-
8/10/2019 Mol. Hum. Reprod. 1996 Bamberger 457 61
1/5
Molecular Human Reproduction vol.2 no.6 pp.
4 5 7 -4 61 ,
1996
Expression of steroidogenic factor-1 (SF-1) mRNA and protein
in
the human placenta
Ana-Maria Bamberger
1
-
5
, Shereen Ezzat
2
, Bruce Cao
2
, Margaret W ong
3
, Keith L.Parker
3
,
Heinrich M.Schuhte and Sylvia L.Asa
4
institute for Hormone and Fertility Research, University of
Hamburg, Grandweg 64, 22529 Hamburg, Germany,
de pa rtm en t of Medicine Endoc rinology), Wellesley Hospital,
University of Toronto, Toronto, Onta rio, Canada,
3
Department of Medicine and Biochemistry, Howard Hughes Medical
Institute, Duke University Medical Center, Durham,
NC,
USA and
4
Department of Pathology, Mount Sinai Hospital, Samuel Lunenfeld
Research Institute, University of
Toronto, Toronto, Ontario, Canada
^ o whom correspondence should be addressed
Steroidogenic factor-1 (SF-1), also known as adrenal-4-binding
protein (Ad4BP), is a recently-described
transcription factor, which has been shown to be important for
the differentiation of steroidogenic tissues.
In addition, SF-1 has been implicated in regulating the
glycoprotein hormone oc-subuntt gene in a pituitary
gonadotroph cell line. Considering that the human placenta
produces both steroids and human chorionic
gonadotrophin (HCG), we studied the expression of SF-1 in this
tissue. Human first trimester and term
placentas were collected at the time of therapeutic abortion and
birth respectively. Messenger RNA was
extracted, reverse transcribed, and used for polymerase chain
reaction (PCR) amplification with primers
specific for the human SF-1 cDNA sequence. A band of the
expected size was obtained from both first and
third trimester samples, indicating that SF-1 expression in the
human placenta starts early in pregnancy and
is maintained until birth. In addition to normal placental
samples, JEG3 and JAR choriocarcinoma cells were
also analysed and found to express SF-1 mRNA. The identity of
the amplified products was confirmed by
diagnostic restriction digest and Southern hybridization. SF-1
protein was localized mainly to the nuclei of
the cyto- and syncytiotrophoblast and to some mesenchymal
villous nuclei by immunocytochemistry using
a specific antibody. We conclude that SF-1 is expressed in human
first trimester and term placenta, where it
could be implicated in the regulation of HCG production, in
steroidogenesis, or both.
Keywords
mRNA/placenta/protein/steroidogenic factor-1 (SF-1)
Introduction
Steroidogenic factor-1 (SF-1), also known as
adrenal-4-binding
protein (Ad4BP) is a recently-described transcription factor
encoded by the mammalian homologue of the
Drosophila
FTZ-F1 gene (Ikeda
et al.
1993). SF-1 was first identified
through its ability to bind to and coordinately regulate the
expression of several genes encoding enzymes of the steroid
hormone biosynthesis pathway (Rice
et al.
1991; Lala
et al.
1992;
Morohashi
et al
1992; Lynch
et al.
1993; Morohashi
et al.
1993). Subsequently, SF-1 was shown to belong to the
nuclear steroid receptor family of transcription factors
(Honda
et al.
1993) and to regulate several other genes, such as the
Mullerian inhibiting substance (MIS) gene (Shen
et al.
1994),
the oxytocin gene in the bovine ovary (Wehrenberg
et al.
1994), and the glycoprotein a-subunit gene in the aT3-l
pituitary gonadotroph cell line (Bamhart and Mellon, 1994).
Targeted disruption of the
Ftzfl
gene in mice demonstrated
that SF-1 is essential for adrenal and gonadal development
(Luo
et al.
1994; Sadovsky
et al.
1995), as well as for the
differentiation of pituitary gonadotrophs (Ingraham
et al.
1994), and the formation of the ventromedial nucleus of the
hypothalamus (Ikeda
et al.
1995). Recent data from our
laboratory (Asa et al. 1996) indicate that SF-1 may also be
implicated in regulating cytodifferentiation of gonadotrop hs
in
the human pituitary, since SF-1 was found to be expressed
exclusively in this cell type, both in normal pituitaries and
in
pituitary adenomas.
