tử mARN khi chuẩn bị cADN (ADN bổ trợ hoặc ADN bản sao) để tách dòng, mặc dù nó cũng có tác động trên khuôn ADN. 2.1.4. T4 ADN-polimerase Enzyme T4 ADN-polimerase có nguồn gốc từ phage T4 kí sinh trong tế bào vi khuẩn. Hoạt tính của enzyme này giống như hoạt tính đoạn Klenow và được sử dụng trong tổng hợp mẫu dò có độ phóng xạ cao. Bởi vì hoạt tính exonuclease 3'-5' mạnh hơn T4 ADN-polimerase. 2.1.5. Terminal transferase Enzyme Terminal transferase được li trích từ tuyến ức bò, có chức năng xúc tác phản ứng gắn các deoxynucleotit vào đầu 3'-OH tự do của ADN. Terminal transferase được sử dụng khi thêm đuôi polinucleotit để tạo đầu so le cho ADN trong kĩ thuật tạo dòng. 2.1.6. Enzyme Alkaline phosphatase Enzyme Alkaline phosphatase được phân lập từ E. coli hay từ ruột bò. Chức năng của enzyme này là loại bỏ nhóm 5' phossphate của ADN, ARN và
58

các nucleotit tự do. Enzyme Alkaline phosphatase sử dụng để loại bỏ 5'phossphate của ADN và ARN trước kho đánh dấu phóng xạ bằng 32P; sử dụng trong kĩ thuật tạo dòng, khi vector được "mở" bởi RE và enzyme làm mất 5' phosphate để vector không thể nối lại được. 2.1.7. Enzyme T4 polinucleotit kinase Enzyme T4 polinucleotit kinase có nguồn gốc từ phagene T4 kí sinh trong E. coli. Chức năng của enzyme T4 polinucleotit kinase ngược lại với Alkaline phosphatase, xúc tác cho phản ứng thêm vào nhóm phosphate ở đầu 5' của ADN hay ARN. T4 polinucleotit kinase sử dụng để đánh dấu phóng xạ ở đầu 5' của ADN trong kĩ thuật xác định trình tự ADN và phosphoryl hoá các trình tự ADN không có nhóm phosphate ở đầu 5' để chuẩn bị cho phản ứng nối của kĩ thuật tạo dòng. 2.2. Enzyme nối ADN-ligase Enzyme nối ADN-ligase xúc tác sự hình thành liên kết nối hai đoạn ADN và ARN. ADN-ligase là một enzyme nối quan trọng trong tế bào vì chức năng của nó là sửa chữa các mối liên kết photphodiester bị đứt gãy một cách ngẫu nhiên hoặc trong tái bản ADN hay tái tổ hợp ADN. ADN-ligase được sử dụng trong kĩ thuật tạo dòng và thường được kết hợp với các enzyme Alkaline phosphatase và polinucleotit kinase. Nhóm enzyme ligase được sử dụng nhiều nhất trong các thí nghiệm với ADN và ARN chủ yếu gồm E. coli ADN-ligase, T4 ADN-ligase và T4 ARN-ligase. 2.2.1. E. coli ADN-1igase Enzyme E. coli ADN-ligase được tách chiết từ E. coli, xúc tác cho các phản ứng nối hai trình tự ADN có đầu dính (so le).
Hình 4.9. Enzyme E. coli xúc tác phản ứng nối hai trình tự có đầu dính 2.2.2. T4 ADN - ligase Enzyme T4 ADN-ligase chiết từ phage T4 kí sinh ở E. coli, xúc tác cho phản ứng nối hai trình tự có đầu bằng, được sử dụng nhiều trong kĩ thuật tạo 59

dòng. 2.2.3. T4 ARN-ligase Enzyme T4 ARN-ligase chiết từ phage T4 kí sinh ở E. coli, xúc tác cho phản ứng nối hai trình tự ARN bằng liên kết phosphodiester. Enzyme T4 ARN-ligase thường được sử dụng để đánh dấu phóng xạ ở đầu 3' của ARN trong kĩ thuật sản xuất mẫu dò (probe). Như vậy, enzyme ligase có hiệu quả trong sự gắn các khe hở của các trình tự có đầu dính hoặc đầu bằng với nhau trong những điều kiện thích hợp. Nhiệt độ thích hợp là 370C, nhưng nó có thể hoạt động ở nhiệt độ thấp hơn nhiều (4 15÷ 0C).
§3. ENZYME NUCLEASE Enzyme nuclease là nhóm enzyme phân cắt ADN (ADNse) và ARN (ARNse) 3.1. DN-ase I Enzyme DN-ase I có khả năng cắt liên kết ngay sau một bazơ pyrimidine trên ADN mạch đơn hay mạch kép. Trong thực nghiệm DN-ase I được sử dụng để loại ADN trong chế phẩm ARN và protein, tạo điểm đứt trong kĩ thuật đánh dấu mẫu dò. 3.2. Nuclease S1 Enzyme nuclease S1 tách chiết từ Aspergillus oryzae có tác dụng phân giải ADN mạch đơn, mạch kép. Enzyme nuclease S1 được sử dụng phân tích đặc điểm cấu trúc phân tử lai ADN - ARN, loại bỏ đầu so le để tạo đầu bằng, loại bỏ các nút vòng trên ARN trong phản ứng phiên mã ngược tạo cADN. 3.3. Exonuclease III Ezyme Exonuclease III chiết từ E. coli xúc tác phản ứng phân giải tuần tự các nucleotit từ đầu 3'- OH tự do của ADN theo chiều từ 3'→5' tạo ra những vùng mạch đơn trên ADN mạch kép. Enzyme Exonuclease III có ứng dụng tạo cấu trúc mạch đơn của một vùng trên phân tử ADN, tạo đột biến mất đoạn trên ADN khi phối hợp với nuclease Sĩ 3.4. RN-ase A Enzyme RN-ase A có hoạt tính rất mạnh, có thể chịu đựng được nhiệt độ 900C trong 60 phút. Enzyme RN-ase A có khả năng cắt liên kết phosphodiester ngay sau một bazơ pyrimidine của ARN. Enzyme RN-ase A được sử dụng để loại bỏ ARN trong chế phẩm ADN hoặc protein, loại bỏ các đoạn không bắt cặp
60

trên ARN của phân tử lại ARN -ADN. 3.5. RN-ase H Enzyme RN-ase H có khả năng loại bỏ ARN trong phân tử lai ARN - ADN và thường được sử dụng để loại ARN sau phản ứng phiên mã ngược để tiếp tục tổng hợp mạch thứ hai của CANAtạo ADN mạch kép. THẢO LUẬN
1. Thế nào là enzyme cắt và enzyme cắt hạn chế? Các nhóm enzyme giới hạn.
2. Vấn đề đặc điểm của các loại enzyme dùng trong kĩ thuật sinh học phân tử.
3. Trình bày các kiểu cắt của enzyme giới hạn, cho ví dụ. Nêu những ứng dụng cơ bản của RE.
4. Thế nào là bản đồ cắt giới hạn? cho ví dụ minh hoạ. 5. Nhóm enzyme nhân ADN và nối ADN. 6. Nghiên cứu chỉ thị sinh học phân tử bằng phương pháp sử dụng các loại
enzyme cắt hạn chế.
61

Chương 5: XÁC ĐỊNH MỐI LIÊN QUAN GIỮA PROTEIN VÀ ĐẶC TÍNH Ở SINH VẬT Tóm tắt: Protein lả sản phẩm biểu hiện gene, protein biểu hiện thành tính trạng và đặc tính ở sinh vật. Trong tế bào có nhiều loại protein chức năng quan trọng và những hiểu biết về đặc điểm của những protein này là cơ sở cho những ứng dụng vào công tác chọn giống và bảo vệ sức khoẻ con người. Những loại protein đề cập trong chương này là protein liên quan đến tính chống chịu của sinh vật và protein ức chế hoạt động ribosome. Xác định mốt liên quan giữa protein và tính trạng, đặc tính ở sinh vật lập thành các chỉ thị protein cho đặc tính và tính trạng. Các chỉ thị protein có thể là chỉ thị hàm lượng protein, enzyme, phân tử protein hay phổ isozyme và xác định các loại chỉ thị này được thực hiện theo quy trình kĩ thuật nhất định. Hàm lượng protem, đường và hoạt độ enzyme liên quan đến áp suất thẩm thấu trong tế bào và quan hệ với tính chống chịu của sinh vật. Có nhiều đặc tính và tính trạng của sinh vật được quan tâm nghiên cứu hiện nay là tính chống chịu của sinh vật và việc tìm ra các chỉ thị protein cho đặc tính này có ý nghĩa cao trong chọn tạo giống. Nội dung của chương gồm 6 vấn đề cơ bản: (1). Một số loại protein chức năng; (2). Quy trình xác định chỉ thị protein; (3). Kĩ thuật phân tích protein; (4). Kĩ thuật phân tích enzyme; (5). Kĩ thuật phân tích đường, (6). Nghiên cứu RIPs bằng Westem blot.
§1. MỘT SỐ LOẠI PROTEIN CHỨC NĂNG 1.1. Protein liên quan đến tính chống chịu Khả năng chống chịu của sinh vật nói chung và của thực vật nói riêng liên quan đến cấu trúc và chức năng của các thành phần trong tế bào. Các vùng tế bào và các quá trình xảy ra trong mỗi vùng liên quan đến khả năng chịu hạn, nóng, mặn, phèn của thực vật (hình 5.1).
62
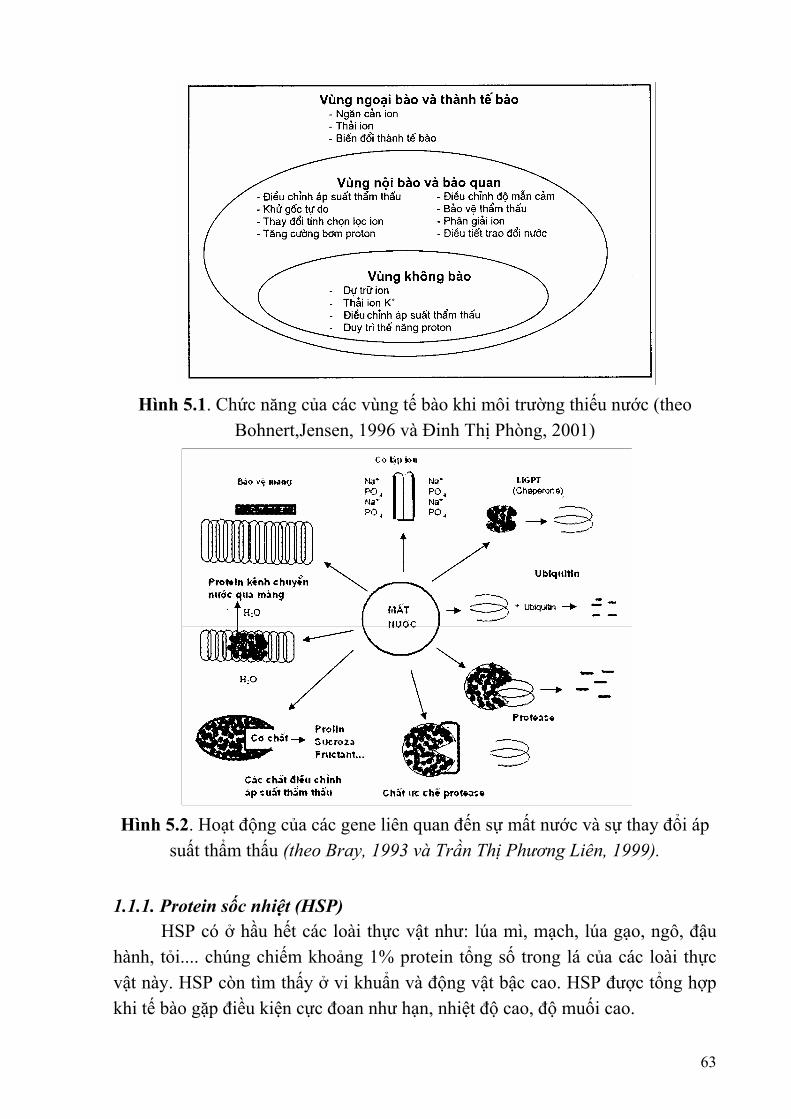
Hình 5.1. Chức năng của các vùng tế bào khi môi trường thiếu nước (theo
Bohnert,Jensen, 1996 và Đinh Thị Phòng, 2001)
Hình 5.2. Hoạt động của các gene liên quan đến sự mất nước và sự thay đổi áp
suất thẩm thấu (theo Bray, 1993 và Trần Thị Phương Liên, 1999). 1.1.1. Protein sốc nhiệt (HSP) HSP có ở hầu hết các loài thực vật như: lúa mì, mạch, lúa gạo, ngô, đậu hành, tỏi.... chúng chiếm khoảng 1% protein tổng số trong lá của các loài thực vật này. HSP còn tìm thấy ở vi khuẩn và động vật bậc cao. HSP được tổng hợp khi tế bào gặp điều kiện cực đoan như hạn, nhiệt độ cao, độ muối cao.
63

HSP có thể chia làm 6 nhóm dựa trên cơ sở khối lượng phân tử khác nhau: MW 110, 90, 70, 60, 20, 8,5 kDa. Trong số đó có nhóm HSP70 và HSP60 có nhiều đại diện của chất môi giới phân tử (gọi là chaperonin). HSP8,5 hoa (ubiquytin) có chức năng bảo vệ cho tế bào nhưng không phải là môi giới phân tử. Ubiquytin có hoạt tính protease với chức năng phân giải các protein không có hoạt tính enzyme, ngăn chặn hiện tượng gây độc tế bào của những protein này. Ubiquytin có MW rất thấp, ít chịu ảnh hưởng của nhiệt độ nên ubiquytin có vai trò tự sửa chữa của tế bào khi gặp yếu tố cực đoan, đặc biệt là nhiệt độ. 1.1.2. Môi giới phân tử (MGPT) Theo Ellis (1997) MGPT là một nhóm gồm nhiều loại protein khác nhau, nhưng chúng đều có chức năng tham gia tạo cấu trúc không gian đúng cho protein trong tế bào. Chức năng của MGPT thể hiện ở những nội dung sau: - Giữ ổn định chuỗi polipeptit khi đang tổng hợp và tạo cấu trúc không gian đúng cho chúng. - Tạo lại cấu trúc không gian của protein sau khi vận chuyển qua màng tế bào. - Lắp giáp các chuỗi polipeptit vào phức để tạo cấu trúc bậc 4. - MGPT được tăng cường nhờ quá trình tổng hợp trong điều kiện cực đoan do nhu cầu cấp thiết của tế bào. Hoạt động của MGPT được thực hiện nhờ việc sử dụng năng lượng ATP. MGPT được nghiên cứu đầu tiên ở thực vật và liên quan đến enzyme Ribulose bisphotphat cacboxylase/oxygenelase (RUBISCO). RUBISCO xúc tác các phản ứng tổng hợp chất hữu cơ nhờ sử dụng CO2 tự do trong quang hợp. RUBISCO có cấu trúc bậc 4 gồm từ 4- 8 tiểu đơn vị lớn (MW52) và 8 tiểu đơn vị nhỏ (MW14 kDa). Một loại protein đặc hiệu gắn với những tiểu đơn vị chính tạo phức RUBISCO-binding protein (RBP) và RBP là MGPT. MGPT ở thực vật tham gia tạo cấu trúc không gian đúng cho những phân tử protein mới được tổng hợp, chuyển protein qua màng, duy trì cấu trúc đặc hiệu của protein, ngăn chặn sự huỷ hoại protein chưa tạo cấu trúc đúng, khởi đầu sự phân giải protein bị biến tính. MGPT có thể phân thành một số nhóm chủ yếu sau:
• HSP70 là nhóm MGPT có hoạt tính ATP-ase và có tính bảo thủ cao. Trong điều kiện cực đoan HSP70 có thể ngăn chặn sự co cụm và kết tụ protein.
• HSP60 (chaporonin) là loại protein cấu tạo từ các tiểu đơn vị có khối lượng 60 kDa và cũng có hoạt tính ATP-ase. Chúng có cấu tạo 2 vòng,
64

mỗi vòng có từ 7-8 tiểu đơn vị. Ở tế bào Prokaryot, chaperonin giống với protein tìm thấy ở ti thể, lục lạp và được gọi là GroEL, GroES. Trong tế bào Eukaryot tìm thấy một loại chaperonin trong tế bào chất là CCT. Nghiên cứu về CCT thực vật còn ít được đề cập. Tuy nhiên gần đây đã có những công bố đầu tiến của Nông Văn Hải và cộng sự (1998), Trần Thị Phương Liên và cộng sự (1999) về kết quả phân lập và tách được dòng gene CCT từ cây đậu tương. δ
1.1.3. HSP100, HSP90, SHSP Các nhóm HSP100, HSP90, sHSP có tính bảo thủ cao và có hoạt tính ATP-ase. Phần lớn các protein này được sinh ra khi.gặp nhiệt độ cao. Chức năng của HSP100, HSP90, SHSP là ngăn chặn hiện tượng co cụm của protein và tái hoạt hoá các phân tử protein đã biến tính. 1.1.4. LEA (Late embryogenesis abundant protein) LEA là loại protein được tổng hợp với số lượng lớn trong giai đoạn cuối của quá trình hình thành phôi. LEA có vai trò quan trọng đối với khả năng chịu khô hạn của hạt. Cấu trúc phân tử của LEA nổi bật là giàu axit amin ưa nước, không chứa xystein, tryptophan, có vùng xoắnα và có khả năng chịu nhiệt. Chúng thực hiện các chức năng cô lập ion, bảo vệ màng, huỷ protein biến tính và điều chỉnh áp suất thẩm thấu. LEA được chia làm 6 nhóm: nhóm 1-D19; nhóm 2-D11 (dehydrin); nhóm 3-D7; nhóm 4- D113; nhóm 5-D29; nhóm 6-D95. Nhiều gene LEA đã được nghiên cứu, phân lập và xác định chức năng. Khi gặp hiện tượng mất nước mARN của nó được tổng hợp và dần trở thành một lượng lớn gồm nhiều loại mARN khác nhau trong hạt chín, mARN bị phân giải hết khi hạt nảy mầm. Mức độ phiên mã của gene LEA được điều khiển bởi ABA và độ mất nước của tế bào. 1.2. Protein bất hoạt ribosome (Ribosome inactivating proteins - RIPs) Protein bất hoạt ribosome (ribosome inactivating proteins - RIPs) là những độc tố (toxin) có hoạt tính rRNA N-glicosidase và các đặc tính sinh - dược học rất đa dạng. Cơ chế tác động của RIPs là phân cắt cầu nối N- glicosidic của adenine đặc hiệu trên vùng Sarcin/Ricin Loop (còn được gọi là SRL hay R/S Domain) của rRNA (vị trí A4324 trên rRNA 28S ở gan chuột). Kết quả là ribosome không gắn kết được với EF-2 và GTP, làm ức chế quá trình sinh tổng hợp protein. RIPs được phân loại theo cấu trúc phân tử của chúng theo hai hoặc ba tipe khác nhau: (1) RIPs tipe I chỉ có một chuỗi polipeptit A (đôi khi ở dạng glicosyl hoá) với hoạt tính RIPs N-glicosidase; (2) RIPs tipe II có hai chuỗi: polipeptit A liên kết với chuỗi plipeptit B (là một phân tử có hoạt tính giống lectin) bằng một
65

hoặc hơn các cầu nối disulfide; (3) RIPs tipe III giống tipe I, bao gồm một chuỗi polipeptit, nhưng lại chỉ có hoạt tính sau khi được phân cắt đoạn tín hiệu bằng một loại protease đặc hiệu. Số lượng RIPs tipe I (khối lượng phân tử khoảng 30kDa) nhiều hơn RIPs tipe II và III. Tuy nhiên một số RIPs không thể phân loại theo các tipe kể trên. RIPs được phát hiện ở hơn 50 loài thuộc 13 họ thực vật, bao gồm cả cây một và hai lá mầm. RIPs không chỉ thấy ở họ Bầu bí (Cucurbitaceae) với rất nhiều loài là các cây thuốc quý như Trichosanthes, Luffa, mà còn thấy ở rất nhiều các họ thực vật khác như Cẩm chướng (Caryophylaceae), Thầu dầu (Euphorbiaceae), Hoà thảo (Poaceae), Họ đậu (Fabaceae), họ Hoa phấn (Nygtaginaceae)... RIPs không chỉ tìm thấy ở thực vật, mà còn thấy ở một số loại vi khuẩn đường ruột. Sự có mặt của RIPs ở rất nhiều cơ thể sinh vật khác nhau đã đưa đến một giả định về khả năng tồn tại của loại protein tương tự ở thế giới động vật.
§2. QUY TRÌNH XÁC ĐỊNH CHỈ THỊ PHÂN TỬ PROTEIN Protein là sản phẩm của sự biểu hiện gene. Protein biểu hiện tính trạng của cơ thể. Nghiên cứu xác định cơ sở protein của tính trạng hình thái, cấu tạo, sinh lí, hoá sinh của cơ thể sinh vật được quan tâm một số khía cạnh cụ thể như hàm lượng protein, các tiểu phần protein với kích thước phân tử khác nhau. Quy trình xác định chỉ thị protein cho một tính trạng nào đó có thể được thực hiện theo các bước sau: Bước 1. Nghiên cứu đặc tính hình thái, sinh lí Sàng lọc, phân lập các cá thể theo kết quả đánh giá dựa trên kiểu hình. Những cá thể mang tính trạng quan tâm có thể được tạo ra do lai giống hoặc đột biến thực nghiệm, sàng lọc trong quần thể tự nhiên và cũng có thể do xử lí, đánh giá trong điều kiện nhân tạo. Hiện nay người ta quan tâm chủ yếu các tính trạng chung chịu ở thực vật (khả năng chịu nhiệt: nóng, lạnh; mặn; hạn; phèn; khả năng chịu sâu, kháng bệnh), các bệnh do viêm hoặc ung thư ở động vật. Bước 2. Xác định chỉ thị protein
• Chỉ thị hàm lượng protein - Chiết protein ở giai đoạn sinh trưởng, phát triển quan tâm của cá thể. - Phân tích hàm lượng protein. - Xác định mối liên quan đến tính trạng.
66

• Chỉ thị phân tử protein - Chiết protein dùng cho điện di, tách chiết phân đoạn protein. - Phân tích thành phần điện di. - Phân tích đa hình protein. - Giải trình tự axit amin của protein.
• Western blot - Chiết protein dùng cho điện di. - Tách protein bằng điện di trên gel poliacrylamide. - Chuyển protein từ gel sang màng (PVDF) nhờ điện trường.
• Chỉ thị enzyme - Chiết enzyme.
- Phân tích định lượng và định tính hoạt độ enzyme. - Phân tích điện di entyme.
Hình 5.3. Sơ đồ phân tích xác định chỉ thị protein
§3. KĨ THUẬT PHÂN TÍCH PROTEIN 3.1. Tách chiết protein Protein có tính tan trong nước và muối, do vậy người ta chiết protein bằng một số loại dịch chiết có độ pH khác nhau kết hợp với phương pháp li tâm thu
67

dịch chiết. Sau đây trình bày cách chiết protein sử dựng cho định lượng và điện di protein. 3.1.1. Tách chiết protein dự trữ trong hạt thực vật Protein dự trữ trong hạt có thể được chiết bằng các loại dịch chiết như nước cất, NaCl, dung dịch đệm phosphat xitrat pH = 10... Đối với các loại hạt có dầu thì trước khi chiết protein cần phải loại lipit, ví dụ kĩ thuật loại loại lipit ở hạt đậu tương hoặc đậu xanh: Hạt đậu bóc vỏ, bỏ phần phôi mầm sau đó nghiền nhỏ. Bột nhận được giữ ở 40C. Loại lipit bằng petrolium ete qua đêm ở 40C. Li tâm 14.000 vòng/phút để loại dịch li tâm. Lặp lại một vài lần nữa. Bột đã loại lipit giữ ở 40C. 3.1.1.1. Chiết protein dùng để phân tích định lượng hoặc điện di Bột đã loại lipit được sử dụng chiết protein tổng số bằng dung dịch NaCl 1M pH = 7.0, chiết bằng nước cất pH = 7.0 hoặc chiết bằng đệm photphat pH=10. Bột đã loại lipit được chiết bằng dung dịch đệm qua đêm ở 40C, trong 24 giờ. Ngày hôm sau li tâm 14.000 vòng/phút trong 30 phút thu dịch phía trên. Dịch li tâm thu được chứa protein tổng số bao gồm albumin và globulin. Trong định lượng protein có thể tiến hành lặp lại nhiều lần; còn trong phân tích điện di thì có thể thực hiện một lần. Dung dịch protein được giữ ở -200C. 3.1.1.2. Chiết phân đoạn protein cho phân tích điện di Tách chiết và phân đoạn protein theo phương pháp Osborne và Klimenko. Có thể lấy ví dụ ở cây họ đậu: Hạt được nghiền nhỏ thành bột mịn, loại lipit bằng ete petroleum sau đó tiến hành phân đoạn. Protein được chiết bằng dung dịch NaCl 1M và nước cất. Theo cách thứ nhất, bột đã loại lipit được chiết bằng NaCl 1M sau đó li tâm 14000 vòng/phút trong 20 phút. Dịch li tâm chứa albumin và globulin tổng số (phân đoạn I). Cặn li tâm chứa tinh bột và tạp chất được loại bỏ. Tiếp theo albumin và globulin tổng số được phân đoạn bằng cách thẩm tích, sau đó li tâm. Dịch li tâm thu được chứa phần lớn albumin (phân đoạn II). Cặn li tâm là các protein không tan trong nước mà tan trong NaCl. Phần này được hoà vào NaCl 1M (phân đoạn III). Theo cách thứ hai, protein được chiết bằng nước cất sau đó li tâm. Dịch li tâm chứa phần lớn albumin và một phần globulin (phân đoạn IV).Cặn li tâm được chiết tiếp bằng NaCl 1M và li tâm lấy dịch (phân đoạn VII) Phân đoạn IV được xử lí tiếp bằng thẩm tích. Dịch li tâm chứa phần lớn albumin (phân đoạn V). Cặn li tâm chứa phần lớn globulin tan trong NaCl 1M (phân đoạn VI). 3.1.2. Tách chiết protein trong mô động vật Chiết protein từ mô động vật đơn giản hơn ở thực vật và được tiến hành
68

tương tự như ở thực vật. Protein trong mô động vật được chiết nhờ muối hoặc nước và bằng li tâm. 3.2. Định lượng protein 3.1.1. Phân tích hàm lượng nitơ tổng số bằng phương pháp Keldjal Nguyên tắc : Tất cả các dạng nitơ có trong cơ thể hay trong các mô được gọi là nhờ nitơ tổng số. Nitơ có trong thành phần axit amin của protein là nitơ protein. Nitơ không có trong thành phần protein như muối vô cơ, axit nitric... là nitơ phi protein. Nitơ tổng số = nitơ protein + nitơ phi protein. Oxi hoá nguyên liệu bằng H2SO4 có mặt chất xúc tác và nhiệt độ cao, kết quả của việc công phá mẫu này là đạm trong thành phần chất hữu cơ chuyển thành (NH4)2SO4, dưới tác dụng kiềm mạnh, nóng NaOH, sục NH3 bay lên gặp nước tạo thành NH4OH, NH4OH được H2SO4 thụ, Sau đó chuẩn độ lượng NH3 được H2SO4 bằng NaOH. Kết quả phân tích sẽ thu được hàm lượng mía nitơg số bao gồm cả mùi nitơtein và nitơ phi protein. Do vậy cần tiến hành phân tích hàm lượng nitơ phi protein để xác định hàm lượng nitơ protein mà chúng ta quan tâm trong đánh giá chất lượng sản phẩm sinh học. Xác định hàm lượng nitơ phi protein có thể được xác định theo phương pháp của Barstein trên máy phân tích đạm tự động Vapodest của hãng Gerhardt và máy Kjeldahl. Hàm lượng nhơ pitotein = Hàm lượng nitơ tổng số - Hàm lượng nitow phi protein 3.1.2. Phân tích hàm lượng protein tan tổng số Nguyên tắc chung là sử dụng các thuốc thử như Folin, Bradford, Gordan... với sự hấp thụ ở những bước sóng khác nhau trên máy quang phổ Uvis Đồng thời với sự phân tích mẫu thí nghiệm là sử dụng protein chuẩn đã biết trước hàm lượng để tính hàm lượng protein của mẫu thí nghiệm.
• Phương pháp định lượng protein tan tổng số theo phương pháp Lowry. Cơ sở của phương pháp là sự hình thành phức chất biurê. Phức này tham gia với thuốc khử Folin-ciocalteu để cho màu đặc trưng. Cường độ màu tỉ lệ với hàm lượng protein. Đo cường độ màu của hỗn hợp trong ống nghiệm trên máy so màu quang điện ở bước sóng λ = 750nm. Sau khi đo được mật độ quang thì đem đối chiếu với đồ thị chuẩn để tìm lượng protein tương ứng. Thí nghiệm để xây dựng đồ thị chuẩn thì dịch phân tích là dung dịch của một loại protein cụ thể (ở đây là Albumim) đã biết trước hàm lượng.
• Phương pháp sử dụng thuốc thử Bradford Thuốc thử Bradford gồm: Coomasie Brilliant Blue G, ethanol, H3PO4,
69

nước cất; protein chuẩn. Các bước tiến hành: - Pha thuốc thử Bradford. - Sử dụng protein chuẩn và thuốc thử Bradford để xây dựng đường chuẩn và đo trên máy quang phổ ở bước sóng 595nm. - Căn cứ vào đường chuẩn xây dựng được sẽ xác định được hàm lượng protein của mẫu thí nghiệm. Ví dụ: Từ kết quả đo trên máy quang phổ, sử dụng máy vi tính theo chương trình excel lập đường hồi quy tuyến tính y = ax +b (y : mật độ quang; x : hàm lượng protein). Căn cứ vào đồ thị chuẩn y = 1,039. x + 0,0031 sẽ tính được hàm lượng protein của mẫu thí nghiệm. Giả sử khi đo mẫu thí nghiệm với thuốc khử Folin-Ciocalteu trên máy quang phổ ở bước sóng 750nm được trị số y = 0,234 thì thay vào phương trình y = 1,039. x + 0,0031 ta được hàm lượng protein : x = (0,234 - 0,0031)/1,039 = 0,22 mg/ml. Tuy nhiên, khi phân tích trên máy quang phổ bởi phương pháp Bradford hoặc Lowry thì căn cứ vào các số liệu lập đường chuẩn với albumin nguyên chất máy tự động lập phương trình và tính toán cho ta kết quả về hàm lượng protein. 3.3. Phân tích thành phần axit amin Xác định hàm lượng từng loại axit amin trong mẫu thí nghiệm bằng phương pháp sắc kí trên giấy hoặc hiện nay thường sử dụng máy phân tích axit amin tự động Biochrom 20 của hãng Pharmacia Biotech hoặc trên máy HP- Amino - Quant (Amino quant Series II - Operotor Handbook). Sử dụng OPA (ortho-phthaladehyde) tạo dẫn xuất đối với axit amin bậc I và FMOC (9- Fluoreryl methyl chlorofarmate) đối với các axit amin bậc II. Mẫu được xử lí theo phương pháp thủy phân ở 1050C trong 24 giờ. Tuỳ theo mục đích nghiên cứu mà có thể xác định hàm lượng axit amin tự do hoặc liên kết. Trong mục đích nghiên cứu tìm mối liên quan giữa hàm lượng của một loại axit amin nào đó với đặc tính chống chịu hoặc với tính trạng chất lượng (tính thơm của hạt thực vật chẳng hạn) thì có thể xác định hàm lượng axit amin tự do trong tế bào. Còn nếu mục đích nghiên cứu là đánh giá chất lượng protein của sản phẩm thì xác định hàm lượng axit amin liên kết. Căn cứ vào hàm lượng protein và hàm lượng từ loại axit amin xác định được thành phần axit amin trong protein. Thành phần axit amin trong protein được tính theo gam axit 70

amin/100g protein. 3.4. Kĩ thuật điện di protein Các loại protein có khối lượng phân tử khác nhau, do vậy chúng có tốc độ di chuyển khác nhau trong điện trường, vì thế có thể tách được các loại protein nhờ phương pháp điện di. Từ kết quả phân tích thành phần điện di mà người ta có thể đạt được những mục đích nghiên cứu khác nhau. Kĩ thuật điện di protein bao gồm phương pháp điện di một chiều và điện di hai chiều. Các kĩ thuật này cho phép phân tách hỗn hợp phức hệ protein thành những tiểu phần khác nhau dựa trên nguyên tắc tốc độ dịch chuyển khác nhau theo khối lượng phân tử của các tiểu phần đó. Nhờ kĩ thuật điện di một chiều người ta có thể tách được các tiểu phần protein trong protein tổng số tan trong dung dịch NaCl 1M và protein tan trong nước. Dựa vào các băng điện di có thể so sánh thành phần điện di giữa các mẫu nghiên cứu, xác định tính đa dạng sinh học trên cơ sở hiện tượng đa hình protein. Bằng phương pháp điện di protein SDS-PAGE người ta đã phát hiện những sai khác về các băng điện di ở kích thước phân tử 10-16kDa giữa các thể đột biến ở 5 giống lúa khác nhau, xác định được sự khác nhau về thành phần và hàm lượng protein chiết từ lá cây 10 ngày tuổi của 12 thể đột biến đậu tương. Các kết quả nghiên cứu về thành phần điện di protein dự trữ trong hạt đậu tương và trong hạt nảy mầm, nghiên cứu thành phần diện di enzym protease trong hạt nảy mầm của các giống dậu tương chịu hạn khác nhau cho thấy mối liên quan giữa sự thay đổi phổ điện di với khả năng chống chịu của giống đậu tương này. Kĩ thuật điện di protein hai chiều giúp cho các nhà sinh học phân tử phân tách các tiểu phần protein) định tính và định lượng các loại pohpeptide. Kỹ thuật điện di còn được ứng dụng để xác định các thể đột biến, xác định sự đa hình protein. Về mặt lí thuyết kĩ thuật điện di hai chiều có thể phát hiện hai loại đột biến: (1) đột biến do có sự thay thế một hay nhiều nucleotit, mất cặp nucleotit; (2) đột biến do sự biến đổi trình tự nucleotit. Ngoài ra: kĩ thuật điện di protein 2 chiều trên gel poliacrylamid còn được ứng dụng trong việc nghiên cứu biểu hiện gene ở giai đoạn phát triển sớm của cây. xác định được nhiều chỉ thị protein liên quan đến các tính trạng hình thái khác của cây trồng... Kĩ thuật điện di protein có thể theo các bước sau:
Khi thực hiện phân tích điện di protein, trước hết người ta chế tạo bản gel
71

