Embed Size (px)
Citation preview

III. KLASIFIKACIJA I FILOGENIJA
Prof. dr Jelka Crnobrnja Isailović
ORGANSKA EVOLUCIJA

KLASIFIKACIJA
Opisivanje, imenovanje, razvrstavanje taksona.
FILOGENIJA
Istorija nastanka grupe organizama od zajedničkog pretka

LINNE-ov SISTEM KLASIFIKACIJE
“Systema naturae” 1735
1. BINOMNA
NOMENKLATURA
2. HIJERARHIJSKA KLASIFIKACIJA
(vrsta, rod, familija, red, klasa, filum, kraljevstvo…)

LINNE-ov SISTEM KLASIFIKACIJE
Apis pubescens, thorace subgriseo, abdomine fusco, pedibus posticis glabris utrinque margine ciliatis
= Apis melifera

LINNE-ov SISTEM KLASIFIKACIJE
Hijerarhijski nivo = taksonomska kategorija
Takson = svaka grupa
organizama unutar neke
taksonomske kategorije
Triturus cristatus Artenkreiss

LINNE-ov SISTEM KLASIFIKACIJE
Primata 4 paralelna gornja sekutića
2 grudne bradavice
Homo Simia Lemur Vespertilio

EVOLUCIJA I KLASIFIKACIJA
-Klasifikacija kao odraz stvarnih istorijskih procesa;
-KLADOGENEZA: grananje “loza” ili evolutivne linije u dve ili više potomačkih;
-ANAGENEZA: evolutivna promena različitih osobina kod svake od potomačkih linija
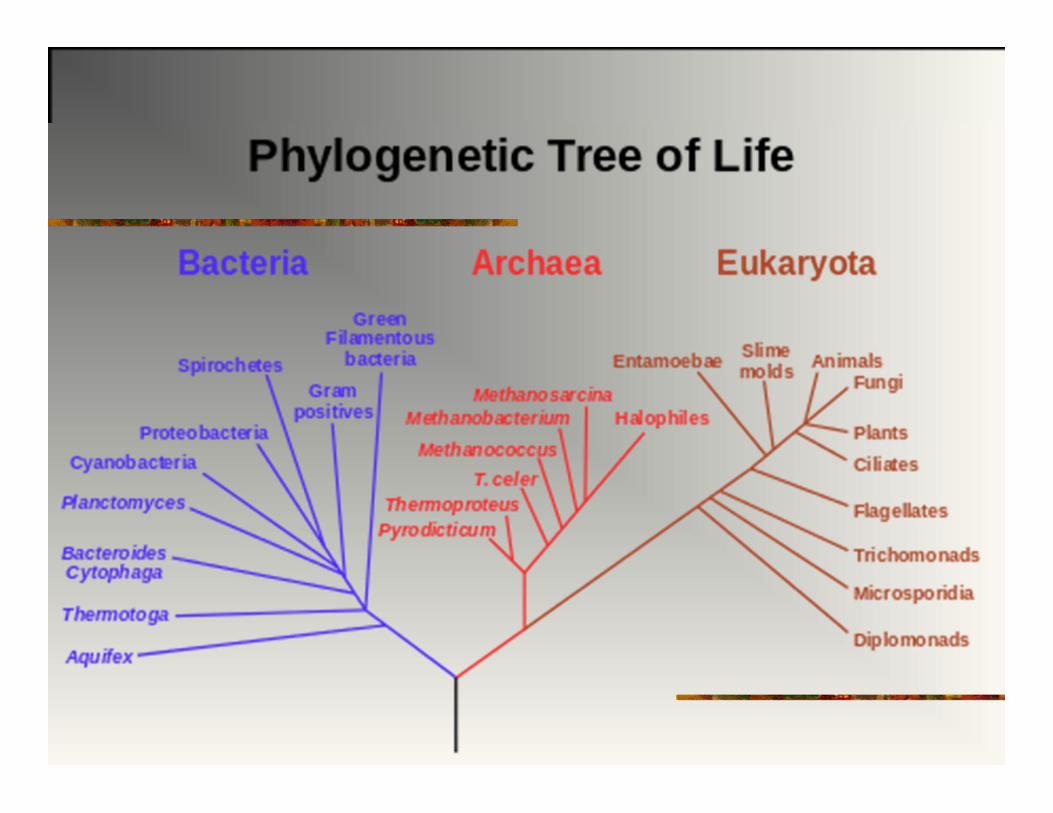

