Embed Size (px)
Citation preview

Pázmány Péter Katolikus Egyetem
Információs Technológiai és Bionikai Kar
Szakdolgozat
Mikroglia sejtek stressz indukálta morfo-
funkcionális elemzése
Gulyás Krisztina
Molekuláris bionika mérnöki BSc
2017
Témavezető:
dr. Kovács Krisztina

2
Alulírott Gulyás Krisztina, a Pázmány Péter Katolikus Egyetem Információs Technológiai és
Bionikai Karának hallgatója kijelentem, hogy ezt a szakdolgozatot meg nem engedett segítség
nélkül, saját magam készítettem, és a szakdolgozatban csak a megadott forrásokat használtam
fel. Minden olyan részt, melyet szó szerint, vagy azonos értelemben, de átfogalmazva más
forrásból átvettem, egyértelműen a forrás megadásával megjelöltem. Ezt a Szakdolgozatot más
szakon még nem nyújtottam be.
___________________________________
Aláírás

3
Tartalomjegyzék
1. Bevezetés ........................................................................................................................ 6
2. Irodalmi áttekintés .......................................................................................................... 8
2.1. Mikroglia sejtek funkciója és morfológiája ............................................................ 8
2.2. Stresszválaszt befolyásoló agyi régiók és hormonok .............................................. 10
2.3. Mikroglia sejtek kapcsolata a gyulladásos és a stresszel járó folyamatokkal ......... 11
2.4. Mikroglia sejtek különböző paramétereinek meghatározása képelemző
eljárásokkal ............................................................................................................. 12
2.5. Képek szegmentálására alkalmazott módszerek áttekintése ................................... 13
2.6. Sejtek szegmentálására használt képfeldolgozási technikák ................................... 15
3. Módszerek ....................................................................................................................... 17
3.1. Kísérleti állatok és kísérleti elrendezés ................................................................... 17
3.2. Perfúzió és szövetfeldolgozás ................................................................................. 19
3.3. Felvételek elemzésére kidolgozott módszer bemutatása ......................................... 21
3.3.1. Sejtek detektálása ........................................................................................... 22
3.3.1.1. Előfeldolgozás .................................................................................... 24
3.3.1.2. Szegmentálás ...................................................................................... 26
3.3.2. Nyúlványrendszer modellezése ...................................................................... 28
3.3.3. Paraméterek meghatározása ........................................................................... 31
3.4. Grafikus felhasználói felület bemutatása ................................................................ 32
3.5. Felvételek elemzése ImageJ szoftver segítségével .................................................. 35
4. Eredmények .................................................................................................................... 37
4.1. Krónikus stressz kísérlet ......................................................................................... 37
4.2. Az automatikus sejtdetekció eredménye ................................................................. 38
4.3. A különböző módszerekkel meghatározott paraméterek kiértékelése .................... 39
4.4. A felvételek elemzésére alkalmazott módszerek összehasonlítása ......................... 41
5. Diszkusszió, konklúzió ................................................................................................... 43
6. Összefoglalás .................................................................................................................. 46
7. Köszönetnyilvánítás ........................................................................................................ 47
8. Irodalomjegyzék ............................................................................................................. 48
9. Mellékletek ..................................................................................................................... 51

4
Kivonat
A mikroglia sejtek a központi idegrendszer rezidens immunsejtjei. Mikrokörnyezetük
megváltozására dinamikusan reagálnak mind funkcionális, mind pedig morfológiai értelemben.
Ennek hátterében állhat sérülés, kórokozók, de az újabb tanulmányok értelmében akár stressz
hatására bekövetkező változások is indukálhatják. A mai napig számos kutatás témája a
mikroglia morfológiája és funkciója közti összefüggés vizsgálata, illetve a csupán morfológia
alapján történő elemzés. A sejtek detektálása illetve szegmentálása kiemelt szerepet játszik a
sejtekre jellemző morfológiai információ kinyerésében. Manuális módon ez a folyamat igen
időigényes, mivel általában nagy adathalmaz áll rendelkezésre, valamint nem túl objektív, így a
feladat automatizálásába nagy erőket fektettek az elmúlt évtizedekben.
Jelen dolgozatban a stressz és a mikroglia aktiváció kapcsolatát vizsgáltuk krónikus
variábilis stressznek kitett patkányok hipotalamuszában. A kísérleti állatokat 3 héten keresztül,
napi két alkalommal véletlenszerűen válogatott stresszoroknak tettük ki, majd megmértük ennek
hatását az állatok testtömeg gyarapodására, cukor preferenciájára és ACTH illetve
kortikoszteron szintjére. Ez után immunhisztokémiai eljárás során egy mikroglia-specifikus
marker (ionizált kálciumkötő adapter molekula, Iba-1) segítségével jelenítettük meg a sejteket,
majd az eredményről fénymikroszkópos felvételeket készítettünk.
Az így kapott felvételek feldolgozásához kifejlesztettem egy MATLAB-alapú automatizált
képelemző eljárást, mely lehetővé teszi sejtek detektálását, nyúlványrendszerük modellezését
és jellemző paramétereik meghatározását, ezzel segítve a morfológiai elemzést. A detekció
alapját logaritmikus képjavítás és intenzitásérték-alapú szegmentáció képzi, míg a nyúlványok
modellezéséhez régió alapú aktív kontúr módszert használtam. A kifejlesztett algoritmus többek
között meghatározza a sejtek számát, az általuk elfoglalt területet és annak a legkisebb konvex
sokszögnek a nagyságát, mellyel a teljes sejt lefedhető (Convex Environmental Area, CEA). A
felvételek elemzését ImageJ programmal is elvégeztem, melyben globális intenzitásküszöb
alapján szegmentáltam a képeket.
A sejtek automatikus detektálására kifejlesztett módszer a sejtek 92.41%-át megtalálta.
A felvételek minőségbeli változatossága, és a mikrogliára jellemző morfológiai sokszínűség
miatt a kifejlesztett eljárás jobb eredményt adott a képen lévő összes sejt morfológiai
információjának egy lépésben történő kinyerésére, mint a globális intenzitás-alapú küszöbölés.
Összességében, a kapott eredmények azt mutatták, hogy a krónikus stressz hatására csökkent az
állatok testtömeg gyarapodása és cukor preferenciája, valamint fokozódott a hipotalamusz-
hipofízis-mellékvesekéreg tengely aktivitása. A mikroglia aktiváció tekintetében a krónikus
stressz jelentősen növelte a sejtek által elfoglalt összterületet a sejtek számában való növekedés
nélkül a hipotalamusz paraventrikuláris magjában (PVN), mely a neuroendokrin stresszválasz
egyik fő szabályzó központja.

5
Abstract
Microglia cells are resident immune cells of the central nervous system. They react with
dynamic morphological and functional changes as a response to any change in their
microenvironment, such as injury, pathogens, and even various systemic stressors due to recent
studies. Several researches focused on the relationship between morphology and function of
microglia, and analysis based solely on cell morphology. Detection and segmentation of cells
plays an important role in extracting morphological information. This process in a manual way
is time-consuming due to the large amount of available data and also not objective, so in the last
decades, many efforts have been devoted to automate this process.
In this study, we investigated the relationship between stress and microglia activation
in various regions of the hypothalamus of rats after chronic variable stress. For 3 weeks, the
experimental animals were subjected to randomly selected stressors twice a day. We determined
the effect of stress on the weight gain, sucrose preference, ACTH and corticosterone levels of
the animals. After that, immunohistochemistry was performed using a microglia-specific marker
(ionized calcium-binding adapter molecule, Iba-1), followed by imaging with a light
microscope.
To evaluate the images, we developed a MATLAB-based automated image analyzing method,
that allows detection of the cells, modeling of their processes and determining their
characteristic parameters thus helping morphological analysis. Detection is based on logarithmic
enhancement and intensity thresholding, while we used region-based active contours to model
the processes. The implemented algorithm among others, determines the number of cells, the
area occupied by the cells and the size of the smallest convex area which can cover the whole
cell (Covered Environmental Area, CEA). I also analyzed the images with ImageJ, using a
segmentation method based on global intensity thresholding.
The developed method for automated cell detection found 92.14% of the cells. Due to
the varying quality of the images and the morphological diversity of microglia, the proposed
method compared to the global intensity-based thresholding gave better results in extracting the
morphological information of all the cells. Overall, present results revealed that chronic stress
induce a decrease in sucrose preference and body weight gain, as well as increased the activity
of the hypothalamic-pituitary-adrenal axis. With regard to microglia activation, we found that
chronic stress significantly increased the total area covered by the cells in the hypothalamic
paraventricular nucleus (PVN), the main regulator of neuroendocrine stress response, without
an increase in the number of cells.

6
1. Bevezetés
A mikroglia sejtek, vagy más néven (felfedezőjük után), Hortega-féle sejtek a központi
idegrendszerben található gliasejtek, melyek fontos szerepet játszanak az idegrendszer
immunológiai védelmében [2]. Kóros folyamatot észlelve a mikroglia sejtek nyugvó
állapotukból morfológiai változásokon keresztül aktív/reaktív állapotba kerülnek [4]. Az elmúlt
időszakban számos tanulmány foglalkozott a mikroglia aktiváció és a stressz kapcsolatával.
Ezek többsége egyetért abban, hogy a stressznek szerepe lehet a mikroglia morfológiai
aktiválásában [4-5][10][35].
A sejtek morfológiájának és a szövetek szerkezetének vizsgálatára széles körben
használják a mikroszkópos felvételek elemzését. Amennyiben egy adott kezelés által kiváltott
morfológiai változásokat szeretnénk vizsgálni, akkor a sejteket nem elég megjeleníteni. Ekkor
a kezelés hatására bekövetkező változások kvantitatív elemzésére van szükség. Néhány kutatás
keretein belül foglalkoztak olyan automatizált képelemző módszerek kifejlesztésével, melyek
segítik a celluláris változások monitorozását, valamint megkönnyítik a sejtek különböző
tulajdonságainak meghatározását, így nagyban felgyorsítják a munkafolyamatot [7-8]. A
képelemzés egy olyan módszer, melyet a képeken lévő releváns információ kiemelésére,
kinyerésére használnak [12]. A tudomány és az ipar számos területén alkalmazzák, ilyen például
az orvostudomány, orvosbiológia, mikroszkópia, űrkutatás, geográfia, ipari automatizálás és a
robotika is. A képek elemzésére használt módszerek jelentősen eltérnek az alkalmazástól
függően. Mikroszkópos felvételek esetén általában a képen található objektumok lehető
legpontosabb meghatározása, számlálása, általuk elfoglalt terület vagy egyéb paraméter
kiszámítása, esetleg egyes objektumok követése a cél. Általánosságban ennek főbb lépései a
képek javítása, majd azok szegmentálása. Előbbi a feladat szempontjából lényegtelen, vagy akár
zavaró információ csökkentésére, illetve a lényeges információ kihangsúlyozására, utóbbi pedig
a hasonló tulajdonságokkal rendelkező régiók kiemelésére szolgál [11].
A manuális módon történő sejtszámlálás és az egyes sejtek különböző paramétereinek
meghatározása igen időigényes és monoton feladat főleg, ha nagyobb mennyiségű felvétel áll
rendelkezésünkre. Ez indokolttá teszi egy olyan algoritmus implementálását, amely segíti és
gyorsítja ezt a folyamatot, ezáltal megkönnyíti a kutatók mindennapos munkáját.
Szakdolgozatom célja a mikroglia aktiváció és a krónikus stressz kapcsolatának
vizsgálata, valamint egy olyan algoritmus implementálása, mely automatikusan detektálja és
elemzi a mikroglia sejtekről készített fénymikroszkópos felvételeket. További célom egy olyan
interaktív grafikus felhasználói felület létrehozása, melynek segítségével a felhasználó
módosíthat a detektált sejteken (hozzáadhat, illetve törölhet közülük), ezzel pontosítva a
végeredményt.

7
A munkám során elvégzendő feladatok a következők voltak:
mikroglia sejtek morfológiai és funkcionális szempontok alapján történő osztályozása,
valamint a morfológia és funkció közötti összefüggés meghatározása
mikroglia sejtek megjelenítése fénymikroszkópiával
mikroglia sejtek automatikus detektálására, valamint nyúlványhálózatuk modellezésére
alkalmas program kifejlesztése
a sejtek különböző paramétereinek meghatározása az implementált algoritmus
segítségével
krónikus stresszen átesett patkányok különböző agyterületein a mikroglia sejtek
elemzése, morfológiai változásainak vizsgálata
A fénymikroszkópos felvételek készítését, és az ezt megelőző munkálatokat a Magyar
Tudományos Akadémia Kísérleti Orvostudományi Kutatóintézet (MTA-KOKI) Molekuláris
Neuroendokrinológia csoportjánál végeztem. A kutatócsoport jelenleg az ImageJ nevű szoftvert
használja a fénymikroszkópos felvételek elemzésére. Az általuk alkalmazott módszer nem
biztosít automatizált elemzést, így munkám során szerettem volna létrehozni egy olyan eszközt,
ami segítheti későbbi munkájukat.

