Embed Size (px)
Citation preview

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
---------------------
NGUYỄN THỊ VÂN
NGHIÊN CỨU ĐẶC ĐIỂM SINH HỌC CHỦNG XẠ KHUẨN
Streptomyces toxytricini (VN08 - A12) KHÁNG BỆNH BẠC LÁ LÚA DO
Xanthomonas oryzae
LUẬN VĂN THẠC SĨ KHOA HỌC
Hà Nội - 2014

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
-----------------------
NGUYỄN THỊ VÂN
NGHIÊN CỨU ĐẶC ĐIỂM SINH HỌC CHỦNG XẠ KHUẨN
Streptomyces toxytricini (VN08 - A12) KHÁNG BỆNH BẠC LÁ LÚA DO
Xanthomonas oryzae
Chuyên ngành: Vi sinh vật học
Mã số: 60420107
LUẬN VĂN THẠC SĨ KHOA HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC: PGS. TS. Dương Văn Hợp
TS. Phạm Thế Hải
Hà Nội - 2014

i
LỜI CẢM ƠN
Lời đầu tiên, em xin gửi lời cảm ơn chân thành tới PGS. TS. Dương Văn Hợp
và TS. Phạm Thế Hải, người thầy đã trực tiếp hướng dẫn em rất tận tình trong cả
quá trình thực hiện đề tài, giúp em vượt qua khó khăn và hoàn thành tốt luận văn
này.
Em xin gửi lời cảm ơn tới các thầy cô giáo trong Khoa Sinh học, trường Đại
Học Khoa học Tự nhiên đã nhiệt tình giảng dạy, hướng dẫn và cung cấp cho em
những kiến thức bổ ích trong suốt hai năm học vừa qua và giúp đỡ em rất nhiều
trong việc nắm bắt kiến thức cũng như động viên em rất lớn về mặt tinh thần.
Em cũng xin gửi lời cảm ơn sâu sắc tới TS. Nguyễn Kim Nữ Thảo Viện Vi
sinh vật và Công nghệ Sinh học, Đại học Quốc gia Hà Nội đã tận tình giúp đỡ, chỉ
bảo, động viên và tạo mọi điều kiện thuận lợi để em hoàn thành tốt luận văn này.
Em cũng xin chân thành cảm ơn TS. Phan Thị Phương Hoa (chủ nhiệm đề
tài Nafosted số 106.03 - 2010.34 “Sàng lọc các chất có hoạt tính sinh học từ xạ
khuẩn dùng cho đấu tranh sinh học và diệt vi khuẩn Xanthomonas oryzae pv.
oryzae gây bệnh bạc lá lúa ở Việt Nam”) và các thành viên tham gia đề tài đã hỗ
trợ cho em kỹ thuật chuyên môn để em hoàn thành luận văn.
Em cũng xin gửi lời cảm ơn Ban lãnh đạo và các anh chị Viện Vi sinh vật và
Công nghệ sinh học đã luôn chia sẻ, giúp đỡ và tạo điều kiện rất lớn trong suốt quá
trình em thực hiện luận văn.
Em xin cảm ơn TS. Kyung Sook Bae, TS. Song Gun Kim và cử nhân
Hyangmi Kim Bảo tàng giống chuẩn (KCTC), Viện Khoa học và Công nghệ sinh
học Hàn Quốc (KRIBB) đã tạo mọi điều kiện thuận lợi giúp đỡ em trong thời gian
học tập ngắn hạn tại Hàn Quốc để em hoàn thành luận văn.
Em xin cảm ơn TS. Hiroshi Kinoshita trường Đại học Osaka Nhật Bản đã hỗ
trợ và giúp đỡ em trong quá trình nghiên cứu để em hoàn thành luận văn này.

ii
Xin cảm ơn sinh viên Vũ Thị Kim Chi đã hỗ trợ em một số công việc thí
nghiệm trong quá trình nghiên cứu để em hoàn thành luận văn.
Cuối cùng, em xin gửi lời cảm ơn đến gia đình thân yêu và bạn bè đã luôn ở
bên, động viên, giúp đỡ em trong suốt thời gian học tập và thực hiện luận văn.
Hà Nội, ngày 2 tháng 10 năm 2014
Học viên
Nguyễn Thị Vân

iii
MỤC LỤC
MỞ ĐẦU ................................................................................................................ 1
MỤC TIÊU NGHIÊN CỨU .................................................................................... 3
CHƢƠNG I. TỔNG QUAN .................................................................................... 4
1.1. Bệnh bạc lá lúa và tác hại của bệnh bạc lá lúa ................................................... 4
1.1.1. Giới thiệu chung............................................................................................4
1.1.2. Tác nhân gây bệnh bạc lá lúa........................................................................5
1.2. Các cách phòng trừ bệnh bạc lá lúa ................................................................... 7
1.2.1. Sử dụng thuốc hóa học..................................................................................7
1.2.2. Chọn giống lúa kháng bệnh...........................................................................8
1.2.3. Khống chế sinh học.....................................................................................11
1.3. Xạ khuẩn ........................................................................................................ 12
1.3.1. Giới thiệu chung về xạ khuẩn......................................................................12
1.3.2. Khả năng sinh chất kháng sinh của xạ khuẩn..............................................14
1.3.3. Sử dụng xạ khuẩn trong khống chế sinh học...............................................16
1.4. Tình hình nghiên cứu xạ khuẩn trong kiểm soát bệnh bạc lá lúa ..................... 17
CHƢƠNG 2. NGUYÊN LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU .................. 19
2.1. Nguyên liệu và hóa chất .................................................................................. 19
2.1.1 Nguyên liệu..................................................................................................19
2.1.1.1. Các chủng Xoo ................................................................................... 19

iv
2.1.1.2. Các chủng xạ khuẩn ........................................................................... 20
2.1.1.3. Vi sinh vật kiểm định ......................................................................... 20
2.1.2. Hóa chất, dụng cụ và thiết bị.......................................................................20
2.1.2.1. Hóa chất ............................................................................................. 20
2.1.2.2. Dụng cụ và thiết bị ............................................................................. 21
2.1.2.3. Môi trƣờng nghiên cứu ....................................................................... 21
2.2. Phƣơng pháp nghiên cứu ................................................................................ 22
2.2.1. Phƣơng pháp thử hoạt tính kháng vi sinh vật..............................................22
2.2.1.1. Phƣơng pháp thỏi thạch ...................................................................... 22
2.2.1.2. Phƣơng pháp đục lỗ thạch .................................................................. 23
2.2.2. Phân loại bằng đặc điểm hình thái...............................................................23
2.2.2.1. Hình thái khuẩn lạc ............................................................................ 23
2.2.2.2. Hình thái chuỗi sinh bào tử và bề mặt bào tử ...................................... 23
2.2.3. Hóa phân loại...............................................................................................24
2.2.3.1. Phân tích thành phần axit amin trong thành tế bào .............................. 24
2.2.3.2. Phân tích thành phần menaquinone .................................................... 24
2.2.3.3. Phân tích thành phần axit béo ............................................................. 25
2.2.3.4. Phân tích thành phần G+C trong ADN ............................................... 25
2.2.4. Phân loại sinh học phân tử giải trình tự 16S - rADN..................................25
2.2.5. Tối ƣu hóa điều kiện nuôi cấy xạ khuẩn......................................................27
2.2.5.1. Lựa chọn môi trƣờng nuôi cấy thích hợp ............................................ 27
2.2.5.2. Lựa chọn thời gian nuôi cấy thích hợp ................................................ 27
2.2.5.3. Lựa chọn thể tích nuôi cấy thích hợp .................................................. 28
2.2.5.4. Lựa chọn nhiệt độ nuôi cấy ................................................................ 28

v
2.2.5.5. Lựa chọn cách cấy giống .................................................................... 28
2.2.6. Tinh sạch hoạt chất kháng Xoo...................................................................28
2.2.6.1. Tách dịch chiết thô ............................................................................. 28
2.2.6.2. Tinh sạch bằng silica gel .................................................................... 28
2.2.6.3. Tinh sạch bằng HPLC ........................................................................ 29
2.2.7. Xác định khối lƣợng phân tử bằng khối phổ (MS)......................................29
CHƢƠNG 3. KẾT QUẢ VÀ THẢO LUẬN.......................................................... 31
3.1. Khả năng kháng vi sinh vật của chủng xạ khuẩn VN08 - A12. ........................ 31
3.2. Phân loại chủng VN08 - A12 .......................................................................... 32
3.2.1. Phân loại bằng hình thái..............................................................................32
3.2.2. Hóa phân loại..............................................................................................33
3.2.2.1. Thành phần axit amin trong thành tế bào ............................................ 33
3.2.2.2. Thành phần menaquinone ................................................................... 34
3.2.2.3.Thành phần axit béo ............................................................................ 35
3.2.2.4. Thành phần G + C trong ADN............................................................ 37
3.2.3. Trình tự 16S rADN......................................................................................38
3.3. Tối ƣu hóa điều kiện nuôi cấy của chủng VN08 - A12 .................................... 39
3.3.1. Môi trƣờng nuôi cấy thích hợp....................................................................39
3.3.2. Thời gian nuôi cấy thích hợp.......................................................................40
3.3.3. Thể tích nuôi cấy thích hợp.........................................................................41
3.3.4. Nhiệt độ nuôi cấy thích hợp........................................................................41
3.3.5. Cách cấy giống thích hợp............................................................................42
3.4. Tinh sạch và phân tích hoạt chất kháng Xoo của chủng VN08 - A12 .............. 43
3.4.1. Tinh sạch hoạt chất kháng Xoo của chủng VN08 - A12.............................43

vi
3.4.2. Phân tích trọng lƣợng phân tử.....................................................................47
KẾT LUẬN VÀ KIẾN NGHỊ ............................................................................... 51
4.1. Kết luận .......................................................................................................... 51
4.2. Kiến nghị ........................................................................................................ 51
TÀI LIỆU THAM KHẢO ..................................................................................... 52
PHỤ LỤC........................................................................................................... - 1 -

vii
DANH MỤC CÁC BẢNG
Bảng 1. Đặc điểm và nguồn gốc của các gen Xa kháng bệnh bạc lá lúa [37] ............ 8
Bảng 2. Trình tự các đoạn mồi đặc hiệu để phát hiện gen Xa trên lúa [15] ............... 9
Bảng 3. Danh sách 10 chủng Xoo phân lập đƣợc ở miền Bắc Việt Nam đƣợc sử
dụng trong nghiên cứu ........................................................................................... 19
Bảng 4. Chƣơng trình phân tích HPLC .................................................................. 29
Bảng 5. Hoạt tính kháng 10 chủng Xoo của chủng xạ khuẩn VN08 - A12 ............. 31
Bảng 6. Hoạt tính kháng vi sinh vật của chủng xạ khuẩn VN08 - A12 ................... 31
Bảng 7. Hoạt tính kháng vi sinh vật có lợi của chủng xạ khuẩn VN08 - A12 ......... 32
Bảng 8 : Kết quả phân tích thành phần menaquinone của chủng VN08 - A12 ........ 35
Bảng 9. Kết quả phân tích thành phần menaquinone của chủng VN08 - A12 ......... 36
Bảng 10. Kết quả phân tích thành phần GC của chủng VN08 - A12 ...................... 38
Bảng 11. Hoạt chất của VN08 - A12 thu đƣợc từ các phân đoạn khi tinh sạch bằng
silica gel ................................................................................................................ 44
Bảng 12. Hoạt chất của VN08 - A12 thu đƣợc từ HPLC từ phân đoạn 4.1 ............. 45
Bảng 13. Chƣơng trình chạy HPLC phân tích thành phần axit amin ................... - 1 -
Bảng 14. Phân tích thành phần menaquinone chuẩn........................................... - 2 -
Bảng 15. Phân tích thành phần axit béo chuẩn .................................................... - 3 -
Bảng 16. Trình tự mồi dùng cho phản ứng đọc trình tự ADNr 16S ..................... - 5 -

viii
DANH MỤC CÁC HÌNH
Hình 1. Lúa bị nhiễm bệnh bạc lá do vi khuẩn Xoo. ................................................. 5
Hình 2. Khuẩn lạc và tế bào của vi khuẩn Xoo [9] ................................................... 6
Hình 3. Tổn thƣơng ít trên lá của giống lúa IRBB21 chứa gen Xa21 do bệnh bạc lá
lúa ở Ấn Độ [19] ................................................................................................... 10
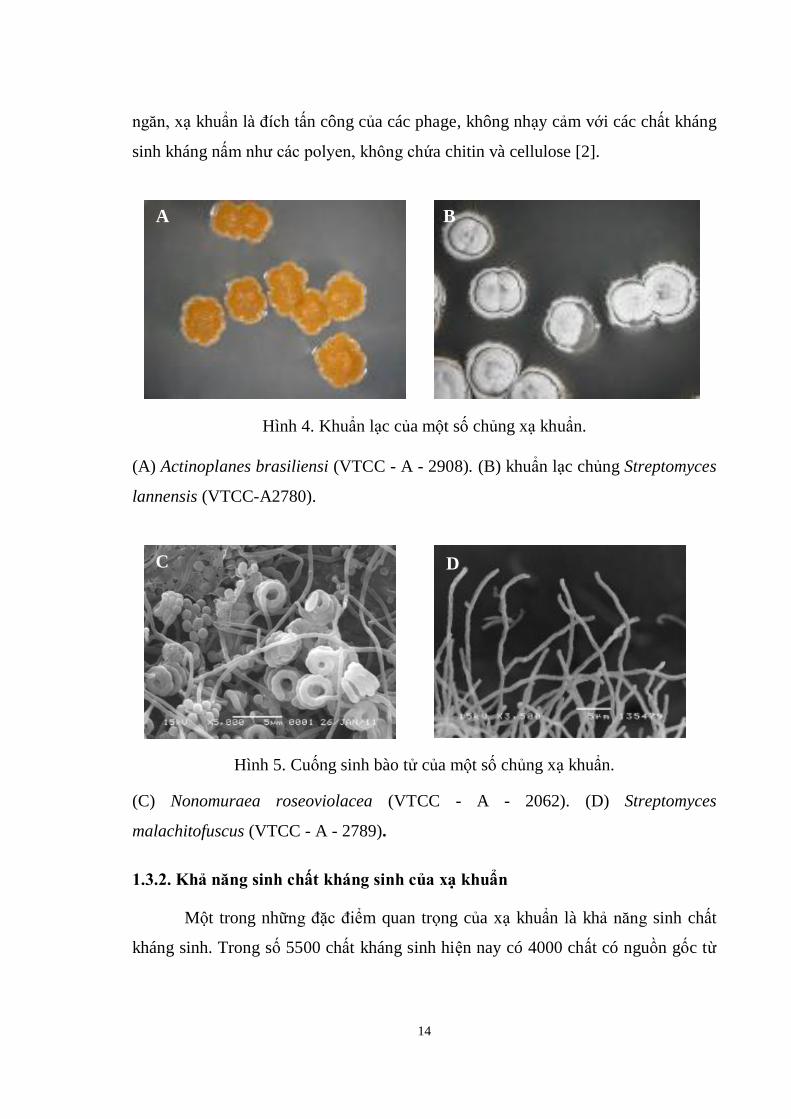
Hình 4. Khuẩn lạc của một số chủng xạ khuẩn. ..................................................... 14
Hình 5. Cuống sinh bào tử của một số chủng xạ khuẩn. ......................................... 14
Hình 6. Bản đồ phân bố của 10 chủng Xoo ở miền Bắc Việt Nam đƣợc sử dụng
trong nghiên cứu .................................................................................................... 20
Hình 7. Khuẩn lạc và chuỗi bào tử của chủng VN08 - A12 .................................... 33
Hình 8. Chuỗi bào tử và bào tử của chủng VN08 - A12 ......................................... 33
Hình 9. Kết quả chạy sắc ký bản mỏng thành phần axit amin trong thành tế bào của
chủng VN08 - A12. ............................................................................................... 34
Hình 10. Kết quả phân tích LC thành phần menaquinone của chủng VN08 - A12. 35
Hình 11. Sắc ký đồ thành phần axit béo của chủng VN08 - A12 ............................ 36
Hình 12. Sắc ký đồ thành phần GC của chủng VN08 - A12 ................................... 37
Hình 13. Cây phân loại chủng VN08 - A12 dựa trên trình tự gen 16S – rADN. ..... 39
Hình 14. Ảnh hƣởng của môi trƣờng nuôi cấy khác nhau đến sự sản sinh hợp chất
kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12 ....................................... 40
Hình 15. Ảnh hƣởng của thời gian cấy khác nhau đến sự sản sinh hợp chất kháng
sinh ức chế vi khuẩn Xoo của chủng VN08 - A12 .................................................. 40
Hình 16. Ảnh hƣởng của thể tích môi trƣờng nuôi cấy khác nhau đến sự sản sinh
hợp chất kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12 (S. toxytricini) .. 41
Hình 17. Ảnh hƣởng của nhiệt độ môi trƣờng nuôi cấy khác nhau đến sự sản sinh
hợp chất kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12 ......................... 42
Hình 18. Ảnh hƣởng của cách cấy giống đến sự sản sinh hợp chất kháng sinh ức chế
vi khuẩn Xoo của chủng VN08 - A12. ................................................................... 43
Hình 19. Quy trình tách dịch chiết thô của chủng VN08 - A12 .............................. 44
Hình 20. Phổ HPLC của hoạt chất chủng VN08 - A12........................................... 46

ix
Hình 21. Phổ HPLC của hoạt chất sau tinh sạch .................................................... 47
Hình 22. Phổ hấp thụ UV của hoạt chất sau tinh sạch ............................................ 47
Hình 23. Kết quả phân tích khối phổ của chủng VN08 - A12 ................................ 48
Hình 24. Cấu trúc hóa học của factumycin [31] ..................................................... 48
Hình 25. Cấu trúc của factumycin [46] .................................................................. 49
Hình 26. Cụm gen sinh tổng hợp factumycin (A) WAC5292 (Streptomyces
toxytricini) và (B) Streptomyces cattleya………………………………………………..49
Hình 27. Sắc ký đồ thành phần menaquinone chuẩn ........................................... - 2 -
Hình 28. Sắc ký đồ thành phần axit béo chuẩn .................................................... - 4 -
Hình 29. Hoạt tính kháng Xoo của VN08 - A12 trong các điều kiện nuôi cấy khác
nhau ................................................................................................................... - 7 -
Hình 30. Hoạt chất kháng sinh của chủng VN08 - A12 ....................................... - 7 -

x
TỪ VIẾT TẮT
ADN Axit deoxyribonucleic
Ala Alanine
DAP Diaminopimelic
DW Distilled Water (Nƣớc cất)
ĐHQGHN Đại học Quốc Gia Hà Nội
GC Gas Chromatography (Sắc ký khí)
Glu Glutamic acid
Gly Glycine
HPLC High Performance Liquid Chromatography (Sắc ký lỏng hiệu năng cao)
Lys Lysine
MQ Milli- Q water
MS Mass Spectrometry ( Phƣơng pháp khối phổ)
MT Môi trƣờng
SKS Sinh kháng sinh
TLC Thin- layer Chromatography (Sắc ký lớp mỏng)
VTCC Vietnam Type Culture Collection (Bảo tàng giống chuẩn Vi sinh vật)
Xoo Xanthomonas oryzae pv. Oryzae

1
MỞ ĐẦU
Bệnh bạc lá lúa do vi khuẩn Xanthomonas oryzae pv. oryzae (Xoo) gây ra,
xuất hiện khá phổ biến trên các khu vực trồng lúa ở Việt Nam, đặc biệt tập trung ở
khu vực đồng bằng sông Cửu Long. Bệnh gây hại trong vụ lúa hè thu nhiều hơn
trong vụ đông xuân do thời tiết ẩm ƣớt, nhiều sƣơng mù, độ ẩm không khí cao.
Bệnh xuất hiện và gây hại từ giai đoạn lúa đẻ nhánh đến đòng trổ và chín. Bệnh bạc
lá lúa gây thiệt hại rất nặng nề cho ngành nông nghiệp, làm giảm năng suất lúa 25 -
50 %, thậm chí mất trắng (Số liệu thống kê của cục bảo vệ thực vật). Do đó, việc
kiểm soát và ngăn chặn sự bùng phát của bệnh bạc lá lúa là vấn đề rất quan trọng và
là mối quan tâm của nhiều nhà nghiên cứu khoa học. Một trong những giải pháp
quan trọng nhất phòng trừ bệnh bạc lá lúa hiện nay là sử dụng các dòng lúa mang
gen kháng bệnh. Tuy nhiên, các dòng lúa chỉ mang một vài gen kháng đơn lẻ, đƣợc
nuôi trồng liên tục trên diện rộng dẫn đến tình trạng các chủng Xoo có khả năng gây
bệnh ngay cả khi có các gen kháng đó [18]. Chính vì vậy, việc kiểm soát bệnh bạc
lá lúa bằng biện pháp sinh học đang thu hút đƣợc sự chú ý của nhiều nhà nghiên
cứu [36]. Một trong những biện pháp sinh học đó là việc sử dụng xạ khuẩn để
khống chế vi khuẩn gây bệnh, đây là biện pháp có triển vọng cao, an toàn với môi
trƣờng và có hiệu quả về kinh tế.
Xạ khuẩn là nhóm vi khuẩn Gram dƣơng, đƣợc biết đến nhƣ nguồn sinh các
chất kháng sinh và các chất có hoạt tính sinh học. Trong số khoảng hơn 10000 loại
thuốc kháng sinh đƣợc phát hiện do vi sinh vật thì có đến hai phần ba đƣợc phân lập
từ xạ khuẩn mà chủ yếu thuộc chi Streptomyces; và nhiều chất trong đó là các thuốc
kháng sinh đang đƣợc ứng dụng và sản xuất rộng rãi hiện nay [6]. Ngoài ra xạ
khuẩn còn là nguồn sinh các loại vitamin, enzym, chất ức chế miễn dịch, hocmôn
sinh trƣởng. Số loài xạ khuẩn trong tự nhiên ƣớc tính lớn hơn rất nhiều so với con
số 1000 loài đƣợc công bố. Đồng thời xạ khuẩn luôn là nguồn vi sinh vật rất giá trị
trong việc tìm kiếm các chất có hoạt tính sinh học mới. Bộ sƣu tập hơn 3000 chủng

2
xạ khuẩn đang đƣợc bảo quản tại Bảo tàng giống chuẩn, Viện Vi Sinh và Công nghệ
sinh học, ĐHQGHN hứa hẹn tiềm năng tìm ra các chất có hoạt tính sinh học mới
dùng cho việc khống chế và tiêu diệt vi khuẩn Xoo gây bệnh bạc lá lúa ở Việt Nam
hiện nay. Trong nghiên cứu này, đƣợc sự tài trợ của quỹ đề tài NAFOSTED, TS.
Phan Thị Phƣơng Hoa và các cộng sự đã sàng lọc 2690 chủng xạ khuẩn và lựa chọn
đƣợc 17 chủng có khả năng kháng tất cả 10 chủng Xoo. Trong đó, lựa chọn chủng
VN08 - A12 là một tác nhân kiểm soát sinh học của bệnh bạc lá lúa. Chủng VN08 -
A12 có nhiều ƣu điểm nổi bật đã đƣợc thử nghiệm ngoài đồng ruộng và thu đƣợc
kết quả rất đáng mừng. Kết quả cho thấy chủng VN08-A12 không chỉ có thể làm
giảm chiều dài tổn thƣơng của cây lúa do nhiễm Xoo, mà còn làm giảm đáng kể tổn
thất năng suất của giống lúa bị nhiễm Xoo [23] [24]. Chính vì vậy, để phát triển một
chế phẩm khống chế sinh học để kiểm soát bệnh bạc lá lúa, chúng tôi đã tiến hành
thực hiện đề tài “Nghiên cứu đặc điểm sinh học chủng xạ khuẩn Streptomyces
toxytricini (VN08 - A12) kháng bệnh bạc lá lúa do Xanthomonas oryzae”.

3
MỤC TIÊU NGHIÊN CỨU
Nghiên cứu này đƣợc thực hiện nhằm thực hiện các mục tiêu cụ thể nhƣ sau:
Phân loại chủng xạ khuẩn VN08 - A12.
Lựa chọn điều kiện nuôi cấy tối ƣu nhất để khả năng sinh hoạt chất của
chủng xạ khuẩn là cao nhất.
Tinh sạch và xác định cấu trúc của hoạt chất kháng Xoo sinh ra bởi chủng xạ
khuẩn VN08 - A12.

