Embed Size (px)
Citation preview

Prof. dr. sc. Darko Grbeša redoviti je sveučilišni profesoru trajnom zvanju, predaje Hranidbu životinja na dodi-plomskom studiju, voditelj i nositelj je dva predmetana diplomskom studiju Hranidba i hrana za životinje naAgronomskom fakultetu Sveučilišta u Zagrebu. Objavio je 175 radova među kojima su neki u prestižnimmeđunarodnim znanstvenim časopisima kao što jeFood Chemistry. Prema Web of Science D. Grbeša je umeđunarodno priznatim časopisima najcitiraniji profesorhranidbe životinja iz RH. Prof. Grbeša autor je tri knjigei pet poglavlja u drugim knjigama. Pod njegovim men-torstvom obranjeno je pet doktorskih teza. Trostruki jestipendist Cochran Fllowship Programa američkog mi-nistarstva poljoprivrede.Savjetnik je tvornica krmnih smjesa i najvećih hrvatskihmliječnih farmi. Održao je preko 150 predavanja o hra-njivosti krmiva i hranidbi krava u Hrvatskoj i inozemstvu.
Bc Institut d.d., Rugvica, Dugoselska 7, 10370 Dugo Selo10000 Zagreb, Hrvatska
Prof. dr. sc. Darko Grbeša
Hranidbena svojstva kukuruza
ISBN 9789535507505

Hranidbena svojstva kukuruza
Zagreb, 2016.
Prof. dr. sc. Darko Grbeša
Nema bolje prakse od dobre teorije

Recenzenti:Prof. dr. sc. Stjepan Mužic
Doc. dr. sc. Goran Kiš
Lektor: Jadranka Vrbnjak Ferenčak
CIP zapis je dostupan u računalnome katalogu Nacionalne i sveučilišne knjižniceu Zagrebu pod brojem 000926156.
ISBN 9789535507505

1. Značenje kukuruza..........................................................................................................7
2. Upotreba kukuruza .........................................................................................................92.1. Hrana za životinje ...................................................................................................................92.2. Bioetanol ...............................................................................................................................11
3. Građa zrna.........................................................................................................................123.1. Endosperm.......................................................................................................................12
3.1.1. Tipovi endosperma ..................................................................................................133.2. Klica .................................................................................................................................153.3. Omotač ............................................................................................................................15
4. Fizikalna svojstva ..........................................................................................................164.1. Hektolitarska masa ..........................................................................................................164.2. Težina 1000 zrna..............................................................................................................174.3. Oštećenja zrna.................................................................................................................18
5. Kemijski sastav ..............................................................................................................195.1. Vlaga................................................................................................................................195.2. Škrob................................................................................................................................19
5.2.1. Sadržaj.....................................................................................................................205.2.2. Građa škroba ...........................................................................................................21
5.2.2.1. Amiloza ..................................................................................................................215.2.2.1.1. Sadržaj amiloze i amilopektina u Bc hibridima ........................................................22
5.2.2.2. Granule škroba .......................................................................................................235.2.2.2.1. Građa granula škroba devet Bc hibrida kukuruza ....................................................23
5.2.2.3. Zeinski omotač granula škroba ..................................................................................245.2.3. Mjesto i brzina probave škroba ................................................................................25
5.2.3.1. Svinje i perad..........................................................................................................255.2.3.1.1. Čimbenici brzine probave škroba ..........................................................................265.2.3.1.2. Brzo probavljiv škrob...........................................................................................275.2.3.1.3. Sporo probavljiv škrob.........................................................................................285.2.3.1.4. Rezistentan škrob...............................................................................................285.2.3.1.5. Udjeli frakcija u ukupnom škrobu Bc hibrida ...........................................................285.2.3.1.6. Brzina probavljivosti............................................................................................295.2.3.1.7. Brzina probave škroba Bc hibrida .........................................................................30
5.2.3.2. Preživači ................................................................................................................315.2.3.2.1. Razgradljivi škrob ...............................................................................................32
5.2.3.2.1.1. Razgradljivost škroba kukuruza .......................................................................335.2.3.3. Kunići ....................................................................................................................34
6. Šećeri...................................................................................................................................35
7. Protein.................................................................................................................................367.1. Protein kukuruza ..............................................................................................................367.2. Sadržaj.............................................................................................................................377.3. Zeini .................................................................................................................................38
7.3.1. Sadržaj zeina u Bc hibridima kukuruza....................................................................397.4. Preživači ..........................................................................................................................417.5. Aminokiseline ...................................................................................................................42
Sadržaj:

8. Ulje ........................................................................................................................................448.1. Sadržaj.............................................................................................................................448.2. Masne kiseline .................................................................................................................45
9. Vlakna..................................................................................................................................479.1. Funkcija............................................................................................................................479.2. Sadržaj.............................................................................................................................48
10. Energija ............................................................................................................................49
11. Minerali .............................................................................................................................54
12. Ukusnost kukuruza ....................................................................................................5513. Fitonutrijenti ..................................................................................................................56
13.1. Fenoli .............................................................................................................................5613.2. Karotenoidi.....................................................................................................................58
13.2.1. Karotenoidi Bc hibrida kukuruza ............................................................................5913.2.2. Pojedini karotenoidi ...............................................................................................6213.2.3. Parametri boje zrna ...............................................................................................6413.2.4. Pigmentacijska svojstva karotenoida .....................................................................6413.2.5. Sadržaj vitamina A .................................................................................................67
13.3. Vitamin E........................................................................................................................6913.3.1. Pojedini tokoferoli ..................................................................................................70
13.4. Antioksidacijska moć......................................................................................................72
14. Klima .................................................................................................................................7414.1. Kemijski sastav ..............................................................................................................7414.2. Fizička svojstva zrna......................................................................................................76
15. Mikotoksini .....................................................................................................................7815.1. Uvod...............................................................................................................................7815.2 Klimatski ekstremi ...........................................................................................................7815.3. Kukuruz..........................................................................................................................8115.4. Bc hibridi u sušnoj 2012.................................................................................................8215.5. Antioksidansi Bc hibrida i otpornost ...............................................................................85
16. Mljevenje .........................................................................................................................8816.1. Svinje .............................................................................................................................8816.2. Perad .............................................................................................................................8916.3. Preživači ........................................................................................................................89
17. Silaža cijele biljke kukuruza...................................................................................9117.1. Stadij zrelosti..................................................................................................................92
17.1.1. Promjene kemijskog sastava .................................................................................9417.2. Siliranje mijenja sastav i probavljivost............................................................................9617.3. Hibrid..............................................................................................................................9817.4. Spremanje silaže .........................................................................................................101
18. Poruke ............................................................................................................................107

Predgovor
Porast stanovništva prati i znatniji porast potrošnje animalnih proizvoda,osobito mesa peradi i svinja čija je glavna hrana kukuruz. Veći porastpotrošnje pak zahtijeva veću proizvodnju i efikasnije iskorištenje energiješkroba kukuruza u animalnoj proizvodnji. Isto tako, preduvjet učinkovite kon-verzije zdravlje su zrnja i životinja koje ovisi i o antioksidansima kukuruza.Zbog svega toga naš je cilj u animalnoj proizvodnji proizvesti što više mesa,mlijeka i jaja sa boljim iskorištenjem kukuruza.
Hranidbena svojstva objedinjuju hranjivu vrijednost, antioksidacijska, pig-mentacijska i provitamin A djelovanja kukuruza. Hranidbena svojstva utječuna efikasnost proizvodnje, zdravlje životinja i konzumenata animalnihproizvoda. Zabluda je svođenje selekcije kukuruza na puki prinos škroba jerzanemaruje genetsko bogatstvo sadržaja bioaktivni tvari kukuruza koje subitne za održivu proizvodnju te zdravlje kukuruza, životinja i čovjeka.
Samo holistički (svestrani i cjelokupan) pristup može dati pravu sliku vri-jednosti pojedinih hibrida kukuruza u proizvodnji i zdravlju životinja. Bitna jeoptimalizacija koja zadovoljava ne jedno nego više svojstava hibrida, koja sučesto suprotnih zahtjeva. Primjerice, amiloza smanjuje probavljivost ali povi-suje otpornost na pojavu aflatoksina pa je potrebno naći razinu amiloze kojapovisuje otpornost, a u znatnoj mjeri ne smanjuje probavljivost.
Često se postavlja pitanje jesu li dovoljno velike genetske razlike hranid-benih svojstva između hibrida. Međutim, kukuruz je biljka jedinstvenegenetske raznolikosti jer se dvije inbred linije više razlikuju nego hominidi kojisu se razdvojili prije 3,5 milijuna godina.
Bc Institut iz Zagreba jedan je od rijetkih proizvođača sjemena u svijetukoji sustavnom primjenom istraživanja fizikalnih, kemijskih i bioloških svojstvau selekciji kukuruz čini boljom kulturom, kako za ratare tako i za stočare.
Neočekivano velike promjene klimatski prilika tijekom jedne vegetacijskesezone uzrokuju nestabilne prinose visokorodnih hibrida pa se traže hibridine toliko visokih koliko stabilnih prinosa. Dodatno, danas je zdravstvena is-pravnost hrane njezino najvažnije svojstvo koje ozbiljno narušavaju češće iopsežnije zaraze mikotoksinima uzrokovane klimatskim promjenama.(Ne)sklonost razvoju mikotoksina, ako nije najvažnije onda je među najvažni-jim svojstvima svakog hibrida kukuruza.
Cilj je ove monografije o hranidbenim svojstvima kukuruza iznijeti dublje isveobuhvatne svjetske znanstvene spoznaje o čimbenicima zrna koji utječu
5

6
na (1) što bolju pretvorbu energije kukuruza u animalne proizvode i (2)zdravstvenu ispravnost kukuruza te zdravlje životinja.
U prvom dijelu publikacije opisane su definicije i značenje te prosječne vri-jednosti fizikalnoga i kemijskog sastava, a koji upućuju na hranidbena svo-jstva kukuruza. Osobiti naglasak stavljen je na utjecaj zeina i struktureendosperma na kinetiku probave škroba koja bitno utječe na konverzijuhrane. Isto tako opisani su i značenje i karakteristične vrijednosti sadržaja fi-tonutrijenata, te njihove antioksidacijske i pigmentacijske vrijednosti u Bc hi-bridima kukuruza.
U drugom dijelu publikacije tablično su prikazane petogodišnje prosječnevrijednosti 75 svojstava 27 Bc hibrida kukuruza. Isto tako, dane su i nam-jenske preporuke za korištenje svakog od 27 hibrida kukuruza.
Knjiga je prvenstveno namijenjena ratarima koji proizvode kukuruz istočarima koji koriste kukuruz, ali isto tako i obiteljskim gospodarstvima testudentima.
Zahvaljujem na uloženom trudu, vremenu i znanju djelatnicama Zavodaza hranidbu životinja Kristini Kljak, Mariji Duvnjak, Aniti Brnić, Kristini Mikuličić,Aniti Kurilić i Vlatki Radanović.
Isto tako zahvaljujem se sljedećim mr. ing. Hranidbe životinja i hrane kojisu diplomirali na temama iz hranidbenih svojstava Bc hibrida kukuruza: Mar-tina Drdić, Martina Faltak, Marija Kos, Tomislav Njari, Siniša Škvorc, MarioGal, Josip Kadić, Maja Jurkas, Marijan Sučija, Igor Bemko.
7
1. Značenje kukuruza
Kukuruz se od svih žitarica najviše, ili 988 milijuna tona, proizvede naoko 160 milijuna hektara u svijetu. Isto tako, najviše od svih žitarica kuku-ruza se potroši u hranidbi životinja, pa u prehrani ljudi te u proizvodnji bioe-tanola, plastike, vlakana i kukuruznog sirupa. Točnije, u hranidbi životinjapotroši se najviše ili 67% svjetske proizvodnje kukuruza pa on od svih ra-tarskih i industrijskih kultura najviše ili 33% podmiruje energetske i čak 13%proteinskih potreba svjetske animalne proizvodnje. Proizvodnja kukuruzaiznimno je važna privredna djelatnost, primjerice u SAD-u je u 2014. ostva-rila prihod od gotovo 62 milijarde dolara, što je više nego u proizvodnji svihostalih žitarica zajedno.
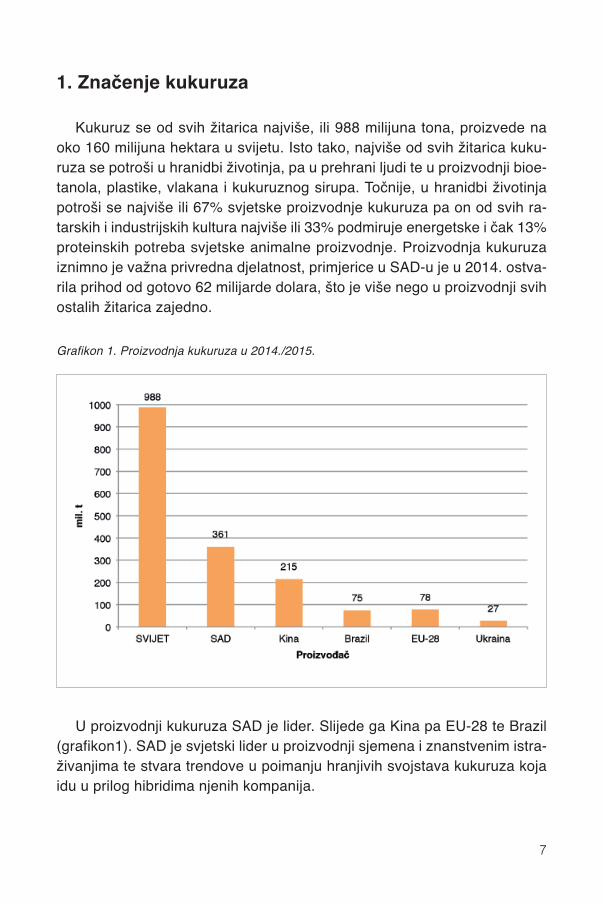
U proizvodnji kukuruza SAD je lider. Slijede ga Kina pa EU-28 te Brazil(grafikon1). SAD je svjetski lider u proizvodnji sjemena i znanstvenim istra-živanjima te stvara trendove u poimanju hranjivih svojstava kukuruza kojaidu u prilog hibridima njenih kompanija.
Grafikon 1. Proizvodnja kukuruza u 2014./2015.

8
EU proizvodi više nego dovoljno pšenice (127%) i ječma (124,4%) i ne-dovoljno kukuruza (87,3%) za zadovoljenje hranidbenih potreba vlastite ani-malne proizvodnje (FEFAC, 2015.). U EU-28 proizvede se 78 milijuna tonakukuruza i još uveze 11,5 milijuna tona žitarica za hranidbu životinja. Naj-veća je proizvodnja (u milijunima tona u 2014.) kukuruza u Francuskoj(18,5), pa Rumunjskoj (12), slijede Italija i Mađarska (9,2). Iako Hrvatskaproizvodi 2,1 milijun tona ili 2,7% europske proizvodnje, mi smo 10. po reduproizvođač kukuruza među 28 država EU-a.
Porast broja stanovnika za 2 milijarde do 2050. Ili 156 000 dnevnopovećava potrebe za hranom za 50-70%. Dodatni porast potrošnje kukuruzauzrokuju zemlje brzorastuće populacije u kojima se uslijed promjena pre-hrambeni navika naglo (8,1%/g) povisuje konzumacije mesa peradi i svinjakojima je glavna kukuruz.
Očekivane klimatske promjene sa sve većim brojem sušnih/poplavnih idana s temperaturama višim od 30 °C smanjivat će prinose i otpornost teproširiti patogene organizme i mijenjati hranjivost kukuruza.
Porast potreba uz očekivane fluktuacije proizvodnje zahtijevaju hibridebolje učinkovitosti u animalnoj proizvodnji. Bez porasta boljeg iskorištenjaenergije i bioaktivnih tvari kukuruza u proizvodnji mesa i mlijeka animalnaće proizvodnja teško pratiti rastuće potrebe za animalnim proizvodima.

9
2. Upotreba kukuruza
2.1. Hrana za životinje
Kukuruz se odlikuje najboljom konzumacijom, najvišim sadržajem ene-rgije u formi škroba i ulja, linolne kiseline, karotenoida i fenola među žitari-cama. Početna hrana za mlade životinje sadržava 55-60%, a hrana za tov60-80% kukuruza koji podmiruje 60% energetskih i do 20% proteinskih po-treba mladih životinja, 75% energije i 30% proteinskih potreba svinja u tovute 75-80% energetskih i 47% proteinskih potreba junadi u tovu. Nedostacikukuruza su nizak sadržaj proteina, esencijalnih aminokiselina lizina i trip-tofana, minerala te sklonost infestacijama mikotoksigenim plijesnima.
Donedavno se smatralo da je zrno kukuruza ujednačenog sastava i hra-njivosti. Međutim, kultivari kukuruza ne razlikuju se samo po osnovnom ke-mijskom sastavu već i po drugim svojstvima koja im daju različitu hranjivuvrijednost. Značajno je različit kemijski sastav kukuruza porijeklom iz razli-čitih svjetskih regija (tablica 1.).
Nadalje, globalne razlike vidljive su i iz istraživanja u Engleskoj, gdje suuvezli kukuruz iz 15 država i hranili piliće do 28 dana hranom koja je sadr-žavala 55% kukuruza. Na kraju tova pilići, iako hranjeni smjesama istogasirovinskog sastava, postignuli su vrlo različite priraste od 680 do 1301 g ipotrošili od 1,43 do 2,67 kg hrane za kilogram prirasta. Ovi rezultati jasnopokazuju vrlo različitu hranjivost kukuruza.
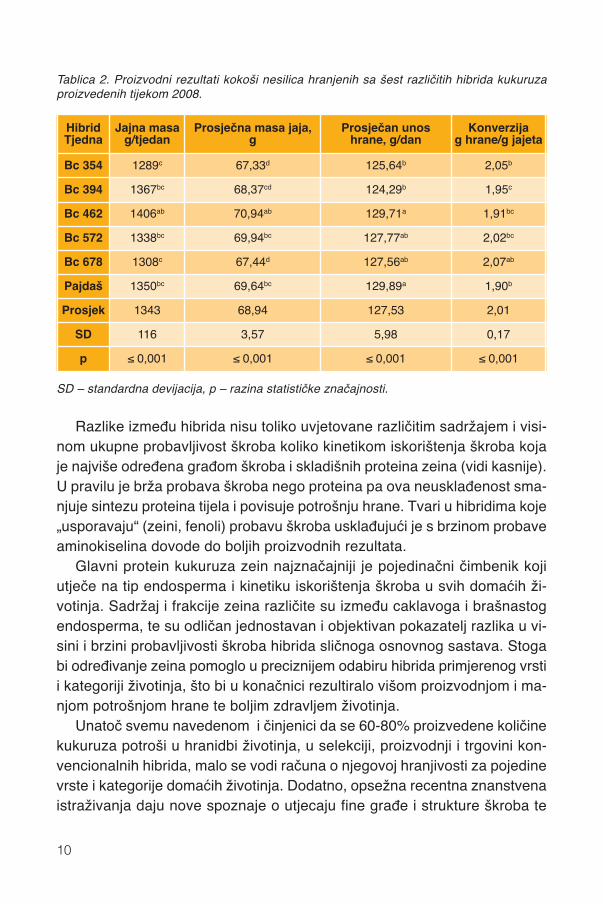
Dodatno, istraživanja pokazuju drugačiju hranjivost kukuruza, koja nepotječe samo iz različitoga osnovnoga kemijskog sastava već i kukuruzusličnoga kemijskog sastava daju vrlo raznoliku proizvodnju životinja. Spo-menuto pokazuje i naš pokus u kojemu su kokoši hranjene istim udjelomBc hibrida sličnoga osnovnoga kemijskog sastava proizvodile najveću masujaja, najkrupnija jaja i najviše pojele potpunu hranu s hibridima Bc 462, Bc572 i Pajdaš (tablica 2.)
Država Broj uzoraka Škrob Sirovi
proteinSirovamast
Sirovavlakna
SAD 367 76,1 ± 1,32 8,41 ± 0,66 3,78 ± 0,11 2,69 ± 0,16Brazil 153 74,9 ± 2,19 9,23 ± 0,70 4,07 ± 0,11 2,67 ± 0,24
Tajland 32 74,6 ± 1,55 9,06 ± 0,40 4,21 ± 0,11 2,70 ± 0,23Indija 38 74,6 ± 1,94 9,44 ± 0,64 3,94 ± 0,12 2,92 ± 0,15Svijet 650 75,5 ± 1,84 8,78 ± 0,79 3,90 ± 0,21 2,84 ± 0,22
Tablica 1. Kemijski sastav suhe tvari zrna kukuruza roda 2009.
10
Razlike između hibrida nisu toliko uvjetovane različitim sadržajem i visi-nom ukupne probavljivost škroba koliko kinetikom iskorištenja škroba kojaje najviše određena građom škroba i skladišnih proteina zeina (vidi kasnije).U pravilu je brža probava škroba nego proteina pa ova neusklađenost sma-njuje sintezu proteina tijela i povisuje potrošnju hrane. Tvari u hibridima koje„usporavaju“ (zeini, fenoli) probavu škroba usklađujući je s brzinom probaveaminokiselina dovode do boljih proizvodnih rezultata.
Glavni protein kukuruza zein najznačajniji je pojedinačni čimbenik kojiutječe na tip endosperma i kinetiku iskorištenja škroba u svih domaćih ži-votinja. Sadržaj i frakcije zeina različite su između caklavoga i brašnastogendosperma, te su odličan jednostavan i objektivan pokazatelj razlika u vi-sini i brzini probavljivosti škroba hibrida sličnoga osnovnog sastava. Stogabi određivanje zeina pomoglo u preciznijem odabiru hibrida primjerenog vrstii kategoriji životinja, što bi u konačnici rezultiralo višom proizvodnjom i ma-njom potrošnjom hrane te boljim zdravljem životinja.
Unatoč svemu navedenom i činjenici da se 60-80% proizvedene količinekukuruza potroši u hranidbi životinja, u selekciji, proizvodnji i trgovini kon-vencionalnih hibrida, malo se vodi računa o njegovoj hranjivosti za pojedinevrste i kategorije domaćih životinja. Dodatno, opsežna recentna znanstvenaistraživanja daju nove spoznaje o utjecaju fine građe i strukture škroba te
HibridTjedna
Jajna masag/tjedan
Prosječna masa jaja, g
Prosječan unos hrane, g/dan
Konverzijag hrane/g jajeta
Bc 354 1289c 67,33d 125,64b 2,05b
Bc 394 1367bc 68,37cd 124,29b 1,95c
Bc 462 1406ab 70,94ab 129,71a 1,91bc
Bc 572 1338bc 69,94bc 127,77ab 2,02bc
Bc 678 1308c 67,44d 127,56ab 2,07ab
Pajdaš 1350bc 69,64bc 129,89a 1,90b
Prosjek 1343 68,94 127,53 2,01
SD 116 3,57 5,98 0,17
p ≤ 0,001 ≤ 0,001 ≤ 0,001 ≤ 0,001
Tablica 2. Proizvodni rezultati kokoši nesilica hranjenih sa šest različitih hibrida kukuruzaproizvedenih tijekom 2008.
SD – standardna devijacija, p – razina statističke značajnosti.

11
zeinskih frakcija omotača na visinu i brzinu probavljivosti i iskorištenješkroba, ali i fitonutrijenata kukuruza na proizvodnju i zdravlje kukuruza i ži-votinja. Zbog klimatskih promjena, sve će se veća pažnja posvećivati sta-bilnosti prinosa u klimatskim ekstremnim, ali i sastojcima koji povećavajuotpornost na patogene i ublažavaju zaraze mikotoksigenim plijesnima zrnakukuruza.
Poznavanje hranjivosti hibrida kukuruza važno je za Hrvatsku koja po prih-vatljivim cijenama proizvodi svu potrebnu količinu kako kukuruza za hranuza životinje (2,100.000 t), tako i sjemena hibrida kukuruza. Hrvatska je poz-nata po proizvodnji i izvozu sjemenskog kukuruza čije su prednosti konven-cionalne metode selekcije i poznata hranjivost te sadržaj fitonutrijenata.
2.2. Bioetanol
Poznavanje ekoloških učinaka etanola važno je za proizvođače kukuruzakao jedne od glavnih sirovina za njegovu proizvodnju. Kukuruz, šećernatrska i uljarice rastu usvajanjem ugljikova dioksida iz atmosfere pa se izga-ranjem biogoriva nastalih iz tih kultura oslobađa prije „zarobljeni“ ugljikovdioksid. Osim toga, izgaranjem etanola u motoru oslobodi se 1,94, a ben-zina 2,45 kg ugljikova dioksida. Istraživanja u Njemačkoj pokazuju da se iz-garanjem litre etanola emisija CO2 smanjuje za čak 1,1 kg ili za 62% manjenego izgaranjem fosilnih goriva. Dodatno, etanol daje 2,3 puta više energijenego što se utroši u njegovu proizvodnju od polja do destilerije. Dosadašnjekorištenje etanola smanjilo je emisiju ugljikova dioksida za 100 milijuna tona.Brojne države, pa tako i Europska unija (EU) i Hrvatska, zakonski su propi-sale da benzin i nafta moraju sadržavati 5,75% bioetanola i biodizela bezobzira na njihovu cijenu. Samo u Njemačkoj, u 2015. korištenjem biogorivaproizvodnja ugljikova dioksida smanjena je za 1,5 milijuna tona.
Bioetanol (alkohol) prve generacije proizvode kvasci fermentacijom mo-nosaharida kako škroba (65-70%) iz kukuruza tako i šećera iz repe ili trske.Preradom 1 tone zrna kukuruza dobije se 420 litara etanola. Danas se sveviše koristi etanol druge generacije proizveden iz lignoceluloze i otpada, tetreće generacije gdje biodizel potječe iz lipida algi.
Svjetska proizvodnja etanola iznosi 93 milijarde litara u 2014., od togapolovinu (54,1 milijarda litara ili 58% ) proizvodi SAD, dok Europa proizvodisamo 5,5 milijardi ili 6% svjetske proizvodnje.
12
3. Građa zrna
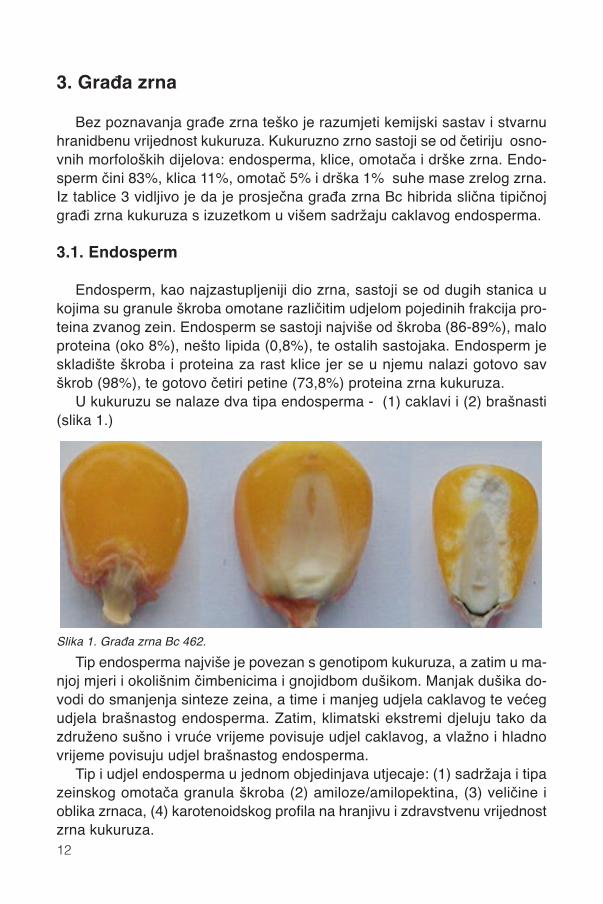
Bez poznavanja građe zrna teško je razumjeti kemijski sastav i stvarnuhranidbenu vrijednost kukuruza. Kukuruzno zrno sastoji se od četiriju osno-vnih morfoloških dijelova: endosperma, klice, omotača i drške zrna. Endo-sperm čini 83%, klica 11%, omotač 5% i drška 1% suhe mase zrelog zrna.Iz tablice 3 vidljivo je da je prosječna građa zrna Bc hibrida slična tipičnojgrađi zrna kukuruza s izuzetkom u višem sadržaju caklavog endosperma.
3.1. Endosperm
Endosperm, kao najzastupljeniji dio zrna, sastoji se od dugih stanica ukojima su granule škroba omotane različitim udjelom pojedinih frakcija pro-teina zvanog zein. Endosperm se sastoji najviše od škroba (86-89%), maloproteina (oko 8%), nešto lipida (0,8%), te ostalih sastojaka. Endosperm jeskladište škroba i proteina za rast klice jer se u njemu nalazi gotovo savškrob (98%), te gotovo četiri petine (73,8%) proteina zrna kukuruza.
U kukuruzu se nalaze dva tipa endosperma - (1) caklavi i (2) brašnasti(slika 1.)
Tip endosperma najviše je povezan s genotipom kukuruza, a zatim u ma-njoj mjeri i okolišnim čimbenicima i gnojidbom dušikom. Manjak dušika do-vodi do smanjenja sinteze zeina, a time i manjeg udjela caklavog te većegudjela brašnastog endosperma. Zatim, klimatski ekstremi djeluju tako dazdruženo sušno i vruće vrijeme povisuje udjel caklavog, a vlažno i hladnovrijeme povisuju udjel brašnastog endosperma.
Tip i udjel endosperma u jednom objedinjava utjecaje: (1) sadržaja i tipazeinskog omotača granula škroba (2) amiloze/amilopektina, (3) veličine ioblika zrnaca, (4) karotenoidskog profila na hranjivu i zdravstvenu vrijednostzrna kukuruza.
Slika 1. Građa zrna Bc 462.

13
3.1.1. Tipovi endosperma
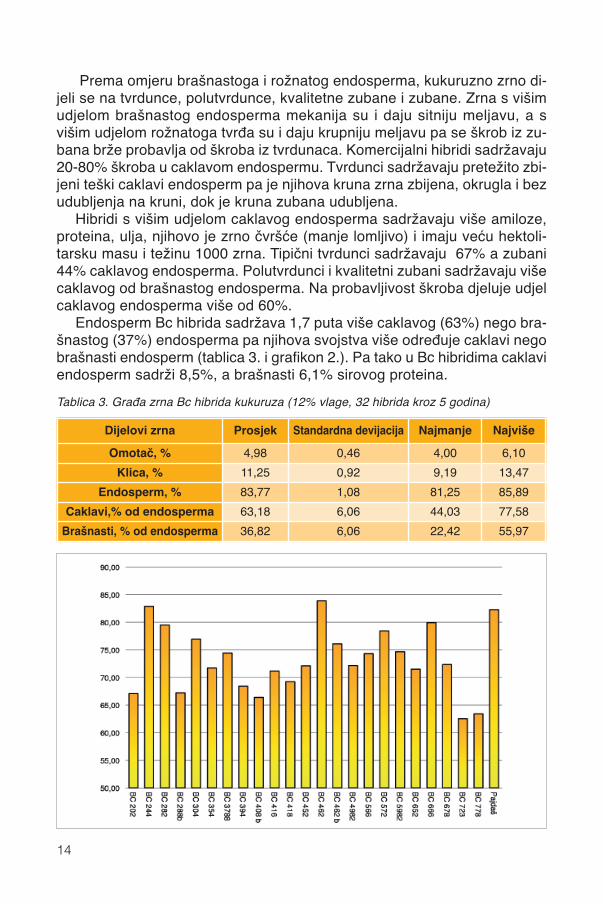
Rožnati ili caklavi endosperm je tvrd i zbijen, sadržava sitne višekutne(poligonalne) granule škroba s više amiloze koje su s višim sadržajem i dru-gačije građenim zeinom tijesno obavijene u zbijene vodootporne granule(slika 2.). Sadržaj i struktura zeina od svih pojedinačnih čimbenika najvišeutječe na tip endosperma te brzinu i probavu škroba, glavnog skladišta ene-rgije u zrnu kukuruza.
Caklavi endosperm smješten je ispod košuljice pa određuje (1) stupanjotpornosti na oštećenja insektima, plijesnima i fizičkim sredstvima, (2) odo-lijevanje stresorima tijekom ubiranja, manipulacije i skladištenja, (3) krup-noću mljevenja, (4) brzinu i mjesto probave škroba, te (5) brojna drugauporabna svojstva zrna i brašna kukuruza. Količina caklavog endospermau zrnu određena je genetski i raste sa sazrijevanjem i gnojidbom dušikom.
Caklavi endosperm sadržava više proteina zeina, amiloze (23:20%), še-ćera, karotenoida, te sitnije granule škroba od brašnastog endosperma. Me-đutim, caklavi endosperm sadržava nešto manje škroba od brašnastogendosperma. Količina caklavog endosperma u korelaciji je s razinom ukup-nih proteina, odnosno zeina i obrnuto razmjera koncentraciji glutelina. Toč-nije, udjel zeina povezan je sa caklavošću zrna (vidi poglavlje Protein).Prema nekim autorima, zubane karakterizira sadržaj zeina do 7,6% uškrobu, a tvrdunce sadržaj veći od 8,8% u škrobu.
Brašnasti endosperm je mekan i rahao, a nalazi se u unutrašnjosti zrnai obavija klicu (slika 2.). Krupne i okrugle granule brašnastog endospermaobavijene su tankom proteinskom ovojnicom i okružene zračnim džepovima.Sastoje se od više od 90% škroba te malo proteina i ulja. Brašnasti endo-sperm skloniji je lomu, njegova je hektolitarska težina manja, sadržavamanje proteina i ulja ali više škroba od rožnatog endosperma.Slika 2. Građa caklavog i brašnastog endosperma.
14
Prema omjeru brašnastoga i rožnatog endosperma, kukuruzno zrno di-jeli se na tvrdunce, polutvrdunce, kvalitetne zubane i zubane. Zrna s višimudjelom brašnastog endosperma mekanija su i daju sitniju meljavu, a svišim udjelom rožnatoga tvrđa su i daju krupniju meljavu pa se škrob iz zu-bana brže probavlja od škroba iz tvrdunaca. Komercijalni hibridi sadržavaju20-80% škroba u caklavom endospermu. Tvrdunci sadržavaju pretežito zbi-jeni teški caklavi endosperm pa je njihova kruna zrna zbijena, okrugla i bezudubljenja na kruni, dok je kruna zubana udubljena.
Hibridi s višim udjelom caklavog endosperma sadržavaju više amiloze,proteina, ulja, njihovo je zrno čvršće (manje lomljivo) i imaju veću hektoli-tarsku masu i težinu 1000 zrna. Tipični tvrdunci sadržavaju 67% a zubani44% caklavog endosperma. Polutvrdunci i kvalitetni zubani sadržavaju višecaklavog od brašnastog endosperma. Na probavljivost škroba djeluje udjelcaklavog endosperma više od 60%.
Endosperm Bc hibrida sadržava 1,7 puta više caklavog (63%) nego bra-šnastog (37%) endosperma pa njihova svojstva više određuje caklavi negobrašnasti endosperm (tablica 3. i grafikon 2.). Pa tako u Bc hibridima caklaviendosperm sadrži 8,5%, a brašnasti 6,1% sirovog proteina.
Dijelovi zrna Prosjek Standardna devijacija Najmanje NajvišeOmotač, % 4,98 0,46 4,00 6,10
Klica, % 11,25 0,92 9,19 13,47Endosperm, % 83,77 1,08 81,25 85,89
Caklavi,% od endosperma 63,18 6,06 44,03 77,58Brašnasti, % od endosperma 36,82 6,06 22,42 55,97
Tablica 3. Građa zrna Bc hibrida kukuruza (12% vlage, 32 hibrida kroz 5 godina)

15
Perad bolje iskorištava (polu)tvrdunce i kvalitetne zubane – hibride svišim udjelom caklavog nego brašnastog endosperma. Svinjama su ukusnijihibridi s višim udjelom ulja, kojeg tvrdunci često sadržavaju više od zubana.
Junad bolje iskorištava škrob hibrida s višim udjelom brašnastog negorožnatog endosperma. Naime, brašnasti se endosperm brže fermentira uburagu do propionske kiseline koja sadržava i više energije nego octena ki-selina. Prevelika količina brašnastog endosperma u obroku krava može do-vesti do jakog pada kiselosti (pH) u buragu. Kada pH padne niže od 5,5javljaju se acidoze i s njima su povezani manje uzimanje hrane i manja ma-snoća mlijeka.
3.2. Klica
Klica u pravilu čini 10-12% mase zrna kukuruza, nalazi se iznad drške isastoji se od skuteluma i embrija, koji je skladište hranjiva i hormona za em-brio. Klica je bogata uljem (33%), proteinom (18,4%), šećerom i pepelom(10,8%). Sadržava 82,1% ulja - pretežito triglicerida, 78,4% minerala i69,3% šećera kukuruza, te 26,2% ukupnog proteina zrna kukuruza. Proteinklice znatno je boljega aminokiselinskog sastava nego protein endosperma,čak je hranjiv kao animalni protein. Iz tablice 3. vidljivo je da Bc hibridi ku-kuruza sadržavaju u prosjeku 11,25% klice, a razlike između hibrida krećuse u rasponu 9,19-13,47%. Veličina klice upućuje na sadržaj kvalitetnogproteina i ulja pa što je više klice u zrnu to je viši sadržaj esencijalnih ami-nokiselina i ulja u zrnu.
3.3. Omotač
Vanjski tanki omotač, košuljica sjemena ili perikarp kukuruza čini 5%suhe mase zrna i pri mljevenju se izdvaja kao kukuruzne posije. Sastoji seod gotovo isključivo vlakana (90%) i štiti unutrašnjost zrna od mehaničkihi bioloških oštećenja.

16
4. Fizikalna svojstva
Suvremena istraživanja pokazuju kako na hranjivost ne utječu samo ke-mijska već i fizikalna svojstva zrna.
4.1. Hektolitarska masa
Hektolitarska masa najčešće se koristi za procjenu kvalitete zrna kuku-ruza. Viša hektolitarska masa povezana je s nižim sadržajem pepela, siro-vih vlakana i sirovog proteina te višom caklavosti endosperma. Izrazito niskevrijednosti hektolitarske mase (< 62 kg/hl) upućuju na niži sadržaj škroba iulja te caklavog endosperma. Hektolitarska masa je visoko nasljedno svoj-stvo (h = 0,75) pa je pokazatelj potencijalnih razlika između hibrida u ke-mijskom sastavu i sadržaju energije u zrnu. Isto tako, hektolitarska je masapokazatelj zrelosti, tvrdoće zrna i loma. Tako primjerice tvrdunci imaju hek-tolitarsku masu > 0,77 kg/L.
Međutim, klimatski ekstremi mijenjaju hektolitarsku masu. Sušno i vrućevrijeme povisuju udjel caklavog endosperma, a vlažno i hladno vrijeme po-visuje udjel brašnastog endosperma i tako smanjuje hektolitarsku masu.Nadalje, hektolitarska masa koristi se i u prometu i trgovini jer upućuje nato koliki volumen zauzima određena masa zrna, a što je način procjenemase zrna u silosima, kamionima, vagonima ili brodovima.
Zrno kukuruza velike hektolitarske mase ima visoku energetsku vrijed-nost. Pri hektolitarskoj masi od 56 do 76 kg/hl mala je razlika (13,9-14,4MJ/kg ME) u sadržaju energije te proizvodnim rezultatima u svinja.
Međutim, zrna lakša od 60 kg/hl imaju veći udjel vlaknastog omotačamanju probavljivost i energetsku vrijednost. Kokoši hranjene kukuruzom vi-soke hektolitarske mase (84 kg/hl) postignule su bolju konzumaciju, proiz-vodnju jajne mase i iskorištenje hrane od kokoši hranjenih hibridimanormalne hektolitarske mase. I u tovu goveda zrna lakša od 51,4 kg/hl sa-državaju 7% manju količinu ukupnih probavljenih hranjivih tvari za goveda.
Može se reći da je vrlo niska ili visoka hektolitarska masa dobar pokaza-telj energetske vrijednosti zrna kukuruza.
Prosječna hektolitarska masa 32 Bc hibrida kroz pet godina iznosi 76,7kg/hl što pokazuje njihovu visoku energetsku vrijednosti (tablica 4.). Utvr-

17
đena hektolitarska masa znatno je viša od minimalno potrebne (65 kg/hl)kako u Hrvatskoj tako i od prve (> 72 kg/hl) klase zrna kukuruza u SAD-u.
Kao što je već istaknuto, hektolitarska masa pokazatelj je udjela cakla-voga i brašnastog endosperma. Hibridi više hektolitarske mase sadržavajuviše caklavog, a hibridi manje hektolitarske mase više brašnastog endo-sperma. Bc 462 sadržava najviše rožnatog endosperma i stoga ima najvišuhektolitarsku masu, a Jambo, koji ima manje caklavog endosperma, imanajmanju hektolitarsku masu.
Iznimka su nezrela zrna kukuruza koja imaju manju hektolitarsku masuod zrelih a ipak imaju istu energetsku vrijednost. Istraživanja pokazuju dakilogram suhe tvari iz suhoga ili vlažnoga (siliranoga) nezrelog zrna kuku-ruza daje slične proizvodne rezultate u tovu svinja, goveda i janjadi kao isuha zrela zrna kukuruza. Pravilno provedeno siliranje košta manje od su-šenja zrna kukuruza za postizanje iste probavljivosti i proizvodnih rezultata.Zato je za hranidbu svinja i goveda na vlastitoj farmi jeftinije silirati nego su-šiti zrno kukuruza.
4.2. Težina 1000 zrna
Težina 1000 zrna dobar je pokazatelj veličine zrna, volumena i specifičnemase. Tovna junad bolje iskorištava hranjive tvari iz hibrida s većom (340 i344 g) nego s manjom (310) težinom 1000 zrna. Prema američkim istraži-vanjima, hibridi s višim udjelom caklavog endosperma teži su od hibrida sviše brašnastog endosperma. Prosječna težina 1000 zrna Bc hibrida kuku-ruza je 355 g (tablica 5.) i viša je od 324 g kolika je prosječna težina 60američkih hibrida. Teža zrna postižu manju razgradnju škroba u buragu pasu dobar pokazatelj razgradljivosti škroba.
Hektolitarskamasa, kg/hl
Prosjek Standardna devijacija
Najmanja, Jumbo
Najviša, Bc 462
76,68 2,37 71,46 80,80
Tablica 4. Hektolitarska masa (kg/hl) Bc hibrida kukuruza (12% vlage, 32 hibrida kroz 5 godina)
Perad treba hraniti hibridima više (> 78 kg/hl) hektolitarske mase.

18
4.3. Oštećenja zrna
Perikarp zrna oštećuju insekti i plijesni u polju i takva zrna mogu biti za-ražena plijesnima, odnosno sadržavati mikotoksine. Nadalje, cijelo zrnooštećuje se tijekom berbe, sušenja, transporta i premještanja. Hibridi s višeloma skloniji su kvarenju masti koje „jedu“ plijesni pa sadržavaju manje ene-rgije. Naime, lom kukuruza ima 3% ili oko 481 kJ/kg manju energetsku vri-jednost, višu koncentraciju opasnih mikotoksina od cijelog zrna kukuruza.Brazilski znanstvenici stvorili su jednadžbu pomoću koje se iz postotnogudjela pojedinih vrsta oštećenja može procijeniti visina energetskih gubi-taka. ME (MJ/kg) = 0,064 + 1,62 x napuknuta zrna + 6,98 x lom i nečistoće
+ 10,06 x pljesnivo zrnje + 12,28 x insektima oštećena zrna+ 5,87 x oštećena raznim uzrocima
Energetsku vrijednost najviše smanjuju pljesniva i insektima oštećenazrna koja su energetski prazna.
Najmanji je lom kada se zrno bere pri vlazi od 22%, a viša ili niža vlagapovisuje lom. Zrna mehanički oštećena tijekom berbe dodatno se lakšeoštećuju u kasnijim manipulacijama. Sušenje, osobito naglo, na tempera-turama višim od 80 ºC, stres je koji izaziva pucanje i lom zrna. Naime, pu-canje tijekom sušenja događa se kada brzina uklanjanja vode s površinezrna premašuje brzinu kretanja vode iz središta na površinu zrna.
Isto tako, kretanje kukuruza kroz transportne sustave uzrokuje udaranjekoje dovodi do loma zrna i stvaranja sitnih čestica koje su sklonije kvarenjui time manje hranjive vrijednosti. Karakteristike su zrna koje su povezane slomom: (1) omjer caklavog i brašnastog endosperma, (2) prosječna težinazrna, (3) kvaliteta i količina perikarpa, (4) gustoća zrna, (5) hektolitarska ilitest težina, (6) veličina i oblik zrna. Većina od ovih svojstava je nasljedna,ali selekcionari kukuruza ne pridaju veliku pažnju ovim svojstvima.
Zrna otpornija na lom imaju viši udjel caklavog a niži brašnastog endo-sperma pa je to razlog zašto se Bc 462 manje lomi od npr. hibrida Dunav.Manji lom imaju i Bc hibridi s višim sadržajem ulja i proteina kao što su novihibridi Kekec, Mejaš i Riđan.
Prosjek Standardnadevijacija Najmanja Najviša
Težina 1000 zrna, g 355,18 26,59 300,14 417,50
Volumen 1000 zrna, ml 447,43 31,54 384,04 529,75
Tablica 5. Težina i volumen 1000 zrna Bc hibrida kukuruza (12% vlage, 32 hibrida kroz 5 godina)

19
5. Kemijski sastav5.1. Vlaga
Kukuruz treba brati pri vlazi od 22%, i to radi smanjenja mehaničkih ošte-ćenja perikarpa zrna tijekom berbe i manipulacije. Viša i niža vlaga zrnočine sklonijim mehaničkim oštećenjima. Kukuruz treba skladištiti na vlazi10-15% kako bi se izbjegnulo kvarenje od strane kukaca, bakterija i plijesni.Vlaga viša od 13% omogućuje aktivnost kukaca koji stvaraju uvjete za rastplijesni koje kukuruz zagrijavaju na više od 65 °C. Nadalje, voda ne sadr-žava energiju pa svaki 1% više vlage od prosjeka smanjuje za 50 kJ/kg zrnakukuruza. Zato se krmiva uspoređuju pri istom sadržaju vlage, a u tablicamaprikazane vrijednosti preračunate su na 12% vlage. Isto tako, svakih 1%više vlage u zrnu povisuje potrošnju energije pri mljevenju, i to 7%.
Visokovlažno zrno kukuruza sa 30-35% vlage silira se ili na drugi načinkonzervira (dodavanjem ugljikova dioksida, kiselina). Vlažno zrno je pro-bavljivije te njegova suha tvar ima najmanje 5% više energije za preživačei svinje od suhe tvari osušenog zrna kukuruza.
5.2. Škrob
Najveći dio (60-80%) pojedene količine hrane životinje koriste kao izvorenergije, a škrob žitarica glavni je izvor energije za visoko proizvodne do-maće životinje. Odrasle monogastrične životinje najviše energije dobivajuiz glukoze oslobođene razlaganjem škroba u crijevima, a preživači iz hla-pljivih masnih kiselina stvorenih fermentacijom škroba u predželucima. Ku-kuruz sadržava najviše škroba(64,1%) među našim žitaricama, te 82%njegove energije potječe iz škroba. Škrob kukuruza najvažniji je (50-75%)sastojak hrane i glavni izvor energije za perad, svinje i visoko proizvodnekrave i junad u Hrvatskoj. Zbog toga hrana odraslih svinja mora sadržavatinajmanje 33% škroba, hrana dojnih krmača 42%, a visoko mliječnih krava(> 30 kg/d mlijeka) treba sadržavati 20-25% škroba. Previše škroba krozdulje razdoblje nije dobro za rasplodne životinje jer se debljaju, a u kravaviše od 30% te junadi 45% u suhoj tvari obroka može izazvati acidoze.