8
2. Irodalmi áttekintés
2.1. A mikroglia sejtek funkciója és morfológiája
A mikroglia sejtek a központi idegrendszer immunsejtjei, melyek a neuronoktól eltérően
mezodermális eredetűek [1]. Fontos szerepet játszanak a gyulladásos folyamatokban és
fertőzésekben, a szinaptikus plaszticitás szabályozásában [3], a neurogenezisben [2], a
sejttörmelék és egyéb anyagok extracelluláris térből történő eltakarításában [3]. Élettani
körülmények között a mikroglia sejtek nyugalmi, „kereső” állapotban vannak. Ekkor jellemzően
kiterjedt és dúsan elágazó nyúlványrendszerrel rendelkeznek, mellyel folyamatosan pásztázzák
környezetüket, így gyorsan tudnak reagálni a különböző káros behatásokra [2]. Nyúlványaik
kiterjesztésével majd visszahúzásával mintavételezik mikrokörnyezetüket és fenntartják a
homeosztázist [3]. Modern, in vivo képalkotó módszerekkel kimutatták, hogy a motilis
mikroglia nyúlványok kapcsolatot hoznak létre az idegsejtek dendritjeivel [2], így a mikroglia
sejtek aktívan közreműködhetnek az információ feldolgozásában az agyban.
Patológiás változások következtében a mikroglia sejtek gyorsan aktiválódnak, mely
során a sejtek morfológiai változásokon mennek keresztül [2][39]. Ezen változás
eredményeképp először rövid nyúlványú, úgynevezett reaktív mikrogliává, szélsőséges esetben
pedig nyúlványokkal nem rendelkező, amőboid, fagocitáló mikrogliává alakulnak. A folyamatra
jellemző továbbá, hogy a sejt a sérülés irányába vándorol [39]. Az aktivációt különböző
extracelluláris jelek indukálhatják, például sejtpusztulás következtében kibocsátott anyagok,
kórokozók vagy sérülés. Az aktivációs folyamat nem csak morfológiai változásokkal jár, hanem
előidézi az intracelluláris jelátviteli és génexpressziós folyamatok megváltozását is, melyek
következtében a sejtek gyulladáskeltő vagy csökkentő citokineket termelhetnek [1][3].
A mikrokörnyezetükből érkező különböző ingerek hatására változás mutatkozik a sejtek
citokin-, receptor- és más egyéb markereinek expressziójában, mely alapján a mikroglia sejteket
különböző funkcionális fenotípus kategóriákba sorolhatjuk. A mikroglia esetében a
makrofágokhoz hasonlóan kétféle aktivált fenotípust különböztet meg az irodalom, melyek a
klasszikusan aktivált M1-es és az alternatívan aktivált M2-es típus [32]. Az M1 típusú
mikrogliára fokozott proinflammatorikus citokin (például interleukin-1β és tumor nekrózis
faktor-α) és szabadgyök (például nitrogén-monoxid) termelés jellemző, így ez a típus szerepet
játszhat a gyulladás következtében fellépő szöveti károsodásban [13][32]. Ezen változások
mellett a klasszikusan aktivált sejtek morfológiája is jelentősen megváltozik. A gyulladásos
válaszokban résztvevő mikroglia esetében megfigyelték, hogy a vékony nyúlványok
fokozatosan visszahúzódnak, megvastagodnak, majd végül a sejt amőboid formát vesz fel [13].
Az M2-es fenotípusú mikroglia antiinflammatorikus citokineket (például interleukin-10) és

9
növekedési faktorokat termel, valamint fokozott fagocitotikus aktivitás jellemzi. Ez a típus részt
vehet a sérülések utáni regenerációban és a sejttörmelék eltakarításában [32].
Összességében, a különböző környezeti változásokra reagálva a mikroglia sejtek
jelentős fenotípusos plaszticitást mutatnak mind morfológiai, mind funkcionális szempontból,
valamint szerepet játszhatnak neurotoxikus és neuroprotektív folyamatokban egyaránt. A
morfológiai változásokból viszont sajnos nem lehet egyértelműen következtetni a sejtek
expressziós profiljára, vagyis funkcionális orientációjára [3][39].
1. ábra: Mikroglia aktiváció fokális központi idegrendszeri sérülést követően [40].
Nekrotikus sejthalál következtében a károsodott szövet és az azt körülvevő asztrociták ATP-t
bocsátanak ki, mely specifikus, mikroglia sejtek által expresszált receptorokat aktivál. Az ábrán
jelölt P2X4R és P2Y12R receptorok aktív állapotban a sérülés felé lévő nyúlványok sérülés
irányába történő kiterjesztését, illetve a többi nyúlvány visszahúzását indukálják. Ezt követően,
a P2Y6R receptor közvetítésével a sejt egyetlen fagocitotikus nyúlványt alakít ki, ami végül
egybeolvad a sejttesttel. Az így kialakult fagocita részt vesz a sejttörmelék extracelluláris térből
történő eltakarításában.

10
2. ábra: A mikroglia sejtek változatos morfológiájának sematikus ábrázolása [41].
Nyugalmi állapotban (A) a sejtekre hosszú és elágazó nyúlványok jellemzők. Enyhén aktivált
állapotban (B) a nyúlványrendszer elágazódása fokozódik, míg közepesen aktivált/reaktív
állapotban (C) megnagyobbodott sejttest, rövidebb és vastagabb (visszahúzódó) nyúlványok
figyelhetők meg. Az amőboid állapotra (D) lekerekített forma, kevés nyúlvány, vagy a
nyúlványok hiánya, valamint nagy proinflammatorikus aktivitás jellemző.
2.2. Stresszválaszt befolyásoló agyi régiók és hormonok
A stressz egy olyan faktor, mely befolyásolni tudja az immunrendszer reakciókészségét,
tartós jelenléte esetén jelentősen fokozhatja a gyulladásos folyamatokat és meghosszabbítja
azok fennállását [4-5], valamint a különösen nagy stresszel járó események a depresszió
elsődleges rizikófaktorai [13]. Felismerték már, hogy stressz hatására számos fiziológiás és
patológiás változás következik be mind a periférián, mind a központi idegrendszer szintjén.
A krónikus stressz okozhat idegi károsodást, illetve negatívan befolyásolhatja a
neurogenezist és a plaszticitást [4]. A stressz központi idegrendszerre gyakorolt hatásai
nagyrészt a keringési rendszerben jelenlévő kortikoszteroidoknak tulajdoníthatók, melyek a
hipotalamusz-hipofízis-mellékvesekéreg tengely stressz által történő aktiválódása során
szabadulnak fel, majd a vér-agy gáton átjutva, specifikus receptorokon keresztül fejtik ki az
idegsejtekre gyakorolt hatásukat [4]. A mellékvesekéreg az emlősök egyik fő stressz szerve,
mely közvetítő szerepet játszik a külső és belső stresszorok által kiváltott stresszreakcióban
glükokortikoidok, például rágcsálók esetében kortikoszteron termelésével. A mellékvesekéreg
kortikoszteroid szintézisének és szekréciójának fő szabályozója az adrenokortikotrophormon
(ACTH) [28]. Az ACTH egy peptidhormon, mely az adenohipofízesben képződik.
Termelődését a hipotalamuszban lévő paraventrikuláris mag parvocelluláris magcsoportja által
termelt kortikotropin felszabadító hormon stimulálja [29], tehát a hipotalamusz-hipofízis-
mellékvesekéreg tengely hormontermelésének egyik fő szabályozója a paraventrikuláris mag,

11
mely stressz hatására serkenti az ACTH szekréciót. Az ACTH stimulálja a kortikoszteron
szintézist, mely végül negatív visszacsatolás révén visszahat a rendszerre [29].
A stresszválaszban további fontos résztvevő a szimpatikus idegrendszer is, mely stressz hatására
adrenalint és noradrenalint szabadít fel. Ezek fiziológiás hatása igen gyors, jelentős szívritmus
emelkedést, perifériás vazokonstrikciót, vérnyomás emelkedést és energia mobilizációt
eredményeznek [13].
Ezen kívül jól ismert, hogy a stressz hatással van a szervezet metabolizmusára, végső
soron befolyásolja a testtömeg változását [4]. A hipotalamuszban található nucleus arcuatus a
táplálékfelvétel és az energia felhasználás egyik fontos központi szabályozó régiója, valamint a
perifériáról érkező metabolikus és humorális szignálok (metabolitok, leptin, inzulin) szenzora
[6][31]. A nucleus aruatusban található neuronok másodlagos neuronoknak továbbítják az
információt, mint például a paraventrikuláris mag parvocellularis sejtjeinek [31], így
befolyásolják a szervezetet érintő stresszre adott neuroendokrin és autonóm választ.
2.3. Mikroglia sejtek kapcsolata a gyulladásos és a stresszel járó
folyamatokkal
Az agyban fellépő gyulladásos reakciók, melyek igazolhatók a mikroglia sejtek
tulajdonságainak változásával a neurodegeneratív betegségek közös jellemzői [32]. Számos
tanulmány vizsgálta már a mikroglia sejteket, mint lehetséges mediátorokat a gyulladással járó
idegi kórfolyamatokban, melyek kimutatták, hogy több neurodegeneratív megbetegedés,
például a Parkinson kór, az Alzheimer kór, a Huntington kór és a sclerosis multiplex esetén is
megfigyelhető kóros mikroglia aktiváció [4]. Kimutatták, hogy az akut fizikai illetve érzelmi
stressz jelentős hatással van mikroglia morfológiai aktiválására a hipotalamusz és hippocampus
területén [4-5], és az aktiváció mértéke arányos a stressz időtartamával [4]. Ezen kívül ismert,
hogy az akut stressz jelentősen növelheti a proinflammatorikus citokinek szintjét a patkányok
agyában, melyek egyik fő forrása a mikroglia [13].
Egy másik kutatásban [5], melyben a krónikus stressz hatását vizsgálták patkányokon arra
jutottak, hogy a krónikus stressz szelektíven növeli a mikroglia sejtek számát bizonyos stressz-
érzékeny agyi területeken, és a nyugalmi állapotban lévő mikroglia sejtek átalakulását indukálja.
Összesen 15 agyterületet vizsgáltak, melyből 9 esetben mutatkozott szignifikáns különbség a
kísérleti és kontroll csoport között. Ebből azt a következtetést vonták le, hogy a mikroglia
aktiváció fontos szerepet játszhat a stresszválasz szabályozásában és a stresszhez való
alkalmazkodásban. Kreisel és mtsai [10] a mikroglia aktiváció dinamikus változásának szerepét
vizsgálták krónikus stressz indukálta depressziószerű állapot kialakulásában, rágcsálókon.
Kutatásuk eredményeképp azt kapták, hogy a mikroglia sejtek működésének zavara etiológiai
szerepet játszik a stressz indukálta depresszióban. Ebből azt a következtetést vonták le, hogy a

12
mikroglia stimulátorok gyorsan ható antidepresszánsként szolgálhatnak a depresszív és stresszel
kapcsolatos állapotok egyes formáiban.
2.4. Mikroglia sejtek különböző paramétereinek meghatározása
képelemző eljárásokkal
Az automatizált képelemző eljárások előnye a manuális módszerekkel szemben, hogy
nagy adathalmazok gyors és objektív elemzését teszik lehetővé. A továbbiakban ismertetem
néhány, a szakirodalomban fellelhető módszer áttekintését. A tanulmányok többségében a
sejteket egy mikroglia-specifikus markerrel, az ionizált kálciumkötő adapter molekulával (Iba-
1) detektálták [5][7-9], de volt, hogy genetikai manipuláció következtében, specifikusan a
mikroglia sejtekben expresszálódó zöld fluoreszcens fehérjét (GFP) használtak [10]. Az
elemzésre szánt felvételeket a fehérje jelölés módjától függően konfokális [7-8][10] vagy
fénymikroszkóppal [4][9] készítették.
Sugama és mtsai. [4] intenzitásérték-alapú szegmentálást alkalmaztak. Minden olyan
pixelt, aminek értéke nem haladta meg a küszöbértéket a sejthez rendeltek, míg ellenkező
esetben a háttérhez. A küszöbértéket minden egyes sejthez manuálisan állították be. Az
eredmény egy bináris kép lett, melyből a WinROOF képelemző szoftver segítségével
kiszámították a sejt felszínét. Tynan és mtsai. [5] szintén az intenzitásértékeket vették alapul a
felvételek szegmentálásához, majd méretalapú szűrést is alkalmaztak a pontosabb eredmény
érdekében. Meghatározták a sejtek számát, valamint az Iba-1 pozitív területek százalékos
arányát a vizsgált régióban. A sejtek méretének becslésére a sejtek által elfoglalt összterületet
elosztották a sejtek számával.
Zhang és mtsai [7] a retinában található mikroglia sejtek térbeli jellemzőit vizsgálták
képfeldolgozó eljárásokkal. Kifejlesztettek egy olyan MATLAB-alapú módszert, melynek fő
építőelemei a képjavítás és szegmentálás, a morfológiai alapú sejtfelismerés és a területszámítás.
A feldolgozás során logaritmikus képjavítást és küszöbérték-alapú szegmentálást alkalmaztak.
Kozlowski és Weimer [8] szintén MATLAB-alapú módszert implementált a mikroglia sejtek
morfológiájának automatikus meghatározására. A szegmentáció első lépéseként lokális
maximumkereső algoritmussal meghatározták a sejtek pozícióját. A sejtmaszkokat az egyes
pozíciók meghatározott környezetében számított iteratív lokális küszöbérték alapján hozták
létre. A sejtenkénti morfológiai elemzéshez lokális iteratív küszöbérték-alapú szegmentálással
különítették el a sejteket a háttértől. Az így kapott sejtek alakjának jellemzéséhez a sejtek
kerületét, kiterjedését, excentricitását és a sejttest méretét vették alapul. Belátták, hogy az olyan
marker segítségével történő elemzés, mely intenzitása változik a mikroglia aktivációs állapotától

13
függően külön kihívást rejt magában, mivel az aktivált sejtek így könnyebben detektálhatók,
mint a nyugalmi állapotban lévők. Ilyen marker például az Iba-1 is.
A következő tanulmányban [9] patkányok prefrontális lebenyében lévő mikroglia sejteket
vizsgáltak NeuroExplorer és MATLAB szoftverek segítségével. Meghatározták a sejttest
kerületét, az elsődleges nyúlványok számát, az elágazások számát, a nyúlványok hosszát,
valamint azt a területet, mely magában foglalja az egész sejtet. Utóbbit abból a konvex
sokszögből határozták meg, melyet a sejt legtávolabbra nyúló nyúlványaira illesztve kaptak.
Egy másik tanulmányban Kriesel és mtsai. [10] a sejtek számát és a sejttestek által elfoglalt
területet Nikon Imaging Elements Software segítségével automatikusan határozták meg. A
nyúlványok hosszának kiszámításához az egyes nyúlványokat manuálisan követték nyomon, és
jelölték.
2.5. Képek szegmentálására alkalmazott módszerek áttekintése
A képek szegmentálása fontos, és gyakran alapvető lépés a képelemzés, objektum
reprezentáció és sok más képfeldolgozási feladat tekintetében is. Ez alatt azt a folyamatot értjük,
melynek során felosztjuk a képet több, az elvégzendő feladat szempontjából érdekes részre.
Főként a felvételen található objektumok kinyerésére használják. Számos szegmentáló eljárást
fejlesztettek ki az elmúlt időszakban, melyeket a következő főbb kategóriákba sorolhatjuk [12]:
1. Küszöbérték alapú szegmentáció
2. Régió alapú szegmentáció
3. Él alapú szegmentáció
4. Klaszterezés
5. Illesztés (matching)
1. Küszöbérték alapú szegmentáció
A küszöbölés egy gyakran használt, nagyon egyszerű és kis számítási teljesítményt igénylő
szegmentációs eljárás, melynek során többnyire az intenzitásértékeket veszik alapul. Ennél
a módszernél feltesszük, hogy a kép egyes részei szétválaszthatók az intenzitásértékeik
alapján, tehát létezik egy t érték, melynél kisebb értékű pixelek a háttérhez (0), nagyobb
értékű pixelek pedig az objektumokhoz (1) tartoznak [7][11] világos előtér objektumokat
feltételezve:
𝑦(𝑛1, 𝑛2) = { 0 𝑥(𝑛1, 𝑛2) ≤ 𝑡
1 𝑥(𝑛1, 𝑛2) > 𝑡
Az optimális küszöbérték, vagyis t meghatározása több módon történhet. Az egyik ilyen az
Otsu által kifejlesztett módszer alkalmazása, mely automatikusan meghatározza az optimális

