Embed Size (px)
Citation preview

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƢỜNG ĐẠI HỌC KHOA HỌC TỰ NHIÊN
-----------------------
Phạm Thu Hằng
NGHIÊN CỨU TẠO DÒNG LÚA CHUYỂN GEN MÃ HÓA
NHÂN TỐ PHIÊN MÃ NHÓM NAC LIÊN QUAN ĐẾN TÍNH
CHỊU HẠN
Chuyên ngành: Hóa sinh học
Mã số: 62420116
(DỰ THẢO) TÓM TẮT LUẬN ÁN TIẾN SĨ SINH HỌC
Hà Nội - 2017

Công trình đƣợc hoàn thành tại:
Trƣờng Đại học Khoa học Tự nhiên, Đại học Quốc gia Hà Nội.
Ngƣời hƣớng dẫn khoa học:
PGS.TS. Phạm Xuân Hội
Phản biện 1: .............................................
Phản biện 2: .............................................
Phản biện 3: .............................................
Luận án sẽ đƣợc bảo vệ tại Hội đồng chấm luận án cấp Đại học
Quốc gia họp tại:
Trƣờng Đại học Khoa học Tự nhiên – Đại học Quốc gia Hà Nội
vào hồi......giờ......., ngày.......tháng.......năm 2017.
Có thể tìm hiểu luận án tại:
- Thƣ viện Quốc gia Việt Nam
- Trung tâm thông tin - Thƣ viện, Đại học Quốc gia Hà Nội

1
MỞ ĐẦU
1. Tính cấp thiết của đề tài
Dƣới tác động của biến đổi khí hậu toàn cầu, tình trạng hạn hán
xảy ra thƣờng xuyên hơn, với mức độ ngày càng trầm trọng, là
nguyên nhân chính làm giảm sản lƣợng cây trồng. Trong khi đó, dân
số toàn cầu tăng nhanh, ƣớc tính năm 1950 sẽ tăng lên 9 tỷ ngƣời,
ảnh hƣởng nghiêm trọng đến an ninh lƣơng thực, tạo ra một áp lực
lớn cho việc đảm bảo lƣơng thực. Vì vậy, an ninh lƣơng thực là vấn
đề then chốt của nền khoa học nông nghiệp thế giới nói chung và
Việt Nam nói riêng.
Lúa (Oryza sativa L.) là một trong ba loại cây lƣơng thực quan
trọng nhất của con ngƣời. Trên thế giới, cây lúa đƣợc xếp vào vị trí
thứ 2 sau cây lúa mì về diện tích và sản lƣợng. Ở Việt Nam, một đất
nƣớc từ lâu đời đã gắn liền với nền văn minh lúa nƣớc, vai trò cây
lúa đặc biệt quan trọng. Cây lúa đóng vai trò chính trong sản xuất
lƣơng thực, đóng góp sản lƣợng cao nhất (60%) và đƣợc canh tác với
diện tích lớn nhất (xấp xỉ 3,8 triệu ha). Chính vì vậy, việc tạo ra các
giống lúa có năng suất cao, đồng thời chống chịu đƣợc với điều kiện
bất lợi ngoại cảnh đặc biệt là hạn có ý nghĩa quan trọng đối với việc
duy trì và tăng năng suất lúa gạo, góp phần giữ ổn định an ninh lƣơng
thực quốc gia.
Các nghiên cứu tạo giống truyền thống chủ yếu dựa trên các
phƣơng pháp lai tạo, chọn giống truyền thống nên hiệu quả đạt đƣợc
không thực sự cao. Một số ứng dụng mới trong chọn giống cây trồng
nhƣ dùng chỉ thị phân tử, gây đột biến... tuy đã đạt đƣợc một số kết
quả nhất định nhƣng vẫn có những hạn chế. Phƣơng pháp chuyển gen
ra đời đƣợc ví nhƣ “chìa khóa đa năng” để mở những nút thắt vốn

2
gây rất nhiều khó khăn cho các nhà chọn tạo giống truyền thống
nhằm tạo ra một giống cây trồng “hoàn hảo” hơn khi chúng đƣợc kết
hợp với nhau. Với hƣớng nghiên cứu này, các gen quy định những
tính trạng quan tâm đƣợc chủ động chuyển vào lúa, từ đó tạo ra các
giống lúa mang các đặc tính nhƣ mong muốn của con ngƣời.
Tính trạng chịu hạn là tính trạng đa gen mà sự biểu hiện của mỗi
gen liên quan chặt chẽ với quá trình phiên mã. Vì vậy, nhóm gen mã
hóa nhân tố phiên mã tham gia điều khiển quá trình phiên mã đang là
trọng tâm trong các nghiên cứu chọn tạo giống nhằm tăng cƣờng tính
chống chịu của cây trồng với điều kiện hạn. Nhóm gen mã hóa nhân
tố phiên mã mặc dù không tham gia trực tiếp vào quá trình đáp ứng
với điều kiện hạn nhƣng sự biểu hiện của chúng lại có vai trò điều
hòa biểu hiện của rất nhiều gen chức năng khác, dẫn tới làm tăng
cƣờng khả năng chịu hạn của thực vật. Nhiều nghiên cứu chuyển
gen mã hóa nhân tố phiên mã vào lúa đã đƣợc chứng minh tăng
cƣờng khả năng chịu hạn so với cây không chuyển gen.
Nhóm gen mã hóa nhân tố phiên mã NAC (tên gọi đƣợc bắt
nguồn từ tên gọi của ba protein đầu tiên đƣợc phát hiện có chứa vùng
NAC là NAM - no apica meristem, ATAF - trancription activation
factor và CUC - cup shaped cotyledon) là họ nhân tố phiên mã lớn
nhất đặc trƣng của thực vật. Nhân tố phiên mã NAC tham gia vào rất
nhiều quá trình sinh lý, sinh hóa khác nhau trong tế bào, bao gồm các
quá trình phát triển, già hóa, tạo thành tế bào thứ cấp và đáp ứng
chống chịu stress môi trƣờng nhƣ hạn, mặn và lạnh của tế bào. Một
số gen đáp ứng stress thuộc nhóm NAC ở lúa nhƣ SNAC1/OsNAC9,
SNAC2/OsNAC6, OsNAC5 và OsNAC10 đã đƣợc chứng minh có liên
quan tới quá trình đáp ứng hạn ở cây lúa. Cây lúa chuyển gen

3
SNAC1/OsNAC9, OsNAC5 và OsNAC10 có khả năng kháng hạn và
năng suất cao hơn so với cây không chuyển gen.
Xuất phát từ thực tế trên, chúng tôi tiến hành đề tài luận án tiến sĩ
“Nghiên cứu tạo dòng lúa chuyển gen mã hóa nhân tố phiên mã
nhóm NAC liên quan đến tính chịu hạn” với mục tiêu thu tạo ra
các dòng lúa chuyển gen chống chịu hạn, đồng thời không gây ảnh
hƣởng đến khả năng sinh trƣởng, phát triển. Các kết quả thu đƣợc sẽ
là nguồn vật liệu, số liệu quan trọng trong việc tạo ra các giống cây
trồng chuyển gen có khả năng chống chịu tốt với điều kiện hạn.
Mục tiêu nghiên cứu
Phân lập, thiết kế vector chuyển gen mã hóa nhân tố phiên mã
nhóm NAC liên quan tính chịu hạn từ giống lúa Việt Nam.
Tạo dòng lúa chuyển gen mã hóa nhân tố phiên mã thuộc nhóm
NAC có khả năng chống chịu hạn.
Đối tƣợng v nội dung nghiên cứu
Đối tượng nghiên cứu của luận án là các gen mã hóa nhân tố phiên
mã nhóm NAC liên quan tới tính chịu hạn của giống lúa Việt Nam.
ác n i ung nghiên cứu ch nh
Nội dung 1: Phân lập các gen mã hóa nhân tố phiên mã nhóm
NAC gồm OsNAC1, OsNAC5, OsNAC10 liên quan đến tính chịu hạn
từ giống lúa Việt Nam;
Nội dung 2: Thiết kế các cấu trúc biểu hiện gen OsNAC1 ở
thực vật dƣới sự điều khiển của promoter liên tục (Ubiquitine, 35S)/
cảm ứng (Lip9, Rd29A).
Nội dung 3: Chuyển gen mã hóa nhân tố phiên mã OsNAC1
vào lúa.
Nội dung 4: Đánh giá sinh trƣởng, phát triển và khả năng
chịu hạn của các dòng lúa chuyển gen.