EVOLUCIJA I KLASIFIKACIJA
MONOFILETIČKA GRUPA:
Svi članovi vode poreklo od zajedničkog pretka.
Grupi pripada i zajednički predak i svi potomci.
Ovakva klasifikacija odslikava evolucionu istoriju
(genealoške odnose među taksonima)
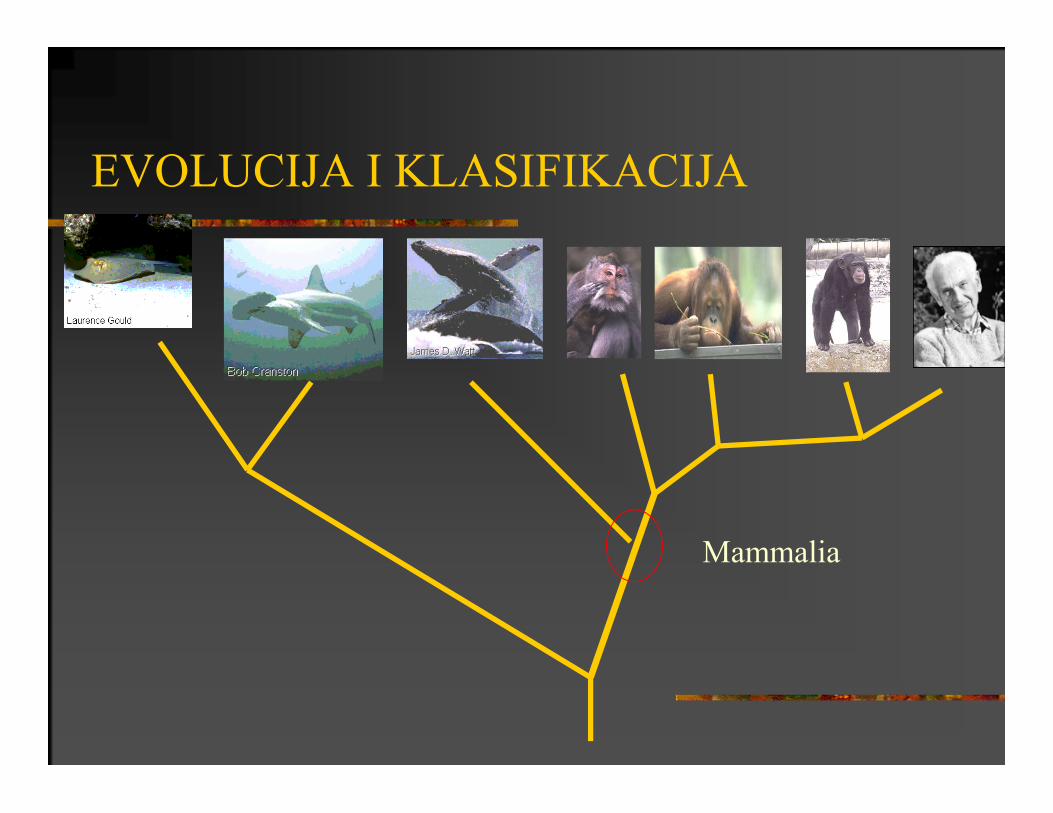
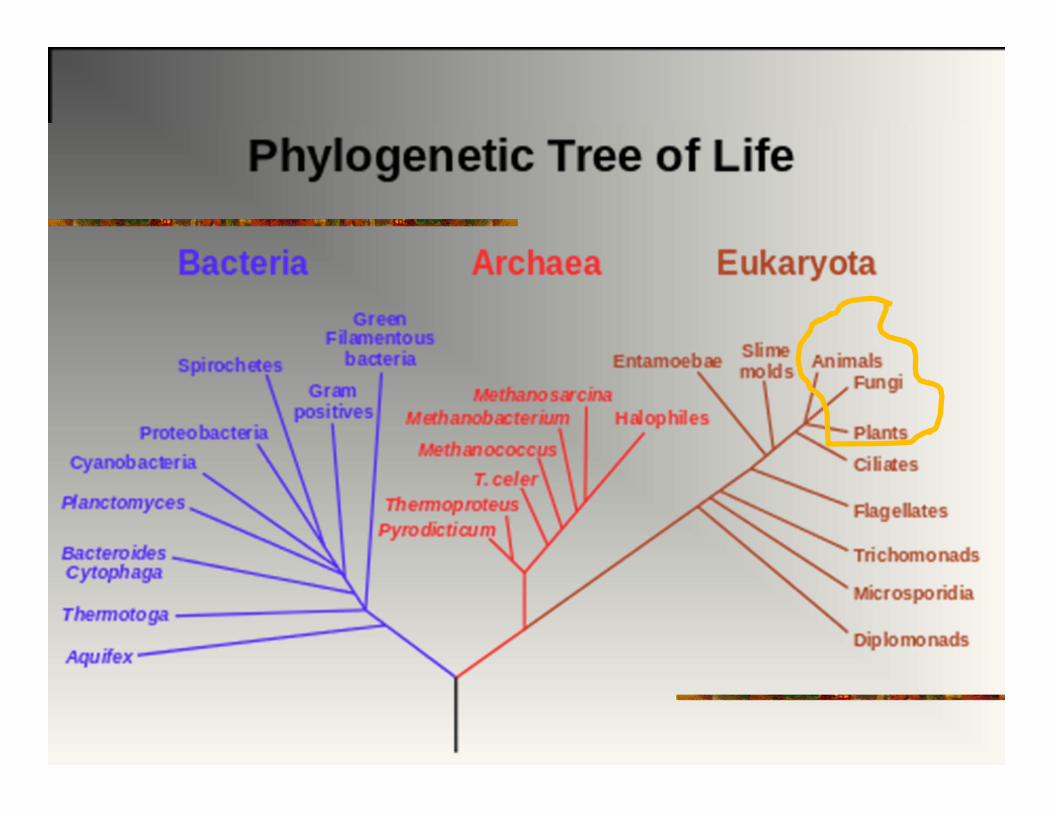
EVOLUCIJA I KLASIFIKACIJA
Mammalia

EVOLUCIJA I KLASIFIKACIJA
PARAFILETIČKA GRUPA:
Monofiletička grupa gde je klasifikacionom metodom izdvojen iz grupe neki od potomaka.
Ovakva klasifikacija ne sadrži potpunu informaciju o filogeniji grupe.
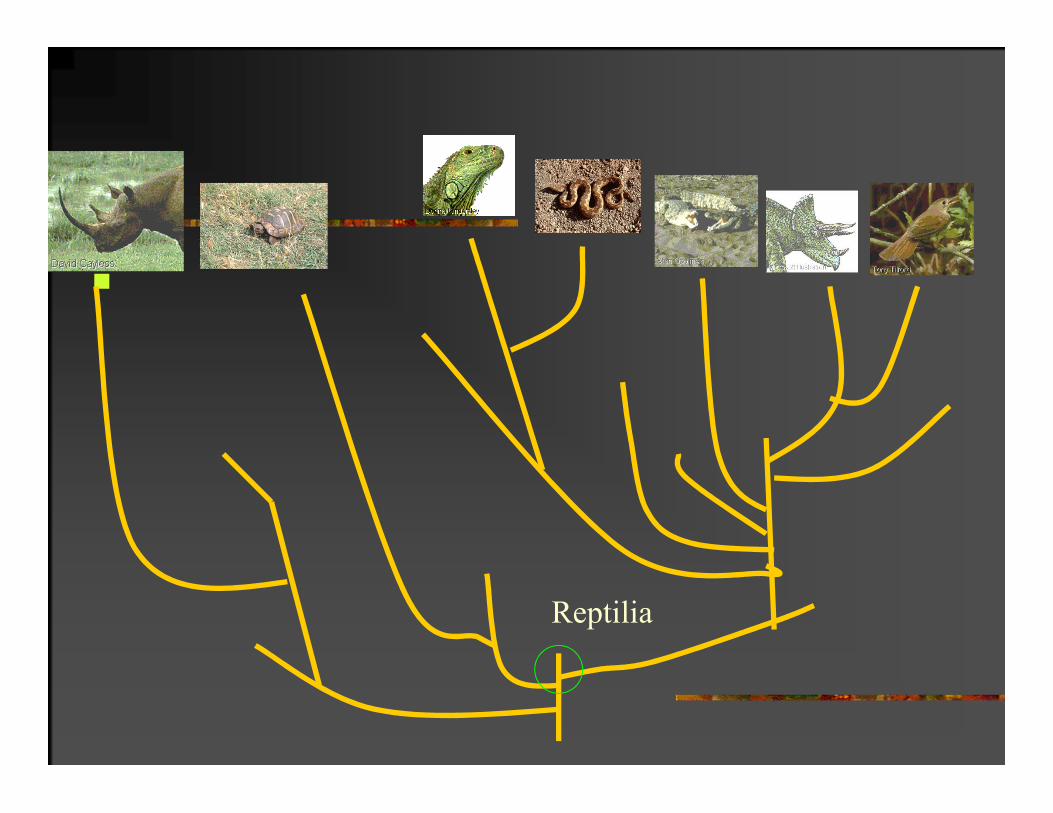
Reptilia

EVOLUCIJA I KLASIFIKACIJA
POLIFILETIČKA GRUPA:
Sastoji se od nesrodnih evolutivnih linija, od kojih je svaka više srodna sa vrstama koje nisu u toj grupi.
Do grupisanja je došlo zbog karakteristika koje su slične zbog konvergentne evolucije.
Kitovi (hidrodinamičan oblik tela Ajkule
&
dorzalno peraje)

FENETIKA I KLADISTIKA
FENETIKA
Michener i Sokal 1950.
Klasifikacija zasnovana ne na nekoliko subjektivno odabranih karakteristika već na stepenu ukupne sličnosti vrsta sa što više karakteristika obuhvaćenih analizom.

