Embed Size (px)
Citation preview

1
TDP-43 の細胞質における神経細胞死誘導機構の解明
Cause of neuronal cell death by TDP-43 in the cytoplasm
北村 朗
北海道大学先端生命科学研究院・細胞機能科学分野・助教
日本学術振興会 国際共同研究加速研究者

2
1.要旨
筋萎縮性側索硬化症 (ALS)には様々な原因型が存在するが,本研究では TARDBP を責任遺
伝子 (翻訳産物は TDP-43)とする ALS10 の病因解明を目的として下記の三点に着目し研究
を行った.第一に, TDP-43の 25 kDaカルボキシル末端断片 (TDP25)が凝集することで細
胞死を引き起こすが,細胞質において配向を持たず形成されたオリゴマーが毒性の元とな
る可能性について検証した.その結果,秩序ある配向性を持つ TDP25 の凝集体は分子シャ
ペロンである HSP70 の結合性が弱く,配向性を持たない TDP25 の凝集体には HSP70 の結
合性が強いことから,配向性の違いが分子シャペロンによる認識度の違いを生むことが示
唆された.さらに,このような構造の差異が細胞毒性につながる可能性も考えられる.第二
に,TDP25 が凝集する機構について検証したところ,TDP25 の凝集には欠損 RRM2 (RNA
と結合する領域の一部)と C 末端のグリシンに富んだ領域(凝集性が伝播するプリオン様の
構造をとる)の両者が必要であることがわかった.第三に,TDP-43 を RNAi によりノック
ダウン(細胞内のタンパク質存在量を減らす操作)した細胞では,ある RNA X の発現量が
減少し,この発現量を外来性発現により校正すると細胞死が抑制できることを明らかにし
た.以上の結果から,TDP-43から生成した TDP25の凝集体が全長の TDP-43を巻き込み凝
集することで細胞質に封入体を形成する.この状態は細胞死を引き起こすが,この時,TDP-
43 の標的となる RNAの発現量減少を引き起こすことにより,細胞死の頻度を上げることも
考えられる.
記載ソフトウェア:MS-Word 2016

3
2.背景
2-1.ALS 研究の歴史
筋萎縮性側索硬化症 (ALS)は,神経変性疾患の一種であり,古典的な定義では,脳神経系
における運動皮質,脳幹および脊髄に存在する上位および下位運動ニューロンが影響され
る疾患である(Rowland & Shneider 2001).近年は,小脳や知覚系などの異常がほとんど無い
にもかかわらず,認知症を合併することがあり,多因子・多系統疾患としても認識される
(Rowland & Shneider 2001).
ALSには,遺伝子に変異が入り起こる常染色体優性遺伝の家族性 (Familial ALS; FALS)と,
孤発性 (Sporadic ALS; SALS)の2種類存在する.FALS では遺伝子に変異が入ることで ALS
を発症するリスクが上昇する.ALS における責任遺伝子として,20 種類以上のものが知ら
れており,現在も増えつつある.一方,SALS における発症機序はほぼ不明であるが,加齢
や持続的なストレスにより長い時間を経て上述の FALS 責任遺伝子を含めた様々な細胞内
外のタンパク質に異常が起こることで発症すると考えられている.
ALS 研究の歴史は,19世紀にフランスの神経医 Jean Martin Charcotが “la sclérose larérale
amyotrophique”(フランス語で「筋萎縮性側索硬化症」)と記述したことに始まる.この時点
で患者の痙攣と麻痺は確認されており,さらに生検の結果から,脊髄の前角において組織の
損傷があることが記されている (Charcot & Joffroy 1869).ただし残念ながら,19世紀の報告
以後,ALS の分子的病因に関する研究進展は残念ながら無いと言える状況であった.
神経病理所見の点から追ってみると,SALS では下位運動ニューロンの萎縮が見られ,か
つ下位運動ニューロンの細胞質にヘマトキシリン・エオジン染色される好酸性の小さな封
入体が観察されることがある.この封入体はブニナ小体と呼ばれ,SALS において特異的な
封入体である(Bunina 1962, Okamoto et al 2008).このブニナ小体の他に,脊髄前角神経にユ