4
Địa điểm nghiên cứu
Luận án đƣợc thực hiện tại 2 địa điểm chính: ộ môn ệnh học
Phân tử, Viện Di truyền Nông nghiệp (Viện Khoa học Nông nghiệp
Việt Nam); Phòng Thí nghiệm tƣơng tác cây trồng - vi sinh vật thuộc
Trung tâm nghiên cứu vì sự phát triển tại Montpellier, Pháp.
Đóng góp mới của luận án
Luận án đã phân lập đƣợc 3 gen mã hóa nhân tố phiên mã nhóm
NAC gồm OsNAC1, OsNAC5 và OsNAC10 từ giống lúa bản địa Việt
Nam. Luận án đã tạo đƣợc các hệ vector biểu hiện gen trong tế bào
thực vật mang gen OsNAC1 đƣợc điều khiển bởi promoter hoạt động
liên tục (Ubiquitin và 35S) và promoter hoạt động cảm ứng (Lip9 và
Rd29A). Luận án đã tạo đƣợc một số dòng lúa chuyển gen biểu hiện
gen đích, sinh trƣởng bình thƣờng trong điều kiện không xử lý hạn và
có khả năng chịu hạn tốt hơn cây đối chứng không chuyển gen.
Ứng dụng thực tiễn của luận án
Kết quả nghiên cứu của luận án đã tạo ra đƣợc các dòng lúa T2
mang một bản sao của gen OsNAC1 ở dạng đồng hợp đặt dƣới sự
điều khiển của promoter hoạt động cảm ứng điều kiện stress Lip9 và
promoter hoạt động liên tục Ubiquitin. Cây chuyển gen có kiểu hình
tƣơng đồng với cây đối chứng không chuyển gen (về khả năng sinh
trƣởng, phát triển và năng suất) trong điều kiện bình thƣờng không
có stress hạn, trong khi tỷ lệ sống sót cao hơn trong điều kiện xử lý
stress hạn ở giai đoạn cây non và các chỉ tiêu về hình thái, sinh lý,
sinh hóa thể hiện khả năng chịu hạn cao hơn ở giai đoạn sinh sản
trong điều kiện xử lý stress hạn. Đây là kết quả quan trọng, chứng tỏ
tiềm năng ứng dụng của gen OsNAC1 nói riêng và các gen mã hóa
nhân tố phiên mã thuộc nhóm NAC liên quan tới đáp ứng stress nói
chung trong công tác tạo giống cây chuyển gen chịu hạn. Trên cơ sở

5
kết quả của luận án, các vector biểu hiện gen OsNAC1 do luận án tạo
ra đang đƣợc nghiên cứu chuyển vào các giống cây trồng quan trọng
nhƣ ngô, bông, đậu tƣơng… để tạo ra các giống cây chuyển gen kháng
hạn nhằm đáp ứng nhu cầu cấp thiết của sản xuất trong điều kiện thay
đổi khí hậu toàn cầu ở Việt Nam.
2. Bố cục của luận án
Luận án gồm 144 trang, bao gồm: Phần mở đầu (04 trang); Tổng
quan tài liệu (42 trang); Nguyên liệu và phƣơng pháp nghiên cứu (15
trang); Kết quả nghiên cứu và thảo luận (47 trang); Kết luận (02
trang); Kiến nghị (01 trang); Các công trình khoa học của tác giả liên
quan đến luận án (01 trang); Tài liệu tham khảo (22 trang) với 215 tài
liệu gồm 2 thứ tiếng: tiếng Việt (11 tài liệu) và tiếng Anh (204 tài
liệu); Phụ lục (7 trang). Luận án có 9 bảng, 27 hình.

6
Chƣơng 1: TỔNG QUAN
1.1. GIỚI THIỆU CHUNG VỀ CÂY LÚA
1.1.1. Đặc điểm nông sinh học của cây lúa Oryza sativa
Lúa Oryza sativa thuộc bộ Hòa thảo (Graminales), họ Hòa thảo
(Graminaceae), chi Oryza. Chi Oryza phân bố rộng trên thế giới và
có từ 19 – 23 loài, trong đó có loài Oryza sativa L (2n = 24) đƣợc
trồng phổ biến ở khắp các nƣớc trên thế giới và tập trung chủ yếu ở
châu Á. Loài Oryza gluberrima S. đƣợc trồng một diện tích nhỏ ở
một số nƣớc thuộc châu Phi.
1.1.2. Ảnh hƣởng của hạn hán đối với ngành sản xuất lúa gạo
Việt Nam là một trong số các quốc gia bị tác động nặng nề nhất
bởi biến đổi khí hậu. Sự xuất hiện thƣờng xuyên của hạn hán tiếp tục
đe dọa nghiêm trọng đến nền sản xuất lúa gạo trong nƣớc. Hệ quả
của các tác động này là áp lực ngày càng tăng đối với giá lúa gạo
trên thị trƣờng thế giới do Việt Nam là một trong những nƣớc xuất
khẩu gạo quan trọng nhất trên thế giới.
1.1.3. Ảnh hƣởng của hạn tới đặc điểm chịu hạn cuả cây lúa
Hạn hán luôn là mối đe dọa nghiêm trọng đối với nền sản xuất
lúa gạo. Hạn ảnh hƣởng tiêu cực đến cây lúa ở tất cả các đặc điểm,
bao gồm hình thái, sinh lý, sinh hóa, phân tử.
1.2. VAI TRÒ CỦA NHÂN TỐ PHIÊN MÃ LIÊN QUAN TỚI ĐÁP
ỨNG STRESS HẠN Ở THỰC VẬT
Nhân tố phiên mã (Transcription factor - TF) là những protein
hoạt động kết hợp với một số yếu tố điều hòa khác, bao gồm các protein
cải biến/thay đổi cấu trúc sợi nhiễm sắc, để điều hòa tƣơng tác của RNA
polymerase với DNA.
NAC là nhân tố phiên mã lớn nhất đặc chƣng của thực vật. Tên gọi
NAC bắt nguồn từ tên của ba protein đầu tiên đƣợc phát hiện có chứa