FENETIKA I KLADISTIKA
Sličnost = određena brojem istih stanja karaktera.
Srodstvo = kada su se dve ili više vrsta odvojile od zajedničkog pretka.

FENETIKA I KLADISTIKA
Dve vrste mogu imati ista stanja karaktera iz tri razloga:
1. dele izvedeno stanje karaktera –
placenta: ljudi i konji su srodniji međusobno nego bilo ko od njih sa gušterima.

FENETIKA I KLADISTIKA
Dve vrste mogu imati ista stanja karaktera iz tri razloga:
2. dele predačko (primitivno) stanje karaktera –petoprstost, koja je primitivno stanje, ujedinjuje ljude i guštere u odnosu na konje (jedan prst)

FENETIKA I KLADISTIKA
Dve vrste mogu imati ista stanja karaktera iz tri razloga:
3. dele slično stanje karaktera koje nije poreklom od zajedničkog pretka, već je evoluiralo nezavisno u nekoliko linija kao rezultat konvergentne evolucije -
dorzalno peraje kod ajkula i kitova.-
HOMOPLAZIJA

FENETIKA I KLADISTIKA
KLADISTIKA
Hennig 1950.
Novi kriterijumi:
klasifikacija treba da počiva striktno na filogenetičkim odnosima, a ne i na nivou adaptivne divergencije ili ukupne sličnosti.
Treba izbaciti homoplazične osobine iz analize, kao i predačke karakteristike.

FENETIKA I KLADISTIKA
KLADA = monofiletička grupa.
PLEZIOMORFAN = primitivan predački karakter.
SIMPLEZIOMORFAN = predački karakter koji je zastupljen u nekoliko taksona.
APOMORFAN = izveden ili napredan karakter.
SINAPOMORFAN = apomorfan karakter zastupljen kod nekoliko taksona.
AUTAPOMORFAN = apomorfan karakter koji poseduje samo jedan od n taksona unutar monofiletičke grupe.
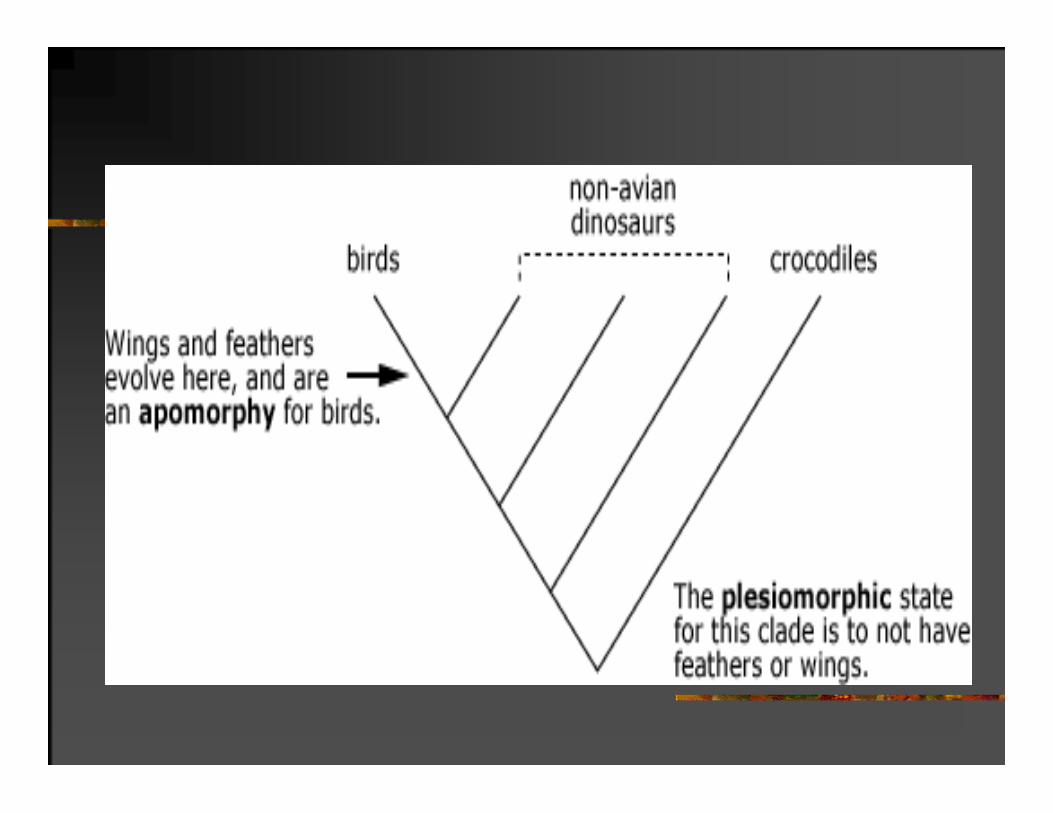

FENETIKA I KLADISTIKA
MONOFILETIČKE GRUPE SU DEFINISANE ZAJEDNIČKIM, JEDINSTVENO NASTALIM IZVEDENIM STANJIMA KARAKTERA.
Amnion izdvaja superklasu Amniota; donja vilica nastala samo od zubne kosti izdvaja klasu Mammalia; stalnorastući očnjaci i krila izdvajaju redove Rodentia i Chiroptera.
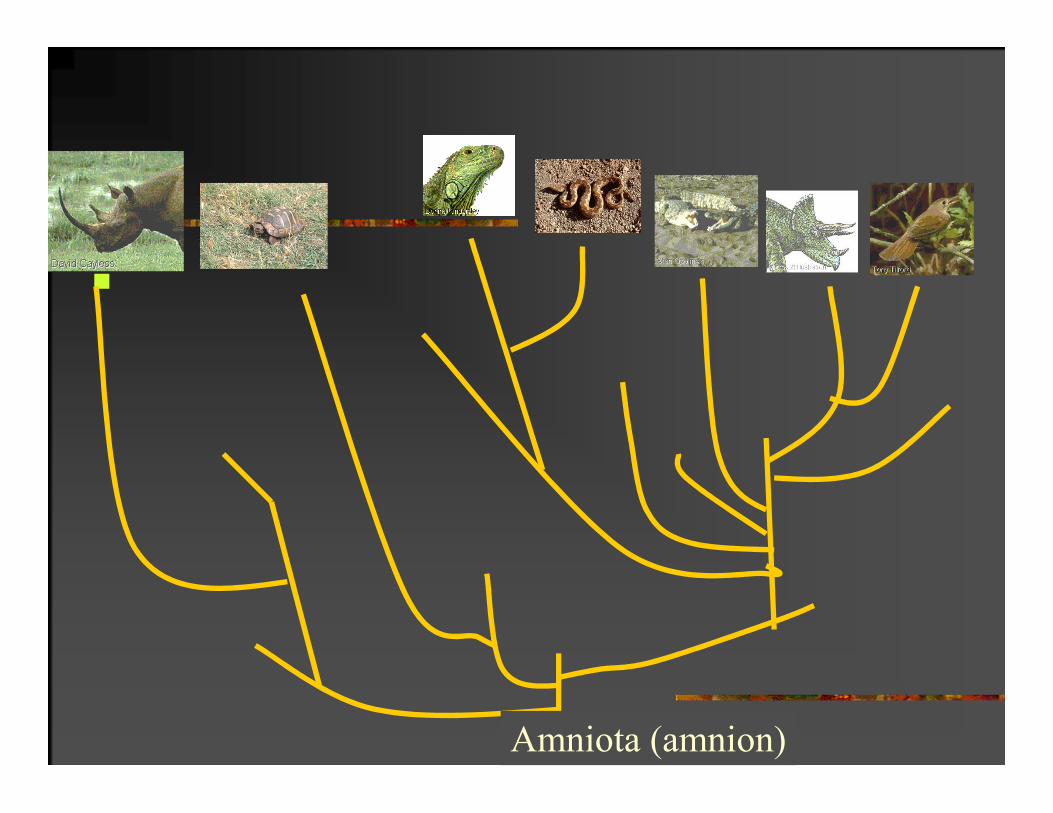
FENETIKA I KLADISTIKA
Amniota (amnion)