4
ビキチン陽性封入体が存在することが発見された(Lowe et al 1988).ユビキチンとは細胞内
において選択的に分解されるタンパク質に対しタグとして標識されるタンパク質であるこ
とから,タンパク質分解系に異常が起こり,元来分解されるべきタンパク質が蓄積すること
で細胞が障害を起こしているのではないかと予想された.また,ユビキチン陽性封入体は糸
くず状に細胞質に存在していることが多く,このような構造はスケイン様封入体と呼ばれ
る.FALS においてはこのようなユビキチン陽性封入体が観察されやすい.しかしながら,
神経病理学のみでは詳細な ALS 病因の分子機構を明らかにすることは困難であった.
このような状況が一転したのは,1993年になってからである.この年,FALS 患者の家系
解析から,Cu/Zn superoxide dismutase 1 (SOD1)タンパク質をコードする遺伝子に一アミノ酸
置換を引き起こす変異が存在することが報告された(Rosen 1993).この ALS 関連遺伝子の発
見により,ALS 研究は,その時期既に成熟していた分子生物学の手法と相まって進歩した
と言える.SOD1 は生体に有害であるスーパーオキシドを酸素と過酸化水素に分解・無毒化
する酵素であることから,当初は SOD1 の活性が低下または不活化することにより細胞障
害が起こると考えられた.しかしその後の研究で,SOD1 を欠損させたマウスの解析では,
運動ニューロンの障害は起こらず,変異型 SOD1 を強制発現したマウスで運動麻痺が起こ
ることがわかった(Gurney et al 1994).さらに,SOD1 は FALS 患者の運動ニューロンにおい
てユビキチン陽性の封入体(凝集したタンパク質が細胞内で隔離されできた構造体)を形成
することが神経病理学的に示された.また,変異の入った SOD1 はミスフォールドして凝集
体を形成しやすいことが生化学的解析から明らかになった.これらのことから,SOD1が構
造異常により何らかの毒性を獲得し FALS の原因となっているのではないかと考えられた.
このように SOD1 を対象とした FALS 病因研究は進んでいたが,全ての ALS 病態を説明
できるものでは無かった.2006 年になり,SOD1 の変異を持たない ALS および前頭側頭葉

5
変性症 (FTLD-U)におけるユビキチン陽性封入体の構成成分として,Transactivation response
DNA/RNA-binding protein 43 kDa (TDP-43)が同定された(Arai et al 2006, Neumann et al 2006).
当初,この TDP-43は他の要因による二次的要因によって封入体を形成することから,ALS
の一次的要因ではないことも疑われたが,SOD1変異によって起こる FALS の運動ニューロ
ンにおいて形成されるユビキチン陽性封入体に TDP-43が含まれないことに加え (Tan et al
2007),TDP-43 に変異を有する ALS が見出されたことから(Sreedharan et al 2008),TDP-43は
ALS における一次的要因であると考えられている.
2-2.TDP-43の局在と機能
TDP-43は 414アミノ酸から構成される 43 kDaのタンパク質である(図1).遺伝子の名
前は TARDBP であるが,ALS の責任遺伝子として分類されているため,ALS10 とも呼ばれ