4
CHƢƠNG I. TỔNG QUAN
1.1. Bệnh bạc lá lúa và tác hại của bệnh bạc lá lúa
1.1.1. Giới thiệu chung
Bệnh bạc lá lúa - hay còn gọi là bệnh cháy bìa lá lúa (Bacterial leaf blight
disease) là một trong những bệnh phổ biến trong các nƣớc trồng lúa. Bệnh bạc lá lúa
đƣợc phát hiện lần đầu ở vùng Fukuoko, Kyushu, Nhật Bản ngay từ những năm
1884 [45]. Hiện nay, bệnh bạc lá đã xảy ra trên rất nhiều quốc gia, đặc biệt ở châu
Á. Ở nhiều nƣớc châu Á, căn bệnh này đã trở thành đại dịch trên lúa. Bệnh có thể
làm giảm sản lƣợng ở mức độ khác nhau, tùy thuộc vào giai đoạn nhiễm bệnh của
cây, mức độ nhạy cảm của giống lúa và ảnh hƣởng của môi trƣờng. Ở một số nƣớc
châu Á và Đông Nam Á, bệnh bạc lá lúa thƣờng làm giảm năng suất 10 - 20 %
nhƣng có thể lên đến 50% [33] [38]. Ở Nhật Bản, thiệt hại năng suất ƣớc tính là 20 -
30% thậm chí đến 50%. Ở vùng nhiệt đới, bệnh phá hoại gây ảnh hƣởng nặng tới
hàng triệu ha lúa. Ở Ấn Độ, thiệt hại về sản lƣợng từ 6 đến 60%. Việc giảm sản
lƣợng chủ yếu là do giảm số lƣợng bông và trọng lƣợng hạt [43].
Ở Việt Nam, tuy khả năng lây lan thành dịch của bệnh bạc lá lúa không cao
nhƣ các bệnh dịch hại khác nhƣ rầy nâu, đạo ôn, hậu quả do bệnh bạc lá lúa gây ra
không hề thua kém những dịch hại khác. Theo số liệu thống kê của cục Bảo vệ Thực
vật, từ năm 1999 - 2003 diện tích lúa bị hại do bệnh bạc lá gây ra trong cả nƣớc là
108.691,4 ha (miền Bắc là 86.429,2 ha; miền Nam là 22.262,2 ha), trong đó diện tích
bị hại nặng nhất là 156,76 ha và diện tích mất trắng là 80 ha. Cho đến năm 2013 diện
tích lúa nhiễm bệnh bạc lá là 135,4 nghìn ha, tăng gấp 6,5 lần so với năm 2010. Từ
năm 2010 đến năm 2011 diện tích cây mắc bệnh tăng nhẹ và đột ngột tăng cao trong
giai đoạn từ năm 2011 đến năm 2012. Diện tích lúa bị mắc bệnh năm 2012 là 90,543
nghìn ha, cao gấp gần 4 lần so với năm 2011 (26 nghìn ha) đến năm 2013 diện tích
lúa bị mắc bệnh đã tăng đến 135,4 nghìn ha. Trong đó diện tích lúa bị nhiễm nặng là
hơn 9,5 ngàn ha. Tình trạng mất trắng vẫn đang diễn ra và tăng cao trong các năm trở

5
lại đây tập trung nhiều tại các tỉnh phía bắc và một số tỉnh vùng đồng bằng sông Cửu
Long. Tuy nhiên, kết quả khảo sát mới đây của chi cục cho thấy, vụ hè thu 2014, lần
đầu tiên bệnh này đã xuất hiện tập trung ở cánh đồng huyện Đạ Tẻh , Lâm Đồng và
diện tích nhiễm bệnh lên đến 25 ha và tỷ lệ gây hại từ 50% - 80% (cao hơn nhiều so
với bình quân chung cả nƣớc).
Bệnh bạc lá phát sinh trong suốt thời kỳ lúa chín nhƣng các triệu chứng bệnh
điển hình thƣờng xuất hiện từ thời kỳ đẻ nhánh đến thời kỳ trổ và chín, đạt đỉnh ở
giai đoạn ra hoa [33]. Bệnh bạc lá lúa đƣợc chia làm hai giai đoạn: giai đoạn
“Kresek” và giai đoạn cháy lá. Kresek là giai đoạn phá hoại nghiêm trọng nhất, toàn
bộ lá của cây chuyển màu vàng nhạt và héo. Nếu xảy ra trong thời kỳ đẻ nhánh sớm
sẽ dẫn đến mất mùa một phần hoặc hoàn toàn. Cây con dƣới 21 ngày tuổi dễ bị
kresek ở nhiệt độ từ 28oC đến 34
oC [5]. Chính vì vậy, bệnh lây lan dễ dàng hơn ở
vùng khí hậu ấm áp và ẩm. Trong giai đoạn cháy lá, lá có màu vàng đặc trƣng với
gợn sóng trên phiến lá (hình 1).
Hình 1. Lúa bị nhiễm bệnh bạc lá do vi khuẩn Xoo.
(www.thaibinhseed.com.vn/benh-bac-la-lua)
1.1.2. Tác nhân gây bệnh bạc lá lúa
Tác nhân gây bệnh bạc lá lúa là vi khuẩn Xanthomonas oryzae pv. oryzae
(Xoo). Vi khuẩn này có tế bào hình que với đầu tròn, với chiều dài từ 0,8 đến 1 µm,

6
chiều rộng 0,4 đến 0,7 μm, bao quanh tế bào là một màng nhầy. Đây là loại vi
khuẩn Gram âm và không sinh bào tử, khuẩn lạc hình tròn, lồi, bề mặt nhẵn, màu
vàng (hình 2) [9]. Vi khuẩn này chủ yếu xâm nhập vào hệ thống mạch dẫn rồi dẫn
đến nhiễm trùng toàn thân cây lúa . Ngoài ra vi khuẩn còn có thể xâm nhập qua lỗ
thùy khổng ở mép lá, đầu mút lá dễ dàng gây tổn thƣơng dọc theo gân lá [39].
Hình 2. Khuẩn lạc và tế bào của vi khuẩn Xoo [9]
Loại vi khuẩn này đầu tiên đƣợc các nhà khoa học Nhật Bản, Hori và Bokura
đặt tên là Bacillus oryzae từ năm 1911 và đƣợc đổi tên nhiều lần [33]. Sau đó, do
phát hiện là tác nhân gây bệnh bạc lá lúa, loài vi khuẩn này đƣợc đổi tên thành
Xanthomonas campestris pv. oryzae để phân biệt với các tác nhân gây bệnh sọc lá
X. campestris pv. oryzicola [51]. Năm 1990, Swings phát hiện ra vi khuẩn gây bệnh
bạc lá và bệnh sọc lá khác biệt với các vi khuẩn gây bệnh X. campestris khác nên
phân loại lại thành một loài riêng biệt X. oryzae, bao gồm pv. oryzae và pv.
oryzicola. Bằng các phân tích giải trình tự DNA, protein và phân tích thành phần
acid béo, vị trí phân loại của Xanthomonas oryzae đã đƣợc khẳng định và công nhận
đến ngày nay [49] [50].
Xoo là một loại vi khuẩn Gram âm, màu vàng, sản sinh lƣợng lớn
polysaccharide ngoại bào (EPS). Các chủng đột biến thiếu EPS không có khả năng
gây bệnh bạc lá lúa [44]. Xoo chỉ có thể tồn tại trong đất từ một đến hai tháng, ngoài
ra Xoo có thể tồn tại trong hạt giống của các cây lúa bị nhiễm, hay trong rơm rạ. Vi
5µm

7
khuẩn này sẽ đƣợc kích hoạt khi gặp độ ẩm thích hợp và sinh trƣởng trong gốc rạ
cũng nhƣ trong một số loại cỏ nhƣ Leersia sp.. Giả thuyết tác nhân gây bệnh là từ
giống lúa hay từ ADN của hạt giống nhiễm bệnh đã bị loại bỏ [16] [48]. Các vi
khuẩn gây bệnh bạc lá xâm nhập vào cây qua các vết thƣơng xây xát ở trên lá do
mƣa bão gây ra và qua các lỗ khí khổng, sau đó sẽ gây bệnh trên các mô và từ đó sẽ
nhân và lây lan toàn bộ cây dẫn đến nhiễm trùng toàn thân [26]. Vi khuẩn lây lan
thông qua nƣớc tƣới, mƣa và gió. Nƣớc tƣới cũng đƣợc coi là tác nhân đóng góp lớn
vào sự lây lan của bệnh này trên diện rộng đất canh tác. Tuy nhiên, vi khuẩn chỉ tồn
tại đƣợc trong nƣớc dƣới 15 ngày [45].
1.2. Các cách phòng trừ bệnh bạc lá lúa
Bệnh bạc lá lúa gây tổn thất nặng nề và ảnh hƣởng nghiêm trọng đến sản
xuất lúa gạo trên toàn thế giới, do vậy đòi hỏi phải có chiến lƣợc quản lý nhằm
chống lại sự bùng phát của dịch bệnh. Bƣớc đầu cần thực hiện để kiểm soát bệnh
bạc lá lúa là làm giảm tác nhân gây bệnh và ngăn chặn phát triển của tác nhân gây
bệnh trên cây chủ. Điều này có thể đƣợc thực hiện thông qua việc sử dụng các hóa
chất, giống kháng bệnh, và tác nhân sinh học.
1.2.1. Sử dụng thuốc hóa học
Thuốc hóa học đƣợc sử dụng để giết chết hoặc ức chế sự nhân lên của các tác
nhân gây bệnh bằng cách ngăn chặn con đƣờng trao đổi chất của vi khuẩn. Một số
hóa chất đƣợc sử dụng để kiểm soát bệnh bạc lá lúa nhƣ Bordeaux có hoặc không
có đƣờng, hỗn hợp đồng xà phòng, thuốc diệt nấm và đồng thủy ngân, dịch phun
oxychloride. Ở Ấn Độ, sử dụng bột Clo trong xử lý nƣớc cũng làm giảm bệnh [11].
Một số chất hữu cơ tổng hợp diệt khuẩn cũng đã đƣợc sử dụng nhƣ niken dimethyl
dithiocarbamate, dithianone, phenazine và phenazine N- oxit. Ngoài ra, thuốc diệt
nấm dithiocarbamate cũng ức chế sự phát triển của Xoo bằng cách ngăn chặn quá
trình sinh tổng hợp acid béo và lipid [54]. Một vài loại thuốc kháng sinh nhƣ
streptocycline và các thuốc diệt nấm nhƣ zineb, carbendazim cũng đƣợc chứng
minh là có khả năng ức chế vi khuẩn gây bệnh in vitro. Một số chất diệt khuẩn nhƣ

8
kasugamycin, phenazin và streptomycin có thể ngăn chặn đƣợc các vi khuẩn gây
bạc lá lúa, nhƣng lại có nhƣợc điểm là giá thành đắt và không thân thiện với môi
trƣờng. Việc sử dụng các thuốc hóa học luôn mang lại hậu quả rất nặng nề đối với
môi trƣờng. Đồng thời, cũng chƣa có phƣơng thức kiểm soát bằng hóa chất nào
mang lại hiệu quả cao bởi vì khả năng tồn tại và phát triển của các chủng kháng
thuốc.
1.2.2. Chọn giống lúa kháng bệnh
Chọn giống lúa kháng bệnh hiện nay cũng là phƣơng pháp đang đƣợc quan
tâm. Hiện nay, 23 gen kháng bệnh bạc lá lúa (Xa) đã đƣợc công bố (bảng 1). Các
đoạn mồi cũng đã đƣợc thiết kế để phát hiện các gen Xa trên các giống lúa khác
nhau (bảng 2).
Bảng 1. Đặc điểm và nguồn gốc của các gen Xa kháng bệnh bạc lá lúa [37]
Tên gen Xa
mới
Tên gen Xa
cũ
Nhiễm sắc thể Giống, dòng lúa đại diện
Xa1 Xa1 NST 4 Kogyoku
Xa1-h Xa-1h NST 4 IR28, IR29, IR30
Xa2 Xa2 NST 4 Rentai Emas2, tẻ tép
Xa3 Xa-w NST11 Wase Aikoku3
Xa-4b NST11 Semora Mangga
Xa-6 NST-5 Zenith
Xa-9 NST-6 Sateng
Xa4 Xa-4 NST11 TKM6, IR20, UR22
Xa5 Xa-5 NST 4 DZ192,IR1545-339
Xa7 Xa-7 NST 4 DV85, DZ78
Xa8 Xa-8 NST 4 PI231129
Xa10 Xa-10 NST11 Cas 209
Xa11 Xa-11 RP 9-3, IR8

9
Tên gen Xa
mới
Tên gen Xa
cũ
Nhiễm sắc thể Giống, dòng lúa đại diện
Xa12 Xa-kg NST4 Kogyoku, Java14
Xa12-h Xa-kgh NST 4 IR28, IR29, IR30
Xa13 Xa-13 NST 5 BJ1, Chinsura Boro II
Xa14 Xa-14 Tai chung Native 1
Xa15 (t) Xa-nm (t) M41
Xa16 Xa-16 Tẻ tép
Xa17 Xa-as(t) NST 4 Asominori
Xa18 Xa-18 IR24, Milyang 23
Xa19 Xa19 Đột biến XM5
Xa20 Xa-20 Đột biến M6
Xa21 Xa-21 NST 11 IRBB21
Xa22 (t)b Xa-22 (t) NST 11 Zachanglong
Xa23 (t)b Xa-23 (t) NST 11 WBB1
Bảng 2. Trình tự các đoạn mồi đặc hiệu để phát hiện gen Xa trên lúa [15]
Mồi Trình tự nucleotide Xa-gen
MP1 5'-ATC-GAT-CGA-TCT-TCA-CGA-GG-3' Xa-4
MP2 5'-dTG-CTA-TAA-AAG-GCA-TTC-GGG-3'
RG556 F 5'-TAG-CTG-CTG-CCG-TGC-TGT-GC-3' Xa-5
RG556 R 5'-AAT-ATT-TCA-GTG-TGC-ATC-3'
P3 F 5'-CAG-CAA-TTC-ACT-GGA-GTA-GTG-GTT-3' Xa-7
P3 R 5'-CAT-CAC-GGT-CAC-CGC-CAT-ATC-GGA-3'
Xa21F 5'-ATA-GCA-ACT-GAT-TGC-TTT-GC-3' Xa-21
Xa 21 R 5'- CGA - TCG - GTA - TAA- CAG-CAA-AAC-3'
RG136 F 5' - TCC-CAG-AAA-GCT - ACA-GC-3' Xa-13
RG136 R 5' - GCA-GAC-TCC-AGT=TTG-ACT-TC-3'

10
Gen Xa21 lần đầu tiên đƣợc phát hiện trong một giống lúa hoang dại Oryza
longistaminata và đã dƣợc chuyển vào giống lúa IR24 để tạo ra giống lúa kháng
bệnh IRBB21 [30]. Ở Ấn Độ và Philippin, giống lúa này đã đƣợc chứng minh là có
khả năng kháng với hầu hết các chủng Xoo. Tuy nhiên, cho đến nay một số chủng
Xoo ở một số vùng của châu Á đã đƣợc phát hiện là có thể vƣợt qua đƣợc gen
kháng Xa21 trong giống lúa IRBB21 (hình 3) [18]
Hình 3. Tổn thƣơng ít trên lá của giống lúa IRBB21 chứa gen Xa21 do bệnh bạc lá
lúa ở Ấn Độ [19]
Các nghiên cứu về bệnh Xoo cuối những năm 2000 đã đƣợc phát triển lên từ
việc phân lập, chuẩn đoán và định dạng bằng hình thái vi khuẩn, lên mức sử dụng
các kĩ thuật phân tử (ví dụ nhƣ chuẩn đoán bệnh nhanh bằng PCR, giải trình tự các
gen kháng bệnh Xoo trên lúa, dùng phƣơng pháp đánh dấu gen để chuyển gen kháng
vào các dòng lúa) [3] [47]. Các nhà khoa học Việt Nam kết hợp với Viện nghiên
cứu lúa quốc tế (IRRI) của Nhật Bản và xác định các gen kháng Xa7, Xa21 (trội) và

11
Xa5 (lặn) có tính kháng cao đối với hầu hết các dòng vi khuẩn Xoo gây bệnh bạc lá
lúa ở các tỉnh phía Bắc [47], đồng thời xác định các dạng bệnh gây ra bởi các chủng
Xoo đã phân lập đƣợc từ các mẫu lúa trồng tại đồng bằng sông Cửu Long [13].
Tuy nhiên, việc sử dụng liên tục và trên diện rộng các gen kháng bệnh đơn lẻ
đã dẫn đến sự chọn lọc các dòng vi khuẩn gây bệnh có khả năng phá vỡ sức kháng
bệnh của cây. Vì thế, việc chuyển tổ hợp các gen kháng đƣợc cho là giải pháp lâu
dài cho việc làm chậm lại sự xuất hiện các dòng vi khuẩn gây bệnh miễn dịch với
các dòng lúa có gen kháng bệnh. Các gen khác nhau kháng các dòng, các chủng, các
dạng sinh học của vi khuẩn gây bệnh khác nhau. Tổ hợp các gen đó làm rộng phổ
tác động kháng lại sự đa dạng của các chủng Xoo gây bệnh. Hơn nữa, bằng cách tổ
hợp các gen chính và các gen phụ kháng bệnh bạc lá lúa sẽ làm kéo dài sức kháng
bệnh ở cây lúa [8]. Do vậy, các nghiên cứu hiện nay đang tìm cách chuyển nhiều
gen kháng vào một giống lúa tạo tính kháng ổn định cao.
1.2.3. Khống chế sinh học
Các thuốc hóa học đã đƣợc chứng minh là có hiệu quả trong việc kiểm soát
bệnh bạc lá lúa nhƣng không đƣợc ứng dụng rộng vì những biến đổi khó lƣờng của
các tác nhân gây bệnh. Sự xuất hiện các quần thể kháng thuốc đặt ra mối đe dọa
nghiêm trọng cho chiến lƣợc kiểm soát hóa học lâu dài. Đồng thời, các chất hóa học
thƣờng rất độc hại đối với ngƣời sử dụng, gây ảnh hƣởng đến các loài sinh vật khác,
và tích lũy qua thời gian dài sẽ gây ảnh hƣởng trầm trọng lên hệ sinh thái. Việc
chọn tạo giống kháng bệnh cũng đã đƣợc áp dụng nhƣng các nhóm quần thể kháng
lại gen kháng bệnh cũng phát triển nhanh chóng. Do đó, kiểm soát sinh học đƣợc
cho là một giải pháp sinh thái có hiệu quả cho việc ngăn ngừa bệnh bạc lá lúa. Vì
thế khống chế sinh học kết hợp với các biện pháp kể trên có thể là giải pháp tốt hơn
để điều trị bệnh bạc lá lúa. Islam và Bora (1998) đã đƣa ra biện pháp phòng trừ
bệnh trên lúa bằng việc sử dụng 2 chủng vi khuẩn Rhizobacterial vào việc khử
trùng hạt giống. Kết quả cho thấy việc xử lý hạt giống không chỉ có tác dụng làm
giảm ảnh hƣởng của bệnh bạc lá mà còn có tác dụng làm tăng năng suất lúa [27].

12
Ngoài ra, các nghiên cứu của Gnanamanickam và các cộng sự của ông ở Ấn
Độ và Philipin đã tìm thấy chủng Pseudomonas fluorescens và một số chủng
Bacillus đƣợc phân lập từ các mẫu vùng rễ lúa, có khả năng ức chế sự phát triển của
Xoo trong phòng thí nghiệm [17]. Năm 2008, Ji và cộng sự công bố chủng
Lysobacter antibioticus đƣợc phân lập từ rễ cây lúa ở tỉnh Yunnan, Trung Quốc, có
khả năng ức chế sự phát triển của nhiều loại nấm và vi khuẩn gây bệnh thực vật,
trong đó có Xoo [29].
1.3. Xạ khuẩn
1.3.1. Giới thiệu chung về xạ khuẩn
Xạ khuẩn là nhóm vi khuẩn Gram dƣơng, thƣờng có tỷ lệ GC trong ADN cao
hơn 55%. Trong số khoảng 1000 chi và 5000 loài sinh vật nhân sơ đã công bố có
khoảng 100 chi và 1000 loài xạ khuẩn [12]. Xạ khuẩn thuộc về lớp Actinobacteria,
bộ Actinomycetales, 10 dƣới bộ, 35 họ, 110 chi và 1000 loài. Xạ khuẩn phân bố chủ
yếu trong đất và đóng vai trò rất quan trọng trong chu trình tuần hoàn vật chất trong
tự nhiên. Chúng sử dụng axit humic và các chất hữu cơ khó phân giải khác trong
đất. Trong đất xạ khuẩn chiếm từ 9% - 45% tổng số vi sinh vật, số lƣợng trung bình
khoảng 106 - 10
8 tế bào/ g đất. Số lƣợng xạ khuẩn trong đất không chỉ phụ thuộc
vào loại đất mà còn phụ thuộc vào mức độ canh tác của đất và khả năng bao phủ
của thực vật. Đất giàu chất dinh dƣỡng hữu cơ, khoáng và lớp đất trên bề mặt (đến
40 cm) thƣờng có số lƣợng xạ khuẩn lớn. Trong 1 g đất canh tác có thể có tới 5 x
106 tế bào xạ khuẩn, trong khi đó đất vùng sa mạc, nóng, khô, độ ẩm thấp, nghèo
chất dinh dƣỡng, có số lƣợng xạ khuẩn thấp hơn 10 - 100 lần, dao động trong
khoảng 104 - 10
5 tế bào/ 1 g đất. Sự phân bố của xạ khuẩn trong đất còn phụ thuộc
nhiều vào độ pH của môi trƣờng, thƣờng có nhiều trong lớp đất trung tính và kiềm
yếu hoặc axit yếu, trong khoảng pH 6,0 - 8,0. Xạ khuẩn không có nhiều trong lớp
đất kiềm hay axit và càng hiếm trong các lớp đất rất kiềm. Số lƣợng xạ khuẩn trong
đất cũng thay đổi theo thời gian trong năm [4].

13
Khuẩn lạc của xạ khuẩn không trơn ƣớt nhƣ ở vi khuẩn và ở nấm men mà
thƣờng rắn chắc, thô ráp, dạng vôi, dạng nhung tơ, hay dạng mang dẻo, không trong
suốt. Kích thƣớc khuẩn lạc thay đổi tuỳ loại xạ khuẩn và tuỳ điều kiện nuôi cấy,
đƣờng kính khuẩn lạc trung bình 0,5 - 2 mm. Khuẩn lạc của xạ khuẩn có nhiều màu
sắc khác nhau nhƣ đỏ, da cam, vàng, lam, hồng, nâu, tím. Màu sắc của xạ khuẩn
cũng đƣợc coi là một trong những đặc điểm phân loại quan trọng [1].
Trên môi trƣờng đặc, đa số xạ khuẩn có hai loại khuẩn ty: khuẩn ty khí sinh
(aerial mycelium) và khuẩn ty cơ chất (substrate mycelium). Nhiều loại chỉ có
khuẩn ty cơ chất nhƣng cũng có loại (nhƣ chi Sporichthya) lại chỉ có khuẩn ty khí
sinh. Giữa khuẩn lạc thƣờng thấy có nhiều bào tử màng mỏng gọi là bào tử trần
(conidia hay conidiospores). Nếu bào tử nằm trong bào nang (sporangium) thì đƣợc
gọi là nang bào tử hay bào tử kín (sporangiospores). Bào tử ở xạ khuẩn đƣợc sinh ra
ở đầu một số khuẩn ty theo kiểu hình thành các vách ngăn (septa). Các chuỗi bào tử
trần có thể chỉ là 1 bào tử (nhƣ ở Thermoactinomyces, Saccharomonospora,
Promicromonospora, Micromonospora và Thermomonosspora), có thể có 2 bào tử
(nhƣ ở Microbispora), có thể là chuỗi ngắn (nhƣ ở Nocardia, Pseudonocardia,
Streptoverticillium, Sporichthya, Actinomadura, Microtetraspora,
Streptoalloteichus, Glycomyces, Amycolata, Amycolatopsis, Catellatospora,và
Microellobosporia), có thể là chuỗi dài (nhƣ ở Streptomyces, Saccharopolyspora,
Actinopolyspora, Kibdelosporangium, Kitasatosporia, Saccharothrix, nhiều loài ở
Nocardia, Nocardioides, Pseudonocardia, Amycolatopsis và Streptoverticillium), có
thể các bào tử trần nằm trên bó sợi (synnema), tƣơng tự bó sợi của nấm (nhƣ ở
Actinosynnema và Actinomadura). Các chuỗi bào tử có thể thẳng, xoắn hoặc lƣợn
sóng, có thể mọc đơn hay mọc vòng. Các cuống sinh bào tử (sporophore) và cuống
sinh nang bào tử (sporangiophorres) có thể riêng rẽ hoặc phân nhánh. Các đặc điểm
hình thái này rất quan trọng khi tiến hành định tên xạ khuẩn. Tuy nhiên, xạ khuẩn
hoàn toàn khác biệt so với nấm ở những đặc điểm sau: Xạ khuẩn không có nhân
thật, đƣờng kính khuẩn ty và bào tử nhỏ hơn so với ở nấm, khuẩn ty không có vách
14
ngăn, xạ khuẩn là đích tấn công của các phage, không nhạy cảm với các chất kháng
sinh kháng nấm nhƣ các polyen, không chứa chitin và cellulose [2].
(A) Actinoplanes brasiliensi (VTCC - A - 2908). (B) khuẩn lạc chủng Streptomyces
lannensis (VTCC-A2780).
(C) Nonomuraea roseoviolacea (VTCC - A - 2062). (D) Streptomyces
malachitofuscus (VTCC - A - 2789).
1.3.2. Khả năng sinh chất kháng sinh của xạ khuẩn
Một trong những đặc điểm quan trọng của xạ khuẩn là khả năng sinh chất
kháng sinh. Trong số 5500 chất kháng sinh hiện nay có 4000 chất có nguồn gốc từ
Hình 4. Khuẩn lạc của một số chủng xạ khuẩn.
Hình 5. Cuống sinh bào tử của một số chủng xạ khuẩn.
C D
B A

15
xạ khuẩn. Đa số các chất kháng sinh có nguồn gốc từ xạ khuẩn đều có phổ kháng
rộng, kìm hãm và ức chế đƣợc nhiều loại vi sinh vật khác nhau [35].
Rất nhiều chất kháng sinh đƣợc sinh ra bởi xạ khuẩn đang đƣợc sử dụng rộng
rãi trong thực tế hiện nay nhƣ:
Streptomycin: Có nguồn gốc từ Streptomyces griseus có khả năng kháng các vi
khuẩn Gram dƣơng khá mạnh, đƣợc sử dụng để diều trị các bệnh dịch hạch, ho gà
và quan trọng hơn cả là bệnh lao [40] .
Neomycin: Là chất kháng sinh có hoạt phổ rộng, đƣợc phát hiện từ chủng xạ
khuẩn Streptomyces fradiae, có khả năng kháng lại vi khuẩn Gram dƣơng và Gram
âm, đặc biệt chống đƣợc nhiều loại vi khuẩn kháng penicillin và streptomycin [25]
Gentamycin: Có nguồn gốc từ Micromonospora purpurea, có phổ kháng sinh
rộng, có tác dụng chống lại vi khuẩn Gram dƣơng nhƣ tụ cầu, phế cầu đã kháng lại
penicillin và Gram âm nhƣ màng não cầu, lậu cầu. Trong y học hiện nay,
Gentamycin chủ yếu dùng để diều trị các bệnh nhiễm Pseudomonas [19].
Tetracyclin: Là kháng sinh đƣợc tách chiết từ một số chủng xạ khuẩn thuộc chi
Streptomyces. Loại kháng sinh này có phổ rộng, chống lại đƣợc cả vi khuẩn Gram
dƣơng lẫn Gram âm. Ngoài đƣợc sử dụng trong y học, tetracyclin còn đƣợc sử dụng
trong công nghiệp chế biến thực phẩm [7].
Cloramphenicol: Có nguồn gốc từ xạ khuẩn Streptomyces venezueae, có phổ
kháng sinh rộng với vi khuẩn Gram dƣơng và Gram âm [42].
Erythromycin: Có nguồn gốc từ Streptomyces erythreus, có phổ kháng sinh rộng
đối với các vi khuẩn Gram dƣơng, đƣợc sử dụng để điều trị viêm phổi do
Mycoplasma và viêm họng do liên cầu khuẩn [52].
Novobiocin: Có nguồn gốc từ Streptomyces spheroides và Streptomyces niverus,
có hoạt tính mạnh với các vi khuẩn Gram dƣơng, đặc biệt có khả năng chống các tụ
cầu đã kháng penicillin và một số chất kháng sinh khác [34].