điện di protein (ví dụ gel poliacrylamid). Tra dịch chứa protein đã biến tính vào "giếng" trên bản gel poliacrylamid, đặt hai đầu mép gel lên diện trường, chạy điện di với hiệu điện thế khác nhau trong 1,5 - 2 giờ. Gel được cố định bằng tricloaxetic, ngâm vào dung dịch nhuộm màu coosmasie blue R-250. Sản phẩm điện di protein được chụp ảnh để phân tích. Thiết bị điện di bao gồm: máy điện di và bộ nguồn (hình 5.4). Máy điện di có thể là điện đi đứng hoặc điện di ngang. Gel sử dụng điện di protein hoặc phân tích điện di các enzyme có thể là poliacrylamid chứa SDS hay không có SDS. Ngoài ra có thể là gel thạch agar, gel agarose... Bảng 5.1. Thành phần của gel poliacrylamid Thành phần nhân bản gel
% gel H2O (ml) 30% Degassed acylamid/Bis (ml)
Gel buffer (ml)
SDS (ml)
Gel cô mẫu 4% 6,1 1,3 2,5 0,1 Gel phân tách protein
12% 3,4 4,0 2,5 0,1
Kĩ thuật chế gel điện di protein bao gồm các bước: (1) Lắp ráp khuôn gel; (2) tạo gel; (3) đổ gel.
Hình 5.4. Thiết bị phân tích điện di protein Phân tích hình ảnh điện di protein cho kết quả thống kê các băng điện di của các đối tượng nghiên cứu, từ đó xác định được tính đa hình protein của các đối tượng nghiên cứu. Dựa vào thang protein chuẩn sẽ xác định được các băng
72
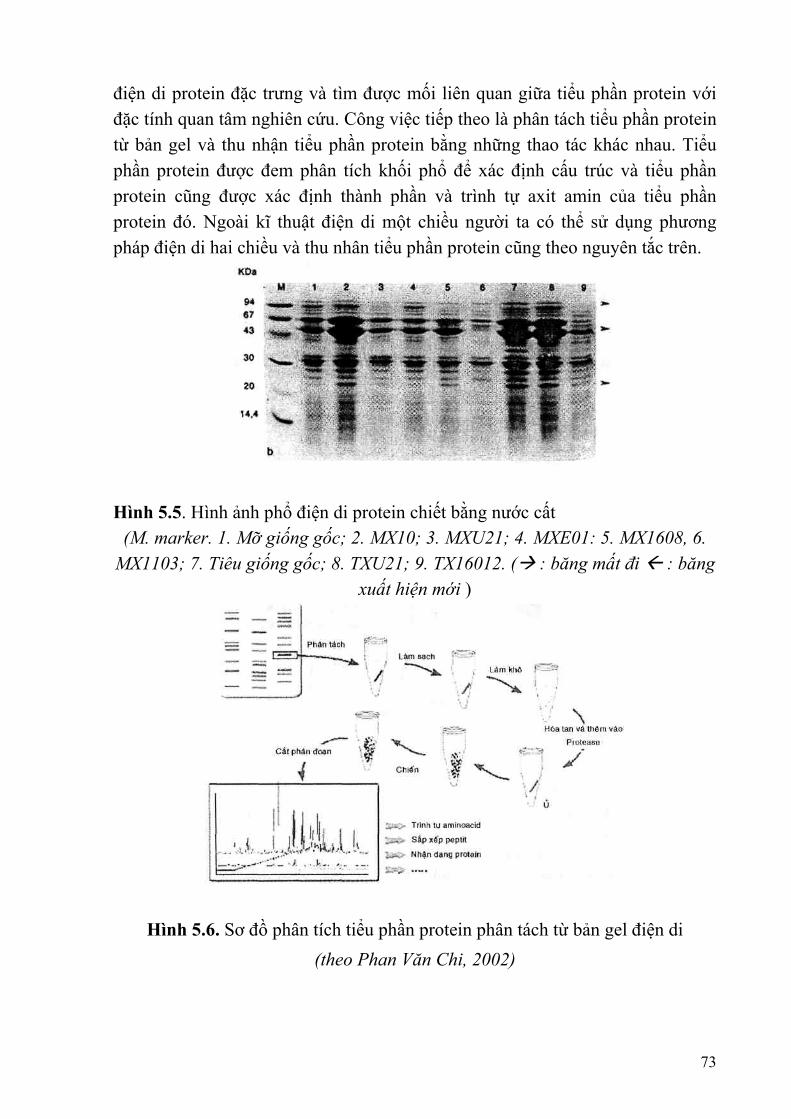
điện di protein đặc trưng và tìm được mối liên quan giữa tiểu phần protein với đặc tính quan tâm nghiên cứu. Công việc tiếp theo là phân tách tiểu phần protein từ bản gel và thu nhận tiểu phần protein bằng những thao tác khác nhau. Tiểu phần protein được đem phân tích khối phổ để xác định cấu trúc và tiểu phần protein cũng được xác định thành phần và trình tự axit amin của tiểu phần protein đó. Ngoài kĩ thuật điện di một chiều người ta có thể sử dụng phương pháp điện di hai chiều và thu nhân tiểu phần protein cũng theo nguyên tắc trên.
Hình 5.5. Hình ảnh phổ điện di protein chiết bằng nước cất (M. marker. 1. Mỡ giống gốc; 2. MX10; 3. MXU21; 4. MXE01: 5. MX1608, 6.
MX1103; 7. Tiêu giống gốc; 8. TXU21; 9. TX16012. ( : băng mất đi : băng xuất hiện mới )
Hình 5.6. Sơ đồ phân tích tiểu phần protein phân tách từ bản gel điện di (theo Phan Văn Chi, 2002)
73

§4. KĨ THUẬT PHÂN TÍCH ENZYME Phân tích enzyme nói chung theo hai cách tiếp cận, đó là xác định hoạt độ enzyme và phân tích phổ enzyme. Có thể tiến hành xác định hoạt độ của enzyme thông qua định lượng hoặc định tính. Đó là xác định lượng cơ chất bị phân giải trong một đơn vị thời gian, hoặc xác định thời gian phân giải trong cùng lượng cơ chất, đo vòng phân giải.... Kết quả xác định hoạt độ enzyme sẽ cho thấy mối quan hệ giữa hoạt độ của loại enzyme nào đó với đặc tính nghiên cứu. Dưới đây giới thiệu một vài phương pháp xác định hoạt độ của enzyme. Ở thực vật, khi hạt nảy mầm thì enzyme hoạt động mạnh. Hoạt độ của enzyme tăng dẫn đến sự biến đổi các chất dự trữ. Mức độ hoạt hoá các enzyme phụ thuộc vào tính chất và thành phần hoá học của hạt. Đối với các cây họ Đậu chất dự trừ chủ yếu là protein cho nên hoạt tính protease mạnh hơn các enzyme khác. Kết quả là protein bị phân giải thành các axit amin, các axit amin có thể được sử dụng tổng hợp nên các protein thứ cấp cấu trúc nên chất nguyên sinh của phôi hạt sinh trưởng và cây non, cũng có thể kết hợp với NH3 để tạo nên các amít, đặc biệt là sự nảy mầm của hạt trong tối. Ngoài protein, trong hạt đậu xanh còn có tinh bột, do đó hoạt tính α - amylase ở cây họ Đậu cũng khá cao. Sự tăng hoạt độ α - amylase sẽ làm tăng hàm lượng đường do đó làm tăng áp suất thẩm thấu và tăng khả năng giữ nước. Người ta đã nghiên cứu thấy α - amylase, hàm lượng đường và áp suất thẩm thấu có mối tương quan thuận. Người ta đã nghiên cứu thấy rằng đặc tính hoá sinh và sinh học phân tử của cây đậu tương có khả năng chịu hạn, chịu nóng có liên quan đến vai trò của a-amylase và protease. Hạt sau khi được ngâm nước với nhiệt độ thích hợp và đủ O2 thì các enzyme bắt đầu hoạt động phân giải các chất hữu cơ. Quá trình phân giải protein và tinh bột diễn ra rất phức tạp. Enzyme α -amylase thuộc nhóm hyđrolaza có trong nước bọt, hạt hoà thảo nảy mầm, tụy tạng, trong nấm mốc, vi khuẩn. Enzyme α -amylase phân giải các liên kết 1,4- glucozit ở giữa chuỗi mạch polisacarit, tạo thành các đextrin phân tử thấp. Do đó dưới tác dụng của enzyme này dung dịch tinh bột nhanh chóng bắt màu với dung dịch iot và bị giảm độ nhớt mạnh. Ion canxi có tác dụng làm bền cấu trúc không gian của phân tử enzyme. Enzyme α - amylase tương đối bền với nhiệt hơn các amylase khác nhưng lại kém bền với axit. Protease cũng thuộc nhóm hydrolaza phân giải các liên kết peptit ở giữa chuỗi mạch polipeptit nên còn gọi là endo peptit hydrolaza. Protease đóng vai trò quan trọng trong cơ thể sống. Chúng không chỉ tham gia vào phân giải protein dự trữ trong hạt nhằm cung cấp các axit amin cho thời kì phát triển đầu tiên của cây mà chúng còn tham gia vào quá trình xử lí phân tử protein và hoạt 74

hoá các enzyme khác. Ngoài ra, protease còn tham gia phân giải các protein lạ hoặc bị biến tính khi gặp điều kiện cực đoan như lạnh, hạn, mặn,... Protease trong hạt có thể được tổng hợp từ trước ở dạng tiền chất và tồn tại song song với protein dự trữ, cũng có thể được tổng hợp mới trong quá trình hạt nảy mầm. Sự có mặt của protease trong hạt dự trữ protein của hạt đang nảy mầm là bằng chứng về sự tham gia của chúng vào quá trình phân giải protein, các sản phẩm peptit trong các bào quan đó. Quá trình phân giải protein dự trữ của các cây họ Đậu đều giống nhau. Khi protein bị phân giải hết những không bào lớn được hình thành trong các tế bào dự trữ. Các axit amin được tạo ra trong quá trình này được sử dụng để sinh tổng hợp các protein mới trong mầm non. Quá trình biến đổi protein dưới xúc tác của protease diễn ra đầu tiên là : protein dự trữ ở dạng nguyên thuỷ bị endopeptidaza đặc hiệu cắt một số mối liên kết trên bề mặt của phân tử globulin theo cơ chế "khoá kéo". Sau đó có sự thay đổi cấu trúc không gian của globulin và làm cho chúng trở thành cơ chất dễ phân giải bởi endopeptidaza khác và các exopeptidaza có sẵn trong protein dự trữ trong hạt, tạo thành các polipeptit, oligopeptit ngắn hơn. Tiếp theo, các sản phẩm này bị aminopeptidaza phân giải đến axit amin. Định lượng hoạt độ enzyme α - amylase theo phương pháp Heinkel được thực hiện theo nguyên tắc là: xác định lượng tinh bột bị phân giải dựa trên cơ sở xác định mức giảm cường độ màu của hỗn hợp phản ứng với dung dịch iot. Đơn vị hoạt độ enzyme là lượng enzyme có khả năng phân giải 1 mg tinh bột sau 30 phút ở 300C. Dựa vào tính chất hoà tan trong dung môi hữu cơ, chiết enzyme a- amylase bằng dung dịch đệm photphat xitrat pH = 6,8. Đem cân mẫu và nghiền nhỏ tròng dung dịch đệm photphat xitrat, li tâm lạnh 12000 vòng/phút trong 15 phút thu dịch chứa enzyme α - amylase cho vào tube nhựa để làm thí nghiệm. Thí nghiệm được tiến hành ở ống thí nghiệm, ống kiểm tra, ống đối chứng. Trong ống thí nghiệm, cho 0,25ml tinh bột 1% vào 0,25ml NaCl 0,1% tiếp tục cho thêm 0,5ml dung dịch đệm photphat xitrat pH = 6,8 và 0,25ml dịch chứa enzyme, sau đó lắc đều và để trong tủ ấm 300C trong 30 phút. Sau khi để tủ ấm lấy ra cho thêm 1,25ml dung dịch axit sunfosalisihc 20%, lắc đều. Lấy 1ml từ hỗn hợp trên cho thêm 9ml iot loãng 150 lần lắc đều để 10 phút rồi đo trên máy quang phổ UV Visible Spectrophotometers ở bước sóng 560 nm. Ống kiểm tra: tương tự ống thí nghiệm chỉ khác là cho 1,25ml dung dịch axit sunfosalisilic 20% vào trước sau đó mới cho dịch chứa enzyme và để trong tủ ấm 30 phút. ống đối chứng: tương tự như ống thí nghiệm chỉ khác là thay dịch chứa enzyme bằng nước cất. 75

Kết quả: lấy kết quả ống kiểm tra trừ đi ống thí nghiệm nhân với độ pha loãng, tìm được số mg tinh bột bị thuỷ phân bởi dịch chiết chứa enzyme α - amylase trong 30 phút ở 300C. Cho hỗn dịch (thạch agar, tinh bột, nước) vào bình nón, đun cách thuỷ cho tan thạch, đổ vào đĩa petri dày 4mm, để nguội, đục lỗ đường kính (d-mm). Nhỏ 100 l dịch chiết chứa enzyme vào mỗi lỗ, để tủ lạnh qua đêm để enzyme khuếch tán, chuyển sang tủ ấm ở 30
μ0C trong 24 giờ. Nhuộm lugol trong 5 phút
và tráng bằng NaCl 1N. Đo đường kính vòng phân giải (D-mm). Hoạt tính enzyme a-amylase được xác định bằng: D - d (mm). Xác định hoạt độ enzyme protease theo phương pháp anson cải tiến theo nguyên tắc: dùng cazein làm cơ chất; sau đó diệt enzyme và kết tủa protein chưa bị thuỷ phân bằng axit tricloaxetic. Xác định hoạt độ phân giải protein của enzyme trên cơ sở định lượng sản phẩm được tạo thành trong phản ứng bằng phản ứng màu với thuốc thử Folin-ciocalteau. Cân mẫu và nghiền nhỏ trong dung dịch đệm photphat pH = 6,5. Li tâm 12000 vòng/phút trong 15 phút ở 40C. Dịch thu được sử dụng làm thí nghiệm như sau: Ống thí nghiệm: cho 1ml cơ chất đã đạt 300C trong 10 phút trộn với 0,5ml dịch chứa enzyme protease đã đạt 300C trong 10 phút, sau đó cho thêm 2,5ml tricloraxetic 5%, lắc đều để 30 phút ở 300C. Lọc lấy 0,5ml dịch lọc và cho thêm 2ml Na2CO3 6%, lắc đều rồi cho 0,5ml thuốc thử Foling- Ciocalteu. Để hỗn hợp trên 30 phút rồi đo trên máy quang phổ UV Visible Spectrophotometers ở bước sóng 720nm. Ống kiểm tra: sau khi cho 1ml dung dịch cơ chất, cho ngay 2,5ml dung dịch axit tricloaxetic 5%, lắc đều rồi mới cho 0,5ml dung dịch chứa enzyme protease. Các bước tiếp theo làm tương tự như ở ống thí nghiệm. ống đối chứng: làm tương tự như trên chỉ thay dung dịch chứa enzyme bằng nước cất. Cho hỗn dịch (thạch agar, cazein, nước) vào bình nón, đun cách thuỷ cho tan thạch, đổ vào đ a petri dày 4mm, để nguội, đục lỗ đường kính (d-mm). Nhỏ 100μ l dịch chiết chứa enzyme vào mỗi lỗ, để tủ lạnh qua đêm để enzyme khuếch tán, chuyển sang tủ ấm ở 300C trong 24 giờ. Nhuộm lugol trong 5 phút và tráng bằng NaCl 1N. Đo đường kính vòng phân giải (D-mm). Hoạt tính protease được xác định bằng: D-d (mm). Phân tích isozyme với mục tiêu là phát hiện hoạt tính của một loại enzyme nào đó. Tuy nhiên trong phân tích di truyền, người ta căn cứ vào kết quả phân tích phổ isozyme mà đánh giá được sự đa dạng sinh học của các loài sinh vật nghiên cứu và xác định được mối quan hệ về di truyền mức độ sai khác giữa
76

các đối tượng nghiên cứu. Đại đa số các enzyme có bản chất là protein, cho nên thông thường người ta phân tích phổ isozyme bằng điện di trên gel acrylamid. Căn cứ vào hoạt động của loại enzyme đó mà người ta trộn thêm cơ chất của enzyme đó vào gel. Trong điều kiện nhiệt độ thích hợp sau khi chạy điện di mà enzyme cần phân tích sẽ hoạt động tương tác với cơ chất. Căn cứ vào kết quả nhuộm gel mà phát hiện ra phổ isozyme của loại enzyme đó. Cũng có thể sử dụng màng nitrocellulose để phát hiện hoạt động của enzyme.
§5. NGHIÊN CỨU RIPS BẰNG WESTERN BLOT Western blot là kĩ thuật phát hiện các tiểu phần protein bằng kháng thể đánh dấu. Western blot bao gồm các bước cơ bản sau: Bước 1: Tách protein bằng điện di trên gel poliacrylamid. Bước 2: Chuyển protein từ gel sang màng (PVDF) nhờ điện trường. Bước 3: Nhuộm màng bằng kháng thể đặc hiệu. Bước 4: Nhuộm màng bằng cộng hợp (kháng thể kháng kháng thể gắn enzyme). Bước 5: Bổ sung cơ chất cho enzyme chuyển hóa để có thể nhìn thấy vệt protein phản ứng với kháng thể đặc hiệu trên màng. Enzyme sẽ chuyển hóa cơ chất thành hợp chất có màu có thể nhìn thấy trên màng. Như vậy chỉ vệt protein nào phản ứng với kháng thể thì mới hiển thị trên màng.
Hình 5.7. Sơ đồ phân tích Western blot
Nếu xét về cấu trúc bậc một và các tính chất hoá sinh thì các protein bất
77

hoạt ribosome (RIPs) tipe I là một họ protein tương đối đồng nhất, kiềm tính, khá bền vững. Tuy nhiên chúng vẫn rất khác nhau về đặc tính enzyme và hoạt tính sinh học. Do có các đặc tính sinh - dược học cũng như khả năng ứng dụng rất đa dạng, cho nên việc tìm kiếm những loại RIPs mới có những tính chất lí thú vẫn đang tiếp diễn ở rất nhiều phòng thí nghiệm trên thế giới. Ở nước ta tuy chưa nhiều nhưng đã bắt đầu có những công bố về loại protein này từ nguồn thực vật. Dưới đây là kết quả khảo sát sự phân bố của RIPs tipe I bằng kĩ thuật Western blot trên một số đối tượng thực vật (thuộc họ Cucurbitaceae) như: cây Trichosanthes sp. Bac Kan 8-98 thu thập ở Bắc Kạn, cây Hồng qua (Mảnh bát - Coccinia cordifolia Cugn.), Gấc (Momordica conchinchinensis), Dưa chuột (Cucumis sativus L.), Mướp đắng (Momordica charantia), Thiên hoa phấn (rễ của Trichosanthes kirilowii Maxim). Để tiến hành việc khảo sát và xác định RIPs, protein tổng số từ các mẫu rễ, hạt được chiết rút, dịch chiết protein được lọc qua màng milipore 0,2μm. Protein được tủa lại bằng acetone lạnh theo tỉ lệ 1:1 (w/v) ở 40C. Các bước điện di protein bằng SDS-PAGE (12,5% acrylamide) được tiến hành theo các phương pháp của Laemmli, sau đó thực hiện phương pháp Western blot. Phản ứng chéo giữa kháng thể kháng Trichobakin tái tổ hợp với các mẫu protein trên là rất đặc hiệu và chỉ có ở vùng 23 - 30 kDa. THẢO LUẬN
1. Trình bày đặc điểm và vai trò của một số protein liên quan đến tính chịu nhiệt ở thực vật.
2. Protein bất hoạt ribosome RIPs, phương pháp khảo sát RIPs và ứng dụng của những nghiên cứu này.
3. Xác định mối liên quan giữa protein và tính trạng có thể tiến hành theo quy trình nào?
4. Phương pháp phân tích chỉ thị phân tử protein được phân tách bằng điện di.
78

Chương 6
KĨ THUẬT SINH HỌC PHÂN TỬ TRONG PHÂN TÍCH HỆ GENE Ở SINH VẬT Tóm tắt: Sinh học phân tử là nòng cốt của công nghệ sinh học. Phân tích sinh học phân tử được bắt đầu từ việc thu nhận dịch chiết ADN từ tế bào sống đủ sạch để thực hiện các phân tích tiếp theo. Tuỳ theo vật liệu nghiên cứu mà có phương pháp tách chiết ADN phù hợp. Dung dịch ADN sau khi kiểm tra hàm lượng và độ sạch được sử dụng để phân tích theo những mục đích khác nhau như Southem blot, Northem blot, xác định trình tự ADN, PCR, RFLP, AFLP,... Nội dung của chương gồm 5 vấn đề cơ bản: (1). Phương pháp tách chiết ADN, (2). Lai phân tử, (3). Xác điịh trình tự nucleotit, (4). Kĩ thuật RFLP, (5). Trình tự nucteotit lặp lại. Thành tựu của sinh học hiện đại và những hiểu biết về di truyền học phân tử làm cơ sở cho việc nghiên cứu, xây dựng các kĩ thuật sinh học hiện đại. Những kĩ thuật sinh học phân tử ứng dụng trong phân tích genome của sinh vật đã được công bố trong các tạp chí khoa học và các sách chuyên khảo.
§1. PHƯƠNG PHÁP TÁCH CHIẾT ADN Mọi nghiên cứu và ứng dụng sinh học phân tử đều bắt đầu bằng việc thu nhận một lượng axit nucleic, đủ tinh sạch để tiến hành các thí nghiệm tiếp theo. Điều quan tâm hàng đầu là các kĩ thuật tách chiết axit nucleic là thu nhận các phân tử ở trạng thái nguyên vẹn không bị phân huỷ bởi các tác nhân cơ học hoặc hoá học. Có thể lấy ví dụ các bước tách ADN từ mầm hạt đậu dưới dây:
79
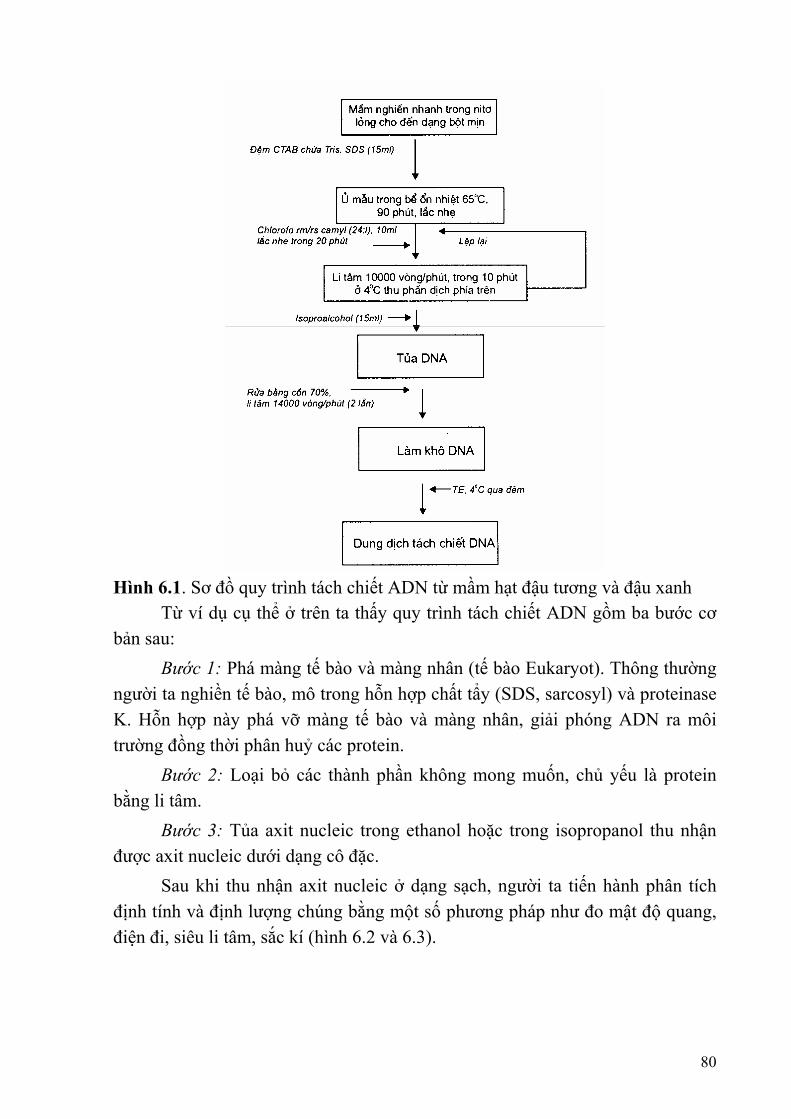
Hình 6.1. Sơ đồ quy trình tách chiết ADN từ mầm hạt đậu tương và đậu xanh Từ ví dụ cụ thể ở trên ta thấy quy trình tách chiết ADN gồm ba bước cơ bản sau: Bước 1: Phá màng tế bào và màng nhân (tế bào Eukaryot). Thông thường người ta nghiền tế bào, mô trong hỗn hợp chất tẩy (SDS, sarcosyl) và proteinase K. Hỗn hợp này phá vỡ màng tế bào và màng nhân, giải phóng ADN ra môi trường đồng thời phân huỷ các protein. Bước 2: Loại bỏ các thành phần không mong muốn, chủ yếu là protein bằng li tâm. Bước 3: Tủa axit nucleic trong ethanol hoặc trong isopropanol thu nhận được axit nucleic dưới dạng cô đặc. Sau khi thu nhận axit nucleic ở dạng sạch, người ta tiến hành phân tích định tính và định lượng chúng bằng một số phương pháp như đo mật độ quang, điện đi, siêu li tâm, sắc kí (hình 6.2 và 6.3).
80
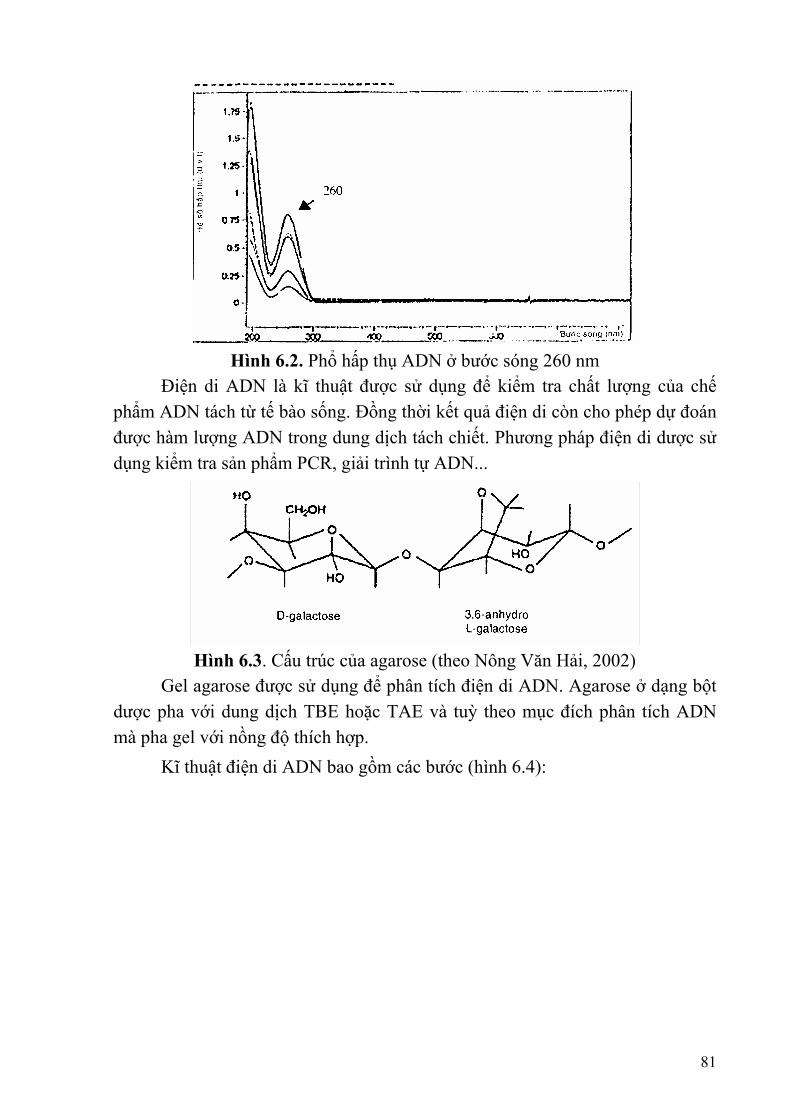
Hình 6.2. Phổ hấp thụ ADN ở bước sóng 260 nm
Điện di ADN là kĩ thuật được sử dụng để kiểm tra chất lượng của chế phẩm ADN tách từ tế bào sống. Đồng thời kết quả điện di còn cho phép dự đoán được hàm lượng ADN trong dung dịch tách chiết. Phương pháp điện di dược sử dụng kiểm tra sản phẩm PCR, giải trình tự ADN...
Hình 6.3. Cấu trúc của agarose (theo Nông Văn Hải, 2002)
Gel agarose được sử dụng để phân tích điện di ADN. Agarose ở dạng bột dược pha với dung dịch TBE hoặc TAE và tuỳ theo mục đích phân tích ADN mà pha gel với nồng độ thích hợp. Kĩ thuật điện di ADN bao gồm các bước (hình 6.4):
81

Hình 6.4. Các bước tạo gel điện di agarose (1, 2) Tạo khuôn ga agarose; (3) Đổ ga trên khuôn ga ; (4) Tra dung dịch ADN ; (5) Chạy điện di.
Sản phẩm điện di ADN được nhuộm bởi EtBr và chụp ảnh dưới ánh sáng đèn cực tím. (1) Nhuộm gel bằng EtBr ; (2) Soi gel trên ánh sáng đèn cực tím để phát hiện sự có mặt của ADN ; (3) Chụp ảnh để phân tích.
82
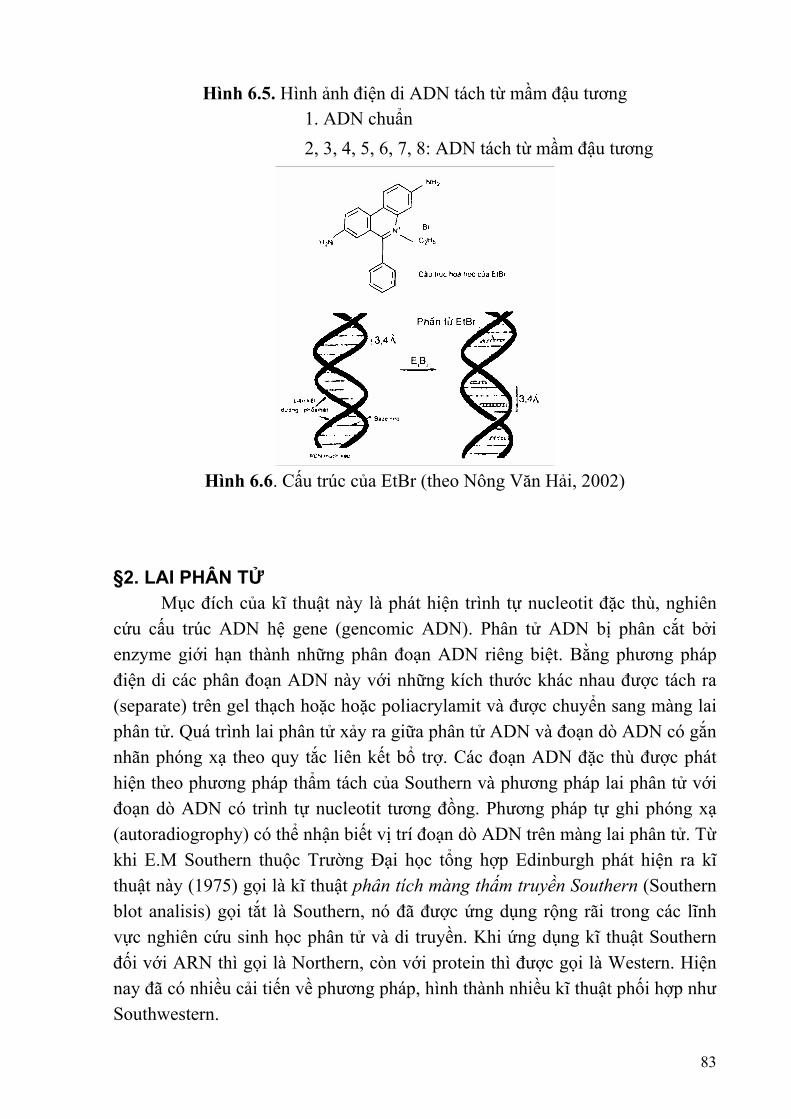
Hình 6.5. Hình ảnh điện di ADN tách từ mầm đậu tương 1. ADN chuẩn 2, 3, 4, 5, 6, 7, 8: ADN tách từ mầm đậu tương
Hình 6.6. Cấu trúc của EtBr (theo Nông Văn Hải, 2002)
§2. LAI PHÂN TỬ Mục đích của kĩ thuật này là phát hiện trình tự nucleotit đặc thù, nghiên cứu cấu trúc ADN hệ gene (gencomic ADN). Phân tử ADN bị phân cắt bởi enzyme giới hạn thành những phân đoạn ADN riêng biệt. Bằng phương pháp điện di các phân đoạn ADN này với những kích thước khác nhau được tách ra (separate) trên gel thạch hoặc hoặc poliacrylamit và được chuyển sang màng lai phân tử. Quá trình lai phân tử xảy ra giữa phân tử ADN và đoạn dò ADN có gắn nhãn phóng xạ theo quy tắc liên kết bổ trợ. Các đoạn ADN đặc thù được phát hiện theo phương pháp thẩm tách của Southern và phương pháp lai phân tử với đoạn dò ADN có trình tự nucleotit tương đồng. Phương pháp tự ghi phóng xạ (autoradiogrophy) có thể nhận biết vị trí đoạn dò ADN trên màng lai phân tử. Từ khi E.M Southern thuộc Trường Đại học tổng hợp Edinburgh phát hiện ra kĩ thuật này (1975) gọi là kĩ thuật phân tích màng thấm truyền Southern (Southern blot analisis) gọi tắt là Southern, nó đã được ứng dụng rộng rãi trong các lĩnh vực nghiên cứu sinh học phân tử và di truyền. Khi ứng dụng kĩ thuật Southern đối với ARN thì gọi là Northern, còn với protein thì được gọi là Western. Hiện nay đã có nhiều cải tiến về phương pháp, hình thành nhiều kĩ thuật phối hợp như Southwestern.
83