14
globális küszöbértéket az intenzitásosztályok közötti variancia maximalizálásával [7]. A kép
felosztásához továbbá hisztogram küszöbölést illetve vágást szoktak használni.
Ez a technika jól alkalmazható olyan képekre, melyek sötétebb háttéren világos
objektumokat tartalmaznak [12] (vagy fordítva), de sajnos sok esetben egy sima küszöbölés
nem elegendő a kívánt cél eléréséhez, például a kép egyenetlen megvilágítása miatt [11]. A
munkám során vizsgált felvételeken a háttér és a sejttesteknek megfelelő régiók
intenzitástartománya jelentősen eltér, így ez a módszer alkalmazható.
Nem csak az intenzitásértékek alapján lehet küszöbölni. A szegmentáció eredményeképp
kapott bináris képen lehet például az objektumok mérete alapján történő küszöböléssel
finomítani az eredményen.
2. Régió alapú szegmentáció
A régió alapú szegmentálás két alapvető művelettel dolgozik, a terület-szétválasztással és a
területegyesítéssel. Mindkét esetben definiálni kell egy hasonlósági kritériumot, mely
alapján az alkalmazott algoritmus eldönti, hogy mit csináljon az aktuális régióval (például
szétválassza a régiót vagy sem) [12]. Ez lehet akár textúra vagy színbéli hasonlóság, például
egyesíthetünk két régiót az alapján, hogy az intenzitásértékeik átlagának különbsége elér-e
egy előre meghatározott értéket [11].
3. Él alapú szegmentáció
Ennél a módszernél azzal a feltevéssel élünk, hogy a képen lévő élek az objektumok
körvonalait reprezentálják, ily módon alkalmasak azok azonosítására. A szegmentáció első
lépése általában egy olyan bináris kép létrehozása, ami csak az éleket tartalmazza [12].
Tekintsünk a képre egy függvényként, melynek értelmezési tartománya a pixelek halmaza,
értékkészlete pedig a felvehető intenzitásértékek halmaza (intenzitásfüggvény). Ekkor az
intenzitásértékekben bekövetkező változás mértékét és irányát a gradiens kiszámításával
tudjuk meghatározni [11]:
𝑱(𝑥, 𝑦) = ∇𝐼(𝑥, 𝑦) = (𝜕𝐼(𝑥, 𝑦)
𝜕𝑥,𝜕𝐼(𝑥, 𝑦)
𝜕𝑦) (𝑥, 𝑦).
Az élek a kép azon részei, ahol az intenzitás hirtelen változik, tehát azokat a pontokat
keressük, ahol a gradiens nagy.
Az élek detektálása után a következő lépés, hogy az észlelt élek közül kizárjuk azokat,
melyek nem zártak, majd a fennmaradó zárt éleket kitölthetjük az eredeti kép megfelelő
részeivel [12]. Így megkapjuk eredményül a szegmentált képet. Ez a módszer érzékeny a
zajra, és nem működik jól azokon a képeken, melyekre alacsony kontraszt, valamint az
objektumok és a háttér közötti sima átmenet jellemző. Mivel a vizsgált felvételek sok esetben

15
zajosak (például fedés közben keletkezett buborékok), jellemző rájuk a sima átmenet és az
alacsony kontraszt, ezért nem ezt a módszert alkalmaztam a munkám során.
4. Klaszterezés
A klaszterező módszerek valamilyen szempontból hasonló minták csoportosítására
hivatottak. Az egyik alapvető módszer a K-közép klaszterezés, melynek célja a kép K darab
szegmensre történő felosztása a klaszteren belüli variancia minimalizálásával. Hátránya,
hogy a klaszterek számát (K) előre meg kell adni [12].
A másik elterjedt módszer az úgynevezett mean-shift algoritmus, ami egy nem paraméteres
iteratív klaszterezési technika, mely egy ismeretlen sűrűségfüggvényből származó mintákkal
modellezi az egyes pixelekhez tartozó jellemzővektorokat (feature vector), majd megpróbál
klasztereket találni az így kapott eloszlásban [11].
5. Illesztés
Ha van előzetes információnk arról, hogy az objektum, amit azonosítani szeretnénk a
képeken hogyan néz ki, akkor ezt felhasználhatjuk a szegmentálás során. Ehhez szükséges
egy sablon, ami jól reprezentálja az adott objektumot. A sablont a kép különböző részeihez
hasonlítjuk mindaddig, amíg meg nem találjuk a kívánt objektumot [12]. Az általunk vizsgált
sejtek alakja elég változatos, így nem állt rendelkezésemre olyan általános sablon, ami
megfelelően reprezentálná a sejtek különböző állapotait, így munkám során nem
alkalmaztam ezt a módszert.
2.6. Sejtek szegmentálására használt képfeldolgozási technikák
A mikroszkópos képek szegmentálásának célja a sejtmagok, sejtek vagy szöveti
komponensek elkülönítése a kép többi részétől [14]. A sejteket ábrázoló képek automatizált
szegmentálása általában nagyobb kihívást jelent, egyrészt a sejtekre jellemző tulajdonságok
(például alak, méret, textúra és sejtsűrűség) nagy változatossága, másrészt a különböző festési
eljárások és mikroszkópok miatt [15].
Az intenzitás alapú küszöbölés a legrégebbi, és a mai napig az egyik leggyakrabban
használt módszer [14-15], mivel a festési eljárások eredményeképp a sejtek jelentősen eltérő
intenzitással rendelkeznek, mint a háttér. Ez a módszer önmagában általában nem szolgáltat jó
eredményt, ezért sok esetben ez csak a szegmentáció első lépése.
Elterjedt módszer továbbá a különböző morfológiai operátorok, mint például az erózió és
dilatáció alkalmazása mind elő, mind utófeldolgozási lépésként. Ezen kívül gyakran
alkalmazzák a watershed transzformációt és az aktív kontúr módszert is. Tekintsünk a képre egy
elárasztott topografikus térképként, ahol az élek jelzik a kiemelkedéseket. A watershed

16
transzformáció megtalálja a képen lévő vízgyűjtő medencéket (vagyis objektumokat), melyeket
vízválasztó vonalak választanak el egymástól. Ezen vonalak összessége adja a watershed
vonalat. Az elárasztás forrásaként szolgáló jelzőpontokat definiálhatjuk a gradiens lokális
minimumaiként, ám ez gyakran túlszegmentáláshoz vezet [11][20][26]. A jelzőpontokat
meghatározhatjuk más módszerekkel is, például morfológiai operátorok alkalmazásával [20].
Az aktív kontúr módszer igen ígéretes technikának bizonyult, mellyel jó eredményeket lehet
elérni, de ugyanakkor kihívást jelenthet a megfelelő kezdeti állapot megválasztása [14].

17
3. Módszerek
Ebben a fejezetben bemutatom azokat a módszereket, melyeket munkám során
alkalmaztam. Ezek három nagy csoportra bonthatók, melyek a következők: (1) kísérleti állatok
stresszelése, melynek célja változás indukálása a mikroglia sejtekben, (2) kísérleti állatok
feldolgozása, mely magában foglalja az állatok agyának előkészítését, a mikroglia sejtek
kimutatását és fénymikroszkóppal történő megjelenítését, (3) fénymikroszkópos felvételek
elemzése, melynek célja a mikroglia sejtek kvantitatív vizsgálata, a stressz kísérlet hatására
esetlegesen bekövetkező változások kimutatása.
3.1. Kísérleti állatok és kísérleti elrendezés
A stressz kísérletben összesen 55 Wistar patkányt (kísérleti csoport: n = 31 ♂, kontroll
csoport: n = 24 ♂) vizsgáltunk. A kísérleti csoport egyedein posztnatálisan anyai szeparációt
végeztünk, vagyis születésüktől számított 2 héten keresztül napi 3 órára elválasztottuk az
anyjuktól őket. Ily módon ezek az egyedek korai, úgynevezett „early life” stressznek lettek
kitéve. A korai stressz után az állatokat csoportosan helyeztük el, és 7 hetes (felnőtt) koruktól
kezdve, 3 héten keresztül, napi két alkalommal különböző enyhe stresszhatásoknak tettük ki
őket. A stresszorokat a kísérlet tervezésekor random módon válogattuk. Ezt a procedúrát
krónikus variábilis stressznek (chronic variable stress, CVS) nevezi az irodalom. Az általunk
alkalmazott krónikus variábilis stressz protokollt az 1. táblázat szemlélteti.
Az állatok testtömeg gyarapodásának követése céljából a CVS előtt és után is
megmértük testtömegüket. A kísérleti és a kontroll csoport állatait feldolgozásuk módja alapján
további két alcsoportra bontottuk. Az első csoport (kísérleti csoport: n = 8 ♂, kontroll csoport:
n = 8 ♂) metabolikus paramétereit (például légzési hányados, energia leadás) a kísérlet 18.
napjától kezdve 3 napon keresztül indirekt kalorimetriával felvételeztük. A metabolikus mérés
végeztével ezen állatokat perfundáltuk, agyukat feldolgoztuk.
A másik csoport (kísérleti csoport: n = 23 ♂, kontroll csoport: n = 16 ♂) egyedein a kísérlet
végeztével viselkedés teszteket, például cukor preferencia tesztet végeztünk. A cukor
preferencia teszt során a kísérleti állatok 24 órán keresztül szabadon választhattak két palack
közül. Az egyik 1%-os szacharóz oldatot, a másik sima csapvizet tartalmazott. A két palackot
felcseréltük 12 óra letelte után. Az állatok folyadék fogyasztását a palackok tömegének
mérésével követtük. A cukor preferencia kiszámításánál az elfogyasztott szacharóz oldat
százalékos arányát viszonyítottuk a teljes elfogyasztott folyadékmennyiséghez. Végül
dekapitáltuk a csoport egyedeit, és vért vettünk tőlük hormonvizsgálat céljából. A levett
vérmintákat egyből centrifugáltuk és az így kapott vérplazmát felhasználásig -20°C-on tároltuk.
A plazma ACTH és kortikoszteron koncentrációját direkt radioimmunoassay segítségével

18
határoztuk meg. Ez a specifikus eljárás antigén-antitest reakción alapszik, segítségével
meghatározható kis koncentrációjú, antigén sajátosságot mutató molekulák mennyisége. A
továbbiakban az általam részletesebben vizsgált, első csoporton alkalmazott módszereket fogom
ismertetni.
Nap Délelőtt Délután
1. cukor preferencia teszt világosság és nedves alom, 1 éjszaka
2. macska vizelet szaganyag, 15 p elektromos sokk (0,5 mA 30 mp-enként),
5 p
3. alom nélküli izoláció zsúfoltság alomtársakkal
4. mozgáskorlátozás, 30 p tiszta ketrec, alja 4°C-ra hűtve
5. stroboszkóp zsúfoltság alomtársakkal
6. predátor szaganyag (róka) és izoláció,
15 p sötétség és izoláció, 1 éjszaka
7. x szociális elkerülés
8. elektromos sokk (0,5 mA 30 mp-enként),
5 p világosság
9. alom nélküli izoláció dőlt ketrec és nedves alom
10. mozgáskorlátozás, 30 p világosság, 1 éjszaka
11. dőlt ketrec szociális kudarc
12. stroboszkóp világosság, 1 éjszaka
13. sötétség sötétség
14. mozgáskorlátozás, 30 p nedves alom, 1 éjszaka
15. elektromos sokk (0,5 mA 30 mp-enként),
5 p alom nélküli izoláció és világosság
16. zsúfoltság és sötétség, 4 ó stroboszkóp
17. tiszta ketrec, alja 4°C-ra hűtve predátor szaganyag (róka), 15 p
18. zsúfoltság dőlt ketrec és nedves alom
19. stroboszkóp világosság, 1 éjszaka
20. elektromos sokk (0,5 mA 30 mp-enként),
5 p izoláció
21. kényszerített úszás teszt cukor preferencia teszt
22. cukor preferencia teszt megemelt keresztpalló teszt
23. x stroboszkóp
1. táblázat: CVS protokoll összefoglalása naponkénti bontásban

19
3.2. Perfúzió és szövetfeldolgozás
A metabolikus mérést követően az első csoport állatainak túlaltatása (200 mg/ttkg
nembutállal), majd transzkardiális perfúziója következett először fiziológiás sóoldattal (0.9%
NaCl), majd 70 ml jéghideg fixálóval (4% paraformaldehid, 0.4 M foszfát pufferben, pH:7). A
perfúzió célja a vérerek kimosása és a szövet fixálása, mely közvetlenül az állat saját keringési
rendszeren keresztül végzett, úgynevezett belső fixálással valósul meg. Előnye, hogy a fixáló
folyadék a kapilláris hálózaton keresztül eljut mindenhova, így a fixálás a célszerv teljes
területén szinte egyszerre kezdődik. További előnye, hogy a szervet/szöveteket még azelőtt
rögzítjük, hogy az állatból eltávolítottuk volna, ezzel nagymértékben csökkentve az esetlegesen
fellépő mechanikai roncsolódást vagy számottevő bomlást [38].
A perfúzió után a koponyából eltávolított agyat 10% szacharózt tartalmazó fixáló folyadékban
tároltuk 3 órán keresztül (poszt-fixálás), majd 10% szacharózt tartalmazó 0.1M KPBS-ben
(kálium-foszfát puffer, pH7.4) tároltuk egy éjszakán át 4oC-on. Ezt követően 25µm-es koronális
metszeteket készítettünk fagyasztó mikrotomon, melyeket felhasználásig fagyálló folyadékban,
-20oC-on tároltunk. Ezután a metszetek immunhisztokémiai festése következett.
Az immunhisztokémiai festés egy olyan általános biokémiai eljárás, mellyel szöveti
antigéneket lehet kimutatni egy adott mintában specifikus ellenanyag segítségével. Ez történhet
enzimreakció következményeképp kiváló csapadék vagy fluorofórok segítségével. Előbbi
esetben az eredmény fénymikroszkóppal, míg utóbbi esetben floureszcens mikroszkóppal
vizsgálható [38]. Az immunhisztokémiai eljárás során lezajló reakció sematikus ábrázolása a 3.
ábrán látható.
Az Iba-1 immunhisztokémiai eljáráslépései az alábbiak voltak:
1. KPBS-el történő mosás 4x5 percig a fagyálló metszetekből történő kimosása céljából
2. Endogén peroxidázok gátlása, 0,3 %-os H2O2 (hidrogénperoxid) oldattal történő mosás
5 percig
3. KPBS-el történő mosás 4x5 percig a H2O2 oldat metszetből történő kimosása céljából
4. Blokkoló szérum (2% normál kecske szérum) 1 óra
5. Primer antitestet tartalmazó oldatban (nyúlban termeltetett anti Iba-1, 1:1000
hígításban) történő inkubálás 4°C-on egy éjszakán át
6. KPBS-el történő mosás 4x5 percig
7. Szekunder antitestet (kecskében termeltetett biotinilált anti-nyúl IgG, Vector,
Burlingame, CA, USA) tartalmazó oldatban történő inkubálás 1 órán keresztül
8. KPBS-el történő mosás 4x5 percig
9. Avidin-biotin-enzim (HRP, torma peroxidáz) komplexet (Elite Plus kit, Vector)
tartalmazó oldatban történő inkubálás 1 órán keresztül