20
5.2.1. Sadržaj
U zrnu kukuruza najviše škroba sadržava endosperm (87,6%), a samomalo klica (8,3%) i omotač (7,3%). Međutim, iz endosperma potječe 98%škroba zrna kukuruza pa što je viši udjel endosperma to je više i škroba uzrnu.
Sadržaj škroba vrlo je ujednačen kako između svih ispitanih Bc hibrida,tako i tipa kukuruza zubana i tvrdunaca te vrlo sličan sadržaju (64%) škrobau francuskim hibridima. U pravilu, rani hibridi sadržavaju tek nešto manješkroba od kasnih hibrida. Među Bc hibridima najviše škroba sadržava Bc778, a najmanje Bc 448 (tablica 6.). Hibridi s višim sadržajem škroba pri-kladniji su za hranidbu svinja i junadi, ali i za proizvodnju etanola kao goriva.
Maksimalna probavljivost škroba visoka je i ujednačena. Varira manjenego njegov sadržaj te je po iznosu slična tabličnim vrijednostima od 97%(tablica 7.). Male razlike u sadržaju i maksimalnoj probavljivosti škroba neobjašnjavaju vrlo različite proizvodnje goveda, svinja i pilića kada se hraneistim udjelima različitih hibrida. Bolji je pokazatelj iskoristivosti škroba, kakoje utvrđeno, ne njegova probavljivost u cijelom probavnom traktu već u tan-kom crijevu u kojem se on hidrolizira do glukoze pa danas govorimo o ileal-noj probavljivosti škroba (vidi kasnije).
Sve ovo upućuje na to da razlike u građi škroba, matrici zein-škrob i ve-ličini granula uzrokuju različitu hranjivost zrna sa sličnim sadržajem i pro-bavljivosti škroba.
Škrob, %Prosjek Standardna devijacija Najmanje Bc 448 Najviša, Bc 778
63.46 1,03 61,48 65.06
Škrob, %Prosjek Standardna devijacija Najmanje Bc 224 Najviša, Bc 462
97,1 0,61 96,1 98,8
Tablica 6. Prosječan sadržaj škroba Bc hibrida kukuruza (12% vlage, 32 hibrida kroz 5 godina)
Tablica 7. Prosječna maksimalna probavljivost škroba Bc hibrida kukuruza (12% vlage, 32hibrida kroz 5 godina

21
5.2.2. Građa škroba
Građa škroba razlikuje se između hibrida te bitno djeluje na visinu i brzinuprobave škroba odnosno ravnomjernost dotoka glukoze u tvorne organe, atime i na proizvodnju životinja i iskorištenje hrane.
Čimbenici građe škroba koji utječu na probavljivost su omjer caklavi/bra-šnasti endosperm, omjer amiloza/amilopektin, udjel ukupnih i pojedinihzeina, veličina i oblik granula.
5.2.2.1. Amiloza
Škrob se sastoji od dvaju polimera vrlo različitog stupnja razgranatosti.U pravilu, normalan kukuruz sadržava 25% škroba u formi amiloze a 75%u formi amilopektina pa porast jedne smanjuje udjel druge forme škroba.Sadržaj amiloze i amilopektina (65-71%) dosta varira (19-35%), ovisno ohibridu, uvjetima rasta i stadiju zrelosti zrna.
Ravna i zbijena molekula amiloze povezana s lipidima i drugim škrobo-vima ograničava probavljivost pa je ona odgovorna za količinu sporo pro-bavljivoga i neprobavljivoga (rezistentnog) škroba. Vanjska amilaza možehidrolizirati 75-95% amiloze i gotovo sav amilopektin. U normalnim granu-lama škroba amilozna struktura smještena je iznad amilopektina te određujenjegovu probavljivost. Kako amiloza s jedne strane ograničava probavljivosta s druge je strane izvor sporo probavljivoga i rezistentnog škroba, tako pra-sad hranjena škrobom s puno amiloze (94,6%) postiže 7% manje prirastenego životinje hranjene s umjerenih 18% amiloze. Naime, koncentracijaamiloze > 35% znatno smanjuje in vitro probavljivost škroba. Međutim, pra-sad hranjena bez amiloze koja je izvor rezistentnog škroba također, dodušenesignifikantno, smanjuje priraste i iskorištenje hrane prasadi. Utvrđeno jeda umjerene koncentracije od oko 5% amiloze ili 20-24% od amilopektinapovoljno djeluje na priraste, konverziju i zdravlje peradi, svinja i nekih vrstariba. Nadalje, amiloza u debelom crijevu povisuje količinu kratkolančanihmasnih kiselina (mliječne, octene i maslačne) i smanjuje broj patogenih bak-terija što povoljno djeluje na zdravlje. Isto tako amiloza kao sastojak rezi-stentnog škroba smanjuje produkciju, a time i emisiju amonijaka iz debelogcrijeva.
Nasuprot tome amilopektin je razgranata molekula škroba, veće povr-šine, pa se potpunije i lakše probavlja enzimima životinja i mikrobima pro-bavila. Nadalje, škroba u formi amilopektina više je u manjim granulama inižeg stupnja kristalizacije ili A-tipa kristalizacije, što ga (amilopektin) doda-tno čini više i brže probavljivim u crijevima i razgradljivijim u buragu. Nasu-
22
prot tome amiloza sadržava više krupnijih i B-tipa ili C-tipa kristalizacije paje s time povezana manja probavljivost. Sadržaj amiloze u škrobu normalnihhibrida kukuruza kreće se u rasponu 22,4-32,5% a amilopektina 67,5-77,6%. U pravilu, amiloze je više u caklavom (23:20%) nego brašnastomendospermu. Nadalje, visoke temperature okoline (> 35 °C) tijekom rastasmanjuju sadržaj amiloze, i to čak 2,2-2,4%.
Hibridi s nešto višim sadržajem amiloze manje su skloni pojavi aflato-ksina, ali pružaju manju probavljivost škroba. Prema mojem mišljenju, možese smatrati dobrim onaj hibrid koji sadržava nešto više amiloze, koja musmanjuje sklonost pojavi aflatoksina, a pritom ne smanji znatno probavljivostškroba.
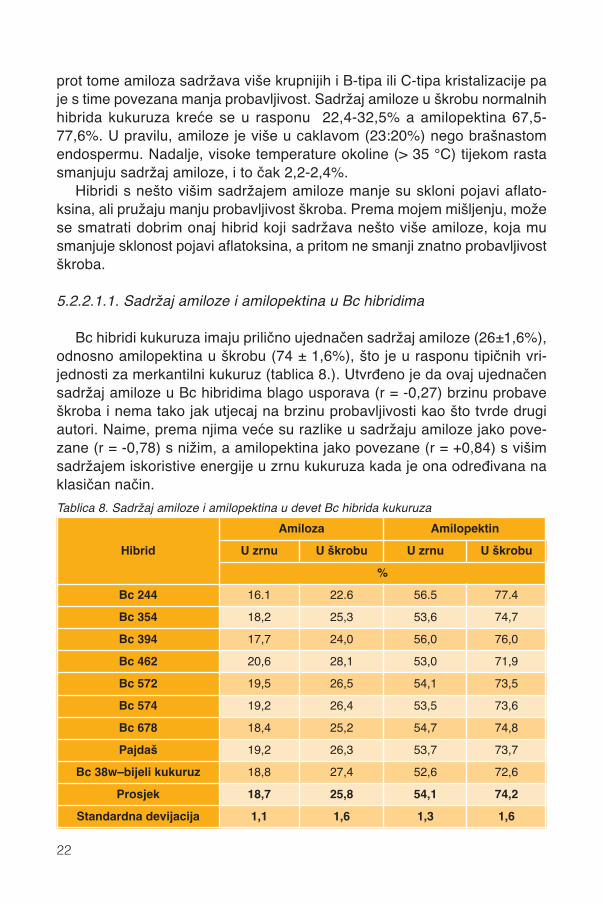
5.2.2.1.1. Sadržaj amiloze i amilopektina u Bc hibridima
Bc hibridi kukuruza imaju prilično ujednačen sadržaj amiloze (26±1,6%),odnosno amilopektina u škrobu (74 ± 1,6%), što je u rasponu tipičnih vri-jednosti za merkantilni kukuruz (tablica 8.). Utvrđeno je da ovaj ujednačensadržaj amiloze u Bc hibridima blago usporava (r = -0,27) brzinu probaveškroba i nema tako jak utjecaj na brzinu probavljivosti kao što tvrde drugiautori. Naime, prema njima veće su razlike u sadržaju amiloze jako pove-zane (r = -0,78) s nižim, a amilopektina jako povezane (r = +0,84) s višimsadržajem iskoristive energije u zrnu kukuruza kada je ona određivana naklasičan način.
HibridAmiloza Amilopektin
U zrnu U škrobu U zrnu U škrobu%
Bc 244 16.1 22.6 56.5 77.4Bc 354 18,2 25,3 53,6 74,7Bc 394 17,7 24,0 56,0 76,0Bc 462 20,6 28,1 53,0 71,9Bc 572 19,5 26,5 54,1 73,5Bc 574 19,2 26,4 53,5 73,6Bc 678 18,4 25,2 54,7 74,8Pajdaš 19,2 26,3 53,7 73,7
Bc 38w–bijeli kukuruz 18,8 27,4 52,6 72,6Prosjek 18,7 25,8 54,1 74,2
Standardna devijacija 1,1 1,6 1,3 1,6
Tablica 8. Sadržaj amiloze i amilopektina u devet Bc hibrida kukuruza

23
Sadržaj zeina i oblik granula škroba više utječu na brzinu probave škrobaBc hibrida kukuruza nego male razlike u sadržaju amiloze.
Međutim, uočeno je da su Bc hibridi s nešto više (1-3%) amiloze kao štosu Bc 462, Bc 572 i Bc 574, Pajdaš manje skloni pojavi aflatoksina.
5.2.2.2. Granule škroba
Granule škroba znatno se razlikuju po dostupnosti enzimatskoj hidrolizi.U pravilu se sitne granule potpunije i brže hidroliziraju (probavljaju) od krup-nih granula škroba. U endospermu zrna kukuruza smještene su granuleškroba „uronjene“ u proteinsku matricu koja ih omata. Kemijski gledano,granule se sastoje od amiloze i amilopektina te zeina. U gramu škroba na-lazi se oko milijardu granula škroba. Hibridi se razlikuju po veličini, obliku istupnju kristalizacije granula škroba.
Generalno, sitnije i okrugle granule sadržavaju manje amiloze i više ami-lopektina i škroba A tipa kristalizacije. Nasuprot tome, krupnije granule sa-državaju više amiloze i škroba jače ili B-tipa i C-tipa kristalizacije. Sitnijegranule više se i brže probavljaju od krupnih granula. Međutim, probavljivostgranula mijenja udjel zeina i krupnoća mljevenja, pri čemu više zeina sma-njuje a sitno mljevenje povisuje probavljivost granula. Caklavi endospermsadržava više sitnih i tijesno povezani višekutnih granula. Nasuprot tome,brašnasti endosperm karakteriziraju krupne okrugle nepovezane granuleškroba.
5.2.2.2.1. Građa granula škroba devet Bc hibrida kukuruza
Kako hibridi imaju prilično ujednačene veličine i oblike granula škroba, takosu signifikantne i male razlike između njih. Među promatranim Bc hibridimaBc 462 sadržava najsitnije i najmanje kuglaste, dok Bc 38w sadržava najkrup-nije i najviše kuglaste granule škroba (tablica 9. i slika 3.). Bc 462 s najsitnijimi nepravilnim kuglastim granulama sadržava najviše amiloze i ima sporiju pro-bavljivost, odnosno više poželjnoga sporo probavljivog škroba.
Prosjek Standardnadevijacija Najmanje Bc 462 Najviše Bc 38w
Veličina, µm 12,02 0,38 11,49 12,77
Kuglastost 1 = kugla 0,914 0,009 0,903 0,917
Tablica 9. Veličina i kuglastost granula devet Bc hibridima

24
5.2.2.3. Zeinski omotač granula škroba
Sadržaj i građa zeina prikazani su u poglavlju 7, dok je u ovom odlomkuprikazan utjecaj zeina na probavljivost škroba. Sadržaj zeina prikazuje sena više načina u ovisnosti o njegovoj promatranoj funkciji. Kako ovdje ob-jašnjavamo utjecaj zeina na probavu škroba, logično je da se njegov sadržajizražava kao udjel u škrobu.
Zein koji izvana obavija granule škroba polagano se probavlja pa sma-njuje neophodan kontakt između enzima i unutarnjeg škroba. Naša istraži-vanja pokazuju da zein više od molekularne građe škroba usporava (r =-0,46) probavu granula škroba jer ga pepsin nepotpuno hidrolizira. Osimtoga, nehidrolizirani zein smanjuje bubrenje granula što je preduvjet njihovehidrolize od strane α-amilaza u tankom crijevu ili buragu.
Među Bc hibridima kukuruza najviše zeina u odnosu na sadržaj škrobasadržava Bc 462 (tablica 10.), koji ujedino sadržava i najviše caklavog en-dosperma i ima najsporiju probavu škroba.
Zein, % od škrobaProsjek Standardna
devijacijaNajmanje Bc 38w Najviše 462
8,0 0,6 6,9 8,9
Slika 3. Granule škroba Bc 462 i Pajdaša
Tablica 10. Sadržaj zeina u % od škroba u Bc hibridima kukuruza (12% vlage)

5.2.3. Mjesto i brzina probave škroba
Kinetika probave škroba više utječe na performanse životinja nego sta-tični koeficijenti probavljivosti. Naime, za sintezu proteina mesa, mlijeka ijaja istovremeno su potrebni glukoza i aminokiseline, a škrob se brže pro-bavlja do glukoze nego protein do aminokiselina. Što je veća brzina probaveškroba od proteina to je lošije iskorištenje hrane te je više treba za jedinicuproizvoda.
5.2.3.1. Svinje i perad
Kako je već navedeno, produkt crijevne hidrolize škroba je glukoza a mi-krobne hlapljive masne kiseline koji su glavni izvor energije za intenzivnohranjene visoko proizvodne životinje. Kako je sadržaj škroba između hibridaprilično ujednačen, samo male razlike u visini, brzini i mjestu probavljivostznatno utječu na količinu energije koju će životinja dobiti iz zrna kukuruza.Probavljivost pokazuje koliki se dio (%) od pojedene količine škroba u cije-lom probavnom traktu apsorbira (upije) kao glukoza ili hlapljive masne ki-seline u organizam životinje.
Probavljivost škroba kukuruza u cijelom probavnom traktu peradi, svinjai goveda je 97%, što je gotovo potpuna probavljivost i nema većih razlikaizmeđu hibrida kukuruza. Međutim, za visoku proizvodnju mlijeka, mesa ijaja osim količine važna je istovremena i ujednačena probava škroba i pro-teina.
Potvrđuju to i istraživanja u proteklih dva desetljeća, koja pokazuju dase različita proizvodnja i zdravlje peradi te svinja ostvaruju pri istom sadržajui probavljivosti kukuruznog škroba (isti sadržaj energije). Utvrđeno je da raz-like potječu iz razlika u mjestu i brzini probave škroba. Tako pilići hranjeniistim udjelom izdvojenog škroba iz graška i kukuruza u usporedbi s onimaizdvojenim iz tapioke prirastaju više, bolje iskorištavaju hranu, energiju iaminokiseline, a imaju i više poželjnih bakterija u slijepom crijevu.
Važnost mjesta i brzine probave škroba – kinetike probave odavno supoznati u hranidbi preživača i prehrani ljudi – glikemijski indeks. Poznato jeda hrana sa sporijom i ujednačenijom hidrolizom škroba osigurava ravno-mjerniju razinu glukoze u krvi pa je ona energetski kvalitetnija, „hranjivija“za organizam. Mjesto probave škroba određuje brzina probavljivosti u cri-
25

26
jevima monogastričnih životinja pa se prema brzini probave škrob dijeli nabrzo probavljivi, sporo probavljivi i rezistentni škrob.
Danas se probavljivost škroba i proteina u svinja i peradi prikazuje kaoilealna probavljivost (%) jer se glukoza i aminokiseline apsorbiraju samo utankom crijevu, a u debelom crijevu neprobavljeni škrob i protein bakterijepretvore u spojeve koji imaju 20% manje energije nego glukoza i aminoki-selne. Mjerenjima je utvrđeno da probava škroba u tankom crijevu peradi isvinja traje do 4 sata, te se u debelom crijevu fermentira škrob koji nije bioprobavljen u tankom crijevima (slika 4.).
Osim što je važno da se što više škroba probavi u tankom crijevu, važnoje i koliko ga se probavi u pojedinim dijelovima tankog crijeva. Naime, oko75% energije održanja crijeva potroši tanko crijevo i ono ima prednost u op-skrbi energijom pred tijelom. Prvi su dio tankog crijeva duodenum (dvanaes-nik) i jejunum (vito crijevo) koji čine 65% dužine, te troši više energije negodrugi dio crijeva ili ileum (ravno crijevo) koji čini 35% dužine tankog crijeva.
5.2.3.1.1. Čimbenici brzine probave škroba
Probavljivost škroba žitarica u tankom crijevu najviše određuje sadržajvlakana, amiloze i zeina u zrnu. U pravilu, probavljivost svih tvari pa i škrobaopada s porastom količine vlakana u krmivu. Međutim, endosperm kukuruzasadržava vrlo malo vlakana(< 2%) a najveći dio škroba (98%) i proteina zrna(74%) pa probavu škroba kukuruza znatno više određuje zein od struktureškroba (amilopektin/amiloza i veličina granula). Naime, zein omata zrnca
Slika 4. Mjesta probave škroba u crijevima peradi i svinja
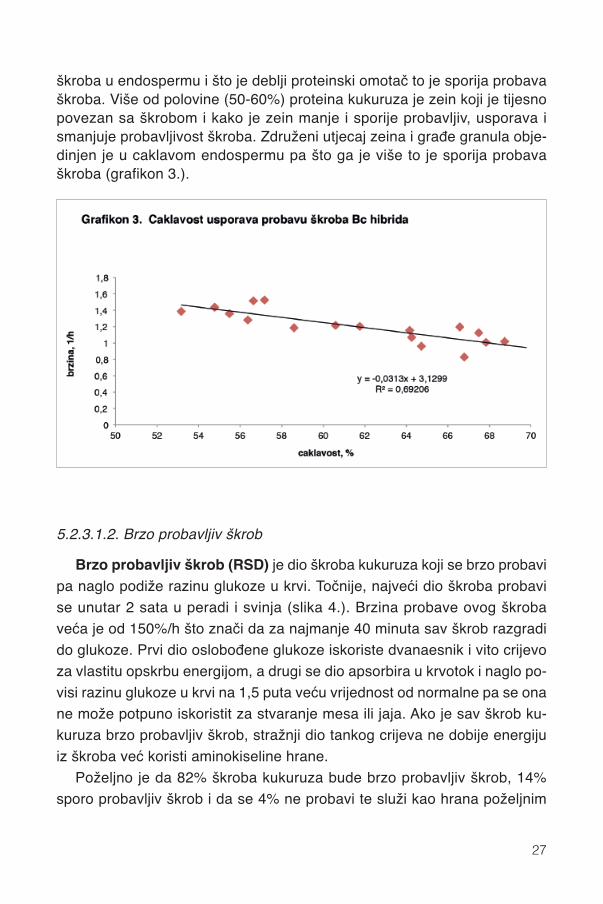
škroba u endospermu i što je deblji proteinski omotač to je sporija probavaškroba. Više od polovine (50-60%) proteina kukuruza je zein koji je tijesnopovezan sa škrobom i kako je zein manje i sporije probavljiv, usporava ismanjuje probavljivost škroba. Združeni utjecaj zeina i građe granula obje-dinjen je u caklavom endospermu pa što ga je više to je sporija probavaškroba (grafikon 3.).
5.2.3.1.2. Brzo probavljiv škrob
Brzo probavljiv škrob (RSD) je dio škroba kukuruza koji se brzo probavipa naglo podiže razinu glukoze u krvi. Točnije, najveći dio škroba probavise unutar 2 sata u peradi i svinja (slika 4.). Brzina probave ovog škrobaveća je od 150%/h što znači da za najmanje 40 minuta sav škrob razgradido glukoze. Prvi dio oslobođene glukoze iskoriste dvanaesnik i vito crijevoza vlastitu opskrbu energijom, a drugi se dio apsorbira u krvotok i naglo po-visi razinu glukoze u krvi na 1,5 puta veću vrijednost od normalne pa se onane može potpuno iskoristit za stvaranje mesa ili jaja. Ako je sav škrob ku-kuruza brzo probavljiv škrob, stražnji dio tankog crijeva ne dobije energijuiz škroba već koristi aminokiseline hrane.
Poželjno je da 82% škroba kukuruza bude brzo probavljiv škrob, 14%sporo probavljiv škrob i da se 4% ne probavi te služi kao hrana poželjnim
27

28
bakterijama u debelom crijevu peradi i svinja. Pri prosječnom udjelu od 727g škroba u kg suhe tvari u zrnu treba biti 596 g/kg brzo probavljivog škroba.
5.2.3.1.3. Sporo probavljiv škrob
Sporo probavljiv škrob (SDS) dio je škroba kukuruza koji se nakon 2sata nije probavio u prednjem dijelu tankog crijeva pa se postupno probavljau donjem dijelu tankog crijeva ili ileumu kroz 4 sata nakon uzimanja. Povoljnisu učinci sporo probavljivog škroba: (1) postupnom probavom metabolizamujednačeno dobiva glukozu pa se ona učinkovitije iskorištava u sintezi pro-teina, (2) polagana je hidroliza proteina do aminokiselina pa je potrebnaistovremena ili polagana hidroliza škroba do glukoze za njihovu ugradnju utjelesni protein tijela, (3) sporo probavljiv škrob daje energiju donjem dijelutankog crijeva pa štedi aminokiseline koje se u slučaju manjka glukoze ko-riste kao izvor energije. Naime, probavni trakt troši visokih 20% konzumi-rane energije. Perad hranjena sporo probavljivim u odnosu na brzoprobavljiv škrob ima u portalnoj cirkulaciji visokih 49,0% više lizina i 32,5%triptofana, a upravo su ove dvije aminokiseline limitirajuće u kukuruzu.
Poželjno je da krmne smjese ili potpuna brašnasta hrana sadržavaju 50g/kg umjereno probavljivog škroba, što znači da ga kukuruz koji sudjeluje uhrani sa 60% treba sadržavati (50/0,6) 83 g/kg. Istraživanjem je utvrđenoda Bc hibridi kukuruza u prosjeku sadržavaju idealnu količinu od 82 gSDS/kg suhe tvari.
5.2.3.1.4. Rezistentan škrob
Neprobavljiv ili rezistentan škrob (RS) je kao i vlakna neprobavljivnakon 4 sata probave u tankom crijevu te dospijeva u slijepo i debelo crijevogdje postaje hrana za poželjne mliječno- kisele bakterije koje njegovom fer-mentacijom proizvode kratkolančane masne kiseline, među kojima je oso-bito važna maslačna kiselina. Rezistentan škrob sastoji se od amiloze ineprobavljenog amilopektina.
U prosjeku, kukuruzno zrno sadržava 4-6% rezistentnog škroba. Prasadi pilići najbolje rastu kada potpuna hrana sadržava oko 4% rezistentnogškroba.
5.2.3.1.5. Udjeli frakcija u ukupnom škrobu Bc hibrida
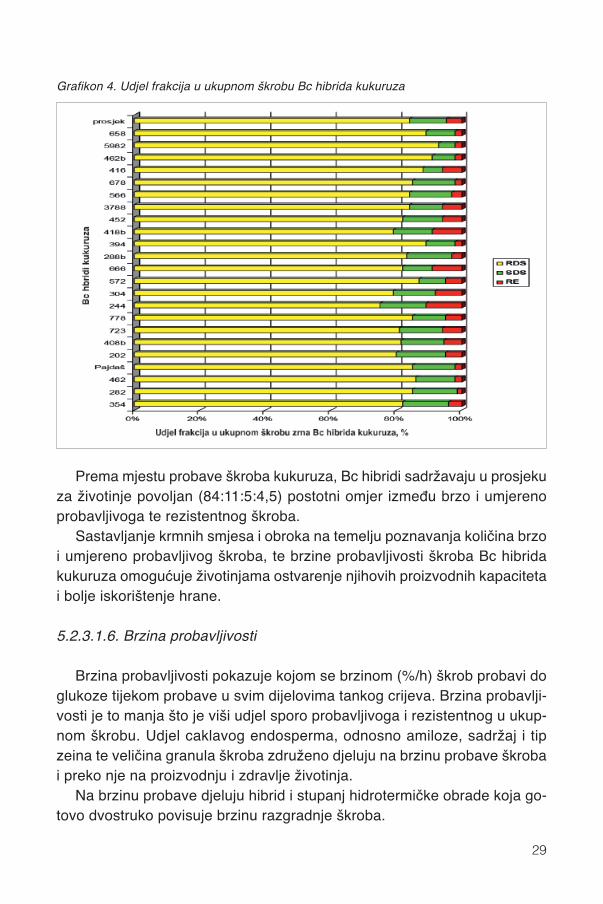
Udjeli (% od škroba) brzo probavljivog (RDS), sporo probavljivoga (SDS) irezistentnog (RS) škroba 22 Bc hibrida kukuruza navedeni su u grafikonu 4.
29
Prema mjestu probave škroba kukuruza, Bc hibridi sadržavaju u prosjekuza životinje povoljan (84:11:5:4,5) postotni omjer između brzo i umjerenoprobavljivoga te rezistentnog škroba.
Sastavljanje krmnih smjesa i obroka na temelju poznavanja količina brzoi umjereno probavljivog škroba, te brzine probavljivosti škroba Bc hibridakukuruza omogućuje životinjama ostvarenje njihovih proizvodnih kapacitetai bolje iskorištenje hrane.
5.2.3.1.6. Brzina probavljivosti
Brzina probavljivosti pokazuje kojom se brzinom (%/h) škrob probavi doglukoze tijekom probave u svim dijelovima tankog crijeva. Brzina probavlji-vosti je to manja što je viši udjel sporo probavljivoga i rezistentnog u ukup-nom škrobu. Udjel caklavog endosperma, odnosno amiloze, sadržaj i tipzeina te veličina granula škroba združeno djeluju na brzinu probave škrobai preko nje na proizvodnju i zdravlje životinja.
Na brzinu probave djeluju hibrid i stupanj hidrotermičke obrade koja go-tovo dvostruko povisuje brzinu razgradnje škroba.
Grafikon 4. Udjel frakcija u ukupnom škrobu Bc hibrida kukuruza

30
Spora brzina probavljivosti (< 70%/h) pokazuje da se škrob nije potpunoprobavio do glukoze pa životinja ne dobije svu dostupnu energiju škroba zarast te su manji prirasti. Velika brzina probavljivosti (> 150%/h) pokazujeda se škrob u potpunosti probavio, ali se u metabolizmu neučinkovito isko-risti za sintezu proteina pa su manji prirasti životinja. Optimalna ili umjerenabrzina (100-135%/h) pokazuje da se gotovo sav škrob postupno probaviodo glukoze i da je ona gotovo potpuno iskorištena za sintezu proteina mi-šića pa su viši prirasti životinja.
Jasno je utvrđena razlika u brzini probave škroba unutar sorata pšenicei unutar hibrida kukuruza. Pri istom sadržaju škroba u hibridima kukuruzamože biti različita proizvodnja životinja, i to zbog različite brzine njegoveprobave.
5.2.3.1.7. Brzina probave škroba Bc hibrida
Testirali smo brzinu probave devet Bc hibrida koji su se u pokusu poka-zali dobrima u tovu pilića.
In vitro brzina probave škroba istraživanih Bc hibrida kukuruza kreće seu rasponu 119- 146%/h, s prosjekom 135%/h, i nalazi se u tipičnom rasponuza žitarice 111-159%/h. Optimalna je probavljivost za najviši prirast i naboljeiskorištenje hrane 120-140%/h pa su s energetskog stanovišta za tov pilićadobri sljedećih devet hibrida (tablica 11.).
Životinje hranjene kukuruzom optimalne brzine probave jedu ga radije iviše, rastu brže i bolje iskorištavaju hranu. Isto tako umjerena ili optimalnabrzina probavljivosti škroba pomaže životinjama da bolje iskoriste aminoki-seline pa se one mogu hraniti s nešto manje skupih proteinskih krmiva, kao
Hibrid Bc 244 Bc 354 Bc 572 Bc 394 Bc 462 Bc 574 Bc 678 Pajdaš Bc 38w bijeli
Probavlji-vost,
%94,8 95,0 93,9 95,4 93,4 94,7 93,7 93,7 95,7
Brzina, %/h 137 127 137 130 119 140 141 141 146
Tablica 11. Brzina probave škroba odabranih Bc hibrida pilića

31
što je sojina sačma. Nadalje, male količine rezistentnog škroba dobre su zarazvoj dobrih bakterija i bolje zdravlje životinja.
5.2.3.2. Preživači
Pri hranidbi preživača kukuruzom odnosno škrobom treba imati na umu da jenjihov burag prilagođen probavi vlakana a ne škroba pa svaka nepravilnost uhranjenju velikim količinama škroba i malim količinama vlakana dovodi u opasnostproizvodnju i zdravlje životinje. Tako prevelika količina (> 30%) ukupnog i naročitofermentirajućeg škroba (> 22%), odnosno zrna žitarica (> 65%), a mala količinavlakana (< 20-23%), osobito dugih (< 3-4 cm), ili pufera (natrij bikarbonata imagnezijeva oksida), te hranjenje velikom količinom žitarica (> 3 kg) u jednomhranjenju ili gladovanje 12-24 h, te neprilagođenost buraga žitaricama, mogudovesti do pada pH i pojave acidoze i s njom povezanih poremetnji. Kako jeopasnost od acidoze veća u krava nego u junadi u tovu, one trebaju u obrokuimati najmanje 30% a junad najmanje 10% voluminozne krme, najbolje sijenalucerne.
Škrob je u završnom tovu junadi prvi (40%), a u visoko mliječnih kravadrugi (25-30%) najzastupljeniji sastojak obroka. Škrob je glavni i najbogatijiizvor energije za mikrobe buraga i kravu, te njegovom fermentacijom nastalapropionska kiselina za sintezu mliječnog šećera. Zbog svega navedenog,škrob mora imati svojstva koja mu omogućuju da podmiri potrebe mikrobaburaga (razgradnja) i potrebe krava.
Glavno je mjesto (80-90%) probave škroba i apsorpcije glukoze u peradi isvinja gornji dio tankog crijeva (duodenum i ileum), u preživača su to burag(60%) i tanko crijevo (30%), te debelo crijevo (10%). Probavljivost škroba ucijelom probavnom traktu krava znatno varira (91% ± 7,4%), a uzrokovana jerazličitom građom zrna hibrida (brašnasti > caklavi), konzerviranjem (visokovlažno > suho zrno), preradom (mljeven > flekičan), te mljevenjem (sitan >krupno mljeveni).
Pojedeni škrob prvo razgrade mikroorganizmi u buragu preživača za vla-stiti rast, a produkte mikrobne razgradnje – hlapljive masne kiseline, osobitopropionsku kiselinu, i mikrobni protein koristi preživač kao izvor energije i gra-đevnih tvari. Tako se po svakom kilogramu škroba koji probave – fermentirajumikrobi buraga krava dobije 540 g hlapljivih masnih kiselina i 150 g mikrobnogproteina ili 96 g aminokiselina. Fermentacijom škroba nastane veća masahlapljivih masnih kiselina i mikroba nego fermentacijom vlakana pa životinjehranjene s više škroba dobiju više hlapljivih masnih kiselina (energije) i ami-nokiselina, odnosno postižu više priraste i višu mliječnost. Preciznije, preko

32
sinteze mikrobnog proteina goveda dobiju više aminokiselina nego iz poje-dene hrane pa škrob umjereno povisuje sadržaj proteina u mlijeku i prirastu.Zato se visok proizvodne krave i junad hrane obrokom s visokim udjelomškroba koji mogu pojesti mikrobi. Primjerice, analizom 140 pokusa u SAD-uutvrđeno je da prosječna krava koja pojede 20 kg suhe tvari/d i u njoj 25%škroba ili 5,4 kg/d, mikrobi buraga „pojedu“, odnosno razgrade najviše (60%)ili 3,14 kg, u tankom crijevu se probavi 32% ili 1,65 kg, dok se ostatak raz-gradi u debelom crijevu.
5.2.3.2.1. Razgradljivi škrob
Dio škroba koji se razgradi u buragu naziva se razgradljivi škrob, a dioškroba koji ne razgrade mikrobi buraga, ali se probavi u crijevima, naziva senerazgradljivi ili rezistentni škrob. Za maksimalnu sintezu mikrobnog proteinabitna je optimalna fermentacija u buragu koja se postiže kada obrok sadržava12-16% fermentirajućeg škroba. Visoki udjel fermentirajućeg škroba smanjujefermentaciju vlakana i dovodi u opasnost zdravlje buraga i životinje, dok niskakoncentracija smanjuje moguću sintezu mikrobnog proteina. Acidoze u pro-sjeku smanjuju mliječnost za 3 kg/d, masnoću na niskih 2,9%, a sadržaj pro-teina na 3,4% te opasno povisuju učestalosti acidoze (kiselost manja od5,5-5) i posljedično laminitisa (šepavost) životinja. Nadalje, tovna junad hra-njena visokim udjelom škroba u obroku ima niski pH-sadržaja buraga i crijevašto tijekom čekanja životinja za klanje povisuje rizik od pojave krvarenja uzro-kovanog bakterijom Escherichiom colli koja je opasna za ljude, konzumentejunećeg mesa.
Optimalna koncentracija fermentirajućeg škroba je između najviših 22% inajmanjih 16% u visoko mliječnih krava. Visoke energetske potrebe visokomliječnih krava koje nisu podmirene iz dopuštene koncentracije fermentira-jućeg škroba a do dopuštene količine ukupnog škroba 25-30%, podmiruju seiz rezistentnog škroba.
U preživača je rezistentan škrob neprobavljiv u buragu, a za razliku od pe-radi i svinja probavljiv je u crijevima. Dodatno, taj se škrob ne fermentira pahidrolizom u crijevima daje glukozu koja sadržava 1,3 puta više energije odhlapljivih kiselina i glavna je sirovina za sintezu mliječnog šećera. Naime, ge-netski je određena sinteza mliječnog šećera pa nedostatak glukoze za njezinusintezu dovodi do manje mliječnosti i moguće pojave ketoze mliječnih krava.Njemački i nizozemski normativi preporučuju da obrok krava sadržava naj-manje 5% nefermentirajućega ili rezistentnog škroba u suhoj tvari. Najbolji je

33
izvor rezistentnog škroba suho zrno kukuruza, osobito tip (polu)tvrdunaca isirak. Nasuprot tome, malo rezistentnog škroba sadržavaju sitnozrne žitaricei silaža kukuruza.
5.2.3.2.1.1. Razgradljivost škroba kukuruza
Razgradnja škroba u buragu znatno (41-78%) se razlikuje između (1)hibrida kukuruza (2) uvjeta uzgoja i (3) načina prerade. Tako je 61%razgradnja škroba standardnog kukuruza sa 60% caklavog endosperma, tekod kukuruza bez caklavog endosperma i sa samo 2,5% zeina/škrobu čak92%. Ovako visoka razgradnja (92%) škroba smanjuje probavljivost vlakana(NDF) sa 60 na 50% i potencijalno uzrokovati acidoze i s njima povezaneporemetnje.
Kada se gleda hibrid tada je najveća razlika između tipičnih zubana itvrdunaca. Razgradnja škroba tipičnih zubana 5% je viša nego kod polutvrdunacai kvalitetnih zubana (61:66%). Među svojstvima hibrida, caklavost zrna kukuruzanajznačajnije djeluje na razgradnju škroba (r = 0,86) i suhe tvari (r = 0,82) uburagu. Naime, zeini su slabo i sporo topljivi u blago kiselom (pH 6,2-6,5)buragu pa oni kao omotač granula najviše ograničavaju razgradnju škroba uburagu. Jačina povezanosti zeinske matrice sa zrncima škroba te tip i udjelzeina u endospermu snažno djeluju na razgradnju škroba pa tako viši sadržajzeina u hibridima s više caklavog endosperma znatno (r = -0,87) smanjujerazgradnju škroba. Sazrijevanjem zrna raste caklavost, opada udjel vode igranule škroba postaju sve zbijenije, što je povezano s porastom koncentracijenetopljivih zeina u matrici koja obavijanjem granula škroba čini kemijskuzapreku mikrobiološkoj razgradnji u buragu. Može se reći da je uloga zeinazaštitna u sprečavanju razaranja škrobnih zrnaca, kao što vlakna igraju uloguu zaštiti cjelovitosti zrna. Dugotrajnim stajanjem silaže otapa se ili razgrađujezeinski omotač pa je znatno viša razgradnja siliranog nego suhog zrnakukuruza.
Veća je razlika u razgradnji škroba između sitno i krupno mljevenog zrna(15%) nego između hibrida kukuruza (5%). Sitnije (0,7 mm) u usporedbi skrupnim (3,5 mm) mljevenjem povisuje razgradnju škroba polutvrdunaca sa50 na 62%, a zubana sa 50 na 65%. Sitnijim mljevenjem povisuje serazgradnja u buragu i posljedično viša proizvodnja propionske kiseline isinteza mikrobnog proteina. Krupnije mljevenje smanjuje razgradnju u buragui povisuje dotok škroba u crijeva pa posljedično u metabolizam ulazi više

34
energijom bogatija glukoza nego propionska kiselina. Isto tako krupnijemljevenje sprečava pojavu acidoza, nadama i laminitisa.
Prosječna razgradnja škroba suhog zrna kukuruza u buragu je 60% odškroba.
Mikrobi buraga najbolje probavljaju hranu i rastu kada je visina razgradnješkroba slična visini razgradnje proteina u buragu. Prema američkim istraži-vanjima, optimalna razgradnja proteina u buragu je 60-65%. Kako je vidljivoiz tablice 12, prosječna razgradnja škroba suhog zrna Bc hibrida u buragudovoljno je visoka za maksimalnu mikrobnu sintezu i dovoljno niska da spriječipojavu acidoza u pravilnoj hranidbi. Razgradnja škroba Bc hibrida slična jefrancuskim (60%) te znatno manja nego kod (74%) američkih hibrida kuku-ruza, a razlog je viši sadržaj caklavog endosperma u našim nego američkimhibridima kukuruza.
Preporučuje se krave hraniti Bc polutvrduncima/kvalitetnim zubanima napočetku laktacije i kada obrok sadržava dosta razgradljivog škroba iz silažekukuruza, silaže zrna i/ili sitozrnim žitaricama. Kada obrok sadržava nedo-voljno razgradljivog škroba tada se krave hrane zrnom Bc hibrida u tipu zu-bana. Isto tako, junad u fazi prilagodbe ili početku tova treba hraniti suhimzrnom polutvrdunaca/kvalitetni zubana, a u završnoj fazi tova Bc hibridima svišim udjelom brašnastog endosperma.
5.2.3.3. Kunići
Probavni sustav kunića prilagodio se iskorištenju hranjivih tvari iz volu-minozne, a ne koncentratne krme kao što je kukuruz. Visoko škrobni obroknepotpuno se probavi u tankom crijevu pa ga dosta dospije u slijepo crijevogdje omogućuje iznimno brz rast mikroba. Ako se tamo nalaze mikrobi kojiproizvode toksine, prvenstveno Clostridium siroforme, velika količina škrobadovodi do pojave enteritisa i moguće visoke smrtnosti kunića. Hrana mladihkunića može sadržavati do 13, a tovnih do 18% škroba.
U hranidbi kunića trebamo koristiti hibride kukuruza s puno brašnastogai malo rezistentnog škroba, a zrno se ne smije fino samljeti.
Razgradljivi škrob u buragu, %
Prosjek Standardna devijacija
Najmanja Bc 462
Najviša Bc 778
60.15 4.61 49.20 74.72
Tablica 12. Visina razgradnje škroba (%ukupnog škroba) Bc hibrida kukuruza u buragu (12%vlage, 32 hibrid kroz 5 godina)

6. ŠećeriZrna Bc hibrida sadržavaju u prosjeku 1,5% šećera u 88% suhe tvari i
nema bitne razlike (+/-0,5%) između hibrida namijenjenih za hranidbu živo-tinja. Sastoje se od saharoze, glukoze i fruktoze, brzo i potpuno su probav-ljivi, a prema dosadašnjim spoznajama pospješuju uzimanje hrane u svinja,kako fermentaciju silaža tako i suhe tvari u buragu, i nemaju bitan utjecajna proizvodnju pilića u tovu. Iako šećer sadržava manje energije od škrobapa svaki 1% više šećera smanjuje sadržaj energije on povisuje konzumacijuhrane u svinja i goveda te omogućuje lakše siliranje zrna. Viši sadržaj še-ćera od prosjeka sadržavaju Bc 306 (3,3%), Kekec (2,7%) Riđan (2,8%).
35

36
7. Protein
Životinje trebaju protein hrane kao izvor aminokiselina iz kojih one gradei obnavljaju vlastiti protein. Zanimljivo je da bezmasno tijelo svih životinja usvim stadijima života sadržava stalnih 20-22% proteina. Mladunci brzo rastui jedu manje hrane nego odrasle životinje pa njihova hrana treba sadržavativiše proteina. Tako mladunci trebaju najmanje 20% proteina, tovne životinje14-18%, a za odrasle bređe svinje i krave najmanje 12% proteina u zrako-suhoj tvari obroka.
Svi raznovrsni proteini životinje sastoje se od istih aminokiselina, a raz-likuju se jedino po koncentraciji pojedine aminokiseline. Prva je funkcija pro-teina hrane da budu izvor esencijalnih aminokiselina (nezamjenjive ilinenadoknadive) koje životinja treba za sintezu vlastitih proteina a ne možeih sintetizirati. Životinja treba 9-10 bitnih od ukupno 22 aminokiseline u pro-teinu. Esencijalne su aminokiseline lizin, metionin, triptofan, tireonin izoleu-cin, histidin, fenilalanin, arginin i valin.
Druga je funkcija proteina osigurati dovoljno dušika i ugljika za sintezunebitnih aminokiselina. Nebitne – neesencijalne (zamjenjive ili nadoknadive)aminokiseline životinja može sintetizirati jednu iz druge, a poluesencijalneaminokiseline samo iz esencijalnih aminokiselina (cistin iz metionina, tiroziniz fenilalanina) ili su potrebne samo u određenom razdoblju života pojedinimkategorijama životinja (glutamin, arginin, taurin).
Treća je funkcija proteina održanje zdravlja. Probavom zeina oslobađajuse peptidi koji vežu žučne kiseline i smanjuju razinu kolesterola u krvi ljudi.Isto tako koncentrat peptida kukuruza smanjuje hipertenziju u ljudi.
7.1. Protein kukuruza
Kukuruz sadržava 8-9% proteina, od čega je 80-85% smješteno u endo-spermu i 20-25% u klici. Međutim, klica sadržava visokih 30-35% proteinavisoke biološke vrijednosti, a endosperm 9% proteina lošega aminokiselin-skog sastava. Kako klica čini tek 15-20% a endosperm 80-85% mase zrna,protein endosperma čini 75% proteina zrna. Najveći dio (70%) proteina ku-kuruza čini zein koji je siromašan bitnim aminokiselinama, osobito lizinom itriptofanom. Međutim, kako potpuna hrana sadržava najviše (oko 60%) ku-kuruza, tako on podmiruje dobar dio (20-47%) proteinskih potreba domaćihživotinja.

37
U kukuruzu najvišu koncentraciju protein sadržava klica (17-20%), osre-dnju endosperm (8-9%) i najmanju perikarp (4-6%).
U endospermu je skladište prolamina (zein) i gultelina. Dvostruko je većiudjel zeina nego glutelina u proteinu endosperma zrna kukuruza.
Protein klice (glutelin) je boljeg aminokiselinskog sastava od proteina en-dosperma (zein), ali kako je endosperm najzastupljenija komponenta u zrnu,iz njega potječe 75% a iz klice 25% proteina zrna. Zein se dvostruko sporijeprobavlja (0,027:0,06%/h) od ostalih frakcija proteina kukuruza pa kao omo-tač granula smanjuje i usporava probavu i razgradnju škroba.
Glavni protein kukuruza zein više utječe na iskoristivost škroba nego naaminokiselinsku opskrbu životinja.
7.2. Sadržaj
Varijabilniji je sadržaj proteina od drugih tvari u zrnu kukuruza (koeficijentvarijacije 8,7%) te su razlike određene genotipom, godinom proizvodnje,gnojidbom dušikom, agrotehnikom i klimom.
Iz tablice 13 vidi se da Bc hibridi u prosjeku sadržavaju nešto više pro-teina (8,37%) nego francuski (8,10%) i američki (8,27%). Iako je kukuruzsiromašan proteinom, zbog visokog prinosa i zbog visokog udjela u hranion je najjeftiniji izvor nebitnih aminokiselina jer podmiruje 20% proteinskihpotreba peradi, 30% potreba svinja i gotovo polovinu (50%) potreba junadiu tovu.
Na razlike u sadržaju proteina nisu djelovali okolišni čimbenici jer su istra-živani Bc hibridi proizvedeni primjenom slične umjereno intenzivne agroteh-nike i rasli su u sličnim agroekološkim uvjetima (tablica 13.). Viši sadržajproteina u zrnu povezan je s višim udjelom klice od endosperma, višim udje-lom rožnatog od brašnastog endosperma i drugim manje važnim razlikamaizmeđu endosperma i klice. (Polu)tvrdunci sadržavaju više proteina jer suu njima male granule škroba okružene proteinskim omotačem i još sadrža-
Sirovi protein, % Prosjek Standardna
devijacijaNajmanje
Bc 778Najviše Bc 462
Sadržaj 8,37 0,62 7,26 9,86Probavljivost,
% 83,54 0,91 Najmanje Najviše 78,78 85,80
Tablica 13. Sadržaj i probavljivost sirovog proteina u Bc hibridima kukuruza (12% vlage, 32hibrida kroz 5 godina)

38
vaju oko 4% vrlo gustog perikarpa koji je bogat proteinom (27,7%). Hibridi s višim sadržajem proteina dobar su i jeftin izvor nebitnih amino-
kiselina za sve vrste i kategorije životinja kada se primjereno nadopune so-jinom sačmom ili drugim izvorom esencijalnih aminokiselina (lizina,triptofana i metionina). Među Bc hibridima kukuruza visok sadržaj sirovogproteina (% u 88% suhe tvari) sadržavaju pored Bc 462, Mejaš (9,4), Kekec(9,4) i Riđan.
7.3. Zeini
Zein je najzastupljenija proteinska frakcija kukuruza koja ima brojne bio-loške uloge, pa tako određuje njegov aminokiselinski sastav, tip endo-sperma, brzinu i visinu probave i razgradnje škroba i osobito sadržajmetaboličke energije. Sadržaj zeina određen je genotipom i okolišnim čim-benicima. Manjak dušika dovodi do manje sinteze zeina i manje caklavog,odnosno više brašnastog endosperma.
Sadržaj zeina bolji je pokazatelj tvrdoće zrna i probavljivosti škroba odsadržaja caklavog endosperma u zrnu, a pritom se jednostavnije i preciznijeodređuje od caklavosti endosperma. Udjel zeina u suhoj tvari veći je kodhibrida s višim udjelom caklavog endosperma i amiloze (tablica 14.). Sadr-žaj zeina, osim vlažnosti, krupnoće mljevenja i hidrotermičke prerade imavelik utjecaj na probavljivost suhog zrna i mali na probavljivost škroba u od-stajaloj silaži zrna i biljke kukuruza.
Zbog brojnih raznovrsnih uloga, sadržaj zeina prikazuje se na više načinau ovisnosti o njegovoj promatranoj funkciji pa se tako prikazuje kao udjel u:(1) zrnu, (2) suhoj tvari zrna, (3) proteinu i (3) škrobu.
Prema nekim autorima, zubane karakterizira niži sadržaj zeina, do 7,6%u škrobu, a tvrdunce viši sadržaj, > 8,8%, zeina u škrobu. Zatim, više jezeina u vanjskim nego unutarnjim dijelovima endosperma.
U zrnu kukuruza nalaze se četiri glavne frakcije zeina (α, β, γ i δ), odkojih svaku sintetiziraju drugi geni i svaka ima specifičnu ulogu u mikrostruk-turi endosperma a time i u iskorištenju škroba (slika 5.). α-zein je najzastu-pljeniji (70%) zein koji obavija granule škroba zajedno s δ -zeinom. Nadalje,β i γ zeini nalaze se oko proteinskih tjelešaca prvih dvaju zeina. Gama (γ)zein je povezan sa caklavosti zrna i nižom probavljivošću škroba.
Sadržaj zeina u šest Bc hibrida kukuruza snažno usporava brzinu pro-bave škroba (r = -0,70) i povisuje (r = 0,81) metaboličku energiju za peradpa se pri sličnom kemijskom sastavu može koristiti kao razlikovni pretkazi-vač kvalitete kukuruza za perad.
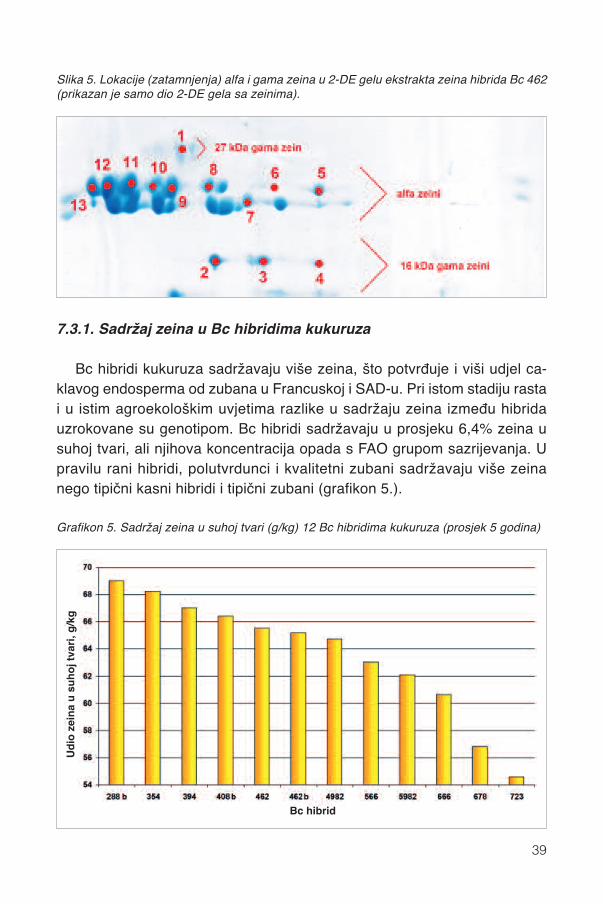
7.3.1. Sadržaj zeina u Bc hibridima kukuruza
Bc hibridi kukuruza sadržavaju više zeina, što potvrđuje i viši udjel ca-klavog endosperma od zubana u Francuskoj i SAD-u. Pri istom stadiju rastai u istim agroekološkim uvjetima razlike u sadržaju zeina između hibridauzrokovane su genotipom. Bc hibridi sadržavaju u prosjeku 6,4% zeina usuhoj tvari, ali njihova koncentracija opada s FAO grupom sazrijevanja. Upravilu rani hibridi, polutvrdunci i kvalitetni zubani sadržavaju više zeinanego tipični kasni hibridi i tipični zubani (grafikon 5.).
39
Slika 5. Lokacije (zatamnjenja) alfa i gama zeina u 2-DE gelu ekstrakta zeina hibrida Bc 462(prikazan je samo dio 2-DE gela sa zeinima).
Grafikon 5. Sadržaj zeina u suhoj tvari (g/kg) 12 Bc hibridima kukuruza (prosjek 5 godina)
Udio
zein
a u
suho
j tva
ri, g
/kg
Bc hibrid