16
Amphoterycin: Có nguồn gốc từ Streptomyces nodosus, đƣợc dùng để điều trị
các bệnh ngoài da do nấm Candida abbicans gây ra [10].
Actinomycin: Có nguồn gốc từ Streptomyces antibiticus có hoạt tính kìm hãm sự
phát triển của các khối u ác tính, đƣợc dùng để điều trị một số bệnh ung thƣ [53].
Daunorubixin: Có nguồn gốc từ Streptomyces coeruleorubidus, đƣợc dùng để
điều trị các bệnh bạch cầu cấp tính, bệnh Hodgkin [41].
1.3.3. Sử dụng xạ khuẩn trong khống chế sinh học
Một trong những phƣơng pháp kiểm soát sinh học đƣợc tập trung nghiên cứu
đó là sử dụng các chủng xạ khuẩn. Các chủng xạ khuẩn có khả năng thúc đẩy tăng
trƣởng thực vật bằng cách tác động trực tiếp nhƣ thải ra sắt, photpho hòa tan và sản
sinh các hormone thực vật hoặc gián tiếp nhƣ ức chế tác nhân gây bệnh hoặc cảm
ứng các cơ chế kháng của thực vật chống lại mầm bệnh [14] [22].
Một số chủng thuộc chi Streptomyces đã đƣợc chứng minh là không những
hỗ trợ thực vật trong việc hấp thu các chất dinh dƣỡng mà còn kiểm soát các tác
nhân gây bệnh cho cây trồng [20]. Năm chủng Streptomyces spp. (CAI - 24, CAI -
121, CAI - 127, KAI - 32 và KAI - 90) đƣợc phân lập từ cỏ ở các đống ủ đã đƣợc
báo cáo là có hoạt tính chống lại bệnh héo lá ở đậu xanh do Fusarium oxysporum
gây ra. Ngoài ra, các chủng xạ khuẩn Streptomyces đƣợc chọn lọc này cũng có khả
năng sinh siderophore, indole acetic acid (trừ KAI - 90), hydrocyanic acid, cellulase
(chỉ KAI - 32 và KAI - 90) và protease (chỉ CAI - 24 và CAI - 127) [20].
Năm 2006, Prapavathy và cộng sự công bố hiệu quả của chủng Streptomyces
sp. PM5 trong việc ức chế sự tăng trƣởng của nấm gây bệnh đạo ôn Pryricularia
oryzae và nấm gây bệnh bạc lá lúa Rhizoctonia solani. Ngoài ra, một số chủng
Streptomyces spp. phân lập từ 21 mẫu đất Indonesia đƣợc công bố có khả năng ức
chế sự sinh trƣởng của vi khuẩn Gram dƣơng Bacillus subtilis và Gram âm
Xanthomonas axonopodis [32].

17
Trƣớc đây, các nghiên cứu về khống chế sinh học để kiểm soát các bệnh trên
lúa chƣa đƣợc chú ý nhiều, nhƣng gần đây, đã có một vài thành tựu có ý nghĩa.
Trong đó có sử dụng các vi khuẩn có mối quan hệ với thực vật nhƣ là các tác nhân
khống chế sinh học. Ví dụ nhƣ các nghiên cứu của Gnanamanickam và các cộng sự
của ông tại Ấn Độ và Philipin đã tìm thấy chủng Pseudomonas fluorescens có khả
năng ức chế sự phát triển của Xoo gây bệnh bạc lá lúa [17]. Ngoài ra chủng còn cho
thấy có khả năng ức chế sự phát triển của nhiều vi sinh vật gây bệnh trên cây nhƣ vi
khuẩn, nấm, virus và giun sống kí sinh. Các kết quả thử nghiệm đã chứng minh
chủng P. fluorescens này có vai trò quan trọng trong việc kiểm soát các bệnh cây củ
cải đƣờng, lúa mạch và cây thuốc lá. Các nghiên cứu của Ji và cộng sự năm 2008 đã
cho thấy chủng Lysobacter antibioticus 13-1 có khả năng ức chế sự phát triển của
nhiều loài nấm và vi khuẩn gây bệnh thực vật, trong đó có Xoo [29].. Chủng này có
khả năng kìm hãm sự phát triển của Xoo hiệu quả lên tới 69.7%. Những thử nghiệm
trên đồng ruộng cho thấy hiệu quả làm giảm sự có mặt của Xoo trên lúa từ 59.1-
r73.5%. Hiệu quả ức chế Xoo của chủng Lysobacter antibioticus 13-1 thử trên các
dòng lúa khác nhau có biến động tùy theo từng dòng lúa. Thêm vào đó hiệu quả ức
chế này còn phụ thuộc vào từng chủng Xoo gây bệnh [29].
1.4. Tình hình nghiên cứu xạ khuẩn trong kiểm soát bệnh bạc lá lúa
Tại Viện Vi sinh vật và Công nghệ sinh học, TS. Phan Thị Phƣơng Hoa và
cộng sự đã sàng lọc 2690 chủng xạ khuẩn đang đƣợc lƣu giữ tại Bảo tàng giống
chuẩn Vi sinh vật (VTCC) và tìm đƣợc 167 chủng đƣợc lựa chọn với khả năng
kháng cao nhất cả hai chủng kiểm định Micrococcus luteus NBRC 13867 và
Escherichia coli NBRC 14237 [23]. Các chủng xạ khuẩn này đƣợc tiếp tục sàng lọc
khả năng kháng 10 chủng vi khuẩn gây bệnh bạc lá lúa Xanthomonas oryzae pv.
oryzae (Xoo) R1, R2, R3, R4, R5, R6, R7, R8, R9 và R10 do PGS. TS. Phan Hữu
Tôn (Trƣờng Đại học Nông nghiệp Hà Nội) và cộng sự phân lập. Trong số 167
chủng xạ khuẩn này, 17 chủng xạ khuẩn có khả năng kháng cả 10 chủng Xoo. Để
nghiên cứu khả năng kháng đặc hiệu đối với Xoo của các 17 chủng xạ khuẩn, các

18
chủng vi sinh vật Bacillus subtilis NBRC 3134, Saccharomyces cerevisiae NBRC
10217, Azotobacter sp. VTCC - B - 106 và Pseudomonas putida VTCC - B - 657
đƣợc sử dụng. 5 chủng VN06 - A -353, VN08 - A - 306, VN08 - A - 352, VTCC -
A - 99, VN10 - A - 44 và VN08 - A12 có tính đặc hiệu cao với Xoo, chúng chỉ ức
chế một loại vi sinh vật kiểm định. Trong số đó, chủng VN08 - A12 sinh trƣởng tốt
nhất và không gây ức chế những vi sinh vật có lợi nhƣ Azotobacter sp. (VTCC - B -
106) và Pseudomonas putida (VTCC - B - 657). Những thử nghiệm ngoài đồng
ruộng trên 2 giống lúa Oryza sativa L. SS1 và Oryza sativa L. KD18 cho thấy dịch
nuôi cấy của chủng VN08 - A12 làm giảm đáng kể tổn thƣơng do Xoo gây ra trên lá
lúa [23]. Chính vì vậy, trong nghiên cứu này, chúng tôi tập trung nghiên cứu các
đặc điểm sinh học và phân tích hoạt chất kháng Xoo do chủng VN08 - A12 sinh ra.
19
CHƢƠNG 2. NGUYÊN LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU
2.1. Nguyên liệu và hóa chất
2.1.1 Nguyên liệu
2.1.1.1. Các chủng Xoo
10 chủng vi khuẩn Xoo gây bệnh bạc lá lúa ở miền Bắc Việt Nam đã nhận
đƣợc từ PGS. TS. Phan Hữu Tôn và cộng sự (trƣờng Đại Học Nông Nghiệp Hà
Nội). Danh sách và nơi phân bố của 10 chủng vi khuẩn Xoo trên đƣợc trình bày
trong bảng 3 và hình 6.
Bảng 3. Danh sách 10 chủng Xoo phân lập đƣợc ở miền Bắc Việt Nam đƣợc sử
dụng trong nghiên cứu
STT Chủng Phân lập từ giống lúa Địa điểm thu thập
1 R1 Khang dân, Bắc thơm 7 Bắc Ninh, Hà Nội
2 R2 Nếp tân, HT1 Sơn La, Hải Dƣơng
3 R3 Nhị ƣu 838, Hƣơng cốm Nghệ An, Thái Bình
4 R4 Thục Hƣng Thái Bình
5 R5 Khang dân, Bắc thơm Nghệ An, Hải Dƣơng
6 R6 Nhị ƣu 383 Yên Bái
7 R7 Q5, Nàng hƣơng Hải Dƣơng, Thái Bình
8 R8 Nếp thơm Hải Dƣơng
9 R9 Q5 Hải Dƣơng
10 R10 Hybrid rice Yên Bái

20
Hình 6. Bản đồ phân bố của 10 chủng Xoo ở miền Bắc Việt Nam đƣợc sử dụng
trong nghiên cứu
2.1.1.2. Các chủng xạ khuẩn
Chủng xạ khuẩn VN08 - A12 đƣợc phân lập năm 2008 hiện đang đƣợc lƣu
giữ tại Bảo tàng Giống chuẩn Vi sinh vật học (VTCC), Viện Vi sinh vật và Công
nghệ Sinh học, Đại học Quốc Gia Hà Nội.
2.1.1.3. Vi sinh vật kiểm định
Sáu chủng vi sinh vật kiểm định đƣợc sử dụng hiện đang đƣợc bảo quản tại
VTCC bao gồm:
- Pseudomonas putida (VTCC - B - 657)
- Saccharomyces cerevisiae (VTCC - Y - 62)
- Bacillus subtilis (VTCC - B - 888)
- Azotobacter sp. (VTCC - B - 106)
- Fusarium oxysporum (ATCC - 7601)
- Staphylococcus aureus (ATCC - 29923)
2.1.2. Hóa chất, dụng cụ và thiết bị
2.1.2.1. Hóa chất
- Các loại đƣờng chuẩn: glucose, saccarose, lactose, fructose, mantose ... đƣợc
mua từ hãng Merck (Đức).

21
- Các loại muối: K2HPO4, KH2PO4, KI, MgSO4. 7H2O, KNO3, NaCl, FeSO4.
7H2O, (NH4)2SO4, CaCO3, MnCl2, Na2CO3, ZnCl2, ZnSO4 ... đƣợc nhập từ các hãng
của Trung Quốc.
- Cao thịt, cao nấm men, cao malt, peptone... của hãng Merck (Đức).
- Các loại hóa chất khác nhƣ thạch, tinh bột tan, casein, CMC (Carboxyl
Methyl Cellulose) đƣợc sản xuất ở Nhật, Trung Quốc, Việt Nam.
- Các dung môi nhƣ ethanol, iso - propanol, methanol, acetone, ethylacetate
đƣợc mua từ hãng Fisher (Mỹ).
2.1.2.2. Dụng cụ và thiết bị
- Kính hiển vi quang học Olympus (Nhật) - Tủ ấm, tủ sấy Memmert (Đức)
- Kính hiển vi điện tử Joel (Nhật) - Cân phân tích, cân kỹ thuật
- Máy lắc (Hàn Quốc) - Nồi khử trùng (Đài Loan)
- Box cấy vô trùng Nuaire (Mỹ) - Máy đo pH 151 Martini
- Máy ly tâm Hettich (Đức) - Máy cất nƣớc Hamilton.
- Các dụng cụ thủy tinh của Trung Quốc, Đức, Việt Nam.
2.1.2.3. Môi trƣờng nghiên cứu
Môi trƣờng nuôi cấy xạ khuẩn:
+ Môi trƣờng 2M (g/ l): tinh bột - 20, bột đậu tƣơng - 15, cao nấm men - 2, CaCO3
- 4, pH 6.2
+ Môi trƣờng No. 8(g/ l): casitone - 7.5, cao nấm men - 7.5, glycerol - 15, NaCl -
2.5
+ Môi trƣờng 301 (g/ l): tinh bột - 24, glucose - 1, peptone - 3, cao thịt - 3, cao nấm
men - 5, CaCO3 - 4, pH 7.0
+ Môi trƣờng A - 3M g/l): glucose - 5, glycerol - 20, tinh bột - 20, pharmamedia -
15, cao nấm men - 3, Diaion HP - 20 - 10, pH 7,0

22
+ Môi trƣờng A - 16 (g/ l): glucose - 20, pharmamedia - 10, CaCO3 - 5
+ Môi trƣờng SKS (g/ l): tinh bột - 10, glucose - 10, bột đậu tƣơng - 10, peptone -
5, CaCO3 - 3
+ Môi trƣờng YS (g/l): cao nấm men - 2, tinh bột - 10, thạch - 16
Môi trƣờng thử hoạt tính kháng vi khuẩn:
+ Môi trƣờng PSA (g/ l): Khoai tây - 300, Na2HPO4 - 2, Ca(NO3)2 - 0.5, pepton - 5,
sucrose - 15, thạch - 16, pH 7,0
+ Môi trƣờng YM (g/ l): Glucoza - 10, pepton - 5, cao nấm men - 3, cao malt - 3,
thạch - 15
+ Môi trƣờng thạch thƣờng (g/ l): Pepton - 5, cao thịt - 2, NaCl - 1, thạch - 15, pH –
7,0
+ Môi trƣờng Muller - Hinton (g/l): Cao thịt - 3 ; casein thuỷ phân – 17,5; tinh bột
tan – 1,5; thạch 17; pH 7,4 + 0,2.
2.2. Phƣơng pháp nghiên cứu
2.2.1. Phƣơng pháp thử hoạt tính kháng vi sinh vật
2.2.1.1. Phương pháp thỏi thạch
Vi sinh vật kiểm định đƣợc nuôi trên môi trƣờng Muller - Hinton, PSA, YM,
hoặc thạch thƣờng. Dịch nuôi vi sinh vật kiểm định (1 - 5 ml/ l) đƣợc thêm vào môi
trƣờng đã khử trùng và để nguội đến 40 - 45C và đổ ra đĩa petri. Sau đó các thỏi
thạch có xạ khuẩn đƣợc đặt lên đĩa, ủ ở 30oC trong 24 giờ. Hoạt tính kháng vi sinh
vật đƣợc đánh giá bằng khả năng tạo vòng ức chế xung quanh thỏi thạch theo công
thức D - d (D: đƣờng kính vòng kháng trên đĩa thạch, d: đƣờng kính của lỗ đục)
(mm).

23
2.2.1.2. Phương pháp đục lỗ thạch
Môi trƣờng chứa vi sinh vật kiểm định đƣợc chuẩn bị nhƣ trong phƣơng pháp
thỏi thạch. Sau đó, 4 - 5 lỗ trên đĩa thạch đƣợc tạo bằng khoan đục lỗ. 100 µl dịch
nuôi hoặc mẫu đƣợc nhỏ vào mỗi lỗ, ủ ở 30oC trong 24 giờ. Khả năng ức chế đƣợc
đánh giá bằng khả năng tạo vòng ức chế xung quanh lỗ, theo công thức D - d (mm).
2.2.2. Phân loại bằng đặc điểm hình thái
2.2.2.1. Hình thái khuẩn lạc
Hình thái khuẩn lạc xạ khuẩn đƣợc xác định về kích thƣớc, bề mặt khuẩn lạc,
màu sắc của hệ sợi khí sinh và hệ sợi cơ chất bằng cách cấy ria 3 pha các chủng xạ
khuẩn trên các môi trƣờng (ISP1, ISP2, ISP3, ISP4, ISP5, ISP6, ISP7 và YS), ủ ở
30oC trong 7 - 14 ngày.
2.2.2.2. Hình thái chuỗi sinh bào tử và bề mặt bào tử
Hình thái chuỗi sinh bào tử được quan sát dưới kính hiển vi quang học
sử dụng phương pháp cắm lamen nghiêng 45oC trên đĩa thạch YS đã cấy chủng
xạ khuẩn, ủ ở 30oC trong 7 - 14 ngày.
Ngoài ra, chuỗi bào tử và bề mặt bào tử cũng đƣợc quan sát dƣới kính hiển
vi điện tử quét qua các bƣớc sau:
- Cố định mẫu: Đặt thỏi thạch có bào tử xạ khuẩn lên lam kính nghiêng 45oC
trong bình có chứa osmium oxide 2% trong 2 - 3 giờ.
- Rửa mẫu: Mẫu đƣợc rửa lần lƣợt bằng ethanol ở các nồng độ 50%, 60%,
70%, 80%, 90%, 95% và 100% trong 15 phút. Sau đó, mẫu đƣợc ngâm trong
isopentyl acetate 100% trong 12 giờ.
- Làm khô mẫu: Mẫu đƣợc làm khô bằng thiết bị có phun ngập khí nitơ trong
20 phút.
- Quan sát dƣới kính hiển vi điện tử quét.

24
2.2.3. Hóa phân loại
2.2.3.1. Phân tích thành phần axit amin trong thành tế bào
Chuẩn bị thành tế bào: Tế bào ƣớt (1 - 2 ml) + 5 ml nƣớc cất vô trùng + hạt
thủy tinh (loại lớn và nhỏ, đƣờng kính 0, 11 - 0, 12 mm). Phá tế bào bằng sóng siêu
âm ca.180 trong 1 - 1,5 giờ cho đến khi dung dịch trở nên trong. Ly tâm với tốc độ
10000 vòng/ 30 phút. Bỏ dịch trên. Thêm 3 ml SDS 4%. Giữ ở 100oC trong 40 phút.
Ly tâm với tốc độ 10000 vòng/ phút trong 30 phút ở 30oC loại bỏ dịch trên. Rửa với
nƣớc ít nhất 3 lần với điều kiện giống trên (17000 vòng/ phút trong 30 phút). Đông
khô qua đêm.
2 - 3 mg thành tế bào đƣợc bổ sung 200 μl HCl 4 N, ủ ở 100oC qua đêm.
Mẫu đƣợc làm khô bằng hút chân không (khoảng 24 giờ). Thêm 200 μl nƣớc cất,
chuyển sang ống eppendorf, ly tâm với tốc độ 12000 vòng/ phút trong 5 phút. Dịch
trên đƣợc thu và lọc qua màng lọc.
Phân tích mẫu bằng sắc ký bản mỏng TLC: Mẫu (3 - 5 μl) và các axit amin
chuẩn (1 μl) DAP, Ala, Gly, Glu, Lys đƣợc chấm lên bản sắc ký cellulose (phụ lục
1). Bản sắc ký đƣợc nhuộm bằng ninhydrin ở 100oC trong 5 phút. Các axit amin sẽ
hiện lên dƣới dạng các chấm màu tím.
Phân tích mẫu bằng HPLC: 10 - 20 μl mẫu và các axit amin chuẩn (25 nmol)
đƣợc làm khô bằng hút chân không trong 2 giờ, thêm 20 μl Ethanol/ DW/
Triethylamine 2/ 2/ 1, làm khô bằng hút chân không trong 2 giờ. Thêm 20 μl
Ethanol/ DW/ Triethylamine/ Phenylisothiocyanate 7/ 1/ 1/ 1, giữ ở nhiệt độ phòng
trong 20 phút, làm khô bằng hút chân không trong 2 giờ. Mẫu đƣợc hòa tan bằng
0,5 ml dung dịch A và đƣợc phân tích bằng HPLC (phụ lục 1).
2.2.3.2. Phân tích thành phần menaquinone
100 - 500 mg tế bào khô đƣợc hòa tan trong 10 - 15 ml Chloroform:
methanol (2: 1), trộn bằng khuấy từ chậm trong 3 giờ. Ly tâm 3000 vòng/ phút

25
trong 10 phút, lấy dịch trên, làm khô bằng cô quay. Hòa tan bằng 1,5 ml acetone,
làm khô bằng hút chân không.
Mẫu đƣợc hòa tan bằng 50 μl ethanol, chấm lên bản gel silica. Menaquinone
đƣợc phát hiện bằng tia UV, sau đó đƣợc cạo ra chuyển vào ống eppendorf, thêm
1,5 ml acetone, ly tâm 3000 vòng/ phút trong 5 phút, lấy dịch trên, làm khô bằng
nitơ lỏng. Thêm 50 - 100 μl ethanol, lọc và dùng làm mẫu chạy HPLC và khối phổ
(MS) (phụ lục 2).
2.2.3.3. Phân tích thành phần axit béo
5 mg tế bào khô + dung dịch kiềm (R1) 1 ml, vortex 5 - 10 giây, giữ ở 100oC
trong 5 phút, vortex 5 - 10 giây, giữ tiếp ở 100oC trong 25 phút, để nguội xuống
nhiệt độ phòng, có thể đặt trực tiếp dƣới vòi nƣớc sau khi hạ xuống 80oC. Thêm 2
ml dung dịch methyl hóa (R2) giữ ở 80oC trong 10 phút. Thêm 3 ml dung dịch tách
(R3) trộn đều, loại bỏ lớp dƣới. Thêm 3 ml dịch rửa (R4), ly tâm 2000 vòng/ phút
trong 3 phút, chuyển 2/3 lớp trên sang ống đựng mẫu. Mẫu đƣợc phân tích bằng sắc
ký khí (hệ thống MIDI).
2.2.3.4. Phân tích thành phần G+C trong ADN
25 μl ADN (2 - 25 μg) giữ ở 60oC trong 1 giờ, giữ ở 100
oC trong 2 phút,
chuyển lên đá. Thêm 25 μl đệm acetate (phụ lục 5), giữ ở 37oC trong 1 giờ. Thêm
25 μl đệm Glycine (0,1 M, pH 10,4) và 2 μl alkaline photphatase (0,35 U/ μl), giữ ở
37oC trong 1 - 2 giờ. Phân tích bằng HPLC (phụ lục 4).
2.2.4. Phân loại sinh học phân tử giải trình tự 16S – rADN
Tách chiết ADN
Tế bào xạ khuẩn thu từ 1,5 ml dịch nuôi cấy đƣợc hòa tan trong 100 l TE,
thêm 0,4 mg lysozyme, ủ 37oC trong 1 giờ. Thêm 100 l SDS 10% (w/v), ủ 37
oC
trong 30 phút. Thêm 8 l ARNase 3 mg/ ml, ủ ở 37oC trong 30 phút. Thêm 12 l

26
proteinase K (5 mg/ ml), ủ 15 phút ở 56oC. Thêm 1 V phenol: chloroform: isoamyl
alcohol (P: C: I = 25: 24: 1), ly tâm 15000 vòng/ phút, 15 phút, thu lớp dịch trên.
Thực hiện bƣớc này 3 lần. Thêm 1/ 10 V natri acetate 3 M và 2,5 V ethanol 100%,
đặt trong đá 30 phút. Ly tâm 15000 vòng/ phút trong 15 phút, thu cặn, rửa bằng
ethanol 70%. Làm khô, hoà tan trong 50 l TE.
Phản ứng khuếch đại ADN
Thành phần phản ứng (l): 10X buffer - 10; dNTP 2 mM - 10; mồi 27F (10
pM) - 2; 1525R (10 pM) - 2; Taq polymerase (5u/l) - 2; ADN khuôn (50 - 100g/
l) - 1- 2; H2O đủ 100 l
Chu trình nhiệt: 35 chu kỳ
95oC - 3 phút
95oC - 30giây
55oC - 30 giây
72oC - 1 phút
4oC -
Kiểm tra các sản phẩm của PCR bằng điện di agarose 1 %. Sản phẩm PCR
sau đó đƣợc tinh sạch bằng kit PCR clean up (QIAgen).
Phản ứng khuếch đại ADN cho đọc trình tự
Thành phần Terminator Ready Reaction Mix (Termix): Buffer 5X - 9 l,
Bigdye Ready Reaction premix -18 l, H2O - 9l.
Thành phần phản ứng PCR: Termix - 8 l, dNTP 2 mM - 10, Mồi - 1 l, ADN
khuôn - 1 l (nồng độ ADN là 40 - 60 g/ ml), H2O - 10 l.
- Chu trình nhiệt cho phản ứng khuếch đại gen:

27
96oC - 1phút
96oC - 10giây 25 chu kỳ
50oC - 5 giây
60oC - 4 phút
- Tinh sạch sản phẩm PCR:
Thêm 5 l EDTA 125 mM và 60 l ethanol 100%, để 15 phút ở nhiệt độ
phòng. Ly tâm 15000 vòng/ phút trong 15 phút, thu cặn, rửa bằng 60 l ethanol
70%. Thêm 10 l HiDi Formamide. Để ở 96oC trong 2 phút sau đó chuyển ngay
mẫu lên đá. Mẫu đƣợc đọc trình tự bằng máy ABI 3100 Avant.
Phân tích trình tự và xây dựng cây phát sinh chủng loại
Trình tự của ADNr 16S đƣợc phân tích bằng phần mềm CLUSTAL W. Các
trình tự tham khảo dùng trong nghiên cứu cây phát sinh chủng loại đƣợc lấy từ dữ
liệu của DDBJ, EMBL, GenBank. Cây phát sinh đƣợc xây dựng theo phƣơng pháp
của Saitou và Nei (1987) với độ lặp lại 1000 lần trên phần mềm ClustalX2 và
ClustalW2.
2.2.5. Tối ƣu hóa điều kiện nuôi cấy xạ khuẩn
2.2.5.1. Lựa chọn môi trường nuôi cấy thích hợp
Chủng VN08 - A12 (3% giống cấp 1) đƣợc nuôi trong 25 ml của 6 môi
trƣờng: 3M, 16M, 2M, 301, N8, SKS trong bình tam giác 250 ml, lắc ở 150 vòng/
phút trong 4 ngày. Dịch nuôi cấy sau ly tâm đƣợc thử hoạt tính kháng Xoo R2 theo
phƣơng pháp đục lỗ thạch.
2.2.5.2. Lựa chọn thời gian nuôi cấy thích hợp
Chủng VN08 - A12 đƣợc nuôi cấy nhƣ trên trong môi trƣờng SKS. Dịch
nuôi cấy (1 ml) đƣợc thu sau 2, 3, 4, 5, 6 và 7 ngày, thử hoạt tính đối kháng Xoo R2
theo phƣơng pháp đục lỗ thạch.

