Embed Size (px)
Citation preview

Mirjana Milošević • Borislav Kobiljski
SEMENARSTVO I
Kljuca žila nevidljivka u nedrima,zrno se klicom rodnom nudi,odušak nadošloj snazi išće.
Život ustreptali žari se u njemuupinje se božanski oganj da ne ugasi.
Brojem se od smrti brani.
Brekće srčano oklopnik mali,štedro mu bije snaga,
izobilje navešćuje.Na put bez povratka se sprema,
život i smrt u istoj šaci nosi,ništeći sebe jednoga stotinu drugih rađa.
(Izvod iz pesme „Seme“ napisane za monografiju, autora Milana Tripkovića, koji će nam svima ostati u sećanju kao poeta i dobar čovek)

Izdavač/Published by:INSTITUT ZA RATARSTVO I POVRTARSTVO, NOVI SAD
Urednici/Editors:prof. dr Mirjana Miloševićprof. dr Borislav Kobiljski
Recenzenti/Reviewers:akademik Rudolf Kastori, redovni profesor u penziji
na predmetu Fiziologija biljaka, Poljoprivredni fakultet, Novi Sad
prof. dr Jovan Crnobarac, redovni profesor na predmetu Posebno ratarstvo, Poljoprivredni fakultet, Novi Sad
prof. dr Mile Ivanović, redovni profesor Biološkog fakulteta Univerziteta u Beogradu
Tehnički urednici/Technical editors:dr Ana Marjanović-Jeromela
Tanja Vunjak-Kvaić
Lektori/Proof-readers:Tamara Šljivić
Dušanka Stojšić
Korice/Cover design:Aleksandar Vojisavljević
Prepress:Grafički Atelje Abraka Dabra, Novi Sad
Press:SP Print, Novi Sad
Tiraž/Printed in:1.000
ISBN 978-86-80417-30-1
Štampanje monografije sufinansiralo je Ministarstvo prosvete i nauke Republike Srbije
CIP - Kaтaлогизациjа у публикациjиБиблиотека Матице српске, Нови Сад631.53.02 SEMENARSTVO. [Vol.] 1 / [autori Mirjana Milošević ... et al.] ; urednici Mirjana Milošević, Borislav Kobiljski. - Novi Sad : Institut za ratarstvo i povrtarstvo , 2011 (Novi Sad : SP Print). - 397 str. : ilustr. ; 25 cmTitaž 1.000. - Bibliografija.ISBN 978-86-80417-30-11. Милошевић, Мирjана [aутор] [уредник]а) СеменарствоCOBISS.SR-ID 267980039

Mirjana Milošević Borislav Kobiljski
SEMENARSTVO
Novi Sad, 2011
I

Autori/Authors:
Prof. dr Mirjana Milošević
Prof. dr Borislav Kobiljski
Mr Gojko Mladenović
Dr Milka Vujaković
Dipl. ing. – master Dušica Jovičić
Mr Maja Ignjatov
Mr Dragana Petrović
Dr Ksenija Taški-Ajduković
Dr Nevena Nagl
Dr Zorica Nikolić
Dr Ankica Kondić-Špika

5
PREDGOVOR
Semenarstvo je, pored oplemenjivanja, jedna od najznačajnijih gra-na poljoprivrede. Za razumevanje samog procesa proizvodnje, sertifika-cije, metoda ispitivanja semena, biotehnoloških metoda koje se prime-njuju u njegovoj proizvodnji, neophodan je veliki fond znanja. Upravo pomenuto znanje i iskustvo ugradilo je 56 naučnih radnika Instituta za ratarstvo i povrtarstvo u pisanje monografije Semenarstvo.
Monografija Semenarstvo je sastavljena iz tri toma koja čine jednu celinu:
I tom čini opšte osnove semenarstva, s akcentom na značaj evrop-skih integracija na tokove domaćeg semenarstva;
II tom sadrži poglavlja iz posebnog semenarstva koja se odnose na proizvodnju semena ekonomski najznačajnijih ratarskih biljnih vrsta kao što su pšenice, kukuruz, soja, suncokret, šećerna repa, krmno bije, konoplja;
III tom se odnosi na posebno semenarstvo u kom je data tehnologi-ja proizvodnje semena povrtarskih biljnih vrsta.
Monografija Semenarstvo je tako koncipirana da je mogu koristiti proizvođači semena, kompanije koje se bave doradom semena, njegovim prometom, studenti i svi oni koji se interesuju za seme i semenarstvo.
Recenzenti, naši eminentni naučni radnici, izrazili su svoje mišlje-nje o monografiji Semenarstvo, dajući pozitivnu ocenu napisanom tekstu. Izvodi iz recenzija glase:
„Ovaj rukopis predstavlja dobro komponovanu, harmoničnu celinu sa svim elementima jedne monografije posvećene tako značajnom pro-blemu kao što je semenarstvo“ (prof. dr Rudolf Kastori).
„Jasno je da iza ovako kompleksno pripremljenog rukopisa stoji višegodišnji uloženi trud i napor urednika i autora teksta da čitaocima približe ovu složenu oblast sa naučnog, tehnološkog, ali i privrednog sta-novišta“ (prof. dr Mile Ivanović).
„Sadržaj ove monografije će biti od velikog značaja ne samo nauč-nim radnicima i specijalistima iz oblasti semenarstva i oplemenjivanja bi-ljaka, već i studentima i doktorantima da obogate svoja saznanja iz oblasti semenarstva“ (prof. dr Jovan Crnobarac).U Novom Sadu, 01. 12. 2011. Autori

6
SADRŽAJ – I
OPŠTE SEMENARSTVO/GENERAL SEED SCIENCE . . . . . . . . . . . . . . . . . . .9Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10Ekonomski značaj semenarstva . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12Definicija i zadaci semenarstva . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 17Izvori genetičke varijabilnosti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 19Pojam sorte i njeno priznavanje . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36Stručni nadzor nad proizvodnjom semena . . . . . . . . . . . . . . . . . . . . . . . . . 61Organizacije koje se bave semenarstvom . . . . . . . . . . . . . . . . . . . . . . . . . . . 76Međunarodne organizacije koje se bave semenarstvom . . . . . . . . . . . . . . 76Organizacija za hranu i poljoprivredu . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76Međunarodna organizacija u trgovini semenom . . . . . . . . . . . . . . . . . . . . 77Međunarodna pravila u trgovini semenom . . . . . . . . . . . . . . . . . . . . . . . . . 78Pravila i postupci u trgovini semenom namenjenog setvi . . . . . . . . . . . . . 78Međunarodna unija za zaštitu novih biljnih sorti. . . . . . . . . . . . . . . . . . . . 79Učestalost postkontrolnih testova kod sertifikovanog semena iz prethodne godine. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86Međunarodna organizacija za ispitivanje kvaliteta semena. . . . . . . . . . . . 88Međunarodna organizacija za standardizaciju . . . . . . . . . . . . . . . . . . . . . . 93Evropska unija . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94Razlog za članstvo Srbije u EU. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 94Šta podrazumevaju EU integracije? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 95Proces prilagođavanja sistemu EU . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 97Sektori koje pokriva Zajednička poljoprivredna politika (ZPP). . . . . . . . 98Reforme sektora semenarstva . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99Evropska semenarska asocijacija (ESA) . . . . . . . . . . . . . . . . . . . . . . . . . . . 100Semenarska asocijacija Srbije. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 102Zakonski propisi u oblasti semenarstva . . . . . . . . . . . . . . . . . . . . . . . . . . . 104Ekologija semena . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 116Agrotehničke mere u proizvodnji semena . . . . . . . . . . . . . . . . . . . . . . . . . 131Skladištenje semena. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 168
ISPITIVANJE KVALITETA SEMENA . . . . . . . . . . . . . . . . . . . . . . . . . . . . .207Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 208Uzorkovanje semena . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 209Pribor za uzorkovanje semena . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211Masa prosečnog uzorka . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 215Priprema radnog uzorka u laboratoriji . . . . . . . . . . . . . . . . . . . . . . . . . . . . 216Metod mehaničkog deljenja uzorka . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 216Metod ručnog deljenja uzoraka . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 219

7
Ispitivanje čistoće semena. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 222Ispitivanje klijavosti semena. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225Mirovanje semena . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 225Metode za prekidanje mirovanja semena. . . . . . . . . . . . . . . . . . . . . . . . . . 228Podloge za ispitivanje klijavosti semena. . . . . . . . . . . . . . . . . . . . . . . . . . . 230Ocena ponika . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 234Ispitivanje sadržaja vlage u semenu. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 241Određivanje mase 1000 semena . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 245Ispitivanje životne sposobnosti semena . . . . . . . . . . . . . . . . . . . . . . . . . . . 246Fiziološki testovi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 249Biohemijski testovi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 253
ISPITIVANJE ZDRAVSTVENOG STANJA SEMENA . . . . . . . . . . . . . . . 261Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 263Metode za ispitivanje zdravstvenog stanja semena . . . . . . . . . . . . . . . . . . 266Metode identifikacije bez prethodne inkubacije semena. . . . . . . . . . . . . 267Metode uz prethodnu inkubaciju semena . . . . . . . . . . . . . . . . . . . . . . . . . 269Novije metode za ispitivanje patogena . . . . . . . . . . . . . . . . . . . . . . . . . . . . 282
PRIMENA GENETSKIH MARKERA U IDENTIFIKACIJI I ODREĐIVANJU GENETSKE ČISTOĆE SORTI I HIBRIDA. . . . . . . . .299
Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 301Sortne primese . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 301Morfološki markeri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 302Biohemijski markeri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 303DNK markeri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 317
GENETIČKI MODIFIKOVANE BILJNE VRSTE – METODE ZA TESTIRANJE . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .337
Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 339Metode za testiranje genetički modifikovanih organizama . . . . . . . . . . . 341Organizacija laboratorije za GMO analize i okruženje . . . . . . . . . . . . . . 343Standardizacija metoda za testiranje GMO . . . . . . . . . . . . . . . . . . . . . . . . 344Referentni materijal i ograničenja u GMO detekciji i kvantifikaciji. . . . 345Procedura analize genetičke modifikacije PCR metodom. . . . . . . . . . . . 347Izolacija DNK. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 348PCR metode . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 349Multipleks PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 352Nested PCR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353Real Time PCR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 354Razvoj metoda u budućnosti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 355

8
Monografija - Semenarstvo
GENOTIPSKA KARAKTERIZACIJA GAJENIH BILJAKA – DNK FINGERPRINTING . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 359
Uvod . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 361Istorijat . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 362DNK fingerprinting tehnike. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363Dominantni markeri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363Kodominantni markeri . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 365Markeri za detekciju polimorfnosti organelarne DNK . . . . . . . . . . . . . . 366Tehnologije za brzu detekciju identiteta. . . . . . . . . . . . . . . . . . . . . . . . . . . 367DNK barkoding . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 368Poređenje različitih marker sistema i izbor tehnike za DNK fingerprinting. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 370Primena DNK fingerprintinga. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 373Identifikacija genotipa. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 373Identifikacija sorti . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 374Očuvanje biodiverziteta . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 377Karakterizacija germplazme. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 378Osnovne kolekcije (core collections) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 381Upravljanje biodiverzitetom. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 382Perspektive primene DNK fingerprintinga . . . . . . . . . . . . . . . . . . . . . . . . 388

Mirjana Milošević, Borislav Kobiljski, Gojko Mladenović
Opšte semenarstvo

10
UVOD
U narednih 25 godina doći će do povećanja broja stanovnika, urba-nizacija će oduzeti još jedan deo poljoprivrednog zemljišta, dok su izvori novih obradivih površina limitirani. Zemljište na poljoprivrednim gaz-dinstvima postaje degradirano u mnogim zemljama. Napred navedeno upućuje na to da je potrebno naći način, na globalnom nivou, kako da se obezbedi dovoljno hrane za ljude i životinje i kako da se koriste obnovljivi izvori energije. To će svakako zahtevati nov pristup biljkama, kako sa bio-loškog tako i ekonomskog stanovišta. U isto vreme, razvijaju se novi pravci istraživanja genoma primenom novih savremenih tehnologija, kako bi se stvorile nove biljne sorte boljih genetičkih i agronomskih karakteristika.
Organizmi koji vrše fotosintezu su sistemi koji obezbeđuju život na planeti zemlji. Oni su primarni izvor energije, sastojci hraniva, gradivne supstance ćelija za veliki broj drugih organizama. Jedinstveni proces foto-sinteze rezultira stvaranjem biomase potrebne svim živim organizmima. To znači da su biljke srce našeg lanca ishrane. Milenijumima je čovečan-stvo prolazilo kroz proces klasičnog oplemenjivanja da bi razvilo biljke koje više odgovaraju njihovim potrebama. Nove tehnologije će pronaći metode tačnije i efikasnije identifikacije gena koji učestvuju u povećanju nivoa proizvodnje i kvaliteta novostvorenih sorti, pa samim tim i kvalite-ta semena, novih gena koji kod biljaka regulišu tolerantnost na biotičke i abiotičke činioce, ili bolje korišćenje inputa. Nove tehnologije će pomoći istraživačima da okarakterišu i koriste genetičku divergentnost i genetičke resurse. Sekvence gena su postale izvor osnovnog znanja o gajenim bilj-kama. Sekvencioniranje gena obezbeđuje ispitivanje genoma, daje kom-pletan uvid u gene za buduća istraživanja. Ono otvara nove prespektive za ispitivanja koja će objasniti razlike između genotipova, vrsta i njihove filogenične odnose čak i kod vrlo udaljenih vrsta (Kolektiv autora, 2003).
Modeliranje i istraživanja vezana za simulaciju nepovoljnih uticaja spoljašnje sredine će doprineti da se predvidi ponašanje biljaka u određe-nom okruženju, a na taj način će pomoći oplemenjivačima da nađu način za stvaranje biljaka prilagođenih uslovima koji ih očekuju u budućnosti (Milošević i sar., 2010).

11
Opšte semenarstvo
Hrana je osnovni preduslov da bi čovek održao svoj opstanak u vre-menu i prostoru. Iz tog razloga neophodno je da se on sve više okreće prirodi. Tako posmatrano, poljoprivredna proizvodnja, a posebno proi-zvodnja semena ima izuzetan značaj, jer se 95% hrane proizvodi od bi-ljaka koje se razmnožavaju semenom. Za proizvodnju dovoljnih količina semena za podmirenje potreba za setvu određene biljne vrste, organizuje se proizvodnja i dorada kvalitetnog, sortnog semena. Te aktivnosti se na-zivaju semenarstvo.

12
Semenarstvo
EKONOMSKI ZNAČAJ SEMENARSTVA
Semenska proizvodnja se ubraja među najprofitabilnije delatnosti u oblasti poljoprivrede. Površine pod semenskom proizvodnjom su male u odnosu na komercijalnu proizvodnju, ali su finansijski efekti značaj-no veći. Podaci koji su naznačeni u tabeli 1 ukazuju na vrednost proi-zvodnje semena u odabranim državama od strane Svetske semenarske federacije (International Seed Federation – ISF) u 2009. godini. Merilo odabira država je bio ekonomski doprinos semenarstva domaćem trži-štu (ISF, 2009).
Ukupna vrednost semenske proizvodnje u svetu iznosi 32 milijarde dolara. Srbija zauzima zavidno visoko mesto u ostvarenju finansijskih sredstava od prometa semenom, koje iznosi 120 miliona dolara godišnje (tab. 1).
Klimatski i edafski uslovi, tradicija, određuju da li će se u nekoj ze-mlji razvijati uspešno semenarstvo, odnosno proizvodnja semena ra-tarskih i/ili povrtarskih biljnih vrsta. Veličina zemlje nema veliki uticaj na sredstva koja se ostvaruju u prometu semenom. Holandija se ubraja, gledano na površinu zemlje, u manje zemlje u Evropi, ali je zato promet semena koji ostvari najveći. SAD, Francuska, Nemačka, Čile, Kanada, Meksiko, Mađarska, Danska, Italija, Kina čine u najveće izvoznike se-mena u svetu, kako ratarskih tako i povrtarskih biljnih vrsta. Srbija se nalazi na 34. mestu najuspešnijih zemalja po finansijskim efektima koje je ostvarila izvozom semena u 2009. godini (tab. 2).
Pored toga što proizvodnja i izvoz semena predstavljaju jednu od najvažnijih ekonomskih pokazatelja u poljoprivredi, semenska industri-ja učestvuje u zapošljavanju velikog broja drugih delatnosti. Pre svega je to mašinska industrija koja proizvodi mašine za proizvodnju i doradu semena, skladišne kapacitete, laboratorijsku opremu za ispitivanje kva-liteta semena i dr (sl. 1, 2).

13
Opšte semenarstvo
Tabela 1. Ostvarena vrednost od semenarstva na domaćem tržištu u odabranim zemljama (ISF, 2009)Table 1.The realized value of seed production on domestic market in selected countries (ISF, 2009)
Vrednost na domaćem tržištu (mil.dolara)Value of domestic market(mil.of dollars)
SAD/USA 8,500 Maroko/Marocco 140Kina/China 4,000 Egipat/Egipt 140Francuska/France 2,150 Bugarska/Bulgaria 120Brazil /Brasil 2,000 Čile/Chile 120Indija/India 1,500 Srbija/Serbia 120Japan/Japan 1,500 Nigerija/Nigeria 120Nemačka/Germany 1,500 Slovačka/Slovakia 110Italija/Italy 1,000 Novi Zeland/New Zeland 100Argentina/ Argentina 950 Švajcarska/Switzerland 90Kanada/Canada 550 Paragvaj/Paraguay 80
Ruska Federacija/Russian Federation 500 Portugalija/Portugal 80
Španija/Spain 450 Irska/Ireland 80Australija/Australia 400 Alžir/Algeria 70Koreja/Korea 400 Urugvaj/Urugvay 70UK/UK 400 Kenija/Kenia 60Meksiko/Mexico 350 Iran/ Iran 55Poljska/Poland 350 Izrael/Israel 50Turska/Turkey 350 Tunis/Tunisia 45Tajvan/Taiwan 300 Kolumbija/Columbia 40Južna Afrika/South Africa 300 Bolivija/Bolivia 40Mađarska/Hungary 300 Slovenija/ Slovenia 40Holandija/Netherland 300 Zimbabve/Zimbabwe 30Češka Republika/Czech Republic 300 Peru/Peru 30
Danska/Denmark 250 Libija/Libia 25Bangladeš/Bangladesh 250 Saudijska Arabija/Saudi Arabia 20Grčka/Greece 240 Zambija/Zambia 20Švedska/Sweden 240 Ekvador/Ecuador 15Rumunija/Romania 220 Tanzanija/Tanzania 15Belgija/Belgium 190 Malavi/Malawi 10Finska/Finland 160 Uganda/Uganda 10
Austrija/Austria 150 Dominikanska Republika/Republica Dominicana 7
Ukupno /Total = 32,002
Podaci čine kompilaciju zvaničnih statističkih podataka i izveštaja o međunarodnom prometu semena ISFISF compilation based on official statistics and international seed trade reports

14
Semenarstvo
Tabela 2. Izvoz semena u odabranim zemljama u 2009. godini (ISF; 2009)Table 2. Seed export of selected countries over 2009
Država/Country
Ratarske biljkeField crops
quantitymetičke tone/metric tonnes
Ratarske biljke
Field Crops value
miloni dolara/USD millions
Povrtarske biljke
Vegetable Crops quantitymetičke tone/metric tonnes
Povrtarske biljke
Vegetable Crops value
miloni dolara/USD millions
Ukupna vrednost
Total value
Holadnija/Netherland 151396 241 11361 1058 1299SAD/USA 225300 746 18495 432 1178Francuska/France 330675 884 9352 278 1162Nemačka/Germany 82466 458 1261 48 506Čile/Chile 52500 261 1912 109 370Kanada/Canada 170200 273 6833 82 355Meksiko/Mexico 35280 244 774 11 255Mađarska/Hungary 77241 221 1250 14 235Danska/Denmark 79173 168 8916 55 223Italija/Italy 80363 123 8940 94 217Argentina/Argentina 140000 163 269 9 172Belgija/Belgium 24102 160 760 4 164Kina/China 36600 72 4130 68 140Austrija/Austria 32075 115 114 3 118Japan/Japan 4215 30 1311 87 117Španija/Spain 40261 62 1620 47 109Izrael/Israel 5821 14 4100 83 97Rumunija/Romania 60991 86 0 0 86Australija/Australia 31350 65 1107 18 83Novi Zeland/ New Zealand 5500 32 6500 32 64Južna Afrika/ South Africa 7217 48 1595 13 61UK/UK 5802 40 1050 21 61Brazil/Brasil 8319 46 169 8 54Tajland/Thailand 1200 3 1896 44 47Turska/Turkey 20000 36 532 11 47Češka Republika/ Czech Republic 67228 41 178 4 45Poljska/Poland 82923 39 450 4 43Slovačka/Slovakia 46324 38 0 0 38Švedska/Sweden 6203 31 96 4 35Indija/India n.a. 16 3870 17 33Republika Koreja/ Rep. of Korea 2320 6 410 20 26Švajcarska/Switzerland 30200 22 12 2 24Bugarska/Bulgaria 38064 18 2000 5 23Srbija/Serbia 30000 19 100 1 20
Naznaka: Odnos izmene valute 1evro=1,4 dolara; n.a.: informacija nedostupnaNote: Conversion rate: 1 Euro = 1.4 US Dollars; n.a: information not available

15
Opšte semenarstvo
Prevoz semena upošljava transportnu industriju, štamparska indu-strija učestvuje u proizvodnji vreća i obezbeđenja ostalih načina pako-vanja semena, štampanju etiketa i dokumentacije koja prati seme.
Izuzetnu dobit od semenske industrije ostvaruje hemijska industri-ja, jer su sredstva za hemijsku zaštitu semena ili zakonom propisana, ili neophodna zbog prisustva štetnih organizama u zemljištu, te se troše u velikim količinama.
U Srbiji ima 5,1 miliona hektara poljoprivrednog zemljišta, od čega je 66% obradivog poljoprivrednog zemljišta (sl. 3). Za setvu pomenu-tih površina potrebno je oko 300.000 tona semena. Navedene potrebe mogu se ostvariti proizvodnjom visoko kvalitetnog semena uz poštova-nje pravila i normi koje su zacrtane međunarodnim propisima i doma-ćom zakonskom regulativom.
Slika 1. Prečistači semena Figure 1. Seed cleaners
(http://spectrumindustries.tradeindia.com/Exporters_Suppliers/
Exporter1583.398659/Vibro-Cleaner.html)
Slika 2. Aparati koji se koriste u ispitivanju
kvaliteta semena Figure 2. The devices used
in seed quality testing(foto: Milošević)

16
Semenarstvo
Slika 3. Struktura poljoprivrednog zemljišta u SrbijiFigure 3. Strucutre of agricultural land in Serbia
(Milošević, 2009)
Osnova razvoja poljoprivrede je stvaranje novih biljnih sorti i pri-mena međunarodno usaglašenog sistema u proizvodnji semena. Doma-će naučne institucije ostvaruju značajne uspehe u oblasti oplemenjiva-nja poljoprivrednog bilja (preko 1.500 novostvorenih sorti) koje po rod-nosti i kvalitetu spadaju u vodeće u svetu (posebno kukuruz, pšenica, suncokret, soja). Uspešan razvoj oplemenjivanja biljaka prati intenzivan razvoj proizvodnje semena za domaće potrebe i za izvoz, s obzirom na već izgrađene kapacitete za doradu semena, povoljne zemljišno–klimat-ske uslove i obučene stručne kadrove. Znatne količine semena kukuru-za, suncokreta, pšenice se izvoze svake godine.
Pašnjaci/Grassland28%
Višegodišnji zasadi/ Permanent crops
6%
Obradivo zemljište/ Arable land
66%

17
Opšte semenarstvo
DEFINICIJA I ZADACI SEMENARSTVA
Definicija semenarstva
Semenarstvo u najširem smislu reči obuhvata proizvodnju, doradu i promet semena. Ono podrazumeva niz faza koje čine jednu celinu, a to su:
– ispitivanje zemljišta (organsko semenarstvo),– zasnivanje i gajenje semenskog useva, – kontrola semenskih useva u polju, – ubiranje semena, – dorada semena, ispitivanje i utvrđivanje semenskih kvaliteta, – skladištenje i čuvanje, – distribucija i promet semena i – njegova upotreba.
Semenarstvo se može smatrati i završnom fazom oplemenjivanja biljaka, jer novostvorene i priznate sorte treba širiti u proizvodnji, a pri tome održati, ili čak poboljšati njihove proizvodne osobine i biološku vrednost semena (Milošević i sar., 1996). Posedovanje dovoljnih koli-čina semena od sorti i hibrida visokog potencijala rodnosti i drugih dobrih agronomskih osobina znači bogatstvo jedne zemlje, neproce-njivu vrednost i više od toga, nacionalni ponos i ugled jedne države (Sarić, 1971).
Polazeći od osnovnih zadataka koje ima, semenarstvu se može dati ovakva definicija: „Semenarstvo je deo poljoprivredne nauke, struke i proizvodnje koji na organizovan način osigurava masovno umnožavanje semena poljoprivrednih biljaka, uz maksimalno očuvanje sortne čistoće i semenskih kvaliteta” (Mihaljev i Dokić, 1986).

18
Semenarstvo
Zadaci semenarstva
Po Mihaljevu i Dokiću (1986) osnovni zadatak semenarstva je pre svega umnožavanje semena priznatih sorti. U momentu priznavanja sor-te, raspolaže se malim količinama semena. U cilju promovisanja i širenja sorte potrebno je seme umnožiti u onim količinama koje su dovoljne za pomenute svrhe, a kasnije i za podmirenje potreba poljoprivredne proi-zvodnje. Interes oplemenjivača, a i distributera, je da se dobre sorte što pre rašire u proizvodnji.
Iz pomenutih razloga, vlasnici sorti ulažu značajna materijalna sredstva u promociju novostvorenih sorti putem propagandnog materijala, kataloga, demonstracionih ogleda. Demonstracioni ogledi se postavljaju na vidnom mestu, obično pored glavnog puta, kako bi bili što dostupniji široj javno-sti, odnosno svim zainteresovanim za saznanja o kvalitetu novih sorti. Dani polja, koje organizuje vlasnik novih sorti, jedan je od najboljih načina za promociju nove sorte, jer se pored vizuelnog utiska, dobijaju odgovori na sva pitanja vezana za njene karakteristike. Propagiranje novih sorti i njihovu prodaju na terenu vrše promoteri, koji pored toga što moraju da imaju boga-to znanje o sortama koje nude u prodaji, treba da su pristupačni i snalažnjivi u svom poslu.

19
Opšte semenarstvo
IZVORI GENETIČKE VARIJABILNOSTI
Genetički diverzitet kao izvor varijabilnosti
Osnovu za dobre oplemenjivačke programe i stvaranje visoko pri-nosnih sorti čini genetički diverzitet. Genetički diverzitet zavisi od bilja-ka, odnosno od biljnog genoma, koji je najkompleksniji živi sistem. On je sastavljen od tri interaktivna genoma. Osim nuklearnog (jedarnog) genoma, ostatak genetičkog sistema je smešten u plastidima i mitohon-drijama. Ove organele su semiautohtona tela i imaju svoje organizacio-ne i funkcionalne celine, ali ne mogu samostalno da sintetizuju sve svoje proteine. Nuklearni genom igra važnu ulogu u biogenezi organela.
Ispitivanje genetičkog diverziteta zasnovano je danas na ispitivanju hromozomskih lokacija gena (sl. 4), gena za povećanje prinosa, kao i
drugih kompleksnih karakteristika važnih za poljoprivredu. Za ispitiva-nje hromozomskih lokacija gena kori-ste se posebne mape vezanih gena, tzv. „linkidž mape“. „Linkidž mape“ daju frekvenciju rekombinacija (frekvenci-ja rekombinacije je frekvencija broja spajanja lokusa na hromozomima ili genima tokom mejoze) između gena na hromozomima i na taj način čine mogućim ispitivanje lokacija gena koji određuju karakteristike biljaka važne za poljoprivredu.
Tehnike molekularnog kloniranja i sekvencioniranja dezoksiribonukle-inske kiseline (DNK) su omogućile izučavanje strukture gena na nivou nu-kleotida. Poznavanje strukture, orga- nizacije i ispoljavanja osobina biljnog ge-noma postignuto je upotrebom tehnike
Slika 4. Hromozomi sadrže gene nosioce naslednih osobina
Figure 4. The chromosomes contain gene carriers of hereditary traits (http://www.ubthenews.com/
topics_summaries.htm)

20
Semenarstvo
rekombinantne DNK. Ova tehnika je omogućila izolaciju i karakteriza-ciju specifičnih delova DNK kloniranjem sekvenci DNK u ćelijama bak-terija u kojima mogu biti umnoženi do potrebne količine, a sa kojima se može vršiti željena analiza (http://www.molecular-plant-biotechnology.info/nuclear-genome/plant-genome.htm). Ispitivanje genoma omogu-ćava konačno sagledavanje genetičkog potencijala kako gajenih biljaka tako i njihovih divljih srodnika, kako bi divlji srodnici bili eventualno iskorišćeni za istraživanje ili gajenje.
Karakterizacija germplazme znači i upotrebu DNK „fingerprinting” tehnika za precizno ustanovljavanje, identifikaciju i kvantitativno odre-đivanje genetičkog diverziteta. Ova ispitivanja su važna zbog opadanja genetičkog diverziteta, kao odgovor na klimatske promene, promena kod populacija patogena ili poljoprivredne prakse. U ispitivanjima 105 sorti argentinske pšenice (Triticum aestivum L.) gajenih od 1932. do 1995. godine izvršena je karakterizacija uzoraka upotrebom SSR (Sim-ple Sequence Repeat) i AFLP (Amplified Fragment Length Polymorphism) markera. Podaci molekularnih ispitivanja su korišćeni da se ustanovi genetički diverzitet sorti koje su upotrebljene u oplemenjivačkim pro-gramima, kao i da se dokaže da se vremenom izgubio deo genetičkog diverziteta, u odnosu na onaj koji se ranije koristio u oplemenjivačkim programima (genetička erozija). Ustanovljene su razlike u dobijenim rezultatima, kod obe primenjene tehnike, između oplemenjivačkih pro-grama sa velikim razlikama u broju primenjenih sorti. Nisu pronađene razlike u genetičkom deverzitetu između sorti stvorenih šezdesetih go-dina i onih stvaranih tri decenije kasnije. Svaki reprezentativni uzorak je sadržao kompletan diverzitet argentinske germplazme (Manifesto et al., 2001).
Vrednost gena i genotipa
Geni i njihove karakteristike, kao osnovica genetičkog diverziteta, vredni su zbog rezultata koji ostvaruju, podrazumevajući pod tim agro-nomske karakteristike kao što je na primer rezistentnost na štetne orga-nizme. Pored toga, od izuzetne su važnosti otpornost na abiotičke stre-sove, kao što su visoke temperature, suša, iskorišćavanje hranljivih ma-terija i visok sadržaj proteina ili specifičan sadržaj masnih kiselina u ulju.

21
Opšte semenarstvo
Oplemenjivači su izmenili tip lista kod graška, iz klasičnog u vitice (aphila tip), da bi obezbedili uspravan položaj biljke. To je delom oboga-ćivanje genofonda, a delom potreba da se ugradi pozitivno svojstvo kod ove biljne vrste (sl. 5 ).
Postoje podaci o stvaranju sorti pšenice efikasnih u korišćenju azo-ta, tj. sorti skromnih u zahtevima prema azotu. Efikasnost ishrane azo-tom meri se povećanjem prinosa ро jedinici primenjene doze azotnog đubriva. Efikasnost đubrenja azotom (EFN) realizuje se kroz usvajanje azota iz đubriva (EAN) i njegovo iskorišćavanje u biljci (EUN) u formi-ranju prinosa i predstavlja njihov proizvod:
EFN = EAN x EUN = ANNf
x YgAN
= YgNf
Na osnovu ove relacije prinos (Yg) se može definisati kao proizvod iz akumulacije i iskorišćavanja azota u biljci:
Yg = AN x EUN ( EFN).
Isti prinosi se mogu dobiti slaganjem visokih vrednosti jednog i niskih vrednosti drugog, ili osrednjih vrednosti oba pokazatelja. Obje-dinjavanjem visokih vrednosti oba parametra u novom genotipu, bez narušavanja balansa fizioloških procesa, mоžе se ostvariti dalje pove-ćanje prinosa i efikasnosti đubrenja azotom. Širok dijapazon variranja
Slika 5. Različit tip lista kod graška dobijen u procesu oplemenjivanja (a-aphila tip, b-intermedijarni,c-klasičan tip)
Figure 5. Different types of peas leafs obtained in the process of breeding (a-aphila type, b-intermediate, c-classic type)
(foto: Mikić, A.)
a) b) c)

22
Semenarstvo
ovih parametara ukazu-je da oplemenjivanje u ovom pogledu ima per-spektivu (Đokić, 1995). Korišćenje širokog dija-pazona sorti, koja pose-duje veliku genetičku va-rijabilnost, daje još veće mogućnosti u dobijanju genotipova koji mogu da doprinesu ne samo bo-ljem iskorišćavanju azota nego i mnogim drugim poželjnim osobinama (sl. 6).
Obogaćivanje genetičke varijabilnosti suncokreta na bazi divljih vr-sta usmereno je na povećanje otpornosti prema bolestima, insektima, poboljšanju kvaliteta ulja i otpornosti prema stresu (suši). „Interspeci-es” i „intergenus” hibridizacijom može se ostvariti povećanje genetičke varijabilnosti. Sadašnja kolekcija u Institutu za ratarstvo i povrtarstvo u Novom Sаdu, koji održava i FAO kolekciju suncokreta, od preko 5.000 inbred linija i preko 1.000 populacija divljih vrsta su garancija za uspe-šan rad u oplemenjivanju suncokreta u vremenu koje dolazi (Škorić i Kovačev, 1995).
Neki genotipovi su od neprocenjive vrednosti za poljoprivrednike u predelima siromašnim genofondom, posebno pojedine kombinacije gena kod dobro adaptiranih populacija, koje je nemoguće ponovo stvo-riti ako se jednom izgube.
Vrednost genetičke varijabilnosti
Za postojanost i eventualno povećanje vrednosti genetičkih resursa od izuzetne je važnosti :
– održavanje genetičke varijabilnosti koja omogućava da se zadrži stabilnost u proizvodnji na poljoprivrednim dobrima na lokalnom, na-cionalnom i globalnom nivou,
Slika 6. Genetički diverzitet između sorata pšeniceFigure 6. Genetic diversity beetween varietys
(foto: Denčić)

23
Opšte semenarstvo
– osiguranje opstanka genetičkog diverziteta za buduće loše klimat-ske uslove, jer genetički diverzitet predstavlja „blagajnu” za potencijalnu vrednost još nepoznatih resursa.
Napred navedeno je razlog za održavanje prirodnog ekosistema i tradicionalnog načina proizvodnje na poljoprivrednim dobrima, jer biljke iz tih staništa mogu da sadrže i iz njih mogu da se razviju važne genetičke osobine.
Uloga individualnih poljoprivrednih proizvođača je velika u očuva-nju genetičkih resursa jer oni, širom sveta, obezbeđuju 15–20% hrane. Kao primer važnosti održavanja starih sorti može poslužiti jedna turska sorta pšenice koja je nosilac gena otpornosti i tolerantnosti na različite rase rđe, gari i drugih gljivičnih patogena. Ovaj gen je korišćen u stva-ranju brojnih novih sorti koje se gaje širom SAD-a. Jedna sorta lucerke, poreklom iz Irana, poslužila je kao izvor gena za otpornost na nemato-de koje predstavljaju izuzetan ekonomski problem u gajenju ove važne krmne biljke.
Divlji srodnici imaju takođe veliki značaj kada se radi o poboljšanju karakteristika sorti, posebno kada su u pitanju štetni organizmi. Kao primer se može uzeti paradajz (Lycopersicon esculentum). Njegovi divlji srodnici su korišćeni kao donori gena za otpornost na gljivična obolje-nja (L. hirsutum, L. pimpinellifolium); rezistentnosti na viruse (L. chilen-se, L. peruvianum); rezistentnost na nematode (L. peruvianum); rezi-stentnost na insekte (L. hirsutum); za kvalitet plodova (L. chmielewskii); i adaptaciju na nepovoljne spoljašnje uslove (L. cheesmanii). Rezisten-tnost krompira na cistične nematode je uneta u krompir (Solanum tu-berosum) iz divljeg srodnika Solanum demissum (William et al., 2007).
Otpornost pšenice na stabljičinu rđu uneta je u pšenicu (Triticum aestivum) iz divljih srodnika Triticum timopheevi i iz Agropyron spp. Neke sorte pšenice su postale tolerantne na sočivastu pegavost prenoše-njem gena otpornosti iz divljeg srodnika Aegilops ventricosa.
Pojava gljivičnog oboljenja Phomopsis skoro je onemogućila gajenje suncokreta u Srbiji sedamdesetih godina prošlog veka (sl. 7). Istraži-vački tim na čelu sa oplemenjivačem Škorićem iz Instituta za ratarstvo i povrtarstvo iz Novog Sada, je napravio pravi podvig u oplemenjivanju. Unošenjem gena divljih srodnika stvorio je hibride visoko tolerantne na ovo obolenje, bez gubljenja dobrih agronomskih svojstava. Osnovu otpornosti na Phomopsis i danas koriste oplemenjivači u celoj Evropi.

24
Semenarstvo
Manipulisanje genetičkim diverzitetom
Čovekove aktivnosti mogu da dovedu do povećanja genetičkog di-verziteta. To se, pre svega, ogleda u njegovom angažovanju na odabiru brojnih biljnih vrsta, sorti, unapređenju tehnologije proizvodnje i dru-gim delatnostima kojima se ljudi bave već više od 10.000 godina. Ukr-štanjem linija roditeljskih parova takođe se menja sastav biodiverziteta. Procenat rekombinacija ostvarenih kod F1 individua mogu varirati od 1% do 50% i uvek su viđeni kao posebni lokusi na razdvojenim hromo-zomima. Najveći procenat rekombinacija se odvija kod roditeljskih pa-rova kod kojih postoji najveća genetička udaljenost na dva lokusa (sl. 8).
Slika 7. Pojava Phomopsis-a na stablu suncokreta (levo), poleganje stabla usled infekcije (desno)
Figure 7. Occurrence of Phomopsis in sunflower stem (left), lodging due to stem infection (right)
(foto: Gulya)
Slika 8. Šema rekombinacije hromozoma Figure 8. Chromosomes recombination scheme
(http://genome.wellcome.ac.uk/ doc_wtd020778.html)

25
Opšte semenarstvo
Oplemenjivački rad i genetički resursi
Osnovni uslov za uspešan oplemenjivački rad je postojanje genetičke varijabilnosti unutar vrste. Ona se oplemenjivačkim radom može obogaćivati ili osi-romašiti. Genetička varijabilnost suncokreta (Helianthus annuus) je dosta uska, a posebno u pogledu gena koji uslovljavaju otpornost prema bolestima (sl. 9). Divergen-tnost se može delimično povećati korišćenjem divljih vrsta putem interspecijes hibridizacije (sl. 10). Rod Helianthus ima 49 vrsta i 19
Slika 9. Gajeni suncokret (H. annuuus) Figure 9. Sunflower plant (H.annuus)
(foto: Milošević)
Slika 10. H. angustifolius, H. tuberosus (s leva na desno gore) Tithonia diversifolia i H. petiolaris (s leva na desno dole)
Figure 10. H. angustifolius, H. tuberosus (from left to right above) Tithonia diversifolia i H. petiolaris (from right to left down) (foto: Sakač, V.)

26
Semenarstvo
podvrsta sa 12 jednogodišnjih i 37 višegodišnjih vrsta i bogatu vаrijabilnost unutar svake vrste, odnosno podvrste. Ova varijabilnost рružа veliku mogućnost za povećanje genetičke varijabilnosti u nared-nom periodu. Interspecies hibridizacija je često оtеžаnа usled različitog broja hromozoma i ploidnosti (2n, 4n, 6n) kod divljih vrsta i prisustva inkompatibilnosti. Iz tog razloga рrevazilaženje problema se može po-stići primenom novih metoda biotehnologije (Škorić i Kovačev, 1995).
Oplemenjivači za svoj rad koriste različite izvore. Najčešće sop-stvene kolekcije namenskog tipa. Tako postoje kolekcije pšenice i sun-cokreta u Institutu za ratarstvo i povrtarstvo u Novom Sadu, kolekcija genotipova kukuruza u Institutu za kukuruz Zemun Polje. Za očuvanje ukupne genetičke varijabilnosti unutar vrste na naučnim osnovama, staraju se banke gena. One vode računa o genetičkim resursima počev od proučavanja terena sa koga ih sakupljaju, kolekcionisanja, čuvanja do razmene informacija i uzoraka.
Oplemenjivači danas raspolažu veoma efikasnim metodama rada i odgovarajućom opemom u procesu stvaranja novih sorti, ali se ne može reći da raspolažu dovoljnim informacijama o genetičkoj varijabilnosti unutar vrsta, posebno u Srbiji, niti da su isti dovoljno iskorišćeni (Pen-čić i sar., 1997). U cilju dobijanja informacija o genetičkoj varijabilnosti danas se koriste savremene metode determinacije DNK (RAPD). Ovaj metod je korišćen za inter i intraspecijes razdvajanje roda Helianthus. Izučavano je oko 35 vrsta i podrvrsta roda Helianthus, kao i 30 inbred linija H. annuus i dva uzorka srodnika i to Titonia speciosa i Simsia foe-tida. Sačinjen je fenogram preko koga su određeni genetički odnosi ga-jenog suncokreta i divljih vrsta. Analiza je pokazala da postoji relativno mala genetička udaljenost između H. annuus i H. laetiflorus, H. salici-folius, H.bolanderi i H. tuberosus, što ukazuje da ti divlji srodnici mogu biti izvor korisnih gena za H. annuus, a koje mogu biti korišćene za po-boljšanje svojstava gajenog suncokreta (Sivolap and Solodenko, 2006).
Najnovija dostignuća u modernoj biotehnologiji, a naročito u pri-meni molekularnih markera, dovela su do razvoja brojnih novih pristu-pa, koji pružaju mogućnost povećanja efikasnosti i ekonomičnosti ople-menjivanja biljaka. Jedan od najznačajnijih pristupa je svakako mar-kerima pomognuta selekcija (Marker Assisted Selection – MAS), koja predstavlja proces gde se markeri koriste pri direktnoj selekciji genetič-ke determinante ili determinanti svojstva od interesa. U osnovi, MAS se bazira na primeni utvrđene i potvrđene povezanosti i međuuslov-

27
Opšte semenarstvo
ljenosti gena, koji kontrolišu ekspresiju nekog svojstava od značaja za oplemenjivački proces i markera. MAS je fokusirana na:
1) povećanje efikasnosti selekcije i to za svojstva koja se nalaze pod kontrolom većeg broja gena i pod jakim su uticajem činioca spo-ljašnje sredine (imaju nisku heritabilnost),
2) selekciju svojstava od ekonomskog značaja, u slučajevima gde je primena uobičajenih ogleda nedovoljno pouzdana, dugotrajna i neekonomična,
3) akumulaciju (piramidiranje) gena otpornosti na prevalentne bo-lesti (Landjeva et al., 2007).
Da bi osigurali optimalnu efikasnost u pogledu troškova, moleku-larni markeri koji se koriste u MAS treba da omoguće efikasan skrining velikih populacija i da imaju visok stepen ponovljivosti u različitim la-boratorijama (Kobiljski i sar., 2008).
U poslednjih dvadesetak godina primenom molekularnih markera mapiran je veliki broj značajnih major gena kao i lokusa za kvantita-tivna svojstva (Quantitative Trait Loci- QTL) kod pšenice (Landjeva et al., 2007). Uprkos činjenici da heksaploidnu pšenicu karakteriše izuzet-no veliki genom (1,8 x 1010 baznih parova), što predstavlja izazov za unapređenje procesa selekcije bilo kojom od dostupnih metoda, među oplemenjivačima pšenice je sve veći broj onih koji veruju da MAS ima kapacitete da poveća efikasnost selekcije u značajnoj meri (Patnaik and Khurana, 2001). Postoji veliki broj primera koji podržavaju ovo gledi-šte, od kojih su verovatno najznačajnija nedavna dostignuća u Australiji, SAD-u, Kanadi i Meksiku (International Maize and Wheat Improvement Center - CIMMYT, Mexico), gde su molekularni oplemenjivački pro-grami implementirani kako u državnom, tako i u privatnom sektoru oplemenjivanja pšenice (Gupta et al., 2010).
U Institutu za ratarstvo i povrtarstvo u Novom Sadu u proteklih deset godina uloženi su značajni napori da bi se ostvarila integracija konvencionalnog i molekularnog oplemenjivanja pšenice. Rad na pri-meni molekularnih markera u Institutu obuhvata nekoliko oblasti i to:
a) fragmentna analiza prisustva major gena i lokusa vezanih za naj-važnija oplemenjivačka svojstva kvantitativno-poligenog karak-tera (prinos, abiotski i biotski stres i tehnološki kvalitet),
b) DNK karakterizacija 190 sorti pšenice gajenih u Srbiji u posled-njih 40 godina,

28
Semenarstvo
c) validacija pažljivo odabranih tzv. „kandidat“ markera sa ciljem evaluacije njihovog potencijala za primenu u našim agroekološ-kim uslovima i
d) zajednički istraživačko-aplikativni rad u okviru međunarodnih projekata sa ciljem daljeg povećanja potencijala za primenu MAS (Marker Asistirane Selekcije) u oplemenjivanju pšenice.
Rezultati desetogodišnjeg rada obuhvataju, počev od molekularne karakterizacije genetičke varijabilnosti u novosadskoj kolekciji genotipo-va pšenice (Kobiljski et al., 2002) i detekciji alelne polimorfnosti u loku-sima major gena i QTL-a za agronomski važna svojstva (Kobiljski et al., 2006; Tošović-Marić et al., 2008; Marjanović, 2005; Barjaktarović et al., 2004; Obreht et al., 2006), preko DNK fingerprintinga 190 sorti pšenice stvorenih u Srbiji (Kondic-Spika et al., 2010), do utvrđivanja statistički značajnih veza između kandidat markera i agronomskih svojstava, odno-sno detekcije kandidat gena od potencijalnog značaja za oplemenjivački program pšenice (Kobiljski et al., 2009; Dodig et al., 2010č Neumann et al., 2010; Brbaklić i sar., 2010; Trkulja i sar., 2011). Dalja istraživanja podrazu-mevaju neprestano uključivanje novih markera u proces validacije, u cilju dobijanja najoptimalnije kolekcije markera, koji bi bili korišćeni za MAS.
Značaj novih tehnologija u očuvanju genetičkih resursa
U narednim dekadama predviđa se povećanje broja stanovnika, ur-banizacija će oduzeti još jedan deo poljoprivrednog zemljišta, dok su izvori novih obradivih površina ograničeni. Obradivo zemljište, zbog intezivne upotrebe i neodgovarajućih uslova održavanja postaje degra-dirano u mnogim zemljama. Napred navedeno upućuje na to da je po-trebno naći način kako da se obezbedi dovoljno hrane za ljude i životi-nje i obnovljive resurse na održiv način. To zahteva upotrebu novog pri-stupa biljkama, kako sa biološkog stanovišta tako i na nivou spoljašnjeg okruženja. U isto vreme, razvijaju se novi pravci istraživanja genoma primenom novih, savremenih tehnologija (European Technology Plat-form, 2007).
Interes za nove selekcione pristupe se javio sa ciljem proširenja ge-netičke osnove sorti, a to je:

29
Opšte semenarstvo
– značajno proširenje genetičke baze razvijene u selecionim pro-gramima,
– smanjenje genetičke erozije primenom populacionog pristupa u selekciji,
– povećanje učestalosti gena, sa minimalnim gubitkom genetičke varijabilnosti preko povratne selekcije i
–primenom metoda biotehnologije.
U savremenim uslovima visokorazvijenih tehnoloških postupaka i naučnih dostignuća, čovek sve više manipuliše genetičkim materijalom brojnih biljnih vrsta koje se odlikuju specifičnim, veoma korisnim karak-teristikama za ljudsku populaciju, kao što je oporavak od različitih bolesti, ili ljudima osiguravaju opstanak u svim delovima sveta. Jedna od delatno-sti koja doprinosi pomenutim poboljšanjima je i genetički inženjering. U okviru njega čovek manipuliše postojećim genetičkim materijalom bilj-nih vrsta poznatih osobina, stvarajući nove genetičke kombinacije veoma preciznim i specifičnim laboratorijskim postupcima.
Metodom rekombinantne DNK, hibridni molekul DNK stvoren in vitro, unosi se u ćelije organizma primaoca u kojima se ponaša kao deo
Slika 11. Cirkularni hromozom Agrobacterium tumefaciensFigura 11. Circular Chromosome Agrobacterium tumefaciens
(http://wishart.biology.ualberta.ca/BacMap/graphs_cgview.html)
Dužina/Lenght: 2,841,581 bp; Gena/Gene 2,728Uzorak/Accession: NC_003062

30
Semenarstvo
njegovog genoma. Osnovne komponente metoda su DNK i vektori za kloniranje (molekuli DNK koji imaju sposobnost umnožavanja, kao što su na primer plazmidi bakterija). Jedna od češće upotrebljavanih bakterija za ovu namenu je Abrobacterium tumefaciens, čija je stuktura hromozo-ma prikazana na slici 11. Osnovne tehnike obuhvataju izolaciju, sečenje i združivanje molekula DNK kao i njegovu transformaciju (unošenje hi-bridnog molekula DNK u ćeliju primaoca) (Milošević i sar., 2009).
Primenom novih tehnologija mogu se ustanoviti geni koji učestvu-ju u povećanju nivoa proizvodnje i kvaliteta novostvorenih sorti, novih gena koji kod biljaka regulišu tolerantnost na biotičke ili abiotičke čini-oce, bolje iskorišćenje hraniva, vode, otpornost na bolesti, insekte i dr. (sl. 12). One pomažu istraživačima da bolje okarakterišu i koriste gene-tičku divergentnost i genetičke resurse.
U praktične svrhe geni se koriste za lakšu selekciju modifikovanih ćelija i njihovu identifikaciju. Novi geni su uključeni u genom i prenose se na potomstvo. Proizvodi genetičkih modifikacija se nazivaju gene-tički modifikovani organizmi (Genetically Modified Organisms - GMO),
Bt gen će pomoći kukuruzu da razvije rezistentnost na insekte/Bt gene will help corn to develop resistance to insecticides
Bt gen unet u kukuruz/Bt gene insert in corn
Bt gen/Bt gene
Kukuruz/Corn
Enzim/EnzymeEnzim se koristi za isecanje gena/ Cutting enzyme
Slika 12. Proces dobijanja Bt kukuruza otpornog na kukuruzni plamenac primenom biotehnologije
Figure 12. The process of obtaining Bt corn resistant to corn pennant using biotechnology
(http://www.ces.ncsu.edu/resources/crops/ag546-1/helixes3.jpg)
Kukuruz/Corn

31
Opšte semenarstvo
a u Kartagena protokolu o biološkoj raznovrsnosti uz Konvenciju o bi-odiverzitetu, živi modifikovani organizmi (Living Modified Organisms-LMO). Na stečenom iskustvu zasnivaju se pravila za sigurnu i efikasnu upotrebu GMO.
Sekvencioniranje gena obezbeđuje ispitivanje složenosti genoma, daje sveobuhvatan uvid u gene za buduća istraživanja kod biljaka, ali to isto može da se vrši i kod biljnih vrsta koje se sada ne gaje, ali su se gajile u prošlosti (sl. 13). Sekvencioniranje gena otvara nove perspektive za ustanovljavanje porekla sorti i može da objasni razlike između njih.
Slika 13. DNKFigure 13. DNA
(http://www.molecularstation.com/molecular-biology-images/ 502-dna-pictures/109-dna.html)
Sa gledišta očuvanja genetičkog diverziteta posebno je važno izvrši-
ti sekvencioniranje i pozicioniranje starih, autohtonih sorti.FAO je pokrenuo inicijativu za procenjivanje razvića biljaka kom-
binacijom molekularnih i morfoloških metoda. Inicijativa je rezultira-la povećanjem baze podataka genofonda. Ključni činilac za pokretanje nove inicijative je pristupačnost uzorku sa odgovorajućim taksonom. Važno je da je molekularna analiza primenjena za isti ili sličan takson kao onaj upotrebljen u morfološkim analizama, kako bi se moglo izvrši-ti upoređivanje (Adams, 1997).

32
Semenarstvo
Uspešnost gajenja krompira i paradajza uslovljena je mogućnošću kontrole patogena Phytophthora infestans, koja prouzrokuje plamenjaču. Iz tog razloga se obavljaju brojna istraživanja na različitom nivou, s ciljem da se dugoročno ostvari kontrola bolesti. Razumevanje mehanizma delo-vanja patogena i njegovog razvoja je jedan od preduslova za dugotrajno rešenje problema. Urađeni su brojni genotipski i fenotipski testovi, ali su oni bili ograničenih mogućnosti do momenta uvođenja novih metoda. Značajan napredak u izučavanju genoma P. infestans ostvaren je prime-nom kodominantnih biomolekularnih markera. Njihovom upotrebom moguće je izučiti populaciju P. infestans i odrediti njenu biologiju, epide-miologiju, ekologiju, genetiku i evoluciju (Cooke and Lees, 2006).
Grahorica (Vicia sativa) je klasičan primer kompleksa dobro razdvo-jenih taksona i izvedenih formi, ona predstavlja različit stepen filogene-tičke divergentnosti. Ispitivanjima na nivou DNK njihova je divergen-tnost još više izražena, što se može ustanoviti primenom biotehnoloških metoda kao što su randomizirano umnožavanje polimorfizma DNK (RAPD) i polimorfizama dužine amplifikovanih fragmenata (AFLP) u odnosu na morfološke razlike (Potokina et al., 2000). Ustanovljeno je da postoji razičitost intraspesies članova grupe Vicia sativa L.senso stricto, obične grahorice, ekonomski važne krmne biljke, ako se njen diverzitet uporedi sa bliskim srodnicima, ali filogenetski odvojenog taksona skupa Vicia sativa. Upotrebom AFLP za filogenetsko razdvajanje DNK „finger printinga“ ispitano je 673 uzorka V.sativa iz Vavilov Instituta (Saint Pe-tersburg) i 450 uzoraka iz Instituta za izučavanje biljnih genetičkih re-sursa (Gatesleben) (sl. 14). Ova istaživanja su prva dokazala intraspecies različitost Vicia sativa čuvanih ex situ.
Mapiranje gena
Poboljšanje osobina biljaka moguće je izvesti boljim razumevanjem njihove molekularne osnove i procesa koji se putem njih dešavaju, kao i putem identifikacije gena, njihove karakterizacije za važna agronomska svojstva. Tokom prošlih petnaestak godina učinjen je značajan napre-dak u molekularnom mapiranju biljnog genoma.
Mape vezanih gena urađene su na osnovu DNK markera za veliki broj biljnih vrsta i one pomažu u savlađivanju različitih istraživačkih zadataka. Razvijeno je i praktično se koristi nekoliko glavnih tipova

33
Opšte semenarstvo
DNK markera – Polimorfizam dužine restrikcionih fragemnata (Restric-tion Fragment Length Polymorphisms – RFLP), Randomizirano umno-žavanje polimorfizma DNK (Random Amplified Polymorphic DNA – RAPD), Ponavljanje pojedinačnih sekvenci (Simple Sequence Repeats – SSR), Polimorfizam dužine amplifikovanih fragmenata (Amplified Fragment Length Polymorphisms – (AFLP), Polimorfizam pojedinačnih nukleotida (Single Nucleotide Polymorphisms (SNP) i Insercija/brisanje
Slika 14. Vrste roda Vicia: V. panonica (gore desno), V. sativa (gore levo), V. narbonensis (dole desno), V.villosa (dole levo)
Figure 14. Vicia species: V. panonica (above right), V. sativa (above left), V. narbonensis (bottom right), V.villosa (below left)
(foto: Mikić, A.)

34
Semenarstvo
(Insertions/Deletions -In/Del). Sledeća generacija mapa biće bazirana na pojedinačnim proteinskim sekvencama (Sihgle Protein Seqeunces – SPS). Kompletno mapiranje gena izvršeno je kod Arabidopsis thaliana 2000. godine, kod pirinča je završeno 2009. godine, a kod kukuruza rad na mapiranju je pri kraju (tab. 3). Mapiranje gena pomaže u razumeva-nju strukture, funkcije i evolucije biljnog genoma. Ono može biti važno oruđe za unapređenje u oblasti oplemenjivanja, jer se mogu locirati geni i za kvalitativna i kvantitativna svojstva, obezbeđujući osnovu za kloni-ranje gena i na kraju za genetičke modifikacije biljnog genoma.
Tabela 3. Ekpresija gena kod pirinča (RiceGE) Table 3. Genomic expres for rice (Gene Expression Atlas Data, 2011)
Potencijalno najinteresantnije nove tehnologije za genetičke analize su oligonukleotidni čipovi, kapilarna elektroforeza i matriks laser de-sorpciona jonizacija. U svakom slučaju će biti interesantno posmatrati uticaj tih novih tehnologija za razumevanje organizacije biljnog geno-ma. Najnovija dostignuća iz oblasti mapiranja gena objavljuju se u pre-stižnim časopisima (sl. 15).

35
Opšte semenarstvo
Naučnici sa Ajova državnog univerziteta koji se bave genetikom i hemijom su uspešno upotrebili nanotehnologiju da probiju zid ćelije i da u isto vreme ubace željeni gen i odgovrajuće hemikalije. Pomoću odgovarajućih hemikalija su izvršili aktiviranje gena sa velikom preci-znošću. Ovi rezultati ukazuju da je moguće koristiti nanotehnologiju na polju biologije i biotehnologije u poljoprivredi, stvarajući novu moćnu alatku za ciljano ubacivanje gena u biljne ćelije (http://www.physorg.com/news98540343.html).
Geni ne samo da mogu da se ispitaju primenom nanotehnologije, nego oni mogu da se i obeleže nanočipovima (biočipovi), koji se mogu koristiti za označavanje i pronalaženje pojedinačnih gena važnih za da-lja istraživanja. Oni se sada uglavnom koriste u humanoj medicini, ali svakako da će u skoroj budućnosti biti primenljivi i kod biljaka. (http://www.ipo.org/AM/Template.cfm?Template=/CM/ContentDisplay.cfm&ContentID=22898).
Slika 15. Publikacije vezane za izučavanje genoma Figure 15. Publications related to the study of genome
http://signal.salk.edu)

36
Semenarstvo
POJAM SORTE I NJENO PRIZNAVANJE
Sorta
Uspeh u oplemenjivanju meri se krajnjim proizvodom, a to je sorta. Da bi sorta bila priznata, mora da ima određene karakteristike. One po-drazumevaju visok prinos, otpornost ili tolerantnost na bolesti i insekte, dobre agronomske karakteristike i visok kvalitet semena. Nove osobine sorte moraju biti prihvatljive za proizvođača, inače sorta neće doprineti povećanju poljoprivredne proizvodnje.
Pojam sorte
Pojam sorte je definisan Međunarodnim kodom za nomenklatu-ru (International Code of Nomenclature of Cultivated Plants), u članu 2, osmog izdanja iz 2009. godine (IHSH, 2009).
Sorta je primarna kategorija gajenih biljaka čija je nomenklatura data u ovom Kodu (član 2.1) koji zatim opisuje sortu kao jedinicu u toj kategoriji.
„Sorta je skup gajenig biljaka koja je stvorena sa posebnim osobina-ma ili kombinacijom osobina i jasno su različite, unifomne i stabilne u tim karakteristikama koje zadržavaju kada se umnožavaju u određenom periodu.” (član 2.2).
Sorta znači podpodelu vrste na nižu sistematsku jedinicu koja je različita, ujednačena i stabilna:
– „različita” u smislu da sorta može da se razlikuje po jednoj ili više morfoloških, fizioloških ili drugih osobina po kojoj se izdvaja od svih ostalih poznatih sorti,
– „ujednačena” u smislu da se varijacije osnovnih osobina po koji-ma se razlikuje mogu opisati; i
– „stabilna” u smislu da će sorta ostati nepromenjena u svojim oso-binama i svojoj ujednačenosti kada se ponavlja bez obzira na ra-zličitost kategorija i sorti.

37
Opšte semenarstvo
Naziv sorte može biti legalno zaštićen regulativom vezanom za zašti-tu prava oplemenjivača. Sorta se identifikuje unikatnim imenom. Nazivi sorti su regulisani Međunarodnim kodom za nomenklaturu i registro-vani u Međunarodnom registru sorti (International Cultivar Registrati-on Authority – ICRA). Naziv sorte sadrži botaničko ime (rod, vrsta, in-traspecifičan takson, interspecifičan takson, interspecifičan hibrid ili in-tergeneričke hibiride) praćene poreklom sorte (date od oplemenjivača).
Davanje novog i originalnog imena sorti nije lako, posebno u gru-pama koje su istorijski imale stotine ili čak hiljade sorti. Srećom, mnoge od ovih grupa su navedene u Međunarodnom registru sorti koji objav-ljuju kontrolne liste i registre imena sorti koja su u upotrebi ili koja su korišćena u prošlosti. U alfabetskim spiskovima ICRA rodova može se pretražiti da li je ime sorte već korišćeno, a zatim se može konsultovati ICRA publikacija ili direktno registar ICRA.
U praksi se najviše gaje hibridne sorte kod stranooplodnih biljaka i to F1 generacija prostih hibrida (kukuruz, sirak, šećerna repa) ali i kod sa-mooplodnih biljaka gde je biološki moguće ili ekonomski opravdano pro-izvoditi hibridno seme (paradajz, krastavac, plavi patlidžan, tikva i dr.).
Hibridna sorta predstavlja F1 generaciju nastalu ukrštanjem dve sa-mooplodne linije (kod stranooplodnih biljaka) ili dve čiste linije (kod samooplodnih biljaka), odnosno populaciju nastalu ukrštanjem tri ili četiri roditelja. Kod nekih stranooplodnih vrsta, raž, lucerka i dr., hi-bridnom sortom se smatra i potomstvo nastalo ukrštanjem sorti ili po-pulacija. Ukoliko se koristi F1 generacija prostih hibrida, hibridna sorta je heterozigotna, ali su sve jedinke istog genotipa, pa se sorta odlikuje skoro potpunom fenotipskom uniformnošću. U svim ostalim slučaje-vima hibridna sorta se sastoji od različitih genotipova pa se stoga ne odlikuje fenotipskom uniformnošću (dvojni, trojni hibridi).
Činioci koji narušavaju osobine sorti
U procesu umnožavanja sorti potrebno je očuvati njene morfološ-ke, biološke i agronomske osobine, genetičku, odnosno sortnu čistoću i kvalitet semena kao i genetički potencijal za prinos na nivou kakav je bio u momentu priznavanja sorte. U toku gajenja kod nekih sorti ili hi-brida može doći do promene ovih osobina i do opadanja genetičkog po-tencijala za prinos. Rezultati naučnih istraživanja pokazuju da su uzroci ove pojave mnogobrojni, a po Dokiću i Mihaljevu (1986) najvažniji su:

38
Semenarstvo
a) Cepanje unutar sorti koje nastaje kao posledica genetičkog cepa-nja unutar sorti, naročito kod samooplodnih ali i kod stranooplodnih biljaka. Javljaju se genotipovi drugačijih morfoloških osobina i manje produktivnosti, a bolje prilagođenosti agroekološkim uslovima, koje onda prirodna selekcija favorizuje, posledica čega je promena genetske kompozicije sorte. Ovo se naročito odnosi na nove sorte koje u nizu kvantitativnih svojstava ne predstavljaju homozigotnu čistu liniju, nego se sastoje od biljaka ili linija nejednakih po svom genetičkom potenci-jalu za prinos.
b) Degeneraciju sorti uslovljavaju činioci spoljne sredine favorizu-jući prirodnu selekciju genotipova bolje prilagođenih spoljnim uslovi-ma, a koji nastaju kao rezultat prirodnih mutacija, koje iako niskih uče-stalosti (oko 10-6) tokom gajenja jedne sorte mogu izmeniti genetičku kompoziciju sorte. Bolje prilagođeni genotipovi spoljnim uslovima su po pravilu slabije rodnosti.
c) Biološko mešanje sorti nastaje zbog prirodne – nekontrolisane stranooplodnje pri čemu se stvaraju genotipovi promenjenih morfološ-kih osobina, slabije rodnosti, a što doprinosi promeni genetičke kom-pozicije sorte. Ova pojava je naročito česta kod stranooplodnih biljaka iako se dešava i kod samooplodnih vrsta (posebno u godinama kada u vreme cvetanja i oprašivanja vladaju uslovi visoke temperature i visoke relativne vlažnosti vazduha, što povećava procenat stranooplodnje kod samooplodnih vrsta).
d) Napad bolesti i štetočina se uglavnom javlja tokom dugog niza godina gajene sorte i hibridi koji su pri stvaranju bili otporni na preo-vlađujuće fiziološke rase nekog patogenog organizma ili insekta, a mogu tu svoju otpornost da izgube zbog pojave novih fizioloških rasa pato-gena i novih biotipova insekata (vertikalna otpornost). Otporna sorta vrši selekcioni pritisak na populaciju parazita, eliminiše određene rase, a druge postaju prevalentne na koje sorta nije otporna. Kao posledica toga smanjuju se proizvodne osobine gajene sorte.
e) Mehaničko mešanje sorti podrazumeva mešanje semena gajene sorte sa semenom drugih vrsti biljaka ili drugih sorti iste vrste. Ono može nastati u raznim fazama procesa proizvodnje semena, a javlja se kao posledica nedovoljne pažnje učesnika pri setvi i žetvi semenskih useva, doradi i skladištenju semena ili pri drugim manipulacijama s ne-dorađenim semenom ili semenskom robom.

39
Opšte semenarstvo
S obzirom da je seme živi materijal, nosilac i prenosilac velikog bro-ja genetički uslovljenih, a po čoveka korisnih, osobina oplemenjenih biljaka, pri svim operacijama u semenarstvu mora se voditi računa o očuvanju kvaliteta semena (čistoća, energija klijanja, klijavost, sadržaj vlage, zdravstveno stanje, vitalnost, ujednačenost i dr.). Do narušavanja kvaliteta semena može doći zbog nepovoljnih uslova spoljne sredine u proizvodnji semena, zbog nepridržavanja agrotehničkih rokova, zbog nepridržavanja propisanih doza hemijskih sredstava za suzbijanje koro-va i zaštitu useva i semena od štetnih organizama, zbog mehaničkih po-vreda semena pri žetvi, doradi, neodgovarajućeg uskladištenja i drugih uzroka. Seme se mora pripremiti za promet i setvu i to se čini u procesu dorade semena.
Osnovni cilj organizovanja proizvodnje semena je da ona bude efi-kasna i da može da obezbedi proizvodnju sortnog i kvalitetnog semena kao i da se zaštiti proizvodnja semena od svih propusta. Zbog toga se svaka zemlja trudi da dobru organizaciju semenarstva obezbedi zakon-skim propisim koji regulišu pojedine faze u procesu proizvodnje seme-na. Po pravilu zemlje sa razvijenom poljoprivrednom proizvodnjom imaju razvijeno, dobro organizovano, savremeno uređeno i efikasno semenarstvo (Milošević i Malešević 2004).

40
Semenarstvo
PRIZNAVANJE SORTI
Ispitivanje sorti i zahtevi za njihovo priznavanje
Pravo na komercijalizaciju sorta dobija onog momenta kada je pri-znata i upisana u sortnu listu. Da bi sotra bila priznata mora da prođe postupak ispitivanja radi ustanovljavanja njene različitosti u odnosu na druge, već priznate sorte, ujednačenosti, odnosno da su sve biljke jedne sorte fenotipski iste u određenom procentu. Stabilnost sorte označava njenu karakteristiku da u različitim uslovima spoljne sredine daje stabil-ne prinose. Pored tih karakteristika, sorta mora da ima dobre agronom-ske osobine kao što su visina prinosa, otpornost na poleganje kod žita, graška, tolerantnost na štetne organizme.
Ispitivanja radi utvrđivanja različitosti, ujednačenosti i stabilnosti (DUS)
Za zemlje sa obaveznom regulativom za priznavanje, postoje dve vrste ispitivanja u cilju priznavanja sorti:
1. Ispitivanje radi utvrđivanja različitositi, ujednačenosti i stabilno-sti (Distinctness, Uniformity, Stability – DUS)
2. Ispitivanje radi utvrđivanja poljoprivredne vrednosti (Value for Cultivation and Use – VCU) (sl. 16).
Ispitivanje sorti radi utvrđivanja različitosti, ujednačenosti i stabilnosti
Od velike je važnosti donošenje odluke da li se nova sorta razlikuje ili ne od ostalih priznatih sorti, jer mogućnost razlikovanja nove sorte predstavlja način kontrole koji je neophodan za zaštitu sorte.

41
Opšte semenarstvo
Za pravilno izvođenje DUS testa mora se :
– razviti tehnički vodič za gajenje, ispitivanje morfoloških karakte-ristika (boja, oblik) i fizioloških karakteristika (otpornost na bole-sti, posebne osobenosti i dr.) sorte,
– izvršiti kolekcionisanje i konzervaciju sorti, uključujući standar-dne sorte, i razvoj njihovih baza podataka,
– sakupljanje podataka o novouvedenim sortama/vrstama u zemlju za izvođenje DUS testova u svrhu zaštite intelektualne svojine (http://www.ncss.go.jp/main_e/functions/dus.html)
DUS testovi imaju dvojnu ulogu. Koriste se u procesu priznavanja sorti i u procesu zaštite prava oplemenjivanja novih biljnih sorti. Proce-dura za DUS test podrazumeva oglede u polju i laboratoriji usklađenih sa tehničkim procedurama postojećih vodiča UPOV (International Uni-on for the Protection of New Varieties of Plants) (UPOV, 2009).
Ispitivanje različitosti, uniformnosti i stabilnosti sorte može da se prepusti državnim organizacijama koje su nezavisne od oplemenjivača. Ispitivanja moraju da se vrše najmanje dve sezone. Uslovi i nivo odgo-vornosti između poverioca posla i ispitivača mora biti određen ugovo-rom. Poverilac posla može poslati ispitivaču tehnički upitnik i prijavu
VREDNOSTI ZA GAJENJE I
KORIŠĆENJEVALUE FOR
CULTIVATION AND USE
RAZLIČITOST, UNIFORMNOST,
STABILNOSTDISTINCTNESS,UNIFORMITY,
STABILITY
DUS testDUS testing
VCU testVCU testing
Priznavanje sortiRealising of plant varieties
Slika 16. Sistem priznavanja sorti u BugarskojFiger 16. Plant variety registration system in Bulgaria
(Atanassov, 2003)

42
Semenarstvo
za zaštitu novih sorti (Commission Directive 2008/83/EC), član 2 regu-lative za procedure za ispitivanje različitosti, ujednačenosti i stabilno-sti (DUS) novih sorti radi zaštite autorskih prava. U Poljskoj je u 2008. godini u DUS testovima bilo ispitano 10611 sorti, uključujući 724 sorte koje su bile u postupku zvaničnog ispitivanja radi priznavanja. DUS te-stove izveo je Istraživački centar za testiranje sorti (The Research Centre for Cultivar Testing – COBORU) (sl. 17) (UPOV, 2009).
U našoj zemlji, ispitivanja sorti u svrhu priznavanja je obavezno i traje dve godine za ratarske i povrtarske biljne vrste. Ako se radi o sor-tama sa izuzetno dobrim agronomskim karakteristikama, Ministarstvo poljoprivrede, trgovine, šumarstva i vodoprivrede Republike Srbije, može izdati dozvolu za proizvodnju te sorte i za kraće vreme od vreme-na koje je zakonom predviđeno. Ako se neke biljne vrste tradicionalno ne gaje u Srbiji, sorte ne moraju da prođu proces priznavanja da bi bile gajene u Srbiji.
Botaničke karakteristike koje se koriste za razlikovanje sorti često je teško korektno proceniti, pa zbog toga procenu mora izvršiti specijali-sta. Takva ispitivanja su teška i zahtevaju dosta vremena. Za ispitivanja je potrebno obezbediti parcele za proveru, kako bi se omogućilo poređenje
Slika 17. Broj DUS testova u 2008. izvedenih u PoljskojFigure 17. Number of varieties in DUS testing in 2008 in Poland
(UPOV, 2009)

43
Opšte semenarstvo
Tabela 4 . UPOV deskriptor za soju (UPOV, 2002)Table 4. UPOV guideline for soybean
Faza ispitivanjaStage of examination
UPOVBroj osobina
Character numbers
Opis osobineCharacter description
Primarna/Primary
Vegetativna/Vegetative 1Hipokotil: obojenost
antocijaninomHipokotil: color anthocyanin
U cvetu/in bloom 3 Biljka:način rastaPlant: the way of growth
5 Biljka: boja malja/Plant: hair color
6 List: oblik lateralne liskeList: a form of lateral coot
7 List: boja /Leaf: color8 Cvet: boja / Flower: color
15Vreme početka cvetanja
(1 otvoreni cvet na 10% biljaka)Start time of flowering (1 open
flower to 10% of plants)U zrelosti/in maturity 4 Biljka: visina / Plant: height
16 Vreme zrelosti Time of maturity
Sekundarna/Secendary
Vegetativna/Vegetative 2 Biljka: vrsta rastaPlant: growth type
U cvetu/in bloom 3 Biljka: način rastaPlant: the way of growth
8 Liska: veličina / leaf: sizeU vreme razvijanja mahuna /At the time of developing pods 10 Mahuna: boja / Legume: color
U zrelosti/in maturity 11 Seme: veličinaSeed:size
12 Seme: oblikSeed: shape
13 Seme: boja ljuske hilumaSeeds: shell color of hylum
14 Seme: boja hilumaSeed: color of hylum
* Reference: UPOV uputstva za izvođenje testova za različitost, homogenost i stabilnost.Doc.No. TG/80/6; Klasifikovana i kao Soja hispida Moench* Reference: UPOV’s instructions for carrying out tests for diversity, homogeneity and
stability.Doc.No. TG/80/6; Classified as Soja hispida Moench

44
Semenarstvo
sorti tokom cele vegetacije. Karakteristike koje se posmatraju opisane su u deskriptorima (ključevima) koje je sačinila Međunarodna organizacija za zaštitu novih biljnih sorti ( International Union for the Protection of New Va-rieties of Plants – UPOV), a Organizacija za ekonomsku saradnju i razvoj, šema za seme (Organization for Economic Co-opration iand Development, Seed Scheme-OECD) za njihovu proveru u proizvodnji (OECD, 2008) (tab. 4). Danas postoje sorte koje je teško razlikovati, pošto su kod njih prisutni pojedinačni geni, recimo za otpornost na bolesti što je jedina karakteristika po kojoj se mogu razlikovati od drugih sorti. Kod sorti pšenice i ječma je ta-kođe teško izvršiti tačno određivanje sortne pripadnosti. Posebnim metoda-ma tečne hromatografije, kao što je RH-HPLC, može da se precizira specifič-nost i originalnost određenog genotipa na brojnim lokusima kod kukuruza, pšenice, ječma, krompira i dr. Još novija metoda je metoda polimorfizma dužine restrikcionih fragmenata (RFLP). Ovom metodom može da se utvrdi preko 200 DNA fragmenata u sorti i tako dobiju tačne informacije o njenom identitetu. Reakcija lančane polimerizacije (PCR) takođe se može koristi kao pouzdana metoda u naznačenu svrhu (sl. 18).
Slika 18. Princip rada PCR-aFigure 18. Principle of PCR
(http://users.ugent.be/~avierstr/principles/pcr.html)
PCR: Reakcija lančane polimerizacije/PCR: Polymerase Chain Reaction
30 - 40 ciklusa u 3 faze/30 - 40 cycles of 3 steps :
Faza 1 : denaturacija/ Step 1 : denaturation
Faza 2 : razgradnja/ Step 2 : annealing
Faza 3 : umnožavanje/ Step 3 : extension
1 minut 94°C/ 1 minute 94°C
45 sekundi 54°C/ 45 seconds 54°C
2 minuta 72°C samo dNTPs/ 2 minutes 72°C only dNTP's
uzlazni i silazni prajmeri/ forward and reverse primers!!!

45
Opšte semenarstvo
Ispitivanja sorti radi ustanovljavanja poljoprivredne vrednosti (VCU)
Poljoprivredna vrednost opredeljuje dalje gajenje novostvorene sor-
te. Zbog toga je od posebne važnosti realno i tačno ustanoviti pokazate-lje poljoprivredne vrednosti.
U postupku pome-nutog ispitivanja sorte se ispitaju na prilago-đenost gajenja na ve-likom broju lokaliteta koji obuhvataju različi-te tipove zemljišta i kli-matske uslove. U najve-ćem broju zemalja ove oglede vrše neutralne organizacije, kako bi se izvršilo objektivno oce-njivanje novih sorti u odnosu na komercijal-ne, koje služe kao stan-dard. Cilj ovih ogleda je da se odaberu samo one sorte koje imaju veću agronomsku vred-nost od već postojećih najboljih sorti (sl. 19).
Agronomska svoj-stva sorte, odnosno spo- sobnost sorte da ostva-ruje, recimo, visok pri-nos, su isto toliko značajna za proizvođača kao i za oplemenjivača. Ra-nostasnost je veoma važna u regionima gde je vegetaciona sezona kratka, ili gde je usev u sastavu višepoljnog plodoreda. Razvijen korenov sistem, čvrstina i visina stabla mogu da utiču na prihvatanje jedne sorte u širo-koj proizvodnji, jer ovi činioci utiču na prilagođavanje sorte, na nivo đu-brenja i dr. Hemijski sastav semena je jedan od činilaca kvaliteta. Veći
Slika 19. Kvalitetna biljka – kvalitetno seme Figure 19. Quality plant - quality seed
(foto: Milošević)

46
Semenarstvo
sadržaj proteina ili bolji odnos aminokiselina kod žita može biti pred-nost određene sorte.
Za sorte koje treba da nađu široku primenu u proizvodnji, nije dovoljno da zadovolje želje proizvođača, već one moraju biti i lake za umnožavanje. Neke visoko prinosne sorte nisu uspele da se rašire u pro-izvodnji zbog toga što su oplemenjivači ili proizvođači imali problema sa proizvodnjom semena.
Agronomska vrednost sorte zavisi od mnogih karakteristika, od ko-jih su najvažnije:
– sposobnost da ostvaruje visok prinos,– sposobnost da pozitivno reaguje na poboljšane uslove obrade i
mere nege, – da daje prinos visokog kvaliteta,– da je otporna, odnosno tolerantna na bolesti i štetočine,– da se lako prilagođava na nepovoljne agroekološke činioce (mraz,
suša, i dr.),– da je pogodna za mehanizovanu obradu i žetvu.
Sortni ogledi
Izvođač sortnih ogleda ima zadatak da izvrši procenu vrednosti no-vih sorti u uporednim ogledima sa standardnim sortama. Pri izvođenju sortnih ogleda vrše se posmatranja u toku vegetacije (vreme nicanja, broj niklih biljaka, vreme cvetanja, formiranje semena i dr.) utvrđuje prinos i vrše morfološka i laboratorijska ispitivanja.
Planiranje sortnih ogleda uključuje izbor načina postavljanja ogle-da, veličine i oblika parcele, broja ponavljanja. Način postavljanja ogle-da uglavnom zavisi od broja sorti koje treba ispitati. Za manji broj sorti (četiri do šest) najpogodniji je latinski kvadrat. Kada se ispituje 10–20 sorti najbolje je oglede izvesti po randomiziranom blok sistemu.
Veličina parcela treba da je takva da se isključe varijacije pojedi-načnih biljaka, znači ne suviše mala, a ni prevelika kako ne bi otežavala izvođenje ogleda. Uobičajeno je da je veličina ogledne parcelice od 5m2 do 15m2 za pšenicu i 15 m2 do 25 m2 za kukuruz. Oblik parcela je pravo-ugaoni. Broj ponavljanja u ogledima kreće se od tri do šest, a u najvećem broju slučajeva je određen statističkom metodom koja je izabrana za izvođenje ogleda.

47
Opšte semenarstvo
Izvođenje sortnih ogleda
Prva priprema parcele u polju je osnovna obrada zemljišta, koja se izvodi na isti način i u isto vreme kao i ona koja se primenjuje u proi-zvodnji. Obrada zemljišta se pažljivo izvodi, ali treba da se vodi računa da se ogledi seju mašinama koje obezbeđuju bolju strukturu zemljišta od onih koje se koriste za setvu komercijalnih useva. Naročito je važno da zemljište ima ravnu površinu i mrvičastu strukturu.
Posle osnovne obrade vrši se obeležavanje parcela, blokova i ponav-ljanja unutar ogledne površine. To može da se uradi ručno, ali je mnogo praktičnije koristiti mašine, kao što su traktori kojima su dodati pogod-ni uređaji za obeležavanje.
Obeležavanje parcela se izvodi postavljanjem etiketa na kojima je naznačen broj parcele i šifra sorte. One se privremeno postavljaju i posle završetka setve se fiksiraju na konačna mesta, obično ispred prvog reda svake parcelice (sl. 20).
Setva se obavlja pažljivo, jer kvalitet obavljanja setve utiče na tačnost
rezultata dobijenih iz ogleda. Setva može da se izvede na različite nači-ne: ručno ili mašinski, red po red s jednorednom sejalicom, sejalicama pogodnog tipa i specijalnim sejalicama koje setvu semena vrše automat-ski. Obavljanje setve sejalicama ima prednost nad ručnom setvom jer
Slika 20. Obeležene parcelice na oglednom poljuFigure 20. Marked plots on experimental field (foto: Sabadoš)

48
Semenarstvo
značajno smanjuje vreme setve, tako da setva može da se obavi brzo kada su uslovi vremena i zemljišta najbolji. Prednost sejalica je i u tome što seme niče ujednačenije i tako značajno smanjuju eksperimentalnu grešku.
Suzbijanje korova na oglednim parcelama se obično izvodi herbi-cidima. Mehaničko suzbijanje korova se vrši u ogledima sa pšenicom, čupanjem pojedinačnih biljaka, dok se mašinski mogu suzbijati u ogle-dima sa kukuruzom i drugim biljnim vrstama sa širokorednom setvom.
Za žetvu eksperimentalnih parcela postoje mašine različitog tipa. Korišćenjem malih kombajna, žetva se izvodi u kratkom roku čime se uštedi vreme i radna snaga. U mnogim slučajevima žetva i vršidba se izvode ručno.
Osmatranja biljaka u polju i laboratorijske analize
Prinos, koji je najvažnji pokazatelj kvaliteta nove sorte, predstavlja samo jednu od karakteristika koje određuju agronomsku vrednost nove sorte. Ostale karakteristike se utvrđuju osmatranjima u polju i labora-torijskim analizama. Osmatranja u polju se vrše beleženjem različitih karakteristika; neke od njih, koje se odnose na pšenicu i kukuruz, date su u narednom pregledu. Za pšenicu se vrše sledeća poljska osmatranja:
– datum pojave ponika,– otpornost na izmrzavanje (niske temperature),– datum bokorenja,– datum klasanja,– otpornost na bolesti kao što su žuta rđa (Puccinia striformis), li-
sna rđa (P. recondita), stabljičina ili crna rđa (P. graminis), pepelnica (Erisiphe graminis), gar (Ustilago nuda, U. tritici), glavnica (Tilletia caries; T. tritici), pegavost lista (Septoria spp.), Fusarium spp. i dr. ,
– otpornost na štetočine kao što je pšenična muva (Oscinella frit) (sl. 21),
– otpornost na poleganje,– tolerantnost na zaslanjena zemljišta,– otpronost na sušu,– intenzitet bokorenja,– otpornost na osipanje semena iz klasa,– datum sazrevanja,– broj dana od nicanja do sazrevanja (dužina vegetacije).

49
Opšte semenarstvo
Uobičajeni sistem vođenja beležaka, za većinu osobina je davanje ocena od 0 do 9, gde je 9 najbolja ocena za posmatranu osobinu. Dobi-jene podatke treba upotpuniti opisom morfoloških karakteristika sorti, uključujući visinu biljke i dužinu klasa, zbijenost klasa i prisustvo ili odsustvo osja, karakteristike glume, boja i veličina semena.
U laboratoriji se analizira kva-liteta zrna, brašna, testa i hleba. Pri tom se ustanovljava:
– masa hiljadu semena,– hektolitarska masa,– sadržaj glutena,– kvalitet glutena (sedimenta-
cioni broj),– osobine meljivosti,– kvalitet testa (farinografom),– kvalitet pecivosti (pečenjem
testa, zapreminom hleba, poroznost hleba) (sl. 22),
– hemijski sastav (pepeo, skrob, protein, lizin).
Slika 21. Oštećenja na pšenici od Oscinella frit (pšenična muva)Figure 21. Damage to wheat from Oscinella frit (wheat flies)
(http://agro.se-ua.net/images/shvmuha1.jpg)
Slika 22. Poroznost hlebaFigure 22. Porosity of bread
(http://sweetthingdesigns.typepad.com/.a/6a00e54f836792883301156fb1d962970c-450wi)

50
Semenarstvo
Kod laboratorijskih analiza zrna kukuruza obično se uključuju sle-deći pokazatelji:
– masa hiljadu semena,– hektolitarska masa,– konzistencija semena,– sadržaj skroba,– sadržaj proteina,– sadržaj ulja,– sadržaj lizina i drugih aminokiselina.
Komisija za priznavanje sorti
Odluke o priznavanju i registraciji sorte se obično poveravaju komi-siji za priznavanje sorti. Komisiju imenuje rešenjem Ministarstvo poljo-privrede, trgovine, šumarstva i vodoprivrede Republike Srbije. U okviru komisije postoje potkomisije za pojedine biljne vrste ili grupu biljnih vrsta. Tako postoji komisija za uljane biljne vrste, za kukuruz i dr. Čla-novi komisije čine članovi delegirani iz državnih organizacija, privatnih institucija, multinacionalnih kompanija. Oni treba da budu nezavisni od oplemenjivača o čijim sortama se odlučuje.
Komisija za priznavanje sorti se obično sastaje u redovnim interva-lima. Rezultati sortnih ogleda, za sortu koja je prijavljena u svrhu ispiti-vanja i priznavanja, se razmatraju i komisija za priznavanje sorti donosi odluku o tome da li neku sortu treba priznati kao novostvorenu ili ne. Stepen u kome komisija za priznavanje uzima u obzir rezultate ispitiva-nja samog oplemenjivača, ukoliko su podneti, razlikuje se od sistema do sistema. U šemama dobrovoljne registracije mnogo pažnje se obično poklanja rezultatima ispitivanja samog oplemenjivača. U šemama oba-vezne registracije, kao što je to u Srbiji i većini evropskih zemalja, u razmatranje se uzimaju samo zvanični rezultati ispitivanja prijavljene sorte za priznavanje. Komisija koja je uvrstila sorte u sortnu listu ima pravo da traži ukidanja prava gajenja, ako je sorta podložna bolestima, pa predstavlja izvor širenja štetnih organizama. Novopriznata sorta se upisuje u sortnu listu. Objavljivanjem odluke sortne komisije stavlja-njem sorte na sortnu listu u Službenom glasniku Republike Srbije, sorta se smatra priznatom.

51
Opšte semenarstvo
Sortne liste
Sortne liste imaju za cilj da obaveštavaju kupca semena o novim sortama koje je priznala sortna komisija, ponovo priznatih već registro-vanih sorti čija je registracija istekla, brisanim sortama sa sortne liste i da ozvaniče ove promene.
Sortna lista koju izdaje Ministarstvo poljoprivrede, trgovine, vodo-privrede i šumarstva Republike Srbije, na predlog Komisije za priznava-nje sorti ima restriktivnu ulogu. U EU, Srbiji i nekim drugim evropskim zemljama prodaja semena sorti koje nisu na zvaničnoj sortnoj listi je zabranjena u saglasnosti sa zakonskom regulativom za priznavanje ili obaveznim pravilima za kontrolu proizvodnje semena.
Pored toga što je sorta priznata na nacionalnoj sortnoj listi, sorta mora biti priznata i na sortnoj listi zemlje u koju se uvozi sa namerom da seme sorte bude namenjeno tržištu zemlje u kojoj se proizvodi ili se seme umnožava za izvoz. Pošto se izvoz vrši po OECD šemi za seme, sorta koja je namenjena međunarodnom prometu mora biti uključena u OECD sortnu listu. U njoj je naznačeno i vreme za koje ovo priznavanje zvanično važi. Po isteku toga vremena sorta se skida sa sortne liste. Po-stoji mogućnost, da ako je sorta dobra, dobije preregistraciju i ponovo se unese u sortnu listu, odnosno katalog priznatih sorti. U katalozima OECD-a u kojima se nalaze sorte namenjene međunarodnom prometu, obavezno se naznačava institucija ovlčašćena za umnožavanje sorti sa OECD sortne liste.
Sorta namenjena izvozu na tržište Evropske unije mora biti prizna-ta i u zemlji u koju se izvozi, odnosno, mora biti na sortnoj listi te ze-mlje ili mora biti priznata na nivou EU i to naznačeno u EU katalogu (EU Common Catalogue) (Council Directive 2002/53/EC) koji ustvari predtsavlja sortnu listu EU. Priznata sorta ima svoj vek trajanja, što je naznačeno u sortnoj listi.
Pored svih napred pomenutih uslova svaka zemlja koja izvozi seme u EU mora imati odobrenu ekvivalenciju za izvoz, od Saveta za poljopri-vredu EU, jer sve zemlje vode računa o svojim potrebama u semenu. EU veoma pažljivo razmatra zahteve za dodelu ekvivalenicije. To je ustvari jedan vid kreiranja ekonomske politike u semenarskoj sferi. Sve zemlje u tranziciji imaju ovakav sistem priznavanja sorti ali i pravo izvoza po OECD šemi.

52
Semenarstvo
Trendovi kod registracije sorti
Mnoge evropske zemlje su već razvile zakonodavni sistem koji do pušta gajenje i prodaju populacija i starih sorti. Poljoprivredna politika Švajcarske prva je uvela klauzulu za korišćenje starih sorti i populaci-ja, ali kao nesertifikovano seme i sadni materijal. Sorte ne moraju biti registrovane u nacionalnom katalogu sorti. Dozvoljeno je izuzeće oko 60 populacija žita i oko 70 populacija krompira čije je gajenje bilo do-zvoljeno još 1999. godine. Uslovi za izuzeće su vrlo jednostavni. Zahtev treba da je u saglasnosti sa osnovnim informacijama kao što su podaci o podnosiocu i populaciji (staroj sorti). Registracija je do danas besplatna.
Finska je uspostavila drugačiji model. Prema njenom predlogu, poljoprivrednik podnosi zahtev za registraciju sorte odeljenju za pri-znavanje sorti, za koje poljoprivrednik plaća naknadu. Ovaj proces je zasnovan na metodi i smernicama UPOV-a, čak i ako sorta ne pose-duje uniformnost ili kriterijume stabilnosti. Manje pažnje se poklanja sposobnosti za klijanje semena i genetičkoj čistoći starih sorti koje su prihvaćene za gajenje. Neki manje pozitivan aspekt ovog predloga je da se stvara relativno strog i fleksibilan regulacioni okvir koji reguliše ispi-tivanje semena.
Druge evropske zemlje imaju propise koji nisu obećavajući za pri-znavanje lokalnih sorti. U Francuskoj, zakonom su zadržane visoke cene priznavanja lokalnih sorti (221 eura po sorti). Osim toga, u cilju registracije, različitost sorte se ne može lako dokazati, a potrebno je i da je sorta starija više od 20 godina, što predstavlja izazov za lokalno sta-novništvo i sputava njihovu želju da koriste lokalne populacije (tab. 5) (Toledo, 2002).
Zaštita sorti
Zaštita sorti znači pravo oplemenjivača da ustupi pravo gajenja sor-te samo onim proizvođačima od kojih može da naplati licencu, odnosno vrati uložena sredstva u stvaranje sorte i ostvari tako materijalne pre-duslove za dalje stvaralaštvo. Za većinu zemalja Evropske unije stvarno saznanje o vrenosti stvorene sorte počinje 1996. godine, kada je zašti-ćeno 1386 sorti. Trend zaštite je rastao, tako da je u 2008. godini broj zaštićenih sorti iznosio 3012 (sl. 23).

53
Opšte semenarstvo
Tabela 5. Upoređenje tri nacionalna zakona o semenu u Evropi, Francuskoj, Švajcar-skoj i Finskoj
Table 5. Comparation of Three National Seed Laws in Europe in France, Switzerland and Finland (Toledo, 2002 )
Francuska/ France
Švajcarska/ Switzerland
Finska/ Finland
Registracija lokalnih sorti/Registration of local varieties Da / Yes Da / Yes Teško/
DifficultRegistracija starih komercijalnih sorti/Registration of old commercial varieties
Ne / No Da / YesSamo sorte starije od
20 godina/ Only varieties >20 years
Plaćanje takse/ Fee payable Ne / No Da / Yes Da, visoko/Yes, high
Morfološke osobine moraju biti registrovane/ Morphological traits must be registered
Ne / No
Da ali ne striktno/
Yes but not strict
Da, DUS kriterijumi/Yes, DUS criteria
Ograničenja za komercallizaciju/Restrictions on commercialisation?
KvantitativnoSamo za domaću
upotrebu/Quantitative
Domestic use only
KvantitativnoSamo
individualno/Quantitative
Only individuals
Samo za domaću upotrebu/
Domestic use only
Slika 23. Povećanje broja zaštićenih sorti u Evropskoj uniji po godinama (UPOV, 2009) Figure 23. Increasing the number of protected varieties in the European Union by the year

54
Semenarstvo
Na nivou EU u Anžeu u Francuskoj, nalazi se agencija koja vrši zaštitu sorti za potrebe evropskih zemalja (Central Plant Variety Office – CPVO), ali i članica država kandidata za članstvo, uz posebno odo-brenje. Iz podataka sa slike 24 se vidi da se najviše štite ukrasne biljke (54,2%), zatim ratarske biljne vrste (26,2%), povrće(13,6%) i voće (6%).
Slika 24. Zaštita sorti po grupama biljnih vrsta Figure 24. Variety protection in groups of plant species (UPOV, 2009)
Posebno je među ukrasnim biljnim vrstama cenjena orhideja, čije se sorte obavezno štite. U tabeli 6 je prikazano koliko vrsta i sorti orhideja je priznato u periodu od 1995. godine do 2008. godine (UPOV, 2009).
Veoma dobru organizaciju zaštite sorti imaju Francuska, Nemačka, Češka, Slovačka, Mađarska, Bugarska. Na primeru Bugarske biće pri-kazan proces zaštite sorti. Oplemenjivač ili oplemenjivačka ustanova prijavljuju sortu kancelariji za patente, radi zaštite autorskih prava. Na osnovu pozitivnog mišljenja agencije za ispitivanje sorti o DUS testu, dobija se privremena ili stalna zaštita sorte. Privremenu zaštitu sorti mogu da imaju samo one zemlje koje su potpisale UPOV konvenciju iz 1991. godine (sl. 25).
Koliko je važno imati dobar sistem zaštite sorti može se videti iz primera Češke Republike. Iz tabele 7 se vidi kolika je vrednost jedne stvorene sorte i koliki je značaj zaštite sorti. Za stvaranje jedne sorte pšenice utroši se 2,5 miliona dolara. Pšenica se seje na oko 100 000 ha. Međutim, sertifikovanim semenom se seje svega 50% površina. Iz tog razloga se, naplaćujući 20 dolara po toni semena za autrostvo godiš-nje, vrati oplemenjivaču ili oplemenjivačkoj ustanovi 200000 dolara. Sva uložena sredstva u stvaranje jedne sorte se vraćaju posle 12 godina.
26,2%
13,6%
54,2%
6%
Slika 25. Zaštita sorti u Bugarskoj (Atanassov,2003)Figer 25. Variety protection in Bulgaria (Atanassov, 2003)

55
Opšte semenarstvo
Tabela 6. Broj prijavljenih aplikacija za zaštitu orhideja (1995–2008)Table 6. Number of applications for orchid protection (UPOV, 2009)
Vrsta orhideja / Kind of orchid Broj prijava / Number of applicaitons
Phalaenopsis 346
xDoritaenopsis 47
Dendrodubium 37
Cymbidium 20
Ludisia discolor 9
Vanda 8
xColmanara 2
Miltonia 2
Oncidium 2
Zygopetalum 2
xBratonia 1
Cypripedium 1
xlonocidium 1
Xlwanagara 1
xMiltonidium 1
xOdontonia 1
Spathoglottis 1
xVuystekeara 1
xZelglossoda 1
xGoodaleara 1
xOdontocidium 1

56
Semenarstvo
Tabela 7. Naplata autorstva za sortu pšenice u Češkoj Republici Table 7. Royality for wheat variety in Czech Republic (Krehlik, 2002)
Ukupna vrednost stvorene sorte pšenice (USD)Total value of developed wheat variety (USD) 2 500 000
Rojaliti – 1t ISR semena (USDRoyality – 1t of seed – Certified seed Ist generation (USD) 20
Površina / Area 100 000
Razmena certifikovanog semena (ha)Exchange of certified seed (ha) 50 000
Ukupna prodaja certifiovanog semena (t)Total of certified seed sold (t) 10 000
Ukupan rojaliti godišnje (USD)Total royality annually (USD) 200 000
S obzirom da je Češka Republika zemlja u kojoj su prisutne stra-ne kompanije, svaka od tih zemalja je zaštitila svoje sorte. Prema broju zaštićenih sorti po pojedinim državama, može se zaključiti i koliko je prisustvo pojedinih stranih sorti u semenskoj proizvodnji u Češkoj. U tabeli 8 prikazana je prijava za zaštitu stranih sorti u Češkoj Republici (Krehlik, 2002). Posle Češke, najveći broj zaštićenih sorti imaju Nemač-ka, Slovačka, Holandija i Francuska.
Slika 25. Zaštita sorti u BugarskojFiger 25. Variety protection in Bulgaria (Atanassov, 2003)
SORTA - AUTORPLANT VARIETY - AUTHOR
PRIJAVAAPPLICATION DOCUMENT
KANCELARIJA ZA PATENTEPATENT OFFICE
POZITIVAN DUS TESTPOSITIVE DUS TESTING
AGENCIJA ZA ISPITIVANJE SORTE, POLJSKU INSPEKCIJU I KONTROLU SEMENA
AGENCY FOR VARIETY TESTING,FIELD INSPECTION
AND SEED CONTROL
IZVEŠTA
JREPO
RTSERTIFIK
AC
IJAC
ERTIFICATIO
N
PRIVREM
ENA
LEGA
LNA
ZAŠTITA
TEMPO
RARY LEG
AL PRO
TECTIO
N

57
Opšte semenarstvo
Tabela 8. Broj domaćih i stranih sorti zaštićenih u Češkoj RepubliciTabela 8. Number of domestic and foreing varyeties protected in Czech Republic
(Krehlik, 2002)
Zemlja /Country Broj sorti / No. of varieties
Čečka Republika/Czech Republic 613
Slovačka/Slovakia 158
Nemačka/Germany 177
Holandija/Netherland 117
Francuska/France 57
SAD/USA 35
Velika Britanija/Great Britain 12
Poljska/Poland 8
Austria/Austria 7
Belgija/Belgium 6
Kanada/Canada 4
Mađarska/Hungary 4
Danska/Denmark 2
Italija/Italy 2
Švajcarska/Switzerland 1
Zaštita sorti je od izuzetnog značaja te je i broj sorti koje se prijav-ljuju za zaštitu veliki. Posebno se vodi računa o zaštiti prava oplemenji-vača u SAD-u. Broj zaštićenih sorti pamuka se značajno povećavao od 1970. godine, kada se i počelo sa zaštitom sorti (Plant Variety Protection – PVP), do 2000. godine. Kako slika 26. pokazuje, od 1980. godine broj stvorenih i zaštićenih sorti je imao paralelan tok (Naseem et al., 2001).
Značaj intelektualne svojine će i u narednim dekadama imati odlu-čujuću ulogu kod opredeljivanja pojedinih kompanija o uvođenju sorti na tržišta koja nemaju uređen pomenuti sistem. Oplemenjivanje koje se vrši sve intenzivnije, uz primenu novih tehnologija, skraćuje srednji vek sorte, pogotovo kod povrtarskih biljnih vrsta i ukrasnog jednogodišnjeg bilja. Na slici 27 jasno se vidi koliko je intenzivniji broj introdukovanih sorti od prosečnog veka trajanja sorte u zemljama EU (Naseem et al., 2001).

58
Semenarstvo
Slika 26. Broj prijavljenih i zaštićenih sorti pamuka u SAD Figure 26. Number of protected and realising variety in USA
(http://www.agbioforum.org/v8n23/v8n23a06-oehmke.htm#F2)
Slika 27. Broj prijavljenih sorti i njihov prosečan vek upotrebe Figure 27. Number of realising variety and varietal life span
(http://www.agbioforum.org/v8n23/v8n23a06-oehmke.htm#F2)
Broj
sort
i/N
umbe
r of v
arie
ties
Broj
sort
i/pro
seča
n ve
kN
umbe
r of v
arie
ties/
vari
etal
life
span
(yea
rs)

59
Opšte semenarstvo
„Seme s tavana“
UPOV konvencija iz 1991. godine reguliše plaćanje „semena s ta-vana” (Farm saved seed). Ova odredba predstavlja važan balans iz-među duge tradicije koje poljoprivredni prozvođači neguju čuvajući sopstveno seme i učešća koje daje savremeno oplemenjivanje svojim korisnicima.
Pravo naplate licene za „seme s tavana“ je utemeljeno u zakonu EU iz 1994. godine. Kasnije je ono ugrađeno u Zakon o semenu UK 1998. godine. „Seme s tavana“ nudi iste genetičke mogućnosti kao i sertifiko-vano seme, te zakonska regulativa nalaže plaćanje licence kod semena s tavana“, ali u značajno manjem iznosu od onog koji se plaća za serti-fikovano seme. Treba napomenuti da se licenca naplaćuje uglavnom za ratarske biljne vrste.
Naplata licence za „seme s tavana“ je određena za svaku pojedinu sortu shodno ugovoru između poljoprivredne organizacije i imaoca sorte. Koncept izuzeća farmera “farmers’ exemption” je uveden da bi uveo u sistem male proizvođače koji proizvode manje od 92 t žita i da bi ih izuzeo od plaćanja licence. Mali proizvođači su jedini autorizovani da koriste seme koje su sami proizveli. Različite zemlje imaju i različit procenat „semena s tavana” koje koriste (tab. 9).
Tabela 9. Upotreba semena iz sopstvene proizvodnje žita u nekim zemljama EU.Table 9. Using Farm-Saved Cereal Seed in Some EU Countries.
(Toledo, 2002 )
Država/Counry % semena sa tavana / % of farm save seed
UK/UK 30
Nemačka/Germany 46
Francuska/France 35
Portugalija/ Portugal 75
Španija/Spain 88
Semenska industrija je veliki protivnik „semena s tavana”. Ona je za uklanjanje dve glavne razlike između zaštite sorte i industrijskih

60
Semenarstvo
patenata. To je početak „lista želja” semenske industrije za novu revi-ziju UPOV konvencije. Posebno su veliki zagovornici ujednačavanja patentnog prava i zaštite sorti primenom UPOV sistema kompanije koje imaju proizvode biotehnologije.
Zaštita biljnih sorti je prva standardizovana po UPOV-oj kon-venciji 1961. godine. PVP je uglavnom slična obliku zaštite autorskih prava intelektualne svojine. Vlasnici sorte imaju isključivo pravo nad komercijalizacijom i marketingom zaštićenih sorti, ali malo kontrole nad drugim korisnicima. Farmeri su mogli da umnožavaju semenski materijal za svoje potrebe onoliko dugo koliko su to želeli. Ostali od-gajivači mogu slobodno da koriste zaštićene sorte da razviju sopstveni materijal.
Ovo je dramatično promenio sam UPOV revizijom iz 1991. godine. Na osnovu uspešnog lobiranja iz semenske globalne industrije, revizija PVP pretvorila se u nešto vrlo blisko patentu. Upotreba „semena s ta-vana” je dozvoljena samo kao opcioni izuzetak, ograničenja su stavljena na dalje gajenje, a monopolska prava su produžena sve do berbe proiz- voda. Ovo je verzija UPOV koje se sada brzo širi u zemlje u razvoju, kao rezultat ugovora Svetske trgovinske organizacije – STO (World Tra-de Organization – WTO) i Trgovinskog aspekta zaštite intelektualne svojine TRIPS-a (Trade Related Aspects of Intellectual Property Rgihts – TRIPS) (Silva and Cavalcanti, 2000).