40
Zein djeluje na tvrdoću zrna pa Bc hibridi s višim sadržajem zeina imajumanje loma i mikotoksina te su zdraviji. Umjereni udjel zeina u proteinu (48-68%) suhog zrna omogućuje ravnomjernu probavu hrane i bolje iskorištenješkroba, preveliki (> 68%) smanjuje energetsku vrijednost hrane, a premali(<35%) dovodi do lošeg iskorištenja škroba i opasnosti od acidoza u preži-vača.
Finim (< 800 μm) mljevenjem i flekičanjem suhog zrna s povišenim sa-držajem zeina neutralizira se njihov negativan učinak na probavljivostškroba. Probavljivost škroba silaže zrna i cijele biljke kukuruza ovisi nesamo o sadržaju zeina već i o krupnoći i vlazi silaže. Tako probavljivostškroba silaža raste s porastom vlažnosti (r = 0,53), finoće mljevenja (česticemanje od 4,75 mm) (r = 0,70), a opada sa sadržajem zeina ( r = - 0,47).Stajanjem silaže (> 4-6 mjeseci) smanjuje se sadržaj zeina i raste probav-ljivost škroba u buragu pa je veća opasnost od acidoza u krava i junadi utovu u proljeće i ljeto kada se hrane starom silažom nego u jesen kada sehrane mladom silažom (vidi kasnije).
Sadržaj zeina viši je u caklavom nego brašnastom endospermu, a rastesa sazrijevanjem biljke i jačinom suše tijekom rasta kukuruza. Kako su go-dine sve češće sušne, odredili smo sadržaj zeina u 12 Bc hibrida kukuruzaproizvedenih istom agrotehnikom u istim agroekološkim uvjetima u Rugvicitijekom pet godina (grafikon 6.).
Grafikon 6 pokazuje da je najviša koncentracija zeina u suhoj tvari u su-šnoj 2003. bila čak 15% viša nego u sljedeće četiri godine. Zaključujemoda će suše sve više utjecati ne samo na prinos već i na hranjivost kukuruza.
% udjel caklavog endosperma
% udjel brašnastog
endosperma
Amiloza, % udjel u
zrnuAmilopektin,
% udjel u zrnuBrzina
probave 1/h
0,50 -0,57 0,68 -0,56 -0,63
Tablica 14. Povezanost sadržaja zeina u suhoj tvari s građom zrna i brzinom in vitro probave6 Bc hibrida kukuruza
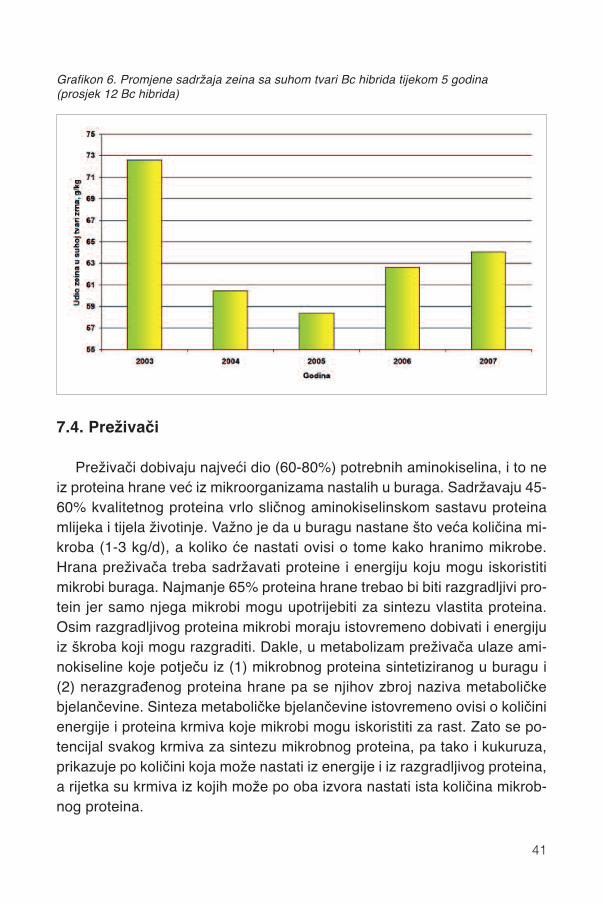
41
7.4. Preživači
Preživači dobivaju najveći dio (60-80%) potrebnih aminokiselina, i to neiz proteina hrane već iz mikroorganizama nastalih u buraga. Sadržavaju 45-60% kvalitetnog proteina vrlo sličnog aminokiselinskom sastavu proteinamlijeka i tijela životinje. Važno je da u buragu nastane što veća količina mi-kroba (1-3 kg/d), a koliko će nastati ovisi o tome kako hranimo mikrobe.Hrana preživača treba sadržavati proteine i energiju koju mogu iskoristitimikrobi buraga. Najmanje 65% proteina hrane trebao bi biti razgradljivi pro-tein jer samo njega mikrobi mogu upotrijebiti za sintezu vlastita proteina.Osim razgradljivog proteina mikrobi moraju istovremeno dobivati i energijuiz škroba koji mogu razgraditi. Dakle, u metabolizam preživača ulaze ami-nokiseline koje potječu iz (1) mikrobnog proteina sintetiziranog u buragu i(2) nerazgrađenog proteina hrane pa se njihov zbroj naziva metaboličkebjelančevine. Sinteza metaboličke bjelančevine istovremeno ovisi o količinienergije i proteina krmiva koje mikrobi mogu iskoristiti za rast. Zato se po-tencijal svakog krmiva za sintezu mikrobnog proteina, pa tako i kukuruza,prikazuje po količini koja može nastati iz energije i iz razgradljivog proteina,a rijetka su krmiva iz kojih može po oba izvora nastati ista količina mikrob-nog proteina.
Grafikon 6. Promjene sadržaja zeina sa suhom tvari Bc hibrida tijekom 5 godina(prosjek 12 Bc hibrida)
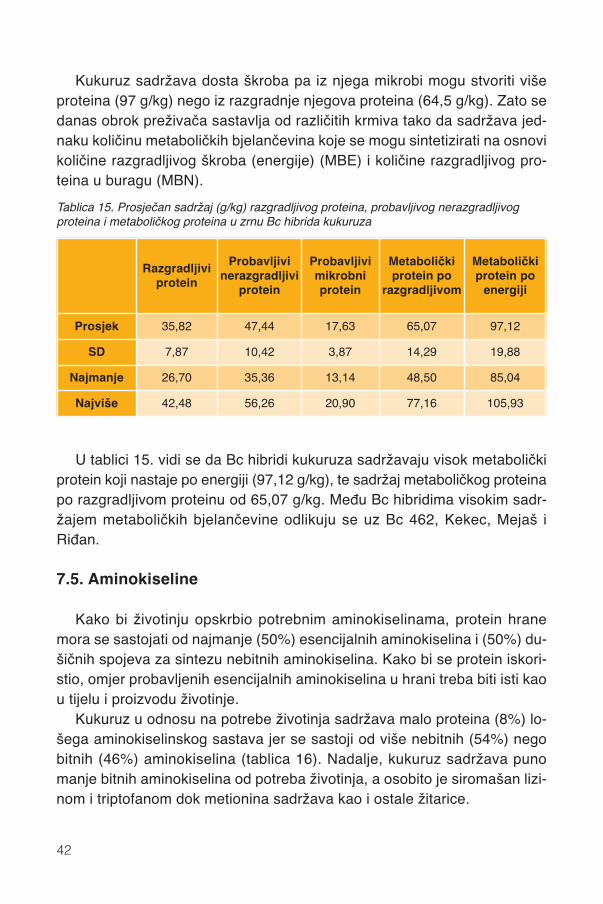
42
Kukuruz sadržava dosta škroba pa iz njega mikrobi mogu stvoriti višeproteina (97 g/kg) nego iz razgradnje njegova proteina (64,5 g/kg). Zato sedanas obrok preživača sastavlja od različitih krmiva tako da sadržava jed-naku količinu metaboličkih bjelančevina koje se mogu sintetizirati na osnovikoličine razgradljivog škroba (energije) (MBE) i količine razgradljivog pro-teina u buragu (MBN).
U tablici 15. vidi se da Bc hibridi kukuruza sadržavaju visok metaboličkiprotein koji nastaje po energiji (97,12 g/kg), te sadržaj metaboličkog proteinapo razgradljivom proteinu od 65,07 g/kg. Među Bc hibridima visokim sadr-žajem metaboličkih bjelančevine odlikuju se uz Bc 462, Kekec, Mejaš iRiđan.
7.5. Aminokiseline
Kako bi životinju opskrbio potrebnim aminokiselinama, protein hranemora se sastojati od najmanje (50%) esencijalnih aminokiselina i (50%) du-šičnih spojeva za sintezu nebitnih aminokiselina. Kako bi se protein iskori-stio, omjer probavljenih esencijalnih aminokiselina u hrani treba biti isti kaou tijelu i proizvodu životinje.
Kukuruz u odnosu na potrebe životinja sadržava malo proteina (8%) lo-šega aminokiselinskog sastava jer se sastoji od više nebitnih (54%) negobitnih (46%) aminokiselina (tablica 16). Nadalje, kukuruz sadržava punomanje bitnih aminokiselina od potreba životinja, a osobito je siromašan lizi-nom i triptofanom dok metionina sadržava kao i ostale žitarice.
Razgradljiviprotein
Probavljivi nerazgradljivi
protein
Probavljivimikrobni protein
Metaboličkiprotein po
razgradljivom
Metaboličkiprotein po
energiji
Prosjek 35,82 47,44 17,63 65,07 97,12
SD 7,87 10,42 3,87 14,29 19,88
Najmanje 26,70 35,36 13,14 48,50 85,04
Najviše 42,48 56,26 20,90 77,16 105,93
Tablica 15. Prosječan sadržaj (g/kg) razgradljivog proteina, probavljivog nerazgradljivog proteina i metaboličkog proteina u zrnu Bc hibrida kukuruza
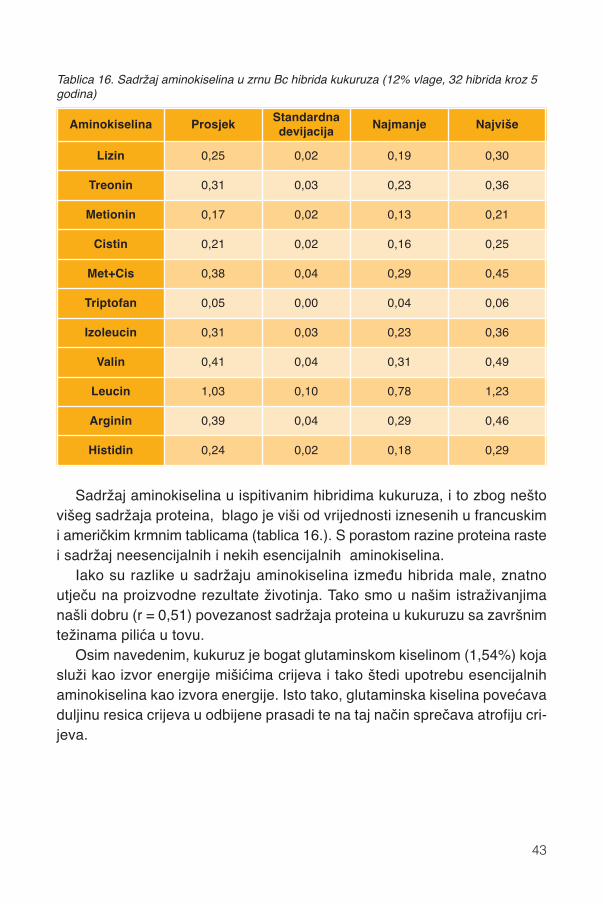
43
Sadržaj aminokiselina u ispitivanim hibridima kukuruza, i to zbog neštovišeg sadržaja proteina, blago je viši od vrijednosti iznesenih u francuskimi američkim krmnim tablicama (tablica 16.). S porastom razine proteina rastei sadržaj neesencijalnih i nekih esencijalnih aminokiselina.
Iako su razlike u sadržaju aminokiselina između hibrida male, znatnoutječu na proizvodne rezultate životinja. Tako smo u našim istraživanjimanašli dobru (r = 0,51) povezanost sadržaja proteina u kukuruzu sa završnimtežinama pilića u tovu.
Osim navedenim, kukuruz je bogat glutaminskom kiselinom (1,54%) kojasluži kao izvor energije mišićima crijeva i tako štedi upotrebu esencijalnihaminokiselina kao izvora energije. Isto tako, glutaminska kiselina povećavaduljinu resica crijeva u odbijene prasadi te na taj način sprečava atrofiju cri-jeva.
Aminokiselina Prosjek Standardnadevijacija Najmanje Najviše
Lizin 0,25 0,02 0,19 0,30
Treonin 0,31 0,03 0,23 0,36
Metionin 0,17 0,02 0,13 0,21
Cistin 0,21 0,02 0,16 0,25
Met+Cis 0,38 0,04 0,29 0,45
Triptofan 0,05 0,00 0,04 0,06
Izoleucin 0,31 0,03 0,23 0,36
Valin 0,41 0,04 0,31 0,49
Leucin 1,03 0,10 0,78 1,23
Arginin 0,39 0,04 0,29 0,46
Histidin 0,24 0,02 0,18 0,29
Tablica 16. Sadržaj aminokiselina u zrnu Bc hibrida kukuruza (12% vlage, 32 hibrida kroz 5godina)

44
8. UljeUlje (mast) iz hrane opskrbljuje životinju koncentriranom energijom za
korištenje ili skladištenje i esencijalnim masnim kiselinama koje životinjatreba a ne može ih sintetizirati. Jedinica (%) ulja kukuruza sadržava 2,25puta više energije od škroba i proteina, te 3,6% ulja daje oko 10% energijekukuruza. Sve masne kiseline, uključujući i esencijalne imaju strukturnefunkcije, a stupanj nezasićenosti daje fluidnost staničnim membranama.Esencijalne masne kiseline imaju dvostruku vezu na 6 (omega 6) ili 3 uglji-kovom atomu lanca (omega 3) od krajnje metilne (CH3) skupine i imaju višefiziološku nego energetsku ulogu u hranidbi životinja.
Omega 6 i omega 3 masne kiseline tekućine su na sobnoj temperaturipa se ugrađuju u staničnu membranu životinjskih stanica i time omogućujuoptimalnu propusnost. Međutim, preveliki udjel omega 6 u odnosu naomega 3 kiseline nije dobar za zdravlje životinja. Pretpostavlja se da je ne-povoljan svaki omjer veći od 6, a što je ujednačeniji udjel omega 6 odomega 3 kiselina to je bolji utjecaj na zdravlje. Kukuruzno ulje ima izrazitonepovoljan omjer jer sadržava dosta omega 6 kiselina (56%) a vrlo maloomega 3 kiselina (oko 1%). Ipak, u Bc institutu provodi se selekcija na po-rast udjela omega 3 masnih kiselina u ulju kukuruza.
Nekoliko omega 3 esencijalnih masnih kiselina služi kao sirovina za sin-tezu staničnih hormona prostaglandina i drugih eikosanoida koji su moćnifiziološki regulatori staničnih funkcija. Eikosanoidi reguliraju grušanje krvi,krvni tlak, kontrakciju mišića i imunosni odgovor.
Kolesterolu slične tvari – fitosteroli i fitostenoli nalaze se u aleuronskomsloju zrna kukuruza i njihova konzumacija za više od 40% smanjuje učesta-lost srčanih udara u ljudi. U prosjeku, klica Bc hibrida kukuruza sadržava16,83% ulja u 88% suhe tvari
Dodavanje ulja povisuje uzimanje hrane i poboljšava njezinu teksturu,zdravlje i plodnost životinja, te smanjuje prašnjavost hrane. Ulje je nosačvitamina topljivih u mastima (A, D, E i K) koji su bitne tvari za sve proceseu stanicama tijekom cijelog života životinje.
8.1. Sadržaj
Kukuruz sadržava više ulja (3,6%) nego pšenica, ječam i tritikale. U zrnukukuruza najviše je masti smješteno u klici (82,6%), pa endospermu(15,4%) i najmanje u omotaču (2%).
Ulje zrna bogato je nezasićenim masnim kiselinama (85%) koje sadrža-vaju više energije nego zasićene masne kiseline. Kukuruz sa 1% više uljaod prosjeka ima više energije, a time i višu vrijednost za stočare.

45
Prosječan sadržaj (3,73%) i varijabilnost (SD ± 0,26%) ulja u zrako-suhojtvari 32 Bc hibrida zrna kukuruza kroz pet godina (tablica 17.) vrlo je sličanprosjeku (3,7 ± 0,4%) od 2634 analize francuskog kukuruza i viši od pro-sjeku (3,48%) od 115 analiza američkog kukuruza. Na variranje sadržajaulja u kukuruzu znatnije djeluje genetski čimbenik – hibrid nego okolišni čim-benici - gnojidba, lokacija i gustoća sklopa kukuruza.
Bc hibridi kukuruza s višim sadržajem ulja (Bc 462, Alibi, Riđan i Kekec)bolji su u hranidbi peradi i svinja nego preživača.
Relativno velike razlike u sadržaju ulja (± 0,5%) odražavaju se na proiz-vodnju peradi zato što svakih 0,1% ulja povisuje sadržaj ME, i to 35 kJ.Pilići i kokoši bolje iskorištavaju hranu s hibridima kukuruza koji sadržavajuviše ulja. Nadalje, ulje pomaže apsorpciji karotenoida iz zrna kukuruza paje bolja boja jaja.
8.2. Masne kiseline
Ulje kukuruza sadrži 85% masnih kiselina i odličan je izvor nezasićenihesencijalnih masnih kiselina - linolne i linolenske i visoko potentne forme vi-tamina E koji su bitni za zdravlje životinja i ljudi.
Kvaliteta ulja određena je omjerom nezasićenih prema zasićenim ma-snim kiselinama.
Ulje (sirova mast), %
Prosjek Standardna devijacija Najmanje Bc 778 Najviše Alibi
3.60 0,80 2.83 4.7
Tablica 17. Sadržaj ulja (%) u Bc hibridima kukuruza (12% vlage, 32 hibrida kroz 5 godina)
Masna kiselina Prosjek Standardna Najmanje NajvišePalmitinska (C16:0) 0.34 0.02 0.27 0.40
Stearinska (C18:0) 0.06 0.00 0.04 0.07
Oleinska (C18:1) 0.82 0.06 0.65 0.98
Linolna (C18:2) 1.73 0.13 1.36 2.06
Linolenska (C18:3) 0.03 0.00 0.02 0.04
Tablica 18. Sadržaj (%) najzastupljenijih masnih kiselina u zrnu Bc hibrida kukuruza (12%vlage, 32 hibrida kroz 5 godina)

46
Tablica 18. pokazuje da kukuruzno ulje obiluje nezasićenim masnim ki-selinama (86,7%) pa tako i zrno Bc hibrida sadržava najviše linolne (1,73%)i oleinske kiseline (0,82%). Nezasićene i dugolančane masne kiseline sa-državaju više energije nego zasićene i pridonose sa 10% ME kukuruza.
Glavna je prednost kukuruznog pred drugim uljima njegova visoka odr-živost. Razlog ovome je visoki sadržaj linolne kiseline koja je zbog smještajana drugom ugljkovom lancu glicerola zaštićena od oksidacije i njezinih štet-nih posljedica po zdravlje životinja. Nadalje, ulje kukuruza stabilno je i zbogvisokog sadržaja prirodnih u mastima topljivih antioksidansa koji obuhvaćajutokole (vitamin E) i druge spojeve.
Linolna kiselina je omega 6 esencijalna masna kiselina koja izgrađujemembrane stanica i lipoproteine, te se iz nje stvaraju tvari bitne za plodnostživotinja. U hrani nesilica treba biti najmanje 1,2-1,5% linolne kiseline kojapovećava krupnoću jaja. Hrana pilića treba sadržavati najmanje 1,2% ovekiseline. Isto tako, hrana za prasad i krmače treba sadržavati 1,5-1,8% li-nolne kiseline. Tipičan udjel Bc hibrida osigurava potrebni udjel linolne ki-seline u potpunoj hrani.
Količina i sastav masti hrane određuju sastav masti trupa svinja i peradi.Naše tržište traži čvrstu slaninu koja se dobije hranjenjem svinja ječmom.Međutim, mast svinja hranjenih kukuruzom umjesto ječmom sadržava višelinolne kiseline (12:9.2%) i ima viši jodni broj pa je trbušna slanina mekana.Viši udjel nezasićenih masnih kiselina u masti trupa, međutim, iako daje me-kanu slaninu povoljno djeluje na zdravlje ljudi.
Utvrđene razlike u sadržaju pojedinih masnih kiselina odraz su razlika usadržaju ulja a ne u sastavu ulja pojedinih Bc hibrida kukuruza (tablica 18).
U hranidbi peradi te prasadi i krmača treba koristiti Bc hibride s višim sa-držajem ulja jer će životinje takvu hranu radije jesti i dobivati više linolne ki-seline, odnosno više energije.

47
9. Vlakna
9.1. Funkcija
Vlakna kukuruza, neškrobni ugljikohidrati, uglavnom su celuloza, hemi-celuloza u manjoj količini pektini, oligosaharidi, te lignin i svi su sastojci su-baleuronskog i aleuronskog omotača te perikarpa. Celuloza se sastoji odlinearno povezanih molekula glukoze, a hemiceluloza od homopolimera: D-ksiloze, D-manoze, D-glukouronske kiseline D-galaktoze, L-arabinize itd.Homopolimeri koji grade hemicelulozu su arabinoksilan, β-glukani, ksiloglu-kani, galaktomanan. Pektini su građeni od poligalaktourana, ramanogalak-tourana. Oligosaharidi (α-galaktozidaze) su rafinoza, stahioza, verbakoza iajugoze.
Kemijski gledano, vlakna su najraznovrsnija hranjiva tvar te ih je teškoodrediti kao jednu tvar. Stoga se određuju na različite načine, ovisno o pri-lagođenosti probavila njihovu iskorištenju. Stara metoda određivanja sirovihvlakana vrlo je neprecizna jer od prisutne količine sastojaka vlakana odre-đuje samo sadržaj celuloze, ali ne i hemicelulozu, lignin i pektin. Zato je sa-držaj ukupnih vlakana (NDF) 2-4 puta veći od sadržaja sirovih vlakana.
Preciznije metode određivanja za preživače su neutralna deterdžent me-toda (NDF), a za monogastrične životinje neškrobni polisaharidi (NŠP). Ana-liza NDF najviše se koristi za voluminoznu krmu kojom se hrane preživači,a sastoji se od celuloze, hemiceluloze, lignina i donekle pektina. Vlaknazrna kukuruza su NŠP koji su zbroj sadržaja NDF+pektina+oligosaharida.Kako zrno kukuruza sadržava malo, samo 0,8% pektina i još manje oligo-saharida - oko 0,3% onda su neškrobni polisaharidi za 1,1% više od NDF.
NŠP se dijele na topljive ugljikohidrate, veći dio hemiceluloze, pektini ioligosaharidi, te netopljiva vlakna, celuloza i lignin (lignoceluloza). Zrno ku-kuruza sadržava 5,5% topljivih i 3,8% netopljivih vlakana.
Ako kukuruz sadržava više vlakana to je manji sadržaj proteina i osobitoškroba u zrnu, odnosno manja je energetska vrijednost krmiva. Osim toga,vlakna, odnosno NŠP neprobavljivi su dio krmiva za enzime peradi i svinjai dobro probavljivi za enzime mikroorganizama u probavnom sustavu. Pilićii prasad ne mogu, a odrasle kokoši i svinje mogu probaviti dio vlakana hranezahvaljujući enzimima mikroorganizama slijepoga i debelog crijeva. S jednestrane vlakna od svih hranjivih tvari najviše (r=-0,43) smanjuju energetskuvrijednost zrna kukuruza za monogastrične životinje, a s druge strane go-tovo najviše pridonose zdravlju probavnog sustava. Preživači, zahvaljujućibrojim mikroorganizmima u buragu, te kunići i konji zahvaljujući mikrobimau slijepom i debelom crijevu, dobro iskorištavaju vlakna obroka pa su pro-

48
dukti fermentacije vlakana važan izvor energije za biljojede. Netopljiva su vlakna inertna, a topljiva su aktivna na više načina. Topljiva
vlakna u tankom crijevu pilića bubre i stvaraju viskoznu masu što nije dobroza rast, iskorištenje hraniva i vlažnost izmeta. Na sreću, kukuruz sadržavanajmanje topljivih vlakana među žitaricama te je i zbog toga odličan izvorenergije.
9.2. Sadržaj
Zrno kukuruza sadržava najmanje vlakana među žitaricama pa je i tojedan od razloga njegove visoke probavljivosti, konzumacije i energetskevrijednosti. Razlog je činjenica da 51% vlakana potječe iz omotača i drškezrna kojih je najmanje (5%) u zrnu, a samo 12% u endospermu kojega jenajviše (84%) od zrna kukuruza. Naime, u zrnu kukuruza najviše vlakanasadržavaju omotač (90%) i drška zrna (95%), slijedi aleuronski endosperm(50%) pa klica (11%), a najmanje škrobni endosperm (1%).
U tablici 19. prikazane su karakteristične vrijednosti sadržaja frakcija vla-kana u zrnu Bc hibrida kukuruza. Kada se gledaju frakcije vlakana, zrno sa-država najviše (9,07%) NDF koji se sastoji od 6,2% hemiceluloze i 2,78%ADF (celuloza+lignin). Vlakna kukuruznog zrna sadržavaju dvostruko višehemiceluloze (%NDF-%ADF) od celuloze (%ADF-%ADL). Vrlo je mali(0,34%) sadržaj lignina u zrnu, a njime je najbogatiji vrh kukuruza. Bc hibridiu usporedbi s američkim (9,11%) i francuskim (10,4%) sadržavaju neštomanje svih frakcija vlakana.
Općenito, zrno kukuruza pa tako i Bc hibrida sadržava malo svih frakcijavlakana, a osobito lignina (tablica 19) koji preko ograničenja probavljivostsmanjuje hranjivu vrijednost i konzumaciju krmiva.
Frakcije vlakana Prosjek Standardnadevijacija Najmanje Najviše
Neutralna deterdžent vlakna (NDF), % 9,07 0,65 7,33 10,39
Hemiceluloza 6,29 0,27 4,98 6,92
Kisela deterdžent vlakna (ADF), % 2,78 0,27 2,35 3,47
Celuloza 2,44 0,24 2,21 2,88
Lignin (ADL), % 0,34 0,10 0,14 0,5
Tablica 19. Sadržaj frakcija vlakana u zrnu Bc hibrida kukuruza (12% vlage, 32 hibrida kroz5 godina)

49
10. Energija
Domaće životinje najveći dio (65-70%) pojedene hrane koriste kao izvorenergije te podmirenje energetskih potreba sudjeluje sa 50-60% u cijeni ko-štanja animalne proizvodnje. Zrno kukuruza energijom je najbogatija žitaricakoja se uzgaja u Europi i samo ono može biti jedini izvor energije svim vr-stama i kategorijama peradi i svinja, te glavni preživačima. Silaža kukuruzaenergijom je najbogatije silirano voluminozno krmivo našeg podneblja.Škrob je glavni izvor energije i zrna (77,5%) i silaže kukuruza (45,0%) pasve što utječe na njegov sadržaj i iskoristivost određuje energetsku vrijed-nost kukuruza.
U potpunoj hrani životinja kukuruz nije jedino krmivo pa njegov tipičanudjel podmiruje sljedeće količine energetskih potreba: 60% mladih životinja,75% svinja u tovu, 45-50% mliječnih krava, te 75-80% junadi u tovu.
Za kukuruz kao glavni sastojak potpune hrane iznimno je važno što toč-nije procijeniti energetsku vrijednost kako bi se ostvarilo što bolje iskorište-nje hrane, primjerna visina i kvaliteta proizvoda, dobro zdravlje a svimeovime ekonomska dobit.
Različita proizvodnja životinja hranjenih istim udjelom kukuruza i sastavompotpune hrane najbolji je pokazatelj neujednačene energetske vrijednostikukuruza. Dobar su primjer dvostruke razlike prirasta (680-1301 g/28 dana)između skupina pilića koje su svaka hranjena kukuruzom iz jedne od 15država umiješanim s jednakim udjelom u potpune smjese istog sastava.Različita energetska vrijednost kukuruza uzrokovana je brojnim čimbenicimakoji se mogu svrstati u sljedeće skupine: genetika, uvjeti rasta i žetve,postupci prerade i sastav hrane. Dio razlika može se pripisati varijacijamau kemijskom sastavu, a dio kinetici probave škroba kukuruza kao glavnogizvora energije. Različite proizvodnje životinja hranjenih hibridima sličnogsastava i ukupne probavljivosti pokazuju da kinetika probave škroba i me-tabolizma glukoze imaju veliki utjecaj na iskorištenje energije zrna kukuruza.Klasična hranidba uzima da je energetska vrijednost statična (jednavrijednost) unutar iste vrste životinja i tipa proizvodnje, dok suvremenaistraživanja pokazuju da je ona promjenljiva vrijednost (vidi poglavlje škrob).Stoga nam tablične i iz kemijskog sastava procijenjene vrijednosti pokazujupotencijalnu, ali ne i stvarnu energetsku vrijednost standardnog zrna zaprosječne životinje u standardnim uvjetima proizvodnje. Nažalost, standardnikukuruz, životinja i uvjeti proizvodnje statističke su kategorije koje ne

50
egzistiraju u stvarnoj proizvodnji. Nadalje, klasična procjena hranjivevrijednosti još ne uzima u račun razlike u kinetici probave škroba imetabolizma glukoze između pošiljki kukuruza. Međutim, u nedostatkustvarne koristimo se potencijalnom energetskom vrijednosti kukuruza uhranidbi peradi, svinja, preživača, konja, kunića i riba.
Energija u zrnu kukuruza naziva se bruto energija i nalazi se u organskojtvari, voda i pepeo ne sadržavaju energiju, a životinja ju crpi nakon konzu-macije samo iz probavljenoga, apsorbiranoga i metaboliziranog dijelaproteina, masti i ugljikohidrata (grafikon 7..). Energetska vrijednost kukuruzamjeri se kao razlika između energije u zrnu kukuruza i energije ostalenakon odbijanja energije u gubicima (grafikon 7.). Tijekom transformacijeenergije kukuruza u energiju proizvoda energija se gubi u: (1) neprobavljenimtvarima kukuruza u fecesu (neprobavljena vlakna, škrob, masti i bjelančevine)),(2) neiskorištenim probavljenim tvarima u plinovima i mokraći, te (3) meta-bolizmom hraniva proizvedenoj toplini (grafikon 7.). Što se manje energijekukuruza gubi tijekom njegove probave i metabolizma to je viša njegovaenergetska vrijednost. Kako se domaće životinje razlikuju po građi probavnogtrakta i metabolizmu, različitu količinu energije iskoriste iz istog zrnakukuruza perad, svinje i goveda. Neto energetska vrijednost kukuruza po-kazuje koliko se kilograma mesa, mlijeka i jaja može proizvesti iz energijekilograma kukuruza kada su ostale potrebne hranjive tvari prisutne u hraniu najmanje dovoljnoj količini.
Grafikon 7. Metabolički put iskorištenja energije iz kukuruza u domaćih životinja
BRUTO ILI UKUPNA ENERGIJA (toplina oslobođena izgaranjem krmiva)
Energija fecesa PROBAVLJIVA ENERGIJA = ENERGIJA U PROBAVLJENIM HRANJIVIM TVARIMA
METABOLIZIRAJUĆA ILI FIZIOLOŠKA ENERGIJA
Energija metana
Energija mokraće
NETO ENERGIJAPrirast topline stvoren probavom i metabolizmom hranjivih tvari
NE za REPRODUKCIJU
NE za PROIZVODNJU
NE za ODRŽAVANJE
Toplinaiskorištenja HT
Ukupna proizvodnja topline

51
Bruto (BE) ili ukupna energija ukupna je kemijska energija u organskojtvari zrna kukuruza koja se oslobodi kao toplina nakon sagorijevanja uzorkau kalorimetru u uvjetima čistog O2 i tlaka od 25 atmosfera. Sadržaj brutoenergije može se predvidjeti iz kemijskog sastava jer svaka jedinica hranjivetvari uvijek ima istu količinu bruto energije u svim krmivima (tablica 20.).
Mala je razlika u sadržaju bruto energije u suhoj tvari između zrna (18,8MJ/kg) i stabljike kukuruza (18 MJ/kg) jer se slična količina topline oslobodispaljivanjem škroba i vlakana (tablica 20). Međutim, mi intuitivno znamo daće pilići hranjeni zrnom kukuruza koje je bogato škrobom (64%) rasti, a hra-njeni kukuruzovinom bogatom vlaknima (70%) umrijeti, i to zato što ne mogurazgraditi vlakna pa iz njih dobiju 0 kJ energije. Preživači mogu iskoristitivlakna, doduše manje nego škrob, pa iz suhe tvari zrna iscrpe samo 8,45ili 45%, a iz stabljike 4,6 MJ/kg ili 25% bruto energija. Zato bruto energijanije dobar pokazatelj energetske vrijednosti kukuruza za životinje. Ona jesamo početak određivanja iskorištenja energije prisutne u hranjivim tvarimau zrnu kukuruza.
Probavljiva energija (PE) definira se kao dio ukupne energije krmivaumanjene za energiju neprobavljivih sastojaka hrane izlučenih fecesom.
PE = Bruto energija – Energija fecesa
Probavljiva energija nalazi se u probavljenim i apsorbiranim tvarima, tj.u glukozi, masnim kiselinama i aminokiselinama. Zato vrsta i količina hra-niva te njihova probavljivost bitno određuju sadržaj probavljive energije ukukuruzu. Kako voda i pepeo ne sadržavaju energiju a netopljiva su vlakna(lignoceluloza) potpuno neiskoristiva za perad i svinje, količina topljivih vla-kana, škroba, ulja i proteina pri istom sadržaju vlage određuju razlike u sa-držaju probavljive energije u kukuruzu.
U pravilu svinje probave visokih 75-80% bruto energije prosječne hranei 90% bruto energije zrna kukuruza pa je probavljiva energija 90% brutoenergije zrna kukuruza. Ulje ima dvostruko više energije od škroba pa po-visuje sadržaj probavljive energije. Kako kukuruz sadržava malo vlakana i
Hranjive tvariProtein Ulje Škrob Šećeri Vlakna
23,6 39,8 17,3 16,0 18,9
Tablica 20. Bruto energija je toplina oslobođena sagorijevanjem 1 g hranjive tvari (kJ/g)

52
prilično nepromjenjivu količinu škroba, samo razlike u sadržaju ulja utječuna razlike u sadržaju energije između hibrida.
Danas se sadržaj probavljive energije koristi za procjenu energetske vri-jednosti hrane za biljojede pa je u tablicama prikazano koliko energije izsvakog Bc hibrida dobiju konji i kunići. Probavljiva energija ne vodi računao daljnjim gubicima energije iz organizma u hranivima izlučenim urinom iplinovima.
Metabolizirajuća energija (ME) dobije se kada se od probavljive energijeoduzmu gubici energije u metanu (0,5% od PE) i dušičnim spojevima mo-kraće (urea, hipurna kiselina, kreatinin, alantonin), te nedušičnim spojevimakao što su glukouronska i limunska kiselina (3,5% od PE).
ME = Probavljiva energija – Energija mokraće – Energija metana
Metabolizirajuća energija je energija krmiva u zadržanim hranjivim tva-rima u životinji i stoji joj na raspolaganju za metaboličke funkcije održavanja,proizvodnje rasta i razmnožavanja, pa se još naziva i zadržana energija, ra-spoloživa ili fiziološka energija. Kako kukuruz sadržava malo vlakana i pro-teina, to je metabolička energija 86% bruto ili 96% probavljive energije zasvinje.
Energiju probavljenih hranjivih tvari kukuruza u metabolizmu najboljeiskorištavaju svinje, slijede perad pa goveda. Razlog je što svinje među do-maćim životinjama najbolje probave škrob, masti i vlakna kukuruza, te iznjih izvuku najviše energije.
Sljedeći je gubitak energije u obliku topline koja nastaje tijekom pretvorbemetaboličke u neto energiju. Neto ili čista energija kukuruza je ona količinaenergije krmiva koja nakon svih gubitaka ostane životinji za podmirenje po-treba održavanja osnovnih životnih funkcija, za stvaranje novih tjelesnih sa-stojaka (rast), ugradnju u plod (razmnožavanje) i stvaranje proizvoda(mlijeko, vuna i jaja). U pravilu se najveći dio energije zadržanih hranjivimtvarima gubi u obliku topline pri njihovoj transformaciji u neto energiju pagoveda 60%, svinje 70%, a perad 80% metaboličke energije iskoriste kaoneto energiju (tablica 21). Goveda najlošije iskorištavaju bruto energijuhrane, oko 35%, što je znatno više nego u automobilima gdje samo 15%energije goriva prijeđe u kretanje. Apsorbirane masti proizvode najmanjetopline tijekom metabolizma pa one imaju i najviše ne samo bruto već i netoenergiju.
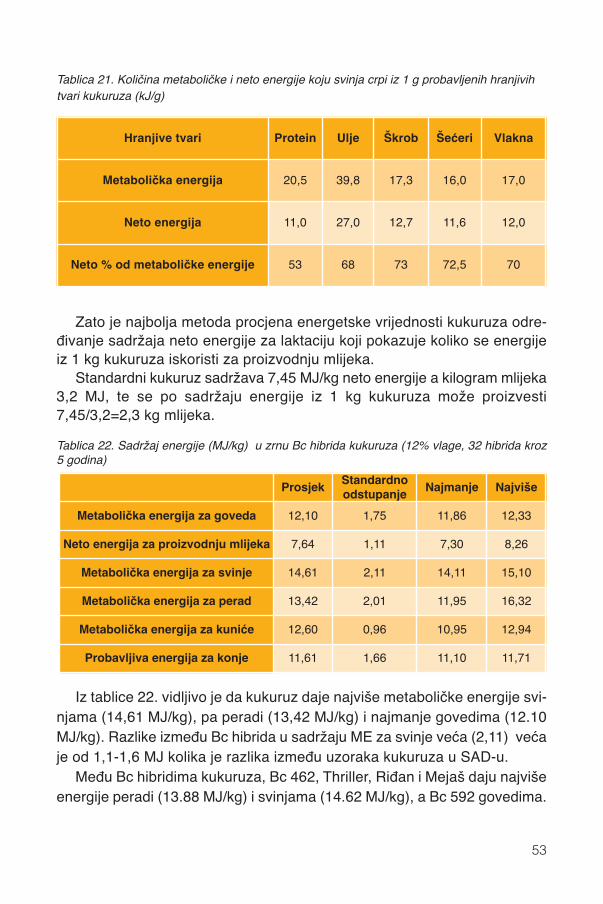
53
Zato je najbolja metoda procjena energetske vrijednosti kukuruza odre-đivanje sadržaja neto energije za laktaciju koji pokazuje koliko se energijeiz 1 kg kukuruza iskoristi za proizvodnju mlijeka.
Standardni kukuruz sadržava 7,45 MJ/kg neto energije a kilogram mlijeka3,2 MJ, te se po sadržaju energije iz 1 kg kukuruza može proizvesti7,45/3,2=2,3 kg mlijeka.
Iz tablice 22. vidljivo je da kukuruz daje najviše metaboličke energije svi-njama (14,61 MJ/kg), pa peradi (13,42 MJ/kg) i najmanje govedima (12.10MJ/kg). Razlike između Bc hibrida u sadržaju ME za svinje veća (2,11) većaje od 1,1-1,6 MJ kolika je razlika između uzoraka kukuruza u SAD-u.
Među Bc hibridima kukuruza, Bc 462, Thriller, Riđan i Mejaš daju najvišeenergije peradi (13.88 MJ/kg) i svinjama (14.62 MJ/kg), a Bc 592 govedima.
Hranjive tvari Protein Ulje Škrob Šećeri Vlakna
Metabolička energija 20,5 39,8 17,3 16,0 17,0
Neto energija 11,0 27,0 12,7 11,6 12,0
Neto % od metaboličke energije 53 68 73 72,5 70
Tablica 21. Količina metaboličke i neto energije koju svinja crpi iz 1 g probavljenih hranjivihtvari kukuruza (kJ/g)
Prosjek Standardnoodstupanje Najmanje Najviše
Metabolička energija za goveda 12,10 1,75 11,86 12,33
Neto energija za proizvodnju mlijeka 7,64 1,11 7,30 8,26
Metabolička energija za svinje 14,61 2,11 14,11 15,10
Metabolička energija za perad 13,42 2,01 11,95 16,32
Metabolička energija za kuniće 12,60 0,96 10,95 12,94
Probavljiva energija za konje 11,61 1,66 11,10 11,71
Tablica 22. Sadržaj energije (MJ/kg) u zrnu Bc hibrida kukuruza (12% vlage, 32 hibrida kroz5 godina)
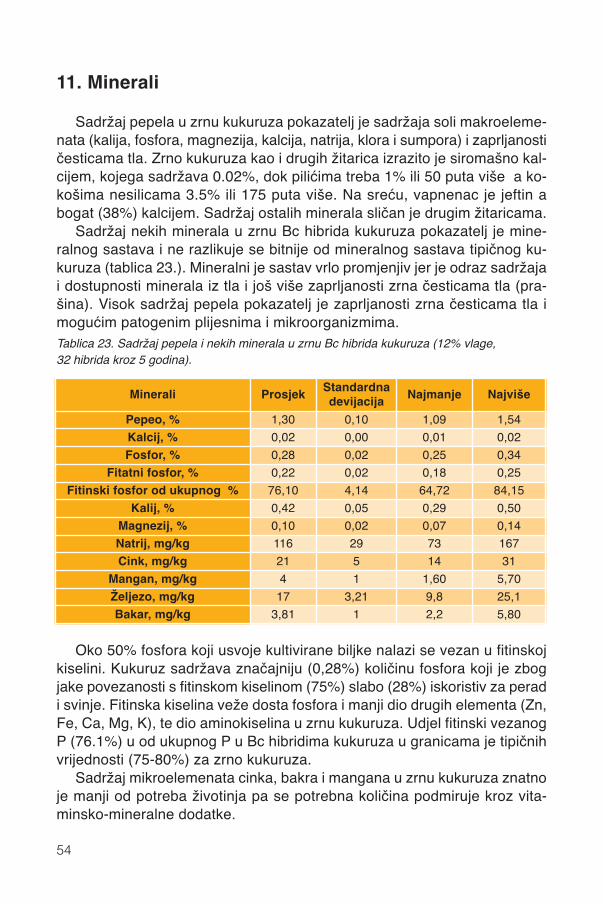
54
11. MineraliSadržaj pepela u zrnu kukuruza pokazatelj je sadržaja soli makroeleme-
nata (kalija, fosfora, magnezija, kalcija, natrija, klora i sumpora) i zaprljanostičesticama tla. Zrno kukuruza kao i drugih žitarica izrazito je siromašno kal-cijem, kojega sadržava 0.02%, dok pilićima treba 1% ili 50 puta više a ko-košima nesilicama 3.5% ili 175 puta više. Na sreću, vapnenac je jeftin abogat (38%) kalcijem. Sadržaj ostalih minerala sličan je drugim žitaricama.
Sadržaj nekih minerala u zrnu Bc hibrida kukuruza pokazatelj je mine-ralnog sastava i ne razlikuje se bitnije od mineralnog sastava tipičnog ku-kuruza (tablica 23.). Mineralni je sastav vrlo promjenjiv jer je odraz sadržajai dostupnosti minerala iz tla i još više zaprljanosti zrna česticama tla (pra-šina). Visok sadržaj pepela pokazatelj je zaprljanosti zrna česticama tla imogućim patogenim plijesnima i mikroorganizmima.
Oko 50% fosfora koji usvoje kultivirane biljke nalazi se vezan u fitinskojkiselini. Kukuruz sadržava značajniju (0,28%) količinu fosfora koji je zbogjake povezanosti s fitinskom kiselinom (75%) slabo (28%) iskoristiv za peradi svinje. Fitinska kiselina veže dosta fosfora i manji dio drugih elementa (Zn,Fe, Ca, Mg, K), te dio aminokiselina u zrnu kukuruza. Udjel fitinski vezanogP (76.1%) u od ukupnog P u Bc hibridima kukuruza u granicama je tipičnihvrijednosti (75-80%) za zrno kukuruza.
Sadržaj mikroelemenata cinka, bakra i mangana u zrnu kukuruza znatnoje manji od potreba životinja pa se potrebna količina podmiruje kroz vita-minsko-mineralne dodatke.
Minerali Prosjek Standardnadevijacija Najmanje Najviše
Pepeo, % 1,30 0,10 1,09 1,54Kalcij, % 0,02 0,00 0,01 0,02Fosfor, % 0,28 0,02 0,25 0,34
Fitatni fosfor, % 0,22 0,02 0,18 0,25Fitinski fosfor od ukupnog % 76,10 4,14 64,72 84,15
Kalij, % 0,42 0,05 0,29 0,50Magnezij, % 0,10 0,02 0,07 0,14Natrij, mg/kg 116 29 73 167Cink, mg/kg 21 5 14 31
Mangan, mg/kg 4 1 1,60 5,70Željezo, mg/kg 17 3,21 9,8 25,1Bakar, mg/kg 3,81 1 2,2 5,80
Tablica 23. Sadržaj pepela i nekih minerala u zrnu Bc hibrida kukuruza (12% vlage,32 hibrida kroz 5 godina).

55
12. Ukusnost kukuruza
Hranjivost je združeno djelovanje kemijskog sastava, probavljivosti i kon-zumacije. Ukusnost hrane bitan je dio hranjivosti preko poticanja konzuma-cije. Sklonost (preferencija) je relativna konzumacija ispitivane u odnosu nastandardnu hranu.
Osobito su svinje osjetljive na ukusnost hrane koja određuje koju ćehranu i koliko će pojedine hrane pojesti. Tri su međusobno isprepletena su-stava u odabiru i visini konzumacije hrane: (1) ukusnost poželjne hrane ineukusnost nepoželjne/toksične hrane, (2) fizikalna svojstva i (3) povratniodgovor razine šećera i drugih hraniva u krvi.
Svinjama je ugodan sladak i umami okus koji potječu iz jednostavnihugljikohidrata i L-aminokiselina. Svinje i perad, zbog visokih energetskih po-treba, skloniji su hrani bogatijoj uljem, šećerima i škrobom nego vlaknima.Toksične i antinutritivne tvari gorka su okusa koji odvraća svinje od konzu-macije. Fizikalna svojstva koja utječu na teksturu digesta i rastezanje zidacrijeva određuju sklonost pojedinoj hrani i visini konzumacije hrane. Divljesvinje i jeleni više odabiru hibrid prema kemijskom sastavu nego prema fi-zikalnim svojstvima. Testiranje sklonosti svinja prema svakom od 25 krmivaprovedenom u Španjolskoj (2014.) pokazalo je da svinje radije jedu krmivas više probavljivog škroba i s višim glikemijskim indeksom nego krmiva sviše vlakana. Isto tako, okus koji dolazi od maltodekstrina kukuruza takođerdjeluje na preferenciju svinja.
Hibridi kukuruza razlikuju se po ukusnosti, a iskustvo i poljski pokusi po-kazuju da divlje svinje rado jedu Bc 462 i Bc 282, te da su sklonije ranimnego kasnijim Bc hibridima kukuruza. Nadalje, vjerojatno će više jesti i hi-bride bogate šećerom (Bc 306, Kekec i Riđan). Isto tako jeleni preferirajuprobavljivije i ranije hibride cijele biljke kukuruza u ranijim stadijima razvojazrna.