20
10. KPBS-el történő mosás 4x5 percig
11. Na-acetátos (0,1M) mosás 2x10 percig az ideális pH beállítása céljából
12. Előhívás (7,5 ml DV (desztillált víz) + 15 mg DAB (3,3´- diaminobenzidin) + 7,5 ml
NAS (nikkel ammónium szulfát) + 225 µl 0.3%-os H2O2 (500 µl DV-be 5 µl 30%-os))
13. Na-acetátos (0,1M) mosás 2x10 percig
14. KPBS-el történő mosás 2x10 percig
15. Festett metszetek felhúzása zselatinos tárgylemezre
16. Dehidrálás felszálló alkoholsorban
17. Depex fedőanyaggal történő fedés
Az ily módon megfestett metszetekről fénymikroszkópos felvételeket készítettünk húszszoros
nagyításban Nikon Eclispe 6000 mikroszkóp és Spot RT színes digitális kamera (Diagnostic
Instruments Inc., IL, USA) segítségével. Dolgozatomban a hipotalamuszban található nucleus
paraventricularisról és a nucleus arcuatusról készített felvételeket vizsgáltam, állatonként
átlagosan 3-3 metszeten bilaterálisan. Ezen agyterületek sematikus ábrázolása a 4. ábrán látható.
A metszeteket patkány agyatlasz [30] alapján válogattuk.
3. ábra: Az immunhisztokémiai eljárás során lezajló reakció sematikus ábrázolása [42]. A
vizsgált fehérje (esetünkben Iba-1) más fajba beoltva antigénként viselkedik, tehát indukálja a
rá specifikus antitestek termelődését. Az antitestekben gazdag vérplazmából készült primer
ellenanyagot használhatjuk az Iba-1 kimutatására. Az avidin-biotin komplex módszer esetén
biotinilált szekunder (primer ellenanyagot specifikusan felismerni képes) antitestek kapcsolják
Biotin
Avidin
Riporter enzim
Szöveti antigén
Primer antitest
Biotinilált szekunder antitest

21
össze a primer antitesteket az avidin-biotin-peroxidáz komplexekkel. Színtelen szubsztrát
hozzáadása után az enzimreakció lezajlik, mely során oldhatatlan, színes csapadék képződik.
4. ábra: A dolgozatban vizsgált agyterületek sematikus ábrázolása [43]. Az ábra bal oldalán
lévő metszeten a nucleus paraventrikularis (PVN), míg a jobb oldalán lévőn a nucleus arcuatus
(ARC) látható, melyeket pirossal jelöltem.
3.3. Felvételek elemzésére kidolgozott módszer bemutatása
Az elkészült felvételeket MATLAB (R2017a) szoftver, és annak képfeldolgozó
eszköztára segítségével elemeztem. A képek eredeti mérete 1600x1200 pixel volt, de mivel
olyan régiókat is tartalmaztak, melyek nem az általam vizsgált területekhez tartoztak, ezért
kivágtam belőlük a vizsgálni kívánt részletet. A kidolgozott eljárás három főbb lépésből áll,
melyek a következők: (1) sejtek detektálása, (2) a sejtekhez tartozó nyúlványrendszer
modellezése, (3) a detektált mikroglia sejtek különböző paramétereinek meghatározása a stressz
kísérlet hatására esetlegesen bekövetkező változások kimutatása céljából. A 16 kísérleti állatból
összesen 178 felvételt készítettünk, ezért létrehoztam egy 30 darab, random módon válogatott
képből álló tesztadatbázist, melyen a különböző kipróbált módszereket teszteltem. Azt a
módszert, mely jó eredményt adott a tesztadatbázison kipróbáltam a teljes adatbázisra lefuttatva
is, majd a legjobb eredményt adó eljárás mellett döntöttem.
3.3.1. Sejtek detektálása
A sejtek detektálására olyan módszerek használhatók, melyek azok alapvető jellemzőit
képesek hatékonyan kiemelni. Mivel a szürkeárnyalatos képek esetén hiányzik a
színinformáció, így a sejteket leginkább az őket leíró, a háttértől különböző intenzitásértékekkel
és a belső textúrákkal tudjuk jellemezni. Ez a korlátozott jellemzőkészlet leszűkíti az
alkalmazható eljárások skáláját is, így a munkám során az intenzitás alapú küszöbölő és régió

22
alapú eljárásokkal, illetve textúra alapú csoportosítással próbálkoztam. Sejtszegmentálás esetén
elterjedt még a sejtek alakjára vonatkozó alaki megkötésekkel dolgozó eljárások alkalmazása
[23], ezek azonban a mikroglia sejtek rendkívül változatos nyúlványrendszere miatt nem
jöhettek szóba. Mivel a sejtekhez tartozó nyúlványrendszer pontos detektálása kulcsfontosságú
a stressz hatására esetlegesen bekövetkező változások követése céljából, így a nyúlványok
modellezésére régió alapú aktív kontúr módszert használtam, mely képes a nagy alaki
változatosság kezelésére.
A különböző módszerek tesztelése során megpróbáltam a képekre a [22]-ben leírt
textúra alapú eljárást alkalmazni. Ez a módszer kisebb egységekre, úgynevezett textúra
atomokra bontja, majd az atomok statisztikai megkülönböztethetősége alapján osztályozza a
képet. Az eljárás négy főbb szakaszra bontható, melyek sorrendben a következők: rotáció
invariáns, szomszédság alapú textúra ábrázolás; ritka textúrák modellezése textúra atom
tanuláson keresztül; az atomok statisztikai megkülönböztethetősége alapján egy teljes, súlyozott
gráf létrehozása, ahol a csúcsok a textúra atomok, az élek súlyai pedig az élek által összekötött
két atom statisztikai megkülönböztethetősége; a textúra atomok előfordulási valószínűsége és
statisztikai megkülönböztethetősége alapján feltűnőségi térkép (saliency map) kiszámítása.
Alapvetően a természetes képeken lévő kiugró régiók detektálására lett kifejlesztve, melyhez az
optimális atomszámot 20-nak találták. Mivel az általam vizsgált képeken jóval kevesebb típusú
textúra fordul elő, mint a természetes képeken, ezért az atomszámot csökkentve teszteltem a
módszert. Ez az eljárás ígéretesnek tűnt a képek egy részén, viszont több esetben egy osztályba
sorolta a sejtmagoknak megfelelő homogénebb régiókat a háttérrel (5. ábra). Mivel több
atomszám kipróbálásával sem sikerült olyan beállítást találni, mely a tesztképeken egységesen
működött volna, így elvetettem ezt a módszert.
Ezen kívül megpróbáltam a sejteket csupán aktív kontúr eljárás segítségével detektálni. Ennél a
módszernél fontos a megfelelő kiindulási állapot megválasztása, így ha nem rendelkezünk
előzetes információval az objektumok vélhető elhelyezkedéséről, az pontatlan eredményre
vezethet. A detekció kezdetén nem volt előzetes információm, ami alapján el tudtam volna
indítani az algoritmust, ezért egy négyzetrácsos maszkkal inicializáltam. Ez sajnos nem vezetett
jó eredményre az esetek egy részében (6. ábra), mivel az algoritmus minden egyes négyzetben
próbált valami struktúrát keresni és ez alapján meghatározni az előteret, így az eljárás
alkalmazása önmagában nem hatékony, szükséges hozzá egy előfeldolgozási lépésben
előállított kezdeti maszk, mely a sejtek pozíciójára jó közelítést ad.
A különböző eljárások kipróbálása során arra jutottam, hogy a sejtek textúra jellemzőjének
változatossága nem teszi lehetővé a textúra, mint önálló sejtjellemző alkalmazását a hatékony
szegmentálás eléréséhez, így a kidolgozott módszer a sejteket az intenzitásértékük alapján
különbözteti meg a háttértől. A következőkben ismertetem a választott módszert, mely alapját
logaritmikus transzformáció és többszintű, küszöbérték-alapú szegmentálás képzi.

23
5. ábra: A textúra alapú eljárás eredményének bemutatása 4-es atomszámmal. Az ábra A
és C részén egy-egy részlet látható két különböző képből, míg a B és D részén ezen képek
osztályozott változata szerepel. Látszik, hogy a második esetben (D-vel jelölt kép) a háttér és a
sejtmagoknak megfelelő régiók is feketével jelennek meg, tehát az algoritmus egy osztályba
sorolta őket, míg az első esetben (B-vel jelölt kép) a sejtmagokat külön osztályba sorolta.

24
6. ábra: Aktív kontúr módszerrel kapott eredmény négyzetrácsos maszkkal történő
inicializálás esetén. Az ábra A és C részén egy-egy részlet látható két különböző képből, míg
a B és D részén ezen képek szegmentált változata szerepel. Látható, hogy az első esetben (B
kép) megfelelő a szegmentálás eredménye, míg a második esetben (D kép) mintha felcserélné
az előteret és a hátteret a nem megfelelő inicializálás következtében.
3.3.1.1. Előfeldolgozás
A feldolgozás első lépéseként adaptív hisztogram kiegyenlítést alkalmaztam
(adapthisteq függvény). A hisztogram megadja, hogy egy képen összesen hány pixel veszi fel
az adott intenzitásértéket. A hisztogram kiegyenlítés célja a kontraszt növelése az
intenzitásértékek teljes dinamikatartományon történő egyenletes elosztásával. Az adaptív
módszer nem globálisan, hanem a képet kisebb részekre osztva, minden részre egymástól
függetlenül alkalmazza a hisztogram kiegyenlítést [11]. Ezeket a kisebb részeket csempének is
szokták hívni. Az adapthisteq függvény a [16]-ban leírt, kontraszt-limitált adaptív hisztogram
kiegyenlítést implementálja, melyet eredetileg orvosi képalkotáshoz fejlesztettek ki. Sima
adaptív hisztogram kiegyenlítés esetén sajnos számolni kellene a módszer jelentős hátrányával,
ami a zaj nagymértékű erősítése. A zaj amplifikáció csökkenthető a homogén területeken történő
kontrasztjavítás korlátozásával. Ezek a területek azonosíthatók egy-egy nagyobb kiugrással a
hisztogramban, mivel az őket alkotó pixelek közel azonos intenzitástartományba esnek. A
kontraszt-limitált módszer korlátozza az egyes csempék lokális hisztogramjaihoz tartozó
25
pixelek maximális számát, majd az így eltávolításra kerülő pixeleket egyenletesen szétosztja a
teljes hisztogramon. Végül, a szomszédos csempék egyesítése bilineáris interpolációval történik
a folyamat során létrejött mesterséges határvonalak megszüntetése céljából.
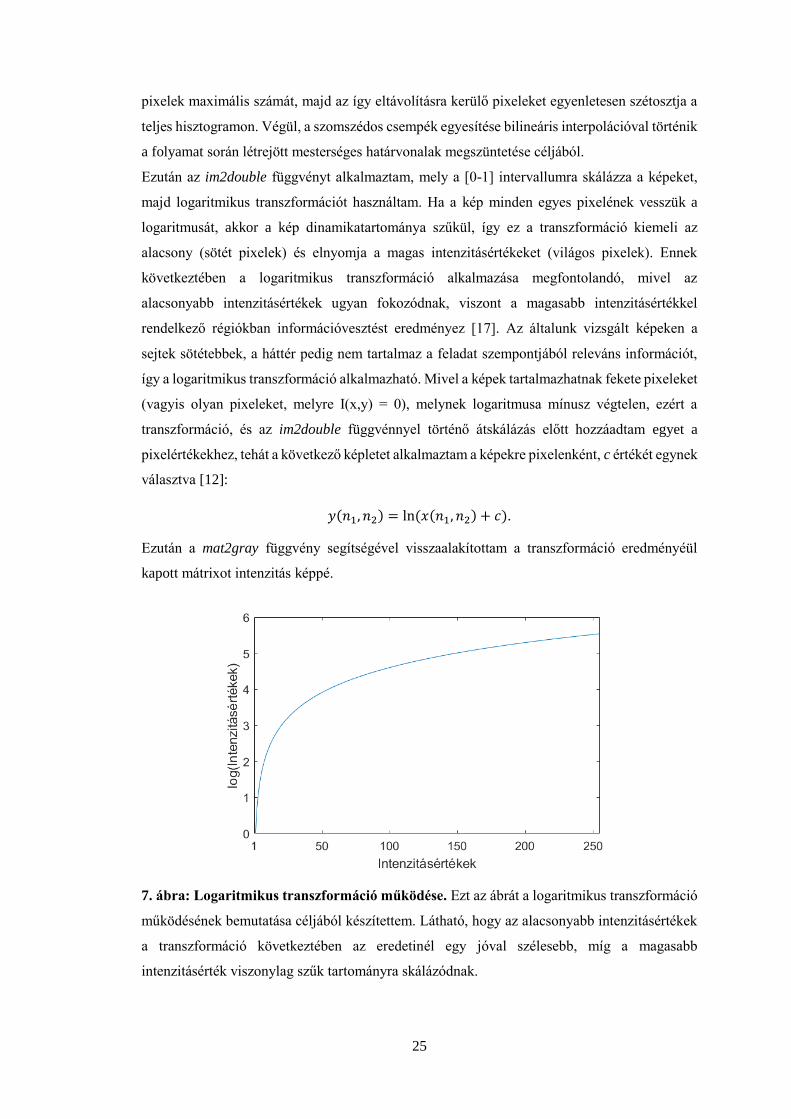
Ezután az im2double függvényt alkalmaztam, mely a [0-1] intervallumra skálázza a képeket,
majd logaritmikus transzformációt használtam. Ha a kép minden egyes pixelének vesszük a
logaritmusát, akkor a kép dinamikatartománya szűkül, így ez a transzformáció kiemeli az
alacsony (sötét pixelek) és elnyomja a magas intenzitásértékeket (világos pixelek). Ennek
következtében a logaritmikus transzformáció alkalmazása megfontolandó, mivel az
alacsonyabb intenzitásértékek ugyan fokozódnak, viszont a magasabb intenzitásértékkel
rendelkező régiókban információvesztést eredményez [17]. Az általunk vizsgált képeken a
sejtek sötétebbek, a háttér pedig nem tartalmaz a feladat szempontjából releváns információt,
így a logaritmikus transzformáció alkalmazható. Mivel a képek tartalmazhatnak fekete pixeleket
(vagyis olyan pixeleket, melyre I(x,y) = 0), melynek logaritmusa mínusz végtelen, ezért a
transzformáció, és az im2double függvénnyel történő átskálázás előtt hozzáadtam egyet a
pixelértékekhez, tehát a következő képletet alkalmaztam a képekre pixelenként, c értékét egynek
választva [12]:
𝑦(𝑛1, 𝑛2) = ln(𝑥(𝑛1, 𝑛2) + 𝑐).
Ezután a mat2gray függvény segítségével visszaalakítottam a transzformáció eredményéül
kapott mátrixot intenzitás képpé.
7. ábra: Logaritmikus transzformáció működése. Ezt az ábrát a logaritmikus transzformáció
működésének bemutatása céljából készítettem. Látható, hogy az alacsonyabb intenzitásértékek
a transzformáció következtében az eredetinél egy jóval szélesebb, míg a magasabb
intenzitásérték viszonylag szűk tartományra skálázódnak.