7
vùng liên kết DNA tƣơng tự nhau (domain NAC), bao gồm NAM (no
apical meristem), ATAF (Arabidopsis transcription activation factor) và
CUC (cup-shaped cotyledon). NAC tham gia vào rất nhiều quá trình sinh
lý, hóa sinh khác nhau trong tế bào, bao gồm các quá trình phát triển, già
hóa, tạo thành tế bào thứ cấp, đáp ứng các yếu tố stress phi sinh học và
hữu sinh. NAC là thành phần rất quan trọng của con đƣờng truyền tín hiệu
trong đáp ứng chống chịu stress của thực vật cho thấy tiềm năng ứng dụng
của họ gen NAC trong nghiên cứu tạo giống cây trồng chống chịu stress.
1.3. AGROBACTERIUM TUMEFACIENS VÀ CƠ CHẾ CHUYỂN
GEN VÀO THỰC VẬT
Stress sinh học và phi sinh học là nguyên nhân chính làm giảm
năng suất trên nhiều đối tƣợng cây trồng quan trọng nhƣ lúa, ngô,
bông, lúa mì, cà chua... Và để đối phó với vấn đề này, công nghệ
chuyển gen thực vật đƣợc sử dụng và đƣợc coi là một công cụ hiệu
quả để chuyển các gen ngoại lai nhằm cải thiện nguồn gen cây trồng.
1.4. TÌNH HÌNH NGHIÊN CỨU TẠO GIỐNG LÚA CHUYỂN
GEN CHỐNG CHỊU HẠN
Một hƣớng giải quyết đƣợc chính phủ, các tổ chức xã hội và các
nhà khoa học đặc biệt quan tâm đó là nghiên cứu tạo ra các giống lúa
mới có năng suất cao, đồng thời có khả năng chống chịu tốt với điều
kiện hạn. Định hƣớng nghiên cứu phổ biến và rất đƣợc quan tâm hiện
nay đó tăng cƣờng biểu hiện các gen đáp ứng hoặc liên quan đến đáp
ứng chống chịu hạn trong cây lúa chuyển gen. Ở Việt Nam, định hƣớng
chuyển gen vào lúa đã bắt đầu hình thành vào những năm 1995 của thế
kỷ trƣớc. Tuy nhiên, chúng ta hầu nhƣ chƣa có một nghiên cứu hoàn
chỉnh nào về các gen liên quan đến đáp ứng chống chịu stress, đặc biệt
là nhóm gen mã hoá nhân tố phiên mã liên quan tới đáp ứng hạn ở lúa.

8
Chƣơng 2. VẬT LIỆU & PHƢƠNG PHÁP NGHIÊN CỨU
2.1. VẬT LIỆU
2.1.1. Đối tƣợng nghiên cứu
Giống lúa J02 đƣợc mua từ Công ty Cổ phần giống – vật tƣ nông
nghiệp công nghệ cao Việt Nam;
Các giống lúa nƣơng và IR64 do Trung tâm Tài nguyên môi
trƣờng – Viện Khoa học Nông nghiệp Việt Nam cung cấp.
2.1.2. Chủng vi sinh vật
Vi khuẩn Escherichia coli chủng DH5α đƣợc mua từ hãng
Thermo Scientifics (Mỹ); vi khuẩn A. tumefaciens chủng LBA4404
đƣợc mua từ Công ty Clontech Laboratories (Mỹ).
2.1.3. Vector và oligonucleotide
Vector pBI-Lip9, pBI-Ubi, pBI-35S và pCAM-Rd29A do nhóm
nghiên cứu của Phạm Xuân Hội (Bộ môn Bệnh học phân tử thực vật –
Viện Di truyền nông nghiệp) thiết kế và cung cấp; vector pGEM-T
mạch thẳng có đầu T đƣợc đóng gói kèm trong bộ kit pGEM®-T
Easy Vector Systems của hãng Promega.
Các cặp oligonucleotide sử dụng làm mồi cho PCR đƣợc thiết kế
dựa trên các trình tự đã đƣợc công bố trên Gen ank và đƣợc đặt mua
từ hãng Invitrogen (Mỹ) và Sigma (Mỹ).
2.2. PHƢƠNG PHÁP NGHIÊN CỨU
- Xử lý mẫu thực vật theo phƣơng pháp của Qin (2007), Dubouzet
(2003) và Tran (2004).
- Tách chiết, định lƣợng DNA/RNA.
- Nhân dòng gen OsNAC1/ OsNAC5/ OsNAC10 vào vector
pGEM-T.
- Thiết kế vector biểu hiện gen OsNAC1.

9
Hình 2.2: Sơ đồ thiết kế vector pCAM-Rd/OsNAC1
Hình 2.3: Sơ đồ thiết kế pBI-Lip9/ OsNAC1, pBI-35S/ OsNAC1 và
pBI-Ubi/ OsNAC1
- Tạo cây lúa chuyển gen biểu hiện OsNAC1.
- Sàng lọc kiểu gen, kiểu hình của các cây chuyển gen
- Đánh giá khả năng chịu hạn của cây chuyển gen ở điều kiện nhà
lƣới bằng các phƣơng pháp định lƣợng sinh lý, sinh hóa ở cây lúa:
hàm nƣớc tƣơng đối theo phƣơng pháp của Schonfeld (2002), hàm
lƣợng chlorophyll theo phƣơng pháp của Robert J. Porra (2006), hàm
lƣợng proline theo phƣơng pháp của Bates (1973), sự sản sinh ROS
theo phƣơng pháp Thorda- Christensen (1997).

10
Chƣơng 3. KẾT QUẢ & THẢO LUẬN
3.1. PHÂN LẬP GEN MÃ HÓA NHÂN TỐ PHIÊN MÃ NHÓM
NAC LIÊN QUAN ĐẾN TÍNH CHỊU HẠN Ở LÚA
3.1.1. Khảo sát khả năng chịu hạn của một số giống lúa Việt Nam
10 giống lúa Việt Nam và giống lúa đối chứng IR64 khảo sát khả
nănng chịu theo phƣơng pháp chuẩn của Viện Lúa quốc tế IRRI, 1996.
Kết quả thể hiện ở Bảng 3.1
Bảng 3.1: Kết quả đánh giá khả năng chịu hạn ở lúa
STT Tên giống Phƣơng pháp đánh
giá theo IRRI
1 Lúa Tẻ đỏ 1,00 ± 0,6
2 Pê đớ 2,33 ± 0,8
3 Khấu le 2,67 ± 0,5
4 Blue xá 2,67 ± 1,1
5 Lốc nghệ 3,33 ± 1,1
6 Lúa ngoi 3,67 ± 0,8
7 Lúa lốc đỏ 4,33 ± 0,8
8 Nếp khau Pí Pột 4,67 ± 0,5
9 Lúa tẻ nƣơng 5,67 ± 0,8
10 Chành trụi 5,67 ± 0,8
11 IR64 5,67 ± 1,1
Giống lúa Tẻ đỏ đƣợc lựa chọn làm nguồn vật liệu thí nghiệm
phân lập các gen mã hóa nhân tố phiên mã nhóm NAC liên quan tới
tính chịu hạn ở lúa.
3.1.2. Phân lập gen mã hóa nhân tố phiên mã nhóm NAC liên
quan tới tính chịu hạn
3.1.2.1. Tách chiết RNA tổng số từ mẫu lúa xử lý hạn
Kết quả điện di RNA trên gel agarose 1% cho thấy, các mẫu RNA

11
tinh sạch phục vụ cho thí nghiệm phân lập gen (Hình 3.1).
Hình 3.1: Tách chiết RNA tổng số từ mẫu lúa xử lý hạn
Ghi chú: giếng 1 – 4: RNA tổng số tách chiết từ mẫu lá lúa xử lý hạn
trong 1, 6, 12 và 24 giờ được điện di trên gel agarose 1%. Giếng M:
thang chuẩn DNA 1 kb.
3.1.2.2. Phân lập các gen mã hóa nhân tố phiên mã nhóm NAC
3.1.2.2.1. Nhân bản gen mã hóa nhân tố phiên mã nhóm NAC
Các đoạn gen đích đƣợc nhân bản bằng phản ứng PCR sử dụng
các cặp mồi đặc hiệu đã thiết kế, sau đó kiểm tra bằng điện di trên gel
agarose 1%.
Hình 3.2: Nhân bản gen OsNAC1, OsNAC5 và OsNAC10 bằng kỹ
thuật PCR từ cDNA lúa Tẻ đỏ xử lý hạn
Ghi chú: Sản phẩm PCR nhân gen đích từ cDNA của các mẫu lúa Tẻ đỏ
được điện di trên gel agarose 1%. (A) PCR nhân bản gen OsNAC1,
giếng 1: mẫu lúa xử lý hạn. giếng 2: đối chứng âm (mẫu lúa không xử lý
hạn); (B) PCR nhân bản gen OsNAC, giếng 2: khuôn là cDNA; giếng 1:
đối chứng âm. (C) PCR nhân bản gen OsNAC10, giếng 2: khuôn là
cDNA; giếng 1: đối chứng âm. Giếng M: thang chuẩn DNA 1kb.