56
13. FitonutrijentiFitonutrijenti su sastojci biljne hrane koji imaju prvenstveno ulogu zaštitite
biljaka i unapređuju zdravlje životinja i čovjeka. Cjelovite su žitarice u od-nosu na voće i povrće često bogatiji izvor nekih i jedini drugih fitonutrijenata.
Ljudi i životinje prvenstveno konzumiraju kukuruz da bi podmirili energet-ske potrebe pa je svako dodatno povoljno djelovanje sastojaka kukuruzadodatna vrijednost za proizvođača i potrošača.
Kukuruz je bogat izvor fitonutrijenata koji ima najveću antioksidacijskuaktivnost među žitaricama. Kukuruz sadržava fitonutrijente u svim dijelovimazrna: fenole, dijetna vlakna i fitinsku kiselinu u omotaču, karotenoide i rezi-stentni škrob u endospermu, te esencijalne masne kiseline, fitosterole i vi-tamin E u klici. Nadalje, kukuruz sadržava već opisane ugljikohidrate,rezistentni škrob i dijetna vlakna koji također unapređuju zdravlje životinja iljudi.
Bc hibridi kukuruza visoke su antioksidacijske aktivnosti i visokog poten-cijala prinosa pa su uz dobra adaptivna svojstva atraktivni za proizvodnju.
13.1. Fenoli
Fenoli su skupina spojeva s benzenskim prstenom i jednom ili više hi-droksilnih skupina koje biljka stvara tijekom rasta radi zaštite od negativnihvanjskih utjecaja patogenih organizama, suše i fizičkih oštećenja. Preciznijerečeno, fenoli su najvećim dijelom nalaze u staničnoj stijenici te predstav-ljaju fizičku i kemijsku barijeru biljke kao unakrsno vezivo vlakana omotača,antioksidansi koji se bore protiv razarajućih radikali patogenih organizama,te sredstva za odbijanje konzumacije od strane insekata. Fenoli su sastojcilignina i tanina te flavonoida antocijana koji daje crvenu boju zrnu kukuruza.Ferulinska (ferulična) kiselina najzastupljeniji je fenol jer čini 85% fenolnihspojeva zrna kukuruza i rijetko se nalazi u voću i povrću. Fenolni spojeviprisutni su u slobodnoj i konjugiranoj formi u žitaricama. Najviša je njihovakoncentracija u aleuronskom sloju žitarica, ali se mogu naći i u embriju iomotaču zrna.
Negativni vanjski utjecaji suše, patogenih mikroorganizama i fizička ošte-ćenja povezani su s oksidacijskim stresom čije učinke ublažavaju fenoli kojisu najsnažniji antioksidansi zrna kukuruza. Osim na zrno kukuruza, fenolipovoljno djeluju na (1) probavu te metabolizam glukoze i lipida, (2) sporim
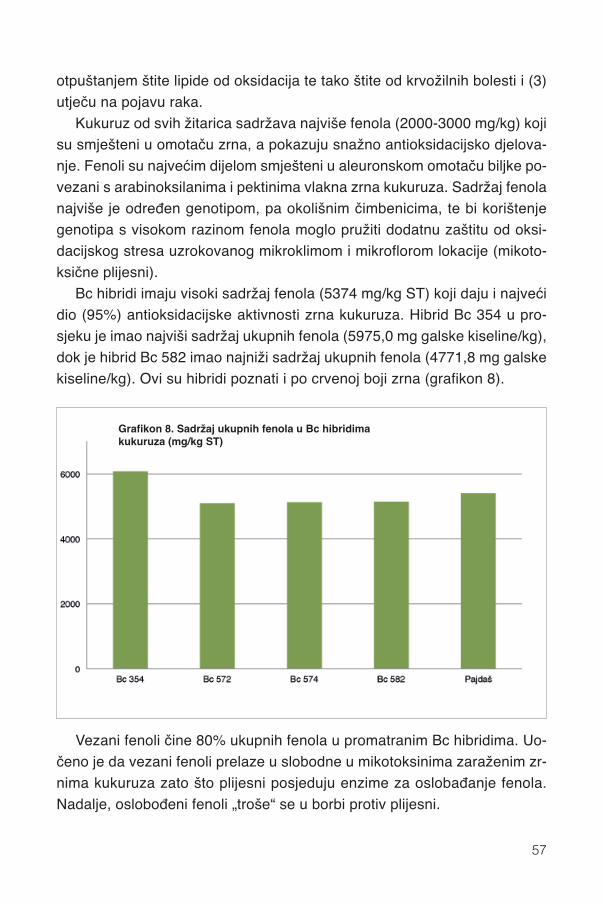
57
otpuštanjem štite lipide od oksidacija te tako štite od krvožilnih bolesti i (3)utječu na pojavu raka.
Kukuruz od svih žitarica sadržava najviše fenola (2000-3000 mg/kg) kojisu smješteni u omotaču zrna, a pokazuju snažno antioksidacijsko djelova-nje. Fenoli su najvećim dijelom smješteni u aleuronskom omotaču biljke po-vezani s arabinoksilanima i pektinima vlakna zrna kukuruza. Sadržaj fenolanajviše je određen genotipom, pa okolišnim čimbenicima, te bi korištenjegenotipa s visokom razinom fenola moglo pružiti dodatnu zaštitu od oksi-dacijskog stresa uzrokovanog mikroklimom i mikroflorom lokacije (mikoto-ksične plijesni).
Bc hibridi imaju visoki sadržaj fenola (5374 mg/kg ST) koji daju i najvećidio (95%) antioksidacijske aktivnosti zrna kukuruza. Hibrid Bc 354 u pro-sjeku je imao najviši sadržaj ukupnih fenola (5975,0 mg galske kiseline/kg),dok je hibrid Bc 582 imao najniži sadržaj ukupnih fenola (4771,8 mg galskekiseline/kg). Ovi su hibridi poznati i po crvenoj boji zrna (grafikon 8).
Vezani fenoli čine 80% ukupnih fenola u promatranim Bc hibridima. Uo-čeno je da vezani fenoli prelaze u slobodne u mikotoksinima zaraženim zr-nima kukuruza zato što plijesni posjeduju enzime za oslobađanje fenola.Nadalje, oslobođeni fenoli „troše“ se u borbi protiv plijesni.
Grafikon 8. Sadržaj ukupnih fenola u Bc hibridimakukuruza (mg/kg ST)

58
Među ispitivanim Bc hibridima najvišu antioksidacijsku aktivnost pokazujuBc 354 i Pajdaš koji sadržavaju i najviše ukupnih fenola (grafikon 9.). An-tioksidacijska aktivnost usko je ( r = 0,81) određena sadržajem fenola.
Naša istraživanja pokazuju da suša povisuje genetski određen udjelukupnih i slobodnih fenola koji ublažavaju njezine nepovoljne učinke. Hibridis višim genetskim predispozicijama za sintezu fenola mogli bi imati većiznačaj u nastupajućim klimatski ekstremnim godinama.
13.2. Karotenoidi
Karotenoidi su velika skupina (> 700 spojeva) u mastima topljivih antio-ksidacijskih spojeva prisutnih u biljkama koji su odgovorni za žutu, naran-častu i crvenu boju voća, cvijeća i žitarica. U biljkama djeluju kao prekursorihormona, bojila i esencijalne su komponente fotosintetskog aparata gdje suuključeni u sakupljanje svjetlosne energije u stanju smanjene količine svjetlakoju prenose na klorofil gdje se koristi u fotosintezi. Također, igraju ključnuulogu kao fotoprotektivni spojevi hvatanjem kisika proizašlog iz viška svjet-losne energije, onemogućujući oštećenje membrana. U biljnom svijetu po-stoji nekoliko stotina različitih karotenoida, a strukturno se razlikuju dvijeskupine: karoteni, koji su čisti ugljikovodici, te ksantofili, koji još sadržavajui kisik – oksigenirani derivati karoten.
Grafikon 9. Antioksidacijska aktivnost ukupnih fenolapet Bc hibrida kukuruza (mmol Troloxa/kg)

59
Životinje i ljudi ne mogu sintetizirati karotenoide pa ih moraju dobivatibiljnom ili animalnom hranom. Karotenoidi su antioksidansi i bojila (kože,potkožne masti i žumanjak jajeta), a neki su i prekusori vitamina A (α i β-karoten, te β-kriptoksantin kao najvažniji iz kukuruza).
U životinjama, karotenoidi su antioksidansi protiv vanjskih oksidansa(reaktivne vrste kisika), kao što je ultravioletno zračenje, te tako štite kožui vid. Isto tako brane životinju od unutrašnjih oksidansa koji se najviše proiz-vode u mitohondrijima i fagocitima. Među karotenoidima najjače antioksi-dacijsko djelovanje imaju α-karoten > β-kriptoksantin > β-karoten > lutein >zeaksantin. Pritom je djelovanje navedenih karotenoida bilo od dva do triputa više od α-tokoferola, tj. vitamina E.
Nedostatak karotenoida odnosno prekursora vitamina A u prehrani ljudii životinja dovodi do različitih poremećaja i bolesti, poput sljepoće, uspora-vanja rasta, slabljenja imunosnog sustava, a služe i u prevenciji raka i kar-diovaskularnih bolesti.
Kada se karotenoidi ne iskoriste kao provitamini vitamina A ili antioksi-dansi nakupljaju se i skladište u različitim organima – jetrima i masnomtkivu, a u peradi su osobito bitni kao bojila žumanjka jajeta, kože, mesa ikljuna. Uobičajeno je mišljenje u nas da intenzivnija boja kože i mesa po-kazuje zdraviju perad koja je „prirodnije“ hranjena (paša). Znak vrsnosti kva-litete jaja u nas je zlatnožuta do narančasta boja žumanjka jajeta. Bojajajeta potječe od karotenoida hrane (kukuruza). Karotenoidi u jajetu zaštitasu od kvarenja i izvor antioksidansa i provitamina A za piliće. Boja jajeta in-strumentalno se određuje tzv. kolorimetrima koji boju klasificiraju u tri di-menzije: (1) L* (mjera svjetline od crnog (0) do bijelog (100)), (2) a* (mjeracrvenosti od crvene (a) do zelene (-a) boje) i (3) b* (mjera žutosti od žute(b) do plave (-b) boje. Vrsta i sadržaj karotenoidnih spojeva u žumanjku ja-jeta obično je odraz vrste i sadržaja karotenoida u hrani nesilica.
Uobičajena primjena sintetskih bojila u hrani životinja i ljudi smanjuje sezbog rizika od stvaranja različitih oblika raka i kožnih bolesti. Zbog toga seekstrakti prirodnih pigmenata sve više koriste u pigmentaciji žumanjaka ja-jeta - lucerna, rajčica, paprika i cvijet nevena ili njihove kombinacije. U tomesvjetlu sadržaj karotenoida u hibridima kukuruza iznimno je važno poznavatiradi manje upotrebe skupih sintetskih i ekstrakata prirodnih bojila jaja i pot-kožne masti peradi.
13.2.1. Karotenoidi Bc hibrida kukuruza
Među krmivima koja se koriste u intenzivnoj hranidbi najviše karotenoidasadržava zrno kukuruza i njegovi su proizvodi, te dehidrirana lucerna. Me-
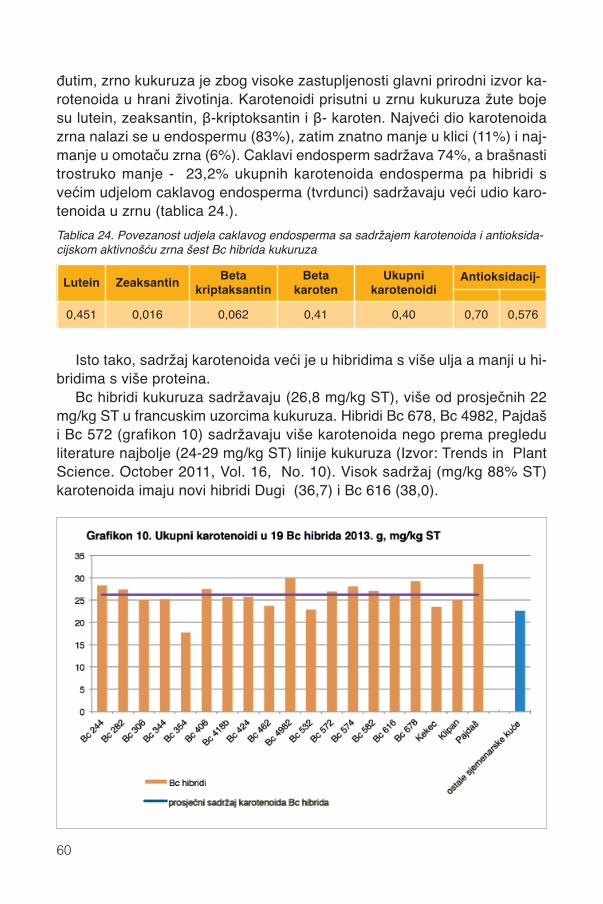
60
đutim, zrno kukuruza je zbog visoke zastupljenosti glavni prirodni izvor ka-rotenoida u hrani životinja. Karotenoidi prisutni u zrnu kukuruza žute bojesu lutein, zeaksantin, β-kriptoksantin i β- karoten. Najveći dio karotenoidazrna nalazi se u endospermu (83%), zatim znatno manje u klici (11%) i naj-manje u omotaču zrna (6%). Caklavi endosperm sadržava 74%, a brašnastitrostruko manje - 23,2% ukupnih karotenoida endosperma pa hibridi svećim udjelom caklavog endosperma (tvrdunci) sadržavaju veći udio karo-tenoida u zrnu (tablica 24.).
Isto tako, sadržaj karotenoida veći je u hibridima s više ulja a manji u hi-bridima s više proteina.
Bc hibridi kukuruza sadržavaju (26,8 mg/kg ST), više od prosječnih 22mg/kg ST u francuskim uzorcima kukuruza. Hibridi Bc 678, Bc 4982, Pajdaši Bc 572 (grafikon 10) sadržavaju više karotenoida nego prema pregleduliterature najbolje (24-29 mg/kg ST) linije kukuruza (Izvor: Trends in PlantScience. October 2011, Vol. 16, No. 10). Visok sadržaj (mg/kg 88% ST)karotenoida imaju novi hibridi Dugi (36,7) i Bc 616 (38,0).
Lutein Zeaksantin Beta kriptaksantin
Beta karoten
Ukupni karotenoidi
Antioksidacij-
0,451 0,016 0,062 0,41 0,40 0,70 0,576
Tablica 24. Povezanost udjela caklavog endosperma sa sadržajem karotenoida i antioksida-cijskom aktivnošću zrna šest Bc hibrida kukuruza
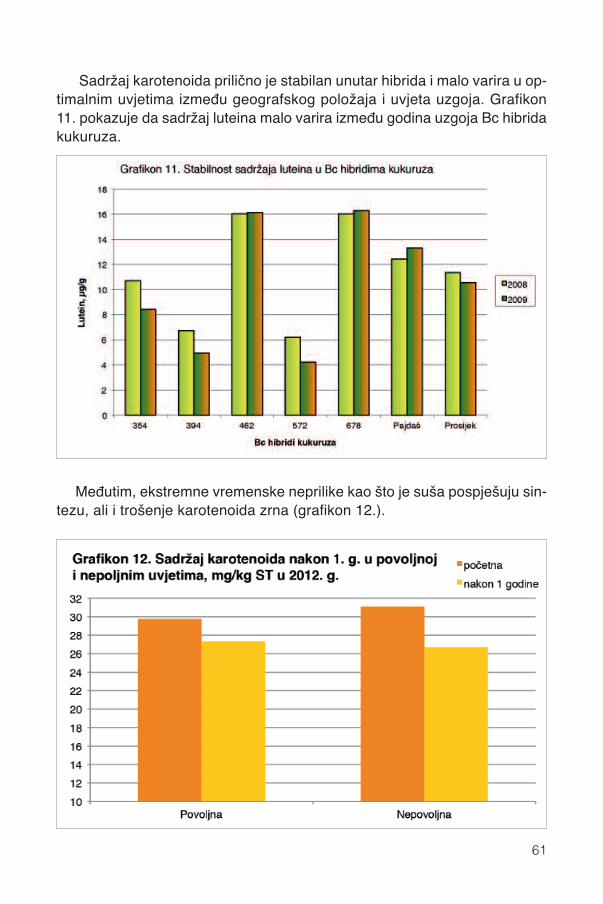
61
Sadržaj karotenoida prilično je stabilan unutar hibrida i malo varira u op-timalnim uvjetima između geografskog položaja i uvjeta uzgoja. Grafikon11. pokazuje da sadržaj luteina malo varira između godina uzgoja Bc hibridakukuruza.
Međutim, ekstremne vremenske neprilike kao što je suša pospješuju sin-tezu, ali i trošenje karotenoida zrna (grafikon 12.).

62
13.2.2. Pojedini karotenoidi
Između kultivara kukuruza postoje velike razlike u sadržaju kako ukupnihtako i pojedinih karotenoida, što je osnova za proizvodnju hibrida povišenogsadržaja jednog ili više karotenoida. U prosjeku hibridi Bc kukuruza sadr-žavaju uobičajenih 10 puta više luteina i zeaksantina nego β-kriptoksantinai β-karotena (tablica 25. i grafikon 13.).
U ljudskom organizmu karotenoidi su antioksidansi, smanjuju pojavu kar-diovaskularnih bolesti, raka pluća, reumatioidnog artritisa i osteoproze. Lu-tein i zeaksantin najviše pridonose antioksidacijskoj aktivnosti kukuruza.
Među 600 karotenoida u prirodi, 10 je u krmi i samo su dva u oku. Luteini zeaksantin su karotenoidi oka. Oba karotenoida bitna su za zdravlje očijui degenerativne bolesti oka starijih osoba i glavni su karotenoidi retine oka(slika 6.). Dnevna konzumacija 6 mg/d štiti i ublažava pojavu navedenih bo-lesti oka, a može se dobiti konzumacijom 300 g/d kukuruznog brašna.
Međutim, om jer luteina i zeaksantina znatno varira između hibrida. U kuku-ruzu postoje tri tipa nasljeđivanja sadržaja luteina i ze a ksa ntina: (1) vi še luteina:
Slika 6. Oko sadržava zeaksantin i lutein.
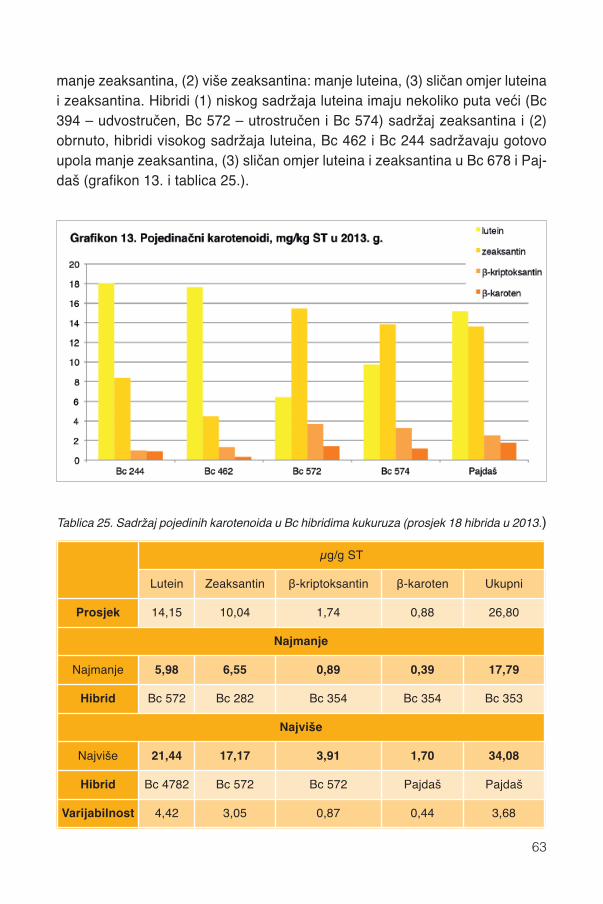
ma nje zeaksantina, (2) vi še zeaksantina: ma nje luteina, (3) sličan omjer luteinai zeaksantina. Hibridi (1) niskog sadržaja luteina imaju nekoliko puta veći (Bc394 – udvostručen, Bc 572 – utrostručen i Bc 574) sadržaj zeaksantina i (2)obrnuto, hibridi visokog sadržaja luteina, Bc 462 i Bc 244 sadržavaju gotovoupola manje zeaksantina, (3) sličan omjer luteina i zeaksantina u Bc 678 i Paj-daš (grafikon 13. i tablica 25.).
63
µg/g ST
Lutein Zeaksantin β-kriptoksantin β-karoten Ukupni
Prosjek 14,15 10,04 1,74 0,88 26,80
Najmanje
Najmanje 5,98 6,55 0,89 0,39 17,79
Hibrid Bc 572 Bc 282 Bc 354 Bc 354 Bc 353
Najviše
Najviše 21,44 17,17 3,91 1,70 34,08
Hibrid Bc 4782 Bc 572 Bc 572 Pajdaš Pajdaš
Varijabilnost 4,42 3,05 0,87 0,44 3,68
Tablica 25. Sadržaj pojedinih karotenoida u Bc hibridima kukuruza (prosjek 18 hibrida u 2013.)
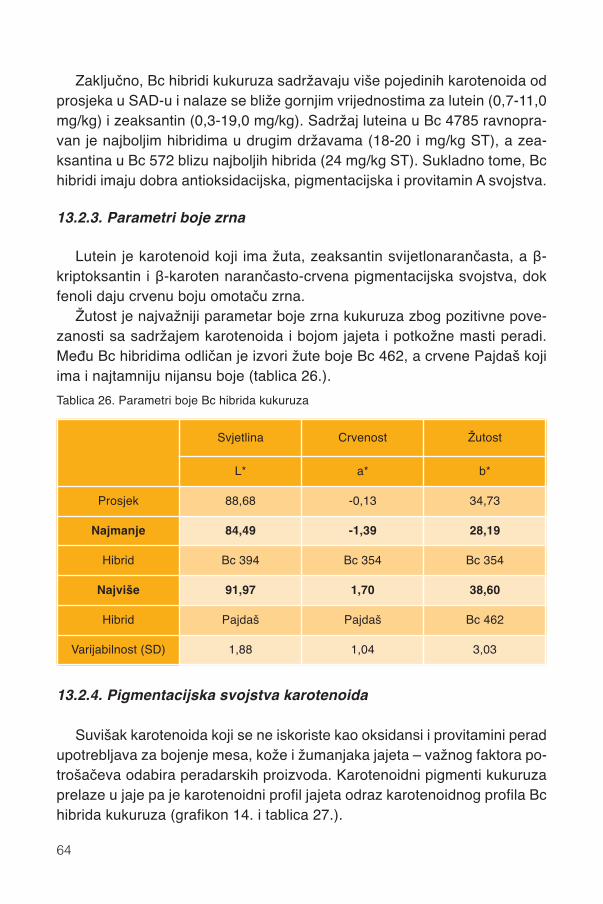
64
Zaključno, Bc hibridi kukuruza sadržavaju više pojedinih karotenoida odprosjeka u SAD-u i nalaze se bliže gornjim vrijednostima za lutein (0,7-11,0mg/kg) i zeaksantin (0,3-19,0 mg/kg). Sadržaj luteina u Bc 4785 ravnopra-van je najboljim hibridima u drugim državama (18-20 i mg/kg ST), a zea-ksantina u Bc 572 blizu najboljih hibrida (24 mg/kg ST). Sukladno tome, Bchibridi imaju dobra antioksidacijska, pigmentacijska i provitamin A svojstva.
13.2.3. Parametri boje zrna
Lutein je karotenoid koji ima žuta, zeaksantin svijetlonarančasta, a β-kriptoksantin i β-karoten narančasto-crvena pigmentacijska svojstva, dokfenoli daju crvenu boju omotaču zrna.
Žutost je najvažniji parametar boje zrna kukuruza zbog pozitivne pove-zanosti sa sadržajem karotenoida i bojom jajeta i potkožne masti peradi.Među Bc hibridima odličan je izvori žute boje Bc 462, a crvene Pajdaš kojiima i najtamniju nijansu boje (tablica 26.).
13.2.4. Pigmentacijska svojstva karotenoida
Suvišak karotenoida koji se ne iskoriste kao oksidansi i provitamini peradupotrebljava za bojenje mesa, kože i žumanjaka jajeta – važnog faktora po-trošačeva odabira peradarskih proizvoda. Karotenoidni pigmenti kukuruzaprelaze u jaje pa je karotenoidni profil jajeta odraz karotenoidnog profila Bchibrida kukuruza (grafikon 14. i tablica 27.).
Svjetlina Crvenost Žutost
L* a* b*
Prosjek 88,68 -0,13 34,73
Najmanje 84,49 -1,39 28,19
Hibrid Bc 394 Bc 354 Bc 354
Najviše 91,97 1,70 38,60
Hibrid Pajdaš Pajdaš Bc 462
Varijabilnost (SD) 1,88 1,04 3,03
Tablica 26. Parametri boje Bc hibrida kukuruza
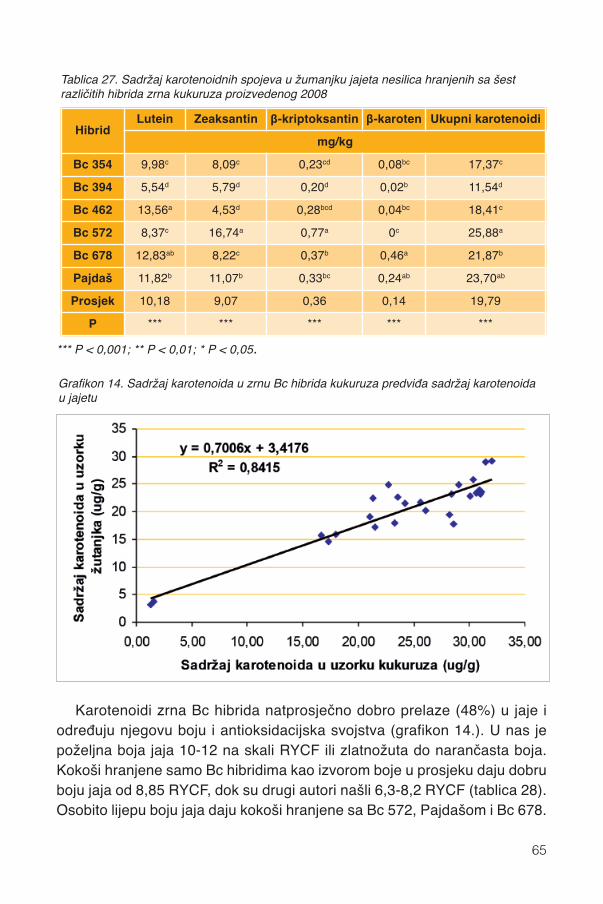
Karotenoidi zrna Bc hibrida natprosječno dobro prelaze (48%) u jaje iodređuju njegovu boju i antioksidacijska svojstva (grafikon 14.). U nas jepoželjna boja jaja 10-12 na skali RYCF ili zlatnožuta do narančasta boja.Kokoši hranjene samo Bc hibridima kao izvorom boje u prosjeku daju dobruboju jaja od 8,85 RYCF, dok su drugi autori našli 6,3-8,2 RYCF (tablica 28).Osobito lijepu boju jaja daju kokoši hranjene sa Bc 572, Pajdašom i Bc 678.
65
HibridLutein Zeaksantin β-kriptoksantin β-karoten Ukupni karotenoidi
mg/kgBc 354 9,98c 8,09c 0,23cd 0,08bc 17,37c
Bc 394 5,54d 5,79d 0,20d 0,02b 11,54d
Bc 462 13,56a 4,53d 0,28bcd 0,04bc 18,41c
Bc 572 8,37c 16,74a 0,77a 0c 25,88a
Bc 678 12,83ab 8,22c 0,37b 0,46a 21,87b
Pajdaš 11,82b 11,07b 0,33bc 0,24ab 23,70ab
Prosjek 10,18 9,07 0,36 0,14 19,79P *** *** *** *** ***
Tablica 27. Sadržaj karotenoidnih spojeva u žumanjku jajeta nesilica hranjenih sa šestrazličitih hibrida zrna kukuruza proizvedenog 2008
*** P < 0,001; ** P < 0,01; * P < 0,05.
Grafikon 14. Sadržaj karotenoida u zrnu Bc hibrida kukuruza predviđa sadržaj karotenoidau jajetu
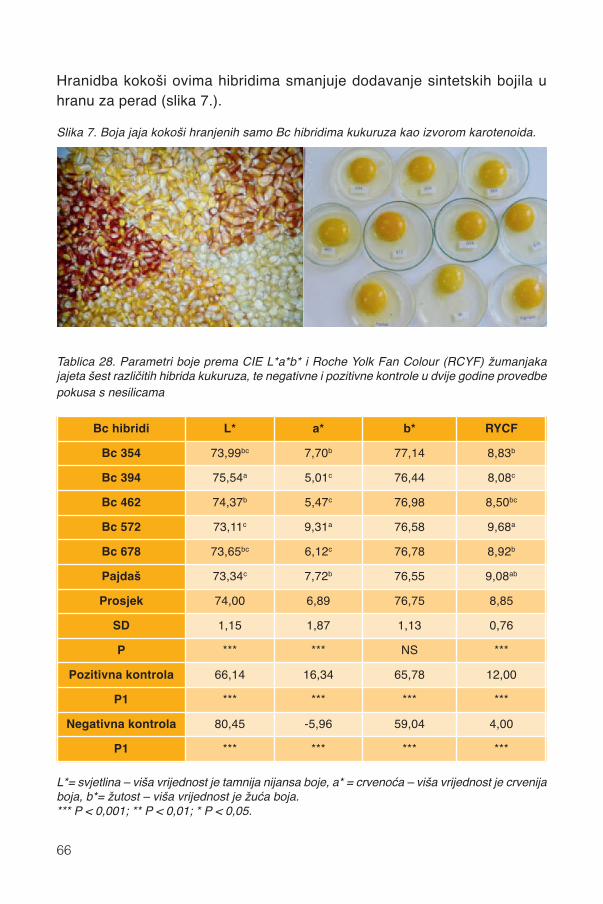
66
Hranidba kokoši ovima hibridima smanjuje dodavanje sintetskih bojila uhranu za perad (slika 7.).
Bc hibridi L* a* b* RYCF
Bc 354 73,99bc 7,70b 77,14 8,83b
Bc 394 75,54a 5,01c 76,44 8,08c
Bc 462 74,37b 5,47c 76,98 8,50bc
Bc 572 73,11c 9,31a 76,58 9,68a
Bc 678 73,65bc 6,12c 76,78 8,92b
Pajdaš 73,34c 7,72b 76,55 9,08ab
Prosjek 74,00 6,89 76,75 8,85
SD 1,15 1,87 1,13 0,76
P *** *** NS ***
Pozitivna kontrola 66,14 16,34 65,78 12,00
P1 *** *** *** ***
Negativna kontrola 80,45 -5,96 59,04 4,00
P1 *** *** *** ***
Slika 7. Boja jaja kokoši hranjenih samo Bc hibridima kukuruza kao izvorom karotenoida.
Tablica 28. Parametri boje prema CIE L*a*b* i Roche Yolk Fan Colour (RCYF) žumanjakajajeta šest različitih hibrida kukuruza, te negativne i pozitivne kontrole u dvije godine provedbepokusa s nesilicama
L*= svjetlina – viša vrijednost je tamnija nijansa boje, a* = crvenoća – viša vrijednost je crvenijaboja, b*= žutost – viša vrijednost je žuća boja. *** P < 0,001; ** P < 0,01; * P < 0,05.
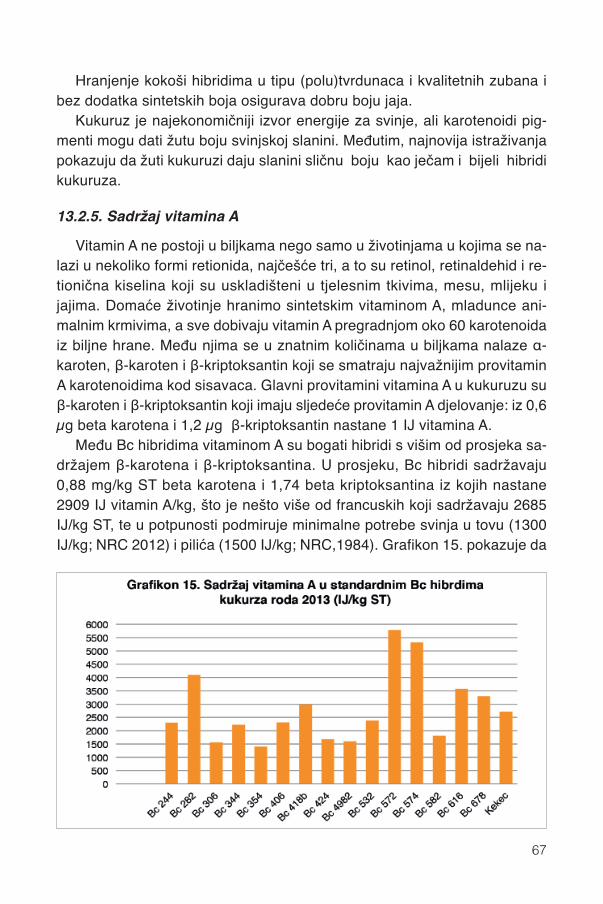
67
Hranjenje kokoši hibridima u tipu (polu)tvrdunaca i kvalitetnih zubana ibez dodatka sintetskih boja osigurava dobru boju jaja.
Kukuruz je najekonomičniji izvor energije za svinje, ali karotenoidi pig-menti mogu dati žutu boju svinjskoj slanini. Međutim, najnovija istraživanjapokazuju da žuti kukuruzi daju slanini sličnu boju kao ječam i bijeli hibridikukuruza.
13.2.5. Sadržaj vitamina AVitamin A ne postoji u biljkama nego samo u životinjama u kojima se na-
lazi u nekoliko formi retionida, najčešće tri, a to su retinol, retinaldehid i re-tionična kiselina koji su uskladišteni u tjelesnim tkivima, mesu, mlijeku ijajima. Domaće životinje hranimo sintetskim vitaminom A, mladunce ani-malnim krmivima, a sve dobivaju vitamin A pregradnjom oko 60 karotenoidaiz biljne hrane. Među njima se u znatnim količinama u biljkama nalaze α-karoten, β-karoten i β-kriptoksantin koji se smatraju najvažnijim provitaminA karotenoidima kod sisavaca. Glavni provitamini vitamina A u kukuruzu suβ-karoten i β-kriptoksantin koji imaju sljedeće provitamin A djelovanje: iz 0,6µg beta karotena i 1,2 µg β-kriptoksantin nastane 1 IJ vitamina A.
Među Bc hibridima vitaminom A su bogati hibridi s višim od prosjeka sa-držajem β-karotena i β-kriptoksantina. U prosjeku, Bc hibridi sadržavaju0,88 mg/kg ST beta karotena i 1,74 beta kriptoksantina iz kojih nastane2909 IJ vitamin A/kg, što je nešto više od francuskih koji sadržavaju 2685IJ/kg ST, te u potpunosti podmiruje minimalne potrebe svinja u tovu (1300IJ/kg; NRC 2012) i pilića (1500 IJ/kg; NRC,1984). Grafikon 15. pokazuje da
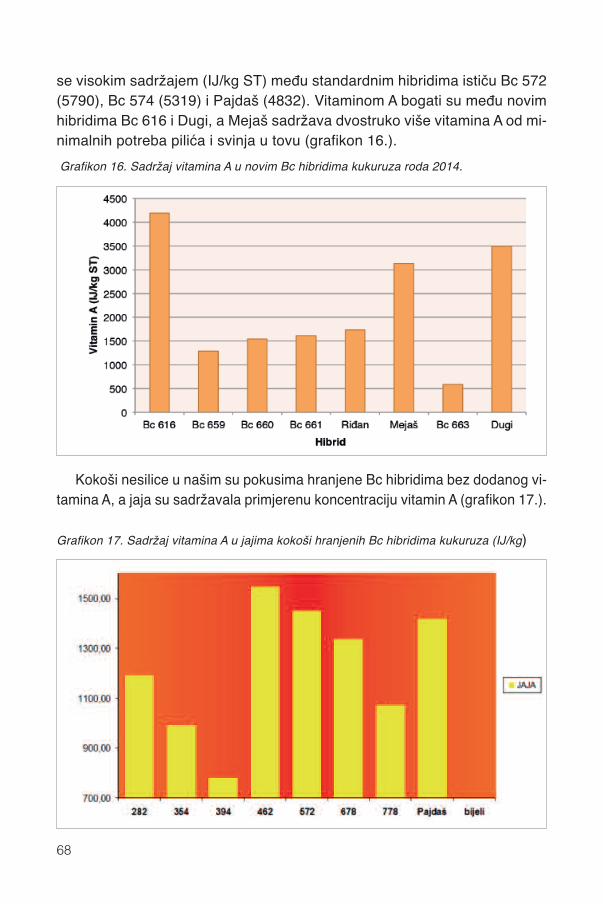
68
se visokim sadržajem (IJ/kg ST) među standardnim hibridima ističu Bc 572(5790), Bc 574 (5319) i Pajdaš (4832). Vitaminom A bogati su među novimhibridima Bc 616 i Dugi, a Mejaš sadržava dvostruko više vitamina A od mi-nimalnih potreba pilića i svinja u tovu (grafikon 16.).
Kokoši nesilice u našim su pokusima hranjene Bc hibridima bez dodanog vi-tamina A, a jaja su sadržavala primjerenu koncentraciju vitamin A (grafikon 17.).
Grafikon 16. Sadržaj vitamina A u novim Bc hibridima kukuruza roda 2014.
Grafikon 17. Sadržaj vitamina A u jajima kokoši hranjenih Bc hibridima kukuruza (IJ/kg)

69
13.3. Vitamin E
Vitamin E je zajednički naziv za skupinu osam prirodnih tokoferola i to-kotrienola koji posjeduju aktivnost α-tokoferola. Zajednički se nazivaju tokolii svi imaju antioksidacijsko djelovanje. Vitamin E je najvažniji antioksidansprve linije obrane koji sprečava oksidacijska oštećenja nezasićenih masnihkiselina membrana svake stanice od oksidansa – peroksil radikala. Naime,peroksil radikali 1000 se puta brže vežu s vitaminom E nego s nezasićenimmasnim kiselinama i na taj ga način neutraliziraju. Svaka infekcija i drugistres povezani su s oksidacijskim stresom koji troši vitamin E iz ograničenihrezervi organizma te ga životinja treba kontinuirano dobivati hranom. Namanjak vitamina E osjetljive su lipidne membrane živaca, mišića (krvne žile,srce, mišići nogu) i jetara, tkiva koja troše puno kisika pa proizvode dostaoksidansa. Svaka vrsta životinja pokazuje različite simptome manjka vita-mina E. U peradi se javlja nekroza jetara eksudativna dijateza, encefalo-malacija, u svinja dudoliko srce, a u preživača muskulaturna distrofija.Vitamin E pomaže u plodnosti životinja te manjak uzrokuje reproduktivneporemetnje. U krava vitamin E ublažava pojave zadržavanja posteljice iupala maternice.
Dodavanje vitamine E povisuje zaštitu od nekoliko zaraznih i virusnih bo-lesti, stimulira sintezu protutijela, popravlja oštećene gene i održava svježuboju mesa.
Nedavna istraživanja jasno pokazuju da dvostruko manje doze prirodnogvitamina E bolje sprečavaju oksidaciju nezasićenih masti (ulja) kukuruza,soje i osobito lana nego sintetski vitamin E. Isto tako prirodni vitamin E boljesmanjuje oštećenja mišića u trkaćih konja i više ga se nakuplja u jetrima odsintetskog vitamina E. Stoga je važno znati sadržaj i profil vitamina E u zrnupojedinih hibrida kukuruza.
Kukuruz među žitaricama sadržava najviše (17-47 mg/kg ST ) vitaminaE koji je najvećim dijelom smješten u klici gdje štiti ulje od oksidacijskogkvarenja. Sadržaj vitamina E je usko povezan sa sadržajem ulja u zrnu ku-kuruza. Velike su genetske razlike u sadržaju i profilu vitamina E izmeđukultivara kukuruza.
Prema našim istraživanjima, Bc hibridi sadržavaju visokih 43,32 mg/kgST (grafikon 18.), dok europske tablice navode 19,7 mg/kg ST, a premaameričkim, prosječni sadržaj je 13,2 mg/kg ST.
Osobito su bogati vitamin E spojevima hibridi Bc 4982 i Bc 424 (grafikon18. i tablica 29.).
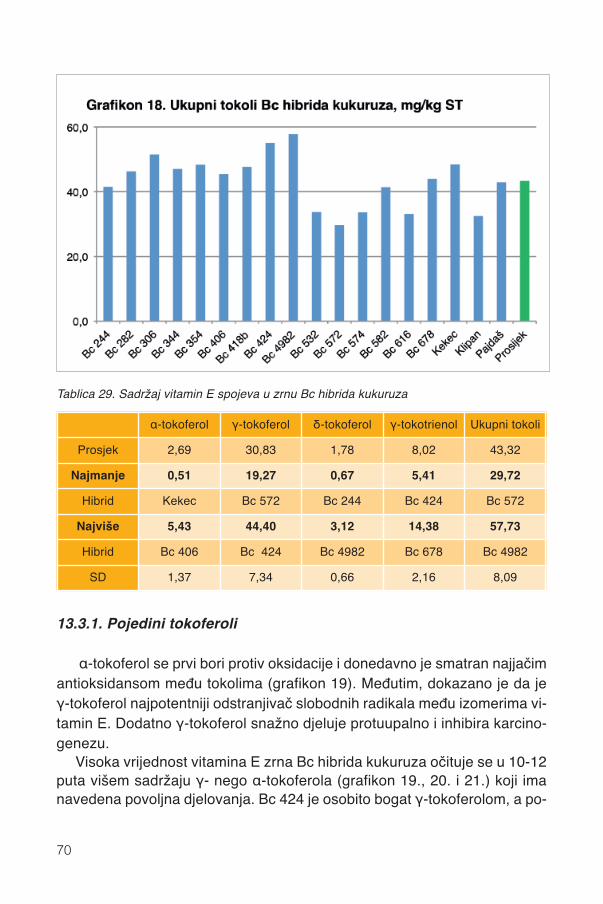
70
13.3.1. Pojedini tokoferoli
α-tokoferol se prvi bori protiv oksidacije i donedavno je smatran najjačimantioksidansom među tokolima (grafikon 19). Međutim, dokazano je da jeγ-tokoferol najpotentniji odstranjivač slobodnih radikala među izomerima vi-tamin E. Dodatno γ-tokoferol snažno djeluje protuupalno i inhibira karcino-genezu.
Visoka vrijednost vitamina E zrna Bc hibrida kukuruza očituje se u 10-12puta višem sadržaju γ- nego α-tokoferola (grafikon 19., 20. i 21.) koji imanavedena povoljna djelovanja. Bc 424 je osobito bogat γ-tokoferolom, a po-
α-tokoferol γ-tokoferol δ-tokoferol γ-tokotrienol Ukupni tokoli
Prosjek 2,69 30,83 1,78 8,02 43,32
Najmanje 0,51 19,27 0,67 5,41 29,72
Hibrid Kekec Bc 572 Bc 244 Bc 424 Bc 572
Najviše 5,43 44,40 3,12 14,38 57,73
Hibrid Bc 406 Bc 424 Bc 4982 Bc 678 Bc 4982
SD 1,37 7,34 0,66 2,16 8,09
Tablica 29. Sadržaj vitamin E spojeva u zrnu Bc hibrida kukuruza
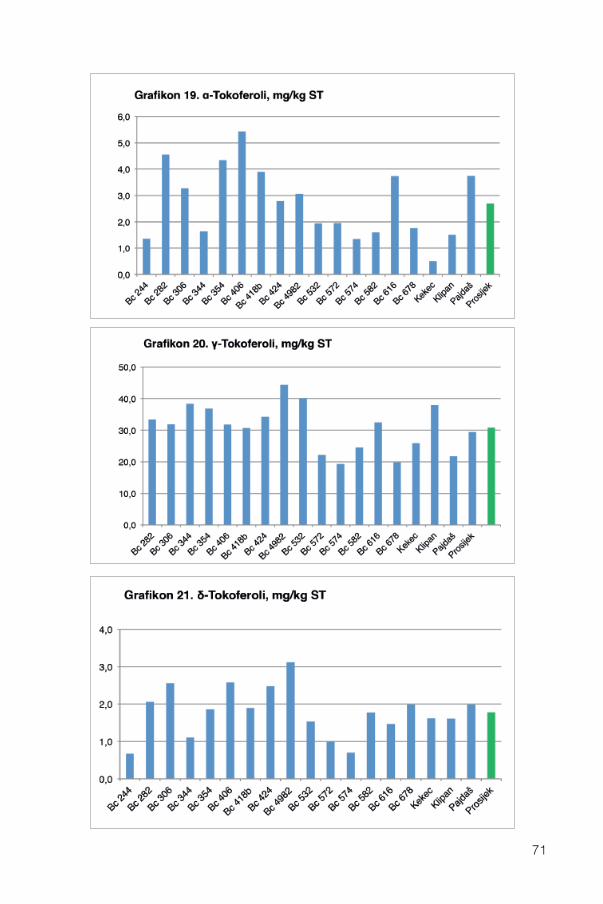
71

72
Tablice 30. Minimalne potrebe peradi i svinja za vitaminom E (mg/kg ST)
pularni hibridi Bc 678 i Pajdaš sadržavaju dvostruko više vitamina E od eu-ropskoga i američkog prosječnog sadržaja u zrnu kukuruza. Tipičan udjelkukuruza od 60% u potpunim smjesama podmiruje minimalne potrebe svi-nja u tovu, te pilenki i kokoši nesilica (tablica 30.).
13.4. Antioksidacijska moć
Životinje trebaju kisik kako bi dobile energiju. Međutim, kisik je i potenci-jalni otrov za životinje i ljude. Slobodni radikali uglavnom su spojevi kisika,ali i dušika s jednim ili više nesparenih elektrona kojega nastoje uzeti oddrugih spojeva u tijelu životinje što dovodi do oštećenja vitalnih funkcija ži-votinje. Slobodne radikale proizvodi: okolina (previsoka temperatura i vlaga,onečišćenje, radijacija), hrana (oksidirane nezasićene masne kiseline, malooksidansa, previše željeza, previše mikotoksina i drugih štetnih tvari) i or-ganizam životinje (bakterije, virusi i obrambena tijela). Slobodni radikaliuključeni su u nastanak i razvoj brojnih bolesti srca, pluća, bubrega, živacai drugih organa ljudi i životinja.
Životinju od stalnog napada slobodnih radikala štite antioksidansi koji ne-utraliziraju slobodne radikale.
Antioksidacijska sposobnost je pokazatelj zaštitnog djelovanja zrna ku-kuruza protiv opasnih oksidacija u organizmu.
Zrnjevlje cjelovitih žitarica sadržava jednako ili više antioksidansa negovoće i povrće. Kukuruz sadržava ne samo hranjive tvari već i antioksidansekoji povoljno djeluju na kvalitetu hrane i zdravlje životinja. Kukuruz ima najvišuantioksidacijsku aktivnost od svih žitarica (181,42 ± 0,86 μmol vitamin Cekvivalenta/g), slijede pšenica (76,70 ± 1,38 μmol vitamin C ekvivalenta/g),zob (74,67±1,49 μmol vitamin C ekvivalenta/g), riža (55,77 ± 1,62 μmol vi-tamin C ekvivalenta/g). Od brojnih prirodnih antioksidansa, kukuruz sadržavapo jačini djelovanja fenole, karotenoide i tokole (vitamin E). Među njima fenolinajviše (> 85%) pridonose antioksidacijskoj aktivnosti zrna kukuruza
Antioksidacijska moć heksanskog ekstrakta karotenoida snažno je pove-zana sa caklavošću jer su karotenoidi smješteni u endospermu (tablica 31.).
Međutim, antioksidansi se tijekom skladištenja troše pa je prikazana an-tioksidacijska moć zrna Bc hibrida kukuruza nakon jedne godine skladište-nja. Teško je procijeniti koliko se antioksidansa troši tijekom skladištenja
Prasad Tov Krmače Brojleri Pilići Kokoši
16 11 44 20 10 10
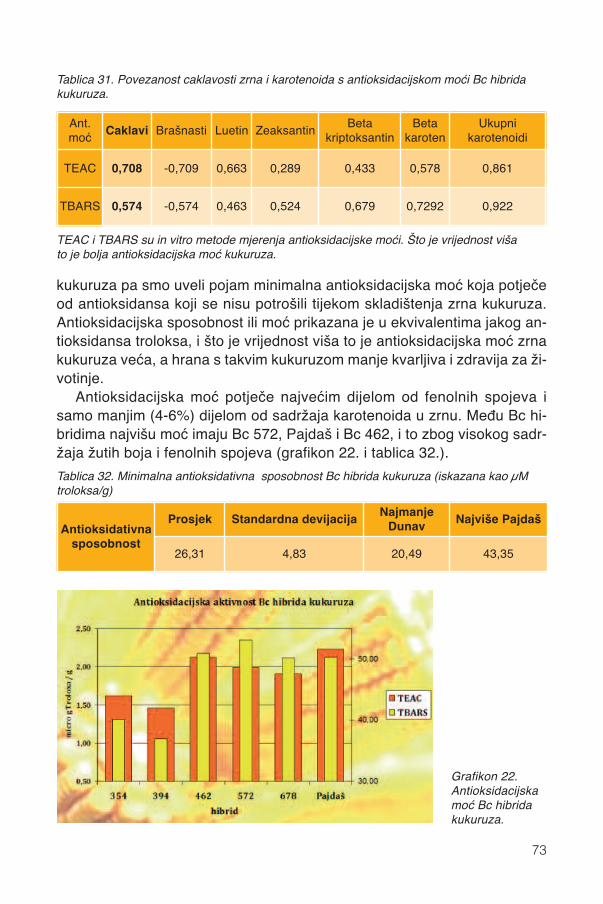
73
kukuruza pa smo uveli pojam minimalna antioksidacijska moć koja potječeod antioksidansa koji se nisu potrošili tijekom skladištenja zrna kukuruza.Antioksidacijska sposobnost ili moć prikazana je u ekvivalentima jakog an-tioksidansa troloksa, i što je vrijednost viša to je antioksidacijska moć zrnakukuruza veća, a hrana s takvim kukuruzom manje kvarljiva i zdravija za ži-votinje.
Antioksidacijska moć potječe najvećim dijelom od fenolnih spojeva isamo manjim (4-6%) dijelom od sadržaja karotenoida u zrnu. Među Bc hi-bridima najvišu moć imaju Bc 572, Pajdaš i Bc 462, i to zbog visokog sadr-žaja žutih boja i fenolnih spojeva (grafikon 22. i tablica 32.).
Tablica 31. Povezanost caklavosti zrna i karotenoida s antioksidacijskom moći Bc hibridakukuruza.
Ant.moć Caklavi Brašnasti Luetin Zeaksantin Beta
kriptoksantinBeta
karotenUkupni
karotenoidi
TEAC 0,708 -0,709 0,663 0,289 0,433 0,578 0,861
TBARS 0,574 -0,574 0,463 0,524 0,679 0,7292 0,922
TEAC i TBARS su in vitro metode mjerenja antioksidacijske moći. Što je vrijednost višato je bolja antioksidacijska moć kukuruza.
Grafikon 22.Antioksidacijskamoć Bc hibridakukuruza.
Tablica 32. Minimalna antioksidativna sposobnost Bc hibrida kukuruza (iskazana kao µMtroloksa/g)
Antioksidativnasposobnost
Prosjek Standardna devijacija NajmanjeDunav Najviše Pajdaš
26,31 4,83 20,49 43,35
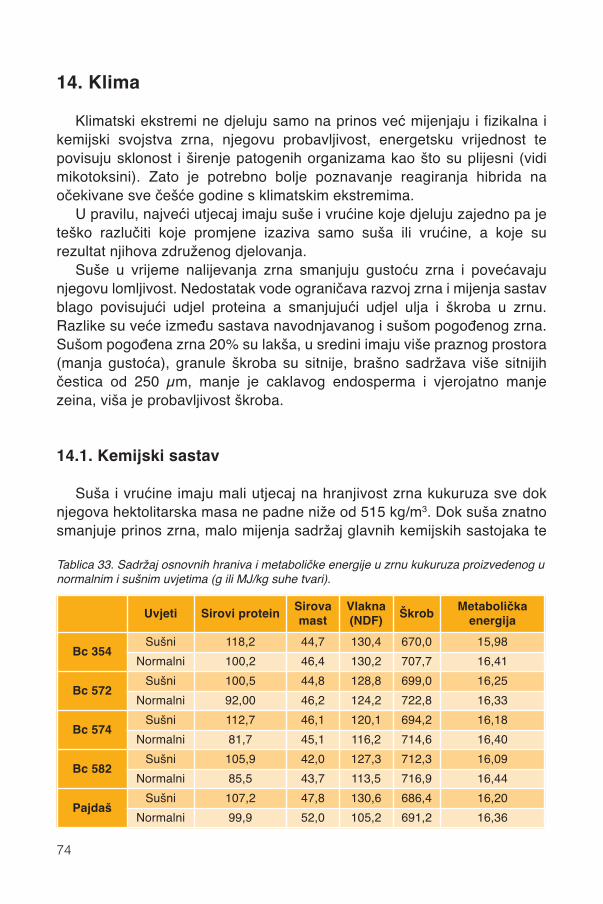
74
14. KlimaKlimatski ekstremi ne djeluju samo na prinos već mijenjaju i fizikalna i
kemijski svojstva zrna, njegovu probavljivost, energetsku vrijednost tepovisuju sklonost i širenje patogenih organizama kao što su plijesni (vidimikotoksini). Zato je potrebno bolje poznavanje reagiranja hibrida naočekivane sve češće godine s klimatskim ekstremima.
U pravilu, najveći utjecaj imaju suše i vrućine koje djeluju zajedno pa jeteško razlučiti koje promjene izaziva samo suša ili vrućine, a koje surezultat njihova združenog djelovanja.
Suše u vrijeme nalijevanja zrna smanjuju gustoću zrna i povećavajunjegovu lomljivost. Nedostatak vode ograničava razvoj zrna i mijenja sastavblago povisujući udjel proteina a smanjujući udjel ulja i škroba u zrnu.Razlike su veće između sastava navodnjavanog i sušom pogođenog zrna.Sušom pogođena zrna 20% su lakša, u sredini imaju više praznog prostora(manja gustoća), granule škroba su sitnije, brašno sadržava više sitnijihčestica od 250 µm, manje je caklavog endosperma i vjerojatno manjezeina, viša je probavljivost škroba.
14.1. Kemijski sastav
Suša i vrućine imaju mali utjecaj na hranjivost zrna kukuruza sve doknjegova hektolitarska masa ne padne niže od 515 kg/m3. Dok suša znatnosmanjuje prinos zrna, malo mijenja sadržaj glavnih kemijskih sastojaka te
Tablica 33. Sadržaj osnovnih hraniva i metaboličke energije u zrnu kukuruza proizvedenog unormalnim i sušnim uvjetima (g ili MJ/kg suhe tvari).
Uvjeti Sirovi protein Sirovamast
Vlakna(NDF) Škrob Metabolička
energija
Bc 354 Sušni 118,2 44,7 130,4 670,0 15,98Normalni 100,2 46,4 130,2 707,7 16,41
Bc 572 Sušni 100,5 44,8 128,8 699,0 16,25Normalni 92,00 46,2 124,2 722,8 16,33
Bc 574 Sušni 112,7 46,1 120,1 694,2 16,18Normalni 81,7 45,1 116,2 714,6 16,40
Bc 582 Sušni 105,9 42,0 127,3 712,3 16,09Normalni 85,5 43,7 113,5 716,9 16,44
Pajdaš Sušni 107,2 47,8 130,6 686,4 16,20Normalni 99,9 52,0 105,2 691,2 16,36
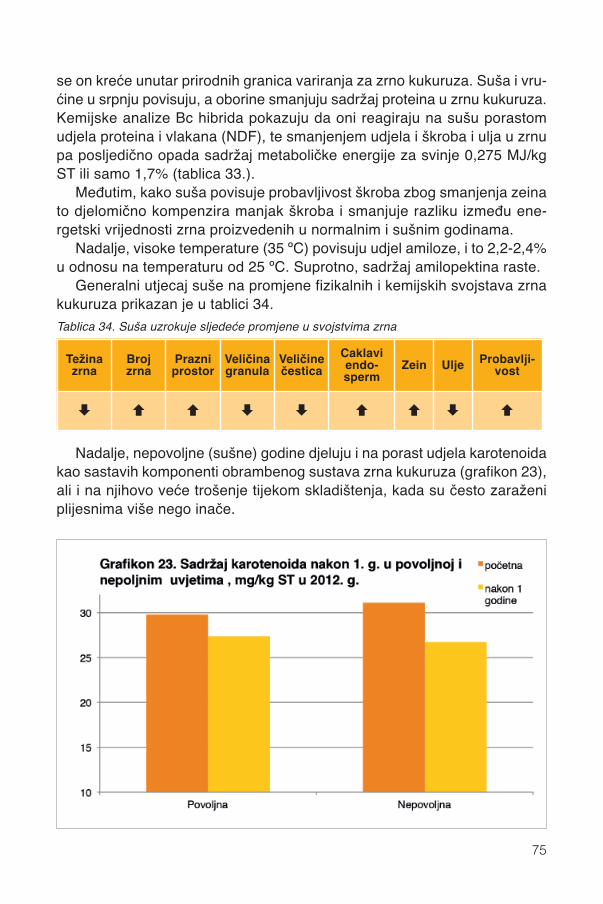
75
se on kreće unutar prirodnih granica variranja za zrno kukuruza. Suša i vru-ćine u srpnju povisuju, a oborine smanjuju sadržaj proteina u zrnu kukuruza.Kemijske analize Bc hibrida pokazuju da oni reagiraju na sušu porastomudjela proteina i vlakana (NDF), te smanjenjem udjela i škroba i ulja u zrnupa posljedično opada sadržaj metaboličke energije za svinje 0,275 MJ/kgST ili samo 1,7% (tablica 33.).
Međutim, kako suša povisuje probavljivost škroba zbog smanjenja zeinato djelomično kompenzira manjak škroba i smanjuje razliku između ene-rgetski vrijednosti zrna proizvedenih u normalnim i sušnim godinama.
Nadalje, visoke temperature (35 ºC) povisuju udjel amiloze, i to 2,2-2,4%u odnosu na temperaturu od 25 ºC. Suprotno, sadržaj amilopektina raste.
Generalni utjecaj suše na promjene fizikalnih i kemijskih svojstava zrnakukuruza prikazan je u tablici 34.
Nadalje, nepovoljne (sušne) godine djeluju i na porast udjela karotenoidakao sastavih komponenti obrambenog sustava zrna kukuruza (grafikon 23),ali i na njihovo veće trošenje tijekom skladištenja, kada su često zaraženiplijesnima više nego inače.
Tablica 34. Suša uzrokuje sljedeće promjene u svojstvima zrna
Težinazrna
Broj zrna
Prazniprostor
Veličinagranula
Veličinečestica
Caklavi endo-sperm
Zein Ulje Probavlji-vost
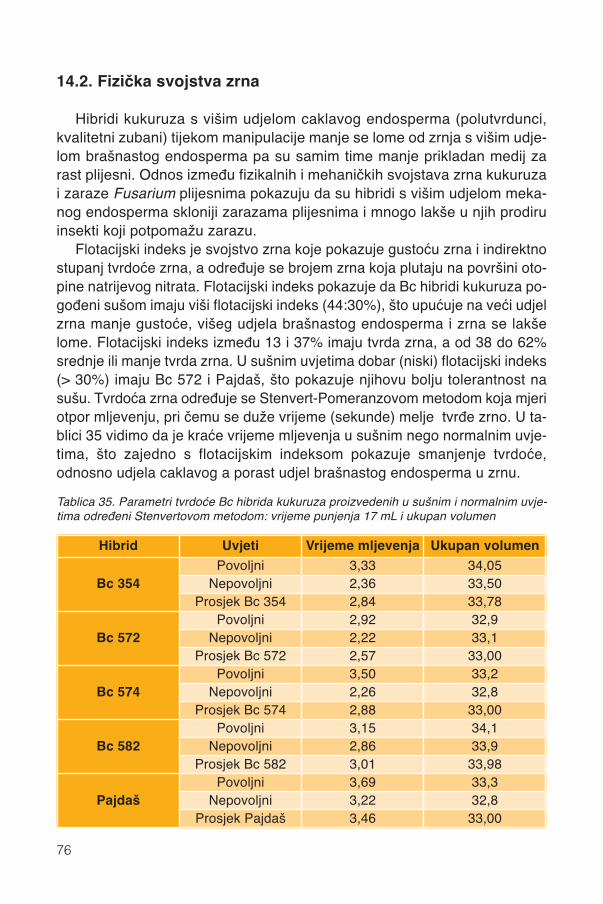
76
14.2. Fizička svojstva zrna
Hibridi kukuruza s višim udjelom caklavog endosperma (polutvrdunci,kvalitetni zubani) tijekom manipulacije manje se lome od zrnja s višim udje-lom brašnastog endosperma pa su samim time manje prikladan medij zarast plijesni. Odnos između fizikalnih i mehaničkih svojstava zrna kukuruzai zaraze Fusarium plijesnima pokazuju da su hibridi s višim udjelom meka-nog endosperma skloniji zarazama plijesnima i mnogo lakše u njih prodiruinsekti koji potpomažu zarazu.
Flotacijski indeks je svojstvo zrna koje pokazuje gustoću zrna i indirektnostupanj tvrdoće zrna, a određuje se brojem zrna koja plutaju na površini oto-pine natrijevog nitrata. Flotacijski indeks pokazuje da Bc hibridi kukuruza po-gođeni sušom imaju viši flotacijski indeks (44:30%), što upućuje na veći udjelzrna manje gustoće, višeg udjela brašnastog endosperma i zrna se lakšelome. Flotacijski indeks između 13 i 37% imaju tvrda zrna, a od 38 do 62%srednje ili manje tvrda zrna. U sušnim uvjetima dobar (niski) flotacijski indeks(> 30%) imaju Bc 572 i Pajdaš, što pokazuje njihovu bolju tolerantnost nasušu. Tvrdoća zrna određuje se Stenvert-Pomeranzovom metodom koja mjeriotpor mljevenju, pri čemu se duže vrijeme (sekunde) melje tvrđe zrno. U ta-blici 35 vidimo da je kraće vrijeme mljevenja u sušnim nego normalnim uvje-tima, što zajedno s flotacijskim indeksom pokazuje smanjenje tvrdoće,odnosno udjela caklavog a porast udjel brašnastog endosperma u zrnu.Tablica 35. Parametri tvrdoće Bc hibrida kukuruza proizvedenih u sušnim i normalnim uvje-tima određeni Stenvertovom metodom: vrijeme punjenja 17 mL i ukupan volumen
Hibrid Uvjeti Vrijeme mljevenja Ukupan volumen
Bc 354Povoljni 3,33 34,05
Nepovoljni 2,36 33,50Prosjek Bc 354 2,84 33,78
Bc 572Povoljni 2,92 32,9
Nepovoljni 2,22 33,1Prosjek Bc 572 2,57 33,00
Bc 574Povoljni 3,50 33,2
Nepovoljni 2,26 32,8Prosjek Bc 574 2,88 33,00
Bc 582Povoljni 3,15 34,1
Nepovoljni 2,86 33,9Prosjek Bc 582 3,01 33,98
PajdašPovoljni 3,69 33,3
Nepovoljni 3,22 32,8Prosjek Pajdaš 3,46 33,00

77
Suša smanjuje prinos i udjel zrna, ali ne uvijek i hranjivost vegetativnogdijela biljke kukuruza. Silaža kukuruza sadržava manje zrna, ali je viši sa-držaj proteina, osobito zeina, frakcija vlakana i šećera. Kako suša smanjujeudjel lignina u stabljici, tako se povisuje 0,5-1,5% probavljivost vlakana(NDF) u silaži kukuruza. Suša, ovisno o količini gnojidbe i stadiju razvojabiljke kada je djelovala često povisuje razinu nitrata do granica opasnih zapreživače.
Suša i vrućine združeno, snažno povisuju rizik infekcija zrna patogenimsojevima plijesni Aspergillus flavus koje proizvode aflatoksin i Fusarium ver-ticillioides koje proizvode fumonisin.