6
る.この ALS10 は遺伝子名のみならず,TDP-43 に責任のある ALS 病態の分類名としても
用いられている.全身の臓器に発現し,細胞内では主に核に局在する.一次構造上の特徴と
して,核局在化シグナル配列と核排出シグナル配列を有し,核と細胞質の間を行き来きでき
る.また,106-176番目,および 192-241番目の領域はそれぞれ RNA/DNAに結合するモチ
ーフであり,それぞれ RRM1 または RRM2 と呼ばれている.274-314 番目の領域はグリシ
ンに富んだ領域であり,さらにこの領域を含む C 末端側は特定のアミノ酸組成に偏ってい
る.またこの領域は特定の構造を取らないため天然変性領域とも呼ばれている.ただし,こ
のようなグリシンに富んだ領域は不均一核内リポタンパク質 (hnRNP)に共通したものであ
り,TDP-43 と hnRNP が相互作用する際,必要な領域 (hnRNP 認識領域)である. FALS お
よび SALS 患者から数多くの TDP-43変異が同定されているが,そのほとんどがこの hnRNP
認識領域に保存されている.FALS において複数の家系で報告されている変異は,315 番目
のアラニンがスレオニンに変異した A315T,また 337 番目のメチオニンがバリンに変異し
た M337Vなどが知られている.この M337V 変異を有する7名中6名が ALS を発症してい
るという報告もあり(Sreedharan et al 2008),FALS における TDP-43変異と病因の関係性は着
目されているものの例数が少ないことから,これら変異の意義については未だ不明な点が
多い.さらに,培養細胞を用いた我々の研究結果からも,ALS 関連変異を有する TDP-43の
野生型との顕著な違いを見いだせていない(未発表データ).
2-3.TDP-43のスプライシング機能と ALS 病態
TDP-43の機能は,その RNA 結合能を利用して mRNA のスプライシングや micro RNAの
プロセッシングを介在することである.実際,TDP-43は pre-mRNA のイントロン領域に頻
繁に存在する UG 塩基を認識し,mRNA のスプライシングに関与することが知られている

7
(Polymenidou et al 2011, Tollervey et al 2011).それら pre-mRNAの中でも,長鎖のものに対し
機能することが示唆されている(Polymenidou et al 2011).また,331番目のグルタミンがリジ
ンに変異した TDP-43Q331Kを発現するマウスにおいて特異的にスプライシング状態の異なる
RNAが見られることから,ALS 関連変異型 TDP-43による RNA スプライシング効率の変化
が実際の ALS への関与が着目されている(Arnold et al 2013).しかしながら,TDP-43の機能
異常による顕著な RNA スプライシングの変化が実際の ALS 患者の中で見いだされている
のは POLDIP3 遺伝子のみであり,RNA スプライシング状態の変化と ALS 病態を直接証明
するには至っていない(Shiga et al 2012).
2-4.TDP-43のカルボキシル末端断片と凝集性
前項で述べたように,RNA スプライシングの異常は,TDP-43をノックダウンさせるなど
その量を著しく減少させたとき,顕著に観察することができる.しかしながら,TDP-43 の
減少や機能損失 (Loss-of-function)が ALS の病因かどうかは明らかになっていない.一方,
TDP-43 が本来の機能は異なる何らかの毒性を獲得する (gain-of-function)ことで神経変性や
ALS の発症に至るという仮説が存在する.ALS10 患者の運動ニューロンでは TDP-43 のユ
ビキチン陽性封入体が観察されることに加え,不溶性画分には 25 kDaや 35 kDaのカルボキ
シル末端側断片が存在することが知られている.これらのカルボキシル末端断片が生成す
る原因として,プロテアーゼであるカスパーゼ 3 またはカルパインによる切断を受けて生
ずるという説(Yamashita et al 2012, Zhang et al 2009)に加え,35 kDaの断片に関しては,当該
箇所にあるメチオニンから選択的翻訳が起こるという説も存在する(Xiao et al 2015).培養細
胞でカスパーゼ 3 を活性化すると,TDP-43 の切断とほぼ同時にカルボキシル末端断片の細
胞質移行および封入体形成が観察される(Kitamura et al 2016).重要なことは,これらカルボ