FENETIKA I KLADISTIKA
Zajednički izveden karakter treba da je evoluirao samo jednom – da bi grupa bila monofiletička. Ako utvrdimo da je evoluirao nekoliko puta unutar ispitivane grupe, to je homoplazičan karakter i nije podesan za analizu.
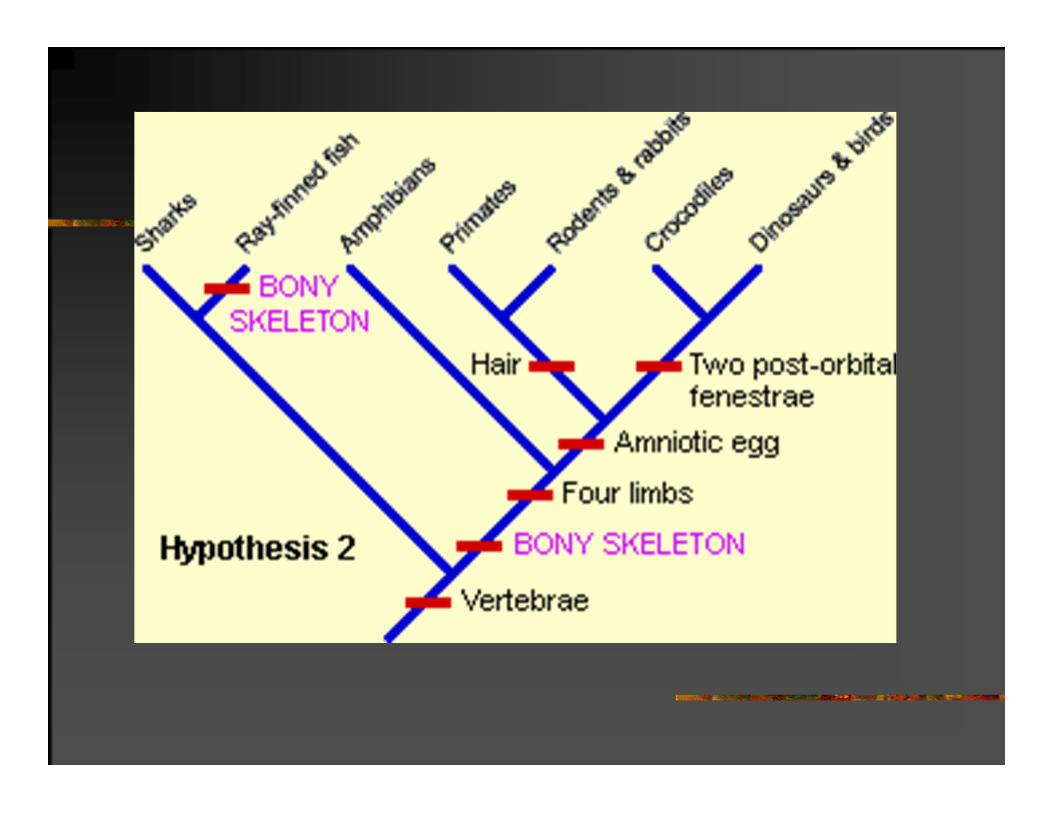
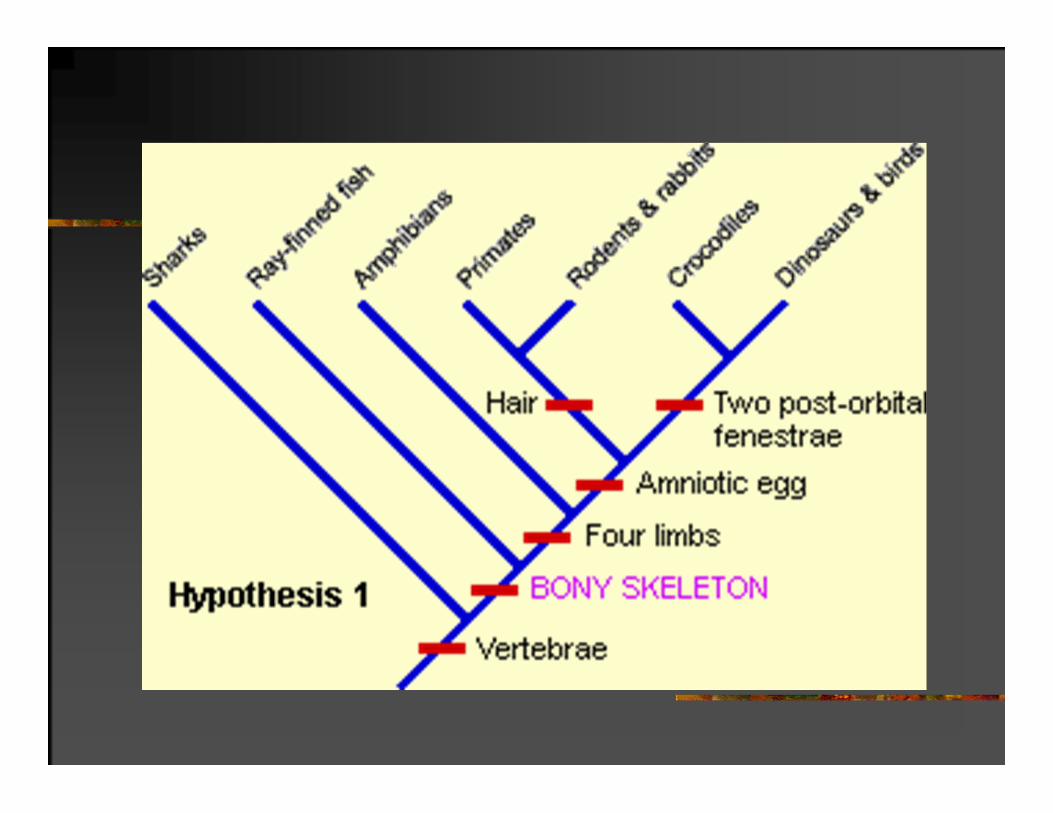