12
Kết quả phân tích trình tự axit amin suy diễn của các gen OsNAC1,
OsNAC5 và OsNAC10 từ lúa Tẻ đỏ cho thấy, cả 3 gen phân lập đƣợc đều
mã hóa cho protein với các vùng đặc trƣng của họ protein NAC. OsNAC1,
OsNAC5 chứa đầy đủ cả 5 domin bảo thủ A, B, C, D và E, trong khi
OsNAC10 thiếu domai E (Hình 3.3). Ngoài ra, cả ba protein này đều chứa
một vùng tín hiệu định vị nhân PRDRKYP phổ biến cho các nhân tố
phiên mã, nằm ở domain C (Hình 3.3). Kết quả này hoàn toàn phù hợp
với các công bố trƣớc đây.
Hình 3.3: Phân tích trình tự axit amin của protein OsNAC1,
OsNAC5 và OsNAC10 của giống lúa Tẻ đỏ
Ghi chú: Vùng tín hiệu định vị nhân được kí hiệu bởi dấu ngoặc; các
vùng bảo thủ của protein NAC được kí hiệu A, B, C, D, E; TĐ: gen
phân lập từ lúa Tẻ đỏ, JA: gen phân lập từ lúa Japonica (Niponbare)
đã công bố trên ngân hàng gen mã số AY596808.1, AB028184.1 và
AK069257.1.

13
3.1.3. Phân tích biểu hiện của OsNAC1, OsNAC5 và OsNAC10
trong điều kiện hạn
Hình 3.4: Mức độ biểu hiện của OsNAC1, OsNAC5 và OsNAC10
trong điều kiện xử lý hạn
Ghi chú: Mức độ biểu hiện gen OsNAC1 (cột màu xanh), OsNAC5
(cột màu đỏ) và OsNAC10 (cột màu vàng) của lúa Tẻ đỏ đã xử lý hạn
(PEG 20%) trong thời gian 0 giờ, 1 giờ, 6 giờ, 12 giờ và 24 giờ được
xác định bằng RT - PCR. Mức độ biểu hiện gen tại thời điểm chưa xử
lý stress (0 h) có giá trị bằng 1. Actin được sử dụng làm gen nội
chuẩn. Giá trị thể hiện trên đồ thị là kết quả trung bình của 3 lần lặp
lại thí nghiệm
Trong nghiên cứu này, chúng tôi sử dụng phƣơng pháp RT-PCR
nghiên cứu mức độ biểu hiện của các gen OsNAC1, OsNAC5 và
OsNAC10 ở giống lúa Tẻ đỏ trong điều kiện xử lý stress hạn. Mức độ
biểu hiện khác nhau ở các gen khác nhau, trong đó OsNAC1 có mức
độ biểu hiện cao hơn rõ rệt và duy trì mức độ biểu hiện cao trong thời
gian dài hơn đáng kể so với hai gen còn lại khi xử lý stress hạn. Điều
này cho phép dự đoán OsNAC1 đóng góp vai trò nhiều hơn trong đáp
ứng chống chịu hạn của cây lúa Tẻ đỏ so với OsNAC5 và OsNAC10,
đặc biệt là đối với stress hạn kéo dài. Chính vì vậy, OsNAC1 đƣợc
lựa chọn làm đối tƣợng nghiên cứu chính cho các thí nghiệm tiếp
theo nhằm tạo ra dòng lúa chuyển gen có khả năng chống chịu hạn.

14
3.2. THIẾT KẾ VECTOR BIỂU HIỆN OsNAC1 TRONG TẾ BÀO
THỰC VẬT
3.2.1. Thiết kế hệ vector chuyển gen pCAMBIA1301
Hình 3.5: PCR v cắt enzyme giới hạn kiểm tra sự có mặt của
OsNAC1 trong plasmid tái tổ hợp pCAM-Rd/OsNAC1
Ghi chú: (A) Sản phẩm PCR plasmid pCAM-Rd/OsNAC1, giếng 1 và 3:
PCR với mồi OsNAC1-Fw/Rv; giếng 3 và 4: PCR với mồi RD-Fw/NAC1-
Rv; giếng 1 và 2: khuôn là pCAM-Rd/OsNAC1; giếng 3 và 4: đối chứng
âm (không có DNA khuôn). (B) Sản phẩm cắt enzyme giới hạn plasmid
pCAM-Rd/OsNAC1, giếng 1: sản phẩm cắt giới hạn bằng enzyme
BamHI; giếng 3: sản phẩm cắt giới hạn bằng enzyme HindIII/EcoRI;
giếng 2: vector nguyên bản. Giếng M: thang chuẩn DNA 1kb.
3.2.2. Thiết kế hệ vector chuyển gen pBI101
Hình 3.6: PCR v cắt giới hạn kiểm tra sự có mặt của OsNAC1
trong plasmid tái tổ hợp pBI-Ubi/OsNAC1, pBI-Lip9/OsNAC1và
pBI-Lip9/OsNAC1
Ghi chú: (A) Sản phẩm PCR plasmid tái tổ hợp; giếng 1, 3: khuôn là
pBI-Ubi/OsNAC1; giếng 5, 7: khuôn là pBI-35S/OsNAC1; giếng 9,
11 : khuôn là pBI-Lip9/OsNAC1; giếng 1-2, 5-6 và 9-10: PCR với
cặp mồi OsNAC1-Fw/ OsNAC1-Rv; giếng 3, 4: PCR với cặp mồi
Ubi-Fw/ OsNAC1-Rv; giếng 7, 8: PCR với cặp mồi 35S-Fw/
OsNAC1-Rv;giếng 11,12: PCR với cặp mồi Lip9-Fw/ OsNAC1-Rv;
giếng 1, 3, 6, 8, 10, 12: đối chứng âm với khuôn là nước. (B) Sản
phẩm cắt giới hạn plasmid tái tổ hợp bằng BamHI; giếng 1 và 2:
pBI-Ubi/OsNAC1; giếng 3 và 4: pBI-35S/OsNAC1; giếng 5 và 6:
pBI-Lip9/OsNAC1; giếng 2, 4 và 6: sản phẩm cắt giới hạn; giếng 1,
3 và 5: plasmid nguyên bản. Giếng M: thang chuẩn DNA 1 kb.