8
キシル末端側断片は全長の TDP-43よりも凝集性が高く,細胞毒性を持ちうるという点にあ
る(Kitamura et al 2016, Zhang et al 2009).また,ALS 患者において TDP-43の 409および 410
番目のセリンが高度にリン酸化されていることが確認されているが,実際これらのカルボ
キシル末端側断片のセリン 409/410 もリン酸化されている(Kitamura et al 2016, Zhang et al
2009).リン酸化と細胞毒性の関係は不明であるが,TDP25 のような TDP-43 のカルボキシ
ル末端断片を発現させると,細胞質に封入体を形成し,細胞死を引き起こす.
2-5.ALS における神経細胞死の原因仮説
我々も実際,この TDP25 発現による細胞死は検証しており,さらに核局在化シグナル配
列を付加した TDP25 (NLS-TDP25)は細胞質封入体を形成せず,かつ細胞死を抑制できるこ
とを報告している(Kitamura et al 2016).以上のことから,TDP25 の細胞質局在が細胞死の原
因になっているという仮説を得た.しかしながら,どのような構造または会合状態の TDP25
が毒性を獲得するのか,そして NLS タグで標識した TDP25 はなぜ低い毒性を示すのか,そ
の分子基盤は明らかになっていない.
さらに,TDP-43 の機能損失による RNA 全体の変調と神経細胞死の関係も依然未解明で
ある.そこで,本研究では,第一に NLS-TDP25と TDP25 の性質比較を行い,第二に TDP25
が凝集する機構について解析した.さらに第三に,TDP-43 をノックダウンしたときの標的
分子を見つけることで,TDP-43 がどのように細胞死に関与するのかを明らかにすることに
した.
3.方法
3-1.細胞培養とトランスフェクション

9
培養細胞はマウス神経芽細胞腫 Neuro2A 細胞を用いた.Neuro2A 細胞は DMEM 培地
(Sigma-Aldrich)に,10% FBS,ペニシリン・ストレプトマイシンを添加した培地で維持した.
プラスミド DNAは,QIAGEN midi prep kitにて回収したものを,吸光分光光度計を用いて
定量した後,3.5 cmディッシュ一枚当たり 1.0 gを Lipofectamine 2000 (Thermo Fisher)を用
いて細胞へ導入した.
3-2.蛍光顕微鏡観察
蛍光顕微鏡法には,全て Carl Zeiss社製レーザー共焦点顕微鏡 (LSM 510 META)を用いた.
対物レンズとしては,C-Apochromat 40×/1.2NA W Korr UV-VIS-IR (Carl Zeiss)を用いた.超解
像顕微鏡としては Structured illumination microscopy (SIM)を使用した.SIM のシステムとし
ては,Nikon社製 N-SIM を使用した.対物レンズとしては,Apo TIRF 100×/1.49NA Oil (Nikon)
を使用した.
3-3.FRET
FRET 効率を計測するためのアクセプターフォトブリーチング法は,全て Carl Zeiss 社製
レーザー共焦点顕微鏡 (LSM 510 META)を用いて行った.対物レンズとしては,C-
Apochromat 40×/1.2NA W Korr UV-VIS-IR (Carl Zeiss)を用いた.ドナー蛍光タンパク質として
はシアン色の mSECFP を使用し,アクセプター蛍光タンパク質としては黄色の mVenus を
利用した.ドナーまたはアクセプター蛍光分子を励起するためには 458 nm または 514 nm
のレーザーをそれぞれ利用した.アクセプターのフォトブリーチングには,514 nm の光を
照射した.
FRET 効率 EFRETは下記の式により求めた.