Considering that the human placenta is an important source
of both steroids and human chorionic gonadotrophin (HCG),
we studied the expression of SF-1 in this tissue.
Materials and methods
Human
placentaland adrenal tissues
Human first trimester and term placentas were obtained at the
time
of therapeutic abortion and birth respectively. Fresh tissue was
divided
for histological and immunocytochemical studies and for
molecular
analysis. Normal human adrenal tissue (used as a positive
control)
was obtained at autopsy from patients with no evidence of
endocrine
abnormality and examined histologically to exclude the
possibility of
incidental pathology.
ll
culture
JEG3 and JAR human choriocarcinoma cells were purchased from
ATCC (Rockville, MD, USA) and maintained in Dulbecco's
minimal
essential medium (DMEM) with 4.5 g glucose/1 and glutamine,
with
10% fetal bovine serum (FBS) added (JEG3), and Roswell Park
European Society for Human Reproduction and Embryology
45 7
-
8/10/2019 Mol. Hum. Reprod. 1996 Bamberger 457 61
2/5
A.M.Bamberger et al.
2 3 0
SF-1
OCATCTTaOOCTOCCTOCAOGAGCCCACCAAAAGCCGCCCCGACCAGCCGGCGGCCT
TCGGCCTCCTGTGCAGAATGGCCGACCAGACCTTCATCTCCATCGTGGACTGGGCACGCA
GGTGCATGGTCTTCAAGGAGCTGGAGGTGGCCGACCAGATGACGCTGCTGCAGAACTGCT
GGAGCGAGCTGCTGGTGTTCGACCACATCTACCGCCAQOTCCAOCACGGCAAOG
Figure 1. Sequence of human SF-I cDNA in the amplified region
(bp 885-1115 in the coding region). The sequences used for
primer
design are in bold letters; the BsrI recognition sequence is
underlined.
Memorial Institute (RPMI) 1640 medium with 10% FBS (JAR).
Both
cell lines were grown in 5% CO
2
and used for RNA extraction when
the cells were 60-80% confluent.
RNA extraction
Fresh frozen human first trimester and term placental tissue
was
homo genized with RN Azol (Tel-Test, Friends wood, TX, USA ;
2 ml/100 mg tissue) in a glass-Teflon homogenizer. Total RNA
was
extracted with chloroform (0.2 ml/2 ml homogenate),
precipitated
with isopropanol, washed with 75% ethanol, and dissolved in
diethylpyrocarbonate (DEPC)-treated, RNase-free water. RNA
con-
centration and purity were determined by spectrophotom etry. The
same
extraction method was used for JEG3 and JAR choriocarcinoma
cells.
Reverse transcription polym erase chain reaction
RT-PCR)
Complementary DNA was synthesized in each case from 5 |ig
total
RNA with Superscript RNase H" Reverse Transcriptase (Gibco
BRL, Gaithersburg, MD, USA) using oligo(dT) primers
(Pharmacia,
Piscataway, NJ, USA). Of the resulting cDNA 5% was used as a
template for polymerase chain reaction (PCR).
A recombinant bacteriophage clone containing the genomic
sequences of the human SF-1 gene has been isolated (Taketo et
al.
1995). Partial sequence of this clone and a second
bacteriophage
clone yielded the coding sequences to the gene. For PCR, the
following oligonucleotide primers were used to identify
SF-1:
upstream 5' GCA TCT TGG GCT GCC TGC AG 3' and downstream
5 '
CCT TGC CGT GCT GGA CCT GG 3'. These primers span one
intron between exons 4 and 5 of the human SF-1 genomic DNA,
generating a 230 bp product from cDNA (Figure 1). PCR was
carried
out in a volume of 25 |il; following an initial denaturing step
(95C
for 120 s), amplification was carried out through 30 cycles at
95C
for 30 s, 60C annealing for 30 s and 72C for 45 s in a
thermal
cycler using GeneAmp PCR reagents (Am plitaq; Perkin Elmer
Cetus,
Norwalk, CT, USA).
PCR products were visualized through electrophoresis in a 1%
agarose gel and ethidium bromide staining. The 230 bp
fragment
generated by PCR was also extracted in chloroform and
precipitated
in ethanol before digestion with the restriction endonuclease
Bsr-I
(Boehringer Mannheim, Indianapolis, IN, US A). Following
digestion,
the expected 120 bp and 110 bp fragments were visualized by
electrophoresis with ethidium bromide staining.