26
8. ábra: Az előfeldolgozás lépéseinek bemutatása. Az ábra A-val jelölt részén az eredeti kép,
B-vel jelölt részén az adaptív hisztogram kiegyenlített, míg C-vel jelölt részén a logaritmikus
transzformáció eredményeképp kapott kép szerepel. A végeredmény egy olyan kép, melyen jól
elkülönül a háttér, a sejttest és a nyúlványok. Ez segíti a későbbi automatikus feldolgozást.
3.3.1.2. Szegmentálás
A következő lépés a képek szegmentálása volt, melyhez a 2.5. fejezetben ismertetett
módszerek közül az intenzitásértékek alapján történő küszöbölést választottam, mivel a vizsgált
képeken a háttér és az objektumok jelentősen eltérő intenzitástartományba estek. A mikroglia
sejtek háttértől való elválasztásához többszintű, küszöbérték-alapú szegmentálást használtam.
A küszöbértékeket a multithresh függvény segítségével határoztam meg, mely tetszőleges
számú küszöbérték számítását végzi el, Otsu módszerét alkalmazva. A meghatározni kívánt
küszöbértékek számát kettőnek választottam, így végül három osztályt kaptam. Az első
osztályba a kép azon részei estek, melyek a legsötétebbek, vagyis főként a sejttestek, így ezt
vettem alapul a képen található sejtek számának meghatározásához. Ehhez a megfelelő
küszöbérték alapján szegmentáltam a képet, így eredményül egy olyan bináris képet kaptam,
melyen fekete a háttér, az objektumok pedig fehéren ábrázolódnak. Innentől kezdve a
sejtszámlálás felfogható úgy, mint a képen lévő összefüggő komponensek (objektumok)
számának meghatározása.
Az objektumokat a bwlabel függvényt alkalmazva jelöltem meg. Ez a függvény egy bináris
képet vár bemenetéül és egy úgynevezett címke mátrix-szal tér vissza, melyben a különböző
összefüggő régiókban lévő pixelek más-más értéket vesznek fel, így a későbbiekben
egyértelműen azonosíthatók. Tehát, ha adott a következő mátrix, mely egy bináris képet
reprezentál:

27
[1 1 01 1 00 0 0
0 0 01 1 11 1 1
],
akkor a bwlabel függvényt alkalmazva a címke mátrix az alábbi mátrix lesz.
[1 1 01 1 00 0 0
0 0 02 2 22 2 2
].
Ezután a regionprops függvény segítségével meghatároztam a sejtek detektálásához szükséges
paramétereket. A regionprops függvény a képen lévő régiók tulajdonságainak mérésére szolgál.
Mivel bemenetéül egy olyan mátrixot adtam meg, melyen előzetesen megjelöltem az
objektumokat, ezért a kívánt paramétereket objektumonként számította ki. Meghatároztam ezen
összefüggő régiókat alkotó pixelek számát és lineáris indexét, valamint a régiók súlypontját.
Ekkor a képen még szerepeltek olyan objektumok, melyek nem tekinthetők teljes sejtnek
(például képbe belógó nyúlványok), ezért kizártam azokat az objektumokat, melyek által
elfoglalt terület egy bizonyos pixelszám alatt volt. Ezt a minimális pixelszámot empirikus
módon határoztam meg, mivel nem volt előzetes információm az átlagos sejtméretről, és az
összes objektum átlagos méretét véve túl kicsi számot kaptam. Néhány érték kipróbálása után a
legjobbnak tűnőn finomítottam tovább és végül a minimális méretet 235 pixelnek választottam.
Ezután meghatároztam a képen maradt objektumok számát. A detekció eredményét a 9. ábra
szemlélteti. A sejtek alakja elég változatos, ezért előfordulhat olyan eset, hogy az így kapott
maszkon szerepelnek konkáv objektumok, melyek súlypontja nem minden esetben esik az
objektumon belülre. Mivel a későbbiekben (például NND meghatározása, lásd 3.3.3. fejezet)
szükségem lesz egy olyan pontra, ami biztosan az adott régióhoz tartozik, úgy módosítottam a
súlypont koordinátáit, hogy a kapott új pont a régión belülre essen.

28
9. ábra: A detekció eredménye. A bal oldalon egy fénymikroszkópos felétel látható, melyen
előzetesen meg lettek jelölve a mikroglia sejtek (ezeket piros ponttal jelöltem). Jobb oldalon a
bemutatott módszerrel történő szegmentálás eredményéül kapott bináris maszk látható.
3.3.2. Nyúlványrendszer modellezése
A következő lépésben a detektált sejtekhez hozzárendeltem a nyúlványrendszerüket,
melyhez aktív kontúr módszert használtam. Az aktív kontúr módszer, vagy másnéven snake
alapötlete, hogy egy olyan görbe segítségével detektálja az objektumokat, melyet iteratív módon
alakít az adott kép függvényében. A módszer célja az alábbi energiafüggvény minimalizálása:
𝐸 = ∫1
2
1
0
(𝛼|𝒗′(𝑠)|2 + 𝛽|𝒗"(𝑠)|2) + 𝐸𝑒𝑥𝑡(𝒗(𝑠)) 𝑑𝑠 ∶ 𝒗(𝑠) = [𝑥(𝑠), 𝑦(𝑠)],
ahol α és β a belső energia rugalmassági és merevségi komponenseinek súlyozására szolgáló
paraméterek, Eext pedig a képből származó külső energia [21]. Az aktív kontúr módszerek
alapvetően két csoportba sorolhatók, megkülönböztetünk parametrikus [24-25], illetve
nemparametrikus (régió alapú) módszereket [19]. A parametrikus módszerek általában
érzékenyebbek a zajra és a paraméter beállításokra, ezen kívül a nagy görbületű kontúrok
detekciója is problémás. A régió alapú aktív kontúr eljárások előnye, hogy kevésbé érzékenyek
a kezdőkontúr kijelölésre és több, akár komplex objektumot is képesek egyszerre megtalálni.
Azonban problémát jelent számukra a nyitott élek és a körvonalon belüli intenzitásváltozás,
valamint a konvergencia sebességük is lassabb.
A fentiek alapján a nyúlványrendszer változatos és komplex objektumainak
körvonalazására a régió alapú módszer lehet alkalmasabb, ráadásul ebben az esetben egy jó

29
inicializálással egy lépésben az összes sejt nyúlványrendszere kinyerhető. Ehhez a beépített
activecontour függvényt használtam, mely a Chan és Vese [19] által leírt módszert
implementálja. Ebben a szerzők egy level-set (szinthalmaz) alapú görbe evolúciós formulát
javasoltak, mely implicit módon írja le a változó kontúrt. A módszer alapvető célja az adott kép
két régióra (háttér és objektum) történő felosztása az által, hogy az objektum határait egy
háromdimenziós level-set függvény nullaszintű görbéjének feleltetik meg, tehát az eljárás során
egy olyan level-set alapú görbét keresünk, mely minimalizálja az energiát (ECV). Legyen ϕ egy
level-set függvény, ekkor a Chan-Vese formula a következőképp írható:
𝐸𝐶𝑉(𝑐1, 𝑐2, 𝜙) = 𝜆1 ∫(𝑓 − 𝑐1)2
𝛺
𝐻(𝜙) 𝑑𝑥 + 𝜆2 ∫(𝑓 − 𝑐2)2
𝛺
(1 − 𝐻(𝜙)) 𝑑𝑥 + 𝜇 ∫|∇𝐻(𝜙)| 𝑑𝑥
𝛺
ahol λ1, λ2 > 0 és μ ≥ 0 az egyes tagok súlyozására szolgáló fix paramétereket, f a vizsgált képet,
H pedig az egységugrás függvényt jelöli. Az első tag az objektumra, a második a háttérre, a
harmadik tag pedig a görbe hosszára vonatkozik. Az objektum (ϕ ≥ 0) és a háttér (ϕ < 0) átlagos
intenzitásértékeit c1, illetve c2 jelöli. A vizsgált kép szegmentálásához ezt az energiafüggvényt
minimalizálni kell c1-re, c2-re és ϕ-re vonatkozóan. A keresett kontúr a level-set függvény
nullaszintű pontjainak halmaza, vagyis azoké a pontoké, melyekre a függvényérték zérus (ϕ =
0).
10. ábra: A Chan-Vese formula alapján a görbe, mint a level-set függvény nullaszintű
vonala. Forrás: [19]
A MATLAB-os activecontour függvénynek az adaptív hisztogram kiegyenlített képeket adtam
bemenetéül, kezdeti kontúrként pedig a detekció eredményeképp kapott maszkot használtam.
Az így kapott bináris képen a nyúlványok látványosan vastagabbak voltak, mint az eredeti
felvételen, ezért morfológiai vékonyítást alkalmaztam. A végső eredmény a 11. ábrán látható.
Sok esetben előfordult, hogy az egymáshoz közel lévő sejtek érintkeztek, vagy éppen
nyúlványaik átfedtek egymással, ezért a maszkon egy objektumként jelentek meg. Mivel
szerettem volna a sejteket külön-külön is elemezni, fontos volt erre a problémára megoldást
találni. Első lépésként kiszűrtem az átfedő sejteket a detekció eredményeképp kapott maszk
alapján. Amennyiben egy adott objektum a detekció során kapott maszkon lévő objektumok

30
közül egynél többet tartalmazott, úgy a sejtek nyúlványrendszere átfedett. A következő lépés
ezen sejtek szétválasztása volt, melyhez gyakran watershed transzformációt használnak. Ez
olyan konvex objektumok esetében működik jól, melyek kismértékben fednek át egymással [26-
27]. Ezt alátámasztották az általam elvégzett tesztek is, mivel a módszer a sejtek szétválasztására
nem adott hatékony eredményt. A mikroglia sejtek nyugalmi állapotában a nyúlványrendszer
igen kiterjedt (tehát konkáv objektumokról beszélünk), ezért a szétválasztást szintén az
activecontour függvénnyel oldottam meg.
A függvény ContractionBias paraméterével lehet befolyásolni, hogy a kontúrok milyen irányba
tendáljanak. Értékét növelve, mely alapértelmezetten nulla, a kezdeti kontúrok egyre kevésbé
terjednek ki, vagyis a megfelelő értéket megválasztva elérhető a kívánt cél, viszont számolni
kell a folyamat következtében fellépő nyúlványveszteséggel is. A szétválasztáshoz kivágtam az
eredeti képből és a detekció eredményeképp kapott maszkból a megfelelő régiót, majd addig
növeltem a ContractionBias paramétert, amíg a szegmentáció eredménye pontosan annyi
objektum lett, amennyi átfedett. Végül lecseréltem az átfedő objektumokat az így kapott
eredményre. Amennyiben a sejtek szétválasztása sikertelen volt (például a vártnál több
objektumot eredményezett), úgy kizártam őket a sejtenkénti analízisből.
11. ábra: Aktív kontúr módszerrel történő szegmentálás eredménye. Bal oldalon az eredeti
fénymikroszkópos felvétel, míg jobb oldalon az aktív kontúr módszerrel történő szegmentálás
eredményéül kapott bináris maszk látható. A kezdeti kontúrokat a 9. ábrán látható maszkkal
adtam meg.