ŠTEDLJIVOST (PARSIMONIJA)I FILOGENETIČKI ZAKLJUČAK
Odabiramo objašnjenje koje zahteva najmanje nedokumentovanih pretpostavki
ili
najuverljivije filogenetičko stablo je ono koje daje rešenje na osnovu najmanjeg broja evolucionih promena.
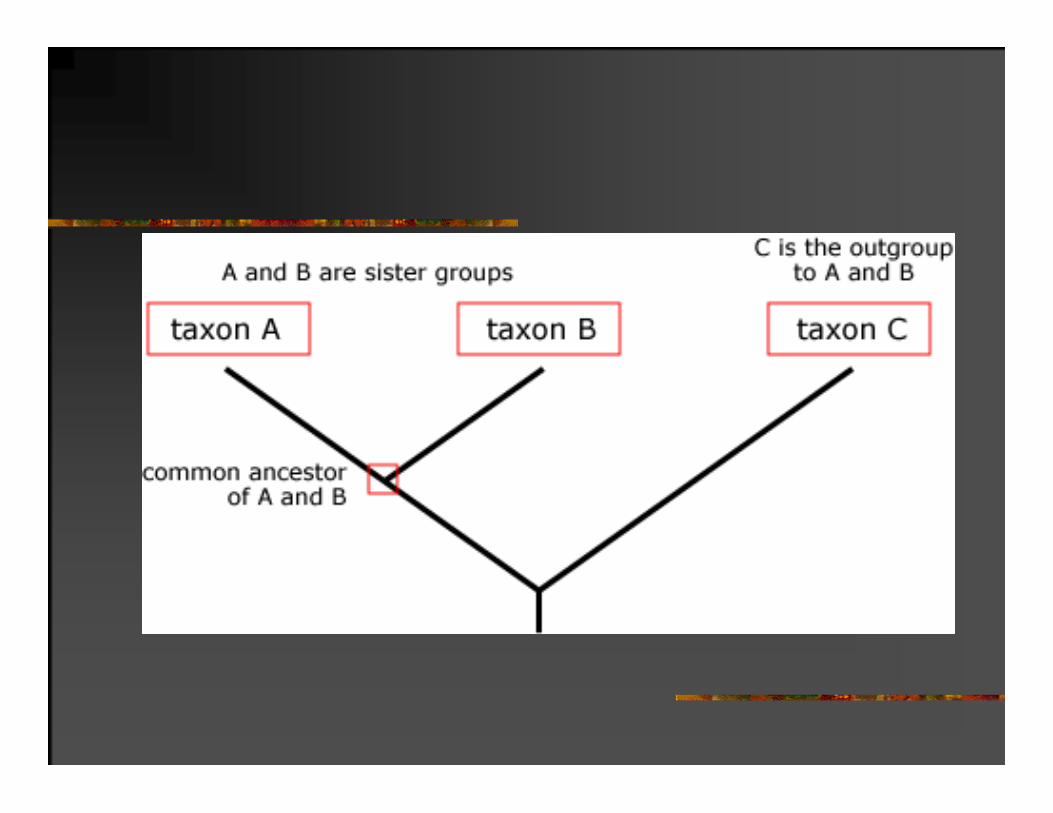

Najbolji način za potvrđivanje filogenetičke hipoteze je njeno slaganje sa nezavisnim podacima.
Neki od tih podataka su fosilni nalazi i geografsko rasprostranjenje.

FILOGENETIČKA ANALIZA I IZUMRLI ORGANIZMI
DNA iz fosila poredi se sa DNA recentnih rođaka. DNA pruža podatke o brzini evolucije DNA sekvenci.

PRINCIPI EVOLUCIONE ISTORIJE
PREUZETI IZ SISTEMATIKE
1. Homologe karakteristike su se razvile od zajedničkih predaka ( stanje karaktera može biti promenjeno)
3
5
5
5 prstiju
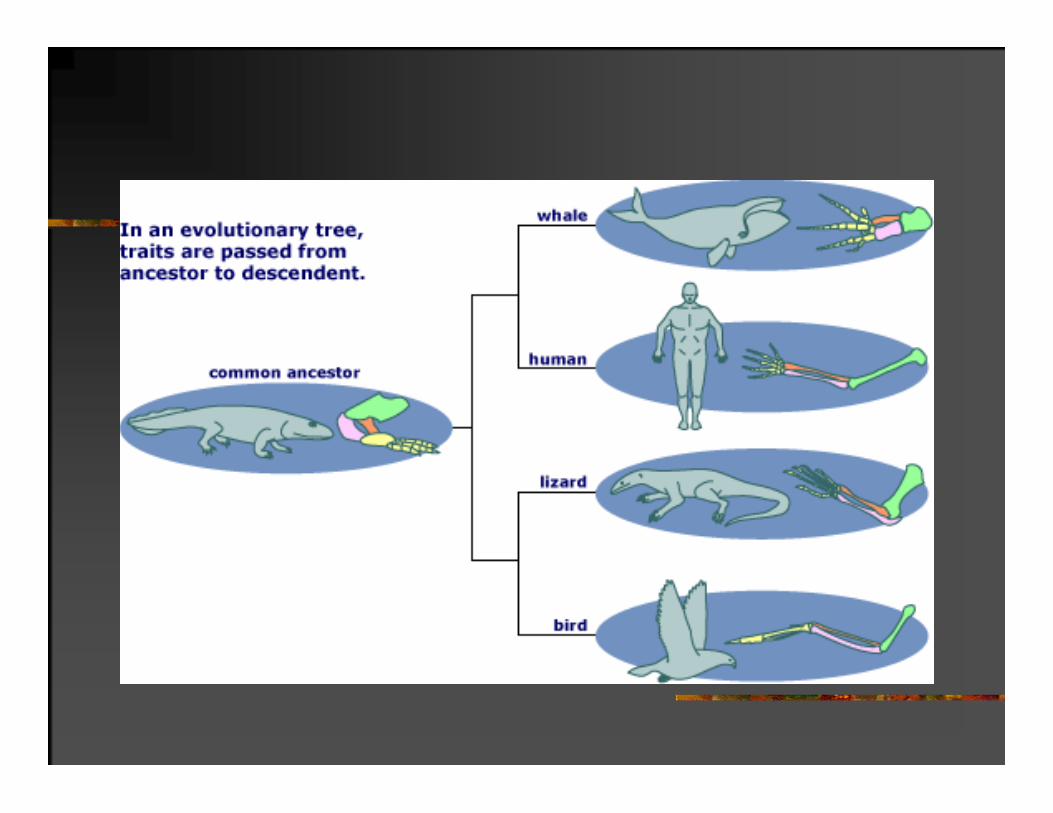