78
15. Mikotoksini
15.1. Uvod
Procjenjuje se da je 25% svjetske proizvodnje hrane zaraženo mikoto-ksinima, a čak 72% krme sadržava detektibilne koncentracije mikotoksinakoji su jedan od glavnih javnozdravstvenih problema u svijetu u 21. stoljeću.Tri su najvažnija roda mikotoksigene plijesni Aspergillus, Fusarium (13vrsta) i Penicillium koje proizvode četiri najčešća mikotoksina: aflatoksin(Aspergillus), ohratoksin (Aspergillus i Pencillium), trihotocene i fumonisin(Fusarium). Godišnji gubici procjenjuju se na 190 milijuna $ od aflatoksinai 40 milijuna $ od fumonisina u SAD-u. Kukuruz je najmanje otporan na za-razu plijesnima i posljedično kontaminaciji mikotoksinima u RH.
Zdravstvena sigurnost hrane osnovni je preduvjet njezina korištenja uEU. Zdravstveni su rizici ukupnih mikotoksina visoki, a samo višim dozamaaflatoksina izloženo je 55 milijuna ljudi u svijetu. Sve viša razina svijesti omikotoksinima i prijetnje koje oni predstavljaju za zdravlje životinja i naročitoljudi snažno usmjerava zahtjeve javnosti na zakonsko smanjenje njihovekoncentracije na najmanju moguću mjeru, te tjera poljoprivrednika u EU upravcu iznalaženja i uvođenja mjera ublažavanja njihovih nepovoljnih uči-naka. Mikotoksini iz hrane za životinje neposredna su opasnost za životinjui posredna za ljude koji konzumiraju animalne proizvode (mlijeko, meso,jaja).
15.2. Klimatski ekstremi
Nema načina za potpuno sprečavanje pojave plijesni na hrani niti lijekaprotiv mikotoksina pa su oni najveća i najraširenija opasnost hrane za živo-tinje za zdravlje ljudi i životinja. Stoga je presudan holistički pristup smanje-nja njihove koncentracije putem (1) smanjenog razvoja patogenih plijesni iprodukcije mikotoksina na kukuruzu tijekom cijelog procesa proizvodnje odpolja do valova, (2) uklanjanja stvorenog mikotoksina s krme ili (3) iz pro-bavnog trakta životinje.
Epizode suša koje se izmjenjuju s visokim vlagama > 75%, te tempera-ture >12 i < 32 ºC najhitniji su čimbenici koji pogoduju infekciji patogenim

79
sojevima plijesni. Porast koncentracije CO2 u atmosferi sa 300 na 7000 ili1000 ppm treći je pogodbeni čimbenik razvoja toksikogenih plijesni (grafikon24.).
Očekivani klimatski ekstremi u sjevernoj Europi, više temperature i za30-40%, češće oborine te u jugozapadnoj Europi uključujući i RH vrućine isuše, osobito ljeti, pogodovat će višoj kontaminaciji aflatoksinom i fumoni-sinom. Među žitaricama kukuruz je najizloženiji djelovanju suše/oborina ivrućina koje u svakom koraku proizvodnje kukuruza znatno utječu na pojavumikotoksina. Stres suše djeluje na tri načina: kao prvo smanjuje prirodnuobranu biljke od infekcija, kao drugo smanjenje aktivnosti vode tla posljedi-čno smanjuje rast i aktivnost bakterija, ameba i kompetitivnih gljiva, i kaotreće potpomaže rast kserofilnih plijesni A. flavus i A. parasiticus.
Grafikon 24. Utjecaj različitih biotskih i abiotskih čimbenika na ekofiziologiju plijesni
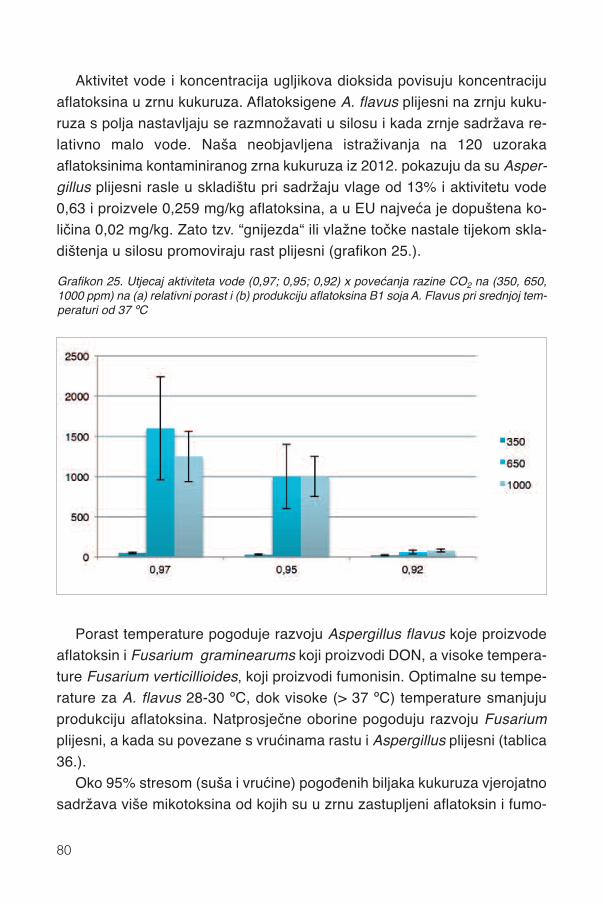
80
Aktivitet vode i koncentracija ugljikova dioksida povisuju koncentracijuaflatoksina u zrnu kukuruza. Aflatoksigene A. flavus plijesni na zrnju kuku-ruza s polja nastavljaju se razmnožavati u silosu i kada zrnje sadržava re-lativno malo vode. Naša neobjavljena istraživanja na 120 uzorakaaflatoksinima kontaminiranog zrna kukuruza iz 2012. pokazuju da su Asper-gillus plijesni rasle u skladištu pri sadržaju vlage od 13% i aktivitetu vode0,63 i proizvele 0,259 mg/kg aflatoksina, a u EU najveća je dopuštena ko-ličina 0,02 mg/kg. Zato tzv. “gnijezda“ ili vlažne točke nastale tijekom skla-dištenja u silosu promoviraju rast plijesni (grafikon 25.).
Porast temperature pogoduje razvoju Aspergillus flavus koje proizvodeaflatoksin i Fusarium graminearums koji proizvodi DON, a visoke tempera-ture Fusarium verticillioides, koji proizvodi fumonisin. Optimalne su tempe-rature za A. flavus 28-30 ºC, dok visoke (> 37 ºC) temperature smanjujuprodukciju aflatoksina. Natprosječne oborine pogoduju razvoju Fusariumplijesni, a kada su povezane s vrućinama rastu i Aspergillus plijesni (tablica36.).
Oko 95% stresom (suša i vrućine) pogođenih biljaka kukuruza vjerojatnosadržava više mikotoksina od kojih su u zrnu zastupljeni aflatoksin i fumo-
Grafikon 25. Utjecaj aktiviteta vode (0,97; 0,95; 0,92) x povećanja razine CO2 na (350, 650,1000 ppm) na (a) relativni porast i (b) produkciju aflatoksina B1 soja A. Flavus pri srednjoj tem-peraturi od 37 ºC

81
nisin B1, a u silaži otrovi Pencillium plijesni. Ovi mikotoksini smanjuju mli-ječnost, narušavaju zdravlje i reprodukciju životinja.
15.3. Kukuruz
Utvrđena je velika razlika između kultivara i hibrida kukuruza u sklonostizarazi Fusarium plijesnima. U kultivarima otpornima na A. flavus pronađenisu antifungalni proteini i peptidi koji se sintetiziraju prije zaraze te tako spre-čavaju razvoj aflatoksigene plijesni, i to u svili, komušini i endospermu. Upravilu hibridi kasnije skupine sazrijevanja skloniji su zarazi Fusarium plije-snima. Kako se zrno zarazi fumonisinom u kasnijim stadijima sazrijevanja(voštanoj zriobi), njegovo ranija berba (> 25% vlage) znatno pomaže pri te-škim zarazama. Isto tako infestacije insektima potpomažu razvoj Fusariumplijesni na zrnu kukuruza pa tako 90% zrna zaraženih kukuruznim moljcemsadržava fumonisin.
Pojava mikotoksina ovisi i o kemijskom sastavu stadija razvoja zrna ku-kuruza. Tako se proizvede više aflatoksina na zrnima u mliječnoj zrelosti,kada su bogata jednostavnim šećerima, a manje na zrnima u caklavoj fazirazvoja kada su bogata složenim ugljikohidratima kao što je škrob u formiamilopektina. Naime, stres uzrokovan sušom usporava sintezu škroba, aline i jednostavnih šećera, pa njihova viša prisutnost u zrnu pogoduje razvoju
Tablica 36. Povoljni uvjeti rasta glavnih plijesni i mikotoksini na kukuruzu
Povoljna okolina Mikotoksin Mikotoksigene plijesni
Topla i suha sezonarasta
Vlažno prije berbe ili nakon berbe
Aflatoksini (B1,B2, G1, G2 i M1 Aspergillus flavus Aspergillus parasiticus
Važno i vrlo toplo Fumonizin Fusarium verticilioidesFusarium proliferatum
Vruće, vlažno i kišovito vrijeme
Deoksinivalenol Zearalenon Fusarium graminearum

82
Fusarium ili Aspergillus plijesni te potencijalnom nakupljanju aflatoksina ifumonisina.
Kada se promatra građa škroba, tada je amilopektin za produkciju afla-toksina pogodniji od amiloze. Naime, Aspergillus flavus plijesni koje proiz-vode aflatoksin imaju znatno veću aktivnost α-amilaze od netoksinogenihsojeva. Stoga selekcija na višu probavljivost dovodi do veće osjetljivosti naprodukciju mikotoksina. Isto tako i sadržaj antioksidansa pomaže zrnu u su-stavu otpornosti na razvoj plijesni i kontaminaciju mikotoksinima.
15.4. Bc hibridi u sušnoj 2012.
Europska agencija za hranu procjenjuje da će u našem klimatskom po-dručju temperature porasti za 2-3 ºC te će se javiti umjereno češće, a akotemperatura poraste za 5 ºC vrlo česte pojave viših od zakonski minimalnodopuštenih koncentracija aflatoksina. Smatramo da će biti sve više kuku-ruza zaraženog i ostalim mikotoksinima, ali u najčešćim koncentracijamamanjim od zakonom minimalno dopuštenim. Aflatoksin je visoko toksičanmikotoksin za ljude i životinje. Stoga u ovom poglavlju iznosimo rezultatepraćenja utjecaja suše na razvoj mikotoksina, osobito aflatoksina, i to napet Bc hibrida kukuruza.
Suša mnogostruko povisuje koncentraciju plijesni na zrnu kukuruza (ta-blica 37). Stres suše djeluje na tri načina: prvo smanjuje prirodnu obranubiljke od infekcija, drugo smanjuje aktivnosti vode tla te posljedično sma-njuje i rast i aktivnost bakterija, ameba i kompetitivnih gljiva, i treće potpo-maže rast kserofilnih plijesni A. flavus i A. parasiticus. Stoga mnogiznanstvenici smatraju da se bez navodnjavanja kukuruza i ostalih predžet-venih mjera teško sprječava razvoj toksigenih Aspergillus plijesni.
Tablica 37. Broj plijesni i kvasca na normalnoj i suhoj lokaciji (cfu*/g)
Hibrid Bc 354 Bc 572 Bc 574 Bc 582 Pajdaš
Normalno 20 200 8 800 30 320 78 800 14 000
Suša 460 000 236 600 309 000 478 000 628 000
Gornja dopuštena koncentracija plijesni u 1 g biljne hrane za životinje je 200 000
*cfu = colony forming unit
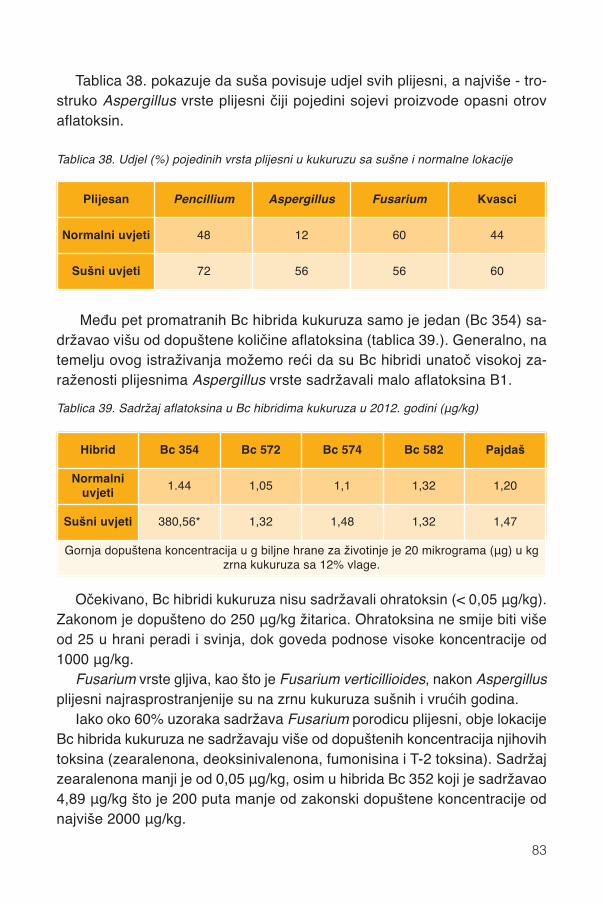
83
Tablica 38. pokazuje da suša povisuje udjel svih plijesni, a najviše - tro-struko Aspergillus vrste plijesni čiji pojedini sojevi proizvode opasni otrovaflatoksin.
Među pet promatranih Bc hibrida kukuruza samo je jedan (Bc 354) sa-državao višu od dopuštene količine aflatoksina (tablica 39.). Generalno, natemelju ovog istraživanja možemo reći da su Bc hibridi unatoč visokoj za-raženosti plijesnima Aspergillus vrste sadržavali malo aflatoksina B1.
Očekivano, Bc hibridi kukuruza nisu sadržavali ohratoksin (< 0,05 μg/kg).Zakonom je dopušteno do 250 μg/kg žitarica. Ohratoksina ne smije biti višeod 25 u hrani peradi i svinja, dok goveda podnose visoke koncentracije od1000 μg/kg.
Fusarium vrste gljiva, kao što je Fusarium verticillioides, nakon Aspergillusplijesni najrasprostranjenije su na zrnu kukuruza sušnih i vrućih godina.
Iako oko 60% uzoraka sadržava Fusarium porodicu plijesni, obje lokacijeBc hibrida kukuruza ne sadržavaju više od dopuštenih koncentracija njihovihtoksina (zearalenona, deoksinivalenona, fumonisina i T-2 toksina). Sadržajzearalenona manji je od 0,05 μg/kg, osim u hibrida Bc 352 koji je sadržavao4,89 μg/kg što je 200 puta manje od zakonski dopuštene koncentracije odnajviše 2000 μg/kg.
Tablica 38. Udjel (%) pojedinih vrsta plijesni u kukuruzu sa sušne i normalne lokacije
Plijesan Pencillium Aspergillus Fusarium Kvasci
Normalni uvjeti 48 12 60 44
Sušni uvjeti 72 56 56 60
Tablica 39. Sadržaj aflatoksina u Bc hibridima kukuruza u 2012. godini (μg/kg)
Hibrid Bc 354 Bc 572 Bc 574 Bc 582 Pajdaš
Normalniuvjeti 1.44 1,05 1,1 1,32 1,20
Sušni uvjeti 380,56* 1,32 1,48 1,32 1,47
Gornja dopuštena koncentracija u g biljne hrane za životinje je 20 mikrograma (μg) u kgzrna kukuruza sa 12% vlage.
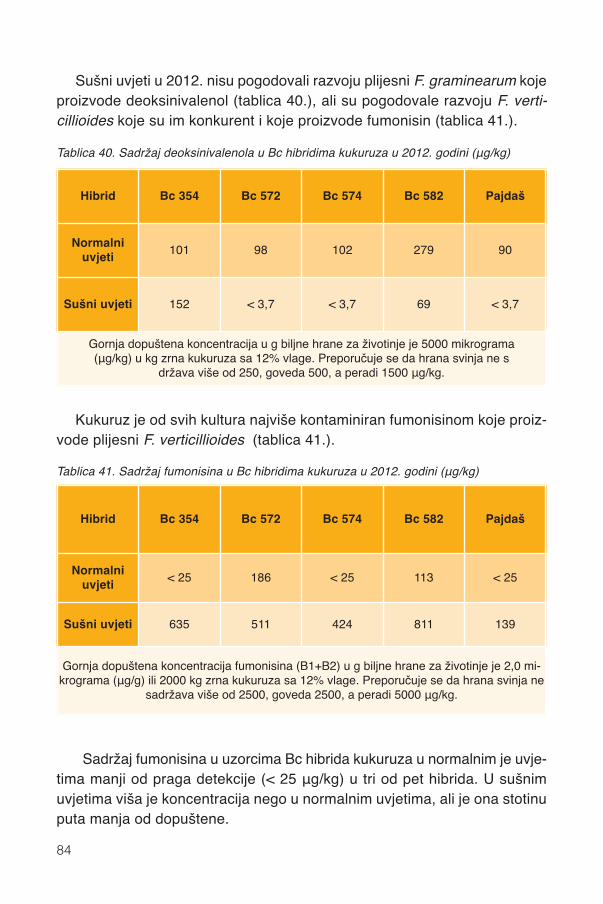
84
Sušni uvjeti u 2012. nisu pogodovali razvoju plijesni F. graminearum kojeproizvode deoksinivalenol (tablica 40.), ali su pogodovale razvoju F. verti-cillioides koje su im konkurent i koje proizvode fumonisin (tablica 41.).
Kukuruz je od svih kultura najviše kontaminiran fumonisinom koje proiz-vode plijesni F. verticillioides (tablica 41.).
Sadržaj fumonisina u uzorcima Bc hibrida kukuruza u normalnim je uvje-tima manji od praga detekcije (< 25 μg/kg) u tri od pet hibrida. U sušnimuvjetima viša je koncentracija nego u normalnim uvjetima, ali je ona stotinuputa manja od dopuštene.
Tablica 40. Sadržaj deoksinivalenola u Bc hibridima kukuruza u 2012. godini (μg/kg)
Hibrid Bc 354 Bc 572 Bc 574 Bc 582 Pajdaš
Normalniuvjeti 101 98 102 279 90
Sušni uvjeti 152 < 3,7 < 3,7 69 < 3,7
Gornja dopuštena koncentracija u g biljne hrane za životinje je 5000 mikrograma(μg/kg) u kg zrna kukuruza sa 12% vlage. Preporučuje se da hrana svinja ne s
država više od 250, goveda 500, a peradi 1500 μg/kg.
Tablica 41. Sadržaj fumonisina u Bc hibridima kukuruza u 2012. godini (μg/kg)
Hibrid Bc 354 Bc 572 Bc 574 Bc 582 Pajdaš
Normalniuvjeti < 25 186 < 25 113 < 25
Sušni uvjeti 635 511 424 811 139
Gornja dopuštena koncentracija fumonisina (B1+B2) u g biljne hrane za životinje je 2,0 mi-krograma (μg/g) ili 2000 kg zrna kukuruza sa 12% vlage. Preporučuje se da hrana svinja ne
sadržava više od 2500, goveda 2500, a peradi 5000 μg/kg.
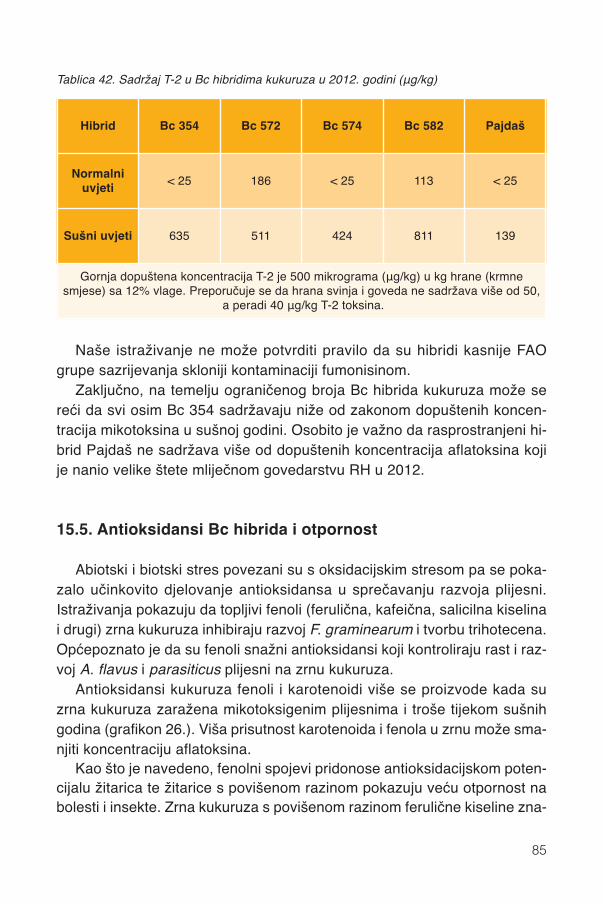
85
Naše istraživanje ne može potvrditi pravilo da su hibridi kasnije FAOgrupe sazrijevanja skloniji kontaminaciji fumonisinom.
Zaključno, na temelju ograničenog broja Bc hibrida kukuruza može sereći da svi osim Bc 354 sadržavaju niže od zakonom dopuštenih koncen-tracija mikotoksina u sušnoj godini. Osobito je važno da rasprostranjeni hi-brid Pajdaš ne sadržava više od dopuštenih koncentracija aflatoksina kojije nanio velike štete mliječnom govedarstvu RH u 2012.
15.5. Antioksidansi Bc hibrida i otpornost
Abiotski i biotski stres povezani su s oksidacijskim stresom pa se poka-zalo učinkovito djelovanje antioksidansa u sprečavanju razvoja plijesni.Istraživanja pokazuju da topljivi fenoli (ferulična, kafeična, salicilna kiselinai drugi) zrna kukuruza inhibiraju razvoj F. graminearum i tvorbu trihotecena.Općepoznato je da su fenoli snažni antioksidansi koji kontroliraju rast i raz-voj A. flavus i parasiticus plijesni na zrnu kukuruza.
Antioksidansi kukuruza fenoli i karotenoidi više se proizvode kada suzrna kukuruza zaražena mikotoksigenim plijesnima i troše tijekom sušnihgodina (grafikon 26.). Viša prisutnost karotenoida i fenola u zrnu može sma-njiti koncentraciju aflatoksina.
Kao što je navedeno, fenolni spojevi pridonose antioksidacijskom poten-cijalu žitarica te žitarice s povišenom razinom pokazuju veću otpornost nabolesti i insekte. Zrna kukuruza s povišenom razinom ferulične kiseline zna-
Tablica 42. Sadržaj T-2 u Bc hibridima kukuruza u 2012. godini (μg/kg)
Hibrid Bc 354 Bc 572 Bc 574 Bc 582 Pajdaš
Normalniuvjeti < 25 186 < 25 113 < 25
Sušni uvjeti 635 511 424 811 139
Gornja dopuštena koncentracija T-2 je 500 mikrograma (μg/kg) u kg hrane (krmnesmjese) sa 12% vlage. Preporučuje se da hrana svinja i goveda ne sadržava više od 50,
a peradi 40 μg/kg T-2 toksina.
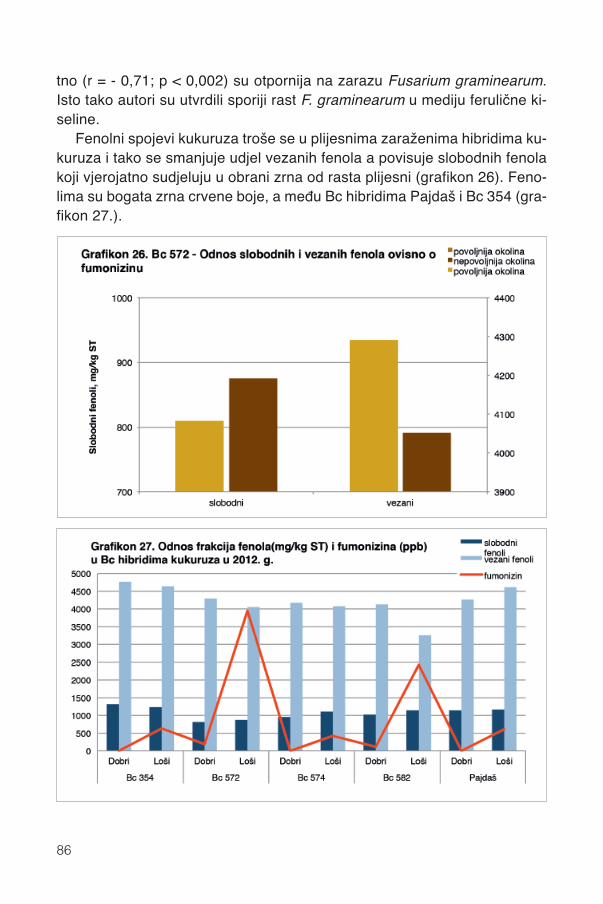
86
tno (r = - 0,71; p < 0,002) su otpornija na zarazu Fusarium graminearum.Isto tako autori su utvrdili sporiji rast F. graminearum u mediju ferulične ki-seline.
Fenolni spojevi kukuruza troše se u plijesnima zaraženima hibridima ku-kuruza i tako se smanjuje udjel vezanih fenola a povisuje slobodnih fenolakoji vjerojatno sudjeluju u obrani zrna od rasta plijesni (grafikon 26). Feno-lima su bogata zrna crvene boje, a među Bc hibridima Pajdaš i Bc 354 (gra-fikon 27.).
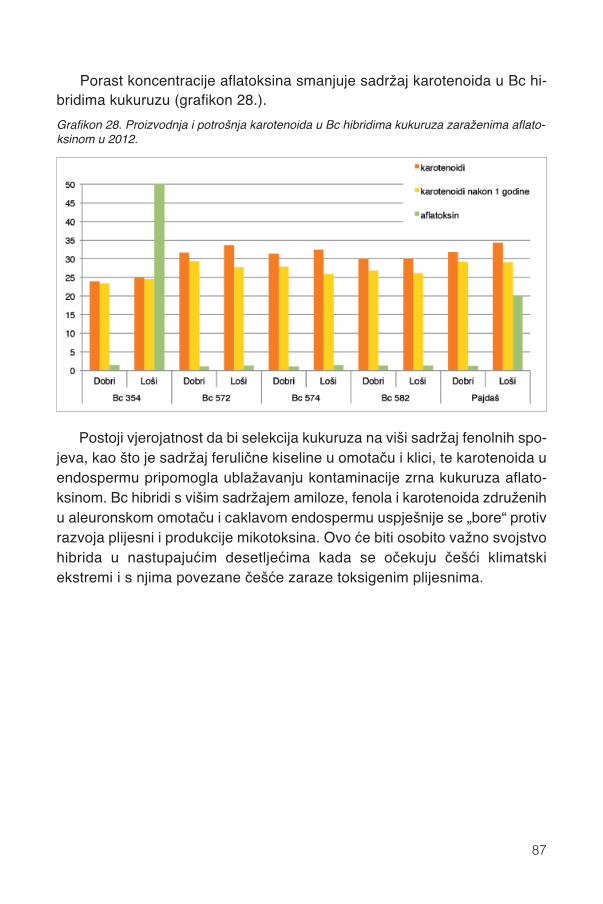
87
Porast koncentracije aflatoksina smanjuje sadržaj karotenoida u Bc hi-bridima kukuruzu (grafikon 28.).
Postoji vjerojatnost da bi selekcija kukuruza na viši sadržaj fenolnih spo-jeva, kao što je sadržaj ferulične kiseline u omotaču i klici, te karotenoida uendospermu pripomogla ublažavanju kontaminacije zrna kukuruza aflato-ksinom. Bc hibridi s višim sadržajem amiloze, fenola i karotenoida združenihu aleuronskom omotaču i caklavom endospermu uspješnije se „bore“ protivrazvoja plijesni i produkcije mikotoksina. Ovo će biti osobito važno svojstvohibrida u nastupajućim desetljećima kada se očekuju češći klimatskiekstremi i s njima povezane češće zaraze toksigenim plijesnima.
Grafikon 28. Proizvodnja i potrošnja karotenoida u Bc hibridima kukuruza zaraženima aflato-ksinom u 2012.

88
16. Mljevenje
Prerada kukuruza znatno mijenja njegova hranidbena svojstva i energet-sku vrijednost. Krupnoća mljevenja različito djeluje na različite vrste životi-nja: Perad bolje iskorištava energiju iz krupno nego fino mljevenogkukuruza, a svinje obrnuto. Funkcija je mljevenja kao najčešćeg oblika pre-rade zrna kukuruza, između ostalog, povećati probavljivost škroba u pro-bavnom traktu i omogućiti njegovu ravnomjernu izmiješanost s ostalimsastojcima krmnih smjesa. Neprimjerena meljava zrna može umanjiti hra-nidbeni potencijal pojedinog hibrida kukuruza. Hibridi s višim udjelom bra-šnastog nego rožnatog endosperma lakše se melju pa se troši manjeenergije. Za smanjenje veličine čestica od 1000 na 600 µm blago raste po-trošnja energije sa 2,7 na 3,8 kW/t, ali daljnje usitnjavanje na 400 µm udvo-stručuje potrošnju energije na 8,1 kW/t. Zato je krupnoća mljevenjakompromis između dobiti ostvarene višom probavljivosti i troškova sitnijegmljevenja.
16.1. Svinje
Krupnoća mljevenja važna je za ekonomičnu svinjogojsku proizvodnju.Ako je zrno krupno mljeveno slabije se iskorištavaju hranjive tvari iz njegapa sporije raste, a ako je brašnasto tada se troši puno energije za mljevenjea brašno može izazvati čireve u svinja. Generalno, svinje iskoriste više MEiz sitno nego krupno mljevenog zrna kukuruza. Vrijedi pravilo - što je svinjamlađa treba joj sitnija meljava za dobro iskorištenje hrane i visoke priraste.Hrana za prasad, osobito ako se peletira, melje se na krupnoću od 0,5 mm,za svinje u tovu na 0,66-0,75 mm, a za krmače na veličinu 0,4-0,6 mm. Čaki kada se hrana peletira nije potrebno mljeti kukuruz sitnije od 675 µm.
Neujednačena veličina čestica dovodi do loše izmiješanosti smjese. Zapravilnu izmiješanost kukuruza s ostalim sirovinama važno je da su česticeujednačene krupnoće. Odstupanja veličine čestica zrna kukuruza od pro-sjeka mjerena koeficijentom varijacije ne bi trebala biti veća od 10%. Zapravilnu izmiješanost smjese trajanje miješanja kukuruza s ostalim kompo-nentama ovisi o vrsti miješalice. Kada se smjese rade u vodoravnoj miješa-lici, tada miješanje traje 15 min., a u okomitoj miješalici 5-10 min. nakondodavanja zadnje komponente.

89
16.2. Perad
Perad bolje iskorištava energiju iz krupno nego sitno mljevenog kuku-ruza. Odabire hranu na temelju njezinih taktilnih (dodirnih) i optičkih svoj-stava, kao što su veličina, oblik i gustoća čestica kukuruza. Kako kukuruzčini 55-60% hrane pilića, njegova taktilna svojstva određuju uzimanje cijelesmjese. Kada se pilići hrane brašnastom hranom kukuruz bi trebalo mljetina krupnoću od 980 do 1170 µm, odnosno tako da je više od 60% česticaveće od 1 mm jer su tada viši prirasti pilića, teži mehanički želudac, manjepatogenih (Clostridium spp., Campylobacterium spp., Bacteroides spp), aviše poželjnih bakterija (Lactobacillus spp. i Bifidobacteria spp.) u slijepomcrijevu. Krupnije mljeveni kukuruz je hrana za poželjne bakterije pa se možesmatrati i prebiotikom. Sitne čestice smanjuju razvoj mišićnog želuca pe-radi, a on je glavni regulator kretanja hrane. Što je kukuruz finije mljeven toje brži prolaz hrane kroz probavni sustav peradi pa je manja probavljivosthranjivih tvari.
Kada se pilići tove peletiranom hranom tada krupnoća kukuruza nemaveći utjecaj na proizvodnju pa se preporučuje krupno mljeveni kukuruz do-dati u krmne smjese nakon peletiranja.
Odlični su rezultati u tovu kada se pilići hrane mješavinom 25% valjanjemrazbijenog zrna kukuruza u tipu kvalitetnih zubana i 75% drobljene peleti-rane smjese.
Preporučuje se kokoši nesilice hraniti krupnije mljevenim krmivima tije-kom toplinskog stresa.
16.3. Preživači
Veća je razlika u razgradnji škroba između sitno i krupno mljevenog zrna(15%) nego hibrida kukuruza (5%). Mljevenje mijenja mjesto probave ku-kuruza u preživača. Što je zrno kukuruza finije mljeveno to je viša razgra-dnja njegova škroba u buragu. Tako je niska (15%) razgradnja cijelog zrnau buragu, a sitno (< 0.34 mm) mljevenog je potpuna ili 100%. Sa svakih 0,1mm većom ili manjom krupnoćom čestica od 1,2 mm smanjuje se ili rasterazgradnja škroba u buragu za 3%. Nadalje, peletranje povisuje razgradnjuškroba sa 60 na 64%, ekstrudiranje na 77%, a flekičanje na 86%.
Sitnijim mljevenjem povisuje se razgradnja u buragu i posljedično viša

90
proizvodnja propionske kiseline i sinteza mikrobnog proteina. Krupnije mlje-venje smanjuje razgradnju u buragu i povisuje dotok škroba u crijeva, a po-sljedično u metabolizam ulazi energijom bogatija glukoza nego propionskakiselina. Isto tako, krupnije mljevenje sprečava pojavu acidoza, nadama ilaminitisa.
Dakle, kada se krave hrane silažom kukuruza i/ili silažom zrna i/ili sitno-zrnim žitaricama, tada životinje dobiju dovoljno razgradljivog škroba pa sedohranjuju krupnije mljevenim suhim zrnom kukuruza.
Krave hranjene obrokom silaža lucerke i trava bogatim visoko razgradlji-vim proteinima (80%) dopunjene sitno mljevenim suhim ili siliranim zrnomdat će više mlijeka nego kada je kukuruz usitnjen valjanjem.
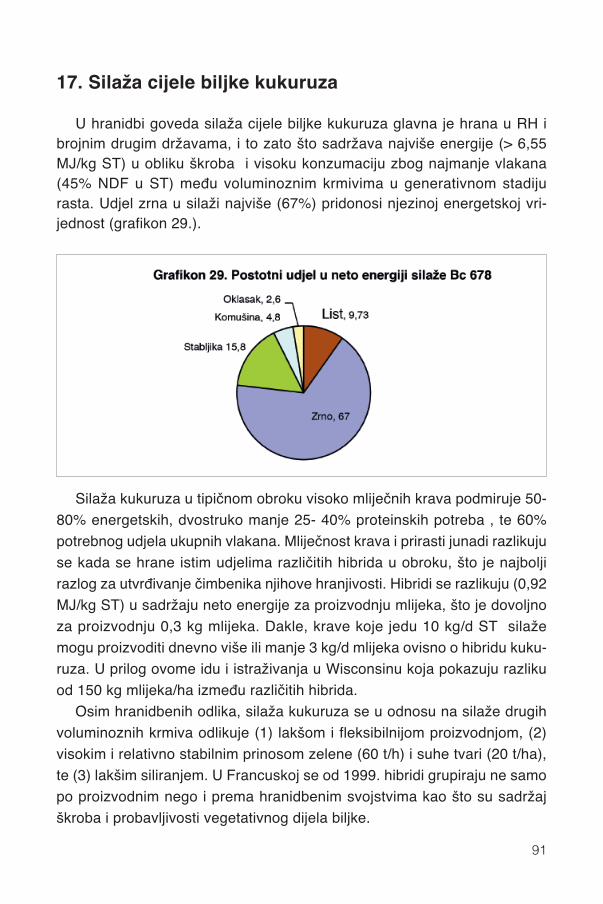
91
17. Silaža cijele biljke kukuruza
U hranidbi goveda silaža cijele biljke kukuruza glavna je hrana u RH ibrojnim drugim državama, i to zato što sadržava najviše energije (> 6,55MJ/kg ST) u obliku škroba i visoku konzumaciju zbog najmanje vlakana(45% NDF u ST) među voluminoznim krmivima u generativnom stadijurasta. Udjel zrna u silaži najviše (67%) pridonosi njezinoj energetskoj vri-jednost (grafikon 29.).
Silaža kukuruza u tipičnom obroku visoko mliječnih krava podmiruje 50-80% energetskih, dvostruko manje 25- 40% proteinskih potreba , te 60%potrebnog udjela ukupnih vlakana. Mliječnost krava i prirasti junadi razlikujuse kada se hrane istim udjelima različitih hibrida u obroku, što je najboljirazlog za utvrđivanje čimbenika njihove hranjivosti. Hibridi se razlikuju (0,92MJ/kg ST) u sadržaju neto energije za proizvodnju mlijeka, što je dovoljnoza proizvodnju 0,3 kg mlijeka. Dakle, krave koje jedu 10 kg/d ST silažemogu proizvoditi dnevno više ili manje 3 kg/d mlijeka ovisno o hibridu kuku-ruza. U prilog ovome idu i istraživanja u Wisconsinu koja pokazuju razlikuod 150 kg mlijeka/ha između različitih hibrida.
Osim hranidbenih odlika, silaža kukuruza se u odnosu na silaže drugihvoluminoznih krmiva odlikuje (1) lakšom i fleksibilnijom proizvodnjom, (2)visokim i relativno stabilnim prinosom zelene (60 t/h) i suhe tvari (20 t/ha),te (3) lakšim siliranjem. U Francuskoj se od 1999. hibridi grupiraju ne samopo proizvodnim nego i prema hranidbenim svojstvima kao što su sadržajškroba i probavljivosti vegetativnog dijela biljke.