31
12. ábra: Érintkező/átfedő sejtek szétválasztásának eredménye. Az ábra A és D részén 2-2
egymással érintkező sejt látható, B és E részén pedig a róluk készült maszk. Az ábra C és F
részén ugyanezen sejtekről készült maszk látható az aktív kontúr módszerrel történő
szétválasztás után. Első esetben (C-vel jelölt kép) a szétválasztáshoz elegendő volt egyszer
növelni a ContractionBias paramétert, így a nyúlványveszteség is minimális. Második esetben
(F-el jelölt kép) a szétválasztás több iteráció alatt zajlott csak le, így a nyúlványvesztés
kifejezettebb.
3.3.3. Paraméterek meghatározása
A vizsgált paraméterek meghatározását a maszkolt képeken végeztem. A detektált
mikroglia sejtek által elfoglalt összterület meghatározásánál még figyelembe vettem az átfedő
sejteket is, így a szétválasztásból fakadó nyúlványveszteség ezt nem befolyásolja. A képeken
egy pixel 1264⁄ mm-nek felel meg, így a területszámításnál az alábbi képletet használtam:
(1264⁄ )
2∗ (𝑓𝑒ℎé𝑟 𝑝𝑖𝑥𝑒𝑙𝑒𝑘 𝑠𝑧á𝑚𝑎).
Az egyes sejtekre vonatkozó paramétereket a regionprops függvény segítségével határoztam
meg. Kiszámítottam sejtenként a lefedett területet (covered area, CA), valamint annak a
legkisebb konvex környezeti területnek a nagyságát, mellyel a teljes sejt lefedhető (covered
environmental area, CEA). Ezt a sejtek legtávolabbra nyúló nyúlványaira illesztett konvex
sokszögből határoztam meg (13. ábra). Ezután kiszámítottam a sejtenkénti terület és konvex
környezeti terület hányadosát, mely egyfajta becslést adhat a sejtek morfológiájára. Ez az
arányszám akkor maximális, hogyha az objektum, amire a sokszöget illesztjük a lehető
legnagyobb mértékben kitölti azt, vagyis megegyezik vele. Tehát minél inkább illeszkedik a
sokszög az objektumra, a hányados annál nagyobb. Ez a vizsgált sejtekre vonatkoztatva annyit

32
jelent, hogy a fagocitáló, amőboid formát mutató sejtekre számolt arányszám magasabb lesz,
mint a nyugalmi állapotban lévő mikroglia sejtekre számított érték.
Ezen kívül meghatároztam minden képre a sejtek legközelebbi szomszédjának átlagos
távolságát (nearest neighbor distance, NND). Ehhez minden sejtre kiszámítottam a hozzá
legközelebb lévő szomszédjától vett távolságot (euklideszi távolság alapján), majd vettem a
kapott eredmények átlagát. A pontokat, melyek alapján a távolságszámítás történt a regionprops
függvény által kiszámított súlypont adta.
Az eredményeket a könnyebb átláthatóság végett szöveges dokumentumba mentettem.
13. ábra: Konvex környezeti terület ábrázolása. Az ábra bal oldalán az eredeti képből
kivágott egyetlen sejt látható, míg jobb oldalán a sejtről készített maszkot ábrázoltam szürkével,
a sejt legtávolabbra nyúló nyúlványaira illesztett konvex sokszöget pedig fehérrel.
3.4. Grafikus felhasználói felület bemutatása
Kitűzött céljaim között szerepelt egy olyan interaktív grafikus felhasználói felület (GUI)
létrehozása, melynek segítségével a felhasználó korrigálhatja a detekció eredményét, ezzel
pontosítva az elemzés végeredményén. Ezt a felületet szintén MATLAB-ban implementáltam.
Az elemezni kívánt kép kiválasztása után a 3.3.2-es pontban ismertetett detekciós
algoritmus lefut a képre, és az eredménye egyből láthatóvá válik a felhasználó számára. Ezt
követően a felhasználónak lehetősége nyílik a módosításra, vagyis törölhet, illetve hozzáadhat
sejteket a detekció eredményéhez (14. ábra). A törlés lehetőség egyszerűen eltávolítja a
kiválasztott sejtre vonatkozó eltárolt adatokat, illetve a detekció eredményéül kapott jelölt
maszk megfelelő régiójában lévő pixelek értékét nullára változtatja (vagyis egyenlővé teszi a
háttérrel). A hozzáadás lehetőséget választva meg kell adni a kiválasztott sejt egy pontját, amit
legegyszerűbben a sejtre történő jobb egérgombbal való kattintással lehet megkapni. A pont
megadása után a maszkon létrejön egy új objektum, melynek súlypontja a felhasználó által
megadott pont, valamint hozzáadódik egy új elem az adatokat tároló struktúrához az új
objektumra vonatkozó adatokkal. A módosítás során a maszkon létrejött változásokat a 15. ábra
szemlélteti.

33
14. ábra: A grafikus felhasználói felület segítségével megvalósítható módosítások
bemutatása. Sárga jelölés látható azokon a sejteken, amiket a program automatikusan
meghatározott. Zölddel jelöltem azokat, melyeket az Add point gombot használva, utólag lettek
hozzáadva a detekció eredményéhez. Amennyiben a felhasználó törölni szeretne a megjelölt
sejtek közül, akkor ezt az eltávolítandó sejt sorszámát megadva, a Delete point gombbal teheti
meg. Ennek hatására a sejt jelölése megszűnik.

34
15. ábra: A felhasználói beavatkozás hatására történő változások bemutatása a
sejtdetekció eredményéül kapott maszkon. A bal oldali képen az automatikus detekció
eredményéül kapott maszk látható. Pirossal jelöltem azokat az objektumokat, melyeket a
grafikus felhasználói felület segítségével el szeretettem volna távolítani. A jobb oldali képen a
már módosított maszk látható. Pirossal jelöltem azokat a régiókat, melyeket a felhasználói
felület segítségével adtam hozzá a detekció eredményéhez. Ezen kívül látszik, hogy az
eltávolítás céljából megjelölt sejtek itt már nem jelennek meg, tehát sikeresen törlődtek.
Miután már csak azok a sejtek vannak megjelölve, amit a felhasználó jónak lát, akkor
elindíthatja a nyúlványrendszer modellezését (Create mask gomb) a módosított maszkkal.
Ekkor lefut a 3.3.2-es pontban leírt, nyúlványhálózat modellezésére alkalmazott eljárás az
érintkező/átfedő sejtek szétválasztása nélkül, melynek eredménye, vagyis a teljes sejtekről
készült maszkolt kép megjelenik egy külön ablakban. Ekkor a felhasználónak lehetősége nyílik
az érintkező sejtek manuális szétválasztására (Separate cells gomb). Ezt a folyamatot a 16. ábra
szemlélteti. Amennyiben a felhasználó az eredményt látva már nem végezne további
módosításokat, akkor elvégezheti a részletes elemzést (Analyse gomb, 3.3.3-as pontban leírt
számítások), majd elmentheti ennek eredményét szöveges dokumentum formájában, valamint a
maszkolt képet is (Save results illetve Save mask gombok). A fejezetben szereplő példa felvétel
részletes elemzésének eredménye megtekinthető a mellékletek között (1. melléklet).

35
16. ábra: A manuális sejt szétválasztás bemutatása. Az ábra A-val jelölt részén két érintkező
sejtről készült maszk látható. A felhasználónak a Separate cells gombra kattintva lehetősége
nyílik a sejtek szétválasztására. Ehhez csupán bele kell kattintani a képbe és húzni kell egy
egyenest, ami átszeli azt a régiót, ahol el szeretnénk választani a két sejtet (ezt a B-vel jelölt kép
szemlélteti). Az ábra C-vel jelölt részén a szétválasztás eredményét jelenítettem meg. Látható,
hogy ezzel a módszerrel alig pár pixelnyit veszítünk a sejtekből, így a nyúlványveszteség
elhanyagolható. Összehasonlítva az automatizált szétválasztással (12. ábra), a manuális módszer
jóval kevesebb információvesztéssel jár, valamint, mivel a felhasználó jelölheti meg a
szétválasztás helyét, ezzel a módszerrel pontosabb eredmény érhető el.
3.5. ImageJ szoftverrel történő elemzés
A kutatócsoport tagjai jelenleg ImageJ programmal elemzik a munkájuk során készített
felvételeket, mely egy Java-alapú, széleskörben használt képelemző alkalmazás. A
dolgozatomban vizsgált képek elemzését megcsináltam ImageJ-vel is, hogy legyen mivel
kvantitatív módon összehasonlítani a kapott eredményeket. Az elemzés során azt a módszert
alkalmaztam, amit ők is szoktak, és a szoftver 1.51j8-as verzióját használtam.
A sejteket a Multi-point Tool segítségével számláltam és jelöltem meg. Ez az eszköz a
megjelölt pontok koordinátáit rögzíti, így ezeket szöveges dokumentum formájában el lehet
menteni (Save as XY Coordinates). Az így kapott koordináták alapján, MATLAB segítségével
a 3.3.3-as pontban ismertetett módon kiszámítottam az NND-t.
A képekre ezután alkalmaztam az Enhance Contrast parancsot, mely hisztogram nyújtással
valósítja meg a kontraszt növelését [33]. Hisztogram nyújtással elérhetjük, hogy az
intenzitásértékek a teljes dinamikatartományt kihasználják. Erre az általános képlet a következő
[18]:
𝑦(𝑛1, 𝑛2) = 255 ∗ (𝑥(𝑛1, 𝑛2) − min
𝑛1,𝑛2
(𝑥(𝑛1, 𝑛2))
max𝑛1,𝑛2
(𝑥(𝑛1, 𝑛2)) − min𝑛1,𝑛2
(𝑥(𝑛1, 𝑛2)))

36
A következő lépés a képek élesítése volt, melyet a Sharpen parancs alkalmazásával értem el.
Ez a művelet az egyes képpontokat az azok 3*3-as szomszédságában lévő pixelek súlyozott
átlagával helyettesíti, mely kiszámításához a következő súlymátrixot használja [33]:
[−1 −1 −1−1 12 −1−1 −1 −1
]
Ezt követően az intenzitásértékek alapján küszöböltem a képet. A Threshold/Auto parancs
automatikusan meghatározza a küszöbértéket a hisztogram alapján, az IsoData algoritmus
segítségével [33]. Sok esetben módosítanom kellett ezen a küszöbértéken, hogy a szegmentálás
megfelelő legyen.
Az így kapott bináris képen méret alapú küszöbölést alkalmaztam. Ehhez az Analyse Particles
parancs kiválasztása után a Size lehetőségnél beállítottam egy minimális pixelértéket, így
kizártam azon objektumokat a további analízisből, melyek területe ezt nem érte el. Ezt a
minimális méretet minden képre manuálisan állítottam be úgy, hogy a legjobb eredményt adja.
Előfordult, hogy a képen így is maradtak olyan objektumok, melyek nem lettek sejtként jelölve.
Végül kiszámítottam a képen maradt objektumok összeterületét a képen lévő, objektumokat
reprezentáló pixelek összegzésével.
17. ábra: ImageJ-vel és a kifejlesztett módszerrel történő szegmentáció eredménye. Az ábra
A-val jelölt részén az eredeti kép egy részlete, a B-vel jelölt részen ImageJ-vel történő
szegmentálás eredménye, míg a C-vel jelölt részen az általam kifejlesztett módszerrel történő
szegmentálás eredménye látható. A B és a C részt összevetve látszik, hogy a C részen a
nyúlványrendszer részletesebb.

37
4. Eredmények
4.1. Krónikus stressz kísérlet
A stressz kísérlet során kapott eredményeken kétmintás t-próbát végeztem, melyet
megelőzően a minták szórásának egyenlőségét F-próbával ellenőriztem.
Az állatok testtömeg gyarapodásában nem mutatkozott szignifikáns különbség a csoportok
között. A kísérleti csoport testtömeg gyarapodását 84.25 ± 8.71g-nak, míg a kontroll csoportét
89.88 ± 16.43g-nak találtuk.
Hogy átfogóbb képet adjak a stressz kísérlet eredményéről, a továbbiakban ismeretem a másik
módszerrel feldolgozott csoport hormonvizsgálatának és cukor preferencia mérésének
eredményeit is. A hormonvizsgálat eredményének kiértékelése során nem mutatkozott
szignifikáns különbség a csoportok között sem az ACTH (kísérleti: 17.86±1.22 fmol/ml,
kontroll: 15.85±1.72 fmol/ml), sem pedig a koritkoszteron (kísérleti: 69.55±17.21 pmol/ml,
kontroll: 45.40±8.90 pmol/ml) szintekben. A cukor preferencia vizsgálat sem mutatott jelentős
eltérést a két csoport között (kísérleti csoport: 86.98 ± 1.01%, kontroll csoport 88.58 ± 1.34%),
viszont ahogy a testtömeg gyarapodás és a hormonvizsgálat esetén, itt is megfigyelhető a
csoportok közti átlagos különbség.
1. diagram: A hormonvizsgálat eredménye (átlag ± szórás). A két csoport között ugyan
szignifikáns különbség nem mutatkozott, de a kísérleti csoport kortikoszteron értékei átlagosan
meghaladják a kontroll csoportét.
0
15
30
45
60
75
90
ACTH szint [fmol/ml] Kortikoszteron szint[pmol/ml]
Kontroll csoport (n = 16)
Kísérleti csoport (n = 23)

38
2. diagram: Az első csoport állatainak testtömeg gyarapodása a stressz kísérlet elejétől a
18. napjáig (átlag ± szórás). Látható, hogy a kísérleti csoport testtömeg gyarapodása elmaradt
a kontroll csoportétól.
3. diagram: Cukorpreferencia teszt eredménye (átlag ± szórás). A kísérleti csoport
átlagosan kevesebbet fogyasztott a szacharóz oldatból, mint a kontroll csoport.
4.2. Az automatikus sejtdetekció eredménye
Az automatikus sejtdetekció eredményének kiértékelésekor az általam elemzett
felvételek manuálisan megjelölt változatát használtam referenciaként. A kifejlesztett módszer
teljesítményének mérésére kiszámítottam a találati arányt (hit rate, vagy másnéven recall) és a
pozitív prediktív értéket (Positive Predictive value, PPV, vagy másnéven precision) a következő
módon:
𝑟𝑒𝑐𝑎𝑙𝑙 =𝑇𝑃
𝑇𝑃 + 𝐹𝑁
𝑝𝑟𝑒𝑐𝑖𝑠𝑖𝑜𝑛 = 𝑇𝑃
𝑇𝑃 + 𝐹𝑃
30
50
70
90
110
Kontroll csoport (n = 8) Kísérleti csoport (n = 8)
Test
töm
eg g
yara
po
dás
[g]
75
80
85
90
95
Kontroll csoport (n = 16) Kísérleti csoport (n = 23)
%

39
ahol TP jelöli az érvényes találatokat, FN a fals negatív, FP pedig a fals pozitív találatokat.
Előbbi 0.9241-nek, míg utóbbi 0.9-nek adódott, tehát a kifejlesztett módszer az előzetesen
megjelölt sejtek 92.41%-át megtalálta, és az összes találat közül 90% volt érvényes.
Sejtszám Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
Manuális 52 ± 4 44 ± 5 49 ± 3 46 ± 7
Automatizált 51 ± 6 47 ± 6 50 ± 6 47 ± 7
2. táblázat: A manuális és az automatizált detekció eredménye. A táblázatban a különböző
módszerekkel detektált sejtek száma (átlag ± szórás) szerepel. Szignifikáns eltérést egyik
módszer esetén sem mutatkozott a két csoport között.
4.3. A különböző módszerekkel meghatározott paraméterek kiértékelése
A felvételek elemzését három módon végeztem el (ImageJ-vel, a kifejlesztett
automatizált módszerrel és az ehhez készült grafikus felhasználói felület segítségével). A kapott
eredményeken kétmintás t-próbát végeztem, melyet megelőzően a minták szórásának
egyenlőségét F-próbával ellenőriztem. A GUI-es elemzés során úgy módosítottam a detekció
eredményét, hogy az teljesen megegyezzen az előzetesen manuálisan megjelölt sejtekkel. Az
eredmények a 2-7. táblázatokban megtekinthetők, melyek a különböző agyterületekre
kiszámított átlagértéket és szórást tartalmazzák.
Az elemzés során a sejtek számát, az általuk elfoglalt összterületet, a legközelebbi
szomszédjuktól vett átlagos távolságukat (NND), a sejtek áltagos méretét, konvex környezeti
területének átlagos méretét (CEA) és az utóbbi két paraméter hányadosát vettem alapul. A
vizsgált paraméterek tekintetében egyedül a PVN területén, a sejtek által elfoglalt összterület
esetén mutatkozott szignifikáns különbség a kísérleti és a kontroll csoport között. A csoportok
közti eltérés mindhárom alkalmazott módszer esetén megfigyelhető (3. táblázat), viszont csak a
GUI-s elemzés során lett szignifikáns (p = 0.042).