Negative controls included mock reverse transcription
without
RNA or without reverse transcriptase (RT). The positive control
was
represented by RNA from human adrenal cortex.
Southern hybridization
Amplified DNA was further analysed by Southern hybridization.
RT-
PCR product from total RNA of the H295 human adrenal tumour
cell line was subcloned into the pCRII vector using the TA
cloning
kit (Invitrogen, San Diego, CA, USA) and sequenced. A 230 bp
fragment of this clone was labelled with [a-
32
P]-ATP using random
primers and used for hybridization.
The cDN A fragments w ere separated by agarose gel
electrophoresis,
1 2 3 4 5 6 7 8 9 1 0 1 1
H
2
O 1st 3rd JEG JAR
trimester trimester JAR -RT Adrenal
Figure 2. Reverse transcription-polymerase chain reaction
(RT-
PCR) and hybridization for SF-1 in human placenta. Upper
panel:
ethidium bromide staining of RT-PCR products shows bands of
the
expected size with SF-1-specific primers. Lanes 1:
water/SF-1
specific primers. Lanes 2, 3, 4: cDNAs from three different
human
first trimester placental samples, showing an amplification
product
of the expected size (230 bp) with SF-1 specific primers. Lanes
5,
6, 7: cDNAs from three different human term placentas,
showing
the amplification product of the expected size with SF-1
specific
primers (the RNA of origin for the cDNA in lane 5 was
slightly
degraded before extraction on account of the placental
tissue
collection conditions). Lanes 8 and 9: cDNAs from JEG3 (lane
8)
and JAR (lane 9) choriocarcinoma cells, showing the
amplification
product of the expected size with SF-1 specific primers. Lane
10:
JAR (-RT) negative con trol. Lane 11: positive control
consisting of
cDNA from human adrenal cortex amplified with SF-1 specific
primers. Lower panel: Southern hybridization of the gel shown
in
the upper panel with a radiolabelled human SF-1 probe
confirms
the specificity of the RT-PCR products. The probe consisted of
a
RT-PCR product from total RNA of the H295 human adrenal
tumour cell line; the lanes are the same as in the upper
panel.
transferred to a nylon membrane (Gene Screen Plus, Du Pont,
Wilmington, DE, USA), and baked for 2 h at 80C. The blots
were
prehybridized for 2 h, then hybridized for 18 h at 42C, washed
at
high stringency, and autoradiographed for 2 h at room
temperature.
Immunohistochemical localization of
SF-1
protein
Formalin-fixed paraffin embedded tissues were sectioned at 5
urn
and rehydrated. For staining of nuclear antigens, sections
were
pretreated for antigen retrieval by microwaving in citrate
buffer (Shi
el al. 1991). Endogenous peroxidase activity was blocked
with
aqueous hydrogen peroxide, and non-specific binding was
prevented
by preincubation in normal goat serum. The avidin-biotin
technique
was performed with a primary polyclonal SF-1 antiserum raised
in
458
-
8/10/2019 Mol. Hum. Reprod. 1996 Bamberger 457 61
3/5
SF-1 in human placenta
."&' >
r
1
B
.
i
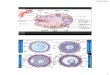
Figure 3. Immunohistochemical localization of SF-1 protein in
human placenta. (A) Strong nuclear immunoreactivity for SF-1 in
a
first trimester placenta sample: positive nuclei in the cyto-
and syncytiotrophoblast and some mesenchymal villous cells; (B)
cytoplasmic a-
human chorionic gonadotrophin (HCG) immunoreactivity in the
trophoblast of a first trimester placenta sample; (C) replacement
of the SF-1
specific antiserum with non-immune serum results in negative
staining in first trimester placenta; (D) positive control for SF-1
staining
represented by a human adrenal cortex sample.
rabbits against the DNA-binding domain of mouse SF-1,
produced
in
Escherichia coli
as a GST fusion protein and partially purified via
GST-sepharose column chromatography (Product code: 06 431,
lot
13702; Upstate Biotechnology Inc., Lake Placid, NY, USA) at
a
dilution of
1:1000.
This antibody shows cross-reactivity only with
human and bovine SF-1. For ot-HCG a monoclonal antibody
(Zymed
Laboratories In c., South San F rancisco, CA, USA ) at 1:4. The
reaction
product was visualized with 3,3'-diamino-benzidine
tetrahydro-
chloride. The positive control for SF-1 consisted of adrenal
gland
fixed and embedded with identical conditions. The specificity of
the
reaction was verified by replacing the primary antibody with
non-
immune rabbit serum.