92
Kako silaža kukuruza može biti energetsko i vlaknasto krmivo, odabir hi-brida za siliranje određuje potreba krava za energijom ili vlaknima. Pri do-statnoj proizvodnji voluminozne krme sijemo hibride s visokim udjelomškroba i probavljivog lista, a pri manjku voluminozne krme koristimo hibrides manje zrna ali s dosta probavljive stabljike i lista.
Hranjivost silaže kukuruza određena je trima bitnim čimbenicima: (1) sta-dijem zrelosti biljke, (2) kvalitetom siliranja i vremenom korištenja, te (3) hi-bridom kukuruza.
17.1. Stadij zrelosti
Kako je sadržaj energije i vlakana najviše ovisan o stadiju rasta kukuru-zne biljke, tako je utvrđeno da su oni najviši pri sadržaju suhe tvari 32-35%kada biljka sadržava isto toliko škroba. Navedeni sadržaj suhe tvari kom-promis je između prinosa zrna i dobre probavljivosti stabljike i lista, te dobresilažnosti biljke što rezultira viskom konzumacijom energije iz silaže kuku-ruza a time i višom mliječnošću krava (tablica 43.).
Sadržaj suhe tvari najjednostavnije se procijeni iz stadija zrelosti zrna ubiljci kukuruza namijenjenoj siliranju (tablica 44.).
Tablica 43. Odnos između sadržaja suhe tvari silaže kukuruza i mliječnosti krava
Sadržaj suhetvari, % 20 25 30 35
Konzumacijasuhe tvari, kg/d 10,5 11,5 13,0 15,5
Mliječnost, kg/d 9,0 12,5 15,5 20,0
Izvor: INRA, 1989. Ruminant Nutrition.
Tablica 44. Stadiji zrelosti zrna i sadržaj suhe tvari u cijeloj biljci kukuruza
Stadij zrelostizrna Mliječni Tjestasti Tjestasto
-caklavi Caklavi
% suhe tvaribiljke < 25 27 -29 30 -35 >40
Izvor: Carpentier i Cabon, 2011.
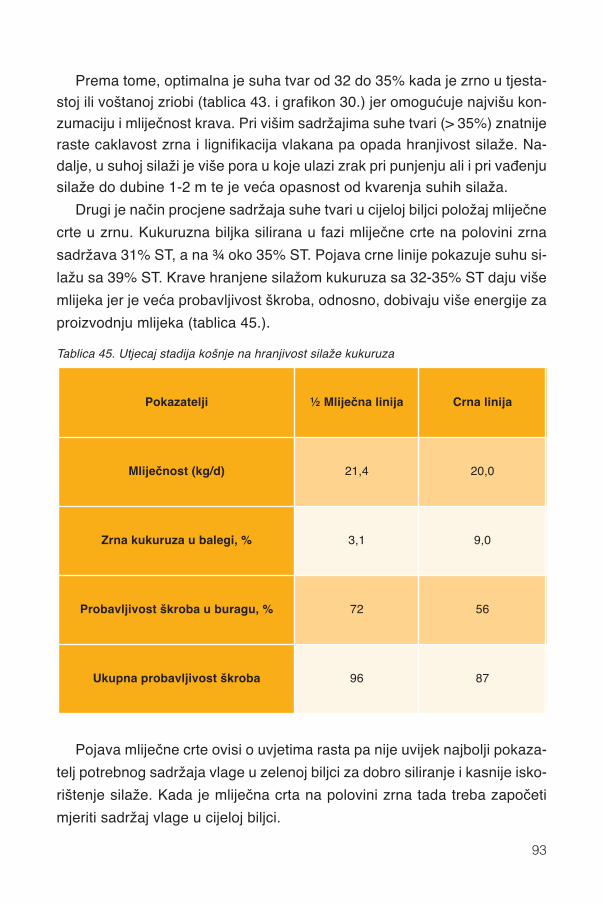
93
Prema tome, optimalna je suha tvar od 32 do 35% kada je zrno u tjesta-stoj ili voštanoj zriobi (tablica 43. i grafikon 30.) jer omogućuje najvišu kon-zumaciju i mliječnost krava. Pri višim sadržajima suhe tvari (> 35%) znatnijeraste caklavost zrna i lignifikacija vlakana pa opada hranjivost silaže. Na-dalje, u suhoj silaži je više pora u koje ulazi zrak pri punjenju ali i pri vađenjusilaže do dubine 1-2 m te je veća opasnost od kvarenja suhih silaža.
Drugi je način procjene sadržaja suhe tvari u cijeloj biljci položaj mliječnecrte u zrnu. Kukuruzna biljka silirana u fazi mliječne crte na polovini zrnasadržava 31% ST, a na ¾ oko 35% ST. Pojava crne linije pokazuje suhu si-lažu sa 39% ST. Krave hranjene silažom kukuruza sa 32-35% ST daju višemlijeka jer je veća probavljivost škroba, odnosno, dobivaju više energije zaproizvodnju mlijeka (tablica 45.).
Pojava mliječne crte ovisi o uvjetima rasta pa nije uvijek najbolji pokaza-telj potrebnog sadržaja vlage u zelenoj biljci za dobro siliranje i kasnije isko-rištenje silaže. Kada je mliječna crta na polovini zrna tada treba započetimjeriti sadržaj vlage u cijeloj biljci.
Tablica 45. Utjecaj stadija košnje na hranjivost silaže kukuruza
Pokazatelji ½ Mliječna linija Crna linija
Mliječnost (kg/d) 21,4 20,0
Zrna kukuruza u balegi, % 3,1 9,0
Probavljivost škroba u buragu, % 72 56
Ukupna probavljivost škroba 96 87

94
17.1.1. Promjene kemijskog sastava
Sazrijevanjem od mliječno-voštane (< 30% ST) do caklave zrelosti (35%ST) zrna povisuje se sadržaj suhe tvari i u njoj udjel škroba, a smanjuje sa-držaj vlakana zbog porasta udjela zrna, odnosno klipa sa 55 na 67% u ST.Tako primjerice sa sazrijevanjem biljke od 28 do 41% suhe tvari opada sa-držaj vlakana za oko 7%, a dvostruko (oko 14%) raste sadržaj škroba pazato raste sadržaj neto energije. Isto tako sa sazrijevanjem od mliječne docaklave zriobe zrna opada sadržaj sirovog proteina sa 12 na 8%, u prosjekuiznosi 7,5% u ST.
Mlade biljke u mliječnoj zriobi sadržavaju 20% topljivih šećera i njihovsadržaj drastično opada na samo 5% u silaži caklavog zrna. Sadržaj šećeratakođer proporcionalno (r = 0,77) opada s porastom udjela škroba tijekomsazrijevanja biljke kukuruza. Šećeri su bitni za dobro siliranje zelene biljkekukuruza.
Lipidi silaže potječu iz zrna i njihov je maleni udjel u biljci 2,5-3,0% panemaju znatan utjecaj na energetsku vrijednost silaže.
Silaža kukuruza pri 32% ST sadržava i malo (4-5%) pepela. Pepeo jeviši u mliječnoj nego caklavoj zriobi i njegova koncentracije tijekom sazrije-vanja biljke opada vrlo malo (0,5-2%). Visoka energetska vrijednost silažeobjašnjava se visokim sadržajem (95%) organske tvari koja je izvor energije.Zbog niskog sadržaja minerala, obroke sa silažom kukuruza treba primje-reno dopuniti mineralnim smjesama.
Visoki sadržaj pepela loš je ne samo zato što smanjuje energetsku vri-
Grafikon 30. Položaj mliječne crte pokazuje sadržaj suhe tvari u biljci kukuruza
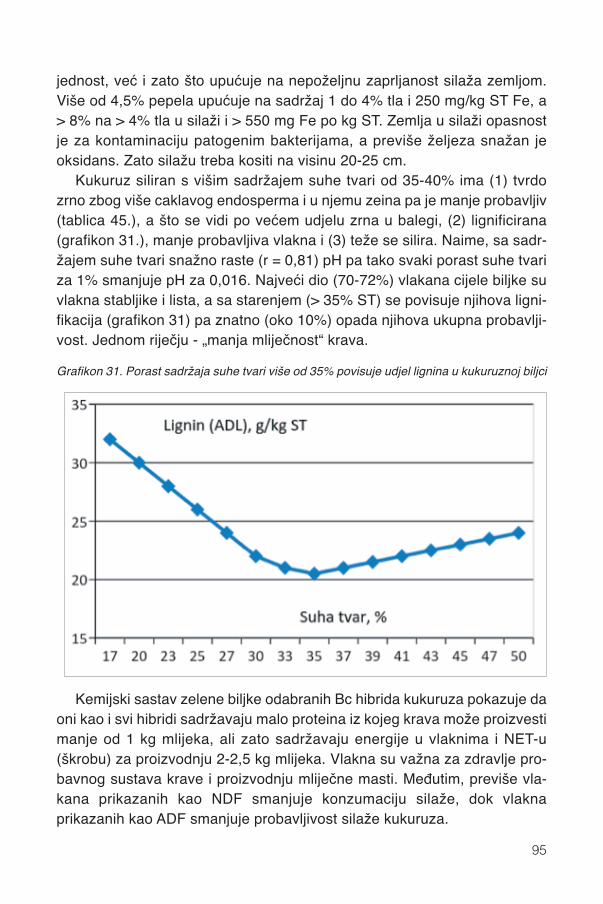
95
jednost, već i zato što upućuje na nepoželjnu zaprljanost silaža zemljom.Više od 4,5% pepela upućuje na sadržaj 1 do 4% tla i 250 mg/kg ST Fe, a> 8% na > 4% tla u silaži i > 550 mg Fe po kg ST. Zemlja u silaži opasnostje za kontaminaciju patogenim bakterijama, a previše željeza snažan jeoksidans. Zato silažu treba kositi na visinu 20-25 cm.
Kukuruz siliran s višim sadržajem suhe tvari od 35-40% ima (1) tvrdozrno zbog više caklavog endosperma i u njemu zeina pa je manje probavljiv(tablica 45.), a što se vidi po većem udjelu zrna u balegi, (2) lignificirana(grafikon 31.), manje probavljiva vlakna i (3) teže se silira. Naime, sa sadr-žajem suhe tvari snažno raste (r = 0,81) pH pa tako svaki porast suhe tvariza 1% smanjuje pH za 0,016. Najveći dio (70-72%) vlakana cijele biljke suvlakna stabljike i lista, a sa starenjem (> 35% ST) se povisuje njihova ligni-fikacija (grafikon 31) pa znatno (oko 10%) opada njihova ukupna probavlji-vost. Jednom riječju - „manja mliječnost“ krava.
Kemijski sastav zelene biljke odabranih Bc hibrida kukuruza pokazuje daoni kao i svi hibridi sadržavaju malo proteina iz kojeg krava može proizvestimanje od 1 kg mlijeka, ali zato sadržavaju energije u vlaknima i NET-u(škrobu) za proizvodnju 2-2,5 kg mlijeka. Vlakna su važna za zdravlje pro-bavnog sustava krave i proizvodnju mliječne masti. Međutim, previše vla-kana prikazanih kao NDF smanjuje konzumaciju silaže, dok vlaknaprikazanih kao ADF smanjuje probavljivost silaže kukuruza.
Grafikon 31. Porast sadržaja suhe tvari više od 35% povisuje udjel lignina u kukuruznoj biljci
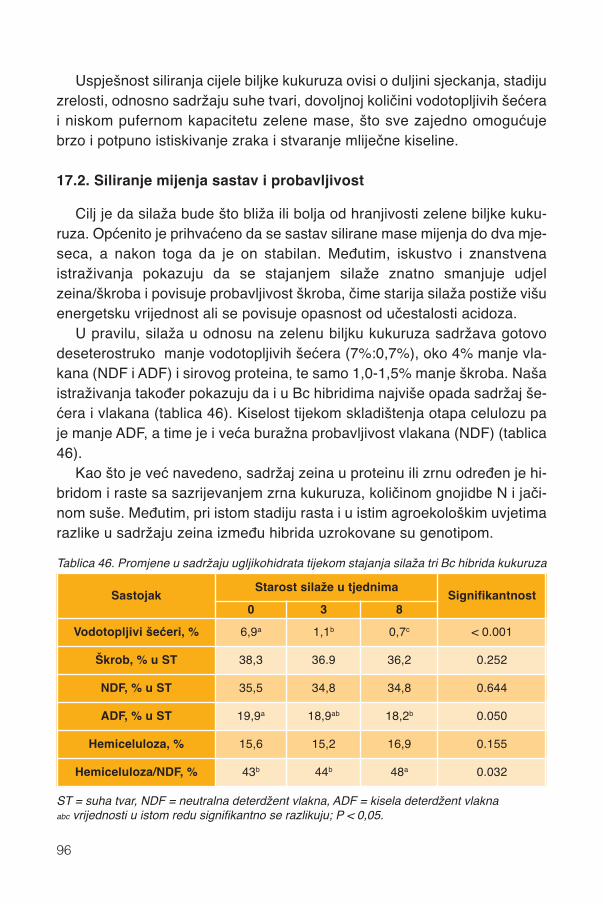
96
Uspješnost siliranja cijele biljke kukuruza ovisi o duljini sjeckanja, stadijuzrelosti, odnosno sadržaju suhe tvari, dovoljnoj količini vodotopljivih šećerai niskom pufernom kapacitetu zelene mase, što sve zajedno omogućujebrzo i potpuno istiskivanje zraka i stvaranje mliječne kiseline.
17.2. Siliranje mijenja sastav i probavljivost
Cilj je da silaža bude što bliža ili bolja od hranjivosti zelene biljke kuku-ruza. Općenito je prihvaćeno da se sastav silirane mase mijenja do dva mje-seca, a nakon toga da je on stabilan. Međutim, iskustvo i znanstvenaistraživanja pokazuju da se stajanjem silaže znatno smanjuje udjelzeina/škroba i povisuje probavljivost škroba, čime starija silaža postiže višuenergetsku vrijednost ali se povisuje opasnost od učestalosti acidoza.
U pravilu, silaža u odnosu na zelenu biljku kukuruza sadržava gotovodeseterostruko manje vodotopljivih šećera (7%:0,7%), oko 4% manje vla-kana (NDF i ADF) i sirovog proteina, te samo 1,0-1,5% manje škroba. Našaistraživanja također pokazuju da i u Bc hibridima najviše opada sadržaj še-ćera i vlakana (tablica 46). Kiselost tijekom skladištenja otapa celulozu paje manje ADF, a time je i veća buražna probavljivost vlakana (NDF) (tablica46).
Kao što je već navedeno, sadržaj zeina u proteinu ili zrnu određen je hi-bridom i raste sa sazrijevanjem zrna kukuruza, količinom gnojidbe N i jači-nom suše. Međutim, pri istom stadiju rasta i u istim agroekološkim uvjetimarazlike u sadržaju zeina između hibrida uzrokovane su genotipom.
Tablica 46. Promjene u sadržaju ugljikohidrata tijekom stajanja silaža tri Bc hibrida kukuruza
Sastojak Starost silaže u tjednima Signifikantnost0 3 8
Vodotopljivi šećeri, % 6,9a 1,1b 0,7c < 0.001
Škrob, % u ST 38,3 36.9 36,2 0.252
NDF, % u ST 35,5 34,8 34,8 0.644
ADF, % u ST 19,9a 18,9ab 18,2b 0.050
Hemiceluloza, % 15,6 15,2 16,9 0.155
Hemiceluloza/NDF, % 43b 44b 48a 0.032
ST = suha tvar, NDF = neutralna deterdžent vlakna, ADF = kisela deterdžent vlaknaabc vrijednosti u istom redu signifikantno se razlikuju; P < 0,05.
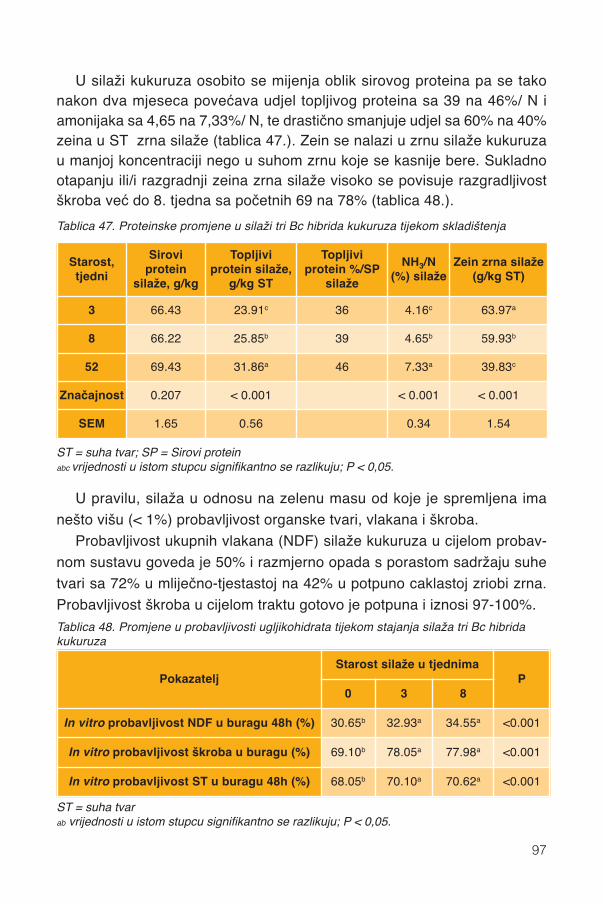
97
U silaži kukuruza osobito se mijenja oblik sirovog proteina pa se takonakon dva mjeseca povećava udjel topljivog proteina sa 39 na 46%/ N iamonijaka sa 4,65 na 7,33%/ N, te drastično smanjuje udjel sa 60% na 40%zeina u ST zrna silaže (tablica 47.). Zein se nalazi u zrnu silaže kukuruzau manjoj koncentraciji nego u suhom zrnu koje se kasnije bere. Sukladnootapanju ili/i razgradnji zeina zrna silaže visoko se povisuje razgradljivostškroba već do 8. tjedna sa početnih 69 na 78% (tablica 48.).
U pravilu, silaža u odnosu na zelenu masu od koje je spremljena imanešto višu (< 1%) probavljivost organske tvari, vlakana i škroba.
Probavljivost ukupnih vlakana (NDF) silaže kukuruza u cijelom probav-nom sustavu goveda je 50% i razmjerno opada s porastom sadržaju suhetvari sa 72% u mliječno-tjestastoj na 42% u potpuno caklastoj zriobi zrna.Probavljivost škroba u cijelom traktu gotovo je potpuna i iznosi 97-100%.
Tablica 47. Proteinske promjene u silaži tri Bc hibrida kukuruza tijekom skladištenja
Starost, tjedni
Sirovi protein
silaže, g/kg
Topljivi protein silaže,
g/kg ST
Topljivi protein %/SP
silažeNH3/N
(%) silažeZein zrna silaže
(g/kg ST)
3 66.43 23.91c 36 4.16c 63.97a
8 66.22 25.85b 39 4.65b 59.93b
52 69.43 31.86a 46 7.33a 39.83c
Značajnost 0.207 < 0.001 < 0.001 < 0.001
SEM 1.65 0.56 0.34 1.54
ST = suha tvar; SP = Sirovi proteinabc vrijednosti u istom stupcu signifikantno se razlikuju; P < 0,05.
Tablica 48. Promjene u probavljivosti ugljikohidrata tijekom stajanja silaža tri Bc hibridakukuruza
PokazateljStarost silaže u tjednima
P0 3 8
In vitro probavljivost NDF u buragu 48h (%) 30.65b 32.93a 34.55a <0.001
In vitro probavljivost škroba u buragu (%) 69.10b 78.05a 77.98a <0.001
In vitro probavljivost ST u buragu 48h (%) 68.05b 70.10a 70.62a <0.001
ST = suha tvarab vrijednosti u istom stupcu signifikantno se razlikuju; P < 0,05.

98
Odležavanje silaže znatno povisuje probavljivost vlakana, i to 4%, a timei njezinu energetsku vrijednost (tablica 48.).
U literaturi se navodi izrazito varijabilna (63-93%) in vitro buražna pro-bavljivost škroba, što je uzrokovano stadijem zrelosti, vlagom, veličinom če-stica zrna u silaži, te osobito sadržajem zeina. Prosječna buražnaprobavljivost škroba je 78% koliko iznosi i u silaži Bc hibrida (78%) većnakon 3. tjedna od siliranja (tablica 48.) i opada sa sadržajem zeina. Zbogranijeg skidanja zelenog kukuruza, zrno u silaži sadržava manje zeina negosuho zrno. Nadalje, kako se zein otapa već u prva tri tjedna skladištenja si-laže buražna probavljivost škroba naraste sa 69 na 78% ili 13% relativno.Međutim, caklavost snažno smanjuje biorazgradljivost škroba sprečavajućikolonizaciju zrnaca škroba mikrobima buraga.
17.3. Hibrid
Genetska varijabilnost energetske vrijednost silaža kreće se u ra-sponu 5,6-6,9 MJ NE/kg ST, što je utvrđeno istraživanjima 273 hibridau eksperimentalnoj stanici INRA Lusignan (Francuska). Hibridi u opti-malnoj zrelosti zrna (32-35% ST) razlikuju se po udjelu i buražnoj pro-bavljivosti škroba i vlakana, a time i energetskoj vrijednosti. Mliječnostkrava razlikuje se 1-3 kg/d, konzumacija suhe tvari 1 kg/d, a prirasti ju-nadi 100 g/d kada se hrane jednakim udjelom različitih hibrida. Jedanod bitnih kriterija za odabir hibrida za siliranje sadržaj je zrna veći od50%. Nadalje, opsežna usporedna testiranja hibrida u tipu tvrdunaca,polutrvdunaca - kvalitetnih zubana i zubana u Francuskoj pokazuje danajveći prinos ST po ha i sadržaj škroba u ST postižu kvalitetni zubani.Isto tako je i ukupna probavljivost škroba viša u kvalitetnih zubana, avlakana slična između tipova endosperma zrna silaža kukuruza. Među-tim, buražna probavljivost (razgradljivost) suhe tvari i osobito škrobaznatnije se razlikuje između hibrida kukuruza (tablica 50). Sukladno na-vedenom, kvalitetni zubani daju i najveći prinos i sadržaj neto energije.
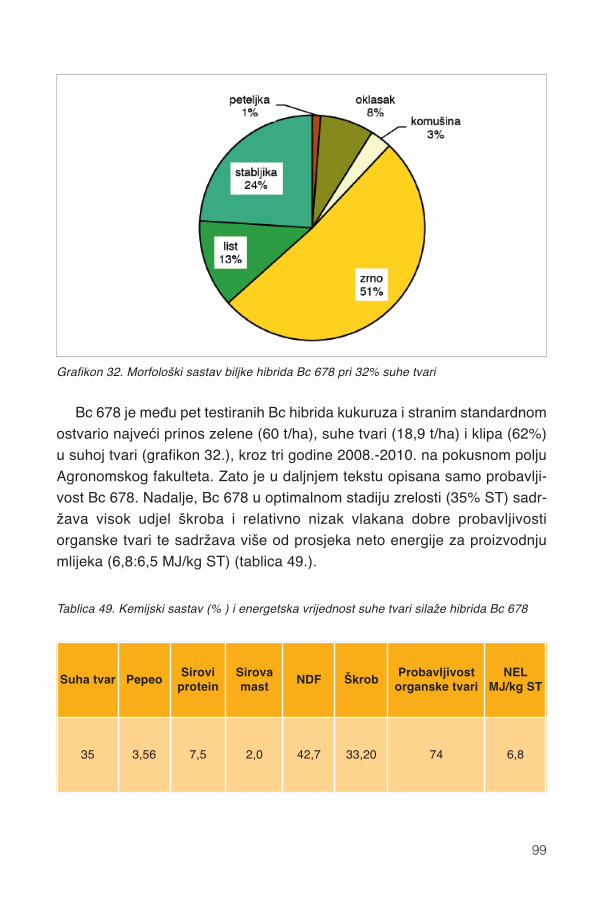
99
Bc 678 je među pet testiranih Bc hibrida kukuruza i stranim standardnomostvario najveći prinos zelene (60 t/ha), suhe tvari (18,9 t/ha) i klipa (62%)u suhoj tvari (grafikon 32.), kroz tri godine 2008.-2010. na pokusnom poljuAgronomskog fakulteta. Zato je u daljnjem tekstu opisana samo probavlji-vost Bc 678. Nadalje, Bc 678 u optimalnom stadiju zrelosti (35% ST) sadr-žava visok udjel škroba i relativno nizak vlakana dobre probavljivostiorganske tvari te sadržava više od prosjeka neto energije za proizvodnjumlijeka (6,8:6,5 MJ/kg ST) (tablica 49.).
Grafikon 32. Morfološki sastav biljke hibrida Bc 678 pri 32% suhe tvari
Tablica 49. Kemijski sastav (% ) i energetska vrijednost suhe tvari silaže hibrida Bc 678
Suha tvar Pepeo Siroviprotein
Sirovamast NDF Škrob Probavljivost
organske tvariNEL
MJ/kg ST
35 3,56 7,5 2,0 42,7 33,20 74 6,8
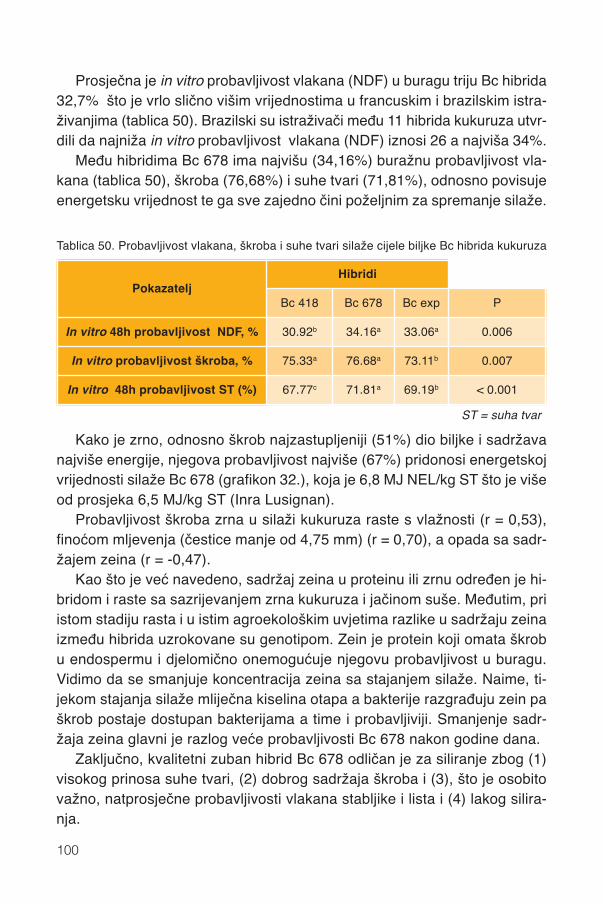
100
Prosječna je in vitro probavljivost vlakana (NDF) u buragu triju Bc hibrida32,7% što je vrlo slično višim vrijednostima u francuskim i brazilskim istra-živanjima (tablica 50). Brazilski su istraživači među 11 hibrida kukuruza utvr-dili da najniža in vitro probavljivost vlakana (NDF) iznosi 26 a najviša 34%.
Među hibridima Bc 678 ima najvišu (34,16%) buražnu probavljivost vla-kana (tablica 50), škroba (76,68%) i suhe tvari (71,81%), odnosno povisujeenergetsku vrijednost te ga sve zajedno čini poželjnim za spremanje silaže.
Kako je zrno, odnosno škrob najzastupljeniji (51%) dio biljke i sadržavanajviše energije, njegova probavljivost najviše (67%) pridonosi energetskojvrijednosti silaže Bc 678 (grafikon 32.), koja je 6,8 MJ NEL/kg ST što je višeod prosjeka 6,5 MJ/kg ST (Inra Lusignan).
Probavljivost škroba zrna u silaži kukuruza raste s vlažnosti (r = 0,53),finoćom mljevenja (čestice manje od 4,75 mm) (r = 0,70), a opada sa sadr-žajem zeina (r = -0,47).
Kao što je već navedeno, sadržaj zeina u proteinu ili zrnu određen je hi-bridom i raste sa sazrijevanjem zrna kukuruza i jačinom suše. Međutim, priistom stadiju rasta i u istim agroekološkim uvjetima razlike u sadržaju zeinaizmeđu hibrida uzrokovane su genotipom. Zein je protein koji omata škrobu endospermu i djelomično onemogućuje njegovu probavljivost u buragu.Vidimo da se smanjuje koncentracija zeina sa stajanjem silaže. Naime, ti-jekom stajanja silaže mliječna kiselina otapa a bakterije razgrađuju zein paškrob postaje dostupan bakterijama a time i probavljiviji. Smanjenje sadr-žaja zeina glavni je razlog veće probavljivosti Bc 678 nakon godine dana.
Zaključno, kvalitetni zuban hibrid Bc 678 odličan je za siliranje zbog (1)visokog prinosa suhe tvari, (2) dobrog sadržaja škroba i (3), što je osobitovažno, natprosječne probavljivosti vlakana stabljike i lista i (4) lakog silira-nja.
Tablica 50. Probavljivost vlakana, škroba i suhe tvari silaže cijele biljke Bc hibrida kukuruza
PokazateljHibridi
Bc 418 Bc 678 Bc exp P
In vitro 48h probavljivost NDF, % 30.92b 34.16a 33.06a 0.006
In vitro probavljivost škroba, % 75.33a 76.68a 73.11b 0.007
In vitro 48h probavljivost ST (%) 67.77c 71.81a 69.19b < 0.001
ST = suha tvar

101
Silaža cijele biljke hibrida Bc 678 ostvarit će svoj visoki potencijal jedinoako se primjereno spremi (tablica 53.). Hranjivost i kvaliteta siliranja svakesilaže uspoređuje se sa standardom koji sadržava poželjne vrijednosti (ta-blica 53.).
17.4. Spremanje silaže
Kukuruznu biljku namijenjenu siliranju treba kositi na visini od 15 cm kadanemamo dovoljno voluminozne krme ili kada nije zaprljana zemljom i obr-nuto, kada imamo dovoljno voluminozne krme i kada je jako zaprljana zem-ljom (> 5,5% pepela) te je tada kosimo na većim visinama.
Masu za siliranje optimalnog sadržaja suhe tvari (32-35% ST) treba sje-ckati na teoretsku dužinu 1,90-2,3 cm, sušu silažu kraće na 1,2 cm.
Razbijanje zrna u silaži kukuruza povisuje iskoristivost škroba pa kravehranjene s tipičnih 30-40% silaže u obroku daju 0,85– +1,25 kg/d više mli-jeka. Preporučuje se razmak između valjaka 2,00, te da je 90-95% razbije-nih od ukupnih zrna u silaži, te da je > 70% čestica manje od ¼ zrna.Poželjno je da > 70% čestica zrna siliranog kukuruza prolazi kroz sito pro-mjera 4,75 mm.
Bolja je mliječnost krava hranjenih silažom kukuruza preporučene duljinesječke i loma zrna (tablica 51.).Tablica 51. Duljina sječke stabljike i krupnoća mljevenja zrna za silažu kukuruza
Duljina sječke biljke, cm 1,95 2,54
Lom zrna, mm 0,0 2,0 8,0 2,0 8,0
Sadržaj suhe tvari, % 43,6 43,6 43,0 42,7 39,9
Krupnoća > 19,0 mm (%) 18,6 6,4 13,1 12,1 25,3
Probavljivost ukupnih vlakana % 20,1 29,7 30,6 35,4 23,2
Probavljivost škroba,% 79,4 83,2 75,8 87,7 75,3
Unos suhe tvari, kg/d 21,9 21,8 22,2 22,1 21,9
Mliječnost,kg/d 35,4 36,1 36,0 37,9 34,1
Suha tvar obroka krave sadržavala je 38% silaže kukuruza, 6% sijena lucerne i 56%koncentrata.
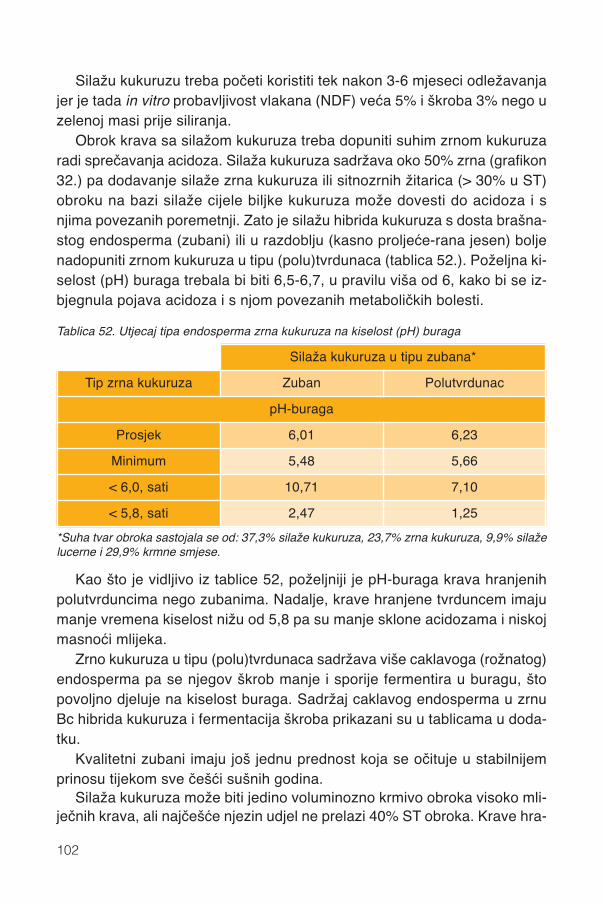
102
Silažu kukuruzu treba početi koristiti tek nakon 3-6 mjeseci odležavanjajer je tada in vitro probavljivost vlakana (NDF) veća 5% i škroba 3% nego uzelenoj masi prije siliranja.
Obrok krava sa silažom kukuruza treba dopuniti suhim zrnom kukuruzaradi sprečavanja acidoza. Silaža kukuruza sadržava oko 50% zrna (grafikon32.) pa dodavanje silaže zrna kukuruza ili sitnozrnih žitarica (> 30% u ST)obroku na bazi silaže cijele biljke kukuruza može dovesti do acidoza i snjima povezanih poremetnji. Zato je silažu hibrida kukuruza s dosta brašna-stog endosperma (zubani) ili u razdoblju (kasno proljeće-rana jesen) boljenadopuniti zrnom kukuruza u tipu (polu)tvrdunaca (tablica 52.). Poželjna ki-selost (pH) buraga trebala bi biti 6,5-6,7, u pravilu viša od 6, kako bi se iz-bjegnula pojava acidoza i s njom povezanih metaboličkih bolesti.
Kao što je vidljivo iz tablice 52, poželjniji je pH-buraga krava hranjenihpolutvrduncima nego zubanima. Nadalje, krave hranjene tvrduncem imajumanje vremena kiselost nižu od 5,8 pa su manje sklone acidozama i niskojmasnoći mlijeka.
Zrno kukuruza u tipu (polu)tvrdunaca sadržava više caklavoga (rožnatog)endosperma pa se njegov škrob manje i sporije fermentira u buragu, štopovoljno djeluje na kiselost buraga. Sadržaj caklavog endosperma u zrnuBc hibrida kukuruza i fermentacija škroba prikazani su u tablicama u doda-tku.
Kvalitetni zubani imaju još jednu prednost koja se očituje u stabilnijemprinosu tijekom sve češći sušnih godina.
Silaža kukuruza može biti jedino voluminozno krmivo obroka visoko mli-ječnih krava, ali najčešće njezin udjel ne prelazi 40% ST obroka. Krave hra-
Tablica 52. Utjecaj tipa endosperma zrna kukuruza na kiselost (pH) buraga
Silaža kukuruza u tipu zubana*Tip zrna kukuruza Zuban Polutvrdunac
pH-buragaProsjek 6,01 6,23
Minimum 5,48 5,66< 6,0, sati 10,71 7,10< 5,8, sati 2,47 1,25
*Suha tvar obroka sastojala se od: 37,3% silaže kukuruza, 23,7% zrna kukuruza, 9,9% silaželucerne i 29,9% krmne smjese.

103
njene s više od 65% silaže kukuruza u obroku smanjuju mliječnost. Bolje jesilažu kukuruza hraniti proteinom bogatijim silažama trava i leguminoza.Krave daju najbolju proizvodnju kada silaža kukuruza čini 50, a trava 50%suhe tvari voluminoznog djela obroka. U obroku krava tijekom tranzicije, tritjedna prije teljenja i poslije teljenja silaža kukuruza ne smije prelaziti 50%ST voluminoznog dijela obroka ili 30% ukupnog obroka.
17.5. Preporuke za siliranje biljke i zrna iz sušnih godina
Silirati biljku kukuruza sušne godine pri istom sadržaju suhe tvari 32-35%nije isto kao i silirati silažu iz normalnih godina proizvodnje. Tijekom sušnegodine lišće uvene, ali sadržaj vode u stabljici ostaje visok te se stječedojam da je silaža suša nego što je u stvarnosti, pa tako dobivamo silaže smanje od željenih 32-35% suhe tvari.
Previše (> 35%) tvari nije dobro jer je iz njih teško istisnuti zrak pa se si-laža grije i pljesnivi. Temperatura suhe silaže s prisutnim zrakom dugo vre-mena nakon zatvaranja viša je od okolne, što znači da kvasci pretvarajuškrob i osobito mliječnu kiselinu u toplinu. Razgradnja mliječne kiseline po-visuje kiselost (pH) na više od 4 što nije dobro za očuvanje silaže jer onatreba pH manji od 3,8. U nas se u suhim silažama vide bijele i crvene plije-sni. Isto tako suhe silaže mogu sadržavati puno opasnih nitrata u donjemdijelu te ih treba rezati na visini 25-40 cm. Suhe silaže treba sjeckati na teo-retsku duljinu od 1 do 2 cm. Silažnu masu za trbuh silaže treba sjeckati sit-nije na 1 cm da bi se jače sabila.
Obavezno treba dodavati i inokulante mliječno-kiselih bakterija jer jemanji njihov broj na suhoj masi za siliranje. Mliječno-kisele bakterije stvarajupoželjnu mliječnu kiselinu, koja povećava kiselost (pad pH) te otapa zeinkoji smanjuje probavljivost škroba.
Suhe silaže osim povišenog sadržaja suhe tvari imaju i povišeni sadržajšećera pa su sklone kvarenju. Dodaci heterofermentativne bakterije stvarajuoctenu kiselinu koja smanjuje broj mikroba koji kvare silaže. Sličan je učinaki dodavanja 1-2 kg/toni puferirane propionske ili benzojeve kiseline suhojmasi za siliranje (> 40% suhe tvari). Treba biti oprezan pri dodavanju vodesuhoj masni za siliranje jer se velike količine vode neće zadržati već će secijediti iz silaže.
Kako suhe mase za siliranje sdržavaju dosta šećera, ne preporučuje sedodavati melasu. Isto tako ne preporučuje se ni dodavanje uree suhim si-lažama.
Najvišu hranjivu vrijednost normalna silaža postiže nakon dva mjeseca,a suha nakon četiri mjeseca odležavanja.
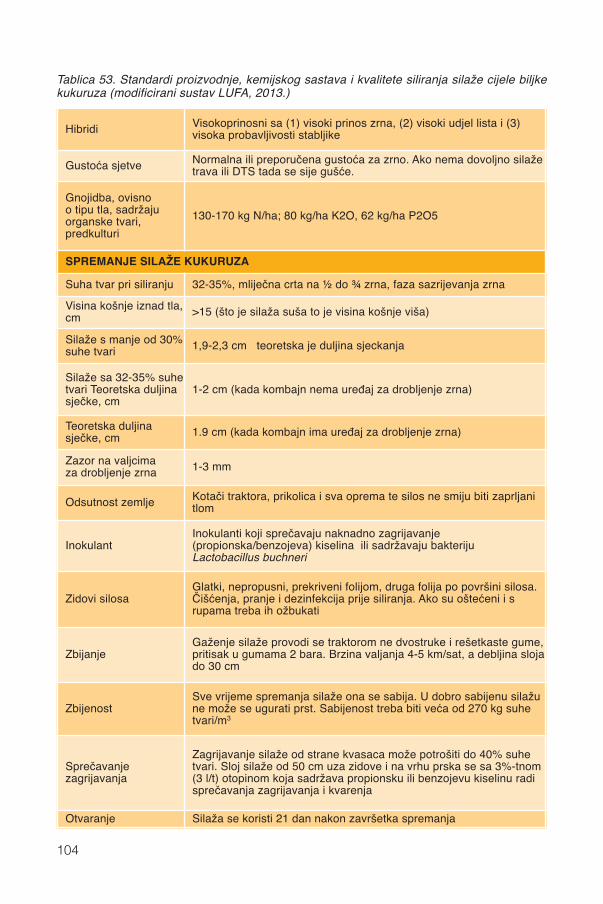
104
Tablica 53. Standardi proizvodnje, kemijskog sastava i kvalitete siliranja silaže cijele biljkekukuruza (modificirani sustav LUFA, 2013.)
Hibridi Visokoprinosni sa (1) visoki prinos zrna, (2) visoki udjel lista i (3)visoka probavljivosti stabljike
Gustoća sjetve Normalna ili preporučena gustoća za zrno. Ako nema dovoljno silažetrava ili DTS tada se sije gušće.
Gnojidba, ovisno o tipu tla, sadržajuorganske tvari,predkulturi
130-170 kg N/ha; 80 kg/ha K2O, 62 kg/ha P2O5
SPREMANJE SILAŽE KUKURUZASuha tvar pri siliranju 32-35%, mliječna crta na ½ do ¾ zrna, faza sazrijevanja zrna Visina košnje iznad tla,cm >15 (što je silaža suša to je visina košnje viša)
Silaže s manje od 30%suhe tvari 1,9-2,3 cm teoretska je duljina sjeckanja
Silaže sa 32-35% suhetvari Teoretska duljinasječke, cm
1-2 cm (kada kombajn nema uređaj za drobljenje zrna)
Teoretska duljinasječke, cm 1.9 cm (kada kombajn ima uređaj za drobljenje zrna)
Zazor na valjcima za drobljenje zrna 1-3 mm
Odsutnost zemlje Kotači traktora, prikolica i sva oprema te silos ne smiju biti zaprljanitlom
InokulantInokulanti koji sprečavaju naknadno zagrijavanje(propionska/benzojeva) kiselina ili sadržavaju bakteriju Lactobacillus buchneri
Zidovi silosaGlatki, nepropusni, prekriveni folijom, druga folija po površini silosa.Čišćenja, pranje i dezinfekcija prije siliranja. Ako su oštećeni i srupama treba ih ožbukati
ZbijanjeGaženje silaže provodi se traktorom ne dvostruke i rešetkaste gume,pritisak u gumama 2 bara. Brzina valjanja 4-5 km/sat, a debljina slojado 30 cm
ZbijenostSve vrijeme spremanja silaže ona se sabija. U dobro sabijenu silažune može se ugurati prst. Sabijenost treba biti veća od 270 kg suhetvari/m3
Sprečavanjezagrijavanja
Zagrijavanje silaže od strane kvasaca može potrošiti do 40% suhetvari. Sloj silaže od 50 cm uza zidove i na vrhu prska se sa 3%-tnom(3 l/t) otopinom koja sadržava propionsku ili benzojevu kiselinu radisprečavanja zagrijavanja i kvarenja
Otvaranje Silaža se koristi 21 dan nakon završetka spremanja
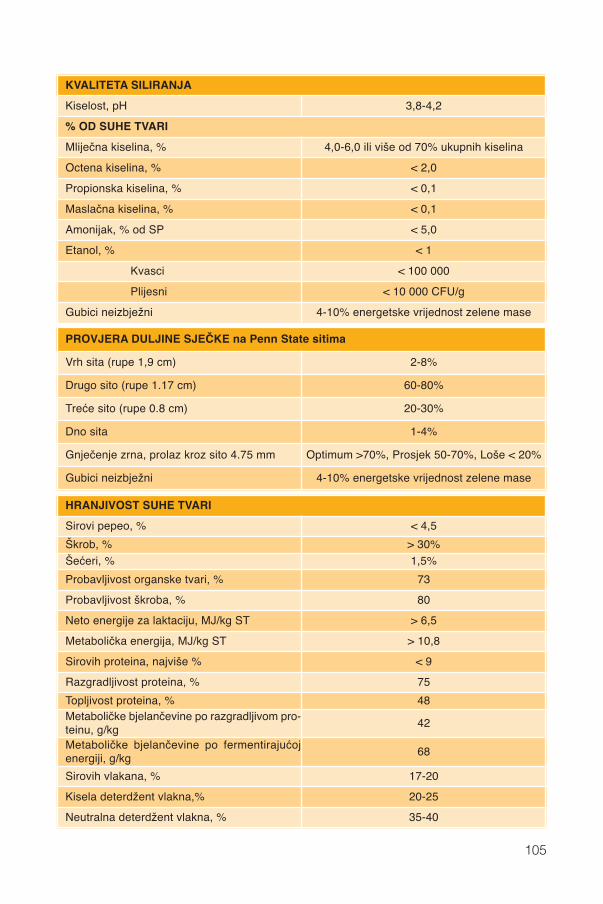
105
KVALITETA SILIRANJAKiselost, pH 3,8-4,2% OD SUHE TVARIMliječna kiselina, % 4,0-6,0 ili više od 70% ukupnih kiselinaOctena kiselina, % < 2,0Propionska kiselina, % < 0,1Maslačna kiselina, % < 0,1Amonijak, % od SP < 5,0Etanol, % < 1
Kvasci < 100 000Plijesni < 10 000 CFU/g
Gubici neizbježni 4-10% energetske vrijednost zelene mase
PROVJERA DULJINE SJEČKE na Penn State sitimaVrh sita (rupe 1,9 cm) 2-8%
Drugo sito (rupe 1.17 cm) 60-80%
Treće sito (rupe 0.8 cm) 20-30%
Dno sita 1-4%
Gnječenje zrna, prolaz kroz sito 4.75 mm Optimum >70%, Prosjek 50-70%, Loše < 20%
Gubici neizbježni 4-10% energetske vrijednost zelene mase
HRANJIVOST SUHE TVARISirovi pepeo, % < 4,5Škrob, % > 30%Šećeri, % 1,5%Probavljivost organske tvari, % 73Probavljivost škroba, % 80Neto energije za laktaciju, MJ/kg ST > 6,5Metabolička energija, MJ/kg ST > 10,8Sirovih proteina, najviše % < 9Razgradljivost proteina, % 75Topljivost proteina, % 48Metaboličke bjelančevine po razgradljivom pro-teinu, g/kg 42Metaboličke bjelančevine po fermentirajućojenergiji, g/kg 68
Sirovih vlakana, % 17-20Kisela deterdžent vlakna,% 20-25Neutralna deterdžent vlakna, % 35-40
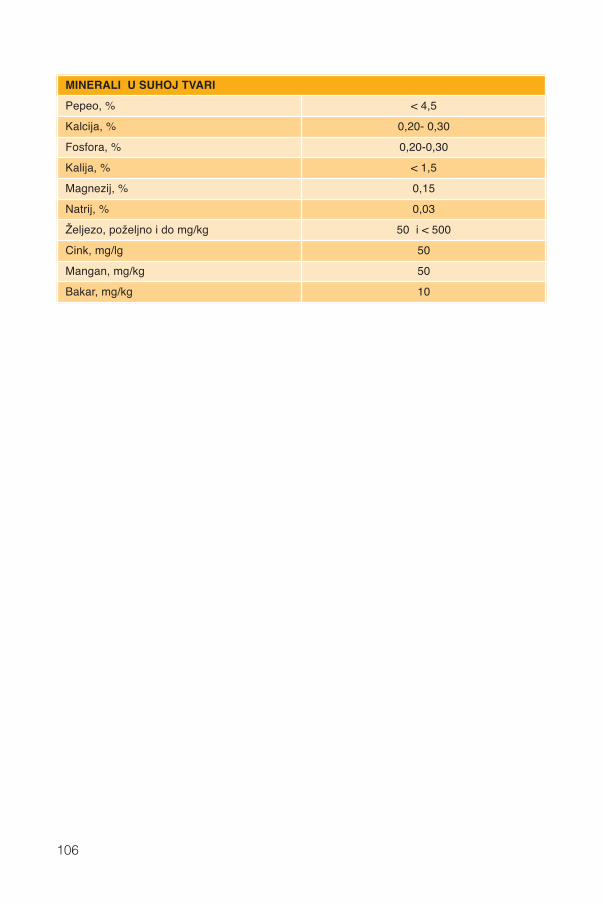
106
MINERALI U SUHOJ TVARIPepeo, % < 4,5Kalcija, % 0,20- 0,30Fosfora, % 0,20-0,30Kalija, % < 1,5Magnezij, % 0,15Natrij, % 0,03Željezo, poželjno i do mg/kg 50 i < 500Cink, mg/lg 50Mangan, mg/kg 50Bakar, mg/kg 10