61
Opšte semenarstvo
STRUČNI NADZOR NAD PROIZVODNJOM SEMENA
Dobro organizovano semenarstvo podrazumeva korišćenje dekla-risanog semena u što višem procentu, kao i sorti visokog genetičkog potencijala za rodnost. Stalnom proizvodnjom svih kategorija semena, održava se određeni nivo čistoće sorti, koji predstavlja osnovni pokaza-telj kvaliteta semena. Ukoliko je kategorija semena koju treba proizvesti viša, utoliko su kriterijumi stroži. Za ispunjenje pomenutih kriterijuma potrebno je imati dobro organizovan sistem sertifikacije semena koji podrazumeva poljski pregled useva tokom vegetacije, laboratorijsko is-pitivanje kvaliteta semena, gde oba pokazatelja uslovljavaju deklarisa-nje, kao osnov za puštanje semena na tržište.
Nove sorte, superiorne po nekim kvantitativnim i kvalitativnim osobinama nad postojećim, neprestano se stvaraju kroz proces opleme-njivanja biljaka. Seme tih sorti trebalo bi da se nađe na raspolaganju proizvođačima u dovoljnim količinama, u oblastima za koje su najpo-godnije. Kupci semena treba da budu sigurni da dobijaju dobro seme željene sorte. Da bi se obezbedila garancija genetske čistoće sorte i kva-litetno seme, razvijen je sistem nadzora nad proizvodnjom semenskih useva (poljska kontrola ili aprobacija). Ovo uključuje vođenje evidencije o umnožavanju sorti i kontrolu useva za proizvodnju semena na njivi i ispitivanje semena koje se sprema za tržište u laboratoriji, što se naziva stručnim nadzorom nad proizvodnjom semena ili sertifikacija.
Svrha sertifikcije semena je da obezbedi genetički identitet i gene-tičku čistoću semenskih useva. U prometu komercijalnog semena kupac dobija informaciju samo o sorti i prodavcu i to prihvata kao dovoljno. Kod kupovine semena sorta i njeno poreklo moraju biti verifikovani od ovlašćenih ustanova za seritikaciju i u tom slučaju može se pratiti trag do proizvođača kroz informacije sa etikete na vreći. Pregled use-va u polju, uzorkovanje semena, laboratorijske analize i odgovarajuće obeležavanje na vreći dokazuju da su ispunjeni svi zahtevi u pogledu sertifikacije. Sertifikacija semena predstavlja najbolje osiguranje dobrog kvaliteta semena, poznate čistoće i porekla. Sertifikovanje semena je si-stem umnožavanja ograničen brojem generacija umnožavanja.

62
Semenarstvo
Uslovi za proizvodnju sertifkovanog semena uključuju: – dokaz o tome da je posejano seme bilo korišćeno (dokumentacija
sa vreće), – polje sa usevom koji se smatra semenskim mora biti kontrolisano
od strane inspektora da bi se dokazalo da usev ima odgovarajuću genetičku čistoću i identitet, izolaciju, shodno predviđenim stan-dardima ( izveštaj o poljskoj kontrol i aprobaciji),
– reprezentativan uzorak uzet iz partije semena mora biti testiran na kvalitet kako bi se ustanovilo da li je dobijeni rezultat u sagla-snosti s predviđenim standardima (izveštaj o kvalitetu semena),
– sertifikat za potrebne kvalitete mora biti prisutan prilikom proda-je semena (deklaracija).
Principi organizacije stručnog nadzora nad proizvodnjom semena
Oplemenjivači proizvode seme nove sorte samo u malim količi-nama, jer moraju usredsrediti rad na stvaranje novih sorti. Posao oko umnožavanja semena preuzimaju stručnjaci za semenarstvo, čija je to uža specijalnost. Njihov zadatak je da u procesu umnožavanja semena održe autentičnost sorte kontrolom semenskih useva u polju.
Organizacione šeme za kontrolu proizvodnje semena (sertifikaciju) jako odstupaju od zemlje do zemlje i između biljnih vrsta, ali se šeme uvek zasnivaju na tri opšta načela:
1. Održavanje sorte
Da bi se osiguralo da seme prodato proizvođačima ima određenu sortnu čistoću, umnožavanje mora početi od čistog semena koje čini osnovu za dalje umnožavanje određene sorte. Ovo prvo načelo podra-zumeva da sav semenski materijal može da se prati unazad, sve od one male, početne količine semena koje proizvode oplemenjivači, a koja se zove „nukleus”, materijal za „održavanje” ili „roditeljski materijal”. Ovaj materijal se proizvodi uklanjanjem svih nepoželjnih atipičnih biljaka nastalih iz stranooplodnje. Male količine genetički čistog semena „nu-kleusa” koriste se kao osnova za održavanje sorti, odnosno, buduće po-novljeno umnožavanje.

63
Opšte semenarstvo
Početni materijal za održavanje sorti treba da bude najčistiji. To je polazni materijal sa najvećim odnosom biljaka koje odgovaraju tipu koji je izabrao oplemenjivač u opisu sorte. Ovaj materijal treba da bude pod kontrolom osoblja koje održava odnos umnoženog materijala sa origi-nalnim, preciznim sistemom izbora biljaka na osnovu osobina koje se pažljivo pregledaju i beleže.
2. Ograničeni broj umnožavanja
Broj umnožavanja semenskog materijala pojedinih sorti, posle pri-znavanja je različit. Propisuje ga država zakonskom regulativom, mada je OECD jasno definisao broj generacija umnožavanja kada se radi o semenu namenjenom izvozu. Na primer, predosnovno seme je materijal odobren iz programa za održavanje sorti i koristi se umnožavanjem za proizvodnju osnovnog semena. Seme se zatim umnožava do utvrđenog broja generacija.
Dve ili više generacija sertifikovanog semena (ili ekvivalenta) mogu biti dozvoljene za umnožavanje u neograničenim ciklusima umnožava-nja materijala na nivou predosnovnog ili osnovnog semena jer je takav semenski materijal količinski ograničen, a čistoća se lakše kontroliše nego kod većih površina pod semenskim usevom.
3. Standardi za proizvodnju semena
Donošenje standarda u semensku proizvodnju doprinosi proverava-nju kvaliteta i sortne čistoće semenskih useva uz pomoć poljske inspek-cije (aprobacije). Pažljivo rukovanje semenom na početku proizvodnog lanca smatra se „održavanjem sorte”, mada se u izvesnom smislu i ceo proces kontrole proizvodnje semena sa ograničenim brojem generacija umnožavanja i utvrđenim standardima čistoće može smatrati procesom za održavanje početnih karakteristika sorte.
Kontrola proizvodnje semena je put za proizvodnju genetički čistog semena određenog kvaliteta, a primenjuje se na sorte koje su priznate. Seme ovih sorti se umnožava preko serije generacija. Izraz predosnovno, osnovno i sertifikovano seme se koristi za određivanje raznih generacija umnožavanja semena. Zahtev za kvalitetom semena u ovim kategorija-ma se razlikuje, naročito u oceni sortne čistoće. Predosnovno i osnov-no seme su namenjeni samo umnožavanju semena. Generacije koje se

64
Semenarstvo
dobijaju umnožavanjem osnovnog semena se nazivaju sertifikovano seme i tu postoji više generacija umnožavanja.
Poljska kontrola semenskih useva
Sertifikacija semena je legalni sistem za kontrolu kvaliteta umnoža-vanja i proizvodnje semena koji se sastoji od sledećih postupaka:
a) kontrola semenskih useva i skladišta, b) pre i posle kontrolnih testova,c) testova kontrole kvaliteta (OECD, 2003). Procena sortnog identiteta i čistoće za vreme proizvodnje semena je
od ključnog značaja za održavanje visokog standarda kvaliteta semena. Proizvođač semena mora da osigura da se ništa ne dogodi u vreme vege-tacije, žetve, dorade, pakovanja i obeležavanja partija semena ili kasnije distribucije, što bi moglo negativno da utiče na kvalitet semena,
OECD šema za sertifikaciju semena ima dve procedure koje su sači-njene tako da proveravaju status sorte u različitim stadijumima njegove proizvodnje:
a) gajenje biljaka na kontrolnim parcelama i izvođenje laboratorij-skih tesova semena i ponika pri čemu se koriste uzorci iz partija semena koje su posejane,
b) poljski pregled semenskog useva tokom vegetacije, jednom ili više puta, radi izdavanja izveštaja (sertifikata) o stanju useva.
Kod vršenja pomenutih testova i poljskih pregleda neophodno je da se usvoje tehnički metodi koji će omogućiti dobijanje rezultata dovoljne tačnosti i pouzdanosti.
Metode opisane u uputstvima OECD-a su izgrađivane tokom niza godina, kako bi dale zadovoljavajuće rezultate i stvorile principe na ko-jima takve metode treba da se baziraju. Metode su usvojile zemalje čla-nice OECD i one koje nisu članice, a koje učestvuju u OECD šemi za sertifikaciju semena i razmenjuju sertifikovano seme u međunarodnoj trgovini. Od nacionalno ovlašćenih ustanova, u svakoj zemlji, za rad OECD šeme za sertifikaciju semena, se zahteva da koristi navedene me-tode ili, ukoliko to neka zemlja želi, da bi pojednostavila postupak, da ih ugradi u sopstvenu regulativu.

65
Opšte semenarstvo
Kontrolne test parcele
Kontrolne test parcele se koriste za posmatranje identiteta i čistoće sorte u različitim stadijumima umnožavanja semena i na taj način se dokazuje da je kvalitet semena proizvedenog po OECD šemi za serti-fikaciju semena na zadovoljavajućem nivou. Organizovane su tako da daju odgovor na dva pitanja:
– da li se karakteristike biljke dobijene iz uzetog uzorka semena slažu sa opisom karakteristika sorte i na taj način potvrđuje njen identitet,
– da li biljke dobijene iz uzetog uzorka semena poseduju sortnu či-stoću predviđenu standardima za tu osobinu.
Na prvo pitanje može da se odgovori vršenjem vizuelnog poređenja između biljaka sa kontrolne parcele posejane semenom iz uzorka koji reprezentuje partiju semena i biljkama sa parcele dobijene sejanjem se-mena iz referentnog uzorka, u daljem tekstu „standardni uzorak”.
Drugo pitanje zahteva identifikaciju biljaka koje odstupaju od sor-tnih karakteristika, tzv. „of type” biljaka unutar kontrolne parcele tako da njihov broj može da se poveže sa standardima predviđenim u OECD šemi za sertifikaciju semena. Ovaj test meri uniformnost partije seme-na i određuje da li su ili ne karakteristike sorte ostale nepromenjene za vreme umnožavanja semena ili nisu. Test takođe označava efektivnost ograničenja broja generacija umnožavanja.
Pred-kontrolni testovi
Kada se seme umnožava radi proizvodnje sledeće generacije, in-formacija dobijena sa kontrolne parcelice je bezvredna zbog toga što ovlašćenoj ustanovi daje podatke o identitetu i kvalitetu koje su bile na raspolaganju otprilike istovremeno ili pre nego što je sledeći semen-ski usev spreman za poljski pregled. U ovoj fazi pred-kontrolni test je postavljen istovremeno kada i semenski usev sledeće generacije. Rezul-tati dobijeni u pred-kontrolnim testovima upoređuju se sa rezultatima dobijenim pri poljskom pregledu useva. Tako se dobija ključna informa-cija o čistoći sorte i rezultati postaju integralni deo procesa sertifikacije semena.

66
Semenarstvo
Posle-kontrolni testovi
Posle-kontrolni testovi ukazuju na kvalitet proizvedenog semena, a rezultati obično nisu na raspolaganju do kraja sledeće sezone, nakon što je seme požnjeveno. Posle-kontrolni tesovi su vredni jer utvrđuju koliko je efikasan ili neefikasan proces proizvodnje semena u održavanju sortne čistoće i identifikovanju načina za poboljšanje sistema proizvodnje. Upo-ređenjem između biljaka gajenih iz proizvedene partije semena i onih ga-jenih iz standardnog uzorka, ovlašćena ustanova može da utvrdi kvalitet i da garanciju da su se proizvođači pridržavali minimalnih standarda.
Za seme koje se dalje umnožava, jedna kontrolna parcela može da ima dve funkcije: posle-kontrolisanje partije semena od poslednje žetve i pred-kontrolisanje semenskog useva za sledeću žetvu. Kod hibridnih sorti, zbog toga što se sortni identitet i čistoća hibrida ne mogu veri-fikovati tokom proizvodnje, neophodno je da se obezbedi proizvodni kvalitet u posle-kontrolnim testovima.
Hibridna sorta posmatrana u posle-kontrolnim testovima mora od-govarati sorti, a biljke moraju da imaju izgled sa karakteristikama hibrida koje je ovlašćena ustanova dobila u vreme njene registracije. Roditeljske komponente za proizvodnju hibrida moraju biti praćene na pred-kontrol-nim parcelama (sl. 28).
Standardni uzorak je uzo-rak oplemenjivačevog semena sa karakteristikama identič-nim sorti koja je prijavljena sortnoj komisiji . Služi za pro-veru sortnog identiteta i njene genetičke čistoće. Svrha stan-dardnog uzorka je da obezbe-di izvorni opis sorte; njegovo održavanje i autentičnost koji
Slika 28. Ogledno polje za posle-kontrolu
Figure 28. Experimental field for post-control test
(foto: Denčić)

67
Opšte semenarstvo
su od ključnog značaja. U pred-kontroli i posle-kontroli standardni uzorak se koristi za poređenje sa biljkama gajenim iz uzoraka partije semena koje su bile na tržištu. Koliko razultati dobijeni u posle-kontrol-nim testovima mogu da odstupaju kod ratarskih i povrtarskih biljnih vrsta prikazano je na slici 29. Odstupanje sortne čistoće je kod ratarskih biljnih vrsta značajno manje nego kod povrtarskih.
Slika 29. Rezultati posle-kontrolnih testova kod ratarskih i povrtarskih biljnih vrsta Figure 29. Results of post-control tests in field and vegetable plant crops
(Milošević i Malešević, 2004)
Prethodni usev – Kod postavljanja kontrolnih parcela, ovlašćena organizacija mora da vodi računa da obezbedi da polje bude pogod-no za postavljanje ogleda. Ne sme postojati rizik od kontaminacije od samoniklih biljaka istih ili srodnih biljnih vrsta. To se radi proverom prethodnog useva sa polja koje će se korstiti i obezbeđivanjem pažljivo planiranog plodoreda.
Laboratorijski testovi
Pored pregleda biljaka na kontrolnim parcelama postoji još nekoliko laboratorijskih tesova koji mogu da se koriste za identifikaciju sorti nekih biljnih vrsta. Prva klasifikacija može ponekad da se izvrši vizuelnim po-smatranjem osobina semena kao što su oblik, boja, šara i drugih fizičkih osobina. Ovim se ne identifikuje samo vrsta, nego i klasifikaciona grupa i čak mogu da se identifikuju pojedinačna semena u mešavini u uzorku.

68
Semenarstvo
Ponik može da pruži dodatne karakteritike, na primer prisustvo ili odsustvo antocijanske pigmentacije u koleoptilu raži. Kod nekih vrsta nivo ploidnosti može da se koristi za klasifikovanje sorti, na primer di-ploidnog i tetraploidnog ljulja (OECD, 2008).
Druga klasifikacija je moguća kod nekih biljnih vrsta korišćenjem hemijskih testova. Kod uljane repice može da se koristi test na prisustvo eruka kiselina i sadržaj glukozinolata; za pšenicu reakcija semena na fenol uspešno se koristi za identifikaciju semena koja su primesa druge sorte.
Elektroforeza i druge hemi-taksonomske tehnike se uspešno koriste za identifikaciju sorti kod nekih useva. Ova tehnika se pokazala kori-snom u svrhu priznavanja sorti gde je broj semena, koji treba da se ispita za određenu sortu, relativno mali. Elektroforeza se koristiti u procesu sertifikacije semena za određivanje čistoće sorte. Ona može da se ko-risti za potvrđivanje identiteta malog broja pojedinačnih semena tamo gde se na osnovu drugih testova ne mogu izvesti pouzdani zaključci. U slučaju hibrida suncokreta, uljane repice ili kukuruza moguće je proce-niti nivo samooplodnje korišćenjem elektroforeze. Sortna čistoća ovih hibrida takođe može da se proceni ovom tehnikom.
Poljski pregled semenskih useva
Svrha poljskih pregleda je provera da li semenski usev pokazuje ka-rakteristike zasejane sorte (sortni identitet) i da obezbedi da ne dođe do okolnosti koje bi mogle biti štetne za kvalitet semena koje će biti po-žnjeveno (sortna čistoća). Semenski usev može više puta da se pregleda tokom vegetacione sezone što zavisi od biologije biljne vrste. Ono što se ne sme izostaviti je da se izvrši bar jedan poljski pregled za vreme perioda cvetanja ili pre prašenja polena koji omogućava ocenu sortnog identiteta. Za neke useve se zahteva poljski pregled u punoj zrelosti, jer je u toj fazi najlakše uočiti eventualne sortne nečistoće.
Mada se tehnike poljskog pregleda razlikuju u detaljima, u zavisno-sti od određnih osobina svake vrste, glavni principi za proveru useva pri poljskoj kontroli su sledeći:
– istorija prethodnog useva na polju treba da bude takva da se rizik od nepoželjnih samoniklih biljaka iste ili srodne vrste, koje bi mo-gle da kontaminiraju semenski usev, svede na minimum,
– semenski usev treba da bude dovoljno izolovan od drugih useva kako bi se smanjio rizik od kontaminacije neželjenim polenom (tab. 10),

69
Opšte semenarstvo
– da bude fizički izolovan kako bi se sprečila mehanička mešavina pri žetvi,
– da bude izolovan od izvora bolesti koje se prenose semenom,– da bude slobodan od korova i semena drugih vrsta, posebno onih
čije bi seme teško moglo da se izdvoji od semena useva tokom dorade,
– da bude slobodan od bolesti koje se prenose semenom,– da ima odgovarajući sortni identitet (tab 11),– ne sme da bude prisutno više atipičnih biljaka od broja dozvolje-
nog po standardu za sortnu čistoću,– ne sme da postoji više biljaka drugih vrsta nego što to standard
dozvoljava, – za hibridne sorte odnos roditeljskih komonenti treba da bude za-
dovoljavajući i definisan od strane održivača.
Pregled semenskog useva treba da bude dopunjen rezultatima sa pred-kontrolnih parcela, koje ovlašćena ustanova stalno nadzire, a koji će poslužiti inspektoru kao pouzdani podatak o sortnom identitetu i aspektima čistoće relevantnim za upotrebljenu partiju semena.
Proizvođači semena traba da zadrže bar jednu etiketu sa vreće svake partije semena korišćene za setvu, da bi se autorizovao identitet pose-janog semena, ukoliko je to potrebno. Proizvođač semena mora da po-kaže etiketu svake semenske partije posejane u polju, tako da inspektor može jasno da je vidi. Za hibride, etikete partija semena korišćenih za liniju oca i liniju majke moraju se čuvati i verifikovati.
Sortna čistoća hibrida dobijena u proizvodnji useva može da se pro-veri samo u posle-kontrolnoj parceli zasejanoj uzorcima proizvedenog semena hibrida. Sortna čistoća može da se obezbedi pod uslovom da postoji:
– adekvatna izolaciona distanca od izvora kontaminirajućeg polena,– dobri uslovi za rasejavanje polena (tab. 12),– visoki nivo muške sterilnosti ženskog roditelja,– nizak nivo srodstva,– visoki nivoi sortne čistoće oba roditelja,– odvojena žetva muške komponente pre roditelja koji nosi seme
(ženskog)

70
Semenarstvo
Tabela 10. Minimalni zahtevi za izolacione distance kod pojedinih biljnih vrstaTable 10. Minimum requirements for isolation distances for some field crops
(OECD, 2008)
Biljna vrsta, porodicaPlant species, family
Za parcele od 2 ha i manjeFor fields of 2 ha or less
Za polja veća od 2 ha
For field larger than 2 ha
LeguminosaeZa proizvodnju / Field to produce:- seme za dalje umnožavanje
seed for further multiplication /- seme za druge svrhe ili stočnu hranu /
seed for anemity purposes or fodder production
200 m
100 m
100 m
50 m
Žita – Cereals (osim hibria/Except hybrids)Za proizvodnju / Field to produce:Za samooplodne vrste- bazno seme / Basic seed- sertifikovano seme / Certified seedZa samoplodni triticale:- bazno seme / Basic seed- sertifikovano seme / Certified seed
300 m250 m
50 m20 m
Za sve veličine polja/All size fields
Helianthus annuusZa proizvodnju / Field to produce:- bazno seme (hibride sorte)
Basic seed (hybrids varietis)- bazno seme (druge vrste osim hibirida/
Basic seed (varieties other than hybrid)- sertifikovano seme / Certified seed
1.500 m
750 m
500 m
Zea maysZa proizvodnju / Field to produce:-bazno seme / Basic seed- sertifikovano seme / Certified seed
200 m200 m

71
Opšte semenarstvo
Tabela 11. Sortna čistoća semenskih useva pojedinih biljnih vrstaTable 11 Varietal purity in seed production for some feld crops
(OECD, 2008)
Biljna vrstaSpecies
Osnovno semeBasic seed(%)
Sertifikovano seme prve generacije
Certified seed first generation
(%)
Sertifikovano seme druge generacije
Certified seed second generation
(%)
Pisum sativum 99.7 99.0 98.0Vicia faba 99.7 99.0 98.0Glycine max 99.5 99.0 99.0Brassica napus var. oleiferaOsim sorti isključivo tipa krmnog bilja kao što je navedeno u OECD listi sortiExcept varieties of strictly the fodder type as indicated in the OECD List of Varieties
99.9 99.7 99.7
Brassica rapaOsim sorti isključivo tipa krmnog bilja kao što je navedeno u OECD listi sortiExcept varieties of strictly the fodder type as indicated in the OECD List of Varieties
99.9 99.7 99.7
Brassica oleracea convar. acephala 99.7 99.0 98.0
Brassica napus var. napobrassica 99.7 99.0 98.0
Sinapis alba 99.7 99.0 98.0Helianthus annuus 99.7 99.0 98.0Pisum sativum 99.7 99.0 98.0Vicia faba 99.7 99.0 98.0Linum usitatissimum 99.7 98.0 97.5Papaver somniferum 99.0 98.0 98.0Triticum aestivum 99.9 99.7 99.0Hordeum vulgare 99.9 99.7 99.0Avena sativa 99.9 99.7 99.0Oryza sativa 99.9 99.7 99.0Uglavnom samooplodne sorte X TriticosecaleMainly self-pollinating varietis of X Triticosecale
99.7 99.0 98.0

72
Semenarstvo
Tabela 12. Minimalni zahtevi i standardi za proizvodnju roditeljskih linija i sertifiko-vanog hibridnog semena šećerne repe po OECD šemi
Table 12. Minimum requirements and stanards for the production of parent lines and certified hybrid seed under OECD sugar beet seed scheme (OECD, 2007)
Usev za proizvodnju / Crop for production
Bazno seme roditeljskih linija: roditeljske linije koje proizvode semeBasic seed of parent lines Parent line for seed pordution
sa biljkama koje praše polen, koje uključuju atipične biljke . . . . . . . . . . . . 99,8%with plant which poninate, incuding pollen grain of parent lines
polen roditeljske linije . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99,8%pollen grain of parent lines
bazno seme roditeljskih hibrida, seme roditeljske linije . . . . . . . . . . . . . . . . 99,7% basic seed of hybrids, seed of parent lines
sa muško fertilnih biljaka, u koje se ubrajaju atipične biljke . . . . . . . . . . . . 99,5%from male fertile plants, including out cross
Sertifikovano seme hibridnih sorti: roditeljske linije koje proizvode semeHybrid certified seed
sortna čistoća. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99,0%varietal purity
muška sterilnost . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99,5%male sterile
polen roditeljske linije . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99,5%pollen grain of parent lines
U post kontroli: sertifikovano seme hibridnih sorti . . . . . . . . . . . . . . . . . . . 95,0%In post control: certified hybrid seed

73
Opšte semenarstvo
Sprovođenje stručnog nadzora (sertifikacije) nad proizvodnjom
Sistemom sertifikacije semena se postiže:
– stalno povećanje zastupljenosti visokoprinosnih i kvalitetnih sorti,– identifikacija novih sorti i povećanje njihove zastupljenosti pod
odgovarajućim i opšte prihvaćenim imenima,– obezbeđivanje neprekidnog snabdevanja tržišta određenim kate-
gorijama semena dobrog kvaliteta.
Važno je napomenuti da je:
– sertifikacija semena samo jedan od podržavajućih elemenata se-menskog programa,
– sertifikacija semena nije umnožavanje semena i njegova proi-zvodnja, ona je sistem za izvršavanje ovih zadataka,
– sertifikacija semena nije službena kontrola i usmeravanje marke-tinga semena, nju ne interesuje ko proizvodi već kako se proizvodi seme.
U većini zemalja sertifikacija semena se postepeno razvijala od skromnih početaka, često bez znanja šta se radi u drugim zemljama, te otuda velike razlike između nacionalnih postupaka za sertifikaciju. Ove razlike potiču od činjenice da su za vreme razvoja sistema sertifi-kacije postupci trebalo da se prilagode lokalnim uslovima i postojećim programima organizacije semenarstva. Na šemi sertifikacije semena u našoj zemlji pokazano je kako ide put novostorene sorte u procesu kon-trolisanog umnožavanja primenom sistema sertifikacije. Instituti, kre-atori sorti održavaju predosnovno seme, jer najbolje poznaju njegove karakteristike. Osnovno seme i niže kategorije semena se umnožavaju sklapanjem ugovora sa prozvodnim organizacijama. Kontrolu ove pro-izvodnje prati država, odnosno, poljoprivredne stručne službe (poljo-privredne stanice). Njihovi stručnjaci vrše ocenjuju useve u polju. Na osnovu zapisnika koje poljoprivredne stručne službe sačine, a broj zapi-snika predstavalja broj poljskih pregleda predviđenih pravilnicima, Mi-nistarstvo poljoprivrede, trgovine, šumarstva i vodoprivrede Republike Srbije izdaje uverenje o poljskoj kontroli (aprobaciono uverenje) (sl. 30).

74
Semenarstvo
Posle žetve i dorade, ovlašćene, akreditovane laboratorije ustanov-ljavaju kvalitet semena. Na osnovu uverenja o poljskom pregledu, koje izdaje poljoprivredna stručna služba i uverenja o kvalitetu semena, koje izdaje ovlašćena, akreditovana laboratorija, doradni centar izdaje dekla-raciono uverenje. Organizacija koja je izvršila doradu, dužna je da o ko-ličinama semena izvesti ovlašćenu organizaciju za izdavanje etiketa za vreće, jer će dobiti toliko etiketa koliku količinu semena i broj pakova-nja semena u partiji je prijavila. Kada dorađivač dobije etikete, deklariše seme (stavljanje etiketa na vreće). Tek dobijanjem deklaracionog uvere-nja (lista) vlasnik semena može deklarisano seme da stavlja u promet.
Slika 30. Šema sertifikacije semena kod nasFiger 30 Organiaztion of seed certification in our country
(Milošević, Malešević, 2004)
INSTITUTIOplemenjivanjeINSTITUTES
Breeding
SERTIFIKACIJAPoljska kontrola
CERTIFICATIONField control
SERTIFIKACIJAKontrola kvaliteta semena
CERTIFICATIONSeed quality control
REGIONALNE ORGANIZACIJE Žita, krmno bilje, povrće
REGIONAL ORGANIZATIONSCereals, forage crops, vegetables
INDUSTRIJA ULJA I ŠEĆERAŠećerna repa, suncokret, soja
SUGAR AND OIL INDUSTRYSugar beet, sunflower, soybean
POLJOPRIVREDNEORGANIZACIJEAGRICULTURAL
ORGANIZATIONS
POLJOPRIVREDNE STANICE Poljska kontrola
EXTENTIONS SERVICIESField control
LABORATORIJAKontrola kvaliteta semena
LABORATORYSeed quality control
DORADADeklaracija
SEED PROCESSINGLabeling
TRGOVINSKE ORGANIZACIJEProdaja i distribucija
TRADE ORGANIZATIONSSale and distribution

75
Opšte semenarstvo
Često u zemlji postoji potreba za uvozom semena bilo da su u pitanju nepovoljni klimatski uslovi ili nedovoljna proizvodnja. Postoji moguć-nost da domaća semenska industrija mora da pronađe mogućnost izvo-za za višak svojih proizvoda. Zbog toga postoji međunarodni postupak sertifikacije semena koji treba imati u vidu kada se razvija nacionalna sertifikaciona šema. Izvoz i uvoz semena je uspešniji ako postoji uzaja-mno priznavanje kategorija sertifikovanog semena i primenjenih kontrol-nih mera.
Setimela i sar. (2004) su dali svoje viđenje o organizaciji održavanja sorte i proizvodnji osnovnog semena na različite načine, odnosno u za-visnosti od toga kako se razvija program proizvodnje semena. U daljem tekstu dat je prikaz četiri načina organizovanja održavanja sorte, odno-sno načina proizvodnje semena:
1. Rad na selekciji biljaka, proizvodnji i prometu semena je u okvi-ru jedne organizacije, s tim što održavanje sorti i proizvodnju osnovnog i komercijalnog semena vrši oplemenjivač, odnosno oplemenjivačka kuća.
2. Rad može da se podeli na oplemenjivanje biljaka, održavanje sor-te i proizvodnju osnovnog semena kao jednu celinu i proizvodnju komercijalnog semena kao drugu. Na ovaj način posluju mnoge institucije koje se kreću u pravcu privatizacije, gde je prethodno državna semenska proizvodnja, kao najatraktivnija oblast za pri-vatizaciju, prodata ili komercijalizovana.
3. Treći način je kada je proces oplemenjivanja odvojen od organi-zacije koja se bavi proizvodnjom semena (osnovnog i komerci-jalnog).
4. Svi učesnici u programu proizvodnje semena su odvojeni.

76
Semenarstvo
ORGANIZACIJE KOJE SE BAVE SEMENARSTVOM
Međunarodne organizacije koje se bave semenarstvom
U cilju obezbeđivanja dovoljnih količina sortnog i kvalitetnog se-mena u svetskim razmerama ustanovljene su međunarodne organiza-cije koje svojom aktivnošću i povezanošću sa drugim organizacijama iz oblasti poljoprivrede ostvaruju pre svega sistem kontrole semena u međunarodnom prometu, kao i potreban kvalitet semena. Ako se ima u vidu da je samo jedna trećina stanovništva zemljine kugle u situaciji da obezbedi odgovarajuću ishranu, a da se dve trećine nalaze na mini-mumu egzistencije, te ako se zna da će stalno rastuća populacija ljudi do 2015. godine iznositi oko osam milijardi, ne treba naglašavati potrebu postizanja visokih prinosa, a prvi uslov za to je korišćenje kvalitetnog sortnog semena za setvu. Radi obezbeđivanja napred pomenutog formi-rane su brojne međunarodne organizacije, koje su razvijanjem svojih si-stema omogućile obezbeđenje standarda kvaliteta na globalnom nivou.
Organizacija za hranu i poljoprivredu Organizacija za hranu i poljoprivredu osnovana je pod pokrovitelj-
stvom Ujedinjenim nacija (FAO – Food and Agricultural Organization of the United Nations). Glavni zadatak FAO-a je da spreči glad u svetu. U FAO-u su jednako zastupljene razvijene zemlje i zemlje u razvoju, koje debatom dolaze do rešenja zajedničkih problema. Pored toga zadatak FAO-a je da pomogne zemljama u tranziciji da unaprede svoju poljopri-vredu, kako bi imale dovoljno kvalitetne hrane. Postoji posebno odeljenje FAO-a koje vodi statističke podatke o proizvedenim količinama hrane. Od 2001. godine održavaju se redovni sastanci predstavnika zemalja Cen-tralne i Istočne Evrope i zemalja u tranziciji, koji se bavi pitanjem razvoja semenarstva. Sagledavanje sveukupnog stanja semenarstva u zemljama u tranziciji, postojeće zakonske regulative iz oblasti semenarstva, postojećih

77
Opšte semenarstvo
institucija za očuvanje biljnih i životinjskih resursa, širenja informacija o načinu upravljanja granom semenarske proizvodnje i prenošenja znanja iz drugih zemalja postavio je sebi u zadatak FAO. Na prvom inicijalnom sastanku održanom u Budimpešti 2002. godine formirane su radne grupe sastavljene od predstavnika iz raznih zemalja u tranziciji čiji je cilj da sažmu informacije i daju ideju ujednačavanja sistema iz gore navedene oblasti.
Međunarodna organizacija za trgovinu semenom
Međunarodna organizacija za trgovinu semenom (International Seed Federation – ISF) predstavlja interese svojih članica na međunarodnom nivou, uspostavlja saradnju između članica i pomaže rešenju nastalih me-đusobnih problema. Razvija i omogućava slobodan promet semena sa pravičnim i korektnim regulativama koji istovremeno pomaže proizvo-đačima i korisnicima semena. Promoviše, uspostavlja i štiti intelektual-nu svojinu koja prati ulaganja u oplemenjivanje, podiže svest o vrednosti oplemenjivačkog doprinosa u smislu obezbeđenja hrane, posmatrano u globalnim razmerama i održive poljoprivrede, posebno sa proizvodnjom i upotrebom visokokvalitetnog semena dobijenog primenom metoda modernih tehnologija. Ova organizacija ima obavezu da unapredi pra-vila o proizvodnji i prodaji semena i ostalog reproduktivnog materijala publikovanjem Pravila o prometu semena na međunarodnom tržištu i za licenciranje nove tehnologije. ISF posreduje u slučajevima arbitraže, po-maže i daje podršku u organizovanju i razvoju nacionalnih i regionalnih semenarskih asocijacija, pomaže i podržava obrazovanje kadrova. ISF je od posebne važnosti za sve one koji se bave semenom i zbog toga što ko-ordinira sa svim drugim međunarodnim organizacijama koje su poveza-ne, direktno ili indirektno, sa semenom, a neke od njih je i sama osnovala.
Forum ISF-a sarađuje sa:– Međunarodnom organizacijom za ekonomsku saradnju i razvoj (OECD)– Međunarodnom organizacijom za hranu i poljoprivredu (FAO)– Međunarodnom unijom za zaštitu novih biljnih sorti (UPOV), – Međunarodnom organizacijom za ispitivanje semena (ISTA), – Svetskom trgovinskom organizacijom (WTO), – Evropskom semenarskom asocijacijom (ESA)– Azijsko–pacifičkom semenarskom asocijacijom (APSA), – Udruženjem latinoameričkih semenarskih asocijacija (FELAS),

78
Semenarstvo
Međunarodna pravila u trgovini semenom
Proceduralna pravila za rešavanje nesuglasica za trgovinu semenom namenjenog setvi i menadžment intelektualne svojine – posredovanje, pomirenje i arbitraža su uspostavljena ranih tridesetih godina prošlog veka i daju uputstva za posredovanje i pomirenje i njihove amandmane usvojene 2001. godine, koji su stupili na snagu januara 2002. godine. Po-sredovanje, pomirenje, arbitraža za nesuglasice između stručnjaka koja se odnose na menadžment prava intelektulne svojine u oblasti opleme-njivanja biljaka i etički kod za arbitre. Ono što je potrebno naglasiti je da su arbitražne ustanove retke u svetu i da postoji samo nekoliko takvih, od kojih je jedna FIS arbitražna komora.
Pravila za ispitivanje semena i međunarodni serifikati za ispitiva-nje kvaliteta semena su ustanovljeni zahvaljujući FIS-u 1931. godine u Vageningenu koji je predvideo važnost donošenja ovakvih pravila.
ISF je imao saradnju i sa organizacijom koja je prethodila OECD-u, OEED (Organizacija za evropsku ekonomsku saradnju) do osnivanja OECD Šeme za sertifikaciju krmnog bilja u međunardnom prometu. ISF je uspostavila plan globalnog osiguranja protiv grešaka i propusta u semenskoj industriji na globalnom nivou. Ovo osiguranje obuhvata sve biljne vrste, uključujući i genetički modifikovane organizme (Gene-tically Modified Organisms – GMO), na svim kontinentima.
Pravila i postupci u trgovini semenom namenjenog setvi
Pravila i postupke u trgovini semenom namenjenog setvi usvojila je generalna skupština FIS-a u Ostendu 1. juna 1994. godine, a stupila su na snagu 1. januara 1995. godine. Ona su zamenila sva prethodna FIS pravila, sa izuzetkom onih koja se odnose na seme drveća i šiblja. Naj-novije izdanje je stupilo na snagu 1. januara 2002. godine.
Pravila ukazuju veoma detaljno i precizno na:– uslove zaključivanja ugovora (potvrđivanje ugovora, ugovor kao
predmet odobravanja uvoza ili izvoza, ugovor kao predmet ugo-varanja setve i umnožavanja, uvozne regulative),
– uslove ugovora (količina, kvalitet, pakovanje, rokovi isporuke, do-kumentacija, osiguranje),

79
Opšte semenarstvo
– izvršavanje ugovora (obaveštenje o nameri za slanje pošiljke sa instrukcijama, nedostaci u instrukcijama kod slanja pošiljke, za-kašnjenje pošiljke, obaveštenje o pošiljci, poništavanje pošiljke, isticanje ugovora, plaćanje),
– proveru kvaliteta i analize (kvaliteta semena, primenjene analize, kontrola identiteta sorte),
– razmirice ( žalbe, viša sila, arbitaža).
Međunarodna unija za zaštitu novih biljnih sorti
Međunarodna unija za zaštitu novih biljnih sorti (The Internatio-nal Union for the Protection of New Varieties of Plants - UPOV) je me-đudržavna organizacija koja primenjuje Međunarodnu konvenciju za zaštitu novih sorti bilja (Convention for the Protection of New Varieties of Plants), prvi put potpisanu u Parizu decembra 1961. godine. Cilj Konvencije je da obezbedi ekskluzivno pravo oplemenjivača na zaštitu njihovih novostvorenih sorti. Konvencija je nekoliko puta menjana, a dve verzije su sada u upotrebi i to verzija iz 1978. godine i 1991. go-dine.
Oba akta UPOV konvencije, iz 1978. godine i 1991. godine, odre-dila su minimalni obim zaštite nove biljne sorte, a pružaju državama članicama mogućnost da u svoju zakonsku regulativu unesu specifič-nosti.
Konencijom iz 1978. godine, obezbeđuje se minimalni obim prava oplemenjivača, autora sorte zahteva neophodnost prethodnog ovlašće-nja imaoca prava za proizvodnju u svrhu konvencionalnog marketin-ga, ponude za prodaju i marketing materijala za umnožavanje zaštićene sorte. Konvencija iz 1991. godine sadrži detaljnije odredbe koje definišu akte koji se odnose na materijal za umnožavanje u vezi sa kojim se au-torizacija imaoca sorte traži. Izuzetno, ali samo tamo gde imalac sorte nije imao opravdanu mogućnost da koristi svoje pravo, u vezi sa mate-rijalom za umnožavanje, njegova autorizacija može da se zahteva u vezi sa bilo kojim specifičnim aktima vezanim za požnjeveni materijal sorte. U tabeli 13 dat je pregled zemalja članica UPOV, vreme pristupanja ovoj organizaciji, kontribucija koju poseduju, kao i Konvencija kojoj su pri-stupile.

80
Semenarstvo
Tabe
la 1
5. D
ržav
e čl
anic
e U
POV-
a (U
POV,
200
9)Ta
ble 1
5. M
embe
rs o
f UPO
V (U
POV,
200
9)
Drž
ava
/org
aniz
acija
Stat
e / o
rgan
izat
ion
Dat
um k
ada
je d
ržav
a /
orga
niza
cija
pos
tala
čl
an U
POV
/D
ate
of U
POV
Mem
ber
ship
Broj
uč
estv
ujuć
ih
jedi
nica
/N
umbe
rs o
f un
its
Usv
ojen
a K
onve
ncija
/C
onve
ntio
n w
hich
is
adop
ted
Dat
um k
ada
je d
ržav
a /
orga
niza
cija
pos
tala
deo
K
onve
ncije
/D
ate
whe
n is
Cou
ntry
/or
gani
zatio
n be
com
e
mem
ber
Alb
anija
/Alb
ania
Okt
obar
/Oct
ober
15,
200
50.
219
91 A
kt/A
ctO
ktob
ar/O
ctob
er 1
5, 2
005
Arg
entin
a/A
rgen
tina
Dec
emba
r/D
ecem
ber 2
5, 1
994
0.5
1978
Akt
/Act
Dec
emba
r/D
ecem
ber 2
5, 1
994
Aust
ralij
a/Au
stra
liaM
art/M
arch
1, 1
989
1.0
1991
Akt
/Act
Janu
ar/Ja
nuar
y 20
, 200
0Au
strij
a/Au
stria
Juli/
July
14,
199
40.
7519
91 A
kt/A
ctJu
li/Ju
ly 1
, 200
4A
zerb
ejdž
an/A
zerb
eija
nD
ecem
bar/
Dec
embe
r 9, 2
004
0.2
1991
Akt
/Act
Dec
emba
r/D
ecem
ber 9
, 200
4Be
loru
sija/
Bela
rus
Janu
ar/Ja
nuar
y 5,
200
30.
219
91 A
kt/A
ctJa
nuar
/Janu
ary
5, 2
003
Belg
ija/B
elgi
um2
Dec
emba
r/D
ecem
ber 5
, 197
61.
519
61/1
972
Akt
/Act
Dec
emba
r/D
ecem
ber 5
, 197
6Bo
livija
/Bol
ivia
(Plu
rinat
iona
l St
ate
of)
Maj
/May
21,
199
90.
219
78 A
kt/A
ctM
aj/M
ay 2
1, 1
999
Braz
il/Br
azil
Maj
/May
23,
199
90.
2519
78 A
kt/ A
ctM
aj/M
ay 2
3, 1
999
Buga
rska
/Bul
garia
April
/Apr
il 24
, 199
80.
219
91 A
kt/A
ctAp
ril/A
pril
24, 1
998
Kan
ada/
Can
ada
Mar
t/Mar
ch 4
, 199
11.
019
78 A
kt/A
ctM
art/M
arch
4, 1
991
Čile
/Chi
leJa
nuar
/Janu
ary
5, 1
996
0.2
1978
Akt
/Act
Janu
ar/Ja
nuar
y 5,
199
6K
ina/
Chi
naAp
ril/A
pril
23, 1
999
0.5
1978
Akt
/Act
3/A
prilA
pril
23, 1
999
Kolu
mbi
ja/C
olom
bia
Sept
emba
r/Se
ptem
ber 1
3, 1
996
0.2
1978
Akt
/Act
Sept
emba
r/Se
ptem
ber 1
3, 1
996
Kost
arik
a/C
osta
Ric
aJa
nuar
/Janu
ary
12, 2
009
0.2
1991
Akt
/ Act
Janu
ar/Ja
nuar
y 12
, 200
9

81
Opšte semenarstvo
Hrv
atsk
a/C
roat
iaSe
ptem
bar/
Sept
embe
r 1, 2
001
0.2
1991
Akt
/Act
Sept
emba
r/Se
ptem
ber 1
, 200
1Č
eška
Rep
ublik
a/C
zech
Rep
ublic
Janu
ar/Ja
nuar
y 1,
199
30.
519
91 A
kt/A
ctN
ovem
bar/
Nov
embe
r 24,
200
2D
ansk
a/D
enm
ark4
Okt
obar
/Oct
ober
6, 1
968
1.5
1991
Akt
/Act
April
/Apr
il 24
, 199
8D
omin
ikan
ska
Repu
blik
a/
Dom
inic
an R
epub
licJu
n/Ju
ne 1
6, 2
007
0.2
1991
Akt
/Act
Jun/
June
16,
200
7Ek
vado
r/Ec
uado
rAv
gust
/Aug
ust 8
, 199
70.
219
78 A
kt/A
ctAv
gust
/Aug
ust 8
, 199
7Es
toni
ja/E
ston
iaSe
ptem
bar/
Sept
embe
r 24,
200
00.
219
91 A
kt/A
ctSe
ptem
bar/
Sept
embe
r 24,
200
0Ev
rops
ka u
nija
/Eur
opea
n U
nion
Juli/
July
29,
200
55.
019
91 A
kt/A
ctJu
li/Ju
ly 2
9, 2
005
Fins
ka/F
inla
ndAp
ril/A
pril
16, 1
993
1.0
1991
Akt
/Act
Juli/
July
20,
200
1Fr
ancu
ska/
Fran
ce5
Okt
obar
/Oct
ober
3, 1
971
5.0
1978
Akt
/Act
Mar
t/Mar
ch 1
7, 1
983
Gru
zija
/Geo
rgia
Nov
emba
r/N
ovem
ber 2
9, 2
008
0.2
1991
Akt
/Act
Nov
emba
r/N
ovem
ber 2
9, 2
008
Nam
ačka
/Ger
man
yAv
gust
/Aug
ust 1
0, 1
968
5.0
1991
Akt
/Act
Juli/
July
25,
199
8M
ađar
ska/
Hun
gary
April
/Apr
il 16
, 198
30.
519
91 A
kt/A
ctJa
nuar
/ Jan
uary
1, 2
003
Isla
nd/Ic
elan
dM
aj/M
ay 3
, 200
60.
219
91 A
kt/A
ctM
aj/M
ay 3
, 200
6Ir
ska/
Irel
and
Nov
emba
r/N
ovem
ber 8
, 198
11.
019
78 A
kt/A
ctN
ovem
bar/
Nov
embe
r 8, 1
981
Izra
el/Is
rael
Dec
emba
r/D
ecem
ber 1
2, 1
979
0.5
1991
Akt
/Act
April
/Apr
il 24
, 199
8Ita
lija/
Italy
Juli/
July
1, 1
977
2.0
1978
Akt
/Act
Maj
/May
28,
198
6Ja
pan/
Japa
nSe
ptem
bar/
Sept
embe
r 3, 1
982
5.0
1991
Akt
/Act
Dec
emba
r/D
ecem
ber 2
4, 1
998
Jord
an/Jo
rdan
Okt
obar
/Oct
ober
24,
200
40.
219
91 A
kt/A
ctO
ktob
ar/O
ctob
er 2
4, 2
004
Keni
ja/K
enya
Maj
/May
13,
199
90.
219
78 A
kt/A
ctM
aj/M
ay 1
3, 1
999
Kirg
istan
/Kyr
gyzs
tan
Juni
/June
26,
200
00.
219
91 A
kt/A
ctJu
n/Ju
ne 2
6, 2
000
Leto
nija
/Lat
via
Avgu
st/A
ugus
t 30,
200
20.
219
91 A
kt/A
ctAv
gust
/Aug
ust 3
0, 2
002
Litv
anija
/Lith
uani
aD
ecem
bar/
Dec
embe
r 10,
200
30.
219
91 A
kt/A
ctD
ecem
bar/
Dec
embe
r 10,
200
3M
eksik
o/M
exic
oAv
gust
/Aug
ust 9
, 199
70.
7519
78 A
kt/A
ctAv
gust
/Aug
ust 9
, 199
7M
arok
o/M
oroc
coO
ktob
ar/O
ctob
er 8
, 200
60.
219
91 A
kt/A
ctO
ktob
ar/O
ctob
er 8
, 200
6

82
Semenarstvo
Hol
andi
ja/N
ethe
rland
sAv
gust
/Aug
ust 1
0, 1
968
3.0
1991
Akt
/Act
6Ap
ril/A
pril
24, 1
998
Nov
i Zel
and/
New
Zea
land
Nov
emba
r/N
ovem
ber 8
, 198
11.
019
78 A
kt/A
ctN
ovem
bar/
Nov
embe
r 8, 1
981
Nik
arag
va/N
icar
agua
Sept
emba
r/Se
ptem
ber 6
, 200
10.
219
78 A
kt/A
ctSe
ptem
bar/
Sept
embe
r 6, 2
001
Nor
vešk
a/N
orw
aySe
ptem
bar/
Sept
embe
r 13,
199
31.
019
78 A
kt/A
ctSe
ptem
bar/
Sept
embe
r 13,
199
3O
man
/Om
anN
ovem
bar/
Nov
embe
r 22,
200
91.
019
91 A
kt/A
ctN
ovem
bar/
Nov
embe
r 22,
200
9Pa
ragv
aj/P
arag
uay
Febr
uar/
Febr
uary
8, 1
997
0.2
1978
Akt
/Act
Febr
uar/
Febr
uary
8, 1
997
Poljs
ka/P
olan
dN
ovem
bar/
Nov
embe
r 11,
198
90.
519
91 A
kt/A
ctAv
gust
/Aug
ust 1
5, 2
003
Port
ugal
/Por
tuga
lO
ktob
ar/O
ctob
er 1
4, 1
995
0.2
1978
Akt
/Act
Okt
obar
/Oct
ober
14,
199
5Ko
reja
/Rep
ublic
of K
orea
Janu
ar/Ja
nuar
y 7,
200
20.
7519
91 A
kt/A
ctJa
nuar
/Janu
ary
7, 2
002
Mol
davi
ja/R
epub
lic o
f Mol
dova
Okt
obar
/Oct
ober
28,
199
80.
219
91 A
kt/A
ctO
ktob
ar/O
ctob
er 2
8, 1
998
Runu
nija
/Rom
ania
Mar
t/Mar
ch 1
6, 2
001
0.2
1991
Akt
/Act
Mar
t/Mar
ch 1
6, 2
001
Rusij
a/Ru
ssia
n Fe
dera
tion
April
/Apr
il 24
, 199
80.
519
91 A
kt/A
ctAp
ril/A
pril
24, 1
998
Sing
apur
/Sin
gapo
reJu
li/Ju
ly 3
0, 2
004
0.2
1991
Akt
/Act
Juli/
July
30,
200
4Re
publ
ika
Slov
ačka
/Slo
vaki
aJa
nuar
/Janu
ary
1, 1
993
0.5
1991
Akt
/Act
Juni
/June
12,
200
9Sl
oven
ija/S
love
nia
Juli/
July
29,
199
90.
219
91 A
kt/A
ctJu
li/Ju
ly 2
9, 1
999
Južn
a A
frik
a/So
uth
Afr
ica
Nov
emba
r/N
ovem
ber 6
, 197
71.
019
78 A
kt/A
ctN
ovem
bar/
Nov
embe
r 8, 1
981
Špan
ija/S
pain
Maj
/May
18,
198
02.
019
91 A
kt/A
ctJu
li/Ju
ly 1
8, 2
007
Šved
ska/
Swed
enD
ecem
bar/
Dec
embe
r 17,
197
11.
519
91 A
kt/A
ctAp
ril 2
4, 1
998
Švaj
cars
ka/S
witz
erla
ndJu
li/Ju
ly 1
0, 1
977
1.5
1991
Akt
/Act
Sept
emba
r/Se
ptem
ber 1
, 200
8Tr
inid
ad i
Toba
goTr
inid
ad a
nd T
obag
oJa
nuar
/Janu
ary
30, 1
998
0.2
1978
Akt
/Act
Janu
ar/Ja
nuar
y 30
, 199
8
Tuni
s/Tu
nisia
Avgu
st/A
ugus
t 31,
200
30.
219
91 A
kt/A
ctAv
gust
/Aug
ust 3
1, 2
003
Turs
ka/T
urke
yN
ovem
bar/
Nov
embe
r 18,
200
70.
519
91 A
kt/A
ctN
ovem
bar/
Nov
embe
r 18,
200
7

83
Opšte semenarstvo
1 “196
1/19
72 A
kt” o
znač
ava
međ
unar
odnu
kon
venc
iju za
zašt
itu n
ovih
bilj
nih
sort
i od
2. d
ecem
bra
1961
, sa
doda
tnim
am
andm
anim
a za
kona
od
10
nove
mbr
a 19
72; “
1978
zako
n” o
znač
ava
Akt
od
23 o
ktob
ra 1
978.
kon
venc
ije; “
1991
zako
n” o
znač
ava
Akt
od
19 m
atra
199
1, k
onve
ncije
. / “1
961/
1972
A
ct” m
eans
the
Inte
rnat
iona
l Con
vent
ion
for t
he P
rote
ctio
n of
New
Var
ietie
s of P
lant
s of D
ecem
ber 2
, 196
1, a
s am
ende
d by
the
Add
ition
al A
ct o
f N
ovem
ber 1
0, 1
972;
“197
8 A
ct” m
eans
the
Act
of O
ctob
er 2
3, 1
978,
of t
he C
onve
ntio
n; “1
991
Act
” mea
ns th
e A
ct o
f Mar
ch 1
9, 1
991,
of t
he C
onve
n-tio
n.2 sa
not
ifika
cijo
m iz
član
a 34
(2) 1
978
zako
na /
With
a n
otifi
catio
n un
der A
rtic
le 3
4(2)
of t
he 1
978
Act
3 sa d
ekla
raci
jom
da
Akt
iz 1
978
nije
prim
enlji
v u
pose
bnim
adm
inist
rativ
nim
regi
onim
a H
ong
Kong
a /
With
a d
ecla
ratio
n th
at th
e 19
78 A
ct is
not
appl
icab
le to
the
Hon
g Ko
ng S
peci
al A
dmin
istra
tive
Regi
on4 sa
dek
lara
cijo
m d
a ko
nven
cija
iz 1
961,
uz d
odat
ak za
kona
iz 1
972,
Akt
197
8 i 1
991
zako
n ni
su p
rimen
ljivi
u G
renl
andu
i Fa
rski
m O
strv
ima
/ W
ith a
dec
lara
tion
that
the
Con
vent
ion
of 1
961,
the
Add
ition
al A
ct o
f 197
2, th
e 19
78 A
ct a
nd th
e 19
91 A
ct a
re n
ot ap
plic
able
to G
reen
land
and
the
Faro
e Is
land
s5 sa
dek
lara
cijo
m iz
197
8. p
rimen
ljiva
na
terit
oriji
Fra
ncus
ke u
klju
čuju
ći p
reko
mor
ske
terit
orije
/ W
ith a
dec
lara
tion
that
the
1978
Act
appl
ies t
o th
e te
rrito
ry o
f the
Fre
nch
Repu
blic
, inc
ludi
ng th
e O
vers
eas D
epar
tmen
ts a
nd T
errit
orie
s6 ra
tifika
cija
za K
ralje
vstv
o u
Evro
pi /
Ratifi
catio
n fo
r the
Kin
gdom
in E
urop
e7 sa
reze
rvac
iojo
m k
orisn
ika
na čl
an 3
5(2)
Kon
venc
ije iz
199
1 / W
ith a
rese
rvat
ion
purs
uant
to A
rtic
le 3
5(2)
of t
he 1
991
Act
.
Ukr
ajin
a/U
krai
neN
ovem
bar/
Nov
embe
r 3, 1
995
0.2
1991
Akt
/Act
Janu
ar/Ja
nuar
y 19
, 200
7U
jedi
njen
o K
arlje
vstv
oU
nite
d K
ingd
omAv
gust
/Aug
ust 1
0, 1
968
2.0
1991
Akt
/Act
Janu
ar/Ja
nuar
y 3,
199
9
SAD
/USA
Nov
emba
r/N
ovem
ber 8
, 198
15.
019
91 A
kt/A
ct7
Febr
uarF
ebru
ary
22, 1
999
Uru
gvaj
/Uru
guay
Nov
emba
r/N
ovem
ber 1
3, 1
994
0.2
1978
Akt
/Act
Nov
emba
r/N
ovem
ber 1
3, 1
994
Uzb
erki
stan
/Uzb
ekist
anN
ovem
bar/
Nov
embe
r 14,
200
40.
219
91 A
kt/A
ctN
ovem
bar/
Nov
embe
r 14,
200
4SR
. Vije
tnam
/Vie
t Nam
Dec
emba
r/D
ecem
ber 2
4, 2
006
0.2
1991
Akt
/ Act
Dec
emba
r/D
ecem
ber 2
4, 2
006

84
Semenarstvo
Postajući član UPOV-a, država pokazuje nameru da zaštiti ople-menjivače i njihov rad na osnovu međunarodno priznatih principa. Ono pruža mogućnost oplemenjivačima da ostvare zaštitu sorti i u drugim državama članicama, a podstiče i strane oplemenjivače da ula-žu u oplemenjivačke programe i proizvodnju semena na sopstvenoj teritoriji.
Srbija bi trebalo da postane član UPOV organizacije u oktobru 2011. godine. Zakon o zaštiti prava oplemenjivača biljnih sorti je pri-hvaćen konsenzusom na redovnom aprilskom zasedanju UPOV-a 2011. godine. Isti zakon je usvojen u Skupštini Republike Srbije u maju 2009. godine. Zahtev Srbije za pristupanje Konvenciji iz 1991. godine je podnet.
Da bi sorte mogle biti zaštićene, moraju da su:
– različite od postojećih, opšte poznatih sorti,– dovoljno homogene, ujednačene,– stabilne u svojim osobinama,– nove u smislu da nisu bile komercijalizovane pre datuma utvrđe-
nog na prijavi za zaštitu, odnosno godinu dana od prve komerci-jalizacije u zemlji i četiri godine u inostranstvu.
Kao i kod svih prava na intelektualnu svojinu, prava oplemenjiva-ča se daju za ograničen period vremena na kraju kojeg njima zaštićene sorte prelaze u domen javnosti (public line). Prava su predmet kontrole, u interesu javnosti, protiv bilo kakve mogućnosti zloupotrebe. Važno je napomenuti da se autorizacija imaoca prava zaštite ne traži radi kori-šćenja njegove sorte u svrhu istraživanja, uključujući njenu upotrebu u oplemenjivanju novih sorti.
Stvaranje novih sorti zahteva značajna ulaganja u smislu kreativno-sti, materijalnih sredstava, vremena. Mogućnost da se dobiju izvesna ek-skluzivna prava koja se odnose na njegovu novu sortu pruža uspešnom oplemenjivaču veću šansu da pokrije uložene troškove i akumulira sred-stva neophodna za dalje investiranje. Ukoliko oplemenjivač nije zaštitio sortu ne postoji ništa što bi sprečilo druge da manipulišu selekcionim materijalom u smislu njegove komercijalizacije, čime se rad oplemenji-vača pokazao nesvrsishodnim. Naplata licence prema podacima EU za 2000/2001. godine, a koje se ni do danas nisu značajnije menjale, napla-ćuje se kao što je naznačeno u tabeli 14.
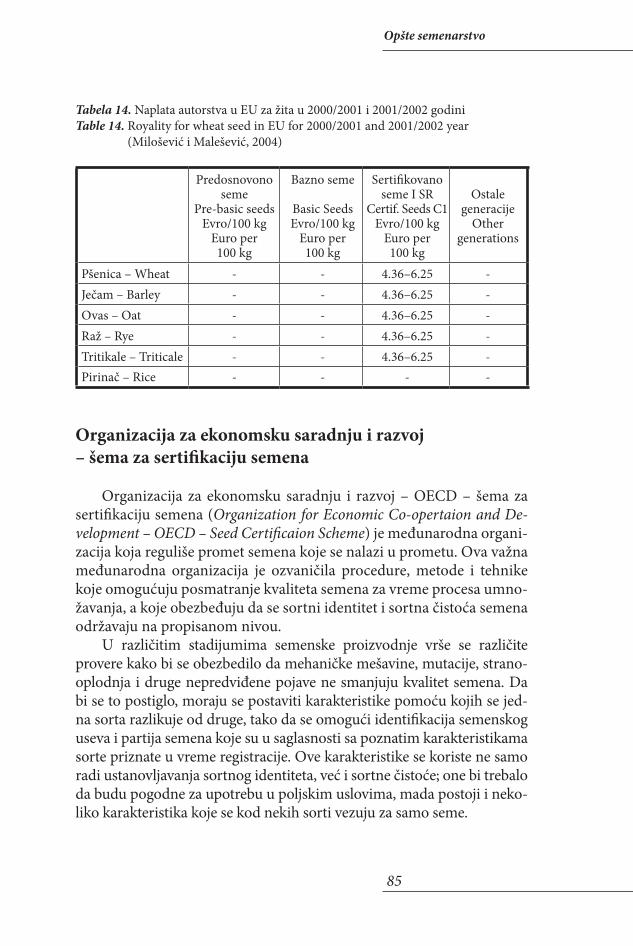
85
Opšte semenarstvo
Tabela 14. Naplata autorstva u EU za žita u 2000/2001 i 2001/2002 godiniTable 14. Royality for wheat seed in EU for 2000/2001 and 2001/2002 year
(Milošević i Malešević, 2004)
Predosnovono seme
Pre-basic seeds Evro/100 kg
Euro per 100 kg
Bazno seme
Basic Seeds Evro/100 kg
Euro per 100 kg
Sertifikovano seme I SR
Certif. Seeds C1Evro/100 kg
Euro per 100 kg
Ostale generacije
Other generations
Pšenica – Wheat - - 4.36–6.25 -Ječam – Barley - - 4.36–6.25 -Ovas – Oat - - 4.36–6.25 -Raž – Rye - - 4.36–6.25 -Tritikale – Triticale - - 4.36–6.25 -Pirinač – Rice - - - -
Organizacija za ekonomsku saradnju i razvoj– šema za sertifikaciju semena
Organizacija za ekonomsku saradnju i razvoj – OECD – šema za sertifikaciju semena (Organization for Economic Co-opertaion and De-velopment – OECD – Seed Certificaion Scheme) je međunarodna organi-zacija koja reguliše promet semena koje se nalazi u prometu. Ova važna međunarodna organizacija je ozvaničila procedure, metode i tehnike koje omogućuju posmatranje kvaliteta semena za vreme procesa umno-žavanja, a koje obezbeđuju da se sortni identitet i sortna čistoća semena održavaju na propisanom nivou.
U različitim stadijumima semenske proizvodnje vrše se različite provere kako bi se obezbedilo da mehaničke mešavine, mutacije, strano-oplodnja i druge nepredviđene pojave ne smanjuju kvalitet semena. Da bi se to postiglo, moraju se postaviti karakteristike pomoću kojih se jed-na sorta razlikuje od druge, tako da se omogući identifikacija semenskog useva i partija semena koje su u saglasnosti sa poznatim karakteristikama sorte priznate u vreme registracije. Ove karakteristike se koriste ne samo radi ustanovljavanja sortnog identiteta, već i sortne čistoće; one bi trebalo da budu pogodne za upotrebu u poljskim uslovima, mada postoji i neko-liko karakteristika koje se kod nekih sorti vezuju za samo seme.

86
Semenarstvo
OECD pravila se odnose na sedam šema. Svaka od njih ima spe-cifična pravila jer se radi o biljnim vrstama koje pripadaju različitim porodicama, koje imaju svoje biološke i proizvodne specifičnosti. To su:
– trave i leguminoze, – krstašice, ostale uljane biljke i biljke za vlakno, – žita,– stočna i šećerna repa, – subteraneanska detelina i slične biljne vrste, – kukuruz i sirak, – povrće. Kao primer može poslužiti deo OECD šeme za sertifikaciju semena
žita. Kada sorta treba da se nađe u međunarodnom prometu od strane OECD-a prihvaćene su sledeće kategorije semena:
– predosnovno,– sertifikovano seme (Council Directve (2000) 146/FINAL, 2009)
Učestalost post-kontrolnih testova kod sertifikovanog semena iz prethodne godine
Kod ustanovljenih nepravilnosti < 0,5% proverava se 5% svih partija semena koje su bile na tržištu. Kod ustanovljenih nepravilnosti većih od 0,5% proverava se 3,0% i 10% svih partija semena koje su bile na tržištu.
Poštovanje jednog od osnovnih parametara koje uvažava OECD, genetičke čistoće prikazano je u tabeli 15. Parametri su uslov za to da se seme nađe u međunarodnom prometu.
Tabela 15. Minimalan procenat genetičke čistoće kod žitaTable 15. Minimum of genetic purity for small grains (OECD, 2007)
Vrste/SpeciesOsnovno
seme/ Basic Seed
Sertifikovano seme – prva generacija/
Certified Seed - first generation
Sertifikovano seme – druga generacija/
Certified seed -second generation
Triticum aestivum, Hordeum vulgare, Avena spp. i Oryza sativa
99.9% 99.7% 99.0%
Većinom samooplodnie sorte x Triticosecale/ Mainly self-pollinating varieties of X Triticosecale
99.7% 99.0% 98.0%

87
Opšte semenarstvo
Slika 31. Izgled OECD sertifikataFigure 31. OECD certificat

88
Semenarstvo
Izgled OECD sertifikata koji izdaje Laboratorija za ispitivanje seme-na Instituta za ratarstvo i povrtarstvo prikazan je slici 31 kao deo kvali-tata koji se ustanovljava kod semenskih useva. Za žita je on naznačen u tabeli 16.
Tabela 16. Dozvoljeni broj biljaka drugih biljnih vrsta u usevuTable16. Permited number of other plants in crop (OECD, 2008)
Vrste/Species Osnovno seme/Basic seed
Sertifikovano seme/Certified seed
Stranooplodne sorte Secale cereale × Triticosecale/ Cross-pollinating varieties of Secale cereale and × Triticosecale
1 u 30 m2
1 in 30 sq. m1 u 10 m2
1 in 10 sq. m
Osnov za proizvodnju semena čine OECD sortne liste. Ukoliko neka sorta nije na sortnoj listi OECD-a, ne može da se uključi u među-narodni promet. Svake godine, zemlja članica OECD šeme za sertifika-ciju semena dužna je da inovira OECD sortnu listu, ukoliko želi da se neka nova sorta nađe u međunarodnom prometu.
Međunarodna organizacija za ispitivanje kvaliteta semena
Međunarodna organizacija za ispitivanje kvaliteta semena (Interna-tional Seed Testing Asociation – ISTA) osnovana je 1921. godine sa sedi-štem u Cirihu, Švajcarska. ISTA je povezana u radu sa napred pomenu-tim organizacijama. Po rečima jednog od predsednika ISTA-e Norberta Leista „ISTA je samo faza u globalizaciji svetskog tržišta, ona nastavlja sa tradicionalnom odgovornošću i efikasnom ulogom u tehničkoj or-ganizaciji uspešno već 90 godina. ISTA je kvalifikovana i spremna da se suoči sa novim razvojnim izazovima, jer je to asocijacija vladinih, privatnih i kompanijskih laboratorija”.
Osnovni cilj ISTA-e je ujednačavanje metoda rada, kako bi se me-đunarodni promet semena nesmetano odvijao. Standardi za akreditaciju laboratorija moraju biti ispunjeni da bi jedna laboratorija bila ovlašće-na za izdavanje ISTA sertifikata. Prethodno laboratorija mora da pro-đe, sa zadovoljavajućim rezultatom, pred-akreditacione programe koji dokazuju znanje iz jedne oblasti, npr. čistoće semena, klijavosti semena

89
Opšte semenarstvo
Slika 32a. Primer statističke obrade podataka ISTA referee testa − rezultati ispitivanja čistoće semena pšenice
Figure 32a. Example of statistic data processing of ISTA referee test − resultes of purity test of wheat seed

90
Semenarstvo
Slika 32b. Primer statističke obrade podataka ISTA referee testa − rezultati ispitivanja klijavosti semena pšenice
Figure 32b. Example of statistic data processing of ISTA referee test − resultes of germination test of wheat seed

91
Opšte semenarstvo
(proficiency tests), a koji moraju demonstrirati tokom obilaska komisije za akreditaciju. Svake treće godine ISTA vrši reakreditaciju laboratori-ja, da bi se uverila u održavanje nivoa kompetentnosti, usaglašenosti sa njihovim zahtevima, koji su nastupili u međuvremenu. Akreditacija se vrši po sistemu ISO 9001, EN 45000 sistemu 17025. ISTA akreditaciju potvrđuje predsednik ISTA-e posle odluke Izvršnog komiteta.
Pored toga ISTA organizuje kružne analize, upoređuje rezultate svake laboratorije sa stvarnim (kontrolnim) i o tome izveštava svaku laboratoriju, kako bi pomenute laboratorije mogle da izvrše korektivne mere (sl. 32a, 32b).
ISTA ima sistem u koji ulaze:– međunarodna pravila za ispitivanje semena,– standardi za akreditaciju laboratorija,– program kružnih analiza (Ring test),– ISTA međunarodni sertifikati (sl. 33).
Pomenuti sistem obezbeđuje ujednačenost u ispitivanju kvaliteta semena i pouzdanost prikazanih rezultata.
U Međunarodnim pravilima za ispitivanje semena date su metode ispitivanja koja su sve laboratorije, koje imaju ISTA akreditaciju, dužne da primenjuju. U njima su po odeljcima naznačene metode za:
– uzorkovanje semena,– čistoću semena,– broj primesa drugih biljnih vrsta u osnovnoj masi semena,– klijavost semena,– tetrazolijum test,– zdravstveno stanje semena,– verifikaciju vrsta i sorti,– određivanje sardžaja vlage u semenu,– određivanje mase 1000 semena,– testiranje obloženog semena,– ispitivanje vitalnosti embriona semena,– testiranje semena pomoću merenja proba (za šumsko seme),– test pomoću X zraka,– tolerancije,– sertifikati (ISTA Rules, 2011).

92
Semenarstvo
Slika 33. Međunarodni ISTA oranž sertifikatFigure 33. International ISTA orange certification

93
Opšte semenarstvo
Međunarodna organizacija za ispitivanje semena ima za viziju da bude međunarodna asocijacija čiji se rad zasniva na naučnim dostignu-ćima iz oblasti semenarstva i dorade semena, razvijajući i usavršavajući metode uzorkovanja i ispitivanja kvaliteta semena, akreditacije labora-torija i osiguranjem sertifikata kao organizacija van politike, neprofita-bilna, legalna i nezavisna kako od vlade tako i od kompanija.
ISTA je za našu zemlju od izuzetne važnosti jer se celokupan među-narodni promet semena ispituje po njenim metodama i na taj način se ostvaruje značajan dohodak kod izvoza semena. Veoma tesno sarađuje sa Međunarodnom organizacijom za očuvanje biodiverziteta (Biodiver-sity International – BI) dajući joj informacije o naučnim dostignućima iz oblasti očuvanja genofonda, jer se osnove rada ISTA-e i BI-a podudara-ju. Obe rade na ispitivanju kvaliteta semena, istina sa različitih osnova, kao i načina njegovog održavanja, odnosno čuvanja.
Međunarodna organizacija za standardizaciju
Međunarodna organizacija za standardizaciju (International Standar-dization Organization - ISO) je nevladina organizacija osnovana 1947. go-dine sa ciljem razvijanja svetskih standarda, unapređenja međunarodne komunikacije i saradnje, kao i promovisanja pravične raspodele trgovine. Njen cilj je zaštita potrošača u pogledu kvaliteta preko poboljšanja sistema upravljanja. Svet je otišao daleko napred u tom segmentu. Preko šezdeset zemalja sveta je uključeno u proces standardizacije.
Veza između međunarodnih organizacija ISO-ISTA je neophodna jer ukoliko ISTA ne bi prihvatila ISO standardizaciju, izgubila bi akreditaciju. To dalje znači da zemlje članice ISTA moraju da prihvate ISO standardi-zaciju, zbog akreditacije laboratorija ISTA u okviru svojih institucija.
Ova organizacija je još jedna važna karika za koju se vezuje ISTA. ISO standarde koriste izvoznici, uvoznici u svim segmentima proizvod-nje i ispitivanja kvaliteta. Ovaj prepoznatljivi međunarodni standard je prihvaćen i kao deo nacionalnih standarda u mnogim zemljama. Bez obzira na to što je u zaostatku procesa standardizacije, naša zemlja se preko Akreditacionog tela Srbije (ATS) uključila u međunarodne toko-ve standardizacije i sačinila skup specifičnih uputstava za izbor i kori-šćenje odgovarajućeg sistema kvaliteta.

94
Semenarstvo
Evropska unija
Evropska unija (European Union – EU) je ekonomska i politička uni-ja 27 zemalja članica koje su locirane u Evropi. Zalaže se za regionalne integracije. Formirana je 1993. godine Mastritskim sporazumom, pod pokroviteljstvom Evropske ekonomske zajednice, prethodnice Evropske unije. U EU živi oko 500 miliona stanovnika, ima oko 30% učešća svet-skog nominalnog bruto proizvoda (US $ 18,4 milijarde u 2008).
EU je razvila jedinstveno tržište putem standardizovanog sistema zakona koji se primenjuju u svim državama članicama, čime se obez-beđuje slobodno kretanje ljudi, robe, usluga i kapitala. To omogućava održavanje zajedničke politike o trgovini, poljoprivredi, ribarstvu i regio-nalnom razvoju. Šesnaest država članica su prihvatile zajedničku valutu, evro i ona predstavlja evropsku monetarnu uniju (European Monetary Union – EMU). EU je razvila ograničenu ulogu u spoljnoj politici, pošto je član Svetske trgovinske organizacije STO i G20 zemalja u UN.
Kao međunarodna organizacija, Evropska unija funkcioniše kroz hibridni sistem supranacionalizama i međudržvnosti. U pojedinim po-dručjima, odluke se donose kroz pregovore između država članica, dok su u drugima, odgovorne nezavisne nadnacionalne institucije. Važne institucije Evropske unije su Evropska komisija, Savet Evropske unije, Evropski savet, Sud pravde Evropske unije i Evropska centralna banka. Evropski parlament biraju građani država članica na svakih pet godina., (http://europa.eu/index_en.htm).
Razlog za članstvo Srbije u EU
Srbija bi članstvom u Evropskoj uniji obezbedila pripadnost zajednici država koje svojim potencijalom proizvodnje i kapitalom koji poseduju, mogu da vrše usmerenje ka podizanju konkurentnosti proizvodnje poljo-privrednih proizvoda svojih članica. To bi bio istorijski korak za Srbiju jer bi došlo do promene načina rada i poslovanja na svim nivoima. U oblasti poljoprivrede formirale bi se neophodne institucije za saradnju sa EU.
Prvi važan korak koji je Srbija preduzela u saradnji sa EU je zaklju-čivanje Sporazuma o stabilizaciji i pridruživanju (SSP), jer je prvi put uspostavljen ugovorni odnos između Republike Srbije i Europske unije. Sporazum predstavlja pravni osnov i sadržajni okvir odnosa RS i EU.

95
Opšte semenarstvo
On je stoga prvi korak ka institucionalizovanju odnosa sa Evropskom unijom i omogućava postupne pripreme Republike Srbije za ostvarenje punopravnog članstva u Uniji.
Šta podrazumevaju EU integracije?
Postupak integracije Srbije u Evropsku uniju može se uslovno po-deliti u tri dela, koja podrazumevaju različite aktivnosti:
I deo – liberalizacija tržišta, uklanjanje trgovinskih barijera,II deo – institucionalno prestrukturiranje,III deo – usklađivanje zakonske regulative.Pod liberalizacijom tržišta podrazumeva se proces smanjenja ca-
rina, čime se otvara put za uvoz jeftinijih proizvoda, pa i semena na naše tržište. Samim tim pojačava se konkurencija semenarstvu u Srbi-ji, mada su i danas skoro sve multinacionalne kompanije prisutne na srpskom tržištu. Razlika će biti u ceni semena jer se procesom libera-lizacije predviđa postepeno sniženje zaštitnih carina koje su postojale do potpisivanja Sporazuma.
Kada je u pitanju seme u tabelama 17, 18, 19, 20 i 21 navedeno je postepeno snižavanje carina po godinama za seme pojedinih biljnih vrsta.
Tabela 17. Postepeno snižavanje carinskih stopa za seme kukuruza, suncokreta šećer-ne repe
Table 17. Gradual lowering of tariff rates for seed corn, sunflower, sugar beet (Milošević, 2010)
Vrsta semena/Seed MFN 2009 2010 2011 2012 2013-2014Semenski kukuruz – hibridniHybrid corn seed 30 24 21 15 12 9
Semenski kukuruz – ostaliCorn seed -other 30 24 21 15 12 9
Seme suncokretaSunflower seed 20 16 16 14 12 10
Seme šečerne repeSugar beet seed 10 8 6 4 2 0
MNF tarifa- nivo tarife koje članice GATT/WTO troše na robu drugih članicaMNF tariff - The tariff level that a member of the GATT/WTO charges on a good to other members.

96
Semenarstvo
Tabele 18. Dinamika sniženja carina po godinama (%) Table 18. Dynamics decrease tariffs by the year (%) (Milošević, 2010)
Vrsta semena/Seed MFN 2009 2013- 2014
Seme soje/Soybean seed 5 0
Slačica/Mustard 5 0
Krompir/Potato 5 0
Mak/Popy 10 0
Konoplja/Hemp 10 0
Tabele 19. Dinamika sniženja carina po godinama (%)Table 19.Dynamics decrease tariffs by the year (%) (Milošević, 2010)
Vrsta semena/Seed MFN 2009 2010 2011 2012 2013-2014
Raž/Ray 20 0
Tvrda pšenicaDurum wheat 30 0
Obična pšenica/Wheat 30 0
Ječam, ovas/Barley, oat 20 0
Tritikale/Titicale 10 0
Tabele 20.Dinamika sniženja carina po godinama (%)Table 20. Dynamics decrease tariffs by the year (%) (Milošević, 2010)
Vrsta semena/Seed MFN 2009 2010 2011 2012 2013-2014
Lucerka/Alfalfa 10 80 60 40 20 0Crvena detelinaRed clover 5 0
Detelina/Clover 5 0
Seme trava/Grass seed 5 0
Seme krmnogbilja – ostaloForage crops – others
5 0

97
Opšte semenarstvo
Zajednička poljoprivredna politika – ZPP, (Common agricultural policy – CAP) je skup zakona i prakse koje je Evropska unija usvojila radi postizanja zajedničke, unificirane politike u sektoru poljoprivrede – ZPP je najobuhvatnija od svih ekonomskih politika EU.
Institucionalno prestrukturiranje podrazumeva fomiranje institu-cija koje će pratiti process evropskih integracija, kao što je Uprava za javna plaćanja pri Ministarstvu poljoprivrede, trgovine, šumarstva i vo-doprivrede Republike Srbije, preko koje se usmeravaju sva sredstva iz pretpristrupnih fondova EU. Uprava za javna plaćanja mora biti akre-ditovana od strane komisije EU, kako bi se uskladio način platnog pro-meta i korišćenja sredstava pretpristupnih fondova IPA, što je urađe-no tokom 2010. godine. Direkcija za bezbednost hrane je neophodna u smislu zaštite pava potrošača za korišćenje hrane visokog kvaliteta. Sistem javnih skladišta omogućava berzansko poslovanje poljoprivred-nim proizvodima. Na taj način, vlasnik može da proda svoju robu u momentu i po ceni koja mu najviše odgovara. Seme nije predmet ber-zanskog poslovanja.
Usaglašavanje zakonodavnog okvira sa zakonodavstvom EU je ve-oma obiman i mukotrpan posao. Srbija treba da uskladi svoje zakono-davstvo iz oblasti poljopirvrede sa 8.600 direktiva EU. Usvajanjem 15 zakona i 53 podzakonska akta iz oblasti poljoprivrede u 2009. godini počeo je taj proces.
Proces prilagođavanja sistemu EU
Istorijiski gledano, zbog postupka prilagođavanja EU i njenom si-stemu funkcionisanja poljoprivrede prvo su uvedeni posebni programi kao što je bio Phare (Programme of Community aid to the countries of Central and Eastern Europe – Phare) za jačanje institucija, ulaganja u regulatornu infrastrukturu, ulaganja u poljoprivrednu i socijalnu kohe-ziju, zatim SAPARD, poseban program za poljoprivredu i ruralni razvoj zemalja kandidata za članstvo u EU (Special Accession Programme for Agriculture and Rural Development – Sapard). Od 2007. su pomenuti programi zamenjeni novim, jedinstvenim pretpristupnim programom IPA (Instrument for Pre-Accession), a poseban program predstavlja IPARD (Instrument for Pre-Accession for Rural Development) koji se od-nosi na razvoj sela.

98
Semenarstvo
Cilj ovih programa nije samo konkretno pružanje pomoći, nego pre svega „učenje“ pravila i uvođenje novih propisa kako se ne bi dogodi-lo da nova država članica, nakon pristupa Europskoj uniji, ne uspe da iskoristi sredstva koja su za nju predviđena zbog loše pripreme vlastitih struktura (Milošević, 2010).
Sektori koje pokriva Zajednička poljoprivredna politika (ZPP)
ZPP vrši intervencije u cenovnoj podršci, subvencijama, samo za neke sektore i to za:
– žita, pirinač, krompir,– šećer,– grašak, boraniju,– hmelj,– duvan,– konoplju,– sve vrste semena,– (meso, voće i povrće).
Seme se proizvodi u svim zemljama članicama EU, a najveći proi-zvođači su: Holandija, Danska, Francuska, Nemačka, Španija i Italija. Površina pod semenskom proizvodnjom u svih 25 EU država članica je oko 450 000 ha sa Bugarskom i Rumunijom. Proizvodnja sertifiko-vanog semena u 2004. i 2005. je bila 480.000 t, uključujući pirinač sa 70.000 t, lan i konoplja 20.000 t i krmno bilje koje je bilo gajeno na oko 380.000 ha.
Seme je podeljeno u četiri grupe biljnih vrsta:– žita (Triticum spelta i Oriza sativa ),– uljane biljne vrste (lan, konoplja),– seme trava,– leguminoze (grašak, detelina, pasulj).

99
Opšte semenarstvo
Reforme sektora semenarstva
Sektor semenarstva je uključen u ZPP kao set regulativa pod brojem (EC) No. 1782/2003 (Counncil Regulation (EC) No 1782/2003 ) koja uključuje specifične potrebe sektora. Promene koje je uvela EU u sek-toru semenarstva regulisane su i regulativom (EC) broj 1947/2005 od 23. novembra 2005. godine (Counncil Regulation (EC) No. 1947/2005) Evropske komisije koja uključuje organizaciju tržišta semenom. Pored toga regulativa uključuje trgovinu semenom sa trećim zemljama, klau-zule remećenja tržišta i opšte odredbe vezane za pomoć pojedinih drža-va, uključujući i posebne završne klauzule.
U EU, kao organizaciona jedinica odgovorna za pitanje semenar-stva, postoji Komitet za marketing semena i komunikaciju u razmeni informacija u okviru Komisije EU.
U semenarskom sektoru uzimaju se u obzir kategorije semena (ba-zno i sertifikovano) koje su predviđene za subvencionisanje u EU, ugo-vori između kompanija registrovanih za semensku proizvodnju i proi-zvođača semena kao i uslovi za sertifikaciju i proizvodnju.
Visina podrške varira i zavisi od biljne vrste. Svaka država odlučuje gde će se primeniti ovakav sistem podrške, a to zavisi od važnosti biljne vrste za region, odnosno državu koja daje podsticaj. Kao primer može se uzeti tvrda – durum pšenica. Podrška ovoj vrsti pšenice se menjala na sledeći način:
– Smanjena subvencija sa €344,50/ha na €285/ha u toku tri godine (2002, 2003, 2004), a zatim je izmenjen režim u 2005. godini. U sledeće tri godine podrška je bila €138,90/ha. U narednom periodu podrška je značajno smanjena, na €40/ha i to samo za područja koja tradicionalno gaje tvrdu pšenicu.
Sekcija Evropske semenarske asocijacije – (ESA) za žita je preporu-čila za durum pšenicu:
– da se održi aktuelan nivo podrške uz obavezno korišćenje sertifi-kovanog semena,
– da se kod svih žita obavezno koristi sertifikovano seme (small gra-ins),
– da se osigura premija za kvalitet.

100
Semenarstvo
Kod procesa ugovaranja semenskih useva predloženo je da predu-slov bude upotreba 100% sertifikovanog semena. Dalja inicijativa ove semenarske asocijacije je bila da se zaštiti semenarski sektor kroz više-godišnje ugovaranje proizvodnje, uz obavezu daljeg umnožavanja kate-gorija semena do poslednje dozvoljene generacije umnožavanja pred-viđene OECD šemom za sertifikaciju semena, odnosno nacionalnom zakonskom regulativom.
Ako se posmatraju proteinske biljne vrste cenovna podrška se kre-tala na sledeći način:
– Podrška za grašak i pasulj koja je plaćana po prinosu, konvertova-će se na plaćanje po hektaru i iznosiće €55,75/ha, do 1,4 milliona hekta-ra, mada će i dalje subvencije ostati vezane za visinu prinosa.
Evropska semenarska asocijacija (ESA)
Ranih šezdesetih godina dvadesetog veka, 1961. godine formirana je asocijacija koja se bavila trgovinom semena (Seed Committe of the Common Market – COMESCO), 1964. godine organizacija oplemenji-vača krompira (Association of Common Market Potato Breeders – ASSO-PO-MAC), 1970. godine asocijacija proizvođača krmnog bilja (Forage Seed Production – AMUFOC) i 1977. godine Asocijacija oplemenjivaća biljaka Evropske ekonomske zajednice (Association of Plant Breeders of the European Economic Community – COMASSO). Svaka od ovih or-ganizacija, predstavljala je važan segment oplemenjivanja biljaka, proi-zvođača semena i lanca marketinga. Evropska semenarska asocijacija je formirana 1998. godine, kao krovna organizacija koja je objedinila već postojeće četiri organizacije i individualne proizvođače povrća. Uskoro se osetila potreba, s obzirom na promene političkog i administrativnog okruženja i mnoštva zajedničkih zadataka i interesa u EU, za promenu načina rada ESA. Nova ESA formirana je 2000. godine. Uključila je već postojeće članove. Spojila je sve do tada postojeće organizacije koje po-sluju sa semenom u jednu, koju prepoznaje EU i sada predstavlja evrop-sku semenarsku industriju, koja je aktivna u istraživanjima, oplemenji-vanju, proizvodnji i marketingu semena (http://www.euroseeds.org/who-we-are). Snaga ESA i jeste u tome što postoji dobra organizovanost i povezanost svih učesnika u semenskoj industriji. ESA je pored Američ-ke semenarske asocijacije (American Seed Trade Organization – ASTA)

101
Opšte semenarstvo
najjača organizacija te vrste u svetu. Njene članice uživaju pune povla-stice članstva. Ne postoji nijedna nacionalna semenarska asocijacija u EU koja nije njen član. Na taj način se dobijaju potpune informacije o inovacijama i napretku u oblasti semenarstva.
ESA je u 2010. godini dobila još jednog značajnog člana. Regional-na asocijacija EESNET (East European Seed Network) pridružila se ESA, posle 10 godina postojanja. Članovi EESNET su bile Češka, Slovačka, Srbija, Crna Gora, Makedonija, Rusija, Litvanija, Ukrajina, Poljska i Bu-garska (sl. 34). Zadatak ove organizacije je da udružuje semenare koji na svojim godišnjim sastancima razmenjuju informacije o proizvod-nji semena, novinama koje su se desile na tom polju. Susreti su važni i zbog toga što se rađaju nova poznanstva, sklapaju novi poslovi, pri-maju novi članovi. To je neprofitabilna organizacija, sa dobrovoljnim načinom učlanjenja. Ideja formiranja ove organizacije došla je iz Češke Republike. Za ostvarenje ciljeva formiranja regionalne organizacije češ-ka semenarska asocijcija CMSS je uložila mnogo truda i znanja. Preko ove organizacije EU ima kontakt sa semenarima zemalja u tranziciji, obaveštavanje o najnovijim događanjima iz specifičnih oblasti vezanih posredno ili neposredno za semenarstvo.
Slika 34. Zemlje članice EESNET-aFigure 34. EESNET member countries
EESNETUkrajina Ukraine
Makedonija Macedonia
Albanija Albania
Rusija Russia
Bugarska Bulgaria
Slovačka Sloavakia
Rumunija Romania
MađarskaHungary
Srbija i Crna GoraSerbia and Montenegro
LitvanijaLithuania
Hrvatska Croatia
Bosna i Hercegovina Bosnia and Herzegovina
Poljska Poland
Češka Republika Czech Republic

102
Semenarstvo
Semenarska asocijacija Srbije
Semenarska asocijacije Srbije (SAS) osnovana je sa ciljem udruži-vanja semenara na nivou tadašnje Jugoslavije. Politička situacija, for-miranje Srbije kao samostalne države, dovelo je i do promena u pro-storu delovanja i nazivu semenarske asocijacije. Do tada se ona zvala Jugoslovenska semenarska asocijacija – YUSEA, te je promenila ime u Semenarska asocijacija Srbije – SAS. YUSEA je osnovana 18. maja 2001. godine za vreme održavanja novosadskog poljoprivrednog sajma u No-vom Sadu, na inicijativu Instituta za ratarstvo i povrtarstvo iz Novog Sada. Na osnivačkoj Skupštini su definisani ciljevi udruženja, a oni su:
– prezentacija i zaštita poslovnih interesa njenih članova, naročito ispred državnih organa, nacionalnih i međunarodnih organizacija,
– učestvovanje u stvaranju zakonske regulative koja se odnosi na ga-jenje biljaka i proizvodnju semena, akata poreske i fiskalne politi-ke i drugih regulativa i njeno predlaganje resornom ministarstvu,
– praćenje i primena gore navedenih zakonskih akata,– analiza efekata gore pomenutih regulativa i primena zakonske ini-
cijative,– razvoj i modernizacija gajenja biljaka i proizvodnje semena,– udruživanje oplemenjivača, proizvođača i korisnika semena i se-
menskih preduzeća,– davanje podrške poslovnim projektima članova Udruženja i po-
maganje saradnje sa stranim partnerima,– zaštita prava oplemenjivača i pružanje pomoći u ostvarivanju tih
prava,– određivanje i podsticanje standardne i poslovne etike koja se od-
nosi na poslovne transakcije,– sprovođenje ostalih zajedničkih interesa članova Udruženja u
skladu sa zakonom i drugim regulativama. U okviru SAS postoje i posebne grupe, čiji je zadatak da rešavaju
specifična pitanja vezana za oblast oplemenjivanja, iniciranja i predla-ganja zakonske regulative kao i druge grupe koje se formiraju kada se za to ukaže potreba.

103
Opšte semenarstvo
SAS je formirana radi toga da organizuje i poveže sve strukture koje u svojoj delatnosti imaju semenarstvo zbog izuzetnog značaja koji se-menarstvo ima u okviru poljoprivrede i u našoj ukupnoj privredi. Na slici 35 prikazani su članovi osnivači YUSEA-e. Neki od njih više ne postoje, ali treba znati da su te organizacije bile vizionarske i predvidele su značaj udruživanja na polju semenarstva.
Slika 35. Članovi i osnivači YUSEA (SAS)Figure 35. Memebers and founders
of YUSEA (SAS)
Institut za ratarstvo i povrtarstvo, Novi Sad,Institute of field and vegetable crops, Novi Sad,Institut za kukuruz »Zemun Polje«, Zemun,Institute for maize »Zemun Polje«, Zemun,Institut za istraživanja u poljoprivredi »Srbija« Beograd,Institute for agricultural investigations »Srbija« Beograd,»Hibrid« Beograd,»Semenarstvo« Novi Sad,»Produktiva« Novi Sad,»Agrocoop« Novi Sad,»Agroseme« Sremska Mitrovica,»Seme-Pančevo«, Pančevo,»Tamiš« AD Pančevo,»Banat-Seme« Zrenjanin,»PIK-Bečej« AD Poljoprivreda,HK »Seme-Sombor« Sombor,»Agrobačka« Bačka Topola,»Agroseme-Kikinda«»Agroseme-Panonija« Subotica
Novi članovi: / New members:- VST trade, Banja Luka- BV Comerce, Novi Sad- Atel, Novi Sad- Crompton, Novi Sad

104
Semenarstvo
ZAKONSKI PROPISI U OBLASTI SEMENARSTVA
Znak jednakosti se može staviti između kvalitetnog i deklarisanog (sertifikovanog) semena. Kvalitet se obezbeđuje kroz dug proizvodni proces koji počinje zasnivanjem semenskog useva, negom useva, struč-nim pregledom useva, a završava preko ubiranja semena, njegove dora-de, ispitivanja kvaliteta i deklarisanja partije semena.
Za postizanje pravih vrednosti prethodno navedenih operacija po-trebno je primeniti mere koje preporučuju i nameću obaveze za odre-đeni način izvođenja tih operacija. To se postiže donošenjem adekvatne zakonske regulative.
Zakonski propisi u zemljama EU
Evropska unija nema jedinstven set zakona vezanih za semenarstvo. Svaka zemlja članica ima sopstveni zakon o semenu, zakon o priznavanju sorti, zakon o zaštiti novih biljnih sorti i dr.. Ono što im je zajedničko je da su u osnove svih tih zakona ugrađena pravila relevantnih međunarodnih organizacija. Pošto govorimo o semenarstvu onda su to regulative, pravi-la odnosno standardi koje propisuje OECD šema za sertifikaciju semena kada je poljski pregled semenskih useva u pitanju. Zakoni o zaštita novih biljnih sorti u osnovi poštuju UPOV konvenciju. Deo zakona o semenu koji se odnosi na kvalitet semena zasniva se na pravilima međunarodne organizacije ISTA. Ono što Evropska unija reguliše za sve zemlje članice je jedinstveno tržište semenom. Kroz taj sistem uređuje se pitanje finansij-ske podrške proizvođačima semena, što je jedan od mehanizama regula-cije površina pod pojedinim biljnim vrstama, odnosno ukupne semenske proizvodnje. Ako je podrška po hektaru proizvedenog semena veća to će usloviti i veće površine ukupno pod tom biljnom vrstom. Tržište seme-nom se reguliše, osim cenovnom podrškom i uređenjem sistema unutraš-nje organizacije tržišta i carinskog mehanizma.

105
Opšte semenarstvo
O međunarodnim organizacijama vezanim za semenarstvo je već bilo reči, te će ovde biti izneta osnova regulative Saveta Evropske komi-sije (EC) broj 1947/2005 od 23. novembra 2005. godine o zajedničkom organizovanju tržišta semena u Evropskoj uniji.
Regulativa Saveta EC broj 1947/2005 (Council regulation EC no. 1947/2005) od 23. novembra 2005. godine reguliše svojim odredbama zajedničko tržište semena i u isto vreme stavlja van važnosti propis EEC broj 2358/71(Council Regulation EEC no. 2358/71) i EEC broj 1674/72 (Council Regulation EEC no. 2358/71). Regulativa je objavljena u Služ-benom listu Evropske unije L 312 , 29/11/2005 P. 0003 – 0007. Pri izra-di regulative uzet je u obzir Ugovor o osnivanju Evropske zajednice, predlog Evropske komisije, mišljenje Evropskog parlamenta i mišljenje Evropskog ekonomskog i socijalnog komiteta, što ukazuje na široko obavljenu raspravu vezanu za regulative.
Regulativa Saveta EC broj 1947/2005 posebno uzima u obzir rad i razvoj jedinstvenog tržišta poljoprivrednih proizvoda koji treba da prati zajedničku poljoprivrednu politiku (Common Agricultural Policy) Evropske unije, da uključi zajedničku organizaciju poljoprivrednih tr-žišta koja mogu imati različite oblike zavisno od proizvoda koji je u pitanju. Osnova regulative je i u tome da speči turbulencije na tržištu semenom u Evropskoj uniji. Neophodne odrednice za sprovođenje ove regulative treba da su u skladu sa Odlukom Saveta 1999/468/EC od 28. juna 1999. godine (Council Decision 1999/468/2004) koja u opštim odredbama daje područja kojom regulativa pokriva biljne vrste nazna-čene u tabeli 22.
Regulativa Saveta EC broj 1947/2005 određuje vreme prodaje seme-na koje počinje 1. jula svake godine i traje do 30. juna naredne godine. Istom regulativom su određena pravila trgovine sa trećim zemljama, u koje spada i Srbija. Svaki član Zajednice daje listu sorti koje, zajedno sa onima koje su dali drugi članovi, sačinjavaju Zajednički katalog ra-tarskog i povrtarskog bilja EU (EU Common Catalogue). Samo sorte iz zajedničkog kataloga mogu da se nađu na tršištu u EU.
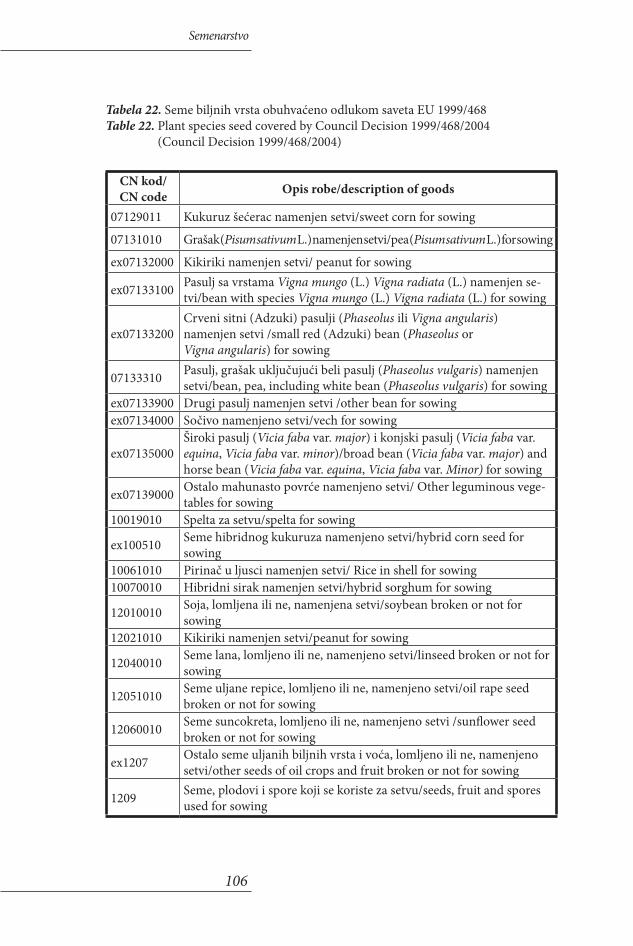
106
Semenarstvo
Tabela 22. Seme biljnih vrsta obuhvaćeno odlukom saveta EU 1999/468Table 22. Plant species seed covered by Council Decision 1999/468/2004
(Council Decision 1999/468/2004)
CN kod/ CN code Opis robe/description of goods
07129011 Kukuruz šećerac namenjen setvi/sweet corn for sowing
07131010 Grašak (Pisum sativum L.) namenjen setvi/pea (Pisum sativum L.) for sowing
ex07132000 Kikiriki namenjen setvi/ peanut for sowing
ex07133100 Pasulj sa vrstama Vigna mungo (L.) Vigna radiata (L.) namenjen se-tvi/bean with species Vigna mungo (L.) Vigna radiata (L.) for sowing
ex07133200Crveni sitni (Adzuki) pasulji (Phaseolus ili Vigna angularis) namenjen setvi /small red (Adzuki) bean (Phaseolus or Vigna angularis) for sowing
07133310 Pasulj, grašak uključujući beli pasulj (Phaseolus vulgaris) namenjen setvi/bean, pea, including white bean (Phaseolus vulgaris) for sowing
ex07133900 Drugi pasulj namenjen setvi /other bean for sowingex07134000 Sočivo namenjeno setvi/vech for sowing
ex07135000Široki pasulj (Vicia faba var. major) i konjski pasulj (Vicia faba var. equina, Vicia faba var. minor)/broad bean (Vicia faba var. major) and horse bean (Vicia faba var. equina, Vicia faba var. Minor) for sowing
ex07139000 Ostalo mahunasto povrće namenjeno setvi/ Other leguminous vege-tables for sowing
10019010 Spelta za setvu/spelta for sowing
ex100510 Seme hibridnog kukuruza namenjeno setvi/hybrid corn seed for sowing
10061010 Pirinač u ljusci namenjen setvi/ Rice in shell for sowing 10070010 Hibridni sirak namenjen setvi/hybrid sorghum for sowing
12010010 Soja, lomljena ili ne, namenjena setvi/soybean broken or not for sowing
12021010 Kikiriki namenjen setvi/peanut for sowing
12040010 Seme lana, lomljeno ili ne, namenjeno setvi/linseed broken or not for sowing
12051010 Seme uljane repice, lomljeno ili ne, namenjeno setvi/oil rape seed broken or not for sowing
12060010 Seme suncokreta, lomljeno ili ne, namenjeno setvi /sunflower seed broken or not for sowing
ex1207 Ostalo seme uljanih biljnih vrsta i voća, lomljeno ili ne, namenjeno setvi/other seeds of oil crops and fruit broken or not for sowing
1209 Seme, plodovi i spore koji se koriste za setvu/seeds, fruit and spores used for sowing

107
Opšte semenarstvo
Zakonski propisi iz oblasti semenarstva u Srbiji
Postojanje zakonskih propisa o semenu u srednje razvijenim ze-mljama i zemljama u razvoju je opravdano kada su sve zainteresovane strane za proizvodnju i promet semena svesne značaja njegovog kva-liteta, obezbeđuju kontinuitet razvojnih programa, marketinga i po-većavaju potrošnju semena dobrog kvaliteta (deklarisanog semena). Propisi o kvalitetu semena i domaći standardi oslanjaju se na propise i standarde međunarodnih organizacija i to: UN/EU, ISTA, OECD, ISO, UPOV i dr.
Zakonski propisi u oblasti semenarstva regulišu:– organizovanje proizvodnje sortnog semena tako da se obezbedi
dobijanje semena visokog kvaliteta,– izvođenje procesa dorade, ispitivanje, deklarisanje i puštanje u
promet sortnog semena, – određivanje odgovornih lica za izvođenje pomenutih radnji,– priznavanje sorti i hibrida,– zaštitu autorskih prava stvaraoca sorti i hibrida.
Zakon o semenu
Zakon o semenu (Službeni glasnik RS, broj 45/05) u osnovnim odredbama definiše pojam semena i sorte i određuje kategorije semena. Zakon uređuje uslove i način proizvodnje, dorade, korišćenja, prometa, uvoza i ispitivanja kvaliteta semena poljoprivrednog bilja, rasada poljo-privrednog bilja i micelija jestivih i lekovitih gljiva. Na osnovu podnetih zahteva određuje se ko može proizvoditi i dorađivati seme, uz obavezu ispunjenja propisanih uslova. Proizvodnja semena podleže obaveznoj stručnoj kontroli čime se utvrđuje poreklo upotrebljenog semena, vrsta, sorta i kategorija. U pogledu kvaliteta semena zakonom se propisuje ko može da vrši ispitivanje kvaliteta i koje uslove treba da ispunjava, kao i uslove za pakovanje, deklarisanje i obeležavanje. Zakon uređuje prizna-vanje novostvorenih domaćih sorti i odobravanje uvođenja u proizvod-nju stranih sorti, kao i upis sorti poljoprivrednog bilja u Registar sorti poljoprivrednog bilja.

108
Semenarstvo
Za prateću regulativu, koja bliže određuje primenu Zakona, mogu se izdvojiti Pravilnik o kvalitetu semena poljoprivrednog bilja (Službe-ni list SFRJ broj 47/87) (poslednje izmene – Službeni glasnik RS, broj 23/09), Pravilnik o kontroli proizvodnje semena, sadržini i načinu vo-đenja evidencije o proizvodnji rasada poljoprivrednog bilja i obrascu izveštaja o proizvodnji micelija jestivih i lekovitih bilja (Službeni glasnik RS, broj 60/06).
U postupku revidiranja Zakona o semenu i usklađivanja sa naj-novijim propisima u EU, planira se preraspodela oblasti čime bi se važeći zakon o semenu raščlanio na dva zakona, Zakon o priznavanju sorti i novi Zakon o semenu. Novi zakon o semenu će se razlikovati od trenutno važećeg po tome što neće obuhvatati samo seme poljo-privrednog bilja, već i sadni materijal poljoprivrednog i ukrasnog bi-lja, odnosno celokupni reprodukcioni materijal (osim šumskih biljnih vrsta). Do sada su ovu oblast pokrivala tri Zakona: Zakon o semenu (Službeni glasnik RS, broj 45/05) i Zakon o semenu i sadnom mate-rijalu (Službeni glasnik RS, broj 54/93). Kada se ove oblasti objedine, dobiće se novi Zakon o semenu i sadnom materijalu poljoprivrednog bilja. Neke od novina ovog zakona bi bile: uvođenje kategorizacije se-mena u skladu sa direktivama EU, obaveza licenciranja lica koja vrše poslove stručne kontrole u polju (aprobatori), formiranje i vođenje je-dinstvenog Registra svih subjekata u poslovima proizvodnje, dorade i prometa semena poljoprivrednog bilja prema delatnostima, definisa-nje obaveze prijavljivanja i kontrole predosnovnog semena, obaveza pribavljanja sertifikata za seme, pravila za deklarisanje semena u pro-metu. Deo zakona koji se odnosi na micelije jestivih i lekovitih gljiva, neće biti predmet ovog Zakona.
Na osnovu člana 6. stav 1. Zakona o tehničkim zahtevima za pro-izvode i ocenjivanje usaglašenosti (Službeni glasnik RS, broj 36/09) donet je Pravilnik o izmenama i dopunama pravila o kvalitetu semena poljoprivrednog bilja (Službeni glasnik RS, broj 72/10). U Pravilniku se daju metode ispitivanja kvaliteta semena, poljoprivrednog bilja, koji je usaglašen sa pravilima ISTA (ISTA Rules, 2011). Osnovu ovog Pravilni-ka čini Pravilnik o ispitivanju kvaliteta semena (Službeni glasnik SFRJ, broj 47/87).

109
Opšte semenarstvo
Zakon o priznavanju sorti poljoprivrednog bilja
Zakon o priznavanju sorti poljoprivrednog bilja (Službei glasnik RS, broj 30/10) reguliše priznavanje novostvorenih domaćih sorti ratarskog i povrtarskog bilja, voćaka i vinove loze i odobravanje uvođenja u proi-zvodnju stranih sorti ratarskog i povrtarskog bilja. U periodu do 2005. godine, oblast priznavanja sorti je bila regulisana Zakonom o priznavanju sorti poljoprivrednog i šumskog bilja (Službeni glasnik SRJ, broj 12/98 i Službeni glasnik SRJ, broj 37/02) i Zakonom o sadnom materijalu voćaka, vinove loze i hmelja (Službeni glasnik RS, broj 18/05). S obzirom da ovi zakoni regulišu različite oblasti, pojavili u se problemi prilikom njihove primene, jer u delu koji se odnosi na postupak priznavanja sorti i upisa u Registar sorti, zakoni nisu u popunosti usaglašeni. Zakoni nisu usaglašeni ni sa pravnim tekstovima EU. Donošenjem ovog zakona prevazilaze se na-pred navedeni problemi i oblast priznavanja sorti poljoprivrednog bilja je definisana jednim zakonom koji je usaglašen sa relevatnim propisima EU, kao i sa UPOV konvencijom. Zakonom se reguliše izvođenje DUS i VCU testova, definiše pojam odomaćene sorte, održavanje sorti, referentnih ko-lekcija. Definisani su pojmovi sorta, sorta standard i standardni uzork.
Uz zakon su doneta i brojna podzakonska akta, koja praktično omogućavaju primenu Zakona o priznavanju sorti poljoprivrednog bi-lja. Pravilnik o potrebnoj količini semena, odnosno sadnog materijala koji se dostavlja radi ispitivanja, kao i vreme dostavljanja tog semena, odnosno sadnog materijala, u zavisnosti od vrste bilja (Službeni glasnik RS, broj 11/10). Pravilnik propisuje potrebnu količinu semena i sadnog materijala za ispitivanje proizvodne i upotrebne vrednosti sorte i za is-pitivanje različitosti, uniformnosti i stabilnosti sorte kao i vreme dostav-ljanja semena odnosno, sadnog materijala.
Pravilnik o vrstama poljoprivrednog bilja za koje se mogu vršiti do-datna ispitivanja sorte i metodama ispitivanja sorti tog bilja (Službeni glasnik RS, broj 6/11) daje mogućnost da se dodatno mogu ispitati sorte kukuruza, suncokreta i šećerne repe. Ograničen je broj sorti koje mogu dodatno da se ispituju i to za kukuruz do pet sorti i to po jedna sorta iz svake grupe zrenja FAO od 300 do 700. Za suncokret to iznosi do šest sorti (iz grupe zrenja rani i srednje rani) i za šećernu repu do šest sorti. Ispitivanja se vrše za proizvodnu vrednost sorti koje su upisane u Regi-star sorti poljoprivrednog bilja.