2.1. Phương pháp Southern blot Sử dụng một trình tự ADN biết trước lai với đoạn ADN cần tìm. Các bước tiến hành có thể thực hiện theo các bước sau: - Cắt ADN bởi enzyme giới hạn thành các đoạn ADN nhỏ có thể dịch chuyển trên gel agasore. - Biến tính các sợi ADN kép trên gel agarose bằng phương pháp xử lí với kiềm. - Chuyển ADN đã biến tính sang màng nylon hoặc nitrocellulose. - Lai với trình tự ADN đã được đánh dấu bằng phóng xạ hoặc huỳnh quang.
Hình 6.7. Sơ đồ mô tả phương pháp Southern blot
Phương pháp Southern blot cho phép xác định được trình tự đoạn ADN cần tìm, có thể phát hiện được đột biến mất đoạn hoặc thêm đoạn khi so sánh cá thể đột biến với cá thể bình thường. 2.2. Phương pháp Northern blot Phương pháp Northern blot tương tự như phương pháp Southern blt, nhưng mục tiêu của phương pháp Northern blot là nghiên cứu cấu trúc của mARN. Đó là kỹ thuật lai giữa mARN với trình tự ADN biết trước, đánh dấu phóng xạ. (1) Tách chiết ARN tổng số và biến tính. (2). ARN được chuyển sang màng và lai với trình tự ADN đặc hiệu. (3). Vị trí và số lượng các đoạn của ARN lai với ADN được phát hiện trên phim phóng xạ. Phương pháp Northern blot cho phép nghiên cứu cấu trúc ARN, phát hiện đột biến.
84

§3. XÁC ĐỊNH TRÌNH TỰ NUCLEOTIT CỦA ADN Có nhiều kĩ thuật xác định trình tự các đôi bazơ trong một chuỗi ADN: - Kĩ thuật điện đi phân đoạn đánh dấu đầu cuối cho phép xác định trình tự chuỗi ADN sợi đơn. - Xác định trình tự nucleotit thông qua trình tự amino axit dựa vào các bộ ba mã hoá, tuy nhiên số mã của các amino axit thường có một số nucleotit trùng lặp hoặc một số bộ ba cùng quy định một loại amino axit, cho nên trình tự ADN thu được chỉ là tương đối. - Xác định trực tiếp thông qua phản ứng kết thúc đầu cuối. Nguyên tắc chính là tạo ADN sợi đơn, mà phân tử sau có cùng trình tự và chỉ dài hơn phân tử trước một nucleotit ở một đầu cuối. Gene ở tế bào tiền nhân có trình tự liên tục, còn gene ở sinh vật nhân thực có thể gián đoạn. Đoạn nucleotit mã hoá protein được gọi là exon, còn đoạn không mã hoá gọi là intron. 3.1. Phương pháp hoá học của Maxam-Gilbert Nguyên tắc của phương pháp hoá học của Maxam-Gilbert là sử dụng phương pháp đánh dấu phóng xạ (P32), xử lí hoá học để phá huỷ 1 nucleotit tạo ra hàng loạt các đoạn ADN có kích thước khác nhau.
Các đoạn DNA bị phá huỷ 1 nucleotit Điện di trên gel poliacrylamid
Hình 6.8. Sơ đồ minh hoạ phương pháp Maxam-Gilbert Các khâu trong phương pháp Maxam-Gilbert : (1). ADN sợi kép biến tính thành dạng sợi đơn. (2). Đánh dấu phóng xạ (P32) ở đầu 5'. (3). Sử dụng 4 loại hoá chất khác nhau, mỗi loại hoá chất chỉ phá huỷ một loại nucleotit nhất định và tiến hành 4 phản ứng độc lập, kết qủa tạo ra các đoạn ADN kích thước khác nhau.
85
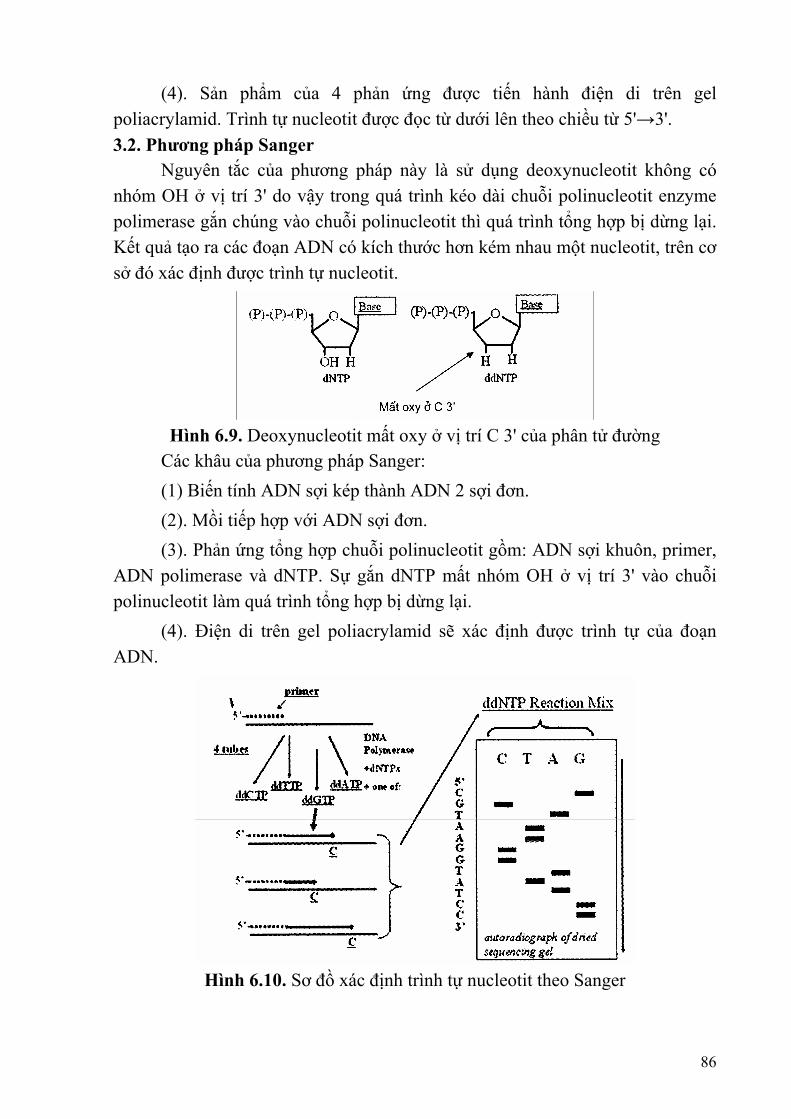
(4). Sản phẩm của 4 phản ứng được tiến hành điện di trên gel poliacrylamid. Trình tự nucleotit được đọc từ dưới lên theo chiều từ 5'→3'. 3.2. Phương pháp Sanger Nguyên tắc của phương pháp này là sử dụng deoxynucleotit không có nhóm OH ở vị trí 3' do vậy trong quá trình kéo dài chuỗi polinucleotit enzyme polimerase gắn chúng vào chuỗi polinucleotit thì quá trình tổng hợp bị dừng lại. Kết quả tạo ra các đoạn ADN có kích thước hơn kém nhau một nucleotit, trên cơ sở đó xác định được trình tự nucleotit.
Hình 6.9. Deoxynucleotit mất oxy ở vị trí C 3' của phân tử đường
Các khâu của phương pháp Sanger: (1) Biến tính ADN sợi kép thành ADN 2 sợi đơn. (2). Mồi tiếp hợp với ADN sợi đơn. (3). Phản ứng tổng hợp chuỗi polinucleotit gồm: ADN sợi khuôn, primer, ADN polimerase và dNTP. Sự gắn dNTP mất nhóm OH ở vị trí 3' vào chuỗi polinucleotit làm quá trình tổng hợp bị dừng lại. (4). Điện di trên gel poliacrylamid sẽ xác định được trình tự của đoạn ADN.
Hình 6.10. Sơ đồ xác định trình tự nucleotit theo Sanger
86

§4. RFLP TECHNOLOGY (Kĩ thuật phân tích hiện tượng đa hình của độ dài các phân đoạn ADN) Trong những năm gần đây kĩ thuật phân lập gene đã dần thúc đẩy cơ bản trong lĩnh vực thú vị nhất là việc sử dụng các đoạn ADN phân lập như các chỉ thị phân tử. Kĩ thuật phân tích hiện đa hình của độ dài các phân đoạn ADN là một kĩ thuật mới mẻ đầy hứa hẹn làm thay đổi hoàn toàn một số lĩnh vực trong đi truyền và chọn giống. Thông tin di truyền được chứa trong các chuỗi ADN của các nhiễm sắc thể nhân và bộ gene của các cơ quan tử. Cây trồng có khả năng sao chép phân tử ADN với chế độ chính xác cao và mau lẹ song có nhiều tác động tự nhiên và nhân tạo gây ra biến dị trong trình tự ADN. Những biến đổi về cặp bazơ nitơ có thể xuất hiện hoặc do biến đổi chuỗi ADN trên phạm vi rộng lớn như là kết quả của sự đảo đoạn, chuyển đoạn và khuyếtđoạn hoặc do chuyển chỗ trong nhiễm sắc thể. Có một số cách xác định biến dị của chuỗi ADN, đó là xác định trình tự ADN và so sánh từng đoạn; sử dụng enzyme cắt hạn chế để cắt ADN thành những phân đoạn có chiều dài khác nhau.
Hình 6.11. Thiết bị tự động xác định trinh tự ADN mao quản Biến đổi trình tự nucleotit tại một locus dẫn đến xuất hiện bằng ADN mới trên gel thạch hoặc poliacrylamid. Hiện tượng này được gọi là đa hình của độ dài các phân đoạn ADN. Hiện tượng đa hình độ dài phân đoạn ADN là các chỉ thị phân tử. Bằng cách sử dụng các chỉ thị phân tử đó (chỉ thị phân tử RFLP) các nhà sinh học phân tử và di truyền chọn giống có thể thiết lập các bản đồ gene liên kết di truyền và định vị các gene có tầm quan trọng về kinh tế. Kĩ thuật RFLP được tiến hành theo các bước sau:
87

(1). Tách ADN từ mô sống. (2). Sử dụng các enzyme cắt hạn chế để cắt ADN thành các đoạn có kích thước khác nhau. (3). Điện di trên gel agarose. (4). So sánh các đoạn ADN giữa các đối tượng nghiên cứu. 4.1. Bản đồ RFLP 4.1.1. Nguyên lí Nguyên lí của kĩ thuật này dựa trên sự khác biệt tự nhiên của các chuỗi nucleotit trong phân tử ADN và khi phân tử ADN bị cắt thành nhiều đoạn nhỏ bởi enzyme cắt hạn chế thì các đoạn ADN có thể khác nhau về kích thước hay chiều dài. Sự khác nhau này có thể được khai thác để phân tích sự phân li của các đoạn nhiễm sắc thể khi lai tạo. Các phân tử ADN có kích thước nhỏ như ADN lục lạp có thể sử dụng enzyme (RE) giới hạn để lập bản đồ RFLP, tuy nhiên khả năng ứng dụng ADN lục lạp trong tạo giống rất hạn chế, vì hầu hết các gene có ý nghĩa kinh tế quan trọng lại nằm trong nhân. Phân tích RFLP có thể ứng dụng với ADN nhân, nhưng rất phức tạp vì ADN nhân có kích thước lớn và hàm lượng nhiều. Khi sử dụng RE đối với ADN thì hàng triệu phân đoạn ADN được tạo thành. 4.1.2. Thư viện chỉ thị RFLP Người ta dùng những đoạn nhiễm sắc thể làm chỉ thị đánh dấu để nhận biết các phân đoạn cắt hạn chế. áp dụng phương pháp lai ADN - ADN có tính đặc hiệu cao bởi việc sử dụng các đoạn ADN mẫu để nhận biết các phân đoạn ADN nhân từ hỗn hợp các đoạn ADN tạo thành bởi việc xử lí RE. Tập hợp các đoạn ADN mẫu đó được gọi là thư viện RFLP. Đoạn ADN mẫu có độ dài 2-5 kb ở trạng thái tinh khiết và được sử dụng để lai phân tử. Các đoạn ADN tinh khiết người ta cần phải thực hiện kĩ thuật nhân gene (gene cloning). Kĩ thuật nhân đoạn ADN có thể được hiện như sau: Đoạn ADN được gắn vào plasmit rồi biến nạp vào vi khuẩn. Vi khuẩn sinh sản dẫn đến đoạn ADN được nhân lên tạo nhiều bản sao đoạn ADN. Bằng kĩ thuật nuôi cấy và phân lập plasmit từ vi khuẩn sẽ tạo ra một số lượng lớn các đoạn ADN dùng làm mẫu cho lai ADN. Quy trình xác định RFLP được thực hiện theo các bước sau : (1) Tách chiết ADN nhân ; (2). Xử lí ADN với enzyme cắt giới hạn ; (3). điện di sản phân cắt bởi RE ; (4). Biến tính ADN thành dạng sợi đơn; (5). Chuyển lên màng lai nitrocellulose (Southern blot) để đoạn nADN mẫu lai với đoạn ADN tương đồng theo nguyên tắc bổ sung giữa các cặp bazơ. Người ta sử dụng kĩ thuật phát quang hoá học (Enhanced Chemical Luminesence = ECL) hoặc đánh dấu phóng xạ để xác định kết quả lai bằng phim âm bản hoặc bằng phóng xạ tự ghi. 88

4.1.3. Thiết lập bản đồ di truyền RFLP Người ta tiến hành lai hữu tính được F1 và cho F1 tự thụ cho F2. Khi F1 giảm phân hình thành giao tử thì các nhiễm sắc thể có sự tái tổ hợp thông qua trao đổi chéo. Trao đổi chéo dẫn đến tái tổ hợp là cơ sở cho việc thiết lập bản đồ di truyền truyền thống (bản đồ liên kết). Kĩ thuật RFLP cho phép quan sát trực tiếp các phân tử chỉ thị ADN trên đoạn nhiễm sắc thể. Các chỉ thị RFLP phân li hoàn toàn chính xác như các chỉ thị gene và tuân theo các định luật Menđen một cách nghiêm ngặt. Vì thế bản đồ RFLP được thiết lập theo nguyên tắc như đối với các chỉ thị truyền thống. Quy trình thiết lập bản đồ di truyền bằng chỉ thị RFLP được thực hiện theo các bước sau: (1) Chọn cặp bố mẹ cùng loài xa nhau về mặt di truyền tạo F1. Tách ADN và sử dụng RE xử lí ADN. Sàng lọc tính đa dạng và cặp bố mẹ nào có mực độ đa dạng thích hợp được chọn làm cặp lai. (2). Tạo quần thể lập bản đồ. Cho cặp bố mẹ lai với nhau được F1, cho F1 tự phối được F2 hoặc có thể lai ngược. Một quần thể F2 hoặc lai ngược có 50 cá thể thì có thể đủ xây dựng bản đồ di truyền chi tiết. (3). Đánh giá RFLP trong quần thể. Tách ADN từng cá thể và tiến hành kĩ thuật RFLP. Quá trình lập bản đồ được tiến hành theo 3 quá trình sau: - Sàng lọc RFLP: Mẫu được thử với cặp bố mẹ để phát hiện ra loại enzyme giới hạn thích hợp tạo ra sự đa dạng RFLP rõ rệt nhất giữa hai cá thể bố mẹ. - Chọn cặp chỉ thị 1enzyme: Chọn cặp chỉ thị /enzyme thích hợp, đó là loại enzyme cắt ADN. Có thể tiến hành như sau: tách ADN ở cây bố, cây mẹ và từng cây con để lập bản đồ. Sử dụng từng loại enzyme cắt, điện di, thấm truyền lên màng lai (mỗi enzyme dùng một bộ màng lai riêng), chọn cặp chỉ thị / enzyme thể hiện sự đa dạng về kiểu gene sẽ dùng để lập bản đồ. - Đánh giá mức độ liên kết: Khi so sánh hai chỉ thị thu được hai kết quả, (1). Chúng có xu hướng cùng phân li và hai chỉ thị có mối liên kết. Ví dụ: F2 dị hợp về chỉ thị 1 và dị hợp về chỉ thị 2 trong khi bố mẹ đồng hợp về cả hai chỉ thị. (2). Sự phân bố tính đồng hợp hay dị hợp của hai chỉ thị là không theo quy luật mà hoàn toàn ngẫu nhiên, như vậy chúng không liên kết. Sử dụng nhiều chỉ thị sẽ xác định được mức độ liên kết khác nhau của từng chỉ thị và nhóm chỉ thị. 4.2. Ứng dụng của RFLP
Việc phát triển kĩ thuật phân tính hiện tượng đa hình của độ dài các phân đoạn ADN đã mở rộng cánh cửa đi tới phát hiện, theo dõi và điều khiển sự biến dị di truyền ở nhiều loại cây trồng mà trước đây không thể làm được (Tanksley
89

và cộng sự, 1989). Những nghiên cứu gần đây về lúa (Mc Couch và cộng sự, 1988), cà chua (Pettersun và cộng sự, 1988, 1990, 1991), đậu tương (Keim và cộng sự, 1990), lúa mì (Sharp và cộng sự, 1989), Ngô (Burr và cộng sự, 1988, Grant và cộng sự 1989, Beavis và cộng sự. 1991, Stuber và cộng sự, 1992) đã chứng tỏ rằng việc lập bản đồ các chỉ thị phân tử RFLP là một phương pháp có sức mạnh lớn lao trong việc xác định các locus điều khiển các tính trạng số lượng - các đặc tính có tầm quan trọng về mặt nông nghiệp. Người ta cũng xác định nhiều tính trạng đơn gene ở cây lúa thông qua liên kết với các chỉ thị phân tử RFLP (Mackill và cộng sự, 1993, Ronald và cộng sự, 1992, Mc Couch và Tanksley, 1991, Yu và cộng sự, 1991, Yoshimura và cộng sự, 1992). Kĩ thuật phân tích hiện tượng đa hình của độ dài các phân đoạn ADN cho phép phát hiện hiện tượng đa hình của bộ gene thông qua so sánh các bản đồ phân cắt ADN giữa các cá thể khác nhau. Các chỉ thị phân tử RFLP mang tính chất riêng biệt, đồng trội và không gây hại. Các chỉ thị phân tử này không chịu tác động bởi điều kiện môi trường (kể cả sự thay đổi về mùa vụ và vị trí địa lí và giai đoạn phát triển của cây. Việc phân tích hiện tượng đa hình của độ dài các phân đoạn ADN được tiến hành trên phân tử ADN tách chiết từ lá non, tránh việc phải chăm sóc cây còn non tới lúc trưởng thành. Người ta đã áp dụng các chỉ thị phân tử RFLP vào công tác chọn giống gián tiếp, làm rút ngắn quá trình chọn lọc các gene quý hiếm, các gene có tầm quan trọng về kinh tế. Bởi lẽ các chỉ thị phân tử RFLP có thể liên kết chặt chẽ với các gene trên và vẫn được duy trì trong quá trình phân li. Vì vậy người ta có thể theo dõi và chọn lọc các gene thông qua các chỉ thị phân tử đó. N. R. Apuyru và cộng sự (1988) đã xác định hiện tượng đa hình của độ dài các phân đoạn ADN của hai giống đậu tương hoang dại Minsoy và Norin 1 bằng cách sử dụng các dòng ADN chọn lọc ngẫu nhiên làm đoạn dò ADN. Các tác giả đã nhận thấy rằng một trong số năm đoạn dò thử nghiệm đã bộc lộ tính đa hình. Hơn nữa số băng đa hình xuất hiện là do có sự sắp xếp lại phân tử ADN. P. Kềnh và cộng sự (1989) đã khảo sát 58 giống đậu tương chọn lọc từ Geneus Glicine và Subgeneus Soja bằng 17 chỉ thị phân tử RFLP để đánh giá mức độ đa dạng phân tử, tính hữu hiệu của các chỉ thị phân tử được thể hiện ít nhất trong số các giống đậu tương dạng trồng và nó được biểu hiện nhiều nhất giữa các giống thuộc các loài đậu tương khác nhau như Glicine max (L) Merr, G. soja Sieb và Zucc. Các chỉ thị phân tử RFLP được áp dụng cho việc lập bản đồ gene, nghiên cứu đa dạng sinh học, phân loại phát sinh chủng loại cho đến điều khiển vật nuôi cây trồng. Nó là một công cụ đắc lực trong công tác lai tạo và chọn giống thực 90

vật. Đồng thời việc xác lập các chỉ thị phân tử RFLP đã mở ra một hướng mới là phân lập các gene mà vẫn chưa biết sản phẩm của nó. THẢO LUẬN
1. Thế nào là lai phân tử? Phân biệt phương pháp Southern blot, Northern blot. Những ứng dụng của các phương pháp này.
2. Vấn đề xác định trình tự ADN. Nguyên tắc và phương pháp. 3. Vấn đề xác định chỉ thị RFLP và bản đồ RFLP.
91

Chương 7
PHẢN ỨNG CHUỖI POLIMERASE(POLIMERASE CHAIN REACTION - PCR) Tóm tắt: Kĩ thuật phản ứng chuỗi dùng enzyme polimerase (Polimerase chain reaction = PCR hay PCR Technology) lần đầu tiên được Mullis và cộng sự mô tả năm 1986 và cũng chính do sự phát minh này mà Mullis đã xứng đáng nhận giải thưởng Nobel về Y học và Sinh lí học năm 1993. Phản ứng chuỗi polimerase (PCR) là kĩ thuật cơ bản và rất quan trọng trong Sinh học phân tử. Phản ứng PCR gồm 3 giai đoạn chính: (1). Biến tính ADN mẫu; (2). Tiếp hợp mồi, (3). Tổng hợp đoạn ADN mới. PCR cũng như các kĩ thuật sinh học phân tử khác cần phải có ADN tinh sạch và chịu ảnh hưởng của nhiều yếu tố. Hiện nay đã có nhiều phiên bản của PCR như RT-PCR, RAPD, AFLP... PCR và các kĩ thuật cải tiến từ PCR có nhiều ứng dụng trong thực tiễn, như: sản xuất mẫu dò, khuếch đại số lượng các đoạn ADN và ARN, định lượng bằng PCR, phân tích đột biến, phân tích ADN đa hình, phân lập gene và tách dòng gene, phân tích sinh vật chuyển gene, xác định quan hệ di truyền... Nội dung của chương gồm 3 vấn đề cơ bản: (1). Phản ứng chuỗi polimerase (PCR); (2). Phân tích tính đa hình của ADN được nhân bản ngẫu nhiên (Random Amplified Polimorphic DNA - RAPD); (3). Phân tích tính đa hình chiều dài các phân đoạn ADN được nhân bản có chọn lọc (Amplifed Fragment Length Potimorphism - AFLP).
§1. PHẢN ỨNG CHUỖI POLIMERASE (PCR) 1. Nguyên tắc Phản ứng chuỗi polimerase là một kĩ thuật đơn giản, cho phép khuếch đại invitro một trình tự ADN lên hàng triệu lần trong vài giờ. Điều này rất thuận lợi cho việc phát hiện những trình tự hiếm trong các bộ gene (đột biến điểm, tái tổ hợp, retrovirus) từ những đoạn ADN ngắn được phân lập hoặc từ một lượng rất nhỏ lấy ra được khuếch đại cho các thử nghiệm sinh học, y học và nông nghiệp. Đồng thời nó giúp cho các nhà sinh học phân tử nghiên cứu cấu trúc trình tự ADN trong bộ gene của các cơ thểsinh vật khác nhau. Phản ứng chuỗi PCR được thực hiện khi có ADN mẫu, 2 đoạn mồi (mỗi đoạn mồi dài 18-30 bazơ), Taq polimerase (ensyme chịu nhiệt được tách từ vi 92

khuẩn Thermus aquaticus):,bốn loại deoxyronucleotit triphotphate ( dATP, dTTP, dGTP, dCTP), dung dịch đệm và con Mg2+ Phản ứng chuỗi PCR được thực hiện trên thiết bị nhân ADN (máy PCR).
Hình 7.1. Thiết bị nhân ADN (Máy PCR) Kĩ thuật PCR là một phương pháp hoàn toàn mới trong việc nghiên cứu và phân tích hệ gen. Sử dụng kĩ thuật PCR có thể tạo ra một số lượng lớn các bản sao của đoạn ADN cần tổng hợp mà không phải tách và nhân dòng. Thực chất PCR là một phương pháp invitro cho phép nhân bản nhanh một đoạn ADN nào đó mà chỉ cần một khối lượng mẫu ban dầu hạn chế. Các thành phần cơ bản của một phản ứng PCR gồm: ADN khuôn, hai đoạn mồi để xác định các điểm bắt đầu tổng hợp ADN, Taq polimerase, hỗn hợp bốn tiền chất deoxynucleotit, dung dịch đệm chứa một số cation hoá trị 1, ion Mg2+ và dung môi (nước khử ion khử trùng). Enzyme Taq polimerase là enzyme chịu nhiệt, được tách từ vi khuẩn suối nước nóng Thermus aquaticus. Enzyme Taq có trọng lượng phân tử là 94 kDa và không mất hoạt tính ở nhiệt độ cao trong giai đoạn gây biến tính ADN. Hoạt tính của enzyme Taq giảm 50% sau 150 phút ở nhiệt 92,50C; sau 40 phút ở nhiệt độ 950C và sau 5-6 phút ở nhiệt độ 970C. Nếu nhiệt độ tăng lên tới 950C trong 20 giây để biến tính ADN kép thành sợi đơn thì hoạt tính enzyme Taq còn lại 65% sau 50 chu kì phản ứng. Nồng độ enzyme Taq tối ưu cho phản ứng PCR là 0,5-2,5 đơn vị, nhưng nếu nồng độ enzyme quá cao sẽ làm giảm hiệu suất xúc tác của phản ứng. Ngoài ra, nồng độ Mg2+ và dNTP cũng ảnh hưởng đến hoạt tính của enzyme. Hiện nay, có nhiều loại polimerase chịu nhiệt khác có chức năng riêng biệt và hoàn thiện hơn như Tth polimerase tách chiết từ Thermus thermophilus, có khả năng hoạt động như một enzyme phiên mã ngược khi có mặt của ARN khuôn và ion Mg2+. ADN khuôn (template) ADN khuôn là vật liệu khởi đầu cho phản ứng PCR nên đòi hỏi có độ tinh
93

sạch cao. ADN khuôn có thể là sợi đơn hoặc sợi đôi của chuỗi ADN được biết trước trình tự ở hai đầu để thiết kế mồi. Thông thường, nồng độ của ADN khuôn được đưa vào phản ứng PCR khoảng 10-500ng. Đoạn mồi (primer) Các đoạn mồi thực chất là các oligonucleotit dài khoảng 4-10 bazơ (đối với mồi ngẫu nhiên) hoặc khoảng 12-24 bazơ (đối với mồi đặc trưng). Nồng độ mồi thích hợp để tiến hành phản ứng PCR là từ 0,1-0,5μM. Lượng mồi đưa vào phản ứng PCR phải phù hợp với lượng ADN cần tổng hợp (thường 106 phân tử ADN cần 108 phân tử mồi). Nếu nồng độ mồi quá thấp thì mồi sẽ hết trước khi số chu kì kết thúc, còn nồng độ mồi quá cao thì sẽ dễ làm tăng sản phẩm không đặc hiệu. Các đoạn mồi cần có tính đặc thù để làm tăng tính đặc hiệu của phản ứng. Mồi phải được thiết kế sao cho không được bổ trợ lẫn nhau, số lượng 4 loại bazơ nitơ trong mồi nên xấp xỉ nhau, lượng G-C giữa hai mồi phải bằng nhau, tránh những vùng trình tự không bình thường như polipurine hoặc poliprimidine hay các trình tự lặp. Các nucleotit (dNTPs) dNTPs là hỗn hợp của bốn loại deoxyribonucleotit (dATP, dTTP, dGTP, dCTP) làm nguyên liệu cho phản ứng tổng hợp ADN. Tuỳ thuộc vào mục đích nghiên cứu, các nhà khoa học còn có thể sử dụng một số nucleotit đã được thay đổi như gắn thêm biotin hoặc digoxygenin... Nồng độ phản ứng của các dNTP thường dùng vào khoảng 200mM mỗi loại, tuy nhiên ở nồng độ dNTP thấp (10-100mM) Taq ADN polimerase hoạt động chính xác hơn. Hơn nữa, nồng độ tối ưu của chúng phụ thuộc vào rất nhiều yếu tố như: Nồng độ Mg2+ nồng độ chất mồi, độ dài của sản phẩm được khuếch đại số chu kì của phản ứng. Thành phần của dung dịch đệm phụ thuộc vào loại enzyme polimerase được sử dụng. Trong dung dịch đệm quan trọng nhất là ion Mg2+ làm tăng nhiệt độ nóng chảy (Tm) của ADN mạch đôi, tạo ra phức chất tan với dNTPs để hình thành cơ chất mà enzyme polimerase có thể nhận ra, điều này rất cần thiết cho quá trình liên kết của các dNTP. Nồng độ Mg2+ trong hỗn hợp phản ứng cuối cùng biến đổi từ 0,5-5,5mM (nồng độ này có thể thay đổi khi cần thiết). Nồng độ lớn Mg2+ quá thấp sẽ làm giảm khả năng tổng hợp của enzyme polimerase, còn nếu nồng độ quá cao sẽ tạo ra những phân đoạn không đặc hiệu. Nhìn chung, nồng độ MgCl2 có ảnh hưởng nhiều tới hiệu quả và tính đặc thù của phản ứng. Ngoài Mg2+ còn một số chất khác trong dung dịch đệm như AMSO, DMSO, formamide được thêm vào như các chất phụ gia nhằm tạo ra các sản phẩm PCR có kích thước lớn. 94

Mỗi một chu kì PCR gồm 3 giai đoạn cơ bản có nhiệt độ khác nhau: (1) Giai đoạn tách chuỗi ADN: ADN bị biến tính từ dạng sợi kép thành dạng sợi đơn ờ nhiệt độ 94-950C trong khoảng thời gian rất ngắn (khoảng 1-1,5 phút).
(2). Giai đoạn gắn mồi: Nhiệt độ phản ứng được hạ thấp xuống 400- 650C cho phép hai đoạn mồi gắn vào ADN mẫu ở vị trí tương đồng theo nguyên tắc bổ sung (A - T, G - C) ở hai đầu ADN cần nhân.
(3) Giai đoạn kéo dài: ở nhiệt độ 720C ADN Taq polimerase hoạt động kéo dài đoạn ADN từ đoạn mồi và hai đoạn ADN mới được tổng hợp theo nguyên tắc bổ sung từ hai đầu đoạn ADN khuôn ban đầu.
Sau một chu kì gồm ba giai đoạn như trên, một đoạn ADN khuôn được nhân lên thành hai, các đoạn ADN được nhân bản trong mỗi chu kì lại được coi là ADN khuôn cho chu kì nhân bản tiếp theo. Chính vì vậy sau k chu kì nhân bản sẽ tạo ra 2K đoạn ADN giống đoạn ADN khuôn ban đầu.
95
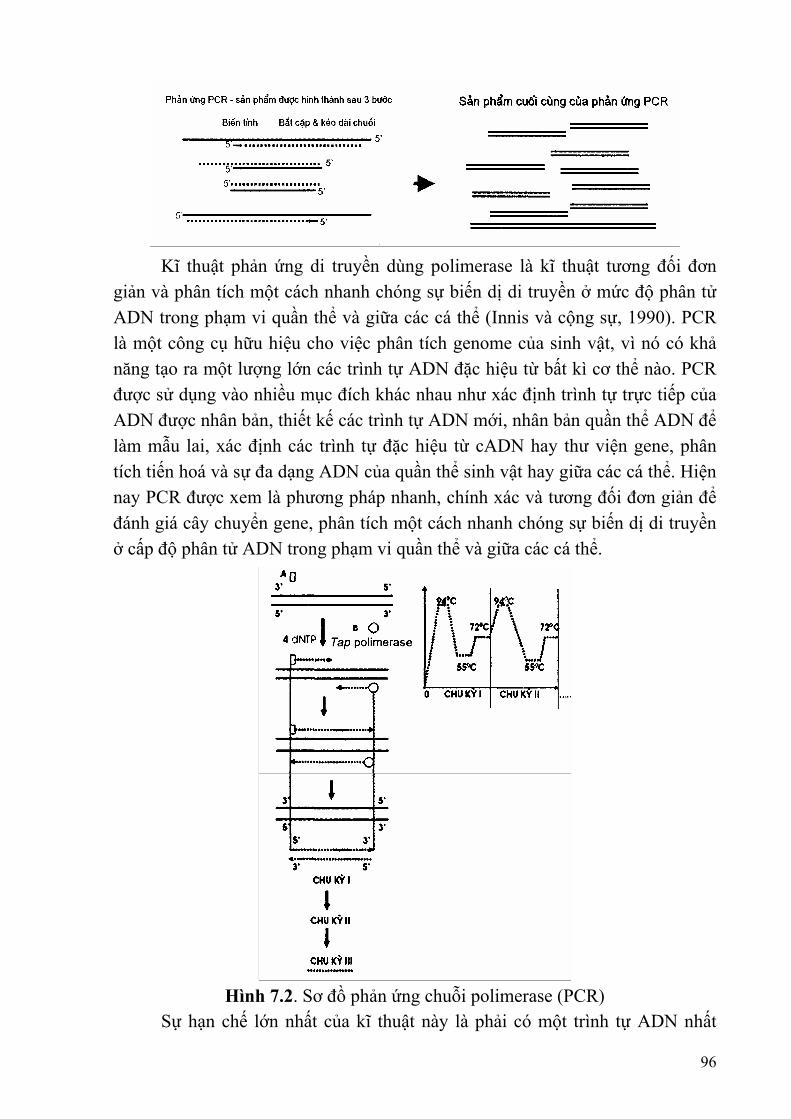
Kĩ thuật phản ứng di truyền dùng polimerase là kĩ thuật tương đối đơn giản và phân tích một cách nhanh chóng sự biến dị di truyền ở mức độ phân tử ADN trong phạm vi quần thể và giữa các cá thể (Innis và cộng sự, 1990). PCR là một công cụ hữu hiệu cho việc phân tích genome của sinh vật, vì nó có khả năng tạo ra một lượng lớn các trình tự ADN đặc hiệu từ bất kì cơ thể nào. PCR được sử dụng vào nhiều mục đích khác nhau như xác định trình tự trực tiếp của ADN được nhân bản, thiết kế các trình tự ADN mới, nhân bản quần thể ADN để làm mẫu lai, xác định các trình tự đặc hiệu từ cADN hay thư viện gene, phân tích tiến hoá và sự đa dạng ADN của quần thể sinh vật hay giữa các cá thể. Hiện nay PCR được xem là phương pháp nhanh, chính xác và tương đối đơn giản để đánh giá cây chuyển gene, phân tích một cách nhanh chóng sự biến dị di truyền ở cấp độ phân tử ADN trong phạm vi quần thể và giữa các cá thể.
Hình 7.2. Sơ đồ phản ứng chuỗi polimerase (PCR)
Sự hạn chế lớn nhất của kĩ thuật này là phải có một trình tự ADN nhất
96