28
2.2.5.3. Lựa chọn thể tích nuôi cấy thích hợp
Chủng VN08 - A12 (3% giống cấp 1) đƣợc nuôi trong 25, 50, 75, 100, 125,
150 ml môi trƣờng SKS trong bình tam giác 250 ml, lắc 150 vòng/ phút trong 4
ngày. Dịch nuôi cấy sau ly tâm đƣợc thử hoạt tính kháng Xoo R2 theo phƣơng pháp
đục lỗ thạch.
2.2.5.4. Lựa chọn nhiệt độ nuôi cấy
Chủng VN08 - A12 đƣợc nuôi trong môi trƣờng SKS, lắc 150 vòng/ phút ở
các nhiệt độ 25, 30, 35, 40oC trong 4 ngày. Dịch nuôi cấy sau ly tâm đƣợc thử hoạt
tính kháng Xoo R2 theo phƣơng pháp đục lỗ thạch.
2.2.5.5. Lựa chọn cách cấy giống
Chủng xạ khuẩn VN08 - A12 đƣợc cấy bằng các cách khác nhau: giống cấp
1, giống cấp 2 và bào tử (1x108, 2x10
8, 3x10
8, 4x10
8, 5x10
8, 6x10
8 bào tử/ ml), lắc
150 vòng/ phút ở 30oC trong 4 ngày. Dịch nuôi cấy sau ly tâm đƣợc thử hoạt tính
kháng Xoo R2 theo phƣơng pháp đục lỗ thạch.
2.2.6. Tinh sạch hoạt chất kháng Xoo
2.2.6.1. Tách dịch chiết thô
Dịch nuôi cấy (100 ml) đƣợc ly tâm ở 8000 vòng/phút trong 10 phút, thu
dịch trong. Thêm 50 ml ethylacetate, lắc đều, ly tâm ở 4oC vận tốc 8000 vòng/ phút
trong 10 phút. Thu lớp dịch phía trên, bổ sung khoảng 0,02 g Na2SO4, lắc đều, lọc
lấy dịch trong, cô đến khô, hòa tan lại bằng 1 ml methanol.
2.2.6.2. Tinh sạch bằng silica gel
Silica gel (20 ml) hòa vào hexan và nhồi vào cột. Các dung môi cần sử dụng:
(1). Hexan 100%
(2). 80% Hexan: 20% ethyl acetate
(3). 50% Hexan: 50% ethyl acetate

29
(4). 20% Hexan: 80% ethyl acetate
(5). 0% Hexan: 100% ethyl acetate
(6). 50% Methanol: 50% ethyl acetate
Chuyển dịch chiết thô lên cột. Các dung môi (1), (2), (3), (4), (5), (6) đƣợc
lần lƣợt thêm vào cột (40 ml), các phân đoạn đƣợc thu tƣơng ứng, cô quay đến khô,
hòa tan trong 1 ml methanol. Các phân đoạn đƣợc thử hoạt tích kháng Xoo R2 bằng
phƣơng pháp đục lỗ thạch.
2.2.6.3. Tinh sạch bằng HPLC
Các phân đoạn có hoạt tính đƣợc phân tích và tinh sạch bằng phƣơng pháp
HPLC sử dụng cột Cadenza CD - C18 (7,5 x 4,6 mm). Nồng độ acetonitrile đƣợc
tăng tuyến tính từ 15% đến 85% trong 22 phút với tốc độ 1,2 ml/ phút.
Kênh A: Nƣớc MQ: Axit formic = 1000: 1
Kênh B: Acetonnitrile
Bảng 4. Chƣơng trình phân tích HPLC
Thời gian (phút) Dịch A (%) Dịch B (%) Tốc độ (ml/ phút)
0 85 15 1
3 85 15 1
25 15 85 1
29 15 85 1
32 85 15 1
35 85 15 1
2.2.7. Xác định khối lƣợng phân tử bằng khối phổ (MS)

30
Chất kháng Xoo đã đƣợc tinh sạch qua máy HPLC đƣợc phân tích trọng
lƣợng phân tử bằng phƣơng pháp khối phổ. Mẫu đƣợc bơm trực tiếp vào máy
khối phổ ở trạng thái tích điện dƣơng.

31
CHƢƠNG 3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Khả năng kháng vi sinh vật của chủng xạ khuẩn VN08 - A12
Chủng xạ khuẩn VN08 - A12 đƣợc tiến hành thử hoạt tính kháng với 10
chủng Xanthomonas oryzae pv. oryzae (Xoo) từ R1 đến R10 bằng phƣơng pháp
thỏi thạch. Kết quả đƣờng kính vòng hoạt tính kháng Xoo đƣợc thể hiện trong bảng
5. Bảng 5. Hoạt tính kháng 10 chủng Xoo của chủng xạ khuẩn VN08 - A12
Chủng Đƣờng kính vòng kháng khuẩn (D-d: mm)
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
VN08-
A12
10±1 14±1 18±1 12±4 8±1 14±2 8±1 5±1 8±1 10±1
Kết quả trong bảng 5 cho thấy chủng VN08 - A12 có khả năng kháng cả 10
chủng Xoo. Trong đó, chủng này có khả năng kháng mạnh đối với R2, R3 và R6
nhƣng kháng yếu với R8.
Mục đích của nghiên cứu này là lựa chọn chủng có khả năng kháng đƣợc tất
cả 10 chủng Xoo nhƣng không gây hại cho vi sinh vật khác. Chính vì vậy, khả năng
ức chế đối với 5 vi sinh vật kiểm và 2 vi sinh vật hữu ích của chủng xạ khuẩn này
đƣợc tiếp tục thử nghiệm. Kết quả đƣợc trình bày trong bảng 6 và bảng 7.
Bảng 6. Hoạt tính kháng vi sinh vật của chủng xạ khuẩn VN08 - A12
STT Chủng Vi sinh vật kiểm đinh Đƣờng kính vòng kháng
khuẩn (D-d: mm)
1 Pseudomonas putida VTCC - B-657 -
2 Fusarium oxysporum ATCC - 7601 -
3 Staphylococcus aureus ATCC - 29923 -
4 Saccharomyces cerevisiae VTCC - Y - 62 -
5 Bacillus subtilis VTCC - B - 888 -

32
Kết quả cho thấychủng xạ khuẩn VN08 - A12 không ức chế vi sinh vật kiểm
định nào. Ngoài ra, chủng VN08 - A12 có khả năng tăng trƣởng khá nhanh. Vì vậy,
chủng VN08 - A12 đƣợc tiếp tục kiểm tra khả năng ức chế đối với hai vi sinh vật có
lợi Azotobacter sp. (VTCC - B - 106) và Pseudomonas putida (VTCC - B - 657).
Kết quả thử hoạt tính cho thấy chủng VN08 - A12 không kháng cả hai loại vi khuẩn
đƣợc kiểm tra này (Bảng 7). Nhƣ vậy, chủng VN08 - A12 có khả năng kháng rất
đặc hiệu với vi khuẩn Xoo.
Bảng 7. Hoạt tính kháng vi sinh vật có lợi của chủng xạ khuẩn VN08 - A12
Chủng xạ khuẩn Đƣờng kính vòng kháng khuẩn (D - d, mm)
Azotobacter sp.
(VTCC - B - 106)
Pseudomonas putida
(VTCC - B - 657)
VN08 - A12 - -
3.2. Phân loại chủng VN08 - A12
3.2.1. Phân loại bằng hình thái
Sau 7 ngày nuôi cấy trên đĩa thạch YS, chủng VN08 - A12 tạo khuẩn lạc lồi,
mép tạo viền có dạng sợi tia nhỏ, hệ sợi khí sinh có màu từ trắng đến nâu. Hệ sợi cơ
chất có màu nâu nhạt (hình 7A). Đƣờng kính khuẩn lạc 0,5 - 2,5 mm, không tiết sắc
tố vào môi trƣờng.
Chuỗi bào tử của chủng VN08 - A12 có dạng thẳng, dài, thƣờng có trên 20
bào tử trong một chuỗi (hình 7B và hình 8C). Khi quan sát dƣới kính hiển vi điện tử
quét (SEM) bề mặt bào tử có dạng nhẵn (hình 8D).

33
3.2.2. Hóa phân loại
3.2.2.1. Thành phần axit amin trong thành tế bào
Thành phần axit amin trong thành tế bào của chủng VN08 - A12 đƣợc xác
định bằng sắc ký bản mỏng nhƣ đã mô tả trong phần phƣơng pháp 2.2.3.1 (hình 9).
Kết quả so sánh với chất chuẩn cho thấy chủng VN08 - A12 chứa LL - DAP. (DAP:
Diaminopimelic).
Hình 7. Khuẩn lạc và chuỗi bào tử của chủng VN08 - A12
(Ảnh SEM ở 15KVx 5,000)
(Ảnh SEM ở 10KVx 30,000)
Hình 8. Chuỗi bào tử và bào tử của chủng VN08 - A12
1µm
A B
D C

34
Hình 9. Kết quả chạy sắc ký bản mỏng thành phần axit amin trong thành tế bào của
chủng VN08 - A12.
(1). Chất chuẩn Meso - DAP và LL - DAP
(2). Mẫu VN08 - A12
3.2.2.2. Thành phần menaquinone
Thành phần menaquinone của chủng VN08 - A12 đƣợc phân tích theo
phƣơng pháp đã đƣợc trình bày trong mục 2.2.3.2. Mẫu menaquinone sau khi đƣợc
phân tách bằng sắc ký bản mỏng đƣợc thu lại để phân tích bằng LC - MS. Hai đỉnh
ở phút 14,019 và 15, 689 trên LC đƣợc xác định tƣơng ứng là MK - 9(H6) và MK -
9(H8) bằng phân tích khối phổ đi kèm (hình 10). Kết quả phân tích diện tích của hai
đỉnh này đƣợc thể hiện trong bảng 8, cho thấy thành phần menaquinone của chủng
VN08 - A12 là MK - 9 (H6) 51,9% và MK-9(H8) 48,1%.
1 2
Meso - DAP
LL - DAP Chủng VN08 -A12:
LL - DAP

35
Hình 10. Kết quả phân tích LC thành phần menaquinone của chủng VN08 - A12.
Bảng 8 : Kết quả phân tích thành phần menaquinone của chủng VN08 - A12
Peak Thời
gian
lƣu
Diện
tích
peak
Chiều
cao
peak
Phần trăm
diện tích
peak (%)
Phần trăm
chiều cao
peak (%)
Loại
menaquinone
của VN08 -A12
1 14,019 840142 44224 51,961 55,145 MK - 9 (H6)
2 15,689 776728 35972 48,039 44,55 MK - 9(H8)
Tổng 1616869 80196 100,00 100,000
3.2.2.3.Thành phần axit béo
Thành phần axit béo của chủng VN08 - A12 đƣợc phân tích bằng hệ thống
sắc ký khí kết hợp với phần mềm MIDI theo phƣơng pháp đƣợc trình bày trong mục
2.2.3.3. Kết quả phân tích đƣợc thể hiện trong hình 11 và bảng 9, cho thấy chủng
MK - 9 (H6) MK - 9 (H8)

36
VN08 - A12 có thành phần axit béo gồm anteios - (anteios - C15 : 0), iso - (iso - C16 :
0) và (n - C16 : 0).
Hình 11. Sắc ký đồ thành phần axit béo của chủng VN08 - A12
Bảng 9. Kết quả phân tích thành phần menaquinone của chủng VN08 - A12
Thời gian lƣu Tên peak Tỷ lệ phần trăm
6,974 14: 0 iso 1,77
7,503 14: 0 0,77
8,478 15: 0 iso 9,18
8,619 15: 0 anteiso 19,25
9,064 15: 0 1,13
9,838 16: 1 iso H 2,45
10,119 16: 0 iso 16,02
10,441 16:1 cis 9 11,16
10,744 16: 0 10,09

37
Thời gian lƣu Tên peak Tỷ lệ phần trăm
11,472 16:0 9 methyl 5,41
11,661 17: 1 anteiso 4,89
11,840 17: 0 iso 4,19
12,003 17: 0 anteiso 11,53
12,127 17: 1 0,75
12,438 17: 0 10 methyl 0,45
13,205 18: 1 iso H 0,41
3.2.2.4. Thành phần G+C trong AND
Thành phần GC của chủng VN08 - A12 đƣợc phân tích bằng phƣơng pháp
HPLC nhƣ đƣợc trình bày trong mục 2.2.3.4. Kết quả đƣợc thể hiện trong hình 12
và bảng 10. Kết quả phân tích cho thấy rằng thành phần GC trong ADN của chủng
VN08 - A12 là 75%.
Hình 12. Sắc ký đồ thành phần GC của chủng VN08 - A12
A
C G
T

38
Bảng 10. Kết quả phân tích thành phần GC của chủng VN08 - A12
Peak Thời gian
lƣu
Diện tích
peak
Chiều
cao peak
Phần
trăm
diện tích
peak (%)
Phần trăm
chiều cao
peak (%)
Thành
phần
GC
1 2,299 1063153 162556 38,736 40,537 C
2 4,058 1019074 161599 37,130 40,298 G
3 6,302 327432 40445 11,930 10,086 T
4 6,841 334930 36411 12,203 9,080 A
Tổng 2744589 401012 100,000 100,000
3.2.3. Trình tự 16S rADN
Kết quả trình tự gen 16S rADN của chủng VN08 - A12 (phụ lục 8).
Căn cứ vào các kết quả phân tích về hình thái đƣợc thể hiện trong phần 3.2.1.
cho thấy chủng VN08-A12 có đặc điểm điển hình của chi Streptomyces. Phân tích
thành phần axit amin trong thành tế bào của chủng VN08 - A12 đặc trƣng bởi LL -
DAP. Các thành phần chính của axit béo là anteios- (anteios - C15 : 0), ISO - (iso -
C16 : 0) và bình thƣờng (n- C16 : 0) axit. Các menaquinones là MK-9 (H6) 51,9%
và MK-9 (H8) 48,1%. Thành phần GC là 75%..
Kết quả Blast search trên GenBank cho thấy chủng VN08 - A12 có trình tự
16S rADN hoàn toàn tƣơng đồng với loài Streptomyces toxytricini với tỉ lệ 100%.
Cây phân loại của chủng VN08 - A12 đƣợc dựng bằng phần mềm ClustalX2 và
ClustalW2 cho thấy chủng nghiên cứu nằm trong cùng nhóm với nhóm chủng
Streptomyces toxytricini (hình 13). Nhƣ vậy căn cứ vào các kết quả phân tich về
hình thái, đặc điểm sinh lý sinh hóa, hóa phân loại và trình tự 16s rADN của chủng
VN08 - A12 cho thấy chủng VN08 - A12 thuộc chi Streptomyces toxytricini.

39
Hình 13. Cây phân loại chủng VN08 - A12 dựa trên trình tự gen 16S – rADN.
3.3. Tối ƣu hóa điều kiện nuôi cấy của chủng VN08 - A12
3.3.1. Môi trường nuôi cấy thích hợp
Để lựa chọn môi trƣờng nuôi cấy của chủng VN08 - A12 để đạt hoạt tính
kháng Xoo cao nhất, 6 loại môi trƣờng đã đƣợc lựa chọn để thử nghiệm bao gồm
3M, 16M, 2M, 301, N8 và SKS. Chủng VN08 - A12 đƣợc nuôi trong 25 ml của mỗi
loại môi trƣờng trong bình tam giác 250 ml, lắc ở 150 vòng/ phút trong 4 ngày ở
nhiệt độ 30oC. Dịch nuôi cấy sau ly tâm đƣợc thử hoạt tính kháng Xoo R2 theo
phƣơng pháp đục lỗ thạch. Kết quả đƣợc trình bày trong hình 14 và hình 29C. Kết
quả cho thấy, môi trƣờng SKS là môi trƣờng tốt nhất cho chủng VN08 - A12 để sản
sinh chất kháng Xoo.

40
Hình 14. Ảnh hƣởng của môi trƣờng nuôi cấy khác nhau đến sự sản sinh hợp chất
kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12
3.3.2. Thời gian nuôi cấy thích hợp
Chủng VN08 - A12 đƣợc nuôi lắc trong môi trƣờng SKS ở 30oC, dịch nuôi
cấy đƣợc thu và thử hoạt tính kháng Xoo ở các ngày khác nhau. Kết quả đƣợc trình
bày trong hình 15 và hình 29B. Kết quả cho thấy rằng, thời gian nuôi cấy ảnh hƣởng
đến sự sản sinh lƣợng chất kháng Xoo. Khả năng ức chế Xoo (R2) đạt cao nhất sau 4
ngày nuôi cấy và giảm dần khi tăng thời gian nuôi cấy.
Hình 15. Ảnh hƣởng của thời gian cấy khác nhau đến sự sản sinh hợp chất kháng
sinh ức chế vi khuẩn Xoo của chủng VN08 - A12
0
5
10
15
20
25
30
35
3M 16M 2M 301 N8 SKS
Đư
ờn
g k
ính
vò
ng
kh
án
g k
hu
ẩn
(D
-d
:mm
)
Môi trường nuôi cấy
0
5
10
15
20
25
30
3 4 5 6 7
Đư
ờn
g k
ính
vò
ng
kh
án
g k
hu
ẩn
(D
:mm
)
Thời gian nuôi cấy (ngày)

41
3.3.3. Thể tích nuôi cấy thích hợp
Để xác định thể tích nuôi cấy thích hợp, chủng VN08 - A12 đƣợc nuôi cấy
trong môi trƣờng SKS với các thể tích khác nhau ở 30oC. Sau 4 ngày, dịch nuôi cấy
đƣợc thu để thử hoạt tính kháng Xoo. Kết quả đƣợc thể hiện trong hình 16 và hình
29A. Kết quả cho thấy hoạt tính kháng Xoo cao nhất ở thể tích 25 ml trong bình 250
ml và giảm dần với sự gia tăng của thể tích môi trƣờng. Kết quả này khẳng định oxy
là nhân tố quan trọng ảnh hƣởng khả năng sinh hoạt chất kháng Xoo của chủng
VN08 - A12.
Hình 16. Ảnh hƣởng của thể tích môi trƣờng nuôi cấy khác nhau đến sự sản sinh
hợp chất kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12 (S. toxytricini)
3.3.4. Nhiệt độ nuôi cấy thích hợp
Chủng VN08 - A12 đƣợc nuôi trong môi trƣờng SKS, ở các nhiệt độ khác
nhau. Dịch nuôi cấy sau 4 ngày đƣợc thử hoạt tính kháng Xoo. Kết quả đƣợc trình
bày trong hình 17 và hình 29D. Nhiệt độ nuôi cấy ảnh hƣởng rất lớn đến sự sản sinh
hoạt chất kháng Xoo. Kết quả cho thấy rằng chủng VN08 - A12 đƣợc nuôi ở nhiệt
30oC là tốt nhất.
0
5
10
15
20
25
30
25 50 75 100 125 150
Đư
ờn
g k
ính
vò
ng
kh
án
g k
hu
ẩn
(D
-d:m
m)
Thể tích nuôi cấy (ml)

42
Hình 17. Ảnh hƣởng của nhiệt độ môi trƣờng nuôi cấy khác nhau đến sự sản sinh
hợp chất kháng sinh ức chế vi khuẩn Xoo của chủng VN08 - A12
3.3.5. Cách cấy giống thích hợp
Cách cấy giống thƣờng ảnh hƣởng rất lớn đến khả năng sinh các chất có hoạt
tính sinh học. Chính vì vậy, chúng tôi khảo sát các cách cấy giống khác nhau để tìm
phƣơng pháp thu đƣợc hoạt chất kháng Xoo cao nhất, bao gồm: giống cấp 1 ở 24,
48, 76, 92 giờ và bào tử (1x108, 2x10
8, 3x10
8 và 4x10
8)
bào tử. Dịch nuôi cấy sau 4
ngày đƣợc thử hoạt tính kháng Xoo (R2). Kết quả đƣợc trình bày trong hình 18
hình 29E và hình 29F. Kết quả cho thấy rằng phƣơng pháp cấy bằng giống cấp 1 tốt
hơn cấy bằng bào tử. Giống cấp 1 ở các thời điểm khác nhau không ảnh hƣởng đến
khả năng sinh hoạt chất.
0
5
10
15
20
25
30
35
25 30 35 40
Đư
ờn
g k
ính
vò
ng
kh
án
g k
hu
ẩn
(D
-d:m
m)
Nhiệt độ nuôi cấy (o C)

43
Hình 18. Ảnh hƣởng của cách cấy giống đến sự sản sinh hợp chất kháng sinh ức chế
vi khuẩn Xoo của chủng VN08 - A12.
3.4. Tinh sạch và phân tích hoạt chất kháng Xoo của chủng VN08 - A12
3.4.1. Tinh sạch hoạt chất kháng Xoo của chủng VN08 - A12
Dịch chiết thô của chủng VN08 - A12 đƣợc tách chiết bằng 1/2 thể tích ethyl
acetate nhƣ đã mô tả trong mục 2.2.6.1 và tóm tắt ở sơ đồ hình 19. Hoạt tính kháng
Xoo của dịch chiết thô đƣợc kiểm tra bằng phƣơng pháp đục lỗ thạch và khẳng định
hoạt chất đƣợc tách trong dung môi ethylacetate (hình 30A).
Dịch chiết thô tiếp tục đƣợc tinh sạch bằng cột silica gel, các phân đoạn đƣợc
thu và thử hoạt tính kháng Xoo (R2) nhƣ đƣợc trình bày trong mục 2.2.6.2. Hoạt
tính của các phân đoạn đƣợc thể hiện trong bảng 11 và hình 30B. Các phân đoạn từ
4.1 trở đi đều có hoạt tính chứng tỏ hoạt chất kháng Xoo chỉ có thể thôi khỏi cột với
nồng độ ethylacetate trên 80%.
23
24
25
26
27
28
29
24 48 72 96 1x108 2x108 3x108 4x108
Đư
ờn
g k
ính
vò
ng
kh
án
g k
hu
ẩn
(
D-d
:mm
)
Cách cấy giống
(1x108) (2x108) (3x108) (4x108)
Bào tử
Giống cấp I
( Thời gian nuôi – giờ)

44
Hình 19. Quy trình tách dịch chiết thô của chủng VN08 - A12
Bảng 11. Hoạt chất của VN08 - A12 thu đƣợc từ các phân đoạn khi tinh sạch bằng
silica gel
Phân đoạn
mẫu chủng
VN08 - A12
Nồng độ
dung môi
Thể tích
(ml)
Đƣờng kính
vòng kháng
(D-d, mm)
1.1 Hexan 100% 20 -
1.2 Hexan 100% 20 -
2.1 80% Hexan: 20% ethyl acetate 20 -
2.2 80% Hexan: 20% ethyl acetate 20 -
3.1 50% Hexan: 50% ethyl acetate 20 -
3.2 50% Hexan: 50% ethyl acetate 20 -
4.1 20% Hexan: 80% ethyl acetate 20 15
4.2 20% Hexan: 80% ethyl acetate 20 10
5.1 0% Hexan: 100% ethyl acetate 20 5
5.2 0% Hexan: 100% ethyl acetate 20 4
5.3 50% Methanol: 50% ethyl acetate 20 3
Dịch trong
(Dịch chiết thô) Dịch
nuôi
cấy (Ly tâm)
Dịch trong
(1V)
1/2V Ethyl
acetate
Ly tâm