110
Semenarstvo
Pravilnik o načinu određivanja imena sorte poljoprivrednog bi-lja (Službeni glasnik RS, broj 76/10) bliže određuje način određiva-nja imena sorte poljoprivrednog bilja. Pravilnik određuje da ime sorte može biti određeno u obliku zamišljenog naziva ili u obliku koda od-nosno šifre, u skladu sa zakonom kojim se uređuje priznavanje sorti poljoprivrednog bilja. Naziv sorte može da se sastoji od najviše tri reči, odnosno celine. Izuzetno može da sadrži više reči ako je sorta priznata u nekoj drugoj zemlji. Pravilnik o sadržini i načinu vođenja registra sorti poljoprivrednog bilja (Službeni glasnik RS, broj 73/10) defini-še se način vođenja Registra za domaće i odomaćene sorte ratarskog i poljoprivrednog bilja, strane sorte voća i vinove loze, odomaćenim sortama ratarskog i povrtarskog bilja, voća i vinove loze, brisanim sor-tama poljoprivrednog bilja i privremeno priznatim sortama povrtar-skog bilja.
Vode se tri Registra:I - lista priznatih sorti poljoprivrednog bilja;II -lista brisanih sorti poljoprivrednog bilja;III - lista privremeno priznatih sorti povrtarskog bilja.
Pravilnik o sadržini i obrascu zahteva za priznavanje sorte poljopri-vrednog bilja, kao i dokumentaciji koja se uz taj zahtev prilaže (Službeni glasnik RS, broj 53/10) reguliše postupak priznavanja sorte, koji vlasnik sorte, odnosno njegov ovlašćeni zastupnik podnosi ministarstvu nad-ležnom za poslove poljoprivrede u skladu sa zakonom kojim se uređuje priznavanje sorti poljoprivrednog bilja.
Zakon o zaštiti prava oplemenjivača biljnih sorti
Zakon o zaštiti prava oplemenjivača biljnih sorti i šumskog bilja iz 2000. godine (Službeni list SRJ, broj 28/00) je imao suštinske ne-dostatke u pogledu definisanja prava i obaveza nosioca prava ople-menjivača, kao i usklađenosti sa UPOV Konvencijom, zbog čega do danas nije imao primenu u praksi i u skladu sa njim, nije uspostavljen sistem zaštite prava oplemenjivača. U 2009. godini usvojen je novi Za-kon o zaštiti prava oplemenjivača biljnih sorti (Službeni glasnik RS, broj 41/09) kojim se uređuju uslovi, način i postupak za zaštitu prava

111
Opšte semenarstvo
oplemenjivača biljnih sorti. Ovaj zakon je usklađen sa UPOV Konven-cijom iz 1991.godine, odredbama TRIPS sporazuma (Agreement on Trade -Related Aspects of Intellectual Property Rights) koji se odnose na zaštitu intelektualne svojine, kao i sa ostalim relevantnim propisima Evropske unije. Ovim će se omogućiti pristupanje Republike Srbije UPOV Konvenciji i članstvo u UPOV-u, a ispuniće se i jedan od uslo-va da Republika Srbija pristupi Svetskoj trgovinskoj organizaciji i EU. Uspešna međunarodna trgovina zahteva usklađenost pravila. Dono-šenjem novog zakona i članstvom u UPOV-u uz praktičnu primenu sistema zaštite prava oplemenjivača, intenzivirala bi se međunarodna saradnja usmerena ka ispitivanju novih biljnih sorti. Ovo je važno i zbog plasmana domaćeg semena i sadnog materijala na inostrana tr-žišta.
Trenutno su na snazi osim zakona i sledeći pravilnici iz ove obla-sti: Pravilnik o sadržini i načinu vođenja Registra zahteva za dodeljiva-nja prava oplemenjivača biljnih sorti, Registra zaštićenih biljnih sorti, Registra prenesenih prava oplemenjivača i Registra ugovora o licenci (Službeni glasnik RS, broj 70/09) i Pravilnik o obrascu i sadržini zahteva za dodeljivanje prava oplemenjivača biljnih sorti i dokumentaciji koja se prilaže uz ovaj zahtev i načinu dostavljanja uzoraka reprodukcionog materijala sorte (Službeni glasnik RS, broj 82/09), kao i Pravilnik o listi vrsta poljoprivrednog bilja na koje se odnose izuzeci od prava opleme-njivača i o elementima za određivanje malih poljoprivrednih proizvođa-ča (Službeni glasnik RS, broj 38/10).
Zakon o potvrđivanju međunarodne konvencije o zaštiti novih biljnih sorti
Ovim zakonom se potvđuje Međunarodna konvencija o zaštiti no-vih biljnih sorti koja je doneta u Parizu 1961. godine, a izmenjena u Ženevi 1972., 1978. i 1991. godine. U zakonu se nalazi originalan tekst Konvencije iz 1991. godine na engleskom i srpskom jeziku. Donošenje ovog Zakona je uslov za učlanjenje Srbije u UPOV i on sa Zakonom o zaštiti prava oplemenjivača biljnih sorti čini celinu vezanu za polje za-štite sorti.

112
Semenarstvo
Zakon o zdravlju bilja
U 2009. godini, usvojen je novi Zakon o zdravlju bilja (Službeni gla-snik RS, broj 41/09) kojim se uređuje zaštita i unapređenje zdravlja bilja; mere za sprečavanje unošenja, otkrivanje, sprečavanje širenja i suzbijanje štetnih organizama; fitosanitarna kontrola; uslovi za proizvodnju, prera-du, doradu, uvoz, skladištenje i promet bilja, biljnih proizvoda i propisa-nih objekata, kao i uslovi za pružanje usluga u oblasti zaštite zdravlja bi-lja. Ovim zakonom obezbeđuje se sprovođenje međunarodnih obaveza u skladu sa preporukama IPPC (International Plant Protection Convention), sporazumom o primeni sanitarnih i fitosanitarnih mera (SPS agreement), međunarodnim konvencijama i drugim međunarodnim sporazumima i drugim nacionalnim organizacijama odgovornim za zdravlje bilja. Zakon je usklađen sa glavnim principima EU (Council Directive 2000/29/EC).
Sprovođenje Zakona omogućavaju doneti pravilnici. Pravilnik o zdravstvenom pregledu useva i objekata za proizvodnju semena, rasada i sadnog materijala i zdravstvenom pregledu semena, rasada i sadnog ma-terijala (Službeni list SRJ, broj 66/99) reguliše način pregleda useva u polju sa zdravstvenog aspekta, isto tako i semena u laboratoriji. Pravilnik je pre-trpeo najnovije izmene u 2010. godini (Službeni glasnik RS, broj 107/10).
Pravilnik o listama štetnih organizama i listama bilja, biljnih pro-izvoda i propisanih objekata (Službeni glasnik RS, broj 7/10) utvrđuju liste štetnih organizama i liste bilja, biljnih proizvoda i propisanih obje-kata i to: Lista IA deo I Štetni organizmi za koje nije poznato da su pri-sutni na teritoriji Republike Srbije i čije je unošenje i širenje u Republiku Srbiju zabranjeno, Lista IA deo II Štetni organizmi za koje je poznato da su prisutni na ograničenom području Republike Srbije i čije je unošenje i širenje u Republiku Srbiju zabranjeno, Lista IIA i deo II Štetni orga-nizmi za koje je poznato da su prisutni na ograničenom području Re-publike Srbije i čije je unošenje i širenje u Republiku Srbiju zabranjeno, ako su prisutni na određenom bilju, biljnim proizvodima i propisanim objektima, Lista IIIA Vrste bilja, biljnih proizvoda i propisanih objekata čije je uvoz zabranjen u Republiku Srbiju, Lista IVA deo I Vrste bilja, biljnih proizvoda i propisanih objekata za koje su propisani specifični fi-tosanitarni uslovi pri uvozu, Lista VA deo I Vrsta bilja, biljnih proizvoda i propisanih objekata za koje je obavezan fitosanitarni pregled, radi iz-davanja biljnog pasoša i Lista VB deo I Vrsta bilja, biljnih proizvoda i propisanih objekata za koje je pri uvozu obavezan fitosanitarni pregled

113
Opšte semenarstvo
koje mora da prati fitosanitarni sertifikat, koje su odštampane uz ovaj Pravilnik i čine njegov sastavni deo.
Pravilnik o fitosanitarnoj kontroli bilja, biljnih proizvoda i propi-sanih objekata u međunarodnom prometu (Službeni glasnik RS, broj 32/10) propisuju se način najavljivanja pošiljke bilja, biljnih proizvoda i propisanih objekata, obrazac zahteva za pregled pošiljke bilja, biljnih proizvoda i propisanih objekata, rok za njegovo podnošenje i uslovi koje uvoznik mora da obezbedi radi obavljanja fitosanitarnog pregleda, uslo-vi, način i postupak vršenja fitosanitarnog pregleda pošiljke bilja, bilj-nih proizvoda i propisanih objekata, način uzorkovanja i slanja uzoraka, kao i broj i veličina uzoraka, sadržaj i veličina pečata kojim se overava carinska dokumentacija, oblik i sadržina pečata kojim se poništava fito-sertifikat, odnosno fitosertifikat za reeksport, izgled i sadržina obrasca obaveštenja o preduzimanju fitosanitarnih mera, način, mesto i postu-pak vršenja fitosanitarnog pregleda pošiljaka bilja, biljnih proizvoda i propisanih objekata pri izvozu, obrazac i sadržaj zahteva za izdavanje fitosertifikata, sadržaj i obrazac fitosertifikata i fitosertifikata za reek-sport, uslovi pod kojima se prihvata elektronska forma fitosertifikata, kao i način postupanja sa pošiljkama bilja, biljnih proizvoda i propisanih objekata u prevozu i način vršenja fitosanitarnog pregleda tih pošiljaka.
Pravilnik o higijensko-tehničkim, radnim i drugim uslovima koje mo-raju da ispunjavaju granični prelazi na kojima postoji organizovana fitosa-nitarna inspekcija (Službeni glasnik RS, broj 37/10) propisuje higijensko-tehnički, radni i drugi uslovi koje moraju da ispunjavaju granični prelazi na kojima postoji organizovana fitosanitarna inspekcija, a preko kojih se vrši uvoz, prevoz i izvoz bilja, biljnih proizvoda i propisanih objekata, uvoz i tranzit sredstava za ishranu bilja i oplemenjivača zemljišta, kao i uvoz i tran-zit sredstava za zaštitu bilja i aktivne supstance, odnosno osnovne supstance.
Pravilnik o utvđivanju programa mera zaštite zdravlja bilja za 2010. godinu (Službeni glasnik RS, broj 42/10) utvrđuje Program mera zaštite zdravlja bilja za 2010. godinu, koji je odštampan uz Pravilnik i čini nje-gov sastavni deo.
Konkretne mere zaštite zdravlja bilja, rokovi, način sprovođenja tih mera, subjekti koji će ih sprovoditi, izvori i način obezbeđivanja i ko-rišćenja sredstava, kao i način kontrole sprovođenja mera utvrđeni su programom iz člana 1. ovog pravilnika. Pravilnik o načinu upisa u regi-star pružalaca usluga u oblasti zaštite zdravlja bilja, sadržaju i obrascu zahteva, sadržini, kao i načinu vođenja tog registra (Službeni glasnik RS,

114
Semenarstvo
broj 46/10) bliže se propisuje način upisa u Registar pružalaca usluga u oblasti zaštite zdravlja bilja, sadržaj i obrazac zahteva, sadržina, kao i način vođenja Registra pružalaca usluga.
Pravilnik o uslovima u pogledu objekata, opreme i stručno ospo-sobljenog kadra koje mora da ispunjava pravno lice i preduzetnik koji se upisuje u registar pružalaca usluga u oblasti zaštite zdravlja bilja, u zavisnosti od vrste usluga u oblasti zaštite zdravlja bilja (Službeni gla-snik RS, broj 46/10) bliže propisuje uslove u pogledu objekata, opre-me i stručno osposobljenog kadra koje mora ispunjavati pravno lice i preduzetnik koji se upisuje u Registar pružalaca usluga u oblasti zaštite zdravlja bilja, u zavisnosti od vrsta usluga u oblasti zaštite zdravlja bilja.
Usluge u oblasti zaštite bilja iz stava 1. ovog člana jesu:1) laboratorijske analize i testiranje bilja, biljnih proizvoda i propi-
sanih objekata, radi utvrđivanja prisustva štetnih organizama,2) praćenje i prognoza pojave štetnih organizama, razvoj i kretanja
njihove populacije i utvrđivanje optimalnih rokova za njihovo suzbija-nje i davanje preporuka u skladu sa tim,
3) primenjena i druga istraživanja u oblasti zaštite zdravlja bilja,4) edukacija i davanje saveta i preporuka o štetnim organizmima i
njihovom suzbijanju,5) suzbijanje štetnih organizama primenom dezinfekcije, dezinsekcije,
deratizacije, dekontaminacije i primenom drugih postupaka tretiranja,6) izdavanje biljnog pasoša.Pravilnik o službenoj legitimaciji i službenom odelu fitosanitarnog
inspektora, kao i načinu vođenja evidencije o izdatim službenim legitima-cijama (Službeni glasnik RS, broj 48/10) bliže propisuje obrazac i sadržina službene legitimacije fitosanitarnog inspektora i graničnog fitosanitarnog inspektora, izgled službenog odela fitosanitarnog inspektora i graničnog fitosanitarnog inspektora, oblik znaka graničnog fitosanitarnog inspek-tora, kao i način vođenja evidencije o izdatim službenim legitimacijama fitosanitarnog inspektora i graničnog fitosanitarnog inspektora.
Pravilnik o utvrđivanju vrsta bilja, biljnih proizvoda i propisanih objekata koje se ne nalaze na listi VA deo I i listi VA deo II (Službeni glasnik RS, broj 64/10) utvrđuju određene vrste bilja, biljnih proizvoda i propisanih objekata koji se ne nalaze na Listi VA deo I i Listi VA deo II, a za koje se pravna lica, preduzetnici i fizička lica koja se bave proizvod-njom, preradom, doradom, uvozom, skladištenjem i prometom upisuju u Registar proizvođača, prerađivača, dorađivača, uvoznika, skladištara i prometnika bilja, biljnih proizvoda i propisanih objekata.

115
Opšte semenarstvo
Pravilnik o uslovima i načinu vršenja pregleda i uzorkovanja pošilj-ke pri uvozu, načinu najavljivanja prispeća pošiljke, obrascu i sadržini zahteva za pregled pošiljke i uslovima koje uvoznik mora da obezbedi radi obavljanja fitosanitarnog pregleda, kao i načinu dostavljanja uzora-ka, broju i veličini uzoraka radi ispitivanja i načina postupanja sa odu-zetom pošiljkom (Službeni glasnik RS, broj 86/10) bliže propisuje uslove i način vršenja pregleda i uzorkovanja pošiljke pri uvozu, način najav-ljivanja prispeća pošiljke, obrazac i sadržina zahteva za pregled pošiljke i uslovi koje uvoznik mora da obezbedi radi obavljanja fitosanitarnog pregleda, kao i način dostavljanja uzoraka, broj i veličina uzoraka radi ispitivanja i način postupanja sa oduzetom pošiljkom.
Pošiljka u smislu ovog pravilnika, jeste količina jedne iste vrste sredstava za ishranu bilja, oplemenjivača zemljišta ili sirovine za njihovu proizvodnju koja je pristigla na granični prelaz.
Zakon o genetički modifikovanim organizmima
Na snazi je Zakon o genetički modifikovanim organizmima iz 2009.godine (Službeni glasniku RS br. 41/09) kojim je uvedena zabrana stav-ljanja u promet genetički modifikovanih organizama i proizvoda od ge-netički modifikovanih organizama (član 2). Trenutno se radi na izmena-ma i dopunama aktuelnog zakona, čiji će potom predmet uređivanja biti postupci i procedure za upotrebu u zatvorenim sistemima, za namerno uvođenje u životnu sredinu, za stavljanje u promet odnosno za prevoz GMO i proizvoda od GMO, kao i uslovi za obavljanje navedenih nači-na upotrebe. Novi zakon će regulisati i obeležavanje, rukovanje, prevoz i pakovanje GMO i proizvoda od GMO, od značaja za GMO. Zakon propisuje da je osnovno i obavezno da se pre upotrebe GMO sprovede procena rizika i to po principu „slučaj po slučај” a poštujući princip predostrožnosti, kako bi se postavile mere za sprečavanje i otklanjanje neželjenih efekata prilikom upotrebe GMO. Zakon je usaglašen sa di-rektivom EU 2001/18/EC (Cuncil Directive EU 2001/18/EC).
Podzakonska akta iz 2002. godine biće zamenjena kada se nakon usvajanja novog zakona pripreme novi pravilnici koji će biti usklađeni sa relevantnim propisima EU iz ove oblasti (2001/18/EC, 1946/2003/EC, 65/2004, EC 1829/2003, 1830/2003 i dr.)(Council Decision C(2000) 146/Final incl. 2003, 2004, 2005, 2006, 2007 and 2008, Council Regu-lation (EC) 1946/2003, Commision Regulation (EC) 65/2004, Council Regulation (EC) 1829/2003, Council Regulation (EC) 1830/2003).

116
Semenarstvo
EKOLOGIJA SEMENA
Integralno ratarenje, prinos i kvalitet semena
Neophodno je poznavanje ciklusa kruženja energije, hranljivih ma-terija, kretanje semena i polena, međusobni odnosi između prinosa i uticaja uslova spoljne sredine da bi se razumelo i upravljalo prirodnim resursima (Environment Plant Inspection, http://www.scri.uk/research/epi/agroecology/fieldandlansscape).
Integralno ratarenje promoviše viziju usmerene poljoprivredne proizvodnje, koja se zasniva i odvija na kvalitetu, a ne na količini proi-zvedenog semena. Ova vrsta ratarenja ne prihvata čisto tehnički pristup proizvodnji. Njeni glavni principi kombinuju novine u proizvodnji sa ponovno uspostavljenim značajem koji se daje individualnom proizvo-đaču, njegovoj ekonomskoj i socijalnoj ulozi.
Integralno ratarenje se samoprilagođava specifičnim karakteristika-ma svakog regiona i na taj način štiti postojeće resurse (zemljište, vodu i dr.). Zemljište sadrži prirodne supstance (organske i mineralne mate-rije) u tolikoj meri da može da zaštiti biljku od eventualnih spoljašnjih negativnih uticaja i smanji zagađenja, ekonomsku opterećenost proi-zvođača u kupovini komercijalnih hemijskih supstanci. Iz tog razloga je važno izvršiti analizu zemljišta svake parcele pre nego što se krene u proizvodnju semena.
Integralno ratarenje dozvoljava da se kombinuju ekonomski uslovi, uslovi okruženja i bezbednost hrane. Ono igra važnu ulogu u promociji hrane jer favorizuje metode proizvodnje koji su održivi u odnosu na okruženje, profitabilni za proizvođača, a potrošačima obezbeđuju kva-litetnu hranu, a seme je osnov proizvodnje hrane (http://www.fdh.org/-Agro-ecology-.html?lang=fr).
Spoljašnji uslovi tokom vegetacije mogu značajno da utiču na pri-nos semena, njegov kvalitet i vigor.
Prema Dornbos (1995) postoje četiri važna činioca koji utiču na prinos i vigor semena:

117
Opšte semenarstvo
1) pravilna primena agrotehničkih mera u proizvodnji semenskih useva koji umanjuju posledice stresa,
2) proizvodnju semena locirati u regionima koji imaju povoljne uslove za razvoj određene biljne vrste,
3) birati parcele za proizvodnju koje je moguće navodnjavati, 4) kod upotrebe nekih sorti u proizvodnji voditi računa da su one
stabilne, uniformne i da mogu davati stabilne prinose u različitim kli-matskim uslovima.
Iz napred navedenog, može se zaključiti da se sa sigurnošću ne mogu predvideti spoljašnji uslovi za proizvodnju semena u određenom području. Zbog toga je veoma značajno pratiti variranja spoljašnjih či-nilaca i njihov uticaj na fiziološke procese koji određuju kvalitet semena.
Uslovljenost proizvodnje semena spoljašanjim činiocima zavisi od:– uslova u kojima se razvijala majčinska biljka,– dostizanje najvišeg kvaliteta semena tačnim ustanovljavanjem
momenata pune zrelosti i procena gubitka kvaliteta,– određivanje posledica stresa na kvalitet semena i uslovljenost iz-
među kvaliteta semena, prinosa i komponenti prinosa,– uticaj fizioloških promena na kvalitet semena,– uzroci koji dovode do fizioloških promena, a utiču na umanjenje
kvaliteta semena izazvanog stresnim uslovima u toku razvoja se-mena na majčinskoj biljci,
– mere obrade i nege useva koje mogu da umanje delovanje stresnih uslova tokom vegetacije.
Kvalitet semena salate i njegova sposobnost da održi kvalitet u vre-me skladištenja uslovljavaju genotip i uslovi spoljne sredine. Contreras i sar. su 2009. godine ispitivali kako niske temperature u kojima je gajena majčinska biljka utiču na kvalitet semena. Seme sorte salate Tango je proizvedeno u komorama pod različitim uslovima:
– naizmenično visokoj emperaturi 30/20oC dan/noć i – naizemnično niskoj temperaturi 20/10oC.Seme proizvedeno na niskoj temperaturi je bilo teže 37% u odnosu
na seme gajeno pri visokoj temperaturi (optimalna temperatura za uz-goj salate je 20oC uz prisustvo svetlosti). Klijavost uz prisustvo svetlosti i temperaturi 30oC i 20oC je bila slična za seme u oba tretmana.
Seme šećerne repe je izuzetno osetljivo na nedostatak bora. Ukoliko u vreme gajenja biljaka nema dovoljno bora dolazi do smanjenja prino-sa i kvaliteta semena (Dordas et al, 2007).
118
Semenarstvo
Fiziološki model održavanja i gubitka kvaliteta semena
Kvalitet semena može biti ograničen spoljašnjim uslovima do fizi-ološke zrelosti kada ono dostiže najveću masu, klijavost i vigor. Model gubljenja kvaliteta semena soje u funkciji vremena, tokom razvoja biljke i tokom skladištenja semena prikazano je na slici 36. Kada je seme soje nedozrelo i kada se žanje sa masom od 62, 100, 143 i 167 mg semena-1 (fiziološka zrelost), klijavost semena je 67, 97 i 99%, a 6, 34, 37, 73 i 99% daje tipičan ponik (Miles et al., 1988).
Činioci spoljne sredine imaju veći uticaj na kvalitet semena u fazi nalivanja semena nego u fazi vegetativnog porasta. Stres koji izazivaju dugotrajne visoke temperature tokom razvoja semena soje mogu imati negativan uticaj na njegov kvalitet. Uslovi suše između druge i četvrte faze organogeneze kod soje imaju veći uticaj na kvalitet semena nego u petoj etapi kada se klijavost semena soje umanjuje za oko 10%. Stres koji se ispoljava u šestoj fazi umanjuje klijavost za oko 8%. Tada dolazi do pojave atipičnog ponika i to uglavnom do jakog skraćivanja hipokotila (Dornbos, 1995).
Slika 36. Porast klijavosti semena soje tokom razvoja i njegov pad tokom skladištenja
Figure 36. The increase in germination of soybean seeds during development and its decline during storage
(Miles et al., 1988)
Proc
enat
/Per
cent
(%)
Vreme/Time
Razvoj/Development Skladištenje/Storage
Vigor/VigourTipičan ponik/Typical seedling
Klijavost/Germination
100
80
60
40
20
0
PM

119
Opšte semenarstvo
Suša i visoke temperature dovode do manjih gubitaka u prinosu ukoliko se ispolje u vegetativnoj nego u generativnoj fazi, mada to zavisi od biljne vrste. Suša kod useva soje, ako do nje dođe u ranoj fazi vege-tativnog razvoja, ne utiče na prinos (Meckel et al., 1980). Kod kukuruza pak stresni uslovi tokom ranog vegetativnog perioda mogu dovesti do smanjenja prinosa zbog toga što razvoj reproduktivnih primordija tada određuju broj redova semena u klipu, kao i broj semena u redu. Među-tim uslovi stresa u toku vegetacije malo utiču na kvalitet semena. Dok stres tokom cvetanja i oprašivanja može smanjiti prinos. Direktan efe-kat na klijavost ili vigor nije dokazan. Malo je podataka o uzajamnom dejstvu između spoljašnjih činilaca tokom vegetacije i kvaliteta semena tokom perioda između oprašivanja i formiranja mahuna kod legumino-za i ranih faza formiranja semena kod trava.
Biljke soje su izlagane vodnom stresu u periodu nalivanja semena do fiziološke zrelosti u polju i staklari. Stres je umanjio prinos semena (5–38%) i krupnoću semena (11–35%), ali nije uticao na broj semena. Stres je umanjio produktivnost fotosinteze u oba ispitivanja, te bi se pre moglo diskutovati o uticaju efekta vodnog stresa na efekat formiranja lisne mase, nego na prinos (Egli i Bruening, 2004).
Stres koji izazivaju dugotrajne visoke temperature kod razvoja se-mena soje mogu imati negativan uticaj na njegov kvalitet. Uticaj visoke temperature 37/30oC dan/noć, je ispitivan na zrelom suvom semenu, koje je poticalo sa biljaka gajenih u kontrolisanim uslovima staklare. Seme koje se razvilo pod uslovima visoke temperature je imalo niži kva-litet u odnosu na seme koje je dobijeno sa biljaka gajenih pri tempera-turi 27/18 oC dan/noć. Uticaj visoke temperature se ogledao u tome što je dobijeno sitnije seme niže klijavosti, sa većim procentom atipičnih ponika. Visoka temperatura je negativno uticala na ćelijske membra-ne što je rezultiralo gubitkom elektorlita. Zastupljenost pojedinih ma-snih kiselina u zrelom semenu dobijenog gajenjem biljaka pri visokim temperaturama su imale povećan sadržaj palmitinske (16:0), stearinske (18:0), oleinske (18:1) kiseline i smanjen sadržaj linolne (18:2) i lino-lenske (18:3) kiseline u odnosu na seme dobijeno od biljaka gajenih na nižim temperaturama (Kirnak et al., 2008).
Visoke temperature tokom nalivanja semena soje u kontrolisanim uslovima smanjuju klijavost i vigor, ali efekat visokih temperatura u po-lju do sada nije bio određivan. Seme dve sorte soje (Hutcherson, grupe

120
Semenarstvo
zrenja V i DP4690, grupe zrenja IV) je proizvedeno na poljima Kenta-kija, Misisipija, Arkanzasa i Teksasa od 2000. godine do 2002. godine. Merena je temperatura vazduha od perioda nalivanja zrna do fiziološ-ke zrelosti. Mahune su ručno brane, odstranjeno je smežurano seme i atipično seme, pre ustanovljavanja klijavosti i izvođenja testa ubrzanog starenja. Srednja temperatura tokom nalivanja zrna se kretala od 24,0oC (Kentaki) do 37,6oC (Teksas). Standardna klijavost i klijavost pri izvo-đenju testa ubrzanog starenja su značajno opadali kako je srednja tem-peratura tokom nalivanja semena rasla. Standardna klijavost je opadala linearno (r2=0,49) od blizu 100% na 24oC na 85% na 36oC, dok je opa-danje vezano za test ubrzanog starenja bilo u obliku krive (R2=0,86), a klijavost se svela na 11% na 36oC (sl.37) (Egli et al., 2005).
Postoje genetičke razlike između sorti u pogledu reagovanja na stre-sne uslove i održavanja kvaliteta semena u takvim uslovima, ali su ra-zlike male u odnosu na efekte koje stres izaziva uopšte gledano. Očite razlike u reagovanju na stresne uslove suše konstatovane su kod različi-tih inbred linija kukuruza. Dok su jedne dobro podnosile sušu i održale prinose na odrđenom nivou uz dobar kvalitet semena, kod drugih je došlo do značajnog pada i prinosa i kvaliteta semena. U testu stabilnosti 18 inbred linija na održavanje kvaliteta semena, preko hladnog testa, konstatovano je da polovina varijabilnosti u pogledu kvaliteta semena
Slika 37. Odnos između standardne klijavosit ili klijavosti dobijene testom ubrzanog starenja i srednje dnevne temperature tokom nalivanja zrna soje.
Figure 37. Relationship between standard germination or accelerated-aging germination and mean daily maximum temperature during soybean seed filling.
(Egli et al., 2005)

121
Opšte semenarstvo
pripada genetičkoj kompoziciji inbred linija kukuruza, a druga polovina uticaju spoljašnjih činilaca.
Efekti stresa na razvoj semena, prinos i komponente prinosa uglav-nom su poznati i posvećena im je veća pažnja nego osobinama kvaliteta semena kao što su klijavost i vigor.
Navodnjavanje tokom kasnijih faza formiranja mahuna i ranih faza nalivanja zrna ima najveći uticaj na prinos, ali nema neki značajniji uti-caj na kvalitet semena (Kadham et al., 1985). Jaka suša u tom periodu razvoja soje može da umanji prinos 32–42% ali ima veoma mali efekat na klijavost semena i vigor.
Uprkos brojnim istraživanjima, nije ustanovljena čvrsta veza između mase semena i vigora. Neki drugi činioci koji mogu indirektno da utiču na smanjenje energije klijanja i mase semena mogu dovesti do zabune. Pokorica u vreme nicanja soje može značajno da umanji energiju klija-nja kao i smanjenje veličine kotiledona i njihove mase. Seme veće mase podložnije je mehaničkim oštećenjima tokom mehanizovane berbe i dr.
Ekstremni činioci stresa koji dovode do značajnog umanjenja mase semena, njegove naboranosti i deformacije, može da ima negativan uti-caj na vigor, ali ne i na klijavost semena. Međutim mala masa seme-na nastala usled stresnih uslova u toku njegovog formiranja i nalivanja mogu da se odraze negativno na proces klijanja (Dornbos and McDo-nald, 1986).
Stresni uslovi mogu da dovedu do velikih promena u fiziološkim procesima majčinske biljke i razvoja semena i tako direktno ili indirek-tno utiču na prinos i kvalitet semena. Te promene se ogledaju u nedo-voljnoj količini asimilata potrebnih za nakupljanje rezervnih materija u semenu. Sadržaj proteina, masti i oligosaharida naglo opada, a i ne odlažu se u semenu u dovoljnoj količini.
Nedostatak u hranjljivim elementima u toku vegetacije može tako-đe da dovede do umanjenja kvaliteta semena. Sadržaj kalcijuma je u po-zitivnoj korelaciji sa kvalitetom semena – njegovom klijavosti, dok fos-for, cink i gvožđe pokazuju negativnu korelaciju u odnosu na procenat klijavosti. Primarni efekat nedostatka kalcijuma u ćelijama biljke ogleda se u narušavanju strukture membrana (Hecht-Buchholz, 1979). Sadržaj natrijuma u semenu je u direktnoj vezi sa otpornošću biljaka na bolesti (Chrittenden and Svec, 1974).
Način obrade zemljišta igra jednu od veoma važnih uloga u posti-zanju visokih prinosa, dobrog i kvalitetnog semena. Njime mogu da se

122
Semenarstvo
izbegnu ili umanje uslovi stresa. Uobičajeni klimatski kriterijumi koje bi trebalo da prate proizvođači semena su: količina toplotnih jedinica to-kom fiziološkog dozrevanja, dužina dana, učestalost ekstremno visokih temperatura, mogućnost navodnjavanja i količina vode koja se navod-njavanjem daje biljci.
Postoje razlike u uzgoju biljaka za merkantilnu i semensku proi-zvodnju. Proizvođači semena moraju da vode više računa o napred na-vedenim činiocima. Zbog toga se preporučuje semenska proizvodnja u regionima gde je rizik od stresnih uslova sveden na minimum i gde je prilagođen način obrade zemljišta u cilju dobijanja kvalitetnijeg seme-na. To podrazumeva izbor najboljeg načina obrade, izbor sorte, optima-lan datum setve, obezbeđenje visokog nivoa đubrenja, kontrolu korova, bolesti i insekata i smanjenje činilaca koji dovode do stresa pogodnim merama.
Seme je potrebno požnjeti što pre nakon fiziološke zrelosti. Rana žetva umanjuje kvalitet semena. Kasnije izvedena žetva, u povoljnim uslovima spoljne sredine ne utiče na kvalitet semena direktno, ali može doći do osipanja semena ili do mehaničkih povreda zbog sniženog sa-držaja vlage. Seme kukuruza se žanje sa 30–35% vlage, a seme soje sa 15-17%. Značajnija mehanička oštećenja se događaju već kod sadržaja vlage od 12% i niže kod većine semena ratarskih biljnih vrsta.
Uticaj ekoloških činilaca na osobine semena
Mnogo se zna o fiziološkim i biološkim procesima koji se dešavaju u semenu. Učinjeno je više pokušaja da se ovi procesi povežu sa ekološkim uslovima. Složenost ekoloških činilaca je u tome što oni imaju višestruki uticaj na pokazatelje kvalitetnih svojstava semena. Postoji interakcija i povratni mehanizam pojedinih činilaca. Svako kompleksno fiziološko, fizičko ili biohemijsko objašnjenje razvoja semena od oplodnje do nje-govog klijanja je nemoguće. Svetlost, temperatura, voda, patogeni mi-kroorganizmi, razne hemijske materije, i dr. imaju uticaja na formiranje semena, određuju faze mirovanja semena, klijavost, porast biljke i dr.
Klimatski činioci najviše utiču na procenat klijavosti semena, dok na brzinu klijanja utiču hranljive materije. Strategija za uspešnu semen-sku proizvodnju bi bila da se proizvođači drže optimalnog roka setve i

123
Opšte semenarstvo
obezbede dovoljne količine azota, fosfora i kalijuma, kao i mikroeleme-nata u proizvodnji pšenice (Sharma and Anderson, 2003)
Temperatura i vlažnost
Temperatura i vlažnost vazduha i zemljišta su dva ekološka činio-ca koja najčešće utiču na formiranje i kvaliteta semena. Najznačajnija posledica vodnog stresa je umanjenje rasta biljaka zbog umanjene fo-tosinteze. Smanjenje lisne površine automatski utiče na smanjenje pri-nosa semena (Pearson, 2003). Kvalitet semena soje je veoma umanjen ukoliko postoji deficit vode u vreme formiranja mahuna. Smanjenje nastaje zbog abortivnosti mladih mahuna. Osim što se smanjuje broj semena, umanjuje se i krupnoća semena. Smanjena akumulacija suve materije u semenu u uslovima nedostatka vlage u zemljištu je rezultat iz prevremenog gubljenja lisne površine i skraćenog perioda nalivanja semena. Ukoliko se biljke gaje uz prisustvo viška vode prinosi su takođe smanjeni. U ovakvim uslovima se povećava procenat ulja u semenu, a opada sadržaj proteina. Ukoliko je biljka u toku 15–30 dana posle po-četka cvetanja imala suviše vode u zemljištu, klijavost semena može da se smanji i do 50%.
Ouda i sar. (2010) su sačinili model po kome je moguće izračuna-ti potrebnu količinu vode koja se dodaje biljci soje u sušnim uslovima kako bi se ostvarili dobri prinosi. Zemljišna vlaga je određena pre na-vodnjavanja kao bi se mogla izračunati potrebna količina vode dodata navodnjavanjem da bi se izračunao poljski vodni kapacitet zemljišta. Potrebna količina vode je izračunata po formuli:
CU = (_2 - _1) * Bd * ERZ (1)
gde je: CU = potrebna količina vode (mm), _2 = procenat vlage u ze-mljištu posle navodnjavanja, _1 = procenat vlage u zemljištu pre navod-njavanja, Bd = gustina biljaka (zemljišta) (g/cm3), ERZ = efektivna zona korena.
Najveća zavisnost između količine padavina, temperature i osobina semena je u vremenu sazrevanja i žetve. Visoka temperatura i relativ-na vlažnost vazduha tokom sazrevanja semena i neposredno pred žetvu

124
Semenarstvo
nepovoljno utiču na kvalitet semena. Nepovoljan uticaj naizmeničnog povećanja i smanjenja sadržaja vlage u semenu soje, dok se seme nalazi u mahuni, je još pojačan visokim temperaturama. Rane sorte soje često sazrevaju dok su temperature visoke, a relativna vlažnost dovoljno vi-soka da izazove jaka oštećenja semena. Iz tog razloga kasnostasne sorte daju uglavnom seme boljeg kvaliteta.
Grašak je biljna vrsta koja se odlikuje vrlo neujednačenim sazreva-njem, posebno genotipovi sa intermedijarnim rastom stabla. Zbog toga je žetva proteinskog graška praćena značajnim gubicima i oštećenjima semena. Gubici i oštećenja najmanji su ako se žetva obavi blagovreme-no, pri optimalnoj vlažnosti semena. Međutim, ne postoji jedinstveno mišljenje o optimalnoj vlažnosti za žetvu s obzirom na velike razlike između genotipova proteinskog graška u morfološkoj građi biljaka, tipu porasta stabla, ranostasnosti, uniformnosti sazrevanja semena, obliku i krupnoći semena, građi semenjače (Milošević, 2009).
Problemi odložene, zakasnele žetve i povećane vlage semena prote-inskog graška dovode do:
– poleganja biljaka,– žetva se sporije obavlja,– dolazi do pojave kasnih korova, koji otežavaju žetvu,– heder prilikom podizanja biljaka zahvata više nečistoća,– smanjuje se kvalitet semena.Smanjenje kvaliteta semena ogleda se u sledećem:– semeni omotač gubi boju, najviše u delu izloženom suncu,– dolazi do značajnih mehaničkih oštećenja,– smanjuje se životna sposobnost semena (vigor),– graškov žižak napušta seme pre skladištenja, odnosno fumigacije
(NSW, 2005).Suvi vetrovi, nedostatak vlage i visoka temperatura u prvoj fazi nali-
vanja semena suncokreta povećavaju procenat šturih semena i uzrok su niskom procentu ulja. Isti uslovi u drugoj fazi nalivanja semena dovode do smanjenja apsolutne mase semena, ali ne utiču značajnije na sadržaj ulja. Optimalni uslovi za formiranje semena suncokreta i postizanje vi-sokog sadržaja ulja su vlažno zemljište i srednje dnevne temperature od 20oC do 25oC, kao i umerena vlažnost vazduha. Pri ovakvim uslovima se odvija dalje nakupljanje suve materije, što obezbeđuje veliku masu 1000 semena (Ćupina i Sakač, 1988).

125
Opšte semenarstvo
Uticaj vodnog stresa, koji podrazumeva ili suviše veliku, ili nedo-voljnu količinu vode, utiče na razvoj i sazrevanje semena različitim in-tenzitetom, u zavisnosti od vremena u kome se dešava. Uopšteno gleda-no, razvoj semena je brži ukoliko se vodni stres ispolji. Ukoliko se uzmu kao primer žita, formiranje semena i prinos semena zavise od tri ključna momenta (Slatyer, 1969):
1) začetak i razvoj cveta čime je određen potencijalni broj semena,2) oplodnja koja određuje broj semena,3) nalivanje semena tokom koga seme povećava masu.Prateći rast i razvoj cvasti, evidentno je da manji nedostatak vode
smanjuje broj cvetnih primordija kod pšenice, a verovatno i kod drugih žita broj primordija više zavisi od vodnog stresa nego od razvoja posto-jećih primordija. Ako je vodni stres osrednji i relativno kratak (do 4 ne-delje) brzina formiranja primordija je mnogo veća nego u optimalnim uslovima, a broj primordija nije poremećen. Ako je stres jače izražen ili produžen ukupan broj klasića može biti značajno smanjen.
Kod semena pšenice postoji negativna zavisnost između količine padavina i kvaliteta semena u vreme njegovog formiranja od vlatanja do voštane zrelosti. Velika količina vlage u tom periodu dovodi do smanje-nja sadržaja proteina. Ukoliko je visoka temperatura vazduha, a nizak sadržaj vode u zemljištu u periodu nalivanja semena, pojačava se proces disanja, dolazi do utroška ugljenih hidrata što rezultira povišenim sadr-žajem belančevina u semenu. Povišen sadržaj belančevina se objašnjava i time što je u uslovima suše u vreme nalivanja semena smanjeno pre-meštanje ugljenih hidrata iz vegetativnih u generativne organe (Miloše-vić i Malešević, 2004).
U aridnim i semiaridnim regionima, kao što je Vojvodina, za ozimu pšenicu su najpovoljniji uslovi za dobijanje visokokvalitetnog semena u godinama kada prolećna vegetacija počinje s dovoljno rezervi vlage u zemljištu u sloju od 1 m (140–150 mm vode). U periodu od vlatanja do klasanja bi trebalo da vlada toplo vreme i da ima dovoljno padavina, a da formiranje semena protiče pri temperaturi od 25oC do 30oC i pri umerenom nedostatku vlage. U godinama kada se u proleće u zemljištu nakupi dovoljno nitrata, biljke se dobro razvijaju i intenzivno koriste azot. U periodu klasanja tkivo biljke sadrže i dovoljno azotnih materija, tako da su u mogućnosti da formiraju seme optimalne apsolutne mase (Jeftić, 1992).

126
Semenarstvo
Svetlost
Biljke soje koje ranije cvetaju tokom kratkog dana daju manju koli-činu semena jer je proizvedeno manje fotosintata. Prema istraživanjima urađenim u SAD u 2009. godini formiranje prinosa soje Glycine max (L.) Merr. u poljskim uslovima zavisi od asimilacionog kapaciteta maj-činske biljke tokom vegetacije (High Beam Research, 2009). Kod sunco-kreta i drugih ratarskih biljaka nakupljanje asimilata je najintenzivnije u vreme nalivanja semena. Količina akumuliranih asimilata određena je nešto ranije u periodu formiranja ćelija u klicinim listićima i endos-permu. Što se formira više ovakvih ćelija u fazi razvića kotiledona, to se akumulira više ulja u fazi nalivanja semena. Zbog toga period od cveta-nja do završetka deljenja ćelija u semenu predstavlja kritičan period, jer se uspostavlja ravnoteža između broja i krupnoće semena.
Sorte soje različito reaguju na dužinu dana, koja uslovljava broj dana do cvetanja, period zrenja, masu semena. Konstatovano je da su sorte sa dugom vegetacijom osetljivije na dužinu dana (Norman, 1978). Wallace i Yan (1998) su izveli ogled sa osam sorti pasulja sa ciljem da je vreme cvetanja uslovljeno fotoperiodima. Imali su četiri fotoperioda (9,12,14 i 16h), četiri noćne temperature (18, 21, 24 i 27oC) i dnevne temperatu-re koje su bile za 3 i 6oC više u odnosu na noćne. Sorte su podeljene u dve grupe i to kao ranostasne i kasnostasne. Konstatovao je da se sorte mogu podeliti na dve grupe u odnosu na njihovo reagovanje na fotope-riod i temperaturu i to one kod kojih se dešavaju intenzivne promene koje su se ispoljavale u značajnom odlaganju vremena cvetanja, kako se dužina dana skraćivala ili produžavala.
Dobar primer uticaja fotoperiodizma je postignut kod Chenopodi-um album. Seme biljaka izloženih dugom danu imalo je nižu klijavost od onih sa biljaka koje su bile izložene kratkom danu. Fotoperiod utiče i na razvoj semenog omotača. Seme luka koje je bilo izloženo kratkom danu imalo je propustljivu semenjaču, dok ono koje je bilo izloženo delovanju dugog dana nije. Kod semena krastavca čiji je plod izlagan delovanju crvenog svetla klijavost je bila 92%, dok je izlaganje dugom crvenom svetlu bila sa 23–28% klijavosti.
Gutterman (1982) je pokazao da je zahtev za svetlošću povezan sa klijavošću semena kod mnogih biljnih vrsta. U vreme formiranja seme-na materinsko tkivo okružuje semeni zametak. Ako strukture semena kao što su zid plodnika, brakteje, ostaju zelene sve vreme sazrevanja se-mena zahtev za svetlošću će biti formiran pre nego što se seme požanje.

127
Opšte semenarstvo
Ovo je zbog toga što je fitohrom u semenu vezan u neaktivnom Pr obli-ku, a svetlosni stimulans je potreban da bi ga vratio u aktivan Pfr oblik koji omogućava da se klijanje semena nastavi. Raspored fitohroma u raznim delovima ponika prikazan je na slici 38 (http://plantphys.info/plant_physiology/phytochrome.shtml).
Slika 38. Koncentracija fitohroma varira u zavisnosti od dela biljkeFigure 38. Concentration of phytochrome varies within a plant ( http://plantphys.info/plant_physiology/phytochrome.shtml)
Ako tkivo semena izgubi hlorofil pre nego što je seme potpuno zre-lo, fitohrom u semenu će biti u aktivnom Pfr obliku, tako da će ono klijati u mraku. Cresswell i Grime (1981) su pokazali da je sposobnost semena da klija u mraku u obrnutoj srazmeri povezana sa količinom hlorofila u ispitivanim delovima nekih vrsta semena u vreme zrenja (sl. 39).
Kvalitet svetlosti koje biljka apsorbuje u vreme nakupljanja fotosin-tata je veoma važan. List koji je direktno izložen delovanju crvene sve-tlosti može normalno da obavlja fotosintezu. Međutim, lisna površina koja se nalazi na nižim etažama je zasenjena, pa je odnos crvene i du-gotalasne crvene svetlosti umanjen. Klijanje mnogih semena je podsta-knuto prisustvom crvene svetlosti, dok dalekocrvena svetlost inhibira klijanje semena jer transformiše fitohrom u neaktivan oblik koji spreča-va klijanje (Garden Guide, 2010). Koning (1994) je izveo eksperiment sa naklijavanjem semena salate pod uticajem crvene i dalekocrvene svetlo-sti. U odnosu na kontrolu koja je naklijavana u mraku, najbolji rezultat je dobijen kada je seme naklijavano pod uticajem svetlosti crvenog dela
pojava epikotila, rast lista, kompaktan rast zelenog stabla/ epicotyl uncoiling leaf expansion, greening stem compact growth
kotiledoni dobijaju zelenu boju imobilizacija hraniva/cotyledon greening nutrient mobilization
Koncentracija fitohroma varira u biljci/ The concentration of phytochrome varies within a plant
Koncentracija fitohroma/ Phytochrome Concentration
negativni foto-tropski rast/negative phototropic growth

128
Semenarstvo
spektra (sl. 40). U ogledu koji je izveo Fenner (1987) ustanovljeno je da su sve biljke koje zahtevaju svetlost za klijanje bile inhibirane svetlom koje je prodiralo kroz lisnu masu.
Slika 39. Uticaj količine hlorofila na klijavost semena na svetlu i u mraku Figure 39. Influence of chlorophyll amount on seed germination on light and dark
(Cresswell and Grime,1981)
Klij
avos
t u m
raku
(% o
d kl
ijavo
sti n
a sv
etlo
sti)
Ger
min
atio
n in
the
dark
(% th
an g
erm
inat
ion
in th
e lig
ht)
Količine hlorofila u ispitivanoj građi (mg/g suve materije) Amount of chlorophyll (mg/g dry matter)
100
80
60
40
20
0 0.0 0.6 1.2 1.8 2.4 3.0
He Ae
Hm
Le
Sd
Sn
Pl
St
LhBs
AoAhHp
Sde
MaDp Me
TpSs
DmSp
Ae – Arrheanaterum elatius, Ah – Arabis hirsutaAo – Autoxanthum odoratum,Dm – Draba muralisDp – Digitalis purpureaHe – Helianthemum
chamaecitusHm – Hordeum murinumHp – Hypericum perforatumLc – Lotus corniculatus
Ma – Myosotis arvensisMe – Millium effusumPl – Plantago lanceolataSd – Silence diodica Sde – Sieglingia decumbensSp – Succisa pratensisSs – Seneccio squalidusSt – Serratula tinctoriaTp – Tragopogon pratensisZh – Zeontodon hispidus
Slika 40. Uticaj crvenog i dalekocrvenog dela spektra na
klijavost semena salate Figure 40. Influence of red and farred part of light spectar on
lettuce seed germination(http://plantphys.info/plant_
physiology/phytochrome.shtml)
Crvena/RedKontrola u mraku/
Dark control
Daleko crvena/Far-red
Daleko crvena crvena/Far-red red
Crvena daleko crvena/Red far-red

129
Opšte semenarstvo
Ekološki značaj mirovanja semena
Mirovanje semena može se definisati kao privremeno odlaganje vid-ljivog klijanja bilo koje biljne vrste koja sadrži meristem (Lang ,1987). Mirovanje semena je genetički uslovljeno, a pod snažnim je uticajem ra-zličitih uslova spoljne sredine. Ukoliko je seme u stanju mirovanja, ono ne klija pod uticajem brojnih uslova spoljne sredine. Mirovanje semena može biti indukovano ili uklonjeno brojnim činiocima spoljne sredine, a seme koje nije u stanju mirovanja takođe klija pod uticajem različitih uticaja spoljne sredine. Činioci spoljašnje sredine mogu da izmene sta-nje mirovanja i seme koje je u stanju mirovanja prevednu u oblik koji može da klija (Graeber et al., 2009) (sl.41).
Slika 41. Uticaj spoljašnjih činilaca na klijanje semenaFigure 41. Influence of environmental conditions on seed germination
(http://farm4.static.flickr.com/3038/2869593563_ba0b187064.jpg)
Seme može da se rasejava u prostoru (vetar, insekti i dr.) u vremenu (mirovanje). One vrste semena koje se zadržavaju u zemljištu se poste-peno razvijaju tokom određenog perioda vremena, jer umesto drugog staništa one očekuju da dođe do promena nekih od spoljnih činilaca, koje će im dati dodatni podstrek za otpočinjanje procesa klijanja. Dva najvažnija činoca za prekidanje stanja mirovanja su temperatura i vlaž-nost (Merritt et al., 2007).
Seme ne klija pod uticajem bilo kog činioca spoljne sredine/Seeds do not germinate under any set of environmental conditions
Dormantno/Dormant
Spoljašnji uslovi dovode do gubitka dormantnosti/
Environmental conditions prom
ote dormancy lossSp
olja
šnji
uslo
vi in
duku
ju d
orm
antn
ost/
Envi
ronm
enta
l con
ditio
ns in
duce
dor
man
cy
Privremeno dormantno/Conditionally dormant
Nije dormantno/Nondormant
Klijanje semena pod uticajem velikog broja spoljnih činilaca/ Seeds germinate under a wide range of environmental conditions
Privremeno dormantno/Conditionally dormant

130
Semenarstvo
Slika 42. Uticaj temperatura na klijavost semena koje je u stanju mirovanjaFigure 42. Temperature influence on dormant seed germination(http://farm4.static.flickr.com/3144/2867107023_3822b2212f.jpg)
Seme koje se nalazi u zemljištu, a u stanju je mirovanja delom klija
pod uticajem nekih od činilaca spoljne sredine, uglavnom temperature, a jedan deo semena propada zbog starenja ili zbog nepovoljnih uslova u zemljištu kao što su niska temperatura, neodgovarajuća vlažnost, pri-sustvo predatora i dr. (sl. 42). Zemljište šire gledano može da se prikaže kao „banka gena” u prirodnim uslovima.
Biljne vrste za koje je karakteristično da im se seme zadržava u ze-mljištu u stanju mirovanja su korovi. Ono što naglašava mirovanje se-mena je vlažno zemljište. Hladna zemljišta su karakteristična po tome što ne održavaju seme u stanju mirovanja. U tundrama i subarktičkim oblastima nije zabeleženo mirovanje semena. Tropski predeli su veo-ma slični umerenim i u tim uslovima je mirovanje semena izraženo (Warwick, 1984).
Postojanje prirodnog ciklusa klijanja semena ukazuje na to da je seme koje miruje u zemljištu u stanju neprekidnih fizioloških promena, koje obezbeđuju da njihovo mirovanje bude uvek odgovarajuće za odre-đene vremenske uslove.
Temperatura zemljišta/Soil temperature
Raspoređenost semena/Seed dispersal
Posle zrenja/After ripeningRaslojavanje uslovljeno toplotom/Warm stratification
Raslojavanje uslovljeno niskom temperaturom/Cold stratification
Stimulatori klijanja/Germination stimulans
Klijanje/Germination
Vlažnost zemljišta/Soil moisture
Dormantno seme/Seed dormancy
Leto/SummeLeto/Summer Jesen/Autumn Zima/Winter Proleće/Spring

131
Opšte semenarstvo
AGROTEHNIČKE MERE U PROIZVODNJI SEMENA
Agrotehnika, unutar ekologije obuhvata sve pokazatelje koji se od-nose na uzgajanje biljnih vrsta, u svrhu postizanja što viših i sigurnijih prinosa. Agrotehničkim merama se smanjuju negativni i pojačavaju po-zitivni uticaji pojedinih ekoloških činilaca.
Smatra se da u suštini nema razlika u tehnologiji – agrotehnici pro-izvodnje semenskih i komercijalnih useva. U savremenom semenarstvu se takvo shvatanje ne može prihvatiti kao ispravno, što je i ranije istica-no (Ćirović, 1990). Razlozi za to su mnogobrojni, a najvažniji su:
– što postoji veliki broj sorti različitih bioloških osobina, koji zahte-vaju „sortnu agrotehniku”,
– što pojedine agrotehničke mere imaju neposredan uticaj na pri-nos i kvalitet semena čija se životna sposobnost i biološke osobine moraju sačuvati,
– što je proizvodnja semena rizičan posao, koji podleže zakonskoj regulativi, pa pri projektovanju tehnologije proizvodnje treba sve elemente tako postaviti da se rizik proizvodnje svede na mini-mum, a ekonomski efekti naglase.
Za proizvodnju semena potrebno je sve agrotehničke mere sprove-sti uz najveću pažnju kako bi se dobili naveći mogući prinosi što kvali-tetnijeg semena. Mere koje se primenjuju su:
– plodored i plodosmena,– osnovna obrada zemljišta,– predsetvena priprema,– setva (semenom iz organske proizvodnje, ili netretiranim semenom),– đubrenje (u organskoj proizvodnji stajskim đubrivom),– mere nege, – specifične mere nege semenskih useva, – žetva.Ovde će biti obrađene samo neke od mera koje imaju najveći uticaj
na semensku proizvodnju.

132
Semenarstvo
Setva semenskih useva
Setvu semena treba obaviti u dobro pripremljeno zemljište, koje po-seduje sitno-mrvičastu strukturu, radi lakšeg i ujednačenijeg nicanja se-mena. Prethodnom usevu se poklanja velika pažnja, kako se ne bi došlo u situaciju da su prethodni usev i usev koji se zasniva imali zajedničke patogene, koji bi bili potencijalna opasnost za semenski usev.
Setva semena se obavlja kada temperature zemljišta dostigne odgo-varajuću optimalnu vrednost. Kako temperatura zemljišta utiče na kli-javost semena prikazano je na slici 43. Broj dana potrebnih za klijanje se značajno smanjivao kako je temperatura zemljišta rasla, a povećavao se procenat klijalih semena.
Slika 43. Uticaj temperature zemljišta na klijanje semenaFigure 43. Influence of soil temperature on seed germination.
(http://www.bing.com/images/search?q=seed+++germination+of+dormant+seed+in+soil+images++&view=detail&id=FB7E7CF5DEA1991B5DBB9883D5F9949BB0154C3C&first=61&FORM=IDFRIR&qpvt=seed+++germination+of+dormant+seed+in+soil
+images)
Za svaku vrstu semena postoji optimalna temperatura zemljišta, kada će klijati najveći broj semena u najkraćem vremenskom period. Posebno je seme povrća osetljivo na temperature zemljišta prilikom ni-canja (tab. 23). Brzo i ujednačeno nicanje je od velikog značaja za us-postavljanje optimalnog sklopa biljaka, što u krajnjem utiče na visinu prinosa i kvalitet semena.
Dan
a po
sle k
lijan
ja/
Day
s to
emer
genc
e
Opadanje od/ Decling from
Procenat klijavosti/Percent germination
Optimalna temperatura/ Optimum Temperature
Porast od/ Increasing from

133
Opšte semenarstvo
Tabe
la. 2
3. P
roce
nat t
ipič
nih
poni
ka d
obije
nih
pri r
azlič
itim
tem
pera
tura
ma
Tabl
e 23.
Per
cent
age o
f nor
mal
vege
tabl
e see
dlin
gs p
rodu
ced
at d
iffer
ent t
empe
ratu
res*
**(h
ttp://
ww
w.bi
ng.co
m/im
ages
/sea
rch?
q=se
ed++
+ger
min
atio
n+of
+dor
man
t+se
ed+i
n+so
il+im
ages
++&
view
=det
ail&
id=F
B7E7
CF5D
EA19
91B5
DBB
9883
D5F
9949
BB01
54C3
C&fir
st=61
&FO
RM=I
DFR
IR&
qpvt
=see
d+++
germ
inat
ion+
of+d
orm
ant+
seed
+in+
soil+
imag
es++
)
Biljn
a vr
sta/
Plan
t spe
cies
0ºC
6ºC
10ºC
12ºC
17ºC
22ºC
26ºC
31ºC
36ºC
Asp
arag
us/A
spar
agus
0 0
61(
53)
80(
24)
88(
15)
95(
10)
79(
12)
37(
19)
0Pa
sulj/
Bean
s 0
0 1
52(
31)
82(
18)
90(
7) 8
8(7)
2 0
Repa
/Bee
ts 0
53(
42)
72(
17)
88(
10)
90(
6) 9
7(5)
89(
5) 3
5(5)
0Ku
pus/
Cab
bage
0 2
7 7
8(15
) 9
3(9)
0(6
) 9
9(5)
0(4
) 0
0M
rkva
/ Car
rots
0 4
8(51
) 9
3(17
) 9
5(10
) 9
6(7)
96(
6) 9
5(6)
74(
9) 0
Kar
fiol/C
aulifl
ower
0 0
58(
20)
60(
10)
0(6
) 6
3(5)
45(
5) 0
0C
eler
/Cel
ery
0 7
2(41
) 7
0(16
) 4
0(12
) 9
7(7)
65
0 0
0K
rast
avac
/Cuc
umbe
r 0
0 0
95(
13)
99(
6) 9
9(4)
99(
3) 9
9(3)
49
Plav
i pat
lidža
n/Eg
gpla
nt 0
0 0
0 2
1(13
) 5
3(8)
60(
5) 0
0Sa
lata
/Let
tuce
98(
49)
98(
15)
98(
7) 9
9(4)
99(
3) 9
9(2)
12(
3) 0
0D
inja
/Mus
kmel
on 0
0 0
0 3
8(8)
94(
4) 9
0(3)
0 0
Luk/
Oni
ons
90(
136)
98(
31)
98(
13)
98(
7) 9
9(5)
97(
4) 9
1(4)
73(
13)
2Pe
ršun
/Par
sley
0 0
63(
29)
0(1
7) 6
9(14
) 6
4(13
) 5
0(12
) 0
0Pa
štrn
ak/P
arsn
ips
82(
172)
87(
57)
79(
27)
85(
19)
89(
14)
77(
15)
51(
32)
1 0
Gra
šak/
Peas
0 8
9(36
) 9
4(14
) 9
3(9)
93(
8) 9
4(6)
86(
6) 0
0Pa
prik
a/Pe
pper
s 0
0 1
70(
25)
96(
13)
98(
8) 9
5(8)
70(
9) 0
Rotk
vica
/Rad
ish 0
42(
29)
76(
11)
97(
6) 9
5(4)
97(
4) 9
5(3)
0 0
Span
ać/S
pina
ch 8
3(63
) 9
6(23
) 9
1(12
) 8
2(7)
52(
6) 2
8(5)
32(
6) 0
0Ku
kuru
z šeć
erac
/Sw
eet C
orn
0 0
47(
22)
97(
12)
97(
7) 9
8(4)
91(
4) 8
8(3)
10
Para
dajz
/Tom
atoe
s 0
0 8
2(43
) 9
8(14
) 9
8(8)
97(
6) 8
3(6)
46(
9) 0
Lube
nica
/Wat
erm
elon
0 0
0 1
7 9
4(12
) 9
0(5)
92(
4) 9
6(3)
0
* Ene
rgija
klij
anja
(u d
anim
a) N
umbe
rs in
day
s to
seed
ling
emer
genc
e.
** C
rven
i bro
jevi
=opt
imal
ne d
nevn
e te
mep
erat
ure
zem
ljišt
a za
mak
simal
an b
roj p
onik
a u
najk
raće
m v
rmen
u**
Num
ber i
n re
d =
optim
al d
aytim
e so
il te
mpe
ratu
re fo
r max
imum
pro
duct
ion
in th
e sh
orte
st ti
me.

134
Semenarstvo
Specifične mere nege semenskih useva
Ovde će biti naznačene specifične mere nege, jer je njihova primena vezana za semensku proizvodnju. Takođe će biti reči i o žetvi jer ona najviše utiče na kvalitet dobijenog semena.
Sortno plevljenje i uklanjanje metlica
Ove se mere mogu smatrati biološko-agrotehničkim, a svrha im je obezbeđenje genetičke čistoće i identiteta sorte, odnosno obezbeđenje ukrštanja odabranih roditeljskih parova i dobijanje genetički dobrog semena određenog hibrida (mora se sprečiti nekontrolisana oplodnja).
Kod semenskih useva strnih žita, sortnim plevljenjem se uklanjaju primese drugih vrsta, drugih sorti i atipične biljke sorte čije se seme proizvodi. Ova se mera mora obavezno obavljati i to u više navrata (oba-vezno četiri puta), u onim fazama rasta i razvića kada se osobine osnov-ne sorte najbolje uočavaju (pre klasanja, u punom klasanju, u voštanoj zrelosti i u punoj zrelosti). Posebno treba ukazati na potrebu svesnog, kvalitetnog i stručnog obavljanja sortnog plevljenja (primese se čupaju sa korenom, obavezno iznose sa parcele). Ova se mera ne bi smela izo-staviti, a to se nekada čini zbog smanjenja troškova proizvodnje i pove-ćanja finansijskih efekata u proizvodnji semena. Jasno je da u takvim slučajevima dolazi do smanjenja biološko produktivnih osobina sorte čije se seme proizvodi.
Kod semenskih useva kukuruza neophodno je ukloniti samonikle i štrčeće biljke kod roditelja u onim fazama rasta i razvića kada se one mogu uočiti, a ukoliko je potrebno to se čini u nekoliko navrata. Među-tim, mnogo je važnije i rizičnije pitanje obezbeđenja ukrštanja roditelj-skih komponenti. To se može postići mehaničkim uklanjanjem metlica na semenskom roditelju pre oprašivanja (ručno ili kombinacijom ruč-no-mašinski).
Mehaničko uklanjanje metlica treba obavljati savesno, blagovreme-no i kvalitetno. Ako se uklanjanje metlica čini ručno onda se čupaju cele metlice pre nego što izađu iz lisnog rukavca. Priroda samooplodnih linija, koje ponekad metliče i preko 20 dana ili praše polen pre nego što metlice izađu iz lisnog rukavca, zahteva da se ovaj posao obavlja u ne-koliko navrata (2–3 puta).

135
Opšte semenarstvo
Mašinsko uklanjanje metlica povećava ekonomičnost rada u odno-su na ručno. Međutim, upotreba mašina zahteva visok stepen unifor-mnosti majčinske komponente, veoma preciznu setvu i gotovo idealno ravnu površinu parcele. Osim toga, kod korišćenja mašina uvek je po-trebna i korekcija ručnim radom, jer se mašinom ne mogu ukloniti sve metlice. Analize Veličkovića i Budčarovskog (1987) pokazale su da je pri mašinskom uklanjanju metlica (čupačem tipa „Castri”) kod hibrida BC 6625 uklonjeno 86%, a kod hibrida BC 6661 samo 78% metlica. Ćirović (1990) navodi da je opravdan rad mašinama ako se ukloni 75% metlica.
Ako se pri ručnom radu zbog nepažnje uklone pored metlice i vršni listovi, umanjuje se prinos semena za 5–14%, a kod nekih inbred linija i do 25% (tab. 24). Pri mašinskom radu redovno se sa metlicom uklanjaju i vršni listovi. Prema nalazima Veličkovića i Budčarovskog (1987) u pro-seku se ukloni od 24 do 26% listova, zavisno od hibrida.
Tabela 24. Efekat uklanjanja metlica i vršnih listova kod majčinskih inbred linija na prinos hibridnog semena kukuruza
Table 24. Effect of elimination tasells and top lives from femail inbred lines on seed yield (Veličković i Budčarovski, 1987)
Majčinksa inbred linija/Mother
inbred line
Relativan odnos (kontorla = 100%)Relative ratio (control = 100%)
Metlica+1 listTasell+1 lief
Metlica+2 listaTasell+2 liefs
Metlica+3 listaTasell+3 liefs
Metlica+4 listaTasell+4 liefs
88/2 99 99 91* 86*
1982/III NS 94 88 77** 69**
65/IVNS 96 91* 84** 74**
B 37 E 99 94 90** 87**
B 73 100 98 93* 90**
796 NS 93* 87** 77** 74**
Prosek*/Average 97 93 85 80
*,** Značajno za P=0,05 i P= 0,01 * ,**Significant for P=0,05 i P= 0,01
Dopunsko oprašivanje
Dopunsko oprašivanje je biološka mera kojom se poboljšava pro-cenat oplođenih cvetova što dovodi do formiranja većeg broja semena. Ova mera se danas izvodi i kada su povoljni vremenski uslovi. Pose-ban efekat ove dopunske oplodnje se pokazao kod suncokreta, gde se ono preporučuje u sistemu proizvodnje semena. Za poboljšanje efekta

136
Semenarstvo
oprašivanja koriste se pčele. Košnica sa pčelama se postavlja pored use-va suncokreta, kako bi u toku cvetanja prenele polen sa cveta na cvet. Dokazano je da pojedine pčele mogu da prenesu 100–200 mg polena odjednom, što iznosi polovinu njihove ukupne mase. Jedna pčela može da prenese 250.000 do 6.000.000 polenovih zrna, jedan deo, mehanički (nogama), a veći deo kroz digestivni trakt. Već posle 120 minuta od konzumiranja polena pčela izbacuje polenova zrna na sledeći cvet (sl. 44). Uspešnost dopunskog oprašivanja pčelama zavisi od temperature i svetlosti. Što su temperature više količina sakupljenog polena je veća. U toku vedrog dana pčele mnogo intenzivnije sakupljaju polen nego kada je vreme oblačno. Razni drugi insekti prenošenjem polena povećavaju oplodnju, a time i formiranje semena.
Prenošenje polena na biljke može da se izvrši na više načina. Prema podacima Morto-na i saradnika (2009), polinatori na povrću su uglavnom insekti, vetar i samooplodnja (tab. 25). Intenzivna upotreba pesticida smanjila je značajno broj polinatora, tako da se danas rade brojni projekti kojima bi se iznašao način pove-ćanja njihove brojnosti. Pored toga nepovoljni vremenski uslovi mogu biti uzrok nedovoljne oplodnje (visoka temperatura u vreme oplod-nje, kiša, vetar i dr.). U praksi se primenjuje do-punsko, mehaničko oprašivanje. Kod kukuruza polen se ručno stresa sa metlice jedne biljke na svilu druge ili se polen prikuplja pa se smeša polena nanosi na svilu. Kod useva suncokreta može da se trlja glava o glavu. Nekada je do-
punsko oprašivanje potrebno i kod strnih žita. To se postiže zatezanjem kanapa iznad useva, a radnici zatezanjem konopca istresaju polen. Ova mera, ako se primenjuje, primenjuje se kod krmnog bilja, ali u poslednje vreme retko.
Ukoliko se neka od ovih bioloških mera preduzima, važno je da se obavi u određeno vreme kada su prašnici zreli i pucaju. U toku dana dopunsko oprašivanje najpovoljnije je posle rose, a primenjuje se kod proizvodnje osnovnog semena.
Slika 44. Pčela kao oprašivač Figure 44. Bee as a polinator
(http://www.flickr.com/photos/autanex/519742656)

137
Opšte semenarstvo
Tabe
la 2
5. M
ehan
izm
i opr
ašiv
anja
uob
ičaj
en k
od se
men
a po
vrća
Tabl
e 25.
Pol
linat
ing
mec
hani
sms a
nd sy
stem
s in
com
mon
veg
etab
le cr
ops (
Mor
ton,
200
9)
Biljn
a vr
sta
Cro
p C
omm
on N
ame
Latin
ski n
aziv
Cro
p Sp
ecie
s
Meh
aniz
am o
praš
ivan
jaPr
imar
y Po
llina
ting
Mec
hani
sm(s
)
Sist
e op
raši
vanj
aPo
llina
ting
syst
em
Ukr
štaj
u se
sa d
ivlji
m
srod
nici
ma
u SA
DW
ild C
ross
able
Spe
cies
in
US
Luk
/Oni
on
Alliu
m ce
paIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
N
Beli
luk/
Gar
lic
Alliu
m sa
tivum
Inse
kti/i
nsec
tsVe
ćina
je st
erila
na/
Mos
t are
ster
ileN
Cel
er/ C
eler
yAp
ium
gra
veol
ens
Inse
kti/i
nsec
tsU
kršt
anje
/cro
ssY
Repa
/Bee
tBe
ta v
ulga
ris
Veta
r/w
ind
Ukr
štan
je/C
ross
Y
Repa
/Sw
iss C
hard
Beta
vul
gari
sVe
tar/
Win
dU
kršt
anje
/Cro
ssY
Slač
ica/
Mus
tard
Bras
sica
junc
eaIn
sekt
i/ in
sect
sU
kršt
anje
/Cro
ssY
Kelj/
Kal
eBr
assic
a na
pus
Inse
kti/i
nsec
tsU
kršt
anje
/Cro
ssY
Brok
oli/B
rocc
oli
Bras
sica
oler
acea
Inse
kti/I
nsec
tsU
kršt
anje
/Cro
ssN
Kelj
pupč
ar/B
russ
els S
prou
tsBr
assic
a ol
erac
eaIn
sekt
i/ins
ects
Ukr
štan
je/C
ross
N
Kupu
s/C
abba
geBr
assic
a ol
erac
eaIn
sekt
i/ins
ects
Ukr
štan
je/C
ross
N
Kar
fiol/C
aulifl
ower
Bras
sica
oler
acea
Inse
kti/i
nsec
tsU
kršt
anje
/cro
ssN
Kelj/
Kal
eBr
assic
a ol
erac
eaIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
N
Kin
eski
kup
us/C
hine
se C
abba
geBr
assic
a ra
paIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
N
Kin
eska
slać
ica/
Mus
tard
, Chi
nese
Bras
sica
rapa
Inse
kti/i
nsec
tsU
kršt
anje
/cro
ssY
Repa
uga
rnja
ča/T
urni
pBr
assic
a ra
paIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
Y

138
Semenarstvo
Papr
ika
/Pep
per
Caps
icum
ann
uum
Sam
oopl
odnj
a/se
lfSa
moo
pplo
nja
self
#3N
Endi
vija
/Esc
arol
e/ E
ndiv
eCi
chor
ium
endi
via
Sam
oopl
odnj
a/Se
lfSa
moo
plon
jase
lf #2
N
Lube
nica
/Wat
erm
elon
Citr
ullu
s lan
atus
Inse
kti/I
nsec
tsU
kršt
anje
/cro
ssN
Korij
ande
r/C
ilant
roCo
riand
rum
sativ
umIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
N
Din
ja/C
anta
loup
e M
elon
Cucu
mis
melo
Inse
kti/I
nsec
tsU
kršt
anje
/cro
ssN
Din
ja/H
oney
dew
Mel
onCu
cum
is m
eloIn
sekt
i/ins
ects
Ukr
štan
je/c
ross
N
Mus
katn
a tik
va/M
usk
Mel
onCu
cum
is m
eloIn
sekt
i/Ins
ects
Ukr
štan
je/c
ross
N
Kra
stav
ac/C
ucum
ber
Cucu
mis
sativ
usIn
sekt
i/ins
ects
Ukr
štan
je/C
ross
N
Bund
eva/
Pum
pkin
Cucu
rbita
pep
oIn
sekt
i/Ins
ects
Ukr
štan
je/C
ross
YZi
msk
a tik
va i
bund
eva
Win
ter S
quas
h an
d Sn
ow p
umpk
ins
Cucu
rbita
max
ima
Inse
kti/i
nsec
tsU
kršt
anje
/cro
ssY
Zim
ska
mus
katn
a tik
vaW
inte
r Squ
ash
Cucu
rbita
mos
hata
Inse
kti/I
nsec
tsU
kršt
anje
/Cro
ssY
Letn
ja ti
kva
Sum
mer
Squ
ash
and
Fall
Squa
sh
Cucu
rbita
pep
oIn
sekt
i/Ins
ects
Ukr
štan
je/C
ross
Y
Mrk
ava/
Car
rot
Dau
cus c
arot
aIn
sekt
i/Ins
ects
Ukr
štan
je/C
ross
Y
Kom
orač
/Fen
nel
Foen
iculu
mvu
lgar
eIn
sekt
i/Ins
ects
Ukr
štan
je/C
ross
N
Sala
ta/L
ettu
ceLa
ctuc
a sa
tiva
Sam
oopl
odnj
a/Se
lfSa
moo
plod
nja
self
#1Y
Para
dajz
/Tom
ato
Sola
num
escu
lentu
mSa
moo
plod
nja/
Self
Sam
oopl
odnj
ase
lf #1
/#2
N
Bosil
jak/
Basil
Ocim
um b
asili
cum
Inse
kti/I
nsec
tsU
kršt
anje
/cro
ssN
Pašt
rnak
/Par
sley
Petro
selin
ium
crisp
umIn
sekt
i/Ins
ects
Ukr
štan
je/c
ross
N

139
Opšte semenarstvo
Pasu
lj/C
omm
on B
ean
Phas
eolu
s vul
gari
sSa
moo
plod
nja/
Self
Sam
oopl
odnj
ase
lf #1
N
Gra
šak/
Pea
Pisu
m sa
tivum
Sam
oopl
odnj
a/se
lfSa
moo
plod
nja
self
#1N
Rotk
vica
/Rad
ishRa
phan
us sa
tivus
Inse
kti/i
nsec
tsU
kršt
anje
/cro
ssY
Plav
i pal
idža
n/Eg
gpla
ntSo
lanu
m m
elong
ena
Sam
oopl
odnj
a/se
lfSa
moo
plod
nja
self
#2N
Span
ać/S
pina
chSp
inac
ea o
lerac
eaVe
tar/
win
dU
kršt
anje
/cro
ssN
Bob/
Faba
Bea
nVi
cia fa
baSe
lfSa
moo
plod
nja
self
#2N
Crn
i pas
ulj,
Vig
na/C
owpe
aVi
gna
ungu
icula
taSe
lfSa
moo
plod
nja
self
#2N
Kuku
ruz/
Cor
nZe
a m
ays
Veta
r/W
ind
Ukr
štan
je/c
ross
N
Y-da
N –
ne
Self
#1: s
amoo
plod
ne b
iljke
,stra
noop
lodn
ja je
obi
čno
< 1%
Self
#2: s
amoo
plod
ne b
iljke
,stra
noop
lodn
ja je
obi
čno
oko
2–5%
Self
#3: s
amoo
plod
ne b
iljke
,stra
noop
lodn
ja či
ji st
epen
ukr
štan
ja >
5%
Proc
enju
je s
e da
uku
pna
vred
nost
opr
ašiv
anja
u a
grar
u ko
je o
bave
pče
le iz
nosi
2–8
mili
jard
i dol
ara
godi
šnje
(1,
3–5,
2 m
ilija
rdi
evra
), št
o je
uje
dno
i pot
enci
jaln
a št
eta
od iz
umira
nja
pčel
a. Z
ato
pom
or p
čela
izaz
van
upot
rebo
m n
ekih
pes
ticid
a i d
rugi
m č
inio
cim
a ni
je št
eta
sam
o za
pče
lare
, neg
o za
celu
dru
štve
nu za
jedn
icu
(Ben
jam
in, 2
008)
.
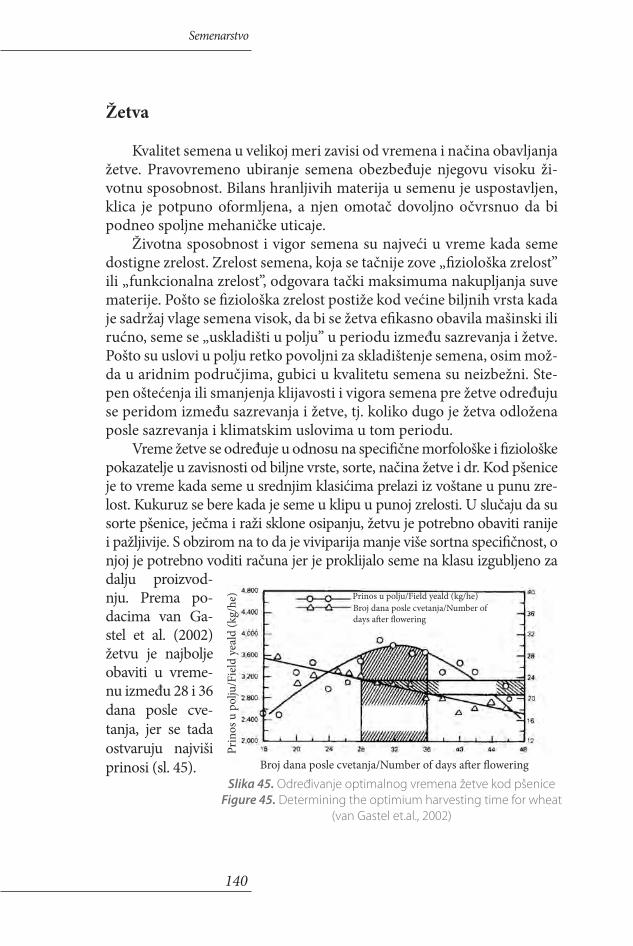
140
Semenarstvo
Žetva
Kvalitet semena u velikoj meri zavisi od vremena i načina obavljanja žetve. Pravovremeno ubiranje semena obezbeđuje njegovu visoku ži-votnu sposobnost. Bilans hranljivih materija u semenu je uspostavljen, klica je potpuno oformljena, a njen omotač dovoljno očvrsnuo da bi podneo spoljne mehaničke uticaje.
Životna sposobnost i vigor semena su najveći u vreme kada seme dostigne zrelost. Zrelost semena, koja se tačnije zove „fiziološka zrelost” ili „funkcionalna zrelost”, odgovara tački maksimuma nakupljanja suve materije. Pošto se fiziološka zrelost postiže kod većine biljnih vrsta kada je sadržaj vlage semena visok, da bi se žetva efikasno obavila mašinski ili rućno, seme se „uskladišti u polju” u periodu između sazrevanja i žetve. Pošto su uslovi u polju retko povoljni za skladištenje semena, osim mož-da u aridnim područjima, gubici u kvalitetu semena su neizbežni. Ste-pen oštećenja ili smanjenja klijavosti i vigora semena pre žetve određuju se peridom između sazrevanja i žetve, tj. koliko dugo je žetva odložena posle sazrevanja i klimatskim uslovima u tom periodu.
Vreme žetve se određuje u odnosu na specifične morfološke i fiziološke pokazatelje u zavisnosti od biljne vrste, sorte, načina žetve i dr. Kod pšenice je to vreme kada seme u srednjim klasićima prelazi iz voštane u punu zre-lost. Kukuruz se bere kada je seme u klipu u punoj zrelosti. U slučaju da su sorte pšenice, ječma i raži sklone osipanju, žetvu je potrebno obaviti ranije i pažljivije. S obzirom na to da je viviparija manje više sortna specifičnost, o njoj je potrebno voditi računa jer je proklijalo seme na klasu izgubljeno za dalju proizvod-nju. Prema po-dacima van Ga- stel et al. (2002) žetvu je najbolje obaviti u vreme-nu između 28 i 36 dana posle cve-tanja, jer se tada ostvaruju najviši prinosi (sl. 45).
Slika 45. Određivanje optimalnog vremena žetve kod pšenice Figure 45. Determining the optimium harvesting time for wheat
(van Gastel et.al., 2002)
Broj dana posle cvetanja/Number of days after flowering
Broj dana posle cvetanja/Number of days after flowering
Prin
os u
pol
ju/F
ield
yea
ld (k
g/he
) Prinos u polju/Field yeald (kg/he)

141
Opšte semenarstvo
Kod šećerne repe sazrevanje semena počinje od osnove rodnih gra-na prema vrhu. Kod određivanja vremena žetve, posebno se mora obra-titi pažnja na osipanje semena, jer se čekanjem da seme sazri na bočnim granama može izgubiti najkvalitetnije seme sa centralnog stabla i se-kundarnih grana. Najsigurnije je momenat žetve odrediti otvaranjem plodova. Pre toga se pregledom celog polja ustanovljava da li je parcela ujednačeno žuta, odnosno da li su biljke svetložute boje. Na centralnom stablu boja semena (ploda) je mrke boje, vrhovi bočnih grana zeleni, stablo je svetlozeleno i sočno. Seme ima tamnomrk omotač, a endos-perm je brašnast. To je momenat prelaska iz mlečno-voštane u punu fiziološku zrelost.
Suncokret se žanje kada mu stablo odr-veni, a jezičasti cvetovi osuše. Žetva soje se obavlja kada stabljika, lišće i mahune dobiju žutomrku boju (sl 46).
Slika 46. Izgled zrele biljke soje spremne za žetvu Figure 46. Mature soybean plant ready for harvesting
(http://www.superstock.com/stock-photography/soybean+plant)
Pomenuti pokazatelji nisu dovoljni, oni jesu informativni, ali bez orijentacionog odre-
đivanja sadržaja vlage u semenu teško da će se bilo koji agronom odluči-ti za određivanje momenta žetve. Ako je sadržaj vlage visok potrebno je dodatno sušenje semena veštačkim putem, što iziskuje velike materijal-ne troškove, a u krajnjem povećava cenu semena. Osim toga u momen-tu postupka sušenja može doći do umanjenja kvaliteta semena. Visoke temperature i brzo gubljenje vlage utiču na životnu sposobnost semena. Dodatna manipulacija semenom izlaže ga mehaničkim oštećenjima, a oštećeno seme je dobra podloga za razvoj štetnih mikroorganizama.
S druge strane pri žetvi semena sa niskim sadržajem vlage dolazi već u prvoj mehanizovanoj radnji do mehaničkih povreda. Svaka do-datna radnja koja podrazumeva uslovno rečeno, pokretanje semena, povećava procenat mehaničkih oštećenja. Već po svojoj prirodi, da bi posedovala namenjeni joj zadatak, semenjača mora da bude elastična. Snižen procenat vlage joj oduzima ovo svojstvo.

142
Semenarstvo
Odlaganje žetve može dovesti do smanjenja kvaliteta semena i zbog uticaja različitih činilaca spoljne sredine, kao što su visoka temperatura, visoka vlažnost vazduha, padavine neposredno pred žetvu, pojava bo-lesti i oštećenja od insekata (Siddique and Wright, 2003). Ako grašak, koji je bio spreman za žetvu pokisne, a zatim se osuši nekoliko puta pre žetve, kvalitet semena će biti umanjen, pre svega zbog niže klijavosti (Mattews, 2008).
Seme soje je izuzetno osetljivo i sklono mehaničkim oštećenjima, prilikom manipulacije s njim, u vreme pre i posle žetve. Najosetljiviji je semeni omotač. Semeni omotač ima ulogu da štiti klicu jer njenim oštećenjem seme gubi klijavost. Mehanička oštećenja su veća ukoliko je sadržaj vlage niži. Mehaničke povrede do kojih dođe u vreme žetve i posle nje utiču na gubitak životne sposobnosti i klijavosti semena kako sveže požnjevenog tako i tokom skladištenja, jer kvalitet semena u toku čuvanja zavisi od njenog početnog kvaliteta. Mehanička oštećenja za-vise u velikoj meri od brzine obrtanja bubnja prilikom žetve soje što je ustanovio Shelar (2008) (sl. 47). Za smanjenje mehaničkih oštećenja je važna brzina kretanja kombajna, a i kvalitet rada hedera. Soja se kom-bajnira kada je sadržaj vlage u semenu nešto ispod 20%, kako bi se sma-njila mehanička oštećenja. Oštećenja mogu biti veoma značajna ukoliko se soja žanje kod sadržaja vlage od 12%.
Slika 47. Opadanje klijavosti oštećenog semena soje kao posledica brzine okre-tanja bubnja prilikom žetve
Figure 47. Effect of thresher speed on demage and germination of soybean seed (http://www.aragriculture.org/crops/soybeans/harvesting.htm)
100
80
60
40
20
0500 600 700 800 900 1000 1100
Procenat oštećenja ili klijanja/Per cent demaged or germination
Oštećeno seme soje/Cracked soybeans
Preporučena brzina
cilindra/Recommen-ded clyinder
speed
Brzina cilindra/Clyinder speed (rpm)
Klijanje/Germination

143
Opšte semenarstvo
Nije seme svih biljnih vrsta podjednako osetljivo na pomenuti po-kazatelj. U izuzetno osetljivu grupu spada seme soje, graška, pasulja. Manje osetljivo je seme žita, sitnozrnih leguminoza, a seme suncokreta je pancirnim slojem zaštićeno od mehaničkih povreda.
Oblik semena takođe igra svoju ulogu kada je reč o mehaničkim oštećenjima. Ako se uporede dve frakcije semena kukuruza pljosnata i okrugla može se konstatovati da je kod okrugle frakcije klica semena više izložena mehaničkom udaru pa samim tim i oštećenjima.
Žetva može da se vrši mehanički i ručno, u zavisnosti od toga koja kategorija semena je u pitanju i koja je količina semenskog useva u pita-nju. Manje parcele baznog semena mogu da se ubiraju ručno. Za ostale se obično koriste kombajni. Kvalitet rada kombajna određuje kvalitet semena, koji podrazumeva čistoću i oštećenja semena, kao i veličina gubitaka. Pošto je pri žetvi semenskih useva potrebna posebna pažnja zbog važnosti materijala o kojem se radi, to se obično posebno pode-šava rad kombajna, bubanj se oblaže gumom ili nekim drugim mekšim materijalom.
Žetva može biti jednofazna ili dvofazna. Najčešće se ona obavlja jednofazno kao što je slučaj kod pšenice, ječma, kukuruza, suncokre-ta, soje. Dvofazni način žetve se primenjuje kod šećerne repe. Kod že-tve šećerne repe prva faza je košenje. Pokošena cvetna stabla se polažu po ostatku stabla da se prosuše, pa se teži tome da se biljke seku što je to moguće više. Vršidba semena se obavlja kada ono ima sadržaj vlage između 18 i 20%. Ovršeno seme je potrebno raširiti po zatvorenim ili otvorenim skladištima i lopatati kako bi se sadržaj vlage umanjio, ukoli-ko se ne raspolaže sušarama za sušenje semena.

144
Semenarstvo
DORADA SEMENA
Primarna dorada semena
Dorada semena je važan činilac koji obuhvata brojne postupke koji su povezani sa manipulacijom semena posle žetve. Nemoguće je uzgaja-ti i žeti useve, a da ne dođe do izvesnih mešavina između useva uprkos brižljivoj nezi koja se usevu poklanja. Ove mešavine mogu da uključe seme korova i drugih useva, različite inertne materije i nerazvijeno seme osnovne biljne vrste.
Primarna dorada semena predstavlja otklanjanje krupnih nečistoća iz semena, grubim prečistačima, koje mogu da budu delovi biljaka, ze-mlja. Delovi biljaka i ostale zelene delove koji mogu da potiču od korova i zemlju treba ukloniti da ne bi odavala vlagu semenu i na taj način je povećavala. U procesu primarne dorade otklanja se komušina sa klipova semenskog kukuruza komušaljkama i dr.
U najširem smislu dorada semena obuhvata sve radnje kojima se seme posle žetve priprema za tržište: vršidba, transport, sušenje, čišćenje od mehaničkih primesa, selektiranje, kalibriranje, tretiranje i pakovanje. U praksi nije uvek moguće slediti utvrđenu šemu za doradu određene vrste semena, zbog toga što seme kao biološki materijal može da ima različit razvoj u zavisnosti od vremenskih uslova, zemljišnih i drugih činilaca tokom vegetacije. Ovo saznanje nameće potrebu za poznava-njem tehnika dorade i morfologije semena. Seme posle dorade mora imati utvrđen nivo kvaliteta za svaku biljnu vrstu, a cilj treba da bude da se unaprede tehnike za postizanje što boljih i ekonomičnijih rezultata u procesu dorade.
Dorada semena podrazumeva primenu različitih tehnoloških procesa koji se obavljaju na osnovu razlika u fizičkim osobinama semena kao što su mehaničke, termičke, optičke, pa čak i akustične (Babić i Babić, 2000). Na slici 48 prikazan je dijagram procesa dora-de suncokreta.

145
Opšte semenarstvo
Slika 48. Šematski prikaz dorade semena suncokretaFigure 48. Scheme of sunflower seed processing (Madsen, Langkilde, 1988)
Fizičke osobine semena Physical properties of the seed
Širina/Width . . . . . . . . . . . . . . . . . . . . . . . 3,5–6,5 mmDebljina/Thickness . . . . . . . . . . . . . . . . . . 2,8–4,9 mmDužina/Length. . . . . . . . . . . . . . . . . . . . . . 8,0–12,3Brzina lebdenja/Floating velocity . . . . . . 6,6–8,4 m/secMasa 1000 semena/Wieght 1000 seeds . 55,8gSpecifična težina/Specific gravity . . . . . . 465,6g/lOblik . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . duguljastiShape . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . obovateTekstura . . . . . . . . . . . . . . . . . . . . . . . . . . . glatkaTexture . . . . . . . . . . . . . . . . . . . . . . . . . . . . smoothBoja . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . crna do tamnosivaColour . . . . . . . . . . . . . . . . . . . . . . . . . . . . . black to dark grey
Druge karakteristike: neke sorte imaju bele linearne prugeOther characteristics: some cultivars with white linear markings
Dijagram protoka (male mašine)/Flow diagram (small scale machines)
Radni uzorakWorking sample
1000 g
7,0
9,0 r8,5 r8,0 r2,4 s
Vazdušni sitasti prečišćivač
Air-screen cleaner
Zubčasti cilindarIndented cylinder
Vazdušni separatorAir separator
r = sita sa okruglim otvorimar = screens with round perforationss = sita sa duguljastim prorezimas = screens with slotted (oblong) perforationsSve dimenzije su u milimetrimaAll dimensions are in millimeters

146
Semenarstvo
Sušenje semena
Sušenje prirodnim putem ili u specijalnim uređajima je najzastu-pljeniji vid očuvanja kvaliteta semena. Osnova ovog postupka je od-stranjivanje vlage iz semena, radi sprečavanja razvoja mikroorganizama i delovanja insekata koji prouzrokuju narušavanje kvaliteta. Osnovna prednost sušenja je u tome što seme može da se čuva u dužem vremen-skom periodu, lakše se transportuje na veća rastojanja i u svakom mo-mentu može da se iznese na tržište, kako bi se ostvarila određena dobit (Babić i Babić, 2000).
Svi poljoprivredni materijali su zbog kapilarno-porozne struktu-re higroskopni, što znači da upijaju ili odaju vlagu okolnom vlažnom vazduhu, u čijem se okruženju nalaze. Intenzitet razmene vodene pare sa okolinom zavisi pre svega od građe i hemijskog sastava materijala. Seme je nehomogenog sastava (skrob, belančevine, masti). Ovi različiti anatomski delovi semena imaju različitu higroskopnost, što izaziva ne-homogen raspored vlage u materijalu. Tako na primer, najveću higro-skopnost poseduje klica, manju omotač, a još manju endosperm. Zbog toga je u sveže ubranom semenu klica najvlažnija (Babić i Babić, 2000).
Visoki sadržaj vlage je najveći pojedinačni razlog gubitka klijavosti semena. Zbog toga što sadržaj vlage predstavlja glavni uzrok gubitka kli-javosti kod semena moraju se poznavati posledice koje potiču od viso-kog sadržaja vlage i kako da se kontrolišu ili odstrane ove nepovoljnosti.
Seme vrlo često, posle žetve ima visok sadržaj vlage. Čak i dovoljno suvo seme u skladištu može da nakupi vlagu do nivoa kada seme nije bezbedno, ukoliko uslovi za čuvanje semena nisu pod kontrolom. Zbog toga se mora poznavati koji su optimalni nivoi vlage, kako ih dostići, a zatim kako da se uskladišti i upakuje seme, da bi održalo te nivoe vlage.
Ako je vlaga u semenu veća od 45 do 60% onda dolazi do prevre-menog klijanja i gubljena kvaliteta semena. Od ovog nivoa vlage pa do 18–20 % razlika je veoma velika, disanje semena je intenzivno kao i razmnožavanje mikroorganizama. Tamo gde postoji malo ili nimalo provetravanja može da dođe do samozagrevanja semena. Ovo sponta-no zagrevanje može da podigne temperaturu dovoljno visoko da uništi klijavost. Sterilnost semena, slaba ispunjenost semena, smanjen prinos kao i prisustvo mikotoksina mogu biti posledica kontaminacije useva i semena gljivom Fusarium graminearum (Argyris et al., 2003).

147
Opšte semenarstvo
Između 14% i 18% vlage može da dođe do razmnožavanja pato-genih gljiva i na taj način oštećenja semena, naročito mehanički ošte-ćenog. Pored toga postoji pojačano disanje semena koje izaziva nagli gubitak vigora i na kraju gubitak klijavosti. Insekti često predstavljaju ozbiljnu opasnost za uskladišteno seme. Kada je sadržaj vlage manji od 8–9 % žišci naročito, a i mnogi drugi insekti, ne mogu da se razvi-jaju (sl. 49)
Tabela 26. Uticaj vlage semena i temperature sušenja na klijavost semena (%) kukuru-za (po hladnom testu)
Table 26. Influence of seed moisture content and temperature of drying on seed corn germination (%) (under cold test) (J.S.Burris,1980)
Samooplodne linije/Self polinated linesA 632 Mo 17
Vlaga u zrnu (%)/
Moisture content (%)
Temperatura sušenja (oC)/
Drying temperature (0C)
Vlaga u vreme berbe (%)/
Moisture content inharvest time (%)
Temperatura sušenja (oC)/
Drying temperature (0C)
50 45 40 35 50 45 40 3552 50 87 96 97 49 7 68 82 8840 78 96 99 98 36 17 61 83 8932 42 87 96 99 34 17 59 75 9026 63 95 99 98 29 61 84 90 9518 99 97 99 100 25 71 89 98 99
L.S.D. 0.05% = 9 i 13 za A632 i Mo 17.
Slika 45. Žitni žižci: a) Sitophilus granarius, b) Sitophilus oryzaeIFigure 45. Wheat weevil: a) Sitophilus granarius, b) Sitophilus oryzaeI
a) b)

148
Semenarstvo
U tabeli 26 prikazan je uticaj vlage i temperature sušenja na klijavost semena kukuruza primenom hladnog testa. Kod visokog sadržaja vlage od 32%, 40% i pri temperaturi sušenja od 50oC i 45oC klijavost semena kod linije A 632 je bila za 49% odnosno 13% niža u odnosu na seme koje je ubrano sa 18% odnosno 26% vlage. Slična zakonomernost je bila i kod linije Mo 17. Uopšteno gledano, što je niži početni sadržaj vlage u semenu i niža temperatura sušenja, procenat klijavosti se povećava.
Kod procesa sušenja je najvažnije da se obezbedi takav način suše-nja, koji će dovesti do najmanjih oštećenja semena. Predviđeni period sušenja zavisi od biljne vrste, i početnog sadržaja vlage u semenu. Po-stoji razlika u predviđanju procesa sušenja kod ratarskih i povrtarskih biljnih vrsta, kao što je prikazano na slici 50.
Slika 50. Predviđanje dužine perioda sušenja kod ratarskih (gore) i povrtarskih (dole) biljnih vrsta
Figure 50. Predicting of period for seed drying of field (above) and vegetables (down) plant species
(http://www.bing.com/images/search?q=seed++drying+images&view=detail&id=C8C6BAE81C2DBAB248242ADFC23409BEDC16F9E7&first=211&FORM=IDFRIR&qp
vt=seed++drying+images)
predviđena/predicted
željena/desired
Predviđen period sušenja/Predicted drying period
razlika/difference: 7.75
12%
5%
Dani sušenja/Day of drying
Sred
nja
vred
nost
sadr
žaja
vla
ge/
Mea
n pe
rcen
tage
moi
stur
e co
nten
t
Sred
nja
vred
nost
sadr
žaja
vla
ge/
Mea
n pe
rcen
tage
moi
stur
e co
nten
t
Vreme (dani)/Time (days)
0 1 2 3 4 5 6 7 8 9 10 11
16
14
12
10
8
6
4
2

149
Opšte semenarstvo
Pod oštećenjima semena može da se podrazumeva gubitak vitalno-sti ili mehanička makro i mikro oštećenja. Vitalnost u tom slučaju gube pojedinačna semena u različito vreme. Procenat preostalog vitalnog se-mena u uzorku je pokazatelj ukupnog oštećenja. Ali gubitak vitalnosti semena ne zavisi samo od vremena. Stepen oštećenja zavisi od sadržaja vlage u semenu i temperature.
Sušenje semena je visoko specijalizovana, tehnička operacija. Si-stem za sušenje mora da bude takav da na zadovoljavajući način deluje u određenim klimatskim uslovima. Za sušenje semena različitih biljnih vrsta koriste se različiti sistemi.
Sistem za sušenje vazduhom najpogodniji je za sušenje semena. Nezagrejani (prirodni) ili zagrejani vazduh se koristi u zavisnosti od klimatskih uslova i količine vlage koja treba da se odstrani iz semena. Uopšteno gledano, nezagrejani vazduh je dovoljan za uklanjanje sadr-žaja vlage od 2 do 4%, osim kada je visoka relativna vlaga, koja smanju-je kapacitet sušenja nezagrejanog vazduha do stepena kada se sušenje produžava. U svim ostalim slučajevima vazduh za sušenje treba da bude zagrejan (Milošević i Malešević, 2004).
Sistem za sušenje pomoću vazduha se sastoji od nekoliko kompo-nenti:
– ventilatora za duvanje vazduha koji usmeravaju vazduh kroz seme i mora da obezbedi traženu količinu vazduha u zavisnosti od vlage se-mena. Vazduh koji prolazi kroz seme, prenosi toplotu na seme pri čemu voda isparava i izbacuje se izvan mase semena;
– grejač obezbeđuje grejnu energiju potrebnu za sušenje čime se povećava kapacitet sušenja vazduha. Grejač može da radi na prirodan gas, naftu, biomasu ili na neki drugi način;
– komore za sušenje, koje su veoma različite u zavisnosti od vrste semena, najčešće su sa perforiranim podom. Zagrejani vazduh prolazi kroz slojeve semena u komori pomoću statičkog pritiska;
– uređaja za kontrolu procesa sušenja koji proces kontrolišu pomo-ću termostata ili kombinacijom termostata i vlagomera da bi se osigura-lo da temperatura vazduha za sušenje ne prelazi 43–45 oC. Postoji i kon-trola koja aktivira različite sigurnosne uređaje da bi opasnost od požara bila svedena na minimum. Za neke biljne vrste (soja) ne preporučuje se viša temperatura sušenja od 38oC (Misra, 1983, Samarah et al., 2009). Viša temperatura značajno smanjuje klijanje jer dolazi do mehaničkih oštećenja i promene ugljenih hidrata.

150
Semenarstvo
Sistem za sušenje je smešten unutar kompleksa koji uključuje pro-storije za doradu i skladištenje. Ovaj položaj je zadovoljavajući pod uslo-vom da seme može da se smesti u sušaru u roku od nekoliko sati.
Bitno je da sušenje počne ubrzo posle žetve. Sušenje je jedna od operacija koja se ne sme odlagati, a kada jednom počne mora da se zavr-ši. To znači neprekidno sušenje, dok se žetva ne završi i seme ne osuši do sadržaja vlage koji je bezbedan za skladištenje. Čak i kod iste biljne vrste, sušenje se mora prilagoditi genotipu. Ako se soja žanje sa mahunama, to je potrebno uraditi u fazi zelenožutih mahuna i sušiti ih na temperaturi vazduha (oko 25oC), ili toplim vazduhom na oko 29oC. U tom slučaju neće doći do mehaničkih oštećenja, a neće se izgubiti ni na vigoru ni na procentu klijavosti. Ovaj eksperiment su izveli Samarah et al., 2009. godine na državnom univerzitetu u Ajovi (Iowa State University).
Precizna organizacija posla je važnija za sušenje od bilo koje druge operacije sa semenom. Kada se sušenje ne uradi kako treba i na vreme, sav trud uložen u umnožavanje semena, proizvodnju i žetvu može da propadne u roku od nekoliko sati.
Dorada semena
Operacije prečišćavanja koje slede posle sušenja ne moraju da se izvode takvom brzinom kao sušenje. Iz tog razloga nije neophodno ni poželjno da kapacitet dorade bude jednak kapacitetu sušenja. Prečišća-vanje može da se izvodi u mnogo dužem vremenskom periodu. Pošto je vreme prečišćavanja duže, potrebno je privremeno skladište za ču-vanje suvog semena, dok ne bude moglo da se doradi i upakuje. Silo ćelije sa aktivnim provetravanjem su dobro rešenje za rinfuzno skladi-štenje semena. Jedinice za sušenje ili provetravanje mogu se periodično uključivati da se dosuši seme ako se vlaga semena povećala zbog visoke atmosferske vlage ili da bi se sprečilo premeštanje vlažnog vazduha i povećanje vlage unutar mase semena do kojih može doći zbog velikih kolebanja temperature.
Posle sušenja, potrebno je izvršiti nekoliko operacija prečišćavanja da bi se seme pripremilo za završno skladištenje i prodaju; krunjenje, prečišćavanje, razvrstavanje po krupnoći, zaštita semena, pakovanje. Kako će se izvesti pomenuti postupci zavisi od vrste semena.