10
𝐸FRET = 1 −𝐼DA
𝐼D
ここで,IDAはアクセプターをフォトブリーチする前のドナーの蛍光強度,IDはアクセプ
ターをフォトブリーチした後のドナーの蛍光強度である.
3-4.FCS
蛍光相関分光法は Carl Zeiss LSM 510 META + ConfoCor2を用いて行った.得られた自己
相関関数は,下記の式(三重項緩和および2成分並進拡散モデル式)に従ってカーブフィッ
ティング解析を行い,拡散時間 (i; i = 1 or )を求めた(Pack et al 2006, Rigler et al 1993).
𝐺(τ) = 1 + [1 +𝑇
1 − 𝑇exp (−
τ
τtriplet)]
1
𝑁[∑ 𝐹𝑖
2
𝑖
(1 +τ
τ𝑖)
−1
(1 +τ
𝑠2τ𝑖)
−12
]
ここで,は変数としての時間,Tは三重項の割合,tripletは三重項緩和時間,Nは検出領
域内の平均分子数,Fiは成分 iの割合である.
上式から得られた拡散時間とは,蛍光分子のゆらぎを計測する微小領域(共焦点体積
と呼ばれる)を蛍光分子が通過する平均時間であるため,装置固有の値である.そのた
め,より一般的な物理量である拡散係数を算出するために下記のように計算した.拡散
係数が既知の基準物質として Rhodamine 6G (拡散係数 DRh6G = 414 m2·s-1)を同条件で測
定し,下記の関係式によって目的試料の拡散係数 (Dsample)を得た.
𝐷sample = (τRh6G
τsample) ∙ 𝐷Rh6G
ここで,Rh6Gは,目的試料と同じ光学条件で測定された Rhodamine 6G水溶液の拡散時
間である.
また,拡散係数から試料の分子量 (MSample)を計算するために,Stokes-Einstein 関係式

11
(Kitamura et al 2014, Pack et al 2006)から下記の改変式を得,さらに単量体 GFPの測定さ
れた拡散係数とそのアミノ酸組成から計算された理論分子量 (MGFP = 27 kDa)を用いて
これに従って計算した.
𝑀Sample = (𝐷Sample
𝐷GFP)
3
∙ 𝑀GFP
3-5.免疫沈降
Neuro2A 細胞の細胞抽出液を得るために,培地を除去した後,室温の PBS を用いて洗浄
した.次に,細胞破砕緩衝液(50 mM Hepes/KOH (pH 7.5), 150 mM NaCl, 1% TritonX-100, プ
ロテアーゼ阻害剤)中で細胞を破砕し,遠心した後,溶解液を得た.この溶解液に抗 GFP 抗
体結合型アガロースビーズ (MBL)を加え,4℃,一時間混和した.プロテアーゼ阻害剤を
含まない破砕液中でビーズを三回洗浄した後,ビーズに結合したタンパク質を回収するた
めに,DTT を含まない Laemmli サンプル緩衝液を加え,室温で5分静置した.液体成分を
回収した後,終濃度 100 mM となるよう DTT を加え,98℃で2分処理し,SDS-PAGE とそ
れに続くウェスタンブロットを行った.ウェスタンブロットに用いた一次抗体は,マウスモ
ノクローナル抗 GFP 抗体 (GF200; ナカライ)とウサギモノクローナル抗 HSPA8 抗体
(Abcam)である.二次抗体として,西洋ワサビペルオキシダーゼ結合型抗マウスまたはウサ
ギ IgG 抗体 (Jackson Laboratory)を用いた.検出は,ECL detection kit (GE)を用いて発色させ,
LAS4000 mini (富士フイルム)を用いて画像取得した.
3-6.RT-PCR
Neuro2A 細胞からの RNA 抽出には,TRizol (Thermo Fisher)を用いた.抽出された RNA の

12
精製には,Pure Link RNA purification kit (Ambion)を用いた.吸光光度計で RNA を定量した
後,Micro RNA first strand synthesis kit (Clontech)を用いて低分子 RNA を鋳型とし逆転写を行
った.定量的リアルタイム PCRのために,検出蛍光試薬として SYBR Green Iを用いる PCR
混合溶液 (SYBR Premix Ex-Taq II; TaKaRa)を利用した.PCR 装置は,北海道大学オープンフ
ァシリティ共同利用機器である Mx3005P (Agilent)を用いた.
4.結果
4-1.TDP-43のカルボキシル末端断片の凝集性
4-1-1.核局在化シグナル配列を付加した TDP-43の 25 kDa カルボキシル
末端断片 (TDP25)の凝集特性
TDP-43のカルボキシル末端断片の一つである TDP25 は,全長の TDP-43 よりも凝集性が高
い.我々はこの TDP25 に核局在化シグナル配列 (NLS)(Kitamura et al 2015)を付加し,核へ
局在させたところ,細胞質封入体形成が無くなり,細胞死を抑制できることを示した
(Kitamura et al 2016).この結果を踏まえ,NLS-TDP25 が凝集形成しにくい原因について詳細
に解析することにした.
まず,緑色蛍光タンパク質 GFP を付加した NLS-TDP25 (GFP-NLS-TDP25)を Neuro2A 細
胞に発現させたところ,核へ高効率に局在し細胞質への局在率は極めて低かった.この時,
この GFP-NLS-TDP25は,核質のみならず核小体にも局在した.この核小体への局在は,タ
グである GFP-NLS も局在したことから,TDP25 ではなくタグの影響であると考えられる.
この核小体への局在を詳細に観察するために,従来型の蛍光顕微鏡よりも分解能の高い超
解像蛍光顕微鏡法の中で,構造化照明法 (Structured illumination microscopy)を用いた(Hirano
et al 2015).GFP-NLS-TDP25 は核小体全般に局在化するものの,核小体と核質との境界付近