15
3.3. NGHIÊN CỨU CHUYỂN GEN OsNAC1 TRONG CÂY LÚA
3.3.1. Biến nạp vector biểu hiện vào A. tumefaciens LBA4404
Hình 3.7: PCR trực tiếp khuẩn lạc mang pCAM-Rd/OsNAC1,
pBI-35S/OsNAC1, pBI-Ubi/OsNAC1và pBI-Lip9/OsNAC1
Ghi chú: Sản phẩm PCR khuẩn lạc A. tumefaciens với cặp mồi OsNAC1-
Fw/ OsNAC1-Rv được điện di trên gel agarose 1%. (A) Thể biến nạp pBI-
35S/OsNAC1, pBI-Ubi/OsNAC1và pBI-Lip9/OsNAC1, giếng 5, 6 và 13:
đối chứng âm (khuôn là nước); giếng 1 – 4: khuôn là thể biến nạp pBI-
35S/OsNAC1; giếng 7 – 9: khuôn là thể biến nạp pBI-Ubi/OsNAC1;
giếng 10 – 12: khuôn là thể biến nạp pBI-Ubi/OsNAC1. (B) Thể biến nạp
pCAM-Rd/OsNAC1; giếng 2 – 4: khuôn là thể biến nạp pCAM-
Rd/OsNAC1; giếng 4: đối chứng âm. Giếng M: thang chuẩn DNA 1 kb.
3.3.2. Nghiên cứu chuyển gen OsNAC1 vào lúa
Cấu trúc biểu hiện gen OsNAC1 đƣợc chuyển vào mô sẹo của lúa
J02 (Oryza sativa L. Japonica) thông qua vi khuẩn Agrobacterium
tumefaciens. Mô sẹo hình thành từ phôi lúa đƣợc lần lƣợt chuyển các
cấu trúc biểu hiện gen OsNAC1 (Ubi:OsNAC1, Lip9:OsNAC1,
35S:OsNAC1, Rd29A:OsNAC1) và các cấu trúc vector nhị phân không
mang gen đích (pCAM IA1301, p I101). Sau đó, mô sẹo đƣợc chọn
lọc và tái sinh trên môi trƣờng nuôi cấy in vitro.
Bảng 3.2: Kết quả chuyển gen vào lúa thông qua vi khuẩn A.
tumefaciens
Cấu trúc gen
chuyển Callus Chọn lọc
Tái sinh
chồi
Tái sinh
rễ
Lip9: OsNAC1 ~100 38 32 29
Ubi: OsNAC1 ~100 50 37 36
35S: OsNAC1 ~100 51 44 41
16
Rd29A:
OsNAC1 ~100 38 14 14
pBI101 ~100 27 17 17
pCAMBIA1301 ~100 41 27 25
3.3.3. Chọn lọc cây lúa mang gen chuyển OsNAC1 có một bản
copy đời T0
Các cây tái sinh trƣớc khi đƣa ra bầu đất sinh trƣởng trong nhà
lƣới đƣợc kiểm tra khả năng mang gen cần chuyển bằng các phản
ứng PCR với ba cặp mồi là Actin-Fw/ Actin-Rv, Hyg-Fw/ Hyg-Rv
và OsNAC1-Fw/ Nos-Rv (đối với dòng chuyển cấu trúc gen
OsNAC1) và xác định số lƣợng bản copy cấu trúc gen chuyển bằng
phƣơng pháp phân tích PCR định lƣợng thời gian thực (qRT-PCR)
với cặp mồi Hyg-RT-Fw/ Hyg-RT-Rv.
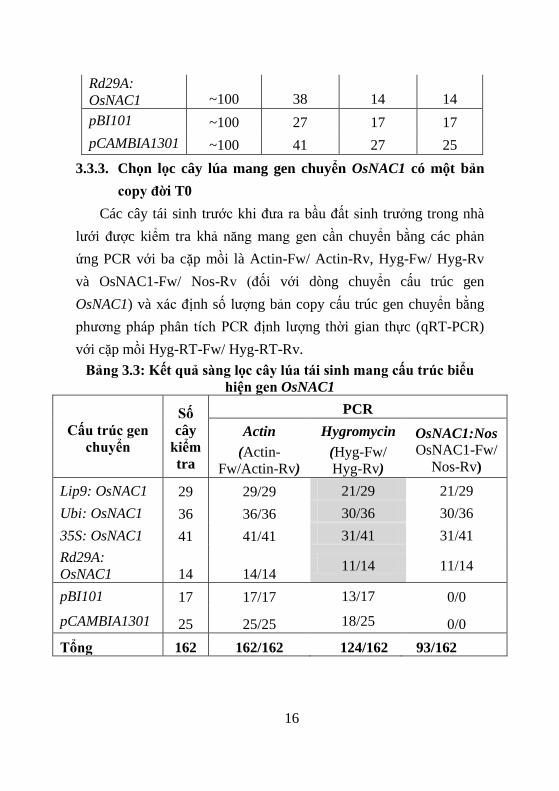
Bảng 3.3: Kết quả s ng lọc cây lúa tái sinh mang cấu trúc biểu
hiện gen OsNAC1
Cấu trúc gen
chuyển
Số
cây
kiểm
tra
PCR
Actin
(Actin-
Fw/Actin-Rv)
Hygromycin
(Hyg-Fw/
Hyg-Rv)
OsNAC1:Nos
OsNAC1-Fw/
Nos-Rv)
Lip9: OsNAC1 29 29/29 21/29 21/29
Ubi: OsNAC1 36 36/36 30/36 30/36
35S: OsNAC1 41 41/41 31/41 31/41
Rd29A:
OsNAC1 14 14/14 11/14 11/14
pBI101 17 17/17 13/17 0/0
pCAMBIA1301 25 25/25 18/25 0/0
Tổng 162 162/162 124/162 93/162
17
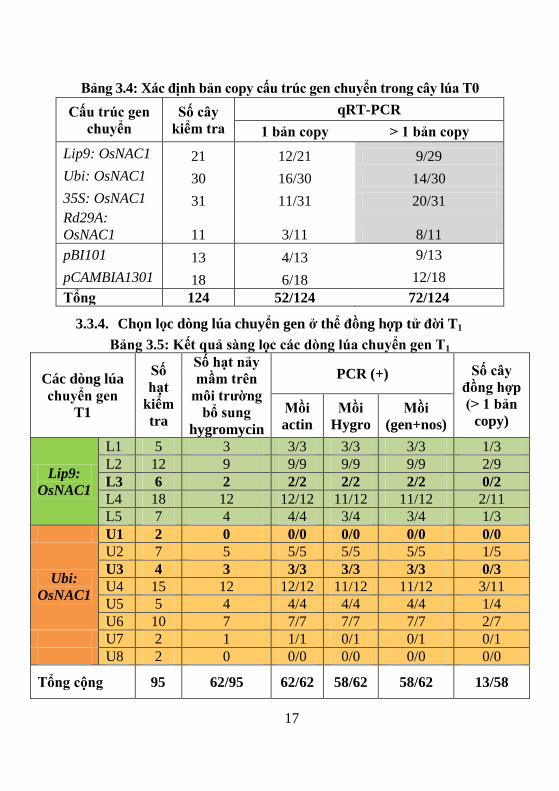
Bảng 3.4: Xác định bản copy cấu trúc gen chuyển trong cây lúa T0
Cấu trúc gen
chuyển
Số cây
kiểm tra
qRT-PCR
1 bản copy > 1 bản copy
Lip9: OsNAC1 21 12/21 9/29
Ubi: OsNAC1 30 16/30 14/30
35S: OsNAC1 31 11/31 20/31
Rd29A:
OsNAC1 11 3/11 8/11
pBI101 13 4/13 9/13
pCAMBIA1301 18 6/18 12/18
Tổng 124 52/124 72/124
3.3.4. Chọn lọc dòng lúa chuyển gen ở thể đồng hợp tử đời T1
Bảng 3.5: Kết quả s ng lọc các dòng lúa chuyển gen T1
Các dòng lúa
chuyển gen
T1
Số
hạt
kiểm
tra
Số hạt nảy
mầm trên
môi trƣờng
bổ sung
hygromycin
PCR (+) Số cây
đồng hợp
(> 1 bản
copy) Mồi
actin
Mồi
Hygro
Mồi
(gen+nos)
Lip9:
OsNAC1
L1 5 3 3/3 3/3 3/3 1/3
L2 12 9 9/9 9/9 9/9 2/9
L3 6 2 2/2 2/2 2/2 0/2
L4 18 12 12/12 11/12 11/12 2/11
L5 7 4 4/4 3/4 3/4 1/3
U1 2 0 0/0 0/0 0/0 0/0
Ubi:
OsNAC1
U2 7 5 5/5 5/5 5/5 1/5
U3 4 3 3/3 3/3 3/3 0/3
U4 15 12 12/12 11/12 11/12 3/11
U5 5 4 4/4 4/4 4/4 1/4
U6 10 7 7/7 7/7 7/7 2/7
U7 2 1 1/1 0/1 0/1 0/1
U8 2 0 0/0 0/0 0/0 0/0
Tổng cộng 95 62/95 62/62 58/62 58/62 13/58
18
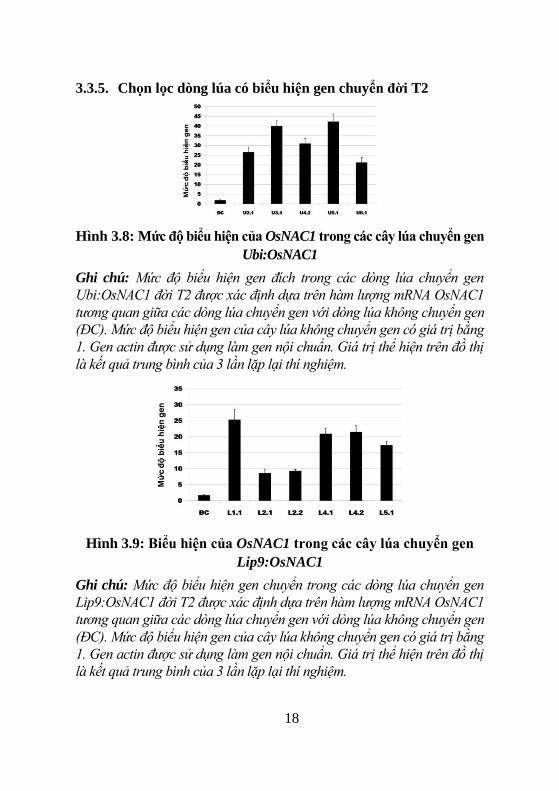
3.3.5. Chọn lọc dòng lúa có biểu hiện gen chuyển đời T2
Hình 3.8: Mức độ biểu hiện của OsNAC1 trong các cây lúa chuyển gen
Ubi:OsNAC1
Ghi chú: Mức độ biểu hiện gen đích trong các dòng lúa chuyển gen
Ubi:OsNAC1 đời T2 được xác định dựa trên hàm lượng mRNA OsNAC1
tương quan giữa các dòng lúa chuyển gen với dòng lúa không chuyển gen
(ĐC). Mức độ biểu hiện gen của cây lúa không chuyển gen có giá trị bằng
1. Gen actin được sử dụng làm gen nội chuẩn. Giá trị thể hiện trên đồ thị
là kết quả trung bình của 3 lần lặp lại thí nghiệm.
Hình 3.9: Biểu hiện của OsNAC1 trong các cây lúa chuyển gen
Lip9:OsNAC1
Ghi chú: Mức độ biểu hiện gen chuyển trong các dòng lúa chuyển gen
Lip9:OsNAC1 đời T2 được xác định dựa trên hàm lượng mRNA OsNAC1
tương quan giữa các dòng lúa chuyển gen với dòng lúa không chuyển gen
(ĐC). Mức độ biểu hiện gen của cây lúa không chuyển gen có giá trị bằng
1. Gen actin được sử dụng làm gen nội chuẩn. Giá trị thể hiện trên đồ thị
là kết quả trung bình của 3 lần lặp lại thí nghiệm.