107
18. Poruke
Porast stanovništva prati povišeni porast potrošnje mesa peradi i svinjate je bolja konverzija biljne pa tako i kukuruza u animalnu hranu jedan odstupova osiguravanja dovoljne količine hrane za ljude. Cilj nam je proizvestišto više mesa, mlijeka i jaja sa što boljim iskorištenjem energije i fitonutrije-nata kukuruza.
Kukuruz je u velikom broju država glavni sastojak hrane domaćih životi-nja koja čini 60-70% troškova stočarske proizvodnje, te se svaka nepravil-nost u procjeni hranjivosti očituje manje učinkovitom proizvodnjom ismanjenom zaradom stočara.
U hranidbi životinja potroši se najviše ili 67% svjetske proizvodnje kuku-ruza od 988 milijuna tona, pa on od svih ratarskih i industrijskih kultura naj-više ili 33% podmiruje energetske i čak 13% proteinske potreba svjetskeanimalne proizvodnje.
Opsežna, recentna, znanstvena istraživanja daju nove spoznaje o utje-caju fine građe i strukture škroba te zeinskih frakcija omotača na visinu i br-zinu probavljivosti, iskorištenje škroba, ali i fitonutrijenata kukuruza naproizvodnju i zdravlje kukuruza i životinja. Danas se prikladnost hibrida ku-kuruza mjeri kroz potencijalnu proizvodnju zrna, otpornosti na bolesti i pri-lagodbu klimatskim ekstremima, te hranjivost, zdravstvenu ispravnost izdravstvenu vrijednost za životinje i ljude.
Kinetika probave škroba u buragu preživača i tanom crijevu peradi i svi-nja određna ponajviše sadržajem i frakcijama zeina više od probavljivostiodređuje energetsku vrijednost kukuruza. Sadržaj zeina u kukuruzu je jed-nostavan i pouzdan pokazatelj enegetske iskoristivosti škroba kukuruza.
Bc hibridi kukuruza odlikuju se raznolikim fizičkim, energetskim, fitonu-tritivnim i pigmentacijskim svojstvima, kinetikama probave škroba, te namnjihovo poznavanje olakšava pronalaženje pravog hibrida za našu animalnuproizvodnju.
Bc hibridi kukuruza objedinjuju visoku hranjivost i antioksidativnu aktiv-nost s dobrim potencijalom prinosa i adaptivnim svojstvima na klimatskeekstreme.
Monografija pruža relevantne spoznaje u vrednovanju kukuruza i podatkeo hranidbenim svojstvima Bc hibrida koji omogućuje precizniji uzgoj i efika-sniju pretvorbu (konverziju) hrane u animalne proizvode.
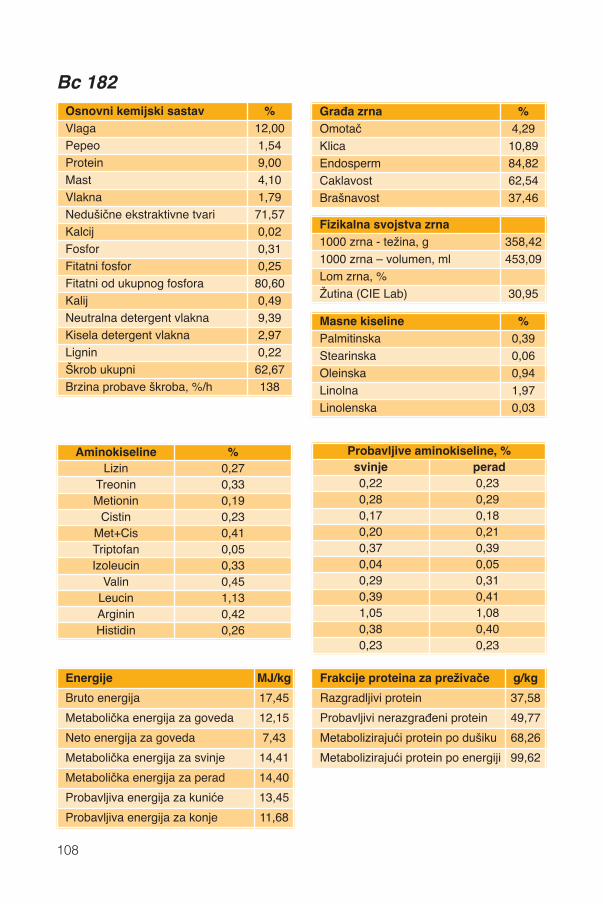
108
Bc 182Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,54Protein 9,00Mast 4,10Vlakna 1,79Nedušične ekstraktivne tvari 71,57Kalcij 0,02Fosfor 0,31Fitatni fosfor 0,25Fitatni od ukupnog fosfora 80,60Kalij 0,49Neutralna detergent vlakna 9,39Kisela detergent vlakna 2,97Lignin 0,22Škrob ukupni 62,67Brzina probave škroba, %/h 138
Aminokiseline %Lizin 0,27
Treonin 0,33Metionin 0,19
Cistin 0,23Met+Cis 0,41Triptofan 0,05Izoleucin 0,33
Valin 0,45Leucin 1,13Arginin 0,42Histidin 0,26
Energije MJ/kgBruto energija 17,45Metabolička energija za goveda 12,15Neto energija za goveda 7,43Metabolička energija za svinje 14,41Metabolička energija za perad 14,40Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,68
Frakcije proteina za preživače g/kgRazgradljivi protein 37,58Probavljivi nerazgrađeni protein 49,77Metabolizirajući protein po dušiku 68,26Metabolizirajući protein po energiji 99,62
Građa zrna %Omotač 4,29Klica 10,89Endosperm 84,82Caklavost 62,54Brašnavost 37,46
Fizikalna svojstva zrna1000 zrna - težina, g 358,421000 zrna – volumen, ml 453,09Lom zrna, %Žutina (CIE Lab) 30,95
Masne kiseline %Palmitinska 0,39Stearinska 0,06Oleinska 0,94Linolna 1,97Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,22 0,230,28 0,290,17 0,180,20 0,210,37 0,390,04 0,050,29 0,310,39 0,411,05 1,080,38 0,400,23 0,23
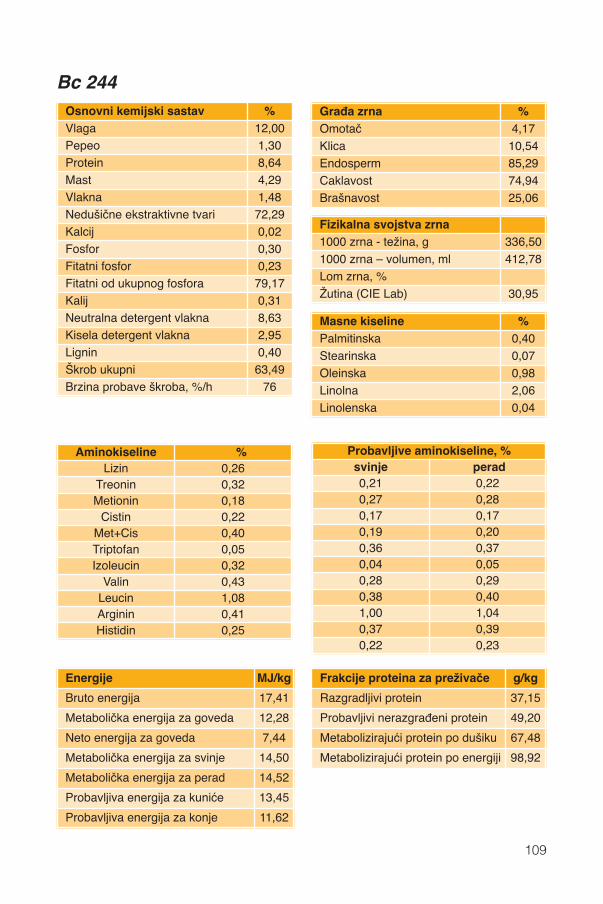
109
Bc 244Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,30Protein 8,64Mast 4,29Vlakna 1,48Nedušične ekstraktivne tvari 72,29Kalcij 0,02Fosfor 0,30Fitatni fosfor 0,23Fitatni od ukupnog fosfora 79,17Kalij 0,31Neutralna detergent vlakna 8,63Kisela detergent vlakna 2,95Lignin 0,40Škrob ukupni 63,49Brzina probave škroba, %/h 76
Aminokiseline %Lizin 0,26
Treonin 0,32Metionin 0,18
Cistin 0,22Met+Cis 0,40Triptofan 0,05Izoleucin 0,32
Valin 0,43Leucin 1,08Arginin 0,41Histidin 0,25
Energije MJ/kgBruto energija 17,41Metabolička energija za goveda 12,28Neto energija za goveda 7,44Metabolička energija za svinje 14,50Metabolička energija za perad 14,52Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,62
Frakcije proteina za preživače g/kgRazgradljivi protein 37,15Probavljivi nerazgrađeni protein 49,20Metabolizirajući protein po dušiku 67,48Metabolizirajući protein po energiji 98,92
Građa zrna %Omotač 4,17Klica 10,54Endosperm 85,29Caklavost 74,94Brašnavost 25,06
Fizikalna svojstva zrna1000 zrna - težina, g 336,501000 zrna – volumen, ml 412,78Lom zrna, %Žutina (CIE Lab) 30,95
Masne kiseline %Palmitinska 0,40Stearinska 0,07Oleinska 0,98Linolna 2,06Linolenska 0,04
Probavljive aminokiseline, %svinje perad0,21 0,220,27 0,280,17 0,170,19 0,200,36 0,370,04 0,050,28 0,290,38 0,401,00 1,040,37 0,390,22 0,23
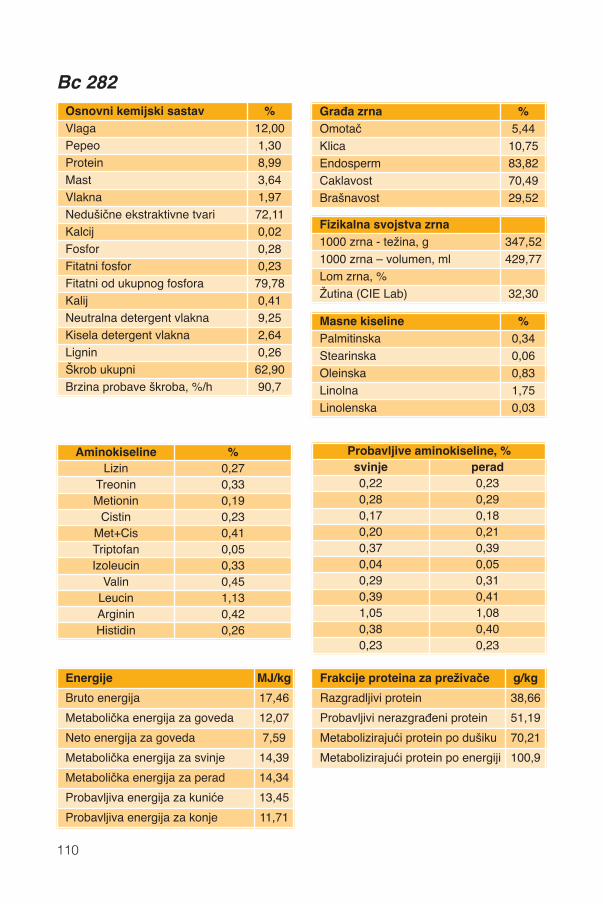
110
Bc 282Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,30Protein 8,99Mast 3,64Vlakna 1,97Nedušične ekstraktivne tvari 72,11Kalcij 0,02Fosfor 0,28Fitatni fosfor 0,23Fitatni od ukupnog fosfora 79,78Kalij 0,41Neutralna detergent vlakna 9,25Kisela detergent vlakna 2,64Lignin 0,26Škrob ukupni 62,90Brzina probave škroba, %/h 90,7
Aminokiseline %Lizin 0,27
Treonin 0,33Metionin 0,19
Cistin 0,23Met+Cis 0,41Triptofan 0,05Izoleucin 0,33
Valin 0,45Leucin 1,13Arginin 0,42Histidin 0,26
Energije MJ/kgBruto energija 17,46Metabolička energija za goveda 12,07Neto energija za goveda 7,59Metabolička energija za svinje 14,39Metabolička energija za perad 14,34Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,71
Frakcije proteina za preživače g/kgRazgradljivi protein 38,66Probavljivi nerazgrađeni protein 51,19Metabolizirajući protein po dušiku 70,21Metabolizirajući protein po energiji 100,9
Građa zrna %Omotač 5,44Klica 10,75Endosperm 83,82Caklavost 70,49Brašnavost 29,52
Fizikalna svojstva zrna1000 zrna - težina, g 347,521000 zrna – volumen, ml 429,77Lom zrna, %Žutina (CIE Lab) 32,30
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,83Linolna 1,75Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,22 0,230,28 0,290,17 0,180,20 0,210,37 0,390,04 0,050,29 0,310,39 0,411,05 1,080,38 0,400,23 0,23
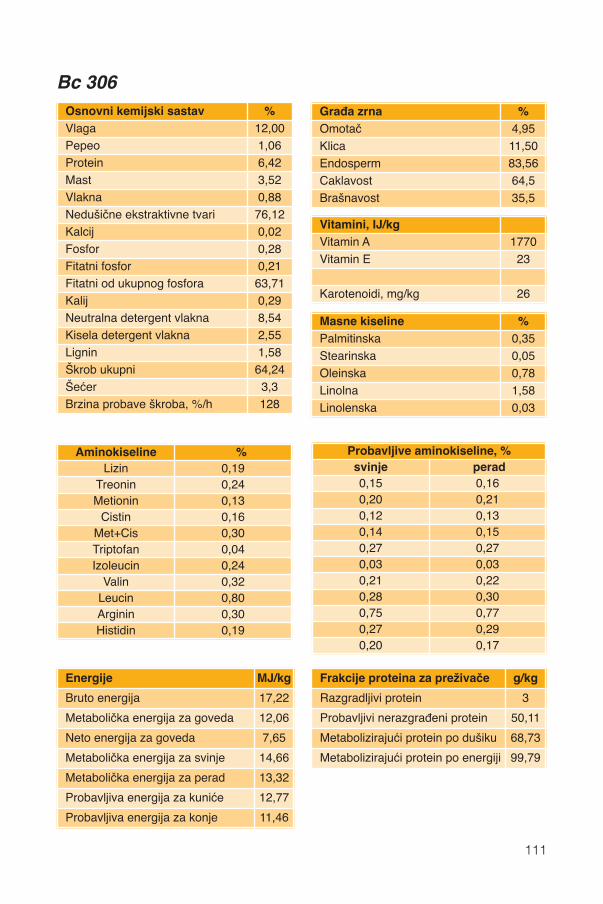
111
Bc 306Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,06Protein 6,42Mast 3,52Vlakna 0,88Nedušične ekstraktivne tvari 76,12Kalcij 0,02Fosfor 0,28Fitatni fosfor 0,21Fitatni od ukupnog fosfora 63,71Kalij 0,29Neutralna detergent vlakna 8,54Kisela detergent vlakna 2,55Lignin 1,58Škrob ukupni 64,24Šećer 3,3Brzina probave škroba, %/h 128
Aminokiseline %Lizin 0,19
Treonin 0,24Metionin 0,13
Cistin 0,16Met+Cis 0,30Triptofan 0,04Izoleucin 0,24
Valin 0,32Leucin 0,80Arginin 0,30Histidin 0,19
Energije MJ/kgBruto energija 17,22Metabolička energija za goveda 12,06Neto energija za goveda 7,65Metabolička energija za svinje 14,66Metabolička energija za perad 13,32Probavljiva energija za kuniće 12,77Probavljiva energija za konje 11,46
Frakcije proteina za preživače g/kgRazgradljivi protein 3Probavljivi nerazgrađeni protein 50,11Metabolizirajući protein po dušiku 68,73Metabolizirajući protein po energiji 99,79
Građa zrna %Omotač 4,95Klica 11,50Endosperm 83,56Caklavost 64,5Brašnavost 35,5
Vitamini, IJ/kgVitamin A 1770Vitamin E 23
Karotenoidi, mg/kg 26
Masne kiseline %Palmitinska 0,35Stearinska 0,05Oleinska 0,78Linolna 1,58Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,15 0,160,20 0,210,12 0,130,14 0,150,27 0,270,03 0,030,21 0,220,28 0,300,75 0,770,27 0,290,20 0,17
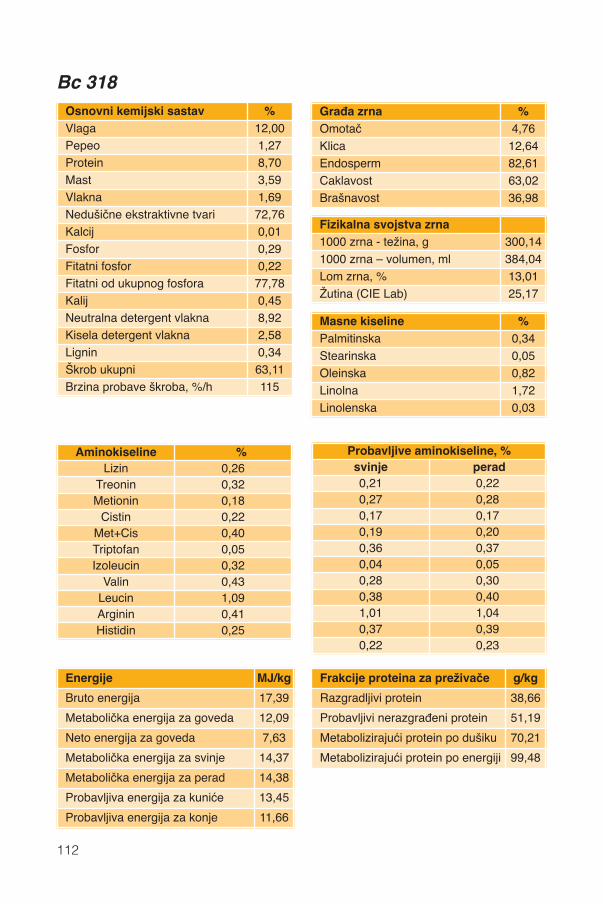
112
Bc 318Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,27Protein 8,70Mast 3,59Vlakna 1,69Nedušične ekstraktivne tvari 72,76Kalcij 0,01Fosfor 0,29Fitatni fosfor 0,22Fitatni od ukupnog fosfora 77,78Kalij 0,45Neutralna detergent vlakna 8,92Kisela detergent vlakna 2,58Lignin 0,34Škrob ukupni 63,11Brzina probave škroba, %/h 115
Aminokiseline %Lizin 0,26
Treonin 0,32Metionin 0,18
Cistin 0,22Met+Cis 0,40Triptofan 0,05Izoleucin 0,32
Valin 0,43Leucin 1,09Arginin 0,41Histidin 0,25
Energije MJ/kgBruto energija 17,39Metabolička energija za goveda 12,09Neto energija za goveda 7,63Metabolička energija za svinje 14,37Metabolička energija za perad 14,38Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,66
Frakcije proteina za preživače g/kgRazgradljivi protein 38,66Probavljivi nerazgrađeni protein 51,19Metabolizirajući protein po dušiku 70,21Metabolizirajući protein po energiji 99,48
Građa zrna %Omotač 4,76Klica 12,64Endosperm 82,61Caklavost 63,02Brašnavost 36,98
Fizikalna svojstva zrna1000 zrna - težina, g 300,141000 zrna – volumen, ml 384,04Lom zrna, % 13,01Žutina (CIE Lab) 25,17
Masne kiseline %Palmitinska 0,34Stearinska 0,05Oleinska 0,82Linolna 1,72Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,220,27 0,280,17 0,170,19 0,200,36 0,370,04 0,050,28 0,300,38 0,401,01 1,040,37 0,390,22 0,23
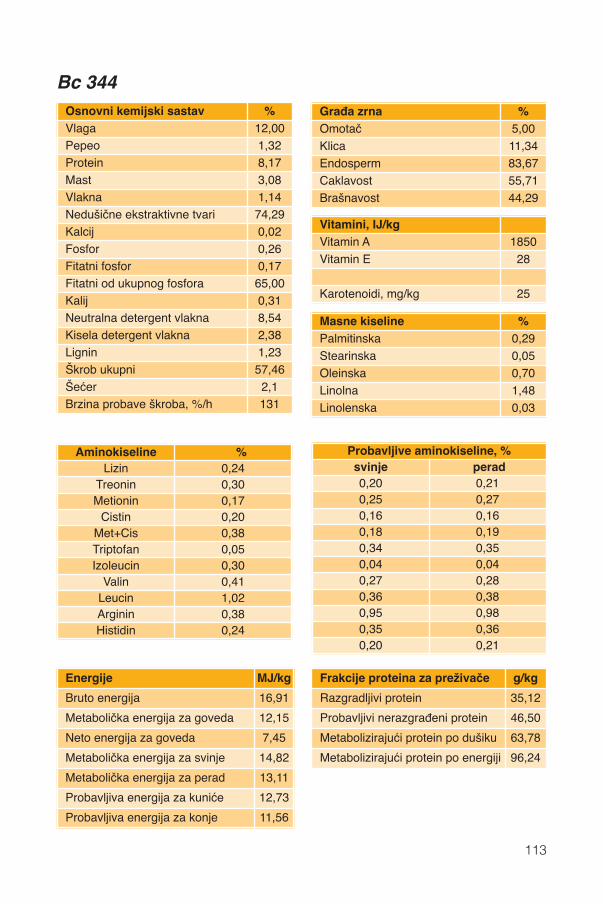
113
Bc 344Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,32Protein 8,17Mast 3,08Vlakna 1,14Nedušične ekstraktivne tvari 74,29Kalcij 0,02Fosfor 0,26Fitatni fosfor 0,17Fitatni od ukupnog fosfora 65,00Kalij 0,31Neutralna detergent vlakna 8,54Kisela detergent vlakna 2,38Lignin 1,23Škrob ukupni 57,46Šećer 2,1Brzina probave škroba, %/h 131
Aminokiseline %Lizin 0,24
Treonin 0,30Metionin 0,17
Cistin 0,20Met+Cis 0,38Triptofan 0,05Izoleucin 0,30
Valin 0,41Leucin 1,02Arginin 0,38Histidin 0,24
Energije MJ/kgBruto energija 16,91Metabolička energija za goveda 12,15Neto energija za goveda 7,45Metabolička energija za svinje 14,82Metabolička energija za perad 13,11Probavljiva energija za kuniće 12,73Probavljiva energija za konje 11,56
Frakcije proteina za preživače g/kgRazgradljivi protein 35,12Probavljivi nerazgrađeni protein 46,50Metabolizirajući protein po dušiku 63,78Metabolizirajući protein po energiji 96,24
Građa zrna %Omotač 5,00Klica 11,34Endosperm 83,67Caklavost 55,71Brašnavost 44,29
Vitamini, IJ/kgVitamin A 1850Vitamin E 28
Karotenoidi, mg/kg 25
Masne kiseline %Palmitinska 0,29Stearinska 0,05Oleinska 0,70Linolna 1,48Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,20 0,210,25 0,270,16 0,160,18 0,190,34 0,350,04 0,040,27 0,280,36 0,380,95 0,980,35 0,360,20 0,21
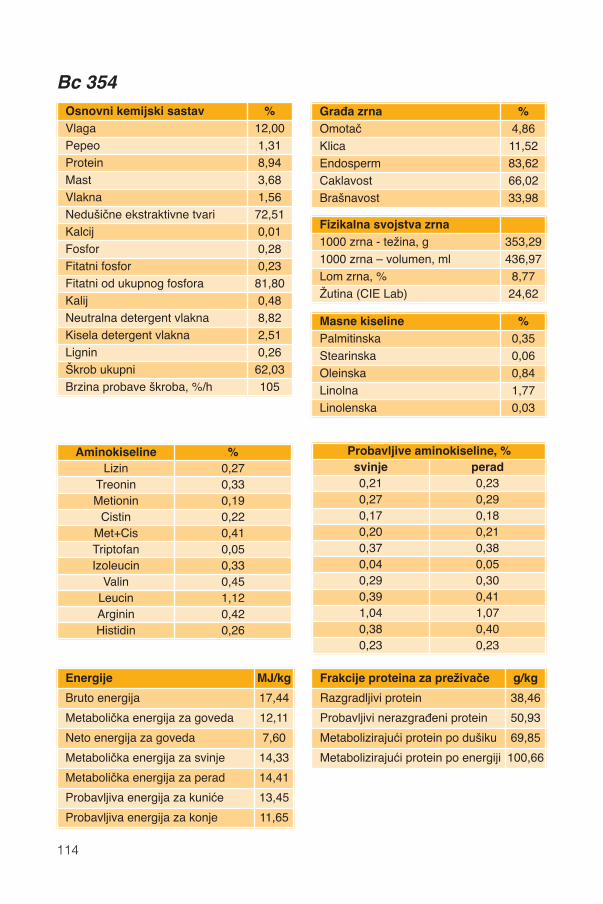
114
Bc 354Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,31Protein 8,94Mast 3,68Vlakna 1,56Nedušične ekstraktivne tvari 72,51Kalcij 0,01Fosfor 0,28Fitatni fosfor 0,23Fitatni od ukupnog fosfora 81,80Kalij 0,48Neutralna detergent vlakna 8,82Kisela detergent vlakna 2,51Lignin 0,26Škrob ukupni 62,03Brzina probave škroba, %/h 105
Aminokiseline %Lizin 0,27
Treonin 0,33Metionin 0,19
Cistin 0,22Met+Cis 0,41Triptofan 0,05Izoleucin 0,33
Valin 0,45Leucin 1,12Arginin 0,42Histidin 0,26
Energije MJ/kgBruto energija 17,44Metabolička energija za goveda 12,11Neto energija za goveda 7,60Metabolička energija za svinje 14,33Metabolička energija za perad 14,41Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,65
Frakcije proteina za preživače g/kgRazgradljivi protein 38,46Probavljivi nerazgrađeni protein 50,93Metabolizirajući protein po dušiku 69,85Metabolizirajući protein po energiji 100,66
Građa zrna %Omotač 4,86Klica 11,52Endosperm 83,62Caklavost 66,02Brašnavost 33,98
Fizikalna svojstva zrna1000 zrna - težina, g 353,291000 zrna – volumen, ml 436,97Lom zrna, % 8,77Žutina (CIE Lab) 24,62
Masne kiseline %Palmitinska 0,35Stearinska 0,06Oleinska 0,84Linolna 1,77Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,230,27 0,290,17 0,180,20 0,210,37 0,380,04 0,050,29 0,300,39 0,411,04 1,070,38 0,400,23 0,23
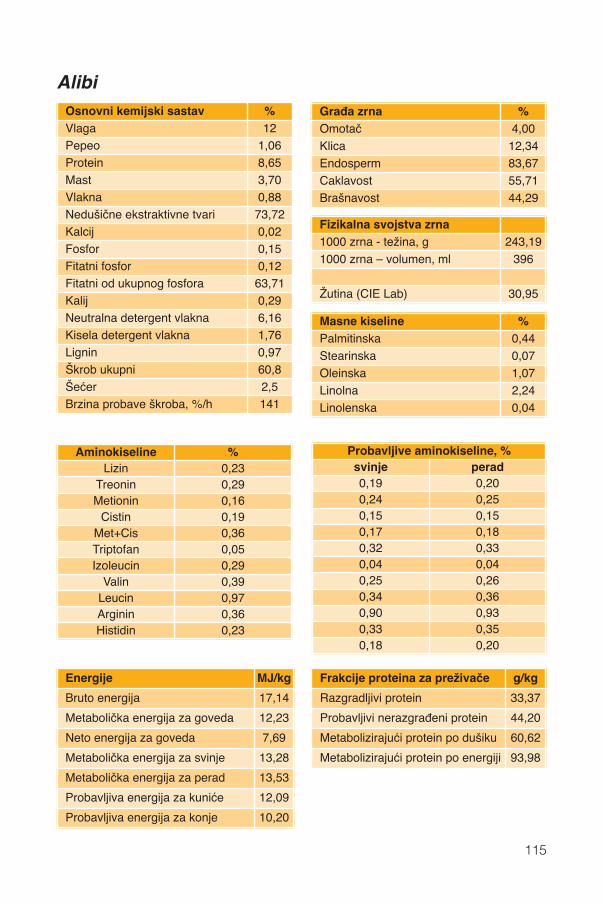
115
AlibiOsnovni kemijski sastav %Vlaga 12Pepeo 1,06Protein 8,65Mast 3,70Vlakna 0,88Nedušične ekstraktivne tvari 73,72Kalcij 0,02Fosfor 0,15Fitatni fosfor 0,12Fitatni od ukupnog fosfora 63,71Kalij 0,29Neutralna detergent vlakna 6,16Kisela detergent vlakna 1,76Lignin 0,97Škrob ukupni 60,8Šećer 2,5Brzina probave škroba, %/h 141
Aminokiseline %Lizin 0,23
Treonin 0,29Metionin 0,16
Cistin 0,19Met+Cis 0,36Triptofan 0,05Izoleucin 0,29
Valin 0,39Leucin 0,97Arginin 0,36Histidin 0,23
Energije MJ/kgBruto energija 17,14Metabolička energija za goveda 12,23Neto energija za goveda 7,69Metabolička energija za svinje 13,28Metabolička energija za perad 13,53Probavljiva energija za kuniće 12,09Probavljiva energija za konje 10,20
Frakcije proteina za preživače g/kgRazgradljivi protein 33,37Probavljivi nerazgrađeni protein 44,20Metabolizirajući protein po dušiku 60,62Metabolizirajući protein po energiji 93,98
Građa zrna %Omotač 4,00Klica 12,34Endosperm 83,67Caklavost 55,71Brašnavost 44,29
Fizikalna svojstva zrna1000 zrna - težina, g 243,191000 zrna – volumen, ml 396
Žutina (CIE Lab) 30,95
Masne kiseline %Palmitinska 0,44Stearinska 0,07Oleinska 1,07Linolna 2,24Linolenska 0,04
Probavljive aminokiseline, %svinje perad0,19 0,200,24 0,250,15 0,150,17 0,180,32 0,330,04 0,040,25 0,260,34 0,360,90 0,930,33 0,350,18 0,20
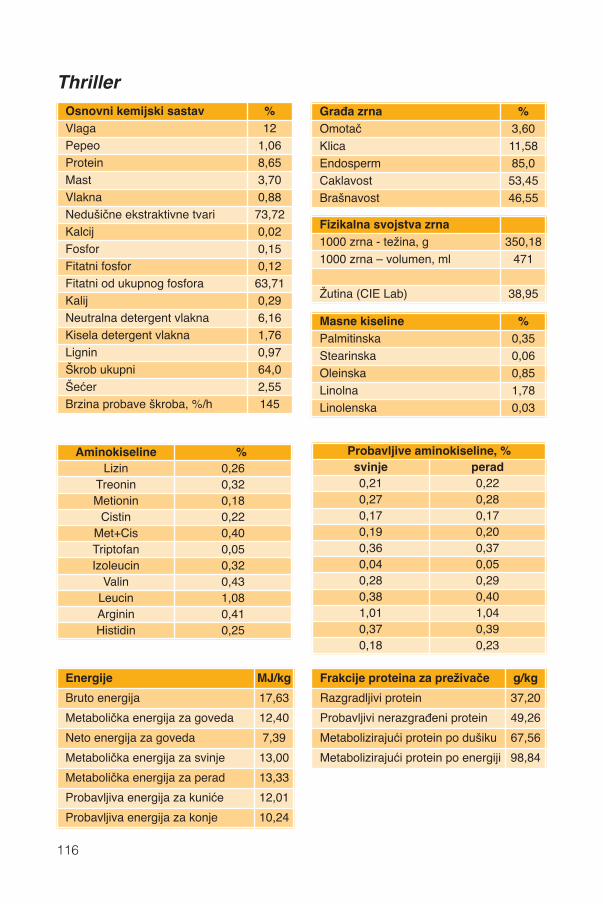
116
ThrillerOsnovni kemijski sastav %Vlaga 12Pepeo 1,06Protein 8,65Mast 3,70Vlakna 0,88Nedušične ekstraktivne tvari 73,72Kalcij 0,02Fosfor 0,15Fitatni fosfor 0,12Fitatni od ukupnog fosfora 63,71Kalij 0,29Neutralna detergent vlakna 6,16Kisela detergent vlakna 1,76Lignin 0,97Škrob ukupni 64,0Šećer 2,55Brzina probave škroba, %/h 145
Aminokiseline %Lizin 0,26
Treonin 0,32Metionin 0,18
Cistin 0,22Met+Cis 0,40Triptofan 0,05Izoleucin 0,32
Valin 0,43Leucin 1,08Arginin 0,41Histidin 0,25
Energije MJ/kgBruto energija 17,63Metabolička energija za goveda 12,40Neto energija za goveda 7,39Metabolička energija za svinje 13,00Metabolička energija za perad 13,33Probavljiva energija za kuniće 12,01Probavljiva energija za konje 10,24
Frakcije proteina za preživače g/kgRazgradljivi protein 37,20Probavljivi nerazgrađeni protein 49,26Metabolizirajući protein po dušiku 67,56Metabolizirajući protein po energiji 98,84
Građa zrna %Omotač 3,60Klica 11,58Endosperm 85,0Caklavost 53,45Brašnavost 46,55
Fizikalna svojstva zrna1000 zrna - težina, g 350,181000 zrna – volumen, ml 471
Žutina (CIE Lab) 38,95
Masne kiseline %Palmitinska 0,35Stearinska 0,06Oleinska 0,85Linolna 1,78Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,220,27 0,280,17 0,170,19 0,200,36 0,370,04 0,050,28 0,290,38 0,401,01 1,040,37 0,390,18 0,23
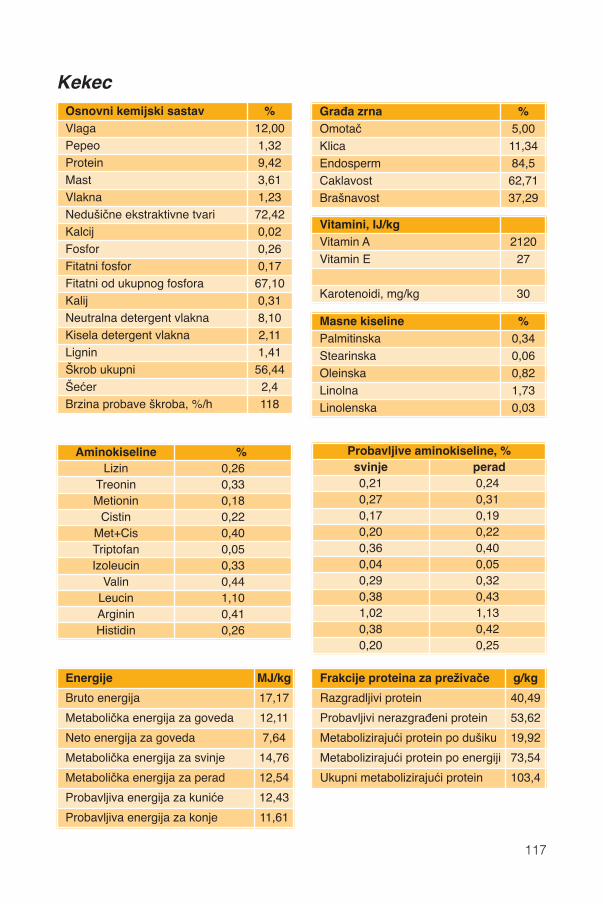
117
KekecOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,32Protein 9,42Mast 3,61Vlakna 1,23Nedušične ekstraktivne tvari 72,42Kalcij 0,02Fosfor 0,26Fitatni fosfor 0,17Fitatni od ukupnog fosfora 67,10Kalij 0,31Neutralna detergent vlakna 8,10Kisela detergent vlakna 2,11Lignin 1,41Škrob ukupni 56,44Šećer 2,4Brzina probave škroba, %/h 118
Aminokiseline %Lizin 0,26
Treonin 0,33Metionin 0,18
Cistin 0,22Met+Cis 0,40Triptofan 0,05Izoleucin 0,33
Valin 0,44Leucin 1,10Arginin 0,41Histidin 0,26
Energije MJ/kgBruto energija 17,17Metabolička energija za goveda 12,11Neto energija za goveda 7,64Metabolička energija za svinje 14,76Metabolička energija za perad 12,54Probavljiva energija za kuniće 12,43Probavljiva energija za konje 11,61
Frakcije proteina za preživače g/kgRazgradljivi protein 40,49Probavljivi nerazgrađeni protein 53,62Metabolizirajući protein po dušiku 19,92Metabolizirajući protein po energiji 73,54Ukupni metabolizirajući protein 103,4
Građa zrna %Omotač 5,00Klica 11,34Endosperm 84,5Caklavost 62,71Brašnavost 37,29
Vitamini, IJ/kgVitamin A 2120Vitamin E 27
Karotenoidi, mg/kg 30
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,82Linolna 1,73Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,240,27 0,310,17 0,190,20 0,220,36 0,400,04 0,050,29 0,320,38 0,431,02 1,130,38 0,420,20 0,25
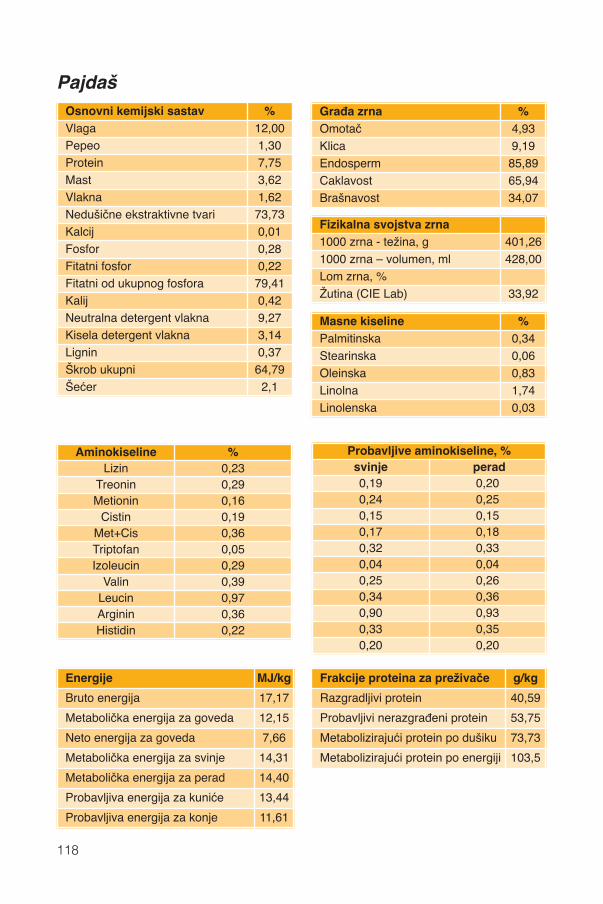
118
PajdašOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,30Protein 7,75Mast 3,62Vlakna 1,62Nedušične ekstraktivne tvari 73,73Kalcij 0,01Fosfor 0,28Fitatni fosfor 0,22Fitatni od ukupnog fosfora 79,41Kalij 0,42Neutralna detergent vlakna 9,27Kisela detergent vlakna 3,14Lignin 0,37Škrob ukupni 64,79Šećer 2,1
Aminokiseline %Lizin 0,23
Treonin 0,29Metionin 0,16
Cistin 0,19Met+Cis 0,36Triptofan 0,05Izoleucin 0,29
Valin 0,39Leucin 0,97Arginin 0,36Histidin 0,22
Energije MJ/kgBruto energija 17,17Metabolička energija za goveda 12,15Neto energija za goveda 7,66Metabolička energija za svinje 14,31Metabolička energija za perad 14,40Probavljiva energija za kuniće 13,44Probavljiva energija za konje 11,61
Frakcije proteina za preživače g/kgRazgradljivi protein 40,59Probavljivi nerazgrađeni protein 53,75Metabolizirajući protein po dušiku 73,73Metabolizirajući protein po energiji 103,5
Građa zrna %Omotač 4,93Klica 9,19Endosperm 85,89Caklavost 65,94Brašnavost 34,07
Fizikalna svojstva zrna1000 zrna - težina, g 401,261000 zrna – volumen, ml 428,00Lom zrna, %Žutina (CIE Lab) 33,92
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,83Linolna 1,74Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,19 0,200,24 0,250,15 0,150,17 0,180,32 0,330,04 0,040,25 0,260,34 0,360,90 0,930,33 0,350,20 0,20
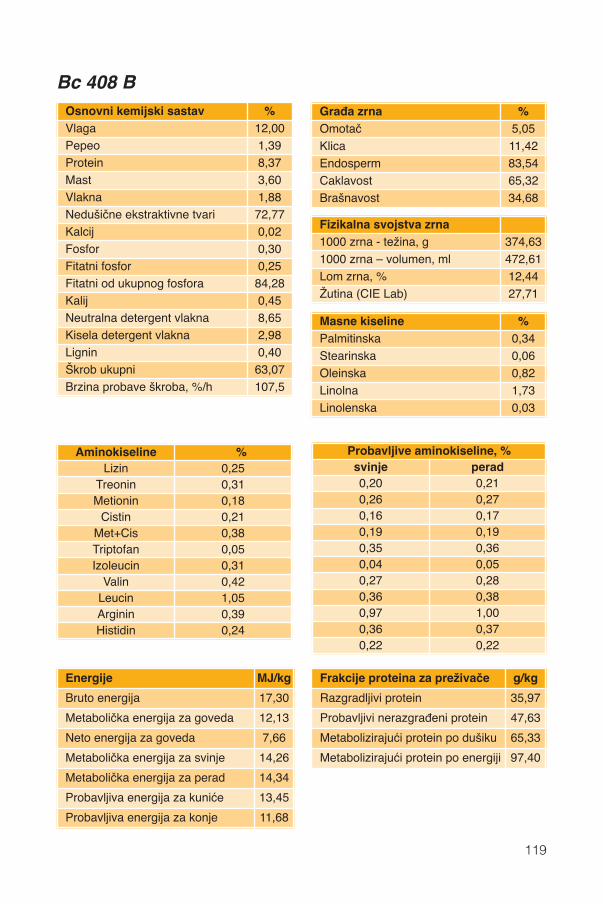
119
Bc 408 BOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,39Protein 8,37Mast 3,60Vlakna 1,88Nedušične ekstraktivne tvari 72,77Kalcij 0,02Fosfor 0,30Fitatni fosfor 0,25Fitatni od ukupnog fosfora 84,28Kalij 0,45Neutralna detergent vlakna 8,65Kisela detergent vlakna 2,98Lignin 0,40Škrob ukupni 63,07Brzina probave škroba, %/h 107,5
Aminokiseline %Lizin 0,25
Treonin 0,31Metionin 0,18
Cistin 0,21Met+Cis 0,38Triptofan 0,05Izoleucin 0,31
Valin 0,42Leucin 1,05Arginin 0,39Histidin 0,24
Energije MJ/kgBruto energija 17,30Metabolička energija za goveda 12,13Neto energija za goveda 7,66Metabolička energija za svinje 14,26Metabolička energija za perad 14,34Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,68
Frakcije proteina za preživače g/kgRazgradljivi protein 35,97Probavljivi nerazgrađeni protein 47,63Metabolizirajući protein po dušiku 65,33Metabolizirajući protein po energiji 97,40
Građa zrna %Omotač 5,05Klica 11,42Endosperm 83,54Caklavost 65,32Brašnavost 34,68
Fizikalna svojstva zrna1000 zrna - težina, g 374,631000 zrna – volumen, ml 472,61Lom zrna, % 12,44Žutina (CIE Lab) 27,71
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,82Linolna 1,73Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,20 0,210,26 0,270,16 0,170,19 0,190,35 0,360,04 0,050,27 0,280,36 0,380,97 1,000,36 0,370,22 0,22
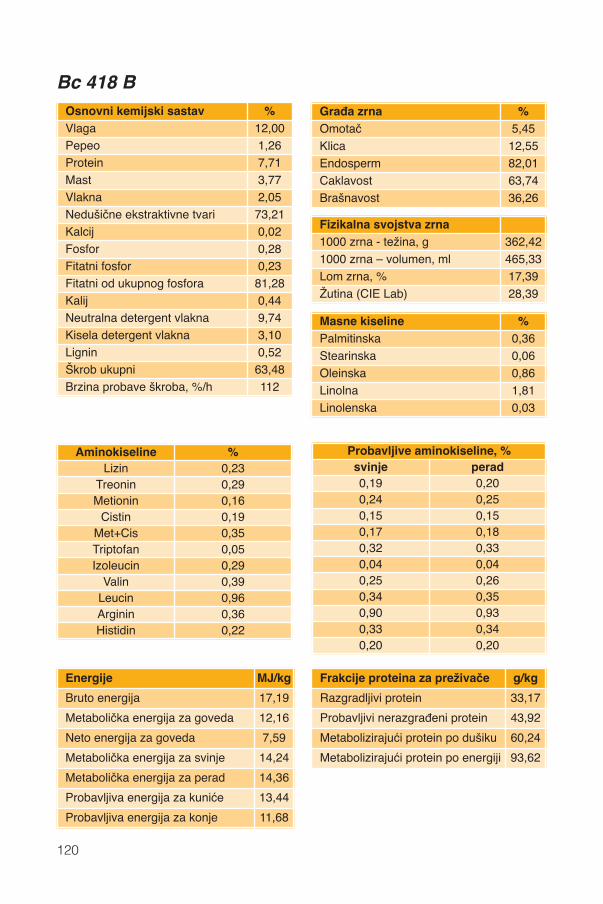
120
Bc 418 BOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,26Protein 7,71Mast 3,77Vlakna 2,05Nedušične ekstraktivne tvari 73,21Kalcij 0,02Fosfor 0,28Fitatni fosfor 0,23Fitatni od ukupnog fosfora 81,28Kalij 0,44Neutralna detergent vlakna 9,74Kisela detergent vlakna 3,10Lignin 0,52Škrob ukupni 63,48Brzina probave škroba, %/h 112
Aminokiseline %Lizin 0,23
Treonin 0,29Metionin 0,16
Cistin 0,19Met+Cis 0,35Triptofan 0,05Izoleucin 0,29
Valin 0,39Leucin 0,96Arginin 0,36Histidin 0,22
Energije MJ/kgBruto energija 17,19Metabolička energija za goveda 12,16Neto energija za goveda 7,59Metabolička energija za svinje 14,24Metabolička energija za perad 14,36Probavljiva energija za kuniće 13,44Probavljiva energija za konje 11,68
Frakcije proteina za preživače g/kgRazgradljivi protein 33,17Probavljivi nerazgrađeni protein 43,92Metabolizirajući protein po dušiku 60,24Metabolizirajući protein po energiji 93,62
Građa zrna %Omotač 5,45Klica 12,55Endosperm 82,01Caklavost 63,74Brašnavost 36,26
Fizikalna svojstva zrna1000 zrna - težina, g 362,421000 zrna – volumen, ml 465,33Lom zrna, % 17,39Žutina (CIE Lab) 28,39
Masne kiseline %Palmitinska 0,36Stearinska 0,06Oleinska 0,86Linolna 1,81Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,19 0,200,24 0,250,15 0,150,17 0,180,32 0,330,04 0,040,25 0,260,34 0,350,90 0,930,33 0,340,20 0,20
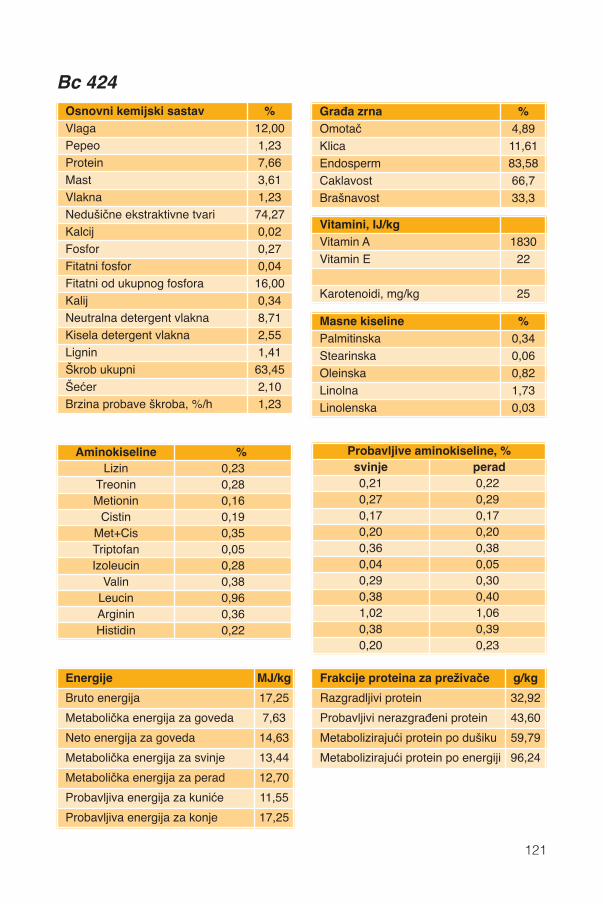
121
Bc 424Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 7,66Mast 3,61Vlakna 1,23Nedušične ekstraktivne tvari 74,27Kalcij 0,02Fosfor 0,27Fitatni fosfor 0,04Fitatni od ukupnog fosfora 16,00Kalij 0,34Neutralna detergent vlakna 8,71Kisela detergent vlakna 2,55Lignin 1,41Škrob ukupni 63,45Šećer 2,10Brzina probave škroba, %/h 1,23
Aminokiseline %Lizin 0,23
Treonin 0,28Metionin 0,16
Cistin 0,19Met+Cis 0,35Triptofan 0,05Izoleucin 0,28
Valin 0,38Leucin 0,96Arginin 0,36Histidin 0,22
Energije MJ/kgBruto energija 17,25Metabolička energija za goveda 7,63Neto energija za goveda 14,63Metabolička energija za svinje 13,44Metabolička energija za perad 12,70Probavljiva energija za kuniće 11,55Probavljiva energija za konje 17,25
Frakcije proteina za preživače g/kgRazgradljivi protein 32,92Probavljivi nerazgrađeni protein 43,60Metabolizirajući protein po dušiku 59,79Metabolizirajući protein po energiji 96,24
Građa zrna %Omotač 4,89Klica 11,61Endosperm 83,58Caklavost 66,7Brašnavost 33,3
Vitamini, IJ/kgVitamin A 1830Vitamin E 22
Karotenoidi, mg/kg 25
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,82Linolna 1,73Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,220,27 0,290,17 0,170,20 0,200,36 0,380,04 0,050,29 0,300,38 0,401,02 1,060,38 0,390,20 0,23
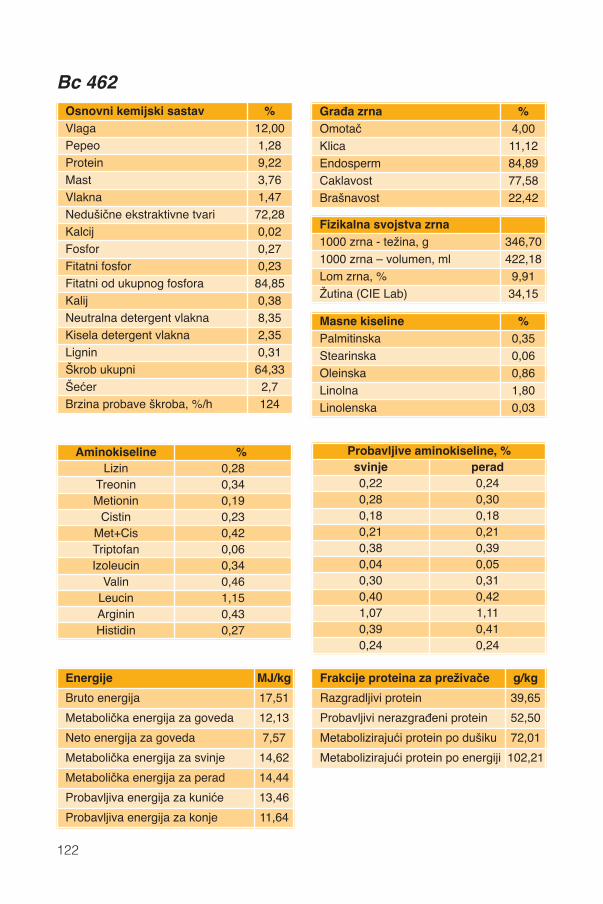
122
Bc 462
Aminokiseline %Lizin 0,28
Treonin 0,34Metionin 0,19
Cistin 0,23Met+Cis 0,42Triptofan 0,06Izoleucin 0,34
Valin 0,46Leucin 1,15Arginin 0,43Histidin 0,27
Energije MJ/kgBruto energija 17,51Metabolička energija za goveda 12,13Neto energija za goveda 7,57Metabolička energija za svinje 14,62Metabolička energija za perad 14,44Probavljiva energija za kuniće 13,46Probavljiva energija za konje 11,64
Frakcije proteina za preživače g/kgRazgradljivi protein 39,65Probavljivi nerazgrađeni protein 52,50Metabolizirajući protein po dušiku 72,01Metabolizirajući protein po energiji 102,21
Građa zrna %Omotač 4,00Klica 11,12Endosperm 84,89Caklavost 77,58Brašnavost 22,42
Fizikalna svojstva zrna1000 zrna - težina, g 346,701000 zrna – volumen, ml 422,18Lom zrna, % 9,91Žutina (CIE Lab) 34,15
Masne kiseline %Palmitinska 0,35Stearinska 0,06Oleinska 0,86Linolna 1,80Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,22 0,240,28 0,300,18 0,180,21 0,210,38 0,390,04 0,050,30 0,310,40 0,421,07 1,110,39 0,410,24 0,24
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,28Protein 9,22Mast 3,76Vlakna 1,47Nedušične ekstraktivne tvari 72,28Kalcij 0,02Fosfor 0,27Fitatni fosfor 0,23Fitatni od ukupnog fosfora 84,85Kalij 0,38Neutralna detergent vlakna 8,35Kisela detergent vlakna 2,35Lignin 0,31Škrob ukupni 64,33Šećer 2,7Brzina probave škroba, %/h 124
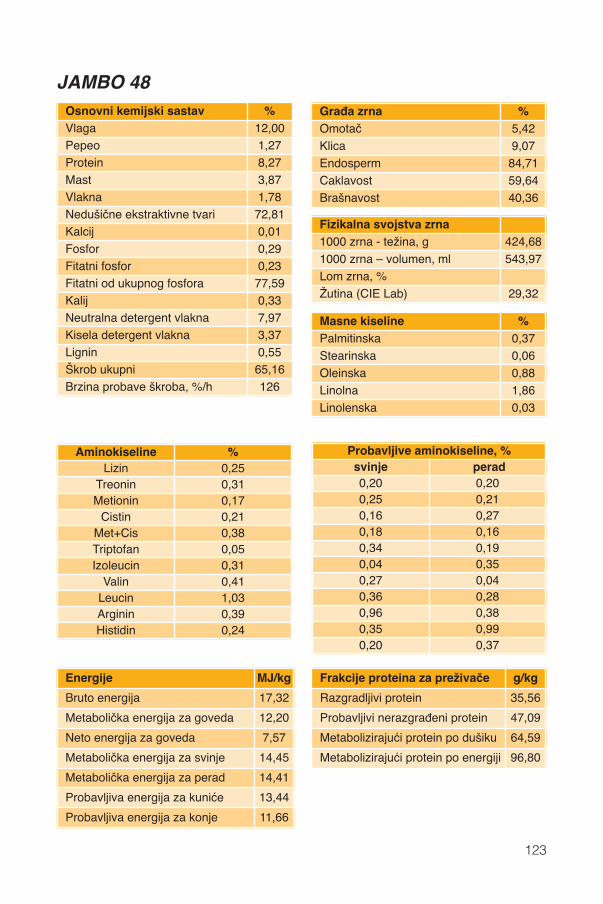
123
JAMBO 48Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,27Protein 8,27Mast 3,87Vlakna 1,78Nedušične ekstraktivne tvari 72,81Kalcij 0,01Fosfor 0,29Fitatni fosfor 0,23Fitatni od ukupnog fosfora 77,59Kalij 0,33Neutralna detergent vlakna 7,97Kisela detergent vlakna 3,37Lignin 0,55Škrob ukupni 65,16Brzina probave škroba, %/h 126
Aminokiseline %Lizin 0,25
Treonin 0,31Metionin 0,17
Cistin 0,21Met+Cis 0,38Triptofan 0,05Izoleucin 0,31
Valin 0,41Leucin 1,03Arginin 0,39Histidin 0,24
Energije MJ/kgBruto energija 17,32Metabolička energija za goveda 12,20Neto energija za goveda 7,57Metabolička energija za svinje 14,45Metabolička energija za perad 14,41Probavljiva energija za kuniće 13,44Probavljiva energija za konje 11,66
Frakcije proteina za preživače g/kgRazgradljivi protein 35,56Probavljivi nerazgrađeni protein 47,09Metabolizirajući protein po dušiku 64,59Metabolizirajući protein po energiji 96,80
Građa zrna %Omotač 5,42Klica 9,07Endosperm 84,71Caklavost 59,64Brašnavost 40,36
Fizikalna svojstva zrna1000 zrna - težina, g 424,681000 zrna – volumen, ml 543,97Lom zrna, %Žutina (CIE Lab) 29,32
Masne kiseline %Palmitinska 0,37Stearinska 0,06Oleinska 0,88Linolna 1,86Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,20 0,200,25 0,210,16 0,270,18 0,160,34 0,190,04 0,350,27 0,040,36 0,280,96 0,380,35 0,990,20 0,37
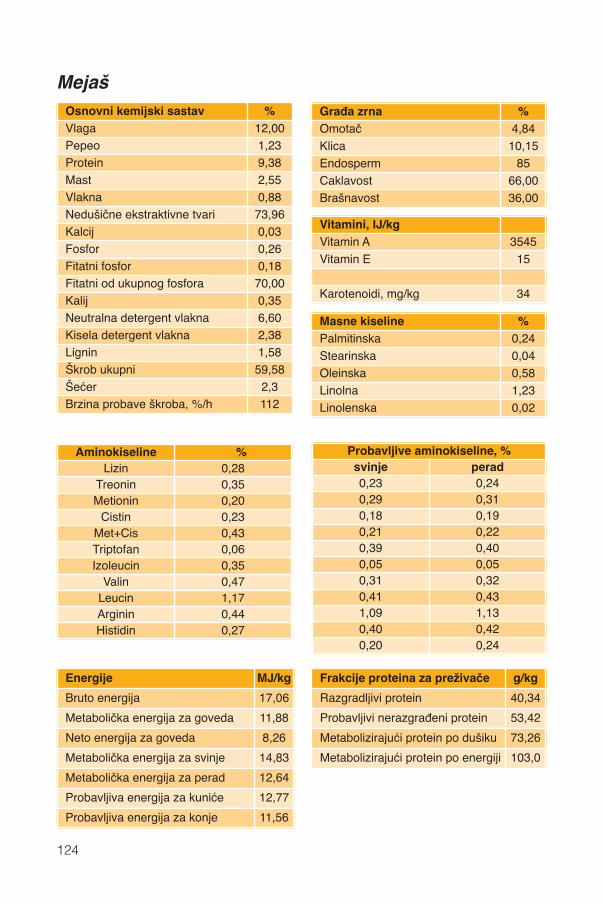
124
MejašOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 9,38Mast 2,55Vlakna 0,88Nedušične ekstraktivne tvari 73,96Kalcij 0,03Fosfor 0,26Fitatni fosfor 0,18Fitatni od ukupnog fosfora 70,00Kalij 0,35Neutralna detergent vlakna 6,60Kisela detergent vlakna 2,38Lignin 1,58Škrob ukupni 59,58Šećer 2,3Brzina probave škroba, %/h 112
Aminokiseline %Lizin 0,28
Treonin 0,35Metionin 0,20
Cistin 0,23Met+Cis 0,43Triptofan 0,06Izoleucin 0,35
Valin 0,47Leucin 1,17Arginin 0,44Histidin 0,27
Energije MJ/kgBruto energija 17,06Metabolička energija za goveda 11,88Neto energija za goveda 8,26Metabolička energija za svinje 14,83Metabolička energija za perad 12,64Probavljiva energija za kuniće 12,77Probavljiva energija za konje 11,56
Frakcije proteina za preživače g/kgRazgradljivi protein 40,34Probavljivi nerazgrađeni protein 53,42Metabolizirajući protein po dušiku 73,26Metabolizirajući protein po energiji 103,0
Građa zrna %Omotač 4,84Klica 10,15Endosperm 85Caklavost 66,00Brašnavost 36,00
Vitamini, IJ/kgVitamin A 3545Vitamin E 15
Karotenoidi, mg/kg 34
Masne kiseline %Palmitinska 0,24Stearinska 0,04Oleinska 0,58Linolna 1,23Linolenska 0,02
Probavljive aminokiseline, %svinje perad0,23 0,240,29 0,310,18 0,190,21 0,220,39 0,400,05 0,050,31 0,320,41 0,431,09 1,130,40 0,420,20 0,24
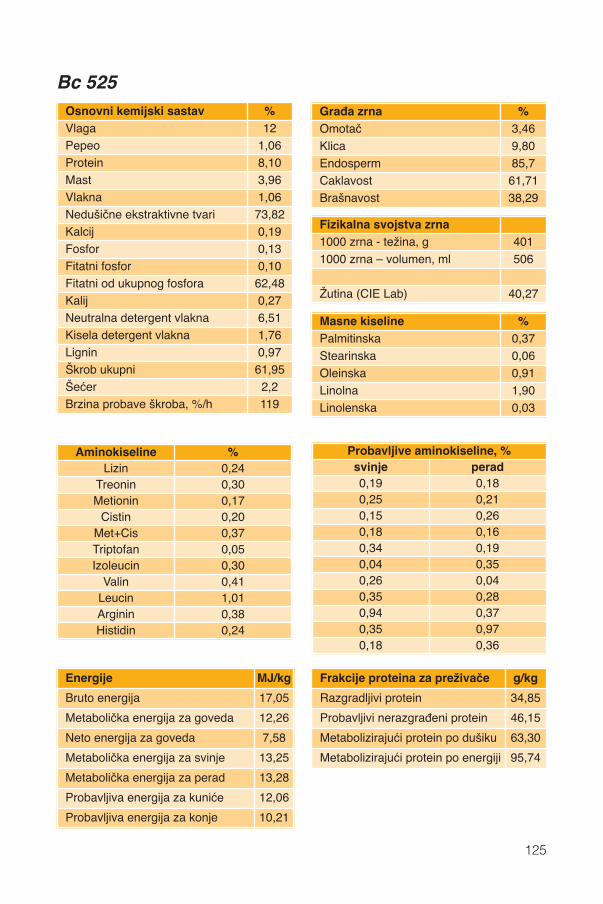
125
Bc 525Osnovni kemijski sastav %Vlaga 12Pepeo 1,06Protein 8,10Mast 3,96Vlakna 1,06Nedušične ekstraktivne tvari 73,82Kalcij 0,19Fosfor 0,13Fitatni fosfor 0,10Fitatni od ukupnog fosfora 62,48Kalij 0,27Neutralna detergent vlakna 6,51Kisela detergent vlakna 1,76Lignin 0,97Škrob ukupni 61,95Šećer 2,2Brzina probave škroba, %/h 119
Aminokiseline %Lizin 0,24
Treonin 0,30Metionin 0,17
Cistin 0,20Met+Cis 0,37Triptofan 0,05Izoleucin 0,30
Valin 0,41Leucin 1,01Arginin 0,38Histidin 0,24
Energije MJ/kgBruto energija 17,05Metabolička energija za goveda 12,26Neto energija za goveda 7,58Metabolička energija za svinje 13,25Metabolička energija za perad 13,28Probavljiva energija za kuniće 12,06Probavljiva energija za konje 10,21
Frakcije proteina za preživače g/kgRazgradljivi protein 34,85Probavljivi nerazgrađeni protein 46,15Metabolizirajući protein po dušiku 63,30Metabolizirajući protein po energiji 95,74
Građa zrna %Omotač 3,46Klica 9,80Endosperm 85,7Caklavost 61,71Brašnavost 38,29
Fizikalna svojstva zrna1000 zrna - težina, g 4011000 zrna – volumen, ml 506
Žutina (CIE Lab) 40,27
Masne kiseline %Palmitinska 0,37Stearinska 0,06Oleinska 0,91Linolna 1,90Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,19 0,180,25 0,210,15 0,260,18 0,160,34 0,190,04 0,350,26 0,040,35 0,280,94 0,370,35 0,970,18 0,36
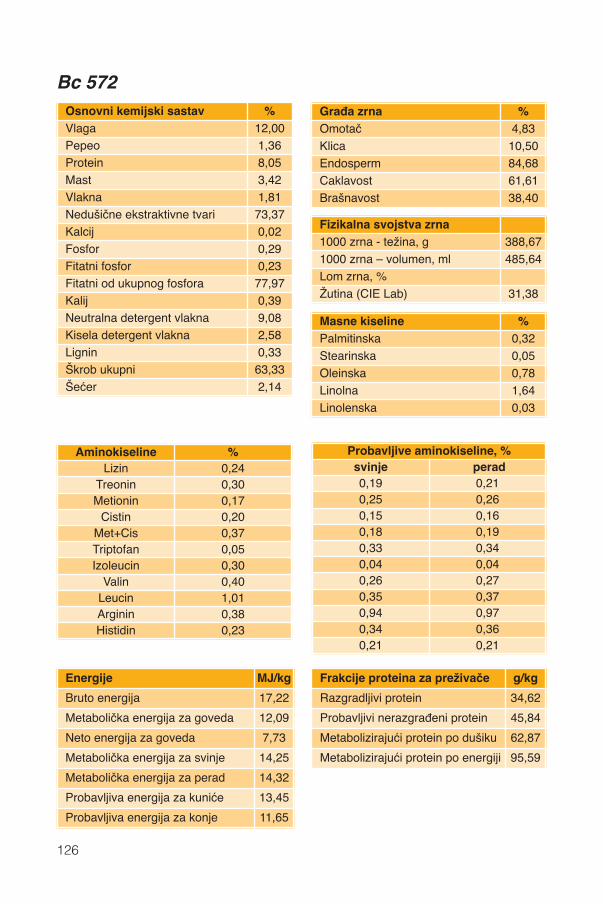
126
Bc 572Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,36Protein 8,05Mast 3,42Vlakna 1,81Nedušične ekstraktivne tvari 73,37Kalcij 0,02Fosfor 0,29Fitatni fosfor 0,23Fitatni od ukupnog fosfora 77,97Kalij 0,39Neutralna detergent vlakna 9,08Kisela detergent vlakna 2,58Lignin 0,33Škrob ukupni 63,33Šećer 2,14
Aminokiseline %Lizin 0,24
Treonin 0,30Metionin 0,17
Cistin 0,20Met+Cis 0,37Triptofan 0,05Izoleucin 0,30
Valin 0,40Leucin 1,01Arginin 0,38Histidin 0,23
Energije MJ/kgBruto energija 17,22Metabolička energija za goveda 12,09Neto energija za goveda 7,73Metabolička energija za svinje 14,25Metabolička energija za perad 14,32Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,65
Frakcije proteina za preživače g/kgRazgradljivi protein 34,62Probavljivi nerazgrađeni protein 45,84Metabolizirajući protein po dušiku 62,87Metabolizirajući protein po energiji 95,59
Građa zrna %Omotač 4,83Klica 10,50Endosperm 84,68Caklavost 61,61Brašnavost 38,40
Fizikalna svojstva zrna1000 zrna - težina, g 388,671000 zrna – volumen, ml 485,64Lom zrna, %Žutina (CIE Lab) 31,38
Masne kiseline %Palmitinska 0,32Stearinska 0,05Oleinska 0,78Linolna 1,64Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,19 0,210,25 0,260,15 0,160,18 0,190,33 0,340,04 0,040,26 0,270,35 0,370,94 0,970,34 0,360,21 0,21
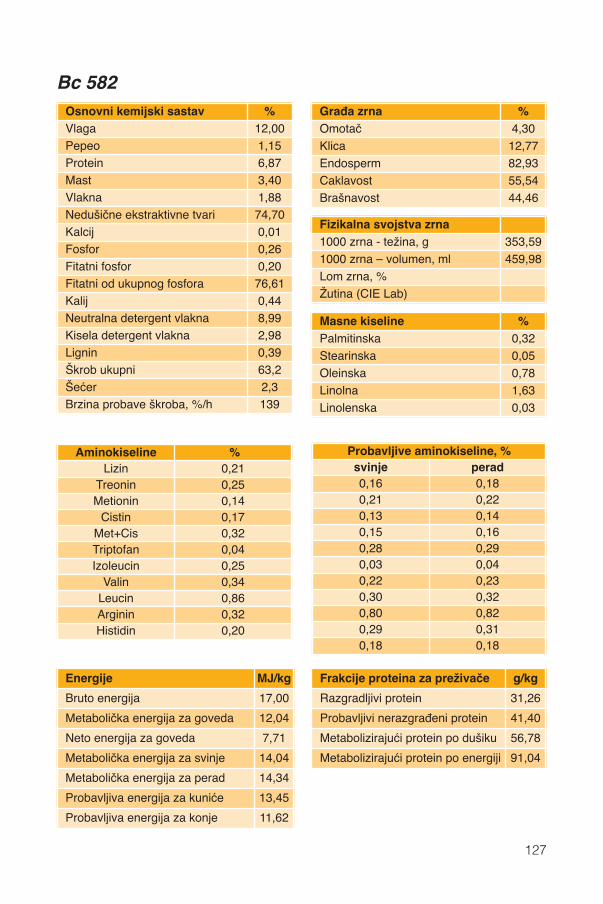
127
Bc 582Građa zrna %Omotač 4,30Klica 12,77Endosperm 82,93Caklavost 55,54Brašnavost 44,46
Energije MJ/kgBruto energija 17,00Metabolička energija za goveda 12,04Neto energija za goveda 7,71Metabolička energija za svinje 14,04Metabolička energija za perad 14,34Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,62
Frakcije proteina za preživače g/kgRazgradljivi protein 31,26Probavljivi nerazgrađeni protein 41,40Metabolizirajući protein po dušiku 56,78Metabolizirajući protein po energiji 91,04
Fizikalna svojstva zrna1000 zrna - težina, g 353,591000 zrna – volumen, ml 459,98Lom zrna, %Žutina (CIE Lab)
Masne kiseline %Palmitinska 0,32Stearinska 0,05Oleinska 0,78Linolna 1,63Linolenska 0,03
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,15Protein 6,87Mast 3,40Vlakna 1,88Nedušične ekstraktivne tvari 74,70Kalcij 0,01Fosfor 0,26Fitatni fosfor 0,20Fitatni od ukupnog fosfora 76,61Kalij 0,44Neutralna detergent vlakna 8,99Kisela detergent vlakna 2,98Lignin 0,39Škrob ukupni 63,2Šećer 2,3Brzina probave škroba, %/h 139
Aminokiseline %Lizin 0,21
Treonin 0,25Metionin 0,14
Cistin 0,17Met+Cis 0,32Triptofan 0,04Izoleucin 0,25
Valin 0,34Leucin 0,86Arginin 0,32Histidin 0,20
Probavljive aminokiseline, %svinje perad0,16 0,180,21 0,220,13 0,140,15 0,160,28 0,290,03 0,040,22 0,230,30 0,320,80 0,820,29 0,310,18 0,18
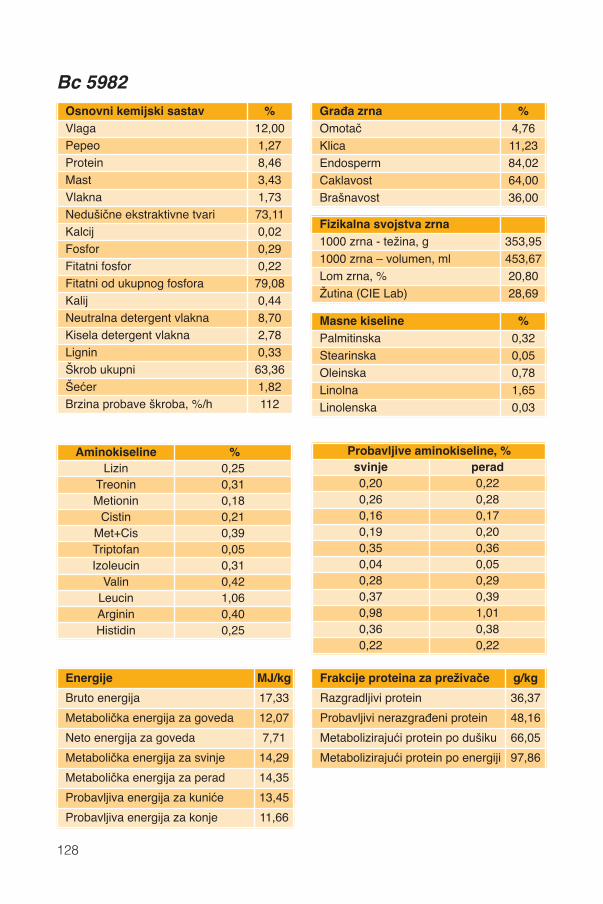
128
Bc 5982Građa zrna %Omotač 4,76Klica 11,23Endosperm 84,02Caklavost 64,00Brašnavost 36,00
Energije MJ/kgBruto energija 17,33Metabolička energija za goveda 12,07Neto energija za goveda 7,71Metabolička energija za svinje 14,29Metabolička energija za perad 14,35Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,66
Frakcije proteina za preživače g/kgRazgradljivi protein 36,37Probavljivi nerazgrađeni protein 48,16Metabolizirajući protein po dušiku 66,05Metabolizirajući protein po energiji 97,86
Fizikalna svojstva zrna1000 zrna - težina, g 353,951000 zrna – volumen, ml 453,67Lom zrna, % 20,80Žutina (CIE Lab) 28,69
Masne kiseline %Palmitinska 0,32Stearinska 0,05Oleinska 0,78Linolna 1,65Linolenska 0,03
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,27Protein 8,46Mast 3,43Vlakna 1,73Nedušične ekstraktivne tvari 73,11Kalcij 0,02Fosfor 0,29Fitatni fosfor 0,22Fitatni od ukupnog fosfora 79,08Kalij 0,44Neutralna detergent vlakna 8,70Kisela detergent vlakna 2,78Lignin 0,33Škrob ukupni 63,36Šećer 1,82Brzina probave škroba, %/h 112
Aminokiseline %Lizin 0,25
Treonin 0,31Metionin 0,18
Cistin 0,21Met+Cis 0,39Triptofan 0,05Izoleucin 0,31
Valin 0,42Leucin 1,06Arginin 0,40Histidin 0,25
Probavljive aminokiseline, %svinje perad0,20 0,220,26 0,280,16 0,170,19 0,200,35 0,360,04 0,050,28 0,290,37 0,390,98 1,010,36 0,380,22 0,22
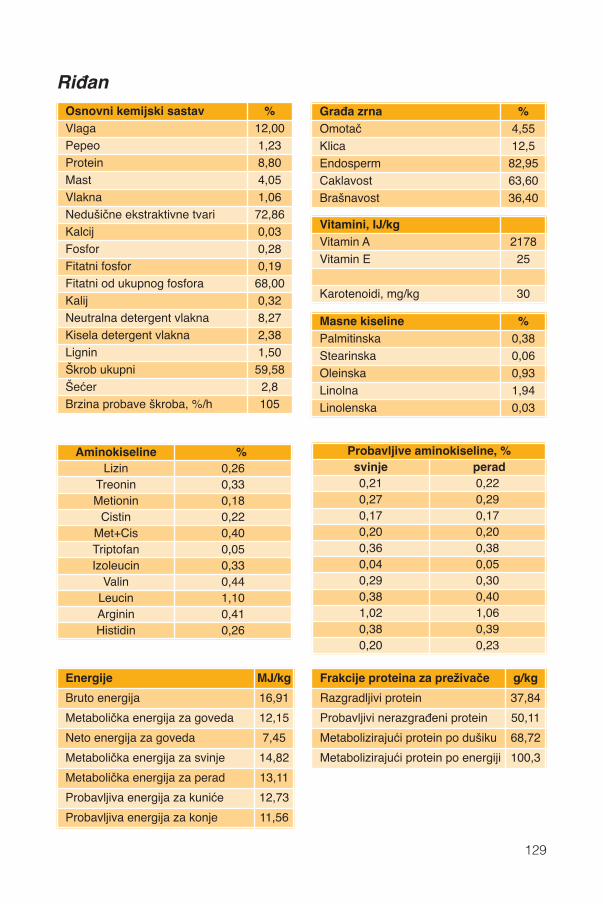
129
RiđanOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 8,80Mast 4,05Vlakna 1,06Nedušične ekstraktivne tvari 72,86Kalcij 0,03Fosfor 0,28Fitatni fosfor 0,19Fitatni od ukupnog fosfora 68,00Kalij 0,32Neutralna detergent vlakna 8,27Kisela detergent vlakna 2,38Lignin 1,50Škrob ukupni 59,58Šećer 2,8Brzina probave škroba, %/h 105
Aminokiseline %Lizin 0,26
Treonin 0,33Metionin 0,18
Cistin 0,22Met+Cis 0,40Triptofan 0,05Izoleucin 0,33
Valin 0,44Leucin 1,10Arginin 0,41Histidin 0,26
Energije MJ/kgBruto energija 16,91Metabolička energija za goveda 12,15Neto energija za goveda 7,45Metabolička energija za svinje 14,82Metabolička energija za perad 13,11Probavljiva energija za kuniće 12,73Probavljiva energija za konje 11,56
Frakcije proteina za preživače g/kgRazgradljivi protein 37,84Probavljivi nerazgrađeni protein 50,11Metabolizirajući protein po dušiku 68,72Metabolizirajući protein po energiji 100,3
Građa zrna %Omotač 4,55Klica 12,5Endosperm 82,95Caklavost 63,60Brašnavost 36,40
Vitamini, IJ/kgVitamin A 2178Vitamin E 25
Karotenoidi, mg/kg 30
Masne kiseline %Palmitinska 0,38Stearinska 0,06Oleinska 0,93Linolna 1,94Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,21 0,220,27 0,290,17 0,170,20 0,200,36 0,380,04 0,050,29 0,300,38 0,401,02 1,060,38 0,390,20 0,23
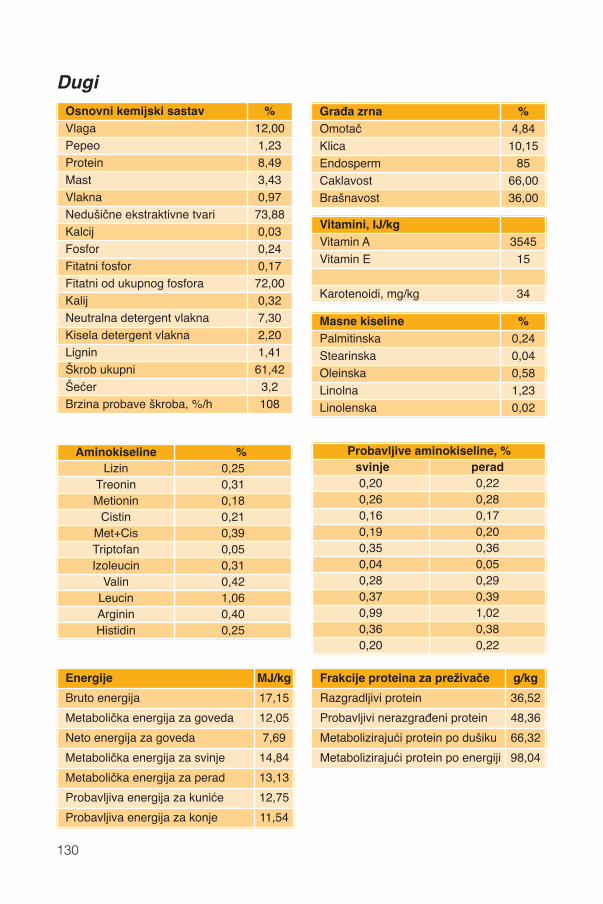
130
DugiOsnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 8,49Mast 3,43Vlakna 0,97Nedušične ekstraktivne tvari 73,88Kalcij 0,03Fosfor 0,24Fitatni fosfor 0,17Fitatni od ukupnog fosfora 72,00Kalij 0,32Neutralna detergent vlakna 7,30Kisela detergent vlakna 2,20Lignin 1,41Škrob ukupni 61,42Šećer 3,2Brzina probave škroba, %/h 108
Aminokiseline %Lizin 0,25
Treonin 0,31Metionin 0,18
Cistin 0,21Met+Cis 0,39Triptofan 0,05Izoleucin 0,31
Valin 0,42Leucin 1,06Arginin 0,40Histidin 0,25
Energije MJ/kgBruto energija 17,15Metabolička energija za goveda 12,05Neto energija za goveda 7,69Metabolička energija za svinje 14,84Metabolička energija za perad 13,13Probavljiva energija za kuniće 12,75Probavljiva energija za konje 11,54
Frakcije proteina za preživače g/kgRazgradljivi protein 36,52Probavljivi nerazgrađeni protein 48,36Metabolizirajući protein po dušiku 66,32Metabolizirajući protein po energiji 98,04
Građa zrna %Omotač 4,84Klica 10,15Endosperm 85Caklavost 66,00Brašnavost 36,00
Vitamini, IJ/kgVitamin A 3545Vitamin E 15
Karotenoidi, mg/kg 34
Masne kiseline %Palmitinska 0,24Stearinska 0,04Oleinska 0,58Linolna 1,23Linolenska 0,02
Probavljive aminokiseline, %svinje perad0,20 0,220,26 0,280,16 0,170,19 0,200,35 0,360,04 0,050,28 0,290,37 0,390,99 1,020,36 0,380,20 0,22