13
に集積していることがわかった.一方,タグのみの GFP-NLS では核小体に均一に局在して
いた.次に,細胞内の凝集性タンパク質の品質管理に直接関わるタンパク質分解の複合体酵
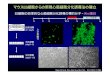
素であるプロテアソームの分解阻害剤で処理すると,GLS-NLS-TDP25は核小体と核質との
境界領域により顕著に局在した(図2).次に,GFP-NLS-TDP25の核小体集積が封入体であ
るかどうかを確かめるために,核小体と核質の間の交換反応を fluorescence recovery after
photobleaching (FRAP)法を用いて生細胞内で調べた.細胞質に形成した封入体において GFP-
TDP25 は 11%程度しか動いていなかったのに対し,GFP-NLS-TDP25 は核小体において約
55%の分子種が交換していた.このことは,GFP-NLS-TDP25 が核小体で封入体を形成して
いるのではなく,何らかの因子と結合し集積していることを示唆する.さらに,プロテアソ
ーム阻害剤を処理すると,核小体における GFP-NLS-TDP25はほぼ不動となった.これらの
ことは,プロテアソームにより分解されなくなった GFP-NLS-TDP25が蓄積するための構造
が,核小体と核質の境界領域にあるのかもしれない.最近の報告から,核小体近傍に凝集性

14
タンパク質を隔離し,タンパク質の品質管理を行うための区画が真核生物である酵母にお
いて存在することがわかっている(Miller et al 2015).そのため,ここで形成された核小体と
核質境界の構造は,このような品質管理の区画形成と関わりがあることも考えられる.
4-1-2.NLS-TDP25 のオリゴマー形成
次に,NLS-TDP25と TDP25
がどのように異なる構造を
持つのかを検証するために,
細胞質の封入体または核小
体集積における分子間の配
向を調べることにした.この
分子間配向を調べるために,
蛍光分子間の距離と配向に依存して起こる蛍光共鳴エネルギー移動 (Fluorescence/Förster
resonance energy transfer; FRET) 法を用いることにした(図3左).FRET 法の原理は以下,
シアン色と黄色の二色の蛍光分子を利用するとして説明する.これら蛍光分子の間に FRET
が起こっていなければ,例えばシアン色の蛍光分子のみを励起したとき(FRET ドナーを励
起する),黄色の蛍光分子(FRET アクセプター)は発光しない.ところが,FRET が起こる
条件において,シアン色を励起したエネルギーの一部が黄色の蛍光分子に共鳴移動すると,
シアン色の蛍光強度は低くなり,黄色の蛍光強度はエネルギー移動の効率に依存して高く
なる.これが FRET である.次に,細胞内における FRET 効率を調べるための方法として,
アクセプターフォトブリーチング法を用いた.アクセプターフォトブリーチング法の原理
についても,上述のようにシアン色と黄色の蛍光分子を例えとして説明する.シアン色を励