45
Phân đoạn 4.1 có hoạt tính cao nhất nên đƣợc tiếp tục tinh sạch bằng phƣơng
pháp HPLC sử dụng cột sắc ký phản pha C18. Các phân đoạn (1 phút/ phân đoạn)
sau HPLC đƣợc thu lại và thử hoạt tính kháng Xoo (bảng 12 và hình 20). Kết quả
cho thấy phân đoạn 14 có hoạt tính cao nhất, tƣơng ứng với phút 14. Vì vậy, chất ở
phút 14 đƣợc tinh sạch (hình 21). Phổ hấp thụ của hoạt chất này đƣợc thể hiện trong
hình 22. Bƣớc sóng hấp thụ cực đại của chất là 360 nm.
Bảng 12. Hoạt chất của VN08 - A12 thu đƣợc từ HPLC từ phân đoạn 4.1
Phân đoạn mẫu
chủng VN08 - A12
Đƣờng kính vòng kháng Xoo (R2)
(D-d, mm)
1 -
2 -
3 -
4 -
5 -
6 -
7 -
8 -
9 -
10 -
11 -
12 -
13 -
14 14

46
Phân đoạn mẫu
chủng VN08 - A12
Đƣờng kính vòng kháng Xoo (R2)
(D-d, mm)
15 12
16 9
17 8
18 5
19 -
20 -
21 -
22 -
23 -
24 -
25 -
26 -
Hình 20. Phổ HPLC của hoạt chất chủng VN08 - A12
min12 13 14 15 16 17
mAU
0
250
500
750
1000
1250
1500
1750
2000
DAD1 A, Sig=254,4 Ref=450,100 (20130228NT 2013-02-28 14-11-30\1AA-0201.D)
11.
945
12.
401
12.
836
12.
992
13.
183
13.
467
13.
568
13.
857
14.
024
14.
156
14.
576
14.
723
14.
939
15.
170
15.
311
15.
425
15.
735
16.
064
16.
221
16.
651 1
7.24
6
17.
397
17.
636
17.
799

47
Hình 21. Phổ HPLC của hoạt chất sau tinh sạch
Hình 22. Phổ hấp thụ UV của hoạt chất sau tinh sạch
3.4.2. Phân tích trọng lƣợng phân tử
Trọng lƣợng phân tử của hoạt chất kháng Xoo tinh sạch từ chủng VN08 -
A12 đƣợc xác định bằng phƣơng pháp khối phổ (hình 23). Kết quả cho thấy hoạt
chất có trọng lƣợng phân tử là 778,43. Dựa trên phổ hấp phụ và trọng lƣợng phân
min5 10 15 20 25 30
mAU
0
10
20
30
40
50
60
70
DAD1 A, Sig=254,4 Ref=450,100 (2013086NT 2013-08-06 09-48-35\014-0401.D)
0.9
31
1.0
60
12.2
22
12.5
85
12.9
07
14.3
68
26.1
94
26.2
94
26.9
10
nm250 300 350 400 450 500 550
mAU
0
50
100
150
200
250
300
350
400
*DAD1, 12.380 (449 mAU, - ) Ref=11.447 & 12.967 of 012-0201.D

48
tử, chúng tôi bƣớc đầu tạm xác định hoạt chất là factumycin với cấu trúc hóa học
nhƣ trong hình 24.
Hình 23. Kết quả phân tích khối phổ của chủng VN08 - A12
Hình 24. Cấu trúc hóa học của factumycin [31]
Factumycin đƣợc phát hiện lần đầu tiên vào năm 1982 nhƣ một chất kháng
sinh thuộc cùng nhóm với aurodox, kirromycin, efrotomycin, mocimycin,
heneicomycin [21]. Factumycin có trọng lƣợng phân tử là 778,97048 g/ mol và
801.438
[M+Na]+

49
công thức phân tử là C44H62N2O10 (hình 25). Sự khác biệt giữa factumycin với các
chất trong nhóm là sự có mặt của một vòng tetrahydrofuran mở và liên kết đôi với
gốc ketone. Thêm vào đó bƣớc sóng hấp thụ của factumycin là 355 nm, dài hơn so
với các chất kháng sinh khác trong cùng nhóm (~322 nm).
Hình 25. Cấu trúc của factumycin [46]
Factumycin có khả năng kháng cả các vi khuẩn Gram âm và Gram dƣơng
[46] [21]. Năm 2011 Kyota Kimura và cộng sự đã chứng minh môi trƣờng lên men
của chủng Streptomyces spp. K07-0034 có khả năng ức chế T3SS (hệ thống tiết típ
3) của các vi khuẩn Gram âm [31]. Năm 2012, Thaker và cộng sự phát hiện ra
factumycin từ chủng xạ khuẩn Streptomyces toxytricini (WAC 5292) có hoạt tính
kháng một số loại vi khuẩn Gram âm thƣờng đƣợc tìm thấy ở mầm bệnh đa kháng
thuốc trong y tế nhƣ: Acinetobacter baumannii, Enterococcus faecalis, Escherichia
coli, Klebsiella pneumoniae, Pseudomonas aeruginosa và Staphylococcus aureus
[46]. Trong thử nghiệm của Kimura năm 2011, đã lây nhiễm trên chuột một liều gây
chết của Citrobactor rodentium - một loại vi khuẩn gây bệnh cho ngƣời tƣơng tự
Escherichia coli. Kết quả quan sát cho thấy những con chuột đƣợc uống audorox
liều lƣợng 25 mg/ kg có khả năng sống sót sau khi bị nhiễm Citrobactor rodentium
[31]. Trong một nghiên cứu in vivo về tác dụng của UK 69,753 - một glycoside của
factumycin trên chuột bị nhiễm Treponema hyodysenteriae, kết quả cho thấy không

50
còn tìm thấy Treponema hyodysenteriae trong phân của chuột thí nghiệm thậm chí
ở thời điểm 12 ngày sau khi thử nghiệm [28]. Tuy nhiên, factumycin chƣa từng
đƣợc sử dụng để kháng Xoo gây bệnh bạc lá lúa.
Những nghiên cứu về cơ chế tổng hợp factumycin ở Streptomyces sp. cho
thấy cụm gen sinh tổng hợp factumycin có kích thƣớc gần 76kb gồm 23 khung đọc
mở: facAVI, facAV, facAIV, facAIII, facAII và facAI (gen quy định PKS và NRPS–
PKS); facHIV, facHV và facHI (Hypothetical protein) và các gene khác: facP,
facRI, facT, facCII, facM, facD, facOI, facOII, facCI, facMII và facB (hình 26).
Trong đó facT là gen quan trọng nhất trong việc sinh tổng hợp factumycin [46].
Trong nghiên cứu này, factumycin đƣợc tìm thấy có trong dịch nuôi cấy
chủng VN08-A12. Thử nghiệm kháng vi sinh vật cho thấy, dịch nuôi cấy của chủng
VN08-A12 có khả năng ức chế sinh trƣởng của cả vi khuẩn Gram âm (Escherichia
coli NBRC 14237 và Xanthomonas oryzae pv. oryzae) và vi khuẩn Gram dƣơng
(Micrococcus luteus NBRC 13867). Tuy nhiên, dịch nuôi cấy của chủng VN08-
A12 có chứa factumycin không thể kháng đƣợc các vi sinh vật kiểm định khác.
Chúng tôi giả thuyết rằng factumycin sinh ra từ chủng VN08-A12 có một phổ
kháng vi sinh vật tƣơng đối hẹp. Điều này sẽ giúp ích cho việc ứng dụng chế phẩm
factumycin trong xử lý bệnh bạc lá lúa trên đồng ruộng trong khi không gây ảnh
hƣởng đến hệ vi sinh vật trong đất.
Hình 26. Cụm gen sinh tổng hợp factumycin (A) chủng WAC5292 (Streptomyces
toxytricini) và (B) Streptomyces cattleya.

51
KẾT LUẬN VÀ KIẾN NGHỊ
4.1. Kết luận
- Bằng phƣơng pháp hóa phân loại, hình thái học và phân tích trình tự gen
16S rADN, chủng VN08 - A12 đƣợc xác định là Streptomyces toxytricini.
- Điều kiện nuôi cấy tối ƣu để chủng VN08 - A12 sinh hoạt chất kháng Xoo
cao nhất đƣợc xác định nhƣ sau: 25 ml môi trƣờng SKS trong bình định mức 250
ml trong 4 ngày ở 30oC, cấy giống cấp 1 tốt hơn cấy bằng bào tử.
- Hoạt chất kháng Xoo sinh ra bởi chủng VN08 - A12 đã đƣợc tinh sạch và
xác định là factumycin.
4.2. Kiến nghị
Kết quả trong nghiên cứu cho thấy chủng VN08 - A12 (Streptomyces
toxytricini) có tiềm năng ứng dụng trong kiểm soát sinh học bệnh bạc lá lúa. Chính
vì vậy, các nghiên cứu tiếp theo sẽ tập trung vào việc cải biến di truyền chủng
VN08 - A12 để tăng khả năng sinh hoạt chất kháng Xoo và phát triển chế phẩm
kiểm soát bệnh bạc lá lúa ở Việt Nam.

52
TÀI LIỆU THAM KHẢO
TIẾNG VIỆT
1. Kiều Hữu Ảnh (1999), Giáo trình Vi sinh vật học công nghiệp, tập 1, Nhà
xuất bản Khoa học và Kỹ thuật, Hà Nội.
2. Nguyễn Lân Dũng (2012), Giáo trình Vi sinh vật học, phần 3, Nhà xuất bản
giáo dục Việt Nam.
3. Lê Lƣơng Tề, Bùi Trọng Thủy (2007). “Một số nghiên cứu về các Xa-gen
kháng bệnh bạc lá lúa”, Tạp chí Bảo vệ thực vật, 1(1), tr. 1-7.
4. Nguyễn Xuân Thành, Dƣơng Đức Tiến (2003), Vi sinh vật học nông nghiệp,
Nhà xuất bản Đại học sƣ phạm, Hà Nội.
TIẾNG ANH
5. Adhikari, T. B., Basnyat, R. C. (1999), “Virulence of Xanthomonas oryzae
pv. oryzae on rice lines containing single resistance genes and gene
combinations”, Plant Disease, 83, pp. 46-50.
6. Baltz, R. H. (2006), “Antibiotic discovery from actinomycetes: will a
renaissance follow the decline and fall?”, SIM News, 55, pp. 186-196.
7. Basavaraj, M., Shivayageeswar, E. N. (2011), “Production and optimisation
of tetracycline by various strains of Streptomyces under solid state
fermentation using pineapple peel as a novel substrate ”, Recent Research in
Science and Technology 3,pp. 01-08.
8. Bharathkumar, S., David, P. R. S., Brindha, P. V., Kavitha, S.,
Gnanamanickam, S. S. (2008), “ Improvement of bacterial blight resistance
in rice cultivars Jyothi and IR50 via markerassisted backcross breeding”,
Journal of Crop Improvement, 21, pp. 101-116.
9. Bulletin, B. O. E. (2007), “Xanthomonas oryzae”, 37, pp. 543-553.
10. Caffrey, P., Lynch, S., Flood, E., Finnan, S., Oliynyk, M. (2001),
“Amphotericin biosynthesis in Streptomyces nodosus: deductions from

53
analysis of polyketide synthase and late genes”, journal of Biology
Chemistry, 8, pp. 713-723.
11. Chand, T., Singh, N., Singh, H., Thind, B. S. (1979), “Field efficacy of stable
bleaching powder to control bacterial leaf blight”, journal of Rice Research
Newsletter, 4, pp. 12-13.
12. Das, S., Lyla, P. S., Khan, S. A. (2008), “Distribution and generic
composition of culturable marine actinomycetes from the sediments of
Indian continental slope of Bay of Bengal”, Chinese Journal
Oceanology Limnology, 26, pp. 166-177.
13. Dinh, H. D., Oanh, N. K., Toan, N. D., Du, P. V., Loan, L. C. (2008),
“Pathotype profilce of Xanthomonas oryzae pv. oryzae isoaltes from the rice
ecosystem in Cuulong river Delta”, Omonrice, 16, pp. 34-40.
14. Fravel, D. R. (2005), “Commercialization and implementation of
biocontrol”, journal of Phytopathology, 43, pp. 337-359.
15. Furuya, N., Taura, S., Thuy, B. T., Ton, P. H., Hoan, N. V., Yoshimura, A.
(2003), “Experimental technique for Bacterial Blight of Rice”, HAU-JICA
ERCB Project, 25, pp. 42-50.
16. Gnanamanickam, S. S., Sakthivel, N., Alvarez, A. M., Benedict, A. A.,
Leach, J. E. (1996), “Serological and molecular probes for the detection of
seedborne bacterial pathogens of rice and lack of demonstrable evidence for
seed transmission of Xanthomonas oryzae pv. oryzae”, journal of plant
Disease, 8, pp. 24-29.
17. Gnanamanickam, S. S., Velusamy, P., Immanuel, J. E., Thomashow, L. S.
(2006), “Biological control of rice bacterial blight by plant-associated
bacteria producing 2,4-diacetylphloroglucinol”, Canadian Journal of
Microbiology, 52, pp. 56-65.
18. Gnanamanickan, S. S. (2009), “Biological control of rice diseases”,
Biological control of rice diseases, 8, pp. 29-31.

54
19. Gonzalaz, R. I., Obregon, A. M., Escalante, L., Sanchez, S. (1995),
“Gentamicin formation in Micromonospora purpurea: stimulatory efect of
amanonium”, Journal of antibiotic, 48, pp. 479-483.
20. Gopalakrishnan, S., Kumar, R., Humayun, P., Srinivas, V., Ratnakumari, B.,
Vijayabharathi, R., Singh, A., Surekha, K., Padmavathi, C., Somashekar, N.,
Rao, R. P., Rao, L. V. S., Babu, V. R., Viraktamath, B. C., Goud, V. V.,
Loganandhan, N., Gujja, B., Rupela Oh, Y. Y. (2013), “Assessment of
different methods of rice (Oryza sativa, L) cultivation for growth parameters,
soil chemical, biological and microbiological properties, water saving and
grain yield in rice-rice system”, Paddy Water Environment, 13, pp. 362-366.
21. Gullo, V. P., Zimmerman, S. B., Dewey, R. S., Hensens, O., Cassidy, P. J.
(1982), “Factumycin, a new antinbiotic (A40A): Fermentation, isolation and
antibacterial spectrum”, Journal of Antibiotic, 12, pp. 1705- 1707
22. Hao, D., Gao, P., Liu, P., Zhao, J., Wang, Y., Yang, W., Lu, Y., Shi, T.,
Zhang, X. (2011), “A novel secretory protein, inhibits Elk1 transcriptional
activity via ERK pathway”, Molecular Biology Reports, 38, pp. 1375–1382.
23. Hoa, P. T. P., Hop, D. V., Quang, N. Q., Ton, P. H., Ha, T. H., Hung, N. V.,
Van, N. T., Hai, T. V., Quy, N. T. K., Dao, N. T. A., Thom, V. T., (2014),
“Biological Control of Xanthomonas Oryzae pv. Oryzae Causing Rice
Bacterial Blight Disease by Streptomyces toxytricini (VN08-A-12), Isolated
from Soil and Leaf-litter Samples in Vietnam ”, Biocontrol science, 19, pp.
3.
24. Hoa, P. T. P., Quang, N. D., Sakiyama, Y., Hop, D. V., Hang, D. T., Ha, T.
H., Van., N. T., Quy, N. T. K., Dao, N. T. A., (2012), “Screening for
actinomyces isolated from soil with the ability to inhibit Xanthomonas
oryzae pv. oryzae causing rice bacterial blight disease in Vietnam”, African
Journal of Biotechnology 10, pp. 1684–5315
25. Howard, T. D. (1998), “The Production of Neomycin by Streptomyces
fradiae in Synthetic Media ”, Applied Microbiology, 1, pp. 103-106.

55
26. Huang, N., Angeles, E. R., Domingo, J., Magpantay, G., Singh, S., Zhang, G.
(1997), “Pyramiding of bacterial blight resistance genes in rice; marker-
assisted selection using RFLP and PCR”, Theoretical and Applied Genetic,
95, pp. 313-320.
27. Islam, N., Bora, L. C. (1998), “Biologycal management of Bacterial leaf
blight of rice (Oryza Santiva) with plant grow promoting Rhizobacteria”,
Indian Journal of Agricultural Univercity, 12, pp. 50-63.
28. Jefson, M. R., Bordner, J., Reese, C. P., Whipple, E. B. (1989), “UK-69, 753,
a novel member of the efrotomycin family of antibiotics II. Structure
determination and biological activity”, Journal of Antibiotic, 11, pp. 1610-
1618.
29. Ji, G. H., Wei, L. F., Wu, Y. P., Bai, X. H. (2008), “Biological control of rice
bacterial blight by Lysobacter antibioticus strain 13-1”, Biological control,
45, pp. 288-296.
30. Khush, G. S., Bacalango, E., Ogawa, T. (1990), “A new gene for resistance
to bacterial blight from O. longistaminata”, Rice Genetics Newsletter, 7, pp.
121-122.
31. Kimura, K., Iwatsuki, M., Nagai, T., Matsumoto, A., Takahashi, Y., Shiom,
K., Mura, S. O., Abe, A. (2011), “A small-molecule inhibitor of the bacterial
type III secretion system protects againstin vivo infection with Citrobacter
rodentium”, Journal of Antibiotic, 64, pp. 197-203.
32. Lestari, Y. (2006), “Short communication : Identification of indigenous
Streptomyces spp. Producing antibacterial compounds”, Journal of
Mikrobiology Indonesia, 11, pp. 99-101.
33. Mew, T. W., Alvarez, A. M., Leach, J. E., Swings, J. (1993), “Focus on
bacterial blight of rice”, Plant Disease, 77, pp. 5-11.
34. Mitchell, J. I., Logan, P. G., Cushing, K. E., Ritchie, D. A. (2006),
“Novobiocin resistance sequences from the novobiocin producing strain
Streptomyces niveus ”, Molecular Microbiology, 10, pp. 1365-2958.

56
35. Miyadoh, S. (2005), “ Actinomycetes: Isolation and their antibiotic
screening”, VNU-CBT and NITE cooperation project
36. Ndonde, M. J. M., Semu, E. (2000), “ Preliminary characterization of some
Streptomyces species from four Tanzanian soils and their antimicrobial
potential against selected plant and animal pathogenic bacteria”, World
Journal of Microbiology and Biotechnology, 16, pp. 595-599.
37. Ogawa, T., Sekizawa, K. (1993), “Studies on the breeding of rice varieties
resistant to bacterial leaf blight and test of the quantitative resistance of rice
native varieties in Japan by clipping inoculation methods”, Bulletin of
Chugoku National Agricultual Experiment, 27, pp. 19-36.
38. Ou, S. H. (1985), “Rice diseases”, Biological control, 380, pp. 295-486.
39. Rafi, A., Akhtar, M. A., Akmal, M., Bibi, A., Ali, M., Rahman, H., Junaid,
M. (2013), “Effect of planting dates on bacterial leaf blight incidence
and yield performance of rice cultivars in different location of khyber
pakhtunkhwa, pakistan”, Sarhad Journal of Agriculture, 29, pp. 408-414.
40. Schatz, A., Bugie, E., Waksman, S. A. (1994), “Streptomycin, a substance
exhibiting antibiotic activity against gram-positive and gram-negative
bacteria”, journal of Biology and Medicine, 55, pp. 66-69.
41. Shang, K., Hu, H., Zhu, C., Zhu, B. (2008), “Production of 4′-
epidaunorubicin by metabolic engineering of Streptomyces coeruleorubidus
strain SIPI-1482”, Journal of Microbiology and Biotechnology, 24, pp. 1107-
1113.
42. Shapiro, S., Vining, L. C. (1983), “Nitrogen metabolism and
chloramphenicol production in Streptomyces venezuelae”, Canadian Journal
of Microbiology, 29, pp. 1706-1714.
43. Srivastava, D. N. (1972), “ Bacterial leaf blight of rice”, Phytopathol, 26, pp.
1-16.
44. Subramoni, S., Jha, G., Sonti, R. V. (2009), “Virulence functions of
xanthomonas,” Biological Control of Rice Diseases, 8, pp. 535-571.

57
45. Tagami, Y., Mizukami, T. (1962). (1962), “Historical review of the
researches on bacterial blight of rice caused by Xanthomonas oryzae (Uyede
and Ishiyama) Dowson”, Special Report of Plant Disease and Insect Pests
Forecasting Service, 10, pp. 1-112.
46. Thaker, M. N., Garcıa, M., Koteva, K., Waglechner, N., Sorensen, D.,
Medinab, R., Wright, R. D. (2012), “Biosynthetic gene cluster and
antimicrobial activity of the elfamycin antibiotic factumycin”, Medicinal
Chemistry Communications, 3, pp. 1020–1026
47. Thanh, D. T. (2006), “Identification of rice bacterial leaf blight Xanthomonas
oryzae pv. oryzae by PCR”, journal of plant science and biotechnology, 6,
pp. 44-47.
48. Unnamalai, N. (1987), “Seedborne nature of Xanthomonas campestris pv.
oryzae (Ishiyama) Dye, the bacterial leaf blight pathogen of rice”, Ph.D.
dissertation, University of Madras.
49. Vauterin, L., Hoste, B., Kesters, K., and Swings, J. (1995), “Reclassification
of Xanthomonas”, International Journal of Systematic Bacteriology, 45, pp.
472–489.
50. Vauterin, L., Rademaker, J., Swings, J. (2000), “Synopsis of the taxonomy of
the genus Xanthomonas”, Phytopathology, 90, pp. 677–682.
51. Vera Cruz, C. M., Gossele, F., Kersters, K., Segers, P., Mooter, V. D. M.,
Swings, J. (1984), “Differentiation between Xanthomonas campestris pv.
oryzae, Xanthomonas campestris pv. oryzicolaand the bacterial “brown
blotch” pathogen on rice by numerical analysis of phenotypic features and
protein gel electrophoregrams”, Journal of General Microbiology, 130, pp.
2983–2999.
52. Wang, Y. G., Davies, J. E., Hut Chinson, C. R. (1982), “Plasmid DNA in
the erythromycin producing microorganism streptomyces erythreus NBRC
2338”, Journal of Antibiotic,, 35, pp. 335-342.

58
53. Weissbach, H., Redfield, B., Beaven, V., Katz, E. (1965), “Actinomycin
synthesis in washed cells of Streptomyces antibioticus”, journal of Biology
Chemistry, 240, pp. 4377-4381.
54. Yoneyama, K., Sekido, S., Misato, T. (1978), “Studies on the fungicidal
action of dithiocarbamates 3 effect of sodium dimethyldithiocarbamate on
fatty acid synthesis of Xanthomonas oryzae”, Annals of the
Phytopathological Society of Japan, 44, pp. 313-320.

- 1 -
PHỤ LỤC
1. Phân tích thành phần axit amin
Dung môi chạy sắc ký bản mỏng TLC (ml): Methanol - 80; nƣớc cất - 17,5;
6 N HCl - 4, Pyridine - 10.
Điều kiện chạy HPLC: Sử dụng UV detector ở bƣớc sóng 254 nm. Dung
dịch (A) 6% CH3CN, dung dịch (B) 60% CH3CN, tốc độ dòng chảy: 1 ml/ phút
Chƣơng trình chạy HPLC:
Bảng 13. Chƣơng trình chạy HPLC phân tích thành phần axit amin
Thời gian (phút) Nồng độ dung dịch B (%)
0 0
15 70
25 100
30 100
0 Dừng
2. Phân tích thành phần menaquinone
Dung môi chạy TLC: Toluen.
Chất chuẩn: Vitamin K. Vạch ngang với vitamin K là ubiquinone.