định mà từ đó người ta thiết kế và tổng hợp các đoạn mồi (đoạn khởi đầu cho tổng hợp ADN). Tuy vậy sau này các nhà sinh học phân tử đã cải tiến kĩ thuật này mà không cần thiết phải biết trước trình tự chuỗi ADN. John Welsh và Michael Mc Clelland đã sử dụng kĩ thuật PCR với các đoạn mồi ngẫu nhiên trong việc xác định in dấu genome. Kĩ thuật PCR được ứng dụng để phát hiện nhanh các đột biến điểm và phân tích ADN đa hình. PCR còn được ứng dụng phân lập gene, phân tích sinh vật chuyển gene. Ở Việt Nam các nhà khoa học đã thành công trong việc phân lập các gene liên quan đến khả năng chống chịu ở cây trồng, như gene chịu hạn ở cây lúa (Lê Trần Bình và cộng sự, 1998), gene chịu nóng ở đậu tương (Trần Phương Liên và cộng sự, 1999),... và hàng loạt các gene đã được tách và giải trình tự nucleotit.
Hình 7.3. Hình ảnh điện di sản phẩm PCR (Đinh Duy Kháng, 2002) 1.2. Phản ứng RT-PCR (PCR ngược) 1.2.1. Nguyên lí Thực chất của RT-PCR là phản ứng nhân một trình tự từ khuôn ARN, gồm 2 giai đoạn: (1) Tổng hợp trình tự ADN 2 sợi từ sợi ARN làm khuôn; (2) Trình tự ADN 2 sợi lại làm khuôn để thực hiện phản ứng PCR. Phản ứng biến đổi ARN thành ADN phải nhờ enzyme phiên mã ngược (Reverse-trancriptase) được gọi là giai đoạn chuyển ngược (RT) thực hiện ở nhiệt độ 500C - 550C trong 30 phút. Sau khi có sản phẩm ADN 2 sợi thì phản ứng tiếp theo là PCR thông thường gồm 3 giai đoạn. RT-PCR được sử dụng để nhân ARN của retrovirut hoặc mARN tách từ tế bào chất. 1.2.2. Tiến hành phản ứng RT- PCR - Giai đoạn 1 (RT): 1 chu kì, thực hiện sao chép ngược từ ARN sợi đơn sang ADN sợi kép ở nhiệt độ 500C- 550C, trong 30 phút.
- Giai đoạn 2 (PCR): (1). Biến tính ở nhiệt độ 940-950C /1phút; (2).Tiếp hợp mồi ở 40-650C; (3). Tổng hợp ADN ở 720C/1 phút. 1.3. Kiểm tra sản phẩm PCR Sản phẩm PCR cùng với ADN marker được kiểm tra bằng phương pháp điện ới trên gel agarose 0,8-2%. Tiến hành chụp ảnh trên ánh sáng đèn UV để xác định sự có mặt của đoạn ADN được nhân bởi PCR.
97

Hình 7.4. Hình ảnh điện di sản phẩm RT-PCR (Đinh Duy Kháng, 2002) 1.4. Những yếu tố ảnh hưởng đến phản ứng PCR Kết quả của phản ứng PCR chịu ảnh hưởng của các yếu tử như: ADN khuôn, Taq polimerase, primer và nhiệt độ nóng chảy của mồi, ton Mg++, nguyên liệu là các dNTP, nước, dung dịch đệm,... 1.5. Các lĩnh vực ứng dụng của phản ứng PCR Phản ứng chuỗi trùng hợp PCR, từ khi được Kary Mullis đề xuất hoàn thiện, đã có những ứng dụng hết sức rộng rãi trong nhiều lĩnh vực khác nhau, cả trong khoa học công nghệ và trong đời sống xã hội. Trong nghiên cứu, kĩ thuật PCR giúp ích hữu hiệu cho việc xác định trình tự nucleotit của các đoạn ADN được nhóm có thể sử dụng PCR để phát hiện đột biến, PCR cho phép phân tích liên kết gene từ những tế bào riêng lẻ, giúp nghiên cứu quá trình tiến hoá ở mức phân tử, thậm chí phục hồi những gene đã tồn tại cách đây hàng chục triệu năm. Trong chọn giống vật nuôi và cây trồng, đặc biệt đối với động vật và thực vật bậc cao, vốn trước đây việc lựa chọn cặp cha mẹ làm giống là rất khó khăn. cần nhiều năm, thì nay kĩ thuật PCR cho phép thực hiện chỉ trong vài ngày, thậm chí vài giờ. Trong y học PCR có thể chẩn đoán chính xác các bệnh nhiễm trùng từ virut đến vi khuẩn và các bệnh do nấm, kể cả HIV-AIDS, chẩn đoán sớm và theo dõi điều trị ung thư... Trong tư vấn di truyền y học PCR cho phép chẩn đoán nhanh, chính xác các bệnh di truyền kể cả chẩn đoán trước sinh giới tính và các dị tật bẩm sinh có thể có từ khi thai mới được 8 tuần tuổi và điều quan trọng là không cần chọc ối, chỉ cần lấy một giọt máu ngoại vi của mẹ để làm mẫu thí nghiệm. Ngày nay kĩ thuật PCR là không thể thiếu trong khoa học hình sự, giúp chẩn đoán nhanh, chính xác thủ phạm chỉ từ một vết máu khô, một sợi tóc hoặc một sinh phẩm nào đó mà thủ phạm để lại trên hiện trường. Ngoài ra kĩ thuật này còn cho phép xác định chính xác quan hệ huyết thống cha - con, ông - cháu... cũng chỉ trong vài giờ. 98

Trong bảo vệ môi trường, việc xác định mức độ ô nhiễm sinh học có thể thực hiện rất hiệu quả và nhanh chóng bằng kĩ thuật PCR. Những ứng dụng thực tiễn đa dạng của phản ứng chuỗi trùng hợp PCR thật vô cùng tận. Nếu như năm 1985 mới có 3 công trình được công bố về PCR thì 5 năm sau kĩ thuật này đã được sử dụng trong hàng nghìn phòng thí nghiệm trên khắp thế giới. Ngay ở nước ta, kĩ thuật PCR cũng đã và đang được dùng trong nhiều trường đại học, viện nghiên cứu và ngày càng trở nên phổ biến. Phản ứng chuỗi trùng hợp thật sự đã đưa lại một cuộc cách mạng trong lĩnh vực ứng dụng thực tế của di truyền học phân tử. Tóm lại, những ứng dụng cơ bản của PCR có thể tóm tắt như sau:
- Sản xuất mẫu dò. - Khuếch đại số lượng các đoạn ADN và ARN. - Định lượng bằng PCR. - Phân tích đột biến. - Phân tích ADN đa hình. - Phân lập gene và tách dòng gene. - Phân tích sinh vật chuyển gene.
§2. PHÂN TÍCH TÍNH ĐA HÌNH CỦA ADN ĐƯỢC NHÂN BẢN NGẪU NHIÊN (RANDQM AMPLIFIED PQLIMORPHIC DAN - RAPD) Williams và cọng sự (1990) đã xác định hiện tượng đa hình của phân tử ADN dựa trên phản ứng khuếch đại phân đoạn ADN ngẫu nhiên bằng các đoạn mồi có trình tự ADN tuỳ ý. Kĩ thuật này được gọi là kĩ thuật khuếch đại ngẫu nhiên các đoạn ADN đa hình (RAPD technology). Hiện tượng đa hình các đoạn ADN khuếch đại ngẫu nhiên xuất hiện là do có sự biến đổi trình tự nucleotit tại vị trí các đoạn mồi liên kết (chẳng hạn đột biến điểm) hoặc do biến đổi các nhiễm sắc thể trong khu vực gene khuếch đại (lồng đoạn, mất đoạn). Phương pháp này đơn giản về mặt kỹ thuật, tiến hành nhanh chóng và chỉ cần một lượng rất ít phân tử ADN (nanogram). Các đoạn ADN đa hình được khuếch đại là một loại chỉ thị phân tử đặc thù mà chúng có thể phát hiện hiện tượng đa hình giữa các cá thể khác nhau ở mức độ phân tử ADN. Các mồi có trình tự nucleotit ngắn (10 nucleotit) liên kết bổ trợ với nhiều locus khác nhau và có khả năng khuếch đại nhiều đoạn ADN khác nhau (từ 2
99
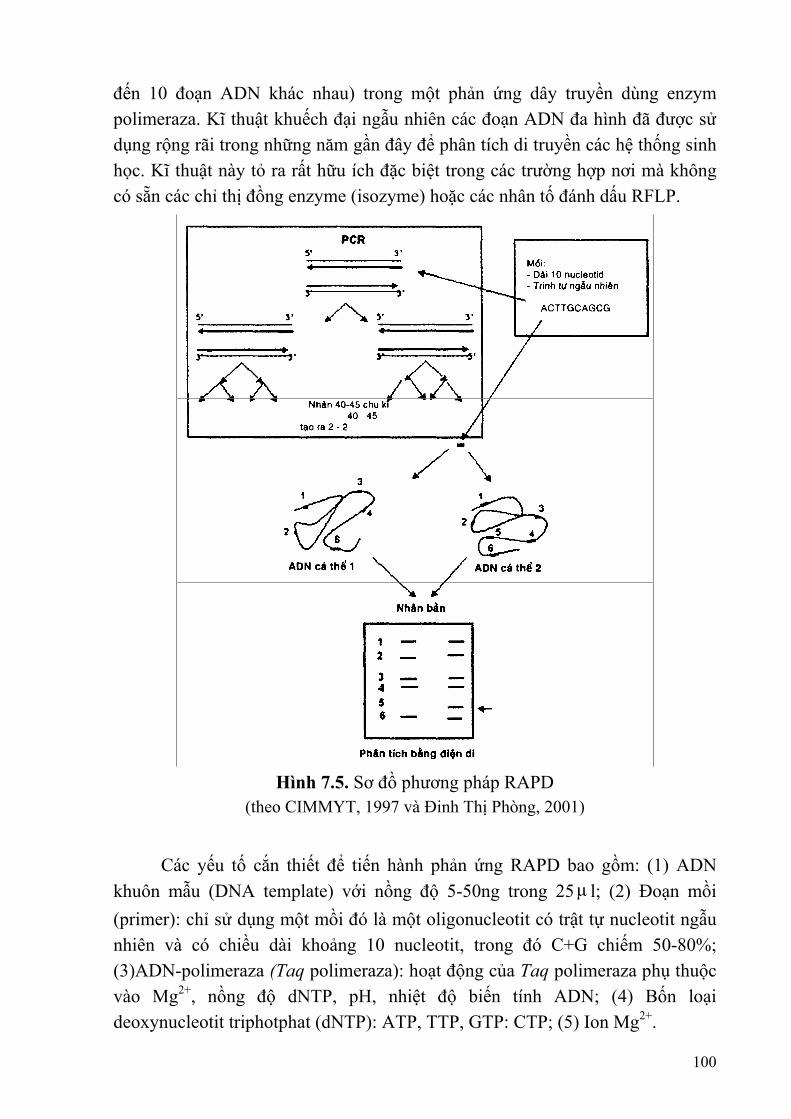
đến 10 đoạn ADN khác nhau) trong một phản ứng dây truyền dùng enzym polimeraza. Kĩ thuật khuếch đại ngẫu nhiên các đoạn ADN đa hình đã được sử dụng rộng rãi trong những năm gần đây để phân tích di truyền các hệ thống sinh học. Kĩ thuật này tỏ ra rất hữu ích đặc biệt trong các trường hợp nơi mà không có sẵn các chỉ thị đồng enzyme (isozyme) hoặc các nhân tố đánh dấu RFLP.
Hình 7.5. Sơ đồ phương pháp RAPD
(theo CIMMYT, 1997 và Đinh Thị Phòng, 2001)
Các yếu tố cắn thiết để tiến hành phản ứng RAPD bao gồm: (1) ADN khuôn mẫu (DNA template) với nồng độ 5-50ng trong 25 l; (2) Đoạn mồi (primer): chỉ sử dụng một mồi đó là một oligonucleotit có trật tự nucleotit ngẫu nhiên và có chiều dài khoảng 10 nucleotit, trong đó C+G chiếm 50-80%; (3)ADN-polimeraza (Taq polimeraza): hoạt động của Taq polimeraza phụ thuộc vào Mg
μ
2+, nồng độ dNTP, pH, nhiệt độ biến tính ADN; (4) Bốn loại deoxynucleotit triphotphat (dNTP): ATP, TTP, GTP: CTP; (5) Ion Mg2+.
100

Phản ứng RAPD được tiến hành qua các giai đoạn giống như PCR: - Giai đoạn biến tính ADN: ở nhiệt độ 950C trong khoảng 30-60 giây làm cho hai mạch khuôn ADN tách nhau. - Giai đoạn tiếp hợp mồi: Khi nhiệt độ hạ xuống 32-400C mồi tiếp hợp và bám vào sợi ADN khuôn. - Giai đoạn tổng hợp: Nhiệt độ được nâng lên 720C thì các đoạn mồi đã bắt cặp với các mạch đơn sẽ được kéo dài với sự tham gia của Taq polimeraza. Sau một chu kì gồm 3 giai đoạn như trên, một đoạn ADN khuôn được nhân lên thành hai, các đoạn ADN được nhân bản trong mỗi chu kì lại được coi là ADN khuôn cho chu kì nhân bản tiếp theo. Vậy sau k chu kì nhân bản sẽ tạo ra 2k các đoạn ADN giống đoạn ADN khuôn ban đầu. RAPD có thể thực hiện từ 40-45 chu kì. Kĩ thuật này khác với kĩ thuật PCR ở chỗ sử dụng các mồi ngẫu nhiên, số chu kì nhiều hơn và ở giai đoạn 2 nhiệt độ hạ thấp xuống 32-360C, không đòi hỏi thông tin về trình tự ADN được nhân bản. Dựa trên sự xuất hiện hay biến mất của các phân đoạn ADN khi điện di sản phẩm RAPD được quan sát thấy ở các cá thể khác nhau và được đánh giá theo quy ước 1 = xuất hiện và 0 = không xuất hiện. Một bảng gồm các giá trị 0 và 1 được thiết lập từ các cá thể nghiên cứu sẽ cho phép tính ra hệ số tương đồng di truyền của các cặp theo Nei và Li. Trong quá trình thiết lập mối quan hệ giữa các loài hay nhóm loài, một số tác giả (Apostol và CS, 1993) đã xây dựng kĩ thuật phân nhóm thông qua các biểu đồ RAPD (RAPD Lot). Thực chất kĩ thuật này gồm 3 bước: Bước 1: So sánh từng cặp đối tượng trong nghiên cứu bằng cách tính toán khoảng cách quan hệ giữa chúng. Bước 2: Lập một ma trận gồm tất cả những giá trị tính toán được trước đó. Bước 3: Giải ma trận và biểu diễn thành một biểu đồ đặc trưng. Ngày nay, các nhà nghiên cứu đã thiết lập được phần mềm máy tính để tự động vẽ nên biểu đồ mối quan hệ hay độ tương đồng di truyền của các đối tượng nghiên cứu sau khi nhập dữ liệu về các phân đoạn được nhân bản của các cá thể. NTSYS pc version 2.0 (Applied Biostatistics lúc., USA., 1998) là tên của một chương trình thuộc kiểu trên để lập ra biểu đồ hình cây Biểu đồ hình cây thu được sẽ thể hiện mức độ gần nhau của các cá thể cho phép đánh giá được mối quan hệ di truyền giữa các cá thể được nghiên cứu. Chương trình này cho phép giảm bớt thời gian nghiên cứu và có độ chính xác cao nên nó là một phần mềm 101

có hiệu quả trong việc phân tích kĩ thuật RAPD. Đối với đa số thực vật, các đoạn mồi dài khoảng 10 nucleotit với 60% GC sẽ tạo ra 2-10 sản phẩm được nhân bản. Hiện tượng đa hình các đoạn ADN được nhân bản ngẫu nhiên xuất hiện là do có sự biến đổi trình tự nucleotit tại vị trí các đoạn mồi liên kết. Vì vậy các đa hình thường được nhận ra do có mặt hay vắng mặt của một sản phẩm nhân bản từ một locut. Phương pháp này đơn giản, nhanh chóng, chỉ cần một lượng nhỏ (nanogram) phân tử ADN có nguồn gốc từ hệ gen. Các đoạn ADN đa hình được nhân bản là một loại chỉ thị phân tử đặc thù mà chúng ta có thể phát hiện hiện tượng đa hình giữa các cá thể khác nhau ở mức độ phân tử ADN. Kĩ thuật RAPD được sử dụng rộng rãi trong những năm gần đây để phân tích di truyền hệ thống sinh học. Nó là phương pháp hiệu quả trong việc xác định kiểu gen, phân tích quần thể và nguồn gốc loài, nghiên cứu di truyền và lập bản đồ di truyền. Người ta đã áp dụng thành công kĩ thuật khuếch đại ngẫu nhiên các đoạn ADN đa hình vào nhiều lĩnh vực khác nhau, như lập bản đồ gen, in dấu ván ADN, chọn giống... Lập bản đồ gene (Gene Mapping) Các nhân tố RAPD cũng phân li theo quy luật Mendel và có thể được sử dụng như các gene đánh dấu di truyền trong việc thiết lập các bản đồ phân tử (Williams và cộng sự, 1990; Foolad và cộng sự, 1993). So với kĩ thuật phân tích hiện tượng đa hình độ dài các phân đoạn ADN (RFLF Technology) thì kĩ thuật này có một số ưu điểm nổi bật là: có thể phát hiện hiện tượng đa hình trong phân tử ADN trên một phạm vi rộng lớn hơn; tính đơn giản và nhanh gọn; cần ít lượng ADN. Kĩ thuật này đã được áp dụng thành công vào nhiều loại cây trồng khác nhau để thiết lập các bản đồ di truyền phân tử như Brassica (Quyros và cộng sự 1991), Arabidopsis thaliana (Reiter và cộng sự, 1992) cà chua (Klein Lankhorst và cộng sự, 1991; Foolad và cộng sự, 1993)... In dấu vân ADN(ADN Fingerprinting) xác định bản đồ giống ở mức độ phân tử và tính đa dạng sinh học.
102
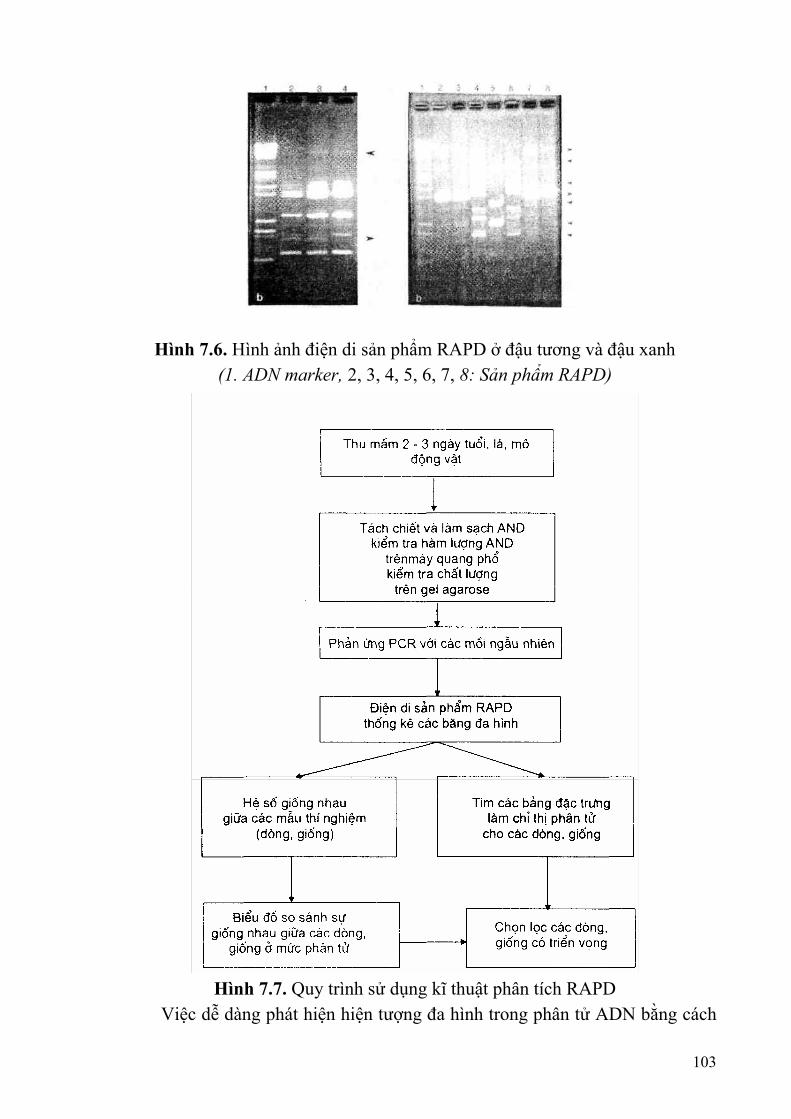
Hình 7.6. Hình ảnh điện di sản phẩm RAPD ở đậu tương và đậu xanh (1. ADN marker, 2, 3, 4, 5, 6, 7, 8: Sản phẩm RAPD)
Hình 7.7. Quy trình sử dụng kĩ thuật phân tích RAPD
Việc dễ dàng phát hiện hiện tượng đa hình trong phân tử ADN bằng cách
103

sử dụng tác nhân tố RAPD làm cho kĩ thuật này trở hấp dẫn trong nghiên cứu nhận dạng các cá thể, chẩn đoán các bệnh di truyền, phân loại thực vật và tiến hoá ở mức độ phân tử, các mối quan hệ phát sinh chủng loại trong giới tính. Trong phạm vi giới thực vật kĩ thuật khuếch đại ngẫu nhiên các đoạn ADN đa hình đã được sử dụng để nhận dạng các giống cây trồng (cây hoa lơ - Hu và Quyros, 1991; cây cần tây - Yang và Quyros, 1993), phát hiện mối quan hệ phát sinh chủng loại (bèo hoa dâu - Van Coppennolle và cộng sự, 1993; Scroch và cộng sự, 1992; ngũ cốc - Dweikat và cộng sự, 1993), đánh giá giống và sự đa dạng di truyền trong tập đoàn giống cây trồng (lúa mì - Vierling và Nguyen, 1992; Caetano - Anolles và cộng sự, 1991). Zheng và cộng sự (1991) khi sử dụng hai đoạn mồi khác nhau đã phát hiện các chỉ thị phân tử RAPD trong tập đoàn 19 giống lúa và chỉ ra tính hữu hiệu lớn lao của các chỉ thị phân tử đó trong việc nhận dạng giống.
Hình 7.8. Hình ảnh điện di sản phẩm RAPD ở lạc (a) và lúa cạn (b)
Fukuoka và cộng sự (1992) đã xác định các đoạn ADN đa hình được khuếch đại ngẫu nhiên ở 16 giống lúa khác nhau và cho rằng chúng khác biệt nhau bởi ít nhất là một băng ADN đa hình và phân bố trong ba nhóm lúa khác nhau tương ứng với các loại phụ Japonica, Javanica và Indica. L. X. Yu và H. T. Nguyen (1993) đã nghiên cứu sự biến dị di truyền của các giống lúa cạn thông qua sử dụng tới 42 đoạn dò ADN khác nhau. Các tác giả đã thu được 260 đoạn gene khuếch đại khác nhau, trong số đó thì 208 đoạn gene (tức là 80%) thể hiện sự đa hình và xác định mối quan hệ di truyền giữa các giống lúa đó. David. J. Mackill (1995) cũng bằng kĩ thuật này đã thành công xác định sự đa dạng di truyền của 134 giống lúa có nguồn gốc khác nhau và chỉ ra rằng sự biến dị di truyền của các nhân tố RAPD là rất quan trọng bổ sung thêm cho việc lập bản đồ gene và in dấu vân ADN. Dán nhãn gene (tagging Genes) Kĩ thuật khuếch đại ngẫu nhiên các đoạn ADN đa hình cũng đã được sử
104

dụng một cách thành công để đánh dấu (dán nhãn) các gene điều khiển các tính trạng quan trọng của cây trồng, chẳng hạn như gene quy định tính chống chịu bệnh đốm vi khuẩn ở cây cà chua (Martin và cộng sự, 1991), gene quy định khả năng chống bệnh giun tròn ở cây cà chua (Klein - Lankhorst và cộng sự, 1991), gene chống bệnh gỉ sắt. Kĩ thuật RAPD còn được ứng dụng trong chọn giống. Việc xác định các chỉ thị RAPD đặc trưng. làm cơ sở cho chọn giống gián tiếp. Căn cứ vào sản phẩm điện di RAPD người ta có thể thiết lập bảng thống kê sự có mặt các phân đoạn ADN được nhân (số 1 : có mặt đoạn ADN; số 0: đoạn ADN không được nhân). Trên cơ sở đó xác định dược hệ số đồng dạng di truyền giữa các đối tượng nghiên cứu, lập sơ đồ phả hệ (cây phát sinh). Từ kết quả đó sẽ xác định được sự sai khác về bộ gene của các đối tượng nghiên cứu. Bảng 7.1. Các phân đoạn ADN được nhân bản ngẫu nhiên trong phản ứng RAPD
Các dòng đột biến và giống gốc Các đoạn mồi
Ước lượng kích thước
(kb)
Đoạn được nhân
Giống gốc ML81 ML10 ML61 ML70 ML59 ML48
8,0 1 1 1 0 0 0 1 1 7,0 2 1 1 0 0 0 1 1 6,0 3 1 1 0 0 0 1 1
RA 31 5,1 4 1 1 0 0 0 1 1 5,0 5 0 0 1 1 1 0 0 4,9 6 1 1 1 1 1 1 1 4,8 7 0 0 1 1 1 0 0 2,5 8 0 0 1 0 1 0 1 2,0 9 0 0 1 0 1 0 0 1,9 10 0 0 1 0 1 0 0 1,5 11 1 1 1 1 1 1 1 1,49 12 0 0 0 1 0 0 0 1,48 13 0 0 0 1 0 1 1 1,43 14 1 1 1 0 1 0 0 1,40 15 0 0 1 0 1 0 0 1,38 16 0 0 1 0 1 0 0 1,35 17 0 0 1 0 1 0 0 1,33 18 0 1 1 1 0 1 1 1,2 17 0 1 0 1 0 0 1 0,98 20 0 0 1 0 1 1 0 0,83 21 0 0 1 0 1 0 0 0,60 22 0 1 0 0 0 1 1
105

Bảng 7.2. Hệ số đồng dạng di truyền của các dòng đậu tương
Dòng ĐB và G.gốc
Giống gốc Lạng Sơn
ML81 ML10 ML61 ML70 ML59 ML48
G.Gốc 1.0000000
ML81 0.7954545 1.000000
ML10 0.5000000 0.5681818 1.000000
ML61 0.5681818 0.7272727 0.704545 1.0000000
ML70 0.5909091 0.4772727 0.7727273
0.5227273
1.0000000
ML59 0.7954545 0.7727273 0.431818 0.690909 0.522727 1.000000
ML48 0.7500000 0.8636364 0.477272 0.6918182
0.5227273
0.8181818
1.0000000
Hình 7.9. Sơ đồ phả hệ của các dòng đậu tương đột biến chọn lọc từ giống gốc Các vệ tinh cũng như chỉ thị phân tử khác như đa hình độ dài các phân đoạn ADN (RFLP), các đoạn ADN đa hình khuếch đại ngẫu nhiên (RAPD) đã được sử dụng rộng rãi để thiết lập bản đồ phân tử chi tiết và định vị các gene điều khiển các tính trạng giá trị ở cây đậu tương (Keim và cộng sự, 1990; Diers và cộng sự, 1992; Yu và cộng sự, 1994). Shoemaker and Specht (1995) đã thông báo sáp nhập các nhóm liên kết cổ điển với các nhóm liên kết di truyền phân tử ở cây đậu tương bằng cách sử dụng 20 1ocus cổ điển, 8 1ocus chỉ thị phân tử RAPD và 110 1ocus chỉ thị phân tử RFLP.
§4. PHÂN TÍCH TÍNH ĐA HÌNH CHIỀU DÀI CÁC PHÂN ĐOẠN ADN ĐƯỢC NHÂN BẢN CÓ CHỌN LỌC (Amplified Fragment Length Polimorphism - AFLP) Zabeau và Vos (1993), Vos và cộng sự (1995) đã đưa ra và mô tả một kĩ thuật mới về in dấu vân ADN. Đó là kĩ thuật khuếch đại chọn lọc các đoạn ADN đa hình (AFLP - Technology). Kĩ thuật này dựa trên cơ sở siêu sao chép chọn 106

lọc các đoạn ADN đa hình từ hệ gene. Kĩ thuật khuếch đại có chọn lọc các đoạn ADN hiện nay là một kĩ thuật có sức mạnh lớn và đầy tiềm năng về in dấu vân ADN. Nó tạo ra một cách nhanh chóng nhất, hiệu quả nhất một bản đồ di truyền dày đặc các chỉ thị phân tử nhằm chọn lọc các tính trạng mong muốn thông qua chọn giống nhờ các chỉ thị phân tử đồng thời là một công cụ lí tưởng đối với việc xác định và đánh giá cây trồng ở mức độ phân tử ADN. Các nhân tố AFLP là những chỉ thị phân tử, nhanh chóng được áp dụng vào việc nghiên cứu di truyền, lập bản đồ gene, dán nhãn gene quan trọng ở nhiều loài thực vật khác nhau như khoai tây (Ballvoro và cộng sự, 1995; Folkertsma và cộng sự, 1996), lúa mạch (Becker và cộng sự, 1995). Kĩ thuật phân tích ADN đa hình có chọn lọc (hay là kĩ thuật khuếch đại chọn lọc các đoạn ADN đa hình - AFLP) gồm 3 bước cơ bản sau đây: (1) Phân cắt phân tử ADN bằng enzyme giới hạn tiếp theo là gắn kết chất gá (adapter). (2) Phản ứng chuỗi sử dụng enzyme polimerase để khuếch đại các đoạn ADN. (3) Phân tích các đoạn ADN trên gel poliacrylamid. Do có sự phối hợp các loại enzyme, giới hạn khác nhau với nucleotit chọn lọc của chất mồi nên kĩ thuật sinh học phân tử hiện đại này cho phép tạo ra nhiều các chỉ thị phân tử hơn so với các phương pháp trước đó. Bằng các nhân tố RFLP và AFLP K. Meksem và cộng sự (1995) trong chiến lược phân lập các diễn chống chịu bệnh sương mai (Phytophthora Infestans) ở cây khoai tây đã thành công thiết lập bản đồ gene có độ phân giải cao về vùng phụ cận locus R1 trên nhiễm sắc thể số 5. Các tác giả đã góp phần chọn lọc một cách gián tiếp các giống khoai tây có khả năng chống bệnh sương mai thông qua chọn giống nhờ các chỉ thị phân tử (Marker Assisted Selection - MAS). D. J. Mackill (1996) đã sử dụng các chỉ thị phân tử AFLP, RAPD và trình tự nucleotit lặp lại đơn giản (Microsatellite) để nghiên cứu mức độ đa hình trên 14 giống lúa khác nhau và cho rằng các chỉ thị phân tử AFLP và trình tự nucleotit lặp lại đơn giản có thể được xem như là các chỉ thị phân tử bổ sung cho việc lập bản đồ gene ở nhiều loại cây trồng nói chung cũng như ở cây lúa nói riêng, đồng thời các chỉ thị phân tử này cũng tỏ ra thích hợp cho việc nghiên cứu bản đồ các locus điều khiển tính trạng số lượng thông qua con lai bắt nguồn từ 2 loài phụ Japonica và Indica. Ngoài ra các chỉ thị phân tử AFLP trở nên cần thiết đối với việc lập bản đồ gene có chất lượng cao nơi mà cần phải làm bão hoà một
107