PRINCIPI EVOLUCIONE ISTORIJE
PREUZETI IZ SISTEMATIKE
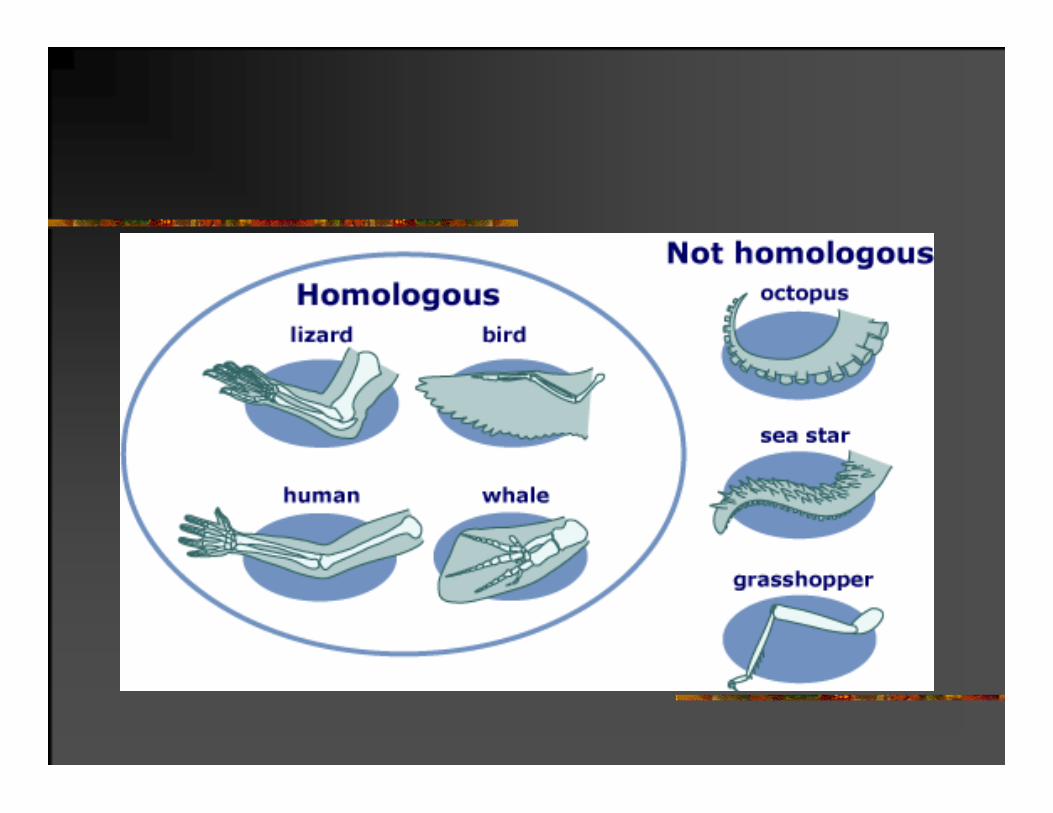
Kriterijumi homologosti:
-položaj karaktera u odnosu na druge delove tela;
-struktura:
-sličnost (građa) embrionalnih stupnjeva
Karakteristike organizma skoro uvek evoluiraju iz prethodno postojećih karakteristika njihovih predaka.
PRINCIPI EVOLUCIONE ISTORIJE
PREUZETI IZ SISTEMATIKE
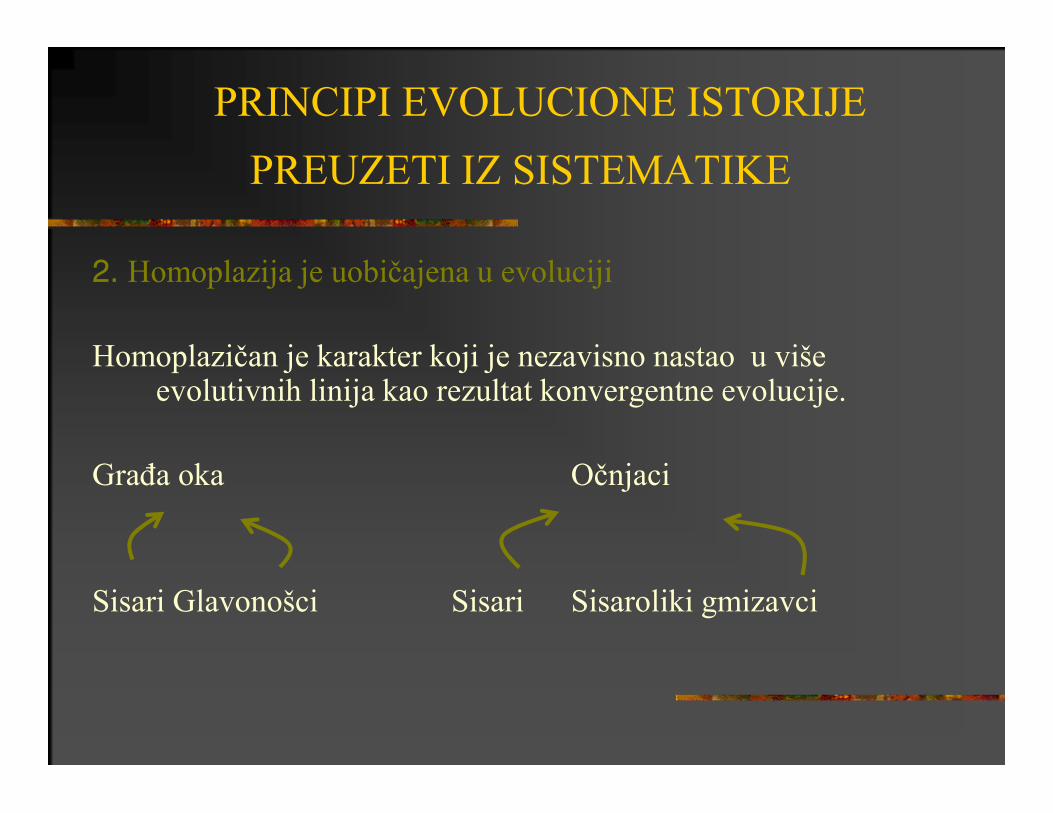
2. Homoplazija je uobičajena u evoluciji
Homoplazičan je karakter koji je nezavisno nastao u više evolutivnih linija kao rezultat konvergentne evolucije.
Građa oka Očnjaci
Sisari Glavonošci Sisari Sisaroliki gmizavci
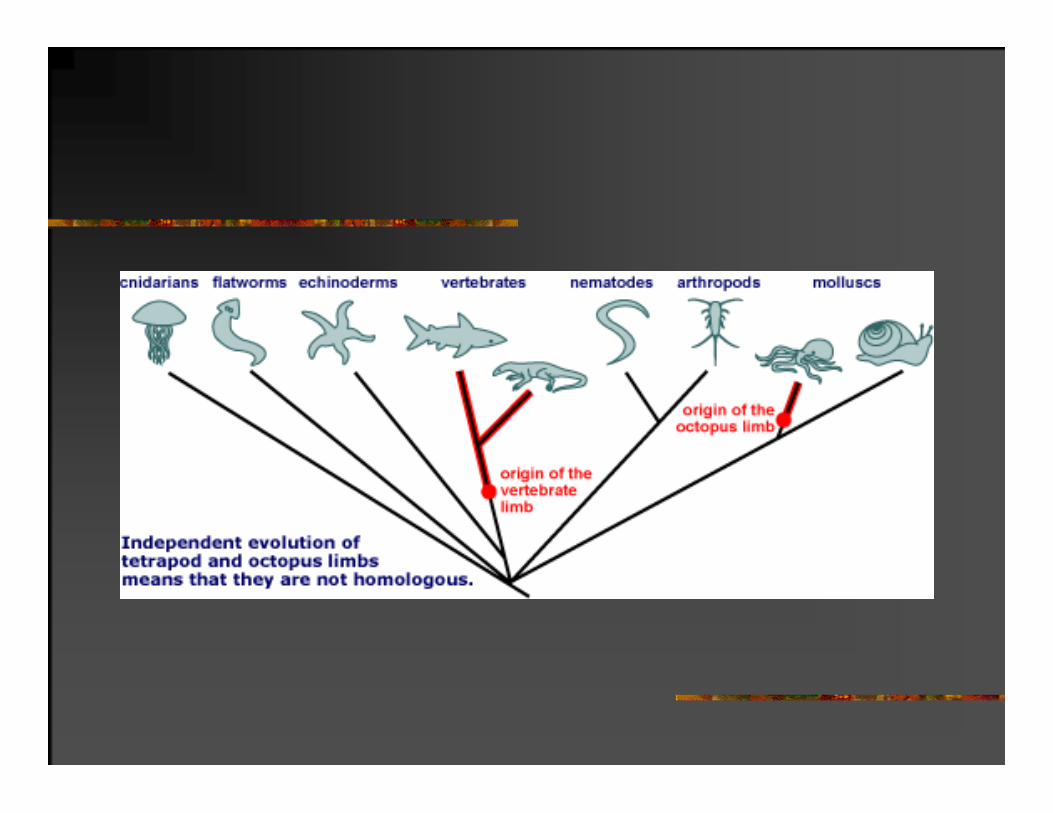