19
3.4. ĐÁNH SINH TRƢỞNG, PHÁT TRIỂN VÀ KHẢ NĂNG
KHÁNG HẠN DÒNG LÚA CHUYỂN GEN
3.3.1. Đánh giá sinh trưởng, phát triển của các dòng lúa chuyển gen
Hình 3.10: Đánh giá sinh trƣởng của các dòng lúa chuyển gen
5 dòng lúa chuyển gen có khả năng sinh trƣởng và phát triển
tƣơng đƣơng với cây đối chứng tiếp tục đƣợc sử dụng trong thí
nghiệm đánh giá khả năng kháng hạn.
3.3.2. Đánh giá khả năng chịu hạn của các dòng lúa chuyển gen T2
3.3.2.1.1. Đánh giá khả năng chịu hạn theo thang điểm chuẩn IRRI
Bảng 3.6: Đánh giá khả năng chịu hạn theo thang chuẩn IRRI ở
các dòng lúa chuyển gen v dòng lúa đối chứng
STT Dòng lúa Thang điểm Thứ tự chịu hạn
1 Đối chứng (ĐC) 7,8 ± 0,96 6
2 L1.1 4,2 ± 0,96 1
3 L4.1 5,8 ± 1,44 3
4 L5.1 6,2 ± 1,28 4
5 U4.2 6,6 ± 0,64 5
6 U6.1 5,4 ± 1,28 2
Nhƣ vậy, xét theo thang điểm chuẩn IRRI, thứ tự chịu hạn giảm
dần của các dòng lúa nhƣ sau: L1.1, L4.1, L5.1, U4.2, U6.1, ĐC.
20
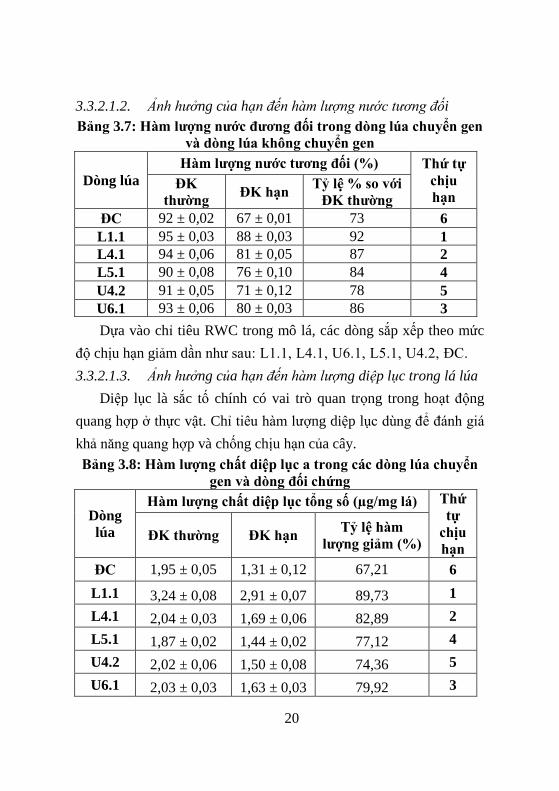
3.3.2.1.2. Ảnh hưởng của hạn đến hàm lượng nước tương đối
Bảng 3.7: H m lƣợng nƣớc đƣơng đối trong dòng lúa chuyển gen
v dòng lúa không chuyển gen
Dòng lúa
H m lƣợng nƣớc tƣơng đối (%) Thứ tự
chịu
hạn ĐK
thƣờng ĐK hạn
Tỷ lệ % so với
ĐK thƣờng
ĐC 92 ± 0,02 67 ± 0,01 73 6
L1.1 95 ± 0,03 88 ± 0,03 92 1
L4.1 94 ± 0,06 81 ± 0,05 87 2
L5.1 90 ± 0,08 76 ± 0,10 84 4
U4.2 91 ± 0,05 71 ± 0,12 78 5
U6.1 93 ± 0,06 80 ± 0,03 86 3
Dựa vào chỉ tiêu RWC trong mô lá, các dòng sắp xếp theo mức
độ chịu hạn giảm dần nhƣ sau: L1.1, L4.1, U6.1, L5.1, U4.2, ĐC.
3.3.2.1.3. Ảnh hưởng của hạn đến hàm lượng diệp lục trong lá lúa
Diệp lục là sắc tố chính có vai trò quan trọng trong hoạt động
quang hợp ở thực vật. Chỉ tiêu hàm lƣợng diệp lục dùng để đánh giá
khả năng quang hợp và chống chịu hạn của cây.
Bảng 3.8: H m lƣợng chất diệp lục a trong các dòng lúa chuyển
gen v dòng đối chứng
Dòng
lúa
H m lƣợng chất diệp lục tổng số (µg/mg lá) Thứ
tự
chịu
hạn ĐK thƣờng ĐK hạn
Tỷ lệ h m
lƣợng giảm (%)
ĐC 1,95 ± 0,05 1,31 ± 0,12 67,21 6
L1.1 3,24 ± 0,08 2,91 ± 0,07 89,73 1
L4.1 2,04 ± 0,03 1,69 ± 0,06 82,89 2
L5.1 1,87 ± 0,02 1,44 ± 0,02 77,12 4
U4.2 2,02 ± 0,06 1,50 ± 0,08 74,36 5
U6.1 2,03 ± 0,03 1,63 ± 0,03 79,92 3
21
Từ kết quả nghiên cứu trên cho thấy, nếu chỉ dựa vào chỉ tiêu
hàm lƣợng diệp lục để đánh giá khả năng chịu hạn thì các dòng lúa
chuyển gen đều có khả năng chịu hạn tốt hơn dòng lúa đối chứng.
3.3.2.1.4. Hàm lượng proline
Bảng 3.9: H m lƣợng proline trong các dòng lúa
Dòng
lúa
H m lƣợng proline (%) Thứ tự
chịu
hạn ĐK thƣờng ĐK hạn H m lƣợng
proline tăng
ĐC 121,09 ± 19,02 215,81 ± 28,94 174,92 6
L1.1 142,93 ± 9,84 347,57 ± 41,04 243,18 1
L4.1 131,95 ± 6,10 268,29 ± 16,46 203,32 3
L5.1 129,52 ± 6,26 254,38 ± 7,32 196,40 5
U4.2 132,09 ± 19,02 265,81 ± 28,94 201,23 4
U6.1 137,24 ± 2,75 282,10 ± 6,87 205,55 2
Nhƣ vậy, chỉ xét chỉ tiêu sự tăng tích lũy hàm lƣợng proline
trong cây có thể khẳng định, các dòng lúa chuyển gen tích lũy hàm
lƣợng proline cao hơn dòng đối chứng thì có khả năng thích nghi và
có sức chống chịu hạn tốt hơn dòng đối chứng.
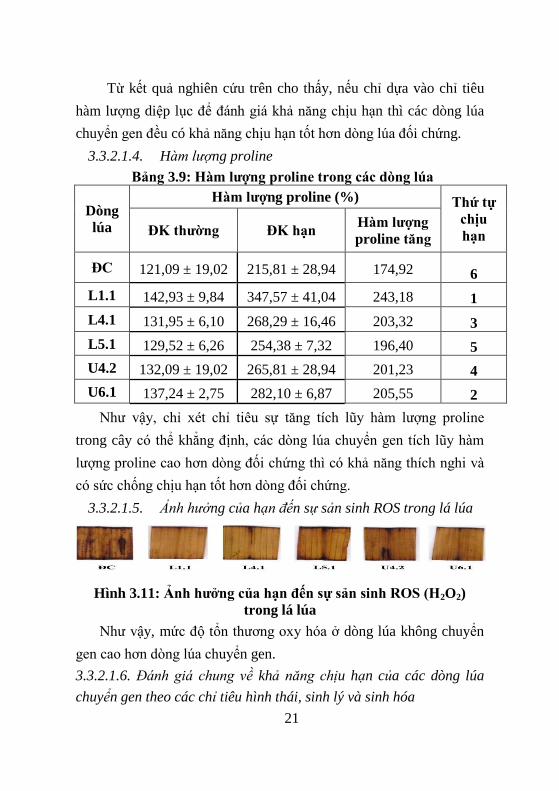
3.3.2.1.5. Ảnh hưởng của hạn đến sự sản sinh ROS trong lá lúa
Hình 3.11: Ảnh hƣởng của hạn đến sự sản sinh ROS (H2O2)
trong lá lúa
Nhƣ vậy, mức độ tổn thƣơng oxy hóa ở dòng lúa không chuyển
gen cao hơn dòng lúa chuyển gen.
3.3.2.1.6. Đánh giá chung về khả năng chịu hạn của các dòng lúa
chuyển gen theo các chỉ tiêu hình thái, sinh lý và sinh hóa
22
Bảng 3.10: Đánh giá chung khả năng chịu hạn dựa trên chỉ tiêu
hình thái, sinh lý v sinh hóa của các dòng lúa
Dòng
lúa
Chỉ tiêu đánh giá khả năng chịu hạn
Tổng
điểm
Thứ
tự
chịu
hạn
Độ
cuốn
lá
RWC
H m lƣợng
diệp lục Hàm
lƣợng
proline
Sự
sản
sinh
ROS a b Tổng
số
ĐC 6 6 6 6 6 6 6 42 6
L1.1 1 1 1 1 1 1 1 7 1
L4.1 3 2 2 2 2 3 3 17 2
L5.1 4 4 4 4 4 5 5 30 4
U4.2 5 5 5 5 5 4 4 33 5
U6.1 2 3 3 3 3 2 2 18 3
Trong nghiên cứu này, dựa trên các chỉ tiêu hình thái, sinh lý và
sinh hóa, cho thấy các dòng lúa chuyển gen có khả năng chịu hạn tốt
hơn so với dòng lúa đối chứng ở giai đoạn sinh sản.
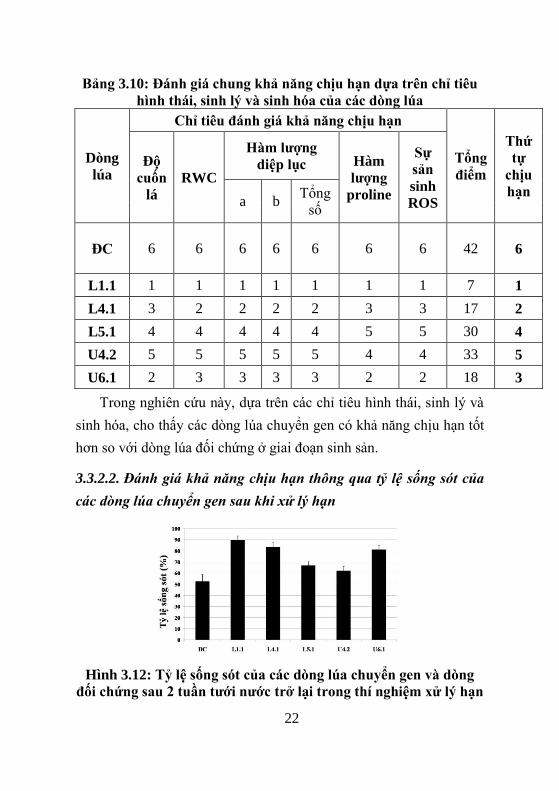
3.3.2.2. Đánh giá khả năng chịu hạn thông qua tỷ lệ sống sót của
các òng lúa chuyển gen sau khi xử lý hạn
Hình 3.12: Tỷ lệ sống sót của các dòng lúa chuyển gen v dòng
đối chứng sau 2 tuần tƣới nƣớc trở lại trong thí nghiệm xử lý hạn