đoạn nhiễm sắc thể đặc hiệu với nhiều số lượng các chỉ thị phân tử (Thomas và cộng sự, 1995). Sự khám phá ra các đại phân tử axit nucleic và protein - cơ sở vật chất chủ yếu của sự sống và nghiên cứu ứng dụng vào thực tiễn bằng các kĩ thuật hiện đại trên cơ sở sự phát triển của sinh học phân tử. Trong công nghệ sinh học thực vật. việc phân loại genome trong tế bào thực vật và mối quan hệ giữa chúng làm sáng tỏ vai trò vật liệu di truyền thực vật, những hiểu biết về axit nucleic và hệ gene trong tế bào thực vật là nền tảng cho các kĩ thuật sinh học phân tử. Các kĩ thuật sinh học phân tử được ứng dụng trong phân tích genome thực vật. Trong các kĩ thuật sinh học phân tử ứng dụng vào chọn tạo giống cây trồng thì các kĩ thuật PCR và RAPD có thể được ứng dụng rộng rãi và, hiệu quả trong điều kiện Việt Nam. Bằng kĩ thuật RAPD, dựa vào sản phẩm điện di lập bảng thống kê các phân đoạn ADN được nhân bản, từ đó thiết lập biểu đồ so sánh sự khác nhau ở mức phân tử giữa các dòng và các giống với nhau. Những kết quả bước đầu về ứng dụng kĩ thuật PCR và RAPD cho phép khẳng định hiệu quả của sự kết hợp phương pháp chọn giống truyền thống với những phương pháp sinh học phân tử hiện dại trong tạo giống cây trồng, vật nuôi.
THẢO LUẬN 1. Trình bày cơ sở, nguyên tắc của phản ứng chuỗi PCR và những ứng dụng
của PCR. 2. PCR trong phân tích hệ gene của sinh vật và trong công tác chọn giống. 3. RAPD là gì? Phương pháp xác định dấu chuẩn RAPD và quan hệ di
truyền giữa các đối tượng nghiên cứu bằng phương pháp RAPD. 4. Thế nào là AFLP? Các bước của kĩ thuật AFLP. Ứng dụng của AFLP.
108

Chương 8
TẾ BÀO CHỦ VÀ VECTOR Tóm tắt: Trong chương này, chúng ta sẽ bàn đến khía cạnh sinh học của việc tách dòng gene bao gồm việc sử dụng các phân tử vector và hệ thống sống thích hợp, tức vật chủ mà ở đó vector có thể được nhân lên. Các loại tế bào chủ sẽ được mô tả đầu tiên, tiếp theo lả các hệ thống vector và các phương pháp dùng để đưa ADN vào các tế bào. Tế bào chủ và vector là hai đối tượng cực kì quan trọng trong kĩ thuật tách dòng. Có hai loại tế bào chủ, đó là: Tế bào chủ nhân sơ, Tế bào chủ nhân chuẩn và E. coli là vật chủ được sử dụng nhiều nhất trong kĩ thuật tách dòng. Vector tách dòng được phân biệt với vector chuyển gene ở một số đặc điểm cơ bản. Có nhiều loại vector được sử dụng trong kĩ thuật di truyền, nhưng được sử dụng rộng rãi nhất là plasmid. Nội dung của chương gồm 3 vấn đề chính là: (1) Tế bào chủ; (2). Vector, (3). Phương pháp chuyển vector tái tổ hợp vào tế bảo chủ.
§1. TẾ BÀO CHỦ Loại tế bào chủ sử dụng trong một ứng dụng cụ thể chủ yếu phụ thuộc vào mục đích của thí nghiệm tách dòng. Nếu mục đích đó là tách ra một gene để phân tích cấu trúc thì yêu cầu ở dây chỉ là một hệ thống đơn giản, dễ sử dụng. Nếu mục đích là biểu hiện thông tin di truyền ở một sinh vật nhân chuẩn bậc cao, chẳng hạn ở thực vật, thì đòi hỏi phải có một hệ thống đặc thù hơn. Thường thì một vật chủ ban đầu đơn giản được sử dụng để tách một đoạn trình tự mà đoạn này sau đó được đưa vào một hệ thống phức tạp hơn để biểu hiện. Các loại tế bào chủ yếu được nêu trên bảng 8.1. 1.1. Các vật chủ nhân sơ Một tế bào chủ lí tưởng là một tế bào dễ nuôi cấy và dễ nhân giống, dễ thu nhận được nhiều loại vector. Vi khuẩn E. coli đáp ứng các yêu cầu này và được sử dụng trong rất nhiều kĩ thuật tách dòng. E. coli được nghiên cứu rất tỉ mỉ, rất nhiều loài khác nhau đã được các nhà di truyền học vi sinh vật phân lập khi nghiên cứu các cơ chế di truyền của sinh vật nhân sơ này. Các nghiên cứu đó đã cung cấp những thông tin khoa học giá trị làm cơ sở cho kĩ thuật di truyền.
109
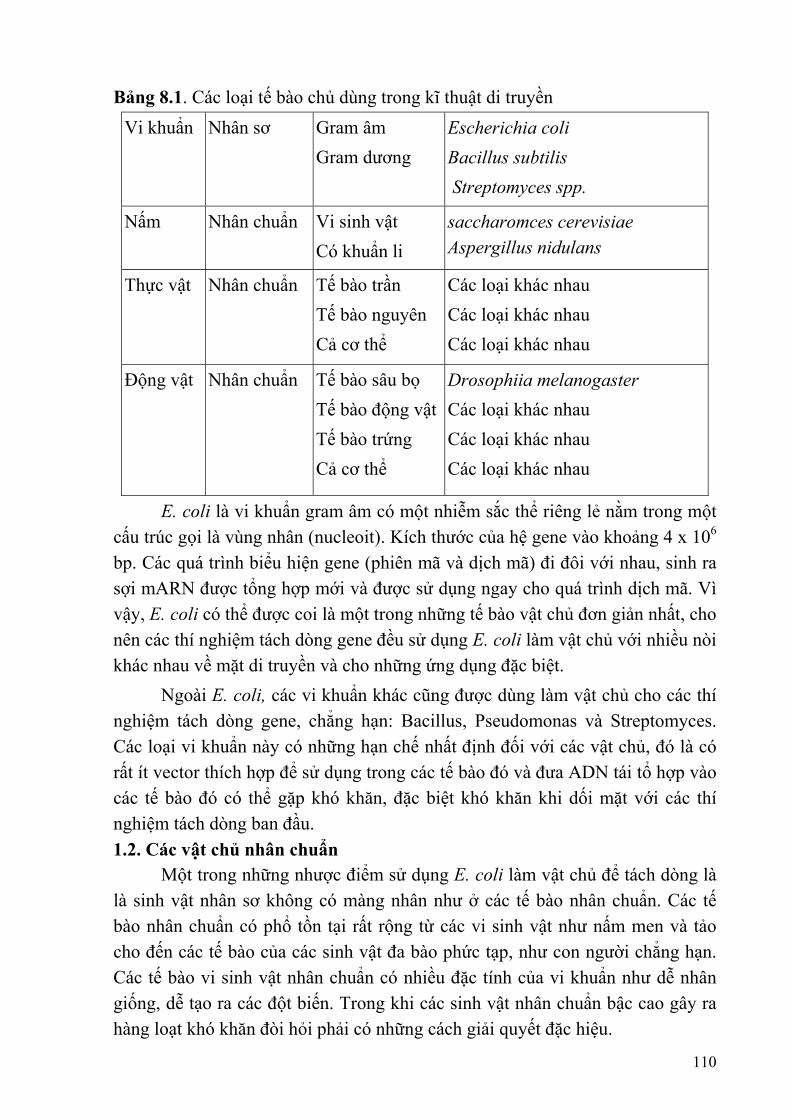
Bảng 8.1. Các loại tế bào chủ dùng trong kĩ thuật di truyền Vi khuẩn Nhân sơ Gram âm
Gram dương Escherichia coli Bacillus subtilis Streptomyces spp.
Nấm Nhân chuẩn Vi sinh vật Có khuẩn li
saccharomces cerevisiae Aspergillus nidulans
Thực vật Nhân chuẩn Tế bào trần Tế bào nguyên Cả cơ thể
Các loại khác nhau Các loại khác nhau Các loại khác nhau
Động vật Nhân chuẩn Tế bào sâu bọ Tế bào động vật Tế bào trứng Cả cơ thể
Drosophiia melanogaster Các loại khác nhau Các loại khác nhau Các loại khác nhau
E. coli là vi khuẩn gram âm có một nhiễm sắc thể riêng lẻ nằm trong một cấu trúc gọi là vùng nhân (nucleoit). Kích thước của hệ gene vào khoảng 4 x 106
bp. Các quá trình biểu hiện gene (phiên mã và dịch mã) đi đôi với nhau, sinh ra sợi mARN được tổng hợp mới và được sử dụng ngay cho quá trình dịch mã. Vì vậy, E. coli có thể được coi là một trong những tế bào vật chủ đơn giản nhất, cho nên các thí nghiệm tách dòng gene đều sử dụng E. coli làm vật chủ với nhiều nòi khác nhau về mặt di truyền và cho những ứng dụng đặc biệt. Ngoài E. coli, các vi khuẩn khác cũng được dùng làm vật chủ cho các thí nghiệm tách dòng gene, chẳng hạn: Bacillus, Pseudomonas và Streptomyces. Các loại vi khuẩn này có những hạn chế nhất định đối với các vật chủ, đó là có rất ít vector thích hợp để sử dụng trong các tế bào đó và đưa ADN tái tổ hợp vào các tế bào đó có thể gặp khó khăn, đặc biệt khó khăn khi dối mặt với các thí nghiệm tách dòng ban đầu. 1.2. Các vật chủ nhân chuẩn Một trong những nhược điểm sử dụng E. coli làm vật chủ để tách dòng là là sinh vật nhân sơ không có màng nhân như ở các tế bào nhân chuẩn. Các tế bào nhân chuẩn có phổ tồn tại rất rộng từ các vi sinh vật như nấm men và tảo cho đến các tế bào của các sinh vật đa bào phức tạp, như con người chẳng hạn. Các tế bào vi sinh vật nhân chuẩn có nhiều đặc tính của vi khuẩn như dễ nhân giống, dễ tạo ra các đột biến. Trong khi các sinh vật nhân chuẩn bậc cao gây ra hàng loạt khó khăn đòi hỏi phải có những cách giải quyết đặc hiệu.
110

Nấm men Saccharomyces cerevisiae là một vi sinh vật nhân chuẩn được ưa thích để nghiên cứu kĩ thuật di truyền. Nó đã được sử dụng nhiều thế kỉ trong sản xuất bánh mì và bia, và được nghiên cứu rất tích cực. Sinh vật này có thể sử dụng dễ dàng để phân tích di truyền học kinh điển, chúng có nhiều loại tế bào đột biến. Hệ gene của S. cerevisaae có khoản 1,35 x 107; bazơ nitơ, nhiều hơn E.coli khoảng 3,5 lần. Các nấm khác cũng được dùng trong các thí nghiệm tách dòng gene là Aspergillus nidulans và Neurospora crassa. Các tế bào thực vật và động vật cũng có thể được sử dụng như các vật chủ trong các thí nghiệm thao tác gene. Các tế bào thực vật và động vật khác thường được nuôi cấy trong các môi trường thích hợp và ở điều kiện này, để sử dụng trong các thí nghiệm thao tác gene so với các tế bào trong cơ thể.
§2. VECTOR 2.1. Khái niệm Để thu nhận gene dưới dạng tinh sạch với hàm lượng lớn người ta phải tách dòng (clone) gene đó. Việc tách dòng cơ bản là tiến hành gắn trình tự ADN vào vector. Vector cũng là một ADN dạng vòng, có khả năng xâm nhập vào tế bào vi khuẩn và mượn tế bào vi khuẩn để tạo ra những bản sao khác giống hệt vector ban đầu.
Hình 8.1. Sơ đồ plasmid
Vector có các ứng dụng chủ yếu sau: - Tạo dòng nhằm khuếch đại số lượng bản sao một trình tự ADN xác định. - Tạo sinh vật chuyển gene (chuyển các gene vào tế bào hay cơ thể). - Sản xuất ARN với số lượng lớn từ ADN được tạo dòng. - Sản xuất protein với số lượng lớn từ ADN được tạo dòng. Đặc tính cần có của một vector:
111

- Vector có khả năng tự sao chép tích cực trong tế bào vật chủ. Độc lập với sự sao chép của bộ gene vật chủ. - Vector có kích thước càng nhỏ càng tốt. - Vector phải được phát hiện rõ ràng trong vật chủ. - Phải tồn tại trong tế bào chủ qua nhiều thế hệ. - Vector chỉ tồn tại một vị trí nhận biết cho mỗi enzyme. Vector được sử dụng trong kĩ thuật tách dòng phân tử ADN đối với tế bào E. coli: (1) Plasmids; (2) Bacteriophage l, (3) Cosmids; (4) Bactenophage M13; (5) Phagemids và một số loại vector đặc biệt khác như: (i) Ti plasmid; (ii) YACS; (iii) BACS; (iv) Retrovirus; (iiv) Baculovirus. (iiiv) Yeast 2 micron plasmid. Việc lựa chọn sử dụng một loại vector dựa vào kích thước của đoạn ADN cần tạo dòng và mục đích tạo dòng. Vector được sử dụng với mục đích tách dòng gene (vector tách dòng) và chuyển gene (vector chuyển gene).
Hình 8.2. Plasmid pBR322 (a) và plasmid pUC18 (b)
(theo Nông Văn Hảí, 2002) 2.2. Các vector plasmid dùng cho E. coli Có một số đặc điểm quan trọng mà các vectơ cần phải có, chúng phải những phân tử ADN nhỏ giúp cho việc phân tách và bảo quản chúng dễ dàng. Phải có điểm khởi đầu sao chép (origin of replication), để cho ADN nó được nhân lên và do vậy được duy trì trong quần thể tế bào khi sinh vật chủ sinh trưởng và phân chia. Cần có một loại dấu chuẩn, chọn lọc (selectable marker) tạo điều kiện cho vector được dễ dàng phát hiện và cũng cần phải có ít nhất một điểm cắt bởi enzyme giới hạn để tạo điều kiện cho ADN xen vào trong quá trình tạo ra các thể tái tổ hợp. Các plamid có các đặc điểm phù hợp với yêu cầu trên và chúng được sử dụng thường xuyên như các vector trong các thí nghiệm tách dòng. 2.2.1. Plasmid 112

Nhiều loại plasmid được tìm thấy trong tự nhiên ở các vi khuẩn và trong một số nấm men. Chúng là những phân tử AND mạch vòng tương đối nhỏ so với nhiễm sắc thể của tế bào vật chủ. Các plasmid tồn tại chủ yếu ở trạng thái ngoài nhiễm sắc thể. Plasmid cos khả năng truyền các tính trạng cho vật chủ (chẳng hạn: tính bền vững với kháng sinh), do vậy giúp cho việc lựa chọn dễ dàng trong những điều kiện xác định. Các gene bền vững với kháng sinh do AND plasmid mã hóa thường được sử dụng trong việc thiết kế các vector dùng cho kĩ thuật di truyền vì chúng cung cấp một phương tiện thuận lợi để lựa chọn các tế bào có mang plasmid. Các plasmid có thể phân chia thành 2 nhóm: tiếp hợp và không tiếp hợp. Các plasmid tiếp hợp có thể tự truyền từ vi khuẩn này sang vi khuẩn khác thông qua quá trình tiếp hợp. Quá trình này đòi hỏi các chức năng của các đoạn đặc thù tra (transfer) và mob (mobilising) nằm trên plasmid. Các plasmid không tiếp hợp thì không tự truyền đi được. Một cách phân loại khác dựa trên số lượng các bản sao của plasmid có trong tế bào chủ. Các plasmid có số bản sao thấp có khuynh hướng kiểm soát chặt chẽ việc sao chép ADN, thường ADN plasmid loại này phải sao chép bắt buộc cùng với sự sao chép ADN nhiễm sắc thể của vật chủ. Các plasmid có số bản sao cao gọi lào các plasmid thoải mái, việc sao chép ADN của chúng không phụ thuộc vào sự sao chép ADN nhiễm sắc thể của tế bào chủ. Nói chung, các plasmid tiếp hợp thường lớn, có cơ chế kiểm soát chặt chẽ việc sao chép ADN và tồn tại trong tế bào với số lượng bản sao thấp. Trong khi đó, các plasmid không tiếp hợp thường nhỏ, sao chép ADN một cách thoải mái và tồn tại trong tế bào với số bản sao cao. 2.2.2. Các plasmid cơ bản dùng để tách dòng Trong kĩ thuật di truyền, các plasmid gặp trong tự nhiên thường được sửa đổi nhiều để tạo ra các vector có đặc điểm mong muốn. Để đặt tên plasmid, chữ p được dùng để chỉ plasmid và tiếp theo đó thường là tên của những người đã phân lập hoặc thiết kế plasmid. Các con số cũng có thể được sử dụng để phân loại các chủng cụ thể. Một trong những plasmid được sử dụng rộng rãi là pBR322, nó được hoàn thiện bởi Francisco Bolivar và đồng nghiệp. Việc thiết kế pBR322 bao gồm một loạt những thao tác để có được những đoạn ADN cần thiết cùng nối với nhau với kết quả cuối cùng là một ADN có 3 nguồn gốc. Plasmid này có tất cả các đặc tính của một vector tốt, chẳng hạn như: trọng lượng phân tử thấp, có các gene kháng kháng sinh, có điểm khởi đầu sao chép, và có một số điểm cắt riêng biệt bởi các enzyme giới hạn. Có một số plasmid trong nhóm pBR mang các đặc điểm khác nhau chút ít. Tuy nhiên, plasmid bắt nguồn từ pBR322 được sử dụng rộng rãi, vì chúng có
113

một số đặc điểm ưu việt so với plasmid ban đầu. Ví dụ: plasmid pAT153, là một dẫn xuất mất đoạn của pBR322. Plasmid này được tạo ra bằng các loại bỏ 2 đoạn ADN từ pBR322 sau khi dùng enzym giới hạn Hael. Lượng ADN bị loại bỏ rất nhỏ (705 cặp bazơ), nhưng hiệu quả đã làm tăng số bản sao lên 3 lần và đã loại bỏ các đoạn trình tự cần thiết cho quá trình huy động (mobilisation). Như vậy, pAT153 về một số phương diện là vector "tốt hơn" pBR322, vì nó có nhiều bản sao hơn trong tế bào và nó có mức độ bền vững sinh học cao hơn do mất khả năng huy động.
Hình 8.3. Vector pBluescript (theo Nông Văn Hải, 2002)
Sự có mặt của 2 gene kháng kháng sinh (Apr và Tcr) cho phép lựa chọn các tế bào có mang plasmid, vì các tế bào đó sẽ kháng với cả ampicilin và tetracylin. Một ưu việt nữa là các điểm cắt giới hạn duy nhất nằm trong các gene kháng sinh cho phép lựa chọn được các ADN tách dòng, và do vậy được gọi là bất hoạt do xen đoạn (insertional inactivation), khi mà đoạn ADN xen vào làm gián đoạn đoạn mã hoá của gene kháng và vì vậy làm thay đổi kiểu hình của tế bào mang ADN tái tổ hợp. 2.2.3. Các vectorplasmid khác Mặc dù các plasmid pBR322 và pAT153 được sử dụng rộng rãi trong nhiều ứng dụng của tách dòng gene, còn có các vector plasmid khác cũng được sử dụng. Nói chung, chúng dược thiết kế sao cho có được những đặc tính cụ thể, không thấy có trong các vector đơn giản. Chúng có thể mang những khởi điểm đặc hiệu để biểu hiện các gene xen vào hoặc chúng có thể có những ưu việt khác như tạo ra khả năng chọn lọc trực tiếp các thể tái tổ hợp. Một nhóm các vector plasmid cũng rất phổ biến là các plasmid thuộc họ PUC. Các plasmid này có 114

một đoạn ngắn chứa một số các điểm riêng lẻ bị cắt bởi enzyme giới hạn. Đoạn này gọi là đoạn đa liên kết (polilinker) hoặc điểm đa tách dòng (multiple cloning site), chúng rất hữu hiệu bởi vì có thể lựa chọn điểm cắt mong muốn để xen đoạn ADN trong quá trình tạo ra ADN tái tổ hợp. Bản đồ của một trong các vector pUC có các điểm cắt giới hạn nằm trong vùng polilinker, các plasmid pUC còn có vùng mang gene - blaceosidaza, gene này mã hoá β α - peptit. Đoạn trình tự này có chứa vùng polilinker, và việc một đoạn ADN vào một trong những điểm tách dòng sẽ dẫn đến một α - peptit không hoạt động chức năng. Việc đó đặt cơ sở cho một phương pháp sàng lọc trực tiếp các thể tái tổ hợp rất hữu hiệu sử dụng cơ chất tạo màu X-gal. Mặc dù các vector plasmid có nhiều tính chất hữu hiệu và chúng rất quan trọng cho việc thao tác gene, nhưng chúng vẫn mang hàng loạt nhược điểm.
Hình 8.4. Tạo vector tái tổ hợp (theo Nông Văn Hải, 2002)
2.3. Các vector phage dùng cho E. coli Mặc dù các vector dựa trên cơ sở phage về nhiều phương diện đặc hiệu hơn các vector plasmid, nhưng về thực chất chúng thực hiện cùng một chức năng, có nghĩa là chúng hoạt động như các phân tử mang các đoạn ADN. Có 2 loại phage(λ và M13) được hoàn thiện tích cực cho các mục đích tách dòng. 2.3.1. Phage Trong những năm 40, Max Delbruck và "nhóm phage" do ông thành lập
115

đã đặt nền móng cho sinh học phân tử hiện đại bằng cách nghiên cứu các phage Đó là những "thể ăn khuẩn", và là các virut chỉ nhân lên được bên trong vi khuẩn. Vì chúng có kích thước rất nhỏ nên người ta còn gọi chúng là hạt (particle). Về mặt cấu trúc, các phage chia thành 3 nhóm chính: (1) không đuôi;(2) có đầu và đuôi, và (3) có lông. Vật chất di truyền có thể là ADN mạch đơn, mạch kép, hoặc ARN. Thường thấy là các ADN mạch kép (DdsDN) (double-stranded ADN). Ở các phage không đuôi và có đuôi, hệ gene được bao bọc trong vỏ protein gọi là capsid (đôi khi được biết đến như vỏ hoặc đầu của phage. Trong các phage, dsADN điển hình hệ gene là những hệ gene bao gồm khoảng 50% toàn bộ khối lượng của hạt phage. Như vậy, phage là những hệ thống tương đối đơn giản nếu so sánh với vi khuẩn, vì nguyên nhân này chúng được sử dụng rất rộng rãi như các mô hình để nghiên cứu sự biểu hiện của gene.
Hình 8.5. Hình ảnh phage λ
Các phage có thể phân thành phage độc (virulent) và phage ôn hoà (temperate), tuỳ thuộc vào chu trình sống của chúng. Khi một phage chui vào trong một tế bào vi khuẩn, nó có thể sinh ra nhiều phage và giết chết tế bào đó, đó là chu trình sinh tan (litic) hoặc nó có thể nhập vào nhiễm sắc thể và ở lại đó mà không giết chết tế bào, đó là chu trình tiềm tan (lisogeneic). Phage độc là phage chỉ có chu trình sinh tan. Phage ôn hoà là phage có chu trình tiềm tan, nhưng cũng có thể có phản ứng sinh tan trong những điều kiện thích hợp. Hệ gene của phage λ có chiều dài 48,5kb, mã hoá cho 46 gene. Toàn bộ hệ gene đã được giải trình tự, và tất cả các điểm điều hoà đều đã được biết. Ở hai đầu của hệ gene mạch dài có các đoạn ngắn (12bp) sợi đơn, chúng bổ trợ cho nhau. Các đoạn này là những "đầu dính", chúng tạo điều kiện cho hệ gene trở thành mạch vòng sau khi nhiễm vào vi khuẩn. Đoạn của hệ gene được sinh ra bằng cách gắn các đầu dính gọi là điểm cos (cos site).
116

Hình 8.6. Hệ gene của Phage λ
Quá trình xâm nhiễm của phage vào vi khuẩn bắt đầu bằng việc phage bám vào dinh dưỡng điểm nhận trên bề mặt vi khuẩn. Khi phage bị hấp thụ thì ADN được tiêm vào bên trong tế bào và chu trình sống có thể bắt đầu. Hệ gene trở thành mạch vòng và phage bắt đầu chu trình sinh tan hoặc tiềm tan tuỳ thuộc vào một loại yếu tố như dinh dưỡng và trạng thái trao đổi chất của tế bào chủ và phụ thuộc vào tính đa nhiễm (số lượng phage xâm nhập vi khuẩn trong quá trình hấp thụ). Nếu chu trình tiềm tan bắt đầu thì hệ gene của phagenhập vào nhiễm sắc thể vật chủ và chúng được duy trì ở dạng prophage. Sau đó, nó sao chép cùng với ADN nhiễm sắc thể và được chuyển tới các tế bào thế hệ con trong một dạng ổn định. Nếu chu trình sinh tan bắt đầu thì một loạt các sự kiện phiên mã sẽ diễn ra, phage hoàn toàn làm chủ tế bào vật chủ và sinh ra nhiều bản sao của hệ gene phage cũng như các protein cấu trúc. Sau đó các thành phần này được lắp ráp lại hay còn gọi là bọc gói để tạo thành các phage trưởng thành, sau đó chúng thoát ra khỏi tế bào chủ khi tế bào này bị tan. Để xác định số lượng các phage có trong dịch tan, người ta pha loãng nhiều lần dịch phage và trộn lẫn với một số lượng dư thừa vi khuẩn chỉ thị rồi cấy lên mặt thạch. Sau khi ủ, vi khuẩn sẽ mọc và tạo thành cái gọi là thảm vi khuẩn. Phage mọc trên thảm này sẽ làm cho các tế bào bị phage xâm nhiễm tiêu tan, tạo nên các vết tan (plaque) trên thảm. Sau đó, người ta đếm số lượng các vết tan và xác định được số lượng phage trong dịch huyền phù ban đầu. Vết tan có thể được lấy ra cho mọc và phân tích tiếp. Phage có thể nhân lên trong dịch lỏng bằng cách cho nhiễm giống đang mọc của tế bào chủ và ủ cho đến khi các tế bào bị tan hoàn toàn, số lượng các hạt phage sinh ra phụ thuộc vào độ đa nhiễm và vào trạng thái của chu trình sinh sản vi khuẩn mà chúng nhiễm vào. Khi ADN chui vào tế bào thì nó chuyển sang phân tử mạch đôi gọi là dạng sao chép (RF) (replicative from), nó sao chép cho đến khi có khoảng 1000 bản sao trong tế bào. Tại thời điểm này, việc sao chép ADN trở nên bất đối xứng (asymmetric) và các bản sao mạch đơn của hệ gene được sinh ra và thoát khỏi tế bào như các hạt M13. Vi khuẩn không bị tan và vẫn còn sống trong suốt quá trình này, mặc dù quá trình sinh sản và phân chia có chậm hơn so với các tế bào không bị xâm nhiễm. 117

2.3.2. Các vector dựa trên cơs ở phage λ Việc sử dụng phage λ làm vector tách dòng phụ thuộc vào một thực tế là không phải tất cả hệ gene của λ đều cần thiết cho phage hoạt động chức năng. Do vậy, có một phạm vi để đưa ADN ngoại lai vào, mặc dù một số đòi hỏi nhất định cần phải có trong quá trình hoàn thiện các vector tách dòng dựa trên cơ sở phage λ . Một là sự sắp xếp các gene trên hệ gene λ sẽ quyết định phần nào cần phải loại bỏ đi hoặc cần phải được thay thế để đưa ADN ngoại lai vào. Vùng trung tâm của hệ gene λ nằm giữa các vị trí 20 và 35 trên bản đồ, là một đoạn lớn có thể bỏ qua mà không cần có sự sắp xếp lại invitro một cách phức tạp đối với hệ gene. Vùng trung tâm này chủ yếu kiểm soát các tính chất tiềm tan của phage, và phần lớn vùng đó có thể cắt bỏ mà không làm suy yếu các chức năng cần thiết cho chu trình sinh tan. Hai là, phage λ kiểu dại nói chung có nhiều điểm nhận biết của các enzyme giới hạn thường được sử dụng trong kĩ thuật tách dòng. Điều đó có thể là một khó khăn lớn, vì nó giới hạn sự lựa chọn các điểm để xen ADN vào. Trong thực tế sẽ tương đối dễ dàng lựa chọn đối với phage có số lượng giới hạn các điểm bị cắt bởi các enzyme giới hạn cụ thể, và kĩ thuật gây đột biến in vitro có thể được sử dụng để sửa đổi các điểm còn lại - tức các điểm không cần thiết. Như vậy, có thể nhận được phage mang tổ hợp mong muốn các điểm nhận biết bởi enzyme giới hạn.
Hình 8.7. Vector Phage λ
118

Một trong các nhược điểm lớn của các vector λ là capsid chỉ có thể bọc gói một số lượng ADN nhất định trong quá trình lắp ráp phage, do vậy, nó giới hạn kích thước của các đoạn ADN ngoại lai cần tách dòng. Trong quá trình bọc gói, các hạt phage có khả năng sống có thể sinh ra với chiều dài ADN nằm trong khoảng 38kb đến 52kb. 2.3.3. Các vector dựa trên cơ sở phage M13 Có hai điểm trong việc gây nhiễm M13 có giá trị đối với kĩ thuật di truyền. Một là, dạng sao chép RF rất giống với plasmid, vì vậy có thể tách chiết và thao tác bằng cùng những kĩ thuật như nhau. Ưu điểm thứ hai là ADN sợi đơn sinh ra trong quá trình nhiễm rất hữu ích trong các thí nghiệm như xác định trình tự theo phương pháp dùng dideoxynucleotit. Chỉ riêng về mặt này đã khiến cho M13 trở thành một vector có tiềm năng hết sức hấp dẫn. Khác với phage λ M13 không mang các gene không quan trọng. Hệ gene 6407 cặp bazơ nằm ở vùng giữa các gene. Vùng này đã được dùng để thiết kế một loạt các vector M13mp bằng cách xen một đoạn polinker/lac Z α - peptit vào đó. Việc này đã tạo điều kiện cho việc sử dụng hệ thống sàng lọc X-gal để phát hiện các thể tái tổ hợp, chẳng hạn như trường hợp với các plasmid pUC. Khi M13 mọc trên thảm vi khuẩn thì các vết tan xuất hiện do làm giảm sự sinh trưởng của các tế bào (chúng không bị tan), và có thể lấy ra phân tích tiếp.
Hình 8.8. Tách dòng gene bởi vector Phage hai sợi M13
(1). AND của Vector M13RF bị cắt bởi HindIII (2). Đoạn AND được cắt bởi Hind III (3). Ligase
119

(4). Biến nạp (5). AND Tái bản
Ưu việt thứ hai của các vector M13 là chúng không hoạt động chức năng một cách đầy đủ khi các đoạn ADN dài được xen vào vector. Mặc dù về mặt lí thuyết không có một giới hạn nào đối với kích thước của các đoạn có thể tách dòng, vì cấu trúc capsid được xác định bởi kích thước hiệu quả tách dòng đối với các đoạn lớn hơn 1,5kb. Trên thực tế, đó không phải là vấn đề chính vì các vector M13 được sử dụng chủ yếu khi tách dòng các đoạn ADN nhỏ để xác định trình tự. Trong ứng dụng này, việc sản sinh ra ADN sợi đơn cùng với sự dễ dàng trong việc tinh sạch ADN từ mô tế bào đã làm lu mờ những hạn chế về kích thước. 2.4. Các vector khác Cho đến nay, chúng ta mới tập trung vào các plasmid và phage để dùng trong các vật chủ E. coli, vi khuẩn này cho đến nay vẫn là đại diện chủ yếu được sử dụng trong phần lớn các công nghệ thao tác gene. Tuy nhiên, còn tồn tại các hệ thống vật chủ / vector khác cũng rất quan trọng, mặc dù chúng chuyên dụng hơn. Trong phần này, chúng tôi sẽ giới thiệu những đặc điểm của các vector vi khuẩn bổ sung và một số vector được sử dụng trong các sinh vật khác. 2.4 1. Các vector plasmid / phage Một đặc tính của các vector phagelà ở chỗ kĩ thuật bọc gói invitro độc lập về mặt trình tự, không đòi hỏi phải có các điểm cos nằm tách biệt bởi ADN với kích thước có thể bọc gói được (38-51kb). Điều đó đã được sử dụng để thiết kế các vector hình thành từ các đoạn trình tự plasmid kết nối với các điểm cos của phage λ . Các vector như thế gọi là các cosmid. Chúng có kích thước nhỏ (4-6kb) và vì vậy, có thể chứa các đoạn ADN tách đòng dài tới 47kb. Vì chúng không có các gene của phage nên chúng hoạt động như các plasmid khi đưa vào E. coli bằng cơ chế bọc gói/ nhiễm của λ . Bởi vậy, các vector cosmid cung cấp một hệ thống hết sức lí tưởng - một phương pháp đặc biệt và có hiệu quả cao để đưa ADN tái tổ hợp vào trong tế bào chủ, và khả năng tách dòng này lớn hơn khoảng 2 lần so với các vector λ tốt nhất. Tuy nhiên, chúng cũng có những nhược điểm và thường thì việc sử dụng cosmid thay cho các vector phage làm mất đi sự dễ dàng trong sử dụng và xử lí các trình tự tách dòng này.
120