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
3.BRZINE EVOLUCIJE KARAKTERA SE RAZLIKUJU.
U odnosu na morfologiju zajedničkog pretka, svaki takson poseduje mešavinu predačkih i izvedenih stanja karaktera.
KONZERVATIVNI KARAKTERI
MOZAIČNA EVOLUCIJA

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
4.EVOLUCIJA JE ČESTO POSTEPENA
Nedostatak prelaznih ili intermedijarnih formi ne znači i da one nisu postojale.
Takođe, ne možemo dogmatski tvrditi da je postepena evolucija jedini vid evolucije.

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
5. KARAKTERISTIKE ČESTO DUGUJU SVOJU PROMENU FORME PROMENI FUNKCIJE.
Jedan od razloga zašto homolog karakter može mnogo da varira medju taksonima – zbog promenjenefunkcije.
Slušne koščice sisara – povezuju donju vilicu sa lobanjom kodgmizavaca-skeletni elementi škrga kod ribolikih predaka
Žaoka kod ženki Hymenoptera-legalica-zadnje noge

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
6. FILOGENETIČKA ANALIZA DOKUMENTUJE EVOLUCIONE TRENDOVE
EVOLUCIONI TREND – unutar evolucione linije kontinuirano menjanje od predačkog ka savremenim stanjima.
Scrophulariaceae – trend ka smanjenju broja prašnika:
5 4+sterilni 4 2

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
7. VEĆINA KLADA POKAZUJE EVOLUTIVNU RADIJACIJU
divergentna evolucija mnoštva srodnih evolutivnih linija za relativno kratko vreme.
Modifikovane za različite načine života
različite okoline
različita hrana
različit način izbegavanja predatora.
Adaptivna radijacija

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
MOLEKULARNI PODACI U FILOGENETIČKOJ ANALIZI
DNA sekvence koje brzo evoluiraju korisne su za analizu odnosa među blisko srodnim vrstama.
DNA sekvence koje sporo evoluiraju korisne su za prepoznavanje srodnosti među veoma starim grupama.

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
GENEALOGIJE GENA
Filogenetska stabla gena predstavljaju istorijske odnose između varijanti DNK sekvenci(=haplotipova) odabranog gena kod različitih taksona.
Ona ne odražavaju obavezno evolucionu istoriju vrsta.

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
MOLEKULARNI SAT
Koncept po kome makromolekuli evoluiraju konstantnom stopom
Sat može biti kalibrisan putem podataka od fosilnih nalaza o apsolutnom vremenu divergencije određenih taksona.

PRINCIPI EVOLUCIONE ISTORIJE PREUZETI IZ SISTEMATIKE
RELATIVNA BRZINA EVOLUCIJE
KALIBRISANA BRZINA
Korelacija između molekularnih razlika i starosti je prilično velika, ali ne potpuna.
Na primer, smatra se da se evolucija ljudske linije odvijala sporije od procene.

DOKAZI EVOLUCIJE
1.HIJERARHIJSKA ORGANIZACIJA ŽIVOTA
2.HOMOLOGIJA
3.EMBRIOLOŠKA SLIČNOST
4.PREDAČKE KARAKTERISTIKE. Slepo crevo, repna kost, rudimentarni mišići pomoću kojih neko može da mrda ušima ili skalpom, umnjaci, mnoštvo nefunkcionalnih DNA sekvenci kao pseudogeni.

DOKAZI EVOLUCIJE
5.KONVERGENCIJA – funkcionalno slične karakteristike koje se razlikuju po strukturi.
6.SUBOPTIMALNI DIZAJN. Ukrštanje puteva hrane i vazduha kod farynxa; "slepa mrlja" u ljudskom oku. Funkcionalno besmislen aranžman aksona ćelija retine koji idu do oka, konvergiraju u optički nerv koji prodire kroz retinu nazad u mozak.

DOKAZI EVOLUCIJE
7.GEOGRAFSKA RASPROSTRANJENOST . Ne bi imala smisla da ne postoji zajednički predak. Marsupialia su raspoređeni na južnoj hemisferi zbog predaka koji su naseljavali južnu kopnenu masu koja je počela da se deli u Mezozoiku.
8.PRELAZNE FORME. Nove činjenice potvrđuju realnost procesa evolucije i bez fosilnih nalaza.