151
Opšte semenarstvo
Krunjenje i vršidba uvek predstavljaju moguće izvore oštećenja se-mena, bez obzira na to da li su urađeni mehanički ili ručno. Zbog toga se mora paziti da se mehanička oštećenja semena, povezana sa ovih po-stupcima, svedu na najmanju moguću meru.
Oprema za prečišćavanje semena se kreće u rasponu od jednostav-nog sita za vejanje – koje se i danas koristi u mnogim delovima sveta – do kompleksa visokospecijalizovane opreme kao što je električna ma-šina za sortiranje semena. Mada se razlikuje po tipu i dizajnu celokupna oprema za doradu semena ima jedan zajednički cilj a to je razdvajanje semena od primesa na osnovu fizičkih svojstva semena.
Osobine semena prema kojima mašine za doradu čiste i razdvajaju seme po osobinama, nazivaju se selektori, koji rade na više principa. Najvažnije osobine za klasiranje semena su brzina padanja ili lebdenja, debljina semena, širina i dužina semena, struktura površine, poroznost, elastičnost, boja i dr.
Brzina pada semena
Za brzinu slobodnog pada semena važi zakon gravitacije. Seme veće specifične mase ima veću brzinu pada. Ako se isto seme pusti više puta sa određene visine ustanoviće se da je isto vreme potrebno za pad se-mena pod uslovom da seme ne menja oblik zbog eventualnih oštećenja. Sa brzinom pada kod tehnike dorade može se naći najveća brzina koju seme može da postigne. Brzina pada se meri u m/s. Ako se seme pšenice pušta kroz vazdušni tunel brzinom od 8 m/s, a brzina vazdušne struje je 8 m/sec seme će se održavati (lebdeti) u struji vazduha. Kod povećanja brzine strujanja vazduha na 8,15 m/s, seme pšenice će padati brzinom od 0,15 m/s. To pokazuje da je brzina padanja jednaka razlici brzine pada semena i brzine protoka vazdušne mase. Na ovom principu rade pneumatski kalibratori.
Brzina padanja tela zavisi ne samo od njegove specifične mase nego i od oblika. Seme oblika lopte pada najbrže, dok ostali oblici sporije pa-daju jer se povećava otpor vazduha. Pored oblika i razni izraštaji na se-menu mogu da uspore brzinu pada. U tabeli 27 su izmerene brzine pada i lebdenja semena, kao i koeficijent trenja istog semena na gumenoj tra-ci. Naznačene su tri brzine kretanja vazduha kada je podignuto prvo
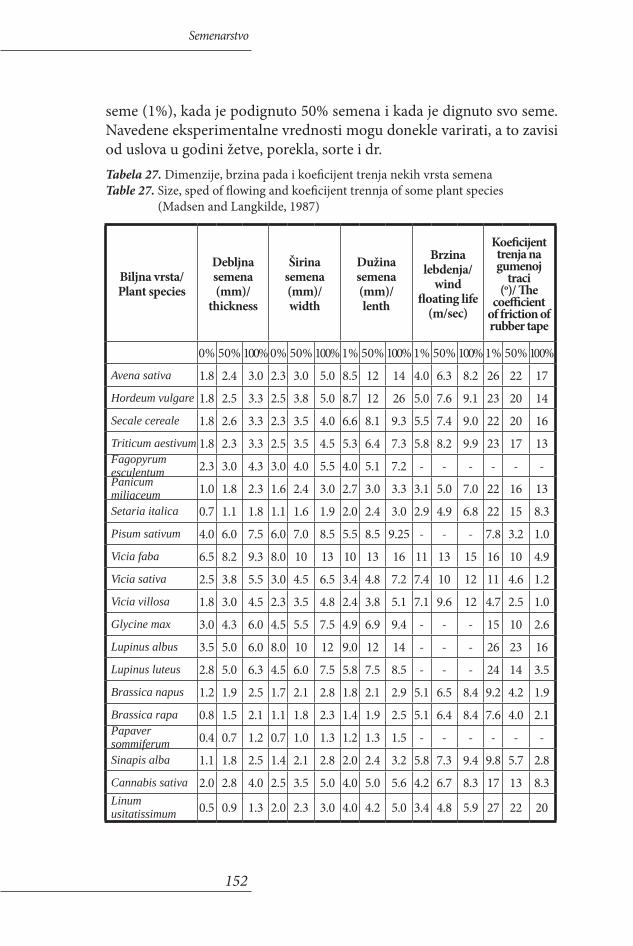
152
Semenarstvo
seme (1%), kada je podignuto 50% semena i kada je dignuto svo seme. Navedene eksperimentalne vrednosti mogu donekle varirati, a to zavisi od uslova u godini žetve, porekla, sorte i dr.Tabela 27. Dimenzije, brzina pada i koeficijent trenja nekih vrsta semenaTable 27. Size, sped of flowing and koeficijent trennja of some plant species
(Madsen and Langkilde, 1987)
Biljna vrsta/Plant species
Debljna semena(mm)/
thickness
Širina semena(mm)/ width
Dužinasemena (mm)/ lenth
Brzina lebdenja/
wind floating life
(m/sec)
Koeficijent trenja na gumenoj
traci(o)/ The
coefficient of friction of rubber tape
0% 50% 100% 0% 50% 100% 1% 50% 100% 1% 50% 100% 1% 50% 100%
Avena sativa 1.8 2.4 3.0 2.3 3.0 5.0 8.5 12 14 4.0 6.3 8.2 26 22 17
Hordeum vulgare 1.8 2.5 3.3 2.5 3.8 5.0 8.7 12 26 5.0 7.6 9.1 23 20 14
Secale cereale 1.8 2.6 3.3 2.3 3.5 4.0 6.6 8.1 9.3 5.5 7.4 9.0 22 20 16
Triticum aestivum 1.8 2.3 3.3 2.5 3.5 4.5 5.3 6.4 7.3 5.8 8.2 9.9 23 17 13Fagopyrum esculentum 2.3 3.0 4.3 3.0 4.0 5.5 4.0 5.1 7.2 - - - - - -Panicum miliaceum 1.0 1.8 2.3 1.6 2.4 3.0 2.7 3.0 3.3 3.1 5.0 7.0 22 16 13
Setaria italica 0.7 1.1 1.8 1.1 1.6 1.9 2.0 2.4 3.0 2.9 4.9 6.8 22 15 8.3
Pisum sativum 4.0 6.0 7.5 6.0 7.0 8.5 5.5 8.5 9.25 - - - 7.8 3.2 1.0
Vicia faba 6.5 8.2 9.3 8.0 10 13 10 13 16 11 13 15 16 10 4.9
Vicia sativa 2.5 3.8 5.5 3.0 4.5 6.5 3.4 4.8 7.2 7.4 10 12 11 4.6 1.2
Vicia villosa 1.8 3.0 4.5 2.3 3.5 4.8 2.4 3.8 5.1 7.1 9.6 12 4.7 2.5 1.0
Glycine max 3.0 4.3 6.0 4.5 5.5 7.5 4.9 6.9 9.4 - - - 15 10 2.6
Lupinus albus 3.5 5.0 6.0 8.0 10 12 9.0 12 14 - - - 26 23 16
Lupinus luteus 2.8 5.0 6.3 4.5 6.0 7.5 5.8 7.5 8.5 - - - 24 14 3.5
Brassica napus 1.2 1.9 2.5 1.7 2.1 2.8 1.8 2.1 2.9 5.1 6.5 8.4 9.2 4.2 1.9
Brassica rapa 0.8 1.5 2.1 1.1 1.8 2.3 1.4 1.9 2.5 5.1 6.4 8.4 7.6 4.0 2.1Papaver sommiferum 0.4 0.7 1.2 0.7 1.0 1.3 1.2 1.3 1.5 - - - - - -
Sinapis alba 1.1 1.8 2.5 1.4 2.1 2.8 2.0 2.4 3.2 5.8 7.3 9.4 9.8 5.7 2.8
Cannabis sativa 2.0 2.8 4.0 2.5 3.5 5.0 4.0 5.0 5.6 4.2 6.7 8.3 17 13 8.3Linum usitatissimum 0.5 0.9 1.3 2.0 2.3 3.0 4.0 4.2 5.0 3.4 4.8 5.9 27 22 20

153
Opšte semenarstvo
Strane primese iz semena se mogu izdvojiti uz pomoć vazduha. Sve one koje imaju sporiju brzinu padanja od semena mogu se izdvojiti. Če-sto šturo seme u partiji semena ima najmanju brzinu pada, tako da ga je lako razdvojiti i tako povećati kvalitet semena.
Debljina semena
Debljina semena različitih biljnih vrsta se određuje pomoću sita, a ne merenjem. Ako je debljina semena Lolium multiflorum 0,5–0,7–1,1 mm, to znači da nijedno seme Lolium multiflorum sa glumama neće proći kroz sito otvora 0,5 mm, polovina semena će proći kroz sito od 0,7 mm, a svo seme će proći kroz sito otvora 1,1 mm. Za doradu semena u praksi to zna-či da je kod čišćenja potrebno uzeti donje sito od 0,5 mm, a gornje od 1,1 mm. Ukoliko se seme želi razdvojiti na sitniju i krupniju frakciju može, se postaviti sito od 0,7 mm (Madsen and Langkilde, 1987).
Širina semena
Seme propada kroz sito sa okruglim otvorima kada nije šire od prečnika otvora. Zbog toga se širina semena raznih vrsta ustanovljava pomoću sita sa okruglim otvorima. Širina semena je najveća dimenzi-ja poprečnog preseka. Seme iste širine može da ima potpuno različite oblike na poprečnom preseku, međutim to neće uticati na to da seme propada kroz otvore na situ sa okruglim otvorima. Duguljasto seme, na primer Lolium perene će morati da se postavi vertikalno da bi prošlo kroz sito sa okruglim otvorima.
Sita sa kvadratnim otvorima ne mogu da razdvajaju tačno seme prema širini. U ovom slučaju oblik semena igra važnu ulogu. Mada po-prečni presek sita sa okruglim otvorima odgovara potpuno dijagonali veličine otvora na situ sa kvadratnim otvorima, mnoge vrste semena ne mogu da prođu kroz njega.
Dužina semena
Prema najdužem i najkraćem semenu u partiji semena se određu-ju sita koja će se koristiti u procesu dorade. Međutim, ne odlučuje samo

154
Semenarstvo
dužina semena o razdvajanju, ukoliko se to vrši u nazubljenom cilindru, gde duže seme ispada iz udubljenja, a kraće ostaje u njima, već i centar sile gravitacije (s). Na primer, seme Bromus inermis i Apera spica-venti je iste dužine. Centar sile gravitacije je za Bromus oko 5 mm, a kod Apera spica-venti samo 1,75 mm, od osnove semena. Kada se seme obe trave stavi u nazubljeni cilindar sa ćelijama od 3,0 mm, seme Apera spica-venti će se podići i odvojiti od Bromus inermis bez osja. Ovo je moguće izvesti u laboratoriji ali kod praktičnog čišćenja semena nije moguće razdvojiti seme u nazubljenom cilindru jer će dugačko osje kod Apera preći ivicu ćelije, tako da će klizajuće seme u cilindru izbaciti seme Apera iz ćelije. Zbog toga prilikom merenja dužine semena treba uzeti u obzir i dodatke kao što su osje, papus i dr.
Koeficijent trenja
Značajan broj tehnoloških operacija u poljoprivredi odvija se na bazi trenja. Pojavom većih skladišta i mehanizovanja operacija, uočena je potreba poznavanja ove fizičke osobine. Kasnije kada je došlo do ra-zvoja njivske mehanizacije ustanovljeno je da i ovde postoji potreba za takvim informacijama (krunjenje kukuruza je tipična operacija koja se bazira na trenju) (Babić i Babić, 2000).
Seme različitog oblika će različitom brzinom klizati preko nagnute trake. Okruglo seme će imati najbrži pad niz traku koja se kreće u su-protnom pravcu u odnosu na pravac kretanja (kotrljanja) semena, dok će ostalo seme padati sporije. Za svaku vrstu semena potrebno je odre-đeno iskošenje trake ili platna. Pljosnato ili dlakavo seme će padati samo na jako iskošenoj ploči. U praksi trenje prilikom kotrljanja je mnogo manje od trenja klizanja. Iz iskustva se zna da trenje kotrljanja iznosi 2% od trenja klizanja.
Trenje tela po određenom predmetu koji se nalazi ispod njega je jednako sili potiska neophodnoj da se to telo pomeri napred preko hori-zontalne površine. U svrhu čišćenja semena moguće je odrediti koefici-jent trenja za razne vrste semena, postepenim smanjenjem ugla iskoše-nja trake koja se kreće prema gore pri određenoj brzini.
Koeficijent trenja semena ne zavisi samo od njegovog oblika, građe i krupnoće, nego i od površine po kojoj se seme kreće. Kod čišćenja se-

155
Opšte semenarstvo
mena promena koeficijenta trenja se uspešno koristi. Zbog toga se za či-šćenje semena šećerne repe koristi kanafas, rotirajuća mašina za odvaja-nje Rumex spp. od Trifolium spp. koja ima plišane valjkove, gumeni kaiš je najpogodniji za razdvajanje Matricaria maritima od Phleum pratense, a spiralni separator glatke metalne površine se koriste za razdvajanje Vicia villosa od Secale cereale.
Osobine površine semena
Poznavanje spoljašnje površine semena značajno je kako za gene-tičare, stručnjake ratarskog prifila, tako i za inženjere poljoprivredne tehnike. Intenzitet disanja zrna tokom skladištenja zavisiće i od ukupne površine materijala, a prilikom analize transporta toplote i mase kod procesa zagrevanja ili hlađenja, bez poznavanja ove veličine ne može se doći do merodavnih analiza (Babić i Babić, 2000).
Razlike u strukturi površine su važna osobina razdvajanja. One se koriste kod čišćenja semena elektromagnetnim putem. Dobro razvijeno seme lucerke je potpuno glatko, što je posebno uočljivo pod mikrosko-pom. Površina većine korovskog semena pronađena u semenu lucerke nije glatka. Ono ima mala udubljenja ili pukotine, lepljivo je, granulisano, dla-kavo; kod dugog korovskog semena (Plantago lanceolata) glatka semenjača kada se navlaži vodom postaje sluzavo-lepljiva. Ako se zapraši slabo navla-ženo neočišćeno seme lucerke gvozdenim prahom, prah neće prionuti za glatku površinu semena lucerke, ali će se gvozdeni prah zalepiti za većinu korovskog semena zbog njegove dlakave ili lepljive površine. Ako se neoči-šćeno seme stavi na magnet, seme sa gvozdenim prahom će biti uklonjeno magnetom, dok će se glatko seme lucerke kretati po magnetnom bubnju.
Boja semena
U posebnim slučajevima boja semena može da služi kao osobina za klasifikaciju. „Sortex” delitelj radi na ovom principu. Od svetlog semena moguće je razdvojiti tamno seme ili bolesno seme sa tamnim mraljama (sl. 51). Seme mora da pada i jedno po jedno prođe kroz fotoćelije. Ta-mno seme ili seme sa tamnim mrljama će se odmah napuniti elektro-

156
Semenarstvo
statičkim nabojem i magnetnom silom ukloniti za vreme pada. Na taj način se vrši razdvajanje od svetlog semena. Fotoćelije, obično tri, se stavljaju u prostoriju za razdvajanje u mašinu na taj način da celokupna površina svakog semena bude obuhvaćena fotoćelijom. One se mogu menjati tako da mogu da se adaptiraju prema stepenu obojenosti seme-na koje treba da se razdvaja. U drugom tipu mašine, tamno seme i ono sa tamnim mrljama se oduva još u prostoriji za razdvajanje i na taj način ukloni sa trake za padanje.
Navedeno fizičko svojstvo semena ukazuje na mogući postupak dorade. Češće, međutim, zadovoljavajuće uklanjanje stranog semena ili materijala iz semena zahteva da seme bude dorađeno na više specifič-nih mašina - pri čemu svaka mašina uklanja određeni deo nepoželjnog materijala.
Linije za pečišćavanje semena
U svom najjednostavnijem obliku, sistem za prečišćavanje semena se sastoji od elevatora, koji podiže seme u prijemni koš selektora koji seme prečišćava i drugog elevatora koji nosi čisto seme na dalju doradu. U zavisnosti od karakteristika različitih materijala koje treba ukloniti, ostale mašine za prečišćavanje se dodaju ovom osnovnom sistemu za prečišćavanje da bi se formirala linija za čišćenje ili doradu. U ove spa-daju kalibratori, aspiratori, gravitacioni separatori itd. Svaka mašina je povezana sa onom ispred i iza u liniji dorade pomoću prenosnika, koji omogućavaju da seme teče neprekidno kroz celu liniju. Otuda različite
Slika 51. Tretirano seme mrkveFigure 51. Carrot treted seed

157
Opšte semenarstvo
mašine za čišćenje moraju da imaju dobro podešen kapacitet ili inten-zitet protoka.
Jedna od najčešćih i vrlo dobro urađenih linija je linija za doradu semena kukuruza, koja može da posluži za primer gde je većina načela dorade semena zastupljena. Postupci u toj doradi idu sledećim hrono-loškim redom:
– prijem nedorađenog semena sa krunjača koje se odgovarajućim unutrašnjim transportom usmerava na aspirator;
– unutrašnjim transporterima se daje mogućnost upućivanja seme-na u odgovarajuću silo ćeliju ili na liniju finalne dorade.
Oprema za finalnu doradu semenskog kukuruza se satoji od:– selektora za čišćenje,– trijer cilindara,– kalibratora (potreban broj cilindara za kalibriranje semena kuku-
ruza u 6 frakcija po širini, debljini i dužini zrna),– gravitacionih separatora,– zaprašivača,– automatske vage (sa odgovarajućim prekidačem za merenje 5-50 kg)
sa linijom za uvrećavanje semena.Šematski prikazi dorade semena pšenice, kukuruza, šećerne repe
i suncokreta dati su na slikama od 52 do 55 (Mirić i Brkić, 2002). Pro-ces dorade prikazan na malim laboratorijskim mašinama, mada nema mnogo odstupanja od procesa dorade u velikim doradnim centrima. Na slikama su date osnovne fizičke karakteristike semena koje, u krajnjoj liniji i određuju šemu dorade.
Slika 52. Simptomi infekcije gljivom Fusarium sp. na poniku pšenice Figure 52. Symptoms of infection caused by
Fusarium sp. on wheat seedling (www.plantpath.wisc.edu)

158
Semenarstvo
Za vreme ili posle prečišćavanja može biti poželjno da se određe-na vrsta semena kalibrira po veličini da bi se poboljšao setveni kvalitet. Kalibriranje semena po veličini postaje važnije kada se setva vrši na po-treban sklop i obično se vrši kod kukuruza. Sitno, nezrelo i smežurano seme se delimično uklanja od ostalog semena u toku osnovnog čišćenja.
Specijalne mašine, kalibratori, se posebno koriste za kalibriranje semena po sve tri dimenzije, dužine, širine i debljine, a gravitacioni
Slika 53. Simptomi infekcije gljivom Sclerotinia sclerotiorum: 1) na semenu suncokreta;
2) gljiva izolovana u čistu kulturuFigure 53. Symptoms of infection caused by Sclerotinia sclerotiorum:
1) on sunflower seed; 2) the fungus isolated in pure culture
(foto: Milošević, 2004)
1) 2)
Slika 54. Simptomi infekcije gljivom Fusarium sp:1) na semenu kukuruza;
2) gljiva izolovana u čistu kulturuFigure 54. Symptoms of infection caused by Fusarium sp:
1) on corn seed; 2) the fungus isolated in pure culture
(foto: Milošević, 2004)
1) 2)

159
Opšte semenarstvo
separator za uklanjanje, insektima oštećenog ili nezrelog semena. Gra-vitacioni separatori su posebno efektni za poboljšanje klijavosti i vigora semenskih partija tako što razdvajaju oštećeno, bolesno seme, seme male mase, ali oni zahtevaju veliko radno iskustvo.
Zaštita semena
Seme, pored toga što je nosilac genetičkog potencijala za razvoj biljaka i ostvarenje visokih prinosa, nosilac je i raznih prouzrokovača bolesti i insekata koji se njima hrane. Rani razvoj biljaka, odnosno faza nicanja je najosetljivija faza, kako u odnosu na spoljašnje uslove tako i na pojavu patogena. Zemljište u koje se seme seje može biti kontamini-rano patogenim organizmima koji oštećuju seme ili ponik.
Iz navedenih razloga koncept zaštite semena podrazumeva upotre-bu hemijskih ili bioloških preparata da bi se kontorlisali štetni organzmi u zemljištu i semenu. Štetni organizmi bez obzira odakle dolaze imaju negativne posledice po kvalitet semena.
Korist od tretmana semena su:– povećanje klijavosti, – postizanje uniformnog nicanja biljaka,– zaštita semena i ponika od bolesti i insekata koji se javljaju počet-
kom vegetacije, brže nicanje i rast,– upotreba biljnih hormona rasta doprinosi bržem i boljem rastu
biljaka,– inokulacija bakterijama Rhizobium pospešuje fiksaciju azota
kod leguminoznih biljaka, doprinosi većoj produktivnosti biljke (Rabi, 2007).
Tretiranje semena fungicidima se koristi iz tri razloga:– da kontroliše bolesti semena koji prouzrokuju trulež, propadanje
ponika, trulež korena,– da kontrolišu patogene koji se nalaze na površini semena, kao što
su gari i– da kontrolišu patogene koji se nalaze u semenu kao što je prašna
gar kod žita (sl. 56).

160
Semenarstvo
Slika 56. Razlozi za tretiranje semenaFigure 56. Reasons for seed treatmen
(http://www.ag.ndsu.edu/pubs/plantsci/crops/pp447w.htm)
Hemijska zaštita semena je prvi put primenjena kod žita u svrhu za-štite od gljiva, rđe i gari (Tillet, 1775; Tesier, 1779), a zatim i drugih pato-gena. Od tada do danas fungicidi imaju vodeću ulogu u zaštiti semena.
Ranije je tehnologija aplikacije preparata bila isključivo u vidu praha za suvo tretiranje semena, zatim su u praksu uvedeni vodeni ratsvori praha, kao tečna formulacija (suspenzija). Kod nekih vrsta semena, kao što je po-vrće i šećerna repa korišćeno je peletiranje kako bi se ujednačila krupnoća i oblik semena, a u masu za piliranje uključivali razni hemijski preparati. Oblaganje semena specijalnim filmom sa dodatkom pesticida (inkrustaci-ja) omogućilo je između ostalog i zaštitu semena od mehaničkih oštećenja.
Hronološki gledano u praksu su prvo uvedeni preparati na bazi neselektivnih organiskih materija, uglavnom na bazi žive, zatim organ-skih selektivnih, na kraju preparati sa sistemičnim delovanjem (sl. 57).
Slika 57. Tretirano semeFigure 57. Treted seed
(http://www.agriculture.com/news/technology/bayer-monsto-reach-seed-
treatment-deal_6-ar8941)
Kontrola tretiranja semena/Seed treatment to control
Patogene sa površine semena/Surface Borne Pathogens
Patogeni iz zemljišta/ Soil Borne Pathogens
Patogeno seme (Gar)/Internally Seed Borne
Pathogens (Loose Smut)

161
Opšte semenarstvo
Insekti mogu oštetiti seme u skladištu ili posle setve. Iz tog razlo-ga skladišta bi morala biti bez insekata sve do setve. Za seme koje je posejano postoji opasnost da bude oštećeno insektima koji se nalaze u zemljištu, pogotovo u nepovoljnim uslovima za nicanje, kao što su niske temperature, prisustvo suviška vlage.
Fungicidi su uglavnom specijalizovani za suzbijanje pojedinih pato-genih gljiva na semenu. Stepen zaštite koji fungicid pruža semenu zavisi od doze preparata, spoljašnjih uslova i patogena koji je prisutan. Neki sistemični fungicidi mogu osigurati zaštitu u ranoj fazi razvoja biljke. U fazi nicanja biljka je najosetljivija, kako u odnosu na spoljašnje uslove, tako i na pojavu patogena. Zbog toga je zaštita semena hemijskim ili biloškim sredstvima veoma važna (Rabi, 2007).
Kao nova sredstva za zaštitu semena uvedeni su biološki preparati, koji imaju posebno mesto u organskoj proizvodnji semena. To su pre-parati koji sadrže žive bakterije ili gljive, antagoniste, a primenjuju se za suzbijanje patogena koji se razvijaju u semenu i zemljištu. Prvi preparat, namenjen tržištu, sadržavao je Bacillus subtilis, zatim Pseudomonas ce-pacia i dr. (Schwinn, 1994).
Kod semena sa čvrstim perikarpom tretiranje semena nema velikog efekta. Posledice tretiranja mogu biti negativne ako je perikarp oštećen, jer seme reaguje više na posledice tretiranja, od onoga sa kojim se pažlji-vo rukovalo. Činioci koji utiču na rezultate tretiranja semena uključuju: vrstu semena, starost, sposobnost zadržavanja vode, oštećenje semena, način sušenja i dr. Veoma su važne osobine preparata koji se nanose na seme. Tako na primer dobar fungicid za tretiranje semena treba da bude :
– efikasan protiv bolesti ponika koje ga parazitiraju, – jeftin i lak za primenu,– ne sme biti štetan za seme i kada se ono skladišti,– kompatibilan sa inokulantima na leguminozama,– nisko toksičan za ljude i životinje.Danas se primenjuju nove tehnologije kod oblaganja semena hemij-
skim sredstvima u procesu dorade. Tehnika „Intellicoat” koristi senzor za temperaturu, tako da seme kukuruza može da se seje 3-4 nedelje ra-nije. Kada zemljište dostigne optimalnu teperaturu za nicanje kukuruza, nešto iznad 12,5oC, omotač semena dozvoljava vodi da prodre u seme jer se menja struktura polimera koji je sastavni deo hemijskog omotača semena (Tec Agrinews, 2003) (sl. 58).

162
Semenarstvo
Za nanošenje preparata na seme koriste se razni aparati u zavisnosti od toga koji je sastav preparata, količina semena koju je potrebno zaštiti i drugih parametara (sl. 59). Uopšteno gledano idealan uređaj za tretira-nje semena treba da ima:
– uređaj za doziranje semena,– uređaj za doziranje sredstva koja se nanose na seme,– mogućnost finog nanošenja sredstva na površinu semena,– komoru za mešanje semena i primenjenog preparata.Tretiranje semena se obično vrši za vreme dorade pre pakovanja.
Potrebno je voditi računa o tome da su hemijska sredstva za tretiranje semena obično štetna po ljudsko zdravlje i da se tretirano seme hemij-skim preparatima ne sme koristiti, kao stočna hrana ili u industrijske svrhe. Zbog toga se preporučuje tretiranje samo dela semena koje će se koristiti za setvu, a seme koje se ne koristi odmah, ne treba tretirati.
Slika 58. Korišćenje sredstva „Intellicoat” omogućava setvu kukuruza 3-4 nedelje ranijeFigure 58. „Intellicoat” seed coating lets corn growers plant 3-4 weeks earlier
(http://www.ccimarketing.com/CCINews/Landec/landec2.asp)
Seme hibridnog kukuruza/Hybrid seed corn
Seme/Seed
Barijera omotača/
Coating barrier
Ispod 12,5°C/Below 55°F Iznad 12,5°C/Above 55°F
Izmena omotača/Coating changes
Voda se odvaja u kapi/
Water is repelled
Voda je dostupna za korišćenje/Water is allowed to permeate
Seme upija vodu/Seed absorbs the water
Intellicoat rano oblaganje semena /Intellicoat early
plant coating

163
Opšte semenarstvo
Slika 59. Mašina za tretiranje semena sojeFigure 59. Seed treatment machine for soybean
(http://www.frontiernet.net/~vohsseeds/)
O efektu tretiranja semena postoje brojni podaci. Manje poznati su efekti biološkog tretiranja semena. Rhodes i Powel (1994) su objavili po-datke koji ukazuju na osnovu bioloških preparata u tretiranju semena. U tabeli 28 dat je prikaz trivijalnih naziva preparata, aktivnih supstanci i biljnih vrsta kod kojih se preporučuje upotreba pomenutih preparata. Neki od preparata su univerzalni, a mikroorganizmi od kojih su sačinje-ni su bakterije.Tabela 28. Biološki preparati za tretiranje semena raznih biljnih vrstaTable 28. Biological matters for seed treatment for different plant species
(Rhodes, Powel, 1994)
Aktivna materija (mikroorganizam)
Active matterBiljna vrsta / Plant species
Bacillus subtillis razne vrste / different species
Bacillus subtillis pamuk, kikiriki, pasulj / cotton,peanut,bean
Streptomyces griseoviridis povrće, cveće / vegetable, flower
Trichoderma harzianum razne vrste / different species
Pseudomonas cepacia razne vrste / different species
Agrobacterium radiobacter seme drvenastog bilja / forest species

164
Semenarstvo
Osim što imaju efekat na suzbijanje patogena u i na semenu bio-loški preparati deluju i na parazite i štetočine u zemljištu. U tabeli 29 dat je pregled patogena i štetočina koje biološki preparati mogu da suzbiju kod određenih biljnih vrsta. Kao što se iz navedenih podataka može videti
Tabela 29. Važnije štetočine i bolesti koje se kontrolišu kod semena primenom bio-loških preparata
Table 29. Important pests and diseases which are controled by biological matters (Rhodes and Powel, 1994)
Biljna vrsta /Plant species
Štetočina ili patogen /Pest or disease
Šećerna repa, pšenica, kukuruz, povrće, leguminoze /
Sugar beet, corn,vegetable, forage species
Štetočine u zemljištu, vaši /soil pests, aphids
Žita / Cereals Tilletia caries, Ustilago spp., Pyrenophora spp., Fusarium spp., Septoria spp.
Pamuk / Cotton Pythium spp., Rhizoctonia solaniXanthomonas campestris
Povrće / Vegetable Pythium spp., Rhizoctonia solani
Kukuruz / Corn Pythium, Fusarium spp.
Šećerna repa / Sugar beet Pythium, Aphanomyces spp., Phoma betae
Krompir / Potato Fusarium spp., Rhizoctonia solani
Grašak, pasulj, sojaPea,bean, soybean
Pythium spp., Rhizoctonia solaniPeronospora spp.
biološki preparati suzbijaju na žitima Tilletia caries, Ustilago spp., Pyre-nophora spp., Fusarium spp., Septoria spp., patogene koji inače mogu da limitiraju ukupnu proizvodnju semena. Kod šećerne repe može da se kontroliše parazit Phoma betae i dr.
Biloški preparati sigurno imaju budućnost u savremenom pri-stupu zaštiti semena od štetnih organizama, jer doprinose očuvanju

165
Opšte semenarstvo
biodiverziteta*, koji je poselednjih decenija značajno osiromašen, de-lom zahvaljujući i hemijskoj zaštiti semena (ISF, 2000).
Pakovanje semena
Način pakovanja zavisi od količine semena koje se pakuje, a mo-menat ekonomičnosti je vrlo značajan jer svakako ne bi bilo opravdano seme pšenice u količini od 50 kg pakovati u aluminijske višeslojne folije. Veće količine semena, što se uglavnom odnosi na ratarske biljne vrste se pakuju u papirne ili jutane vreće, dok se kod semena povrća mogu upotrebiti različiti materijali. Za sada je najbolji način sitnog pakova-nja semena povrća u lamelirane aluminijske folije. One su sačinjene od tri sloja. Spoljašnji sloj je od sintetičkog terilena, srednji od aluminijske folije, a unutrašnji od polietilena. Veoma praktičan i uspešan način pa-kovanja semena povrća je u limenkama, mada je taj način prilično skup.
Pri pakovanju se odmeri određena količina čistog semena i pakuje u tekstilne, papirne, plastične vreće ili u neki drugi pomenuti materijal. Vreće se zatvaraju prišivanjem ili lepljenjem. Za veće količine semena, kod ratarskih biljnih vrsta vreće se obično prošivaju, dok se kod lameli-ranih folija primenjuje lepljenje.
Održavanje vitalnosti semena zavisi od ambalaže u koju se seme pakuje, pogotovo ako seme ne ide odmah u prodaju. Osim od vrste am-balaže održavanje najvažnijeg fiziološkog pokazatelja kvaliteta semena, klijavosti, zavisi od temperature i vlažnosti semena što ilustruje tabe-la 30. Seme pirinča sa početnom vlagom od 9,65% je stavljeno u četiri različita pakovanja i čuvano 5 meseci u kontrolisanim uslovima tem-perature od 16oC i relativne vlažnosti od 65%. Tokom perioda čuvanja semena sadržaj vlage u semenu se povisio posebno kod pakovanja u polipropilenskim vrećicama. Procenat klijavosti nije opao posle pet me-seci skladištenja, ali se smanjila životna sposobnost semena, što su po-kazali rezultati testa ubrzanog starenja. Povećanje količine elektrolita pri izvođenju testa električne provodljivosti ukazuje na lošiji vigor semena kukuruza (Abba and Lovato, 1999).
* Biodiverzitet (biološka raznovrsnost) je kompleksan pojam, koji pokriva mnoge aspekte biološkog variranja. Najčešće se reč biodiverzitet koristi da opiše sve vrste organizama koji žive u određenoj oblasti. Gledano sa niova planete biodiverzitet se može definisati kao „život na Zemlji“.

166
Semenarstvo
Tabela 30. Srednje vrednosti sadržaja vlage, klivosti i životne sposobnosti (ustanovlje-nog testom ubrzanog starenja) i vigor testa (ispitivanog električnom por-vodljivošću) na semenu pirinča u četiri različita pakovanja tokom 5 meseci
Table 30. Means seed moisture content, germination, storability (investigated by acce-lerated aging technique) and vigour test (investigated by electrical conduc-tivity) of rice seeds stored in 4 different containers after 5 months storage (Muangkaeo i sar., 2005)
Tip pakovanjaConteriner type
Sadržaj vlage
Moisture content
KlijavostGermination
Životna sposobnost (AA)
klijavost%Storability (AA) germination%
Vigor (električna porvodljivost)
Vigour (Electrical conductivity)
PA PE MPETWP
9,819,839,7910,4
95959695
92939292
35,3835,9235,2935,29
PA – Poliamid / Polyamide PE – Polietilen / Polyethylene MPET – Metalom obložen polietilen terepfalat /Metallized Polyethylene Terepthalate WP – Tkani polipropilen / Woven Polypropylene
Za pakovanje semena bi trebalo koristiti mašine za pakovanje i merenje. One mogu biti relativno jednostavni i jeftini uređaji sa por-tlabl zatvaračima za vreće. Uređaji za pakovanje mogu biti i sistemi s integrisanim mašinama za punjenje, merenje, zatvaranje i prenoše-nje vreća.
Vreće u koje se seme pakuje treba da budu atraktivne, na prednjoj strani obično imaju vidno označenog proizvođača, odnosno semensku kuću njenim prepoznatljivim logom, biljnu vrstu i dr. Na poleđini vreće kao dopunske informacija može da se naznači setvena norma, potreban broj zrna po hektaru i dr.
Kao propratna dokumentacija na vreću se prišiva ili lepi atest koji sadrži podatke o broju partije semena, biljnoj vrsti, neto masi pakova-nja, da li je seme tretirano, ko je dorađivač i dr.
U slućaju da se seme prepakuje, potrebno je da ustanova koja je iz-dala aprobaciono uverenje, odnosno stavila ateste na vreće, da pismeno odobrenje za ovakav postupak.

167
Opšte semenarstvo
Ukoliko se seme izvozi, a izvoz se obavlja u zemlje koje su članice OECD-a, postoje posebna pravila, koja u pogledu pakovanja, podrazu-mevaju obeležavanje vreća atestima određene boje i to za:
– osnovno seme – atesti bele boje,– aprobirano seme prve generacije – atesti plave boje,– aprobirano seme druge i ostalih generacija – atesti crvene boje.Na svakom atestu crvene boje mora biti naznačena generacija
umnožavanja.Dorađeno, upakovano i obeleženo predviđenim atestom (etiketom)
seme se odlaže u skladišta (sl. 60), ili ide direktno u prodaju. Radi si-gurnije manipulacije semenom vreće jedne ili dela partije stavljaju se na palete i obmotavaju polietilenskim omotačem. Omotač štiti seme od mehaničkih oštećenja, usvajanja vlage, predstavlja izvesnu vrstu zaštite od insekata i glodara.
Slika 60. Seme u skladištuFigure 60. Seed in storage

168
Semenarstvo
SKLADIŠTENJE SEMENA
Seme je živi organizam koje zahteva specifične uslove skladištenja kako bi očuvalo kvalitet do setve i omogućilo dobijanje zdrave i vigo-rozne biljke. Visoko kvalitetno seme je preduslov uspešne biljne proi-zvodnje. Kvalitetno seme je neophodno da osigura dobar sklop biljaka, sa predviđenom količinom semena pri setvi, pod određenim uslovima u polju. Kvalitet semena je rezultat zajedničkog efekta činilaca spoljne sredine tokom vegetacije i uslova kojima je seme izloženo tokom žetve i skladištenja. Nepovoljni spoljašnji činioci (temperatura, padavine, re-lativna vlažnost vazduha i zemljišta) tokom vegetacije mogu značajno smajiti klijavost i vigor semena (Egli et al., 2005). Cilj skladištenja je da održi seme u dobrom fizičkom i fiziološkom stanju od vremena žetve do vremena setve.
Skladištenje mase semena je ekosistem kreiran od strane čove-ka, u kome su živi organizmi i neživa priroda (okruženje) u stalnoj interakciji. Najvažniji živi organizam je seme samo po sebi, ali tu su i razne vrste insekata i mikoflora. U neživu okolinu spadaju različiti fizički i hemijski uticaji, kako je to prikazano na slici 65 (Babić i Ba-bić, 2000).
Najbolji način skladištenja semena je onaj koji u najmanjoj meri utiče na promene bioloških osobina semena što se postiže efikasnom regulacijom vlage i temperature. I najosetljivije seme će održati vital-nost na visokom nivou tokom više godina ako se čuva sa niskim sa-držajem vlage i na niskoj temperaturi (tab. 30). Pivski ječam je bio uskladišten na različitim temperaturama sa različitim sadržajem vla-ge. Na temperaturi od 0oC i pri vlažnosti semena od 10% seme može da se čuva 16 godina. Kako su se temperatura čuvanja i sadržaj vlage u semenu povećavali, tako se smanjivalo vreme životne sposobnosti semena (Aastrup et. al., 1989).

169
Opšte semenarstvo
Tabela. 30. Dužina života semena pivarskog ječma u zavisnosti od temperature čuva-nja i sadržaja vlage u semenu
Table 30. The longevity of seeds of malting barley as affected by temperature storage and moisture content in seeds (Aastrup et.al., 1989)
Temperatura Temperature
Sadržaj vlage u semenuSeed moisture content
10% 12% 14% 16% 18%
0°C 16 godina/years
6 godina/ years
2 godine/ years
1 godina/ year
190 dana/days
2°C 14 godina/years
5 godina/years
1.8 godina/ godina/year
315 dana/dana /days
160 dana/dana /days
4°C 11 godina/ years
4 godine/ years
1.5 godina/ year
260 dana/days
130 dana/days
6°C 9 godina/ years
3 godine/ years
1.3 godine/year
210 dana/days
105 dana/days
8°C 7.5 godina/ years
2.5 godina/ years
1 godina/ year
170 dana/ days
89 dana/ days
10°C 6 godina/ years
2 godina/ years
300 dana/days
140 dana /days
70 dana /days
12°C 5 godina/ years
1.6 godina/ year
240 dana /days
110 dana /days
55 dana/ days
14°C 3.8 godina/ years
1.3 godina/ year
190 dana /days
85 dana /days
45 dana /days
16°C 3 godina/ years
1 godina/ year
150 dana /days
65 dana /days
35 dana /days
18°C 2.3 godina/ years
290 dana/days
115 dana /days
50 dana /days
25 dana /days
20°C 1.8 godina/ year
220 dana/days
90 dana /days
40 dana /days
20 dana /days
22°C 1.4 godine/ year
170 dana/days
70 dana /days
30 dana /days
15 dana/ days
24°C 1 godina/year
130 dana/ days
55 dana /days
25 dana /days
12 dana/ days
26°C 290 dana/days
100 dana/days
40 dana /days
18 dana /days
9 dana /days
28°C 210 days 70 dana /days
30 dana /days
13 dana /days
7 dana/ days
30°C 160 days 55 dana /days
22 dana /days
10 dana /days
5 dana /days

170
Semenarstvo
Postoji Harrington-ovo pravilo za sadržaj vlage u semenu koje glasi: za svako sniženje procenta vlage u semenu životni vek semena se duplira (http:/agritech.tnau.ac.in/seed_certification/seed%20Storage%20factors.htm).
Gubitak klijavosti semena, pod povoljnim uslovima skladištenja, za-visi od biologije semena, a do toga može proći i nekoliko godina. Mnoge vrste semena povrća zadržavaju klijavost veću od 50% i posle 10 i više godina čuvanja. Za komercijalnu povrtarsku proizvodnju, navedena lista, koja ukazuje na dužinu skladištenja pojedinih vrsta semena, može poslu-žiti kao vodič:
– 1 godina – kukuruz šećerac, luk, peršun, paštrnak,– 2 godine – repa, paprika, praziluk,– 3 godine – asparagus, pasulj, mrkva, celer, salata, grašak, spanać,
paradajz,– 4 godine – kupus, karfiol, kelj, kelj pupčar, tikvica, bundeva, bli-
tva, rotkvica, – 5 godina – krastavac, endivija, dinja, lubenica (Victory Seed, 2009).Ellis i sar. (2008) su ispitivali kolika je dugoveđnost semena povrća
ukoliko se ono čuva u povoljnim uslovima skladištenja. Različite vrste po-vrća pokazale su i različito vreme održavanja životne sposobnosti semena (tab. 31).Table 31. Minimalni standardi za klijavost u SAD i državi Kolorado i relativna dužina
životne sposobnosti semena povrćaTable 31. Federal and Colorado minimum germination, seed count and relative
longevity of selected vegetable seed (Ellis i sar., 2008)
Vrsta semenaKind of seed
Minimalna klijavost (%)
Minimum germination (percentage)
Prosečan boroj semena po
gramu i unciAverage number
of seed per:
Relatvna životna sposobnost (godine)Relative longevity
(years)Gram Ounce
Asparagus /Asparagus 60 50 1,400 3
Pasulj /Beans 70 4 100 3
Repa / Beets 65 70 2,000 4
Brokoli / Broccoli 75 290 8,100 3Kelj pupčar
Brussels sprouts 70 300 8,500 4
Kupus / Cabbage 75 280 7,700 4
Mrkva / Carrot 55 790 22,000 3

171
Opšte semenarstvo
Kineski kupus Cabbage, Chinese 75 250 7,000 3
Karfiol / Cauliflower 75 310 8,600 4Celer korenaš
Celeriac 55 1,800 50,000 3
Celery / Celer 55 2,700 76,000 3
Cikorija / Chicory 65 710 20,000 4Kukuruz šećerac
Sweet Corn 75 5 140 2
Krastavac Cucumber 80 40 1,100 5
Plavi patlidžanEggplant 60 260 7,200 4
Endivija / Endive 70 610 17,000 5
Kelj / Kale 75 360 10,000 4
Keleraba / Kohlrabi 75 330 9,200 3
Praziluk / Leek 60 350 9,900 2
Salata / Lettuce 80 930 26,000 1Muskatna dinja
Muskmelon 75 40 1,100 5
Luk / Onion 70 300 8,500 1
Peršun / Parsley 60 640 18,000 1
Paštrnak / Parsnip 60 240 6,800 1
Grašak / Pea 80 7 200 3Paprika Pepper 55 160 4,500 2
Bundeva Pumpkin 75 7 200 4
Rotkva / Radish 75 110 3,100 4
Spanać / Spinach 60 100 2,900 3
Spanać, novozelandski Spinach,
New Zealand40 20 430 3
Bundeva / Squash 75 10 300 4
Repa / Turnip 80 500 14,000 4
Lubenica/Watermelon 70 10 300 4

172
Semenarstvo
Prema podacima Basra (2006) čuvanje semena na temperaturama višim od -20oC je veoma dobro za očuvanje početnog kavliteta semena. Razlog tome je usporavanje fizioloških procesa u semenu, koji su uzrok gubljenja kvaliteta semena.
Dokaz da je seme kvalitetno je njegova sposobnost da klija. Is-pitivanje klijavosti semena se uglavnom vrši u optimalnim uslovima u laboratoriji, tako da su razni nepovoljni uslovi koji dovode do sla-bljenja ponika, kojima je seme izloženo u poljskim uslovima, ovde izbegnuti. Razlika između klijavosti semena u laboratoriji i njegove klijavosti u polju je često velika. Uprkos tome, određivanje klijavosti u optimalnim uslovima je obezbedilo osnovni oslonac u većini istra-živanja o starenju semena.
Uticaj temperature od 10oC na seme kenafe tokom skladištenja ispitivali su Meints i Smith (2003). Ispitivana je energija klijanja, kli-javost, vigor i prinos semena. Seme je čuvano četiri godine na ambi-jentalnoj relativnoj vlažnosti. Klijavost je ispitivana pri standardnoj temperaturi (20-30oC) i nižoj temperaturi (20oC). Vigor semena je proveravan s vremena na vreme. Poljska klijavost je bila ista za razli-čite uslove skladištenja tokom četiri godine, ali je bila dirtektno pod uticajem sušnih uslova u vreme kada je seme sejano u polju. Klija-vost semena se održala iznad 80% tokom vremena skladištenja na 10oC, te se moglo komercijalno koristiti. Gubitak elektrolita meren je konduktometrijski i bio je 30-50% niži kod svežeg semena nego kod onog čuvanog četiri godine na 10oC.
Brzina klijanja, koja se smatra indikatorom vigora semena, je oset-ljivija kao ocena oštećenja semena nego gubitak klijavosti. Primećeno je da se rast primarnog korena usporava, čak i pre nego što klijavost počinje da opada (Guy, 1982). U nekim slučajevima je primećeno da je povećanje klijavosti proporcionalno sa vremenom skladištenja semena do određenog vremena, verovatno zbog uticaja endogenih inhibitora klijanja. Takvi efekti su ustanovljeni kod uskladištenog semena trava i leguminoza, kod kojih mirovanje povećava klijavost do izvesnog vreme-na (Harty et al., 1983).

173
Opšte semenarstvo
Promene u razvoju biljke i prinos semena
Atipičnosti u razvoju ponika se obično javljaju posle klijanja semena koje je duže uskladišteno. Istraživanja ukazuju i na to da na morfologiju relativno zrelih biljaka može da utiče vreme čuvanja semena iz kojeg je biljka ponikla. Kod Arabidopsis thaliana, na primer, broj listova koji se javlja na glavnom stablu pre cvetanja se smanjuje ako je biljka ponikla iz duže čuvanog semena. Sejanje oštećenog, duže uskladištenog semena, može čak da utiče i na morfologiju cvetova (Milošević i Malešević, 2004).
Odnos između dužine uskladištenja semena i prinosa je komplek-san. Prvo, postoji razlika uslovljena genetičkim osobinama između vr-sta. Drugo, razlika u vigoru između partija semena je obično mnogo više izražena u nepovoljnim uslovima spoljne sredine. Treće, prinos po biljci se često povećava kod nekih biljnih vrsta, ako zbog dugog čuvanja semena nikne manji broj biljaka jer je povećan životni prostor.
Proučavanja na kukuruzu ukazuju na to da su slabije biljke i sma-njeni prinosi rezultat delimično ostarelog semena, čak i kada su nivoi klijanja relativno visoki. Sejanje starijeg semena kukuruza dovelo je do sporijeg nicanja, biljke su formirale listove manje lisne površine i ma-nji broj visokoproduktivnih biljaka je dobijen u sledećoj generaciji. Kod sirka prinos po biljci opadao je posle primene testa ubrzanog starenja, čak i kada vrednost laboratorijskog klijanja nije značajno opala.
Produženi uticaj skladištenja soje na prinos istraživali su Eđe i Burris (1970). Produktivnost se vrlo malo smanjila kod semena kod koga je vigor bio umanjen. U ispitivanjima sa semenom soje podvrgnute testu ubrzanog starenja je ustanovljeno da je rani razvoj ponika bio sporiji u partijama semena sa nižom energijom klijanja uzrokovanom dužim čuvanjem ali se razlika između atipičnog i tipičnog ponika smanjila ra-zvojem biljaka. Konstatovan je niži prinos kod semena soje čuvanog dve ili tri godine od onog posle žetve.
Vlaga i temperatura kao činioci oštećenja semena
Relativna vlaga vazduha i temperatura su dva spoljna činioca koja vrše najveći uticaj na starenje semena. McCormack (2004) navodi da sadržaj vlage u semenu ima veći uticaj na dugovečnost semena nego temperatura.

174
Semenarstvo
Ukoliko se posmatra uticaj relativne vlažnosti vazduha na sadržaj vlage u semenu može se videti da se povećanjem relativne vlažnosti vazduha povećava sardžaj vlage u semenu (sl. 61). Odnos između relativne vlažno-sti vazduha i semena određuje i vreme u kome se seme može bezbedno skladištiti kao što je prikazano na slici 62. Odgovarajuća kontrola ova dva činioca obezbeđuje uslove za uspešno i sigurno skladištenje.
Slika 61. Povećanje vlažnosti semena u zavi-snosti od relativne vlage vazduha
Figure 61. Decreasing of seed moisture con-tent depending of ear huminity
Slika 62. Bezbedno skladištenje u odnosu vlažnosti semena i vazduha
Figure 62. Safety storage of seed depending of seed and ear moisture
content
Opšte načelo je da što je suvlje i hladnije skladište to seme duže održava klijavost. Seme sa povišenim sadržajem vlage može se oštetiti na niskim temperaturama zbog smrzavanja, što je povezano sa prisu-stvom slobodne vode. Ovaj problem se ne javlja kod suvog semena, čak ni kada temperatura padne ispod 0oC. Međutim, kod niskog sadržaja vlage seme je mnogo osetljivije na mehanička oštećenja.
Evidentno je da različite vrste semena gube vitalnost u različitoj meri pod istim uslovima skladištenja. Tako su Ellis i Roberts (1981) ustanovili da na stalnoj temperaturi od 10oC različite biljne vrste različi-to reaguju zbog toga što imaju različit temperaturni koeficijent (sl. 62). Najniži temperaturni koeficijent ima seme luka, koje se teško čuva, a
25
20
15
10
5
00 20 40 60 80 100
Relativna vlaga (%)/ Relative humidity (percent)
Vla
ga se
men
a (%
)/Se
ed m
oist
ure
(per
cent
)
1. Skladištenje semena nije sigurno/not safe for seed storage2. Kratkoročno skladištenje /short-term storage3. Srednjeročno skladištenje /medium-term storage4. Dugoročno skladištenje /long-term storage
25
20
15
10
5
00 20 40 60 80 100
Relativna vlaga (%)/ Relative humidity (percent)
Vla
ga se
men
a (%
)/Se
ed m
oist
ure
(per
cent
) 1.
2.
3.
4.

175
Opšte semenarstvo
najviši ječam, koji veoma dobro održava klijavost u različitim uslovima.Vlaga i temperatura sigurno određuju mogućnost dužine skladi-
štenja semena. Primer dužine skladištenja u odnosu na sadržaj vlage i visine temperature skladištenja prikazan je na slici 61. Slika predstva-lja model po kome treba održavati temperaturu i sadržaj vlage kako bi seme održalo odgovarajući nivo kvaliteta kod uljane repice. Ako se tem-peratura ili vlaga uljane repice nalaze u delu koji nije šrafiran na slici 64, treba preduzeti mere za smanjenje jednog od pomenutih činilaca, vlage, temperture ili oba (Canadian Grain Commision, 2009).
Naglo oštećenje semena usled izlaganja visokoj temperaturi i visokoj vlažnosti u laboratorijskim uslovima ima poseban značaj za rad u oblasti starenja semena. Testovi ubrzanog starenja su korisni, a ubrzano starenje podrazumeva izlaganje semena temperaturi od 40oC i visokoj relativnoj vlažnosti vazduha u toku nekoliko dana (najčešće 4), jer daje informaciju
Slika 63. Temperaturni koeficijent za promenu gubitka klijavosti za povećanje temperature od 10°C Q10 za seme navedenih vrsta
Figure 63. Temperature coefficient for change of loss of germination for temperature increase 10°C Q10 for seed of following species
(Ellis and Roberts, 1981)
Tem
pera
turn
i koe
ficije
nt/T
empe
ratu
re co
effici
ent (
Q10
)
Temperatura/Temperature (0C)
22
20
18
16
14
12
10
8
6
4
2
0-20 -10 0 10 20 30 40 50 60 70 80 90 100 110 120
Allium cepaFestuca pratensisGlycine maxHordeum vulgareLactuca sativaLolium pereneOryza sativaPisum sativumTrifolium incarnatumTriticum aestivumVicia fabaAvena sativa, Secale cerealeBegonia semperflores, Myosotis alpestris, Taraxacum dens-leonis, Viola tricolorCallistephus chinensis, Daucus carota, Primula obconica

176
Semenarstvo
o vigoru semena za vrlo kratko vreme, umesto da se istraživanja obavljaju godinama. Korišćenje ubrzanog starenja u istraživanjima fiziologije ošte-ćenja semena je odavno poznato. Na slici 65 prikazan je potencijal vigora kod hibridnog semena suncoktreta (levo) i kukurza (desno) korišćenjem testa ubrzanog starenja. Može se videti da se kod semena suncokreta vigor smanjivao značajno brže nego kod semena kukuruza, mada je smanjenje bilo evidentno kod semena obe biljne vrste (Stan, 1997).
Slika 64. Karta uticaja temperature i vlage na vreme skladištenja uljane repiceFigure 64. Map effect of temperature and moisture ma storage rapeseed
(Canadian Gain Commission, 2009)
Slika 65. Gubitak vigora kod semena suncokreta i kukuruza primenom testa ubrzanog starenja
Slika 65. Vigour potential of sunflower (left) and corn hybrid seeds (right) determined by accelerated ageing method
(http://www.incda-fundulea.ro/rar/nr78/rar7_12.pdf )
Temperatura u skladištu u °C/Temperature °C at binning
Vreme (h)/Time (h)
Vreme (h)/Time (h)
Vlažnost (%)/Humidity (%)
Vlažnost (%)/Humidity (%)
Klij
avos
t (%
)/G
erm
inat
ion
(%)
Klij
avos
t (%
)/G
erm
inat
ion
(%)
sadr
žaj v
lage
% u
skla
dišt
u/m
oist
ure
cont
ent %
, at b
inni
ng
man
je d
ana
za si
gurn
o sk
ladi
šten
je/
few
er d
ays o
f saf
e st
orag
e
manje dana za sigurno skladištenje/fewer days of safe storage
fewer days of safe storage
promene/spoilage
bez promene najmanje 5 meseci/no spoilage for at
least 5 months

177
Opšte semenarstvo
Čuvanje semena na veoma niskim temperaturama je pogodno za čuvanje kolekcija u bankama gena. Čuvanje semena u kontejnerima na -15 ili -20oC je sada već rutinski postupak (Fraser, 1980). Dosadaš-nja iskustva govore da je moguće uspešno uskladištenje mnogih vrsta semena i po nekoliko decenija. Koliko je čuvanje semena pouzdanije na nižim temperaturama, najbolje je prikazano na slici 66. Prave koje označavaju čuvanje semena na -13oC i sadržaja vlage 7% obezbeđuju životnu sposobnost semena više od 65 godina. Ako je sadržaj vlage 12% pri istoj temperaturi životni vek je smanjen na 27 godina. Na 2oC period čuvanja je bio 5 godina. Za seme sa 12% vlage i 13 godina za seme sa 7% vlage.
Slika 66.Uticaj temperature i vlage na preživljavanje semenaFigure 66. Influence of temparature and moisture on seed survival (Tompsett, 1983)
Efekti genotipa na dugovečnost semena
Starenje smanjuje vigor i dugovečnost semena kod inbred linija kukuruza, zbog nenaslednih genetičkih promena. Pored nenaslednih genetičkih promena usled hromozomskih aberacija i oštećenja u DNK sekvencama, nasledne promene tokom skladištenja kukuruza su evi-dentne. Genetička varijabilnost starenja inbred linija semena se može
Klij
avos
t/Ger
min
atio
n (%
)
Godine/Years
12% vlage-moisture 2oC12% vlage-moisture -13oC7% vlage-moisture 2oC7% vlage-moisture -13oC
95
90
80706050403020
10521
0.50 5 10 15 20 25 30 60 65

178
Semenarstvo
koristiti u istraživanjima vezanim za identifikaciju gena vezanih za kli-janje starog semena. Inbred linija kukuruza šećerca P39 i inbred linija EP44 korišćeni su kao materijal za istraživanje. Mešavina živog i mr-tvog semena posle 20 i 22 godine čuvanja su upoređeni upotrebom SSR tehnike. Primenom markera ustanovljeni su genotipovi pojedinačnih semena. Razlika između mrtvog i živog semena može se objasniti zao-stalom varijabilnošću pojedinih semena, spontanim mutacijama ili sta-renjem. Varijabilnost je bila veća na hromozomu broj 7 nego kod dru-gih hormozoma, što ukazuje na vezu između položaja gena u genomu i vitalnosti semena. Polimorfizam je pronađen kod šest poznatih gena upotebom SSR tehnike, koji ukazuje na razlike između mrtvog i živog semena, uključujući i protein za patogenezu 2, superoksid dismutazu 4, katalazu 3, neprozirni endosperm 2 i metilotionin 1 koji koji su vezani za proces klijanja. Pored toga su identifikovani pet kandidat gena, tri od njih mogu biti uključeni u otpornost na bolesti, jedan u detoksikaciju ekeltrofilnih jedinjenja i jedan u regulaciju transkripcije. Genetička va-rijabilnost linija starog semena kukuruza je bila korisna za preliminarne analize identifikacije gena kandidata (Revilla et al., 2009).
Problem čuvanja semena soje u tropskim uslovima je prisutan, te je iz tog razloga Kueneman (1983) izvršio povratna ukrštanja superior-nih i inferiornih genotipova u odnosu na mogućnost dugog čuvanja u skladištima. Ustanovio je da je majčinska biljka vršila značajan uticaj na dugovečnost semena F1 generacije. Kueneman je procenio sposobnost genotipova za skladištenje testom ubrzanog starenja. Ustanovljeni uticaj majčinske biljke se verovatno vrši preko karakteritika semene opne, jer se ona nasleđuje od majke.
Starenje i oštećenje semena
Najbolja definicija starenja i oštećenja semena je da je to gubitak kvaliteta u funkciji vremena. Gubitak kvaliteta može nastati usled fizi-oloških poremećaja i mehaničkih oštećenja. Ovako stroga podela može da postoji, međutim, u najvećem broju slučajeva, starenje semena nasta-je kao posledica interakcije brojnih činilaca.
Definicija starenja semena glasi: starenje ili oštećenje semena je ne-povratan proces. Kada jednom dođe do oštećenja katalitički procesi se ne

179
Opšte semenarstvo
mogu vratiti. Tačno je da seme ne može samo obnoviti neki narušeni si-stem, ali u povoljnim uslovima ima mehanizama kojima se može zaštiti.
Postavlja se pitanje kada je seme toliko oštećeno da gubi životnu sposobnost i šta do toga dovodi. Brojni autori (Woodstock and Tao, 1989; Coolbear, 1995) navode da nepovoljni uslovi setve i značajno smanjenje količine vode mogu drastično da umanje klijavost semena u odnosu na onu koja bi bila u povoljnim uslovima za nicanje.
Dve osnovne biohemijske promene se dešavaju prilikom starenja semena Vigna radiata kod dva različita sadržaja vlage i temperature. Po rezultatima Narayana i sar. (2003) menjao se status lipidne peroksidaze i neenzimskog proteina glikolize. Dva ispitivana pokazatelja su varirala u zavisnosti od promene temperature i vlage. Biohemijske promene i vitalnost semena su značajno izmenjeni i opali u semenu ispod kritične temperature (iznad 40oC, iznad transiracione temperature, kada seme ulazi u staklastu fazu). Podaci pokazuju da je oštećenje semena bilo ve-oma usporeno pre nego što je ušlo u staklastu fazu.
Prema istraživanjima Kranner i sar. (2005) identifikovana su dva univerzalna markera za određivanje dugovečnosti semena baziranih na redukovanom glutation statusu suvog semena (glutation redox sta-te). Tokom oštećenja semena praćena je uloga telomera i drugih DNK oštećenja. Određen je prvi korak u metodi za određivanje dugovečnosti semena. Utvrđena je uloga ćelijske membrane i gubljenje dugovečnosti kod suvog semena. Određen je i sadržaj vode koji je graničan za održa-vanje dugovečnosti semena.
Probert et.al. (2009) su dali model koji ukazuje na uslove koji do-vode do starenja, odnosno oštećenja semena. Oni ovaj proces opisuju kao deo u seriji biohemijskih promena. Posebnu važnost daju oštećenju ćelijskih membrana kao uzroku promena biohemijskih reakcija, a zatim gubljenju ostalih osobina koje počinju smanjenjem klijavosti, posebno poljske, povećanju broja atipičnih ponika i na kraju, potpunom gubitku životne sposobnosti semena.
Uobičajeno je da se kaže da je seme najkvalitetnije kada dostigne maksimalnu masu suve materije, kada se smatra fiziološki zrelim. Posle navedene faze dešavaju se promene u semenu koje utiču na poboljšanje njegovog kvaliteta. Na primer, seme mnogih biljnih vrsta posle dostiza-nja maksimalnog sadržaja suve materije nastavlja sa nakupljanjem spe-cifičnih rezervnih materija, ili se sadržaj vode svodi na niži nivo, što čini čuvanje semena bezbednijim (Kermode i sar., 1986).

180
Semenarstvo
Oštećenja semena u skladištu
O uslovima koji dovode do oštećenja ili starenja semena, ima mno-go podataka. Obično je to početni kvalitet semena, posebno sadržaj vode u semenu, zatim uslovi skladištenja koji podrazumevaju određenu temperaturu i relativnu vlažnost vazduha. O ovome je više rečeno u po-glavlju o doradi i skladištenju semena. Ono što je ovde potrebno istaći i objasniti su mehanizmi koji dovode do starenja, odnosno oštećenja semena.
Važno je ustanoviti šta su primarni, a šta sekundarni prouzrokovači oštećenja. Kao primer mogu se navesti promene u ćelijskim membrana-ma. Veoma slične njima su promene uslovljene genetičkim činiocima. Opadanje vigora semena može biti uslovljeno:
a) otpornošću ćelijskih membrana na oštećenja i
b) mogućnošću obnavljanja ćelijske membrane.
Mogućnost gubljenja vigora kod pojedinačnog semena može biti rezultat nakupljanja toksičnih metabolita u njemu (na primer masnih kiselina kratkih lanaca). Različiti su činioci kod raznih vrsta semena koji dovode do oštećenja semena u raznim uslovima skladištenja. Uvek se mora imati na umu da svi činioci deluju zajednički i da uslovljavaju jedan drugog. Oštećenje membrana, mitohondrija može se odražavati na intenzitet disanja. Oštećenje plazmaleme – endoplazmatičnog reti-kuluma – Goldžijevog aparata smanjuje količinu proteina koji se sinte-tišu u ćelijama. Ova i još neka mehanička oštećenja semena data su na slici 67 (Osborn, 1980).
Pokušaj određivanja uzroka i posledica oštećenja semena, odnosno njihov mehanizam učinio je Basra (1995). U tabeli 32 iznete su katego-rije oštećenja, način ustanovljavanja oštećenja i posledice koje nastaju u ovom procesu.

181
Opšte semenarstvo
“Živ
o se
me”
“A
live
seed
”
Gub
itak
inte
grite
ta m
embr
ane
Loss
of m
embr
ane
inte
grity
Gub
itak
aktiv
nost
i enz
ima
Loss
of e
nzym
atic
act
ivity
N
epre
kidn
a ak
tivno
st n
ukle
aza
Perm
anen
t nuc
leas
e ac
tivity
Ošt
ećen
je sl
obod
nih
radi
kala
D
amag
e of
free
radi
cals
Pero
ksid
acija
lipi
da
Lipi
d pe
roxi
datio
n
Ned
osta
tak
deob
e će
lija
Lack
of
cell
div
ision
Ned
osta
tak
mito
hond
rijal
neak
tivno
sti
Loss
of m
itoho
ndria
l act
ivity
Ribo
nukl
eaza
Ri
bonu
clea
sePo
veća
na a
ktiv
nost
end
odez
oksir
ibon
ukle
aze
Hig
er a
ctiv
ity o
f end
odes
oxiri
bonu
clea
se
Ned
osta
tak
obna
vlja
nja
i pol
imer
aze
enzi
ma
DN
K
Lack
of r
egen
erat
ion
of p
olym
eras
e ez
yme
DN
A Gub
itak
inte
grite
ta D
NK
Lo
ss o
f DN
A in
tegr
ityG
ubita
k in
tegr
iteta
ribo
zom
a Lo
ss o
f rib
osom
e in
tegr
ity
Ned
osta
tak
sinte
ze fu
nkci
onal
ne D
NK
La
ck o
f fu
nctio
nal D
NA
synt
hesis
Ned
osta
tak
dehi
doge
nazn
e tr
ansf
eraz
e tR
NK
-sin
teta
ze
Lack
of d
ehid
roge
nase
tran
sfer
ase
tRN
A sy
thet
ase
Ned
osta
tak
sinte
ze p
rote
ina
Lack
of p
rote
in sy
nthe
sis
“Mrt
vo se
me”
“D
ead
seed
”
Slik
a 67
. Utic
aj fi
ziol
oški
h pr
omen
a na
gub
itak
vigo
ra se
men
aFi
gure
67.
Influ
ence
of p
hysio
logi
cal c
hang
es o
n se
ed v
igou
r (O
sbor
n, 1
980)

182
Semenarstvo
Tabela 32. Mehanizam oštećenja semena u skladištu Table 32. Seed demages mechanism intore (Basra,1995)
Kategorija promena/Category of changes
Mera ustanovljavanja promena/
Measurement for establishing changes
Posledice promena/Consequences caused
by changes
Enzimske i proteinske promene/Enzyme and protein changes
Merenja aktivnosti enzima,analiza proteina,promene sinteze proteina,promene metabolizma RNK/Measurement of enzyme activity, protein analysis, changes of protein synthesis, changes of RNA metabolism
Oštećenje nastalo slobodnim radikalima, npr. hidrolitičko oštećenje,slabljenje sinteze proteina,dezorganizacija ćelijskih membrana/Damages caused by free radicals such as hydrolytic damage, weakening protein synthesis, disorganization of cell membranes
Hormonske promene/Hormone changes
PGR tretmani starog semena,demonstracije promenaosetljivost na PGRS/PGR treatments for old seed, demonstration of changes, sensitivity to PGR
Prekid metabolizma,gubitak osetljivosti na PGRS preko oštećenja membrana ili smanjenog metabolizma u ćelijama/Metabolism interruption, sensitivity to PGRS lost via damaged membranes or decreased metabolism inside cells
Toksični metaboliti/Toxic metabolits
Direktno otkrivanje/Direct revealing
Primarni ili sekundarni proizvodi, reakcija oštećenja,promene u sistemu detoksikacije/Primary or secondary products, rection of demage, changes in detoxication system
Mikroorganizmi/Microorganisms
Direktno otkrivanje oštećenjaeksperimenti inokulacije/Increased moisture content of seed and/or temperature
Povećanje sadržaja vlage semena i/ili temperatureIncreased moisture content of seed and/or temperature

183
Opšte semenarstvo
Insekti kao prouzrokovači umanjenja kvaliteta semena u skladištu
Uzrok propadanja semena su glodari i insekti koji uništavaju ili oštećuju seme već od početka oplodnje. Kod većine krmnih biljaka kontaminacija insektima dolazi iz polja. Kao primer može poslužiti graškov žižak. Graškov žižak (Bruchus pisorum) (sl. 68) predstavlja ekonomski najznačajniju štetočinu u proizvodnji zrna i semena pro-teinaskog graška u oblastima sa aridnijim klimatom, posebno mediteranskim i ze-mljama jugoistočne Evrope i Australije. Oko 90% šteta prouzokovanih graškovim žiškom bi moglo da se predupredi žetvom u optio-malnom roku i fumigacijom odmah nakon žetve (Milošević et al., 2010).
U tabeli 33 je dat pregled insekata koji su ekonomski najštetniji, njihove osnovne karakteristike kao što su veličina imaga, du-žina životnog ciklusa, da li mogu da lete ili ne, kao jedna od osobina po kojoj se razliku-ju, vrsta semena koje oštećuje, i vrsta ošteće-nja koja prouzrokuju. Kao što se iz podataka može videti razvoj generacija je kratak po-sebno kod žižaka (Sitophilus granarius 30 do 50 dana, Sitophilus zeamaise 30–40 dana) (sl. 69), pa posle nekoliko meseci dolazi do nji-hovog značajnog umnožavanja. Većina žiža-ka završava životni ciklus u semenu tako da je njihovu pojavu teško okularno primetiti u skladištu. Ostali skladišni insekti su manje specijalizovani. Najštetniji od njih za žita su Sitophilus granarius i Sitophilus oryzae. Po-java svih insekata je vezana u velikoj meri za sadržaj vlage u okolnoj sredini i semenu i uslovljena je temperaturom. Optimalna
Slika 68. Graškov žižak (Bruchus pisorum)
Figure 68. Pea weevil (Bruchus pisorum)
Slika 69. Kukuruzni žižak (Sitophylus zeamais)
Figure 69. Maize weevil(http://upload.wikimedia.org/
wikipedia/commons/1/1a/ Maize_weevil.jpg)

184
Semenarstvo
temperatura za razvoj većine insekata je 24–37oC, dok do smanjenja in-tenziteta pojave dolazi na temperaturi od 15 do 24oC. Na temperaturi vi-šoj od 15oC većina insekata nije aktivna (Milošević i Malešević, 2004).
Tabela 33. Insekti na semenu u skladištuTable 33. Insects on seed in store (Milošević, Malešević, 2004)
Naziv insekta/Name of
insect
Dužina odraslog insekta/lenght of
imago
Oštećeno seme/
Damage seed
Životni ciklus/
Life cyrcle
Da li može da
leti/ possibility
to fly
Komentar/ Comment
Žižak/weevil mm Dani/days
Sitophilus granariusŽitni žižak/Wheat weevil
3–5 Žita 30-50 ne
Jaja polažu u semenu, lar-ve i lutke se hrane unutar semena. Otporni na hlad-noću/ Eggs pass in the seed, and the larvae feed on the doll inside seeds. Resistant to cold
Sitophilus oryzaePirinčani žižakRise weevil
2–3 žita 25-30 da
Jaja polažu u semenu. Lar-ve i lutke se hrane unutar semena / Eggs pass in the seed. larvae feed on the doll and seeds inside
Sitophilus zeamaiseKuuruzni žižakCorn weevil
2–4 žita 30-40 da
Isto kao pirinčani žižak, osim što je veći / Eggs pass in the seed. larvae feed on the doll and seeds inside
Acanthoscelides obtectusPasuljev žižak Bean weevil
3 pasulj, grašak 21-80 da
Ova pozicija na mahu-nama u polju ili među semenom, larve i lutke se hrane unutar semena, bez nosa. Adaptirani na rela-tivno nisku temperaturu./ Ovipozition the pods in the field or among seed, and larvae feed on the doll inside seeds. without a nose. Adapted to relati-vely low temperatures.

185
Opšte semenarstvo
Patogeni kao prouzrokovači umanjenja kvaliteta semena u skladištu
Pod mikropopulacijom koja se javlja kod semena u skladištima uglavnom se podrazumeva gljivična flora. Kontaminacija semena i zrna gljivičnim organizmima može da rezultira nezadovoljavajućim klija-njem semena, vigorom ili ukupnim kvalitetom semena. Skladišne gljive kontaminiraju obično seme tokom skladištenja i obično nisu prisutne u većem procentu pre žetve u polju.
O prisustvu i ulozi bakterija i virusa ima veoma malo podataka. Pretpostavlja se da fitopatogene bakterije u skladištima nemaju dovolj-no slobodne vode da bi se mogle razvijati. Pored toga, uslovi umanjene vlažnosti iniciraju porast skladišnih gljiva koje svojim razvijanjem one-mogućavaju intenzivnu pojavu fitopatogenih bakterija, odnosno uspo-stavlja se inkopatibilan odnos između ove dve grupe patogena (Bewley and Black, 1983).
Postoje dve grupe gljiva koje se javljaju na semenu. To su tzv. poljske i skladišne gljive. Poljske gljive se javljaju, kao što samo ime govori, u polju i zaražavaju seme koje se razvija u polju. Gljive za-htevaju visok sadržaj vlage za razvoj. Tako, na primer, za žita je ta vlažnost 30–33%. Ovako visoka vlaga umanjuje se u toku sazreva-nja semena, u procesu sušenja semena posle žetve, a pre skladištenja. Kod semena zaraženog poljskim gljivama dolazi do promena kao što su: obezbojavanje, gubljenje funkcije embriona, produkcija toksina. Među ekonomski najznačajnije ubrajaju se gljive iz roda Drechslera, Fusarium, Alternaria i dr.
Gljive koje se ne javljaju u skladištima u većoj meri, ne naseljavaju seme pre žetve. Procenat zaraženosti mesec dana pre žetve iznosi kod semena kukuruza svega 5% ukupne mikoflore. Tokom berbe i tran-sporta na površini zrna može se konstatovati povećanje broja mikro-organizama, a i procenat gljiva koje se javljaju u skladištima time se znatno povećava. Skladišne gljive uglavnom pripadaju rodu Penicilli-um, Aspergillus, Rhizopus, Mucor i dr. (tab. 34) (Milošević, 1983).

186
Semenarstvo
Tabela 34. Mikopopulacija izolovana sa semena kukuruza uskladištenog u siloseTable 34 Seed borne fungi isolated from corn stored in silo (Milošević, 1979)
Vrsta gljiva/ Fungi Vrsta gljiva/Fungi
Rod Aspergillus/Genus AspergillusAspergillus flavusAspergillus clavatusAspergillus fumigatusAspergillius glaucusAspergillus ustusAspergillus candidusAspergillus niger
Genus penicillium/Rod PenicilliumPenicillium cyclopiumPenicillium purpurogenumPenicillium multicolorPenicillium brevicompactumPenicillium notatum
Ostale gljive / Other fungiFusarium oxysporumHelminthosporium carbonumAlternaria tenuisCladosporium cladosporoidesNigrospora oryzae
Intenzitet kojim se odvijaju promene zavisi od više činilaca, a među najznačajnije ubrajaju se temperatura i vlažnost kako samog semena, tako i okoline u kojoj je seme skladišteno. Uslovi koji utiču na razvoj skladišnih gljiva u skladištu su:
– sadržaj vlage u semenu,– temeratura,– početni kvalitet semena,– dužina skladištenja, – aktivnost insekata i grinja na semenu.
Promene koje nastaju na semenu kao posledica zaraženosti skladiš-nim gljivama su brojne, a u najvažnije se ubrajaju:
– gubitak klijavosti,– obezbojavanje semena (kličnog dela i endosperma),– zagrevanje semena,– biohemijske promene u semenu,– gubici u masi semena,– produkcija mikotoksina.

187
Opšte semenarstvo
Tabe
la 3
5. M
ikot
oksin
i u zr
nu i
sem
enu
žita
Tabl
e 35.
Myc
otox
ins i
n w
heat
gra
in a
nd se
ed (h
ttp://
ww
w.fa
o.or
g/W
aird
ocs/
X50
08E/
X50
08e0
1.ht
m)
Mik
otok
sin/
Myc
otox
in B
iljna
vrs
ta/
Plan
t spe
cies
Glji
va iz
vor/
Fu
ngal
sour
ce(s
)Ef
ekti
pris
ustv
a/Eff
ects
of i
nges
tion
Niv
alen
olde
oxyn
ival
enol
/ni
vale
nol
Pšen
ica,
kuk
uruz
, ječ
am
/whe
at, m
aize
, bar
ley
Fusa
rium
gra
min
earu
mTo
ksič
an za
ljud
e In
dija
, Kin
a, Ja
pan
i Kor
eja.
Otr
ovne
za
živo
tinje
, pos
ebno
svin
je /H
uman
toxi
cose
s Ind
ia, C
hina
, Ja
pan,
and
Kor
ea. T
oxic
to a
nim
als,
espe
cial
ly p
igs
Fusa
rium
croo
kwell
ense
Fusa
rium
culm
orum
Zera
leno
neZe
aral
enon
eKu
kuru
z, pš
enic
a/m
aize
, whe
atF.
gra
min
earu
mId
entifi
kova
ne o
d st
rane
Međ
unar
odne
age
ncije
za
istra
živa
nje
raka
(IA
RC),
kao
mog
ući l
juds
ki k
arci
noge
n.
Utič
e na
repr
oduk
tivni
sist
em že
nki s
vinj
a /
Iden
tified
by
the
Inte
rnat
iona
l Age
ncy
for R
esea
rch
on
Can
cer (
IARC
) as a
pos
sible
hum
an c
arci
noge
n. A
ffect
s re
prod
uctiv
e sy
stem
in fe
mal
e pi
gs
F. cu
lmor
umF.
croo
kwell
ense
Och
rato
xin
AJe
čam
, pše
nica
i dr
uge
vrst
e/ba
rley,
whe
at, a
nd
man
y ot
her s
peci
es
Asp
ergi
llus o
chra
ceus
Sum
nja
da je
IARC
kao
ljud
ska
karc
inog
en. K
ance
roge
ne
kod
labo
rato
rijsk
ih ži
votin
ja i
svin
ja /
Susp
ecte
d by
IARC
as h
uman
car
cino
gen.
Car
cino
geni
c in
labo
rato
ry a
nim
als a
nd p
igs
Peni
cilliu
m v
erru
cosu
m
Fum
onisi
nfu
mon
isin
B1Ku
kuru
z/M
aize
Fusa
rium
mon
ilifo
rme
plus
seve
ral l
ess c
omm
on
spec
ies
Sum
nja
da je
IARC
kao
org
ansk
i kar
cino
gen
za
ljude
. Tok
sičan
za sv
inje
i ži
vinu
. Uzr
ok k
onjsk
i eu
coen
cefa
lom
alac
ija (E
LEM
), fa
taln
a bo
lest
kon
ja /
Susp
ecte
d by
IARC
as h
uman
car
cino
gen.
Tox
ic to
pig
s and
po
ultr
y. C
ause
of e
quin
e eu
coen
ceph
alom
alac
ia (E
LEM
), a
fata
l dise
ase
of h
orse
s
Afla
toks
inafl
atox
in B
1, B2
Kuku
ruz,
kiki
riki i
m
noge
dru
ge v
rste
/m
aize
, pea
nuts
, and
m
any
othe
r spe
cies
Asp
ergi
llus fl
avus
Afla
toks
in B
1 i p
rirod
na m
ešav
ina
Afla
toks
ina,
iden
tifiko
van
kao
pote
ncija
lni l
juds
ki k
arci
noge
ni p
o IA
RC. N
ežel
jeni
h ef
ekat
a u
razn
im ži
votin
jam
a, p
oseb
no p
ilići
ma
/ A
flato
xin
B1, a
nd n
atur
ally
occ
urrin
g m
ixtu
res o
f afla
toxi
ns,
iden
tified
as p
oten
t hum
an c
arci
noge
ns b
y IA
RC. A
dver
se
effec
ts in
var
ious
ani
mal
s, es
peci
ally
chic
kens
Afla
toks
inafl
atox
in B
1, B2,
G1, G
2
Kuku
ruz,
kiki
riki/
mai
ze, p
eanu
tsA
sper
gillu
s par
asiti
cus

188
Semenarstvo
Seme soje inokulisano različitim vrstama skladišnih gljiva (Asper-gillus niger, A. Flavus, Fusarium solani i Rhizopus sotlonifer) je čuvano u skladištu tri meseca pri relativnoj vlažnosti vazduha 60%, 70%, 80%, 90% i 100%. Pri sadržaju vlage od 60% nije došlo do promena u sadržaju ulja, proteina, šećera i masnih kiselina. Kako je sadržaj vlage vazduha rastao sadržaj ulja i ukupnih šećera je opadao, ali su proporcionalno rasli sadržaj proteina, jer su gljive uticale na konverziju masti i uglje-nih hidrata u proteine. U osnovi je došlo do promene u sastavu masnih kiselina, povisio se sadržaj oleinske kiseline, a proprocionalno je opao sadržaj linolne i linolenske kiseline (Kakde and Chavan, 2011)
U tabeli 35 prikazani su mikotoksini koji se najčešće sreću u zrnu i semenu žita, kao i njihov štetan efekat na zrno i seme. Aflatoksin se najčešće javlja i jedan je od najpoznatijih mikotoksina kontaminenata zrna i semena. Aflatoksin je sekundarni metabolit koji proizvode gljive Aspergillus flavus i A. parasiticus.
Aflatoksin B1 je najvažniji toksin sa toksikološke tačke gledišta. On je visokopotentni karcinogen za ljude i životinje, nalazi se prvenstveno u kukuruzu, semenu pamuka i stablu oraha. Međunarodna organizacija za istraživanje raka je aflatoksin B1 stavila na listu humanih kancero-gena. Aflatoksin B2 je toksikološki manje značajan jer se nalazi u mno-go nižim koncentracijama nego aflatoksin B1, ali je uvek prisutan gde i aflatoksin B1. Aflatoksini G1 i G2 takođe potiču od gljive A. flavus, a najviše ga ima u kikirikiju, ali se retko nalazi u kukuruzu (http://www.fao.org/Wairdocs/X5008E/X5008e01.htm).

189
Opšte semenarstvo
LITERATURA
Aastrup S, Riis P, Hansen J R ( 1989): Storage Conditions. Proceedings of the European Brewery Convention Congress, Zurich, 171-178.
Abba J E, Lovato A (1999): Effect of seed storage temperature and rela-tive humidity on maize (Zea mays L.) seed viability and vigour. http://cat.inist.fr/?aModele=afficheN&cpsidt=1895615
Adams, R P (1997): Conservation of DNA: DNA banking. In Callow, J.A, Ford-Lloyd, B.V, Newbury, H.J, editors. Biotechnology and Plant Genetic Resources: Conservation and Use. Biotechno-logy in Agriculture Series 19. CAB International, Wallingford, UK. Pp. 163 – 174.
Argyris J, Van Sanford D, TeKrony D M (2005): Fusarium graminearum infexctio during wheat seed development and its effect on seed quality. Seed Physiology, Production and technology, Crop Sci-ence, 43:1782-1788.
Atanassov A (2003): Plant Seed Activities in Bulgaia. 3rd EESNET Mee-ting on GMO and Seed Industry, Belgrade, 4-6 November, 2003.
Babić M, Babić Lj (2000): Dorada semena, Poljoprivredni fakultet, Novi Sad.
Barjaktarović R, Kobiljski B, Obreht D, Vapa LJ (2004): Evaluation of wheat dormancy using microsatellites. Proc. XXXIV Ann. Meeteng “European Soc. For New Methods in Agric. Res.”, 29 Aug.-2 Sept, 2004, Novi Sad, Serbia and Montenegro. 228-230.
Basra A S (1995): Seed Quality Basic Mechanisms and Agricultural Im-plication. Food Products Press. An inprint of the Haworth Pre-ss, Inc, New York.
Basra A S (2006): Handbook of Seed Science and Technology. Food Pro-duct Press.
Bench ALR, Sanchez RA (2004): Handbook of Seed Physiology. Food Product Press.

190
Semenarstvo
Benjamin A (2008): Germany bans chemical linked to honeybean dev-stiation. Guardian. http//: www.guardian.co.uk/environment /2008/may/23/wildlife.endangeredspecies.
Bewley J D, Black M (1983): Physiology and Biochemistry of Seeds. Springler-Verlag, Berlin, Heidelberg, New York.
Brbaklić Lj, Kondić-Špika A, Trkulja D, Kobiljski B (2010): Asocijativna analiza između mikrosatelitskih markera i agronomski važnih svojstava pšenice. Ratarstvo i povrtarstvo. 47(2): 505-510.
Burris S J (1980): Drying High Moisture Corn Seed, 35th Annual Corn and Sorghum research Conference, Washington, USA.
Canadian Grain Commission (2009): Spoilage andheating of stored agri-cultural products. Changes that occur during storage. http://www.tbs-sct.gc.ca/rpp/2009-2010/inst/cgc/cgc02-eng.asp
Chittenden H W, Svec L V (1974): Effect of pottassium on the incidence of Diaporthe sojae in soybean. Agron. Jour, 66, 696-697.
Commision Regulation (2004): (EC) No 65/2004 establishing a system for the development and assignment of unique identifiers for genetically modified organisms, Official Journal of the Europe-an Union L 10/5.
Council Decision (1999): 1999/468/EC laying down the procedure for the excercise of implementing power conferrer on the Commi-sion, EUR Lex.
Council Decision (2009): C(2000) 146/FINAL incl. 2003, 2004, 2005, 2006, 2007 and 2008 amendments OECD Seed Scheme, Deci-sion of the Council revised the OECD Scheme or the Control of Seed Moving in Interantional Trade. http://www.oecd.org/dataoecd/30/46/40203758.pdf
Council Directive (2001): 2001/18/EC Of the European Parliament and of the Council on the deliberate release into the environment of genetically modified organisms and repealing Council Directi-ve 90/220/EEC, 2001L0018— EN— 21.03.2008 — 003.001— 1. http://ws500.ifvcns.ns.ac.rs/cgi-bin/patience.cgi?id=49af5b89-25f7-4057-a792-0a511b089952
Council Directive (2000): 2000/29/EC on protective measures against the intirduction into the Community harmful plant and plant products and against their spred within the Community, EUR Lex. http://eurlex.europa.eu/LexUriServ/LexUriServ.do?uri=CONSLEG:2000L0029:20060414:EN:PDF

191
Opšte semenarstvo
Council Directive (2002):/ 2002/53/EC on the common catalogue of varieties of agrcultural plant species, Official Journal of the Eu-ropean Communities L 193/1. http://eur-lex.europa.eu/LexU-riServ/LexUriServ.do?uri=OJ:L:2002:193:0001:0011:EN:PDF
Commission Directive 2008/83/EC: Examination and the minimum conditions for examining certain varieties of vegetable species. http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2008:219:0055:0057:EN:PDF
Council Regulation (1971): (EEC) no.2358/71 of the Council on common organization of the market in seeds. http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=CONSLEG:1999D0468:20060723:EN:PDF
Council Regulation (1972): (EEC) no 1674/72 of the Council laying down general rules for granting and finansing aid for seed, EUR Lex.
Counncil Regulation (2003): Counncil Regulation (EC) No 1782/2003 establishing common rules for direct support schemes un-der the common agricultural policy and establishing certain support schemes for farmers and amending Regulations (EEC) No 2019/93, (EC) No 1452/2001, (EC) No 1453/2001, (EC) No 1454/2001, (EC) 1868/94, (EC) No 1251/1999, (EC) No 1254/1999, (EC) No 1673/2000, (EEC) No 2358/71 and (EC) No 2529/2001, Official Journal of the European Union L 270/1.
Council Regulation (2003): (EC) No 1829/2003 of the European Parlia-ment and of the Council of 22 September on genetically modi-fied food and feed. http://www.ciaa.be/documents/brochures/GMguidelines.pdf
Council Regulation ( 2003): (EC) No 1830/2003 concerning the tracea-bility and labelling of genetically modified organisms and the traceability of food and feed products produced from geneti-cally modified organisms. http://www.ciaa.be/documents/bro-chures/GMguidelines.pdf
Council Regulation (2003): (EC) No.1946/2003 on transboundary movements of genetically modified organisms. http://www.doeni.gov.uk/index/protect_the_environment/natural_envi-ronment_/genetically_modified_organisms.htm
Council Regulation (2005): (EC) no.1947/2005 on the Common Organ-zation of the Market in Seed and repealing regulation (EEC)

192
Semenarstvo
no.2358/71 and (EEC) no.1647/72, Official Journal of the Eu-ropean Union L 312/3.
Cooke E L, Lees A K (2006): Markers, old and new, for examining Phytophthora infestans diversity, Scottish Crop Research In-stitute, Invergowrie, Dundee, DD2 5DA, UK, CABI Abstract. http://www.cababstractsplus.org/abstracts/Abstract.aspx? AcNo=20043212859
Coolbear P (1995): Mechanism of Seed Deterioration. In Seed Quality Basic Mechanisms and Agricultural Implication. Basra, A.S. Food Products Press. Haworth Press, Inc. New York.
Contreras S, Tay D, Bennett M (2009): Effects of temperature during seed development in Lactuca sativa and Helianthus debilis. http://hcs.osu.edu/students/retreat/abstracts/contreras1.pdf
Coolbear P (1995): Mechanism of Seed Deterioration. In: Seed Qua-lity Basic Mechanisms and Agricultural Implication, Basra, A.S.Food Products Press, Haworth Press, Inc. New York.
Cresswell E G, Grime J P (1981): Induction of a light requirement du-ring seed development and its ecological consequences. Nature 91, 583-585.
Ćirović M (1990): Mogućnost proizvodnje semena hibridnog kukuruza u Jugoslaviji za domaće tržište i izvoz. Zbornik radova jugoslo-venskog savetovanja „Dugoročni program proizvodnje i mar-keting kukuruza”. SPITJ, Beograd, 183-195.
Ćupina T, Sakač Z (1988): Fiziološki aspekti formiranja prinosa sunco-kreta. Poljoprivredni fakultet, Novi Sad.
Delouche J C (1980): Environmental effects on seed development and seed quality. Hart. Scien. 15, 775-780.
Dodig D, Zorić M, Kobiljski B, Šurlan-Momirović G, Quarrie S A (2010): Assessing drought tolerance and regional patterns of genetic diversity among spring and winter bread wheat using simple sequence repeats and phenotypic data. Crop and Pastu-re Science 61(10) 812-824 doi:10.1071/CP10001.
Dordas C, Apostoloides G E, Goundra O (2007): Boron application affects seed yield and seed quality of sugar beets. The Journal of Agricultural Science 145: 377-384.
Dornbos D L J, McDonald M B J (1986): Mass and Composition of De-veloping Soybean Glycine-Max Cultivar Williams-79 Seeds at Five Reproductive Growth Stages. Crop Science 26, 624-30.

193
Opšte semenarstvo
Contact: DORNBOS D L JR ; DEP AGRONOMY, OHIO STA-TE UNIV, COLUMBUS, OH 43210, USA.
Dornbos D L (1995): Production Environment and Seed Quality.In: Basra A.S.: Seed Quality Basic Mechanisms and Agricultural Implication. Food Products Press. An inprint of the Haworth Press, Inc, New York.
Đokić D (1995): Fiziološke osnove selekcije pšenice na efikasnost ishra-ne azotom. Prvi simpozijum za oplemenjivanje organizama. Vrnjačka Banja. Zbornik abstrakata.
Egli D B, Bruening W P (2004): Water stress, photosynthesis, seed sucrose levels, and seed growth in soybean. Journal of Agricul-tural Science 142:1-8.
Egli D B, TeKrony D M, Heitholt J J, Rupe J (2005): Air temperature du-ring seed filling and soybean seed germination and vigor. Crop Sci 45: 1329-1335.
Eđe O T, Burris J C (1970): Physiological and biological changes in de-teriorating soybean seeds. Seed Sci. and Technol, 60, 158-166.
Ellis R H, Roberts E H (1981): The quantification of ageing and survival in orthodox seeds. Seed Science and Technology 9: 373–409.
Ellis R H, Hong T D, Roberts E H (1981): The development of Dessca-tion-tolrance uring Seed Maturation in Six Grain Legumes. Annals of Botany 59:23-29.
Ellis R H, Hong T D (2006): Temperature Sensitivity of the Low-moistu-re-content Limit to Negative Seed Longevity–Moisture Con-tent Relationships in Hermetic Storage. Ann Bot. 2006 May; 97(5): 785–791.
Ellis R H, Bass I N, Whriting D (2008): Storing vegetable and flower seed. Extention no 7: 221.
Envirnment Plant Inspection. http://www.scri.uk/research/epi/agroe-cology/fieldandlansscape
European Technology Platform (2007): Plant fot the Future, Detailed Stategic Research Agenda 2025 and Action Plan 2007-2012. www.plantforthefuture.eu
Fenner M (1987): Seed Ecology. Ed. Chapman and Hall, London.Garden Guides (2010): How Different Lights Make Plants Grow, Gar-
den Guide Research. http://plantphys.info/plant_physiology/phytochrome.shtml

194
Semenarstvo
Gene Expresion Atlas Data Sources (2011): Rice genome. http//:signal.salk.edu/cgi-bin/RiceGE?CHROMOSOME=chr01&LOCATION=24001&INTERVAL=120
Graeber K, Linkies A, Muller K (2009): Cross-species approaches to seed dormancy and germination: conservation and biodiver-sity of ABA-regulated mechanisms and the Brassicaceae DOG1 gene, Springer Science+Business Media B.V.
Gupta P K, Langridge P, Mir R R (2010): Marker-assisted wheat bree-ding: present status and future possibilities. Mol Breeding. 26:145-161.
Gutterman Y (1982): Phenotypic effect of photoperiod on seed germina-tion. In: Elsevier: The Phisiology and Biochemistry of Seed De-velopment, Dormancy and germination, Amsterdam, 67-79.
Guy R (1982): Influence du stockage sur la duree de germination des semences. Rev. Suisse Vitic. Arbovic. Hotic. 14, 99-101.
Hanelt P, Mettin D (1989): Biosystematics od the genus Vicia L. (Legu-minosae) Ann. Rep. Ecolog.Syst. 20, 199-223.
Harty R, Hopkinson J M, English B H, Adler J (1983): Germination, dormancy and longevity in stored seed of Panicum maximum. Seed Sci. and Technol. 11, 341-351.
Hecht-Buchholz C (1979): Calcium deficiency and plant ultrastructure. Soil. Sci. Plant Anal. 10, 67-81.
High Beam Research (2009): Data on soil science discussed by research-ers at University of Minnesota. Agriculture Week. http://www.highbeam.com/doc/1G1205802674.html
http://www.ishs.org/sci/icraname.htmhttp://growingtaste.com/storage.shtmlhttp://spectrumindustries.tradeindia.com/Exporters_Suppliers/Expor-
ter1583.398659/Vibro-Cleaner.html http://www.ubthenews.com/topics_summaries.htmhttp://www.molecular-plant-biotechnology.info/nuclear-genome/
plant-genome.htmhttp://genome.wellcome.ac.uk/doc_wtd020778.htmlhttp://davesgarden.com/guides/pf/b/Asteraceae/Helianthus/none/cul-
tivar/0/http://wishart.biology.ualberta.ca/BacMap/graphs_cgview.htmlhttp://www.ces.ncsu.edu/resources/crops/ag546-1/helixes3.jpg

195
Opšte semenarstvo
http://www.molecularstation.com/molecular-biology-images/502-dna-pictures/109-dna.html
http://signal.salk.edu/cgi-bin/RiceGE?GENE=Os01g01070http://signal.salk.eduhttp://europa.eu/index_en.htmhttp://www.physorg.com/news98540343.htmlhttp://www.ncss.go.jp/main_e/functions/dus.htmlhttp://www.ipo.org/AM/Template.cfm?Template=/CM/ContentDis-
play.cfm&ContentID=22898http://users.ugent.be/~avierstr/principles/pcr.htmlhttp://agro.se-ua.net/images/shvmuha1.jpghttp://www.agbioforum.org/v8n23/v8n23a06-oehmke.htm#F2http://sweetthingdesigns.typepad.com/.a/6a00e54f836792883301156fb
1d962970c-450wihttp://plantphys.info/plant_physiology/phytochrome.shtmlhttp://www.gardenguides.com/75678-different-lights-make-plants-
grow.html http://farm4.static.flickr.com/3144/2867107023_3822b2212f.jpghttp://www.aragriculture.org/crops/soybeans/harvesting.htm
http://www.ag.ndsu.edu/pubs/plantsci/crops/pp447w.htmhttp://www.agriculture.com/news/technology/bayer-monsto-reach-
seed-treatment-deal_6-ar8941http://www.frontiernet.net/~vohsseeds/http:/agritech.tnau.ac.in/seed_certification/seed%20Storage%20fac-
tors.htmhttp://www.fao.org/docrep/006/y4011e/y4011e0v.htmhttp://www.fdh.org/-Agro-ecology-.html?lang=frhttp://www.syngentafoundation.org/__temp/DG_SFSA_SeedProce-
ssingandStorage.pdfhttp://www.fao.org/Wairdocs/X5008E/X5008e01.htmhttp://www.bing.com/images/search?q=seed+++germination+of+dor
mant+seed+in+soil+images++&view=detail&id=FB7E7CF5DEA1991B5DBB9883D5F9949BB0154C3C&first=61&FORM=IDFRIR&qpvt=seed+++germination+of+dormant+seed+in+soil+images
http://www.euroseeds.org/who-we-are

196
Semenarstvo
http://www.bing.com/images/search?q=seed++drying+images&view=detail&id=C8C6BAE81C2DBAB24 242ADFC23409BEDC16F9E7&first=211&FORM=IDFRIR&qpvt=seed++drying+images
http://www.incda-fundulea.ro/rar/nr78/rar7_12.pdfhttp://upload.wikimedia.org/wikipedia/commons/1/1a/Maize_weevil.jpgHartmann, W. (1970): der Einfluss Samenalters auf die Vernaliserba.keit
von Oenothera biennis L. Biochem. Physiol. Planz. 161, 469-471ISHS (2009): International Code of Nomenclature for Cultivated Plants.
Acta Horticulturae Vol. 647 and as Regnum Vegetabile Vol. 144. http://www.ishs.org/sci/icraname.htm
IKISAN (2000): Seed material. http//:www.ikisan.com/links/ap_seeds.shtml#Seed%20Storage
IRRI (2009): Grain Storage - Fungi, CYMMIT. http//:www.knowled-ge-bank.irri.org/storage/index.php/storge-pests-mainme-nu-286/2-fungi-mainmenu-288
ISF (2000): Biological Control Agent Seed Treatment. In ISF: Seed tre-atment and Environment Committe (STEC).
ISF (2009): Seed Statistics, ISF. http://www.worldseed.org/isf/seed_sta-tistics.html
ISTA (2011): International Rules for Seed Testing, ISTA Secreteriat, Zu-rich, Switzeland. http://www.seedtest.org/en/international-ru-les-_content---1--1083.html
Jevtić S (1992): Posebno ratarstvo. Nauka, Beograd.Kadham F A, Speeht J E, Williams J H (1985): Soybean irrigation seri-
ally timed during storage R1 to R6. J. Agronomic responses. Agron. Journ. 77, 291-298.
Kakde R B, Chavan A M (2011): Extracellular Lipase Enzyme Produc-tion by Seed-Borne Fungi under the Influence of Physical Fac-tors, International Journal of Biology. http://ccsenet.org/jour-nal/index.php/ijb/article/view/ 7020
Kirnak H, Dogan E, Alpaslan M, Celik S, Boydak E, Copur O (2008):Drought Strees Imposed at Different Reproductive Stages Influences
Growth, Yield and Seed Composition of Soybean, The Philip-pine Agricultural Scientist, Vol 91, No 3.
Kobiljski B, Denčić S, Pilipović J (2006): Molecular screening of dome-stic germplasm for allelic variants at the dwarfing gene Rth8 locus in wheat. Genetika. 38 (1): 67-74.

197
Opšte semenarstvo
Kobiljski B, Denčić S, Kondic-Špika A, Lohwasser U, Börner A (2009): Locating Stable Across Environment QTL Involved in the De-termination of Agronomic Characters in Wheat. Cereal Rese-arch Communication. Vol. 37, No.3, 327-333.
Kobiljski B, Kondić-Špika A, Denčić S, Kačavenda D, Brbaklić Lj (2008): Novi pristup u molekularnom oplemenjivanju pšenice na pri-nos zrna. Zbornik apstrakata Petog naučno-stručnog simpo-zijuma iz selekcije i semenarstva Društva selekcionara i seme-nara. 25-28. Maj, 2008. Hotel “Breza“ (Vrnjačka Banja). str. 94.
Kobiljski B, Quarrie S, Dencic S, Kirby J, Iveges M (2002): Genetic diver-sity of the Novi Sad wheat core collection revealed by microsa-tellites. Cell.Mol. Biol. Lett. Vol.7, No.2B: 685-694.
Kolektiv autora (2003): Enciklopedija životne sredine i održivog razvo-ja, izdavači Ecolibri, Beograd i Zavod za udžbenike i nastavna sredstva, Srpsko Sarajevo.
Kondic-Spika A, Kobiljski B, Dencic S, Mladenov N, Hristov N, Brba-klic Lj, Trkulja D (2010): Genetic diversity in a collection of serbian wheat (Triticum aestivum L.) cultivars released in 20th century as revealed by microsatellite markers. In: Proceedings of the 12th World Congress of the International Association for Plant Biotechnology, 6-11. June 2010, St.Louis, USA, P-095.
Kermode A R J, Bewley J D , Dasgupta J, Misra S (1986): The transition from seed development to germination: A key role for dessica-tion. Hort Science 21: 1113-1118.
Koning R E (1994): Phytochrome. Plant Physiology Information Website. http://plantphys.info/plant_physiology/phytochrome.shtml. (5-3-2011)
Kranner I, Birtić S, Anderson K M, Pritchard H W (2005): Glu-tathione half-cell reduction potential: A universal stress marker and modulator of programmed cell death? Free Radical Biology and Medicine Volume 40, Issue 12, Page 2071-2244.
Krehlik L (2002):PVP Expirience of the Czsec Republic, FAO Meeting, Poznan (nepublikovano).
Kuchel H, Fox R, Reinheimer J, Mosionek L, Willey N, Bariana H, Jeffe-ries S (2007): The successful application of a marker-assisted wheat breeding strategy. Mol. Breeding. 20: 295-308.

198
Semenarstvo
Kueneman E A (1983): Genetic Contorl of Seed Longetivity in Soybean. Crop Science 23, 5-8.
Lang G A (1987): Dormancy: a new universal terminology. Hort.Scien-ce;22: 817-820.
Landjeva S, Korzun V, Börner A (2007): Molecular markers – actual and potential contributions to wheat genome characterization and breeding, Euphytica. 156: 271-296.
Madsen E, Langkilde N E (1987): Cleaning of Agricultural and Hor-ticultural Seed on Smallscale Machines, ISTA. https://www.seedtest.org/en/productdetail---1--1082--203--26.html
Manifesto M M, Schlatter A R, Hopp H E, Suárez E Y, Dubcovsky J (2001): Quantitative Evaluation of Genetic Diversity in Wheat Germplasm Using Molecular Markers, Crop Science 41:682-690, Crop Breeding, Genetics & Cytology. http://crop.scijour-nals.org/cgi/content/abstract/41/3/682
Marjanović M (2005): Testiranje tolerantnosti pšenice na bor prime-nom in vitro i tehnike molekularnih markera – mikrosatelita. Poljoprivredni fakultet, Univerzitet u Novom Sadu. Magistar-ska teza. 59 strana.
Matthews S (2008): Changes in developing pea (Pisum sativum) seeds in relation to their ability to withstand desiccation, Willey, on line library. http://onlinelibrary.wiley.com/doi/ 10.1111/j.1744-7348.1973.tb01341.x/abstract;jsessionid=D85F6E4E71FA92A8697824FC313155C9.d01t03
McCormack (2004): Seed Processing and Storage. http://www.syngen-tafoundation.org/__temp/DG_SFSA_SeedProcessingandSto-rage.pdf
Meckel L, Egli D B, Phillis A D (1980): The effect of moisture stress on seed growth in soybean. Agron. Jour, 76, 647-650.
Meints P D, Smith C A (2003): Kenaf seed storage duration on germi-nation, emergence, and yield. *Industrial Crops and Products Volume 17, Issue 1, 9-14.
Merritt D J, Turner S R, Clarke S, Dixon K W (2007): Seed dormancy and germination stimulation syndromes for Australian tempe-rate species. Australian Journal of Botany 55: 336-344.
Mihaljev I, Dokić P (1986): Semenarstvo, skripta, Poljoprivredni fakul-tet, Novi Sad.

199
Opšte semenarstvo
Miles D F, Tekrony D M, Egli D B (1988): Changes in viability, germina-tion and respiration of freshly harvested soybean seed during development. Crop. Sci. 28, 700-704.
Milošević M (1983): Mikoflora na semenu kukuruza uskladištenog u silose. Magistarski rad, Poljoprivredni fakultet, Novi Sad.
Milošević M, Mihaljev I, Ćirović M, Dokić P (1996): Opšte semenar-stvo, Feljton, Novi Sad.
Milošević M, Malešević M (2004): Semenarstvo, Institut za ratarstvo i povrtarstvo i Nacionalna laboratorija za ispitivanje semena, Novi Sad.
Milošević M, Dragin S, Stegić M (2009): Biljni genetički diverzitet u po-ljoprivredi, Poljoprivredni fakultet, Novi Sad.
Milošević M (2010): Srbija na putu evropskih integracija. Saopštenje na skupu „Srbija na putu Evropske unije“, PKS, Novi Sad.
Milošević B, Karagić Đ, Mihailović V, Mikić A, Vasiljević S, Pataki I, Vujaković M (2010): Kvalitet semena proteinskog graška u za-visnosti od vlažnosti semena u žetvi i sorte. Ratarstvo i povr-tarstvo, 47, 529-534.
Mirić M, Brkić M (2002): Dorada semena. Beograd: Društvo selekcio-nara i semenara Srbije.
Misra K M (1983): Drying of Soybean Seed. Seed Technology Conferen-ce. Iowa state University, AMES Iowa, USA.
Morton F, Colley M, Brewer L, Stone A, Navazion J (2009) Pollination and Fertilization in Organic Seed Production,Oranic agricul-ture. http://www.extension.org/article/18434
Narayana M, Kumar U M, Sun W Q (2003): Mechanism of seed age-ing under different storage condition for Vigna radiata (L.) Wilcyer: lipid peroxidation, sugar hydrolysis. J.Exp.Botany 54:1057-106.7
Naseem A, Oehmke J F, Schimmelpfennig D E (2001): Does Plant Vari-ety Intellectual Property Protection Improve Farm Productivity? Evidence from Cotton Varieties, AgBio Forum, Journal of Agro-biotechnology Management and Ecomomists vol.8, no2-3, art.6.
Neumann K, Kobiljski B, Denčić S, Börner A (2010): Genome wide asso-ciation mapping - a case study in bread wheat (Triticum aesti-vum L.). Molecular Breeding, doi: 10.1007/s11032-010-9411-7.
Norman A G (1978): Soybean physiology, agronomy and utilization. Academic Press, New York, San Francisco, London.