- 2 -
Điều kiện chạy HPLC: Dung môi (Methanol : Propanol = 2 : 1). Tốc độ dòng
chảy 0,2 ml/ phút. Nhiệt độ cột 40oC. Nhận biết bằng UV detector với bƣớc sóng
275 nm.
Bảng 14. Phân tích thành phần menaquinone chuẩn
Pick Thời
gian
lƣu
Diện
tích
peak
Chiều
cao peak
Phần trăm
diện tích
peak (%)
Phần trăm
chiều cao
peak (%)
Thành phần
menaquinone
chuẩn
1 11,326 201944 12461 11,182 13,675 MK - 9 (H2)
2 12,709 570803 28753 31,605 31,555 MK - 9 (H4)
3 14,031 732861 36762 40,578 40,344 MK – 9 (H6)
4 15,682 300428 13146 16,635 14,427 MK - 9 (H8)
Tổng 1806036 91122 100,000 100,000
Hình 27. Sắc ký đồ thành phần menaquinone chuẩn
MK - 9 (H6) MK - 9 (H8) MK - 9 (H4) MK - 9 (H2)

- 3 -
3. Phân tích thành phần axit béo
Dung dịch: R1: NaOH 15 g, Methanol 50 ml, MQ 50 ml
R2: 6N HCl 65 ml, Methanol 55 ml, pH 1,5
R3: n- hexane 50 ml, methyl-ter-butyl ethel 50 ml
R4: NaOH 10,8 g, MQ 900 ml
R5: 100 ml MQ + 40 g NaCl
Bảng 15. Phân tích thành phần axit béo chuẩn
Thời gian lƣu Axit béo Tỷ lệ phần trăm (%)
6,962 14:0 iso 2,11
7,491 14:0 1,15
8,462 15:0 iso 7,71
8,603 15:0 anteiso 22,49
9,049 15:0
9,820 16:1 iso H 4,85
10,101 16:0 iso 17,32
10,421 12,98
10,723 16:0 6,96
11,453 4,13
11,641 17:1 anteiso 6,90
11,820 17:0 iso 1,83
11,982 17:0 anteiso 9,03

- 4 -
Thời gian lƣu Axit béo Tỷ lệ phần trăm (%)
12,107 17:1 w8c 0,79
13,184 17:0 10methyl 0,59
13,291 18:1 iso H 0,68
13,917 0,48
Hình 28. Sắc ký đồ thành phần axit béo chuẩn
4. Phân tích thành phần G+C trong ADN
Đệm acetate (40 mM, pH 5,3): 2 mM ZnSO4 25 μl + dung dịch nuclease P1
(20 U/ ml) 25 μl.
5. Trình tự các mồi đƣợc sử dụng trong phân tích trình tự 16S rADN

- 5 -
Bảng 16. Trình tự mồi dùng cho phản ứng đọc trình tự ADNr 16S
Mồi Trình tự 5’-3’
27F AGAGTTTGATCCTGGCTCAG
1525R AAAGGAGGTGATCCAGCC
780F GAATTGATACCCTGGTAG
350R CTGCTGCCTCCCGTAG
1100F GCAACGAGCGCAACCC
920R GTCAATTCCTTTGAGTTT
6. Tối ƣu hóa điều kiện nuôi cấy của chủng VN08-A12

- 6 -
3M
SKS
N8
301 2M
16M ĐC
7N
6N 5N
4N
3N
ĐC
125ml
150ml
ĐC
25ml
75ml
100ml
50ml
35o
C
30oC
40oC
ĐC
1x108 2x10
8
3x108
4x108
24h 48h
72h 96h
A) B)
B)
C)
)
D)
E) F)

- 7 -
Hình 29. Hoạt tính kháng Xoo của VN08 - A12 trong các điều kiện nuôi cấy khác
nhau
(A). Thể tích nuôi cấy khác nhau (25, 50, 75, 100, 125, 150 ml), ĐC (-)
(B). Thời gian nuôi cấy khác nhau ( 3, 4, 5, 6, 7 ngày), ĐC (-)
(C). Môi trƣờng nuôi cấy khác nhau (3M, 16M, 2M, 301, N8, SKS), ĐC (-)
( D). Nhiệt độ nuôi cấy khác nhau ( 30, 35, 40oC), ĐC (-)
(E). Cấy giống bằng bào tử (1x108, 2x10
8, 3x10
8, 4x10
8 bào tử)
(F). Giống cấp I ( 24, 48, 72, 96 giờ)
7. Tách chiết và tinh sạch
(A). Dịch chiết thô sau tách bằng ethylacetate ( thử hoạt tính lặp lại)
(B). Các phân đoạn sau cột silica gel
8. Trình tự gen 16S rADN của chủng VN08 - A12:
TGCAAGTCGAACGATGAACCTCCTTCGGGAGGGGATTAGTGGCGAACG
GGTGAGTAACACGTGGGCAATCTGCCCTTCACTCTGGGACAAGCCCTGG
Hình 30. Hoạt chất kháng sinh của chủng VN08 - A12
4.1
4.2
5.1
5.2
5.3
B) A)
2
1
3
4
5
6

- 8 -
AAACGGGGTCTAATACCGGATACGACTGCGGAAGGCATCTTCCGCGGTG
GAAAGCTCCGGCGGTGAAGGATGAGCCCGCGGCCTATCAGCTTGTTGGT
GGGGTAATGGCCTACCAAGGCGACGACGGGTAGCCGGCCTGAGAGGGC
GACCGGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGC
AGCAGTGGGGAATATTGCACAATGGGCGAAAGCCTGATGCAGCGACGC
CGCGTGAGGGATGACGGCCTTCGGGTTGTAAACCTCTTTCAGCAGGGAA
GAAGCGAAAGTGACGGTACCTGCAGAAGAAGCGCCGGCTAACTACGTG
CCAGCAGCCGCGGTAATACGTAGGGCGCAAGCGTTGTCCGGAATTATTG
GGCGTAAAGAGCTCGTAGGCGGCCAGTCACGTCGGATGTGAAAGCCCG
AGGCTTAACCTCGGGTCTGCATTCGATACGGGCTGGCTAGAGTGTGGTA
GGGGAGATCGGAATTCCTGGTGTAGCGGTGAAATGCGCAGATATCAGG
AGGAACACCGGTGGCGAAGGCGGATCTCTGGGCCATTACTGACGCTGA
GGAGCGAAAGCGTGGGGAGCGAACAGGATTAGATACCCTGGTAGTCCA
CGCCGTAAACGTTGGGAACTAGGTGTTGGCGACATTCCACGTCGTCGGT
GCCGCAGCTAACGCATTAAGTTCCCCGCCTGGGGAGTACGGCCGCAAGG
CTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGCGGAGCATG
TGGCTTAATTCGACGCAACGCGAAGAACCTTACCAAGGCTTGACATATA
CCGGAAAGCATTAGAGATAGTGCCCCCCTTGTGGTCGGTATACAGGTGG
TGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTGGGTTAAGTCCCGC
AACGAGCGCAACCCTTGTCCTGTGTTGCCAGCATGCCCTTCGGGGTGAT
GGGGACTCACAGGAGACCGCCGGGGTCAACTCGGAGGAAGGTGGGGAC
GACGTCAAGTCATCATGCCCCTTATGTCTTGGGCTGCACACGTGCTACA
ATGGCCGGTACAATGAGCTGCGATACCGTGAGGTGGAGCGAATCTCAA
AAAGCCGGTCTCAGTTCGGATTGGGGTCTGCAACTCGACCCCATGAAGT
CGGAGTCGCTAGTAATCGCAGATCAGCATTGCTGCGGTGAATACGTTCC
CGGGCCTTGTACACACCGCCCGTCACGTCACGAAAGTCGGTAACACCCG
AAGCCGGTGGCCCAACCCTTGTGGAGG

African Journal of Biotechnology Vol. x(xx), pp. xxx-xxx, xx xxxxx, 2012 Available online at http://www.academicjournals.org/AJB DOI: ISSN 1684–5315 ©2012 Academic Journals
Full Length Research Paper
Screening for actinomyces isolated from soil with the ability to inhibit Xanthomonas oryzae pv. oryzae causing rice bacterial blight disease in Vietnam
Phan Thi Phuong Hoa1,4*, Nguyen Duc Quang2, Yayoi Sakiyama3, Duong Van Hop1, Dinh Thuy Hang1, Trinh Hoang Ha4, Nguyen Thi Van1, Nguyen Thi Kim Quy1 and Nguyen Thi Anh Dao1
1Institute of Microbiology and Biotechnology (IMBT), Vietnam National University, Hanoi (VNUH), Vietnam.
2University of Engineering and Technology, Vietnam National University, Hanoi, Vietnam.
3NITE Biological Resource Center (NBRC), National Institute of Technology and Evaluation (NITE), Kazusakamatari
2-5-8, Kisarasu, Chiba, 292-0818, Japan. 4School of Medicine and Pharmacy, Vietnam National University, Hanoi, Vietnam.
Accepted 3 August, 2012
Bacterial blight disease caused by Xanthomonas oryzae pv. oryzae (Xoo) is one of the major diseases in rice culture of Northen Vietnam, as well as other rice-growing regions of the world. In this study, we isolated and screened for actinomycete strains from Vietnam with the ability to inhibit Xoo isolates from northern Vietnam. From 90 actinomycete strains taken from soil in northern Vietnam in 2010, we screened for their antagonistic activity against 10 races of Xoo causing rice bacterial blight disease. Three actinomycete strains were found to inhibit all 10 Xoo races. Among the three strains, a strain namely VN10-A-44 was shown not to have the ability to produce toxic compounds and was selected for further study. The strain was identified as Streptomyces virginiae by 16S ribosomal RNA gene sequencing. We replaced soybean meal with tofu waste in antibiotic producing medium to improve antagonistic activity of VN10-A-44 against the Xoo pathogen and to make use of tofu waste for large-scale fermentation of VN10-A-44. We found that replacing soybean meal with 20 and 30 g of tofu waste/litter in the antibiotic producing medium gave the largest inhibition zone against the Xoo pathogen. Key words: Xanthomonas oryzae pv. oryzae, rice bacterial blight disease, Streptomyces virginiae, Vietnam.
INTRODUCTION Rice is one of the most important food crops of the world. However, its yield is constantly affected by several major diseases such as bacterial blight (BB), blast and tungro (Dai et al., 2007). Among these diseases, BB caused by Xanthomonas oryzae pv. oryzae (Xoo) is one of the major diseases in Vietnam as well as other rice-growing regions of the world. Bacterial blight disease in rice has reduced Asia’s annual rice production by as much as 60% (Dai et al., 2007). There are many methods and strategies which aim at reducing losses in rice yield and *Corresponding author. E-mail: [email protected] or [email protected].
avoiding outbreaks of disease (such as rice breeding programs and developing BB-resistant rice cultivars, and using chemicals to treat the bacterial pathogen). Chemical control of rice bacterial blight was evidently an ineffective method (Gnanamanickam, 2009; Hastuti et al., 2012). In addition, breeding for BB-resistance, which uses a single major gene, has proved unsuccessful. This is due to rapid evolution of sub-populations overcoming the resistance genes of rice (Gnanamanickam, 2009).
Research by Gnanamanickam (2009) in India and the Philippines have shown that Pseudomonas fluorescens (Velusamy et al., 2006) and some Bacillus (Bacillus spp., Bacillus lentus, Bacillus cereus và Bacillus circulans) (Velusamy, 2002; Velusamy and Gnanamanickam, 2003) strains isolated from rice rhizo-shere samples, inhibited

Table 1. List of races of Xoo used for antagonistic testing.
S/N Xoo race Isolated from rice cultivar Collected from province
1 R1 Khang dan, Bac thom 7 Bac Ninh, Ha Noi
2 R2 Nep tan, HT1 Son La, Hai Duong
3 R3 Nhi uu 838, Huong com Nghe An, Thai Binh
4 R4 Thuc Hung Thai Binh
5 R5 Khang dan, Bac thom Nghe An, Hai Duong
6 R6 Nhi uu 383 Yen Bai
7 R7 Q5, Nang huong Hai Duong, Thai Binh
8 R8 Nep thom Hai Duong
9 R9 Q5 Hai Duong (appeared in Japan)
10 R10 Hybrid rice Yen Bai (appeared in Japan)
the growth of Xoo in the laboratory. From Bacillus strains, Gnanamanickam (2009) and Vasudevan (2002) showed that they can reduce BB in rice from 21 to 59% (as compared to untreated plots). In 2008, Ji and his colleagues (2008) reported that Lysobacter antibioticus isolated from the rhizoshere of rice in the Yunnan province of China can significantly inhibit the growth of various phytopathogenic fungi and bacteria including Xoo. This strain is able to inhibit Xoo growth, with disease suppression efficiency up to 69.7%. Disease suppression efficiency by Lysobacter antibioticus 13-1 on Xoo varied significantly among various ice cultivars. Additionally, this efficiency of bio-control is affected by different pathogens of Xoo isolates. These results suggest that antibiotics and density of L. antibioticus 13-1 colonization on rice leaves may be involved in biocontrol of rice BB (Ji et al., 2008).
Recently, research has shown that some species of the genus Streptomyces fight against some plant pathogenic microorganisms, including Xoo (Cazorla et al., 2007; Chithrashree et al., 2011; Ndonde and Semu, 2000; Park et al., 2011; Rizk et al., 2007). For examples, bottromycin A2 and dunaimycin D3S from Streptomyces bottropensis suppressed rice bacterial blight (Park et al., 2011). Applying selected actinomycete strains and their bioactive compounds for biological control (using a balance of microorganisms and their natural components to suppress pathogens and to favour plants) of bacterial blight in rice, therefore, appears to be cost-effective and ecologically conducive strategy (Gnanamanickam, 2009).
In this study, we aimed to isolate and screen for action-mycete strains that can inhibit races of Xoo from northern Vietnam and those less harmful to other microorganisms. Additionally, we tested the possibility of replacing soybean meal in antibiotic-producing medium with tofu waste to improve inhibition ability of the actinomycete strain and to make use of the waste byproduct excreted in tofu production.
MATERIALS AND METHODS
Isolation of actinomycete strains
In 2010, 12 soil samples were collected from Lang Biang mountain,
Da Lat City and Cat Tien National Park, near Ho Chi Minh City, and a plant root from the Vietnam National University, Hanoi City, Vietnam. We isolated actinomycete strains by using the sodium dodecyl sulfate-yeast extract dilution method and the rehydration-
centrifugation method (Hayakawa et al., 1989; Hayakawa et al., 2000; Hop et al., 2011).
There were a total of 90-actinomycete strains isolated. These strains were preserved at the Vietnam Type Culture Collection in Vietnam as well as NITE in Japan. They were used to screen for actinomycete strains that can inhibit races of Xoo causing rice bacterial blight disease in Vietnam. Xoo races A total of 10 races of Xoo, namely R1, R2, R3, R4, R5, R6, R7, R8, R9 and R10, that were provided by Prof. Phan Huu Ton from Hanoi University of Agriculture, were used as the test microogarnisms. The 10 races were isolated from rice leaves in northern Vietnam. Distribution of the races are listed in Table 1 and Figure 1. It was reported that pathovars R2 and R3 were the most widely distributed in many provinces in northern Vietnam. R5 has caused the greatest loss in rice production; and R9 and R10 were isolated in Vietnam and appeared in Japan (Unpublished data). Media
Five media were used in this study. For culture and preservation of the 10 races of Xoo, Wakimoto medium was used with the following
contents: potato, 300 g; NaHPO4.12H2O, 2 g; Ca(NO3)2.4H2O, 0.5 g; peptone, 5 g; saccarose, 15 g; agar, 15 g; distilled water, 1 L and pH, 7.0 while yeast extract–soluble starch medium (YS medium) (soluble starch, 10 g; yeast extract, 2 g; agar, 15 g; distilled water, 1 L and pH, 7.3) and antibiotic producing medium (APM) (starch, 10 g; glucose, 10 g; soybean meal, 10 g; CaCO3, 3 g; peptone, 10 g; agar, 20 g; Tween 80, one drop; distilled water, 1 L; pH, 7.0) were used for the actinomycete strains. Yeast-extract malt medium (YM
medium) (malt extract, 3 g; yeast extract, 3 g; glucose, 10 g; peptone, 5 g; agar, 17 g; distilled water, 1 L; pH, 7.0) was used for culture of Candida albicans and Muller Hinton agar (MHA) was used for culture of Micrococcus luteus and Escherichia coli in antagonism and toxic-testing assays. All the chemicals and media were purchased from the companies of Sigma and BD.
Assay for antagonism to Xoo races
Antimicrobial activity of actinomycete strains was carried out by the agar-piece method (φ 6 mm) on Wakimoto medium. A total of 90

Figure 1. Distribution of the 10 Xoo races in north Vietnam. Pathogen types: 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 are races of Xoo: R1,
R2, R3, R4, R5, R6, R7, R8, R9 and R10, respectively.
actinomycete strains were screened for their antogonism to races of Xoo. Pure cultures of actinomycete isolates were inoculated on YS medium at room temperature (25 to 30°C) for 7 to 14 days. After adequate growth of the isolates, the pieces of agar (φ 6 mm) that contained actively-growing actinomycete isolates were cut and placed onto the test plates culturing races of Xoo. The plates were incubated at 28 ± 1°C for 2 to 4 days and the inhibitory effect of the actinomycete isolates on Xoo growth was evaluated by measuring
inhibition zones (cm). To prepare the test plates, races of Xoo were cultured in
Wakimoto broth to a concentration of ca 108 CFU/ml. The 150 ml of
Wakimoto agar was autoclaved and cooled down to 45 to 50°C and then mixed with 10 ml of the Xoo cultures. This mixture was poured into Petri disks and used as the test plates.
Since race R2 and R3 were the most abundant races causing rice bacterial blight disease in northern Vietnam, their inhibition by
actinomycete isolates was first tested and measured (Unpublished data). Subsequently, the actinomycete isolates inhibiting both the 2 races of R2 and R3 were tested continuously for their inhibition to the remaining 8 races of pathogens. Isolates that were able to inhibit all the 10 races of Xoo were selected for futher study.
Primary test for toxicity producer
To understand whether the actinomycete isolates inhibiting all the 10 races of pathogen produced toxic coumponds or not, we checked their inhibition of four microogarnisms: Micrococcus luteus (NBRC 13867), E. coli (NBRC 14237), Bacillus subtilis (NBRC 3134) and Saccharomyces cerevisiae (NBRC 10217) (these micro-organisms were kindly provided by NITE, Japan) by using the agar-piece method, as mentioned above. Once the isolate inhibited all the four test microorganisms, it indicated the possibility of producing
toxic compounds (Blackburn and Avery, 2003; Fredrickson and Elliott, 1985). Following the toxicity production test, we found that a specific strain, namely VN10-A-44, inhibited B. subtilis only and it
was therefore selected for further study. Replacing soybean meal with tofu waste in antibiotic producing medium
Tofu waste was replaced with soybean meal in the original antibiotic producing medium (APM) at the same rate (1:1). VN10-A-
44 was cultured on MAPM broth for 7 days at 30°C with shaking for 150 cycles/minute. The cultured broth (100 μl per well) was placed on the test medium seeded with R2 with the method mentioned previously in the materials and methods. After incubation for 24 to 72 h, the results were quantified by the diameter of the inhibition zone (cm).
Preliminary extraction tests of antibiotic produced by VN 10-A44
VN10-A44 was cultured in 50 ml of APM broth at 28 to 30°C using a shaker with 120 rpm rotation for 7 days. The culture was centrifuged at 4°C, 8000 rpm for 15 min to separate the cells and the supernatant. First, we examined whether there were antibiotic remains in the cells or were excreted to culture broth. 100 μl of the supernatant and only cells were placed in wells of the test plates
seeded with the R2 and R3 pathogen. We found that the active compound remaining in cells and the supernatant failed to inhibit any of the race pathogens. Therefore, cells of VN10-A-44 were extracted by 50% acetone. Thereafter, preliminary tests such as dilution assay, stability test and a solvent extraction test were performed to examine the following from the cell-extracted solution:
Dilution assay of the cell extract
The cell extract was diluted 2 and 4 times and 100 μl of original cell extract and the diluted treatments were placed on test plates

Table 2. Abilities of actinomycete strains to inhibit races of Xoo.
Number of Xoo races 0 1 2 3 4 5 6 7 8 9 10
Number of actinomycete isolates
77 (85.6%)
0
(0%)
1
(1.1%)
0
(0%)
1
(1.1%)
0
(0%)
0
(0%)
5
(5.6%)
2
(2.2%)
1
(1.1%)
3
(3.3%)
Figure 2. Inhibition of actinomycete isolates to Xoo races
No
. o
f ac
tin
om
yce
teis
ola
tes
inh
ibit
each
Xo
ora
ce
Xoo races
0
2
4
6
8
10
12
14
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
Figure 2. Inhibition of actinomycete isolates to Xoo races
No
. o
f ac
tin
om
yce
teis
ola
tes
inh
ibit
each
Xo
ora
ce
Xoo races
0
2
4
6
8
10
12
14
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
No
. o
f ac
tin
om
yce
teis
ola
tes
inh
ibit
each
Xo
ora
ce
Xoo races
0
2
4
6
8
10
12
14
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
Xoo races
No
. o
f acti
no
mycete
iso
late
s
that
inh
ibit
ed
each
Xo
o r
ace
Figure 2. Inhibition of Xoo races by actinomycete isolates.
seeded by R2 race in triplicate. After incubation for 24 to 72 h, results were read by the diameter of the inhibition zone.
pH and heat stability test A volume of 1 ml cell extract/1.5 ml-Eppendorf tube was boiled at 100°C for 10 min. On the other hand, the cell extract solution was adjusted to pH 2, 7 and 10. 100 μl of the treated solutions were used for antagonistic activity assays in the R2 race. Solvent extraction test
The cell extract by acetone was extracted again by ethyl-acetate and n-butanol at pH 2 and 10. 100 μl of the treated solution (top and bottom of ethyl acetate extract at pH 2 and 10; top and bottom of n-butanol extract at pH 2 and 10) were used for antagonistic activity assays in the R2 race. Identification of VN 10-A44
Identification of VN-A-44 was based on morphological, chemical and molecular approaches (Sakiyama et al., 2009). The VN10-A-44 was identified by 16S rRNA gene sequencing. Genomic DNA extraction was carried out using a Promega (Madison,WI, USA) extraction kit according to the manufacturer’s protocol. The 16S ribosomal RNA (rRNA) gene was amplified by PCR using TaKaRa Ex Taq (Takara Bio, Otsu City, Shiga, Japan) with the primers, 9F
(5'-GAGTTTGATCCTGGCTCAG-3') and 1541R (5'-AAGGAGGTGATCCAGCC-3'), or occasionally 1510R (5'-GGCTACCTTGTTACGA-3'). Almost all the entire sequence of the
16S rRNA gene (1300 to 1400 bp) was amplified by PCR as reported by Tamura et al. (1999) and directly sequenced using an ABI Prism BigDye Terminator cycle sequencing kit (Applied Biosystems, Foster City, CA, USA) and an ABI Model 3730
automatic DNA sequencer. The 16S rRNA gene sequence was compared with other sequences in the EMBL/GenBank/DDBJ database using BLAST searches and in the EzTaxon16 database, which includes only type strain sequences.
RESULTS Screening for the actinomycete strains capable of inhibiting Xoo races Results are summarized in Table 2 and Figure 2. We found that 13 among 90 isolates (14.4%) investigated were able to inhibit at least two races of Xoo. Three (3.3%) isolates were able to inhibit all the 10 races of pathogen (Tables 2 and 3).
All the 13 (100%) selected strains were able to inhibit pathogen races R1 and R3; whereas 12/13 (92.3%) inhibited R4 and R5; and 11/13 (84.6%) inhibited R7 and R8. Six among 13 selected strains inhibited R6 and four inhibited R9 (Figure 2). Only 3/13 actinomycete strains could inhibit the pathogen race R2. The actinomycete strains that inhibited race R2 (VN10-A-23, VN10-A-44 and VN10-A-54) were also able to inhibit race R9 (Table 3).

Table 3. Antagonistic activities of actinomycete-tested isolates.
Actinomycete strain
Inhibition zones against to a Xoo race (D-d) cm
R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
VN10-A-15 2.0 - 1.2 2.8 3.5 - 2.0 2.6 - 2.0
VN10-A-16 0.9 - 1.5 1.2 1.9 0.2 0.3 1.2 0.1 0.1
VN10-A-19 0.9 - 1.1 0.4 0.3 - - - - -
VN10-A-23 0.9 0.3 2.3 1.1 1.8 0.1 0.9 1.2 0.1 0.2
VN10-A-24 2.4 - 2.7 2.5 1.7 0.1 1.0 1.5 - 2.0
VN10-A-30 1.1 - 1.2 1.1 1.5 - 0.7 0.5 - 1.0
VN10-A-38 1.8 - 1.8 1.5 1.7 - 1.3 1.3 - 1.3
VN10-A-39 1.1 - 0.7 0.7 1.3 - 0.8 0.8 - +
VN10-A-44 2.5 0.1 2.5 1.5 2.5 0.2 1.9 1.9 0.3 2.0
VN10-A-58 2.6 0.1 0.1 1.8 1.8 0.5 1.7 1.3 0.1 0.1
VN10-A-74 0.5 - 1.8 - - - - - - -
VN10-A-76 3.5 - 0.8 1.1 0.9 0.1 1.5 0.8 - 0.9
VN10-A-77 2.0 - 1.1 1.8 2.3 - 1.1 1.1 - +
The inhibition zones were measured by (D-d) cm; the plus denotes that actinomycete strains have positive results but these were not large enough to be measured; the minus denotes that actinomycete strains did not inhibit the pathogen races.
Table 4. Antagonistic activities of the selected strains.
Microorganism Actinomycete strain
VN10-A-44 (D-d) cm VN10-A-23 (D-d) cm VN10-A-58 (D-d) cm
B. subtilis 1.4 ± 0.2 1.3 ± 0.4 2.1 ± 0.1
E. coli - + +
S. cerevisiae - 4.7 ± 0.3 0.9 ± 0.1
M. luteus - + 3.0 ± 0.2
The inhibition zones were measured by (D-d) cm; the plus denotes that actinomycete strains have positive results but these were not large enough to be measured; the minus denotes that actinomycete strains did not
inhibit the microorganisms; ±, indicates standard deviation.
Primary tests for toxicity producer We selected 3 strains that inhibited all the 10 races of Xoo for primary toxicity production test. The 3 strains were tested for their inhibition of 4 indicator micro-organisms (B. subtilis, M. luteus, E. coli and S. cerevisiae). We found that VN10-A-23 and VN10-A-58 could inhibit all the 4 tested microorganisms; whereas VN10-A-44 inhibited B. subtilis only and was therefore chosen for further study (Table 4). Enhancing antibiotic production of VN10-A-44 by replacing soybean meal with waste of tofu in antibiotic producing medium VN10-A-44 inhibits all the 10 pathogen races; however, the inhibition zone for R2 was very small (Figure 3). Tofu waste is abundant in Vietnam and some other countries in Asia. The APM with tofu waste is called modified APM
(MAPM) and it is used to test whether tofu waste can be recycled for producing antibiotics of VN10-A-44. If so, tofu waste may be used for large-scale production of the antibiotics.
Result shows that the inhibition zone was improved (Table 5) when VN10-A-44 was cultured in broth APM (Figure 3B), and largest in broth MAPM (Figure 3C). The amount of tofu waste for replacing soybean meal was determined (Table 6). We found that 20 and 30 g of tofu waste/litter gave the highest inhibition zones for R2. On a daily basis, large amounts of tofu waste were excreted at tofu production sites in Vietnam. We examined the influence of the life-time of tofu waste on the results of R2 inhibition. The results are recorded in Figure 4. We found that 4-day old tofu waste was the best to produce antibiotics against race R2.
Research on preliminary extraction tests of antibiotics produced by VN 10-A-44 showed that the active compound was in n-butanol cells and a pH of 2 was the best conditions for extract. The active compound lost its