23
Kết quả, các dòng lúa chuyển gen Ubi:OsNAC1 và
Lip9:OsNAC1 có khả năng chống chịu stress hạn cao hơn so với cây
lúa không chuyển gen. Đặc biệt, ở các dòng U4.2, U6.1 và L1.1 có tỷ
lệ sống sót ~ 90%, trong khi dòng đối chứng ~ 50%.
KẾT LUẬN
1. Đã phân lập thành công 3 gen mã hóa nhân tố phiên mã
OsNAC1, OsNAC5, OsNAC10 từ giống lúa Việt Nam. Các gen phân
lập đƣợc có kích thƣớc lần lƣợt 951 bp, 993 bp, 1182 bp và giống
99% (AY596808.1), 98% (AB028184.1) và 92% (AK069257.1) ở
giống lúa Japonica (Niponbare) đã đƣợc công bố trên Ngân hàng Gen
thế giới. Trình tự protein của các gen phân lâp đƣợc chứa đầy đủ các
vùng peptide đặc trƣng cho nhóm protein NAC và vùng tín hiệu định
vị nhân. Các gen OsNAC1, OsNAC5 và OsNAC10 đều biểu hiện trong
điều kiện xử lý hạn giả định, trong đó gen OsNAC1 có mức độ biểu
hiện cao hơn rõ rệt và duy trì mức độ biểu hiện cao trong thời gian dài
hơn đáng kể so với hai gen còn lại.
2. Đã thiết kế thành công hai hệ vector biểu hiện gen trong tế
bào thực vật: pCAMBIA1301 và pBI101 mang gen OsNAC1 đƣợc
điều khiển bởi promoter hoạt động liên tục Ubiquitin và 35S;
promoter hoạt động cảm ứng stress Rd29A và Lip9. Các vector mang
cấu trúc biểu hiện gen OsNAC1 đƣợc biến nạp thành công vào các
chủng vi khuẩn Agrobacterium tumefaciens EHA105.
3. Đã chuyển thành công cấu trúc Ubi:OsNAC1:Nos và
Lip9:OsNAC1:Nos vào giống lúa J02. Sự có mặt của cấu trúc biểu
hiện gen đích trong các dòng lúa chuyển gen đƣợc kiểm tra bằng
phƣơng pháp PCR. Các dòng lúa chuyển gen đồng hợp mang một