Results
RT PCR analysis
RT-PCR using cDNA derived from multiple different human
first trimester and term placentas demonstrated the presence
of
SF-1
mRNA in this tissue. A band of the predicted size
(230 bp) was visualized after amplification with the human
SF-1 specific primers (Figure 2). A band of the expected
size was also identified using cDNA from JEG3 and JAR
choriocarcinoma cells (Figure
2).
Human adrenal cortex (Figure
2) was used as a positive control, showing an amplification
product which was of the same size as the placental product.
RT-PCR with omission of reverse transcriptase and with water
replacing template were both negative (Figure 2). Further
negative controls represented by thyroid tissue also yielded
no
band with the SF-1 primers (not shown). Diagnostic digestion
of the placental RT-PCR products with Bsr-I resulted in
fragments of 120 and 110 bp which were of the expected size
and identical to those obtained with the adrenal product
(positive control).
Southern hybridization
To further characterize the 230 bp placental amplification
product, Southern hybrization was performed as described.
Both the positive control and the placental products
hybridized
with the human SF-1 probe (Figure 2, lower panel).
Immunohistochemistry
To assess whether
SF-1
mRNA was translated in human
placenta, immunohistochemical analysis was performed. SF-1
protein immunoreactivity was detected mainly in the nuclei
of
placental cyto- and syncytiotrophoblast, as well as in some
mesenchymal villous cell nuclei (Figure 3A). The immuno-
reactive expression pattern was compared to the ot-HCG
459
-
8/10/2019 Mol. Hum. Reprod. 1996 Bamberger 457 61
4/5
A.M.Bamberger et al.
expression pattern (Figure 3B). The positive control is
repre-
sented by human adrenal cortex showing SF-1 immuno-
reactivity (Figure 3D). The negative control (Figure 3C) was
produced by replacing the SF-1 specific primary antiserum
with non-immune rabbit antiserum.
Discussion
The results of our study clearly demonstrate that SF-1 mRNA
is expressed in human first trimester and term placenta as
well as in JEG3 and JAR human choriocarcinoma cells.
Immunohistochemistry indicates that the mRNA is translated
into the SF-1 protein, which is localized mainly to the
nuclei
of the placental trophoblast, as well as some mesenchymal
villous cells.
The human placenta is, besides the pituitary, the other
important source of gonadotrophin, producing HCG at high
concentrations soon after fertilization and implantation.
HCG
is also produced in trophoblastic malignancy and is a useful
marker of these conditions (Kurman et al. 1984). The regula-
tion of the HCG a- and (i-subunit genes has been the focus
of recent investigation (for review, see Jameson and
Hollenberg,
1993), but SF-1 has not so far been implicated as a possible
regulator of these genes in the placenta. Our data indicate
that
SF-1 is expressed in both normal human placenta and in
choriocarcinoma cells. It will be interesting to determine
whether SF-1 plays a role in regulating one or both these
genes in the normal human placenta and in trophoblastic
tumours producing HCG.
It has been suggested that SF-1 probably does not play a
role in rodent placental function, since no placenta
abnormali-
ties have been reported for the SF-1 knock-out mice (Luo
et al. 1994; Sadovsky et al. 1995). SF-1 mRNA has been
found in mouse placenta (Sadovsky et al. 1995) but not in
rat placenta or Rcho-1 trophoblast cells (Yamamoto et al.
1995). These data do not contradict the possibility of a
role
for SF-1 in the regulation of HCG in the human placenta.
On the contrary, it is a known fact that rodent placentas
do not produce chorionic gonadotrophin, which has so far
been unequivocally demonstrated only in primate and equine
placentas (Roberts
et al.
1994). Thus, expression of SF-1 in
the human, but not the rodent placenta, might be essential
to understand the species-specific expression of chorionic
gonadotrophin.
In addition to producing gonadotrophin, the placenta is
also one of the most important sources of steroid hormones,
producing large amounts of progesterone and oestrogen
(Simpson and MacDonald, 1981). As mentioned previously,
the first observation of SF-1 was based on its capacity to
regulate the expression of steroidogen ic enzyme s (for a
review,
see Parker and Schimmer, 1993). A recent report based on
careful analysis of SF-1-/- mice indicated that, although
SF-1
is expressed in the normal mouse placenta from oestrus day
14 on, placentas of mice lacking SF-1 express both P450scc
and P450cl7. This suggests that SF-1 might not be essential
for expression of these genes in mouse placenta (Sadovsky
et al.