Hình 8.9. Vector Cosmid
Các vector lai plasmid/ phage được hoàn thiện để khắc phục những hạn chế về kích thước của hệ thống tách dòng M13, và đến nay chúng được sử dụng rộng rãi cho các ứng dụng như giải trình tự ADN và sản xuất các mẫu dò để sử dụng trong các nghiên cứu về lai axit nucleic. Các vector này là những plasmid quan trọng, chúng chứa điểm khởi đầu sao chép fl của phage M13. Khi các tế bào có chứa plasmid này bị siêu nhiễm bởi phagethì chúng sản sinh ra các bản sao sợi đơn của ADN plasmid và tiết chúng ra môi trường ở dạng các hạt giống M13. Các vector, chẳng hạn như pEMBL9 hoặc pBluescript có thể tiếp nhận các đoạn ADN dài tới 10kb. 2.4.2. Các vector dùng trong các tế bào nhân chuẩn Khi xem xét các tế bào chủ nhân chuẩn thì các yêu cầu về vector trở nên rất phức tạp. Trong khi các vi khuẩn tương đối đơn giản về mặt di truyền thì các tế bào nhân chuẩn lại có nhiều nhiễm sắc thể nằm trong nhân có vỏ bao bọc. Vì các sinh vật nhân chuẩn hết sức đa dạng, nên lẽ tự nhiên là các vector có xu hướng đặc hiệu cao và cần thiết kế riêng biệt cho các mục đích cụ thể. Nhiều vector đã được hoàn thiện để sử dụng trong các tế bào nấm men và tuỳ thuộc vào từng ứng dụng cụ thể mà lựa chọn vector. Các plasmid episom của nấm men (Yeps) được hoàn thiện trên cơ sở plasmid nấm men thường gặp trong tự nhiên, chúng có thể tự sao chép hoặc hoà nhập vào nhiễm sắc thể. Các plasmid hoà nhập của nấm men (Yips) được thiết kế để gắn vào nhiễm sắc thể bằng cách tương tự như các plasmid YEp, còn các plasmid tự sao chép của nấm men (Yrps) vẫn ở trạng thái độc lập, không hoà nhập vào nhiễm sắc thể. Các plasmid có chứa các đoạn trình tự trong vùng tâm động đúng như các nhiễm sắc thể mini. Cuối cùng, các nhiễm sắc thể nhân tạo của nấm men (YACs) là các vector nấm men phức tạp nhất. Chúng có những vùng của tâm động và điểm mút, và có thể được sử dụng để tách dòng các đoạn ADN có kích thước rất lớn, bởi vì thế tái tổ hợp được duy trì chủ yếu như một nhiễm sắc thể nấm men. Trên thực tế, độ bền vững của các
121
YAC dường như tăng lên khi kích thước của đoạn tách dòng tăng lên, điều này không giống với nhiều hệ thống vector khác. Bảng 8.2. Một số vector có thể dùng cho các tế bào động vật và thực vật
Loại tế bào
Loại vector
Loại hệ gene
Thí dụ
Plasmid ADN Plasmid Ti của Agrobacterium tumefaciens Thực vật Virut ADN
ARN Virut khảm cải hoa, Geminivirut Virut khảm thuốc lá
Plasmid ADN Các loại vector plasmid khác nhau. Nhiều loại là vector lại có chứa một phần hệ gene của SV40
Virut ADN Baculovirut Virut Papilloma Virut Simian 40 (SV40)
Virut ARN Retrovirut
Động vật
Gene nhảy ADN Các phần từ P trong Drosophilla melanogaster
Khi làm thí nghiệm với các sinh vật nhân chuẩn bậc cao đa bào, như thực vật và động vật, các vấn đề đưa ADN tái tổ hợp vào trong cơ thể có khác biệt chút ít so với trường hợp các sinh vật nhân chuẩn đơn bào như nấm men. Mục tiêu của kĩ thuật di truyền ở các sinh vật nhân chuẩn bậc cao bao gồm: (1) Đưa ADN tái tổ hợp vào các tế bào động vật và thực vật trong mô nuôi cấy nhằm nghiên cứu cơ bản về sự biểu hiện của gene hoặc nhằm sản xuất các protein hữu ích. (2). Biến đổi cấu trúc di truyền của sinh vật và tạo ra một vận chuyển gene. Các vector dùng cho các tế bào thực vật và động vật có thể được đưa vào các tế bào trực tiếp bằng các kĩ thuật hoặc chúng có thể có cơ chế xâm nhập sinh học nếu dựa trên các virut hoặc các tác nhân xâm nhiễm khác như Agrobacteria.
§3. PHƯƠNG PHÁP CHUYỂN VECTOR TÁI TỔ HỢP VÀO TẾ BÀO CHỦ Việc thao tác vector và xen ADN để tạo ra các vector tái tổ hợp được thực
122

hiện trong ống nghiệm, và sau đó là đưa ADN tái tổ hợp vào tế bào chủ để nhân lên. Hiệu quả của bước này thường là yếu tố quyết định sự thành công của kĩ thuật tách dòng. Các phương pháp hiện có phụ thuộc vào kiểu loại của hệ thống vật chủ/ vector và bao gồm các kĩ thuật đơn giản đến những kĩ thuật rất phức tạp và đặc thù. 3.1. Biến nạp và lây nhiễm Các kĩ thuật biến nạp và lây nhiễm là những phương pháp đơn giản nhất để đưa ADN tái tổ hợp vào tế bào chủ. Trong trường hợp các tế bào E. coli thì biến nạp có nghĩa là đưa ADN plasmid vào trong tế bào, còn lây nhiễm là đưa ADN phage vào trong các tế bào. Biến nạp còn được sử dụng với nghĩa rộng hơn, cụ thể là: đưa bất kì ADN nào vào bất kì tế bào nào. Để cho biến nạp ở E. coli có hiệu quả các tế bào cần trở nên khả biến (competent). Để đạt được điều này, người ta ngâm các tế bào trong dung dịch calcium chlorid lạnh đóng đá, khi đó, các tế bào trở nên khả biến theo một cách mà cho đến nay chưa biết rõ nguyên nhân. Việc biến nạp các tế bào khả biến được thực hiện bằng cách trộn ADN plasmid với các tế bào, rồi ủ trong đá 20 đến 30 phút, sau đó tạo một sốc nhiệt ngắn (2 phút ở 420C), làm như vậy ADN sẽ chui vào tế bào. Các tế bào biến nạp thường được ủ trong nước thịt dinh dưỡng ở 370C trong vòng 60 đến 90 phút để làm cho các plasmid ổn định và biểu hiện các tính trạng của chúng ra kiểu hình. Sau đó, các tế bào được đưa lên môi trường chọn lọc để nhân lên các tế bào có plasmid. Trong quá trình biến nạp, chỉ có một phần rất nhỏ tế bào khả biến được biến nạp. Mặc dù có nhược điểm rất lớn, biến nạp vẫn là một kĩ thuật quan trọng, nó có thể tạo ra tới 109 các tế bào biến nạp (tức các thể biến nạp) khi đưa 1gr ADN vào thí nghiệm. Thường trên thực tế với 1μ g ADN người ta tạo ra được 106 đến 107 tế bào biến nạp. Lây nhiễm là một quá trình tương tự với biến nạp, chỉ khác ở chỗ ADN plasmid được thay thế bằng ADN phage. Quá trình này cũng không diễn ra với 100% các tế bào, và vì vậy chúng thường được thay thể bằng các bọc gói invitro trong các thí nghiệm đòi hỏi phải đưa ADN của phage vào bên trong các tế bào E. coli. 3.2. Bọc gói ADN của phage invitro Trong quá trình sinh tan của phage λ , ADN của phage được sao chép để tạo thành cái gọi là chuỗi khảm (concatemere). Đó là một phân tử ADN rất dài bao gồm nhiều bản sao của hệ gene λ được nối liền với nhau. Khi các hạt phage được lắp ráp thì ADN được bọc gói thành các capsid, quá trình này bao gồm việc cắt ADN tại các điểm cos bằng enzyme endonucleaza do phage mã hoá. 123

Các hạt phage trưởng thành sau đó dược sinh ra, và thoát ra ngoài khi các tế bào tan, sau đó chúng lại có khả năng nhiễm vào các tế bào khác. Quá trình này thường diễn ra invitro dưới sự điều khiển của các gene phage. Tuy nhiên, cũng có thể thực hiện trong ống nghiệm. Để bọc gói invitro thì các thành phần của capsid λ , và endonucleaza phải có trong ống nghiệm. Trên thực tế, hai nòi vi khuẩn được sử dụng để tạo ra dịch tan gọi là dịch chiết bọc gói (packaging extract). Mỗi nòi mang đột biến về một chức năng trong quá trình phát sinh hình thái của phage). Như vậy, các dịch chiết bọc gói sẽ không hoạt động riêng rẻ. Khi trộn hai cái với nhau với ADN tái tổ hợp không hoạt động khảm (concatemer) trong những điều kiện thích hợp và với đầy đủ các thành phần thì các hạt phage sẽ được sinh ra. Các hạt này sau đó có thể dược sử dụng để gây nhiễm các tế bào E. coli và tạo ra các vết tan. 3.3. Các phương pháp truyền ADN khác
Các phương pháp dùng để đưa ADN vào các tế bào vi khuẩn không dễ áp dụng cho các loại tế bào khác. Hệ thống bọc gói đặc thù cho phage không dùng được cho các hệ thống khác, và việc hiến nạp bằng các phương pháp thông thường có thể không thực hiện được hoặc hiệu quả rất thấp. Tuy nhiên, có những phương pháp khác dùng để đưa AND vào các tế bào. Thông thường, các phương pháp này có những đòi hỏi về kĩ thuật và kém hiệu quả so với các phương pháp dùng cho vi khuẩn. Tuy nhiên, người ta đã đạt được kết quả trong một số trường hợp. Vấn đề khó khăn nhất trong việc đưa ADN vào các tế bào không phải là vi khuẩn thường xảy ra với các tế bào thực vật. Các tế bào động vật lại được biến nạp tương đối dễ dàng. Còn việc biến nạp các tế bào thực vật gặp khó khăn vì vỏ tế bào của chúng rất cứng, ngăn chặn không cho ADN chui vào. Có thể khắc phục việc này bằng cách tạo ra các tế bào trần (protoplasts) là những tế bào mà thành của chúng đã được loại bỏ bằng enzym. Sau đó, các tế bào trần có thể được biến nạp bằng các kĩ thuật như mở lỗ bằng điện, khi một cung được sử dụng để tạo ra các lỗ trên màng tế bào mà qua đó ADN có thể xâm nhập. Sau đó các tế bào trần có thể được tái sinh (tạo vỏ mới). Ngoài ứng dụng này, các tế bào trần còn có vai trò quan trọng trong việc tạo ra các tế bào lai thực vật khi dung hợp với nhau. Mọi loại phương pháp biến nạp khác có thể đưa ADN vào bên trong tế bào là các phương pháp vật lí. Một cách để làm việc đó là sử dụng một cái kim rất mảnh để tiêm ADN trực tiếp vào trong nhân. Kĩ thuật này gọi là vi tiêm và đã được sử dụng thành công cả đối với các tế bào động vật và thực vật. Tế bào được giữ trên đầu một ống thuỷ tinh đầu tầy, còn chiếc kim nhọn được dùng để đâm xuyên qua màng tế bào. Kĩ thuật này đòi hỏi phải có một máy vi
124

thao tác cơ học, một kính hiển vi và đặc biệt phải có kĩ năng thực hành tinh vi. Gần đây người ta đã hoàn thiện một phương pháp tỏ ra rất hữu hiệu để biến nạp tế bào thực vật, gọi là kĩ thuật bắn ADN vào tế bào. ADN được dùng để bao bọc các hạt vonfram cực nhỏ gọi là vi đạn. Khi bắn, chúng được tăng tốc trên viên đạn lớn là bộ phận được bắn đi do ngòi thuốc nổ cháy. Tại một đầu của "súng" có một lỗ nhỏ chặn viên đạn lớn lại nhưng vẫn cho vi đạn chui qua. Khi nhằm bắn vào tế bào, các vi đạn sẽ đưa ADN vào trong tế bào và trong một số trường hợp biến nạp sẽ thực hiện thành công.
THẢO LUẬN 1. Nêu các loại tế bào chủ và ưu, nhược điểm trong kĩ thuật tách dòng gene. 2. Plasmid là gì? các loại plasmid. Trình bày về các vector plasmid dùng cho
E. coli. 3. Trình bày về vector phage. 4. Các vector dùng trong các tế bào nhân chuẩn. 5. Trình bày đặc điểm cơ bản của một số phương pháp chuyển vector tái tổ
hợp vào tế bào chủ. 6. E. coli và sử dụng E. coli trong kĩ thuật di truyền. 7. Vector tách dòng và vector biểu hiện gene. 8. Vấn đề tách dòng gene. 9. Vấn đề chuyển gene và sinh vật chuyển gene.
125

Chương 9
PHÂN LẬP GENE TÁCH DÒNG PHÂN TỬ
VÀ BIỂU HIỆN GENE Tóm tắt: Công nghệ ADN tái tổ hợp bao gồm các kĩ thuật tách dòng gene và biểu hiện gene. Bắt đầu từ sự phân lập gene và sau đó là tạo vector tái tổ hợp, biến nạp vào tế bào chủ, tách dòng và xác định trình tự gene. Protein là sản phẩm biểu hiện gene. Quá trình biểu hiện gene ngoại lai bắt đầu từ việc đưa gene vào vector biểu hiện và biến nạp vào tế bào chủ. Vector biểu hiện gene được thiết kế bao gồm promotor mạnh và điều khiển được. E. coli là vi khuẩn được sử dụng rộng rãi trong công nghệ ADN tái tổ hợp và sự biểu hiện gene thành công nhất hiện nay vẫn là ở E. coli. Phân lập, tách dòng và biểu hiện gene được minh hoạ bằng ví dụ về sự phân lập, xác định trình tự gene mã hoá cho trichobakin-RIP và quá trình biểu hiện gene này. Nội dung của chương gồm 3 vấn đề chính là: (1).Phân lập gene; (2). Tách dòng gene; (3). Biểu hiện gene.
§1. PHÂN LẬP GENE Phân lập gene hay phân lập đoạn ADN theo mục đích có thể được tiến hành theo phương pháp sử dụng enzyme cắt hạn chế hoặc bằng phản ứng chuỗi PCR. Phân lập gene mang ý nghĩa trong kĩ thuật giải trình tự ADN hay tạo dòng ADN tái tổ hợp trong kĩ thuật chuyển gene. 1.1. Phân lập gene bằng kĩ thuật cắt hạn chế Enzyme giới hạn (restrition enzyme) có khả năng cắt ADN ở những điểm đặc hiệu. Với đặc tính đó người ta đã vận dụng để xác định những đoạn ADN mã hoá cho các tính trạng của sinh vật và phân lập đoạn ADN đó. Kĩ thuật phân lập gene nhờ enzyme giới hạn có thể gồm các bước sau: (1). Tách chiết và tinh sạch ADN. (2). Cắt ADN bởi enzyme giới hạn. (3). Điện di trên gel agarose. (4). Dựa vào ADN marker xác định được đoạn ADN cần phân lập. (5). Biến tính ADN thành 2 mạch đơn ngay trên gel, rồi chuyển lên màng
126

lai phân tử. Vị trí các đoạn ADN được giữ nguyên. (6). ADN cố định trên màng được lai với mẫu dò có đánh dấu phóng xạ Rửa để loại bỏ các mẫu dò không bắt cặp. (7). Dùng kĩ thuật phóng xạ tự ghi để định vị các phân tử lai ADN- mẫu dò. 1.2. Phân lập gene bằng kĩ thuật PCR Phản ứng chuỗi polimerase được tiến hành với các primers có trình tự biết trước. Cặp primer gồm primer xuôi (sens primer) và ngược (antisens primer) được thiết kế theo trình tự gene từ ngân hàng gene. Cặp mồi chuyên biệt này được bắt cặp bổ sung với hai đầu của một trình tự ADN. Sử dụng cặp mồi chuyên biệt cùng với ADN mẫu, enzyme, dNTP, Mg++, bufpher... trong thiết bị nhân ADN (máy PCR) thì đoạn gene từ phân tử ADN mẫu sẽ được khuếch đại. Phân lập gene bằng PCR có thể được tiến hành theo các bước sau: (1) Tách chiết và tinh sạch ADN. (2). Thiết kế primers. (3). Chọn điều kiện phản ứng PCR. (4). Nhân đoạn ADN trong máy PCR để phân lập đoạn gene theo mục đích.
§2. TÁCH DÒNG GENE 2.1. Mục đích - Tạo ra một lượng lớn bản sao một trình tự ADN xác định. - Chọn lọc một thư viện gene. 2.2. Các bước của kĩ thuật tách dòng - Chọn và xử lí vector. - Xử lí ADN cần tạo dòng.
- Tạo vector tái tổ hợp. - Chuyển vector tái tổ hợp vào tế bào chủ. - Phát hiện dòng cần tìm trong thư viện gene.
127

Hình 9.1. Quy trinh chung của kĩ thuật tách dòng gene 2.3. Thư viện bộ gene Thư viện bộ gene là tập hợp tất cả các trình tự ADN cấu thành bộ gene đã được gắn vào vector. Về nguyên tắc, thư viện bộ gene được thiết lập từ bất cứ tế bào nào của sinh vật nghiên cứu. Các bước thiết lập thư viện bộ gene: (1) Tách chiết ADN. (2). Cắt ADN thành những đoạn có kích thước xác định bằng RF. (3). Gắn đoạn các ADN vào vector để tạo vector tái tổ hợp. (4). Biến nạp vector tái tổ hợp vào tế bào chủ. (5). Tế bào chủ nuôi cấy trên môi trường đặc và hình thành nên các dòng.
128

Hình 9.2. Tách dòng ADN tái tổ hợp
Ứng dụng: - Giải mã thông tin di truyền chứa trong bộ gene, đặc biệt cấu trúc itron, exon của một gene xác định. - Tạo dòng các trình tự không mã hoá nằm cạnh các gene và đóng vai trò quyết định trong sự điều hoà biểu hiện gene. 2.4. Phân lập, tách dòng và xác định trình tự gene chaperonin ở đậu tương Một ví dụ về xác định trình tự gene chaperonin liên quan đến tính chịu nóng hạn ở cây đậu tương Glicine max (L.) Merrill trên thiết bị tự động theo
129

nguyên tắc của phương pháp Sanger. Glicine max (L.) Merrill là loại cây trồng có vị trí quan trọng hàng đầu trong các loại cây họ đậu. Hạt đậu tương có hàm lượng protein cao từ 28% - 45%, dễ tan và chứa hầu hết các loại axit amin, đặc biệt là các loại axit amin không thay thế. Ngoài protein, hạt đậu tương còn có lipit và nhiều chất hữu cơ khác. Rễ đậu tương có nốt sần chứa vi khuẩn cố định đạm Rhizobium. Các dòng đậu tương M48, M10 và M61 được tạo ra bằng phương pháp đột biến thực nghiệm (Chu Hoàng Mậu, 2001) đã được đánh giá và chọn lọc qua nhiều thế hệ. Ngoài các đặc điểm về năng suất và chất lượng, tác giả còn quan tâm nghiên cứu mối liên quan giữa khả năng chịu hạn với đặc điểm hoá sinh và sinh học phân tử của các dòng đậu tương đột biến này. Dòng đậu tương đột biến chịu hạn ML61 có nguồn gốc từ giống đậu tương Lạng Sơn và được tạo ra bằng đột biến thực nghiệm với công thức xử lí phối hợp (16 kr γ + 0,01%EI). Trần Thị Phương Liên (1999) đã nghiên cứu, phân lập và xác định trình tự gene chaperonin từ giông đậu tương chịu nóng M103, genome của các dòng đậu tương đột biến ML10, ML48 và ML61 có tồn tại gene chaperonin và cấu trúc của nó có điểm gì khác với gene chaperonin đã được nghiên cứu. Kết quả nhân và tách dòng gene chaperonin của các dòng đậu tương đột biến M48, M10 và M61 và xác định trình tự gene chaperonin của dòng đậu tương chịu hạn M61 được thực hiện nhằm mục đích trên. Phát hiện gene chaperonin theo phương pháp của Kary Mullis và cộng sự (1985) với việc sử dụng cặp mồi Chap_N/chap_C thiết kế dựa trên trình tự ADN bổ sung (cDNA) của giống đậu tương Nhật Bản.
Chap_N: 5_GCC ATA TGT CGG CAA TCG CGG CCC C Chap_C: 5_CGG GAT CCC TAC CTC ACA GTT ACA GTT ACA ATA TCA TC
Chu trình nhiệt phản ứng: 950C : 1 phút; 550C : 1 phút; 720C : 1 phút 20 giây: 32 chu kì; 720C : 8 phút; Kết thúc ở 40C. Để tiến hành tách dòng gene chaperonin sản phẩm PCR được cắt thành các đoạn nhỏ hơn bằng SacI. Sau đó đem sản phẩm cắt gắn vào vector pBluescript KS (-) đã được mở vòng. ADN tái tổ hợp được biến nạp vào chủng E. coIi-DH5α . Môi trường nuôi cấy, các phương pháp gắn, tạo tế bào khả biến, biến nạp, tách ADN plasmid, kiểm tra đoạn gene bằng cách xử lí với enzyme giới hạn dược tiến hành theo sách cẩm nang chọn dòng phân tử của Sambrook và cộng sự 1989.
130

2.4.1. Kết quả tách chiết và tinh sạch ADN tổng số Mầm 2-3 ngày tuổi của các dòng dậu tương ML10, ML48 và ML61 được chia nhỏ từ 0,5-1g và giữ ở -700C. Khi tách chiết, mầm được nghiền trong nitơ lỏng và ADN tổng số được chiết ra bằng dung dịch đệm có chứa 10 mM Tris-HCl pH 8.0,1 mM EDTA, 100 mM NaCl, SDS 2,1%. Sử dụng enzyme proteinaza, hỗn hợp phenol : chlorofoc : isoamylalcohol (25 : 24 : 1) để loại protein và các hợp chất không mong muốn, EDTA có tácdụng cố định các ion Mg++ - là yếu tố cần thiết cho sự hoạt động của các nucleaza, do đó ADN trong dung dịch không bị phân giải bởi các enzyme này. Thực hiện quy trình tách chiết ADN như đã mô tả [1], thu được dung dịch có chứa ADN. Dung dịch ADN thu được vẫn tồn tại cả ARN, loại bỏ ARN bằng cách xử lí với ARNaza, ủ ở 370C trong 2-3 giờ. Sau đó tiến hành kiểm tra sản phẩm bằng điện di trên gel agarose 0,8%. Bản gel sau khi điện di được nhuộm bằng EtBr (ethidium bromide) trong 10-15 phút. Do cấu trúc của ADN có các liên kết hydro giữa hai mạch đơn nên khi nhuộm, các phân tử EtBr sẽ bám vào các liên kết này. Phân tử EtBr có khả năng phát quang dưới ánh sáng của tia tử ngoại (UV) nên có thể phát hiện băng vạch khi chiếu bản gaeldưới đèn UV.
Hình 9.3. Kết quả điện di ADN tổng số trên gel agaroza 0,8%
2.4.2. Nhân gene chaperonin từ 3 dòng đậu tương bằng kĩ thuật PCR Chaperonin tế bào chất là một dạng protein tồn tại trong tất cả các tế bào đóng vai trò quan trọng trong việc tạo cấu trúc không gian đúng cho protein tạo khung tế bào là actin và tubulin. Chúng cùng đồng thời thực hiện nhiều chức năng khác như tham gia tái tạo cấu trúc không gian đúng cho protein bị biến tính khi gặp điều kiện bất lợi. Các tác giả đã sử dụng đoạn mồi được tổng hợp dựa vào trình tự cDNA của chaperonin tế bào chất giống đậu tương chịu lạnh Nhật Bản để tiến hành nhân gene chaperonin của các dòng đậu tương đột biến ML10, ML48 và ML61 bằngkĩ thuật pPC.
131

Hình 9.4. Điện di đồ sản phẩm PCR gene chaperonin của 3 dòng đậu tương
Sau 32 chu kì phản ứng, sản phẩm PCR được kiểm tra bằng điện di trên gel agarose 0,8% và kết quả cho thấy ở cả ba dòng đậu tương đều thu được sản phẩm PCR đặc hiệu. Kích thước phân tử (KTPT) của sản phẩm PCR vào khoảng 1,6 kb. Đồng thời sản phẩm PCR nhận được có KTPT giống với đoạn tương ứng ở đoạn cDNA của gene chaperonin trong giống đậu tương Nhật Bản. Điều đó chứng tỏ rằng các dòng đậu tương nghiên cứu đều có gene chaperonin. Như vậy, trong trình tự của nó chỉ có những vùng được dịch mã sang protein (exon), mà không có những vùng không dịch mã (intron). 2.4.3. Phân tích gene chaperonin bằng enzim giới hạn Theo các nghiên cứu đã công bố, trình tự gene chaperonin có điểm nhận biết cho một số enzyme giới hạn như: SacI, AluI, SauI. Trong đó SacI cắt trình tự cDNA thành 5 đoạn có kích thước phân tử tương ứng là 260, 189, 198, 630 và 325 bp. Như vậy, để so sánh gene chaperonin này với các trình tự công bố. Sản phẩm cắt được điện di trên gel agaroza 1,2%.
Hình 9.5. Phổ điện di ADN plasmid
Kết quả trên diện di đồ cho thấy không có sự khác biệt về vị trí điểm cắt SacI ở các dòng đậu tương đột biến so với các gene chaperonin đã được công bố trước đây. Điều này chứng tỏ sản phẩm PCR nhận được có thể chính là đoạn gene chaperonin. Đây là một trong những bằng chứng khẳng định sản phẩm
132

PCR thu được chính là gene chaperonin đã được công bố. Tuy nhiên để có thể khẳng định chắc chắn về gene này cần tiếp tục nghiên cứu tách dòng sản phẩm PCR của một đại diện là dòng đột biến ML61 - dòng được đánh giá là có năng suất cao, chín sớm và khả năng chống chịu hạn tốt để tiến hành đọc trình tự ADN của đoạn gene này làm cơ sở cho việc so sánh với trình tự của gene chaperonin đã được công bố trên ngân hàng gene thế giới. 2.4.4. Tách dòng một số đoạn ADN của gene chaperonin ở dòng ML61 Nhằm mục đích nghiên cứu đọc trình tự gene, cần phải gắn một số đoạn ADN của gene chaperonin ở dòng ML61 đã được cắt bằng enzyme SacI vào vector pBluescript KS (-). Dựa vào các đặc điểm của vector pBluescript KS (-) có chứa gene lacZ và vị trí cắt của SacI... Đồng thời trên gene chaperonin cũng chứa vị trí các điểm cắt đặc hiệu của SacI. Vì vậy, sử dụng SacI để xử lí sản phẩm PCR của dòng đậu tương ML61 và mở vòng vector pBluescirpt KS (-). Phản ứng gắn của vector với các đoạn cắt của sản phẩm PCR được tiến hành ở 160C trong vòng 24 giờ, sử dụng enzyme T4 ligaza. Sau đó, dịch phản ứng được biến nạp vào chủng E. coli - DH5α và cấy trải trên môi trường chọn lọc có ampixillin, X-gal và IPTG. Kết quả đã nhận được một số khuẩn lạc trắng và khuẩn lạc xanh trên môi trường chọn lọc. Những khuẩn lạc trắng có thể đã mang đoạn ADN ngoại lai, còn khuẩn lạc xanh mang plasmid không chứa đoạn gắn vào. Để kiểm tra kết quả của quá trình chọn dòng 9 khuẩn lạc màu trắng (kí hiệu các khuẩn lạc từ 1 đến 9) và một khuẩn lạc màu xanh (làm đối chứng) được sử dụng để tách chiết ADN plasmid. Sau khi tách chiết và làm sạch, sản phẩm ADN plasmit được kiểm tra trên gel agaroza 0,8%. Trên ảnh điện di đồ cho thấy cả 9 khuẩn lạc đều có ADN plasmid với độ di động điện di cao hơn đối chứng. Để xác định được những khuẩn lạc cần tìm có gắn đoạn sản phẩm PCR cắt bằng SacI, chọn ADN plasmid từ bơn khuẩn lạc 1, 4, 7 và 8 để tiến hành phản ứng cắt kiểm tra bằng enzyme SacI. Sản phẩm sau khi cắt được kiểm tra bằng điện di trên gel agaroza 0,8 %. Kết quả trên ảnh điện di dòng số 8 đã gắn được một đoạn ADN khoảng 0,6 kb; dòng số 7 đã gắn được đoạn ADN khoảng 0,4 kb; dòng số 4 đã gắn dược đoạn ADN khoảng 0,2 kb. Riêng sản phẩm cắt của dòng số 1 cho thấy có cả 2 đoạn ADN khoảng 0,6 kb và 0,2 kb được gắn vào vector. Trong quá trình tách dòng, sử dụng enzyme SacI cắt gene chaperonin thành 5 đoạn. Vì SacI là enzyme cắt tạo đầu dính nên sản phẩm cắt chỉ cho được 3 đoạn có đầu dính tương ứng với kích thước là 189, 198 và 630 bp. Như vậy, rõ ràng là dòng số 8 đã gắn được đoạn tương ứng có kích thước là 630 bp. Dòng số 4 thì gắn dược hoặc là đoạn 189 bp hoặc 198 bp (vì hai đoạn
133

này rất khó phân biệt trên diện di đồ). Còn dòng số 1 gắn được cả hai đoạn 630 bp và một trong hai đoạn ngắn nói trên. Đối với dòng số 7 đoạn gắn có kích thước khoảng 400 bp có thể là tổng của hai đoạn ngắn 189 và 198 bp và cũng có thể là đoạn lẫn tạp trong sản phẩm cắt ngẫu nhiên. Để có những kết luận chính xác về các dòng thu được, cần tiến hành đọc trình tự ADN. 2.4.5. Xác định trình tự gene Chaperonin ở cây đậu tương Vector tách dòng TA cloning và các hoá chất cần thiết của hãng Invitrogen, vi khuẩn E. coli chủng DH5α . Tạo dòng phân tử được tiến hành theo phương pháp của Sambrook và Russell. Xác định trình tự gene chaporonin được thực hiện trên máy đọc trình tự tự động ABI PRISM 3100 Avant Genetic Analizer. Nhân đoạn gene chaperonin từ ADN tổng số, tiểu phần CTTδ của dòng đậu tương đột biến ML61. Sau đó, sản phẩm PCR được đưa vào vector TA cloning, được biến nạp vào vi khuẩn E. coli DH5α . Kiểm tra vector tái tổ hợp bằng xử lí với enzyme EcoRI đã thu được dòng mang đoạn gene mong muốn để tiến hành đọc trình tự. Để kiểm tra kết quả đọc trình tự gene chaperonin, sử dụng phần mềm Sinh học Clustalx để so sánh trình tự gene này với các gene chaperonin của giống Cúc Vàng (còn gọi là Cúc Lục Ngạn), một giống địa phương có khả năng chịu hạn, giống M103 đột biến có khả năng chịu nóng và giống đậu tương chịu rét Nhật Bản - Bonminori.
134
135

Trình tự gene chaperonin của dòng đậu tương đột biến ML61 có sự sai khác 41 vị trí nucleotit so với giống M103, 57 vị trí nucleotit so với giống Cúc Vàng và 25 nucleotit so với giống đậu tương Bonminori. Sự sai khác này tạo nên sự thay đổi trình tự 9, 10 và 11 axit amin sai khác của ML61 tương ứng so với Bominori, M103 và Cúc Vàng.
Bảng 9.1: Trình tự axit amin của gene chaperonin ML61 sai khác so với Bonminori, Cúc Vàng và M103
STT Vị trí Bonminori Cúc Vàng M103 MI61 1 8 Gln (Q) Gln (Q) Gln (Q) His (H) 2 24 Glu (E) Asp (D) Asp (D) Glu (E) 3 29 Gli (G) Ala (A) Ala (A) Ala (A) 4 43 Thr(T) Thr(T) Thr(T) Ala (A) 5 54 Ile (1) Thr(T) Thr (I) Ile (I) 6 76 Val (V) Gli (G) Val (V) Val (V) 7 99 Ser (S) Thr(T) Thr (T) Thr (T) 8 110 His(H) Leu (L) His(H) Leu (L) 9 128 Glu Asp (D) Asp Asp 10 129 Pro (P) Ala (A) Pro (P) Ala (A) 11 191 Pro (P) Pro (P) Pro (P) Ala (A) 12 204 Val (V) Ile(I) Val (V) Val (V) 13 220 Leu (L) Pro (P) Leu (L) Leu (L) 14 280 Ser (S) Gli (G) Gli (G) Ser (S) 15 308 Ser (S) Ser (S) Ala (A) Ser (S) 16 360 Phe (F) Val (V Phe (F) Phe (F) 17 447 Asp (D) Ala (A) Ala (A) Ala (A) 18 506 Leu (L) Leu (L) Pro (P) Leu (L) 19 514 Thr(T) Met (M) Thr (T) Thr (T)
Các trình tự nucleotit sai khác còn lại đều do sự dột biến điểm có tính chất đồng chuyển (hoán vị của các nucleotit trong nhóm với nhau) mà không làm thay đổi trình tự axit amin. Mặt khác ở dòng đậu tương MI61 còn có sự sai khác ở 3 vị trí axit amin 8Gln-His, 29Thr-Ala và 191Pro-Ala so với cả 3 giống so sánh trên. ML61 là một dòng đậu tương đã được xử lí đột biến phối hợp với 16 kr γ + 0,01%EI, có thể chính việc xử lí phối hợp tia γ và EI đã gây nên sự đột biến trình tự nucleotit của dòng đậu tương này. 136

Xem xét đến 3 vùng chức năng quan trọng của chaperonin để nghiên cứu đánh giá trình tự nucleotit của dòng ML61 đến việc tạo cấu trúc không gian của chaperonin. Vùng hoạt tính ATPaza và vùng gắn với ATP, đây là hai vùng bảo thủ của gene. Vùng hoạt tính ATPaza vẫn giữ dược tính bảo thủ, còn vùng gắn với ATP ở ML61 sai khác với Bominori ở vị trí 99Thr-Ser và M103 99Thr - Ile. Cúc Vàng có trình tự vùng này giống ML61. Một vùng thứ 3 được kiểm tra là vùng gắn cơ chất với phức CCT, đây là vùng đa hình nhất so với các vùng chức năng khác của chaperonin tiểu phần CCTδ . Trong vùng này ML61 sai khác với M103 ở vị trí axit amin 280Gli-Ser, Cúc Vàng ở hai vị trí 220Pro-Leu và 280Gli-Ser. Không có sự thay đổi của vùng này so với Bominori. Ngoài ra để đánh giá khả năng chịu hạn của ML61, trình tự gene chaperonin này được xem xét theo quy luật biến đổi các axit amin trong các enzim về khả năng chịu lạnh sang nóng. Dựa trên trình tự axit amin, dòng ML61 cũng có sự biến đổi tương tự ở vị trí axit amin 29 Gli-Ala, 99Ser-Thr (Bominori) giống như Cúc Vàng và M103. Hai giống đậu tương đã được đánh giá là có khả năng chịu hạn. 2.5. Thư viện cADN Thư viện ADN là tập hợp các bản sao cADN từ tất cả các mARN của tế bào. Khác với thư viện bộ gene, thư viện cADN được thiết lập từ một tế bào xác định trong đó gene nghiên cứu biểu hiện thành mARN. Mục đích của thư viện cADN là nghiên cứu biểu hiện một gene xác định cùng với những vấn đề liên quan như biểu hiện gene, mối tương tác giữa các gene trong quá trình sống.
Các bước thiết lập thư viện cADN: (1) Chọn vector. (2). Tách chiết mARN. (3). Tổng hợp cADN nhờ enzyme sao mã ngược. (4) Tạo vector tái tổ hợp gồm plasmid và cADN. (5) Biến nạp vector tái tổ hợp vào tế bào vi khuẩn.
(6) Nuôi cấy vi khuẩn trong môi trường lỏng, sau chuyển sang đặc. Các dòng vi khuẩn sẽ hình thành các khuẩn lạc. (7). Phát hiện dòng cần tìm.
137