24
bản copy gen chuyển OsNAC1 đƣợc xác định bằng phƣơng pháp
qRT-PCR. Biểu hiện của gen chuyển OsNAC1 trong cây chuyển gen
T2 đã đƣợc chứng minh bằng phƣơng pháp RT-PCR.
4. Đã đánh giá sinh trƣởng, phát triển thông qua các chỉ tiêu
nông học (chiều cao cây, số nhánh, số hạt chắc/ bông, thời gian sinh
trƣởng) và khả năng chịu hạn của các dòng lúa chuyển gen thông qua
các chỉ tiêu về hình thái (độ cuốn lá), sinh lý (hảm lƣợng nƣớc tƣơng
đối, hàm lƣợng chất diệp lục) và sinh hóa (hàm lƣợng proline, sự sản
sinh chất hoạt hóa oxi hóa). 2 dòng lúa chuyển cấu trúc biểu hiện gen
Ubi:OsNAC1:Nos và 3 dòng lúa chuyển cấu trúc biểu hiện gen
Lip9:OsNAC1:Nos có khả năng sinh trƣởng tƣơng đƣơng với cây đối
chứng trong điều kiện bình thƣờng. Trong điều kiện xử lý stress hạn,
các dòng lúa chuyển gen có các chỉ tiêu hình thái, sinh lý, sinh hóa
thể hiện khả năng chịu hạn cao hơn dòng lúa đối chứng ở thời kỳ
sinh sản. Đồng thời, tỷ lệ sống sót của các dòng lúa chuyển gen thời
kỳ cây non cao hơn so với dòng đối chứng khi xử lý hạn. Cụ thể, các
dòng lúa chuyển gen L1.1, L4.2, U6.1, L5.1, U4.2 có tỷ lệ sống sót
là 90%, 84%, 81%, 67%, 62% cao hơn dòng đối chứng với tỷ lệ sống
sót chỉ đạt 53%.
KIẾN NGHỊ
Tiếp tục nghiên cứu, phân tích các dòng cây chuyển gen
OsNAC1 ở các thế hệ T3, T4… (tính bền vững di truyền, khả
năng duy trì tính chống chịu, năng suất…)..
Mở rộng nghiên cứu chuyển gen OsNAC1 vào các đối tƣợng
cây trồng khác với các promoter điều khiển hoạt động cảm
ứng điều kiện stress để phục vụ công tác chọn tạo giống.

DANH MỤC CÁC CÔNG TRÌNH KHOA HỌC CỦA TÁC GIẢ
LIÊN QUAN ĐẾN LUẬN ÁN
1. Phạm Thu Hằng, Nguyễn Duy Phƣơng và Phạm Xuân Hội
(2014), “Phân lập gen OsNAC10 liên quan tới tính chống chịu
hạn từ giống lúa Indica”, Tạp chí Công nghệ Sinh học 12(2), tr
319-326
2. Nguyễn Duy Phƣơng, Phạm Thu Hằng và Phạm Xuân Hội
(2014), “Phân lập gen mã hóa nhân tố phiên mã OsNAC5 liên
quan tới tính chống chịu stress từ giống lúa Indica”, Tạp chí
Khoa học ĐHQGHN: Khoa học Tự nhiên và Công nghệ 30(4), tr
40-47.
3. Phạm Thu Hằng, Nguyễn Duy Phƣơng, Trần Lan Đài, Phan
Tuấn Nghĩa và Phạm Xuân Hội (2014), “Thiết kế vector biểu
hiện mang gen OsNAC1 đƣợc điều khiển bởi promoter cảm ứng
điều kiện bất lợi RD29A”, Tạp chí Khoa học ĐHQGHN: Khoa
học Tự nhiên và Công nghệ 30(4), tr 1-10.
4. Phạm Thu Hằng, Nguyễn Duy Phƣơng, Phan Tuấn Nghĩa,
Phạm Xuân Hội (2016), “Nghiên cứu chuyển gen OsNAC1 liên
quan đến tính chịu hạn vào giống lúa Japonica”, Tạp chí Nông
nghiệp và phát triển nông thôn 1, tr 17-22.
5. Phạm Thu Hằng, Đàm Quang Hiếu, Phan Tuấn Nghĩa, Phạm
Xuân Hội (2016), “Thiết kế vector và chuyển gen OsNAC1 liên
quan đến tính chịu hạn vào giống lúa J02 (Oryza sativa L.
japonica), Tạp chí Công nghệ sinh học 14(2), tr 271 – 277.





