40
Összterület [%] Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
ImageJ 4.13 ± 0.30 4.08 ± 0.48 3.87 ± 0.35 3.84 ± 0.80
Automatizált 4.00 ± 0,63 4.75 ± 0.90 3.58 ± 0.63 5.20 ± 1.22
GUI 4.05 ± 0.46 4.38 ± 0.76 3.46 ± 0.44 4.92 ± 1.27
3. táblázat: A különböző módszerekkel meghatározott sejtek által elfoglalt összterület.
Szignifikáns különbség csak a grafikus felhasználói felülettel történő elemzés során mutatkozott
két csoport között, kizárólag a PVN esetében (vastaggal szedve).
NND [μm] Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
ImageJ 380 ± 16 350 ± 20 391 ± 10 343 ± 24
Automatizált 385 ± 23 338 ± 22 392 ± 22 339 ± 24
GUI 380 ± 16 350 ± 19 389 ± 10 342 ± 24
4. táblázat: A különböző módszerekkel meghatározott sejtekre kiszámított NND. Bár az
ImageJ-s és a GUI-s eredmények ugyanazon sejtek alapján számítódtak, mégis látható köztük
némi eltérés. Ennek magyarázata, hogy a GUI-s elemzéskor a sejtek egy jó részét az algoritmus
automatikusan detektálta, és az objektumok súlypontja (mely az NND számítás alapját adta)
nem pontosan oda esett ahova az ImageJ-s elemzés során a jelölés.
CA [μm2] Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
ImageJ - - - -
Automatizált 12296 ± 676 11592 ± 684 11307 ± 925 12381 ± 1581
GUI 12740 ± 1065 12064 ± 870 11667 ± 1150 12858 ± 1819
5. táblázat: A különböző módszerekkel meghatározott sejtek átlagos területe. A két
módszer által kapott eredményben való eltérés két dologból adódhat. Az egyik, hogy az
automatikus detekcióból fakadóan volt eltérés a vizsgált sejtek tekintetében a két módszer között
(lásd 4.1-es fejezet). A másik pedig, hogy az automatikus szétválasztás jelentős
nyúlványveszteséggel, így a sejt által elfoglalt terület csökkenésével járhat (12. ábra), míg a
GUI-es elemzés során a manuális szétválasztás során fellépő információvesztés minimális (16.
ábra).

41
CEA [μm2] Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
ImageJ - - - -
Automatizált 47368 ± 4438 40219 ± 1976 43561 ± 5820 42487 ± 6846
GUI 50537 ± 5659 42364 ± 1961 45701 ± 7422 43932 ± 7635
6. táblázat: A különböző módszerekkel meghatározott sejtek átlagos konvex környezeti
területe. A két módszer kissé eltérő eredményeinek hátterében az 5. táblázathoz tartozó
leírásban megfogalmazott okok állnak. A kísérleti és a kontroll csoportot összehasonlítva
látható, hogy a PVN területén átlagosan nagyobb CEA-t mértünk a stresszelt állatokon, de a
különbség nem szignifikáns.
CA/CEA Kísérleti csoport Kontroll csoport
Módszer PVN ARC PVN ARC
ImageJ - - - -
Automatizált 0.30 ± 0.03 0.33 ± 0.02 0.29 ± 0.02 0.33 ± 0.02
GUI 0.29 ± 0.03 0.32 ± 0.02 0.29 ± 0.02 0.33 ± 0.02
7. táblázat: A különböző módszerekkel meghatározott sejtek átlagos terület és konvex
környezeti terület aránya. Ez az arányszám információt ad a sejtek morfológiai értelemben
vett tömörségéről (solidity).
4.4. A felvételek elemzésére alkalmazott módszerek összehasonlítása
A három alkalmazott módszer összehasonlítására páros t-próbát alkalmaztam. Az
összehasonlításkor a sejtek által elfoglalt összterületet vettem alapul, mivel a sejtek számán
kívül ez az egyetlen paraméter, amit mindegyik módszerrel meghatároztam.
A PVN esetében csak az ImageJ-s és a GUI-s elemzés során kapott eredmények között
mutatkozott szignifikáns különbség (p = 0.038), viszont az ARC területén számított értékek
eltérése mindhárom esetben szignifikáns volt (ImageJ és GUI: p = 0.0002; ImageJ és
automatizált módszer: p = 0.0001; GUI és automatizált módszer: p = 0.0098).
Az automatizált és a többi módszer között fennálló eltérés a vizsgált sejtek
különbözőségével magyarázható. Az ImageJ-s és a GUI-s eredmények közti különbség több
okból adódhat. Az ImageJ-s elemzés során sokszor előfordult, hogy a szegmentálás alapjául
szolgáló automatikusan meghatározott intenzitásértéket nagymértékben módosítanom kellett
ahhoz, hogy megfelelő eredményt kapjak. Előfordult, hogy a háttérből nagyobb, összefüggő
részek teljesen egybeolvadtak az objektumokkal, ezért a globális küszöbértéket növelnem
kellett, ami viszont a kép egyes részein jelentős nyúlványvesztést eredményezett. Számos

42
alkalommal előfordult az ellenkező eset is, mikor az elemezni kívánt sejtek egy részéből jelentős
részletek hiányoztak, ezért a küszöbértéket csökkentenem kellett. Ez a többi, eredetileg szépen
kirajzolódó sejt esetében is az objektumok növekedését eredményezte. Ezen kívül számolni kell
azzal is, hogy a kifejlesztett módszer a nyúlványok modellezésekor hozzáveszi a sejthez azokat
a nyúlványrészleteket is, amik ugyan nem lettek előzetesen detektálva, de érintkeznek a sejttel.

43
5. Diszkusszió, konklúzió
A stressz kísérlet eredményeképp megfigyeltük, hogy a kísérleti állatok testtömeg
gyarapodása és cukorpreferenciája kismértékben elmaradt a kontroll csoportétól. A
cukorpreferencia csökkenése enyhe anhedóniás állapot kialakulására enged következtetni. Bár
a csoportok közti különbség nem volt szignifikáns, de az eredmény jól tükrözi, hogy ezek a
tünetek általában krónikus stressz hatására alakulnak ki [5] [10]. A hormonszintekben való
eltérés a hipotalamusz-hipofízis-mellékvesekéreg tengely fokozott aktivitására utal, és
összhangban van más krónikus stresszel foglalkozó kutatások eredményével [10] [34].
A PVN területén történő stressz-indukálta mikroglia aktivációval több tanulmány is
foglalkozott az elmúlt időszakban [5] [34] [35], de az ARC-ban stressz hatására történő
morfológiai aktiváció egyelőre feltérképezetlen terület. A sejtek számában nem találtunk
szignifikáns eltérést egyik vizsgált agyterület esetében sem a kísérleti és a kontroll csoport
között. Ez összhangban van Tynan és mtsai [5] által kapott eredménnyel, miszerint az általuk
vizsgált 15 stressz-reszponzív agyterület közül a PVN-ben nem növekedett jelentős mértékben
a sejtek száma krónikus stressz hatására. Fontos megemlíteni, hogy az általuk és az általunk
alkalmazott stressz protokoll eltérő, viszont Kopp és mtsai [34] CVS-t is alkalmaztak munkájuk
során, és ők sem találtak eltérést a sejtek számában a PVN területén.
A sejtek által átlagosan elfoglalt terület (CA) mértékében csak átlagos különbséget mutattunk
ki a két csoport között, viszont a GUI-s elemzés során a sejtek által elfoglalt összterület a PVN-
ben szignifikánsan nagyobbnak bizonyult a kísérleti csoportban. Ennek egyik oka lehetne
például, hogy a kísérleti csoportban lényegesen magasabb volt a sejtek száma. Esetünkben ez
nem teljesül, így a különbség inkább a nyúlványrendszer nagyobb mértékű elágazódására,
esetleg kiterjedésére utalhat, ami a sejtek nyugalmi állapotból enyhén aktivált állapotba való
lépését jelezheti. Ezt alátámaszthatja az a megfigyelés is, miszerint az Iba-1 intenzitása az
aktivációval fokozódik [4-5] [34]. Wohleb és mtsai [35] is hasonló eredményre jutottak, bár ők
egereken végezték a vizsgálatot, és az általuk alkalmazott stressz időtartama csak
szubkrónikusnak minősül [13]. A többi krónikus stresszt alkalmazó kutatás [5] [34] során nem
találtak számottevő különbséget a két csoport között a PVN területén. Az ARC területén pont
ellenkező változásokat figyeltünk meg, de a csoportok közti különbség egyik esetben sem volt
jelentős.
A CA/CEA értékben akkor figyelhető meg jelentős különbség, hogyha vizsgált csoportok a
sejtek morfológiájában nagymértékben eltérnek, mivel ez a szám a sejtek morfológiai
aktivációjával arányosan nő [36]. Ebben az értékben nem találtunk eltérést a csoportok között
egyik vizsgált agyterület esetében sem, ami arra enged következtetni, hogy a CVS hatására nem
alakult ki jelentős morfológiai aktiváció sem a PVN, sem az ARC területén.

44
Az NND-ben sem mutatkozott számottevő különbség a csoportok között, vagyis stressz hatására
nem indult meg a sejtek migrációja. Ez várható volt, mivel az amőboid formát öltő sejtek
jellemzője, hogy a sérülés irányába vándorolnak [39], az általunk vizsgált felvételeken pedig
nem ez a forma volt a jellemző (lásd CA/CEA arány).
Összességében, a kapott eredmények azt mutatták, hogy a CVS hatására csökkent az
állatok testtömeggyarapodása és cukor preferenciája, valamint fokozódott a hipofízis-
mellékvesekéreg tengely aktivitása. A mikroglia aktiváció tekintetében a CVS jelentősen
növelte a sejtek által elfoglalt összterületet a PVN-ben, a sejtek számában való növekedés
nélkül. Bár az Iba-1 expressziója növekszik a központi idegrendszerben történő fertőzés és
gyulladás következtében [37], a kapcsolódó kutatások, melyek kimutatták a sejtek morfológiai
aktivációját, nem találtak gyulladásra utaló változásokat stressz következtében [4-5].
Esetünkben, a sejtek citokin expressziós profiljának meghatározásához egyéb vizsgálatok is
szükségesek lennének.
A munkám során kifejlesztett detekciós módszer az előzetesen megjelölt sejtek 92.41%-
át megtalálta, viszont ezen felül talált olyan objektumokat is, melyek nem lettek megjelölve.
Ezen objektumok egy része tekinthető sejtnek, mivel a gyakorlatban sokszor nehéz eldönteni,
hogy a fénymikroszkópos felvételeken látható bonyolultabb sejtek/sejtrészletek közül mit
jelöljünk meg további analízisre. Az eljárás néhány esetben sejtként érzékelt olyan
objektumokat is, melyek nem tekinthetők annak, például nagyobb nyúlványok, melyek
belógnak a képbe. Ennek ellenére a detekció pontossága (precision) 90% lett, ami egy jó
kiindulási alapot biztosíthat a felvételek elemzéséhez a GUI adta lehetőségekkel kiegészítve.
A mikroglia sejtek szegmentálása esetén nagyon fontos az alak minél pontosabb
megőrzése a sejt morfológiai tulajdonságainak meghatározásához. Az alkalmazott módszerek
összehasonlításakor megfigyelt különbség mögött több dolog állhat. Az egyik, hogy
amennyiben a háttérben nagymértékű változás figyelhető meg a kép egyes részei között, úgy
globális intenzitásküszöb alkalmazása során eltérő lesz a szegmentáció jósága a sejtek
elhelyezkedésétől függően. Az ImageJ-s elemzés során alkalmazott módszer globális
intenzitásküszöb alkalmazásával szegmentálja a képet, míg a kifejlesztett eljárás régió-alapú
aktív kontúr módszert használ a morfológiai információ kinyerésére. A másik, hogy bár az aktív
kontúr módszerrel történő szegmentáció eredménye sokkal részletgazdagabbnak és
pontosabbnak bizonyult, sajnos ez sem volt tökéletes. A módszer a nyúlványok modellezésekor
hozzávette a sejthez azokat a nyúlványrészleteket is, amik ugyan nem lettek előzetesen
detektálva, de érintkeznek a sejttel.
Összességében elmondható, hogy a felvételek minőségbeli, és a sejtek morfológiai
változatossága miatt az intenzitás-alapú küszöbölés a kifejlesztett módszerrel összehasonlítva
kevésbé effektív a képen lévő összes sejt morfológiai információjának egy lépésben történő

45
kinyerésére. Ezzel együtt, az implementált módszeren még szükséges dolgozni, további
fejlesztési lehetőségei a következők:
1) a sejtdetekcióra használt technika finomítása a pontosabb eredmény elérése végett
2) az átfedő sejtek automatikus szétválasztásának fejlesztése, hogy az minél kevesebb
információvesztéssel járjon
3) további paraméterek meghatározása, például nyúlványok száma és hossza
4) a GUI fejlesztése, például lehetőség a detekció elvetésére, oda nem illő
nyúlványrészletek törlésének lehetősége, illetve felhasználói visszajelzés alapján
történő egyéb fejlesztések/módosítások