(A) (B) (C)
Figure 3. The inhibition of VN10-A-44 to Xoo R2
A, broth YS medium; B, broth APM; C, broth MAPM
(A)(A) (B)(B) (C)(C)
Figure 3. The inhibition of VN10-A-44 to Xoo R2
A, broth YS medium; B, broth APM; C, broth MAPM
Figure 3. The inhibition of Xoo R2 by VN10-A-44. A, Broth YS medium; B, broth APM; C, broth MAPM.
Table 5. Inhibition zones (D-d) cm of VN10-A44 against 10 Xoo races in YS, APM and MAPM.
Medium R1 R2 R3 R4 R5 R6 R7 R8 R9 R10
YS 2.5 0.1 2.5 1.5 2.5 0.2 1.9 1.9 0.3 2.0
APM 1.9 0.6 1.5 1.8 2.2 1.0 1.3 1.7 1.0 1.7
MAPM 2.5 1.4 2.2 2.0 2.5 1.0 2.0 2.0 1.2 2.0
YS, Yeast-extract medium; APM, antibiotic-producing medium; MAPM, modified antibiotic producing medium.
Table 6. Antagonistic activities of VN10-A-44 against Xoo R2 with different amounts of tofu waste in broth
MAPM.
Medium and different amounts of tofu waste Inhibition zone (D-d) cm
Test 1 Test 2 Test 3 Average
YS 0.1 0.1 0.1 0.1
APM (10 g/ littler or 1% soybean meal) 0.7 0.8 0.8 0.8
MAPM (10 g/ littler or 1% tofu waste) 1.3 1.3 1.4 1.3
MAPM (20 g/ littler or 2% tofu waste) 1.3 1.4 1.4 1.4
MAPM (30 g/ littler or 3% tofu waste) 1.3 1.4 1.5 1.4
MAPM (40 g/ littler or 4% tofu waste) 1.4 1.4 1.4 1.4
MAPM (50 g/ littler or 5% tofu waste) 1.2 1.3 1.4 1.3
MAPM (60 g/ littler or 6% tofu waste) 1.2 1.2 1.3 1.2
MAPM (70 g/ littler or 7% tofu waste) 1.2 1.3 1.3 1.3
MAPM (80 g/ littler or 8% tofu waste) 1.2 1.2 1.2 1.2
MAPM (90 g/ littler or 9% tofu waste) 1.4 1.3 1.3 1.3
MAPM (100 g/ littler or 10% tofu waste) 0.9 0.9 1.0 0.95
MAPM (130 g/ littler or 13% tofu waste) 0.9 0.9 1.2 1.0
MAPM (150 g/ littler or 15% tofu waste) 1.0 1.0 1.0 1.0
MAPM (200 g/ littler or 20% tofu waste) 0.9 0.9 1.0 0.9
inhibitionactivity against the pathogen when being treated at 100°C.
Identification of VN-A-44 by analyzing the 16S- DNA sequence showed that the VN10-A-44 strain was closest to Streptomyces virginiae (100%, 1464/1464 bp, identity to accession number AB184175) (Figure 6). Morphology of VN10-A-44 was recorded and is shown in Figure 5. It belongs to Streptomyces with spore chains.
DISCUSSION Biocontrol of Xoo was recently reported in Asia (Gnanamanickam 2009; Ji et al., 2008; Park et al., 2011). With an attempt to select a biological agent from strains of actinomycete, in this study, we screened for actinomyce strains inhibiting all the 10 races of Xoo, whereas at the same time selecting for strains that were

0
0.5
1.0
1.5
2.0
2.5
1 2 3 4 5 6 7 8 9
Days
Inh
ibit
ion z
on
es
(cm
)
Figure 4. Antagonistic actives of VN10-A-44 depend on life-time points of tofu waste after production
0
0.5
1.0
1.5
2.0
2.5
1 2 3 4 5 6 7 8 9
Days
Inh
ibit
ion z
on
es
(cm
)
0
0.5
1.0
1.5
2.0
2.5
1 2 3 4 5 6 7 8 9
Days
Inh
ibit
ion z
on
es
(cm
)
Figure 4. Antagonistic actives of VN10-A-44 depend on life-time points of tofu waste after production
Days
Figure 4. Antagonistic actives of VN10-A-44 depending on life-time points of tofu waste after production.
Figure 5. Morphology of VN10-A-44. a, colony; b, spore
b) a)
Figure 5. Morphology of VN10-A-44. a, Colony; b, spore.
not harmful to other micro-biota and humans. Among 90-tested strains, we selected 13 actinomyces isolates that were able to inhibit from one to 10 races of Xoo. This number of actinomyces strains inhibiting races of Xoo (14.4%) was not high, and this result is in agreement with another study showing lower numbers of Streptomyces strains capable of inhibiting Xoo; whereas most of streptomyces isolates inhibited Xanthomonas vascatoria and Clavibacter michiganensis ssp. michiganensis (Ndonle and Semu, 2000). In the present study, we found that the R1 and R3 races of Xoo were inhibited by 13 actinomyces strains; whereas R2 and R9 were the most difficult to inhibit. Most of the actinomyces strains can inhibit at least 2 races of Xoo (R1 and R3), however only 3 strains (VN10-A-23; VN10-A-44; VN10-A-58) were able to inhibit all the 10 races.
Antibiotic production of actinomycete-inoculated soils
was demonstrated in soils supplemented with a suitable organic source. Stevenson (1959) found that fresh grass (3%), clover (3%) and soybean meal (2%) were suitable supplements for antibiotic production by the majority of the actinomycetes. In this study, we showed that replacing soybean meal in APM with tofu waste (from 2 to 4%) gave the largest inhibition zones (1.4 cm) against race R2; especially the 4-day old tofu waste (inhibition zones up to 2.0 cm). This could be due to the fact that the nutrient values obtained from fresh soybean found in tofu waste were much higher than those of soybean meal. To make soybean meal, soybean was dried by heating and then made into powder. This process reduced many vitamins and some essential amino acids which are very important for the development of actinomycetes from fresh soybean. If tofu waste is used for fermentation of VN10-A-44, it may reduce the cost of large-scale

Streptomyces virginiae (AB184175)
VN-10A44
Streptomyces subrutilus (X80825)
Streptomyces setonii (D63872)
Streptomyces bikiniensis (X79851)
Streptomyces galbus (X79852)
Streptomyces lincolnensis (X79854)
Streptomyces diastatochromogenes (D63867)
Streptomyces bottropensis (D63868)
Streptomyces scabies (D63862)
Streptomyces acidiscabies (D63865)
Streptomyces griseocarneus (X99943)
Streptomyces rimosus (X62884)
Streptomyces mashuensis (X79323)
Streptomyces sampsonii (D63871)
Streptomyces glaucescens (X79322)
Streptomyces pseudogriseolus (X80827)
Streptomyces coelicolor (X60514)
Streptomyces ambifaciens (M27245)
Streptomyces caelestis (X80824)
Streptomyces bluensis (X79324)
Streptomyces espinosus (X80826)
Streptomyces megasporus (Z68100)
81
99
90 76
73
80
72
90
92
98
99 0.005
Figure 6. Phylogeny tree of VN10-A-44 and its relationship with some Streptomyces species.
antibiotic production and help recycle tofu waste. Identification of VN10-A-44 by 16S-rDNA sequencing
showed that VN10-A-44 is identical to S. virginiae. This species is reported to produce VirginiamycinM1 (Suzuki et al., 1998) and it has a high activity in vitro against Gram-positive and negative bacteria, yeast and fungi (Rifaat and Kansoh, 2005). ACKNOWLEDGEMENTS This work was supported by the National Foundation for Science and Technology Development, Vietnam and con-ducted as a joint research project between the Institute of Microbiology and Biotechnology, Vietnam National University Hanoi, Vietnam (VNUH-IMBT) and the Biological Resource Center, National Institute of Technology and Evaluation (NBRC), Japan. REFERENCES Blackburn AS, Avery SV (2003). Genome-wide screening of
Saccharomyces cerevisiae to identify genes required for antibiotic
insusceptibility of eukaryotes. Antimicrob. Agents Chemother.
47:676-681. Dai LY, Liu XL, Xiao YH, Wang GL (2007). Recent advances in cloning
and characterization of disease resistance genes in rice. J. Int. Plant
Biol. 49:112-119. Fredrickson JK, Elliott LF (1985). Effects on winter wheat seedling
growth by toxin-producing rhizobacteria. Plant Soil. 83:399-409.
Gnanamanickam SS (2009). Biological control of bacterial blight of rice, In Biological control of rice diseases. Springer pp.67-78.
Hastuti RD, Lestari Y, Saraswati R, Suwanto A, Chaerani (2012). Capability of Streptomyces spp. In controlling bacterial leaf blight
diseases in rice plants. Am. J. Agric. Biol. Sci. 7(2):217-223 Hayakawa M, Nonomura H (1989). A new method for the intensive
isolation of actinomycetes from soil. Actinomycetologica 3:95-104. Hayakawa M, Otoguro M, Takeuchi T, Yamazaki T, Iimura Y (2000).
Application of a method incorporating differential centrifugation for
selective isolation of motile actinomycetes in soil and plant litter. Antonie Van Leeuwenhoek 78:171-185.
Hop DV, Sakiyama Y, Binh CTT, Otoguto M, Hang DT, Miyadoh S,
Luong DT, Ando K (2011). Taxonomic and ecological studies of actinomycetes from Vietnam: isolation and genus- level diversity. J. Antibiot. (Tokyo):1-8.
Ji GH, Wei LF, Wu YP, Bai XH (2008). Biological control of rice bacterial blight by Lysobacter antibioticus strain 13-1. Biol. Control.
45: 288-296.
Ndonde MJM, Semu E (2000). Preliminary characterization of some Streptomyces species from four Tanzanian soils and their
antimicrobial potential against selected plant and animal pathogenic
bacteria. W. J. Microb. Biotech. 16:595-599.

Park SB, Lee IA, Joo WS, Jeong GK, Lee HW (2011). Screening and
Identification of Antimicrobial Compounds from Streptomyces bottropensis Suppressing Rice Bacterial Blight. J. Microbiol.
Biotechnol. 21:1236-1242. Chithrashree A, Udayashankar C, Chandra NS, Reddy MS, Srinivas C
(2011). Plant growth-promoting rhizobacteria mediate induced
systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. Oryzae. Biol. Control 59:114-122.
Rifaat HM, Kansoh AL (2005). Streptomyces verginiae: Taxonomic,
Identification and biological activities. Arab. J. Biotech. 8:29-34. Rizk M, Abdel RT, Metwally H (2007). Screening of Antagonistic Activity
in Different Streptomyces Species Against Some Pathogenic
Microorganisms. J. Biol. Sci. 7:1418-1423. Sakiyama Y, Thao NKN, Giang NM, Miyadoh S, Hop DV, Ando K
(2009). Kineosporia babensis sp. nov., isolated from plant litter in
Vietnam. Int. J. Syst. Evol. Microbiol. 59:550-554.
Suzuki N, Lee CK, Nihira T, Yamada Y (1998). Purification and
characterization of Virginiamycin M1 Reductase from Streptomyces virginiae. Antimicrob. Agents Chemother. 42:2985-2988.
Vasudevan P (2002). Isolation and characterization of Bacillus spp.
from the rice rhizosphere and their role in biological control of bacterial blight of rice caused by Xanthomonas oryzae pv. oryzae.
Ph.D. dissertation, University of Madras. Velusamy P, Gnanamanickam SS (2003). Identification of 2,4-
diacetylphloglucinol (DAPG) production by plant-associated bacteria
and its role in suppression of rice bacterial blight in India. Curr. Sci. (Bangalore). 85:1270-1273.
Velusamy P, Immanuel JE, Gnanamanickam SS, Thomashow LS
(2006). Biological control of rice bacterial blight by plant-associated bacteria producing 2,4-diacetylphloroglucinol. Can. J. Microbiol. 52:56-65.

Original
*Corresponding author. Tel and Fax: +84-437547407, E-mail : phuonghoa(a)gmail.com
Biological Control of Xanthomonas Oryzae pv. OryzaeCausing Rice Bacterial Blight Disease by
Streptomyces toxytricini VN08-A-12, Isolated fromSoil and Leaf-litter Samples in Vietnam
DUONG VAN HOP1, PHAN THI PHUONG HOA1,4*, NGUYEN DUC QUANG2, PHAN HUU TON3, TRINH HOANG HA4, NGUYEN VAN HUNG3, NGUYEN THI VAN1,
TONG VAN HAI3, NGUYEN THI KIM QUY1, NGUYEN THI ANH DAO1, AND VU THI THOM4
1 Institute of Microbiology and Biotechnology (IMBT), Vietnam National University, Hanoi (VNUH)E2 Building, 144 Xuan Thuy road, Cau Giay District, Hanoi Province, Vietnam2 University of Engineering and Technology, Vietnam National University, Hanoi
3 Faculty of Biotechnology, Hanoi University of Agriculture4 School of Medicine and Pharmacy, Vietnam National University, Hanoi
Received 13 March, 2013/Accepted 13 December, 2013
A total of 2690 actinomycete strains were screened as potential biological control agents in controlling rice bacterial blight (BB) in Vietnam. From these microorganisms, seventeen actino-mycete strains were found to be capable of inhibiting all 10 major Xoo races isolated from Xoo-infected rice leaves. One strain, namely VN08-A-12, contained effective characteristics in selectively inhibiting all 10 Xoo races in vitro, but did not inhibit most of the other tested micro-organisms. Therefore, VN08-A-12 was subsequently selected for rice field trials for two seasons on two rice cultivars SS1 and KD18. Results showed VN08-A-12 was not only able to reduce Xoo lesion lengths in the two rice cultivars (lesion length reduction of up to 38.3 %), but it also significantly reduced Xoo-related yield loss in infected rice cultivars from the field (yield loss reduction of up to 43.2 %). Interestingly, the culture of this strain also increased the rice yield in healthy rice cultivars (from 2.66 % to 16.98 % for SS1 and from 3.11 % to 5.94 % for KD18 cultivar). The strain VN08-A-12 was shown to be identical to Streptomyces toxytricini. To our knowledge, this is the first study reporting S. toxytricini as a beneficial biological agent for the control of BB in rice.
Key words : Xoo / Streptomyces toxytricini / Vietnam.
Biocontrol Science, 2014, Vol. 19, No. 3, 103-111
varietal resistance, researchers have recently attempted to control BB in rice by using biological agents and their bioactive compounds (Chithrashree et al., 2011; Ndonde and Semu, 2000). The use of microorganisms for controlling Xoo has shown significant promise in being an ecologically-conscious and cost-effective solu-tion in rice cultivation (Gnanamanickam, 2009; Park et al., 2011). Actinomycetes are gram-positive bacteria that are well-known as a source of natural products (Anzai et al., 2008). It has been reported that of the approxi-mately 10,000 antibiotics discovered from microorgan-isms, two-thirds were isolated from actinomycetes
INTRODUCTION
Bacter ia l b l igh t (BB) o f r ice i s caused by Xanthomonas oryzae pv. oryzae (Xoo), and is one of the most destructive diseases of this essential crop. The disease can lead to yield losses of up to 50 or 60 % in tropical regions throughout Asia (Adhikari et al., 1994; Dinh et al., 2008; Gnanamanickam, 2009) and it has occurred in many rice cultivating areas of Vietnam (Dinh et al., 2008). In addition to using chemicals and

P. T. PHUONG HOA ET AL.104
oil-separation method (Hop et al., 2011). Among these strains, a total of 2690 actinomycete strains were used to screen for antibiotic-producers by assay for antago-nism to M. luteus NBRC 13867 and E. coli NBRC 14237. The method was previously mentioned in other studies (Ichikawa et al., 1971; Hoa et al., 2012).
Xoo races In the present study, a collection of 10 pathogen races of Xoo were tested. These were named R1, R2, R3, R4, R5, R6, R7, R8, R9 and R10 (Ton et al., 2003; Hoa et al., 2012). Of the 10 races, eight were found from many provinces of northern Vietnam, with two (R9 and R10) from Japan. The races were isolated from rice BB-infected leaves that were collected from the main rice-cultivating areas of northern Vietnam. Among the 10 Xoo races, R2 and R3 were the most widely distributed.
Media In addition to the media [Wakimoto medium (WM), Yeast extract–soluble starch medium(YS), Antibiotic producing medium (APM), Yeast-extract malt medium (YM), Muller Hinton agar (MHA); Ashby medium (AM) and Nutrient Agar (NA)] described in our previous paper (Hoa et al., 2012), we used five other media [Medium 2M, Medium 301, Medium A-16, Medium no. 8, Medium A-3M] in the present study.
Detection of actinomycetes capable of inhibiting Xoo races in vitro The antibacterial activity of actinomycetes against 10 pathogenic Xoo races was tested by the agar-piece method (6 mm) on Wakimoto medium as previously described (Hoa et al., 2012). In brief, the actinomy-cete isolates were inoculated on YS medium at room temperature (25 to 30℃) for 7 to 14 days. The pieces of agar (6 mm) that contained actively-growing actino-mycete isolates were cut and placed onto the test plates culturing races of Xoo. The plates were incu-bated at 28 ± 1℃ for two to four days and the inhibi-tory effect of the actinomycete isolates on Xoo growth was evaluated by measuring inhibition zones (cm). To prepare the test plates, races of Xoo were cultured in Wakimoto broth to a concentration of ca. 108 CFU/ml.
(Okami and Hotta, 1988). Actinomycetes, mainly the genus Streptomyces, have produced a wide variety of bioactive compounds, including antibiotics (Anzai et al., 2008). In recent studies, Streptomyces activity against Xoo strains has been reported (Ndonde and Semu, 2000; Yan et al., 2000; Park et al., 2011) and some bioactive compounds have been identified (Park et al., 2011), suggesting that actinomycetes are potential biological agents for the control of Xoo. In the present study, from a collection of 2690 actino-mycete strains, which had been isolated from 2005 to 2010 and preserved at the Vietnam Type Culture Collection (Institute of Microbiology and Biotechnology, Vietnam National University Hanoi), we screened for actinomycete strains that would be able to inhibit all 10 major Xoo races occurring in northern Vietnam and Japan (Ton et al., 2003; Hoa et al., 2012). An actino-mycete strain named VN08-A-12 was selected as a possible biological control agent of BB. This strain was selected for its potential to inhibit Xoo races, reduce Xoo lesion lengths in rice leaves and reduce Xoo-related yield losses in infected rice cultivars from field trials. The strain was identified to the species level.
MATERIALS AND METHODS
Experimental design A collection of 2690 actinomycete strains was used to test for inhibitory actions against Escherichia coli or Micrococcus luteus. 167 strains that had high inhibition activity against E. coli or M. luteus were selected. We then screened for the strains that inhibited all 10 Xoo races among those 167 strains. 17 strains that inhibited all 10 Xoo races were selected. Then, the antagonistic tests against indicator strains [M. luteus NBRC 13867, E. coli NBRC 14237, Bacillus subtilis NBRC 3134 and Saccharomyces cerevisiae NBRC 10217] and useful microogarnisms [Azotobacter sp. (VTCC-B-106) and Pseudomonas putida (VTCC-B-657)] were done for the 17 selected strains. The strain named VN08-A12 was selected as a potential strain for rice field trials over two cropping seasons; then we identified this strain to the species level.
Actinomycete strains During 2005 to 2010 at the Vietnam Type Culture Collection (Institute of Microbiology and Biotechnology, Vietnam National University Hanoi), over 3000 actino-mycete strains were isolated from soil and leaf-litter samples (Hop et al., 2011; Hoa et al., 2012) by using four methods in conjunction with humic acid-vitamin agar as an isolation medium: the rehydration-centrif-ugation (RC) method, sodium dodecyl sulfate-yeast extract dilution method, dry-heating method and the
TABLE 1. Collection of actinomycete strains from 2005 to 2010Name of strains
Year of isolation
Obtained and tested strains References
VTCCVN07VN08VN10Total
2005200720082010
600100050090
2690
Hop et al., 2011
Hoa et al., 2012

BIOLOGICAL CONTROL OF XANTHOMONAS ORYZAE 105
Actinomycetes were cultured in MAPM broth with shaking at 150 cycles/minute for five days. From this, 250 ml of the cultured solution was used for spraying over an area of 1m2 in each experimental square.Treatments Each of the two rice cultivars (Oryza sativa L. SS1 and Oryza sativa L. KD18) was planted in 45 experi-mental blocks within an experimental rice field at Hanoi University of Agriculture. The biocontrol trials at the rice field were conducted during two growing seasons in 2011, which consisted of planting in October and harvest in May and another planting in the end of May and harvest in November. Three squares were used for each treatment as follows.Treatment 1: (Positive control) rice cultivars were not infected by the Xoo races and were not treated with VN08-A-12. This was done to measure the non-infected/non-treated baseline yield of each paddy for each rice cultivar.Treatment 2: Rice cultivars were not infected by the Xoo races, but they were treated with VN08-A-12. This experiment was conducted to know the effect of VN08-A-12 on the rice yield of each rice cultivar. Treatment 3: Infected rice cultivars were treated as follows: CT1 denotes spraying VN08-A-12 after 1-day infection by Xoo races; CT2 denotes spraying VN08-A-12 after 3-day infection by Xoo races; CT3 denotes spraying VN08-A-12 after 7-day infection by Xoo races; and CT4 denotes spraying VN08-A-12 3 times: after 1, 3 and 7-day infection by Xoo races.Treatment 4: Rice cultivars were infected by Xoo races but were not treated with the VN08-A-12 broth culture. This was done to measure the effect of Xoo races on the host rice cultivars.Measurement of the yield of a paddy To measure and estimate paddy yield for each treat-ment, the rice grain dry weight from three individual rice plants per rice cluster was measured (Ton et al, 2003).
Identification of VN08-A-12 Identification of VN08-A-12 was based on morpho-logical, chemical and molecular methods as mentioned by Sakiyama et al. (2009).
Statistical analysis Data from laboratory and rice field experiments were analyzed and subjected to analysis of Variance (ANOVA) and correlation (SPSS Sofware). The signifi-cance of each treatment through two rice seasons and for each rice cultivar were determined by ANOVA (Tukey's posthoc test) (P values ≤ 0.05).
150 ml of Wakimoto agar was autoclaved and cooled down to 45 to 50℃ and then mixed with 10 ml of the Xoo cultures. This mixture was poured into Petri dishes that were used as the test plates (Hoa et al., 2012).
Test for antibiosis to microorganisms The actinomycete isolates inhibiting all 10 Xoo races were tested for their antibiosis to four indicator microog-arnisms, M. luteus NBRC 13867, E. coli NBRC 14237, B. subtilis NBRC 3134 and S. cerevisiae NBRC 10217 (indicator microorganisms were provided by NITE, Japan), and to two beneficial bacteria, Azotobacter sp. (VTCC-B-106), which is well-known in nitrogen fixation, and P. putida VTCC-B-657 which degrades phosphors into accessible forms for plants. Antibiosis tests were performed by using the agar-piece method as mentioned previously ( Ichikawa et al., 1971; Fredrickson and Elliott, 1985; Blackburn and Avery, 2003).
Selection of medium for antibiotic production of the potential actinomycete strain Seven media [Antibiotic producing medium (APM), Modified-APM (MAPM), Medium 2M, Medium 301, Medium A-16, Medium no. 8, Medium A-3M] were used for selection of a suitable antibiotic inducing medium of actinomycetes. The method was described previously (Hoa et al., 2012).
Biocontrol of BB in rice by actinomycetes in field trialsArtificial infection of Xoo races R2 and R3 on rice leaves The rice cultivars Oryza sativa L. SS1 and Oryza sativa L. KD18 were chosen for field trials for two crop-ping seasons in 2011 because they are widely culti-vated across Vietnam and have susceptibility to the Xoo races. The BB pathogens R2 and R3, being the most prevalent in northern Vietnam, were inoculated in Wakimoto medium flasks and incubated at 28℃ for 48 hours. The Xoo bacterial cells were then diluted in steril-ized water to reach a concentration of 108 CFU/ml to infect the two rice cultivars. This method was described by Ji et al., (2008). The two rice cultivars were planted in the field. Artificial infection was conducted at the rice booting stage because this stage is the most susceptible to BB and infection during this stage subsequently impacts rice yields. Infection was performed by dipping the scis-sors into a Xoo bacterial suspension and using them to clip the tip of the rice leaves (Kauffman et al., 1973). After 14 days, the length of lesions on 15 leaves per rice cluster was measured at the field. Three rice clusters per treatment formula were examined.Xoo treatment by actinomycetes

P. T. PHUONG HOA ET AL.106
R2 expressed the highest levels of inhibition relative to the others (Figure1). Only 25.7, 27.1 and 36.5% of the actinomycete strains inhibited races R9, R6 and R2, respectively. Inhibition of the other races of actinomy-cete strains accounted for at least 49.7%. Most of the tested actinomycete isolates (73%) were able to inhibit more than 2 Xoo races. A total of 17 strains (10.2%) were found to be able to inhibit all 10 Xoo races (Table 2). The inhibition zones of the 17 actinomycete strains inhibiting 10 Xoo races varied from 0.2 to 2.6 cm, depending on the strains and Xoo races (data not shown).
Test of antibiosis to microorganisms The purpose of this study was to select actinomycete strains capable of inhibiting all 10 Xoo races while not being harmful to other microorganisms. The 17 actino-mycete strains capable of inhibiting all 10 Xoo races chosen above were tested for their inhibition activity against four indicator microorganisms (M. luteus, E. coli, B. subtilis and S. cerevisiae) and two useful micro-organisms (Azotobacter sp. and P. putida).
RESULTS
Screening for actinomycete isolates capable of inhibiting Xoo races in vitro From 2690 actinomycete strains, a total of 167 strains showed high activity against M. luteus or E. coli and were subsequently chosen for further antagonistic tests against the 10 Xoo races. The inhibition pattern against each Xoo race showed that races R9, R6 and
TABLE 2. Ability of 167 actinomycete strains to inhibit tested Xoo racesNumber of Xoo strainswhich actinomycete
strains can inhibit0 1 2 3 4 5 6 7 8 9 10
Percentage of actinomycete
Strains17.9% 8.4% 7.8% 5.9% 10.2% 2.4% 8.4% 10.8% 11.9% 5.9% 10.2%
FIG. 1. proportion of the actinomycete strains that inhibits each Xoo race
TABLE 3. Inhibition activity of the 17 actinomycete strains against tested microorganisms
No. Name Inhibition zones (cm)M. luteus E. coli S. cerevisiae B. subtilis Azotobacter sp. P. putida
1234567891011121314151617
VN06-A-353VN06-A-379VN06-A-1098VN06-A-1457VN06-A-1477VN08-A-12VN08-A-306VN08-A-352VN08-A-395VTCC-A-99VTCC-A-289VTCC-A-363VTCC-A-367VTCC-A-378VTCC-A-456VTCC-A-465VN10-A-44
-0.8 ± 0.11.8 ± 0.21.1 ± 0.10.6 ± 0.31.6 ± 0.11.2 ± 0.12.3 ± 0.11.2 ± 0.3+
1.0 ± 0.1+
1.2 ± 0.2++
2.3 ± 0.1-
-2.0 ± 0.21.5 ± 0.42.4 ± 0.21.1 ± 0.1---
0.8 ± 0.2-
0.5 ± 0.1-
0.5 ± 0.2----
1.4 ± 0.1-+-+-----+--
1.0 ± 0.21.2 ± 0.1--
-+
1.0 ± 0.2+
1.0 ± 0.1--++-+
1.8 ± 0.11.0 ± 0.21.0 ± 0.1-
2.0 ± 0.31.4 ± 0.1
n.dn.dn.dn.dn.d-n.dn.dn.dn.dn.dn.dn.dn.dn.dn.d-
n.dn.dn.dn.dn.d-n.dn.dn.dn.dn.dn.dn.dn.dn.dn.d
1.0 ±0.1Note: - indicates that the actinomycete strain did not inhibit the indicator microorganism; + indicates that the strain has antagonistic activity, but the inhibition zone was too small to measure. N.d. denotes not done. Each value is an average of a 3 time-repeated assay.