131
Bc 616Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 7,66Mast 3,61Vlakna 1,23Nedušične ekstraktivne tvari 74,27Kalcij 0,02Fosfor 0,27Fitatni fosfor 0,04Fitatni od ukupnog fosfora 16,00Kalij 0,34Neutralna detergent vlakna 8,71Kisela detergent vlakna 2,55Lignin 1,41Škrob ukupni 63,45Šećer 2,10Brzina probave škroba, %/h 1,23
Aminokiseline %Lizin 0,23
Treonin 0,28Metionin 0,16
Cistin 0,19Met+Cis 0,35Triptofan 0,05Izoleucin 0,28
Valin 0,38Leucin 0,96Arginin 0,36Histidin 0,22
Energije MJ/kgBruto energija 17,25Metabolička energija za goveda 7,63Neto energija za goveda 14,63Metabolička energija za svinje 13,44Metabolička energija za perad 12,70Probavljiva energija za kuniće 11,55Probavljiva energija za konje 17,25
Frakcije proteina za preživače g/kgRazgradljivi protein 32,92Probavljivi nerazgrađeni protein 43,60Metabolizirajući protein po dušiku 59,79Metabolizirajući protein po energiji 96,24
Građa zrna %Omotač 4,89Klica 11,61Endosperm 83,58Caklavost 66,7Brašnavost 33,3
Vitamini, IJ/kgVitamin A 1830Vitamin E 25
Karotenoidi, mg/kg 26
Masne kiseline %Palmitinska 0,34Stearinska 0,06Oleinska 0,82Linolna 1,73Linolenska 0,03
Probavljive aminokiseline, %svinje perad0,18 0,200,24 0,250,15 0,150,17 0,180,32 0,330,04 0,040,25 0,260,33 0,350,89 0,920,33 0,340,20 0,20
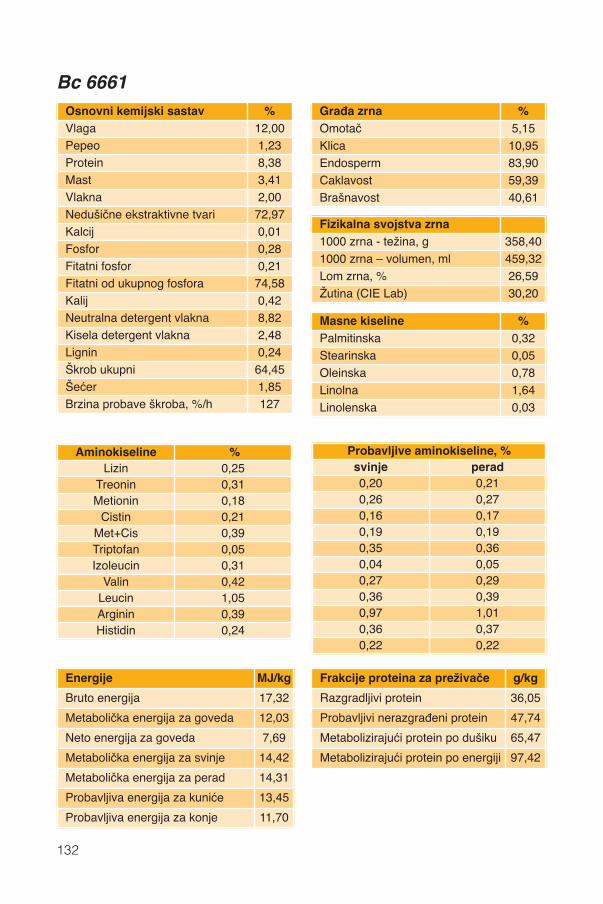
132
Bc 6661Građa zrna %Omotač 5,15Klica 10,95Endosperm 83,90Caklavost 59,39Brašnavost 40,61
Energije MJ/kgBruto energija 17,32Metabolička energija za goveda 12,03Neto energija za goveda 7,69Metabolička energija za svinje 14,42Metabolička energija za perad 14,31Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,70
Frakcije proteina za preživače g/kgRazgradljivi protein 36,05Probavljivi nerazgrađeni protein 47,74Metabolizirajući protein po dušiku 65,47Metabolizirajući protein po energiji 97,42
Fizikalna svojstva zrna1000 zrna - težina, g 358,401000 zrna – volumen, ml 459,32Lom zrna, % 26,59Žutina (CIE Lab) 30,20
Masne kiseline %Palmitinska 0,32Stearinska 0,05Oleinska 0,78Linolna 1,64Linolenska 0,03
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,23Protein 8,38Mast 3,41Vlakna 2,00Nedušične ekstraktivne tvari 72,97Kalcij 0,01Fosfor 0,28Fitatni fosfor 0,21Fitatni od ukupnog fosfora 74,58Kalij 0,42Neutralna detergent vlakna 8,82Kisela detergent vlakna 2,48Lignin 0,24Škrob ukupni 64,45Šećer 1,85Brzina probave škroba, %/h 127
Aminokiseline %Lizin 0,25
Treonin 0,31Metionin 0,18
Cistin 0,21Met+Cis 0,39Triptofan 0,05Izoleucin 0,31
Valin 0,42Leucin 1,05Arginin 0,39Histidin 0,24
Probavljive aminokiseline, %svinje perad0,20 0,210,26 0,270,16 0,170,19 0,190,35 0,360,04 0,050,27 0,290,36 0,390,97 1,010,36 0,370,22 0,22
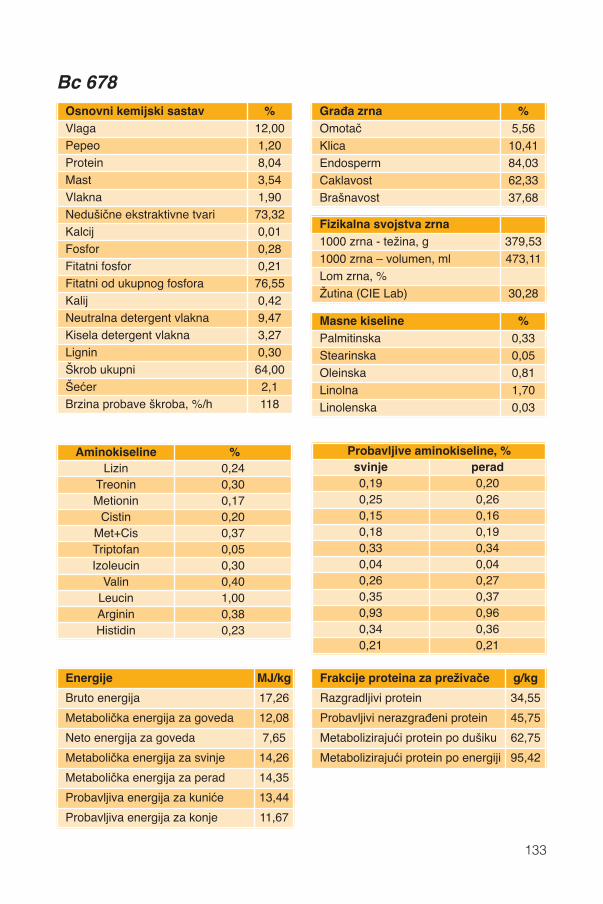
133
Bc 678Građa zrna %Omotač 5,56Klica 10,41Endosperm 84,03Caklavost 62,33Brašnavost 37,68
Energije MJ/kgBruto energija 17,26Metabolička energija za goveda 12,08Neto energija za goveda 7,65Metabolička energija za svinje 14,26Metabolička energija za perad 14,35Probavljiva energija za kuniće 13,44Probavljiva energija za konje 11,67
Frakcije proteina za preživače g/kgRazgradljivi protein 34,55Probavljivi nerazgrađeni protein 45,75Metabolizirajući protein po dušiku 62,75Metabolizirajući protein po energiji 95,42
Fizikalna svojstva zrna1000 zrna - težina, g 379,531000 zrna – volumen, ml 473,11Lom zrna, %Žutina (CIE Lab) 30,28
Masne kiseline %Palmitinska 0,33Stearinska 0,05Oleinska 0,81Linolna 1,70Linolenska 0,03
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,20Protein 8,04Mast 3,54Vlakna 1,90Nedušične ekstraktivne tvari 73,32Kalcij 0,01Fosfor 0,28Fitatni fosfor 0,21Fitatni od ukupnog fosfora 76,55Kalij 0,42Neutralna detergent vlakna 9,47Kisela detergent vlakna 3,27Lignin 0,30Škrob ukupni 64,00Šećer 2,1Brzina probave škroba, %/h 118
Aminokiseline %Lizin 0,24
Treonin 0,30Metionin 0,17
Cistin 0,20Met+Cis 0,37Triptofan 0,05Izoleucin 0,30
Valin 0,40Leucin 1,00Arginin 0,38Histidin 0,23
Probavljive aminokiseline, %svinje perad0,19 0,200,25 0,260,15 0,160,18 0,190,33 0,340,04 0,040,26 0,270,35 0,370,93 0,960,34 0,360,21 0,21
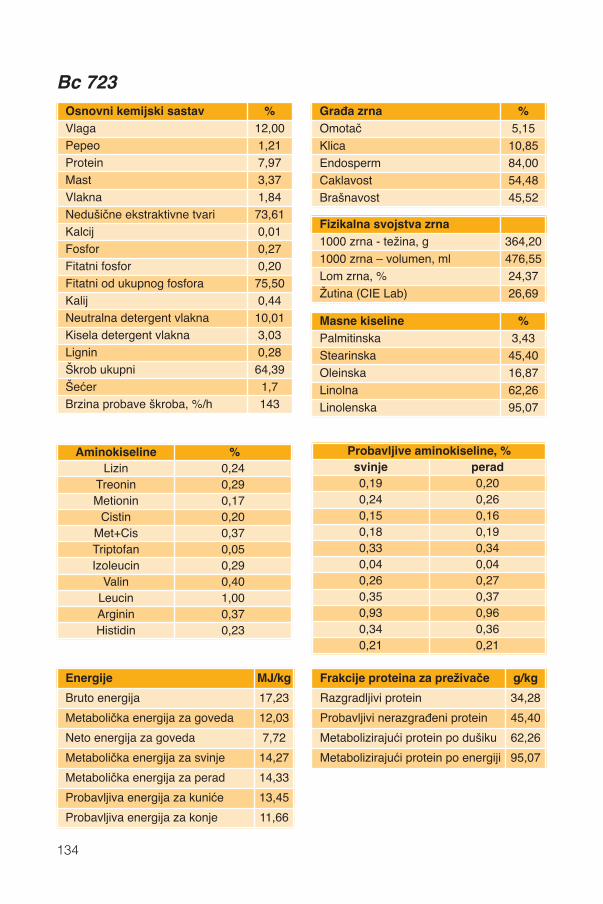
134
Bc 723Građa zrna %Omotač 5,15Klica 10,85Endosperm 84,00Caklavost 54,48Brašnavost 45,52
Energije MJ/kgBruto energija 17,23Metabolička energija za goveda 12,03Neto energija za goveda 7,72Metabolička energija za svinje 14,27Metabolička energija za perad 14,33Probavljiva energija za kuniće 13,45Probavljiva energija za konje 11,66
Frakcije proteina za preživače g/kgRazgradljivi protein 34,28Probavljivi nerazgrađeni protein 45,40Metabolizirajući protein po dušiku 62,26Metabolizirajući protein po energiji 95,07
Fizikalna svojstva zrna1000 zrna - težina, g 364,201000 zrna – volumen, ml 476,55Lom zrna, % 24,37Žutina (CIE Lab) 26,69
Masne kiseline %Palmitinska 3,43Stearinska 45,40Oleinska 16,87Linolna 62,26Linolenska 95,07
Osnovni kemijski sastav %Vlaga 12,00Pepeo 1,21Protein 7,97Mast 3,37Vlakna 1,84Nedušične ekstraktivne tvari 73,61Kalcij 0,01Fosfor 0,27Fitatni fosfor 0,20Fitatni od ukupnog fosfora 75,50Kalij 0,44Neutralna detergent vlakna 10,01Kisela detergent vlakna 3,03Lignin 0,28Škrob ukupni 64,39Šećer 1,7Brzina probave škroba, %/h 143
Aminokiseline %Lizin 0,24
Treonin 0,29Metionin 0,17
Cistin 0,20Met+Cis 0,37Triptofan 0,05Izoleucin 0,29
Valin 0,40Leucin 1,00Arginin 0,37Histidin 0,23
Probavljive aminokiseline, %svinje perad0,19 0,200,24 0,260,15 0,160,18 0,190,33 0,340,04 0,040,26 0,270,35 0,370,93 0,960,34 0,360,21 0,21

135
1. Adom, K.K., R.H. Liu (2002): Antioxidant activity ofgrains. J. Agric. Food Chem. 50: 6182-6187.
2. Agama-Acevedo, E., Barba de la Rosa, A.P., Mén-dez-Montealvo, G., Bello-Pérez1, L.A. 2008. Physi-cochemical and biochemical characterization ofstarch granules isolated of pigmented maize hybrids.Starch – Stärke 60, 433–441.
3. Amerah A.M., Ravindran V., Lentle R.G., ThomasD.G. (2007): Feed particle size: Implications on thedigestion and performance of poultry. World’s PoultryScience Journal, Vol. 63, September 2007.
4. Bach Knudsen, K.E., Nygaard Lærke, H., SteenfeldtS., Skou Hedemann M., Jørgensen, H. (2006): Invivo methods to study the digestion of starch in pigsand poultry. Animal Feed Science and Technology,130: 114-135.
5. Barea, J.M. (2015). Interactions among plants, arbu-scular mycorrhizal and mycotoxigenic fungi related tofood crop health in a scenario of climate change. In.Climate Change and Mycotoxins. (Eds. Botana,L.M.,Saintz, M.J.). Walter de Gruyter GmbH, Berlin/Bo-ston. 54-70.
6. Bhuiyan, M.M., Islam, A.J., Cowieson, A.J., Iji, P.A.2012. Effect of sources and processing on maizegrain quality and nutritional value for broiler chickens.I. Heat treatment and physicochemical properties.Poult. Sci. 6, 101-116.
7. Burt A. J., Grainger C. M., Young J. C., Shelp B. J.,Lee E. A. (2010). Impact of postharvesting handlingon carotenoid concentration and composition in high-carotenoid maize (Zea mays L.) kernels. Journal ofAgricultural and Food Chemistry. 58: 8286-8292.
8. Combs, Jr.,G.F. (2008): The Vitamins . FundamentalAspects in Nutrition and Health. Third Edition. Else-vier Academic Press 30 Corporate Drive, Suite 400,Burlington, MA 01803, USA.
9. Čuklić, D., Svečnjak, Z. Grbeša, D. (2015). Relations-hip between the physical properties of maize kerneland metabolizable energy for pigs. Agriculturae Con-spectus Scientificus. 80(2) 125-131.
10. D’Alfonso, T.H., 2003. Factors affecting ileal digesti-ble energy of maize in poultry diets. In: Corbett, J.L.(Ed.), Recent Advances in Animal Nutrition in Austra-lia, Volume 14, University of New England, Armidale,pp.151-156.
11. Dhital, S., Shrestha, A. K., Gidley, M. J., 2010. Rela-tionship between granule size and in vitro digestibilityof maize and potato starches. Carbohydr. Polymer.82, 480-488.
12. Dona, A.C., Pages, G., Gilbert, R.G., Kuchel, P.W.,2010. Digestion of starch: In vivo and in vitro kineticmodels used to characterise oligosaccharide or glu-cose release. Carbohydr. Polymer. 80, 599–617
13. FAOSTAT, Food and Agricurture Organization of theUnited Nations. <http//:www.faostat.fao.org.>Pristu-pljeno 16.12.2015.
14. Ferraretto L. F., Crump P. M., Shaver R. D. (2015a).Effect of ensiling time and exogenous protease addi-tion to whole-plant corn silage of various hybrids, ma-turities, and chop lengths on nitrogen fractions andruminal in vitro starch digestibility. Journal of DairyScience 98: 8869-8881.
15. Ferraretto L.F., Shaver R.D., Massie S., Singo R.,Taysom D.M., Brouillette J.P., (2015b). Effect of ensi-ling time and hybrid type on fermentation profile, ni-trogen fractions, and ruminal in vitro starch and ne-
utral detergent fiber digestibility in whole-plant cornsilage. The Professional Animal Scientist 31: 146–152.
16. Fox G., Manley M. (2009). Hardness methods for te-sting maize kernels. Journal of agricultural and foodchemistry 57: 5647-5657.
17. Fox G., Manley M. (2009). Hardness Methods for Te-sting Maize Kernels. Journal of Agricultural and FoodChemistry 57:5647-5657-
18. Giuberti G., Gallo A., Masoero F., Ferraretto L. F.,Hoffman P. C., Shaver R. D. (2014). Factors affectingstarch utilization in large animal food production sy-stem: a review. Starch/Stärke 66: 72-90.
19. Grbeša D. (2004): Metode procjene i tablice kemij-skog sastava i hranjive vrijednosti krepkih krmiva, Hr-vatsko agronomsko društvo, Zagreb.
20. Grbeša D. (2008): Kemijski sastav i fizikalna svojstvazrna Bc hibrida kukuruza u hranidbi životinja, Bc insti-tut za oplemenjivanje i proizvodnju bilja d.d., Zagreb.
21. Grbeša, D. (2013): Aflatoksični kukuruz u hranidbimliječnih krava. Mljekarski list, br. 02 (7), specijalniprilog, 2013; str. 2-8
22. Grbeša, D., Duvnjak, M., Kljak K. (2014): Ublažava-nje pojave mikotoksina u krmi i njihovih učinaka na ži-votinje. Krmiva 56 (2014), Zagreb, 1:15-32
23. Gutierrez del Alamo, A., Verstegen, M.W.A., Den Har-tog, L.A., Perez de Ayala, P., Villamide, M.J. (2009):Wheat starch digestion rate affects broiler performan-ce, Poultry Science 88: 1666-1675.
24. Gehring, C.K., Bedford, M. R., Cowieson, A. J., Do-zier III, W.A., 2012. Effects of maize source on the re-lationship between in vitro assays and ileal nutrientdigestibility. Poult. Sci. 91, 1908-1914.
25. Hasjim, J., Srichuwong, S., Scott, M. P., & Jane, J. L.(2009). Kernel composition, starch structure, and en-zyme digestibility of opaque-2 maize and quality pro-tein maize. Journal of agricultural and food chemistry,57: 2049-2055.
26. Hoffman P.C., Esser N.M., Shaver R.D., Coblentz W.,Scott M.P., Bodnar A.L., Schmidt R., Charley B.(2011). Influence of ensiling time and inoculation onalteration of the starch-protein matrix in high-moisturecorn. Journal of Dairy Science 94: 2465–2474.
27. Kljak K. (2012). Antioksidacijska i pigmentacijskasvojstva karotenoida zrna hibrida kukuruza u proizvo-dnji jaja. Doktorska disertacija. Agronomski fakultet.Zagreb.
28. Kljak K., Grbeša D. (2015). Carotenoid content andantioxidant activity of hexane extracts from selectedCroatian corn hybrids. Food Chemistry. 167: 402-408.
29. Kljak K., Grbeša D., Aleuš D. (2011). Relationshipsbetween kernel physical properties and zein contentin corn hybrids. Bulletin of University of AgriculturalSciences and Veterinary Medicine Cluj-Napoca. Agri-culture, 68: 188-194.
30. Kljak K., Grbeša D., Karolyi D. (2009). Estimation ofcarotenoid content from colour analysis of corngrains. Stočarstvo. 63:103-111.
31. Kotarski S. F., Waniska R. D., Thurn, K. K. (1992).Starch hydrolysis by the ruminal microflora. The Jour-nal of nutrition 122: 178-190.
32. Kovalsky Paris, M.P., Liu, Y.J., Nahrer, K., Binder,E.M. (2015). Climate change impacts on mycotoxinproduction. In. Climate Change and Mycotoxins.(Eds. Botana,L.M., Saintz, M.J.). Walter de GruyterGmbH, Berlin/Boston. 133-151.
Popis najvažnije korištene literature

136
33. Kung L., Windle M. C., Walker N. (2014). The effectof an exogenous protease on the fermentation andnutritive value of high-moisture corn. Journal of dairyscience 97: 1707-1712.
34. Kurilich, A.C., J.A. Juvik (1999): Quantification of ca-rotenoid and tocopherol antioxidants in Zea mays. J.Agri. Food Chem. 47:1948-1955.
35. Lee, C.-Y., B.-D. Lee, J.-C. Na, G. An (2010): Carote-noid accumulation and their antioxidant activity inspent laying hens as affected by polarity and feedingperiod. Asian-Aust. J. Anim. Sci. 23: 799-805.
36. Li,X., Yang, X.B. (2015). Impact of climate change ongenetically engineered plants and mycotoxigenic fun-gi in the north central region of the US. In. ClimateChange and Mycotoxins. (Eds. Botana,L.M., Saintz,M.J.). Walter de Gruyter GmbH, Berlin/Boston. 30-51.
37. Liu, S.I., Selle, P.H., Gill, R.J., Cowienson, A.J., 2013.Starch and nitrogen digestion kinetics influencegrowth performance and nitrogen retention in redsorghum-based broiler diets. In: Proceedings of 24thAnnual Australian Poultry Science Symposium, Uni-versity of Sydney, Camden, pp.115-118.
38. Lozano-Alejo, N., G. Vázquez Carrillo, K. Pixley, N.Palacios-Rojas (2007): Physical properties and caro-tenoid content of maize kernels and its nixtamalizedsnacks. Innovat. Food Sci. Emerg. Tech. 8:385-389.
39. Masoero, F., Gallo, A., Zanfi, C., Giuberti, G., Spang-hero, M. 2011. Effect of nitrogen fertilization on che-mical composition and rumen fermentation of differentparts of plants of three maize hybrids. Anim. FeedSci. Technol. 164, 207–216
40. Matilla, P., J.-M. Pihlava, J. Hellström (2005): Con-tents of phenolic acids, alkyl- and alkenilresorcinols,and avenanthramides in commercial grain products.J. Agric. Food Chem. 83: 8290-8295.
41. Moore, S.M., Stadler, K.J., Beitz, D.C., Stahl, C.H.,Fithian, W.A., Bregendahl, K. (2008). The correlationof chemical and physical corn kernel traits with pro-duction performance in broiler chickens and layinghens. Poultry Science. 87:665-676.
42. Oba M., & Allen M. (2008) The impact of improvingNDF digestibility of corn silage on diary cow perfor-mance. http://dairy.ifas.ufl.edu/rns/2011/10Oba.pdf.
43. Pangga, I.B. Salvacion, A.R., Cumagun, C.J.R.(2015). Climate change and plant diseases causedby mycotoxigenic fungi: implications for food security.In. Climate Change and Mycotoxins. (Eds.Botana,L.M., Saintz, M.J.). Walter de Gruyter GmbH,Berlin/Boston. 2-28.
44. Paulis J. W., Wall J. S. (1971). Fractionation and pro-perties of alkylated-reduced corn glutelin proteins.Biochimica et Biophysica Acta (BBA)-Protein Structu-re 251: 57-69
45. Paulsen M. R., Watson S. A., Singh M., White P. J.,Johnson L.A. (2003). Measurement and maintenanceof corn quality. U: Corn: Chemistry and technology,(Ed. 2) (P. J. White, L. A. Johnson ured.) AmericanAssociation of Cereal Chemists, St. Paul, Minnesota,SAD,159-219
46. Peyrat J., Nozière P., Le Morvan A., Férard A., ProtinP. V., Baumont R. (2014). Effects of ensiling maizeand sample conditioning on in situ rumen degradationof dry matter, starch and fibre. Animal Feed Scienceand Technology 196: 12-21.
47. Philippeau, C., Landry, J., & Michalet‐Doreau, B.,2000. Influence of the protein distribution of maizeendosperm on ruminal starch degradability. J. Sci.Food Agr. 80, 404-408.
48. Philippeau C., Michalet-Doreau B. (1998). Influenceof genotype and ensiling of corn grain on in situ de-gradation of starch in the rumen. Journal of dairyscience 81. 2178-2184
49. Pietta, P. G. (2000). Flavonoids as antioxidants. J.Nat. Prod. 63: 1035-1042.
50. Prasad, P. V. V., Staggenborg, S. A., Ristic, Z. (2008).Impacts of drought and/or heat stress on physiologi-cal, developmental, growth, and yield processes ofcrop plants. In: Response of crops to limited water:Understanding and modeling water stress effects onplant growth processes Vol. 1 (Ahuja, L., Ed.), Ameri-can Society of Agronomy, Crop Science Society ofAmerica, Soil Science Society of America, Madison,301-355.
51. Saffa, H.M., Jiménez-Moreno, E., Valencia, D.G.,Frikha, M., Serrano, M.P., Mateos, G.G. (2009). Effectof main cereal of the diet and particle size of the ce-real on productive performance and egg quality ofbrown-laying hens in early phase of production. Poul-try Science. 88:608-614.
52. Sauvant, D., Perez, J.M. and G.Tran (2004). Tablesof Composition and Nutritional Value of Feed Mate-rials. Wageningen Academic Publisher & INRA Paris.
53. Sezer ì., Balkaya A., Karaağaç O., Öner F. (2011).Moisture dependent of some physical and morpholo-gical properties of dent corn (Zea mays var. indentәtәSturt) seeds. African Journal of Biotechnology Vol.10(15), pp. 2857-2866.
54. Sigh J., Dartois A., Kaur L. (2010): Starch Digestibilityin food matrix: a review. Treds in Food Science andTechnology. 21: 168-180.
55. Singh S., Singh G., Singh p., Singh N. (2008): Effectof water stress at different stages of grain develop-ment on the characteristics of starch and protein ofdifferent wheat varities. Science Direct. 108: 130-139.
56. Sundheim, L., Rafossm T. (2015). Fumonisin in mai-ze in relation to climate change. In. Climate Changeand Mycotoxins. (Eds. Botana,L.M., Saintz, M.J.).Walter de Gruyter GmbH, Berlin/Boston. 109-132.
57. Watson, S.A. 2003. Description, development, struc-ture, and composition of the corn kernel. In P. J. Whi-te and L. A. Johnson (Eds.) Corn: Chemistry andTechnology. Amer. Assoc. Cer. Chem., Inc. St. Paul,Minnesota, USA, pp. 68-106.
58. Weurding, R.E. (2002): Kinetics of starch digestionand performance of broiler chickens. Doktorska diser-tacija, Wageninen Institute of Animal Science, Wage-ninen University Wageninen, Nizozemska.
59. Weurding, R.E., Enting, H., Verstegen, M.W.A(2003):The effect of site of starch digestion on performancebroiler chickens. Aminal Feed Scients and Technolo-gy 110: 175-184.
60. White P. J., & Johnson L. A. (2003). Corn: chemistryand technology (No. Ed. 2). American Association ofCereal Chemists.
61. Wilfart A., Jaguelin-Peyraud Y., Simmins H., NobeltJ., Van Milgen J., Montagne L. (2008): Kinetics of en-zymatic digestion of feeds as estimated by a stepwisein vitro method. Animal Feed Science and Technolo-gy. 141: 171-183.
62. Zhao, Z., Egashira, Y., Sanada, H. (2005). Phenolicantioxidants richly contained in corn bran are slightlybioavailable in rats. J. Agric. Food Chem. 53: 5030-5035.
63. Zuleta, A., M.J. Esteve, A. Frígola (2009): ORAC andTEAC assays comparison to measure the antioxidantcapacity of food products. Food Chem. 114: 310-316.
64. ςakir, R. (2004). Effect of water stress at different de-velopment stages on vegetative and reproductivegrowth of corn. Field Crops Res. 89: 1–16.

Prof. dr. sc. Darko Grbeša redoviti je sveučilišni profesoru trajnom zvanju, predaje Hranidbu životinja na dodi-plomskom studiju, voditelj i nositelj je dva predmetana diplomskom studiju Hranidba i hrana za životinje naAgronomskom fakultetu Sveučilišta u Zagrebu. Objavio je 175 radova među kojima su neki u prestižnimmeđunarodnim znanstvenim časopisima kao što jeFood Chemistry. Prema Web of Science D. Grbeša je umeđunarodno priznatim časopisima najcitiraniji profesorhranidbe životinja iz RH. Prof. Grbeša autor je tri knjigei pet poglavlja u drugim knjigama. Pod njegovim men-torstvom obranjeno je pet doktorskih teza. Trostruki jestipendist Cochran Fllowship Programa američkog mi-nistarstva poljoprivrede.Savjetnik je tvornica krmnih smjesa i najvećih hrvatskihmliječnih farmi. Održao je preko 150 predavanja o hra-njivosti krmiva i hranidbi krava u Hrvatskoj i inozemstvu.
Bc Institut d.d., Rugvica, Dugoselska 7, 10370 Dugo Selo10000 Zagreb, Hrvatska
Prof. dr. sc. Darko Grbeša
Hranidbena svojstva kukuruza
ISBN 9789535507505






