200
Semenarstvo
NSW (2005): Field Pea: Western NSW Planting Guide, NSW Depar-tment of Primary Indusrties. http://www.dpi.nsw.gov.au/__data/assets/pdf_file/0007/157507/field-pea-western-NSW-planting-guide.pdf
Obreht D, Kobiljski B, Denčić S, Đan M, Vapa Lj (2006): Assessing gene-tic potential for bread-making quality in wheat by marker-trait association. Cereal Research Communication. Vol 34. No 1-2: 895-902.
OECD (2003): OECD Scheme or the Control of Seed Moving in Interantio-nal Trade. http://www.oecd.org/dataoecd/31/15/40205490.pdf
OECD (2008): OECD Scheme or the Control of Seed Moving in Interantio-nal Trade. http://www.oecd.org/dataoecd/31/15/40205490.pdf
Osborn D J (1980): Senesence in seeds. In Semences in plants, ed. K.V. Thimann (Boca Raton, Florida: CRC Press, pp.13-37.
Ouda S A, Shreif M A, Elenin R A (2010): Simulation of the effect of water stress on soybean yield and water use efficiency , Four-teenth International Water Technology Conference, IWTC 14 2010, Cairo, Egypt.
Patil K B, Shivamurthy S C, Badami R C (2006): Effect on the lipid com-position of soybean during storage at different level of humi-dity. Int.Science, vol.88, issue1, 18-19.
Patnaik D, Khurana P (2001): Wheat biotechnology: A minireview. Electronic Journal of Biotechnology, 4, 2. http://www.ejbio-technology.info/content/vol4/issue2/full/4/reprint.html
Pearson K (2003): How and When Does Water Stress Impact Plant Growth and Development? American Society of Agronomy.Annual Meetings held in Denver, Colorado.
Penčić M, Dumanović J, Radović G, Jelovac D (1997): Značaj banke biljnih gena za selekciju, Selekcija i semenarsto, vol.IV, br.1-2, 7-18, Novi Sad.
Potokina E K, Vaughan E E, Tomoola N (2000): Population diversity of the Vicia satva agg. (Fabaceae) in the flora of the former USSR deduced from RAPD and seed protein analyses. Genet. Re-sources Crp Evol. 47-2, 171-183.
Probert R J, Daws M I, Hay F R (2009): Ecological correlates of ex situ seed longevity: a comparative study on 195 species, Oxford University Press. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2706723/

201
Opšte semenarstvo
Rabi A (2007): Campain for 100% Seed Treatment. http://dacnet.nic.in/ppin/SeedTreatment_Cont.htm
Rakita C, McCormac C (2004): Seed Processing and storage. Principle and practice of seed growers in the Mid-Atlantic and Southern U.S. http://www.syngentafoundation.org/__temp/DG_SFSA_SeedProcessingandStorage.pdf
Revilla P, Butrón A, Rodríguez V M, Malvar R A, Ordás A (2009): Iden-tification of genes related to germination in aged maize seed by screening natural variability, Journal of Experimental Botany Volume 60, Issue14.
Rhodes D J, Powell K A (1994): Biological Seed Treatments. The De-velopment Progress. In: Seed Treatment - Progress and Pros-pects, 303-314.
Samarah N H, Mullen R E, Goggi S, Gaul A (2009): Effect of drying tre-atment and temperature on soybean seed quality during ma-turation. Seed Science and Technology, Volume 37, Number 2, pp. 469-473(5).
Sarić M (1971): Fiziologija biljaka, Univerzitet u Novom Sadu, Novi Sad.Schwinn F J (1994): Seed treatment a panacea for crop protection. In:
Seed Treatment. Progress and Prospects. Trevor Martin. Bri-tish Crop Protection Council Cotenbury.
Setimela P S, Mongo E, Bonziger M (2004): Sucessful Community-Ba-sed Seed Production Strategies, Mexico, D. F. CIMMYT.
Sharma D L, Anderson W K (2003): The influence of climatic factors and crop nutrition on seed vigour in wheat. http://www.regio-nal.org.au/au/asa/2003/c/6/sharma.htm?print=1
Shelar W R (2008): Role of mechanical damage in deterioration od soybean seed quality during storage, Agricultural rewiews, vol 29, issue 3. http://www.indianjournals.com/ijor.aspx?target=ijor:ar&volume=29&issue=3&article=003
Siddique A B, Wright D (2003): Effects of different seed drying met-hods on moisture Percentage and Seed Quality (Viability and Vigour) of Pea Seeds (Pisum sativum L.) Pakistan Journal of Agronomy 2 (4): 201-208.
Silva R R, Cavalcanti M (2000): Multilateral Trade Negotiations On Agriculture - A resource Manual / TRIPS, IV. Agreement on Trade-Related Aspects of Intellectual Property Rights (TRIPS), FAO Corporative Repository. http://www.fao.org/documents/

202
Semenarstvo
Sivolap Y M, Solodenko A E (2006): Inter - and intraspecies differen-tiation in the genus Helianthus by RAPD analysis. CAB Inter-national. http://www.cababstractsplus.org/abstracts/Abstract.aspx?AcNo=19991603182
Slayter R O (1969): Physiological significance of internal water relation to crop yield. In Physiological Aspect of Crop Yield, J.D. Eastin Medison: Amer. Soc. Agron. 53-83.
Službeni list SFRJ (1987): Pravilnik o ispitivanju kvaliteta semena, Služ-beni glasnik SFRJ, broj 47/87.
Službeni glasnik RS (1993): Zakon o semenu i sadnom materijalu Služ-beni glasnik RS, broj 54/93.
Službeni glasnik SRJ (1998): Zakon o priznavanju sorti poljoprivrednog i šumskog bilja, Službeni glasnik SRJ, broj 12/98.
Službeni glasnik SRJ (1999): Pravilnik o zdravstvenom pregledu useva i objekata za proizvodnju semena, rasada i sadnog materijala i zdravstvenom pregledu semena, rasada i sadnog materijala, Službeni list SRJ, broj 66/99.
Službeni glasnik SRJ (2002): Zakon o zaštiti prava oplemenjivača biljnih sorti i šumskog bilja, Službeni list SRJ, broj 28/00.
Službeni glasnik SRJ (2002): Zakon o priznavanju sorti poljoprivrednog i šumskog bilja, Službeni glasnik SRJ, broj 37/02.
Službeni glasnik SRJ (2005): Zakon o sadnom materijalu voćaka, vinove loze i hmelja, Službeni glasnik RS, broj 18/05.
Službeni glasnik R.S.(2005): Zakon o semenu, Službeni glasnik RS, broj 45/05.Službeni glasnik RS (2006): Pravilnik o kontroli proizvodnje semena,
sadržini i načinu vođenja evidencije o proizvodnji rasada poljo- privrednog bilja i obrascu izveštaja o proizvodnji micelija jes-tivih i lekovitih bilja, Službeni glasnik RS, broj 60/06.
Službeni glasnik RS (2009): Zakon o zaštiti prava oplemenjivača biljnih sorti Službeni glasnik RS, broj 41/09.
Službeni glasnik RS (2009): Pravilnik o obrascu i sadržini zahteva za dodeljivanje prava oplemenjivača biljnih sorti i dokumentaciji koja se prilaže uz ovaj zahtev i načinu dostavljanja uzoraka re-produkcionog materijala sorte Službeni glasnik RS, broj 82/09.
Službeni glasnik RS (2009): Zakona o tehničkim zahtevima za proizvo-de i ocenjivanje usaglašenosti Službeni glasnik RS, broj 36/09.
Službeni glasnik RS (2009): Zakon o zdravlju bilja, Službeni glasnik RS, broj 41/09.

203
Opšte semenarstvo
Službeni glasnik RS (2009): Pravilnik o kvalitetu semena poljoprivred-nog bilja, Službeni glasnik RS, broj 23/09.
Službeni glasnik RS (2009): Zakon o genetički modifikovanim organiz-mima, Službeni glasnik RS, broj 41/09.
Službeni glasnik RS (2009): Pravilnik o sadržini i načinu vođenja Regi-stra zahteva za dodeljivanja prava oplemenjivača biljnih sor-ti, Registra zaštićenih biljnih sorti, Registra prenesenih prava oplemenjivača i Registra ugovora o licenci, Službeni glasnik RS, broj 70/09.
Službeni glasnik RS (2009): Registar ugovora o licenci Službeni glasnik RS, broj 70/09.
Službeni glasnik RS (2009): Zakon o genetički modifikovanim organiz-mima iz 2009. godine, Sl. glasniku RS br. 41/09.
Službeni glasnik RS (2010): Pravilnik o sadržini i obrascu zahteva za priznavanje sorte poljoprivrednog bilja, kao i dokumentaciji koja se uz taj zahtev prilaže, Službeni glasnik RS, broj 53/10.
Službeni glasnik RS (2010): Pravilnik o zdravstvenom pregledu useva i objekata za proizvodnju semena, rasada i sadnog materijala i zdravstvenom pregledu semena, rasada i sadnog materijala, Službenom glasniku RS, broj 107/10.
Službeni glasnik RS (2010): Pravilnik o listama štetnih organizama i li-stama bilja, biljnih proizvoda i propisanih objekata, Službeni glasnik RS, broj 7/10.
Službeni glasnik RS (2010): Pravilnik o fitosanitarnoj kontroli bilja, bilj-nih proizvoda i propisanih objekata u međunarodnom prome-tu, Službeni glasnik RS, broj 32/10.
Službeni glasnik RS (2010): Pravilnik o higijensko-tehničkim, radnim i drugim uslovima koje moraju da ispunjavaju granični prelazi na kojima postoji organizovana fitosanitarna inspekcija, Služ-beni glasnik RS, broj 37/10.
Službeni glasnik RS (2010): Pravilnik o listi vrsta poljoprivrednog bilja na koje se odnose izuzeci od prava oplemenjivača i o elementi-ma za određivanje malih poljoprivrednih proizvođača, Službe-ni glasnik RS, broj 38/10.
Službeni glasnik RS (2010): Pravilnik o utvđivanju programa mera za-štite zdravlja bilja, Službeni glasnik RS, broj 42/10.
Službeni glasnik RS (2010): Pravilnik o načinu upisa u registar pruža-laca usluga u oblasti zaštite zdravlja bilja, sadržaju i obrascu

204
Semenarstvo
zahteva, sadržini, kao i načinu vođenja tog registra, Službeni glasnik RS, broj 46/10.
Službeni glasnik RS (2010): Pravilnik o službenoj legitimaciji i službe-nom odelu fitosanitarnog inspektora, kao i načinu vođenja evi-dencije o izdatim službenim legitimacijama, Službeni glasnik RS, broj 48/10.
Službeni glasnik RS (2010): Pravilnik o uslovima u pogledu objekata, opreme i stručno osposobljenog kadra koje mora da ispunja-va pravno lice i preduzetnik koji se upisuje u registar pružalaca usluga u oblasti zaštite zdravlja bilja, u zavisnosti od vrste usluga u oblasti zaštite zdravlja bilja, Službeni glasnik RS, broj 46/10.
Službeni glasnik RS (2010): Pravilnik o utvrđivanju vrsta bilja, biljnih proizvoda i propisanih objekata koje se ne nalaze na listi VA deo I i listi VA deo II, Službeni glasnik RS, broj 64/10.
Službeni glasnik RS (2010): Pravilnik o izmenama i dopunama pravila o kvalitetu semena poljoprivrednog bilja Službeni glasnik RS, broj 72/10.
Službeni glasnik RS (2010): Zakon o priznavanju sorti poljoprivrednog bilja, Sl. glasnik RS, broj 30/10.
Službeni glasnik RS (2010): Pravilnik o potrebnoj količini semena, od-nosno sadnog materijala koji se dostavlja radi ispitivanja, kao i vreme dostavljanja tog semena, odnosno sadnog materijala, u zavisnosti od vrste bilja, Službeni glasnik RS, broj 11/10.
Službeni glasnik RS (2010): Pravilnik o načinu određivanja imena sorte poljoprivrednog bilja Službeni glasnik RS, broj 76/10.
Službeni glasnik RS (2010): Pravilnik o uslovima i načinu vršenja pre-gleda i uzorkovanja pošiljke pri uvozu, načinu najavljivanja prispeća pošiljke, obrascu i sadržini zahteva za pregled pošiljke i uslovima koje uvoznik mora da obezbedi radi obavljanja fito-sanitarnog pregleda, kao i načinu dostavljanja uzoraka, broju i veličini uzoraka radi ispitivanja i načina postupanja sa oduze-tom pošiljkom, Službeni glasnik RS, broj 86/10.
Službeni glasnik RS (2010): Pravilnik o sadržini i načinu vođenja regi-stra sorti poljoprivrednog bilja, Službeni glasnik RS, broj 73/10.
Službeni glasnik RS (2011): Pravilnik o vrstama poljoprivrednog bilja za koje se mogu vršiti dodatna ispitivanja sorte i metodama ispiti-vanja sorti tog bilja, Službeni glasnik RS, broj 6/11.
Stan O (1997): The Test of Accelerated Ageing to Estabish the Vigour Potential in Maize and Sunflower Hybrid Seed. http://www.incda-fundulea.ro/rar/nr78/rar7_12.pdf

205
Opšte semenarstvo
Škorić D, Kovačev L (1995): Germplazma u oplemenjivanju suncokre-ta i šećerne repe u narednih deset godina. Zborniku abstrakta „Prvi simpozijum za oplemenjivanje organizama“, Vrnjačka Banja, 8-11. novembar 1995.
TecAgrinews New (2003): “Intellicoat” Seed Coating Lets Corn Growers Plant 3-4 Weeks Earlier. http://www.ccimarketing.com/CCI-News/Landec/landec2.asp
Tekrony D M, Egli D B, Phillis A D (1980): The Effect of Field Weathe-ring on the viability and vigor of soybean Seed. Agron. Jour. 72, 749-753.
Tessier A H (1779): Traite des maladies des grains, La veuve Herrissant, Paris.Tillet M (1755): Dissertation sur la cause qui carrompt et moireit les
grains de bled dans les epis; et sur les moyens de prevenis ces accident. P. Bruin,Bordeaux.
Toledo A (2002): Saving the seed: Europe’s challenge, Irish Seed Savers.http://irishseedsavers.digino.virtual.tibus.net/research/2009/seeds/saving-the-seed europes-challenge-by-alvaro-toledo/
Tompsett P B (1983): The influence of gaseous environment on the storage life of Araucaria hunsteinii seed, Annual Botany, 52, 229-237.
Tošović-Marić B, Kobiljski B, Obreht D, Vapa LJ (2008): Evaluation of wheat Rht genes using molecular markers. Genetika, Vol. 40, No. 1, 24-31.
Trkulja D, Kondić-Špika A, Brbaklić Lj, Kobiljski B (2011): Analiza veze marker – svojstvo za vreme klasanja i cvetanja pšenice kori-šćenjem pojedinačne marker regresije. Ratarstvo i povrtarstvo. 48(1): 113-120.
UPOV (2002): UPOV’s instructions for carrying out tests for distinctne-ss, homogeneity and stability. Doc.No. TG/08/6.
UPOV (2009): Document C/43/12/2009. http://www.upov.int/export/si-tes/upov/en/documents/c/43/c_43_12.pdf
UPOV (2011): Members of the UPOV, International Council for the Protection of New variety of Plants. http://www.upov.int/en/about/members/pdf/pub423.pdf
Van Gastel A J G, Bishaw Z, Gregg B R (2002): Wheat Seed Production, 463-481. In Bread Wheat: Improvement and Production. (eds. B.C. Cur-tis, S. Rajaram and H. Gomez Macpherson) FAO Plant Production and Protection Series No. 30, FAO, Rome, Italy.

206
Semenarstvo
Veličković M, Budžanovski D (1987): Rezultati ispitivanja rada čupača metlica „Castry” na IPP „Banat” Kikinda. Semenarstvo, br. 2-3, 112.
Victory Seed (2009): Seed Storage and Its Affects on Quality, Viability, and Ger-mination. http://www.victoryseeds.com/information/storing.html
Wallace D H, Yan W (1998): Plant breeding and whole system crop physi-ology, improving adaptation, maturity and yield. Wallingford, UK: CAB Int.
Warwick M A (1984): Buried seeds in arable soils in Scotland, Weed Research 24, 261-268.
William M, Roca C, Ynouye I, Manrique C, Arbizu R G (2007): Indige-nous Anddean Root abd Tuber crops. In: New Food for New MIllenium. Chronica Horticulturae vol.47, no 4, 13-19.
Woodstock L W, Tao K L J (1989): Prevention of imbibitional injury in low vigor soybean embryonic axes by osmotic control of water uptake. Phys. Plant. 51, 133-139.

Milka Vujaković, Dušica Jovičić
Ispitivanje kvaliteta semena

208
UVOD
Seme obeležava početak biljne proizvodnje i zato je obezbeđivanje njegovog kvaliteta prioritet savremenog semenarstva i preduslov za vi-soke prinose svih biljnih vrsta. Izučavanje mehanizma funkcionisanja semena, zatim redovne fiziološke, biohemijske i u novije vreme mole-kularne analize semenskog materijala treba da pruže uvid u kvalitet i daju čvrstu osnovu za unapređenje oplemenjivačkih programa i jača-nje sistema kontrole u procesu semenske proizvodnje (Milošević et. al., 2007). Utvrđivanje kvaliteta semena vrši se primenom standardizova-nih metoda, koje se menjaju u skladu sa naučnim saznanjima iz oblasti fiziologije semena. Pored toga novi tehnološki procesi dorade semena mogu da utiču na promenu metode, u skladu sa zahtevima određenih biljnih vrsta. Seme posle žetve, u procesu dorade, treba da se očisti od fizičke nečistoće, semena drugog kulturnog bilja i korova, kao i od se-mena lošeg kvaliteta i odredi u partije (Basra 2006). Partija je specifična količina semena, koja se fizički može identifikovati i za koju se, na osno-vu toga, može izdati Sertifikat o ispitivanju. Sertifikat o ispitivanju je pokazatelj kvaliteta semena, a on obuhvata ispitivanje čistoće, klijavosti i sadržaj vlage, određivanje mase 1000 semena, ispitivanje zdravstvene ispravnosti i utvrđivanje životne sposobnosti semena. Da bi se ovi ele-menti utvrdili potrebno je izvršiti uzorkovanje semena.

209
Ispitivanje kvaliteta semena
UZORKOVANJE SEMENA
Svrha uzorkovanja je uzeti reprezentativan uzorak određene mase iz koje se može izvršiti ispitivanje kvaliteta semena. Pouzdanost dobi-jenih rezultata zavisi od: tačnosti sa kojom uzorak predstavlja partiju semena i preciznosti izvršenih laboratorijskih analiza (ISTA 2004).
Količina semena koja se ispituje u laboratoriji je srazmerno mala u odnosu na veličinu partije semena koju predstavlja. Uzorak se dobija iz partije semena uzimanjem malih količina semena, po principu slučaj-nosti, sa različitih mesta u partiji i njihovim kombinovanjem. Da bi se ispitivanjem dobili ujednačeni i tačni rezultati, osnovno je da se uzorci uzimaju i pripremaju pažljivo i u saglasnosti sa propisanim metodama (sl. 1). Prilikom uzorkovanja formiraju se sledeći uzorci:
Slika 1. Uzorci semena koji se formiraju u magacinu i laboratoriji
Figure 1.Samples of seed that are formed in the warehouse and laboratory
Tabela 1. Masa partije semena, prosečnog i radnog uzorka i uzorka za determinaciju drugih
semena za neke biljne vrste (ISTA, (2010) i Pravilnik o kvalitetu semena poljoprivrednog bilja
(Sl. list SFRJ 47/87))
Partija semena/Seed lot
Pojedinačni uzorci/Primary samples
Zbirni uzorak/composite sample
Uzorak za vlagu/Moisture sample Prosečan uzorak/
Submitted sample
U magacinu/In the warehouse
Ispitivanje vlage/Moisture content Broj drugih
semena/Other seed count
Radni uzorak/Working samples
Drugo seme/Other seed
Inertne materije/Inert matter
Čisto seme/Pure seed
Klijavost/Germination
Mase 1000 semena/Seed weight
U laboratoriji/In the laboratory
Slika 1. Vrste uzoraka semena koji se formiraju u magacinu i laboratoriji Figure 1. Types of samples of seed that are formed in the warehouse and laboratory

210
Semenarstvo
Tabela 1. Masa partije semena, prosečnog i radnog uzorka i uzorka za determinaciju drugih semena za neke biljne vrste (ISTA, (2010) i Pravilnik o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87))
Table 1. Mass of seed lot, average and working samples and samples for other seeds determination for some plant species (ISTA, (2010) and the Regulations on the quality of seeds of agricultural plants (Official Gazette of SFRY 47/87))
Biljn
a vr
sta
Plan
t spe
cies
Max
. mas
a pa
rtije
sem
ena
(kg)
/ M
axim
um
wei
ght
of lo
t (k
g) (I
STA
, 201
1)
Max
. mas
a pa
rtije
sem
ena
(kg)
(Sl.
list S
FRJ
47/8
7) /
Max
imum
wei
ght o
f lot
(kg
) (O
ffici
al G
azet
te o
f SFR
Y 4
7/87
)
Min
. mas
a za
pro
seča
n uz
orak
(g)
Min
imum
w
eigh
t of s
ubni
tted
sam
ple
(g)
Min
. mas
a za
radn
i uzo
rak
(g)
Min
imum
w
eigh
t of w
orki
ng s
ampl
e (g
)
Min
. mas
a za
uzo
rak
za d
eter
min
aciju
dru
gih
sem
ena
(g)
Min
imum
wei
ght
of w
orki
ng s
ampl
e fo
r ot
her s
eed
dete
rmin
atio
n (g
)
Allium cepa L 10.000 10.000 80 8 80
Aster alpinus L 5.000 5.000 20 5 -
Beta vulgaris L. 20.000 20.000 500 50 500
Daucus carota L. 10.000 10.000 30 3 30
Glycine max. (L) Merr. 30.000 20.000 1000 500 1000
Helianthus annuus L. 25.000 20.000 1000 200 1000
Hordeum vulgare L. 30.000 20.000 1000 120 1000
Medicago sativa L. 10.000 10.000 50 5 50
Pisum sativum L. 30.000 20.000 1000 900 1000
Poa annua L. 10.000 10.000 25 1 10
Secale cereale L. 30.000 20.000 1000 120 1000Sorghum bicolor (L.) Moench 30.000 10.000 900 90 900
Trifolium pratense L. 10.000 10.000 50 5 50
Triticum aestivum L. 30.000 20.000 1000 120 1000
Zea mays L. 40.000 20.000 1000 900 1000

211
Ispitivanje kvaliteta semena
– Pojedinačni (primarni) uzorak je mala količina semena uzeta iz partije semena u toku jedne manipulacije.
– Zbirni uzorak se formira sastavljanjem i mešanjem svih primarnih uzoraka uzetih iz partije semena.
– Prosečan uzorak je uzorak koji se dostavlja laboratoriji za ispitivanje. On mora biti najmanje one mase koja je navedena u Pravilniku o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87) ili u ISTA Pravilima (2011) (tab. 1) i predstavlja bilo ceo zbirni uzorak, bilo njegov poduzorak.
– Radni uzorak je poduzorak formiran u laboratoriji iz prosečnog uzorka, na kome se utvrđuje kvalitet semena.
Veličina partije, za domaći promet, ne sme da prelazi količine naznačene u Pravilniku o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87), a veličine partija za izvoz su date u ISTA Pravilima (2011), sa dozvoljenim odstupanjem od 5%.
U vreme uzorkovanja partija mora da je dorađena, tako da je što je moguće više ujednačena (homogena) i upakovana u pakovanja jednaka po masi i zatvorena jednim štepom. Svako pojedinačno pakovanje mora biti obeleženo jedinstvenom oznakom, koja je prošivena jednim štepom ili nalepljena na pakovanje. Partija semena mora biti dostupna sa svih strana. Ako neki od navedenih elemenata nije ispunjen, uzorkovanje se odbija.
Pribor za uzorkovanje semena Šiljasta sonda i njena upotreba
Šiljasta sonda se sastoji od unutrašnje (nepokretne) i spoljašnje (pokretne) cevi sa prorezima u svojim zidovima, zašiljenog vrha i ručke (sl. 2). Prečnik cevi je oko 25 mm za sve biljne vrste. Sonda se koristi za horizontalno, vertikalno ili dijagonalno uzorkovanje. Ako se sonda koristi za vertikalno uzorkovanje ona mora imati pregrade koje je dele na više odeljaka i sprečavaju mešanje semena. Bez obzira da li se koristi horizontalno ili vertikalno, sonda se dijagonalno ubada u vreću ili rinfuzu. Za seme u rinfuzi, praktičniji je vertikalni položaj sonde. Sonda je pri ubadanju u vreću zatvorena, zatim se otvori i okrene nekoliko puta ili se blago protrese da bi se potpuno napunila. Zatim se ponovo zatvori, izvlači i prazni u odgovarajući sud za seme, ili na parče impregniranog papira ili neku sličnu podlogu.

212
Semenarstvo
Posebno treba biti pažljiv prilikom zatvaranja sonde da se seme ne bi oštetilo.
Sonda se može koristiti za većinu semena, osim za seme nekih izrazito plevičastih vrsta. Do određenog prečnika cevi, može se ubadati kroz zidove vreće od grubo pletene jute ili sličnog materijala. Po izvlačenju sonde, vrhom sonde se zatvaraju otvori. Zatvorene papirne vreće takođe se mogu uzorkovati sondom, a otvori se zatvaraju specijalnom lepljivom trakom.
Slika 2. Šiljasta sonda / Figure 2. Stick trier (Jovičić, 2010)
Nobbe šilo i njegova upotreba
Nobbe šilo je zašiljena cev dovoljno duga da dopre do sredine vreće, sa ovalnim otvorom do zašiljenog kraja. Prečnik cevi je oko 10 mm za sitno seme, oko 14 mm za žita i oko 20 mm za kukuruz (sl. 3).
Nobbe šilo je pogodno za uzorkovanje semena u vrećama, ali ne i za semena u rinfuzi. Šilo se ubada u vreću pod uglom od oko 30o sa otvorom usmerenim na dole, dok ne dopre do sredine vreće. Šilo se zatim rotira za 180o tako da otvor bude usmeren na gore i lagano izvlači, s tim da se brzina izvlačenja smanjuje, da bi se količina semena u šilu postepeno povećavala od sredine ka periferiji vreće. Cev koja je dovoljno duga da može da dopre do druge strane vreće izvlači se relativno konstantnom brzinom. Dok se šilo izvlači treba ga blago protresati da se tok semena održi ujednačenim, što omogućuje glatka unutrašnja površina šila.
Uzorkovanje treba da se vrši naizmenično sa vrha, sredine i dna vreće. Otvori koji su šilom napravljeni u vrećama mogu se zatvoriti na način opisan za sonde.
Slika 3. Nobbe šilo/ Figure 3. Nobbe trier (Jovičić, 2010)

213
Ispitivanje kvaliteta semena
Uzorkovanje rukom
Uzorkovanje rukom se vrši kod plevičastog semena, kao što je: Agropyron, Agrostis, Alopecurus, Anthoxanthum, Arrhenatherum, Axonopus, Bromus, Chloris, Cynodon, Cynosurus, Dactylis, Deschampsia, Digitaria, Elymus, Elytrigia, Festuca, Holcus, Lolium, Melinis, Panicum, Pascopyrum, Paspalum, Poa, Psathyrostachys, Pseudoroegneria, Trisetum, Zoysia, kod semena koje može biti oštećeno upotrebom šila (npr. krupno seme leguminoza), seme sa izraštajima (npr. seme sa krilcima, osjem i slično) ili seme sa niskim sadržajem vlage.
Ovom metodom se uzorci mogu uzimati samo iz otvorenih pakovanja do dubine od 400 mm. Prilikom uzorkovanja prsti šake moraju biti stisnuti da ne dođe do mešanja semena unutar vreće (sl. 4).
Slika 4. Uzorkovanje rukom / Figure 4. Sampling by hand (ISTA, 2004)
Uzorkovanje automatskim uzorkovačem
Najbrži i najracionalniji način uzorkovanja za semenske kompanije je automatski uzorkovač semena. Prednost ovog načina uzorkovanja je u tome što je uzorkovanje nepristrasno, efikasno, izbegavaju se štete koje mogu nastati u toku ručnog uzorkovanja semena na plombiranim pakovanjima, štedi se vreme, a smatra se da sistemično uzorkovanje u redovnim vremenskim razmacima iz protoka mase semena bolje predstavlja partiju semena od uzorkovanja po principu slučajnosti.
Slika 5. Automatski uzorkovač / Figure 5. Automatic sampler (Shauveau and Maill, 2001)

214
Semenarstvo
Automatski uzorkovači moraju da se akredituju i proveravaju (Kojić et. al., 2010). Uzorkovači se sastoje od cevi, spirale koja prolazi kroz cev i elektromotora koji pokreće spiralu. Jedan kraj cevi se nalazi u usipnom košu u masi semena, a na drugom kraju se nalazi trofazni elektromotor. Elektromotor pokreće spiralu koja u zadatim vremenskim intervalima uzima pojedinačne uzorke iz protoka mase semena (sl. 5).
Intenzitet uzorkovanja
Intenzitet uzorkovanja zavisi od broja pakovanja i mora se strogo poštovati da bi uzeti uzorak bio reprezentativan (tab. 2). Tabela 2. Intenzitet uzorkovanja prema ISTA (2011) i Pravilniku o kvalitetu semena
poljoprivrednog bilja (Sl. list SFRJ 47/87), za partije upakovane u vrećeTable 2. The intensity of sampling according to ISTA (2011) and the Regulations on
the quality of seeds of agricultural plants (Official Gazette of SFRY 47/87), for lots packed in bags
Za pakovanja od 15 kg do 100 kgFor packing of 15 up to 100 kg
(ISTA, 2011)
Za partije u vrećamaFor lots packed in bags
(Sl. list SFRJ 47/87)
Br. pakov./Number of packings
Broj pojedinačnih uzorakaNumber of primary samples
Br. pakov./Number of packings
Broj pojedinačnih uzorakaNumber of primary samples
1–4
3 pojedinačna uzorka iz svakog pakovanja3 primary samples from each container
1–5
1 uzorak iz svakog pakovanja, ali ne manje od 5 pojedinačnih uzoraka1 primary sampl from each packing but not less than 5
5–8
2 pojedinačna uzorka iz svakog pakovanja2 primary samples from each container
6–30
1 uzorak iz svakog trećeg pakovanja, ali ne manje od 5 pojedinačnih uzoraka1 primary sampl from each of the third package but not less than 5
9–15
1 pojedinačan uzorak iz svakog pakovanja1 primary samples from each container
31–400
1 uzorak iz svakog petog pakovanja, ali ne manje od 10 pojedinačnih uzoraka/1 primary sampl from each of the fifth package but not less than 10
16–30
15 pojedinačnih uzoraka iz partije semena15 primary saples in total from the seed lot
>400
1 uzorak iz svakog sedmog pakova-nja, ali ne manje od 80 pojedinačnih uzoraka1 primary sampl from each of the seventh package but not less than 80
31–5920 pojedinačnih uzoraka iz par-tije semena/20 primary saples in total from the seed lot
>6030 pojedinačnih uzoraka iz partije semena/30 primary saples in total from the seed lot

215
Ispitivanje kvaliteta semena
Za seme u pakovanjima manjim od 15 kg, pakovanja treba kom-binovati u jedinice za uzorkovanje koje ne prelaze 100 kg i te jedinice treba, prema napred prikazanom intenzitetu, uzorkovati.
Pri uzorkovanju semena iz pakovanja preko 100 kg i semena u rin-fuzi intenzitet uzorkovanja je dat u tabeli 3.Tabela 3. Intenzitet uzorkovanja prema ISTA, (2011) i Pravilniku o kvalitetu semena poljo-
privrednog bilja (Sl. list SFRJ 47/87), za pakovanja preko 100 kg i seme u rinfuziTable 3. The intensity of sampling according to ISTA (2011) and the Regulations on
the quality of seeds of agricultural plants (Official Gazette of SFRY 47/87), for packages over 100 kg of seed in bulk
Veličina partijeSize of lot (kg)
Broj pojedinačnih uzorakaNumber of primary samples
< 500 Najmanje pet pojedinačnih uzoraka/At least 5 primary samples
501–3.000 Jedan pojedinačan uzorak na svakih 300 kg, ali ne manje od 5 uzoraka / One primary sample for each 300 kg, but not less than 5
3.001–20.000 Jedan pojedinačan uzorak na svakih 500 kg, ali ne manje od 10 uzoraka / One primary sample for each 500 kg, but not less than 10
>20.001 Jedan pojedinačan uzorak na svakih 700 kg, ali ne manje od 40 uzoraka / One primary sample for each 700 kg, but not less than 40
Masa prosečnog uzorka
Najmanja masa prosečnih uzoraka definisana je Pravilnikom o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87), za domaći promet i ISTA Pravilima za seme namenjeno međunarodnom prome-tu. Masa prosečnog uzorka, za neke biljne vrste, je navedena u tabeli 1. Prosečan uzorak mora da bude upakovan u platnenu ili papirnu kesu i obeležen etiketom na kojoj mora da stoji oznaka partije, biljna vrsta i sorta/hibrid.
Masa uzorka za određivanje sadržaja vlage u semenu je 50 g ili 100 g. Za biljne vrste koje se melju prilikom određivanja sadržaja vlage masa je 100 g, a za biljne vrste koje se ne melju je 50 g. Uzorak za vlagu mora da bude upakovan u PVC ili staklenu ambalažu da ne bi došlo do usva-janja ili odavanja vlage iz semena.
Upakovan uzorak za vlagu se stavlja u prosečan uzorak. Prosečan uzo-rak se zatvara, obeležava i dostavlja u laboratoriju za ispitivanje semena.

216
Semenarstvo
PRIPREMA RADNOG UZORKA U LABORATORIJI
U laboratoriju za ispitivanje semena dostavlja se prosečan uzorak od koga se, jednom od metoda za polovljenje uzoraka, formira radni uzorak. Metode koje se koriste za pripremu radnog uzorka moraju da obezbede da svako seme iz uzorka ima istu verovatnoću da bude izabrano (Bould 1986). Metode koje se koriste su metod mehaničkog i ručnog deljenja uzoraka. Navedene metode su opisane od strane ISTA (2011).
Metod mehaničkog deljenja uzorka
Ovaj metod je pogodan za sve vrste semena, izuzev za ekstremno plevičasto seme. Princip rada delitelja je da uzorak koji prolazi kroz njega, deli uzorak na dva ili više približno jednakih delova. Uzorak se smanjuje uklanjanjem jedne polovine uzorka pri svakom propuštanju kroz delitelj. Ovaj proces se nastavlja dok se ne dobije približna masa radnog uzorka ili uzorka za određivanje drugih semena (tab. 1). Kao pogodni smatraju se sledeći delitelji: konusni delitelj, zemljišni delitelj, centrifugalni delitelj, rotacioni delitelj i varijabilni delitelj.
Konusni delitelj
Konusni delitelj (tipa Boerner) se proizvodi u dve veličine, manji za vrste sa sitnijim semenom, i veći za vrste sa krupnijim semenom (seme veličine pšenice i veće). Osnovni delovi su: levak i niz skretnih lopatica, raspoređenih u krug, koje usmeravaju seme u dva kanala za ispuštanje (sl. 6). Nedostatak ovog delitelja je što je teško proveravati njegovu čistoću.

217
Ispitivanje kvaliteta semena
Zemljišni delitelj
Delitelj napravljen po istom principu kao i konusni je takozvani ze-mljišni delitelj. Kanali su postavljeni u pravom redu. Zemljišni delitelj se sastoji od levkastog suda, pregrada za deljenje (pregrade su različite debljine u zavisnosti od krupnoće semena) i dve posude za prijem semena (sl. 7).
Prilikom korišćenja delitelja seme treba ravnomerno isipati iz lev-ka, u približno jednakim količinama duž čitave njegove dužine. Ovaj de-litelj je pogodan za krupnosemene i plevičaste vrste, ali se takođe mogu napraviti i tipovi pogodni za sitnose-mene vrste.
Slika 6. Konusni delitelj / Figure 6. The conical divider (Boerner type) (Vujaković, 2004)
Slika 7. Zemljišni delitelj Figure 7. The soil riffle divider
(Vujaković, 2004)
levak/hopper
ventil/valve
kupa/cone
levak koji vodi u posudu na levoj strani funnel to container on left side
posude za uzorke sample container

218
Semenarstvo
Centrifugalni delitelj
Centrifugalni delitelj (tipa Gamet) koristi centrifugalnu silu za me-šanje i raspoređivanje semena preko razdelne površine. Kod ovog deli-telja seme curi silazno kroz levak za punjenje do plitke gumene kupe ili prstena. Posle rotacije prstena, koga pokreće električni motor, seme se izbacuje centrifugalnom silom i pada (sl. 8). Krug ili površina na koju seme pada je podeljen na dva jednaka dela pomoću fiksirane skretne lopatice tako da približno polovina semena pada u jedan, a druga polo-vina u drugi otvor za ispuštanje.
Centrifugalni delitelj pokazuje različite rezultate ako se njime ne rukuje pažljivo.
Slika 8. Centrifugalni delitelj / Figure 8. The centriphugal divider (Vujakovic, 2004)
Rotacioni delitelj
Rotacioni delitelj ima rotacionu jedi-nicu od 6 do 10 posuda, vibraciono grlo i levak. Seme se sipa u levak, rotacioni deli-telj uključi, tako da se rotaciona jedinica okreće oko 100 o/min (sl. 9). Brzina pu-njenja i trajanje deljenja se može regulisati
Slika 9. Rotacioni deliteljFigure 9. The rotari divider (Jovičić, 2010)
zavrtnji za podešavanje
screwelektromotorelectric motor
posuda za uzoraksample container
izlazna cev stationary baffle
disk spirit level
levakhopper
prekidačswitch

219
Ispitivanje kvaliteta semena
podešavanjem razdaljine između kanala otvora levka i vibracionog grla i podešavanjem intenziteta vibracija.
Delitelj je pogodan za sitnosemene vrste i za većinu plevičastih semena (npr. trave i cveće). Samo izrazito plevičasto seme (npr. Tri-setum flavescens) se ne može deliti ovim deliteljem jer se zaglavljuje u levku.
Varijabilni delitelj
Varijabilni delitelj dozvoljava promenu odnosa deljenja, u zavisno-sti od veličine semena, koji može biti između 1:2 i 1:15. Na ovaj način se jednom operacijom redukuje uzorak poznate mase do potrebne mase radnog uzorka (poduzorak). Sastoji se od usipnog levka i tube koja se okreće sa oko 40 o/min. Tuba distribuira protok semena od usipnog lev-ka na unutrašnju površinu drugog levka koji je dobro uklopljen u treći levak. Levkovi su koncentrično postavljeni. U drugom i trećem levku se nalaze kanali koji zauzimaju 50% ivice levka. 50% semena će proći kroz dva levka u sud koji sakuplja seme. Drugih 50% će ostati unutar levkova i otići u drugi sud za seme. Dva levka se mogu okrenuti jedan nasuprot drugom, čime se kanali sužavaju, što rezultira prolaskom manje količine semena kroz njih. Kao traženi uzorak se može koristiti bilo manji uzo-rak izvan levka, bilo veći uzorak unutar levka. Položaj dva levka jednog u odnosu na drugi se može precizno podesiti, čime se unapred određuje veličina poduzorka.
Metod ručnog deljenja uzoraka
Za pripremu radnog uzorka jako plevičastog semena i semena sa puno primesa, metod mehaničkog deljenja uzoraka nije pogodan pa se preporučuje upotreba jednog od metoda ručnog deljenja. Isto tako pri-prema radnog uzorka za ispitivanje zdravstvene ispravnosti semena se vrši metodom ručnog deljenja, da ne bi došlo do zaražavanja drugog uzorka semena. Metode ručnog deljenja su: modifikovan metod polov-ljenja, metod kašike i metod ručnog polovljenja.

220
Semenarstvo
Modifikovani metod polovljenja
Oprema koja se koristi je tacna i rešetka sa četvorouga-onim pregradama iste veličine, koje su otvorene na vrhu i svaka druga nema dno (sl. 10). Posle mešanja, seme se ravnomerno sipa preko rešetke. Kada se re-šetka podigne, približno po-lovina uzorka ostane na tacni. Prosečan uzorak se sukcesivno polovi dok se ne dobije radni uzorak približne, ali ne manje od propisane mase.
Metod kašike
Ovaj metod se kori-sti za smanjenje uzorka za vlagu ili uzorka za ispitivanje zdravstvenog stanja semena. Potrebni su tacna, špatula i kašika sa ravnim ivicama. Po-sle prethodnog mešanja, seme se ravnomerno sipa preko tacne. Sa kaši-kom u jednoj i špatulom u drugoj ruci, treba uze-ti malu količinu semena sa najmanje pet nasu-mično izabranih me- sta (sl. 11). Uzeta količi-na mora da bude približ-ne, ali ne manje mase od propisane.
Slika 10. Modifikovani metod polovljenjaFigure 10. Modified method of dividing
(Mathuer, Kongsdal, 2003)
Slika 11. Metod deljenja kašikomFigure 11. Spoon method (Vujaković, 2004)

221
Ispitivanje kvaliteta semena
Metod ručnog polovljenja
Ovaj metod se primenjuje kod sledećih rodova plevičastog semena: Agrimonia, Andropo-gon, Anthoxanthum, Arrhenathe-rum, Astrebla, Beckmanni, Boute-lou, Brachiari, Briz, Cenchrus, Chloris, Dichanthium, Digitaria, Echinohcloa, Ehrharta, Elymus, Eragrostis, Melinis, Oryza, Penni-setum, Psathyrostachys, Scabiosa, Sorghastrum, Stylosanthes, Tae-niatherum, Gomphrena i Trise-tum. Za sve druge vrste može se koristiti samo za dobijanje rad-nog uzorka u laboratoriji za ispi-tivanje zdravstvenog stanja. Seme se ravnomerno sipa na glatku čistu površinu. Pomoću špatule ravnih ivica seme se izmeša, po-tom se deli na dva dela i svaka polovina se ponovo polovi da bi se dobila četiri dela (sl. 12). Svaki od ova četiri dela se ponovo polo-vi dajući osam delova. Svaki dru-gi deo se uzima (npr. 1, 3, 5 i 7) i postupak se ponavlja dok se ne dobije propisana masa.
Slika12. Metod ručnog polovljenjaFigure 12. Hand halving method
(Jovičić, 2010)

222
Semenarstvo
ISPITIVANJE ČISTOĆE SEMENA
Čistoća semena je značajni pokazatelj kvaliteta semena. Ispitivanje čistoće semena se vrši na random uzorku koji je pripremljen jednom od navedenih metoda za pripremu radnog uzorka. Masa radnog uzorka za čistoću semena je definisana Pravilnikom o kvalitetu semena poljopri-vrednog bilja (Sl. list SFRJ 47/87), za domaći promet i ISTA Pravilima za seme namenjeno međunarodnom prometu (tab. 1). Za izuzetno skupo seme i za male partije semena prihvata se i masa semena koja sadrži najmanje 2500 semena.
Prilikom određivanja čistoće semena iz radnog uzorka određuje se procenat čistog semena, procenat drugog semena i procenat inertnih materija (sl. 13).
Slika 13. Komponente čistoće semena / Figure 13. Components of seed purity
Čisto seme predstavlja seme vrste koju je korisnik naveo, ili seme vrste za koju je
utvrđeno da u ispitivanju preovlađuje, i treba da sadrži sve botaničke varijetete i kultivare date
vrste, zatim nedovoljno zrelo, zakržljalo, smežurano, zaraženo i proklijalo seme. U čisto seme
ulazi i deo semena veći od polovine njegove originalne veličine. Kod familije Poaceae
(Gramineae) čisto seme su klasići kod kojih kariopsa sadrži endosperm i slobodne kariopse. Kod
vrsta Triticum spelta, Avena, Bromus, Dactylis, Festuca, x Festulolium, Koeleria i Lolium
određuje se procenat višesemenih jedinica (MSU) prema slici 14 i sterilni cvetići se ne uklanjaju.
Čistoća semena/Purity testing
Čisto seme/Pure seed
Drugo seme/Seed of other species
Inertne materije/Inert matter
Seme drugog gajenog bilja/Other crops seed
Seme korova/Weed seed
Slika 13. Komponente čistoće semena Figure 13. Components of seed purity (Vujaković, 2004)

223
Ispitivanje kvaliteta semena
Čisto seme predstavlja seme vrste koju je korisnik naveo, ili seme vrste za koju je utvrđeno da u ispitivanju preovlađuje, i treba da sadrži sve botaničke varijetete i kultivare date vrste, zatim nedovoljno zrelo, zakržljalo, smežurano, zaraženo i proklijalo seme. U čisto seme ulazi i deo semena veći od polovine njegove originalne veličine. Kod familije Poaceae (Gramineae) čisto seme uključuje klasiće kod kojih kariopsis očigledno sadrži endosperm i slobodne kariopse. Kod vrsta Triticum spelta, Avena, Bromus, Dactylis, Festuca, x Festulolium, Koeleria i Lolium procenat višesemenih jedinica (MSU) određuje se prema slici 14, pri čemu se sterilni cvetići ne uklanjaju.
Slika 14. Višesemene jedinice (šrafirane površine predstavljaju fertilne cvetiće, dok nešravfirane površine predstavljaju sterilne cvetiće)
Figure 14. Multiple seed units (stippled portion represents fertile flowers, clear portion represents sterile flowers) (ISTA, 2011)
Pod drugim semenom se podrazumevaju jedinice semena bilo koje biljne vrste drugačije od date vrste čistog semena. Za determinaciju dru-gih semena koriste se kolekcije semena i razni atlasi od kojih su neki „Poznavanje semena najčešćih korova u semenarstvu” (Kronaveter and Pal 1994), „Semena sornih rastenij” (Dobrokov 1961) i „Manual for Te-sting Agricultural and Vegetable Seeds” (Agriculture Handbook 1952).
Inertne materije obuhvataju jedinice semena i sve druge materije i strukture koje nisu definisane kao čisto ili drugo seme, kao što su: delovi semena manji od polovine semena originalne veličine, semenjača, de-lovi semenjače, oljuštena zrna–kod suncokreta preko 1%, drugi delovi biljke, primese koje nisu seme, seme bez klubeta kod vrsta Beta sp., ple-ve i plevice kod trava, seme Cuscuta sp. koje je lomljivo ili pepeljastosivo do kremasto belo, seme Fabaceae (Leguminosae), Brassicaceae (Cruci-ferae), Cupressaceae, Pinaceae, Taxaceae i Taxodiaceae bez semenjače, zemlja, kamenčići, pesak, sklerocije gljiva i uginuli insekti.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
SINGLE MSU

224
Semenarstvo
Rezultati za čistoću semena izražavaju se u procentima na jedno decimalno mesto. Pravilnikom o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87) definisane su minimalne vrednosti za čistoću seme-na i maksimalne vrednosti za prisustvo gajenog bilja i korova koje seme treba da ispunjava da bi se stavilo u promet (tab. 4).
Tabela 4. Norme kvaliteta za neke gajene biljne vrste (Sl. list, SFRJ, 47/87)Table 4. Quality standards of some cultivated plant species
Biljna vrstaPlant species
Minimalna čistoća semena
Milnimal seed
purity(%)
Maksi-malno
prisustvo drugih vrsta
Maximal presence of other species
(%)
Maksi-malno
prisustvo korovskih
vrstaMaximal
presence of weed
species (%)
Minimalna klijavost semena
Minimal seed ger-mination
(%)
Maksi-malan sadržaj vlage u semenu
Maximal seed mois-ture con-
tent(%)
Allium cepa L. 96 0,2 0,3 65 11
Avena sativa L. 97 0 0 82 15
Beta vulgaris L. 97 0,3 0,1 80 15
Capsicum spp. 97 0 0 65 12
Festuca rubra L. 90 3 1 70 13
Glycine max (L) Merr. 96 0 0 75 14
Helainthus annuus L. 97 0 0 80 11
Lolium perenne L. 94 2 1 70 13Lycopersicum esculentum Mill. 97 0 0 75 12
Medicago sativa L. 95 2 0,5 70 13
Oryza sativa L. 96 0,5 0,2 85 14
Phaseolus vulgaris L. 97 0 0 70 14
Poa annua L. 82 2 1 65 12
Spinacia oleracea L. 94 0,2 0,5 65 13
Triticum aestivum L. 97 0 0 88 14
Vicia faba L. 97 0 0 75 15
Zea mays 98 0 0 90 13

225
Ispitivanje kvaliteta semena
ISPITIVANJE KLIJAVOSTI SEMENA
Klijavost semena je najbitnija komponenta kvaliteta semena. Cilj is-pitivanja klijavosti je da utvrdi maksimalni potencijal klijavosti partije semena, koji se zatim može iskoristiti za upoređivanje kvaliteta različi-tih partija, kao i za procenu potencijala za nicanje u polju.
Ispitivanje u poljskim uslovima nije pouzdano, pošto poljski uslovi nisu ponovljivi. Zbog toga su razvijene laboratorijske metode, u kojima su spoljašnji uslovi kontrolisani. Uslovi su standardizovani da bi rezul-tati bili u granicama ponovljivosti.
Klijavost semena u laboratorijskim ispitivanjima predstavlja nicanje i razvoj klijanaca do stadijuma u kome njegove osnovne strukture uka-zuju na to da li je ili nije sposoban da se pod odgovarajućim uslovima u zemljištu dalje razvije u normalnu biljku.
Mirovanje semena
Mirovanje ili dormantnost semena (lat. dormio – spavati, mirovati) je pojava pri kojoj seme ne klija ni u povoljnim uslovima sredine (Grbić 2003). Uzroci dormantnosti mogu biti vrlo različiti, a ona se može javiti kao primarna ili sekundarna. Tek ubrano seme odlikuje se primarnom dormantnošću, dok se sekundarna dormantnost može pojaviti zbog ne-povoljnog delovanja nekog spoljašnjeg faktora na seme koje je „preva-zišlo“ blokade primarne dormantnosti. Ovaj tip dormantnosti može se otkloniti zasecanjem ili oštećivanjem omotača jer će na taj način usvaja-nje vode biti omogućeno.
Nikolaeva (1977) u okviru dormantnosti razlikuje sledeće grupe:I. Primarna dormantnostA. Egzogena dormantnost (izazvana uticajem faktora van embriona):1. fizička,2. mehanička,3. hemijska.B. Endogena dormantnost (izazvana faktorima unutar embriona):

226
Semenarstvo
1. morfološka, 2. fiziološka, 3. kombinovana:a. morfološko–fiziološka,b. fiziološko–fizička dormantnost.II. Sekundarna dormantnost
Primarana dormantnost
Egzogena dormantnost Fizička dormantnosti prouzrokovana je nepropustljivošću omota-
ča (semenjače, perikarpa i/ili endosperma) za vodu i kiseonik, pa seme može proklijati tek nakon omekšavanja ili propadanja semenjače. Ka-rakterističan je za leguminoze, čije seme obično ima mali sadržaj vlage i semenjaču koja je nepropustljiva za vodu.
Mehanička dormantnost nastaje kada je semenjača toliko čvrsta da stvara mehanički otpor rastu embriona, što usporava klijanje. Sama mehanička dormantnost je retka i obično se javlja zajedno sa drugim oblicima dormantnosti. Ovaj tip mirovanja semena obično je karakte-rističan za semena drvenastog i žbunastog bilja (Gleditsia spp., Robinia spp.) i u prirodnim uslovama se prevazilazi delovanjem različitih fak-tora (mikroorganizmi, prolaskom semena kroz crevni trakt životinja, šumskim požarima i slično).
Hemijska ili inhibitorna dormantnost nastaje usled prisustva ra-zličitih inhibitora u semenu kao što su fenoli, apcisinska kiselina, kuma-rin i slično. Ove štetne materije se iz semena mogu ukloniti ispiranjem ili potapanjem u vodi.
Endogena dormantnostFiziološka dormantnost semena je najzastupljeniji oblik, koji se
dalje može podeliti na tri nivoa: dugotrajna (duboka), prelazna i krat-kotrajna dormantnost (Baskin and Baskin, 2004). Duboku fiziološku dormantnost odlikuje nemogućnost klijanja ili razvoj abnormalnih kli-janaca (nanizam) ukoliko seme nije bilo izloženo određeno vreme hlad-noj (tip a) ili toploj (tip b) stratifikaciji.
Morfološku dormantnost se često sreće u semenu čiji je embrion nedovoljno razvijen, ali normalno diferenciran. Takav embrion nije fizi-ološki dormantan već zahteva određeno vreme za rast i klijanje.

227
Ispitivanje kvaliteta semena
Kombinovana dormantnost Morfološko-fiziološka dormantnost se takođe javlja u semenu čiji
embrion nije dovoljno razvijen, ali koje pored toga sadrži i fiziološke komponente dormantnosti (Baskin and Baskin, 2004).
Fiziološko-fizička domantnost javlja se u semena, koje pored fizi-ološki dormantnog embriona, ima i omotač slabo propustljiv za vodu i gasove.
Sekundarna dormantnost
Sekundarna dormantnost nastaje nakon odvajanja semena od bilj-ke, usled delovanja nepovoljnih uslova sredine, kod semena koja su već savladala primarnu klijavost.
Faktori koji kontrolišu mirovanje semenaNivo dormantnosti je veoma varijabilan i zavisi od nekoliko činilaca:- genotipa,- uticaja faktora spoljašnje sredine, - interakcije genotipa i faktora spoljašnje sredine,- hormona i- prisustva mikroorganizama na semenu (Milošević et. al., 1996a,
Pržulj et. al., 1999 prema Barjaktarović et. al., 2004).Faktori spoljašnje sredine tokom perioda sazrevanja semena mogu
povećati ili smanjiti nivo mirovanja semena. Mnogi autori smatraju da je, pored vode i dužine trajanja fotoperioda, temperatura veoma važan faktor sredine koji ima uticaja na dormantnost semena (Schuurink et al. 1992). Ranije se smatralo da temperatura reguliše dormantnost i klijanje, a svetlost samo klijanje semena. Međutim, novija istraživanja ukazuju na značajan uticaj svetlosti i na mirovanje semena (Baskin and Baskin 2004). Prema tome, na prekid mirovanja semena bitno utiču ekološki činioci kao što su temperatura, vlaga, kiseonik i svetlost, a uslovi za pre-kid ove pojave značajno se razlikuju među pojedinim biljnim vrstama.
Niske temperature tokom sazrerevanja semena indukuju duboku i produženu dormantnost, dok niske temperature tokom klijanja je pre-kida. Smatra se da niske temperature tokom klijanja utiču na propust-ljivost semenjače za kiseonik. Istovremeno, niske temerature smanjuju intenzitet disanja i na taj način kiseonik postaje dostupan za druge pro-cese (Nyachiro 2002).

228
Semenarstvo
Metode za prekidanje mirovanja kod semena
Kod određenih biljnih vrsta seme mora da prođe period fiziološkog dozrevanja da bi bilo sposobno da klija. Dužina perioda fiziološkog do-zrevanja je različita i zbog toga je u laboratoriji razvijeno više metoda za njegovo prekidanje.
Suvo skladištenje se primenjuje kod vrsta kod kojih period mirova-nja prirodno kratko traje. Dovoljno je čuvanje uzorka na suvom mestu tokom kratkog vremenskog perioda.
Prethodno hlađenje se koristi kod semena poljoprivrednog bilja i cveća, gde je period mirovanja nešto duži. Pravilnikom o kvalitetu se-mena poljoprivrednog bilja (Sl. list SFRJ 47/87) i ISTA Pravilima (2011) definisano je kod kojih vrsta se koriste metode za prekidanje mirovanja semena (tab. 3). Seme se stavi na vlažan filter papir ili u pesak i izlaže temperaturi 5–10oC u periodu od 3 do 7 dana. U nekim slučajevima je neophodno da se period prethodnog hlađenja produži ili da se izvrši ponovno hlađenje. Seme drveća i žbunja se prethodno hladi obično na temperaturi 1–5oC, zavisno od vrste, u trajanju od 2 nedelje do 12 me-seci, pre ispitivanja klijavosti, ali treba paziti da se izbegne smrzavanje semena. Za seme za koje je potreban dug period prethodnog hlađenja, a ispitivanje klijavosti ne može da se završi za 2 meseca, preporučuje se tetrazolijum test.
Prethodno zagrevanje se vrši na temperaturi od 30–35oC uz cirku-laciju vazduha, u periodu od 3 do 7 dana, pre stavljanja na podlogu za ispitivanje. U nekim slučajevima potrebno je ovaj period prethodnog zagrevanja produžiti. Za određene tropske i suptropske vrste prethodno zagrevanje može da se vrši na temperaturi 40–50oC (npr. Arachis hypo-gea: 40oC; Oryza sativa: 50oC).
Svetlost bi trebala da se koristi kod svih uzoraka koji se ispituju u trajanju od 8 časova dnevno, a kod naklijavanja na temperaturi 20-30oC tokom perioda visoke temperature. Za prekidanje mirovanja semena svetlost se preporučuje naročito za određene tropske i suptropske trave (npr. Chloris gayana, Cynodon dactylon). Intenzitet svetlosti koji daje hladna bela lampa treba da je oko 750–1250 luxa.

229
Ispitivanje kvaliteta semena
Kalijum nitrat (KNO3) se koristi umesto vode, na početku ispiti-vanja. Podlogu koja se koristi za naklijavanje navlažiti 0,2% rastvorom KNO3.
Giberelinska kiselina (GA3) se preporučuje za Avena sativa, Horde-um vulgare, Secale cereale, x Triticosecale, Triticum aestivum , Valeriane-lla locusta i duge vrste u skladu sa Pravilnikom. Podloga koja se koristi za naklijavanje se navlaži sa 0,05% rastvorom GA3. Kada je period miro-vanja kraći dovoljan je 0,02% rastvor GA3, a kada je period duži koristi se 0,1% rastvor. Kada se traži koncentracija veća od 0,08% preporučuje se rastvaranje GA3 u rastvoru fosfatnog pufera. Puferni rastvor se pri-prema rastvaranjem 1,7799 g Na2HPO4 x 2 H2O i 1,3799 g NaH2PO4 x H2O u jednoj litri destilovane vode.
Plombirane polietilenske „koverte” se koriste tamo gde se nakon standardnog ispitivanja utvrdi visok udeo svežeg neklijalog semena (npr. kod Trifolium spp.) preporučuje se ponovno ispitivanje u plombi-ranim polietilenskim kovertama, čija veličina treba da bude dovoljna da bi se ispitivanje izvelo na zadovoljavajući način. Ovaj način ispitivanja obično provocira klijanje takvog semena.
Metode za uklanjanje tvrdosemenosti koristi se kod vrsta koje imaju tvrdu semenjaču, a može da se koristi metod natapanja, mehaničke ska-rifikacije i skarifikacija kiselinom.
- Natapanje u vodi u trajanju 24–48 sati. Na primer Avena sativa se potapa u skoro ključalu vodu zapremine tri puta veće od za-premine samog semena i drži u vodi do njenog hlađenja. Ispiti-vanje klijanja započinje odmah nakon natapanja.
- Mehaničkom skarifikacijom vrši se pažljivo probijanje, rasceplji-vanje, struganje ili šmirglanje semenjače pomoću peska. Treba voditi računa da se semenjača probije na odgovarajućem mestu kako bi se izbeglo oštećenje embriona i samim tim klijanca koji će se iz njega razviti. Najbolji položaj za mehaničku skarifikaciju je odmah iznad vrhova kotiledona.
- Skarifikacija kiselinom koristi se koncentrovana sumporna ki-selina (H2SO4) za neke vrste (npr. Macroptilium sp., Brachiaria sp.). Seme se potapa u kiselinu dok opna ne postane rupičasta. Potapanje traje nekoliko minuta do 1 sata. Posle potapanja, a pre ispitivanja klijavosti, seme treba dobro isprati u tekućoj vodi.

230
Semenarstvo
Kod Oryza sativa skarifikacija može da se izvrši natapanjem se-mena u 1M azotnoj kiselini (HNO3) u trajanju od 24 časa (posle prethodnog zagrevanja na 50oC).
Metode za uklanjanje inhibitornih supstanci se koristi kod vrsta koje u perikarpu ili semenjači sadrže različite inhibitore klijanja. Može da se koristi metod prethodnog ispiranja i metod uklanjanja struktura oko semena.
- Prethodno ispiranje se koristi kod vrsta (npr. Beta vulgaris) gde su inhibitori klijanja prisutni u perikarpu ili semenjači. Seme se ispira u tekućoj vodi na temperaturi od 25oC pre nego što se iz-vrši ispitivanje klijavosti. Posle ispiranja seme treba da se osuši na temperaturi od najviše 25oC.
- Uklanjanje struktura oko semena se koristi kod nekih Poaceae vrsta gde se uklanjaju spoljašnje strukture kao što su čekinja ili pleve i plevice.
Podloge za ispitivanje klijavosti semena
Podloge koje se koriste za ispitivanje semena, temperatura naklija-vanja i dužina ispitivanja su propisane Pravilnikom o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87) i ISTA Pravilima (2011) (tab. 5). Dužina ispitivanja se može produžiti za ½ od dužine trajanja ispi-tivanja. Vreme potrebno za prekidanje perioda mirovanja pre i u toku ispitivanja se ne uključuje u dužinu ispitivanja.
Za ispitivanje semena kao podloga koristi se filter papir, pesak i kompost.
Metod sa filter papirom
Filter papir, kao podloga, ne sme da bude zaražen gljivama, bak-terijama i toksičnim supstancama koje mogu da utiču na rast ili ocenu klijanaca. Tekstura filter papira treba da je porozna, kako bi se korenov sistem razvijao na filter papiru, a ne kroz papir. Filter papir treba da ima takav kapacitet za vlagu da zadržava dovoljnu količinu vode za ceo peri-od ispitivanja i na taj način obezbedi kontinuirano snabdevanje semena vlagom. pH vrednost filter papira treba da bude 6,0–7,5.

231
Ispitivanje kvaliteta semena
Tabe
la 5.
Uslo
vi za
ispi
tivan
je kl
ijavo
sti se
men
a gaje
nih
bilja
ka /
Tabl
e 5. T
erm
s for
seed
germ
inat
ion
testi
ng o
f cul
tivat
ed p
lant s
pecie
s (IS
TA, 2
010)
Biljn
a vr
sta
Plan
t spe
cies
Podl
oga
Med
ium
Tem
pera
tura
Tem
pera
ture
(º
C)
Traj
anje
ispi
tivan
ja
(dan
i)/D
urat
ion
of
test
ing
(day
s)Po
stup
ak z
a pr
ekid
anje
stan
ja m
irov
anja
Proc
edur
e fo
r dor
man
cy re
mov
ing
Alliu
m ce
pa L
.N
F; IF
; P20
; 15
6-12
Pret
hodn
o hl
ađen
je/p
rech
llAv
ena
sativ
a L.
IF; P
205-
10Pr
etho
dno
grej
anje
/pre
heat
(30-
35ºC
)Pr
etho
dno
hlađ
enje
/pre
chll
Beta
vul
gari
s L.
NF;
IF; P
20-3
0;15
-25;
204-
14Is
pira
nje
(mul
iger
mno
sem
e: 2
h; m
onog
erm
no se
me:
4 h
)/Pr
ewas
h (m
ultig
erm
: 2 h
; gen
etic
mon
oger
m: 4
h)
Suše
nje
na m
ax 2
5ºC
/ dr
y at
a m
axim
um o
f 25
ºCCa
psicu
m sp
p.N
F; IF
; P20
-30
7-14
KN
O3
Festu
ca ru
bra
L.N
F20
-30;
15-2
57-
21Pr
etho
dno
hlađ
enje
/pre
chll
KN
O3
Gly
cine m
ax (L
.) M
err.
IF; N
FP; P
20-3
0;25
5-8
–
Hela
inth
us a
nnuu
s L.
IF; N
FP; P
; S20
-30;
25;
204-
10Pr
etho
dno
grej
anje
/pre
heat
Pret
hodn
o hl
ađen
je/p
rech
ll
Loliu
m p
eren
ne L
.N
F20
-30;
15-2
5; 2
05-
14Pr
etho
dno
hlađ
enje
/pre
chll
KN
O3
Lyco
persi
cum
escu
lentu
m M
ill.
NF;
IF; P
20-3
05-
14K
NO
3M
edica
go sa
tiva
L.N
F; IF
204-
10Pr
etho
dno
hlađ
enje
/pre
chll
Ory
za sa
tiva
L.N
F; IF
; P20
-30;
255-
14Pr
etho
dno
grej
anje
/pre
heat
(50
ºC)
Poto
piti
u /s
oak
in H
2O il
i/or K
NO
3 (2
4h)
Phas
eolu
s vul
gari
s L.
IF; N
FP; S
20-3
0; 2
5; 2
05-
9-
Poa
annu
a L.
NF
20-3
0;15
-25
7-21
Pret
hodn
o hl
ađen
je/p
rech
ll K
NO
3Sp
inac
ia o
lerac
ea L
.N
F; IF
15; 1
07-
21Pr
etho
dno
hlađ
enje
/pre
chll
Triti
cum
aes
tivum
L.
NF;
IF; P
204-
8Pr
etho
dno
grej
anje
/pre
heat
(30-
35ºC
)Pr
etho
dno
hlađ
enje
/pre
chll,
GA
3Vi
cia fa
ba L
.IF
; P; S
204-
14Pr
etho
dno
hlađ
enje
/pre
chll
Zea
may
s L.
IF; N
FP; P
20-3
0; 2
5; 2
04-
7-
IF –
izm
eđu
filte
r pap
ira/b
etw
een
pape
r, N
F –
na fi
lter p
apiru
/top
of p
aper
, P –
pes
ak/s
and,
S
– su
pstra
t / o
rgan
ic g
row
ing
med
ia, N
FP –
na
filte
r pap
iru p
rekr
iven
o pe
skom
/top
of p
aper
cov
ered
with
san
d, G
A3 –
gib
erel
insk
a ki
selin
a / g
iber
elic
aci
d, K
NO
3 – 0
,2%
kal
ijum
nitr
a / p
otas
sium
nitr
ate

232
Semenarstvo
Postoje dva načina naklijavanja semena sa filter papirom: metod na filter papiru i metod između filter papira.
Metod na filter papiru se koristi kod naklijavanja sitnog semena i semena kome je potrebno prisustvo svetlosti za prekidanje perioda mi-rovanja. Metod se izvodi u plastičnim ili staklenim posudama, različitih dimenzija. Na dno posude stavlja se 1 ili 2 sloja filter papira (zavisnosti od debljine filter papira i kapaciteta za vlagu). U procesu naklijavanja nije poželjno dodavanje vode, jer je dodata količina obično neujedna-čena po ponavljanju i može da se dobije pogrešan podatak o klijavosti semena (sl. 15).
Slika 15. Test klijavosti na filter papiru Figure 15. Seed germination test on top of paper (Jovičić, 2010)
Metod između filter papira koristi se za naklijavanje sitnog i kru-pnog semena. Postoje tri tipa ovog metoda: tip rolne, tip pisma i faltani papir (sl. 16).
Tip pisma koristi se kod sitnijeg semena. Uzorci se u aparat stavlja-ju u horiznotalnom položaju.
Tip rolne se koristi kod krupnijeg semena gde je potrebno obezbe-diti bolji kontakt između semena i podloge. Uzorci se u aparat za ispi-tivanje klijavosti semena stavljaju vertikalno, što obezbeđuje normalan porast ponika zbog pozitivnog geotropizma.
Faltani papir se koristi kod naklijavanja semena Beta vulgaris, zbog potrebe utvrđivanja monogermnosti i semena zaraženog fitopatogenim gljivama, jer se sprečava sekundarna infekcija semena.

233
Ispitivanje kvaliteta semena
Fitotoksično dejstvo pojedinih preparata na seme i ponik mogu se odrediti primenom metode ispitivanja klijavosti semena korišćenjem filter papira kao podloge. Ovaj metod je uvek rigorozniji u proceni jer je fitotoksični efekat u pozitivnoj korelaciji sa asorptivnosti podloge. Ako ovim metodom dobijemo upozoravajuće rezultate pristupa se proveri fitotoksičnosti u pesku ili zemlji (Klokočar – Šmit and Inđić 1991, Vu-jaković et. al. 2003a).
Slika 16. Metod između filter papira (tip pisma, tip rolne, tip falte)Figure 16. Method between paper (pleated paper, paper roll) (Jovičić, 2010)
Metod sa peskom
Pesak treba da bude ujednačen. Preporučuje se da skoro sve čestice treba da prođu kroz sito sa otvorima (okruglim ili četvrtastim) prečnika 0,8 mm i da se zadrže na situ sa otvorima prečnika 0,05 mm. Pesak ne sme da sadrži drugo seme, gljive, bakterije ili toksične materije, što sve može da utiče na klijanje semena, rast klijanaca ili njihovo ocenjivanje. Čestice peska treba da imaju takav kapacitet vlaženja da zadrže dovoljnu količinu vode, kako bi obezbedile kontinuiran dotok vode do semena i klijanaca, ali i da imaju dovoljnu poroznost za aeraciju, radi optimalnog klijanja i rasta korena. pH vrednost peska treba da bude 6,0–7,5. Pesak može da se koristi više puta, ali pre nego što se ponovo upotrebi mora da se ispere, osuši i resteriliše. Pesak koji se koristi za ispitivanje hemijski tretiranog semena se baca.
Kao kod filter papira i kod peska postoje dva metoda za ispitivanje klijavosti semena: metod na pesku i metod u pesku.
Metod na pesku se koristi za naklijavanje sitnijeg semena. Seme se stavi na sloj vlažnog peska 1–2 cm i blago se utisne. Tako pripremljeni uzorci se stavljaju u aparat za ispitivanje klijavosti semena.

234
Semenarstvo
Metod između peska se koristi za naklijavanje krupnijeg semena. Na sloj vlažnog peska debljine 1-2 cm stavi se seme da se ne dodiruje i pre-krije se vlažnim peskom debljine 1 cm (sl. 17).
Slika 17. Test klijavosti u peskuFigure 17. Seed germination test in sand (Jovičić, 2010)
Metod sa kompostom
Kompost se ne preporučuje kao rutinska podloga za ispitivanje kli-javosti semena. Može da se koristi za neke biljne vrste, za koje je to pro-pisano ISTA Pravilima (2011) i kada klijanac ispolji fitotoksične simp-tome ili kada je ocena klijanaca u pesku ili na papiru sumnjiva. Obično se koristi za komparativne ili istraživačke svrhe. Kompost sadrži or-ganske materije (najčešće treset) i mineralne komponente (pesak, perlit, vermikulit) kojih treba da bude oko 20%. pH vrednost treba da bude 6,0–7,5. Ponovna upotreba komposta nije dozvoljena.
Postoje dve varijante upotrebe komposta za ispitivanje klijavosti se-mena: metod na kompostu i metod u kompostu. Postupak u metodama je identičan kao i kod upotrebe peska, kao podloge.
Ocena ponika
Iz frakcije čistog semena se po principu slučajnosti izbroji 4 x 100 semena koja se raspoređuju na vlažnu podlogu bez međusobnog do-dirivanja. Ponavljanja se mogu podeliti u podponavljanja od 50 ili 25 semena, zavisno od veličine semena i potrebnog prostora između seme-na. Nakon perioda naznačenog u Pravilniku o kvalitetu semena poljo-

235
Ispitivanje kvaliteta semena
privrednog bilja (Sl. list SFRJ 47/87) i ISTA Pravilima (2011) pristupa se ocenjivanju klijanaca. Prilikom ocenjivanja osnovne strukture moraju biti dobro razvijene da bi se klijanci mogli oceniti kao tipični ili atipični. Osnovne strukture ponika su:
- korenov sistem (primarni koren u nekim slučajevima sekundar-ni korenovi),
- osa izdanaka (hipokotil, epikotil, kod određenih Poaceae mezo-kotil i vršni pupoljak),
- kotiledoni (jedan do nekoliko) i- koleoptil (kod svih Poaceae ).
Tipičan klijanac
Tipični klijanci pokazuju sposobnost da se u zemljištu dobrog kvali-teta, pod povoljnim uslovima vlage, temperature i svetlosti, kontinuira-no razvijaju u normalnu biljku. Tipičnim se smatraju sledeće kategorije klijanaca:
Neoštećeni klijanci: klijanci sa svim svojim osnovnim strukturama koje su dobro, potpuno i proporcionalno razvijene i koje nisu zaražene patogenima. Neoštećen klijanac, zavisno od vrste koja se ispituje i ima specifičnu kombinaciju nekih od sledećih osnovnih struktura:
a) Dobro razvijen korenov sistem koji se sastoji od:– dugačkog i tankog primarnog korena, obično pokrivenog brojnim
korenovim dlačicama, koji se završava finim vrhom,– sekundarnih korenova kada se formiraju u vremenu propisanom
za ispitivanje i– nekoliko seminalnih korenova (primarni korenčić kod žita) ume-
sto jednog primarnog korena, kod određenih rodova uključujući Avena, Hordeum, Secale, Triticum, x Triticosecale, Cyclamen.
b) Dobro razvijene ose izdanaka, koja se sastoji od: – pravog i obično tankog i izduženog hipokotila, kod vrsta sa epige-
alnim klijanjem,– dobro razvijenog epikotila, kod vrsta sa hipogealnim klijanjem
(sl. 18),– izduženim hipokotilom i epikotilom, kod nekih vrsta sa epigel-
nim klijanjem (sl. 18) i– izduženim mezokotilom, kod određenih rodova Poaceae.

236
Semenarstvo
c) Specifičan broj kotiledona, tj.– jedan kotiledon, kod monokotila ili izuzetno kod dikotila (može
da bude zelen i listolik ili modifikovan i može ceo ili delimično da ostane unutar semena),
– dva kotiledona, kod dikotila (kod vrsta sa epigelnim klijanjem su zeleni i listoliki, veličina i oblik variraju zavisno od vrste koja se ispituje; kod vrsta sa hipogelnim klijanjem oni su poluokrugli i mesnati i ostaju unutar semenjače).
d) Zeleni primarni listovi u razvoju:– jedan primarni list, ponekad mu prethodi nekoliko ljuspastih li-
stova ili
prvi zeleni listthe first green leaf
prvi zeleni listthe first green leaf kotiledoni
cotyledons
primarni koren/primary root
kotiledoncotyledonradikula
radicula
plumuleplumele
epikotilepicotile
A. Hipogealni tip klijanja boba/Hipogel type of germation (bean)
prvi zeleni listthe first green leaf
epikotilepicotile
hipokotilhipocotile
kotiledoni/cotyledonc
hipokotilhipocotile
kotiledonicotyledons
testa
B. Epigealan tip klijanja pasulja/Epigel type of germation (bean)
Slika 18. Hipogealno i epigealno klijanje i struktura ponikaFigure 18. Hipogeal and epigeal germination and seedling structure (Kastori, 1984)

237
Ispitivanje kvaliteta semena
– dva primarna lista, kod vrsta sa naspramnim listovima.e) Vršni pupoljak ili vrh klijanca.
f) Dobro razvijen prav koleoptil, kod Poaceae koji obavija zeleni list sve do vrha ili list viri iz koleoptila.
Klijanci sa blagim oštećenjima: klijanci koji pokazuju određene male nepravilnosti svojih osnovnih struktura, pod uslovom da u poređe-nju sa neoštećenim klijancima iz istog ispitivanja, imaju zadovoljavajući i ujednačen razvoj. U ovu grupu klijanaca ubrajaju se oni kod kojih je:
– primarni koren sa ograničenim oštećenjem i blago usporenim ra-stom,
– primarni koren oštećen, ali sa dovoljno dobro razvijenim sekun-darnim korenovima, kod specifičnih rodova Fabaceae (posebno rodovi sa krupnim semenom, kao što su Phaseolus, Pisum, Vicia) i Poaceae (npr. Zea) i kod svih rodova Cucurbitaceae (npr. Cucumis, Cucurbita, Citrullus) i Malvaceae (npr. Gossypium),
– samo jedan jak seminalni (primarni korenčić) koren kod: Avena, Hordeum, Secale, Triticum, x Triticosecale, i dva kod Cyclamen,
– hipokotil, epikotil ili mezokotil, sa ograničenim oštećenjima,– kotiledoni sa ograničenim oštećenjem (ako je polovina ukupne
površine tkiva funkcionalna (pravilo 50%) i ako ne postoji vidlji-vo oštećenje ili truljenje vršnog pupoljka ili okolnog tkiva),
– samo jedan normalan kotiledon kod dikotila (ako ne postoji vid-ljivo oštećenje ili truljenje vršnog pupoljka ili okolnog tkiva),
– tri kotiledona umesto dva (pravilo 50%),– primarni listovi sa ograničenim oštećenjem, ako je polovina ili
više od pola ukupnog tkiva funkcionalno (pravilo 50%),– samo jedan normalan primarni list, npr. kod Phaseolus (ako ne
postoje tragovi oštečenja ili truljenja vršnog pupoljka), – primarni listovi Phaseolus koji su pravilno formirani ali smanjene
veličine, sve dok su veći od jedne četvrtine normalne veličine,– tri primarna lista umesto dva npr. kod Phaseolus (pravilo 50%),– koleoptil sa ograničenim oštećenjem, – koleoptil sa pukotinom od vrha koja se širi na dole, ne više od jed-
ne trećine dužine (za Zea mays klijanci sa oštećenjima koleoptila se smatraju normalnim ako je prvi list neoštećen ili je sa malim oštećenjem),

238
Semenarstvo
– koleoptil blago uvrnut ili formira petlju (zato što je zarobljen pod lemom i paleom ili omotačem ploda) i
– koleoptil sa zelenim listom koji se ne proteže do vrha, već dostiže bar polovinu visine koleoptila.
Klijanci sa sekundarnom infekcijom: klijanci iz gore navedenih stavki ali napadnuti gljivama ili bakterijama koje ne potiču sa tog se-mena.
Atipični klijanci
Atipični klijanci ne pokazuju potencijal da se u dobrom zemljištu i povoljnim uslovima vlažnosti, temperature, svetlosti razviju u normal-nu biljku. Atipičnim se smatraju sledeći klijanci:
Klijanac u celini ako je: deformisan, slomljen, kotiledoni izbijaju pre primarnog korena iz semenjače, sastoji se od dva spojena klijanca, nije otpala „kragna“ endosperma, žut ili beo, vretenasto uvijen, staklast, truo zbog primarne infekcije i pokazuje fitotoksične simptome.
Primarni koren ako je: zakržljao, zade-bljao, zaostao, nedostaje, polomljen, rascepljen od vrha, sužen, vretenasto uvijen, zarobljen u semenjači, sa negativnim geotropizmom, sta-klast i truo zbog primarne infekcije (sl. 19).
Slika 19. Ponici pšeniceFigure 19. Wheat seedlings (ISTA, 2003)
Sekundarni korenovi ako su: samo jedan zadebljao, slab ili nijedan seminalni koren.
Hipokotil, epikotil, mezokotil ako je: kratak i debeo (osim kod Cyclamen), ne formiran gomolj (kod Cyclamen), jako raspuknut ili slo-mljen, rascepljen po sredini, nedostaje, presavijen ili formira petlju/omču, spiralno uvijen, jako uvijen/uvrnut, vretenasto uvijen, staklast i truo zbog primarne infekcije.
Vršni pupoljak i okolno tkivo ako je: deformisano, oštećeno, ne-dostaje, nekrotičano (truli ili izumire) i trulo zbog primarne infekcije.
tipičnitypic
atipični/atypic

239
Ispitivanje kvaliteta semena
Kotiledoni ako su više od 50%: uvećani ili uvijeni, deformisani, polomljeni ili na drugi način oštećeni, razdvojeni ili nedostaju, izgubili boju ili nekrotični (truli ili izumiru), staklasti i truli kao rezultat primar-ne infekcije.
Ponik kod Allium spp. ako su: krataki, debeli, suženi, presavijeni, formira petlju ili spiralu, bez formiranog „kolenca” i vretenasto uvijeni (sl. 20).
Slika 20. Ponici luka / Figure 20. Onion seedlings (ISTA, 2003)
Primarni listovi ako su više od 50%: deformisani, oštećeni, nedo-staju, izgubili boju, nekrotični, normalnog oblika ali manji od ¼ nor-malne veličine i truli zbog primarne infekcije.
Koleoptil i primarni list kod Poacea ako je: zadebljao ili na dru-gi način deformisan, slomljen, nedostaje, sa vrhom koji je oštećen ili nedostaje, jako presavijen ili formira petlju, formira spiralu, jako uvi-jen, rascepljen od vrha više od jedne trećine dužine, vretenasto uvijen, truo zbog primarne infekcije i rascepljen u osnovi. Primarni list ako je: kraći od polovine dužine koleoptila, nedostaje, izreckan ili na drugi način deformisan, probija iz donjeg dela koleoptila, žut ili beo (nema hlorofila) i truo zbog primarne infekcije.
Neklijalo seme
Neklijalo seme je seme koje ne klija do kraja perioda ispitivanja propisanog Pravilnikom o kvalitetu semena poljoprivrednog bilja

240
Semenarstvo
(Sl. list SFRJ 47/87) i ISTA Pravilima (2011) i može biti: tvrdo seme, sveže seme, mrtvo seme i ostale kategorije neklijalog semena.
Tvrdo seme je oblik mirovanja semena. Čest je kod mnogih vrsta Fabaceae, ali može da se javi i kod drugih familija. Ovo seme nije spo-sobno da upije vodu do kraja perioda ispitivanja.
Sveže seme je seme sposobno da upija vodu u uslovima naznače-nim u pravilniku, ali je proces klijanja blokiran.
Mrtvo seme je obično meko, obezbojeno, često napadnuto gljivama i ne pokazuje znake razvoja klijanaca.
Ostale kategorije neklijalog semena su:– prazno seme (seme koje je potpuno prazno ili sadrži samo rezi-
dualna tkiva),– seme bez embriona (seme koje sadrži svež endosperm ili game-
tofitno tkivo u kome očigledno nema embrionalne šupljine niti embriona) i
– seme oštećeno insektima (seme koje sadrži larve insekata, ostatke od insekata ili pokazuje druge znakove napada insekata što sve utiče na sposobnost semena da klija).
Rezultat ispitivanja klijavosti se izračunava kao prosek 4 ponavlja-nja od po 100 semena (podponavljanja sa 50 ili 25 semena se kombinuju u ponavljanja od 100 semena). Rezultat se izražava kao procenat tipič-nih klijanaca. Procenat se zaokružuje na najbliži ceo broj (0,5 se zaokru-žuje na veću cifru). Na isti način se izračunava i % atipičnih klijanaca, tvrdog, svežeg i mrtvog semena. Zbir svih procenata mora da bude 100.

241
Ispitivanje kvaliteta semena
ISPITIVANJE SADRŽAJA VLAGE U SEMENU
Sadržaj vlage u semenu je kompleksan sistem (Leopold and Ver-tucci 1989). On ima veoma bitnu ulogu u procesu formiranja semena, doradi i čuvanju semena. Poznato je da različite biljne vrste imaju ra-zličit optimum vlage za uspešno čuvanje (tab. 2). Sadržaj vlage u se-menu predstavlja gubitak njegove mase prilikom sušenja (Nijenstein et al. 2007). Propisane metode za određivanje sadržaja vlage u semenu smanjuju oksidaciju, razlaganje ili gubitak ostalih isparljivih supstanci uz istovremeno odstranjivanje što više vlage.
Prosečan uzorak za vlagu je merodavan ako se nalazi u vodootpor-noj ambalaži iz koje je vazduh maksimalno istisnut.
Određivanje vlage počinje što je pre moguće po prispeću uzorka. Za vreme određivanja vlage izlaganje uzorka atmosferi u laboratoriji mora biti smanjeno na apsolutni minimum, a za vrste kojima nije po-trebno mlevenje ne sme da prođe više od 2 min od trenutka uzimanja uzorka iz pakovanja do trenutka kada se radni uzorak zatvara u sud za sušenje.
Tabela 4. Biljne vrste za koje je mlevenje obaveznoTable 4. Plant species for which the grinding required (ISTA, 2011)
Biljna vrsta/Plant species
Amorpha fruticoza Hordeum vulgare Secale cereale
Arachis hypogaea Lathyris spp Sorghum spp.
Avena spp Lupinus spp. Triticum spp.
Cicer arietinum Macroptilium atropurpureum Vicia spp.
Citrullus lanatus Oryza sativa Vigna spp.
Fagopyrum esculentum Phaseolus spp. Zea mays
Glycine max Pisum sativum (svi varijeteti)
Gossupium spp. Ricinus communis

242
Semenarstvo
Tabela 5. Biljne vrste za koje se koristi metod sušenja na visokoj konstantnoj tempe-raturi (130ºC)
Table 5. Plant species used method of drying in the constant high temperature (130ºC) (ISTA, 2011)
Biljna vrsta/Plant species
Agrostis spp Dactylis glomerata Papaver somniferun
Alopecurus pratensis Daucus carota Paspalum spp
Anethun graveolens Deschampsia spp Pastinaca sativa
Anthoxanthum odoratum Elytrigia spp Petroselinum crispum
Anthriscus spp Fagopyrum esculentum Phacelia tanacetifolia
Apium graveolens Festuca spp Phalaris spp
Arrhenatherum spp Galega orientalis Phaseolus spp
Asparagus officinalis Holcus lanatus Phleum spp
Avena spp. Hordeum vulgare (svi varijeteti)
Pisum sativum (svi varijeteti)
Beta vulgaris (svi varijeteti) Lactuca sativa Poa spp
Brachiaria spp Lathyrus spp Scorzonera hispanica
Bromus spp Lepidium sativum Secale cereale
Cannabis sativa Lolim spp Setaria spp
Carum carvi Lotus spp Sorghum spp
Cenchrus spp Lupinus spp Spinacia oleracea
Chloris gayana Lycopersicum lycopersicum Trifolium spp
Cicer arietinum Macroptilium atropurpureum Trisetum flavescens
Cichorium spp Medicago spp Triticum spp
Citrullus lanatus Melilotus spp Varelianella locusta
Cucumis spp Nicotiana tabacum Vicia spp
Cucurbita spp Onobrychis viciifolia Vigna spp
Cuminum cyminum Ornithopus sativus Zea mays
Cynodon dactylon Oryza sativa
Cynosurus cristatus Panicum spp

243
Ispitivanje kvaliteta semena
Određivanje vlage se radi u dva ponavljanja na dva nezavisna pri-premljena radna uzorka u količini koja odgovara veličini prečnika suda koji se koristi: ako je prečnik > 5 cm, a < 8 cm uzima se 4,5 ± 0,5 g; ako je prečnik ≥ 8 cm uzima se 10,0 g ± 1,0 g.
Pojedine vrste semena se pre sušenja melju ili seku (tab. 4). Neop-hodnost za mlevenje zavisi od veličina zrna i vodopropusnosti omotača zrna (poroznost). Karakteristike semena, kao što su visoka vlažnost ili ekstremno tvrd omotač semena, mogu da onemoguće mlevenje. U tim situacijama vrši se sečenje ili drobljenje semena u komadiće do 7 mm prečnika.
Prema Pravilniku o kvalitetu semena poljoprivrednog bilja (Sl. list SFRJ 47/87) i ISTA Pravilima (2011) definisana su dva metoda za ispitivanje sadržaja vlage u semenu: metod u sušnici s visokom kon-stantnom temperaturom i metod u sušnici sa niskom konstantnom temperaturom. Pravilnikom je definisano koje biljne vrste se ispituju na visokoj temperaturi (tab. 5), a koje na niskoj (tab. 6). Postupak sušenja je identičan, razlike su u temperaturi i vremenu sušenja. Kod metode sa visokom konstantnom temperaturom koristi se tempera-tura od 130–133ºC u periodu: 1h ± 3 min, 2h ± 6 min (žita) ili 4h ± 12 min (kukuruz). Metod niske konstantne temperaturi se izvodi na temeraturi 101–105ºC, u trajanju 17h ± 1h. Pre stavljanja poduzora-ka na sušenje izmere se prazne radne posude. U posude se stavi seme (ponovo se izmeri masa) i pripremljeni uzorci se stavljaju u sušnicu na odgovarajuću temperaturu. Nakon završenog sušenja uzorci se vade u eksikator da se prohlade i nakon toga mere i izračunava sadr-
Tabela 6. Biljne vrste za koje se koristi metod sušenja na niskoj konstantnoj tempera-turi (103ºC)
Table 6. Plant species used method of drying in the constant low temperature (103ºC) (ISTA, 2011)
Biljna vrsta/Plant species
Allium spp Capsicum spp Raphanus sativus
Amorpha fruticosa Glycine max Ricinus communis
Arachis hypogaea Gossypium spp Sesamum indicum
Brassica spp Helianthus annuus Sinapis spp
Camelia sativa Linum usitatissimum Solanum melongena

244
Semenarstvo
žaj vlage u semenu prema formuli, na jedno decimalno mesto:
M2 – M3V(%) = ___________________ · 100 M2 – M1
gde je:V - sadržaj vlage u semenu u procentimaM1 – masa posude i poklopca u gramimaM2 – masa posude, poklopca i uzorka u gramima pre sušenjaM3 – masa posude, poklopca i uzorka u gramima posle sušenja.
Kod vrsta kojima je mlevenje neophodno i kod kojih je sadržaj vlage viši od propisanog prethodno sušenje je obavezno. Dva poduzorka, sva-ki težine 25 ± 1 g, sipaju se u izmerene posude. Zatim se ovi poduzorci u posudama suše na 130 ºC 5 do 10 minuta, što zavisi od količine vlage, da bi se sadržaj vlage snizio na propisanu vrednost. Delimično osušen materijal se čuva u laboratoriji najmanje 2 sata i potom se koristi jedan od gore navedenih metoda.
U slučaju veoma vlažnog semena Zea mays (preko 25% sadržaja vlage), seme se širi u slojeve debljine do 20 mm i suši se na 65–75ºC 2–5h, što zavisi od količine vlage. Kod drugih vrsta kod kojih je sadržaj vlage veći od 30%, uzorci se suše preko noći na toplom mestu. Posle predsušenja, poduzorci se ponovo mere u svojim posudama da bi se odredio gubitak mase. Odmah zatim dva dosušena poduzorka se odvo-jeno melju i primenjuje se napred opisana metoda. Prethodno sušenje nije obavezno za semena koja su sečena.
Ako je materijal izložen procesu prethodnog sušenja, sadržaj vlage (V) se izračunava iz rezultata dobijenih u prvoj (predsušenje) i drugoj fazi postupka. Ako je S1 gubitak vlage u prvoj fazi, a S2 gubitak vlage u drugoj fazi, oba izračunata prema gornjoj formuli i izražena procentu-alno, onda se procenat stvarnog sadržaja vlage uzorka obračunava kao:
S1 · S2V(%) = (S1 + S2) – _____________
100

245
Ispitivanje kvaliteta semena
ODREĐIVANJE MASE 1000 SEMENA
Iz frakcije čistog semena rukom ili brojačem, po principu slučajno-sti, izbrojati osam ponavljanja od po 100 semena. Treba izmeriti svako ponavljanje u gramima i izračunati varijansu, standardnu devijaciju i koeficijent varijacije na sledeći način:
Varijansa =
Table 6. Plant species used method of drying in the constant low temperature (103ºC) (ISTA,
2010)
Biljna vrsta/Plant species
Allium spp Capsicum spp Raphanus sativus
Amorpha fruticosa Glycine max Ricinus communis
Arachis hypogaea Gossypium spp Sesamum indicum
Brassica spp Helianthus annuus Sinapis spp
Camelia sativa Linum usitatissimum Solanum melongena
ODREĐIVANJE MASE 1000 SEMENA
Iz frakcije čistog semena rukom ili brojačem, po principu slučajnosti, izbrojati osam
ponavljanja od po 100 semena. Treba izmeriti svako ponavljanje u gramima i izračunati
varijansu, standardnu devijaciju i koeficijent varijacije na sledeći način:
Varijansa = ( ) ( )
( )
22
1−− ∑∑
NNXXn
gde je:
X = masa svakog ponavljanja u gramima
N = broj ponavljanja
Standardana devijacija (S) = Varijansa
Koeficijent varijacije = 100×XS
gde je X = prosečna masa 100 semena
Ako koeficijent varijacije ne prelazi 6,0 za seme plevičastih trava, ili 4,0 za ostala
semena, rezultati se mogu obračunavati. Ako je koeficijent varijacije veći od navedenih vrednosti
analiza se ponavlja.
gde je:X = masa svakog ponavljanja u gramimaN = broj ponavljanjaStandardana devijacija (S) =
Table 6. Plant species used method of drying in the constant low temperature (103ºC) (ISTA,
2010)
Biljna vrsta/Plant species
Allium spp Capsicum spp Raphanus sativus
Amorpha fruticosa Glycine max Ricinus communis
Arachis hypogaea Gossypium spp Sesamum indicum
Brassica spp Helianthus annuus Sinapis spp
Camelia sativa Linum usitatissimum Solanum melongena
ODREĐIVANJE MASE 1000 SEMENA
Iz frakcije čistog semena rukom ili brojačem, po principu slučajnosti, izbrojati osam
ponavljanja od po 100 semena. Treba izmeriti svako ponavljanje u gramima i izračunati
varijansu, standardnu devijaciju i koeficijent varijacije na sledeći način:
Varijansa = ( ) ( )
( )
22
1−− ∑∑
NNXXn
gde je:
X = masa svakog ponavljanja u gramima
N = broj ponavljanja
Standardana devijacija (S) = Varijansa
Koeficijent varijacije = 100×XS
gde je X = prosečna masa 100 semena
Ako koeficijent varijacije ne prelazi 6,0 za seme plevičastih trava, ili 4,0 za ostala
semena, rezultati se mogu obračunavati. Ako je koeficijent varijacije veći od navedenih vrednosti
analiza se ponavlja.
Koeficijent varijacije =
Table 6. Plant species used method of drying in the constant low temperature (103ºC) (ISTA,
2010)
Biljna vrsta/Plant species
Allium spp Capsicum spp Raphanus sativus
Amorpha fruticosa Glycine max Ricinus communis
Arachis hypogaea Gossypium spp Sesamum indicum
Brassica spp Helianthus annuus Sinapis spp
Camelia sativa Linum usitatissimum Solanum melongena
ODREĐIVANJE MASE 1000 SEMENA
Iz frakcije čistog semena rukom ili brojačem, po principu slučajnosti, izbrojati osam
ponavljanja od po 100 semena. Treba izmeriti svako ponavljanje u gramima i izračunati
varijansu, standardnu devijaciju i koeficijent varijacije na sledeći način:
Varijansa = ( ) ( )
( )
22
1−− ∑∑
NNXXn
gde je:
X = masa svakog ponavljanja u gramima
N = broj ponavljanja
Standardana devijacija (S) = Varijansa
Koeficijent varijacije = 100×XS
gde je X = prosečna masa 100 semena
Ako koeficijent varijacije ne prelazi 6,0 za seme plevičastih trava, ili 4,0 za ostala
semena, rezultati se mogu obračunavati. Ako je koeficijent varijacije veći od navedenih vrednosti
analiza se ponavlja.
gde je
Table 6. Plant species used method of drying in the constant low temperature (103ºC) (ISTA,
2010)
Biljna vrsta/Plant species
Allium spp Capsicum spp Raphanus sativus
Amorpha fruticosa Glycine max Ricinus communis
Arachis hypogaea Gossypium spp Sesamum indicum
Brassica spp Helianthus annuus Sinapis spp
Camelia sativa Linum usitatissimum Solanum melongena
ODREĐIVANJE MASE 1000 SEMENA
Iz frakcije čistog semena rukom ili brojačem, po principu slučajnosti, izbrojati osam
ponavljanja od po 100 semena. Treba izmeriti svako ponavljanje u gramima i izračunati
varijansu, standardnu devijaciju i koeficijent varijacije na sledeći način:
Varijansa = ( ) ( )
( )
22
1−− ∑∑
NNXXn
gde je:
X = masa svakog ponavljanja u gramima
N = broj ponavljanja
Standardana devijacija (S) = Varijansa
Koeficijent varijacije = 100×XS
gde je X = prosečna masa 100 semena
Ako koeficijent varijacije ne prelazi 6,0 za seme plevičastih trava, ili 4,0 za ostala
semena, rezultati se mogu obračunavati. Ako je koeficijent varijacije veći od navedenih vrednosti
analiza se ponavlja.
= prosečna masa 100 semenaAko koeficijent varijacije ne prelazi 6,0 za seme plevičastih trava,
ili 4,0 za ostala semena, srednja vrednost se prihvata. Ako je koeficijent varijacije veći od navedenih vrednosti analiza se ponavlja. Mnoga istra-živanja su pokazala da masa 1000 semena zavisi od ispitivane sorte i od godine proizvodnje (Marjanović-Jeromela et. al., 1999, Vujaković et. al., 2010, Jovičić et. al., 2011) (sl. 21).
Slika 21. Seme soje / Figure 21. Soybean seed (Jeromela, 2009)

246
Semenarstvo
ISPITIVANJE ŽIVOTNE SPOSOBNOSTI SEMENA
Međunarodna trgovina semena, kao osnovni pokazatelj kvaliteta uzima klijavost semena, pa se iz tih razloga ulažu veliki napori na ujed-načavanju metoda ispitivanja. Za utvrđivanje klijavosti semena koristi se standardni test klijavosti. Ovaj test je standardizovan za veliki broj biljnih vrsta.
Različiti rezultati ispitivanja klijavosti semena u laboratorijskim uslovima i nicanja u polju podstakli su razvoj koncepta životne spo-sobnosti semena i pojma vigora. Primećeno je da se javljaju razlike u poljskom nicanju između partija semena sa visokom klijavošću, iako su posejane u isto vreme, na istom mestu. Razlike između partija semena ne mogu se uočiti u uslovima povoljnim za klijanje, ali se lako uočavaju kada se seme izloži nepovoljnim uslovima klijanja. Takođe, te razlike se mogu javiti i posle skladištenja pod istim uslovima ili posle transporta do istog odredišta. Posmatranjem klijvosti tih partija postavlja se pitanje da li su rezultati testa klijavosti zaista pouzdani i tačni i kako nasta-je tako izražena razlika u kvalitetu. Ispitivanja su pokazala da današnji standardni test za ispitivanje klijvosti daje korektne rezultate, ali da nije dovoljno precizan da bi ukazao na razlike u kvalitetu između partija sa visokom klijavošću. Ove razlike su izazvane drugim komponentama kvaliteta semena, tkz. životnom sposobnošću semena ili vigorom (Te-Krony 1982, prema Milošević and Zlokolica 1996b).
Životna sposobnost semena ili vigor predstavlja skup onih osobina koje određuju aktivnost i ponašanje partije semena komercijalno pri-hvatljive klijavosti u različitim uslovima spoljašnje sredine (ISTA 2011). Drugim rečima, pojam vigor podrazumeva one osobine semena koje kontrolišu njegovu sposobnost da brzo klija u zemljištu i da toleriše ra-zne, uglavnom negativne spoljašnje činioce (Vujaković 2004).
Vigor semena nije svojstvo koje je moguće pojedinačno izmeriti kao što je moguće klijavost, već je pojam koji opisuje nekoliko osobina koje su povezane sa sledećim aspektima stanja partije semena (ISTA 2011):

247
Ispitivanje kvaliteta semena
1. brzina i ujednačenost klijanja semena i porast ponika;2. sposobnost nicanja semena u nepovoljnim uslovima spoljašnje
sredine i3. stanje semena posle skladištenja, pre svega zadržavanje kapaci-
teta klijanja.
Sazrevanjem semena i nakupljanjem proteina i skroba, klijavost i vi-gor semena se postepeno povećava i maksimum dostiže u fazi fiziološke zrelosti semena (Sun et al. 2007).
Prema Dornbos (1995) postoje četiri važna činioca koji utiču na pri-nos i vigor semena:
1. pravilna primena agrotehničkih mera u proizvodnji semenskih useva koji umanjuju posledice stresa;
2. proizvodnja semena u regionima koji imaju povoljne uslove za ra-zvoj određene biljne vrste;
3. odabir parcela za proizvodnju koje je moguće navodnjavati i4. voditi računa da se koriste stabilne sorte.Sun et al. (2007) smatraju da i pored ovih faktora, na životnu sposob-
nost semena, utiče i način skladištenja (vreme skladištenja, temperatura, vlažnost vazduha, količina kiseonika itd.).
Metodologija za izvođenje standardnog testa klijavosti je stan-dardizovana kako bi se rezultati mogli upoređivati između laborato-rija. Test mora biti sproveden na veštačkoj, standardizovanoj, steril-noj podlozi u vlažnim prostorijama sa kontrolisanom temperaturom. Ovakvi optimalni uslovi gotovo da se ne mogu porediti sa stvarnim uslovima u polju. Uslovi u polju kao što su visoka ili niska tempe-ratura, velika vlažnost zemljišta ili teško zemljište, mogu da dovedu do usporenog klijanja, nicanja, slabog ponika, a ponekad i do gubitka životne sposobnosti semena. U takvim uslovima prinos, a i kvalitet semena mogu biti znatno smanjeni (McDonald 1993). Standardna la-boratorijska klijavost često ne može da predvidi stvarnu klijavost u polju, posebno ako su uslovi nepovoljni (Basak et al. 2006). Kada bi uslovi u polju bili optimalni, standardni laboratorijski test bi mogao tačno da predvidi uspešnost klijanja (Durrant and Gummerson 1990). Zbog toga primenjuju se testovi za ispitivanje vigora semena koji tre-ba da obezbede dodatnu informaciju o fiziološkom kvalitetu semena (Milošević and Ćirović 1994).

248
Semenarstvo
Upotreba vigor testova je veoma značajna, s obzirom da vigor te-stovi daju rezultate koji su često u boljim korelacionim odnosima sa rezultatima poljskog nicanja u nepovoljnim uslovima sredine, od rezul-tata dobijenih primenom standardnog testa klijavosti u laboratorijskim uslovima (Johansen and Wax 1978).
Glavni cilj vigor testova je da odrede koje partije semena bi mogle da podnesu manje povoljne uslove u polju, odnosno da razlikuju par-tije, sa suštinski jednakim procentom klijanja, prema njihovoj sposob-nosti da dobro klijaju uprkos nepovoljnim uslovima sredine (Karrfalt 2004).
Vigor test bi trebao da zadovoljava određene kriterijume kako bi bio prihvatljiv i koristan za proizvođače semena, krajnje korisnike se-mena i analitičare u laboratorijama. Ne postoje univerzalno pirhvaće-ni vigor testovi za sve biljne vrste. Kod izbora vigor testa mora se vo-diti računa o biologiji, fiziologiji i drugim specifičnostima biljne vrste. Iz tog razloga vigor testovi su grupisani na više načina (Milošević et. al. 2010).
McDonald (1980) navodi da vigor testovi treba da budu jednostav-ni, brzi, objektivni, ponovljivi, jeftini i pouzdani.
Vigor testove možemo podeliti u tri grupe (McDonald 1975):
1. Fizički testovi određuju karakteristike semena kao što su frakcio-ni sastav semena, masa hiljadu semena, sadržaj vlage i brzina bubrenja. Ovaj test se zasniva na činjenici da je glavno svojstvo razvoja semena akumulacija hranljivih materija. Ovi testovi su jeftini, brzi i pokazuju pozitivnu korelaciju sa vigorom semena.
2. Fiziološki (direktni) testovi koriste parametre klijanja i porasta ponika. Postoje dva tipa ovih testova:
– testovi u kojima dolazi do klijanja u povoljnim laboratorijskim uslo-vima (standardni laboratorijski metod, test intenziteta porasta),
– testovi u kojima se seme izlaže nepovoljnim uslovima sredine (hladni test, test ubrzanog starenja i Hiltner test);
3. Biohemijski (indirektni) testovi obuhvataju tetrazolijum test i test provodljivosti.