IV. FOSILNI NALAZI
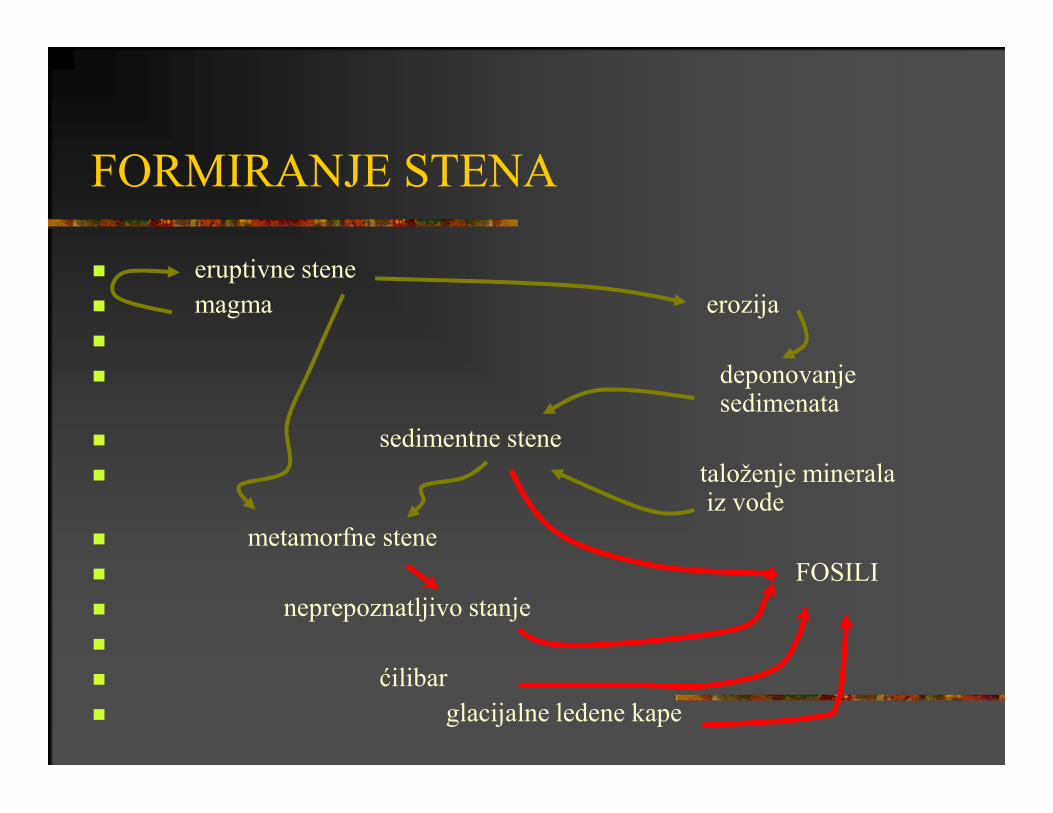
FORMIRANJE STENA
eruptivne stene
magma erozija
deponovanjesedimenata
sedimentne stene
taloženje mineralaiz vode
metamorfne stene
FOSILI
neprepoznatljivo stanje
ćilibar
glacijalne ledene kape

MERENJE GEOLOŠKOG VREMENA
Apsolutni proračuni geoloških događaja vrše se merenjem raspada određenih radioaktivnih elemenata u mineralima eruptivnih stena.
radioaktivni uranijum
U-235 Pb207
nalazi se samo u eruptivnim stenama
poluživot 0.7 biliona godina
Preciznost procene starosti opada sa porastom starosti stena.

MERENJE GEOLOŠKOG VREMENA
Radioaktivni ugljenik
C-14 ima poluživot od 5730 godina
koristan je za datiranje organske materije do
40 000 i 70 000 godina starosti.

SEDIMENTNE STENE
Neki geološki periodi su predstavljeni slojevima na samo nekoliko mesta na Zemlji.
Što je stariji geološki period, teže je naći sloj stena koji je iz tog vremena zbog erozije i metamorfoza ili velike dubine na kojoj se nalaze.
Kanjoni su izuzetak.
Mesta dugih i neisprekidanih nalaza su obično u okeanima i jezerima.

TAKSONOMIJA FOSILNIH NALAZA
HRONOVRSTE – različiti stupnjevi unutar jedne linije – Homo sapiens i H. erectus, na primer. Ime H. erectus može nestati jer se forma promenila unutar linije. To se zove PSEUDOIŠČEZAVANJE.
Paleontolozi sa većom sigurnošću diskutuju rodove i više taksonomske kategorije jer je prepoznavanje da li su dva nalaza dve različite vrste veoma teško.

ODNOSI IZMEĐU FILOGENIJE I FOSILNIH NALAZA
Filogenetski odnosi mogu biti objašnjeni dobijanjem informacije o izumrlim vrstama. U bilo kojoj filogenetskoj analizi, informacija o polarnosti promena filogenetski informativnih karakteristika je neobično važna. Fosili mogu pružiti presudnuinformaciju.
Na primer, evolucija artikularne kosti koja povezuje sisare sa primitivnim gmizavcima bila je skoro nezamisliva dok nisu nađeni fosili therapsida.

BRZINA EVOLUCIJE
BRZINA TAKSONOMSKE UČESTALOSTI
brzina kojom novi taksoni (rodovi) zamenjuju predačke. Zavisi od brzine nastanka taksona i brzine izumiranja (uključujući i taksonomsko „izumiranje").

BRZINA EVOLUCIJE
FILOGENETIČKA (FILETIČKA) BRZINA EVOLUCIJE –
brzina promene pojedinačnih
karaktera ili kompleksa
karaktera unutar određene
linije. Posle visoke početne
brzine promene, te linije
evoluiraju veoma sporo, a njihovi
živi predstavnici često su označeni
kao “živi fosili".

BRZINA EVOLUCIJE
Mera brzine evolucije je darwin = promena za faktor baze prirodnog logaritma (2.718) po milionu godina.
Oprez! Paleontološka literatura koja se tiče brzine evolucije ne daje dobre slučajne uzorke jer supaleontolozi obično usmereni na karaktere koji pokazuju evolutivnu promenu, a ignorišu karaktere koji n i s u d i s k r i m i n a t i v n i
(KONZERVATIVNI KARAKTERI tj oni sa vema malom brzinom evolucije).

BRZINA EVOLUCIJE
Brzina evolucije unutar predačko – potomačke serije fosila.
Prosečne brzine evolucije su ekstremno male.
Računaju se kao razlika između srednjih vrednosti na početku i na kraju vremenskog intervala , podeljeno tim intervalom
(ymean1 – ymean0)/T
T se meri u milionima godina.

ZNAČAJ FOSILNIH NALAZA
JEDINO ONI MOGU DA DAJU INFORMACIJE O:
-organizmima koji vise ne postoje-velikim izumiranjima i diverzifikacijama živog sveta-kretanjima kontinenata-apsolutnoj vremenskoj skali evolucionih događaja-fenotipskim transformacijama unutar pojedinih evolucionih linija-dinamici promene biološke raznovrsnosti od nastanka živih bića do danas





