1995). SF-1 was also found to regulate the expression
of the aromatase gene in rat ovaries and R2C rat Leydig
tumour cells (Lynch
et al.
1993). Aromatase is not exp ressed
in the rat placenta (Means
et al.
1991). Aromatase activity
has been shown to be present in the syncytiotrophoblast of
human placenta, hydatidiform moles, and in choriocarcinoma
cells (Ryan, 1959; MacDonald and Siiteri, 1964; Bahn et al.
1981; Means et al. 1991; Zhou et al. 1992). Further investi-
gation is necessary to determine whether SF-1 plays a role
in
regulating the human aromatase gene, similar to its role in
the
rat (Lynch et al. 1993). The human placenta and chorio-
carcinoma cells are a source of aromatase and as we have
now dem onstrated that they also express S F-1 , we
therefore
predict a possible role for SF-1 in regulating this gene in
placenta. This does not exclude the possiblity that SF-1
might
also regulate other placental steroidogenic enzymes or other
placental genes, including the HCG genes, thus acting at
multiple levels in controlling hormonal mechanisms involved
in the establishment and maintenance of human pregnancy.
Acknowledgements
This work was supported in part by grant MA 12196 of the
Medical Research Council of Canada and the Saul A.Silverman
Family Foundation. The technical assistance of Kelvin So and
Cathy
Grabowski is gratefully acknowledged.
References
Asa, S.L., Bamberger, A.M.. Cao, B. et al. (1996) The
transcription factor
steroidogenic factor-1 is preferentially expressed in the human
pituitary
gonadotroph. J. Clin. Endocrinol. Metab.. in press
Bahn, R.S., Worsham, A.. Speeg, K.V.etal. (1981)
Characterization of steroid
production in cultured human choriocarcinoma cells J. Clin.
Endocrinol
Metab..52 , 447^*50.
Barnhart, K.M. and Mellon. P.L. (1994) The orphan nuclear
receptor,
steroidogenic
factor-1.
regulates the glycoprotein hormone alpha-subunit
gene in pituitary gonadotropes.
Mol. Endocrinol.
8, 878-885.
Honda. S., Morohashi, K., Nomura, M. et al. (1993) Ad4BP
regulating
steroidogenic P450 gene is a member of steroid receptor
superfamily.
J. Biol. Chem. 268, 7494-7502.
Ikeda. Y., Lala, D.S.. Luo. X. et al. (1993) Characterization of
the mouse
FTZ-FI gene, which encodes a key regulator of steroid
hydroxylase gene
expression. Mol. Endocrinol. 7, 852-860.
Ikeda. Y, Luo. X., Abbud. R. et al.(1995) The nuclear receptor
steroidogenic
factor I is essential for the formation of the ventromed ial
hypo thalam ic
nucleus. Mol. Endocrinol. 9, 478-486.
Ingraham. H.A.. Lala. D.S.. Ikeda. Y. et al. (1994) The nuclear
receptor
steroidogenic factor acts at multiple levels of the reproductive
axis.Genes
Dev..8, 2303-2312.
Jameson, J.L. and Hollenberg. A.N. (1993) Regulation of
chononic
gonadolropin gene expression. Endocr. Rev. 14,
203-221.
Kurman, RJ., Young, R.H.. Norris. HJ. et al. (1984)
tmmunocytochemical
localization of placental lactogen and chorionic gonadotropin in
normal
placenta and trophoblastic tumors, with emphasis on
intermediate
trophoblast and the placental site trophoblastic tumor. Int. J.
Gynecol.
Pathol.. 3,
101-121.
LaJa. D.S .. Rice. D.A. and Parker. K.L. (1992) Stero idogen ic
factor I. a key
regulator of steroidogenic enzyme expression, is the mouse
homolog of
fushi tarazu-factor I. Mol. Endocrinol. 6, 1249-1258.
Luo,
X., Ikeda, Y. and Parker. K.L. (1994) A cell-specific nuclear
receptor is
essential for adrenal and gonadal development and sexual
differentiation.
Cell 7 7, 4 8 1 ^ 9 0 .