249
Ispitivanje kvaliteta semena
Fiziološki testovi
Test intenziteta porasta
Test intenziteta porasta se zasniva na činjenici da je dužina ponika posle određenog vremena proizvod vremena potrebnog za klijanje, od-nosno za početni rast, i kasnije brzine rasta (Hampton, TeKrony, 1995).
Ovaj test se izvodi u četiri ponavljanja i potrebno je na filter papiru iscrtati deset paralelnih linija na razmaku od 2 cm Seme se poređa na centralnu liniju i nakon određenog broja dana inkubacije u optimalnim laboratoriskim uslovima za svaku vrstu, vrši se ocenjivanje porasta po-nika, odnosno sagledava se broj vrhova ponika unutar paralelnih linija. Dobijeni rezultati uvršćuju se u sledeću formulu:
(nx1 + nx2 + ... + nx5)L = __________________________________
N
gde je:L – srednja dužina ponika (cm),n – broj vrhova ponika unutar paralelnih linija,x – rastojanje linije od centralne linije (cm),N –broj ocenjivanih ponika. Kod poređenja partija semena iste sorte, one partije koje imaju veću
srednju dužinu imaju i veći vigor (Hampton i TeKrony, 1995). Različite sorte jedne biljne vrste mogu da imaju nasledno različite brzine rasta ponika, koje ne moraju biti povezane sa nicanjem u polju i zbog toga bi upoređivanja trebalo vršiti samo unutar jedne sorte (Perry, 1981).
Danas se ovaj test koristi kod velikog broja vrsta, ali najbolji rezultati dobijaju se kod vrsta koje imaju prav nadzemni deo, kao što su žita i trave.
Hladni (cold) test
Hladni test je najstariji vigor test i primenjuje se od dvadesetih go-dina XX veka. Smatra se da je jedan od najpreciznijih i najčešće korišće-nih vigor testova, posebno u Severnoj Americi. Danas se ovaj vigor test uglavnom koristi za ispitivanje životne sposobnosti semena kukuruza, koji se seje rano u proleće kada su temperature vazduha i zemljišta još uvek niske. Ovaj test se može primenjivati i kod drugih biljnih vrsta. Prema ispitivanjima Vujaković et al. (2011) pozitivan korelacioni odnos između poljske klijavosti i hladnog testa je utvrđen kod stočnog graška,

250
Semenarstvo
pa bi se ovaj test mogao koristiti kao dopunski test za dobijanje što po-uzdanijih rezultata o klijavosti semena.
Hladni test ima i druge mogućnosti upotrebe (AOSA 2002):1. procenu efekta primene fungicida;2. odabir sorte i partije semena pogodne za ranu setvu;3. procenu fizioloških oštećenja nastalih usled produženog skladi-
štenja u nepovoljnim uslovima, oštećenja od mraza ili suše;4. merenje uticaja mehaničkih oštećenja na klijanje u hladnom i
vlažnom zemljištu.Pri izvođenju ovog testa seme se izlaže kombinovanim stresnim
uslovima, niskoj temperaturi i patogenima. Uzorci se inkubiraju na tem-peraturama od oko 10°C u trajanju od sedam dana. Nakon ovog perioda seme se stavlja na 20°C ili 25°C (u zavisnosti od biljne vrste) gde ostaje četiri ili šest dana. Kao supstrat koristi se zemlja sa parcela na kojima je prethodne godine uzgajana ispitivana biljna vrsta, jer ona sadrži odre-đene prouzrokovače bolesti. Ponik se ocenjuje po istom kriterijumu kao kod standardnog laboratorijskog metoda (Hampton and TeKrony 1995).
Postoje i modifikovane metode hladnog testa gde se kao supstrat uzima mešavina zemlje i peska ili samo pesak. Tada je seme izloženo samo niskim temperaturama, dok je uticaj patogena potpuno zanema-ren. Kao supstrat može se koristiti i mešavina sterilisane gline i peska, a seme se tada ređa okretanjem klice ka dole, ka supstratu. Ovakav postu-pak predstavlja ispitivanje otpornosti semena na aerobne uslove.
Veliki nedostatak hladnog testa jeste varijabilnost rezultata zbog primene različitih tipova zemljišta. Nijestein i Kruse (2000) iznose da se najmanje variranje rezultata dobija kada se koristi pesak kao supstrat.
Uslovi proizvodnje utiču na životnu sposobnost semena. Više vred-nosti klijavosti semena primenom hladnog testa dobijene su kod seme-na proizvedenog u uslovima navodnjavanja u odnosu na seme proizve-deno u uslovima suvog ratarenja (Vujaković et al., 2008).
Test ubrzanog starenja
Proizvođači semena se svake godine susreću sa velikim brojem proble-ma vezanih za proizvodnju, doradu, skladištenje i prodaju semena. Veoma važno pitanje koje se mora rešavati svake godine jeste koje partije semena bi trebalo plasirati na tržište, a koje bi se bezbedno mogle sačuvati za nared-nu sezonu. Ova odluka je posebno bitna u slučajevima niskih cena semena na tržištu, ili u slučajevima kada se žele formirati zalihe semena za nared-

251
Ispitivanje kvaliteta semena
nu godinu. Ova odluka se ne može doneti samo na osnovu informacija o procentu klijavosti, jer se uglavnom skladište partije sa sličnom, najčešće visokom, klijavošću. Takođe, klijavost semena se ne čuva podjednako ni pod istim uslovima skladištenja. Dopunske informacije rezultatima klija-vosti može dati test ubrzanog starenja (TUS), kojim se može vršiti provera životne sposobnosti semena. TUS omogućava da se utvrdi da li određena partija semena ima manju ili veću sposobnost očuvanja klijavosti i da rela-tivno precizno odredi period bezbednog čuvanja semena u skladištu.
Danas je test ubrzanog starenja jedan od najčešće korišćenih testova za ispitivanje vigor semena, pre svega zato što je pokazao dobru korela-ciju sa nicanjem u polju (Lovato et. al., 2001).
Kod TUS seme se u kratkom periodu izlaže dvostrukim stresnim uslovima, visokoj temperaturi i visokoj relativnoj vlažnosti, koji pred-stavljaju dva glavna prouzrokovača starenja semena. Seme sa visokim vigorom će se odupreti ovim ekstremnim uslovima i propadaće sporije nego seme sa niskim vigorom.
Ispitivano seme se stavlja u vodeno kupatilo, a vremenski period i temperatura zavise od biljne vrste (tab. 7). Relativna vlažnost vazduha mora biti 97%–100%. Nakon ovog perioda, primenjuje se standardni test klijavosti.Uslovi u kojima se vrši ovaj test moraju biti strogo kontrolisani jer i male promene u temperaturi mogu imati uticaja na rezultate klja-vosti. Prema istraživanjima Vujaković et al. (2011) variranje temperature nije uticalo na klijavost semena, ali je klijavost značajno zavisila od dužine tretmana. Sa povećanjem dužine tretmana došlo je do značajnog smanje-nja klijavosti semena kod sve tri ispitivane vrste grahorice.Tabela 7. Preporučene temerature i vreme za test ubrzanog starenja kod nekih biljnih vrsta Table 7. Recommended temerature and time for the accelerated aging test in some
plant species (Hampton, TeKrony, 1995)
Biljna vrsta Temeratura (°C) Trajanje (h)Allium cepa L. 42 72Glycine max L 41 72Lolium perene L. 41 48Nicotiana tabacum L. 43 72Quercus nigra L. 41 108Trifolium pratense L. 41 72Triticum aestivum L. 41 72Zea mays L. 42 96

252
Semenarstvo
Često se koristi i modifikovani test ubrzanog starenja gde je voda u vodenom kupatilu zasićena solima. Na ovaj način onemogućava se preterano usvajanje vode i samim tim propadanje semena pri izlaganju stresnim uslovima.
Hiltner test
Hiltner test je prvobitno korišćen za utvrđivanje zaraženosti seme-na patogenim gljivama, uglavnom iz roda Fusarium. Nakon što je pri-mećeno da je koleoptil sa inficiranog proklijalog semena bio kratak i nije bio u stanju da probije sloj sitno lomljene cigle bez vidljivog ošteće-nja, ovaj test se počeo razvijati u dva pravca. Prvo, kao test za ispitivanje zdravstvenog stanja semena, kada je otkriveno da temperature niže od 20°C pospešuju razvoj patogenih gljiva. Drugo, kao vigor test, pošto je utvrđeno da test otkriva i oštećenja drugačija od onih koje prouzrokuju gljive, a koja onemogućavaju normalan rast ponika. Hiltner test se kori-sti za ispitivanje vigora semena kod žita (Vujaković et al., 2003b).
Hiltner test se zasniva na činjenici da je oštećeno seme (fiziološka oštećenja, oštećenja od mraza, tretiranje semena fungicidima i slično) često slabo i nesposobno da izdrži nepovoljne uslove za vreme klijanja. Naime, sloj kamenčića ili sitno lomljene cigle koji se koristi u ovom testu izaziva fizički stres i prepreku klijanju. U ovako nepovoljnim uslovima samo seme sa visokim vigorom će obrazovati tipičan ponik (Hampton and TeKrony, 1995) (sl. 22).
Slika 22. Hiltner test / Figure 22. Hiltner test (Jovičić, 2010)

253
Ispitivanje kvaliteta semena
Ovaj test nalazi primenu samo u pojedinim laboratorijama u Evro-pi, a osnovi razlog za to je što je korelacija između dobijenih rezultata i nicanja u polju varijabilna. Takođe, ovaj test ponekad ne uspeva da da dodatne informacije o kvalitetu semena u odnosu na standarni test kli-javosti (Hampton and TeKrony 1995).
Biohemijski testoviTetrazolijum test
Tetrazolijum test je razvijen 1950. godine kao test za dobijanje brze opšte procene vitalnosti semena, posebno kod vrsta kod kojih je izraže-na dormantnost i kod kojih bi test klijavosti trajao suviše dugo. Ovaj test je našao veliku primenu (Hampton and TeKrony 1995):
– kada setva treba da se izvede odmah posle žetve, kod jako dor-mantnog semena;
– kod semena koje sporo klija;– kada je potrebno brzo doći do informacije o potencijalnoj klija-
vosti semena;– radi utvrđivanja različitih tipova oštećenja nastalih pri žetvi ili do-
radi (toplotna, mehanička ili oštećenja od insekata);– radi rešavanja problema koji se mogu javiti prilikom ispitivanja
klijavosti, na primer kada su nejasni razlozi pojave atipičnosti, kada se sumnja na negativan efekat tretiranja pesticidima i slično.
Tetrazolijum test se zasniva na redukciji bezbojnog rastvora 2,3,5 – trifeniltetrazolim hlorida ili bromida u nerastvorljiv 2,3,5 – trifenilfor-mazan crvene boje. Ovaj rastvor se ponaša kao indikator za otkrivanje redukcionih procesa koji se odvijaju u živim delovima semena. Unutar semena tetrazolijum usvaja vodonik iz dehidrogenaze. Hidrogenizaci-jom tetrazolijuma, u živim ćelijama se stvara crvena, stabila supstanca formazan, koja boji žive delove semena (ISTA, 2011).
Seme se natapa u vodi jer se nabubrelo seme manje lomi i lakše seče u odnosu na suvo seme, a i bojenje je ujednačenije. Kod mnogih biljnih vrsta neophodno je tkivo osloboditi pre bojenja da bi se omogućio lak-ši ulazak boje u tkivo. Oslobađanje tkiva moguće je izvršiti skidanjem semenjače, probijanjem, uzdužnim ili poprečnim presecanjem neesen-cijalnog dela semena. Pripremljeno seme potapa se u 0,5%–1% rastvor tetrazolijuma. Važno je da seme bude potpuno prekriveno rastvorom i

254
Semenarstvo
da ne bude izloženo direktnoj svetlosti. Nakon vremena potrebnog za bojenje (zavisi od biljne vrste) vrši se ocenjivanje obojenosti (sl. 23).
Prilikom ispitivanja vitalno seme bi putem biohemijske aktivnosti trebalo da ispolji svoj potencijal za formiranje tipičnih ponika. Nevi-talno seme pokazuje nedostake koji sprečavaju pojavu tipičnog ponika (ISTA 2011).
Vitalno seme treba da pokaže obojenost svih tkiva čija je vitalnost neophodna za normalan razvoj klijanaca. Osim potpuno obojenih, vi-talnih semena, i potpuno neobojenih, nevitalnih semena, mogu se javiti i delimično obojena semena. Zavisno od vrste, mala neobojena mesta nekih delova tkiva se mogu prihvatiti. Mesto, veličina neobojenih obla-sti, a ponekad i intenzitet obojenosti, određuju da li se neko seme sma-tra vitalnim ili ne (sl. 24).
Tetrazolijum test ima i nekoliko nedostataka:1. daje previsoke rezultate vitalnosti, tj. u okviru vitalnog semena ne
može se razdvojiti seme koje će dati tipičan, a koje atipičan ponik;2. ne mogu se jasno uočiti potencijalne atipičnosti ponika (na pri-
mer rascepljen koleoptil, pojava negativnog geotropizna i slično);3. zahteva specijalno obučeno osoblje;4. ne može se otkriti prisustvo patogena niti fitotoksično dejstvo
fungicida.
Slika 23. Tetrazolijum test primenjen na semenu suncokreta (levo) i šematski prikaz nekoliko mogućnosti bojenja semena suncokreta (desno)
Figure 23. Tetrazolium test applied to the sunflower seed (left) (Jovičić, 2008) and schematic view of several options staining sunflower seeds (right) (ISTA, 2008)

255
Ispitivanje kvaliteta semena
Konduktometrijski test
Konduktometrijski test (test električne provodljivosti) je prvi put upotrebljen 1928. godine da bi se utvrdilo zašto grašak u laboratorij-skim uslovima pokazuje visoku, a u polju nisku klijavost. Zasniva se na činjenici da seme koje klija ispušta supstance kao što su šećeri i druge organske materije, kao i elektrolite u zemljišni rastvor, što može povećati aktivnost zemljišnih gljiva. Ta povećana aktivnost gljiva može negativno uticati na klijanje i porast ponika, posebno u hladnim i vlažnim uslovi-ma kada je klijanje i onako otežano (Matthews and Bradnock, 1968).
Kasnije je utvrđeno da biohemijske promene ćelijskih membrana ili fizička oštećenja semena imaju najveći uticaj na količinu otpuštenih elektrolita, a samim tim i na određivanje vigora semena. Što seme ima veću sposobnost da brže uspostavi normalno funkcionisanje ćelijskih membrana, to će otpuštanje elektrolita biti manje, odnosno seme ima veći vigor. Dakle, količina otpuštenih elektrolita iz semena sa visokim vigorom je manja u odnosu na seme sa niski vigorom.
Na električnu provodljivost rastvora utiču različiti faktori. Provod-ljivost rastvora raste sa porastom temerature, za oko 2% za svaki stepen, zbog čega se temeratura pri izvođenju ogleda mora strogo kontrolisati. Na provodljivost utiče i početan sadržaj vlage u semenu, jer veća pro-vodljivost pri manjoj vlažnosti može biti posledica oštećenja zidova će-lija usled natapanja semena vodom.
Slika 24. Tetrazolijum test primenjen na semenu pšenice: potencijalna klijavost (A), fakultativna klijavost (B), nevitalna semena (C)
Figure 24. Tetrazolium test applied to wheat seeds: potential germination (A), facultative germination (B), non-viable seeds (ISTA, 1985)
A B C

256
Semenarstvo
Seme koje se ispituje (vlažnost 10–14%) se potapa u dejonizovanu vodu u trajanju od 24h, na temperaturi od 20°C. Nakon tog perioda potrebno je odmah izmeriti provodljivost rastvora uz pomoć konduk-tometra, a dobijene podatke uvrstiti u formulu (Hampton and TeKrony 1995):
Provodljivost (mS/cmg–1) ____________________________________ = mS/cmg
Masa uzorka (g)
Ako je izračunata vrednost:< 25 mS/cmg – seme ima visok vigor, tj. seme je pogodno za ranu
setvu u nepovoljnim uslovima;25 – 29 mS/cmg – seme može da se koristi za ranu setvu, ali postoji
rizik ako nastanu nepovoljni uslovi;30 – 43 mS/cmg – seme nije pogodno za ranu setvu posebno u nepo-
voljnim uslovima;> 43 mS/cmg – seme ima nizak vigor, tj. nije pogodno za setvu.Konduktometrijski test omogućava brzo dobijanje objektivnih re-
zultata, i može biti sproveden u laboratoriji bez velikih zahteva za opre-mom i obučavanjem ljudi. Našao je veliku primenu kod ispitivanja vi-gora semena, pre svega zato što dobijeni rezultati pokazuju pozitivnu korelaciju sa nicanjem u polju (Matthews and Powell, 1981, prema Mi-lošević i Zlokolica, 1996).
Iako ne mogu da predvide nicanje u polju, vigor testovi su neop-hodni kod ispitivanja životne sposobnosti semena jer pružaju dodatnu informaciju o kvalitetu semena. Dalji razvoj vigor testova bi trebao da ide u pravcu stvaranja kompleksnog testa, koji bi obuhvatio veći broj nepovoljnih činilaca i čiji bi se rezultati prikazali karakteristikama klija-vosti (brzina, ujednačenost, energija, odnos tipičnih i atipičnih ponika) i pokazateljima porasta ponika (dužina, masa, suva materija) u funkciji vremena (Jovičić, 2008).

257
Ispitivanje kvaliteta semena
LITERATURA
AOSA (2002): Seed Vigour Testing Handbook, Contribution No 32 to the Handbook of Seed Testing Association of Official Seed Analysts, NE, USA
Barjaktarović R, Kobiljski B, Obreht D, Vapa Lj (2004): Mirovanje seme-na pšenice i uticaj boje zrna na stepen mirovanja. Sel. Semen., Vol X, br 1-4: 83-87
Basak O, Demir I, Mavi K, Matthews S (2006): Controlled deteriorati-on test for predicting seedling emergence and longevity of pepper (Capsicum annuum L.) seed lots. Seed Sci. Technol., 34: 723-734
Baskin C C, Baskin J M (2004): Determining dormancy - breaking and germination requirements from the fewest seeds. Strategies for Sur-vival, Island Press, Washington, 162-179
Basra A S (2006): Handbook of Seed Science and Technology. USA. Bould A (1986): Handbook of Seed Sampling. International Seed Te-
sting Association, Switzerland Dobrohotov V N (1961): Semena sornmh rasteni. Izdatelstvo seljsko-
hoznoi literaturi, žurnalov i plakatov. MoskvaDornbos D L (1995): Production environment and seed quality. In: Seed
Quality Basi Mechanisms and Agricultural Implications, (ed) Basra A S, Food Products Press. New York, London, Australia, 119-145
Durrant M J, Gummerson R J (1990): Factors associated with germina-tion of sugarbeet seed in the standard test establishment in the field. Seed Sci. and Technol, 18: 1-10
Grbić M (2003): Dormantnost i klijanje semena – mehanizmi, klasifi-kacije i postupci, Glasnik Šumarskog fakulteta, Beograd, 87: 25-49
Hampton J G, Te Krony D M (1995): Handbook of Vigour Test Methods, 3rd Edition, International Seed Testing Association, Switzerland
ISTA (2011): International Rules for Seed Testing. International Seed Testing Association, Switzerland
ISTA (2004): ISTA Handbook on Seed Sampling, 2nd Edition, (ed): Kruse M, International Seed Testing Association, Switzerland

258
Semenarstvo
ISTA (2003): Working sheets on tetrazolium testing Agricultural, Vege-tables and Horticultural Species, Vol I, International Seed Testing Association, Switzerland.
ISTA (1985): ISTA Handbook on Seedling Evaluation. International Seed Testing Association, Switzerland
Johansen R R, Wax L M (1978): Relationship of soybean germination and vigour test to field performance. Agron. J., 70: 273-276
Jovičić D (2008): Ispitivanje životne sposobnosti semena. Master rad, Poljoprivrdni fakultet, Novi Sad
Jovičić D, Marjanović Jeromela A, Vujaković M, Marinković R, Sakač Z, Nikolić Z, Milošević B (2011): Uticaj različitih doza NPK đubriva na kvalitet semena uljane repice. Rat. Pov., 48: 125-130
Karrfalt R (2004): Seed testing, Chapter 5 In: Woody plants seed manual (ed.) Rebecca G. Nisley
Kastori R (1984): Fiziologija semena. Matica srpska, Novi SadKlokočar-Šmit Z, Inđić D (1991): Negative effect of seed treatment an
efekat tretiranja semena. Seed Apotheosis, IV Monography, 107-113Kojić J, Stojadinović J, Mišović M (2010): Accreditation and control of
automatic seed sampler. Journal on Processing and Energy in agri-culture, 14: 58-60
Kronaveter Đ, Pal B (1994): Poznavanje semena najčešćih korova u se-menarstvu. Institut za ratarstvo i povrtarstvo, Novi Sad
Leopold A C, Vertucci C W (1989): Moisture as a regulator of physio-logical reaction in seeds. In. Seed Miosture, (eds. Stanwood P C, McDonald M B), Madison, Wisc, Crop Science Society of America
Lovato A, Noli E, Lovato A F S, Beltrami E, Grassi E (2001): Comparison between three cold test low temperatures, accelerated ageing test and field emergence on maize seed. Seed Symposium, ISTA Con-gres, Angers, France.
Manual for Testing Agricultural and Vegetable Seeds, Agriculture Handbook, No 30, Washington, 1952
Mathur S B, Kongsdal O (2003): Common Laboratory Seed Health Testing Methods for Detecting Fungi. International Seed Testing Association, Switzerland
Matthew S S, Bradnock W K: (1968). Relationship between seed exudation and emergence in peas and French beans. Horticultural Research 8: 89-93

259
Ispitivanje kvaliteta semena
McDonald J M B (1975): A review and evaluation of seed vigor test. Proc. Assoc: Off. Seed Anal., 65: 109-139
McDonald M B (1980): The history of seed vigour testing. J. Seed Tech., Vol 17: 93-100
Milošević M, Ćirović M (1994): Seme. Institut za ratarstvo i povrtar-stvo, Novi Sad
Milošević M, Zlokolica M, Hrustić M, Gvozdenović Đ, Jocković Đ (1996a): Mirovanje semena, Sel. Semen., Vol IV, broj 3-4: 97-104
Milošević M, Zlokolica M (1996b): Vigor semena. Sel. Semen, III, 1-2: 33-43
Milošević M, Vujaković M, Nikolić Z, Ignjatov M (2007): Kontrola kvali-teta semenskog materijala. Nauka osnova održivog razvoja, Društvo genetičara Srbije, Beograd, 95-124
Milošević M, Vujaković M, Karagić Đ (2010): Vigour tests as indicators of seed viability, Genetika, Vol 42, 1: 103-118
Nijenstein J H, Kruse M (2000): The potential for standardisation in cold testing of maize (Zea mays L.). Seed Sci. and Tehnol., 28: 837-851
Nijenstein H, Nydam J, Don R, McGill C (2007): ISTA Handbook on Moisture Determination. International Seed Testing Association. Switzerland
Nikolaeva M G (1977): Factors controlling the seed dormancy pattern. Khan A A (ed.) The Physiology and Biochemistry of Seed Dor-mancy and Germination. North Holland Publishing, Amsterdam, 51–74
Nyachiro J M, Clarke F R, Depaijw R M, Knox R E, Armstrong K C (2002): Temperature effects on seed germination and expression of seed dormancy in wheat. Euphytica, 126 (1): 129-133
Perry D A (1981): Report of Vigour Test Committee 1977-1980, Seed Sci. Technol, Vol 9: 115-126
Pravilniku o kvalitetu semena poljoprivrednog bilja Sl. list SFRJ br. 47/87 Schuurink R C , Van Beckum M M , Heidekamp F (1992): Modulation of
grain dormancy in barley by variation of plant growth conditions. Hereditas, 177: 137-143
Sun Q, Wang J, Sun B (2007): Advances on Seed Vigor Physiologi-cal and Genetic Mechanisms. Agricultural Sciences in China, 6(9): 1060-1066
Tekrony M D (1982): Seed vigor testing, J. Seed Technol., 8: 55-60

260
Semenarstvo
Vujaković M, Jovičić D, Karagić Đ, Katić S, Nikolić Z, Taški-Ajduković K, Milošević B (2010): Testing of field pea seed viability, Biotechno-logy in Animal Husbandry, Vol 26, special issue, Book 2: 209-215
Vujaković M, Jovičić D, Karagić Đ, Mikić A, Nikolić Z, Petrović D, Taš-ki-Ajduković K (2011): Pokazatelji životne sposobnosti semena ozi-mih grahorica (Vicia spp.). Rat. Pov., 48: 131-136
Vujaković M, Marjanović Jeromela A, Jovičić D, Marinković R, Nikolić Z, Crnobarac J, Taški–Ajduković K (2010): Uticaj prihrane na pri-nos i komponente kvaliteta semena uljane repice. Rat. Pov., 47, 2: 539-544
Vujaković M, Milošević M, Nikolić, Z, Taški-Ajduković K, Miladino-vić J, Ignjatov M, Dokić V (2008): Životna sposobnost semena soje proizvedene u uslovima sa i bez navodnjavanja. PTEP časopis za procesnu tehniku i energetiku u poljoprivredi, 1-2: 19-22
Vujaković M, Milošević M, Zlokolica M, Nikolić Z (2003a): Uticaj fun-gicida za tretiranje semena na porast ponika pšenice (Triticum ae-stivum L.) i aktivnosti superoksid dismutaze. Biljni lekar XXXI, 6: 564- 570
Vujaković M, Milošević M, Nikolić Z, Zlokolica M (2003b): Uticaj pre-parata za tretiranje semena na klijavost i vigor pšenice (Triticum aestivum L.). Časopis za procesnu tehniku i energetiku u poljopri- vredi – PTEP, 7, 3-4: 75-78
Vujaković M (2004): Ispitivanje kvaliteta semena. U: Se menarstvo, (ured) Milošević M, Malešević M, Naučni institut za ratarsvo i po-vrtarstvo, Novi Sad, 153-197
Shauveau S, Maill P (2001): Seed Sampling Workshop. ISTA Congress, Angers, Francuska

Maja Ignjatov, Dragana Petrović
Ispitivanje zdravstvenog stanja semena

262
Semenarstvo

263
Ispitivanje zdravstvenog stanja semena
UVOD
Zdravstveno stanje semena podrazumeva prisustvo ili odsustvo štetnih organizama prouzrokovača bolesti, kao što su gljive, bakterije i virusi, zatim prisustvo nematoda i insekata. Utvrđivanje zdravstvenog stanja semena je važno iz više razloga:
– inokulum na semenu može da dovede do progresivnog širenja zaraze u polju i da snizi komercijalnu vrednost useva,
– partije semena iz uvoza mogu da unesu zaraze u nove oblasti. Sto-ga, ispitivanja su neophodna da bi se ispunili karantinski zahtevi,
– ispitivanjem zdravstvenog stanja semena se može objasniti ocena klijanaca i uzroci slabe klijavosti ili stanje useva u polju i tako upotpuniti rezultate testa klijavosti i
– rezultati zdravstvenog stanja ukazuju na potrebu tretiranja, radi uništavanja patogena semena ili smanjenja rizika od prenošenja kako infekcije tako i štetočina.
Validne metode za ispitivanje zdravstvenog stanja semena, odobre-ne od strane ISTA (International Seed Testing Association) detaljno su opisane u poglavlju 7 ISTA Međunarodnih pravila za ispitivanje seme-na. Oni se obično zasnivaju na jednom domaćinu i jednom patogenu, ali mogu biti uključeni i multi-patogeni metodi. Stepen zaraženosti se-mena određuje se prema važećim propisima (Pravilnik o zdravstvenom pregledu useva i objekata za proizvodnju semena, rasada i sadnog ma-terijala i zdravstvenom pregledu semena, rasada i sadnog materijala - Službeni glasnik RS 119/07).
Za ispitivanje zdravstvenog stanja semena dostupno je više metoda koje se razlikuju po osetljivosti i ponovljivosti, dužini vremena potreb-nog za njihovo izvođenje kao i po zahtevu za obukom i opremom. Koji će se metod koristiti zavisi od biljne vrste, patogena i uslova koji se is-pituju i svrhe testa. Izbor metode i ocena rezultata zahtevaju znanje i iskustvo u vezi sa postojećim metodama. Kao radni uzorak može se, zavisno od metoda ispitivanja, uzeti ceo prosečan uzorak ili njegov deo. Uzorak treba da bude upakovan i dostavljen laboratoriji na način da se na njegovo zdravstveno stanje ne utiče.

264
Semenarstvo
Kvalitetno seme je jedan od osnovnih preduslova za uspešnu biljnu proizvodnju. U cilju ispitivanja zdravstvenog stanja semena primenju-ju se različite fitopatološke metode. Njihova uspešnost zavisi od stanja semena, virulentnosti patogena, odnosno od količine inokuluma u se-menu, uslova izvođenja testa (dužine inkubacije, vrste podloge, tempe-rature, vlažnosti, svetlosti i dr.).
Prouzrokovači biljnih bolesti koje izazivaju gljive, bakterije i virusi, svake godine nanose velike štete biljnoj proizvodnji. Veliki broj biljnih patogena prenosi se semenom. Utvrđivanje patogena semena je veoma bitno da bi se sprečilo dalje širenje bolesti. U zavisnosti od vrste semena, prirode prouzrokovača bolesti kao i spoljnih činilaca, rezultira i visina štete na gajenim biljnim vrstama (Jovićević and Milošević, 1990).
Od značaja su i genetska svojstva patogena, kao i otpornost, odno-sno osetljivost biljke domaćina. Za mnoge patogene gljive, bakterije a pogotovo viruse potrebno je prisustvo prenosioca bolesti. Veliki značaj u širenju spora mikroorganizama imaju i kišne kapi, vetar, domaće živo-tinje, razna oruđa, poljoprivredne mašine i dr.
Patogeni se mogu razvrstati u nekoliko kategorija, a osnovna podela se zasniva prema prirodi prouzrokovača bolesti kao što su gljive, bakte-rije, virusi i mikoplazme. Najčešća oboljenja semena prouzrokuju gljive. Prouzrokovači bolesti dovode do različitih promena na samom semenu, koje se mogu okularno zapaziti, kao što su: potpuno uništenje semena, promena boje semena, trulež, deformacija i šturost semena, pegavost, sklerotizacija, stromatifikacija i dr. Takođe, bogat razvoj saprofitskih gljiva, plesni, uključujući „skladišne gljive“, u ispitivanju (testu) može da ukaže na to da seme nije dobrog kvaliteta zbog nepovoljnih uslova tokom žetve, dorade, skladištenja ili zbog starenja.
Pored konvencionalnih laboratorijskih metoda koje se koriste u identifikaciji patogena, danas su razvijene i nove savremene metode. U njih spadaju serološke i molekularne metode čija je glavna odlika da su brze, pouzdane, pogodne za analizu velikog broja uzoraka i re-lativno jeftine. Od seroloških metoda najviše je u upotrebi ELISA test, IF test, metod aglutinacije, a od molekularnih PCR metod (Ignjatov et al., 2006).
Početkom devedesetih godina sprovedeni su prvi PCR testovi za biljne patogene, testiranjem biljaka i semena (Rassmusen and Wulff 1991, Prosen et al. 1993). Ova metoda je veoma osetljiva i potencijal-

265
Ispitivanje zdravstvenog stanja semena
no visoko specifična. Početni korak u sprovođenju svih ovih metoda je priprema i izdvajanje RNK ili DNK patogena. Razvijeno je više metoda izdvajanja i prečišćavanja DNK i RNK iz čistih kultura, iz uzoraka bilj-nog tkiva, iz zemljišta, pa i iz uzoraka vazduha.

266
Semenarstvo
METODE ZA ISPITIVANJE ZDRAVSTVENOG STANJA SEMENA
Metode koje se koriste za ispitivanje zdravstvenog stanja semena uslovno se mogu podeliti na one koje se izvode bez prethodne inkubacije i uz inkubaciju semena. U prvu grupu spadaju:
– makroskopski pregled semena,– ispiranje semena,– metod brojanja embriona i– luminiscencija semena.Metode kod kojih je potrebno obaviti inkubaciju semena u određe-
nom vremenskom periodu, mogu se izvoditi na veštačkim i prirodnim podlogama. Pri primeni veštačkih ili sintetičkih podloga razlikuju se:
– metoda na filter-papiru,– metoda s prethodnim zamrzavanjem i– metoda na hranljivoj podlozi (sl. 1).Da bi se pri ispitivanju zdravstvenog stanja semena dobila podloga
što približnija prirodnim uslovima razvoja gljiva, seme se naklijava u sterilnom pesku, zemlji ili nekoj sličnoj sredini. U ovu grupu ubrajaju se:
– test u pesku, – test u zemlji i– Hiltner test.
Slika 1. Izolovana bakterija na hranjivoj podlozi /Figure 1. Isolated bacteria on nutritive media (foto: Courtesy, J. K., Pataky,C. J. K.)

267
Ispitivanje zdravstvenog stanja semena
Metode identifikacije bez prethodne inkubacije semena
Metoda makroskopskog pregleda semena
Ova metoda zasniva se na direktnom posmatranju uzorka semena i otkrivanju pojedinih tipova bolesti. Na taj način određuje se i čisto-ća semena na prisustvo inertnih materija i korova. Pored posmatranja semena golim okom, koristi se stereomikroskop, uvećanja x 40-50. Na semenu mogu se konstatovati različiti organi gljiva prouzrokovača bo-lesti a to su: acervuli, piknidi, peritecije, sklerocije, spore, gale, tumori itd. Neki patogeni stvaraju vrlo specifične i uočljive simptome na se-menu. Tako na primer, na semenu soje pojava ljubičaste boje ukazuje na prisustvo Cercospora kikuchii (sl. 2). Na semenu pšenice može doći do obezbojavanja i pojave mrkih lezija koje ukazuju na gljivu Bipolaris sorokiniana. A pojava ružičaste boje ukazuje na prisustvo vrsta gljiva iz roda Fusarium (npr. Fusarium culmorum, Fusarium graminearum itd.) (Jovićević and Milošević 1990).
Slika 2. Seme soje zaraženo gljivom Cercospora kikuchii
Figure 2. Soybean seeds infected by Cercospora kikuchii (Petrović, 2007)
Metoda ispiranja semena
Ovom metodom određuje se prisustvo spora gljiva na površini semena, kao što su, na pri-mer, hlamidospore glavnica (Tilletia spp.) i spore drugih gljiva (Peronospora manshurica, Puccinia spp., Ustilago spp., Uromyces spp., itd.). Pomoću ove metode ne određuje se kvantitativna, već samo kvalitativna zaraženost semena pojedinim patogenima. Zbog toga ova metoda nema širu praktičnu primenu.

268
Semenarstvo
Metod brojanja embriona
Ovom metodom određuje se prisustvo gljive Ustilago nuda na se-menu ječma (sl. 3, 3a). Postupak ocene zaraženosti embriona sastoji se u prethodnom odvajanju embriona od ostatka semena, pomoću 5% ra-stvora NaOH. Uzorak od 1000 semena je potrebno potopiti u jedan li-tar sveže pripremljenog rastvora NaOH. Inkubacija potopljenog uzorka traje 24 časa na temperaturi od 20ºC. Uzorak se nakon inkubacije pre-bacuje u odgovarajuću posudu pri čemu se seme ispira toplom vodom radi razdvajanja embriona iz semena. Sakupljanje odvojenih embrio-na vrši se pomoću sita promera perforacija od 1 mm. Radi dodatnog prečišćavanja i odvajanja embriona od perikarpa semena, embrioni se prebacuju u prethodno pripremljen rastvor glicerola i vode u odnosu 1:1. Nakon toga vrši se zagrevanje prečišćenih embriona u rastvoru be-zvodnog laktofenola do tačke ključanja, u trajanju od 30 sekundi. Nakon ključanja, radi lakšeg mikroskopiranja embrioni se potapaju u svež ra-stvor glicerola. Nakon toga se pojedinačni embrioni postavljaju na mi-kroskopsku pločicu i mikroskopiraju (Ignjatov et al., 2011a).
Slika 3. Izgled embriona semena ječma zaraženog gljivom Ustilago nuda
Figure 3. The appearance of the embryo of barley seeds infected by Ustilago nuda
(Ignjatov 2008)
Slika 3a. Izgled embriona zdravog semena ječmaFigure 3a. The appearance of the embryo of healthy barley seeds (Ignjatov 2008)

269
Ispitivanje zdravstvenog stanja semena
Metoda luminiscencije semena
Zasniva se na ispoljavanju fotoluminiscentnih svojstava tkiva se-mena i gljiva pod dejstvom ultravioletnih zraka. Razlikuju se dve vrste fotoluminiscencije: fluoroscencija i fosforescencija. Pojava poznata pod imenom fluoroscencija zasniva se na sposobnosti nekih tela da pod uti-cajem svetlosne energije, koja pada na njihovu površinu, emituju sve-tlost i koristi se za otkrivanje patogena na semenu. Tanak sloj semena se rasporedi ispod kvarcne lampe koja je izvor UV svetla na crnoj podlozi. Zaraženo i zdravo seme se razlikuje po sposobnosti primanja i odbijanja UV svetla, pa emituje svetlost različite talasne dužine, odnosno boje. U sličaju prisustva Ascochyta spp. na semenu graška, reflektuje se žutoze-lena svetlost (Maceljski and Kišpatić 1987).
Metode uz prethodnu inkubaciju semena
Metoda na filter papiru
Ova metoda zasniva se na stimulaciji razvića mikroorganizama u zaraženom semenu. Osim određivanja vrste prouzrokovača bolesti, ovom metodom određuje se i stepen zaraženosti semena. U Petri posu-dama prečnika 9 cm sa tri sterilna filter papira navlažena sterilisanom vodom, postavljaju se semena u odgovarajućem broju u zavisnosti od biljne vrste i veličine semena (5, 10, 25 i 50 zrna) u razmaku od 2cm (sl. 4). Potrebno je postaviti 400 semena po uzorku.
Slika 4. Pravilan raspored od 5, 10, 25 ili 50 semena po Petri kutiji.
Figure 4. Regular schedule of 5, 10, 25 or 50 seeds per Petri dishes (Petrović 2009).

270
Semenarstvo
Nakon inkubacije koja traje 7 dana na 22±2ºC, svako seme se pa-žljivo pregleda pomoću stereomikroskopa a po potrebi prave se mikro-skopski preparati radi što tačnije identifikacije patogena (Mathur and Kongsdal 2003).
Identifikacija parazita pomoću metode filter-papira pogodna je za veći broj gljiva, koje se najčešće sreću na semenu iz roda Helminthos-porium (sl. 5), Alternaria, Stemphylium, Ascochyta, Phoma, Drechslera, Fusarium (sl. 6), Cercospora, Cladosporium, Colletotrichum, Botrytis i Sclerotinia sclerotiorum (sl. 7). Veoma često javljaju se saprofitne gljive iz roda Rhyzopus, Aspergillus, Penicillium (sl. 8), Mucor i dr.
Ukoliko nije moguće determinisati patogena ovom metodom pri-stupa se postupku izolacije patogena na hranljivoj podlozi.
Slika 5. Seme ječma zaraženo gljivom iz roda Helminthosporium (filter papir metod)
Figure 5. Barley seeds infected by Helminthosporium spp. (blotter method)
(Ignjatov 2009)
Slika 7. Seme soje zaraženo gljivom Sclerotinia sclerotiorum (filter papir metod)
Figure 7. Soybean seeds infected by Sclerotinia sclerotiorum (blotter method)
(Petrović 2009)
Slika 6 Seme pšenice zaraženo gljivom iz roda Fusarium (filter papir metod)Figure 6. Wheat seeds infected by
Fusarium spp. (blotter method) (Ignjatov 2009)
Slika 8. Seme pšenice zaraženo gljivom iz roda Penicillium (filter papir metod)Figure 8. Wheat seeds infected by Penicillium spp. (blotter method)
(Petrović 2009)

271
Ispitivanje zdravstvenog stanja semena
Metoda na filter papiru sa zamrzavanjem (The deep freeze method)
Modifikaciju metode s filter papirom primenio je Limonard (1966). Seme se stavi na filter papir kako je opisano u prethodnoj metodi. U zavisnosti od biljne vrste, seme se inkubira 2 do 4 dana a zatim se preko noći izlaže temperaturi od -20°C u trajanju od 24 h kako bi došlo do zaustavljanja klijanja i nesmetanog razvoja patogena. Nakon isteka pe-rioda zamrzavanja, narednih pet do sedam dana seme se izlaže tempe-raturi od 20°C na NUV (Near visible ultraviolet) svetlu. Ovaj postupak uspešno se primenjuje za rutinske analize za ispitivanje gljiva iz roda Drechslera i Fusarium na semenu žita i trava, kao i za sitno seme povrća.
Metoda na hranljivoj podlozi
Dezinfekcijom semena i njegovim zasejavanjem na odgovarajuću hranljivu podlogu, omogućuje se mikroorganizmima iz semena da obra-zuju kolonije i da fruktificiraju. Na hranljivim podlogama lako se mogu uo-čavati osobine kolonija mikroorganizama, tj. njihov izgled, boja, intenzitet porasta itd. Detekcija vrste parazita vrši se pregledom preparata uz pomoć mikroskopa većeg uveličanja, pri čemu se posmatraju reproduktivni organi gljive koji su veoma karakteristični, ne samo za pojedine rodove, već i za neke vrste (sl. 9, 10) (Petrović et al., 2010a, 2011, Ignjatov et al., 2011b).
Pre zasejavanja na hranljivu podlogu, seme se dezinfikuje, a u tu svrhu se najčešće koristi natrijum-hipohlorit (NaOCl) u trajanju 1–10 minuta u zavisnosti od biljne vrste i patogena koji se ispituje. Vreme inkubacije i temperatura zavise od bioloških zahteva mikroorganizama.
Slika 9. Konidija od Alternaria dauci Figure 9. Conidia of Alternaria dauci
(Ignjatov 2009)
Slika 10. Konidija od Dreschslera oryzaeFigure 10. Conidia of Dreschslera oryzae
(Petrović 2009)

272
Semenarstvo
Izolacija mikroorganizama obavlja se na nekoj od hranljivih podloga. Pojedine podloge koriste se za gajenje većine patogena pa se stoga smatra-ju univerzalnim, ali postoje i one specifične za određenog patogena pa se smatraju selektivnim podlogama. Podela podloga koje se koriste u labora-torijskom radu može se obaviti na osnovu različitih kriterijuma, ali se kao najvažniji ističe onaj koji ukazuje na prirodu izvora hranljivih materija. Prema tome postoje prirodne, polusintetičke i sintetičke podloge.
Prirodne podloge su sastavljene od različitih delova biljaka (stablo, list, plod, seme). Ove podloge se lako pripremaju, jeftinije su, a što je najvažnije na njima se razvija veliki broj gljiva.
Polusintetičke podloge su delimično sastavljene od prirodnog ma-terijala, a delimično od jedinjenja poznatog hemijskog sastava. Kao primer takve podloge je krompir-dekstrozni agar (PDA), koja se pored malt-agar podloge (MA) smatra najuniverzalnijom i najpogodnijom za razvoj gljiva. PDA podloga koristi se u identifikaciji velikog broja pa-togenih gljiva: Sclerotinia sclerotiorum (sl. 11) (Petrović et al., 2009), Trichotecium spp., Verticillium spp., Aspergillus spp., Rhizopus spp. (Ignjatov et al., 2008a), Fusarium spp. (sl. 12) (Milošević et al., 2007a, 2007b, 2008), Helminthosporim spp. (Milošević et al., 2007a, 2007b).
Hemijski sastav sintetičkih podloga je poznat, te su one pogodne za izučavanje fiziologije gljiva. Pored hemijskog sastava podloge, na razvoj i sporonošenje mikroorganizama utiče njihova pH vrednost, tempera-tura i vlažnost sredine.
Slika 11. Micelija gljive Sclerotinia sclerotiorum na PDA podlozi
Figure 11. Mycelium of Sclerotinia sclerotiorum on PDA medium
(Petrović 2008)
Slika 12. Micelija gljive Fusarium graminearum na PDA podlozi
Figure 12. Mycelium of Fusarium graminearum on PDA medium
(Ignjatov 2008)

273
Ispitivanje zdravstvenog stanja semena
Za razviće i razmnožavanje bakterija služe posebne podloge. Kao osnovna podloga za izolaciju bakterija koristi se mesopeptonski agar (MPA). Selektivne podloge – treba da zadovolje dva kriterijuma: na toj podlozi treba da se razvije veći broj bakterijskih kolonija nego npr. na MPA i da onemogući razvoj drugih mikroorganizama što postižemo dodavanjem antibiotika (streptomycin, cycloxamid, vankomycin i dr.).
Postoji veliki broj podloga koje se koriste u cilju izolacije bakterija ali neke od najčešće korišćenih su: mesopeptonski agar MPA, hranljivi agar - NA, hranljivi agar sa saharozom - NSA, King B (za fluorescen-tne bakterije), dekstrozni agar sa kvaščevim ekstraktom i CaCO3 - YDC, podloga sa dodatkom Tweena 80 - Tween B, podloga sa sukciničnom i kiničnom kiselinom za izolaciju bakterije X. euvesicatoria (mSQ) (sl. 13); trifenil tetrazolijum hloridni agar TTCA, krompir dekstrozni agar sa kalcijum-karbonatom PDA + CaCO3 i dr. (Lelliott and Stead 1987, Schaad et al. 2001, Arsenijević 1997). Nakon spoljne dezinfekcije seme-na, iz suspenzije dobijene gnječenjem semena ili isečenih delova biljnog materijala u sterilisanom avanu, vrši se zasejavanje pomoću bakteriološ-ke petlje na odgovarajuću hranljivu podlogu, standardnom metodom razmaza po površini hranljive podloge, u Petri kutijama prečnika 90 mm. Dužina inkubacije zavisi od roda bakterije. Pojava Erwinia može se pratiti nakon 24 časa, Pseudomonas posle 24–48 časova, Xanthomo-nas od 48 časova do sedam dana, a Corynebacterium između 72 časa i sedam dana (Ignjatov et al., 2010).
Slika 13. Kolonije bakterije X. euvesicatoria na YDC hranljivoj podlozi
Figure 13. Colonies of X. euvesicatoria on YDC
(Ignjatov 2009)
Slika 14. Kolonije bakterije Pseudomonas syringae pv. glycinea na MPA podloziFigure 14. Colonies of Pseudomonas
syringae pv. glycinea on MPA (Ignjatov 2007)

274
Semenarstvo
Izolacijom patogena iz obolelih listova soje, na mesopeptonsku podlogu (MPA) i podlogu od hranljivog agara sa saharozom (NSA), dva do tri dana nakon izvedene izolacije obrazuju su se krupne, sjajne, bele i ispupčene kolonije bakterija karakteristične za rod Pseudomonas (sl. 14). Kasnijim ispitivanjima patogenih i bakterioloških karakteristika dokazano je da se radi o prouzrokovaču bakteriozne pegavosti soje P. s. pv. glycinea (Ignjatov et al., 2007a).
Patogenost ispitivanih izolata (bakterije)
Izdvajanje bakterija u čistu kulturu i ispitivanje njihovih morfoloških i odgajivačkih osobina bez provere patogenosti dobijenih izolata, pred-stavlja samo početnu stepenicu u nizu predstojećih operacija koje je ne-ophodno proslediti u cilju identifikacije parazita do naziva vrste. Patoge-nost, kao osnovno svojstvo fitopatogenih bakterija, ukazuje nam samo na to da li su dobijene kulture parazitnih ili saprofitnih svojstava. Poznato je naime, da izumrle ostatke obolelih biljnih organa i tkiva naseljavaju i razne saprofitne vrste, koje su ponekad vrlo sličnih morfoloških i drugih osobina u poređenju s fitopatogenim bakterijama. Patogenost izolovanih bakterija iz obolelih tkiva možemo proveriti na biljkama na više načina. Osnovno pravilo je da izolovana bakterija dobijena iz obolele biljke pro-uzrokuje iste ili slične simptome na inokulisanoj biljnoj vrsti odakle je mikroorganizam i izdvojen u čistu kulturu. Prema tome, inokulaciju bi trebalo vršiti na biljci domaćinu – vrsti s koje su bakterije i dobijene.
Pomoću hiprsenzibilne reakcije, moguće je veoma brzo uz pomoć lista duvana ili lista nekih drugih biljaka utvrditi patogenost svih pred-stavnika roda Pseudomonas i Xanthomonas (Klement et al. 1990). Sus-penzija bakterija koncentracije 106 cfu/ml (broj ćelija/ml) ubrizgava se u međunervalni deo lisnog tkiva, u tkivo mezofila lista, a pojava hipersen-zibilne reakcije ocenjuje se nakon 24 časa. Kao kontrola koriste se listovi infiltrirani sterilisanom, destilovanom vodom.
Na ovaj način, patogenost velikog broja izolata bakterije P. s. pv. glycinea dobijen iz obolelih biljnih delova biljaka soje poreklom iz razli-čitih proizvođačkih regiona u Vojvodini, potvrđena je kako na biljkama soje (Ignjatov et al., 2008b) tako i na osnovu hipersenzibilne reakcije na biljkama duvana sorte Samsun (Ignjatov et al., 2008c).
Brza provera patogenosti sojeva bakterija može se izvršiti i inokula-cijom metodom uboda, na pojedinim organima biljaka, postavljenim u

275
Ispitivanje zdravstvenog stanja semena
vlažnu sredinu (kotiledoni listići biljaka, mahune pasulja, lišće breskve, zeleni plodovi voćaka i dr.) (Ignjatov et al., 2008d) (sl. 15).
Morfološke i odgajivačke odlike fitopatogenih bakterija
– Oblik (tačkast, okrugao, končast, rizoidan, nepravilan)– Posedovanje i raspored cilija– Bojenje ili reakcija po Gramu (Gram+ , Gram-) – Površina kolonija (glatka i sjajna, rapava i bez sjaja, ravna, ispupčena)– Izgled i boja kolonije – Struktura (zrnasta, pahuljasta, sluzava, voskasta, kožasta)– Pojava fluorescencije
Morfološke odlike, usled jednostavne građe bakterija, imaju ograničen značaj u identifikaciji ovih parazita. Ipak, neka njihova svojstva kao što su posedovanje i raspored cilija ili odsustvo ovih organa, bojenje po Gramu, prisustvo ili odsustvo spora (fitopatogene bakterije su asporogene) može nam korisno poslužiti za određivanje ovih parazita, bar do naziva roda.
Slika 15. Mrke, ugnute pege na zelenim plodovima višnje i trešnje u zoni uboda nakon veštačke inokulacije suspenzijom bakterije P. s. pv. syringae
Figure 15. Brown, buckle spots on the green fruit, cherries and cherry in the zone of penetration after artificial inoculation suspension of bacteria P. s. pv. syringae (Ignjatov 2007)
Višnja Trešnja

276
Semenarstvo
Prisustvo cilija (sl. 16) – 90% fitopatogenih bakterija je pokretno, od-nosno poseduje cilije. Cilije su obično raspoređene na polovima ili po po-vršini čitavog tela bakterije. Razlikujemo monotrih raspored cilija (kada na jednom polu postoji jedna cilija (rod Xanthomonas)); lofotrih (kada na jednom polu postoji više cilija (rod Pseudomonas)) i amfitrih (po nekoliko cilija na oba pola (rod Pseudomonas)). Bez cilija je atrih, a sa cilijama po celoj površini bakterijske ćelije je peritrih raspored cilija (rod Erwinia). Za rodove Corynebacterium i Agrobacterium karakteristično je da mogu imati različite tipove flagelacije tako da se sreću peritrih ili atrih raspored cilija.
Biohemijsko-fiziološke odlike bakterija
Poznavanje biohemijske aktivnosti pojedinih vrsta bakterija je ta-kođe od velikog značaja u pogledu njihove identifikacije. Do nedavno ove osobine, zajedno sa morfološkim i odgajivačkim svojstvima, imale su i najveći značaj kada se radilo o determinaciji pojedinih vrsta. Sero-loška, molekularna i istraživanja bakteriofaga sve više se koriste u cilju identifikacije (Obradović et al., 2000, Obradović et al., 2004a, Todorović et al., 2006, Ignjatov et al., 2006, 2007b).
13
atrih
Corynebacterium monotrih
Agrobacterium Xanthomonas lofotrih
amfitrih peritrih
Pseudomonas ErwiniaAgrobacterium
Corynebacterium
Biohemijsko-fiziološke odlike bakterija
Poznavanje biohemijske aktivnosti pojedinih vrsta bakterija je takođe od velikog
značaja u pogledu njihove identifikacije. Do nedavno ove osobine, zajedno sa morfološkim i
odgajivačkim svojstvima, imale su i najveći značaj kada se radilo o determinaciji pojedinih
vrsta. Međutim, usled toliko izražene sličnosti pojedinih vrsta istog i nekih različitih rodova
bakterija, serološka, molekularna i istraživanja bakteriofaga sve više se koriste (Arsenijević,
1988). Rutinske laboratorijske analize, velikog broja uzoraka, koje se koriste u cilju brze i
pouzdane determinacije prouzrokovača biljnih bolesti nezamislive su bez molekularnih i
seroloških analiza i testova (Obradović i sar., 2000; Obradović i sar., 2004a; Todorović i sar.,
2006; Ignjatov i sar., 2006, 2007b).
O/F test (oksidativno-fermentativni)
Razlaganje ugljenikovih jedinjenja
Razlaganje skroba
Razlaganje želatina
Stvaranje sumpor-vodonika (H2S)
Stvaranje indola
Redukcija nitrata
Reakcije u mleku
Slika 16. Raspored cilija kod fitopatogenih bakterijaFigure 16. The position of cilia on phytopathogenic bacteria

277
Ispitivanje zdravstvenog stanja semena
Razlikujemo sledeće biohemijske fiziološke reakcije:– O/F test (oksidativno-fermentativni)– Razlaganje ugljenikovih jedinjenja– Razlaganje skroba– Razlaganje želatina– Stvaranje sumpor-vodonika (H2S)– Stvaranje indola– Redukcija nitrata– Reakcije u mleku
LOPAT testovi su dosta u upotrebi u identifikaciji roda Pseudomo-nas, koji sadrže sledećih pet reakcija (Tabela 1):
– Stvaranje levana (Levan production)– Aktivnost oksidaze (Oxidase actyvity)– Trulež krtola krompira (Potato rot)– Metabolizam arginina (Arginine dihydrlase activity)– Hipersenzitivna reakcija duvana (Tobacco hypersensitivity reaction)
Stvaranje levana. Ispituje se da li bakterija stvara levan. Levani su polisaharidi koje u znatnim količinama stvaraju neke saprofitne para-zitne bakterije, ako u podlozi ima saharoze. Stavaranje levana se prati na hranljivoj podlozi sa 5% saharoze. Zasejavanje podloge se vrši raz-mazom i ako se posle 2-3 dana obrazuju sitne kolonije – ne stvara levan, ako se obrazuju krupne, ispupčene, sluzave kolonije (5-6 mm)- stvara levan (sl. 17a).
Aktivnost oksidaze. Ispituje se da li bakterija poseduje enzim ok-sidazu. Platinastom petljom, plastičnom ezom ili sterilnim staklenim ili drvenim štapićem, nanese se pun zahvat ispitivane kolonije, stare 24 časa razvijene na kosoj podlozi i razmaže na prethodno pripremljeni, sterilnom vodom ovlaženi disk. U slučaju da bakterija poseduje enzim katalazu presejani deo kolonije postaje tamnoljubičast u vremenu od 30 do 40 sekundi, ukoliko ne poseduje, nema promene boje (sl. 17b).
Trulež krtola krompira. Ispituje se da li bakterija prouzrokuje trulež krompira. Za ispitivanje pektolitičke aktivnosti koriste se opra-ne, oljuštene i pomoću alkohola dezinfikovane krtole krompira, koje se poprečno iseku na manje kriške debljine 7-8 mm. Ovako pripremlje-ne kriške postave se, aseptičnim putem, u Petri posude. Preko površine

278
Semenarstvo
kriški se naliva sterilna voda do polovine debljine kriški. Bakterijskom petljom se zahvata deo kolonije sa kose hranljive podloge i obilato na-nosi u udubljenja pripremljena u središtu kriški. Petri kutije se drže na sobnoj temperaturi, a očitavanje se izvodi nakon 24 časa. Pojava raz-mekšavanja tkiva, tj. vlažne truleži, znak je aktivnosti pektolitičkih fer-menata (sl. 17c).
Metabolizam arginina. U epruvete se sipa po 5 ml podloge kojoj se dodaje L-arginin, a zasejavanje se vrši ubodom bakteriološkom petljom sa svakim ispitivanim izolatom. Jedna epruveta zalije se tečnim steril-nim parafinom (15 mm) da se postignu anaerobni uslovi sredine, dok se druga epruveta drži aerobno. Formiranje baza iz arginina, odnosno nje-govo razlaganje dejstvom stvorenog fermenta arginindehidrolaze pod oba uslova (aerobno i anaerobno) dokazuje se promenom boje indika-tora (phenol red), koji se u neutralnoj sredini boji u bledoružičastu boju, a kao znak pozitivne reakcije boji se u ružičasto-crvenu boju. Promena boje indikatora prati se u vremenu od 4 do 14 dana (sl. 17d).
Hipersenzitivna reakcija duvana. Svi patogeni var. P. s. daju HR osim P. s. pv. tabaci (metod opisan u Patogenost ispitivanih izolata) (sl. 17e).
Tabela 1. Grupe I-IV: Fitopatogene bakterije roda Pseudomonas; Grupa V: saprofitne bakterije roda Pseudomonas
Table 1. Group I-IV Phytopathogenic bacteria of genus Pseudomonas; Group V: sa-prophytic bacteria of genus Pseudomonas
Grupa L O P A T
Ia + - - - +
Ib - - - - +
II - - + - +
III - + - - +
Iva + + + + -
Ivb - + + + -
V - + - + -

279
Ispitivanje zdravstvenog stanja semena
Slika 17a. Levan pozitivne kolonije na NSA podlozi
Figure 17a. Levan positive colonies on NSA medium (Ignjatov 2007)
Slika 17d. Stvaranje arginindehidrolaze- pozitivna reakcija P. fluorescens (leva epruveta);
negativna reakcija (desna epruveta)Figure 17d. Creating arginindehidrolaze-
positive reaction to P. fluorescens (left tube); (right tube) negative reaction (Ignjatov 2007)
Slika 17e. Hipersenzitivna reakcija na listu duvana
Figure 17e. Hypersensitive reaction on tobbaco leaf (Ignjatov 2009)
Slika 17b. a) (+) stvaranje oksidaze- pozitivna reakcija P. fluorescens; b) (−)
negativna reakcija P. s. pv. syringaeFigure 17b. a) (+) the creation of oxidase-
positive reaction to P. fluorescens b) (-) negative reaction P. s. pv. syringae
(Ignjatov 2007)
Slika 17c. Trulež kriški krompira: negativna reakcija (P. syringae) i pozitivna kontrola (E. c. ssp. carotovora)
Figure 17c. Rot potato slices: negative reaction (P. syringae) and positive control (E. c. ssp. carotovora) (Ignjatov 2007)

280
Semenarstvo
Primena bakteriofaga
Bakteriofagi predstavljaju posebnu grupu virusa, čiji su domaćini bakterije. Odmah nakon otkrića, s početka prošlog veka, privukli su pažnju svojom sposobnošću da parazitiraju ćelije bakterije i tako uti-ču na njihovu populaciju u životnoj sredini. Bakteriofagi u zaštiti bilja imaju višestruku ulogu. Mogu da se koriste za determinaciju, dokaziva-nje patogenosti, ali i kao prirodni agensi u zaštiti bilja od patogena. U nedostatku efikasnih baktericida očigledno je da za sprečavanje pojave i širenje bakterioza bilja rešenje treba tražiti u integralnom pristupu, odnosno sintezi znanja o biologiji i epidemiologiji patogena kao i bak-tericidnom efektu pojedinih supstanci. Najnovija istraživanja ukazuju i na mogućnost efikasne primene nekih bioloških agenasa a među njima su i bakteriofagi (Balogh et al. 2003, Obradović et al., 2004b, 2008a, 2008b, Oradović and Ivanović 2007, Obradović 2009). Tretman fagima je deo standardnog programa integralne zaštite paradajza od bakterio-zne pegavosti na Floridi i južnim delovima SAD-a (Momol et al. 2002, Jones et al. 2006).
Biotest
Zbog neravnomerne distribucije virusa u prirodno zaraženim bilj-kama, korišćenje test i indikator biljaka, koje pojavom specifičnih simp-toma reaguju na mehaničke inokulacije, neophodno je da bi dijagnoza bila pouzdana. Mehaničko prenošenje fitopatogenih virusa na indikator ili test biljke podrazumeva prenošenje sokom obolelih biljaka, koji može biti neprečišćen (onakav kakav se dobija ekstrakcijom iz obolelih bilja-ka) ili prečišćen posebnim postupcima.
Pre mehaničkih inokulacija, odabrane indikator-biljke drže se u mraku 24 časa da bi se povećala osetljivost biljaka prema virusima. Ho-mogenizuje se odabrani biljni materijal u hladnom puferu (0,01M fos-fatnom puferu pH 7, u koji se neposredno pre inokulacija dodaje 1% Na2SO3 koristeći hladan avan i tučak. Abraziv, karborundum prah (400-600 meša), se dodaje prilikom homogenizacije ili se vrši zaprašivanje test biljaka (sl. 18). Inokulacija se obavlja prstom, blagim prevlačenjem preko listova test-biljaka od drške ka vrhu lista, pri čemu se stvaraju fine povrede kroz koje se virus unosi u ćelije inokulisanog tkiva (sl. 19) (Krstić& Bulajić 2007).

281
Ispitivanje zdravstvenog stanja semena
Kao test-biljke koriste se mlade biljke, intenzivnog porasta. Posle inokulacije, inokulisani listovi biljaka speru se slabim mlazom vode da bi se sprao abraziv. Po isteku inkubacije, nakon 5–10 dana, na inokulisa-nim listovima, ispoljavaju se karakteristični simptomi (sl. 20).
Postoji širok spektar biljaka koje se koriste kao test-biljke za identi-fikaciju određenih virusa. Tako je inokulacijom test biljaka dokazano da je izolat blagog šarenila parike infektivan prema Nicotiana tabacum var. samsun, White Burley, Datura metel, D. stramonium Chenopodium ama-ranticolor, C. foetidum i dr (Krstić et al., 1996). Identifikaciju virusa iz obolelih biljaka pasulja, pomoću test biljkama Lycopersicoon esculentum, Datura stamonium, Lupinus albus, Medicago sativa, Nicotiana glutinosa, Nicotiana tabacum var. samsun, Glycine max, Vicia faba, Zinnia elegans i Capsicum annuum vršili su Petrović et al., (2007a), dok su Mijatović et al., (2007) za determinaciju Tomato spotted wilt virus na paprici, koristili reakcije test biljke dragoljuba (Tropaeolum majus).
Slika 18. Zaprašivanje biljaka karborundum prahom
Figure 18. Dusting the plants with carborundum powder (Petrović 2007)
Slika 19. Inokulacija biljaka metodom kažiprsta
Figure 19. Inoculation of plants using finger (Petrović 2007)
Slika 20. Simpotomi virusa običnog mozaika pasulja
(Bean common mosaic virus) na biljci Zinnia elegans
Figure 20. Symptoms caused by the Bean common mosaic virus
on Zinnia elegans (Petrović 2007)

282
Semenarstvo
Novije metode za ispitivanje patogena
Serološke metode identifikacije
S obzirom na potrebe za brzom i pouzdanom identifikacijom biljnih patogena, serološke metode nalaze sve veću primenu u identifikaciji vi-rusa, bakterija i nekih gljiva. Serološka metoda se zasniva na antigenim svojstvima virusa da u životinjskom organizmu izazivaju reakciju stva-ranja specifičnih antitela. Antigeni su materije koje unete u organizam životinje stimulišu u njemu stvaranje odgovarajućih antitela. Antitela su globulini krvnog seruma nastali imunološkim odgovorom organizma pod dejstvom unetih antigena. Serum koji sadrži antitela naziva se imu-ni serum bez koga se serološki metod ne bi mogao primeniti (Lelliott and Stead 1987).
ELISA test (Enzyme Linked Immunosorbent Assay – ELISA). Se-rološka metoda koja se najviše primenjuje u detekciji biljnih patogena je ELISA test. Ova metoda je razvijena krajem sedamdesetih godina proš-log veka (Clark and Adams 1977). Ovo je jednostavna metoda, jeftina, ne zahteva skupocenu opremu za njeno izvođenje i podesna je za testi-ranje velikog broja uzoraka. Kod seroloških metoda problem može biti nedovoljna specifičnost antitela ciljanog patogena, posebno kod gljiva.
Najjednostavnija forma ove metode je PTA-ELISA (plate-trapped antigen), ali se u praksi više koristi DAS-ELISA (double antibody sandwich).
Antiserumi se dobijaju injektiranjem prečišćene suspenzije virusa (antigen) u telo eksperimentalnih životinja, koje proizvode specifičan protein (antitelo). Za svaki virus je neophodno imati antiserum. Za do-kazivanje virusa koriste se gotovi kitovi koji se čuvaju u frižideru na +4ºC i polistirenske mikrotitarske ploče sa 96 (8x12) udubljena. ELISA je osetljiva serološka test metoda kojom se određuje prisustvo antige-na na bazi enzimatske reakcije. Koriste se gotovi kompleti antitela za identifikaciju patogena različitih proizvođača (Bioreba AG, Switzer-land; Loewe Biochemica GmbH, Germany; Adgen Neogen Europe Ltd, Skoyland, UK). Standardni Kit komplet za DAS ELISA test sadrži set antiseruma koji se sastoji od IgG antitela, konjugovanog IgG antitela sa enzimom fosfatazom, pozitivne i negativne kontrole. Tokom prvog koraka inkubacije, površina mikrotitar mesta se oblaže sa antigenom

283
Ispitivanje zdravstvenog stanja semena
specifičnog antitela. Tokom drugog koraka inkubacije, antigen veže fik-sirano antitelo formirano od antitelo-antigen kompleksa. Tokom trećeg koraka inkubacije antitelo-antigen kompleks reaguje sa antitelo-konju-gatom formirajući dupli antitelo sendvič. Prisustvo specifičnog antige-na daje pozitivnu reakciju u alkalnoj fosfotazi sa 4 nitrofenilfosfatom. Vrednosti ekstinkcije ispitivanih uzoraka očitavaju se na automatskom ELISA čitaču na talasnoj dužini od 405 nm, a vrednosti dva puta veće od vrednosti ekstinkcije negativne kontrole set-a, smatraju se pozitivnim (sl. 21).
DAS-ELISA test najviše se primenjuje za detekciju virusnih bolesti kao i nekih bakterija. Tako su Krstić et al., (2008) koristili ovu meodu za detekciju i dijagnozu Tomato spotted wilt virus (TSWV) u zaraže-nim biljkama i vektorima. Takođe, korišćena je za identifikaciju viru-
Slika 21. a) Set antiseruma (IgG antitela, konjugovano IgG antitelo sa enzimom fosfatazom, pozitivna i negativna kontrola), b) Multiscan Ascent automatski Elisa čitač,
c) Ploče sa pozitivnim reakcijama.Figure 21. a) Set antiseruma (IgG antibody, IgG antibody conjugate with the enzyme phosphatase, positive and negative control), b) Multiscan Ascent automatically Elisa
reader, c) plates with positive reactions (Petrović 2007)
a)
b)
c)

284
Semenarstvo
sa iz obolelih biljaka pasulja pri čemu su korišćeni antiserumi za Bean common mosaic virus (BCMV), Bean common necrosis virus (BCMNV), Bean yellow mosaic virus (BYMV), Cucumbe mosaic virus (CMV) i Alfalfa mosaic virus (AMV) (Petrović et al., 2007a, 2008a, 2010b). Petrović et al., (2007b) su DAS-ELISA test koristili za identifikaciju virusa na paradajzu (Tomato mosaic virus (ToMV) i TSWV), dok su istu metodu koristili i Milošević et al., (2005a, 2006) u okviru projekta „Monitoring i postka-rantinski nadzor pošiljaka iz uvoza” kada je vršeno ispitivanje prisustva virusa i bakterija u biljnom tkivu krompira i drugih povrtarskih biljnih vrsta. Mijatović et al., (2002) vršili su identifikaciju i utvrđivanje raspro-stranjenosti Potato virus Y (PVY) na paprici u Srbiji na osnovu rezultata DAS ELISA testa. Identifikaciju sojeva bakterija P. s pv glycinea, izolova-nih iz komercijalnih sorti soje kod nas, ELISA testom opisuje Ignjatov et al., (2007c), dok identifikciju Beet necrotic yellow vein virus (BNYVV) and Beet yellows vrus (BYV) ovom metodom opisuje Milošević et al. (2005b).
Ekspres Kit. Test se zasniva na serološkom svojstvu bakterije da u prisustvu specifičnih antitela stvara talog (Weir 1978, Lelliott &Stead 1987). Postupak se sastoji u tome da se ispitivana bakterijska kolonija (stara 24 časa) pomoću drvene čačkalice prenese u pripremljenu kap test reagensa, koji je predhodno nanet na test pločice. Nakon homogeni-zacije kolonije i reagensa, ukoliko je reakcija pozitivna, dolazi do pojave
granuloznog taloga (aglutinacija) u centru kapi, nakon 60 sekundi. U slučaju negativne reakcije, kap test reagensa ostaje prozirna, bez taloga. Rezultati su vidljivi golim okom ili uz korišćenje ručne lupe. Danas su dostupni komercijalni kitovi koji se zasnivaju na sero-loškom svojstvu patogena da u prisustvu specifičnih antitela vrši
Slika 22. Reagensi Ekspres Kit-a (negativna kontrola, test reagens i pozitivna kontrola)Figure 22. Reagents Express Kit-A (negative control, test reagents and positive control) (Ignjatov, 2008)

285
Ispitivanje zdravstvenog stanja semena
aglutinaciju u veoma kratkom vremenskom periodu (sl. 22). Ispitivanja izolata poreklom sa soje iz nekoliko lokaliteta u Vojvodini, metodom aglutinacije (Ekspres Kit NEOGEN Europe Ltd., Scotland, UK), vršili su Ignjatov et al., (2006, 2008e).
Imunofluorescencija (IF) je često korišćena serološka metoda identifikacije bakterija. Ova se tehnika najčešće koristi u bakteriologiji od serološko-dijagnostičkih metoda. Laka je za izvođenje, a od instru-menata potreban je mikroskop sa UV lampom. Za ova ispitivanja koristi se komplet antiseruma, uključujući pozitivnu i negativnu kontrolu (sl. 23). Kao fluorescirajuća komponenta najčešće se koristi konjugovani fluorescin izotiocijanat (FITC), a pojava zelenih fluorescentih ćelija u vidnom polju fluorescentnog mikroskopa predstavlja pozitivnu reakci-ju. Postupak se izvodi prema protokolu proizvođača (npr. ADGEN NE-OGEN EUROPE Ltd., Scotland, UK).
Molekularna identifikacija patogena
Molekularne metode su zasnovane na poznavanju genoma fitopa-togenih gljiva, bakterija i virusa, a veoma su specifične, brze i osetljive. Ključna prednost ovih metoda je u tome što su nezavisne od spoljaš-njih uslova, starosti i fiziološkog stanja ciljanog patogena (Louws and
Slika 23. Komercijalni IF Set antiseruma bakterije P. s. pv. glycinea (levo) i IF test pločice (desno)
Figure 22. Commercial IF Set antiserums of bacteria P. s. pv. glycinea (left) and IF test slides (right) (Ignjatov 2008)

286
Semenarstvo
Cuppels 2001). Osim toga, moguće je detektovati patogene i u veoma niskim koncentracijama, kako u simptomatičnim, tako i u asimpto-matičnim uzorcima, što je posebno značajno u detekciji karantinskih patogena na semenu i sadnom materijalu, kada je izolacijom nemogu-će utvrditi njihovo prisustvo (Darrase et al. 1994; McManus and Jones 1995).
Lančana reakcija polimeraze je metoda koja je razvijena od strane Kary Mullis-a 1984 godine i jedno je od velikih otkrića u nauci (Mullis 1987). Pretstavlja najnapredniju tehniku detekcije gljiva, bakterija i vi-rusa.
Lančana reakcija polimeraze je metoda za veoma brzo sintetisanje kopija specifičnih DNK sekvenci (amplikona). Ovo je najvažnija i veo-ma osetljiva molekularna metoda koja se koristi u dijagnostici. Za njeno izvođenje je potrebna vrlo mala količina početne DNK. Teorijski, do-voljna je samo jedna kopija DNK. PCR reakcija podrazumeva pripremu uzoraka i master mix-a, izvođenje reakcije u PCR aparatu i analizu pro-izvoda reakcije.
Priprema DNK je početni korak u primeni ove metode. Za izolaciju DNK razvijeni su brojni protokoli, a postoje i komercijalni kitovi koji se mogu nabaviti na tržištu. Primenom ovih procedura izdvojena DNK je za-dovoljavajuće ili visoke čistoće u zavi-snosti od potreba reakcije, kao i same vrste biljnog materijala i patogena.
Za izvođenje PCR reakcije neop-hodan je master mix koji čine PCR pufer, MgCl2, dNTPs, prajmeri i DNK polimeraza. Taq DNA polimeraza je enzim koji sintetiše DNK i igra glavnu ulogu u PCR reakciji. Proces se odi-grava u PCR tubicama koje se postav-ljaju u PCR aparatu (sl. 24). Karakteri-stika PCR aparata je brza promena ra-zličitih temperatura na kojima se odi-gravaju pojedini koraci PCR reakcije.
Slika 24. PCR aparat Figure 24. PCR device

287
Ispitivanje zdravstvenog stanja semena
Prvi korak u PCR reakciji je razdvajanje lanaca DNK koje se postiže zagrevanjem na 94–95ºC. Temperatura se zatim smanjuje na 40-65ºC na kojoj je omogućeno vezivanje prajmera na svakom kraju regiona koji se umnožava. Prajmeri su kratke sekvence jednolančane DNK koje se specifično vežu za ciljanu DNK. Specifičnost PCR reakcije je upravo određena prajmerima. Za dizajniranje prajmera potrebno je poznavanje DNK sekvenci odgovarajućeg patogena i to je preduslov za razvijanje PCR protokola detekcije odgovarajućeg patogena.
Na kraju PCR reakcije umnoženi su milioni kopija sekvenci DNK. PCR proizvodi se analiziraju elektroforezom u agaroznom ili poliakri-lamidnom gelu uz bojenje etidijum-bromidom ili srebro-nitratom. Pri-sustvo trake odgovarajuće veličine u gelu je potvrda detekcije ciljanog patogena.
Pored lančane reakcije polimeraze postoje i druge metode koje se zasnivaju na nukleinskim kiselinama (RNK ili DNK), a koriste se za identifikaciju gljiva:
Nested PCR - sastoji se od dve uzastopne reakcije, pri čemu se u dru-goj reakciji koriste prajmeri koji umnožavaju region u okviru fragmenta DNK koji je umnožen u prvoj reakciji. Koristi se u slučajevima kada se direktnom PCR metodom ne dobijaju pouzdani rezultati, odnosno kada nije moguće dizajnirati visoko specifične prajmere. U tom slučaju se prvo sprovodi PCR reakcija sa prajmerima koji odsecaju fragment DNK odgovarajuće veličine koji je polazni materijal druge reakcije. Ove dve reakcije se mogu sprovoditi u istoj PCR tubici ukoliko su tempera-ture vezivanja prajmera različite. Ovo je veoma osetljiva tehnika koja se primenjuje u detekciji fitopatogenih gljiva i pseudogljiva (Bonants et al. 1997, Ma et al. 2003).
PCR-RFLP – za karakterizaciju prisutnog patogena moguće je posle izvedene PCR reakcije primeniti restrikcione enzime, odnosno RFLP (restriction fragment length polymorphism). Odgovarajući enzim će iseći DNK fragmente dobijene u PCR reakciji. Na osnovu RFLP analize donosi se sud o prisutnoj vrsti (ili varijetetu) patogena.
RAPD analiza (Random Amplification of Polymorphic DNA) – ono što čini ovu metodu popularnom i zbog čega se dosta koristi za poređenje DNK patogena koji nisu dovoljno proučeni je to što prajmeri koji se kori-ste u ovoj reakciji su proizvoljne sekvence. Za ovu metodu nije potrebno

288
Semenarstvo
poznavati DNK sekvencu ciljanog patogena jer će se prajmeri vezati neg-de duž lanca DNK. Poslednjih godina RAPD se koristi za karakterizaciju, praćenje i pravljenje filogenetskog stabla različitih biljnih vrsta i njihovih patogena. RAPD-PCR se koristi u analizi, karakterizaciji i identifikaciji fitopatogenih gljiva (Smith et al. 1996, Bieliková et al. 2002).
Multiplex PCR – ova metoda se primenjuje sa ciljem da se u jed-nom koraku detektuje nekoliko patogena. Ona podrazumeva korišćenje više setova prajmera u jednoj reakciji čime se skraćuje vreme detekcije i smanjuju troškovi izvođenja reakcije. Ova metoda je razvijena za detek-ciju nekoliko patogena pšenice: Septoria tritici, Stangospora nodorum, Puccinia striiformis i Puccinia recondita (Fraaije et al. 2001).
Real-time PCR – je veoma osetljiva metoda, precizna i brža tehnika od konvencionalne PCR metode. Koristi se za preciznu dijagnostiku i kvantifikaciju specifičnih biljnih patogena, kao i za praćenje infekcije (Shaad and Frederick 2002). Osnovna karakteristika ove metode je što se umnoženi PCR proizvodi (amplikoni) prate i mere u toku same re-akcije (TaqMan® proba, fluorescent resonance energy transfer (FRET) proba i molecular beacons).
DNA microarray – ovo je tehnika koja se može koristiti za detekciju neograničenog broja patogena u toku jedne reakcije. Prednost ove teh-nike je što kombinuje umnožavanje DNK sekvenci sa neograničenom sposobnošću sortiranja DNK niza. DNA microarray je kolekcija mi-kroskopskih DNK tačaka, koji obično predstavljaju pojedinačne gene, nanizanih na čvrstu podlogu kovalentno povezani za hemijski pode-sne matrice. Princip rada ove tehnologije je da se dati molekul mRNK specifično veže ili hibridizuje za DNK matricu iz koje potiče. Koristeći odgovarajući softver precizno se meri količina vezane mRNK i na taj način generiše profil ekspresije gena u ćeliji. Na ovaj način omogućeno je da se prati veći broj gena odjednom i utvrdi njihova ekspresija. U biljnoj patologiji ova tehnika je sa uspehom primenjena u identifikaciji nematoda, bakterija i pseudogljiva iz klase Oomicetes (Lévesque et al. 1998; Lievens et al. 2005).
Bereswil et al. (1994) razvili su molekularni metod identifikacije bakterija koje produkuju koronatin. Ovaj metod zasnovan je na lanča-nom umnožavanju fragmenata DNK odgovornih za sintezu fitotoksina koronatina i karakterističan je samo za sojeve koje stvaraju fitotoksin koronatin. Pored P. s. pv glycinea sposobnošću stvaranja fitotoksina

289
Ispitivanje zdravstvenog stanja semena
koronatina odlikuju se patogeni varijeteti: tomato, artropurea, maculi-cola i morsprunorum (Cuppels et al. 1990; Wiebe and Campbell 1993). Ovom metodom PCR-a obavljena je determinacija odabranih izolata bakterije Pseudomonas syringae pv. glicinea izolovanih sa najzastuplje-nijih sorti soje koje se gaje u agroekološkim uslovima Vojvodine (Ignja-tov et al., 2008f). Kao rezultat lančane reakcije polimeraze, korišćenjem prajmera 1 i 2, kao i posmatranjem produkata elektroforeze na agaro-znom gelu, detektovani su fragmenti nukleinske kiseline veličine 650bp, specifični za sojeve P. s. pv. glycinea koji stvaraju fitotoksin koronatin. Ignjatov et al., (2007b) su vršili identifikaciju odabranih izolata Pseudo-monas sysingae pv. tomato izolovanih sa paradajza. Milijašević i Obra-dović (2004) izvršili su karakterizaciju izolata Pseudomonas syringae pv. tomato izolovanih sa paradajza poreklom sa nekoliko lokaliteta u Srbiji. Primenom lančane reakcije polimeraze (PCR) u genom proučavanih sojeva detektovan je gen koji determiniše stvaranje fitotoksina korona-tina, što se smatra diferencijalnom karakteristikom pv. tomato. Prime-nom BIO-PCR metode, osetljivost PCR reakcije prilikom identifikacije fitopatogenih bakterija na semenu, kao i iz biljng tkiva može se uvećati i do sto puta (Louws and Cuppels 2001), dok se primenom Nested-PCR metode osetljivost u detekciji Erwinia amylovora povećana je hiljadu puta (McManus and Jones 1995). Najnovija i najosetljivija je Real-time PCR metoda, pomoću koje je moguće istovremeno kvantifikovati i ana-lizirati podukt tokom PCR umnožavanja. Prednost se sastoji u tome što se na ovaj način može eliminisati „Southern blot” koji se koristi za po-tvrdu identiteta umnoženog produkta (Bouzar et al. 1999) Identifikacija bakterija Rep-PCR metodom zasniva se na činjenici da svaki patogen, identifikovan do nivoa vrste ili patogenog varijeteta, sastoji od sojeva koji poseduju jednu ili više različitih REP-PCR molekularnih profila, što je korisno u identifikaciji bakterija koje izazivaju slične simptome, kao i za detekciju diverziteta u okviru vrste (Ivanović et al., 2004).
Molekularne metode detekcije, zasnovane na osobinama virusnog genoma, predstavljaju jako osetljive i pouzdane metode detekcije virusa. Posebno značajnu ulogu imaju u detekciji satelitnih RNK virusa, virusa koji se ne mogu prenositi mehaničkim putem, a čiji vektori nisu poznati, latentnih virusa, defektnih virusnih čestica, koji se ne mogu detektovati serološkim metodama, u identifikaciji sojeva virusa koji se na osnovu antigenih osobina teško razlikuju, kao i u detekciji virioda (Nikolaeva 1995, Gallitelli and Saldarelli 1996).

290
Semenarstvo
Najosetljivija metoda detekcije biljnih virusa koje imaju RNK je kombinacija reverzne transkripcije i lančane reakcije polimeraze (RT-PCR). Ova metoda PCR-a sastoji se od dve faze: pretvaranje RNK u DNK (reverzna transkripcija, početni korak aktivacije) i razdvajanje la-naca DNK i vezivanje prajmera za kraj regiona koji se umnožava.
Poređenjem različitih metoda detekcije virusa infektivne hloroze paradajza (Tomato infectious chlorosis virus), utvrđeno je da je RT-PCR i do sto puta osetljivija u odnosu na ELISA, EBIA i metodu dot-blot hibridizacije nukleinskih kiselina (Li et al. 1998). U slučaju biljnih vi-rusa, PCR tehnika se izvodi pomoću prečišćene RNK. Danas postoje komercijalni kitovi koji se mogu nabaviti na tržištu, a koji su pogodni za izolaciju RNK iz male količine početnog biljnog materijala. Ekstrakcija se uglavnom radi po proceduri koju preporučuje proizvođač.
Osetljivost detekcije biljnih virusa povećana je uvođenjem Real-ti-me RT-PCR metode (Roberts et al. 2000), a razvojem Multiplex PCR metode omogućeno je istovremeno detektovanje više virusa u slučajevi-ma mešane infekcije (Gallitelli and Saldarelli 1996).
U našoj zemlji, prva ispitivanja zasnovana na osobinama genoma biljnih patogena sprovedena su na biljnim virusima. Ispitivanje prisu-stva virus specifičnih dvolančanih RNK potvrdilo je virusnu prirodu oboljenja dunje (Paunović 1995). Molekularne metode primenjene su u detekciji i karakterizaciji virusa ratarskih i povrtarskih biljaka; Y virusa krompira (Potato Y virus) (Milošević et al., 1998; Dukić et al., 2004), virusa mozaika krastavca (Cucumber mosaic virus) (Krstić et al., 2002; Dukić et al., 2002, 2004), virusa običnog mozaika pasulja (Bean common mosaic virus) (Petrović et al., 2007c, 2008b).

291
Ispitivanje zdravstvenog stanja semena
LITERATURA
Arsenijević M (1997): Bakterioze biljaka. S print, Novi Sad.Balogh B, Jones J B, Momol M T, Olson S M, Obradović A, King P, Jack-
son L E (2003): Improved efficacy of newly formulated bacteri-ophages for management of bacterial spot on tomato. Plant dis., 87: 949-954.
Bereswill S, Bugert P, Volksch B, Ullrich M, Bender C, Geider K (1994): Identification and Relatedness of Coronatine-Producing Pseudo-monas syringae Pathovars by PCR Analysis and Sequence Deter-mination of the Amlification Products. Microbiology, p. 2924-2930, vol. 60, No. 8. American Society for Microbiology, USA.
Bieliková L, Landa Z, Osborne L, Čurn V (2002): Characterization and Identification of Entomopathogenic and Mycoparasitic Fungi using RAPD-PCR Technique. Plant Protection Science, 38: 1–12.
Bonants P, Hagenaar-deWeerdt M, Gent-Pelzer M, Lacourt I, Cooke D, Duncan J (1997): Detection and identification of Phytophthora fragariae Hickman by the polymerase chain reaction. Eur. J. Plant Pathol., 103, 4: 345-355.
Bouzar H, Jones JB, Stall, R E, Louws F J, Schneider M, Rademarker J L W, De Brujin F J, Jackson L E (1999): Multiphasic analyses of Xanthomonads causing bacterial spot disease on tomato and pepper in Caribbean and Central America: Evidence for common lineages within and between countries. Phytopathology, 89: 328-335.
Clark A X, Adams M J (1977): Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
Cuppels D A, Moore R A, Morris V L (1990): Construction and use nonrdioactive DNA probe for detection of Pseudomonas syringae pv. tomato on tomato plants. Applied and Envirometal Microbi-logy 56:1743-1749.
Darrasse A, Prious S, Kotoujansky A, Bertheau Y (1994): PCR and Re-striction Fragments Length Polymorphism of a pel Gene as a Tool

292
Semenarstvo
To Identify Erwinia carotovora in Relation to Potato Diseases. Appl. Environ. Microbiol., 60: 1437-1443.
Dukić N, Finetti-Sialer M M, Gallitelli D, Duduk B (2002): Molekularna identifikacija izolata virusa mozaika krastavca u Jugoslaviji-pod-grupe IA. Zbornik rezimea XII simpozijuma o zaštiti bilja i Save-tovanje o primeni pesticida, Zlatibor, 75.
Dukić N, Krstić B, Finetti-Sialer M M, Gallitelli D, Vico I, Berenji J (2004): Metoda „dot-blot“ hibridizacije nukleinskih kiselina u de-tekciji virusa paprike, paradajza i obične tikve. Zbornik rezimea V kongresa o zaštiti bilja, Zlatibor, 104.
Fraaije B A, Lovvel D, Coelho J M, Baldwin S, Hollomon D W (2001): PCR-based assays to assess wheat varietal resistence to blotch (Septoria tritici and Stangospora nodorum) and rust (Puccinia striiformis and Puccinia recondita) diseases. Eur. J. Plant Pathol., 107: 905-917.
Gallitelli D, Saldarelli P (1996): Molecilar identification of phytopatho-genic viruses. In: method an molecular biology. Speciesdiagno-sticts protocols-PCR and Other Nucleic Acid Method (Clapp.J.C., ed). Humana Press Inc.
Ignjatov M, Balaž J, Milošević M, Popović T (2006): Bakteriozna pega-vost soje. Arhiv za poljoprivredne nauke, 3, 67, 11-17.
Ignjatov M, Milošević M, Nikolić Z, Vujaković M, Petrović D (2007a): Characterization of Pseudomonas savanstanoi pv. glycinea isolates from Vojvodina. Phytopatologia Polonica, 45: 43-54. The Polish phytopathological Society, Poznan.
Ignjatov M, Milošević, Petrović D, Medić-Pap S (2007b): Molecular and serological identification of Pseudomonas syringae pv. tomato in commercial tomato seeds. Abstract book Second International Symposium on Tomato Diseases, Kusadasi, Turkey, 136.
Ignjatov M, Balaž J, Milošević M, Vidić M (2007c): Pseudomonas syrin-gae pv. glycinea-Ekonomski štetan patogen soje u Vojvodini. Biljni lekar, Poljoprivredni fakultet, Novi Sad, 6: 589-595..
Ignjatov M, Milošević M, Petrović D, Vujaković M, Nikolić Z (2008a): Sunflower seed borne diseases. 43th Croatian and 3rd Internati-onal Symposium on Agriculture. Book of abstracts, Opatija, 182.
Ignjatov M, Vujaković M, Nikolić Z, Petrović D, Milošević M, Vidić M (2008b): Susceptibility of soybean genotypes to Pseudomonas syringae pv. glycinea, the causal agent of bacterial blight. Book of

293
Ispitivanje zdravstvenog stanja semena
Abstracts of Inter.Conf.“Conventional and Molecular Breeding of Field and Vegetable Crops”, Novi Sad, Serbia, 87.
Ignjatov M, Vidić M, Milošević M, Balaž J, Petrović D, Nikolić Z, Vuja-ković M (2008c): Rase identification of Pseudomonas syringae pv. glycinea on commercial soybean varieties in Serbia. Zaštita bilja, 59 (263-266), 59-67.
Ignjatov M, Vujaković M, Nikolić Z, Petrović D, Milošević M (2008d): Diferencijalni testovi za patogene varijetete Pseudomonas syringae pv. syringae i Pseudomonas syringae pv. glycinea. Zbornik rezimea IX Savetovanja o zaštiti bilja, Zlatibor, 69.
Ignjatov M, Balaž J, Milošević M, Vidić M, Popović T (2008e): Studies of Plant Pathogenic Bacterium Causal Agent of Soybean Bacterial Spots (Pseudomonas syringae pv. glycinea (Coerper) Young et al.) 419-426. M B. Fatmi et al (eds), Pseudomonas syringae Pathovars and Related Pathogens. Springer Science+Business Media B. V. USA.
Ignjatov M, Vujaković M, Nikolić Z, Milošević M, Petrović D (2008f): Biochemical, serological and molecular identification of Pseudo-monas savanstanoi pv. glycinea bacterial blight of soybean in Voj-vodina province. The Second Joint PSU-UNS International Con-ference on BioScience: Food, Agriculture and Environment, Novi Sad, Serbia, 62.
Ignjatov M, Gašić K, Ivanović M, Šević M, Obradović A, Milošević M (2010): Karakterizacija sojeva Xanthomonas euvesicatoria, patoge-na paprike u Srbiji. Pesticidi i fitomedicina, 25 (2), 139-149.
Ignjatov M, Petrović D, Vujaković M, Taški-Ajduković K, Nikolić Z, Jo-vičić D (2011a): Detekcija Ustilago nuda (Jensen) Rostrup u seme-nu ozimog ječma. Ratar. Povrtar., 48: 179-182.
Ignjatov M, Petrović D, Nikolić Z, Jovičić D (2011b): Morfološka i mo-lekularna identifikacija izolata Fusarium spp. poreklom sa para-dajza. Zbornik radova XVI Savetovanja o biotehnologiji, Čačak, 447-452.
ISTA (2010): International Rules for Seed Testing. Annexe to Chapter 7, Seed Health Testing Methods. International Seed Testing Associa-tion, Switzerland.
Ivanović M, Koprivica M, Milijašević S, Dukić N, Duduk B (2004): Pri-mena molekularnih metoda u dijagnostici biljnih bolesti. Pesticidi i fitomedicina, 19, 223-231.

294
Semenarstvo
Jones J B, Iriarte F B, Obradović A, Balogh B, Momol M T, Jackson L E (2006): Management of bacterial spot on tomatoes with bacteri-ophages. Proceedings of the 1 st International Symposium on Bi-ological control of Bacterial Plant Diseases. Dermstadt, Germany, Mitt. Biol. Bundesanst. Land-Forstwirtsch. 408: 154-157.
Jovićević B, Milošević M (1990): Bolesti semena, Dnevnik, Novi Sad 1-290.Klement Z, Rudolph K, Sands D C (1990): Methods in Phytobacterio-
logy. Akadèmiai Kiadò. Budapest.Krstić B, Krnjaja V, Mijatović M, Tošić M (1996): Virus blagog šarenila
parike prisutan u Srbiji. Zbornik kratkih sadržaja, Prvi balkanski simpozijum povrće i krompir, 164.
Krstić B, Vico I, Dovas I C, Constantinos E, Katis I N, Berenji J. (2002): Molekularna detekcija i delimična karakterizacija jugoslovenskih izolata virusa mozaika krastavca. Zbornik rezimea XII simpozi-juma o zaštiti bilja i Savetovanje o primeni pesticida, Zlatibor, 74.
Krstić B i Bulajić A (2007): Karantinski virusi povrća i ukrasnih biljaka u zaštićenom prostoru. Univerzitet u Beogradu – Poljoprivredni fakultet i Ministarstvo poljoprivrede, vodoprivrede i šumarstva, Beograd.
Krstić B, Bulajić A, Đekić I, Berenji J (2008): Virus bronzavosti paradaj-za-jedan od najdestruktivnijih biljnih virusa. Pesticidi i fitomedi-cina, Beograd, 23, 153-166.
Lelliott R A, Stead D E (1987): Methods for the Diagnosis of Bacterial Diseases of Plants. British Society for Plant Pathology Blackwell Scientific Publications. London, UK.
Lévesque C A, Harlton C E Cock AWAM (1998): Identification of some Oomicetes by reverse dot blot hybridization. Phytopathology, 88: 213-222.
Li R H, Wisler G C, Liu H Y and Duffus J E (1998): Comparation of dia-gnostic techniques for detecting Tomato infectious chlorosis virus. Plant Dis., 82: 489-491.
Lievens B, Brouwer M, Vanachter A, Lévesque C A, Cammue B, Thomma B (2005): Quntitative assessment of phytopathogenic fungi in va-rious substrates using a DNA microarray. Environmental Micro-biology, 7(11): 1698-1710.
Limonard T (1966): A modified blotter test for seed health. Neth J Plant Pathol. 72: 319-321.

295
Ispitivanje zdravstvenog stanja semena
Louws F J, Cupples D A (2001): Molecular techniques. In: Laboratory Guide for identification of Plant Pathogenic Bacteria (Shaad N W, Jones J B, Chun W, eds.). APS Press, St. Paul, MN,USA.
Ma Z, Luom Y, Michailides T J (2003): Nested PCR assays for detection of Monilinia fructicola in stone fruit orchards and Botryosphaeria dothidea from pistacios in California. J. Phytopathol., 151: 312-322.
Mathur S B, Kongsdal O (2003): Common Laboratory Seed Health Te-sting Methods for Detecting Fungi. ISTA, Switzerland.
Maceljski M, Kišpatić J (1987): Zaštita povrća, Znanje, Zagreb.McManus P S, Jones A L (1995): Detection of Erwinia amylovora by
Nested PCR and PCR-Dot-Blot and Reverse-Blot Hybridizations. Phytopathology, 85: 618-623.
Mijatović M, Ivanović M, Obradović A, Zečević B (2002): Potato virus Y (PVY) on pepper in Serbia. Proc. 2nd Balkan Symposium on Ve-getable and Potatoes. Acta Horticulture, 579, ISHS 2002, 545-549.
Mijatović M, Zečević B, Cvikić D (2007): Tomato Spotted Wilt Virus on pepper in Serbia. Proceeding of the 13th Eucarpia Meeting on Ge-netics and Breeding of Capsicum and Eggplant. Warsaw, Poland, 121-127.
Milijašević S, Obradović A (2004): Characterization and race determi-nation of Pseudomonas syringae pv. tomato in Serbia. Abstract book of 1st International Symposium on Tomato Diseases. Orlan-do, Florid, USA
Milošević D, Dulić-Marković I, Čuljković B, Romac S (1998): Primena metode lančane reakcije polimeraze (PCR) za unapređenje di-jagnostike Y virusa (PVY). Zbornik rezimea IV jugoslovenskog kongresa o zaštiti bilja i međunarodni simpozijum o integralnoj zaštiti ratarskih biljaka, Vrnjačka Banja, 33.
Milošević M, Ignjatov M, Medić-Pap S, Petrović D (2005a): Monitoring i postkarantinski nadzor pošiljaka iz uvoza. VII Savetovanje o za-štiti bilja, Zbornik rezimea, 85. Društvo za zaštitu bilja Srbije. Be-ograd.
Milošević M, Ignjatov M, Medić-Pap S (2005b): Sugar beet seed health testing of imported consigments.Proceeding of Articles of XXX-th meeting for plant protection in Republic of Macedonia and I th Congress of Plant Protection »Enviromental Concern and Food Safety«, 257-260.

296
Semenarstvo
Milošević M, Ignjatov M, Medić-Pap S, Petrović D (2006): Ispitivanje virusa krompira. Zbornik rezimea III Simpozijuma o zaštiti bilja u Bosni i Hercegovini, Neum, BIH, 10.
Miloševic M, Ignjatov M, Medic-Pap S, Petrovic D (2007a): Lyophiliza-tion as a method for pathogens long term preservation. Zbornik radova Matice Srpske za prirodne nauke 113: 203-210.
Milošević M, Ignjatov M, Medić-Pap S, Petrovic D (2007b): Maize seed-borne diseases. Plant Protection, Society for Plant Protection of Republic of Macedonia, Skopje, vol.XVIII, No. 18, 22-24.,
Milošević M, Ignjatov M, Vujaković M, Petrović D, Nikolić Z, Dokić V (2008): Svojstva gljiva roda Fusarium sp., i mogućnost dugoroč-nog čuvanja metodom liofilizacije. Arhiv za poljoprivredne nau-ke, 69, 245, 89-95.
Momol M T, Jones J B, Otson S, Obradović A, Balogh B, King P (2002): Integrated management of bacterial Spot on Tomato in Florida. Fact Sheet PP110, Florida Cooperative Extension Service, Institu-te of Food and Agricultural Sciences, University of Florida. EDIS Web site http://edis.ifas.ufl.edu.
Mullis K (1987): Process for amplifying nucleic acid sequences. United States Patent No. 4, 683, 202.
Nikolaeva O V (1995): Nucleic acid hybridization for the detection and localization of plant viruses. In: Molecular methods in plant pat-hology (Singh, R.P. and Singh U.S., eds.). CRC Press. Inc., FL, ISA.
Obradović A, Arsenijević M, Mavridis A, Rudolph K (2000): Patogene i biohemijsko fiziološke karakteristike sojeva Xanthomonas campe-stris pv. vesicatoria patogena paprike u Srbiji. Zaštita bilja, vol. 51 (1-2) No 231-232: 157-175.
Obradović A, Mavridis A, Rudolph K, Janse J D, Arsenijević M, Jo-nes J B, Minsavage G V, Wang J F (2004a): Characterization and PCR-based typing of Xanthomonas campestris pv. vesicatoria from peppers and tomatoes in Serbia. Eur. J. Plant Pathol.,110: 285–292.
Obradović A, Jones J B, Momol M T, Balogh B, Olson S M (2004b): Management of Tomato Bacterial Spot in the Field by Foliar Applications of Bacteriophages and SAR Inducers. Plant Dis., 88: 736-740.
Obradović A, Ivanović M (2007): O primeni antibiotika u zaštiti bilja. Biljni lekar, 1: 52-59.

297
Ispitivanje zdravstvenog stanja semena
Obradović A, Gašić K, Stepanović M (2008a): Bakteriofagi u zaštiti bilja. Biljni lekar, 36 (1), 36-44.
Obradović A, Jones J B, Balogh B, Momol M T (2008b): Integrated management of tomato bacterial spot in: Ciancio A, Mukerji K G (eds.), Integrated Management of Diseases Caused by Fungi, Phytoplasma and Bacteria, 211-223. Springer Science+Business Media B. V. USA.
Obradović A (2009): Bakteriofagi kao baktericidi u zaštiti bilja. Pesticidi i fitomedicina, 24: 9-17.
Paunović S (1995): Double-stranded RNA associated with fruit defor-mation of quince. Acta Hortic., 386: 45-50.
Petrović D, Bagi F, Milošević M, Vasić M (2007a): Identifikacija virusa pasulja na području Vojvodine. Zbornik rezimea XIII Simpoziju-ma sa savetovanjem o zaštiti bilja, Zlatibor, 92-93.
Petrović D, Milošević M, Ignjatov M, Medić-Pap S. (2007b): Detection of viruses transmitted by tomato using serological methods. Ab-stract book Second International Symposium on Tomato Disea-ses, Kusadasi, Turkey, 136.
Petrović D, Bagi F, Milošević M, Vasić M, Ignjatov M, Vujaković M, Niko-lić Z (2007c): Determinacija virusa običnog mozaika pasulja u Voj-vodini. Zaštita bilja. Vol. 58, No 259-262, 15-23, (prihvaćeno 2009).
Petrović D, Bagi F, Milošević M, Vasić M (2008a): Identifikacija najzna-čajnijih virusa pasulja u Vojvodini. Biljni lekar, br. 1, 30-36.
Petrović D, Bagi F, Milošević M, Vasić M, Vujaković M, Ignjatov M, Ni-kolić Z (2008b): Determination of Bean common mosaic virus found in Vojvodina region. The Second Joint PSU-UNS Interna-tional Conference on BioScience: Food, Agriculture and Envi-ronment, Novi Sad, Serbia, 63.
Petrović D, Ignjatov M, Vujaković M, Taški-Ajduković K, Nikolić Z, Mi-lošević M (2009): Pojava gljive Sclerotinia sclerotiorum (Lib.) de Bary na semenu soje u periodu od 2005. do 2007. godine. Zrornik radova, 46, 355-360.
Petrović D, Ignjatov M, Vujaković M, Taški-Ajduković K, Nikolić Z, Mi-lošević M, Jovičić D (2010a): Mikopopulacija semena kukuruza (2006-2008). Ratar. Povrtar, 47: 561-566.
Petrović D, Ignjatov M, Nikolić Z, Vujaković M, Vasić M, Milošević M, Taški-Ajduković K (2010b): Occurrence and distribution of

298
Semenarstvo
viruses infecting bean in Serbia. Arch. Biol. Sci., 62 (3), 595-601.
Petrović D, Ignjatov M, Vujaković M, Nikolić Z, Taški-Ajduković K, Jo-vičić D (2011): Izolacija i detekcija gljiva Alternaria dauci (Kühn) Groves et Skolk i Alternaria radicina Meier, Drechsler et Eddy iz semena mrkve. Ratar. Povrtar, 48: 173-178.
Pravilnik o zdravstvenom pregledu semena, rasada i sadnog materijala. Službeni glasnik Republike Srbije, broj 119, 2007.
Prosen D, Hatziloukas E, Schaad N W, Panopoulos N J (1993): Specific detection of Pseudomonas syringae pv. phaseolicola DNA in bean seed by polymerase chain reaction-based amplification of a pha-seolotoxin gene region. Phytopathology, 83: 965–970.
Rasmussen O F, Wulff B S (1991): Detection of Pseudomonas syringae pv. pisi using PCR. In Proceedings 4th International Working Group on Pseudomonas syringae Pathovars. Kluwer Academic Publishers, Dordrecht, The Netherlands, 369–376.
Roberts C A, Dietzgen R G, Heelan L A, Maclean D (2000): Real-time RT-PCR fluorescent detection of tomato spotted wilt virus. J. Vi-rol. Methods, 88: 1-8.
Schaad N, Jones J B, Chun W (2001): Laboratory guide for identification of plant pathogenic bacteria. APS Press, St. Paul, Minnesota, USA.
Schaad N, Frederick R (2002): Real-time PCR and its application for ra-pid plant disease diagnostics. Can. J. of Plant Pathol., 24: 250-258.
Smith W, Viljoen C D, Wingfield B D, Wingfield M, Calitz F J (1996): A new canker disease of apple, pear, and plum rootstocks caused by Diaporrhe ambigua in South Africa. Plant Dis., 80: 1331-1335.
Todorović B, Balaž J, Milijašević S, Duduk B, Rekanović E (2006) Iden-tifikacija Xanthomonas campestris pv. phaseoli (Smith 1897) Dye 1978 b, klasičnim bakteriološkim, serološkim i molekularnim metodama. Pesticidi i fitomedicina, vol. 21, br. 1, 55-63
Weir D M (1978): Handbook of Experimental Immunology. Blackwell Scientific Publications. Oxford.
WiebeW L, Campbell R N (1993): Caracterization of Pseudomonas syringae pv. maulicola and comparison with P.s. pv. tomato. Plant Dis. 77: 414-419.