Hình 9.6. Tạo vector cADN tái tổ hợp (theo Nông Văn Hải, 2002)
2.6. Phát hiện dòng tế bào chủ chứa vector tái tổ hợp Vector và đoạn ADN ngoại lai được trộn lẫn với nhau dưới tác dụng của enzyme nối ADN-ligase. Sau đó hỗn hợp vector và đoạn ADN ngoại lai được trộn lẫn với tế bào chủ để thực hiện biến nạp và có 3 khả năng có thể xảy ra: (1) Tế bào chủ không có plasmid. (2) Tế bào chủ nhận được plasmid không có gene ngoại lai xen vào; (3) Tế bào chủ nhận dược vector tái tổ hợp. Chính vì vậy trong kĩ thuật tách dòng cần phải xác định dược đúng dòng tế bào chủ chứa vector tái tổ hợp. Có thể sử dụng 3 phương pháp cơ bản sau: lai axit nucleic, dựa vào sự biểu hiện kiểu hình, hiện tượng mất hoạt tính do xen đoạn. (a) Sử dụng phương pháp lai axit nucleic có hiệu quả trong việc tách dòng gene mong muốn từ thư viện ADN bằng việc sử dụng mẫu dò được đánh dấu phóng xạ. Có thể thực hiện theo các bước: làm tan các khuẩn lạc trên giấy nitrocellulo giải phóng ADN, lai mẫu dò lai với ADN trên màng lai. Việc phát hiện các phân tử lai thông qua phóng xạ tự ghi. (b) Phát hiện khuẩn lạc chứa plasmid tái tổ hợp thông qua biểu hiện kiểu hình. Các vector được thiết kế mang trình tự điều hoà của gene lacZ, khi biến nạp vào tế bào chủ, làm biến đổi tế bào chủ có kiểu hình Lac- thành Lac+. Các vi khuẩn có kiểu hình Lac+ được nhận biết nhờ sự chuyển hoá cơ chất X-gal làm khuẩn lạc có màu xanh. Nếu đoạn ADN ngoại lai được gắn xen vào trình tự điều hoà của lacZ, làm gene không hoạt động, do vậy không thực hiện được phản ứng biến đổi X-gal thành màu xanh, vì vậy khuẩn lạc có màu trắng. Nhờ vậy người ta có thể nhận biết nhanh chóng bằng mắt thường các khuẩn lạc chứa vector tái tổ hợp.
138

(c) Hiện tượng làm mất hoạt tính do xen đoạn được sử dụng đối với các vector mang gene kháng thuốc ampiciline (hoặc tetracycline), chứa các điểm nhận biết của các enzyme cắt hạn chế. Plasmid được mở vòng dưới tác dụng của một RE mà điểm cắt của nó nằm trên một gene kháng thuốc nào đó và đoạn ADN ngoại lai được xen vào vị trí mở vòng trên plasmid, làm cho gene kháng thuốc trên mất hoạt tính. Hỗn hợp trên được biến nạp vào tế bào chủ E. coli, tế bào chủ có thể có 3 loại kiểu gene và kiểu hình: (1) Tế bào chủ không nhận được vector sẽ không mọc được trên môi trường có tetracyline (hoặc ampiciline); (2) Tế bào chủ nhận được vector nhưng gene ngoại lai không gắn được vào vector, nó sẽ mọc được trên môi trường có ampiciline, tetracycline; (3) Tế bào E. coli nhận được vector tái tổ hợp, gene amp bị mất hoạt tính do gene lạ xen vào, gene kháng tetracyline vẫn hoạt động bình thường. Do vậy các tế bào chủ mang vector tái tổ hợp không mọc được trên môi trường chứa ampiciline, nhưng vẫn mọc được trên môi trường chứa tetracycline.
§3. BIỂU HIỆN GENE 3.1. Khái niệm 3.1.1. Các hệ biểu hiện gene Trong tế bào sống, biểu hiện gene được hiểu là quá trình sinh tổng hợp protein trong mối liên hệ: ADN → ARN → Protein và protein là sản phẩm biểu hiện gene. Trong kĩ thuật di truyền, biểu hiện gene là hiện tượng protein của gene ngoại lai được tổng hợp trong tế bào chủ. Hiện nay có 4 hệ sau hay sử dụng biểu hiện gene như Escherichia coli, Bacillus subtilis, nấm men (Saccharomyces cerevisiae, Pichia pastoris) và tế bào động vật. Tuy nhiên E. coli và nấm men là hai hệ biểu hiện chính hiện đang được sử dụng rộng rãi trong nghiên cứu và ứng dụng. Cho đến nay có thể nói rằng vấn đề biểu hiện gene ngoại lai vẫn còn mang tính thực nghiệm đối với từng gene, chưa có một mô hình chung cho phép biểu hiện tối ưu đối với các loại protein khác nhau trong một biểu hiện. Vì vậy, để đạt được mức độ biểu hiện cao đối với một gene ngoại lai. trong nhiều trường hợp phải nghiên cứu thăm dò với các yếu tố khác nhau để tìm ra diều kiện tối ưu. Các protein biểu hiện trong các tế bào khác nguồn gốc có thể được chia thành 4 nhóm chính: Nhóm thứ nhất bao gồm các peptit nhỏ có độ dài dưới 80 axit amin. Đây là đoạn peptit được biểu hiện dễ dàng nhất dưới dạng protein dung hợp (fusion
139

protein) và thường được biểu hiện trong tế bào E. coli. Nhóm thứ hai là các chuỗi polipeptit thường được tiết ra ngoài tế bào (như enzyme, cytokin, hormon...) có kích thước trong khoảng 80-500 axit amin. Các protein thuộc nhóm này có thể biểu hiện được trong cả 4 hệ biểu hiện. Nhóm thứ 3 gồm các protein tiết và protein đóng vai trò thụ thể trên bề mặt tế bào có kích thước hơn 500 axit amin. Nhóm thứ tư bao gồm tất cả các protein không tiết có kích thước lớn hơn 80 axit amin. Hầu hết các protein đều có thể được biểu hiện trong tế bào E. coli nếu protein đó không quá nhỏ hoặc quá lớn, hoặc quá kị nước, hoặc chứa quá nhiều gốc cystein. Các protein nhỏ và polipeptit thường không bền vững trong tế bào chủ nên muốn biểu hiện tốt trong E. coli thì cần được gắn với một đoàn protein khác. Đoạn protein này sẽ giúp cho đoàn peptit dược bền vững trong tế bào E. coli sau đi được tổng hợp và hiện nay nó có còn được dùng như một dấu nhận biết tinh chế protein lai này. Sở dĩ các protein có chứa nhiều gốc cystein và cầu disulfit không được biểu hiện tốt trong E. coli là do môi trường bên trong tế bào E. coli có tính khử cao. B. subtilis và nấm men mặc dù không phải là hệ được sử dụng rộng rãi trong nghiên cứu để biểu hiện gene ngoại lai như E. coli nhưng những hệ biểu hiện này cũng có nhiều ưu điểm đáng chú ý. Điển hình là B. subtilis có khả năng biểu hiện và tiết rất tốt protein có nguồn gốc từ Prokaryot ra ngoài môi trường nuôi cấy còn nấm men lại có những ưu điểm quan trọng của cả Eukaryot và Prokaryot. Nấm men sinh trưởng nhanh và có khả năng chế biến protein sau khi dịch mã rất tốt để trở thành dạng protein hoạt động mà hệ biểu hiện gene ở vi khuẩn không có. Hơn nữa nấm men có mức độ an toàn sinh học cao, được sử dụng nhiều trong công nghiệp dược liệu và thực phẩm. Vì vậy, nấm men rất được ưa chuộng để biểu hiện các gene của Eukaryot và gene mã hoá cho protein sử dụng trong thực phẩm và chữa bệnh. Hệ biểu hiện trong tế bào động vật có khả năng biểu hiện được tất cả protein của 4 nhóm nhưng tốn thời gian và quá trình chọn lọc, nhân dòng và nuôi cấy dòng tế bào phức tạp hơn nên hệ biểu hiện này thường không được sử dụng rộng rãi. Những đặc tính sinh học phân tử để điều chỉnh sự biểu hiện gene thể hiện ở (1) Đặc điểm tự nhiên của những trình tự promotor và terminator liên quan đến dịch mã; (2) Độ mạnh yếu và vị trí bám của ribosome; (3) Số phiên bản của gene tách dòng; (4) Vị trí tổng hợp protein trong tế bào, (5) Hiệu suất của quá
140

trình giải mã trong tế bào chủ; (6) Sự bền vững của phân tử protein được mã hoá bởi gene tách dòng trong tế bào chủ. Mức độ biểu hiện gene ngoại lai (foreign gene) còn phụ thuộc vào sự nhận biết của cơ thể vật chủ. Hiện nay đã có phổ rất rộng các cơ thể biểu hiện gene ngoại lai (cả Prokaryot và Eukaryot) nhưng đa số các sản phẩm được sản xuất bởi công nghệ ADN tái tổ hợp được tổng hợp ở E. coli. 3.1.2. Biểu hiện gene từ những promotor mạnh và điều khiển được Yêu cầu tối thiểu với một hệ thống biểu hiện gene có hiệu quả là sự có mặt của một trình tự promotor mạnh và có thể điều khiển được (strong and regulatable promotor) nằm ở phía trước gene tách dòng. Promotor mạnh là một promotor có ái lực cao với enzyme ARN polimerase và sẽ bảo đảm cho trình tự nằm sau promotor dược phiên mã với tần suất cao. Khả năng điều khiển promotor đã cho phép tế bào chủ kiểm soát quy mô phiên mã một cách chính xác. Promotor lac-operon đã được sử dụng rộng rãi trong việc biểu hiện gene tách dòng. Ngoài ra nhiều promotor đã được phân lập từ nhiều cá thể khác nhau để sử dụng trong biểu gene. Phương pháp tối ưu hoá sự biểu hiện của gene tách dòng là gắn gene đó vào plasmid dưới sự kiểm soát của promotor phiên mã liên tục và mạnh. Tuy nhiên, sự biểu hiện liên tục và ở mức độ cao của một gene ngoại lai thường gây hại cho tế bào chủ vì nó gây ra sự thiếu hụt năng lượng, mất cân bằng trong tế bào là suy giảm những chức năng thiết yếu của tế bào. Đồng thời một số hay toàn bộ các plasmid chứa gene tách dòng sẽ bị mất đi sau một số lần phân chia tế bào. Những tế bào không chứa các plasmid mang gene tách dòng sẽ phân chia mạnh hơn các tế bào chứa plasmid mang gene tách dòng và các tế bào không chứa các plasmid mang gene tách dòng sẽ chiếm ưu thế. Điều này đã cản trở quá trình biểu hiện và sản xuất các sản phẩm từ gene tách dòng. Để khắc phục khó khăn này người ta có ý tưởng mong muốn kiểm soát được sự phiên mã của gene táchdòng ở mức chỉ cho gene này biểu hiện ở một giai đoạn sinh trưởng hất định của tế bào. Sử dụng một promotor mạnh và điều khiển được sẽ là giải pháp thực hiện mong muốn đó. Promotor mạnh và điều khiển được đang sử dụng rộng rãi là promotor của operon lac và tryp của E. conli Promotor lac được thiết kế và chế tạo invitro chứa: - Vùng -10 của lac promotor. - Vùng -35 của tryp promotor. - Promotor bên trái (pL) của bacteriophage λ - Promotor gene 10 của bacteriophage T7.
141

Mỗi promotor này tương tác với chất ức chế và nhờ đó có thể tắt hoặc mở sự phiên mã của những gene tách dòng. Mỗi promotor lại dược nhận ra bởi cấu trúc của ARN polimerase holoenzyme nhờ nhân tố σ (nhân tố σ bám vào lõi của ARN polimerase để hình thành holoenzyme, định hướng cho enzyme bám vào những vùng promotor). Promotor lac bị ức chế (tắt) bởi protein kìm hãm lac khi môi trường thiếu lactose và được mở bằng cách bổ sung vào môi trường nuôi cấy lactose hoặc isopropyl-β -D-thiogalactopyranoside (IPTG). Lactose và IPTG bán vào promotor lac và quá trình phiên mã sảy ra. Quá trình phiên mã từ promotor lac còn được điều khiển bởi protein hoạt hoá chuyển hoá (catabolite activator protein-CAP). Khi CAP bám vào promotor lac sẽ làm tăng ái lực của promotor với enzyme phiên mã. Promotor tryp bị ức chế bởi phức hệ protein kìm hãm, phức hệ này bám vào operator tryp và ngăn can quá trình phiên mã. Promotor mở bằng việc loại bỏ tryptophan hoặc thêm vào môi trường axit 3-indoleacrylic. Promotor pL được kiểm soát bởi protein kim hãm cI của bacteriophage λ . Promotor của bacteriophage T7 đòi hỏi phải có enzyme polimerase T7 ARN để thực hiện phiên mã. 3.2. Kĩ thuật biểu hiện gene Gene được biểu hiện là gene đã được phân lập và đã biết trình tự. Các gene này có thể được phân lập bằng các phương pháp khác nhau. Sau khi có đoạn gene mong muốn chúng sẽ được biểu hiện trong các hệ thích hợp theo các bước các cạnh. Bước 1: Lựa chọn hệ thích hợp để biểu hiện gene. Bước 2: Dựa vào trình tự gene, thiết kế các đoạn mồi để nhân gene. Tổng hợp mồi. Bước 3: Chuẩn bị ADN không để nhân gene. Bước 4: Nhân gene bằng PCR từ ADN hệ gene của vi khuẩn hoặc từ ngân hàng cADN nhưng tốt nhất là nhân gene trực tiếp từ chính plasmid mang gene đã từng dùng để đọc trình tự gene đó. Bước 5: Đưa gene vào vector tách dòng và biến nạp vào tế bào E. coli để khuếch đại dòng gene. Bước 6: Tách plasmid từ các thể biến nạp E. coli. Kiểm tra gene trong vector tách dòng bằng enzym hạn chế. Kiểm tra gene và đặc biệt là chiều của gene trong vector biểu hiện. Gene cần biểu hiện phải nằm đúng chiều của prromotor. 142

Bước 7: Đưa gene vào vector biểu hiện và biến nạp vào tế bào E. coli.. Bước 9: Biến nạp vector biểu hiện có mang gene vào tế bào chủ (E. coli hoặc nấm men). Bắt đầu từ bước này, các thao tác để biểu hiện gene ở các chủng khác nhau là khác nhau. Bước 10: Kiểm tra sự biểu hiện của gene. 3.2.1. Lựa chọn hệ biểu hiện E. coli và Nấm men là hai hệ được sử dụng nhiều trong kĩ thuật biểu hiện gene. E. coli được dùng để biểu hiện rất nhiều gene có nguồn gốc cả từ Prokaryot và Eukaryot. Nấm men là cơ thể có ích được sử dụng rất rộng rãi để sản xuất nhiều loại protein tái tổ hợp dùng làm dược liệu để chữa bệnh và cả protein dùng trong công nghiệp thực phẩm. Các gene khi được biểu hiện trong tế bào E. coli tránh được hiện tượng protein bị glicosil hoá, trong khi đó ở nấm men lại có hiện tượng glicosil hóa. Tế bào E.coli sinh trưởng nhanh, thuận tiện cho các thao tác sinh học phân tử nên E. coli cũng thường được chọn để biểu hiện gene dùng cho những mục đích nghiên cứu khác nhau. Tuy nhiên, việc biểu hiện các đoạn gene lớn trong tế bào E. coli có những hạn chế như protein tái tổ hợp bị tồn tại ở dạng không tan, dạng thể vùi và không được tiết ra ngoài môi trường. Ở nấm men các protein tái tổ hợp này thường được tiết ra ngoài môi trường rất hiệu quả, có độ tinh sạch cao và đảm bảo hoạt tính sinh học của chúng. Men có thể biểu hiện và tiết tốt các sản phẩm có nguồn gốc từ động vật, thực vật bậc cao, nấm và vi khuẩn ra ngoài môi trường. 3.2.2. Thiết kế mồi để nhân gene Mồi dùng để nhân gene bằng PCR là một cặp mồi (mồi xuôi và ngược được thiết kế để bám vào vùng tận cùng ở 2 đầu của gene cần nhân. Trước khi thiết kế mồi để nhân gene, việc đầu tiên quan trọng là phải kiểm tra trình tự gene, quyết định đoạn gene cần nhân để đưa vào biểu hiện và kiểm tra các điểm cắt của enzyme hạn chế trong đoạn gene. Để tiện cho việc đưa gene vào vector biểu hiện, các đoạn mồi nhân gene thường được thiết kế có chứa thêm trình tự của enzyme hạn chế ở 2 đầu tận cùng của gene. Khi đó gene có thể được cắt trực tiếp bằng các enzyme này để dưa vào vector biểu hiện. Trình tự enzyme hạn chế được đưa thêm vào mồi cũng được lựa chọn rất kĩ càng. Trước hết các trình tự này phải có mặt trong vùng đa nối (polilinkers, multiple cloning sites) của vector biểu hiện. Tiếp theo, các trình tự này phải là trình tự rất hiếm gặp ở trong gene cần biểu hiện (tốt nhất là trình tự này không có mặt trong gene cần biểu hiện). Trình tự enzyme được thêm vào mồi xuôi nên là trình tự enzyme giới hạn đứng ngay đầu 5' của vùng đa nối còn enzyme hạn chế được thêm vào mồi ngược là trình tự enzyme đứng tận cùng đầu 3' của vùng đa nối. Nếu 2 mồi
143

có thêm 2 enzyme giới hạn khác nhau thì enzyme giới hạn ở mồi xuôi phải đứng trước cnzyme giới hạn ở mồi ngược trong vùng đa nối, tính theo chiều promotor để đảm bảo đoạn gene đưa vào sẽ nằm xuôi chiều với promotor. Mồi luôn được thiết kế và được ghi theo chiều từ 5' đến 3' của gene. Mồi xuôi có trình tự tương đồng với đoạn trình tự của sợi không mã hoá (sợi antisense). Đoạn trình tự tương đồng thường được sử dụng ở mỗi mồi để đảm bảo kết cặp đặc hiệu là khoảng trên 15 nucleotit trở lên. Tận cùng đầu 5' của mồi nên để dư ra ít nhất một nucleotit để đoạn gene nhân lên có trình tự được nguyên vẹn. Nucleotit dư ra ở đầu 5' không nên là nucleotit có khả năng bắt cặp tương đồng tốt với nucleotit ở tận cùng đều 3' trên cùng đoạn mồi, kích thước đoạn mồi cũng không nên quá dài. Tối thiểu chỉ cần 18 nucleotit của đoạn mồi bắt cặp chính xác với đoạn gene cần nhân là được. Tất nhiên độ dài của đoạn mồi có thể thay đổi phụ thuộc vào tỉ lệ các gốc A, T tại vị trí trong đoạn gene mà đoạn mồi cần bắt gặp. Sau khi thiết kế được mồi, việc quan trọng là kiểm tra sự dịch mã của đoạn gene sẽ được nhân lên bằng 2 mồi vừa thiết kế trong vector biểu hiện. Khung đọc của gene phải được đảm bảo một cách nghiêm ngặt. 3.2.3. Chuẩn bị ADN khuôn để nhân gene Để nhân được gene bằng PCR, nguyên liệu đầu tiên cần phải chuẩn bị đó là ADN khuôn. 3.2.4. Nhân gene bằng PCR Trước hết, sợi ADN khuôn phải được biến tính để sợi ADN khuôn trở thành dạng sợi đơn. Sau đó việc tổng hợp và nhân gene diễn ra với khoảng 25 - 30 chu kì. Sau khi kết thúc các chu kì trên, mẫu được ủ ở 750C trong khoảng 3 đến 6 phút, sau đó giữ mẫu ở 40C. Để tiến hành nhân gene bằng PCR, ngoài 2 đoạn mồi và ADN sợi khuôn ra thì phức hợp mẫu được trộn còn có nước khử ion vô trùng, 4 loại deoxyribonucleotit, Taq DNA polimerase hoặc Vent polimerase, đệm thích hợp cho Taq polimerase hoặc Vent polimerase, MgCl2. Thông thường MgCl2 có sẵn trong đệm nên có thể không cần phái bổ sung thêm nữa. Nồng độ MgCl2 ở trong mẫu càng lớn thì càng làm tăng khả năng bắt cặp không đặc hiệu của mồi. Taq polimerase tổng hợp gene có dư ra một nucleotit A ở đầu 3' còn Vent polimerase tổng hợp gene cho đầu bằng. Vì vậy, tuỳ theo điều kiện tách dòng mà chọn Taq hay Vent cho PCR. Nếu sản phẩm PCR được đưa vào vector tách dòng là đầu bằng thì nên chạy PCR bằng Vent. Vent ít gây ra sai sót trong trình tự gene được nhân hơn Taq. Nếu sử dụng một số vector tách dòng được mua từ các hãng, các vector này đã được thiết kế ở dạng thẳng và có dư nucleotit T có khả năng kết
144

cặp tương đồng với nucleotit A dư ra ở sản phẩm PCR thì nên sử dụng Taq polimerase. Kiểm tra sản phẩm PCR trên gel agarose. ADN tích điện âm có khả năng chuyển động trong điện trường theo chiều từ cực âm tới cực dương. Tốc độ chuyển động phụ thuộc vào 2 yếu tố: dạng tồn tại của ADN và kích thước ADN. Các ADN có kích thước lớn sẽ chuyển động chậm trong gel agarose còn ADN có kích thước nhỏ sẽ di chuyển nhanh hơn. CÁC ADN ở dạng siêu xoắn cũng chuyển động nhanh hơn ADN dạng thẳng. Từ một phạm vi nhất định phụ thuộc vào nồng độ gel agarose, kích thước phân tử tuyến tính với quãng đường địch chuyển của ADN trên gel agarose. Thường các đoạn gene có kích thước từ 300 đến 10000bp có thể được phân tách trên gel agarose 0,8%. Đoạn gene càng nhỏ thì nồng độ agarose càng phải tăng lên và để phát hiện các đoạn gene nhỏ tới vài chục nucleotit hoặc một chuỗi nhỏ nucleotit người ta thường phải sử dụng gel poliacrylamid. Các đoạn gene trên gel agarose được phát hiện bằng cách nhuộm với EtBr hoặc bằng đánh dấu phóng xạ. Thông thường nhất là các đoạn gel được nhuộm bằng ethidium bromit và được phát hiện bằng đèn chiếu tia UV. Để xác định được kích thước sản phẩm PCR, thường sản phẩm PCR được chạy song song với đoạn phức hợp ADN chuẩn đã biết kích thước. 3.2.5. Đưa gene vào vector tách dòng và biến nạp vào tế bào E. coli
Đưa gene vào vector tách dòng Sản phẩm PCR gắn trực tiếp với vector và sau đó được biến nạp vào tế bào E. coli DH5α để chọn lọc. Plasmid dùng cho tách dòng dược sử dụng rộng rãi là pCR2.1 và pUC118. Hai vector tách dòng này có khả năng cho số lượng bản sao lớn trong tế bào (200 bản sao/tế bào), có khả năng tải các đoạn gene tốt và ổn định trong tế bào chủ. Để đưa đoạn gene vào vector tách dòng bằng đầu bằng, trước hết plasmid phải được cắt trực tiếp bằng enzyme hạn chế đầu bằng có mặt trong vùng đa nối của vector tách dòng. Sau khi được cắt bằng một enzyme, plasmid rất dễ bị đóng vòng ngay cả khi không có DNA-ligase, vì vậy vector đã được mở vòng phải được xử lí bằng Alkaline photphatase. Enzyme này có khả năng loại gốc phosphate ở đầu 5' của ADN. Sản phẩm PCR không có gốc phosphate. Vì vậy, để có thể gắn với vector đã bị loại phosphate thì sản phẩm PCR phải được gắn thêm gốc phosphate nhờ polinucleotit kanase. Nếu sản phẩm PCR được nhân lên bằng Taq polimerase thì trước khi gắn phosphate, đoạn ADN đó phải được làm bằng đầu bằng enzyme.
Xử lí ADN bằng enzyme hạn chế Phản ứng nối ghép gene
145

Biến nạp sản phẩm lai vào tế bào E. coli: Có hai cách chính để biến nạp sản phẩm lai vào tế bào E. coli là phương pháp sốc nhiệt và phương pháp xung điện. 3.2.6. Tách ADN plasmid và kiểm tra gene trong plasmid
• Tách plasmid Các tế bào E. coli mang plasmid sau khi nuôi cấy đến pha dừng thì được li tâm thư lại. Thành và màng tế bào được phá vỡ bằng kiềm và chất tạo sức căng bề mặt. Sau đó ADN hệ gene và protein được tủa lại cùng với SDS bằng muối axetat kali và bị loại bỏ khỏi ADN plasmid bằng cách li tâm. ADN plasmid được tủa lại bằng cồn.
• Kiểm tra gene trong plasmid Gene ngoại lai trong plasmid được kiểm tra sơ bộ dựa trên trình tự các enzyme hạn chế có trong gene. Enzyme hạn chế được chọn để cắt kiểm tra là enzyme có thể cho ra những đoạn ADN có kích thước thích hợp có thể phát hiện được trên gel agarose. 3.2.7. Đưa gene vào vector biểu hiện Các plasmid tách dòng mang đoạn gene mong muốn được cắt hoàn toàn bằng hai enzyme hạn chế đã được thiết kế ở hai mồi. Vector biểu hiện cũng được cắt bằng enzyme tương tự. Sau đó vector và đoạn gene có kích thước như mong muốn được tinh sạch từ gel agarose và được nối lại với nhau bằng DNA-ligase. Nếu đoạn gene được đưa vào vector biểu hiện bằng một enzyme hạn chế thì phải kiểm tra chiều của gene để tránh hiện tượng các đoạn gene nằm ngược chiều với chiều phiên mã các promotor. 3.2.8. Biến nạp vector biểu hiện vào tế bào chủ Việc biến nạp vector vào tế bào E. coli và tế bào nấm men là khác nhau. Các tế bào E. coli thường được biến nạp bằng phương pháp sốc nhiệt (như đã được trình bày ở phần trên) còn các tế bào nấm men được biến nạp bằng phương pháp xung điện hoặc bằng muối lithium acetat những phương pháp xung điện cho hiệu quả biến nạp tốt hơn và được sử dụng rộng rãi hơn là phương pháp dùng muối. 3.2.9. Kiểm tra sự biểu hiện của gene Bước quan trọng để kiểm tra sự biểu hiện của gene là nuôi cấy các thể biến nạp trong môi trường có chất cảm ứng promotor để protein tái tổ hợp có thể được tổng hợp. Tiếp theo, để kiểm tra protein tái tổ hợp người ta thường dùng phương pháp điện di protein tổng số trên gel acrylamit để phát hiện sơ bộ băng protein tái tổ hợp. 146

3.3. Biểu hiện gene ngoại lai ở vi khuẩn E. coli Trong biểu hiện gene ở E. coli cần chú ý đối với các protein độc và trở ngại lớn nhất là một số protein ngoại lai rất độc đối với E. coli (ví dụ như RIPs) hoặc protein có khả năng gắn vào màng trong vi khuẩn E. coli và phá huỷ phức hợp màng. Như vậy việc tổng hợp những protein này chắc chắn bị hạn chế. Tuy nhiên, E. coli vẫn là một chủng chủ có giá trị để biểu hiện gene ngoại lai và trong nhiều trường hợp nó vẫn là thủng được ưa chuộng sử dụng. Khi biểu hiện bất cứ một gene ngoại lai nào ở E. coli cần quan tâm đến việc lựa chọn plasmid và promolor phù hợp cho quá trình khởi đầu phiên mã. 3.3.1. Cấu trúc vector biểu hiện ở E. coli Vector dùng để biểu hiện gene Prokaryot được thiết kế đầy đủ bao gồm tập hợp tác nhân tố di truyền được sắp xếp hợp lí nhằm điều khiển tối ưu cả quá trình phiên mã, dịch mã để tổng hợp nên protein có chức năng dùng làm dấu hiệu chọn lọc. Gene thường được sử dụng trong vector biểu hiện là gene kháng chất kháng sinh. Gene này tạo điều kiện thuận lợi cho việc nhận biết các dòng tế bào mang plasmid. Đối với plasmid tự sao chép: một trình tự không thể thiếu đó là tâm sao chép. Tâm sao chép quyết định số lượng bản copy của plasmid trong một tế bào. Để tạo điều kiện thuận lợi cho việc đưa các đoạn gene vào vector biểu hiện theo đúng chiều promotor. trên vector luôn phải chứa vùng đa nối (Poilinker region) mang các trình tự của enzyme hạn chế có tần số xuất hiện thấp trên các gene cấu trúc. 3.3.1.1. Promotor Các promotor có khả năng biểu hiện gene cao có những đặc điểm chung sau: (1) promotor mạnh có khả năng tạo protein ngoại lai lớn hơn 10 đến 30% protein tổng số của tế bào. (2) Promotor chỉ hoạt động phiên mã gene ở mức độ cơ sở (phiên mã khi không cảm ứng) với mức độ nhỏ tối đa: (3) Promotor được cảm ứng theo cách đơn giản nhưng hiệu quả cao. Sự cảm ứng bằng nhiệt hoặc bằng hoá chất thường được sử dụng rộng rãi để tạo ra lượng lớn protein mong muốn. Operon lac là một hệ điều hoà hoạt động của gene ở Prokaryot đượcbiết đến sớm nhất và promotor lac được sử dụng rộng rãi để tạo protein ngoại lai. Promotor từ operon tryptophan của E. coli được lựa chọn để biểu hiện protein ở bên trong tế bào. Promotor tryp có một số ưu điểm: (1) là promotor được nghiên cứu rất kĩ; (2) là promotor mạnh, các gene ngoại lai được phiên mã và dịch mã bởi promotor tryp thường tạo ra lượng protein lớn; (3) mức biểu hiện gene cơ bản (khi promotor không được cảm ứng) tương đối thấp, vì vậy việc biểu hiện protein ngoại lai sẽ không ảnh hưởng tới sự sinh trưởng của tế bào và
147

làm cho plasmid tồn tại ổn định hơn trong tế bào; (4) vector biểu hiện sử dụng promotor này có thể biến nạp vào hầu hết các chủng vi khuẩn E. coli, nhờ đó người ta có thể chọn bất kì chủng E. coli nào có đặc điểm mong muốn để biểu hiện gene; (5) việc cảm ứng promotor là rất đơn giản kể cả khi sử dụng bình lắc hoặc nồi lên men. 3.3.1.2. Terminator (vùng kết thúc phiên mã) Quá trình phiên mã được bắt đầu từ promotor, sau đó sợi mARN được kéo dài đến khi gặp một trình tự đặc hiệu, tại đó quá trình phiên mã bị dừng lại. Trình tự đặc hiệu đó là vùng kết thúc phiên mã (terminator). Có hai cơ chế ngừng phiên mã là cơ chế làm giảm tốc độ phiên mã và cơ chế dừng không đặc hiệu. Vị trí giảm phiên mã (attenuator) mang đặc điểm của một terminator đơn giản, được gắn giữa promotor và gene cấu trúc đầu tiên trong một operon. Vì attenuator là yếu tố điều khiển âm (negative regulatory elements) nên thường không được sử dụng trong hệ biểu hiện. Quá trình ngừng phiên mà không đặc hiệu có thể xuất hiện trong bất kì một quá trình phiên mã nào do có chứa trình tự trên ARN thông tin giống với trình tự của terminator một cách ngẫu nhiên. Các yếu tố chống lại sự dừng phiên mã (antiterminator) cũng được đưa vào vector biểu hiện để chắc chắn đoạn ARN thông tin được tổng hợp có độ dài đầy đủ. 3.3.1.3. Peptit tín hiệu (signal peptite) Các protein ngoại lai được tổng hợp trong tế bào E. coli cần thiết phải được tiết ra ngoài vì: (1) E. coli tiết rất ít protein ra ngoài môi trường nên protein ngoại lai nếu được tiết ra sẽ có độ tinh sạch cao. (2) Protein tiết ra ngoài khoang chu chất (perplasmid space) có khả năng hình thành cấu trúc bậc 4 đúng để trở thành protein có hoạt tính sinh học. Quá trình cuộn xoắn và hình thành cầu disulfit của protein nằm trong tế bào thường bị hạn chế và dẫn đến sai hỏng trong cấu trúc protein. (3) Protein khi được tiết ra ngoài sẽ tránh bị phân cắt bởi một số protease có trong tế bào. Để được tiết ra khỏi màng trong tế bào, các chuỗi polipeptit bắt buộc phải có trình tự peptit tín hiệu nằm ở tận cùng đầu N. Thông thường chuỗi peptit tín hiệu được cấu tạo gồm 3 vùng: vùng tận cùng đầu N, vùng tận cùng đầu C và vùng kị nước. Mỗi vùng có tối đa 20 axit amin và tối thiểu là 2 axit amin đối với vùng N, 8 axit amin đối với vùng kị nước và 6 axit amin đối với vùng C. 3.3.1.4. Điểm sao chép (Ori) Số lượng bản sao của plasmid trong tế bào phụ thuộc vào nhân tố khởi đầu sự sao chép ADN tại một vị trí đặc hiệu gọi là tâm sao chép (Ori). Đối với các plasmid sao chép tự do trong tế bào chủ, số lượng bản sao ít nhất cũng bị
148