Lynch. J.P.. Lala. D.S., Peluso. JJ. et al. (1993) Steroidogenic
factor I, an
orphan nuclear receptor, regulates the expression of the rat
aromatase gene
in gonadaJ tissues. Mol. Endocrinol.. 7, 776-786.
MacDonald. P.C. and Siiteri. P.K. (1964) Study of estrogen
production in
uomen with hydatidiform mole. J. Clin. Endocrinol. Melab..24,
685-690.
46 0
-
8/10/2019 Mol. Hum. Reprod. 1996 Bamberger 457 61
5/5
SF-1 in human placenta
Means, CD .. Kilgore. M.W.. Mahendroo. M.S.
et al.
(I99I) Tissue-specific
promoters regulate aromatase cytochrome P450 gene expression in
human
ovary and fetal tissues. Mol. Endocrinol..5 , 20052013.
Morohashi. K.. Honda. S.. Inomata. Y. et al. (I992) A common
transacting
factor. Ad4-binding protein, to the promoters of steroidogenic
P-450s.
J. Biol. Chem..
267 1791317919.
Morohashi. K.. Zanger. U.M.. Honda. S.et al. (1993) Activation
of CYPI IA
and CYPI IB gene promoters by the steroidogenic cell-specific
transcription
factor, Ad4BP. Mol. Endocrinol.. 7, 1196-11204.
Parker. .K.L. and Schimmer. B.P. (1993) Transcriptional
regulation of the
adrenal steroidogenic enzymes.
Trends Endocrinol. Metab..
4 , 46-50.
Rice. D.A.. Mouw. A.R.. Bogerd. A.M. and Parker. K.L. (1991) A
shared
promoter element regulates the expression of three steroidogenic
enzymes.
Mol. Endocrinol.. 5, 1552-1561.
Roberts. R.M. and Anthony. R.V. (1994) Molecular biology of
trophectoderm
and placental hormones. In Findlay. J.K. (ed.). Molecular
Biology of the
Female Reproductive System.Academic Press Inc.. San Diego, pp.
395440.
Ryan. KJ (1959) Biological aromatization of steroids. J. Biol.
Chem.. 234
268-272.
Sadovsky. Y. Crawford. P.A.. Woodson, K.G. et al. (1995) Mice
deficient in
the orphan receptor steroidogenic factor 1 lack adrenal glands
and gonads
but express P450 side-chain-cleavage enzyme in the placenta and
have
normal embryonic serum levels of corticosteroids. Proc.
Natl.
Acad
Sci.
USA.
92, 10939-10943
Shen, W.-H., Moore. C.C.D. Ikeda. Y. et al. (1994) Nuclear
receptor
steroidogenic factor
regulates the Miillerian inhibiting substance gene: a
link to the sex determination cascade Cell. 77, 651-661
Shi,
S. R..
Key. M.E. and Kalra, K.L. (1991) Antigen retrieval in
formalin-
fixed, paraffin em bedded tiss ues: an enhancem ent method
for
immunohistochemical staining based on microwave oven heating of
tissue
sections. J Histochem. Cytochem. 39, 741-748.
Simpson. E.R. and MacDonald, P.C. (1981) Endocrine physiology of
the
placenta. Ann. Rev. Physiol. 43 163-179.
Taketo. M.. Parker. K.L., Howard, T.A.et al.(1995) Homologs
ofDrosophila
Fushi-Tarazu factor I map to mouse chromosome 2 and human
chromosome
9q33. Genomics 25, 565-567.
Wehrenberg, U., Ivell. R.. Jansen, M.
et al.
(1994) Two orphan receptors
binding to a common site are involved in the regulation of the
oxylocin
gene in the bovine ovary. Proc. Nail.
Acad
Sci. USA 91, 1440-1444.
Yamamoto. T., Chapman, B.M.. Clemens. J.W. et al. (1995)
Analysis of
cytochrome P-450 side-chain cleavage gene promoter activation
during
trophoblast cell differentiation. Mol. Cell. EndocnnoL 113
183-194.
Zhou, D., Korzekwa. K.R.. Poulos. T. and Chen. S. (1992) A
site-directed
mutagenesis study of human placental aromatase. J. Biol. Chem.
267
762-768.
Received on December 13. 1995; accepted on March 18. 1996
461


















![Sophismata Buridani ([Reprod.])](https://img.pdfslide.tips/doc/110x75/625d10ffa98da525ef7f60fa/sophismata-buridani-reprod.jpg)