Ksenija Taški-Ajduković, Nevena Nagl
Primena genetskih markera u identifikaciji i određivanjugenetske čistoće sorti i hibrida

300
Semenarstvo

301
Primena genetskih markera u identifikaciji
UVOD
Jedna od važnih komponenti kvaliteta semena je njegov genetski identitet i uniformnost odnosno sortna čistoća. Genetska čistoća stra-nooplodnih biljnih vrsta, osobito ako se proizvodnja zasniva na kori-šćenju hibrida od posebnog je značaja. Uzroci nesortnih primesa mogu poticati od semena nastalog usled nekontrolisane samooplodnje, stra-nooplodnje ili usled mešanja semena različitih sorti/hibrida.
Identitet odnosno genetska čistoća sorti i hibrida se može kontroli-sati putem biotesta u polju, pri čemu se na osnovu fenotipa vrši kontrola semenskog useva. Dugotrajnost kontrole semena na osnovu fenotipskih karakteristika biljaka zahtevali su iznalaženje bržih metoda kontrole ove osobine kvaliteta semena. Na osnovu novih saznanja biologije uvedene su laboratorijske tehnike za identifikaciju i kontrolu genetske uniformnosti sorti i hibrida. Laboratorijska kontrola se zasniva na detekciji polimorfiz-ma proteina ili DNK. Prednost ovih metoda kontrole je u tome što se na ovaj način seme može ispitati pre naredne setve, u toku ili posle dorade, a moguća je kontrola semenskog useva pre žetve pa čak i pre sazrevanja.
Kako su tehnike zasnovane na DNK molekularnim markerima rela-tivno nove i znatno skuplje još uvek su u širokoj upotrebi elektroforetske metode kontrole na osnovu rezervnih proteina i funkcionalnih protei-na/izoenzima.
Sortne primese
Genetska čistoća semena je od različitog značaja za pojedine biljne vrste. Po pravilu za samooplodne biljne vrste i vrste gde se proizvodnja zasniva na sorti, ova čistoća je od manjeg ekonomskog značaja. Među-tim, ukoliko se primese značajno razlikuju po kvalitetu, vremenu sazre-vanja i drugim važnim agronomskim svojstvima od osnovne sorte onda je njihovo prisustvo veoma značajno (Gerić, 1986).
Genetska čistoća stranooplodnih biljnih vrsta, osobito ako se pro-izvodnja zasniva na korišćenju hibrida od posebnog je značaja. Uzroci nesortih primesa mogu biti različiti:

302
a) Primese mogu poticati od „nečistih” roditeljskih linija, pri čemu najveći ekonomski značaj imaju primese u vidu semena nastalog ne-kontrolisanom samooplodnjom. Prinos samooplodnih biljaka je znatno manji od prinosa hibridnih biljaka, pa veći udeo ovih primesa u hibrid-nom semenu značajno smanjuje prinos hibridnog useva. Zbog toga je, na primer u proizvodnji hibridnog semena kukuruza najvažnije blago-vremeno i potpuno ukloniti metlice sa fertilne linije majke.
b) Moguće su i primese poreklom iz nekontrolisane stranooplodnje (out-cross). Zbog toga je u proizvodnji hibridnog semena neophodno obezbediti prostornu izolaciju useva.
c) Pri berbi i preradi semena može doći do mešanja semena različi-tih sorti i hibrida što je veoma značajno ako se oni razlikuju prema grupi zrenja i drugim važnim agronomskim svojstvima (Gerić, 1986).
U praksi je neophodno identifikovati sorte, linije i hibride i odrediti njihovu uniformnost. Ova testiranja se zasnivaju na morfološkim i mo-lekularnim markerima koji mogu biti biohemijski i DNK-markeri. Mar-ker se definiše kao fizička lokacija segmenta DNK na hromozomu, koja se može identifikovati (gen, restrikciono mesto) i čije se nasleđe može pratiti. Prema Tanksley (1983) genetički markeri su tačno određene lo-kacije na hromozomu, koje služe kao obeleživači kod analize genoma.
Morfološki markeri
Identitet i genetska čistoća semenskog materijala se mogu testira-ti u vegetacionom ogledu (sl. 1), praćenjem velikog broja morfoloških karakteristika. U praksi ovaj pristup je vrlo uspešan ali je dugotrajan i skup proces, koji zahteva velike površine zemlje i kvalifikovano osoblje koje je često u situaciji da donese subjektivne ocene. Za ovo testiranje je potrebno gajiti biljke u toku cele vegetacije, pa se proizvedeno seme može koristiti tek naredne godine. Uspešnost ove metode zavisi od toga koliko se sorte, linije i hibridi fenotipski razlikuju. Pojedini genotipovi su veoma slični tako da je teško vizuelno odrediti njihov identitet. Kako je veliki broj morfoloških karakteristika rezultat interakcije genotipa i sredine u kojoj se biljke nalaze, fenotipska ocena nije uvek pouzdana.
Zbog gore navedenih nedostataka morfološke analize uvedene su metode analize biohemijskim markerima, koje su omogućile direktnu kontrolu semena.
Semenarstvo

303
Primena genetskih markera u identifikaciji
Biohemijski markeri
Na osnovu hemotaksonomista razlikuju se dve grupe biomolekula koje se mogu koristiti u klasifikaciji organizama: epismatic ili sekun-darni metaboliti (pigmenti, masne kiseline i dr.) i semantides ili sense-carring molekuli (proteini, nukleinske kiseline). Sekundarni metaboliti se mogu koristiti u svega nekoliko slučajeva dok se proteini i nukleinske kiseline koriste znatno češće.
Analiza sekundarnih metabolita
Postoje mnogobrojni testovi za analizu sekundarnih metabolita u semenu i vegetativnom delu biljke, od jednostavnih kolor testova do kompleksnih hromatografskih analiza antocijana, flavonoida i dr.
Najpoznatiji kolor test je Fenolni test, koji se koristi za identifikaciju na osnovu diferencijalne oksidacije fenola što dovodi do različitog in-tenziteta bojenja semena, od bledo do veoma tamnomrke (ISTA pravila, 2009). Ovaj metod se najčešće koristi za analizu semena pšenice a može se koristiti i za pirinač i Poa pratensis (Cooke, 1995b). Kolor test se ko-risti i za testiranje semena boba na osnovu detekcije tanina pomoću va-nilin reagensa kao i za detekciju tanina Lugolovim reagensom u semenu lupine (ISTA pravila, 2009).
Slika 1. Biotest šećerne repe Figure 1. Field trial of sugar beet (Taški-Ajduković, 2010 )

304
Semenarstvo
Za određivanje identiteta i genetske uniformnosti sorti i hibrida mogu se koristiti i hromatografske metode. Gasno-tečna hromatografija se koristi za razdvajanje masnih kiselina iz semena uljane repice. White and Law (1991) su pokazali da se masne kiseline mogu koristiti za ra-zlikovanje genotipova uljane repice. Pomoću ove vrste hromatografije se mogu analizirati i glikozinolati kod vrsta iz roda Brassica (Morgan, 1989). Visokopritisna tečna hromatografija (HPLC) se može koristiti za identifikaciju hortikulturnih vrsta separacijom antocijanina i flavonoida (Morgan, 1989).
Iako se sekundarni metaboliti mogu koristiti u identifikaciji i odre-đivanju genetske čistoće sorti i hibrida, proteinski, i DNK markeri su, bez sumnje, mnogo značajniji.
Analiza proteina
Proteini se nalaze pod direktnom kontrolom gena te se dugi niz go-dina koriste u određivanju genetskog identiteta određene faze ili gene-racije u procesu oplemenjivanja i utvrđivanju uniformnosti semenskog materijala (Weir, 1990). U identifikaciji varijeteta na osnovu proteina se koriste dva osnovna pristupa (Cooke, 1988):
Slika 2. Fenolni test: Uniformna bojena reakcija kod genetički čiste sorte pšenice (A) i različite bojene reakcije kod mešavine uzorka
Figure 2. Phenol test: Uniform color reaction in a genetically pure wheat variety (A) and a mixture of color reaction in a sample containing more than one variety (B)
(Oregon State University Seed Laboratory) (Taški-Ajduković, 2007)
A B

305
Primena genetskih markera u identifikaciji
a) Direktni (multi-lokusni) pristup se koristi za analizu proteina koji su polimorfni i kodirani multigenskim lokusima, a produkt jednog lokusa može dati nekoliko elektroforetski razdvojivih tra-ka. Kao primer se mogu uzeti rezervni proteini.
b) Indirektni (single-lokusni) pristup se koristi za ispitivanje protei-na koji su polimorfni ali su produkt jednog genskog lokusa (npr. izoenzimi).
Koji će pristup biti korišćen zavisi od načina ukrštanja i genetske strukture ispitivane vrste. Prvi pristup se može koristi za samooplodne biljne vrste (uključujući i vrste koje se vegetativno umnožavaju), dok se drugi uglavnom koristi kod stranooplodnih biljnih vrsta.
Da bi se proteini mogli koristiti kao markeri u identifikaciji i odre-đivanju genetske ćistoće partije semena oni moraju odražavati morfo-loške ili hemijske razlike koje su od interesa kako za uzgajivača tako i za industrijskog korisnika. Tako na primer kod kukuruza, korisnici neće biti zadovoljni semenom koje je genetički čisto prema molekularnim markerima a u polju se razlikuju po visini ili boji zrna. Kako se geni koji kodiraju izoenzime i rezervne proteine i oni koji određuju morfološke osobine najčešće nalaze na različitim, nevezanim lokusima, genetski či-sta partija prema biohemijskim markerima ne mora biti i genetski čista i prema morfološkim pokazateljima. Uprkos ovom riziku evaluacija ge-netske čistoće na osnovu proteinskih markera se koristi preko 20 godina jer, bez sumnje, imajući u vidu preciznost, brzinu analize i cenu ovaj metod omogućuje visok nivo merenja stvarne uniformnosti partije se-mena.
Rezervni proteini
Rezervni proteini semena su specifični za pojedine biljne vrste, vrlo su polimorfni, i zbog toga pružaju široke mogućnosti u identifikaciji i određivanju genetske čistoće sorti i hibrida. Oni su na osnovu osobine rastvorljivosti, podeljeni na sledeće grupe (Osborne, 1924):
– rastvorljivi u vodi (albumini, uključujući većinu enzima),– rastvorljivi u rastvorima soli (globulini),– rastvorljivi u alkoholu (prolamini),– rastvorljivi u kiselinama ili alkalijama (glutelini).

306
Semenarstvo
Rezervni proteini semena su kodirani velikim brojem multigenet-skih lokusa, i ekspresija svakog lokusa obuhvata grupu elektroforetski razdvojivih komponenti proteina. Na ovaj način se može dobiti elek-troforegram koji predstavlja identitet datog varijeteta. Na osnovu ra-zličitosti komponenti prisutnih u elektroforegramu može se utvrditi identitet i genetska čistoća semenskog materijala. Rezervni proteini se češće koriste za identifikaciju i određivanje uniformnosti samooplodnih biljnih vrsta: za pšenicu glijadini, za ječam hordein, za ovas avenin. Kod suncokreta je prisutan značajan nivo polimorfizma glavnog rezervnog proteina (heliantin) te se na osnovu njega mogu utvrditi inbred linije i hibridi suncokreta. Rezervni protein kukuruza zein takođe je veoma polimorfan i može se koristiti za kontrolu linija i hibrida kukuruza.
Izoenzimi
Izoenzimi predstavljaju različite molekularne forme jednog enzima koje su kodirane od strane različitih genskih lokusa, imaju sličnu supstrat-nu specifičnost, a svoju funkciju katalizatora obavljaju u različitim delo-vima ćelije. Izoenzimi se međusobno razlikuju u strukturi i konfiguraciji molekula što uslovljava razlike njihovih fizičko-hemijskih osobina.
Prema strukturi izoenzimi su podeljeni na monomere (jedan poli-peptidni lanac) i polimere (dva ili više peptidnih lanaca), pa mogu biti dimeri, trimeri, tetrameri itd. Polimeri enzima, koji nastaju kao proi-zvod jednog od alela nekog lokusa i koji su građeni od istih subjedinica su homomeri. Heteromeri su izoenzimi koji nastaju kao rezultat inte-rakcije dva različita alela i sastoje se od različitih subjedinica.
U zavisnosti od broja različitih alelnih formi, lokusi koji kontrolišu njihovu sintezu mogu biti monomorfni ili polimorfni. Polimorfizam je pojava održavanja dva ili više alela u istoj populaciji u frekvenciji od 0,01 do 0,99. Genski lokusi čiji se aleli u populacijama javljaju sa učestalošću od 0,99 su monomorfni, a za lokus se kaže da je polimorfan ako se frekven-cija njegovog najčešćeg alela u populaciji kreće od 0,01 do 0,99. Izoenzimi predstavljaju direktne proizvode gena, i kao takvi se tokom poslednje tri decenije intenzivno koriste kao gen markeri za ocenu stepena polimor-fnosti prirodnih populacija, genetičkih razlika unutar i između vrsta, za ispitivanje efekta gena, za konstruisanje hromozomskih mapa, otkrivanje i merenje dejstva selekcije, kao i za određivanje identiteta i čistoće semen-skog materijala (Zlokolica i Taški-Ajduković, 2004).

307
Primena genetskih markera u identifikaciji
Izbor pojedinih izoenzima kao markera je veoma važan, i zavisi od ispitivane biljne vrste, njene ploidnosti i načina oplodnje. Osnovni pre-duslov je polimorfnost markera, njegova alelna varijabilnost, kao i he-terozigotnost lokusa u rezultatima ukrštanja (Stuber, 1992). Do sada je korišćen veliki broj enzimskih sistema među kojima se najčešće koriste: ACP (Acid Phosphatase), ADH (alcohol dehydrogenase), CAT (catala-se), EST (esterase), β-GLU (β-glucosidase), IDH (isocitrate dehydroge-nase), MDH (malate dehydrogenase), PX (peroxidase), PGM (phosp-hoglucomutase), PHI (phosphohexoseisomerase), ACO (aconitase), DIA (diaphorase) (tanksley i orton, 1983).
U cilju utvrđivanja identiteta i genetske uniformnosti sorti i hibrida proteini se mogu analizirati hromatografskim metodama ali se u praksi znatno češće koristi elektroforetsko razdvajanje proteina.
Elektroforeza
Elektroforeza predstavlja proces kretanja koloidnih čestica makro-molekula u električnom polju na određenoj podlozi. Kao podloga se mogu koristiti skrobni gel, agar, poliakrilamid, celulozni acetat i dr. Ove podloge imaju različitu veličinu i oblik pora te deluju kao molekulsko sito koje poboljšava razdvajanje. Molekuli koji su mali u poređenju sa veličinom pora gela olakšano se kreću kroz gel, dok su molekuli koji su mnogo veći od pora gela gotovo inertni. Molekuli srednje veličine se kreću kroz gel različitom brzinom. Elektroforetski se mogu razdvajati naelektrisani makromolekuli poput proteina, DNK i RNK.
Za elektroforetsko ispitivanje neophodno je obezbediti odgova-rajuću disosovanost i pokretljivost molekula. Pokretljivost molekula u toku elektroforeze zavisi od jačine električnog polja, naelektrisanja, mase i oblika molekula koji migrira, te viskoznosti podloge na kojoj se vrši elektroforeza. Postoji rezličita tehnike elektroforeze od kojih će biti opisane samo one koje se najčešće koriste u identifikaciji i određivanju genetske čistoće sorti i hibrida.
Metoda horizontalne elektroforeze na skrobnom gelu
Elektroforeza pri kojoj se gel sa uzorcima nalazi u horizontalnom položaju naziva se horizontalna elektroforeza. Na ovaj način se vrši iden-tifikacija i određivanje genetske uniformnosti stranooplodnih biljnih

308
Semenarstvo
vrsta na osnovu alelnih varijanti izoenzima. Kao podloga se koristi skrobni gel jer on dozvoljava horizontalno presecanje na tanke ploče što omogućava višestruko bojenje. Skrob je podvrgnut procesu hidrolize i liofilizacije. On se nakon kuvanja sa odgovarajućim puferskim siste-mom želatinizuje i postavlja u kalupe (sl. 3).
Kao uzorak za analizu se najčešće koristi koleoptil 5 dana starih po-nika. Oni se maceriraju i u njih potapaju filter papirići (Whatman #3). Papirići se nakon uklanjanja viška ekstrakta nanose na prorez u gelu. Kalup sa gelom se postavlja u kade sa elektrodama. Proces elektroforeze se izvodi na temperaturi 6–10°C, a njegova dužina zavisi od enzima i puferskog sistema na kome se vrši njihovo razdvajanje. Nakon elektro-foreze gel se preseca po dužini na nekoliko delova i boji u zavisnosti od enzima koji se želi detektovati.
Očitavanje izoenzimskih profila (sl. 4) vrši se prema standardima ili verifikovanom protokolu kod kukuruza (Stuber et. al., 1988) ili poređe-njem sa roditeljskim komponentama ili standardom kod biljnih kultura kod kojih nije urađeno mapiranje.
Slika 3. Horizontalna skrobna gel elektroforezaFigure 3. Horizontal starch gel electrophoresis (Taški-Ajduković, 2010 )

309
Primena genetskih markera u identifikaciji
Izoenzimska analiza je korišćena za identifikaciju velikog broja bilj-nih vrsta. Prva upotreba izoenzima kao markera za genetsku čistoću je bila na kukuruzu. Obiman rad Stuber i saradnika omogućio je da ana-liza izoenzima pomoću elektroforeze na skrobnom gelu postane stan-dardna tehnika za identifikaciju i određivanje genetske čistoće kukuru-za (sl. 5). Tokom petnaestogodišnjih istraživanja oni su ispitali hiljade eksperimentalnih ukrštanja i roditelja, determinišući genetsku osnovu kontrole različitih izoenzima. Za više od 30 enzima i oko 70 lokusa na-đena je hromozomska lokacija i genetska kontrola (Goodman i Stuber, 1983), što daje velike mogućnosti u njihovoj primeni u genetskom tu-mačenju ove biljne vrste.
Izoenzimi se dugi niz godina koriste u različitim istraživanjima u našoj zemlji, a najviše u Institutu za ratarstvo i povrtarstvo, Novi Sad. Najčešće su upotrebljavani u procenjivanju divergentnosti i varijabilnosti
Slika 4. Izoenzimska analiza ge-netske čistoće linije suncokreta
Figure 4. Genetic purity of sun-flower inbred line by isozymes
(Taški-Ajduković, 2009 )
Slika 5. Izoenzimska analiza ge-netske čistoće hibrida kukuruza
Figure 5. Genetic purity of maize hybrid by isozymes
(Taški-Ajduković, 2009 )

310
Semenarstvo
selekcijonog materijala i identifikaciji linija kukuruza u cilju ocenjivanja njihove kombinacione sposobnosti (Gerić et. al., 1989; Zlokolica i Milo-sević, 2001). Polimorfizam izoenzima se pokazao pogodan u proceni ge-netičke uniformnosti linija i hibrida suncokreta (Zlokolica et. al., 1996b; Taški-Ajduković i sar, 2010). Izoenzimi su korišćeni i u identifikaciji i karakterizaciji povrtarskih biljnih vrsta kao što su: kupus, krastavac, kar-fiol, mrkva, paradajz i paprika (Zlokolica et. al., 1996a), analizi haploida šećene repe dobijenih ginogenezom i dihaploida dobijenih dejstvom kol-hicina (Mezei et. al., 1996, 2002; Nagl et. al., 2004; Zlokolica et. al., 1994) kao i na biljnom materijalu dobijenom in vitro (Zlokolica et. al., 1996c). Izoenzimi se takođe već dugi niz godina koriste za rutinsku kontrolu ge-netske čistoće kukuruza i suncokreta kao i ostalog poljoprivrednog bilja (Zlokolica i Taški-Ajduković, 2004; Nikolić et. al., 2007).
Elektroforeza na poliakrilamidnom gelu (PAGE)
Najjednostavnija i najstarija tehnika PAGE-a se izvodi u cevčicama sa polimerizovanim gelom na čiji se vrh nanosi određena količina uzor-ka, a zatim se postavlja vertikalno između gornjeg i donjeg rezervoara s puferom. Koristi se pufer čija je pH vrednost ista u oba rezervoara, tako da su svi molekuli proteina negativno naelektrisani i putuju prema anodi u donjem rezervoaru. Primenom električnog polja dolazi do razdvajanja molekula u diskretne zone (trake). Po završetku razdvajanja gel se izvadi iz cevčice a proteinske trake se vizualizuju jednom od metoda detekcije.
Danas se PAGE retko izvodi u cevčicama, već je zamenjena jed-nostavnijim tipom PAGE na pločastim gelovima. Kod ovog tipa elek-troforeze gel debljine 0,5-1 mm se pravi između dve ploče od stakla ili pleksiglasa. Veličina pora i stepen umreženosti pora se podešavaju ko-rišćenjem različitih koncentracija monomera akrilamida i komomera bisa (metilenbisakrilamida). Povećavanjem njihove koncentracije veli-čina pora u gelu se smanjuje što direktno utiče na razdvajanje protei-na. Pored akrilamida i bisa za pravljenje gela su neophodni inicijator TEMED (NNNN tetrametilenamid), amonijum persulfat koji katalizuje reakciju, pufer kojim se obezbeđuje disocijacija proteina i voda. Na vrh smeše za polimerizaciju postavi se tzv. češalj pomoću kojeg se tokom polimerizacije gela formiraju bunari u koje se stavljaju uzorci. Apara-tura za izvođenje PAGE se sklapa tako da se polimerizovani gel donjim delom uroni u donji rezervoar s puferom, a deo prostora iznad mesta

311
Primena genetskih markera u identifikaciji
za nanošenje uzorka, ispuni puferom što čini gornji rezervoar za pufer. U bunare se potom nanosi određena količina uzorka, aparatura se spoji s izvorom struje te vrši proces razdvajanja molekula. Od vrste protei-na koji se analiziraju zavisi koji će se puferski sistemi koristiti i u kom vremenskom periodu će se proteinske komponente razdvajati (Cooke, 1988., 1992; Linskens i Jackson, 1992). Sistem za PAGE elektroforezu se sastoji od kalupa za gel, podloge za fiksiranje kalupa za gel, posude za elektrodni pufer u kome su elektrode, sistema za vodeno hlađenje i ispravljača (sl. 6).
Da bi se dobila bolja i oštrija proteinska slika umesto PAGE-a s kon-tinuiranim gelom, koristi se tzv. disk elektroforeza. Kod ovog tipa elek-troforeze koriste se dva gela. Razdvajajući gel u kojem se odvija separaci-ja molekula nadslojen sa kratkim (1 cm) koncentrujućim gelom velikih pora i za dve jedinice nižeg pH. Primenom električnog polja uzorci se sabijaju u koncentrujućem gelu u oštru zonu i tako ulaze u razdvajajući gel gde se vrši razdvajanje molekula. Vreme i uslovi razdvajanja su slični kao i kod PAGE sa kontinuiranim gelom.
Kod gore opisanih vrsta PAGE elektroforeze ne narušava se struktura proteina te se oni razdvajaju na osnovu naelektrisanja oblika i molekul-ske mase. Kako korišćenjem ove PAGE proteini ostaju nativni i održavaju svoju funkcionalnost mogu se koristiti za razdvajanje kako rezervnih tako i funkcionalnih proteina, izoenzima. Međutim, zbog efikasnosti za raz-dvajanje izoenzima se znatno češće koristi skrobna elektroforeza.
Slika 6. Aparatura za vertikalnu PAGE Figure 6. Vertical PAGE (Taški-Ajduković, 2009 )

312
Semenarstvo
Poseban oblik poliakrilamidne elektroforeze je SDS-PAGE kod koje se razdvajanje proteina odvija u denaturišućim uslovima. Tehnika se ba-zira na tretmanu proteina rastvorom koji sadrži natrijum-dodecilsulfat (eng. Sodium dodecyl sulphate, SDS). SDS je anjonski deterdžent koji ometa gotovo sve kovalentne interakcije u nativnom proteinu izazivaju-ći denaturaciju i razmotavanje molekula proteina. Anjoni SDS se vežu za glavni lanac u razmeri otprilike jedan anjon SDS na svake dve amino kiselinske subjedinice. Ovako nastali kompleks SDS-a sa denaturisanim proteinom ima veliko negativano naelektrisanje, po pravilu mnogo veće od naelektrisanja koje nosi nativni protein. Na taj način se maskiraju in-dividualne razlike u naelektrisanju proteina i oni se razdvajaju samo na osnovu relativne molekulske mase. Pod takvim uslovima razdvajanja, manji molekuli će znatno lakše prolazitii kroz umreženu strukturu i u gelu će doći do raspoređivanja proteinskih molekula u oštre trake tako da će pri vrhu gela biti raspoređeni molekuli velike molekulske mase (HMW, eng. High molecular weight), u sredini molekuli srednje mo-lekulske mase (MMW, eng. Medium molecular weight) i pri dnu gela molekuli male molekulske mase (LMW, eng. Low molecular weight). Bitno je napomenuti da se ova razdvajanja izvode na istim aparaturama kao i nativna PAGE, jedino što se u sklopu poliakrilamidnog gela kao i u sklopu pufera za elektroforezu nalazi SDS.
Po završetku elektroforeze nastale zone (trake) u gelu nije moguće videti, nego ih je potrebno vizuelizovati nekom od tehnika vizuelizaci-je. Najčešće su to tehnike bojenja proteina različitim bojama. Svakako najraširenija tehnika bojenja proteinskih molekula, je bojenje sa Coo-massie brilliant blue (Coomassie plavim). Kod ove se metode gel uroni u kiseli, alkoholni rastvor boje. Pri tome dolazi do fiksiranja proteina u gelu zbog njihove denaturacije i vezivanja boje na molekul proteina. Višak boje se potom ispere rastvorom za obezbojavanje. Ovom se meto-dom može detektovati količina proteina od 1×10-6 g. Trake na gelu koji sadrže manje količine proteina od navedene, potrebno je vizuelizovati primenom osetljive tehnike bojenja – bojenje srebrom, koja je oko 100 puta osjetljivija od Coomassie tehnike bojenja.
Na mestima gde se nalaze molekuli proteina bojenjem se stvaraju obojene zone-trake, i dobija čitav spektar-profil traka (sl. 7. i 8.). Dobije-ni profil (elektroforegram) se analizira u odnosu na poznati standard ili markere poznate pokretljivosti (Metakovsky et. al., 1984.; Wrighley et.

313
Primena genetskih markera u identifikaciji
al., 1985.). Identifikacija vrste se vrši poređenjem učestalosti određenih proteinskih komponenti, a uniformonost na osnovu njihovih razlika.
Nativna PAGE elektroforeza se najčešće koristi za analizu rezervnih proteina cerealija, prolamina, u kiseloj pH, tzv. A-PAGE. A-PAGE meto-da elektroforeze je moćna i veoma korišćena tehnika za ispitivanje cere-alija. Prema podacima Cooke (1995a) postoji 17 publikovanih metoda A-PAGE za analizu glijadina pšenice koji se uglavnom razlikuju u sasta-vu gela i puferskog sistema.
Postojanje ovako velikog broja metoda uzrokuje probleme pri po-ređenju rezultata različitih laboratorija. Zbog toga organizacije kao što je Intenational Seed Testing Association (ISTA) standardizuju metode za verifikaciju sorti i hibrida. Metod za analizu pšenice i ječma zasniva se na analizi proteina rastvornih u alkoholu sa PAGE pH 3.2. (Cooke, 1992). Na taj način se na gelu može jasno razlikovati oko 40 traka glija-dina (sl. 7) i oko 30 hordeinskih traka. Ovako veliki polimorfizam omo-gućuje veliki stepen razlikovanja između varijeteta. Tako Cooke (1987) saopštava da je na osnovu A-PAGE od 150 ispitanih varijeteta pšenice koje se gaje u UK ovom metodom moguće razlikovati 137 varijeteta. A-PAGE korišćena je i za ispitivanje 353 varijeteta ječma koji su na osnovu sastava hordeina podeljeni u 70 grupa (Cooke, 1995a). A-PAGE se može primeniti i na razdvajanje proteina ovsa i tritikale, a uz izvesne modi-fikacije može se primeniti na identifikaciju pirinča, kuku-ruza i sirka.
Slika 7. Elektorforetsko razdvajanje rezervnih proteina semena različitih
sorti pšenice A-PAGEFigure 7. Electrophoretic separati-on of seed storage proteins within
different wheat varieties by A-PAGE(Taški-Ajduković, 2009 )

314
Semenarstvo
SDS-PAGE je veoma jednostavan i efekasan metod za razdvajanja i detekciju polimorfizma ukupnih proteina semena. Ovaj metod se može koristiti za ispitivanje velikog broja biljnih vrsta: graška (sl. 8), soje (Taš-ki-Ajduković et. al., 2008a,b; 2010), pasulja, sočiva, velikog broja trava i krmnog bilja, kafe, pamuka kao i cerealija (Cooke, 1988). Za samooplod-ne biljne vrste, kao što je grašak, SDS-PAGE proteina semena je veoma korisna i usvojena je od strane ISTA kao standardna referendna metoda (ISTA pravila, 2009).
Izoelektrično fokusiranje
Izoelektrično fokusiranje (IEF) (sl. 9) je metoda u kojoj se proteini raz-dvajaju u prisustvu kontinuiranog pH gradijenta. U takvim uslovima, pro-teini migriraju zbog svog naelektrisanja dok ne dostignu zonu s vrednošću pH pri kojoj nemaju neto naelektrisanje (tj. svoju izoelektričnu tačku, pI). Proteini će, stoga, dostići ravnotežno stanje „nulte” migracije i fokusirati se u uske zone. IEF tehnikom moguće je razdvojiti kompo-nente čije se pI vrednosti ra-zlikuju za 0,02 pH jedinice, a ako se koriste uski imobilizi-rani pH gradijenti ta se razlika može smanjiti na svega 0,001 pH jedinica.
Slika 8. Rezervni proteini graška razdvojeni SDS-PAGE
Figure 8. Storage proteins of pea variety separate by SDS-PAGE
(Taški-Ajduković, 2009)
Slika 9. Aparatura za IEFFigure 9. IEF Apparatus
(Taški-Ajduković, 2009)

315
Primena genetskih markera u identifikaciji
Najpopularniji način uspostavljanja gradijenta pH za izoelektrično fokusiranje je ugradnja niskomolekularnih amfoternih molekula u po-liakrilamidni gel. Takve supstance imaju bliske pI vrednosti i pokrivaju određeni raspon pH. Kada ne deluje električno polje, amfoliti su nasu-mično raspoređeni u gelu. Primenom električnog polja amfoliti počinju putovati prema jednoj od elektroda, u zavisnosti od naelektrisanja. Npr. amfolitni molekuli s najnižim pI ima će najnegativnije naelektrisanje i pu-tovati prema anodi sve dok im neto naelektrisanje ne bude jednako nuli. Na tom će se mestu koncentrovati formirajući usku zonu. Na taj način svaki amfolitni molekul migrira do mesta gde mu je neto naelektrisanje jednako nuli, bez obzira na početnu poziciju. Kako svaki od amfolita ima visoki puferski kapacitet, pH okolnog medijuma jednak je pI vrednosti svakog amfolita. Po završetku koncentrovanja amfolita postiže se ravno-težno stanje i u gelu se uspostavlja kontinuirani gradijent pH. Kada se protein nanese na gel, pod uslovom da je njegov pI unutar raspona kojeg pokrivaju amfoliti korišteni za uspostavu pH gradijenta, dolazi do njegove migracije do mesta na kojem se pH podudara s njegovom pI vrednošću. Za razliku od drugih elektroforetskih tehnika, pri izoelektričnom fokusi-ranju uzorak se može naneti bilo gde na gelu jer će molekuli sa istim pI vrednostima uvek putovati na istu poziciju. Zbog toga se proteini visoko koncentrišu u uskim zonama. Ako dođe do njihove difuzije ponovo će postati naelektrisani pa će ih delovanje električnog polja vratiti u zonu u kojoj im je neto naelektrisanje jednako nuli.
Kako IEF razdvaja proteine na osnovu njihovog naelektrisanja, po-trebno je u gelu izbeći efekt sita. Zato se koriste gelovi s nižom kon-centracijom poliakrilamida (3–5% T). IEF se može izvoditi u nativnim uslovima, u gelovima kojima su dodatni samo amfoliti. U denaturišu-ćim uslovima IEF se izvodi uz dodatak uree u visokim koncentracijama.
S obzirom na hemijski sastav i način proizvodnje amfoliti mogu biti dobijeni reakcijom oligoamina s akrilnom kiselinom ili kopolimerizaci-jom amina, aminokiselina i dipeptida s epihlorohidrinom. Tako dobije-ni amfoliti sadrže stotine različitih tipova molekula čije su pI vrednosti jednako raspoređene i pokrivaju široki raspon pH (npr. 3-11). Samo mali broj proteina ima pI vrednosti koje izlaze iz pH raspona 3.5-10, a za većinu proteina su između pH 4-6. IEF se može, zavisno od svojstava proteina koji se analiziraju, izvoditi u gradijentima širokog, ali i vrlo uskog pH gradijenta. Kako bi se kalibrisao pH gradijent u gelu mogu se koristiti standardni proteinski marker poznatih pI vrednosti.

316
Semenarstvo
Iako IEF obezbeđuje visok stepen razdvajanja proteina ova metoda nije široko primenjena za identifikaciju varijeteta (Cooke, 1995a). IEF je počela više da se koristi kada je postalo moguće lako pripremanje veoma tankih gelova (debljine do 0,1mm) (jer je na taj način znatno smanjena potrošnja amfolita, koji su veoma skupi) ili kupovina gotovih gelova koji su se mogli koristiti odmah nakon rehidratacije u odgovarajućem puferu. IEF se češće koristi za identifikaciju i određivanje genetske čistoće strano-oplodnih biljnih vrsta a znatno ređe za glavne samooplodne biljne vrste. Prolamini kukuruza se sastoje od velikog broja polipeptida sličnih mo-lekulskih masa ali značajno različitih u naelektrisanju te se IEF pokazalo kao adekvatna metoda za njihovo testiranje (Taški-Ajduković et. al., 2009) (sl. 10). ISTA je standardizovala ovu metodu za idendifikaciju i određiva-nje stepena čistoće hibrida kukuruza i suncokreta (ISTA pravila, 2009).
Dvodimenzionalna elektroforeza
Za verifikaciju sorti i hibrida može se koristiti dvodimenzionalna elektroforeza. Uzorci se prvo analiziraju u jednoj dimenziji (npr. IEF) a nakon toga se gel rotira pod uglom od 90° stepeni za razdvajanje dru-gom tehnikom (npr SDS-PAGE). Mapa dobijena 2D analizom može se sastojati i od preko sto tačaka što omogućava razlikovanje čak i genet-ski vrlo sličnih genotipova. Međutim, kompleksnost izvođenja tehnike i interpretacije rezultata limitirali su upotrebu 2D elektroforeze u rutin-skim analizama.
Slika 10. Analiza genetske čistoće hibrida kukuruza IEF na tankom sloju Figure 10. Measurement of hybrid purity of maize variety by ultrathin-layer IEF
(Taški-Ajduković, 2009)

317
Primena genetskih markera u identifikaciji
DNK-markeri
Biohemijsko-genetski markeri su veoma pouzdani u oceni genet-ske čistoće različitih hibrida, sorti i linija. Međutim, njihova primena je ograničena u utvrđivanju genetskog identiteta jer se njima postiže veoma mala pokrivenost genoma (Zlokolica i Taški-Ajduković, 2004) te se u poslednjoj deceniji sve češče njihova identifikacija vrši na osno-vu polimorfizma DNK-markera (Smith i Smith, 1992). Kako su tehnike zasnovane na DNK molekularnim markerima relativno nove i znatno skuplje od elektroforetskih tehnika kontrole na osnovu rezervnih pro-teina i izoenzima još uvek nisu u širokoj upotrebi ali ih koristi sve veći broj kompanija osobito kod identifikacije varijeteta.
U osnovi postoje dva opšta tipa DNK-markera: RFLP (Restriction Fragment Lenght Polymorphism) i markeri bazirani na lančanoj reakci-ji polimeraze (PCR - Polymerase Chain Reaction).
RFLP (Restriction Fragment Lenght Polymorphism) markeri
RFLP markeri su zasnovani na polimorfizmu genomske DNK koja se detektuje preko razlike u dužini fragmenata DNK posle isecanja re-strikcionim enzimima i hibridizacije sa odgovarajućim probama. RFLP markeri su kodominantni, odnosno omogućuju razlikovanje hetero-zigota od homozigota. Odlikuju se visokom osetljivošću, jer promena samo jedne baze u redosledu nukleotida na mestu prepoznavanja enzi-ma dovodi do izostajanja prepoznavanja i isecanja putem endonuklea-za, čime se RFLP-profil ispitivane DNK menja.
Primena RFLP markera u ispitivanju polimorfizma zahteva veliku količinu DNK i korišćenje radioaktivno obeleženih proba što je uticalo na razvoj novih tipova molekularnih markera.
Markeri zasnovani na PCR reakciji (Polymerase Chain Reaction)
Ovi markeri se zasnivaju na polimorfizmu između definisanih ge-nomskih DNK sekvenci koji se amplifikuju u PCR reakciji. Razvoj mar-kera zasnovanih na PCR reakciji vezan je za epohalno otkriće reakcije lančane polimerizacije (PCR) od strane Mullis et al. (1986).

318
Semenarstvo
PCR (Polymerase Chain Reaction) odnosno reakcija lančane po-limerizacije je metod koji omogućava selektivno umnožavanje odre-đenog segmenta molekula DNK. Preduslov za izvođenje reakcije je po-znavanje strukture (sekvence) graničnih regiona željenog segmenta, i posedovanje sintetskih jednolančanih oligonukleotida – tzv. prajmera, koji su komplementarni ovim regionima. Reakcija se odvija na principu cikličnog ponavljanja replikacije molekula DNK, pri čemu se prajmeri vezuju za matrične niti i služe DNK polimerazi kao početnice za sin-tezu komplementarnih lanaca nukleotida. Svaka faza PCR reakcije je
Slika 11. Šematski prikaz reakcija lančane polimerizacije (PCR): 1. Denaturacija 2. Hibridizacija 3. Elongacija (P- polimeraza) 4. Kompletiran prvi ciklus
Figure 11. Schematic of the Polymerase Chain Reaction (PCR):1. Denaturing 2. Annealing 3. Elongation (P-Polymerase) 4. The first cycle is complete
(George Rice, Montana State University)

319
Primena genetskih markera u identifikaciji
određena temperaturom na kojoj se odvija i vremenom trajanja. Ključni korak u razvoju metode označila je izolacija termostabilne DNK poli-meraze, koja može da sačuva efikasnost tokom cikličnih temperaturnih promena. Takav enzim dobijen je iz termofilnog bakterijskog soja Ther-mus aquaticus, i poznat je kao Taq polimeraza.
U prvom koraku PCR reakcije (sl. 11) vrši se denaturacija uzorka DNK na temperaturi od 94 do 96°C. U drugoj fazi – fazi hibridizacije, par prajmera se specifično vezuje za komplementarne regione DNK, ograni-čavajuci segment koji treba da bude amplifikovan. Povezivanje prajmera se odvija na temperaturi koja zavisi od njihove sekvence, tj. redosleda nu-kleotida, a iznosi 50–70 °C. Izbor optimalne temperature u ovoj fazi je od ključnog značaja za specifičnost PCR reakcije. Treća faza je faza ekstenzije prajmera. Enzim Taq DNK polimeraza vrši sintezu novih (komplemen-tarnih) lanaca nukleotida, koji se nastavljaju na („produžuju”) prajmere u 5’–3’ smeru. Prekursorski molekuli za sintezu su slobodni nukleotidi u obliku dezoksinukleozid-trifosfata (dNTP), a reakciju katalizuju joni ma-gnezijuma. Taq polimeraza ostva-ruje optimalno dejstvo na 72oC, pa je to uobičajena temperatura na kojoj se odvija faza ekstenzije; vre-me trajanja je određeno dužinom (veličinom) regiona koji se umno-žava. Na završetku ove faze dobi-jaju se dve kopije segmenta DNK koji je ograničen prajmerima. Re-akcija se zatim ciklično ponavlja, a u svakom novom ciklusu kao ma-trice služe i novosintetisani mole-kuli DNK. Ceo proces je automati-zovan i odvija se u PCR aparatu (sl. 12). U aparatu se nalazi termoblok u koji se stavljaju tube sa PCR re-akcionom smešom. Programira-njem PCR aparata da povećava i snižava temperaturu termobloka, obezbeđuje se odvijanje PCR ci-kusa i sinteza DNK fragmenta.
Slika 12. PCR aparat Figure 12. Termocycler(Taški-Ajduković, 2011 )

320
Semenarstvo
Nakon 30 ciklusa željeni segment DNK se umnoži 106 puta, što omoguća-va njegovu vizuelizaciju i analizu nakon gel-elektroforeze i bojenja.
Danas su na raspolaganju različiti tipovi markera zasnovanih na PCR reakciji od kojih su u identifikaciji i određivanju genetske čistoće najviše korišćeni RAPD (Random Amplified Polymorphic DNA), SCAR (Sequence Characterised Amlified Regions), AFLP (Amlified Fragment Length Polymorphism), SSR (Simple Sequence Repeats ili mikrosateliti) i markeri nove generacije SNP (Single Nucleotide Polymophism).
RAPD (Random Amplified Polymorphic DNA) markeri
RAPD metoda se zasniva na korišćenju nespecifičnih oligonukle-otidnih prajmera (najčešće 10 dužine baznih parova) koji se vezuju za genomsku DNK na različitim mestima jednog od komplementarnih la-naca DNK. Gde god da se prajmeri vežu na genomu, DNK sekvenca između njih će se umnožavati. Na primeru datom na slici 13 prajmeri se vezuju na pet mesta na četiri različita hromozoma. Na svakom od me-sta prajmeri se vezuju na različitim rastojanjima jedan od drugog te se
Slika 13. Princip RAPD PCR – Figure 13. Principle of RAPD PCR

321
Primena genetskih markera u identifikaciji
usled toga umnožavaju fragmenti DNK različite dužine. Ako je rastoja-nje između mesta vezivanja prajmera preveliko, nisu pravilno orjentisa-ni ili nema mesta vezivanja u suprotnoj orjentaciji (hromozom 3, sl. 13) do amplifikacije neće doći. Nastali produkti se razdvajaju na agaroznom gelu uz prisustvo etidijum bromida i posmatraju pod UV svetlom.
Genotipovi se mogu razlikovati na osnovu razlike u DNK sekvenci. Ona nastaje usled delecija, insercija ili tačkastih mutacija koje kreiraju ili sklanjaju mesto na koje može da se veže analizirani prajmer. RAPD mar-keri su korišćeni za analizu polimorfizma velikog broja poljoprivrednih biljnih vrsta: soje (Williams et. al., 1990; Zhang i sar 1996. a, b), kukuruza (Marsan et al., 1993; Zhang et al., 1996a), pšenice (Devos i Gale, 1992), suncokreta (Popov et al., 2002; Popović et al., 2008), graška (Samec i Na-sinec, 1996), lucerke (Nagl et al., 2010) (sl. 14), šećerne repe (Nagl et al., 2007, 2008) i mnogih drugih.
Prednost RAPD markera je što se mogu primeniti na sve biljne vr-ste, obezbeđuju kompletnu pokrivenost genoma, vrlo su jednostavni za analizu i nije potrebno znati sekvencu DNK da bi se dizajnirali prajmeri. Veliki broj RAPD prajmera može dati polimorfne DNK produkte, ali svi prajmeri nisu u istoj meri informativni. Cilj je da se pronađu RAPD prajmeri koji će adekvatno opisati veze između genotipova, biti poli-morfni i dati ponovljive rezultate (McDonald, 1996). RAPD markeri su dominantni markeri te nije moguće razlikovati da li je PCR produkt rezultat amlifikacije lokusa koji je hetrozigotan (1 kopija) ili homozigo-tan (2 kopije). Kodominantni markeri, koji proizvode DNK fragmente
Slika 14. Identifikacija genotipova lucerke RAPD-PCRFigure 14. Identification of alfalfa populations using RAPD-PCR (Taški-Ajduković, 2009)

322
Semenarstvo
različite dužine sa istog lokusa se detektuju veoma retko. S obzirom na to da se RAPD markeri zasnivaju na vezivanju prajmera slučajnog re-dosleda baza, njihova ponovljivost je u velikoj meri uslovljena uslovima reakcije, što predstavlja još jedan od nedostataka ove tehnike. Problem ponovljivosti rezultata (čak i u istoj laboratoriji) onemogućava standar-dizaciju ove metode i njenu široku primenu u identifikaciji i određiva-nju genetske čistoće.
Da bi se prevazišli ovi problemi RAPD markeri se prevode u speci-fične prajmere Sequence Characterized Amplified Regions (SCAR). Ovi prajmeri omogućuju specifičnu amplifikaciju određenog lokusa, a praj-merski par se kreira na osnovu odgovarajućeg RAPD fragmenta. Am-plifikovana DNK se izoluje sa gela i umnožava PCR reakcijom. Produkt se klonira i senkvencionira. Na osnovu dobijene sekvence se dizajnira-ju novi, duži specifični prajmeri, najčešće produžavanjem originalnog RAPD prajmera sa 10–15 baza (Schneider K et. al., 1999).
AFLP (Amplified Fragment Lenght Polymorphism) markeri
AFLP metoda kombinuje restrikciono isecanje i PCR (sl. 15). U pr-vom koraku se DNK iseče sa dva tipa restrikcionih enzima: jedan koji prekida lanac češće (prepoznaje restrikciona mesta od 4 bazna para) i drugi koji prekida lanac ređe (prepoznaje restrikciona mesta od 6 ba-znih parova). Na dobijene fragmente se vezuju specifični adapteri, a za-tim se fragmenti umnožavaju PCR reakcijom. Prajmeri koji se koriste za umnožavanje imaju komplementarnu sekvencu adapteru i nasumično dodata jedan, dva ili tri nukleotida (A, T, C ili G). Ovo omugućuje spe-cifičnost prajmera i umnožavanje samo jednog dela fragmenata DNK, na osnovu čega se može utvrditi da li ispitivani uzorak pripada datoj vrsti. PCR umnožavanje se obično izvodi u dva ciklusa. U prvom se adapterima dodaje jedan nukleotid, što dovodi do umnožavanja samo dela fragmenata, a u drugom se dodaju još jedan ili dva, što dovodi do umnožavanja samo dela prethodno umnoženih fragmenata, te se pove-ćava specifičnost reakcije. AFLP metoda omogućava detekciju velikog broja polimorfizma što je čini pogodnom za identifikaciju genotipova. Međutim i kod AFLP markera se dešavaju interlaboratorijske razlike u dobijenom polimorfizmu (Smith i Register, 1998). AFLP markeri su do-minantni markeri te i sa njima nije moguće detektovati sranooplodnju što predstavlja praktičan problem pri detekciji genetske čistoće. Rad sa

323
Primena genetskih markera u identifikaciji
AFLP markerima je veoma spor i osetljiv, a i cena je prilično visoka. Zbog svega navedenog AFLP markeri nisu našli širu primenu u testira-nju genetske čistoće i identifikaciji sorti i hibrida.
Slika 15. Princip AFLP PCR1. Prvi ciklus PCR- preamplifikacija 2. Drugi ciklus PCR-amplifikacija
Figure 15. Principle of AFLP PCR1.1st PCR round-preamplification 2. 2nd PCR round-amplification
(Martinez C., IB-INTA)

324
Semenarstvo
SSR (Simple Sequence Repeats) markeri
Varijabilan broj tandemskih ponovaka VNTR (Variable Number of Tandem Repeat) je osnova na kojoj se zasnivaju mikrosateliti, odnosno SSR markeri. Tandemski ponovci su DNK sekvence koje se sastoje od grupe nukleotida (obično ih ima između 2 i 5), koje se ponavljaju na-dovezujući se u dužinu. Ove male ponovljive sekvence su rasprostranje-ne po celom genomu a DNK sekvence koje ih uokviruju su konstantne za svaku biljnu vrstu. Nakon PCR reakcije sa prajmerima kreiranim na osnovu ovih konzerviranih sekvenci detektuje se polimorfizam u dužini dobijenog fragmenta, jer je broj ponovljivih sekvenci karakterističan za svaki genotip (sl. 16).
Slika 16. Princip SSR – Figure 16. Principle of SSR (IB – INTA)

325
Primena genetskih markera u identifikaciji
Pošto se razlike u dužini fragmenata mogu detektovati čak i između bliskih srodnika mikrosateliti veoma brzo dobijaju na važnosti u identi-fikaciji razlika između različitih genotipova. Za umnožavanje mikrosa-telita koristi se standardni PCR protokol. U PCR reakcijama se koriste mikrosatelitni prajmeri koji mogu biti neobeleženi, ili fluorescentno od-nosno radioaktivno obeleženi. Dobijeni produkti se razdvajaju na razli-čite načine. Produkti koji su nastali u reakcijama sa neobeleženim praj-merima mogu se razdvajati na agaroznom ili poliakrilamidnom gelu. Za razdvajanje produkata SSR-PCR najčešće se koristi PAGE elektroforeza a gel se boji srebronitratom (sl. 17, 18).
Slika 17. SSR analiza genotipova sojeFigure 17. SSR profiels of soybean varietes (Ana Laura Vicario, INASE)
Slika 18. Analiza genetske čistoće paradajza SSR markerimaFigure 18. Assesment of genetic purity of tomato cultivar (Liu et. al., 2007)
Razvoj novih molekularnih metoda genotipizacije je dovelo do ra-zvoja novih sistema za detekciju, zasnovanih na očitavanju fluorescen-cije sa gelova kao što su to na primer sistemi kompanije LI-COR (sl. 19),

326
Semenarstvo
Pharmacia Biotetech i Applied Biosystems. Kod svakog od ovih sistema detekcija DNK fragmenata se odvija pomoću očitavanja laserom, koji izaziva flourescenciju fluorescentno obeleženih produkata u trenutku kada one, tokom elek-troforeze prođu kroz skener.
Kao alternativna tehnika za elektrofo- rezu fragmenata DNA, koristi se kapilarna elektroforeza (CE), koja povećanje brzine reakcije (vreme elek-troforeze se meri mi-nutima) i mogućnosti potpune automatiza-cije procesa elektro-foreze, bez potrebe za pravljenjem gela ili ručnog postavljanja uzoraka. Kao i elektroforeza na gelu i kod CE fra-gmenti DNK se razdvajaju po veličini pomoću umrežene strukture ma-triksa. Nedostatak CE sistema je u tome sto se pre elektroforeze uzorci PCR reakcije moraju podvrgnuti dijalizi, što produžava vreme potrebno za njihovu analizu.
Osnovni elementi CE instrumenta su uska kapilara, dva suda za pufer i dve elektrode povezane za izvor visokonaponske struje. CE in-strumenti sadrže još i laserski izvor pobuđivanja, fluorescentni detektor, automtski uzorkivač (autosampler) koji nosi kivete sa uzorcima i kom-pjuter za kontrolisanje injektovanja uzorka i detekciju (sl. 20).
Kapilare za CE se prave od silicijum-dioksida (stakla) i najčešće im je untrašnji prečnik od 50 do 100 µm i dužinu od 25 do 75 cm. Puferi koji se koriste kod gel-elektroforeze se mogu koristiti i kod CE. Joni pufera za elektrolizu provode struju kroz kapilaru. Međutim, umesto matriks gela kroz koji će molekuli DNA prolaziti kod CE se koristi viskozni rastvor polimera kao medijum, za razdvajanje. Veći molekuli DNA duže ostaju u kontaktu sa fleksibilnim linearnim lancima polimera što je analogno prolasku molekula kroz pore gela. Kako kapilarna elektroforeza obuhvata
Slika 19. Li-Cor DNK analizator Figure 19. Li-Cor DNA Analyzer (Taški-Ajduković, 2011)

327
Primena genetskih markera u identifikaciji
korišćenje uzane kapilare punjene rastvorom polimera umesto gela za se-paraciju DNK, bolji odnos površina/zapremina omogućava mnogo efika-snije oslobađanje toplote koja nastaje tokom procesa elektroforeze, kao i primenu većeg napona za razdvajanje što se rezultuje bržim razdvajanjem.
Pre svakog injektovanja novog uzorka, upumpava se nova porcija ra-stvora polimera u kapilaru. Ovaj proces je analogan automatskom pripre-manju novog gela pre nanošenja novog uzorka. Vrsta i koncentracija ra-stvora polimera koja se koristi određuje rezoluciju (sposobnost razlikova-nja) koja se može postići u velikoj meri na sličan način, kao što se stepen umreženosti poliakrilamidnog gela odražava na sposobnost razlikovanja elektroforetskog sistema. Uzorci DNA se nanose u kapilaru primenom fiksiranog napona za definisani period vremena ili primenom pritiska koji tera uzorak da se „uvuče“ u ulazni deo kapilare.
Mikrosateliti su razvijeni za identifikaciju genotipova velikog broja agronomski važnih biljnih vrsta kao što su pšenica (sl. 20) (Perry, 2004, Röder et al., 1995; Bryan et al., 1997), ječam (Southworth, 2009), soja (Meesang i sar., 2001), kukuruz (Wang et al., 2007), pirinač (Nandakumar et al., 2004,), krompir (Reid i Kerr, 2007), paradajz (Liu, 2007) i dr.
Slika 20. Kapilarni sistem (ABI 3130 Genetic Analyzer i elektroforegram)Figure 20. Capilary system (ABI 3130 Genetic Analyzer and electrophoregram, Applied Biosystems)(Taški-Ajduković, 2011)

328
Semenarstvo
Nasuprot RAPD i AFLP tehnikama inplementacija SSR markera je veoma skupa i prajmeri moraju biti razvijani pojedinačno za najveći broj biljnih vrsta. Kada su već određeni prajmeri nekog mikrosatelita testiranje genetičkog materijala je relativno jeftino. Mikrosateliti su ko-dominantni markeri što znači da se na osnovu njih mogu razlikovati heterozigoti. PCR produkti dobijeni mikrosatelitima su visoko ponov-ljivi (Jones i sar., 1997). Ovo su razlozi zbog kojih je DNA radna grupa, Tehničkog komiteta za ispitivanje genetske čisoće (VARCOM) u okviru ISTA odabrala mikrosatelite za međulaboratorijsko testiranje varijeteta pšenice, pirinča, soje i kukuruza. Ispituje se mogućnost dobijanja istog DNK profila čak i kad se koriste različiti reagensi, oprema i radni pro-tokoli. Planira se standardizacija ovih metoda i njihovo uvođnje u ISTA pravila u poglavlje za testiranje varijeteta.
SNP -Single Nucleotide Polymorphism
SNP je polimorfizam jednog nukleotida u kojem je jedan od četiri nukleotida (A, T, C ili G) zamenjen drugim. SNP-ovi uzrokuju prome-nu sekvence DNK. Postoji veliki broj metoda za detekciju SNP (Kwok, 2001; Syvänen 2001) (sl. 22.). U osnovi sve postojeće metode se zasnivaju na četiri osnovne reakcije: hibridizacija sa alelel-specifičnim probama, ligacija oligonukleotida, ekstenzija prajmera sa pojedinačnim nukleo-
Slika 21. Identifikacija sorti pšenice SSR markerma. Vizuelizacija Li-CorFigure 21. Identification of wheat varieties by SSR on Li-Cor(Taški-Ajduković, 2010 )

329
Primena genetskih markera u identifikaciji
tidima ili restrikcija endonukleazama. Separacija produkata se može odvijati na čvrstoj ili tečnoj podllozi, ili se produkti mogu detektovati u tečnoj podlozi bez separacije pomoću indirektne kolorimetrije, masene spektrometrije, fluorescencijom i dr. Zbog visoke cene ove motede ove metode nemaju široku primenu u identifikaciji i ispitivanju genetske či-stoće sorti i hibrida.
Slika 22. SNP-CAPS ječma – Figur 22. Barley SNP-CAPS (Taški-Ajduković, 2009)
Detekcija prisustva genetičke modifikovanosti (GM)
Kontrola identiteta i genetske čistoće sorti i hibrida uključuje i testira-nje genetičke modifikovanosti. Genetički modifikovane sorte i hibridi su sve prisutniji, kako po broju GM varijeteta tako i po površinama koje zauzi-maju. GM varijeteti mogu biti tolerantni na herbicide i/ili insekticide, mogu proizvoditi biomolekule koji nisu redovno prisutni u tim biljnim vrstama (vitamin A, proteini i dr.) ili pak određene farmaceutske supstance (Taški-Ajduković et al., 2006). Prisustvo GM se određuje iz dvojakih razloga:
a) određivanje slučajnog prisustva GM u konvencionalnim sortama i hibridima (eng. Adventtitious Presence, AP) i
b) određivanje geneske čistoće genetički modifikovanih sorti i hi-brida.
Prisustvo GM se može detektovati na nekoliko načina. GMO tole-rantni na herbicide i insekticide se mogu detektovati na osnovu bioe-seja, imunoeseja ili PCR (Taški et al., 2005). Testiranje genetske čisto-će zahteva kvantitativne metode (procenat semena koji je tolerantan) dok se za AP testiranje mogu koristiti kvalitativan (prisustvo/odsustvo GM), semikvantitativan (procena prisustva GM) i kvantitativan metod (% prisutnosti GM) (Milošević i Taški-Ajduković, 2009).

330
Semenarstvo
LITERATURA
Bryan G J, Collins A J, Stephenson P, Orry A, Smith J B, Gale M D (1997): Isolation and characterisation of microsatellites from hexaploid bread wheat. Theor. Appl. Genet. 94, 557-563
Cooke R J (1987): The classification of wheat cultivars using a standard reference electrophoresis method. J. Nat. Agric. Bot. 17:273–281.
Cooke R J (1988): Electrophoresis in plant testing and breeding. Advan-ces in Electrophoresis, 2: 171-261.
Cooke R J (1995a): Gel electrophoresis for the identification of plant varieties. Journal of Chromatography, 698: 281-29.
Cooke R J (1995b): Variety identification: modern techniques and appli-cations. In: Basra, A. S. (ed.) Seed Quality – Basic Mechanisms and Agricultural Implications. Food Products Press, Binghamton, New York, pp. 279-318.
Cooke R J (Ed.) (1992): Handbook of Variety Testing-Electrophoresis Handbook: Variety Identification. The International SeedTesting Association. Zurich.
Devos K M, Gale M D (1992): The use of random amplified polymor-phic DNA in wheat. Theor. Appl. Genet. 84: 567–572.
Gerić I (1986): Kontrola genetske čistoće hibridnog semena. Semenar-stvo 1-2: 26-31.
Gerić I, Zlokolica M, Gerić C, Stuber C W (1989): Races and popula-tions of maize in Yugoslavia. Isozymes variations and genetic di-versity. Ed. by Intern. Board for plant genetic resources (IBPGR). Systematic and ecogeographic studies on corn genepools, 1-106.
Goodman M M, Stuber C W (1983): Maize. In: Isozymes in Plant Ge-netics and Breeding. S.D. Tanksley and T.J. Orton (Eds.). Elsevier Scientific Publ. Co., Amsterdam, Netherlands.
ISTA (2009): International Rules for Seed Testing. International Seed Testing Association, Switzerland.

331
Primena genetskih markera u identifikaciji
Jones C J, Edwards K J, Castaglione S, Winfield M O, Sa La F, Van De Wiel C, Bredemeijer G, Vosman B, Matthes M, Maly A, Brettsch-neider R, Bettini P, Butatti M, Maestri E, Malcevsci A, Marmroli N, Aert R, Volckaert G, Rueda J, Linaacero R, Vaque A, Karp A (1997): Reproducibility Testing of RAPD, AFLP and SSR Markers in Plants by a Network of European Laboratories. Molecular Bree-ding 3: 381-390.
Kwok P Y (2001): Methods for genotyping single nucleotide polymorp-hisms. Annu Rev Genomics Hum Genet 2:235–258.
Linskens H F, Jackson J F (Eds) (1992): Modern Methods of Plant Analysis. (New Series), V.14 Seed Analysis, Springer, Berlin.
Liu L-W, Wang Y, Gong Y-Q, Zhao T-M, Liu G, Li X-Y, Yu F-M (2007): Assessment of genetic purity of tomato (Lycopersicon esculentu-mL.) hybrid using molecular markers. Scientia Horticulturae 115 (1) : 7-12.
Marsan P A, Egidy G, Monfredini G, Disilvestro S, Motto M (1993): RAPD markers in maize genetic-analysis. Maydica 38:259-264.
McDonald M B (1996): Genetic Purity: From Protein Electrophore-sis to RAPDs. Proceedings of the Annual American Seed Trade Assotiation Corn and Sorghum Research Conference 50: 256 - 271.
Meesang N, Ranamukhaarachchi S L, Petersen M J, Andersen S B (2001): Soybean cultivar identification and genetic purity analysis using microsatellite DNA markers. Seed Science and Technology, 29: 637-645.
Metakovsky E V, Novoselskaya A Y, Kopus M M, Sobko T A, Sozinov A A (1984): Blocks of gliadin components in winter wheat detected by one-dimensional polyacrylamide gel electrophoresis. Theoreti-cal and Applied Genetics 67: 559-568.
Mezei S, Kovacev L, Cacic N, Nagl N (2002): In vitro propagation and development of dihaploids in sugar beet breeding. Biotechnology and Biotechnological Equipment 16(2): 58-61.
Mezei S, Kovačev L, Kuprešanin N, Čačić N, Mrkovački N (1996): Effect of colchicine on some quantitative traits and technologi-cal parametars of sugar beet. Acta Agronomica Hungarica, 44 (4): 1-6.

332
Semenarstvo
Milošević M, Taski-Ajdukovic K (2009): Methods of Transformation and Detection of Genetic Modifications in Field and Vegetable Crops. Acta Agriculturae Serbica XIV: 2897-106.
Morgan A G (1989): Cromatografic application in cultivar identificati-on. Plant & Seeds 2: 35-44.
Mullis K, Faloona S, Scharf S, Saiki R, Horn G, Erlich H (1986): Specific enzymatic amplification of DNA in vitro: the polymerase chain re-action. Cold Spring Harbor Symp. Quant. Biol., 51: 263-273.
Nagl N, Mezei S, Kovačev L, Vasić D, Čačić N (2004): Induction and micropropagation potential of sugar beet haploids. Genetika 36(3): 187-194.
Nagl N, Taški-Ajduković K, Barać G, Milić D, Katić S (2010): Ispitiva-nje mogućnosti primene RAPD markera u detekciji polimorfizma sorti lucerke Ratarstvo i povrtarstvo 47: 511-516.
Nagl N, Vidović D, Kitić M, Kovačev L (2008): Use of RAPD and SRAP markers in sugar beet DNA polymorphism analysis. Proceedings of International Conference “Conventional and Molecular Breeding of Field and Vegetable Crops”, Novi Sad, Serbia, 469-472.
Nagl N, Weiland J, Lewellen R (2007): Detection of DNA polymorphism in sugar beet bulks by SRAP and RAPD markers. 13th European Congress on Biotechnology, Barcelona, Spain, Journal of Biotech-nology 131 (2S): S32.
Nandakumar N, Singh A K, Sharma R K, Mohapatra T, Prabhu K V, Zaman F U (2004): Molecular fingerprinting of hybrids and asse-ssment of genetic purity of hybrid seeds in rice using microsatelli-te markers. Euphytica 136: 257-264.
Nikolić Z, Milošević M, Taški-Ajduković K, Vujaković M (2007): Genetic purity as a component of seed quality. Procidings of the 2nd Interna-tional simposium on agriculture, Opatija, Croatia, 245-248.
Osborne T. J. (1924) The vegetable proteins, Longman Green, London.Perry D (2004): Identification of Canadian durum wheat varieties using
a single PCR. Theor. Appl. Genet. 109:55–61.Popov V N, Urbanovich O Yu, Kirichenko V V (2002): Studying Ge-
netic Diversity in Inbred Sunflower Lines by RAPD and Isozyme Analyses. Russian Journal of Genetics 38:(7) 785–790.

333
Primena genetskih markera u identifikaciji
Popović Andrea, Nagl N, Taški-Ajduković K (2008): RAPD analysis of cultivated sunflower Helianthus annuus L. and its wild types. Pro-cidings of the International conference Conventional and Mole-cular breeding of field and vegetable crops, Novi Sad, Serbia, p 174-176.
Reid A, Kerr E M (2007): A rapid simple sequence repeat (SSR)-based identification method for potato cultivars Plant Genetic Resour-ces: Characterization and Utilization 5(1); 7–13.
Röder M S, Wendehake K, Korzun V, Bredemeijer G, Laborie D, Ber-trand L, Isaac P, Rendell S, Jackson J, Cooke R J, Vosman B, Ganal M W (2002): Construction and analysis of a microsatellite-based database of European wheat varieties. Theor. Appl. Genet. 106, 67-73.
Samec P, Nasinec V (1996): The use of RAPD technique for the identifi-cation and classification of Pisum sativum L. genotypes. Euphytica 89:229-234.
Schneider K, Borchardt D C, Schaefer-Pregl R, Nagl N, Glass C, Jeppson A, Gebhardt C, Salamini F (1999): PCR-based cloning and segre-gation analysis of functional gene homologues in Beta vulgaris. Mol. Genet. Genom. 262(3): 515-524.
Smith J S C, Register J C (1998): Genetic purity and testing technologies for seed quality: a company perspective Seed Science Research 8: 285-293.
Smith J S C, Smith O S (1992): Fingerprinting crop varieties. Advances in agronomy, 47: 85-140.
Southworth C (2009) Barley Variety Identification Using SSRs in Burns R. (ed.), Methods in Molecular Biology, Plant Pathology, vol. 508, Humana Press, LLC.
Stuber C W (1992): Biochemiical and molecular markers in plant bree-ding. Plant Breed. Rev. 9: 37-61
Stuber C W, Wendel J F, Goodman M M, Smith, J S C (1988): Tech-niques and Scoring Procedures for Starch Gel Electrophoresis of Enzymes from Maize (Zea mays L), Technical Bulletin 286, North Carolina Agricultural Research Service.

334
Semenarstvo
Syvänen A C (2001): Accessing genetic variation: genotyping single nucleotide polymorphisms. Nat Rev Genet 2:930–942.
Tanksley S D (1983) Molecular markers in plant breeding. Plant Mole-cular Biology reporter 1:3-8.
Tanksley S D, Orton T J (eds) (1983): Isozemes in Plant Genetics and Breeding, Elsevier, A, B volumes, Amsterdam, Oxford, New York.
Taški K, Milošević M, Zlokolica M, Vujaković M, Nikolić Z (2005): Polja Vojvodine - bez genetički modifikovanih useva. Arhiv za poljopri-vredne nauke 66: 209-213.
Taški-Ajduković K, Milošević M, Nikolić Z, Vujaković M (2006a): Ge-netically modified plants: development and monitoring. IV me-đunarodna Eko-konferencija “Zdravstveno bezbedna hrana” 20-23. septembar, Novi Sad, p 241-246
Taški-Ajduković K, Milošević M, Nikolić Z, Vujaković M (2006b): Upo-treba metoda reakcije lančane polimerizacije (PCR) u agrobioteh-nologiji. Selekcija i semenarstvo XII/ 3-4: 59-63.
Taski-Ajdukovic K, Djordjevic V, Vidic M, Vujakovic M, Milosevic M, Miladinovic J (2008a): The Main Seed Storage Proteins among High-protein Genotypes. Genetika 40: 9 -16.
Taski-Ajdukovic K, Djordjevic V, Vidic M, Vujakovic M, Miladinovic J (2008b): Genotype influence on subunits of main soybean seed storage proteins. 43rd Croatian & 3rd International Symposium on Agriculture, Opatia, Croatia, p109.
Taski-Ajdukovic K, Knoblauch R, Nevena N, Aranka J, Vujakovic M (2009): Genetic purity in seed testing using biomolecular mar-kers. IV Congres of the Serbian genetic society, Tara, Serbia, June 1-5, Book of abstracts 279.
Taski-Ajdukovic K, Djordjevic V, Vidic M, Vujakovic M (2010): Subunit composition of seed storage proteins in high-protein soybean genotypes. Pesquisa Agropecuária Brasileira (PAB), 45: 721-729.
Taški-Ajduković K, Radić V, Jevtić A, Čanak P, Vujaković M, Miklič V (2010): Uporedna analiza genetske čistoće roditeljskih linija sun-cokreta u polju i laboratoriji. Zbornik Instituta za ratarstvo i povr-tarstvo (prihvaćeno za štampu).

335
Primena genetskih markera u identifikaciji
Wang F G, Zhao J R, Dai J R, Yi H M, Kuang M, Sun Y M, Yu X Y, Guo J L, Wang L (2007): Selection and development of represen-tative simple sequence repeat primers and multiplex SSR sets for high throughput automated genotyping in maize. Chinese Science Bulletin 52(2): 215-223. APER.
Weir B S (1990): Genetic data analysis. Methods for discrete population genetics data. Sinauer Association, Sunderland,Massachusetts.
White J, Law J R (1991): Differntiation between varieties of oilseed rape (Brassica napus) on the basis of the fatty acid composition of the oil. Plant Vaerieties and Seed 4: 125-132.
Williams J G, Kubelik A R, Livak K J, Rafalski J A, Tingey S V (1990): DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 25:18(22):6531-5.
Wrighley C W, Autran J C, Bushunk W (1985): Identification of cereal varieties by gel electrophoresis of the grain proteins. Advances in Cereal Science and Technology, 5: 211-259.
Zhang J, McDonald M B, Sweeney P M (1996a): Random amplified polymorphic DNAs (RAPDs) from seeds of differing soybean and maize genoytpes Seed Science and Technology 24: 513-522.
Zhang J, McDonald M B, Sweeney P M (1996b): Soybean cultivar iden-tification using RAPD Seed Science and Technology 24: 589-592.
Zlokolica M, Milosević M (2001): Isozymes as genetic markers in maize breeding. Biologia Plantarum, 44 (2): 207-211.
Zlokolica M, Mezei S, Kovačev L (1994): Genetic variability in haploids and induced dihaploids from unpollinated sugarbeet ovules (Beta vulgaris L.) Jour. Scien. Agric. Research., 56: 198, 3-9.
Zlokolica M, Mezei S, Kovačev L, Nikolić Z, Milošević M, Kuprešanin N (1996c): Genetic variability of sugar beet (Beta vulgaris L.) calli following inoculation with Agrobacterium tumefaciens. Genetika (Yu), Vol. 28, No.2, str. 97-103.
Zlokolica M, Milošević M, Nikolić Z, Graovac M (1996a) The use of genetic marker for identification and estimation of vegetable plant species. Acta Horticulturae, 462: 75-82.
Zlokolica M., Taški K. (2004): Genetska čistoća semana U: M. Miloše-vić, M. Malešević, Eds., Semenarstvo, Ed 1, Vol. 1, Naučni institut za ratarstvo i povrtarstvo i Nacionalna laboratorija za ispitivanje semena, Novi Sad, p 201-240.

336
Semenarstvo
Zlokolica M, Turkav S, Milošević M, Graovac M, Škorić D (1996b): Isozymes variability of selfpollinated sunflower (Helianthus annuus L.) lines. Helia, 19:113-130.

Zorica Nikolić
Genetički modifikovanebiljne vrste –
metode za testiranje

338
Semenarstvo

339
Genetički modifikovane biljne vrste - metode za testiranje
UVOD
Genetički inženjering ili tehnologija rekombinantne DNK obuhvata metode koje omogućavaju prenošenje gena iz jednog organizma u DNK drugog organizma što dovodi do menjanja karakteristika organizma primaoca. Pojava genetički modifikovanih organizama (GMO) predstavlja početak efikasnijeg biološkog puta rešavanja mnogih problema sa kojima se čovek suočava. Transgene biljke, životinje, rekombinantni proteini i vakcine neosporno imaju veliki značaj za čovečanstvo. Primena biotehnologije u poljoprivredi je omogućila povećanje kvaliteta i prinosa gajenih biljnih vrsta, stvaranje useva otpornih na bolesti, insekte i korove, nepovoljne uslove sredine, kao i poboljšanje kvaliteta prehrambenih proizvoda i dr.
Ubrzani razvoj genetički modifikovanih organizama, u poslednjih deset godina, pokrenuo je niz pitanja u vezi sa njihovim oslobađanjem i praćenjem, kao i potencijalnim uticajem na spoljašnju sredinu i potrošače (Mi- lošević et al., 2008).
Površine pod genetički modifikovanim usevima kontinuirano rastu iz godine u godinu. Prema podacima iz 2010. godine GMO biljne vrste su gajene u dvadeset devet zemalja, a po površinama i dalje vode SAD, Brazil i Argentina (tabela 1). Kukuruz otporan prema kukuruznom plamencu, Bt kukuruz, se u 2008. godini komercijalno gajio u sedam zemalja Evropske unije: Španija, Češka Republika, Rumunija, Portugal, Nemačka, Poljska i Slovačka. U 2007. godini gajen je na 88.673 ha, a u 2008. godini površine su povećane na 107.719 ha, što je povećanje za oko 21%. U 2010. godini je šest zemalja EU nastavilo da gaji Bt kukuurz, a Češkoj Republici, Švedskoj i Nemačkoj se po prvi put gajio GMO krompir.
Očekuje se da će se do kraja 2015. godine genetički modifikovane biljne vrste gajiti u 40 zemalja na 200 miliona hektara (James, 2010).
Prema zakonskoj regulativi u Srbiji, u poljoprivrednim proizvodi- ma biljnog porekla koji sadrži jednu ili više autorizovanih genetičkih modifikacija mora se odrediti kvantitet, odnosno procenat genetičke modifikacije, kako bi se sproveo postupak obeležavanja poljoprivrednih proizvoda koji sadrže više od 0,9% GMO. Seme i reproduktivni mate- rijal se ne smatraju genetički modifikovanim ukoliko sadrže do 0,1% primesa genetički modifikovanih organizama (Sl. glasnik 41, 2009).

340
Semenarstvo
Tabela 1. Gajenje genetički modifikovanih biljnih vrsta u svetuTable 1. Global area of genetically modified crops in the world (James, 2010)
R.B.Rank
ZemljaCountry
Površine Area
(mil. ha)
Genetički modifikovana biljna vrstaGenetically modified crop
1● SAD / USA 66,8soja, kukuruz, pamuk, uljana repica, tikvica, papaja, šećerna repa, lucerka /soybean, maize, cotton, canola, squash,papaya, sugarbeet, alfalfa
2● Brazil 25,4 soja, kukuruz, pamuk / soybean, maize, cotton3● Argentina / Argentina 22,9 soja, kukuruz, pamuk / soybean, maize, cotton4● Indija / India 9,4 pamuk / cotton
5● Kanada / Canada 8,8 uljana repica, kukuruz, soja, šećerna repa canola, maize, soybean, sugarbeet
6● Kina / China 3,5 pamuk, paradajz, topola, papaja, slatka paprikacotton, tomato, poplar, papaya, sweet pepper
7● Paragvaj / Paraguay 2,6 soja / soybean8● Pakistan 2,4 pamuk / cotton 9● Južna Afrika / South Africa 2,2 soja, kukuruz, pamuk / soybean, maize, cotton10● Urugvaj / Uruguay 1,1 soja, kukuruz / soybean, maize11● Bolivija / Bolivia 0,9 soja / soybean12● Australija / Australia 0,7 pamuk, uljana repica / cotton, canola13● Filipini / Philippines 0,5 kukuruz / maize14● Mjanmar (Burma) / Myanmar 0,3 pamuk / cotton15● Burkina Faso 0.3 pamuk / cotton16● Španija / Spain 0,1 kukuruz / maize17● Meksiko / Mexico 0,1 pamuk, soja / cotton, soybean18 Kolumbija / Colombia < 0,1 pamuk / cotton
19 Čile / Chile < 0,1 kukuruz, soja, uljana repica /maize, soybean, canola
20 Honduras / Honduras < 0,1 kukuruz / maize21 Portugal < 0,1 kukuruz / maize
22 Češka RepublikaCzech Republic < 0,1 kukuruz, krompir / maize, potato
23 Poljska / Poland < 0,1 kukuruz / maize24 Egipat / Egypt < 0,1 kukuruz / maize25 Slovačka / Slovakia < 0,1 kukuruz / maize26 Kostarika / Costa Rica < 0,1 pamuk, soja / cotton, soybean27 Rumunija / Romania < 0,1 kukuruz / maize28 Švedska / Sweden < 0,1 krompir / potato29 Nemačka / Germany < 0,1 krompir / potato
●17 vodećih zemalja koje gaje 50.000 ha ili više

341
Genetički modifikovane biljne vrste - metode za testiranje
Metode za testiranje genetički modifikovanjihorganizama
Sa pojavom genetički modifikovanih biljaka i hrane, javila se i po- treba za odgovarajućim analitičkim metodama za njihovo testiranje (Anklam i Neuman, 2002).
Paralelno sa stalnim uvećanjem broja autorizovanih i neautorizova- nih genetičkih modifikacija radi se na razvijanju odgovarajućih metoda za njihovu analizu, karakterizaciju i kvantifikaciju. Uspešna strategija za ra- zvoj i primenu metoda podrazumeva stvaranje baza podataka, dostupnih javnosti, koje raspolažu informacijama o sekvencama korišćenim za stva- ranje GMO, metodama, kao i drugim relevantnim podacima o GMO.
Dve osnovne grupe metoda za testiranje GMO su: DNK metode, zasnovane na utvrđivanju prisustva modifovane DNK, i proteinske me- tode, koji identifikuju proizvod genetičke modifikacije – protein (sl. 1). Većina DNK metoda je bazirana na reakciji lančanog umnožavanja – PCR (Anklam et al., 2001).
Svaka od metoda ima svoje prednosti i nedostatke. Kada su u pita- nju proteinski metodi treba imati u vidu da u svim GMO biljnim vrsta- ma nije uvek prisutan „novi” protein ili količina proteina nekada nije dovoljna da bi se otkrila nekim od raspoloživih testova. Pored toga pro- tein može biti specifičan za određeno biljno tkivo, da ima različit nivo ekspresije u toku određenih faza fiziološkog razvoja biljke. Nivo trans- genog proteina se u odnosu na ukupni sadržaj proteina kreće u opsegu od 0 do 2%, čak i ako je ubačen jak promotor. U većini slučajeva nivo transgenog proteina je niži od 2% (Hemmer 1997).
Slika 1. Proteinski testovi za GMO a) ELISA test b) test trakaFigure 1. Protein based GMO detection a) ELISA test b) Lateral flow strip
a)
b)

342
Semenarstvo
GMO molekularna dijagnostika je veoma kompleksna. Visoka he- mijska i termička stabilnost DNK, velika osetljivost i specifičnost meto- de su glavne prednosti DNK metoda testiranja uzoraka. Međutim, upra- vo usled velike osetljivosti metode veliki je broj faktora koji može uticati na rezultat analize. Tako npr. DNK može biti prisutna u veoma maloj koncentraciji u uzorku, ili tokom postupka izolovanja može doći do nje- ne dezintegracije, što će dovesti do lažno negativnih rezultata analize.
Prilikom izbora metoda za testiranje GMO potrebno je razmotriti razlike u pogledu specifičnosti, preciznosti, vremena i cene analize iz- među proteinskih i DNK metoda (Lin et al., 2001). Biramo metodu koja će biti adekvatna, zavisno od postojeće tehničke opremljenosti, raspo- loživog vrema, kao i pitanja na koje želimo odgovor. U tabeli 2 dato je poređenje PCR i proteinskih metoda za GMO detekciju.
Tabela 2. Poređenje PCR i proteinskih metodaTable 2. Comparison of DNA and protein methods
DNK – PCR metod/DNA – PCR method
Proteinski metodi/Protein methods
Test trakama/Lateral flow strip test
ELISA/ELISA
strukturni/regulatorni/specifični geni
structural/regulatory/specific genes
rekombinantni protein/
recombinant protein
rekombinantni protein/
recombinant protein
Vreme / Time 1-3 dana/1-3 day 10-20 min 4-6 sati/4-6 hours
Cena / Cost 90-200 Eur 10-50 Eur 50-100 Eur
Instrumenti Equipment
PCR aparat/PCR machine – ELISA čitač/
ELISA reader
Osetljivost Sensitivity < 0,1% 0,1 - 10% 0,1%
Skreening (pregled)Screening da / yes ne / no ne / no
KvantifikacijaQuantification da / yes ne / no da / yes
IdentifikacijaIdentification da / yes da / yes da / yes

343
Genetički modifikovane biljne vrste - metode za testiranje
Organizacija laboratorije za GMO analize i okruženje
Cilj laboratorije koja se bavi molekularnom dijagnostikom je do- bijanje pouzdanih rezultata, te je jedan od glavnih zahteva obezbediti uslove rada kako bi se verovatnoća kontaminacije i dobijanja lažno po- zitivnih rezultata svela na minimum. U jednoj PCR reakciji se proizvo- de milioni kopija GMO sekvenci, tako da su PCR proizvodi glavni izvor kontaminacije koja se lako može proširiti kroz laboratoriju.
Laboratorija bi trebalo da bude organizovana da obezbedi jedan smer ili tok analize. Svaka faza u procesu GMO detekcije bi trebalo da se izvodi u odvojenoj prostoriji ili bar odvojenim prostorima u okviru jedne velike prostorije. Potrebno je imati odvojene prostore za: prijem uzorka i skladištenje uzoraka, homogenizaciju, izolaciju DNK, pripre- mu PCR reakcija (pre-PCR zona), za PCR instrumente, i za gel elektro-forezu (post-PCR zona) (sl. 2).
Slika 2. Tok uzorka u laboratoriji / Figure 2. Laboratory sample flow
Preventivne mere, kao što su: dekontaminacija UV zračenjem, pro-mena laboratorijske odeće pri prelasku u sledeću laboratoriju, korišće-nje laboratorijske opreme i reakcionih reagenasa u laboratoriji u kojoj se nalaze bez njihovog premeštanja u druge laboratorije, obezbeđuje da ne dođe do kontaminacije između različitih faza procesa detekcije GMO. U nekim laboratorijama u svetu postoji kontrola vazdušnog pritiska u cilju izbegavanja kontaminacija iz vazduha. U pre-PCR laboratoriji bi trebalo da je pozitivan pritisak vazduha, dok u post-PCR smanjen pritisak kako
UzorciSamples
Priprema uzorkaSamples preparation
Priprema rastvoraPreparation of solutions
Čuvanje uzorakaStorage of samples
DNK ekstrakcijaDNA extraction Pre PCR
Pre PCR
Post PCRPCR
Čuvanje DNK uzorakaStorage of DNA samples

344
Semenarstvo
ne bi došlo do disperzije PCR proizvoda. U sam protokol za GMO de-tekciju treba uvrstiti i kontrole za kontaminaciju vazduha.
Uslovi rada moraju biti odgovarajući da bi se korektno izveo test, jer su pojedini delovi procedure temperaturno osetljivi, naročito kada se mere male zapremine hemikalija. Variranja temperature u laboratoriji može uzrokovati smanjenu pouzdanost merenja pipeta, i tako uzroku-jući velike razlike u koncentraciji komponenti u PCR reakciji. Tako je održavanje i beleženje konstantne temperature esencijalno.
Osoblje koje radi mora biti visoko obrazovano i kontinuirano se obrazovati kako bi rezultati bili validni. Provera izvođenja testa se vrši tako što se uz rutinske uzorke dodaju interne kontrole poznatih koncen-tracija GMO.
Standardizacija metoda za testiranje GMO
U cilju standardizacije i validacije GMO metoda (za identifikaci-ju i kvantifikaciju), Evropski komitet za standardizaciju (CEN, Belgija), Francusko udruženje za standardizaciju (AFNOR, Francuska), Među-narodna organizacija za standardizaciju (ISO, Švajcarska), Međunarod-no udruženje laboratorija za testiranje semena (ISTA, Švajcarska) i dr. su pokrenule inicijative na tom planu.
Od 1988. godine CEN je izdao više od 260 evropskih standarda u oblasti hrane, nedavno i na polju GMO detekcije. Marta 2004. godine CEN je publikovao standard EN ISO 21572, koji se odnosi na proteinski metod za detekciju GMO, a 2005. godine standarde pod nazivom: Meto-di analize prisustva genetički modifikovanih organizama i njihovih pro-izvoda. Detaljni zahtevi i validovani postupci u izolovanju DNK dati su u standardu ISO 21571 (Nucleic acid extraction). Kvalitativne metode analize GMO su date u standardu ISO 21569 (Qualitative nucleic acid based methods), a kvantitativne u standardu ISO 21570 (Quantitative nucleic acid based methods) (Nikolić i sar., 2007, Milošević i sar., 2007).
Prema standardu ISO/IEC 17025 laboratorija bi trebalo da odabere metode koji su publikovani u međunarodnim ili nacionalnim standar-dima, ili dati od strane priznate organizacije, ili određeni naučni tekst ili časopis ili metoda koju je opisao proizvođač opreme. Svaki metod, pre nego što počne da se koristi za rutinsko testiranje, mora proći međula-boratorijsku validaciju.

345
Genetički modifikovane biljne vrste - metode za testiranje
Standardna procedura za validaciju analitičke metode uključuje ocenu parametara cele analitičke procedure. Kako se GMO analiza sa-stoji iz velikog broja različitih analitičkih koraka, moguće je validovati pojedinačne korake ili sve zajedno. Tako se mogu nezavisno validovati metoda ekstrakcije DNK, PCR metod za specifičnu sekvencu za pojedi-nu biljnu vrstu ili PCR metod za GM specifičnu sekvencu.
Podaci o validaciji moraju sadržati parametre metode: osetljivost, specifičnost, preciznost i tačnost. Ukoliko je u pitanju kvantitativno odre-đivanje sadržaja GMO, real-time PCR metodom, potrebno je odrediti granicu detekcije i kvantifikacije. Opseg kvantifikacije se može odrediti na osnovu referentnog materijala koji koristimo (Trapmann i sar., 2008).
Učešće u međulaboratorijskim testovima poređenja je od glavnog značaja za procenu rada laboratorije. Ove testove organizuju razne in-stitucije kao što su Institut for Food Research (UK), CSL (Gemma Sche-me) (UK), USDA/GIPSA Proficiency program, ISTA (CH).
Referentni materijal i ograničenja u GMO detekciji i kvantifikaciji
Važna komponenta metoda za testiranje GMO je referentni mate-rijal. Danas je komercijalno dostupan mali broj sertifikovanog referen-tnog materijala. Za GMO vrste koje nisu autorizovane u EU referentni materijal ne postoji.
Prilikom kvantitativnog određivanja sadržaja genetičke modifikacije u uzorku referentni materijal ima direktan uticaj na rezultat analize. Efekti kao što su DNK degradacija, kvalitet i dužina izolovane DNK, sličnost u ponašanju referentnog materijala koji je korišćen za kalibracionu krivu i metod validacije u odnosu na DNK izolovanu iz uzorka, igraju važne uloge u PCR reakciji. Otuda je veoma važna proizvodnja adekvatnog re-ferentnog materijala i njegova primena (Corbisier i sar., 2002).
Pokrenute su inicijative za poboljšanje dostupnosti referentnog ma-terijala uključujući korišćenje kontaminiranog materijala (Holst-Jensen i sar., 2003) kao i primena plazmida kao referentnog materijala (Kuriba-ra i sar., 2002, Taverniers i sar., 2004; Mattarucchi i sar., 2005).
Prema pravilima u EU da bi se novi GMO proizvod registrovao vla-snik mora da Evropskoj Komisiji dostavi materijal, metod za detekciju i

346
Semenarstvo
korišćenu GMO sekvencu, a informacije se dalje prosleđuju ovlašćenim referentnim laboratorijama za GMO.
Postoji mogućnost da genetička modifikacija bude u ekstranuklearnom genomu, npr. u hloroplastima. Broj ekstranuklearnih genoma nije stabilan u odnosu na nuklearni genom, a neki se nasleđuju samo od jednog rodi-telja. Bakterije i mikroorganizmi često imaju ekstrahromozomalnu DNK – plazmidi koji se mogu horizontalan prenositi čak između različitih vrsta. Sve navedeno bi moglo dodatno stvoriti probleme kod kvantifikacije.
Granica detekcije LOD i kvantifikacije LOQ metode su definisani kao najmanja količina koja se može pouzdano detektovati i kvantifikovati. Oba parametra su specifična za metod, ali u isto vreme zavise od uzorka koji je analiziran. Proizvodi kao što su ulje, hidroliziran skrob, rafinisani šećer i sirup, fermentisani proizvodi se karakterišu izrazito degradova-nom DNK ili je prisutna mala koncentracija DNK, u tragovima.
Prema Berdal and Holst-Jensen (2001) možemo razlikovati tri tipa granice: 1. apsolutna granica (najmanji broj kopija koji mora biti prisutan u prvom PCR ciklusu da bi se sa verovatnoćom od 95% korekno uradila detekcija ili kvantifikacija 2. relativna granica (najmanja relativan proce-nat GM materijala koji se može detektovati ili kvantifikovati u optimal-nim uslovima) 3. praktična granica (nivo koji je primenjen u toku analize uzorka uzimajući u obzir količinu DNK uzorka i apsolutni nivo). Predlo-ženo je da se u izveštaju o analizi dostave LOD/LOQ za metod i za analizu.
Veličina genoma varira između pojedinih biljnih vrsta. Približna veličina genoma soje i kukuruza je poznata: kukuruz 2,4–5,0x106 bp, a soje 0,9–1,2x106 bp. Relativna kvantifikacija je određivanje odnosa iz-među broja kopija GMO i broja kopija referentne ili endogene DNK, tako da je moguće izračunati broj kopija genomske DNK u uzorku koji testiramo. Transformisane biljke su heterozigotne, diploidna biljka ima odnos broja kopija specifičnog GMO DNK i specifične biljne DNK 1:2. Međutim, često se dešava da su transformisane biljke samooplodne što dovodi to promene u odnosu na 1:1. Hibridi gajenih biljnih vrsta su često tetra ili poliploidi, što se mora uzeti u obzir prilikom ukrštanja.
U praksi, obično nemamo informacije o nivou ploidnosti materija-la koji testiramo. U programima oplemenjivanja inbred GMO linije se ukrštaju sa nemodifikovanim sortama ili hibridima koji su adaptirani za specifične geografske i klimatske uslove. U tako stvorenoj GMO biljci moguće je da odnos GMO DNK i biljne DNK bude različit 1:2, 1:4 ili drugačiji, što će uticati na verodostojnost rezultata analize.

347
Genetički modifikovane biljne vrste - metode za testiranje
Procedura analize genetičke modifikacijePCR metodom
Detekcija rekombinantne DNK u biljnom materijalu i hrani pomo-ću PCR-a se izvodi u nekoliko koraka: izolacija DNK, umnožavanje cilj-ne sekvence PCR reakcijom – kvalitativan test, a ako je uzorak pozitivan nastavlja se analiza kvantitativnim testom. Ukoliko je sadržaj GMO u uzorku hrane ili hrane za životinje veći od 0,9% prema važećim pravili-ma mora biti obeležen (sl. 3).
Slika 3. Prikaz procedure PCR metode za detekciju GMOFigure 3. Procedure of PCR based method for detection of GMO
PozitivanPositive
NegativanNegative
UzorakSample
Ekstrakcija DNKExtraction of DNA
Sadržaj GMO >0,9%GMO content >0,9%
Obeležavanje uzorkaLabelling of sample
KvantitativanPCR test
QuantitativePCR test
KvantitativanPCR test
QuantitativePCR test

348
Semenarstvo
Veoma važnu ulogu u procesu detekcije GMO ima uzorkovanje se-mena, sirovina ili poluproizvoda u toku procesa proizvodnje. Reprezen-tativnost uzorka se mora očuvati u daljem postupku redukcije uzorka u laboratoriji i pripremi radnog uzoraka. Pravilno uzorkovanje određuje pouzdanost ili reprezentativnost dobijenih rezultata. U cilju ostvarenja reprezentativnosti uzorka, ako se zahteva nizak nivo genetičke modifiko-vanosti uzorka, veličina uzetog uzorka bi trebalo da raste (Gilbert, 1999).
Izolacija DNK
Za izolovanje DNK iz biljnog materijala i hrane biljnog porekla za GMO analizu se primenjuje veliki boj metoda (Anklam i sar., 2002). Njihova pogodnost se ocenjuje na osnovu prinosa, integritet i kvalite-ta izolovane DNK. Predpostavljajući da je laboratorijski uzorak tipičan poljski uzorak i da je adekvatno homogenizovan, za ekstrakciju DNK je obično dovoljno između 100 i 350 mg uzorka.
Postoje tri različita pristupa za izolovanje DNK: CTAB metod, ko-mercijalno dostupni kitovi koji koriste silika kolone za vezivanje DNK, i kombinacija ove dve metode. CTAB metoda je primenljiva za izolovanje DNK iz različitog biljnog materijala i hrane biljnog porekla, posebno zbog dobrog odvajanja polisaharida od DNK. Razvijene su modifikova-ne verzije CTAB metode kako bi se poboljšao prinos i kvalitet izolovane DNK iz specifičnih sastojaka ili hrane. Prilikom izolacije DNK iz skroba uveden je tretman sa enzimom α-amilazom, a kod izolacije iz lecitina upotrebljava se heksan koji uklanjanje lipide (Meyer i sar., 2001, Wurtz i sar., 1998). CTAB metod se može koristiti za ekstrakciju i prečišćavanje DNK iz semena i lista soje (Nikolić i sar., 2009).
Od kvaliteta i čistoće izolovane DNK zavisi uspešnost PCR reakcije. Kvalitet DNK je određen dužinom fragmenata i stepenom oštećenja pri-likom izlaganja zagrevanju, niskom pH, i/ili nukleazama koji uzrokuju hi-drolizu ili degradaciju. Kvalitet DNK varira u zavisnosti od vrste materijala koji se ispituje, stepena obrade uzorka i primenjenog metoda za ekstrak-ciju DNK. DNK koji je izolovan iz namirnica i hrane niskog je kvaliteta, sa prilično kratkim ciljnim sekvencama, u opsegu 100–400 bp. Na čistoću izolovane DNK utiču različite supstance: polisaharidi, lipidi i polifenoli ili hemikalije korišćene u procesu DNK ekstrakcije. Ključni enzim PCR-a, Taq polimeraza, može biti inhibiran od strane polisaharida, EDTA, fenola, SDS-a i mnogih drugih jedinjenja, što vodi lažno negativnim rezultatima analize.

349
Genetički modifikovane biljne vrste - metode za testiranje
PCR metode
PCR metode za testiranje genetički modifikovanih organizama su dostupne u različitim formatima: dupleks, nested PCR; PCR-ELISA, QC-PCR, real time PCR (Anklam i sar., 2002). Imaju široku primenu kako u procesu stvaranja, tako i testiranja genetički modifikovanih or-ganizama.
Metode za GMO detekciju zasnovane na PCR-u se mogu podeliti u četiri nivoa specifičnosti (Holst-Jensen i sar., 2003). Najmanje specifičan je uobičajeno nazvan „screening metod” kojim se detektuju DNK ele-menati, promotor i terminator, koji su prisutni u većini različitih GMO (sl. 4).
Postoji nekoliko DNK elemenata koji su prisutni u većem broj gene-tički modifikovanih biljnih vrsta, na primer: P-35S promoter virus mo-zaika karfiola (CaMV), 5-enol-piruvilšikimat-3-fosfat sintaza (EPSPS) gen, koji kodira CryIA(b) toksin iz Bacillus thuringiensis, pat/bar geni fosfinotrihin acetiltransferaze iz Streptomyces spp., nos5’ terminator no-paline sintaze (Agrobacterium tumefaciens) i, nptII, neomicin fosfotran-sferaza gen, selektovani marker za transformaciju biljnih celija koje su otporne na kanamicin.
Kontrolne laboratorije obično rade analize na prisustvo ciljnih se-kvence koje su zastupljene u mnogim (ali ne i u svim) GM usevima na tržištu, npr. CaMV 35S promoter, nos5’ terminator, i nptII. Treba istaći
Slika 4. Kvalitativan PCR uzorka soje na prisustvo 35S promotora i gena za lektin. M, DNK marker; 1, blank, 2, negativna kontrola, 3-5 GMO standardi: 0.1%, 0.5% 1%,
6-10 uzorci soje Figure 4. Qualitative PCR detection of 35S promoter and lectin in soybean samples.
M, DNA marker, 1, blank, 2, negative control, 3-5 GMO standards: 0.1%, 0.5%, 1%, 6-10 soybean samples
10
metod” kojim se detektuju DNK elemenati, promotor i terminator, koji su prisutni u većini
različitih GMO (Slika 2).
Slika 2. Kvalitativan PCR uzorka soje na prisustvo 35S promotora i gena za lektin. M, DNK marker; 1, blank, 2, negativna kontrola, 3-5 GMO standardi: 0.1%, 0.5% 1%, 6-10 uzorci soje
Postoji nekoliko DNK elemenata koji su prisutni u većem broj genetički
modifikovanih biljnih vrsta, na primer: P-35S promoter virus mozaika karfiola (CaMV), 5-
enol-piruvilšikimat-3-fosfat sintaza (EPSPS) gen, koji kodira CryIA(b) toksin iz Bacillus
thuringiensis, pat/bar geni fosfinotrihin acetiltransferaze iz Streptomyces spp., nos5’
terminator nopaline sintaze (Agrobacterium tumefaciens) i, nptII, neomicin fosfotransferaza
gen, selektovani marker za transformaciju biljnih celija koje su otporne na kanamicin.
Kontrolne laboratorije obično rade analize na prisustvo ciljnih sekvence koje su
zastupljene u mnogim (ali ne i u svim) GM usevima na tržištu, npr. CaMV 35S promoter,
nos5’ terminator, i nptII. Treba istaći da su ovi elementi mogu prirodno biti prisutni u
biljkama koje su ili zaražene virusom mozaika karfiola (Cauliflower Mosaic Virus, CaMV) ili
bakterijom Agrobacterium tumefaciens, tako da ovu činjenicu ne treba zanemariti prilikom
analize dobijenih rezultata. Obzirom da je analiza zasnovana na utvrđivanju prisustva DNK
sekvence koja postoji u prirodi povećan je rizik od lažno pozitivnih rezultata testa.
Ispitivanje na navedene genetičke elemente nije dovoljno, već je za pouzdanu analizu
genetički modifikovanih organizama neophodno dokazati da nisu prisutni i drugi specifični
geni: nptII, nos, EPSPS, pat/bar, CryA(b) i sl..
U tabeli 3. su dati primeri prisustva pojedinih genetičkih elemenata u različitim
genetički modifikovanim biljnim vrstama.
Drugi nivo specifičnosti je metod za identifikaciju specifičnog gena, koji detektuje deo
aktivnog gena i njegovog okruženja, na primer Bt gen za toksin koji deluje protiv insekata ili
EPSPS gen koji kodira protein za toleranciju na specifične herbicide. Ovom metodom
identifikujemo koja je modifikacija prisutna. Ukoliko se radi o autorizovanim genetičkim
modifikacijama isti gen može biti prisutan u nekoliko nezavisnih transformacija.
1500
850
400
200
M 1 2 3 4 5 6 7 8 9 10
1500
850
400
200
M 1 2 3 4 5 6 7 8 9 10 11
195 bp118 bp

350
Semenarstvo
Tabela 3. Primenljivost potencijalnih metoda za detekciju GMO zasnovanih na AgbiosTable 3. Applicability of methods for detection of GMO based on Agbios database
(www.agbios.com)
Biljna vrstaPlant species
Komercijalni naziv
Commercial name
ProizvođačCompany
Prisutan35S
promotorPresence
of 35S promoter
Prisutan nos5 ter-minatorPresence of nos5
termina-tor
Prisutan npt II genPresence of npt II
gene
KukuruzMaize
Maximizer Bt-176
Syngenta Seeds + - -
Bt 11 Syngenta Seeds + + -
Liberty Link T25 Aventis CropScience + - -
StarLink CBH-351 Aventis + + -
Roundup ReadyGA21 Monsanto - + -
Roundup ReadyNK603 Monsanto + + -
Yieldgard Mon 810
Monsanto + - +
SojaSoybean
Roundup ReadyGTS 40-3-2 Monsanto + + -
Uljana repicaCanola
Roundup Ready Monsanto - - -
MS8xRF3 Aventis - + +
LibertyLink InovatorHCN92
Aventis CropScience + - +
ParadajzTomato Flavr Savr Calgene + - +
KrompirPotato NewLeaf Monsanto + + +

351
Genetički modifikovane biljne vrste - metode za testiranje
da su ovi elementi mogu prirodno biti prisutni u biljkama koje su ili za-ražene virusom mozaika karfiola (Cauliflower Mosaic Virus, CaMV) ili bakterijom Agrobacterium tumefaciens, tako da ovu činjenicu ne treba zanemariti prilikom analize dobijenih rezultata. S obzirom da je analiza zasnovana na utvrđivanju prisustva DNK sekvence koja postoji u priro-di povećan je rizik od lažno pozitivnih rezultata testa.
Ispitivanje na navedene genetičke elemente nije dovoljno, već je za pouzdanu analizu genetički modifikovanih organizama neophodno do-kazati da nisu prisutni i drugi specifični geni: nptII, nos, EPSPS, pat/bar, CryA(b) i sl.
U tabeli 3 su dati primeri prisustva pojedinih genetičkih elemenata u različitim genetički modifikovanim biljnim vrstama.
Drugi nivo specifičnosti je metod za identifikaciju specifičnog gena, koji detektuje deo aktivnog gena i njegovog okruženja, na primer Bt gen za toksin koji deluje protiv insekata ili EPSPS gen koji kodira protein za toleranciju na specifične herbicide. Ovom metodom identifikujemo koja je modifikacija prisutna. Ukoliko se radi o autorizovanim genetič-kim modifikacijama isti gen može biti prisutan u nekoliko nezavisnih transformacija.
Treći nivo specifičnosti je konstrukt specifičan metod, koji detek-tuje spoj dva DNK elementa, kao što su promotor i funkcionalan gen. U prirodi ovaj spoj nije prisutan, ali različiti GMO mogu imati neko-liko elemenata sa istim promotorom i genom, ili je isti plazmid kori-šćen u procesu stvaranja GMO (npr. dve različite modifikacije Mon809 i Mon810) (Agbios, 2003). Na slici 5 se može videti identifikacija dve genetičke modifikacije kukuruza T25 i TC 1507 konstrukt specifičnom PCR metodom.
11
Tabela 3. Primenljivost potencijalnih metoda za detekciju GMO zasnovanih na Agbios
(www.agbios.com)
Biljna vrsta
Komercijalni naziv Kompanija Prisutan 35S promotor
Prisutan nos5 terminator
Prisutan nptII gen
kukuruz Maximizer Bt-176 Syngenta Seeds
+ - -kukuruz Bt 11 Syngenta
Seeds + + -kukuruz Liberty Link T25 Aventis Crop
Science + - -kukuruz StarLink CBH-351 Aventis + + -kukuruz Roundup Ready
GA21Monsanto - + -
kukuruz Roundup Ready NK603 Monsanto + + -
kukuruz Yieldgard Mon 810 Monsanto + - +soja Roundup Ready
GTS 40-3-2Monsanto + + -
uljana repica
Roundup Ready Monsanto - - -uljana repica
MS8xRF3 Aventis - + +
uljana repica
LibertyLink Inovator HCN92
Aventis Crop Science + - +
paradajz Flavr Savr Calgene + - +krompir NewLeaf Monsanto + + +
Treći nivo specifičnosti je konstrukt specifičan metod, koji detektuje spoj dva DNK
elementa, kao što su promotor i funkcionalan gen. U prirodi ovaj spoj nije prisutan, ali
različiti GMO mogu imati nekoliko elemenata sa istim promotorom i genom, ili je isti
plazmid korišćen u procesu stvaranja GMO (npr. dve različite modifikacije Mon809 i
Mon810) (Agbios, 2003). Na slici 3 se može videti identifikacija dve genetičke modifikacije
kukuruza T25 i TC 1507 konstrukt specifičnom PCR metodom.
SLIKA 3 Konstrukt specifičan PCR za T25 i TC 1507 kukuruza (Nikolić, 2009)
Najveća specifičnost se postiže kada se detektuje jedinstveni region između
insertovane DNK i genoma domaćina, nazvan “event” specifičan metod (slika 4). Međutim, i
ovaj metod ima svoja ograničenja. Kao rezultat ukrštanja dve GMO linije u rekombinantnoj
liniji će biti prisutno više gena koji regulišu npr. kombinacija otpornosti na herbicide sa
svojstvom tolerantnost na insekte. Oba seta funkcionalnih gena su prisutna u rekombinovanoj
T25
1
TC1507
Slika 5. Konstrukt specifičan PCR za T25 i TC 1507 kukuruza / Figure 5. Construct specific PCR for the dectection of T25 and TC 1507 GMO maize (Nikolic et al., 2009)

352
Semenarstvo
Najveća specifičnost se postiže kada se detektuje jedinstveni region između insertovane DNK i genoma domaćina, nazvan “event” specifi- čan metod (sl. 4). Međutim, i ovaj metod ima svoja ograničenja. Kao rezultat ukrštanja dve GMO linije u rekombinantnoj liniji će biti pri- sutno više gena koji regulišu npr. kombinacija otpornosti na herbicide sa svojstvom tolerantnost na insekte. Oba seta funkcionalnih gena su prisutna u rekombinovanoj liniji, ali ne moraju biti vezani, mogu biti na različitim hromozomima. Otuda postoji mogućnost da će se na ovaj način detektovati samo jedna modifikacija.
Multipleks PCR
Multipleks PCR se izvodi sa više setova prajmera koji simultano umnožavaju različite sekvence. Prisustvo većeg broja prajmera u jednoj reakciji može uzrokovati mnoge probleme, kao što su povećanje formi- ranja pogrešnih PCR proizvoda, prajmer-dimera i amplifikaciju dugih fragmenata (Atlas et al., 1994).
Mogućnost korišćenja nekoliko prajmera, svaki sa određenom specifičnošću, u jednoj reakciji, daje multidimenzionalnu perspektivu dijagnostičkom potencijalu PCR. Ova metoda omogućava simultanu detekciju dve ili više ciljnih sekvenci. Kako se praktično u istoj reakci- onoj zapremini odvija više reakcija umnožavanja, multiplex PCR zah- teva intenzivnu optimizaciju u pogledu koncentracija reakcionih kom- ponenti i programa umnožavanja. Svaka od paralelnih amplifikacija će
Slika 6. Specifična detekcija T45 GMO uljane repice, pozitivni uzorci 11-19 Figure 6. Specific detection of T 45 GMO rapeseed, positive samples 11-19
(Nikolić et al., 2009)
12
liniji, ali ne moraju biti vezani, mogu biti na različitim hromozomima. Otuda postoji
mogućnost da će se na ovaj način detektovati samo jedna modifikacija.
Slika 4. Specifična detekcija T45 GMO uljane repice, pozitivni uzorci 11-19 (Nikolić, 2009)
6.2.1 Multipleks PCR
Multipleks PCR se izvodi sa više setova prajmera koji simultano umnožavaju različite
sekvence. Prisustvo većeg broja prajmera u jednoj reakciji može uzrokovati mnoge probleme,
kao što su povećanje formiranja pogrešnih PCR proizvoda, prajmer-dimera i amplifikaciju
dugih fragmenata (Atlas i sar., 1994).
Mogućnost korišćenja nekoliko prajmera, svaki sa određenom specifičnošću, u jednoj
reakciji, daje multidimenzionalnu perspektivu dijagnostičkom potencijalu PCR. Ova metoda
omogućava simultanu detekciju dve ili više ciljnih sekvenci. Kako se praktično u istoj
reakcionoj zapremini odvija više reakcija umnožavanja, multiplex PCR zahteva intenzivnu
optimizaciju u pogledu koncentracija reakcionih komponenti i programa umnožavanja. Svaka
od paralelnih amplifikacija će imati svoju kinetiku i efikasnost, što rezultira u različitoj
specifičnosti i osetljivosti za svaku ciljnu sekvencu.
Slika 5. Multipleks PCR, simultano umnožavanje gena za zein (277bp), 35S promotor
(195bp) i NOS terminator (118bp) u GMO kukuruzu (Nikolić i sar., 2008)
Multipleks PCR se često koristi u dijagnostici genetički modifikovanih biljaka jer redukuje
vreme potrebno za analizu kao i cenu analize (Nikolić i sar., 2008, 2009; Mafra i sar., 2008).
6.2.2 Nested PCR
277 bp195 bp118 bp
(bp) M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
233bp