46
6. Összefoglalás
A mikroglia sejtek a központi idegrendszer rezidens immunsejtjei, melyek kiemelt
szerepet játszanak annak védelmében és működésében. Nyugalmi állapotban ezek a sejtek
folyamatosan mintavételezik környezetüket, így gyorsan képesek reagálni különböző káros
behatásokra. Ebben az állapotban viszonylag kis sejttest, valamint vékony, kiterjedt és elágazó
nyúlványrendszer jellemző rájuk. Kóros folyamatot észlelve a sejtek aktiválódnak, melynek
során morfológiai és funkcionális változásokon mennek keresztül. Aktiváció során a
nyúlványok visszahúzódnak, a sejttest megnagyobbodik és változás következik be a sejt által
expresszált anyagok tekintetében is.
Több kutatás célja volt már a mikroglia sejtek és a stressz kapcsolatának vizsgálata. Ezen kívül
néhány tanulmányban foglalkoztak automatizált képelemző módszerekkel, melyek segítségével
nyomon lehet követni a mikroglia sejtek változásait mikroszkópos felvételeken. Jelen dolgozat
célja a stressz indukálta mikroglia aktiváció vizsgálata automatizált képelemző eljárásokkal.
A kísérleti állatok 3 hétig, naponta kétszer különböző stresszhatásoknak lettek kitéve,
majd az agyukból készített metszetekről immunhisztokémiai eljárást követően
fénymikroszkópos felvételeket készítettünk hússzoros nagyításban. A képeket MATLAB
(R2017a) szoftver, és annak képfeldolgozó eszköztára segítségével értékeltem ki. A kifejlesztett
módszer lehetővé teszi sejtek detektálásán túl a nyúlványrendszerük modellezését és jellemző
paramétereik meghatározását. Az automatikus detekció alapját logaritmikus képjavítás és
intenzitásérték-alapú szegmentáció, valamint méret alapú küszöbölés képzi. A sejtek
nyúlványrendszerének modellezésére régió alapú aktív kontúr módszert alkalmaztam. Az
elemzés során a sejtek számán kívül meghatároztam az általuk elfoglalt összterület százalékos
arányát, a legközelebbi szomszédjuktól vett átlagos távolságukat, a sejtek áltagos méretét,
konvex környezeti területének átlagos méretét és az utóbbi két paraméter hányadosát, mely
becslést adhat a sejtek morfológiai aktivációjának mértékére. Ezen kívül létrehoztam egy
grafikus felhasználói felületet, mely lehetőséget biztosít a detekció eredményének módosítására
és az érintkező sejtek manuális szétválasztására. A detekcióra használt módszer teljesítményét
a felvételek előzetesen manuálisan megjelölt változata alapján értékeltem. A módszer a
megjelölt sejtek 92.41%-át megtalálta, és az összes találat közül 90% volt érvényes.
A stressz kísérlet következtében csökkent az állatok testtömeggyarapodása és cukor
preferenciája, valamint fokozódott a hipofízis-mellékvesekéreg tengely aktivitása. A vizsgált
paraméterek tekintetében csak a sejtek által elfoglalt összterületben találtunk jelentős változást
stressz hatására, kizárólag a PVN területén, ami csekély mértékű mikroglia aktivációra utalhat.
Összességében, sikerült létrehozni egy olyan algoritmust, mely megkönnyítheti a
változatos morfológiával rendelkező mikroglia sejtek kvantitatív elemzését.

47
7. Köszönetnyilvánítás
Ezúton szeretnék köszönetet mondani konzulensemnek, Dr. Kovács Krisztinának,
amiért elvállalta a témavezetést. Köszönettel tartozom Winkler Zsuzsannának (MTA-KOKI,
Molekuláris Neuroendokrinológia Laboratórium) és Dr. Manno-Kovács Andreának (MTA
Számítástechnikai és Automatizálási Kutatóintézet, Gépi Érzékelés Kutatólaboratórium), akik
folyamatos szakmai támogatásukkal és iránymutatásukkal segítették szakdolgozatom
megvalósulását.

48
8. Irodalomjegyzék
[1] D. Orłowski, Z. Sołtys and K. Janeczko, "Morphological development of microglia in the
postnatal rat brain", International Journal of Developmental Neuroscience, vol. 21, no. 8,
pp. 445-450, 2003.
[2] H. Wake, A. Moorhouse and J. Nabekura, "Functions of microglia in the central nervous
system – beyond the immune response", Neuron Glia Biology, vol. 7, no. 01, pp. 47-53,
2011.
[3] M. Lull and M. Block, "Microglial activation and chronic neurodegeneration",
Neurotherapeutics, vol. 7, no. 4, pp. 354-365, 2010.
[4] S. Sugama et al, "Stress induced morphological microglial activation in the rodent brain:
Involvement of interleukin-18", Neuroscience, vol. 146, no. 3, pp. 1388-1399, 2007.
[5] R. Tynan et al, "Chronic stress alters the density and morphology of microglia in a subset
of stress-responsive brain regions", Brain, Behavior, and Immunity, vol. 24, no. 7, pp.
1058-1068, 2010.
[6] L. Varela and T. Horvath, "Leptin and insulin pathways in POMC and AgRP neurons that
modulate energy balance and glucose homeostasis", EMBO reports, vol. 13, no. 12, pp.
1079-1086, 2012.
[7] Y. Zhang et al, "Image processing methods to elucidate spatial characteristics of retinal
microglia after optic nerve transection", Sci. Rep. 6, 21816, 2016.
[8] C. Kozlowski and R. Weimer, "An Automated Method to Quantify Microglia Morphology
and Application to Monitor Activation State Longitudinally In Vivo", PLoS ONE, vol. 7,
no. 2, p. e31814, 2012.
[9] R. Kongsui et al, "Quantitative assessment of microglial morphology and density reveals
remarkable consistency in the distribution and morphology of cells within the healthy
prefrontal cortex of the rat", Journal of Neuroinflammation 11:182, 2014.
[10] T. Kreisel et al, "Dynamic microglial alterations underlie stress-induced depressive-like
behavior and suppressed neurogenesis", Molecular Psychiatry, vol. 19, no. 6, pp. 699-709,
2013.
[11] R. Szeliski, Computer Vision, 1st ed. London: Springer-Verlag London Limited, 2011, pp.
107-110.
[12] T. Maintz, Digital and Medical Image Processing. 2017, pp. 273-330.
[13] F. Walker, M. Nilsson and K. Jones, "Acute and Chronic Stress-Induced Disturbances of
Microglial Plasticity, Phenotype and Function", Current Drug Targets, vol. 14, no. 11, pp.
1262-1276, 2013.

49
[14] C. Smochina, P. Herghelehiu and V. Manta, "Image processing techniques used in
microscopic image segmentation", Universitatea Tehnică „Gheorghe Asachi” din Iasi
Tomul LVII (LXI), vol. 2, pp. 83-98, 2011.
[15] E. Meijering, "Cell Segmentation: 50 Years Down the Road [Life Sciences]", IEEE Signal
Processing Magazine, vol. 29, no. 5, pp. 140-145, 2012.
[16] K. Zuiderveld, "Contrast Limited Adaptive Histogram Equalization", Graphics Gems, pp.
474-485, 1994.
[17] U. Manikpuri and Y. Yadav, "Image Enhancement Through Logarithmic Transformation",
International Journal of Innovative Research in Advanced Engineering, vol. 1, no. 8, pp.
358-362, 2014.
[18] A. Abdul-Nasir, M. Mashor and Z. Mohamed, "Modified Global and Modified Linear
Contrast Stretching Algorithms: New Colour Contrast Enhancement Techniques for
Microscopic Analysis of Malaria Slide Images", Computational and Mathematical
Methods in Medicine, vol. 2012, pp. 1-16, 2012.
[19] T. Chan and L. Vese, "Active contours without edges", IEEE Transactions on Image
Processing, vol. 10, no. 2, pp. 266-277, 2001.
[20] F. Meyer, "Topographic distance and watershed lines", Signal Processing, vol. 38, no. 1,
pp. 113-125, 1994.
[21] M. Kass, A. Witkin and D. Terzopoulos, "Snakes: Active contour models", International
Journal of Computer Vision, vol. 1, no. 4, pp. 321-331, 1988.
[22] C. Scharfenberger et al, "Statistical Textural Distinctiveness for Salient Region Detection
in Natural Images", 2013 IEEE Conference on Computer Vision and Pattern Recognition,
2013.
[23] B. Misselwitz et al, "Enhanced CellClassifier: a multi-class classification tool for
microscopy images", BMC Bioinformatics, vol. 11, no. 1, p. 30, 2010.
[24] C. Xu and J. Prince, "Gradient Vector Flow: A New External Force for Snakes", IEEE
Proceedings Conference on Computer Vision and Pattern Recognition, pp. 66-71, 1997.B.
[25] Li and S. Acton, "Active Contour External Force Using Vector Field Convolution for
Image Segmentation", IEEE Transactions on Image Processing, vol. 16, no. 8, pp. 2096-
2106, 2007.
[26] Q. Wu, F. Merchant and K. Castleman, Microscope image processing. Amsterdam:
Elsevier/Academic Press, 2008, pp. 146-148.
[27] R. Burduk, Computer recognition systems 4. Berlin: Springer, 2011, p. 631.
[28] C. Mohn et al, "The rapid release of corticosterone from the adrenal induced by ACTH is
mediated by nitric oxide acting by prostaglandin E2", Proceedings of the National Academy
of Sciences, vol. 102, no. 17, pp. 6213-6218, 2005.

50
[29] J. Djordejevic, G. Cvijic and V. Davidovic, "Different Activation of ACTH and
Corticosterone Release in Response to Various Stressors in Rats", Physiological Research,
vol. 52, pp. 67-72, 2003.
[30] L. W. Swanson, Brain maps: structure of the rat brain, 1st ed. Amsterdam: Elsevier, 1992.
[31] S. Bouret, "Formation of Projection Pathways from the Arcuate Nucleus of the
Hypothalamus to Hypothalamic Regions Implicated in the Neural Control of Feeding
Behavior in Mice", Journal of Neuroscience, vol. 24, no. 11, pp. 2797-2805, 2004.
[32] D. Walker and L. Lue, "Immune phenotypes of microglia in human neurodegenerative
disease: challenges to detecting microglial polarization in human brains", Alzheimer's
Research & Therapy, vol. 7, no. 1, 2015.
[33] T. Ferreira, "ImageJ User Guide - IJ 1.46r", Imagej.nih.gov, 2017. [Online]. Available:
https://imagej.nih.gov/ij/docs/guide/index.html. [Accessed: 26- Nov- 2017].
[34] B. Kopp, D. Wick and J. Herman, "Differential effects of homotypic vs. heterotypic chronic
stress regimens on microglial activation in the prefrontal cortex", Physiology & Behavior,
vol. 122, pp. 246-252, 2013.
[35] E. Wohleb et al, "β-Adrenergic Receptor Antagonism Prevents Anxiety-Like Behavior and
Microglial Reactivity Induced by Repeated Social Defeat", Journal of Neuroscience, vol.
31, no. 17, pp. 6277-6288, 2011.
[36] Z. Soltys et al, "Morphology of reactive microglia in the injured cerebral cortex. Fractal
analysis and complementary quantitative methods", Journal of Neuroscience Research,
vol. 63, no. 1, pp. 90-97, 2001.
[37] S. Das Sarma, K. Chatterjee, H. Dinda, D. Chatterjee and J. Das Sarma,
"Cytomorphological and Cytochemical Identification of Microglia", ISRN Immunology,
vol. 2013, pp. 1-10, 2013.
[38] L. Lajos és mtsai, Szövettani és sejtbiológiai vizsgálómódszerek. Budapest: ELTE TTK
Biológiai Intézet, 2012, pp. 15-16, 100-102.
[39] H. Kettenmann, U. Hanisch, M. Noda and A. Verkhratsky, "Physiology of Microglia",
Physiological Reviews, vol. 91, no. 2, pp. 461-553, 2011.
[40] D. Nayak, "Microglia Development and Function", Figure 2 a. 2014.
[41] F. Crews et al, "Molecular Mechanisms of Alcohol Neurotoxicity", Figure 1., 1993.
[42] "IHC detection systems: Advantages and Disadvantages: Novus Biologicals",
Novusbio.com, 2017. [Online]. Available: https://www.novusbio.com/ihc-detection.
[Accessed: 20- Nov- 2017].
[43] Y. Date et al, "Orexins, orexigenic hypothalamic peptides, interact with autonomic,
neuroendocrine and neuroregulatory systems", Figure 2/F, Figure 2/I, 1999.

51
9. Mellékletek
---------------------------INFO---------------------------
Name of the image: 10_Iba1_PVN1b_20x.tif
Size of the image: 1151x805 pixel
CA: covered area
CEA: covered environmental area
Solidity: CA/CEA
Last modified: 24-Nov-2017 20:31:19
---------------------------DATA---------------------------
Number of cells: 30
Total CA: 0.378644 mm^2; percentage: 2.848185 %
Nearest neighbor distance: 111.638475
Number of extracted cells for further measurements: 30
Total CEA: 1.550620 mm^2; percentage: 11.663852 %
Cell CA [mm^2] CEA [mm^2] Solidity
1 0.008968 0.029901 0.299904
2 0.012655 0.040275 0.314214
3 0.016529 0.063820 0.258993
4 0.015195 0.063318 0.239973
5 0.011808 0.053260 0.221713
6 0.012741 0.114210 0.111558
7 0.011579 0.064265 0.180174
8 0.018681 0.084108 0.222108
9 0.007705 0.019599 0.393119
10 0.011005 0.032814 0.335374
11 0.012741 0.052901 0.240846
12 0.011966 0.044508 0.268859
13 0.008996 0.018911 0.475721
14 0.015855 0.073634 0.215316
15 0.009613 0.034694 0.277089
16 0.018150 0.060735 0.298842
17 0.013860 0.087996 0.157509
18 0.010517 0.027046 0.388859
19 0.018523 0.067924 0.272708
20 0.007002 0.014262 0.490946
21 0.016830 0.041681 0.403787
22 0.014735 0.059688 0.246875
23 0.013788 0.070922 0.194416
24 0.019685 0.133221 0.147765
25 0.010431 0.032039 0.325571
26 0.009599 0.030891 0.310729
27 0.006471 0.011005 0.588005
28 0.015955 0.052973 0.301192
29 0.009039 0.031494 0.287016
30 0.008006 0.039228 0.204097
mean 0.012621 0.051711 0.289109
1. melléklet: Egy felvétel részletes elemzésének eredménye.