ảnh hưởng bởi tốc độ sinh trưởng của tế bào. Những plasmid đa phiên bản thường bị mất đi qua các thế hệ tế bào (10-5- 10-6 trên một thế hệ) khi tế bào mang plasmid được nuôi cấy trong môi trường không có áp lực chọn lọc hoặc gene trên plasmid mã hoá cho protein gây độc cho tế bào. 3.3.1.5. Dấu chuẩn chọn lọc Để tiện lợi cho việc nhận ra các tế bào biến nạp có mang plasmid, thông thường người ta sử dụng môi trường nuôi cấy mà ở đó chỉ có tế bào có tính trạng chọn lọc nhất định mới có thể sinh trưởng được. Có một cách giúp dễ dàng nhận biết các thể biến nạp mang plasmid biến nạp đó là đưa các đoạn gene kháng kháng sinh vào plasmid đó. Sự có mặt của plasmid trong tế bào giúp cho tế bào sống sót trên môi trường có chất kháng sinh. Vì vậy, gene kháng kháng sinh có mặt ở hầu hết các vector tách dòng hiện đang được sử dụng như dấu chuẩn chọn lọc. 3.3.1.6. Vùng đa nối (Polilinker region) Vùng đa nối bao gồm trình tự nhận biết đặc hiệu của các enzyme hạn chế hay được sử dụng giúp cho quá trình đưa đoạn gene ngoại lai vào vector biểu hiện trở nên dễ dàng. Hầu hết trình tự của các enzyme hạn chế trong vector biểu hiện đều được đặt theo đúng khung đọc và nằm sau đoạn peptit tín hiệu (Signal peptit). Để tiện cho việc ghép nối các đoạn gene cần biểu hiện, các điểm nhận biết của các enzyme hạn chế trong vùng da nối được thiết kế sao cho chúng chỉ tồn tại như những vị trí duy nhất trên toàn bộ phân tử plasmid. 3.3.2. Quá trình tổng hợp protein ngoại lai Quá trình khởi đầu dịch mã trên phân tử ARN thông tin ở E. coli đòi hỏi một trình tự nhận biết đặc hiệu Shine - Dalgarno (SD) bổ sung với tận cùng 3' của ARN ribosom 16S. Đó là trình tự 5' - UAAGGAGG - 3' đứng trước bộ ba khởi đầu. Bộ ba khởi đầu thông thường nhất là AUG. Có khoảng 8% bộ ba khởi đầu dịch mã GUG, ngược lại trình tự UUG, AUU là trình tự khởi đầu rất hiếm gặp, thường chỉ có mặt ở các gene điều khiển một cách tự phát (ví dụ gene mã hoá cho protein S20 của ribosome, và nhân tố khởi đầu số 3). Khoảng cách từ SD đến mã khởi đầu thường là 8 nucleotit để đảm bảo sự khởi đầu dịch mã tối ưu nhất. Tuy nhiên, sự khởi đầu dịch mã chỉ ảnh hưởng rõ rệt khi khoảng cách đó giảm xuống dưới 4 nucleotit hoặc dài hơn 14 nucleotit. Thường ở Prokaryot, quá trình phiên mã và dịch mã diễn ra rất gần nhau, vì vậy cấu trúc bậc 2 của mARN có thể bao quanh trình tự SD làm cho quá trình biểu hiện gene bị giảm. Điều này được giải quyết bằng cách làm tăng trình tự tương đồng của SD với trình từ 5' UAAGGAGG và tăng số lượng adenin trong vùng khởi đầu. Cấu trúc của đoạn thêm vào của ARN thông tin (Dowstream Box - DB) có ảnh hưởng tới
149

khởi đầu dịch mã được đặt vào đầu mã khởi đầu và bổ sung với bazơ nitơ 1469-1483 của rARN 16S. Các DB có trình tự là 5' - AUGAAUCACAAAGUG 3' đóng vai trò chủ yếu như nhân tố tăng cường dịch mã. Mặc dù khi đưa trình tự DB vào đầu 5' của gene mã hoá cho protein tái tổ hợp có thể làm thay đổi trình tự axit amin của chúng. Trong tế bào tốc độ kéo dài các chuỗi polipeptit không xảy ra với tốc độ như nhau. Ở E. coli các bộ ba trong gene có mức độ sử dụng khác nhau phụ thuộc vào sự có mặt của các ARN vận chuyển tương ứng trong tế bào. Vì vậy các gene được biểu hiện yếu có thể là do gene có chứa nhiều bộ ba chỉ dược nhận biết bởi một số loại ARN vận chuyển hiếm gặp. Trong 6 bộ ba mã hoá cho Leu UUA, UUG, CUU, CUC, CUA, CUG, thì chỉ có CUG được nhận biết tốt và được biểu hiện cao bởi ARN vận chuyển đặc thù cho bộ ba này trong tế bào E. coli. Ở E. coli, chuỗi peptit mới tổng hợp rời khỏi phức hệ ARN thông tin và ribosom nhờ các nhân tố nhận biết mã kết thúc: RF1 nhận biết mã UAA và UAG còn RF2 nhận biết UAA, UGA. Ba mã kết thúc UAA, UAG và UGA có hiệu quả dừng dịch mã khác nhau, trong đó bộ ba UAA được các nhân tố kết thúc dịch mã nhận biết tốt nhất. 3.3.3. Quá trình sửa đối protein sau khi dịch mã 3.3.3.1. Quá trình thuỷ phân do protease Việc biểu hiện protein ngoại lai trong tế bào E. coli nhiều khi không hiệu quả do các protein này bị thuỷ phân bởi protease có trong tế bào chủ. Có nhiều bằng chứng cho thấy quá trình thuỷ phân này phụ thuộc vào năng lượng. Vì vậy, các thể đột biến ảnh hưởng tới protease phụ thuộc năng lượng sẽ đảm bảo ổn định của protein ngoại lai. Bên cạnh đó việc định hướng protein tiết ra ngoài khoang chu chất hoặc ra môi trường nuôi cấy tế bào ở nhiệt độ thấp, cho gene ngoại lai biểu hiện cùng với sự biểu hiện của phân tử "bảo mẫu ("chaperone"). Trong tế bào các protease này có chức năng quan trọng là thuỷ phân các protein bất thường và proteinkhông hoàn thiện. 3.3.3.2. Quá trình tiết protein Một trong những vấn đề hay gặp khi biểu hiện protein ở nội bào của tế bào E. coli là chúng thường tạo thành thể vùi (inclusion bodies). Mặc dù biểu hiện protein dưới dạng thể vùi cũng có một số ưu điểm nhất định nhưng protein tạo ra thường bị mất hoạt tính sinh học và quá trình tái tạo lại cấu trúc (refolding) tương đối phức tạp và tốn kém. Hầu hết protein được biểu hiện trong E. coli đều được thiết kế để sau khi biểu hiện chúng được chuyển ra khoang chu chất (perplasmic space) của tế bào. Một yếu tố không thể thiếu để protein có thể được chuyển qua màng trong ra ngoài khoang chu chất là đoạn peptit tín hiệu. 3.3.3.3. Hình thành cấu trúc không gian của protein
150

Protein sau khi được tổng hợp muốn trở thành protein có hoạt tính sinh học thì chúng phải có cấu trúc không gian đúng. Quá trình hình thành cấu trúc không gian phụ thuộc rất nhiều vào môi trường tbn trong và bên ngoài tế bào. Nếu môi trường có chất gây biến tính mạnh như urê, guanidin - HCl và các tác nhân khử thì quá trình hình thành cấu trúc không gian không thể diễn ra thuận lợi. Sự tạo thành cầu disulfit trong quá trình cuộn xoắn của protein có thể diễn ra bởi oxi hoá trong không khí có sự xúc tác của ion kim loại hoặc phức hợp oxi hoá khử của S.
Một ví dụ về sự biểu hiện gene mã hoá cho RIP ở E. coli. RIP được biểu hiện bằng hệ vector pMAL - C2. Vector biểu hiện protein bất hoạt ribosome tipe I được thiết kế dựa trên cơ sở casette vector pMAL - C2 (RIKEN DNA Bank Japan). Để tiến hành, cặp primer có chứa các điểm cắt của EcoRI và HindIII đã được tổng hợp và sử dụng để nhân đoạn gene mã hoá cho toàn bộ phân tử RIP thành thục (bao gồm 247 amino axit, bắt đầu từ Asp24 và kết thúc bằng Ala27). Đoạn gene nhân được bằng PCR sau xử lí với EcoRI và HindIII được gắn vào vùng cắt gắn đa vị (MCS) của vector pMAL - C2 ở đúng các vị trí cắt của các enzyme kể trên.
THẢO LUẬN 1. Thế nào là sự phân lập gen? Nêu nguyên tắc và quy trình của các phương
pháp phân lập gen. 2. Phân biệt khái niệm thư viện bộ gene và thư viện cADN. 3. Biểu hiện gene là gì? 4. Giải thích kĩ thuật biểu hiện gene. 5. Biểu hiện gene ngoại lai ở vi khuẩn E. coli
151

Tài liệu tham khảo chính 1. Lê Trần Bình và cộng sự. Công nghệ sinh học thực vật trong cải tiến
giống cây trồng. NXB Nông nghiệp H.1997. 2. Lê Trần Bình, Lê Thị Muội. Phân lập gene và chọn dòng chống chịu với
điều kiện bất lợi ở cây lúa. NXB Đại học Quốc gia, 1998. 3. Lê Trần Bình, Phan Văn Chi, Nông Văn Hải, Trương Nam Hải, Lê Quang
Huấn. Áp dụng các kĩ thuật phân tử trong nghiên cứu tài nguyên sinh vật ở Việt Nam. NXB Khoa học và Kĩ thuật, 2003.
4. Phan Văn Chi và cộng sự. Sử dụng một số primer ngẫu nhiên để xác định rapds ở lúa DT-33 và Tám thơm. Tạp chí di truyền và ứng dụng, số 4- 1997, 15-18.
5. Phan Văn Chi. Khối phổ trong nghiên cứu proteomics. Tạp chí Công nghệ sinh học 1(1), 2003.
6. Hồ Huỳnh Thuỳ Dương. Sinh học phân tử. NXB Giáo dục H.1997 7. Helena Curtis. Sinh học. Người dịch: Phan Cự Nhân. NXB Giáo dục.
H.1995. 8. Lê Thị Thu Hiền. Luận án tiến sĩ sinh học, 2003. 9. Lê Thanh Hoà. Sinh học phân tử: Nguyên lí và ứng dụng. Tài liệu giảng
dạy sau dại học, 2003. 10. Phạm Thành Hổ. Di truyền học. NXB Giáo dục H. 1998 11. Kỉ yếu Hội nghị Công nghệ Sinh học toàn quốc. NXB Khoa học và kĩ
thuật H. 1999. 12. Kỉ yếu Hội nghị Công nghệ Sinh học toàn quốc. NXB Khoa học và kĩ
thuật H. 2003. 13. Đinh Duy Kháng. Ứng dụng các kĩ thuật miễn dịch học và SHPT trong
chẩn đoán bệnh ở người. The Second German-Vietnamese Workshop on Genetic Engineering and Bioinformatics, Bài giảng. 2002.
14. Trần Thị Phương Liên. Luận án tiến sĩ Sinh học. 1999. 15. Lê Đình Lương, Phan Cự Nhân. Cơ sở di truyền học. NXB Giáo dục.
1997. 16. Lê Đình Lương. Kĩ thuật di truyền. NXB Khoa học và Kĩ thuật. 2001. 17. Chu Hoàng Mậu. Luận án tiến sĩ sinh học. 2001. 18. Chu Hoàng Mậu. Di truyền học. Bài giảng. 2001. 152

19. Chu Hoàng Mậu, Nguyễn Thị Tâm. Giáo trình Di truyền học. 2004. 20. Những vấn đề nghiên cứu cơ bản trong sinh học. NXB Đại học Quốc gia,
2000. 21. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB khoa học
kĩ thuật, 2003. 22. Đinh Thị Phòng. Luận án tiến sĩ sinh học, 2001. 23. Nguyễn Tiến Thắng. Giáo trình sinh hoá hiện đại. NXB Giáo dục. 1998. 24. Nguyễn Xuân Thụ và cộng sự. Sử dụng dấu chuẩn RAPD để nhận dạng
một số giống chuối trồng ở Việt nam. Tạp chí di truyền và ứng dụng, số 3-1998, 20-25.
25. Phan Hữu Tôn. Phân tích đặc điểm của gene đột biến làm mất đồng thời các subinit a và b của protein b -Congglicinin bằng kĩ thuật PCR ở đậu tương. Tạp chí di truyền và ứng dụng, số 3-1997, 34-38.
26. Từ khoa học sinh học phân tử đến cuộc sống và chăm sóc sức khoẻ. Báo cáo khoa học Hội nghị Sinh học phân tử và hoá sinh toàn quốc, 2003.
27. Nguyễn Văn Uyển. Những phương pháp công nghệ sinh học thực vật. Nhà xuất bản Nông nghiệp H. 1995, tập I, 48-60.
28. Nguyễn Văn Uyển. Những phương pháp công nghệ sinh học thực vật. Nhà xuất bản Nông nghiệp H.1995, tập II, 1995, 56-63.
29. Biotechnology in Agrialture and Forestry. Vol 10, 1990. Legumes and Oilseed Crops Edited hy baiiaj Y.P.S NXB Spriger - Verlag.
30. Phan Văn Chi. Stategy for protein purifcation and identiication. The Second German-vietnamese Workshop on Genetic Engineenng and Bioinfurmatics. Bài giảng, 2002.
31. Foolad M.R., Siva A. and Rodriguer L.R.. Applications of polimerase chain reaction (PCR) to plant genome analisis. I: Tissue and organe culture. Fundamental methods. Spnnger Verlag Berhn, Heidelberg. 1995, 281-298.
32. Nông Văn Hải. Gene Cloning (Molecular Cloning/ Recombinant DNA) Techniques. The Second German-Vietnamese Workshop on Genetic Engineering and Bioinformatics, Bài giảng, 2002.
33. Jena K.K. et al. RFLP analisis ofrice (Oryza sativa L.) introgression lines. Theor. Appl. Genet 1992, 608-616.
34. Karuzina I.P. Giáo trình về cơ sở di truyền học. Người dịch Trịnh Bá Hữu. NXB Mir, 1985.
153

35. Long Mao. Molecular Markers and Their Applications in Rice and Wheat. Dissertation for a degree of Doctor of Philosophy, 1995, 70-78.
36. Monna L. Determination of RAPD markers in rice and their conversion into sequence tagged sites (STSS) and STS..Specirc primer. ADN Research 1, 1994, 139-148.
37. Soybean: Genetics, Molecular Biology and Biotechnology Edited by D. P. S Verma and R. C Shoemaker, CAB INTEARNTIONAL, 1996.
38. Tanksley S.D et al. RFLP mapping in plant breeding: new tools for an old science. Biotechnology vol 7, 1989, 257-263.
39. Vierling R., Nguyen H.T. Use of RAPD marker to determine the genetic relationships of diploid wheat geneotipes. Theor Appl. Genet. 84,1992, 835-838.
40. Weslh J., Mcclelland M. Fingerprinting genome using PCR with arbitraryprimer. Axit nucleics Res. 18, 1991, 7213-7218.
41. Yu J., Nguyen H.T. Genetic variation detected with RAPD markers among upland and lowland rice cultivars (oryza sativa L.). Theor Appl. Genet, 87, 1994, 668-672.
154

NHỮNG CÔNG TRÌNH CỦA TÁC GIẢ
CÙNG CÁC CỘNG TÁC VIÊN ĐÃ CÔNG BỐ 1. Chu Hoàng Mậu, Nông Thị Man (1994). Ảnh hưởng của tia gamma và
hoá chất hoá học nitrozometilure (NMU) đến khả năng nảy mầm và khả năng sống sót của một số giống đậu tương Glicine max (L.) Merrill ở Bắc Thái. Thông báo khoa học, Số 2, Chuyên ngành Sinh học, Trường Đại học Sư phạm Việt Bắc, 23-27.
2. Chu Hoàng Mậu, Nông Thị Man (1995). Hiệu ứng tế bào dưới tác động của tia gamma và hoá chất gây đột biến nitrozometilure ở các giống đậu tương Glicine max (L.) Merr. Thông báo khoa học, Số 1, Trường Đại học Sư phạm - Đại học Thái Nguyên, 62-66.
3. Chu Hoàng Mậu, Nông Thị Man (1995). Ảnh hưởng của tia gamma và hoá chất hoá học nitrozometilure đến tần số và biến dị của các giống đậu tương Glicinemax (L) Merrill. Thông báo khoa học, Số 2, Trường Đại học Sư phạm - Đại học Thái Nguyên.
4. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1995). Khả năng phản ứng kiểu gene của một số giống đậu xanh Vigna radirata (L) ở M1 và M2 đối với các tác nhân đột biến vật lí - hoá học. Thông báo khoa học, Số 3, Trường Đại học Sư phạm - Đại học Thái Nguyên.
5. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1996). So sánh ảnh hưởng của các tác nhân vật lí, hoá học lên quá trình sinh trưởng, phát triển và biến dị của các giống đậu xanh Vigna radiata (L.) địa phương ở M1. Tạp chí Khoa học và Công nghệ, Số 1, Đại học Thái Nguyên, 49-55.
6. Chu Hoàng Mậu, Nông Thị Man (1996). Phổ biến dị hình thái và giá trị trung bình một số tính trạng số lượng của hai giống đậu xanh mục tiêu Bảo Lạc và mỡ Chợ đồn ở M1. Thông báo khoa học, Số 4, Trường Đại học Sư phạm - Đại học Thái Nguyên, 81-88.
7. Chu Hoàng Mậu, Nông Thị Man (1997). Biến dị và di truyền của một số tính trạng số lượng của các giống đậu tương ở M3. Tạp chí Khoa học và Công nghệ, 1(2), Đại học Thái nguyên, 28-32.
8. Chu Hoàng Mậu, Đoàn Bích Liên, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1997), Hiệu quả của xử lí thuần và phối hợp các tác nhân đột
155

biến đến các giống đậu tương địa phương ở M1. Tạp chí Khoa học và Công nghệ, 2(3), Đại học Thái nguyên, 25-29.
9. Chu Hoàng Mậu, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1997). Đánh giá một số dòng đột biến từ hai giống đậu tương ĐH4 và Cúc vàng ở thế hệ M4. Tạp Chí sinh học, 19(3), 53-56.
10. Chu Hoàng Mậu, Đoàn Bích Liên, Nông Thị Man, Lê Trần Bình (1997). Đặc điểm phản ứng của giống đậu tương địa phương Lạng Sơn ở M2. Tạp chí di truyền và ứng dụng, Số 3. 24-28.
11. Chu Hoàng Mậu (1997). Hiệu quả gây biến dị ở giống đậu tương Lạng Sơn khi xử lí riêng rẽ và phối hợp tia gamma với các hoá chất ethyleneimine và nitrosomethyl urea. Tạp chí Di truyền và ứng dụng, Số 4, 1-5.
12. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1997). Đặc điểm của sự phát sinh biến dị ở thế hệ thứ hai của hai giống đậu xanh địa phương, miền núi. Kỉ yếu Annual Report, Viện Công nghệ Sinh học, Nhà xuất bản khoa học kĩ thuật, 76-83.
13. Chu Hoàng Mậu (1997). Tạo vật liệu khởi đầu trong chọn giống đậu tương bằng phương pháp đột biến thực nghiệm. Đề tài NCKH cấp cơ sở
14. Chu Hoàng Mậu, Đoàn Bích Liên, Đỗ Như Tiến, Nông Thị Man (1998). Gây biến đổi nhiễm sắc thể ở đậu tương và đậu xanh bằng xử lí riêng rẽ và phối hợp tia gamma với các hoá chất etilen imin và nitrozometilure. Thông báo khoa học, Số 2, Trường Đại học Sư phạm - Đại học Thái Nguyên, 29-35.
15. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man, Đào Việt Bắc (1998). Nghiên cứu hàm lượng protein, lipit và thành phần axit amin của một số dạng đậu dạng đột biến ở M2. Tạp chí Khoa học và Công nghệ, 2(5), Đại học Thái Nguyên, 39-44.
16. Chu Hoàng Mậu (1998). Nghiên cứu lí thuyết về kĩ thuật sinh học phân tử hiện đại ứng dụng trong phương pháp đột biến thực nghiệm, góp phần chọn tạo giống đậu tương và đậu xanh. Tạp chí Khoa học và Công nghệ, 4(7), Đại học Thái Nguyên, 63-68.
17. Chu Hoàng Mậu, Đoàn Bích Liên, Nông Thị Man, Lê Trần Bình (1998). Nghiên cứu hệ số di truyền và một số chỉ tiêu hoá sinh nhằm đánh giá các dòng đột biến đậu tương Lạng Sơn. Tạp chí Di truyền và ứng dụng, Số 1, 6-10.
18. Chu Hoàng Mậu, Đoàn Bích Liên, Nông Thị Man (1998). Đặc điểm 156

của sự phát sinh biến dị ở giống đậu lương Lạng Sơn trong thế hệ đột biến thứ ba. Tạp chí Di truyền và ứng dụng, Số 1, 28-34.
19. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man (1998). Đặc điểm các biến dị số lượng của hai giống đậu xanh địa phương ở thế hệ đột biến M3. Tạp chí Di truyền và ứng dụng, Số 3, 40-45.
20. Chu Hoàng Mậu, Nông Thị Man, Đào Việt Bắc, Lê Trần Bình (1998). Tạo biến dị ở hai giống đậu tương Cúc vàng và ĐH4 bằng tác nhân vật lí hoá học. Tạp chí Sinh học. 20(2), 55-58.
21. Chu Hoàng Mậu, Nông Thị Man, Lê Xuân Đắc, Đinh Thị Phòng, Lê Trần Bình (1999). Ứng dụng kĩ thuật phân tích hiện tượng đa hình các phân đoạn ADN được nhân bản ngẫu nhiên (RAPD) vào việc đánh giá bộ gene của các dòng đậu xanh đột biến. Tạp chí Sinh học. 21(3), 15-23.
22. Chu Hoàng Mậu, Nông Thị Man, Lê Trần Bình (2000). Đánh giá một số tính trạng kinh tế quan trọng và khả năng chịu hạn của các dòng đậu tương (Glicine max (L.) Merrill) đột biến. Tạp chí Khoa học và Công nghệ, 1(13), Đại học Thái Nguyên: 16-21.
23. Chu Hoàng Mậu, Nông Thị Man, Lê Xuân Đắc, Đinh Thị Phòng, Lê Trần Bình (2000). Đánh giá geneom của một số dòng đậu tương đột biến bằng kĩ thuật phân tích tính đa dạng ADN được nhân bản ngẫu nhiên. Tạp chí Sinh học, 22/(1). 21-26.
24. Chu Hoàng Mậu, Nông Thị Man, Nguyễn Huy Hoàng, Trần Thị Phương Liên, Lê Trần Bình (2000). Nghiên cứu hiện tượng đa hình protein của các dòng đậu tương và đậu xanh đột biến. Kỉ yếu Annual Report, NXB Khoa học kĩ thuật Hà Nội.
25. Chu Hoàng Mậu, Đỗ Như Tiến, Nông Thị Man, Lê Trần Bình (1996). Nghiên cứu một số biến dị hình thái và kinh tế quan trọng của các giống đậu xanh Vigna Radiata(L.) ở các thế hệ đột biến M1, M2. Kỉ yếu Annual Report, NXB Khoa học kĩ thuật Hà nội.
26. Đào Xuân Tân, Nông Thị Man, Chu Hoàng Mậu (2000). Đánh giá một số dòng đậu xanh đột biến ở M6. Tạp chí Khoa học và Công nghệ, 3(15), Đại học Thái Nguyên, 46-49.
27. Chu Hoàng Mậu, Nông Thị Man, Lê Trần Bình (2001). Phân tích một số chỉ tiêu hoá sinh trong hạt của các dòng đột biến đậu tương và đậu xanh. Tạp chí Khoa học và Công nghệ, 1(17), Đại học Thái Nguyên, 59-63.
28. Chu Hoàng Mậu, Nguyễn Phúc Chỉnh, Hoàng Mai Phương, Nguyễn 157

Lam Điền (2000). Phòng thí nghiệm Sinh học hiện đại với sự phát triển khoa học và công nghệ phục vụ sản xuất Nông - Lâm nghiệp địa phương. Tạp chí Khoa học và Công nghệ, 1(17), Đại học Thái Nguyên, 64-69.
29. Đỗ Thị Dương, Chu Hoàng Mậu, Nông Thị Man (2000). Sưu tập và đánh giá một số giống lúa cạn địa phương. Tạp chí Khoa học và Công nghệ, 1(17), Đại học Thái Nguyên, 13-16.
30. Hoàng Mai Phương, Chu Hoàng Mậu, Nông Thị Man, Nguyễn Thị Ngọc Lan (2000). Nghiên cứu thành phần điện di protein dự trữ hạt của một số giống lúa cạn và các dòng lạc đột biến. Tạp chí khoa học và Công nghệ, 1(17), Đại học Thái Nguyên, 26-29.
31. Chu Hoàng Mậu, Nông Thị Man, Lê Trần Bình (2001). Đánh giá năng suất, chất lượng hạt và khả năng chịu hạn của các dòng đậu xanh đột biến Vigna Radiata (L.) Willzeck. Tạp chí Sinh học, 23/(1).
32. Chu Hoàng Mậu (2000). Hàm lượng protein và đặc điểm bộ rễ của các dòng đậu tương đột biến ở giai đoạn nảy mầm và cây non. Thông báo khoa học, số 2, Trường Đại học Sư phạm - Đại học Thái Nguyên, 26-31.
33. Chu Hoàng Mậu (2000). Nghiên cứu hiện tượng protein đa hình và một số chỉ tiêu sinh hoá góp phần chọn lọc các dòng đậu tương và đậu xanh đột biến. Đề tài KHCN cấp Bộ, Mã số: B98-03-20.
34. Chu Hoàng Mậu (2000). Nghiên cứu phục vụ phát triển ngành trồng trọt ở khoa Sinh-Kĩ thuật nông nghiệp, trường Đại học Sư phạm Thái Nguyên. Hội thảo công tác chuyển giao khoa học công nghệ Nông - Lâm khu vực Trung du miền núi phía Bắc. Đại học Thái Nguyên, 43-44.
35. Chu Hoàng Mậu, Đỗ Thị Dương, Hoàng Mai Phương, Nông Thị Man (2001). Khả năng chịu hạn của một số giống lúa cạn địa phương ở giai đoạn cây mạ. Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật, 149-156.
36. Đỗ Thị Dương, Chu Hoàng Mậu, Nông Thị Man (2001). Đánh giá chất lượng hạt của một số giống lúa cạn địa phương. Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật, 35-39.
37. Nguyễn Thị Hương, Chu Hoàng Mậu (2001). Nghiên cứu đặc điểm phản ứng kiểu gene của một số giống lúa nếp nương địa phương ở giai đoạn cây mạ trong điều kiện gây hạn nhân tạo. Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật,. 69-75.
38. Nguyễn Thị Ngọc Lan, Chu Hoàng Mậu (2001). Nghiên cứu đặc điểm hình thái, chất lượng hạt và hàm lượng protein, chlorophyll của ba giống
158

lúa cạn địa phương. Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật 122-125.
39. Chu Hoàng Mậu, Hoàng Cao Nguyên, Nguyễn Thu Hiền, Lê Ngọc Công (2001). Nghiên cứu đặc điểm hình thái và một số chỉ tiêu sinh hoá của cây cỏ phượng vĩ (Pteris multifida Poir). Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật. 161-163.
40. Hoàng Mai Phương, Chu Hoàng Mậu, Nguyễn Thu Hiền (2001). Nghiên cứu đặc điểm hình thái, hoá sinh hạt của một số dòng lạc trồng tại Lào Cai. Kết quả nghiên cứu về Sinh học và Giảng dạy sinh học, NXB Khoa học và Kĩ Thuật, 173-176.
41. Chu Hoàng Mậu, Nông Thị Man, Lê Trần Bình (2001). Đánh giá năng suất, chất lượng hạt và khả năng chịu hạn của các dòng đậu xanh đột biến - Vigna radiata (L.) Wilzeck. Tạp chí Sinh học, 23(1). 54-60.
42. Chu Hoàng Mậu, Nông Thị Man, Lê Trần Bình (2001). Hàm lượng axit amin trong hạt của các dòng đậu tương (Glicine max (L.) Merrill và đậu xanh (Vigna radirata (L.) Wilczeck) đột biến. Tạp chí Sinh học, 23(4), 57-61.
42 Chu Hoàng Mậu, Nguyễn Thị Hải Yến, Nguyễn Thu Hà (2002). Kết quả phân tích đặc điểm hình thái, khối lượng và chất lượng hạt của một số giống lúa cạn sưu tập ở Thái nguyên và Hà Giang. Tạp chí Khoa học và Công nghệ, 2(22), Đại học Thái Nguyên, 62-67. 43. Chu Hoàng Mậu, Nguyễn Thị Sâm (2002). Đánh giá một số giống lạc
Arachis hypogea (L.) trồng tại Bắc Giang. Tạp chí Khoa học và Công nghệ, 3(23), Đại học Thái Nguyên, 5-10.
44. Chu Hoàng Mậu, Thân Thị Mỹ Ngọc (2002). Đặc điểm hình thái, khối lượng và hoá sinh hạt của một số dòng lạc đột biến. Tạp chí Khoa học và Công nghệ, 3(23), Đại học Thái Nguyên, 59-62.
45. Chu Hoàng Mậu, Lò Thanh Sơn (2002). Khảo sát đặc điểm hình thái, chất lượng hạt và những biến động hàm lượng protein ở giai đoạn mầm của một số giống lúa cạn địa phương. Tạp chí Khoa học và công nghệ, 3(23), Đại học Thái Nguyên, 68-76.
46. Chu Hoàng Mậu (2002), Nghiên cứu đặc điểm hoá sinh và sinh học phân tử của một số giống lúa cạn địa phương góp phần chọn lọc giống lúa cạn thích hợp cho vùng núi Đông Bắc Việt Nam. Đề tài KH-CN cấp Bộ, mã số B2001-03-14.
47. Nguyễn Thu Hà, Chu Hoàng Mậu, Nguyễn Thị Hải Yến, Đỗ Thị 159

Dương (2003). Đa dạng sinh học cây lúa cạn ở miền núi phía bắc Việt Nam. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 86-89.
48. Nguyễn Thị Kim Dung, Chu Hoàng Mậu (2003). Chất lượng hạt của các giống lạc L05, L08, L12, L14, MD7. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 306-308
49. Nguyễn Vũ Thanh Thanh, Chu Hoàng Mậu (2003). Đánh giá chất lượng hạt của một số giống đậu xanh Vigna radiata (L.) Willzech. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 398-400.
50. Chu Hoàng Mậu, Nguyễn Thị Sâm (2003). Nghiên cứu phổ điện di protein và thành phần axit amin trong protein lạc của một số giống lạc (Arachis hypogaea L.). Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 348-351.
51. Nguyễn Thu Hiền, Chu Hoàng Mậu, Nguyễn Thị Sâm (2003). Nghiên cứu một số đặc điểm hoá sinh trong giai đoạn hạt nảy mầm và cây non của một số giống lạc (Arachis hypogaea L.). Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 315-318.
52. Hoàng Thị Thu Yến, Chu Hoàng Mậu, Nghiêm Ngọc Minh, Nông Văn Hải, Trịnh Đình Đạt (2003). Phân lập gene chaperonill ở các dòng đậu tương đột biến ML10, ML48 và ML61. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống. NXB Khoa học và Kĩ thuật, 1073-1076.
53. Chu Hoàng Mậu (2004). Nghiên cứu về protein và thành phần axit amin trong protein hạt của một số giống lúa cạn địa phương. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống định hướng nông lâm nghiệp miền núi. NXB Khoa học và Kĩ thuật, 517-520.
54. Nguyễn Vũ Thanh Thanh, Chu Hoàng Mậu (2004). Nghiên cứu thành phần điện di protein dự trữ trong hạt của một số giống đậu xanh. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống định hướng nông - lâm nghiệp miền núi. NXB Khoa học và Kĩ thuật, 622-625.
55. Hoàng Thị Thu Yến, Chu Hoàng Mậu, Nguyễn Đình Cường, Trần Thị Phương Liên, Nông Văn Hải (2004). Đặc điểm nông sinh học, hoá sinh hạt và trình tự gene chaperonin của dòng đậu tương đột biến Ml61 có khả năng chịu hạn. Những vấn đề nghiên cứu cơ bản trong khoa học sự sống định hướng nông - lâm nghiệp miền núi. NXB Khoa học và Kĩ thuật, 733-737.
160

56. Phạm Thị Thanh Nhàn, Chu Hoàng Mậu, Vũ Hô (2004). Nghiên cứu đặc điểm huyết học, hoá sinh máu và phổ điện di protein huyết thanh SDS-PAGE của một số bệnh nhân ung thư phốỉ. Những vấn đề nghiên cứu cơ bản, trong khoa học sự sống định hướng y dược học. NXB Khoa học và Kĩ thuật, 291-294.
57. Phạm Thị Thu Nga, Chu Hoàng Mậu, Vũ Thị Thu Thuỷ (2004). Kết quả đánh giá và nghiên cứu tính đa dạng di truyền của một số giống lúa cạn địa phương sưu tập từ hai tỉnh Bắc Cạn và Cao Bằng. Hội nghị khoa học trường Đại học Khoa học Tự nhiên - Đại học Quốc gia Hà Nội.
58. Chu Hoàng Mậu, Nguyễn Thị Kim Dung (2004). Nghiên cứu tính đa hình protein trong hạt của một số giống lạc (Arachis hypogaea L.) bằng kĩ thuật điện di SDS-PAGE. Hội nghị khoa học trường Đại học Khoa học Tự nhiên - Đại học Quốc gia Hà Nội.
161

Chịu trách nhiệm xuất bản: Giám đốc ĐINH NGỌC BẢO Tổng biên tập LÊ A Người nhận xét: PGS.TS NGUYỄN TRỌNG LẠNG TS. LƯƠNG THỊ HỒNG VÂN
Biên tập: TRẦN THỊ THANH BÌNH
Kĩ thuật vi tính: ĐÀO PHƯƠNG DUYÊN
Trình bày bìa: PHẠM VIỆT QUANG
CƠ SỞ VÀ PHƯƠNG PHÁP SINH HỌC PHÂN TỬ
In 1000 cuốn, khổ 17 x 24cm, tại Công ti In Thái Nguyên. Giấy phép xuất bản số: 16 - 1845/XB - QLXB, kí ngày 20/10/2004. In xong và nộp lưu chiểu tháng 7 năm 2005.
162