353
Genetički modifikovane biljne vrste - metode za testiranje
Slika 7. Multipleks PCR, simultano umnožavanje gena za zein (277bp), 35S promotor (195bp) i NOS terminator (118bp) u GMO kukuruzu
Figure 7. Multiplex PCR, simultaneous amplification of zein (277bp), 35S promoter (195bp) and NOS terminator (118bp) in maize GMO (Nikolic et al., 2008)
12
liniji, ali ne moraju biti vezani, mogu biti na različitim hromozomima. Otuda postoji
mogućnost da će se na ovaj način detektovati samo jedna modifikacija.
Slika 4. Specifična detekcija T45 GMO uljane repice, pozitivni uzorci 11-19 (Nikolić, 2009)
6.2.1 Multipleks PCR
Multipleks PCR se izvodi sa više setova prajmera koji simultano umnožavaju različite
sekvence. Prisustvo većeg broja prajmera u jednoj reakciji može uzrokovati mnoge probleme,
kao što su povećanje formiranja pogrešnih PCR proizvoda, prajmer-dimera i amplifikaciju
dugih fragmenata (Atlas i sar., 1994).
Mogućnost korišćenja nekoliko prajmera, svaki sa određenom specifičnošću, u jednoj
reakciji, daje multidimenzionalnu perspektivu dijagnostičkom potencijalu PCR. Ova metoda
omogućava simultanu detekciju dve ili više ciljnih sekvenci. Kako se praktično u istoj
reakcionoj zapremini odvija više reakcija umnožavanja, multiplex PCR zahteva intenzivnu
optimizaciju u pogledu koncentracija reakcionih komponenti i programa umnožavanja. Svaka
od paralelnih amplifikacija će imati svoju kinetiku i efikasnost, što rezultira u različitoj
specifičnosti i osetljivosti za svaku ciljnu sekvencu.
Slika 5. Multipleks PCR, simultano umnožavanje gena za zein (277bp), 35S promotor
(195bp) i NOS terminator (118bp) u GMO kukuruzu (Nikolić i sar., 2008)
Multipleks PCR se često koristi u dijagnostici genetički modifikovanih biljaka jer redukuje
vreme potrebno za analizu kao i cenu analize (Nikolić i sar., 2008, 2009; Mafra i sar., 2008).
6.2.2 Nested PCR
277 bp195 bp118 bp
(bp) M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
233bp
imati svoju kinetiku i efikasnost, što rezultira u različitoj specifičnosti i osetljivosti za svaku ciljnu sekvencu.
Multipleks PCR se često koristi u dijagnostici genetički modifiko- vanih biljaka jer redukuje vreme potrebno za analizu kao i cenu analize (Nikolić et al., 2008, 2009; Mafra et al., 2008).
Nested PCR
U slučajevima kada je ciljna DNK prisutna u nekoliko kopija jednosta-van način za značajno poboljšanje prinosa reakcije je nested PCR (Newton i Graham, 1994). Metod se sastoji iz dve sukcesivne PCR reakcije u kojima proizvod iz prve amplifikacije služi kao matrica za drugu amplifikaciju.
13
U slučajevima kada je ciljna DNK prisutna u nekoliko kopija jednostavan način za značajno
poboljšanje prinosa reakcije je nested PCR (Newton i Graham, 1994). Metod se sastoji iz dve
sukcesivne PCR reakcije u kojima proizvod iz prve amplifikacije služi kao matrica za drugu
amplifikaciju.
Nested i seminested PCR su često veoma efikasni u redukciji ili eliminaciji neželjenih
proizvoda a u isto vreme povećava senzitivnost reakcije.
Slika 6. Nested PCR uzoraka soje na prisustvo EPSPS gena M, DNK marker; 1, blank, 2, negativna kontrola, 3-5 GMO standardi: 0.1%, 0.5% 1%, 6-16 uzorci soje (Nikolić i sar.,2009).
6.2.3 Real Time PCR
Real-time PCR metoda za određivanje sadržaja GMO se zasniva na umnožavanju
transgenih specifičnih sekvenci i endogenih referentnih gena koji su specifični za određenu
biljnu vrstu. Ovaj test je nastao zahvaljujući otkrićima da enzim Taq polimeraza ima u 5'-3'
smeru egzonukleaznu aktivnost i kreiranjem oligonukleotidne probe koja je obeležena sa dve
fluorescentne boje. U toku PCR reakcije Taq polimeraza, seče internu probu i boja reporter
emituje fluorescenciju koju registruje aparat. Kada je proba intaktna fluorescenciju koju
emituje boja reporter apsorbuje tzv.quencher- druga boja. Nivo emisije fluorescencije je
proporcionalan količini transgena u uzorku (šema 3).
Šema 3. Princip »Real Time« PCR testa
(bp) M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
1500850
400200
5050
169bp
E
prajmer proba
E
prajmer
Slika 8. Nested PCR uzoraka soje na prisustvo EPSPS gena M, DNK marker; 1, blank, 2, negativna kontrola, 3-5 GMO standardi: 0.1%, 0.5% 1%, 6-16 uzorci soje
Figure 8. Detection of EPSPS gen in soybean samples by nested PCR. M, DNA marker, 1, blank, 2, negative control, 3-5 GMO standards: 0.1%, 0.5%, 1%, 6-16 soybean samples
(Nikolic et al., 2009)

354
Semenarstvo
Nested i seminested PCR su često veoma efikasni u redukciji ili elimi-naciji neželjenih proizvoda a u isto vreme povećava senzitivnost reakcije.
Real Time PCR
Real-time PCR metoda za određivanje sadržaja GMO se zasniva na umnožavanju transgenih specifičnih sekvenci i endogenih referentnih gena koji su specifični za određenu biljnu vrstu. Ovaj test je nastao za- hvaljujući otkrićima da enzim Taq polimeraza ima u 5’-3’ smeru egzo- nukleaznu aktivnost i kreiranjem oligonukleotidne probe koja je obe- ležena sa dve fluorescentne boje. U toku PCR reakcije Taq polimeraza, seče internu probu i boja reporter emituje fluorescenciju koju registruje aparat. Kada je proba intaktna fluorescenciju koju emituje boja reporter apsorbuje tzv. quencher – druga boja. Nivo emisije fluorescencije je pro- porcionalan količini transgena u uzorku (sl. 9).
Slika 9. Princip „Real Time” PCR testaFigure 9. Real Time PCR principe (RT-PCR) (Nikolic et al., 2004)
Ukoliko je koncentracija GMO DNK visoka aparat će registrovati fluo-rescenciju posle manjeg broja ciklusa umnožavanja. Svaka analiza sadržaja GMO u uzorku uključuje i seriju standarda, tako se procenat genetske mo-difikacije uzorka određuje u odnosu na standardnu krivu (sl. 10).
Ukoliko je GMO hibrid nastao ukrštanjem dve roditeljske GMO li- nije, kvantitativno se određuje sadržaj pojedinačnih roditeljskih gena u uzorku semena. U SAD, ako su oba roditelja autorizovana, ovaj tip GMO hibrida je dozvoljen, dok je u EU potrebna posebna autorizacija za hibrid. Real Time PCR se koristi i za utvrđivanje sadržaja GMO hra- ne koja u svom sastavu ima komponente biljnog porekla (Taški-Ajdu-ković et al., 2009).
Prajmer / Primer
Prajmer / Primer Proba / Probe

355
Genetički modifikovane biljne vrste - metode za testiranje
Razvoj metoda u budućnosti
Metode koje se koriste za testiranje genetički modifikovanih biljnih vrsta i hrane u potpunosti ne zadovoljavaju potrebe korisnika. Očekuje se da će u budućnosti razvoj metoda za testiranje genetički modifikova- nih organizama voditi ka smanjenju cene analize i dobijanju rezultata za kraće vreme. Verovatno će većina metoda u isto vreme identifikovati različite genetičke elemente i gene, ali i omogućiti kvantifikaciju istih (Holst Jensen 2009).
Prisustvo neautorizovanih genetičkih modifikacija, ali koje su oce- njene kao sigurne u nekim zemljama će biti prihvaćene u većini drugih zemalja kako bi se prevazišao problem u svetskoj trgovini.
Nove tehnologije transformacije, odnosno stvaranja genetički mo- difikovanih organizama će voditi ka novim metodama za detekciju, koje uključuju mikročipove, elektohemijske senzore i masenu spektrofoto- metriju.
Međunarodna harmonizacija i razmena informacija u oblasti stva- ranja GMO, eksperimentalnih poljskih ogleda, DNK sekvenci među kompetentnim zemljama i njihovim laboratorijama, će pozitivno uticati na svetsku trgovinu. Razvoj regulative u oblasti biotehnologije i zahtevi za autorizaciju i obeležavanje će možda rezultirati i u menjanju priorite- ta i stvaranju novih izazova za laboratorije za GMO testiranje.
Slika 10. Kvantitativno određivanje GMO u uzorku Figure 10. Quantification of the GMO content in the sample (Nikolic et al., 2004)
14
Ukoliko je koncentracija GMO DNK visoka aparat će registrovati fluorescenciju posle
manjeg broja ciklusa umnožavanja. Svaka analiza sadržaja GMO u uzorku uključuje i seriju
standarda, tako se procenat genetske modifikacije uzorka određuje u odnosu na standardnu
krivu (šema 4).
Šema 4. Kvantitativno određivanje GMO u uzorku
Ukoliko je GMO hibrid nastao ukrštanjem dve roditeljske GMO linije, kvantitativno
se određuje sadržaj pojedinačnih roditeljskih gena u uzorku semena. U USA, ako su oba
roditelja autorizovana, ovaj tip GMO hibrida je dozvoljen, dok je u EU potrebna posebna
autorizacija za hibrid. Real Time PCR se koristi i za utvrđivanje sadržaja GMO hrane koja u
svom sastavu ima komponente biljnog porekla (Taški i sar., 2009).
7. Razvoj metoda u budućnosti
Metode koje se koriste za testiranje genetički modifikovanih biljnih vrsta i hrane u
potpunosti ne zadovoljavaju potrebe korisnika. Očekuje se da će u budućnosti razvoj metoda
za testiranje genetički modifikovanih organizama voditi ka smanjenju cene analize i dobijanju
rezultata za kraće vreme. Verovatno će većina metoda u isto vreme identifikovati različite
genetičke elemente i gene, ali i omogućiti kvantifikaciju istih (Holst Jensen, 2009).
Prisustvo neautorizovanih genetičkih modifikacija, ali koje su ocenjene kao sigurne u
nekim zemljama će biti prihvaćene u većini drugih zemalja kako bi se prevazišao problem u
svetskoj trgovini.
% GMO u uzorku
Broj ciklusa
% GMO
Bro
j cik
lusa
PCR
pro
izvo
d
14
Ukoliko je koncentracija GMO DNK visoka aparat će registrovati fluorescenciju posle
manjeg broja ciklusa umnožavanja. Svaka analiza sadržaja GMO u uzorku uključuje i seriju
standarda, tako se procenat genetske modifikacije uzorka određuje u odnosu na standardnu
krivu (šema 4).
Šema 4. Kvantitativno određivanje GMO u uzorku
Ukoliko je GMO hibrid nastao ukrštanjem dve roditeljske GMO linije, kvantitativno
se određuje sadržaj pojedinačnih roditeljskih gena u uzorku semena. U USA, ako su oba
roditelja autorizovana, ovaj tip GMO hibrida je dozvoljen, dok je u EU potrebna posebna
autorizacija za hibrid. Real Time PCR se koristi i za utvrđivanje sadržaja GMO hrane koja u
svom sastavu ima komponente biljnog porekla (Taški i sar., 2009).
7. Razvoj metoda u budućnosti
Metode koje se koriste za testiranje genetički modifikovanih biljnih vrsta i hrane u
potpunosti ne zadovoljavaju potrebe korisnika. Očekuje se da će u budućnosti razvoj metoda
za testiranje genetički modifikovanih organizama voditi ka smanjenju cene analize i dobijanju
rezultata za kraće vreme. Verovatno će većina metoda u isto vreme identifikovati različite
genetičke elemente i gene, ali i omogućiti kvantifikaciju istih (Holst Jensen, 2009).
Prisustvo neautorizovanih genetičkih modifikacija, ali koje su ocenjene kao sigurne u
nekim zemljama će biti prihvaćene u većini drugih zemalja kako bi se prevazišao problem u
svetskoj trgovini.
% GMO u uzorku
Broj ciklusa
% GMO
Bro
j cik
lusa
PCR
pro
izvo
d

356
Semenarstvo
LITERATURA
Agbios: Biotch Crops Database. http://www.agbios.com. AGBIOS, Merrickville, Ontario, Canada.
Anklam E, Neumann D (2002): Method development in relation to regulatory requirements for detection of GMOs in the food chain. J. AOAC Int. 85(3):754-6
Anklam E, Gadani F, Heinze P, Pijnenburg H, Van Den Eede G (2001): Analytical methods for detection and determination of genetically modified organisms in agricultural crops and plant-derived food products. Eur. Food Res. Technol. 214: 3-26
Anklam E, Heinze P, Kay S, Van den Ede G (2002): ValidationStudies and proficiency testing. J. AOAC Int. 85 (3): 809-815Atlas B, Bey A K (1994): Polymerase chain reaction. In: Gerhardt P,
Murrey R G E, Wood W A, Krieg N R (Eds.) Methods for general and molecular bacteriology. Washington, DC: American Society for Microbiology, 418-435
Berdal K G, Holst-Jensen A (2001): Roundup Ready® soybeanevent-specific real-time quantitative PCR assay and estimation of the
practical detection and quantification limits in GMO analyses. Eur. Food Res. Technol. 213: 432-438
Corbisier P, Trapmann S, Gancberg D, Van Iwaarden P, Hannes L, Catalani P, Le Guern L, Schimmel H (2002): Effect of DNA fragmentation on the quantitation of GMO. Eur. J. Biochem. 269 (1): 40
Gilbert J (1999): Sampling of row materials and proceesed food for the presence of GMOs. Food Control 10: 363-365
Hemmer W (1997): Foods derived from genetically modified organism and detection methods. BATS-Report 2/1997. Agency
for Biosafety Research and Assesment of Technology Impacts of the Swiss Priority Programe Biotechnology of the Swiss National Science Foundation, Basel, Switzerland
Holst-Jensen A, Rønning S B, Løvseth A, Berdal K G (2003):

357
Genetički modifikovane biljne vrste - metode za testiranje
PCR technology for screening and quantification of genetically modified organisms (GMOs). Anal. Bioanal. Chem. 375: 985-993
Holst-Jensen A (2009): Testing for genetically modified organisms (GMOs): Past, future perspectives. Biotechnol. Adv. 27: 1071-1082
James C, Global status of commercialized biotech/GM crops (2010): Brief 42 International Service for the Acquisition of Agri-Biotech Applications (ISAAA)
Kuribara H, Shindo Y, Matsuoka T, Takubo K, Futo S, Aoki N (2002): Novel reference molecules for quantitation of genetically modified maize and soybean, J. AOAC Int. 85:1077-1089
Lin H Y, Chiang J W, Shin D Y (2002): Detection of genetically modified soybeans by PCR method and immunoassay kits. J. Food Drug Anal. 9: 160-166
Mafra I, Ferreira I M P L V O, Oliveira M B P P (2008): Food authentication by PCR-based methods. Eur. Food Res. Technol. 227 (3): 649-665
Mattarucchi E, Weighardt F, Barbati C, Querci M, Van de Eede, G (2005): Development and applications of real-time PCR standards for GMO quantification based in tandem-marker plasmids. Eur. Food Res. Technol. 221: 511-519
Milošević M, Vujaković M, Nikolić Z, Ignjatov M (2007): Kontrola kvaliteta semenskog materijala. Nauka osnova održivog razvoja, Društvo genetičara Srbije, Beograd, 95-124
Milošević M, Nikolić Z, Vujaković M, Taški-Ajduković K, Ignjatov M, Petrović D (2008): Genetically modified crops – general view. The Second Joint PSU-UNS International Conference on BioScience: Food, Agriculture and Environment, Jun 22-24, 2008, Novi Sad, Serbia, 12-17
Meyer K, Rosa C, Hischenhuber C, Meyer R (2001): Determination of locust bean gum and guar gum by polymerase chain reaction and restriction fragment length polymorphism analysis. J. AOAC Int. 84: 89-99
Newton C R, Graham A (1994): PCR, BIOS Scientific Publishers, Limited, Oxford
Nikolić Z, Taški-Ajduković K, Tatić M, Balešević-Tubić S (2009): Monitoring of the Roundup Ready soybean in the Vojvodina province in Serbia. Ind. Crops 29: 638-641
Nikolić Z, Taški-Ajduković K, Jevtić A, Marinković D (2009): Detection

358
Semenarstvo
of GM Soybean in Food Products by Simultaneous Employment of Three Pairs of PCR Primers. Food Res. Int. 42: 349-352
Nikolić Z, Milošević M, Vujaković M, Marinković D, Jevtić A, Balešević-Tubić S (2008): Qualitative triplex PCR for the detection of genetically modified soybean and maize. Biotechnol. Biotec. Eq. 22: 801-803
Nikolić Z, Milošević M, Vujaković M, Ignjatov M (2007): Genetski modifikovani organizmi u svetlu domaćih i svetskih propisa. Sel. Semen. 3-4: 59-63
ISO 21571Foodstuffs — Methods of analysis for the detection of genetically modified organisms and derived products - Nucleic acid extraction
ISO 21569 Foodstuffs — Methods of analysis for the detection of genetically modified organisms and derived products - Qualitative nucleic acid based methods
ISO 21570 Foodstuffs — Methods of analysis for the detection of genetically modified organisms and derived products - Quatitative nucleic acid based methods
Official Collection of Test Method (1998): Detection of a genetic modification of soybeans by amplification of the modified DNA sequence by means of the polymerase chain reaction (PCR) and hybridisation of the PCR product with a DNA probe. German ederalFoodstuffs Act - Food Analysis, article 35
Službeni glasnik RS (2009): Zakon o genetički modifikovanim organizmima, Službeni glasnik RS, broj 41/09.
Taški-Ajduković K, Nikolić Z, Vujaković M, Milošević M, Ignjatov M, Petrović D (2009): Detection of genetically modified organisms in processed meat products on the Serbian food market. Meat Sci. 81: 230-232
Taverniers I, Van Bockstaele E, De Loose M (2004): Cloned plasmid DNA fragments as calibrators for controlling GMOs: Different real-time duplex quantitative PCR methods. Anal. Bioanal. Chem. 378: 1198–1207
Trapmann S, Burst M, Broll H, Macarthur R,Wood R, Zel J (2008): Guidance document on measurement uncertainty for GMO testing laboratories, European Commission, Joint Resesach Centre.

Ankica Kondić-Špika, Borislav Kobiljski
Genotipska karakterizacija gajenih biljaka
– DNK Fingerprinting

360
Semenarstvo

361
Genotipska karakterizacija gajenih biljaka
UVOD
Dezoksiribonukleinska kiselina (DNK) predstavlja osnovnu gra-divnu jedinicu svih živih organizama. Specifični redosled nukleotida (adenin, guanin, timin i citozin) u sekvencama DNK određuje sintezu proteina i enzima, koji dalje utiču na pojavu različitih osobina koje ka-rakterišu svaki organizam. Kod biljaka to mogu biti oblik listova, boja cvetova, boja ploda, ili pak sinteza širokog spektra različitih hemijskih jedinjenja. DNK predstavlja genotip (genetski identitet) jednog organiz-ma, koji određuje fenotip (fizičke karakteristike) tog organizma.
Unutar živih sistema organizacija DNK je uniformna u čitavom or-ganizmu, nezavisno od organa, uslova sredine ili razvojnih faza. Speci-fičnost organizacije DNK zapravo nosi jedinstvenost svakog organizma. DNK profil (sl. 1a), slično kao barkod (sl. 1b), jedinstven je kao otisak prsta (fingerprint) i specifičan za svaku individuu, te može biti korišćen za identifikaciju individua sa apsolutnom sigurnošću, na isti način kao i konvencinalni otisci prstiju.
Slika 1. DNK profili (a), kao i barkodovi (b), jedinstveni su za svaku individuu, te se mogu koristiti za pouzdanu identifikaciju genotipova kod biljaka
Figure 1. DNA profiles (a), same as barcodes (b), are unique for each individual and could be used for certain identification of plant genotypes (Kondić-Špika et al. 2008)

362
Semenarstvo
ISTORIJAT
Termin DNK fingerprinting, u svom originalnom značenju, odnosi se na metod razvijen 1985. godine od strane Sir Alec Jeffreys-a i njego-vih saradnika, zasnovan na Southern blot analizi i RFLP (Restriction Fragment Lenght Polymorphism) tehnici. U istraživanju Jeffreys et al. (1985) proučavan je gen za mioglobin, protein koji skladišti kiseonik u mišićnim ćelijama. Tehnika je veoma brzo našla široku primenu, prvo u sudskoj medicini, a kasnije i u drugim oblastima.
U analizi biljnog genoma tehnika DNK fingerprintiga prvi put je primenjena u istraživanju Ryskov et al. (1988), u kojem su ispitiva-ne razlike između dve sorte ječma (Hordeum vulgare). U njihovom istraživanju korišćena je Southern blot hibridizacija sa M13 probom, na fragmentima DNK isečenim HaeIII restrikcionim enzimom. Istu probu koristili su i Rogstad et al. (1988) u generisanju DNK profila za brojne skrivenosemenice i golosemenice. U istraživanju, objavljenom iste godine, Dallas (1988) je primenio humanu 33.6 minisatelit probu u razdvajanju sorti pirinča. U 1989. godini sintetički oligonukleotidi koji prepoznaju repetitivne DNK sekvence (mikrosatelite) uvedeni su u DNK fingerprinting kod biljaka. Weising et al. (1989) su pokaza-li da je moguće dobiti polimorfne DNK profile kada se DNK ječma ili nauta iseče restrikcionim enzimima, fragmenti razdvoje elektro-forezom na agaroznom gelu, a zatim osušeni gelovi hibridizuju sa radioaktivno obeleženim (GACA ili GATA) probama. Nakon toga objavljen je veliki broj radova (Weising et al. 1991, 1992, Beyermann et al. 1992, Depeiges et al. 1995, Sharma et al. 1995) o ovom, tzv. oligonukleotidnom fingerprinting pristupu kod biljaka, ukazujući na to da nivo detektovanih razlika u velikoj meri zavisi od izabrane pro-be. Više detalja o DNK fingerprintingu biljaka sa mini- i mikrosatelit komplementarnim hibridizujućim probama može se naći u pregled-nim radovima Nybom (1991, 1993), Weising et al. (1991) i Weising and Kahl (1998).

363
Genotipska karakterizacija gajenih biljaka
TRI DNK FINGERPRINTING TEHNIKE
Dominantni markeri
Tokom godina, naučnici su razvili mnoge tehnike za DNK finger-printing, koje se razlikuju po kompleksnosti, troškovima i pouzdanosti. Najčešće se radi o varijacijama i kombinacijama tehnika zasnovanih na koriščenju restrikcionih enzima i lančane reakcije polimeraze (Polyme-rase Chain Reaction-PCR).
Korišćenjem RFLP tehnike i specifičnih proba-enzim kom-binacija moguće je dobiti veoma kvalitetne profile. Međutim, sa pojavom PCR tehnologije, nasta-le su brojne jednostavnije tehnike za molekularnu karakterizaciju. Dve metode koje koriste prajme-re proizvoljne sekvence objavljene su 1990. (Welsh and McClelland 1990, Williams et al. 1990), dok je treća objavljena 1991. godine (Ca-etano-Anollés et al. 1991a, 1991b). Pokazalo se da ovakvi prajmeri, koji generišu nepoznate ampliko-ne iz genomske DNK, rezultiraju polimorfnim prikazima produka-ta, nakon elektroforeze i bojenja. Iz ove grupe izdvajaju se RAPD (Random Amplified Polymorphic DNA) markeri, koje su razvili Williams et al. (1990), kao najpo-godniji za DNK profilisanje. Samo
Slika 2. AFLP profili 19 uzoraka Cynodon spp. Figure 2. AFLP profiles of 19 Cynodon spp.
samples (Wu et al., 2005)

364
Semenarstvo
nekoliko godina kasnije pojavljuje se novi metod, u vidu AFLP (Am-plified Fragment Lenght Polymorphism) markera, predstavljen u radu Zabeau and Vos (1993). Ovaj metod kombinuje elemente od obe pret-hodne grupe markera, RFLP i RAPD. Iako je tehnički mnogo zahtevnija od RAPD metode, primenom AFLP tehnike dobija se veliki broj poli-morfnih traka u DNK profilima (sl. 2).
Treću grupu tehnika za DNK profilisanje, zasnovanih na PCR-u, karakteriše to što usmeravaju PCR amplifikaciju na određeni tip (najče-šće repetitivne) DNK, bez potrebe da se razvijaju prajmeri koji su spe-cifični za svaku vrstu. Najbolji primer ovakvog pristupa jesu ISSR (Inter Simple Sequence Repeats) markeri. Većina navedenih tipova markera (RFLP, RAPD, AFLP) opisani su u četvrtom poglavlju ove knjige (Pri-mena genetskih markera u identifikaciji i određivanju genetske čistoće sorti i hibrida), te će u ovom poglavlju biti opisani samo ISSR markeri, kao i specifičnosti primene nabrojanih markera u DNK fingerprintingu.
ISSR markeri nastaju u PCR reakciji, u kojoj se kao prajmeri koriste jednostavne ponavljajuće sekvence (npr. [AC]n), da bi se amplifikovao region između dve ovakve ciljne sekvence (Kahl 2001). U ovoj tehnici, prema Zietkiewicz et al. (1994), prajmeri na bazi mikrosatelita koriste se da amplifikuju inter-SSR DNK sekvence, odnosno sekvence između dva mikrosatelitska lokusa. Kod ovih markera jedan prajmer ili kombinacija prajmera može da se sastoji od sekvence mikrosatelita kojoj je dodata jedna ili nekoliko drugačijih baza u regionu 3’, tako da amplifikuje regi-on između dva mikrosatelitska lokusa, sa oligonukleotidima vezanim za ponovljivu sekvencu. Mnogo složeniji način vazivanja prajmera može se postići korišćenjem prajmera sa dodatnim bazama na 5’ kraju, kod kojih je SSR region sastavni deo produkta amplifikacija, kao i kombinovanjem 3’ i 5’-prajmera (Zietkiewicz et al. 1994). ISSR markeri nisu uslovljeni poznavanjem sekvence oko SSR lokusa, te su široko primenjeni kod ra-znih biljnih vrsta (Wolfe and Liston 1998). To su uglavnom dominantni markeri, mada povremeno neki od njih ispolje i kodominantnost. Neo-graničen broj prajmera može se sintetisati za različite kombinacije di-, tri-, tetra-i penta-nukleotida, sa dodatkom od nekoliko baza i mogu se iskoristiti za širok spektar primene kod biljaka. ISSR analiza tehnički je jednostavnija od bilo kog drugog sistema markera, pruža visoko po-novljive rezultate i generiše viši nivo polimorfizma, u odnosu na druge markere, u mnogim živim sistemima. Preciznost ovih markera zasniva se na dugim prajmerima i striktno definisanoj temperaturi anilinga.

365
Genotipska karakterizacija gajenih biljaka
Zbog brzine kojom se može obraditi veliki broj uzoraka, metode zasnovane na PCR tehnici postale su veoma brzo popularne. AFLP, RAPD i ISSR markeri još uvek se dosta koriste, iako su RAPD markeri često kritikovani zbog problema sa reproducibilnošću i kompetitivnim vezivanjem prajmera. Ovi problemi manje su prisutni kod AFLP tehni-ke, koja se često smatra za odličan izbor u slučajevima kada je poželjno dobiti veliki broj polimorfnih traka. Sve tri metode obično daju slične rezultate, kada se primene na istom biljnom materijalu, u cilju proce-ne genetičkog diverziteta i genetičke udaljenosti genotipova. Međutim, detektovani lokusi uglavnom su bialelni (prisutan ili odsutan produkt), te su početni pokušaji da se razdvajanje hetero- od homozigota vrši na osnovu intenziteta dobijenih traka ubrzo napušteni. Zbog toga, ovi markeri se smatraju za dominantne markere, čime je značajno umanjen njihov potencijal za primenu u populacionoj genetici i detaljnijim gene-tičkim analizama.
Kodominantni markeri
SSR (Simple Sequence Repeats) markeri ili mikrosateliti zasnovani su na korišćenju PCR tehnike u kojoj se kao prajmeri koriste sekvence komplementarne regionima koji okružuju hipervarijabilne mikrosateli-te (Smeets et al. 1989, Tautz 1989, Weber and May 1989). Ova tehnika postala je veoma atraktivna u animalnoj genetici početkom 90-ih godi-na, da bi ubrzo privukla i botaničare da istražuju potencijal ovog pri-stupa za genetičke analize kod biljaka. Mikrosateliti su prvi put uspešno primenjeni kod biljaka 1992. godine (Akkaya et al. 1992). S obzirom na to da su mikrosateliti po svojim osobinama multialelni i kodominantni markeri, imaju veoma širok spektar primene u genetičkim istraživanji-ma kod biljaka, kao što su utvrđivanje razlika između genotipova, popu-laciona istraživanja, lociranje ekonomski važnih major gena i QTL-ova, kao i linkidž mapiranje (Li et al., 2002; Kobiljski i sar., 2005; 2009, Brba-klić i sar., 2009, 2010; Neumann et al., 2010; Trkulja i sar., 2011). Glav-ni nedostaci mikrosatelitskih markera jesu vreme i troškovi potrebni za razvijanje prajmera specifičnih za svaku vrstu (Squirrell et al. 2003). Međutim, neka istraživanja su pokazala da je moguć transfer mikrosa-telitskih markera između vrsta koje pripadaju istom rodu, ili ponekad čak i između različitih rodova (Arnold et al., 2002; Peakall et al., 1998).

366
Semenarstvo
Potreba da se smanje troškovi i poveća količina informacija po uzorku dovela je do razvoja nove generacije molekularnih tehnologija. SNP (Single Nucleotide Polymorphism) markeri povezani su sa mesti-ma na hromozomima, na kojima se DNK sekvence dva genotipa razli-kuju u samo jednoj bazi. Ove razlike nastaju kao posledica tranzicije (prelazak purinskih u purinske ili pirimidinskih u pirimidinske baze) ili transverzije (prelazak purinskih u pirimidinske baze ili obrnuto). Neke od prednosti SNP markera, na osnovu iskustava sa sekvenciranjem hu-manog genoma, su:
– SNP markeri su stabilniji od SSR, a otuda i pouzdaniji,– SNP su značajno brojniji od SSR markera (Kwok et al. 1996),– frekvencija pojavljivanja SNP je mnogo veća nego kod SSR mar-
kera (na svakih nekoliko desetina kbp),– SNP nemaju uniformnu distribuciju u genomu, – gustina SNP u genskim sekvencama je niska (Brayan et al., 1999),
ali mogu biti značajni izvori varijabilnosti za eksploataciju u ople-menjivačkim programima i
– pogodni su za tehnike koje ne koriste gelove, čime je omogućen značajno veći protok, odnosno broj uzoraka koje je moguće ana-lizirati u jedinici vremena.
Markeri za detekciju polimorfnosti organelarne DNK
Početkom devedestih godina prošlog veka razvijeni su i PCR praj-meri koji omogućavaju amplifikaciju citoplazmatske i mitohondrijalne DNK kod velikog broja biljnih vrsta (Taberlet et al., 1991; Demesure et al., 1995; Dumolin-Lapègue et al., 1997; Duminil et al. 2002). Zbog konzerviranosti mesta vezivanja unutar kodirajućih regiona, mnogi od ovih prajmera su univerzalni, odnosno mogu se koristiti kod različitih vrsta, rodova, pa čak i familija. Produkti amplifikovani sa ovakvim or-ganele-specifičnim prajmerima mogu biti sekvencionirani direktno, ili sečeni restrikcionim enzimima, ukoliko se koriste tzv. CAPS (Cleaved Amplified Polymorphic Sequences) markeri (Konieczny and Ausubel, 1993). Hloroplastni CAPS markeri postali su uobičajen metod koji se koristi za filogeografska istraživanja unutar biljnih vrsta.

367
Genotipska karakterizacija gajenih biljaka
Prvi univerzalni prajmeri za organelarnu DNK bili su usmereni na prethodno identifikovane gene, dok drugi pristup podrazumeva am-plifikaciju hloroplastnih mikrosatelita (Powell et al. 1995, Weising and Gardner 1999). Ovi mikrosateliti su manje varijabilni u pogledu broja ponavljanja u odnosu na njihove nuklearne analoge (Provan et al. 1999), ali ipak sadrže dovoljnu količinu varijabilnosti u okviru određene biljne vrste (Provan et al. 2001).
Tehnologije za brzu detekciju identiteta
U Kanadi je u poslednjih nekoliko godina intezivno rađeno na ra-zvoju brze i jeftine dijagnostičke platforme (Rapid Identity Detected Technology –RIDT) za DNK fingerprinting registrovanih sorti pšeni-ce (Procuiner et al. 2009). Obrada uzorka korišćenjem ove platforme postiže se fingerprintingom iz jednog semena, tako da se i uzorak koji se sastoji od mešavine različitih sorti može analizirati. RIDT platforma omogućava veliku brzinu, visoku propusnu moć, robotizovanu automa-tizaciju laboratorijskog rada i nisku cenu DNK fingerprintinga pšenice na bazi SNP tehnologije. Jeftina ekstrakcija DNK iz jednog semena, brza PCR amplifikacija i jednostavno skoriranje SNP rezultata najvažniji su preduslovi za određivanje DNK profila iz velikog broja pojedinačnih semena godišnje i to po veoma niskoj ceni. Troškovi su svakako domi-nantan problem u svakom programu fingerprintinga, dok su u ovom si-stemu ukupni troškovi za potrošni materijal po jednom markeru proce-njeni na manje od 0,60 USD po semenu. RIDT platforma takođe može razlikovati sorte koje su veoma slične, što je značajno, s obzirom na to da veći broj sorti može imati zajedničku genetičku osnovu. Rezultati „od semena do fingerprinting podataka” iz centralne laboratorije mogu se preneti elektronskim putem na bilo koju lokaciju, korišćenjem labo-ratorijskog sistema za upravljanje informacijama.
Izradu ove Platforme (RIDT) finansirao je konzorcijum sačinjen od vlade Kanade i velikih industrijskih partnera, koji su na neposredan ili posredan način vezani za proizvodnju i promet semena. Sigurno je da će se trend stvaranja brzih, jeftinih i pouzdanih DNK tehnologija u odre-đivanju identiteta sorti gajenih biljaka nastaviti i u budućnosti, kada će ovakve tehnologije imati sve značajniju ulogu i široku primenu.

368
Semenarstvo
DNK barkoding
DNK barkoding je relativno novi koncept koji je razvijen za do-bijanje brze, tačne i automatizovane identifikacije vrsta pomoću stan-dardizovanih kratkih DNK sekvenci (Vijayan and Tsou, 2010). U DNK barkodingu, specifični redosledi nukleotida u malim DNK fragmenti-ma (400–800 bp) koriste se kao posebne referentne kolekcije za identi-fikaciju uzoraka i za razlikovanje vrsta koje izgledaju veoma slično (sl. 3). Početni cilj DNK barkoding procesa jeste formiranje onlajn biblio-teka barkod sekvenci za sve poznate vrste, koje mogu da posluže kao standard, sa kojim će se porediti DNK barkodovi svih identifikovanih ili neidentifikovanih vrsta. U cilju promovisanja upotrebe DNK barko-dinga za sve eukariotske oblike života na našoj planeti, osnovan je Con-sortium for the Barcode of Life (CBOL) u maju 2004. godine, u koji je do 2010. godine bilo uključeno više od 120 organizacija iz 45 zemalja (Vijayan and Tsou, 2010).
Slika 3. Svaka od četiri baze DNK sekvence predstavljena je drugačijom bojom (a) Tip i redosled baza u DNK sekvenci određuje redosled boja u DNK barkodu (b)
Figure 3. Each of the four bases in a DNA sequence is represented by a different colour (a) (Source: http://www.cmarz.org/barcode.html).
The type and order of the DNA bases in a sequence determine the order of colours in the DNA barcode (b)
(Source: http://www.botanicgardens.ie/herb/research/maplebarcodes.htm)

369
Genotipska karakterizacija gajenih biljaka
Barkoding je dobro razrađena tehnika za identifikaciju vrsta kod životinja (sl. 4). Prvo istraživanje izveli su Hebert et al. (2003), koji su pokazali da se mogu identifikovati pojedine vrste iz kolekcije sačinjene od 200 vrsta leptira, sa 100% tačnosti, pomoću mitohondrijalnog gena za citohrom c oksidazu podjedinica I (COI). Međutim, kod biljaka, zbog poteškoća u pronalaženju prihvatljivog univerzalnog barkoda, to tek treba da bude razrađeno. Na osnovu relativne efikasnosti testira-nja, CBOL je nedavno identifikovao nekoliko lokusa kao potencijalnih kandidat barkodova, te je preporučen standardni barkod sa dva loku-sa (rbcL + matK) za identifikaciju biljnih vrsta. Moć razlikovanja vrsta ovog barkoda sa dva lokusa iznosi oko 70%, te je za bolju rezoluciju potrebno uvesti i dodatne lokuse.
Slika 4. DNK barkodovi različitih životinjskih vrsta Figure 4. DNA barcodes of different animal species
(Source: http://sitfu.com/2010/10/every-species-to-get-own-dna-barcode-international-barcode-of-life/)
DNK barkoding može pomoći u rešavanju nekih problema povezanih sa tradicionalnim taksonomskim identifikacijama vrsta na osnovu mor-foloških osobina, kao što je pogrešna identifikacija vrste usled fenotipske

370
Semenarstvo
plastičnosti i genotipske varijabilnosti svojstava, previđanje prikrivenih taksona itd (CBOL-Plant Working Group, 2009). DNK barkoding, da-kle, može da obezbedi onima koji se bave taksonomijom, konzervaci-jom i sl., isplativo i efikasno sredstvo, kao što je i barkod za identifikaciju proizvoda u supermarketu. U kontekstu brzog ekonomskog razvoja i posledica njegovog uticaja na floru i faunu širom naše planete, identi-fikacija vrsta pomoću brzih metoda od suštinskog je značaja za proce-nu biodiverziteta, u cilju očuvanja retkih endemskih i ugroženih vrsta. Ipak, treba imati u vidu da DNK barkoding nije alternativa taksonomiji i ne može da je zameni, ali je veoma koristan jer može poslužiti za dobi-janje dragocenih informacija o nepoznatim taksonima.
Poređenje različitih marker sistema i izbor tehnike za DNK fingerprinting
Brojna komparativna istraživanja su sprovedena u cilju poređenja različitih tehnika i utvrđivanja koja je najpogodnija i najpouzdanija za DNK fingrprinting i identifikaciju sorti (Jeffreys et al. 1991; Powell et al., 1996). Izbor DNK markera zavisi, između ostalih faktora, od obima i svrhe identifikacije sorti. Za testove koji se sastoje od samo nekoliko uzoraka analiziranih istovremeno, gde reproduktivnost i dokumento-vanje nisu posebno važni, prihvatljiv je bilo koji metod koji obezbeđu-je dovoljnu polimorfnost. Brojne studije u ovom pravcu sprovedene su pomoću multilokus metoda, kao što su AFLP, RAPD i ISSR markeri (Prevost and Wilkinson, 1999; Arnau et al., 2002; Bernet et al., 2004).
U istraživanju Prevost and Wilkinson (1999) analizirane su 34 oda-brane sorte krompira sa ISSR markerima. Produkti su razdvajani na sre-brom obojenim poliakrilamidnim gelovima i dobijeni rezultati bili su visoko ponovljivi. Svaki od dva prajmera visoke polimorfnosti bio je u stanju da razlikuje sva 34 genotipa. Autori su takođe predstavili novu meru za kvantifikaciju vrednosti svakog PCR- prajmera za proizvodnju multilokusnih profila u identifikaciji sorti. Ovu meru autori su nazvali Resolving power-Rp (moć razdvajanja) i ona omogućava ekstrapolaciju rezultata od malih eksperimenata do velikih studija. Rp uzima u ob-zir i broj polimorfnih traka (band) u uzorku i informativnu vrednost svake individualne polimorfne trake (Ib – Informative value of band). Ib je maksimalna kada je traka prisutna u polovini ispitivanih uzoraka,

371
Genotipska karakterizacija gajenih biljaka
a odsutna u drugoj polovini. Rp je jednaka zbiru Ib vrednosti za svaku traku u multilokusnom profilu.
Kada se identifikacija sorti vrši kod ekonomski značajanih biljnih vrsta, sa velikim brojem registrovanih sorti i ako je potrebno DNK pro-file za svaku sortu čuvati u bazi podataka, onda je ponovljivost i upo-redivost rezultata DNK fingerprintinga veoma važno pitanje. Trenutno, mikrosateliti su verovatno najbolji metod koji se može korsititi u ovakve svrhe, jer se kod njih javlja veliki broj polimorfnih alela. Primenljivost mikrosatelitskih lokusa za identifikaciju sorti intenzivno je istraživana (Russel et al., 1997; Bredemeijer et al., 1998; Becher et al., 2000; Kondić-Špika et al., 2010).
Mikrosateliti u velikoj meri ispunjavaju četiri kriterijuma koje je pr-vobitno postavio Bailey (1983):
– maksimalno su varijabilni između sorti, – minimalno su varijabilni unutar sorte, – stabilni su u različitim uslovima sredine i – obezbeđuju ponovljivost rezultata. Suviše visoka stopa mutacija mogla bi da učini beskorisnom bilu
koju identifikaciju sorti primenom markera. Poznato je da mnogi fak-tori utiču na stopu mutacija mikrosatelita, kao što su: dužina i sekvenca ponavljajućeg motiva, biologija organizma, hromozomska lokacija, kao i veličina alela (Jin et al., 1996). Stope mutacija kod biljnih, animalnih i humanih mikrosatelita variraju u širokom opsegu, a frekvencije mutaci-ja u dužini mikrosatelitskih alela veoma retko su izučavane kod biljaka (Udupa and Baum, 2001; Thuilet et al., 2002). Poželjno je, ukoliko je moguće, za identifikaciju sorti kod gajenih biljaka birati lokuse koji is-poljavaju relativno niske stope mutacija.
Iako su mikrosateliti u celini veoma korisni, oni takođe imaju odre-đene nedostatke, uključujući relativno visoku cenu razvoja markera, po-trebu za sofisticiranim laboratorijskim protokolima i opremom, proble-me sa ispravnim određivanjem veličine alela (Haberl and Tautz, 1999; Fernando et al., 2001) i povremenom pojavom lažnih proizvoda ampli-fikacije (Bredemeijer et al., 1998). Zbog svih ovih problema potrebno je uvek u istraživanje uključiti poznate standarde, jer npr. promena flu-orescentne boje prajmera, korišćenje različitih instrumenata za detek-ciju, ili čak različite temperature mogu uticati na veličinu generisanih fragmenata.

372
Semenarstvo
Izbor tehnike DNK fingerprintinga zavisi i od infrastrukture, teh-ničke obučenosti i raspoloživih sredstava, kao i od ciljeva samog istra-živanja. Međutim, kod izbora molekularne tehnike za genotipizaciju najznačajnija su dva kriterijuma, a to su sposobnost razdvajanja i pouz-danost. RAPD markeri imaju malu sposobnost razdvajanja, u poređenju sa AFLP i SSR markerima (tab. 1). Pored toga i ponovljivost rezultata dobijenih primenom RAPD markera često je problematična. SSR mar-keri smatraju se dobrim izborom zbog svoje brojnosti, visoke polimor-fnosti i kodominantnog nasleđivanja mikrosatelita. Kod biljnih vrsta gde su ovi markeri već definisani i opisani, primena SSR tehnike veoma je jednostavna. AFLP i SSR markeri ili mikrosateliti su među najviše korišćenim tehnikama, zbog visokog stepena razdvajanja genotipova i njihove pouzdanosti.
Tabela 1. Poređenje tehnika za DNK fingerprintingTable 1. Comparison of DNA fingerprinting techniques
Tehnika/Technique RFLP RAPD SSR AFLP PCR-seq
Troškovi razvojaDevelopment costs
SrednjiMedium
NiskiLow
VisokiHigh
NiskiLow
VisokiHigh
Nivo polimorfnostiPolymorphism level
Nizak-SrednjiLow-Medium
SrednjiMedium
VisokHigh
SrednjiMedium
SrednjiMedium
PouzdanostReliability
VisokiHigh
NiskiLow
VisokiHigh
SrednjiMedium
VisokiHigh
Potreban nivo obučenostiLevel of skill required
SrednjiMedium
NiskiLow
Nizak-VisokLow-High
SrednjiMedium
VisokiHigh
Troškovi automatizacijeAutomation costs
VisokiHigh
SrednjiMedium
VisokiHigh
VisokiHigh
VisokiHigh
Uzorak/danSample/day 20 50 50 50 20
Izvor/Source: http://www.iasri.res.in/ebook/EBADAT/6-Other%20Useful%20Tech-niques/ 9-DNA%20Fingerprinting-IASRI-KVB.pdf

373
Genotipska karakterizacija gajenih biljaka
PRIMENA DNK FINGERPRINTINGA
Pod DNK fingerprintingom podrazumeva se korišćenje različitih tehnika, najčešće zasnovanih na lančanoj reakciji polimeraze (PCR), radi utvrđivanja specifičnog profila DNK za određeni organizam. Slič-nost DNK profila zavisi od genetske bliskosti ispitivanih uzoraka. U prošlom veku, ispitivanja DNK sekvenci korišćena su za identifikaciju osumnjičenih u krivičnim istragama. Takođe, DNK fingerprinting može se koristiti i kao pravno sredstvo za utvrđivanje roditeljstva. Korišćen u kombinaciji sa forenzičkim i medicinskim dokazima, DNK fingerprin-ting povećao je efikasnost sudskih postupaka.
Koncept DNK fingerprintinga sve više se primenjuje u poslednjih nekoliko decenija i kod biljaka, životinja i mikroorganizama, u identifi-kaciji genotipova, populacionoj genetici, sistematici i filogeografiji.
Identifikacija genotipa
Genotipska karakterizacija biljnih vrsta i varijeteta veoma je korisna jer, kod većine biljaka, iako pripadaju istom rodu i vrsti, mogu se javi-ti značajne razlike između varijeteta, sorti i sl. DNK fingerprintingom mogu se utvrditi razlike između biljaka iz različitih familija, rodova, vrsta, sorti, pa čak i sestrinskih linija. Ukoliko pretpostavimo da svaki genotip nastaje fuzijom dve generativne ćelije, iz toga proizilazi da će svaki genotip biti drugačiji, sa izuzetkom individua koje pripadaju istoj inbred liniji visokog stepena homozigotnosti i klonova, nastalih vegeta-tivnim umnožavanjem, koji imaju isti DNK profil kao i majčinske biljke od kojih su nastali.
Brojni su primeri korišćenja ove tehnike u različite svrhe. Sposob-nost da se identifikuje svaka individualna biljka nalazi se u osnovi broj-nih primena DNK fingerprintinga. Tako je npr. ona primenjena u doka-zivanju ilegalne seče šuma u Kanadi, određivanjem podudaranja DNK profila odsečenog drveća i panjeva. DNK fingerprinting takođe je rađen

374
Semenarstvo
na ostacima pomorandže iz voćnih sokova, u cilju dokazivanja da se umesto agruma najviše klase koriste oni lošijeg kvaliteta, od strane ra-zličitih proizvođača sokova od narandže u Britaniji. Takođe je primenji-van i u Francuskoj da se utvrdi falsifikovanje Chianti vina, proizvedenog od grožđa lošijeg kvaliteta (Weising et al., 2005).
Identifikacija sorti
U proteklih 25 godina tehnike DNK fingerprintinga našle su široku primenu u identifikaciji genotipova/sorti kod brojnih gajenih vrsta, uk-ljučujući žita (Triticum spp., Horduem spp., Secale cereale, Oryza sativa, Zea mays, Echinochloa spp.), uljane kulture (Arachys spp., Brassica spp., Glycine spp.), zrnene mahunarke (Cycer spp., Lens culinaris, Pisum sa-tivum, Phaseolus spp., Vigna spp.), biljke od kojih se dobija šećer (Beta spp., Sacharum spp.), povrće (Capsicum spp., Cucumis sativus, Lycoper-sicon esculentum, Solanum spp., Raphanus sativus) i voće (Anacardium occidentale, Cytrus spp., Mangifera indica, Malus spp., Musa spp., Pru-nus spp., Pyrus spp., Rubus spp., Vitis spp.) (Weising e al., 2005).
Prateći svetske trendove i u našoj zemlji se poslednjih godina radi na DNK fingerprintingu sorti i linija gajenih vrsta. Primenom SSR mo-lekularnih markera (mikrosatelita) u Instituta za ratarstvo i povrtarstvo u Novom Sadu urađeno je profilisanje 180 sorti elitne hlebne pšenice, stvorenih u različitim oplemenjivačkim institucijama Srbije (Kondić-Špika et al., 2010). Izvršena je detekcija polimorfnosti u 18 mikrosa-telitskih lokusa. Materijal korišćen za analizu podeljen je u pet grupa, na osnovu oplemenjivačkog centra u kome je nastao i tipa sorte. Sorte stvorene u Instituta za ratarstvo i povrtarstvo u Novom Sadu, kao naj-brojnija kategorija, podeljene su u dve grupe (NS – ozime (121) i NS – jare (28)), sorte stvorene u Centru za strna žita Kragujevac svrstane su u grupu KG (13), dok su sorte iz Centra za poljoprivredna i tehnološ-ka istraživanja u Zaječaru analizirane u okviru grupe ZA (8). U grupu Ostalih (10), ubrajani su genotipovi poreklom iz manjih oplemenjivač-kih centara, zastupljenih sa jednom ili dve sorte. Kao standardi korišće-ne su sledeće sorte: Chinese Spring, Obrij, Fundulea 4, Hays 2, Mexico 3 i Madrigal, a odabir SSR markera izvršen je prema rezultatima Röder

375
Genotipska karakterizacija gajenih biljaka
i sar. (1998, 2002). Dobijeni produkti amplifikacije razdvajani su putem kapilarne elektroforeze na četvorokapilarnom sekvenceru 3130 Applied Biosystems - ABI Prism 3130. Fragmentna analiza (sl. 5) produkata obavljena je uz pomoć GeneMapper Software Version 4.0 (Applied Bi-osystems).
U 18 ispitivanih mikrosatelitskih lokusa detektovan je 151 alel, pri čemu je prosečan broj alela po markeru iznosio 8,4 (tab. 2). Pri-sustvo retkih alela (frekvencije manje od 2%) utvrđeno je u 17 SSR lokusa. Najveći broj alela i većina retkih alela detektovana je u grupi novosadskih, naročito ozimih sorti pšenice. Analiza distribucije alela po lokusima, jasno je ukazala na efekte različitog selekcionog pritiska koji postoji u navedenim oplemenjivačkim centrima (sl. 6). Na osnovu ovih 18 mikrosatelitskih markera sve ispitivane sorte, osim dve sorte jare pšenice, Golije i Jevrosime, mogle su se jasno razlikovati jedna od druge. Dobijeni rezultati ukazali su na pogodnosti primene mikrosa-telitskih markera za detekciju polimorfnosti, u cilju identifikacije ge-notipova i procene genetičkog diverziteta.
Slika 5. Elektroferogram alela veličine 143 bp u lokusu Xgwm389 (crna) i alela od 192 bp u lokusu Xgwm261 (crvena) detektovanih u sorti ozime pšenice Vojvodina.
Figure 5. Electropherogram of 143 bp (bleck) alelle in Xgwm389 locus and 192 bp (red) allele in Xgwm261 locus detected in winter wheat cultivar Vojvodina.(Kondić-Špika et al., 2010)

376
Semenarstvo
Tabela 2. Analiza sorti pšenice poreklom iz različitih oplemenjivačkih centara u SrbijiTable 2. Analysis of wheat varieties originated from different breeding centres in Serbia
KarakteristikeCharacteristics
NS-ozimeNS-winter
NS-jareNS-spring KG ZA Ostali
Others
Broj sorti / No. of varieties 121 28 13 8 10
Ukupan br. alelaTotal no. of alleles 126 76 70 57 64
Prosečan br. alela po markeruAverage no. of alleles per marker
7 4.2 3,9 3,2 3,6
Broj retkih alela (P<2% u ukupnom setu)No. of rare alleles (P<2% in total set)
48 21 29 20 22
Prosečan br. retkih alela po markeruAverage no. of rare alleles per marker
2,7 1,2 1,6 1,1 1,2
Pored pšenice, u Institutu za ratarstvo i povrtarstvo u Novom Sadu urađen je i DNK fingerprinting 96 genotipova soje sa 15 SSR markera. Što se tiče ostalih ratarskih i povrtarskih vrsta, fingerprinting 96 inbred linija kukuruza sa 40 markera i 96 genotipova pasulja sa 20 markera je u toku. Ovaj rad treba da omogući profilisanje sorti i linija gajenih vrsta, stvorenih u Srbiji u proteklih 40-50 godina i pruži mogućnost za pouz-danu i međunarodno priznatu zaštitu naših rezultata.
Širok spektar studija kod mnogih biljnih rodova pokazuje da je sistem oplemenjivanja jedna od najvažnijih determinanti genetičkog diverziteta i njegove distribucije (Weising et al., 2005; Landjeva et al., 2006). Tako, Yuan et al. (2004) koriste RAPD markere za grupisanje ge-notipova uljane repice ne samo na osnovu njihovog geografskog pore-kla, već koriste i metod oplemenjivanja, kao kriterijum za određivanje genetičke udaljenosti. Prema njihovim rezultatima, primenom rekuren-tne selekcije dobija se šira genetička osnova nego korišćenjem pedigre selekcije. Analizirajući veliki broj genotipova primenom molekularnih markera (AFLP i SSR), Seyis et al. (2004) i Hasan et al. (2004) konstru-isali su dendograme u kojima je distribucija genotipova unutar klastera
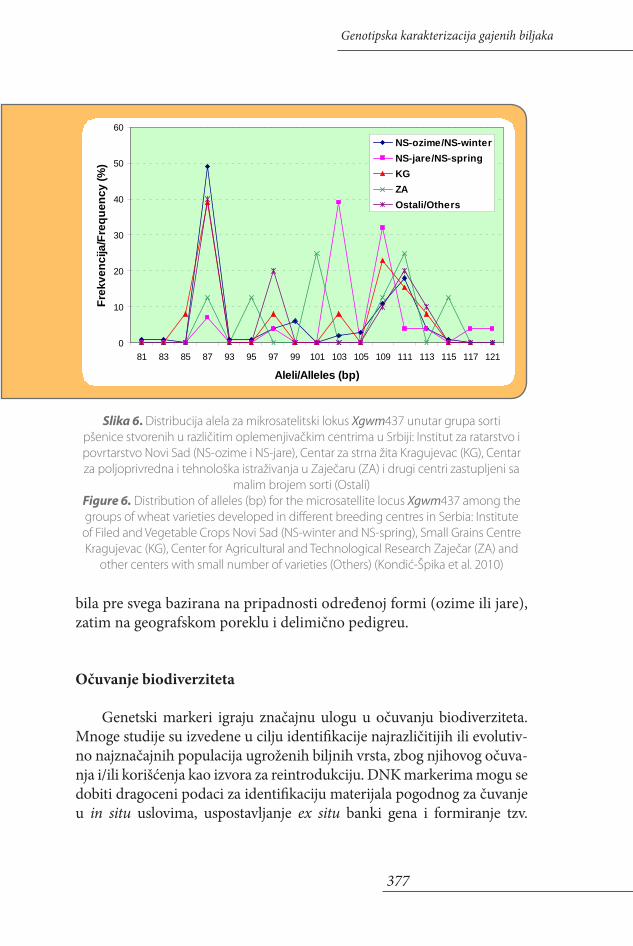
377
Genotipska karakterizacija gajenih biljaka
bila pre svega bazirana na pripadnosti određenoj formi (ozime ili jare), zatim na geografskom poreklu i delimično pedigreu.
Očuvanje biodiverziteta
Genetski markeri igraju značajnu ulogu u očuvanju biodiverziteta. Mnoge studije su izvedene u cilju identifikacije najrazličitijih ili evolutiv-no najznačajnih populacija ugroženih biljnih vrsta, zbog njihovog očuva-nja i/ili korišćenja kao izvora za reintrodukciju. DNK markerima mogu se dobiti dragoceni podaci za identifikaciju materijala pogodnog za čuvanje u in situ uslovima, uspostavljanje ex situ banki gena i formiranje tzv.
0
10
20
30
40
50
60
81 83 85 87 93 95 97 99 101 103 105 109 111 113 115 117 121
Aleli/Alleles (bp)
Frek
venc
ija/F
requ
ency
(%)
NS-ozime/NS-winterNS-jare/NS-springKGZAOstali/Others
Slika 6. Distribucija alela za mikrosatelitski lokus Xgwm437 unutar grupa sorti pšenice stvorenih u različitim oplemenjivačkim centrima u Srbiji: Institut za ratarstvo i povrtarstvo Novi Sad (NS-ozime i NS-jare), Centar za strna žita Kragujevac (KG), Centar za poljoprivredna i tehnološka istraživanja u Zaječaru (ZA) i drugi centri zastupljeni sa
malim brojem sorti (Ostali)Figure 6. Distribution of alleles (bp) for the microsatellite locus Xgwm437 among the groups of wheat varieties developed in different breeding centres in Serbia: Institute of Filed and Vegetable Crops Novi Sad (NS-winter and NS-spring), Small Grains Centre Kragujevac (KG), Center for Agricultural and Technological Research Zaječar (ZA) and
other centers with small number of varieties (Others) (Kondić-Špika et al. 2010)

378
Semenarstvo
osnovnih kolekcija (core collections) maksimalne raznolikosti, kao i za otkrivanje duplikata. Pored toga, dodeljivanje stalnog bar koda svakoj od čuvanih jedinica biodiverziteta omogućava njihovu nedvosmislenu iden-tifikaciju, kako sada, tako i u budućnosti (Weising et al., 2005).
Karakterizacija germplazme
DNK fingerprinting je važan instrument za karakterizaciju germ-plazme, odnosno, ukupnog genetičkog diverziteta prisutnog u svetu za određenu biljnu vrstu, koji obuhvata stare i novostvorene sorte, lokalne populacije, kao i divlje srodnike. Jedan od glavnih ciljeva je da se utvr-di obim i distribucija genetskih varijacija i da se razumeju geografski i ekološki aspekti procesa koji su doveli do utvrđenog stanja varijabilnosti (Hodgkin et al. 2001).
Brojni autori uspešno su koristili različite molekularne markere za određivanje genetičke sličnosti/udaljenosti genotipova u cilju karakteri-zacije germplazme i utvrđivanja stepena genetičke divergentnosti u ko-lekcijama različitih gajenih vrsta. Rezultati dobijeni primenom moleku-larnih markera koriste se za dalje kalkulacije i konstruisanje dendogra-ma, koji omogućavaju da se sagledaju, lakše tumače i uporede rezultati dobijeni pomoću različitih tehnika (Prasad et al., 2009).
U istraživanju Kondić-Špika et al. (2008) mikrosatelitski markeri (19) korišćeni su za karakterizaciju i evaluaciju genetičkog diverziteta u kolekciji sačinjenoj od 40 genotipova pšenice. Amplifikovani fragmenti skorirani su u binarnom formatu, te su ovi podaci dalje korišćeni za izračunavanje koeficijenta sličnosti (Jaccard, 1908). Na osnovu matri-ce koeficijenta sličnosti konstruisan je dendogram (slika 7), primenom UPGMA klaster analize. Na dendogramu se uočava grupisanje genoti-pova u dva glavna klastera, uglavnom prema pripadnosti ozimoj ili jaroj formi. Najsličniji genotipovi bili su sorte Nova Banatka i Vojvođanka, koje su se razlikovale međusobno samo u alelima detektovanim u dva mikrosatelitska lokusa. Na dendogramu je bilo moguće razlikovati sve genotipove jedne od drugih, na osnovu informacija dobijenih prime-nom 19 SSR markera. Mikrosatelitski markeri u ovom istraživanju iza-brani su na osnovu rezultata Röder et al. (2002), koji su pokazali da je njihovom primenom bilo moguće razlikovati 502 evropske sorte pše-nice. Samo nekoliko sorti nastalih iz ukrštanja istih roditelja nije bilo moguće razlikovati primenom ovog seta markera.

379
Genotipska karakterizacija gajenih biljaka
Slika 7. Dendogram 40 genotipova pšenice konstruisan korišćenjem UPGMA klaster analizeFigure 7. Dendogram of 40 wheat genotypes constructed using UPGMA cluster analysis
(Kondić-Špika et al. 2008)
Koristeći sličan pristup, Marjanović-Jeromela et al. (2009) su ispiti-vali genetičku varijabilnost u kolekciji uljane repice (Brassica napus L.) na osnovu morfološke, biohemijske i molekularne (RAPD) karakteri-zacije. Deset morfoloških i biohemijskih osobina (visina biljke, visina do prve grane, prečnik stabla, broj grana, broj listova, broj ljuski, prinos semena po biljci, masa 1000-semena, sadržaj ulja i proteina) analizirano je u trogodišnjem eksperimentu na 30 genotipova ozime uljane repice. RAPD analiza urađena je sa 13 prajmera, izabranih na osnovu njihove polimorfnosti (sl. 8). Značajna genetska varijabilnost između genotipo-va dobijena je na morfološkom i na molekularnom nivou. Upoređujući dendrogram dobijen na osnovu podataka RAPD analize i dendogram konstruisan na bazi deset kvantitativnih svojstava, mogu se uočiti odre-đene sličnosti, ali i razlike. U oba slučaja, genotipovi su grupisani prema geografskom poreklu, mada je ovakvo grupisanje bilo izraženije u den-drogramu dobijenom analizom kvantitativnih osobina. Grupisanje po
Genetic distance
DaraDusanka
MarijaDragicaKostana
SpasenijaMirjana
Jev rosimaNatasa
PetraNev esinjka
LelijaRadinkaBranka
PrincezaKometa
LanaJarka
Iv ankaKraljev ica
Gruzanka KGKragujev acka 58Kragujev acka 56
Nov osadska 6001Becejka
Nov osadska 6238Takov canka
StudenaMaksimaSidjanka
NSR-1Vojv odjanka
Nov a BanatkaPitoma
Alf aNov osadska 6864
PKB KrupnaZrenjaninka
Nov osadska crv enaBiserka

380
Semenarstvo
geografskom poreklu zabeleženo je i od strane drugih autora za različite vrste roda Brassica (Srivastava et al., 2001; Khan et al., 2009).
Vrednosti genetičke distance (GD) između genotipova, kao i nji-hov raspored po klasterima nisu bili slični u poređena dva dendograma (Marjanović-Jeromela et al. 2009). Drugi autori su dobili slične rezulta-te poređenjem morfoloških, biohemijskih i molekularnih analiza kod različitih vrsta (Vollmann et al., 2005; Johnson et al., 2007). Tačnost određivanja genetičke sličnosti/udaljenosti na osnovu molekularnih markera zavisi od mnogih faktora, kao što su broj korišćenih markera, njihova raspodela duž genoma, kao i stepen preciznosti sa kojom su re-zultati analizirani (Schut et al., 1997). Pored toga, molekularni markeri ne mogu se koristiti za izvođenja zaključaka o interalelnim i intraalel-nim interakcijama, koje dovode do ekspresije određenih osobina unu-tar genoma. S obzirom na to da je relativno mali broj RAPD prajmera korišćen u istraživanju Marjanović-Jeromela et al. (2009), razumno je očekivati da značajan deo genoma uljane repice nije bio pokriven anali-zom, te otuda nije bilo većih sličnosti između ispitivanih dendograma.
Postoje i drugi mogući razlozi za nedostatak korelacije između morfoloških i RAPD podataka. Jedan od njih je da RAPD markeri de-tektuju polimorfizam u kodirajućim, kao i ne-kodirajućim regionima, tako da je moguće da je najveći deo utvrđenog polimorfizma poticao od nekodirajućih regiona. Takođe, biljke mogu biti morfološki slične, ali to ne znači nužno i njihovu genetičku sličnost, jer različite genet-ske osnove mogu rezultirati sličnom fenotipskom ekspresijom (Khan et al., 2009). Ovo je posebno izraženo kod kvantitativnih svojstava, na čiju ekspresiju i međusobne odnose značajnog uticaja imaju faktori spoljaš-nje sredine, kao i njihova interakcija sa genotipom (Hristov et al., 2010;
17
Slika 8. RAPD produkti amplifikacije generisani primenom prajmera UBC 64 kod 30 genotipova
uljane repice (Marjanović-Jeromela et al. 2009)
Figure 8. RAPD amplification products generated by primer UBC 64 in 30 rapeseed genotypes
(Marjanović-Jeromela et al. 2009)
Vrednosti genetičke distance (GD) između genotipova, kao i njihov raspored po klasterima nisu
bili slični u poređena dva dendograma (Marjanović-Jeromela et al. 2009). Drugi autori su dobili slične
rezultate poređenjem morfoloških, biohemijskih i molekularnih analiza kod različitih vrsta (Vollmann
et al. 2005, Johnson et al. 2007). Tačnost određivanja genetičke sličnosti/udaljenosti na osnovu
molekularnih markera zavisi od mnogih faktora, kao što su broj korišćenih markera, njihova raspodela
duž genoma, kao i stepen preciznosti sa kojom su rezultati analizirani (Schut et al. 1997). Pored toga,
molekularni markeri ne mogu se koristiti za izvođenja zaključaka o interalelnim i intraalelnim
interakcija, koje dovode do ekspresije određenih osobina unutar genoma. S obzirom na to da je
relativno mali broj RAPD prajmera korišćen u istraživanju Marjanović-Jeromela et al. (2009),
razumno je očekivati da značajan deo genoma uljane repice nije bio pokriven analizom, te otuda nije
bilo većih sličnosti između ispitivanih dendograma.
Postoje i drugi mogući razlozi za nedostatak korelacije između morfoloških i RAPD podataka.
Jedan od njih je da RAPD markeri detektuju polimorfizam u kodirajućim, kao i ne-kodirajućim
regionima, tako da je moguće da je najveći deo utvrđenog polimorfizma poticao od nekodirajućih
regiona. Takođe, biljke mogu biti morfološki slične, ali to ne znači nužno i njihovu genetičku sličnost,
jer različite genetske osnove mogu rezultirati sličnom fenotipskom ekspresijom (Khan et al. 2009).
Ovo je posebno izraženo kod kvantitativnih svojstava, na čiju ekspresiju i međusobne odnose
značajnog uticaja imaju faktori spoljašnje sredine, kao i njihova interakcija sa genotipom (Hristov et
al. 2010, Marjanović-Jeromela et al. 2011). Sličnost u grupisanju genotipova na osnovu molekularnih
markera i fenotipskih osobina može se očekivati jedino u slučajevima kada su markeri izabrani na
osnovu njihove utvrđene povezanosti sa lokusima za kvantitativna svojstva koja su uključena u analizu
(Persson and Gustavsson 2001).
Utvrđivanje nepoznatog porekla vrsta ili sorti još je jedan od načina primene DNK
fingerprintinga u karakterizaciji germplazme. Na primer, Regner et al. (2001) su koristeći AFLP,
Slika 8. RAPD produkti amplifikacije generisani primenom prajmera UBC 64 kod 30 genotipova uljane repice (Marjanović-Jeromela et al. 2009)
Figure 8. RAPD amplification products generated by primer UBC 64 in 30 rapeseed genotypes (Marjanović-Jeromela et al. 2009)

381
Genotipska karakterizacija gajenih biljaka
Marjanović-Jeromela et al., 2011). Sličnost u grupisanju genotipova na osnovu molekularnih markera i fenotipskih osobina može se očekivati jedino u slučajevima kada su markeri izabrani na osnovu njihove utvr-đene povezanosti sa lokusima za kvantitativna svojstva koja su uključe-na u analizu (Persson and Gustavsson, 2001).
Utvrđivanje nepoznatog porekla vrsta ili sorti još je jedan od načina primene DNK fingerprintinga u karakterizaciji germplazme. Na primer, Regner et al. (2001) su koristeći AFLP, RAPD, ISSR i SSR markere u istraživanju 1200 genotipova vinove loze (Vitis spp.), bili u stanju da opišu istoriju više sorti koje su i dalje u upotrebi i to vraćajući se unazad sve do srednjeg veka.
Osnovne kolekcije (core collections)
Takozvane osnovne kolekcije ponekad se uzorkuju ili izdvajaju iz veće kolekcije germplazme, radi intenzivnije karakterizacije. U idealnom slučaju, ovakva kolekcija treba da obuhvati ceo opseg genetske varijabil-nosti jedne biljne vrste. Jedan od načina da se izabere najdivergentniji ma-terijal za osnovnu kolekciju jeste da se primeni klaster analiza, na osnovu matrice udaljenosti, generisane na osnovu podataka o svim raspoloživim uzorcima. Međusobni odnosi između genotipova u dendrogramu su pod uticajem utvrđenih razlika u ispitivanim svojstvima, kao i ekstremnih vrednosti i drugih karakteristika, odnosno upotrebom klaster analize kvalitativnih i kvantitativnih osobina grafički se prikazuju sličnosti i razli-ke između genotipova (Crossa and Cornelius, 1997). Korišćenjem klaster analize i različitih režima uzorkovanja, Hu et al. (2000) su pokazali da je u osnovnoj kolekciji pamuka (Gossipium) zadržana veća količina genetske varijabilnosti, sa uključenim superiornim predstavnicima, kada je izbor vršen na osnovu genotipskih, a ne fenotipskih karakteristika.
U cilju pravljenja osnovne kolekcije pšenice, u Institut za ratarstvo i povrtarstvo u Novom Sadu vršeno je praćenje 710 genotipova iz svetske kolekcije germplazme pšenice, poreklom iz 38 zemalja, u periodu 1993–2000. Tokom tih sedam vegetacionih sezona, ocenjivano je 54 agronom-skih, morfoloških, fizioloških i drugih osobina, u polju i kontrolisanim uslovima. Na osnovu rezultata ove višegodišnje evaluacije, sačinjena je osnovna kolekcija od 96 genotipova sa najvišim fenotipskim variranjima za 26 veoma važnih osobina za programe oplemenjivanja pšenice. Ova kolekcija genotipova korišćenja je za brojna molekularna i fiziološka

382
Semenarstvo
ispitivanja (Kobiljski et al., 2002; Kondić i Šesek, 2000), u cilju identi-fikacije poželjnih genotipova za ukrštanja, kao i utvrđivanja značajnih asocijacija između markera i agronomski važnih svojstava (Dodig et al., 2010), te potencijalnog uključivanja kandidat markera u marker asisti-ranu selekciju pšenice.
Upravljanje biodiverzitetom
Metod DNK fingerprintinga olakšava i upravljanje biodiverzitetom. Nekoliko međunarodnih centara za čuvanje biljne germplazme koristi DNK fingerprinting da im pomogne da usredsrede svoje ograničene resurse na održavanje i propagiranje originalnih i vrednih genotipova u kolekcijama. Kao jedan primer evaluacije banke gena na bazi DNK markera, može se navesti istraživanje Lowe et al. (2003), koji su koristili RAPD analizu za ispitivanje 56 uzoraka germplazme Pennisetum pur-pureum i njegovih hibrida. Ova grupa useva važan je izvor stočne hrane u istočnoj Africi. Unutar kolekcije postojala je veoma mala ili gotovo nikakva genetska varijabilnost, verovatno zbog pretežno vegetativnog razmnožavanja. Kolekcija je ocenjena primenom markera i predložena je racionalizacija broja uzoraka, nakon eliminacije onih sa identičnim profilima.
Očuvanje ex situ kolekcija germplazme često je veoma skupo, po-gotovo za one gajene vrste koje treba da se održavaju vegetativnim raz-množavanjem biljaka u polju. Jedan od takvih useva je i heksaploidna vrsta Ipomoea batatas (slatki krompir). Međunarodni centar za krompir (CIP) poseduje najveću svetsku zbirku slatkog krompira, sa 5526 uzora-ka gajenih sorti poreklom iz 57 zemalja (Zhang et al., 2001). Morfološka merenja ukazala su na to da verovatno postoji značajan broj potenci-jalnih duplikata (~ 1500) među ovim uzorcima. Pokazano je da su se morfološki različite sorte razlikovale i u svojim RAPD profilima, dok su neki od sumnjivih duplikata imali identične RAPD profile, te su mogli biti uklonjeni iz kolekcije.
U još jednom pokušaju da se smanji količina biljnog materijala za čuvanje, Phippen et al. (1997) su pokazali da značajan deo uzoraka Bra-ssica oleracea var. capitata može biti uklonjen iz kolekcije banke gena. Oko 4,6% varijabilnosti je na taj način izgubljeno, ali su troškovi održa-vanja kolekcije smanjeni za 70%.

383
Genotipska karakterizacija gajenih biljaka
DNK fingerprinting u zaštiti prava intelektualne svojine
Pravo oplemenjivača biljaka za novostvorene sorte
Za oplemenjivanje biljaka potrebno je mnogo vremena i truda. U odsustvu svojine nad novim sortama, oplemenjivači imaju malo koristi od novih sorti gajenih biljaka, jer se one mogu lako umnožiti seme-nom ili vegetativnim razmnožavanjem. Međutim, zahtev Svetske tr-govinske organizacije (World Trade Organization – WTO) državama članicama da obezbede zaštitu novih sorti značajno je promenio ovu situaciju. Sprovođenje Sporazuma o trgovinskim aspektima prava inte-lektualne svojine (Trade Related Aspects of Intelectual Property Rights Agreement), pod nadzorom Svetske trgovinske organizacije, dovelo je širom sveta do prelaska sa slobodne razmene i neometane eksploatacije na kontrolisan pristup biljnim genetskim resursima.
Prava intelektualne svojine oplemenjivača i farmera mogu biti zašti-ćena adaptacijom sistema patenata ili primenom nekog od oblika efika-snog sui generis sistema zaštite (http://www.quno.org/geneva/pdf/eco-nomic/Discussion/Sui-Generis-Systems-for-Plant-Variety-Protection-English.pdf), ili pak njihovom kombinacijom. Konvencija Ujedinjenih nacija o Biološkom diverzitetu (UN Convention on Biological Diversity – CBD) iz 1993. godine na sličan način priznaje suverenost nacija nad njihovim biljnim genetskim resursima, kao i pravo oplemenjivača i far-mera da dobiju nadoknadu za direktnu ili indirektnu eksploataciju tra-dicionalnih i novopriznatih sorti.
Nove sorte gajenih biljaka sada mogu da dobiju zaštitu u velikom broju zemalja. Danas opemenjivači imaju pravo intelektualne svojine za novopriznate sorte gajenih biljaka i to pravo je poznato pod nazivom „pravo biljnih oplemenjivača“ (Plant Breeder’s Right-PBR). Kada ople-menjivači dobiju PBR za određene sorte i određeno područje, npr. Ve-lika Britanija ili Evropa, oni mogu da naplate honorar od prodaje mate-rijala koji se koristi za propagaciju. U Evropi, ova prava mogu dobiti od institutcije pod nazivom the Community Plant Variety Office (CPVO). Dve druge organizacije uključene u pravne aranžmane za zaštitu svojih članova od povrede njihovih prava jesu Međunarodna unija za zaštitu novih sorti biljaka (Unija Internationale pour la Protection des Obten-tions vegetales-UPOV) i Međunarodna semenska federacija (Internati-onal Seed Federation-ISF).

384
Semenarstvo
Međutim, za sprovođenje sistema zaštite neophodan je veoma pre-cizan metod za identifikaciju sorti, roditelja hibrida i specifičnih jedinica biodiverziteta. Morfološki deskriptori korišćeni su rutinski u konvenci-onalnom određivanju identiteta sorti. Međutim, ovi deskriptori imaju određene nedostatke, kao što su uticaj spoljašne sredine na ekspresiju svojstava, epistatička interakcija, plejotropni efekti itd. Štaviše, nedovo-ljan broj ovih deskriptora koji nedvosmisleno mogu identifikovati sve veći broj novopriznatih sorti doveo je do potrebe da se traže i druga rešenja. Iz tog razloga postalo je aktuelno ispitivanje potencijalne upo-trebe molekularnih metoda za opis i identifikaciju sorte.
Elektroforeza proteina semena i analiza izozima delimično preva-zilaze ova ograničenja, kao i brojne druge tehnike bazirane na analizi DNK. S obzirom na to da se sve genetske razlike između individua mogu identifikovati u primarnoj strukturi njihove genomske DNK, direktan metod za identifikaciju sorti gajenih biljaka i roditeljskih linija bio bi determinacija sekvenci genoma koji se porede. Sekvenciranje zahteva veoma mnogo uloženog rada i sredstava i praktično je neizvodljivo se-kvencirati kompletne genome svih sorti. Međutim, sa automatizacijom procesa sekvenciranja, moguće je odrediti sekvence kratkih fragmenata amplifikovane DNK. Svakako da je mnogo praktičnija strategija ogra-ničiti poređenje na specifične delove genoma, koji se najčešće razliku-ju među individuama. Za identifikaciju ovakvih regiona, kao i razlika među njima, razvijeni su različiti marker sistemi, o kojima je već bilo reči u ranijim poglavljima. Primenom molekularnih markera mogu se dobiti specifični DNK profili za svaku pojedinačnu biljku, te na taj na-čin DNK fingerprinting može da dokaže da li nova sorta zadovoljava potrebne kriterijume za ostvarivanje PBR, kao što su različitost, unifor-mnost i stabilnost, odnosno kriterijume tzv DUS (distinctness–unifor-mity–stability) testa (Camlin 2001, 2003).
Jedan od glavnih problema sa molekularnim metodama je taj što mora biti definisana minimalna genetska udaljenost između sorti, radi zaštite oplemenjivača već postojećih sorti. Svakako da se razlika od samo jednog baznog para ne može smatrati dovoljnom za registraciju nove sorte (Camlin 2003). UPOV je zbog rešavanja ovakvih problema osnovao radnu grupu (Working Group on Biochemical and Molecu-lar Techniques and DNA Profiling in Particular-BMT), koja je imala

385
Genotipska karakterizacija gajenih biljaka
zadatak da ispita mogućnosti primene molekularnih markera u sistemu registracije sorti. Ova grupa je u sedmoj sezoni svog rada i postojanja (2001, BMT/7/2) došla do zaključka da postoje dva važna momenta u kojima molekularni markeri mogu pomoći DUS testiranje, a to su (1) utvrđivanje različitosti i (2) određivanje koje postojeće sorte treba da budu uključene u referentni set za testiranje u poljskim ogledima. To znači da je u tom momentu veća potreba za primenom markera bila u tzv „pre-screening“ fazi, nego kod donošenja konačne odluke o različi-tosti kandidat sorte u odnosu na druge.
Nove sorte ponekad nastaju od već zaštićenih sorti primenom ra-zličitih metoda kao što su: povratna ukrštanja, genetski inženjering, selekcija iz prirodnih ili indukovanih mutacija ili somaklonalne varija-bilnosti. Takve sorte obično izgledaju veoma slično kao zaštićena sorta od koje su nastale, ali su dovoljno različite da bi mogle da dobiju novo ime (Camlin 2001). Potencijalno, molekularni markeri mogu pomoći u priznavanju ovakvih izvedenih sorti, ispitivanjem da li postoji odre-đeni nivo usklađenosti sa originalnom sortom. S obzirom na to da transformacije i regeneracija biljaka prouzrokuju somaklonalne vari-jacije, koje se mogu ispoljiti i kao promene u agronomskim svojstvima (Bregitzer et al., 1998), postoji određena mogućnost da se primenom molekularnih tehnika proizvedu različiti DNK profili za originalnu i izvedenu sortu. Međutim, ponekad je to veoma teško izvodljivo. Tako npr. Zhang i sar. (2001) nisu bili u stanju da razlikuju izvedenu sortu transgenog slatkog krompira (Ipomoea batatas), koji sadrži intron-β-glucuronidase (Gus) transgen, od originalnog klona, korišćenjem RAPD markera. Na osnovu rezultata AFLP i mikrosatelitskih analiza samooplodnih linija kukuruza (Zea mays), Heckenberger et al. (2003) preporučuju da se kreiraju specifične granične vrednosti za ovakve izvedene sorte za marker sisteme sa različitim stepenom polimorfiz-ma. Ponavljanja laboratorijskih testova su svakako neophodna, jer čak i veoma nizak procenat tehnički indukovane varijabilnosti (npr. zbog nepotpunog sečenja DNK u AFLP analizi) može biti teško razlikovati od pravog genetskog polimorfizma. Heckenberger et al. (2003) takođe su naglasili da treba preduzeti mere opreza kako bi se osigurao visok stepen homogenosti i reproduktivnosti DNK markera pre njihove zva-nične primene u zaštiti biljnih sorti.

386
Semenarstvo
Zbog svega navedenog može se reći da DNK profili mogu postati deo pasoša za dobro opisane sorte gajenih biljaka. Međutim, malo je verovatno da će oni ikada u potpunosti zameniti morfološke deskrip-tore, s obzirom na to da različitost utvrđena na nivou DNK markera možda ne odražava nužno morfološku različitost. Tačnost identifikacije sorte može se kvantifikovati izračunavanjem verovatnoće pronalaženja identičnih profila slučajno (Badouin and Lebrun, 2001; Ibañez, 2001). Za ovu vrstu procene neophodna je korektna referentna grupa sorti, kao i baza podataka koja se odnosi na njih.
Drugi oblici intelektualne svojine
U kontekstu zaštite intelektualne svojine DNK fingerprinting može se primeniti kod patentiranja gena ili genskih fragmenata ili u slučajevima povrede oplemenjivačkih prava i biopiratstva. DNK fin-gerprinting može se koristiti kao dokaz za ili protiv navoda o kršenju prava inteluktualne svojine. Povrede ovih prava mogu nastati u sluča-jevima:
– kada se registrovana sorta gaji/prodaje neautorizovano pod istim ili drugim imenom,
– kada se biljni materijal, uključujući seme, cvetove, plodove i druge biljne proizvode lažno prodaje pod imenom registrovane sorte i
– kada se biljni materijal koji se sakupi iz prirode komercijalno ek-sploatiše bez odobrenja onih koji upravljaju biodiverzitetom.
U svim navedenim slučajevima potrebna je brza i precizna metoda da bi se dokazalo da je do povrede prava došlo. Pored toga, utvrđiva-nje povrede prava oplemenjivača drvenastog voća gotovo je nemoguće konvencionalnim DUS testovima, jer bi to zahtevalo rasadnik drveća i gajenje nekoliko godina dok ne donesu plodove, radi njihovog poređe-nja. Korišćenje DNK fingerprintinga i drugih sličnih metoda veoma je preporučljivo i poželjno u identifikaciji sorti drveća.
Mnogo je primera u kojima je uz pomoć DNK fingerprintinga re-šen slučaj povrede prava intelektualne svojine (Weising et al., 2005). Tako je u Indiji uz pomoć ove tehnologije utvrđeno da je neautorizo-vano komercijalno prodavano seme tri različite sorte čilija obeleženih

387
Genotipska karakterizacija gajenih biljaka
imenom jedne elitne sorte. Fingerprinting je pokazao da su se četiri sorte razlikovale među sobom iako su bile distribuirane pod jednim imenom.
Poznato je da je Basmati pirinač gotovo dva puta skuplji od redov-nog pirinča na evropskom tržištu. Falsifikovanje Basmatija sa drugim pirinčem krupnog zrna veoma je česta pojava. DNK tehnologija sada omogućava da se identifikuju lažna zrna u uzorku Basmati pirinča, te da se njihova količina kvantifikuje. Istraživanje koje je sprovela Bri-tish Food Standards Agency pokazalo je da je 17% od ukupno 363 uzorka, sakupljena iz marketa širom UK, sadržalo preko 20% običnog pirinča, dok je 9% od ovih uzoraka sadžalo čak 60% pirinča koji nije Basmati. Slično istraživanje vršeno je uz pomoć SSR markera na pet sorti Basmati pirinča poreklom iz USA. Podaci o DNK fingerprintin-gu ovih sorti poređeni su sa bazama podataka o alelilma prisutnim u kolekcijama sorti poreklom iz USA i sa područja Indija/Pakistan. Četiri uzorka pokazala su sličnost sa sortama poreklom iz USA, dok je peti uzorak bio bliži indijskoj sorti Pusa Basmati (Weising et al., 2005).

388
Semenarstvo
PERSPEKTIVE PRIMENE DNK FINGERPRINTINGA
S obzirom na visoku preciznost u identifikaciji genotipova biljaka, DNK fingerprinting svakako predstavlja potencijalno veoma korisno oruđe u zaštiti prava intelektualne svojine nad sortama gajenih biljaka i autentičnom germplazmom. Da bi se potencijal ove tehnike u potpu-nosti iskoristio neophodno je organizovati mrežu laboratorija za DNK fingerprinting sa uniformnim, standardizovanim protokolima za sva-ku biljnu vrstu. U ovakvim laboratorijama trebalo bi se fokusirati na primenu mikrosatelistkih markera u fingerprintingu i njihov razvoj za one biljne vrste kod kojih još uvek nisu dostupni. Pored toga, po-trebno je da se kreira i podigne svest, u naučnom i pravnom smislu, o autentičnosti i pouzdanosti DNK fingerprintinga kod biljaka (Weising et al., 2005).
UPOV takođe ispituje mogućnost primene markera za brzu iden-tifikaciju i zaštitu sorti gajenih biljaka ili za verifikaciju identiteta sorti. Ovim je takođe obuhvaćena i elektroforeza proteina semena kod pše-nice i ječma i izozima kod kukuruza, soje i suncokreta, kao dodatnih osobina za utvrđivanje različitosti sorti. Potrebno je posvetiti pažnju brojnim genetičkim, ali i tenhiničkim detaljima i rešiti sve nedoumi-ce i još uvek prisutne probleme, da bi bila moguća široka primena molekularnih markera i DNK fingerprintinga u zaštiti intelektualne svojine.

389
Genotipska karakterizacija gajenih biljaka
LITERATURA
Akkaya MS, Bhagwat AA, Cregan PB (1992): Length polymorphisms of simple sequence repeat DNA in soybean. Genetics. 132: 1131–1139
Arnau G, Lallemand J, Bourgoin M (2002): Fast and reliable strawberry cultivar identification using inter simple sequence repeat (ISSR) amplification. Euphytica. 129: 69–79
Arnold C, Rossetto M, McNally J, Henry RJ (2002): The application of SSRs characterized for grape (Vitis vinifera) to conservation studi-es in Vitaceae. Am. J. Bot. 89: 22–28
Badouin L, Lebrun L (2001): An operational Bayesian approach for the identification of sexually reproduced cross-fertilized populations using molecular markers. Acta Hortic. 546: 81–93
Bailey D C (1983): Isozymic variation and plant breeders’ rights. In: Tanksley SD and Orton JT (eds.), Isozymes in Plant Genetics and Breeding, Part A. Elsevier, Amsterdam, Netherlands, 425–441
Becher S A, Steinmetz K, Weising K, Boury S, Peltier D, Renou J-P, Kahl G, Wolff K (2000): Microsatellites for cultivar identification in Pe-largonium. Theor. Appl. Genet. 101: 643–651
Bernet G P, Mestre P F, Pina J A, Asins M J (2004): Molecular discrimi-nation of lemon cultivars. Hort. Science. 39: 165–169
Beyermann B, Nürnberg P, Weihe A, Meixner M, Epplen J T, Börner T (1992): Fingerprinting plant genomes with oligonucleotide pro-bes specific for simple repetitive DNA sequences. Theor. Appl. Genet. 83: 691–694
Brayan G J, Stephenson P, Collins A, Kirby J, Smith J B, Gale M D (1999): Low levels of DNA sequence variation among adapted genotypes of hexaploid wheat. Theor. Appl. Genet. 99: 192-198
Brbaklić Lj, Kondić-Špika A, Trkulja D, Kobiljski B (2009): Molekularna evaluacija genetičke varijabilnosti elitnog oplemenjivačkog mate-rijala pšenice. Sel. Semen. 15: 33-41

390
Semenarstvo
Brbaklić Lj, Kondić-Špika A, Trkulja D, Kobiljski B (2010): Asocijativ-na analiza između mikrosatelitskih markera i agronomski važnih svojstava pšenice. Ratar. Povrt. / Field Veg. Crop Res. 47: 505-510
Bredemeijer G M M, Arens P, Wouters D, Visser D, Vosman B (1998): The use of semi-automated fluorescent microsatellite analysis for tomato cultivar identification. Theor. Appl. Genet. 97: 584–590
Bregitzer P, Halbert S E, Lemaux P G (1998): Somaclonal variation in the progeny of transgenic barley. Theor. Appl. Genet. 96: 421–425
Caetano-Anollés G, Bassam B J, Gresshoff P M (1991a): DNA amplifi-cation fingerprinting: a strategy for genome analysis. Plant Mol. Biol. Rep. 9: 294–307
Caetano-Anollés G, Bassam B J, Gresshoff P M (1991b): High resolution DNA amplification fingerprinting using very short arbitrary oli-gonucleotide primers. Bio/Technology. 9: 553–557
Camlin M S (2001): Possible future roles for molecular techniques in the identification and registration of new plant cultivars. Acta Hortic. 546: 289–296
Camlin M S (2003): Plant cultivar identification and registration — the role for molecular techniques. Acta Hortic. 625: 37–47
CBOL–Plant Working Group (2009): A DNA barcode for land plants. Proc. Natl. Acad. Sci. USA. 106: 12794–12797
Crossa J, Cornelius P L (1997): Sites regression and shifted multiplica-tive model clustering of cultivar trial sites under heterogeneity of error variances. Crop Sci. 37: 406-415
Dallas J F (1988): Detection of DNA “fingerprints” of cultivated rice by hybridization with a human minisatellite DNA probe. Proc. Natl. Acad. Sci. USA. 85: 6831–6835
Demesure B, Sozi N, Petit R J (1995): A set of universal primers for am-plification of polymorphic non-coding regions of mitochondrial and chloroplast DNA in plants. Mol. Ecol. 4: 129–131
Depeiges A, Goubely C, Lenoir A, Cocherel S, Picard G, Raynal M, Gre-llet, F, Delseny M (1995): Identification of the most represented repeated motifs in Arabidopsis thaliana microsatellite loci. Theor. Appl. Genet. 91: 160–168
Dodig D, Zorić M, Kobiljski B, Šurlan-Momirović G, Quarrie SA (2010): Assessing drought tolerance and regional patterns of ge-netic diversity among spring and winter bread wheat using simple

391
Genotipska karakterizacija gajenih biljaka
sequence repeats and phenotypic data. Crop and Pasture Science. 61: 812-824
Duminil J, Pemonge M-H, Petit R J (2002): A set of 35 consensus primer pairs amplifying genes and introns of plant mitochondrial DNA. Mol. Ecol. Notes. 2: 428–430
Dumolin-Lapègue S, Pemonge M-H, Petit RJ (1997): An enlarged set of consensus primers for the study of organelle DNA in plants. Mol. Ecol. 6: 393–397
Fernando P, Evans BJ, Morales JC, Melnick DJ (2001): Electrophoresis artefacts — a previously unrecognized cause of error in microsa-tellite analysis. Mol. Ecol. Notes. 1: 325–328
Haberl M, Tautz D (1999): Comparative allele sizing can produce inaccurate allele size differences for microsatellites. Mol. Ecol. 8: 1347–1350
Hasan M, Lühs W, Seyis F, Friedt W, Snowdon R (2004): Surveying genetic diversity in the Brassica napus gene pool using SSR mar-kers. Proc 11th Internat. Rapeseed Congress, Copenhagen, Den-mark, 392-394
Hebert P D N, Cywinska A, Ball SL, deWaard JR (2003): Biological iden-tifications through DNA barcodes. Proc. R. Soc. London, Ser. B. 270: 313–321
Heckenberger M, Rouppe van der Voort J, Melchinger AE, Peleman J, Bohn M (2003): Variation in DNA fingerprints among accessi-ons within maize inbred lines and implications for identification of essentially derived varieties: II. Genetic and technical sources of variation in AFLP data and comparison with SSR data. Mol. Breed. 12: 97–106
Hodgkin T, Rovigliono R, De Vicente MC, Dudnik N (2001): Molecular methods in the conservation and use of plant genetic resources. Acta Hortic. 546: 107–118
Hristov N, Mladenov N, Đurić V, Kondić-Špika A, Marjanović-Jerome-la A, Šimić D (2010): Genotype by environment interactions in wheat quality breeding programs in Southeast Europe. Euphytica. 174(3): 315-324.
Hu J, Zhu J, Xu H M (2000): Methods of constructing core collections by stepwise clustering with three sampling strategies based on ge-notypic values of crops. Theor. Appl. Genet. 101: 264–268

392
Semenarstvo
Ibañez J (2001): Mathematical analysis of RAPD data to establish re-liability of varietal assignment in vegetatively propagated species. Acta Hortic. 546: 73–79
Jaccard P (1908): Nouvelles recherches sur la distribution florale. Bull. Soc. Vaudoise. Sci. Nat. 44: 223-270
Jeffreys A J, MacLeod A, Tamaki K, Neil D L, Monckton D G (1991): Minisatellite repeat coding as a digital approach to DNA typing. Nature. 354: 204–209
Jeffreys A J, Wilson V, Thein S L (1985): Individual specific fingerprints of human DNA. Nature. 316: 76-79
Jin L, Macaubas C, Hallmayer J, Kimura A, Mignot E (1996): Mutati-on rate varies among alleles at a microsatellite locus: phylogenetic evidence. Proc. Natl. Acad. Sci. USA. 93: 15285–15288
Johnson R C, Kisha T J, Evans M A (2007): Characterizing safflower germplasm with AFLP molecular markers. Crop Sci. 47: 611-618
Kahl G (2001): The Dictionary of Gene Technology. Wiley-VCH, We-inheim.
Khan M A, von Witzke-Ehbrecht S, Maass BL, Becker HC (2009): Rela-tionships among different geographical groups, agro-morphology, fatty acid composition and RAPD marker diversity in Safflower (Carthamus tinctorius). Genet. Resour. Crop. Evol. 56:19-30
Kobiljski B, Denčić S, Kondić-Špika A, Lohwasser U, Börner A (2009): Locating Stable Across Environment QTL Involved in the De-termination of Agronomic Characters in Wheat. Cereal Res. Commun. 37(3): 327-333
Kobiljski B, Denčić S, Kondić-Špika A, Obreht D (2005): Rezultati prime-ne moderne biotehnologije u oplemenjivanju strnih žita u Nauč-nom institutut za ratarstvo i povrtarstvo-Novi Sad, Zbornik radova Naučnog instituta za ratarstvo i povrtarstvo, Novi Sad 42: 421-425
Kobiljski B, Quarrie S, Denčić S, Kirby J, Ivegeš M (2002): Genetic di-versity of the Novi Sad Wheat Core Collection revealed by micro-satellites. Cell. Mol. Biol. Lett. 7(2B): 685-694
Kondić A, Šesek S (2000): Korišćenje kalusne kulture za ispitivanje to-lerantnosti genotipova pšenice prema suši. Sel. Semen. VII(1-2): 57-59
Kondić-Špika A, Kobiljski B, Denčić S, Mladenov N, Hristov N, Brbaklić Lj, Trkulja D (2010): Genetic diversity in a collection of serbian

393
Genotipska karakterizacija gajenih biljaka
wheat (Triticum aestivum L.) cultivars released in 20th century as revealed by microsatellite markers. In Vitro Cell Dev-An, 46 (Suppl 1), S134 (Poster Abstracts from 12th World Congres of the International Association for Plant Biotechnology, 06-11 June 2010, St. Louis, USA)
Kondić-Špika A, Kobiljski B, Denčić S, Mladenov N, Hristov N, Kača-venda D, Brbaklić Lj (2008): DNA Fingerprinting of Wheat (Tri-ticum aestivum L.) Varieties Using Microsatellite Markers. In: Ko-biljski B (ed.), Conventional and Molecular Breeding of Field and Vegetable Crops (Breeding 08), Conference proceedings. Novi Sad, Serbia,149-152 (CD Ed.)
Konieczny A, Ausubel F M (1993): A procedure for mapping Arabi-dopsis mutations using co-dominant ecotype-specific PCR-based markers. Plant J. 4: 403–410
Kwok P Y, Deng Q, Zakeri H, Taylor S L, Nickerson D A (1996): Increasing the information content of STS-based genome maps: identifying polymorphisms in maped STSs. Genomics. 31: 123-126
Landjeva S, Korzun V, Ganeva G (2006): Evaluation of genetic diversity among Bulgarian winter wheat (Triticum aestivum L.) varieties during the period 1925–2003 using microsatellites. Genetic Re-sources and Crop Evolution. 53:1605–1614
Li Y-Ch, Korol AB, Fahima T, Beiles A, Nevo E (2002): Microsatellites: genomic distribution, putative functions and mutational mecha-nisms: a review. Molecular Ecology. 11: 2453-2465
Lowe A J, Thorpe W, Teale A, Hanson J (2003): Characterisation of germplasm accessions of napier grass (Pennisetum purpureum and P. purpureum x P. glaucum hybrids) and comparison with farm clones using RAPD. Genet. Res. Crop Evol. 50: 121–132
Marjanović-Jeromela A, Kondić-Špika A, Saftić-Panković D, Marinko-vić R, Hristov N (2009): Phenotypic and molecular evaluation of genetic diversity of rapeseed (Brassica napus L.) genotypes. Afri-can Journal of Biotechnology. 8(19): 4835-4844
Marjanović-Jeromela A, Nagl N, Gvozdanović-Varga J, Hristov N, Kon-dić-Špika A, Vasić M, Marinković R (2011): Genotype by envi-ronment interaction for seed yield per plant in rapeseed using AMMI model. Pesq. agropec. bras., Brasília. 46(2): 174-181

394
Semenarstvo
Neumann K, Kobiljski B, Denčić S, Börner A (2011): Genome wide association mapping - a case study in bread wheat (Triticum ae-stivum L.). Mol. Breeding. 27: 37-58
Nybom H (1991): Applications of DNA fingerprinting in plant bree-ding. In: Burke T, Dolf G, Jeffreys AJ and Wolff R (eds.), DNA Fingerprinting: Approaches and Applications. Birkhäuser, Basel, Switzerland, 294–311
Nybom H (1993): Applications of DNA fingerprinting in plant popula-tion studies. In: Pena SDJ, Chakraborty R, Epplen JT and Jeffreys AJ (Eds.), DNA Fingerprinting: State of the Science. Birkhäuser, Basel, Switzerland, 293–309
Peakall R, Gilmore S, Keys W, Morgante M, Rafalski A (1998): Cro-ssspecies amplification of soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume genera: impli-cations for the transferability of SSRs in plants. Mol. Biol. Evol. 15: 1275–1287
Persson H A, Gustavsson B A (2001): The extent of clonality and gene-tic diversity in lingonberry (Vaccinium vitis-idaea L.) reveled by RAPDs and leaf-shape analysis. Mol. Ecol. 10: 1385-1397
Phippen W B, Kresovich S, Candelas F G, McFerson J R (1997): Mole-cular characterization can quantify and partition variation among genebank holdings: a case study with phenotypically similar acce-ssions of Brassica oleracea var. capitata L. (cabbage) “Golden Acre”. Theor. Appl. Genet. 94: 227–234
Powell W, Morgante M, Andre C, Hanafey M, Vogel J, Tingey S, Rafalski A, (1996): The comparison of RFLP, RAPD, AFLP and SSR (micro-satellite) markers for germplasm analysis. Mol. Breed. 2: 225–238
Powell W, Morgante M, Andre C, McNico, JW, Machray G C, Doyle J J, Tingey S V, Rafalsk, J A (1995): Hypervariable microsatellites provide a general source of polymorphic DNA markers for the chloroplast genome. Curr. Biol. 5: 1023–1029
Prasad B, Babar M A, Xu X Y, Bai G H, Klatt A R (2009): Genetic di-versity in the U.S. hard red winter wheat cultivars as reveled by microsatellite markers. Crop Pasture Sci. 60. 16-24
Prevost A, Wilkinson M J (1999): A new system of comparing PCR primers applied to ISSR fingerprinting of potato cultivars. Theor. Appl. Genet. 98: 107–112

395
Genotipska karakterizacija gajenih biljaka
Procuiner J D, Prashar S, Chen G, Wolfe D, Fox S, Ali M L, Gray M, Zhou Y, Shillinglaw M, Roeven R, DePauw R (2009): Rapid ID Technology (RIDT) in Plants: High-Speed DNK Fingerprinting in Grain Seed for the Identification, Segregation, Purity, and Vari-eties Using Labautomation Robotics. JALA. 14: 221-231
Provan J, Powell W, Hollingsworth PM (2001): Chloroplast microsatelli-tes: new tools for studies in plant ecology and evolution. Trends Ecol. Evol. 16: 142–147
Provan J, Thomas WTB, Forster BP, Powell W (1999): Copia-SSR: a sim-ple marker technique which can be used on total genomic DNA. Genome. 42. 363–366
Regner F, Stadbauer A, Eisenheld C (2001): Molecular markers for ge-notyping grapevine and for identifying clones of traditional vari-eties. Acta Hortic. 546: 331–341
Röder M S, Korzun V, Wendehake K, Plaschke J, Tixier M-H, Leroz P, Ganal MW (1998): A microsatellite map of wheat. Genetics. 194: 2007-2023
Röder M S, Wendehake K, Korzun V, Bredemeijer G, Laborie D, Bertrand L, Isaac P, Rendell S, Jackson J, Cooke RJ, Vosman B, Ganal MW (2002): Construction and analysis of a microsatellite-based databa-se of European wheat varieties. Theor. Appl. Genet. 106: 67–73
Rogstad S H, Patton J C, Schaal B A (1988): M13 repeat probe detects DNA minisatellite-like sequences in gymnosperms and angios-perms. Proc. Natl. Acad. Sci. USA. 85: 9176–9178
Russell J R, Fuller J D, Macaulay M, Hatz B G, Jahoor A, Powell W (1997): Direct comparison of levels of genetic variation among barley accessions detected by RFLPs, AFLPs, SSRs and RAPDs. Theor. Appl. Genet. 95. 714–722
Ryskov A P, Jincharadze A G, Prosnyak M I, Ivanov P L, Limborska S A (1988): M13 phage DNA as a universal marker for DNA finger-printing of animals, plants and microorganisms. FEBS Lett. 233: 388–392
Schut J W, Qi X, Stam P (1997): Association between relationships me-saures based on AFLP markers, pedigree data and morphological traits in barley. Theor. Appl. Genet. 95: 1162-1168
Seyis F, Snowdon R, Friedt W, Lühs W (2004): Molecular genetic di-versity in resynthesised and conventional spring rapeseed. Proc.

396
Semenarstvo
of 11th
International Rapeseed Congress, Copenhagen, Denmark, 382-384
Sharma P C, Winter P, Bünger T, Hüttel B, Weigand F, Weising K, Kahl G (1995): Abundance and polymorphism of di-, tri- and tetra-nucleotide tandem repeats in chickpea (Cicer arietinum L.). The-or. Appl. Genet. 90: 90–96
Smeets H J M, Brunner H G, Ropers H H, Wieringa B (1989): Use of variable simple sequence motifs as genetic markers: application to study of myotonic dystrophy. Hum. Genet. 93: 245–251
Squirrell J, Hollingsworth P M, Woodhead M, Russell J, Lowe A J, Gibby M, Powell W (2003): How much effort is required to isolate nucle-ar microsatellites from plants? Mol. Ecol. 12: 1339–1348
Srivastava A, Gupta V, Pental D, Pradhan A K (2001): AFLP-based gene-tic diversity assessment amongst agronomically important natural and some newly synthesized lines of Brassica juncea. Theor. Appl. Genet. 102: 193-199
Taberlet P, Gielly L, Pautou G, Bouvet J (1991): Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 17:1105–1109
Tautz D (1989): Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res. 17: 6463–6471
Thuillet A C, Bru D, David J, Roumet P, Santoni S, Sourdille P, Bataillon T (2002): Direct estimation of mutation rate for 10 microsatellite loci in durum wheat, Triticum turgidum (L.) Thell. ssp. durum. Mol. Biol. Evol. 19:122–125
Trkulja D, Kondić-Špika A, Brbaklić Lj, Kobiljski B (2011): Analiza veze marker – svojstvo za vreme klasanja i cvetanja pšenice korišće-njem pojedinačne marker regresije. Ratar. Povrt. / Field Veg. Crop Res. 48(1): 113-120
Udupa S M, Baum M (2001): High mutation rate and mutational bias at (TAA)n microsatellite loci in chickpea (Cicer arietinum L.). Mol. Genet. Genomics. 265: 1097–1103
Vijayan K, Tsou C H (2010): DNA barcoding in plants: taxonomy in a new perspective. Current Sci. 99(11): 1530-1541
Vollmann J, Grausgruber H, Stift G, Dryzhyruk V, Lelley T (2005): Ge-netic diversity in Camelina germplasm as reveled by seed quality characteristics and RAPD polymorphism. Plant Breeding. 124: 446-453

397
Genotipska karakterizacija gajenih biljaka
Weber J L, May P E (1989): Abundant class of human DNA polymorphi-sms which can be typed using the polymerase chain reaction. Am. J. Hum. Genet. 44: 388–396
Weising K, Beyermann B, Ramser J, Kahl G (1991): Plant DNA fin-gerprinting with radioactive and digoxigenated oligonucleoti-de probes complementary to simple repetitive DNA sequences. Electrophoresis. 12: 159–169
Weising K, Gardner R C (1999): A set of conserved PCR primers for the analysis of simple sequence repeat polymorphisms in chloroplast genomes of dicotyledonous angiosperms. Genome. 42: 9–19
Weising K, Kaemmer D, Weigand F, Epplen J T, Kahl G (1992): Oligo-nucleotide fingerprinting reveals various probe-dependent levels of informativeness in chickpea (Cicer arietinum). Genome. 35: 436–442
Weising K, Kahl G (1998): Hybridization-based microsatellite finger-printing of plants and fungi. In: Caetano-Anollés G and Gress-hoff PM (eds.), DNA Markers: Protocols, Applications, and Over-views. Wiley-VCH, New York, USA, 27–54
Weising K, Nybom H, Wolff K, Kahl G (2005): DNA Fingerprinting in Plants: Principles, Methods, and Applications. CRC Press, Taylor & Francis Group, Boca Raton, USA, 444
Weising K, Weigand F, Driesel A, Kahl G, Zischler H, Epplen J T (1989): Polymorphic simple GATA/GACA repeats in plant genomes. Nucleic Acids Res. 17:10128
Welsh J, McClelland M (1990): Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res. 18: 7213–7218
Williams J G K, Kubelik A R, Livak K J, Rafalski J A, Tingey S V (1990): DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 18: 6231–6235
Wolfe A D, Liston A (1998): Contributions of PCR-based methods to plant systematics and evolutionary biology. In Molecular Syste-matics of Plants II. DNA Sequencing, Soltis, D.E., Soltis, P.S., and Doyle, J.J., Eds., Kluwer, Dordrecht, pp. 43–86
Wu Y Q, Taliaferro C M, Bai G H, Anderson M P (2005): Genetic di-versity of Cynodon transvaalensis Burtt-Davy and its relatedness to hexaploid C. Dactylon (L.) Pers. as indicated by AFLP markers. Crop Sci. 45: 848-853

398
Semenarstvo
Yuan M, Zhou Y, Liu D (2004): Genetic diversity among populations and breeding lines from recurrent selection in Brassica napus as revealed by RAPD markers. Plant Breed. 123: 9-12
Zabeau M, Vos P (1993): Selective restriction fragment amplification: a general method for DNA fingerprinting, European Patent Appli-cation EP 0534858
Zhang D P, Huaman Z, Rodriguez F, Rossel G, Ghislain M (2001): Iden-tifying duplicates in sweet potato (Ipomoea batatas (L.) Lam) cul-tivars using RAPD. Acta Hortic. 546: 535–541
Zietkiewicz E, Rafalski A, Labuda D (1994): Genomic fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reacti-on amplification. Genomics. 20: 176-183

399
IZDAVANJE OVE MONOGRAFIJE POMOGLI SU
––

400