BIOLOGICAL CONTROL OF XANTHOMONAS ORYZAE 107
A12 after a 1-day infection had lesion lengths reduced by 18% in both seasons, and 29.8 % and 24.1 % (in the spring), 15.2 % and 29.4 % (in the summer) after the 3-timetreatments by VN08-A-12; R3- infected KD18 cultivar when treated 3 times with VN08-A12 had Xoo
We found that strains VN06-A-353, VN08-A-12, VN08-A-306, VN08-A-352, VTCC-A-99 and VN10-A-44 only inhibited one of the four indicator microorganisms whereas the others inhibited more (Table 3). It was observed that VN08-A-12 and VN10-A-44 grew faster than the other strains. After three days of cultivation, they showed high inhibition activity against Xoo races while the other strains required from five to seven days of cultivation to express this activity (data not shown). Thus, only two strains VN08-A-12 and VN10-A-44 were checked for their inhibition potential against the two beneficial microorganisms such as Azotobacter sp. (VTCC-B-106) and P. putida VTCC-B-657. The strain VN10-A-44 inhibited P. putida; whereas VN08-A-12 did not inhibit either of the beneficial bacteria tested. Therefore, VN08-A-12 was selected for further analysis and for the rice field trials.
Selection of the best medium for fermentation of VN08-A-12 Seven media were used to find the best medium for antibiotic production of VN08-A-12. From this, MAPM was found to be the best medium for VN08-A-12 to produce bioactive compounds. The inhibition zones measured when cultured in MAPM were significantly higher than those cultured in the other media (data not shown).
Biological control of Xoo races by VN08-A-12 in the rice field trialsXoo lesion length of rice cultivars SS1 and KD18 Inhibition efficacy of VN08-A-12 on rice cultivars was evaluated by Xoo-induced lesion length and rice yield following two rice growing seasons. The Xoo lesion lengths on rice leaves are shown for two rice cultivars SS1 (Figure 2A) and KD18 (Figure 2B). Bacterial blight disease occurred differently in the two rice culti-vars, with SS1 being more severely infected by Xoo races than KD 18, as shown in the Xoo lesion lengths that were from 15.2 to 25.5 cm for SS1 and from 7.2 to 12.9 cm for KD18. Treatment with VN08-A-12 signifi-cantly reduced the Xoo lesion length in rice leaves in both rice seasons. For example, in the R2-infected SS1 cultivar, the treatment after a 1-day infection reduced the Xoo lesion length by 25.3 % and 33.4 % reduc-tions in the spring and summer seasons, respectively; 3-time treatment by VN08-A-12 reached 32.6 % and 38.3 % reductions in the spring and summer seasons, respectively. Lesion lengths in the R3-infected SS1 cultivar were reduced by 27.5 % after a 1-day infec-tion in both rice seasons and by 15.2 % and 29.4 % after 3-time treatments by VN08-A-12 in the spring and summer seasons, respectively. On the other hand, for the R2-infected KD18 cultivar, the treatment by VN08-
FIG. 2. Effect of VN08-A12 on Xoo lesion length on leavesSS1 (A) and Khang dan (B) in spring and summer rice seasons. The leaves were artificially infected by X oo R2 (R2) or X oo R3 (R3) in summer and spring seasons without VN08-A12 treatment (CT0), with a single treatment of VN08-A12 after 1-day infection (CT1), 3-day infection (CT2) and 7-day infection (CT4) or with three-time treatments of VN08-A12 after 1, 3 and 7- day infections. I refers to R2 in Summer; II refers to R2 in Spring; III refers to R3 in Summer; IV refers to R3 in Spring. ANOVA was tested following by Tukey’s posthoc test. *p<0.05, **p<0.005 indicate significant differ-ences between CT1, CT2, CT3, CT4 to the respective control (CT0); # p<0.05 indicates significant difference between CT4 and other groups.

P. T. PHUONG HOA ET AL.108
In summer season, the same trend of reduced rice yields was observed (data not shown). R2 and R3 Xoo caused greater reductions in rice yield of the SS1 cultivar than those in spring season; however, VN08-A-12 also reduced the yield loss of the SS1 cultivar. We found that R2 and R3 races caused more than a 50 % reduction in rice yield of the SS1 cultivar (58.2 % for R2 and 55.1 % for R3). The yield of the paddies of the R2-infected SS1 cultivar increased 20.7 % - 25.5 % (one-time treatment) and 36.6 % (three-time treatment); that of the R3-infected SS1 cultivar increased 20.2 %-25.7 % (one-time treatment) and 38.5 % (three-time treatment) in comparison with the Xoo-infected SS1 cultivar without VN08-A-12 treatment. It was observed that a three-time treatment with VN08-A-12 increased the rice yield by 2.66 % of the healthy SS1 cultivar in comparison with the untreated healthy rice.KD18 cultivar Table 5 shows results on the rice yield of KD18 cultivar. Xoo races R2 and R3 significantly reduced the paddy yield of KD18 by 33.31 % for R2 and 44.05 % for R3. Results showed that VN08-A-12 improved the paddy yield of the rice cultivar. In the spring season, treatment with VN08-A-12 increased the rice yield 9.4 - 29.4 % in the R2-infected KD18 cultivar. On the other hand, the paddy of R3-infected KD18 cultivar treated with VN08-A-12 increased 20.75 - 38.99 % in
lesion lengths in leaves reduced by 27.2 % and 21.2 % in the spring and summer seasons, respectively. The efficacy of VN08-A12 to suppress Xoo races was less when treatment occurred late. Results showed that the treatment after a seven day infection reduced the lesion length from 6.1 % to 12.6 %. This suggests that late treatment by VN08-A-12 resulted in a lower level of inhi-bition of Xoo races.
Paddy yield of rice cultivars: SS1 and KD18SS1 cultivar Results from the SS1 cultivar paddy yield following the spring growing season are shown in Table 4. When infected by Xoo races, the SS1 cultivar paddy yield was reduced by 53.5 % (infected by R2) and 48.4 % (infected by R3). When treated with VN08-A-12, the yield of the paddies of the R2-infected SS1 cultivar increased 24.3 % - 26.8 % (one-time treatment) and 43.2 % (three-time treatment). The yield of the paddies of the R3-infected SS1 cultivars increased 26.8 % - 30.5 % (one-time treatment) and 35.7 % (one-time treatment) in comparison with the Xoo-infected SS1 cultivar without VN08-A-12 treatment. Notably, the VN08-A12 treatment increased the yield of the cultivar paddies from 13.16 % (one-time treatment) to 16.98 % (three-time treatment). Overall, the productivity of SS1 cultivar paddies both healthy and infected with Xoo races was improved when treated with VN08-A-12.
TABLE 4. Effect of VN08-A-12 on the productivity of Oryza sativa L. SS1 in the spring cropping season, 2011Treatment
by VN08-A-12Infected by R2 Infected by R3 Without infection
G/INV. ORY(%)RRY(%) G/INV. ORY(%)RRY(%) G/INV. ORY(%) IRY(%)Positive control Negative controlAfter a 1-day infectionAfter a 3-day infectionAfter a 7-day infectionAt three times treated
55.05 ± 0.93 ⒜25.60 ± 3.14 ⒝40.40 ± 1.22 ⒞40.00 ± 2.00 ⒞39.00 ± 1.00 ⒞49.40 ± 1.00 ⒟
10046.5073.3872.6670.8489.73
0.0053.5026.6227.3429.1610.27
55.05 ± 0.93 ⒜28.60 ± 2.95 ⒝45.20 ± 2.15 (ef)43.60 ± 0.35 (ef)43.20 ± 1.71 (e)48.10 ± 1.15 (df)
10051.6082.1079.2078.4787.37
0.0048.4017.9020.8021.5312.63
55.05 ± 0.93 ⒜n.d.
62.30 ± 0.20 ⒢62.30 ± 0.10 ⒢62.30 ± 0.30 ⒢64.40 ± 0.40 ⒣
100
113.16113.16113.16116.98
13.1613.1613.1616.98
Notes: G/INVD: gram per rice individual; ORY: obtained rice yields; RRY: Reduced rice yields; IRY: Increased rice yields; n.d., not doneStatistical analysis was done by ANOVA following by Tukey’s test for each column and each row; groups sharing at least one common letter were not significantly different (p>0.05); groups sharing no common letter were significantly different (p<0.05)
TABLE 5. Effect of VN08-A-12 on the productivity of Oryza sativa L. KD 18 in the spring cropping season, 2011 Treatment
by VN08-A-12Infected by R2 Infected by R3 Without infection
G/INV. ORY(%)RRY(%) G/INV. ORY(%)RRY(%) G/INV. ORY(%) IRY(%)Positive control Negative controlAfter a 1-day infectionAfter a 3-day infectionAfter a 7-day infectionAt three times treated
63.62 ± 0.75 ⒜42.60 ± 1.51 ⒝61.20 ± 0.26 (af)51.30 ± 0.30 ⒟48.40 ± 1.35 ⒠60.40 ± 0.40 ⒡
10066.9696.1980.6376.0794.93
0.0033.313.81
19.3723.935.06
63.62 ± 0.75 ⒜35.60 ± 0.72 ⒢58.80 ± 1.05 ⒣52.20 ± 1.15 ⒤44.80 ± 1.47 ⒦60.40 ± 0.69 (fh)
10055.9592.4282.0476.7094.93
0.0044.057.58
17.9623.305.06
63.62 ± 0.75⒜n.d.
65.60 ± 0.31 ⒧65.60 ± 0.10 ⒧65.60 ± 0.20 ⒧67.40 ± 0.20 ⒨
100
103.11103.11103.11105.94
3.113.113.115.94
Notes: G/INVD: gram per rice individual; ORY: obtained rice yields; RRY: Reduced rice yields; IRY: Increased rice yields; n.d., not doneStatistical analysis was done by ANOVA following by Tukey’s test for each column and each row; groups sharing at least one common letter were not significantly different (p>0.05); groups sharing no common letter were significantly different (p<0.05)

BIOLOGICAL CONTROL OF XANTHOMONAS ORYZAE 109
and other Asian countries. Our finding that the Streptomyces toxytricini VN08-A-12 was able to strongly inhibit all 10 Xoo races while not being harmful to other tested microorganisms is in agreement with a previous study (Wu and Chen, 1995). They found that Streptomyces toxytricini, isolated from soil in Taiwan was capable of producing antibiotics, but appeared to be non-toxic to mice and tomato seedlings. In this study, the Streptomyces toxytricini VN08-A-12 was isolated from dead leaves collected in Vietnam by using a dry heating method in 2008 (Hop et al., 2011). The leaf source of VN08-A-12 isola-tion suggests that the Streptomyces toxytricini VN08-A-12 may be easily adapted to the rice field both on rice leaves and in the soil. To our knowledge, this is the first report of Streptomyces toxytricini being a poten-tial biological agent for controlling BB in rice whereas Lysobacter antibiocus isolated from the rhizosphere of rice in China (Ji, et al., 2008) and Streptomyces bottro-pensis isolated from soil in Korea (Park et al., 2011) were recently reported. The rice field trials for the two rice cultivars and two rice cropping seasons were evaluated based on lesion lengths and rice yields. In general, the Streptomyces toxytricini VN08-A-12 was able to significantly suppress the Xoo races in natural rice fields. These results were in agreement with a previous study by Hastuti et al. (2012), who showed that Streptomyces spp. may be a potential candidate for biologically controlling BB in rice. On the other hand, Streptomyces toxytricini VN08-A-12 was shown to significantly reduce the yield loss of Xoo infected rice cultivars in the present study. For the SS1 cultivar, which is often heavily affected by BB, the lesion lengths varied from 23.3 cm to 25.5 cm in the two seasons. VN08-A-12 significantly reduced Xoo lesion lengths (Figure 2). In the spring crop-ping season, late treatment by VN08-A-12 in the rice cultivar infected by Xoo R2 (after a seven day infection) reduced Xoo lesion lengths by 6.1 %, whereas early and multiple treatments brought reductions of greater than 32 %. Xoo lesion lengths were shorter when SS1 was infected by R3 compared to R2, and this was simi-larly recorded in the summer cropping season (Figure 1). For the KD18 cultivar, Xoo lesion lengths were less reduced (29.8 % reduction for R2-infected and 27.2 % R3-infected crops in the spring season; 24.1 % and 21.2 % for crops infected by R2 and R3 in the summer season, respectively). The effects of Xoo reduction in the two tested rice cultivars in the spring season were more pronounced than in the summer season. This could have been due to the fact that the summertime climate favors BB and that summer heavy rains could have affected the treatment with VN08-A-12 in the rice
comparison to the Xoo-infected KD18 cultivar without VN08-A-12 treatment. For the healthy KD18 cultivar, a one-time treatment with VN08-A-12 improved the rice paddy 3.11 % while a three-time treatment increased the paddy by 5.94 %. The data on the KD18 paddy in the summer season was not obtained, due to rodent damage to the crop.
Identification of VN08-A-12 Based on the results of morphological and physi-ological characterization of VN08-A-12, we found that the VN08-A-12 strain had typical characteristics of the Streptomyces genus (data not shown). Analysis of the whole-cell hydrolysate of strain VN08-A-12 showed the presence of a chemotype I cell wall character-ized by LLA2pm. No diagnostic sugars were found. The main components of fatty acids were anteios-(anteios-C15:0), iso-(iso-C16:0) and normal (n-C16:0) acids. The menaquinones were MK-9 (H6) 51.9 % and MK-9 (H8) 48.1 %. The GC content was 75 %. The cultural and physiological characterization and the DNA homology suggested strain VN08-A-12 is identical to Streptomyces toxytricini.
DISCUSSION
Bacterial blight disease in rice is one of the major diseases in the rice culture of northern Vietnam as well as other rice-growing regions of the world. Many Xoo races have been identified from many rice cultivation areas of Vietnam (Ton et al., 2003). In this study, actin-omycete strains that were able to inhibit all major Xoo isolates were screened from a collection of 2960 actino-mycete strains. Results showed that actinomycete strains were able to inhibit Xoo races (Figure 1, Table 2) and in particular, 17 strains could inhibit all 10 major Xoo races. Their inhibition assays on indicator microor-ganisms M. luteus NBRC 13867, E. coli NBRC 14237, B. subtilis NBRC 3134, S. cerevisiae NBRC 10217 and useful bacteria Azotobacter sp. (VTCC-B-106) and P. puttida VTCC-B-657 led to our selection of only one strain, namely VN08-A-12, for further rice field study for its potential benefits to the environment. In this study, for cultivation of VN08-A-12 we chose the suitable medium of MAPM in which soybean meal of the original medium (APM) was replaced with tofu waste. Our previous research found that tofu waste could be used instead of soybean meal for the fermen-tation of the Streptomyces virginiae strain VN10-A-44 (Hoa et al., 2012). This medium was good for large-volume fermentation of Streptomycete for large-scale applications in agriculture. Our study thus has the potential to recycle tofu waste which is one of the major sources of waste in the production of tofu in Vietnam

P. T. PHUONG HOA ET AL.110
rice fields should be performed in the future.
ACKNOWLEDGEMENTS
This research was funded by the Vietnam National Foundation for Science and Technology Development (NAFOSTED) under grant number 106.03-2010.34. We would like to thank the leaders of a joint research project between the Institute of Microbiology and Biotechnology, Vietnam National University Hanoi, Vietnam (VNUH-IMBT) and the Biological Resource Center, National Institute of Technology and Evaluation (NBRC), Japan for providing the microorganisms in this study. We would like to thank Dr. Todd W. Miller, Division of Fish and Wildlife, Marianas Protectorate Saipan (USA) for his critical review of this paper. We would like to thank the Korean Collection for Type Cultures (KCTC), Biological Resource Center, Korea Research Institute of Bioscience and Biotechnology, Korea (KRIBB) for organizing an international training course on microbial taxonomy, which provided tech-nical and material support to identify VN08-A-12 to the species level. We also thank Dr. Hiroshi Kinoshita, the International Center for Biotechnology, Osaka University, for his technical support at VNUH-IMBT.
REFERENCES Adhikari, T.B., Mew, T.W., and Teng, P.S. (1994) Progress of
bacterial blight on rice cultivars carrying different Xa genes for resistance in the field. Plant Dis., 78, 73-77.
Anzai, K., Nakashima, T., Kuwahara, T., Suzuki, R., Ohfuku, Y., Takeshita, S., and Ando, K. (2008) Actinomycete bacteria isolated from the sediments at coastal and offshore area of Nagasaki prefecture, Japan: diversity and biological activity. J. Biosci. Bioeng. 2, 215–217.
Blackburn, A.S., and Avery, S.V. (2003) Genome-wide screening of Saccharomyces cerevisiae to identify genes required for antibiotic insusceptibility of eukaryotes. Antimicrob. Agents and Chemother., 47, 676 – 681.
Chithrashree, A., Udayashankar, C., Chandra, N.S., Reddy, M.S., and Srinivas, C. (2011) Plant growth-promoting rhizobacteria mediate induced systemic resistance in rice against bacterial leaf blight caused by Xanthomonas oryzae pv. oryzae. Biol. Control., 59, 114–122.
Dinh, H.D., Oanh, N.K., Toan, N.D., Du, P.V., and Loan, L.C. (2008) Pathotype profile of Xanthomonas oryzea pv oryzea isolated from the rice ecosystem in Cuulong river delta. Omorice, 16, 34-40.
E, L.V., Noda, T., and Du, P.V. (1999) Resistance assess-ment of rice cultivars to Xanthomonas oryzea pv. oryzea and pathogenicity testing of bacterial leaf blight isolates in Vietnam. Omorice, 7, 120-130.
Fredrickson, J.K., and Elliott, L.F. (1985) Effects on winter wheat seedling growth by toxin-producing rhizobacteria. Plant and Soil, 83, 399-409.
Gnanamanickam, S. S. (2009) Biological control of bacterial blight of rice. In Biological control of rice diseases. Springer, pp: 67-78. 108 p.
Hastuti, R.D., Lestari, Y., Saraswati, R., Suwanto, A., and
field. In Vietnam, BB is more severe and causes highest losses in the summer copping season (Ton et al, 2003; E et al., 1999). Figure 2 shows the same trend that earlier and multiple treatments with VN08-A12 leads to effective control of BB in the SS1 and KD18 cultivars. Results from measured rice yields of the tested culti-vars showed that the Streptomyces toxytricini VN08-A-12 reduced BB caused by races R2 and R3 and this corresponded to higher rice yields. This positive effect depended on the rice cultivar, Xoo races, the rice copping season and the timing of the treatment with VN08-A-12. For example, the paddy of cultivar SS1 infected by race R2 had a rice yield ranging from 70.8 to 73.8 %, with a one-time treatment with VN08-A-12 in the spring season (at 1, 3 and 7 day infection); however these were higher than those infected by Race R2 that were not treated with VN08-A-12, which showed 46.5 % paddy yields (reduced 53.5 %). In the case of cultivar SS1 infected by race R3, its yields from paddies were from 78.4 to 82.1 % with a one-time treatment and 51.6 % (48.4 % reduction) without treatment by VN08-A-12. In particular, a three-time treatment with VN08-A12 (with 87.37 % paddy yield) significantly improved the rice yield of cultivar SS1. This was also observed in the summer season (data not shown). Interestingly, it was found that the rice yield of the healthy cultivar SS1 treated with VN08-A12 was significantly higher than that of the untreated one and these results were observed for all the treatments in both the spring and summer seasons. This may be a result of hormesis. It is common in animal/plant physi-ology that the bacteria improvies the physiology of the plant, turns on genes, or produces hormones (Jocelyn, 2003). For cultivar KD18, treatment by VN08-A-12 signifi-cantly reduced the effect of Xoo infection (Table 5). KD18 yields increased from 9.4 to 38.9 % following the treatment by VN08-A-12. In particular, treatments by VN08-A-12 in healthy KD18 showed significantly increased rice yields from 13.6 to 16.9 %. These results suggest that this particular strain could be used as a rice fertilizer. Previous studies showed that plant-promoting rhizobacteria induce systemic resistance in rice against BB (Chithrashree et al., 2011). The improvement of cultivar paddies by treatment with VN08-A-12 could be possibly due to the bioactive compounds that inhibit Xoo and to the nutrient contents of the VN08-A-12 fermented culture. A previous study (Hastuti et al., 2012) also found that Streptomyces isolates increased plant yield in comparison to the chemically-sprayed and non-treated plots. Our results suggested that VN08-A-12 has potential application for the biological control of BB and for the improvement of rice yields. Further examination on large-scale trials of

BIOLOGICAL CONTROL OF XANTHOMONAS ORYZAE 111
soils and their antimicrobial potential against selected plant and animal pathogenic bacteria. World J. Microb. Biotechnol., 16, 595-599.
Okami Y., and Hotta K (1988) Search and discovery of new antibiotics, In: Goodfellow M, Williams ST, Mordarski M (ed). Actinomycetes in Biotechnology. Academic Press, Inc., San Diego, pp. 33-67.
Park, S.B., Lee, I.A., Joo-Won, S., Jeong-Gu, K., and Lee, H.W. (2011) Screening and identification of antimicrobial compounds from Streptomyces bottropensis suppressing rice bacterial bl ight. J. Microbiol. Biotechnol., 21, 1236–1242.
Sakiyama, Y., Thao, N.K.N., Giang, N.M., Miyadoh, S., Hop, D.V., and Ando, K. (2009) Kineosporia babensis sp. nov., isolated from plant litter in Vietnam. Internation J. System. Evolut. Microbiol., 59, 550-554.
Ton, P. H., Thuy, B. T., and Hai, T. V. (2003) Pathogenicity of the Bacterial Leaf Blight Strains from Northern Vietnam. The 2nd National Conference on Plant Pathology and Molecular Biology (In Vietnamese).
Wu, R.Y., and Chen, M.H. (1995) Identification of the Streptomyces strain KS3-5. Bot. Bull. Acad. Sin., 36, 201-205.
Yan, M. V., Da, Q. T., Shi, M. T., and Ding, Z. (2000) The antagonism of 26 strains of Streptomycetes sp. against several vegetables pathogens. Hebei Agr. Univer., 23, 65-68.
Chaerani. (2012) Capability of Streptomyces spp. in controlling bacterial leaf blight disease in rice plants. Am. J. Agr. Biol Sci., 7(2), 217-223.
Hoa, P.T.P., Quang, N.D., Sakiyama, Y., Hop, D.V., Hang, D.T., Van, N.T., Quy, N.T.K., and Dao, N.T.A. (2012) Screening for actinomycetes isolated from soil able to inhibit Xanthomonas oryzae pv. oryzae causing rice bacte-rial blight disease in Vietnam. Afr. J Biotechnol., 11(80), 14586-14594.
Hop, D.V., Sakiyama, Y., Binh, C.T.T., Otoguto, M., Hang, D.T., Miyadoh, S., Luong, D.T., and Ando, K. (2011) Taxonomic and ecological studies of actinomycetes from Vietnam: isolation and genus- level diversity. J. Antibiot (Tokyo), 1-8.
Ichikawa, T., Date, M., Ishikura, T., and Ozaki, A. (1971) Improvement of kasugamycin-producing strain by the agar piece method and the prototroph method. Folia Microbial., 16, 218-224.
Ji, G.H., Wei, L.F., Wu, Y.P., and Bai, X.H. (2008) Biological control of rice bacterial blight by Lysobacter antibioticus strain 13-1. Biol. Control., 45, 288-296.
Jocelyn, K. (2003) Hormesis: Sipping from a poisoned chalice. Science, 302, 376-379.
Kauffman, H.E., Reddy, A.P.K., Hsieh, S.P.Y., and Merca, S.D. (1973) An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzea. Plant Dis. Report., 57, 537-541.
Ndonde, M.J.M., and Semu, E. (2000) Preliminary character-ization of some Streptomyces species from four Tanzanian





