15
起するためのエネルギーの
一部が黄色の蛍光分子に共
鳴移動している時,黄色の
蛍光分子のみが吸収する強
力な光を吸収させること
で,蛍光を発する元の構造
である発色団を破壊する.これをアクセプターフォトブリーチという.発色団が破壊される
と,黄色の蛍光分子はもはや蛍光を発することができなくなると同時に,シアン色からエネ
ルギーが共鳴移動することもできなくなるため,FRET が解消すると共に,シアン色の蛍光
強度が回復する.アクセプターフォトブリーチング法とは,このシアン色分子(FRETドナ
ー)の蛍光強度回復を,黄色の蛍光分子(FRETアクセプター)の褪色前後で比較すること
により,共鳴移動したエネルギーの効率,すなわち FRET 効率を求めるものである(図3
右).
まず,シアン色または黄色の蛍光タンパク質として,それぞれ TFPまたは YFP を選び,
これらを TDP25または NLS-TDP25 の N 末端側に融合し Neuro2A に発現させた.まず,細
胞質の封入体を形成していない区画または核質における TDP25 の分子間で FRET は起こっ
ていなかった.次に,核質における NLS-TDP25の FRET 効率を比較したところ,8.5%とネ
ガティブコントロールに対し有意な FRET 効率を示した(図4).このことは,NLS-TDP25
が少なくとも二量体を形成していることを示唆する.
このように,NLS-TDP25 の分子間では FRET が起きているが,TDP25 では FRET が起き
ていないことがわかった.ただし,FRET効率は蛍光分子間の距離のみならず配向にも依存
するため,FRET効率が計測されないからといって分子間相互作用が無いとは言えない.そ

16
こで次に,分子のオリゴマー化状態を解析できる蛍光相関分光法 (Fluorescence correlation
spectroscopy; FCS) を用いて生細胞内における GFP-TDP25またはGFP-NLS-TDP25の拡散係
数を計測した.分子量が増加すると分子は溶媒などから抵抗を受けやすくなるため,運動性
が下がる.そのため,理論式を用いることで,拡散係数から分子量の計算が可能となる.
GFP-TDP25および GFP-NLS-TDP25 の,核質,細胞質それぞれにおける拡散状態に,変
化は無かった.さらに,速く拡散する分子種と遅く拡散する分子種の二つの成分が存在する
と仮定して解析を行ったところ,速く拡散する分子種の拡散係数は GFP の半分の大きさで
あり,これは,分子量として GFP の 8倍,つまり約 240 kDaであった.この分子量は,GFP-
TDP25 または GFP-NLS-TDP25 単量体の計算分子量 (約 50 kDa)よりも大きく,オリゴマー
形成または内在性分子と相互作用していることが考えられた.
4-1-3.NLS-TDP25 と分子シャペロン HSP70 との相互作用
本助成を受ける以前の解析から,GFP-TDP25 と結合するタンパク質として,細胞質・核
に存在する分子シャペロン HSP70 が同定されていた.分子シャペロンとは,タンパク質の
フォールディングを介助する分子であり,凝集形成の抑制や脱凝集を行う(Hartl et al 2011).
HSP70 は新生ポリペプチド鎖やミスフォールドしたタンパク質にすみやかに結合するシャ
ペロンであり,HSP40 や HSP110 などと共役して脱凝集能を持つことが知られている
(Kampinga & Craig 2010).
TDP25が HSP70 と結合するという結果は,FCS 解析において,内在性分子と相互作用し
て拡散する分子の存在が予想されたこととも一致する.この HSP70 の結合性が,GFP-TDP25
と GFP-NLS-TDP25 で変化するかを,免疫沈降法とウェスタンブロット法により検証した.
抗 GFP 抗体を共有結合したアガロースビーズを用いて,GFP-TDP25 または GFP-NLS-

17
TDP25を発現させた Neuro2A細胞抽出液を処理し得た,沈降液を SDS-PAGE で展開し,抗
HSP70 抗体を用いたウェスタンブロットにより,HSP70 の結合量を比較した.この結果,
TDP25, NLS-TDP25の HSP70 結合量に変化は見られなかった(図5).
次に,HSP70 が細胞内の TDP25 凝集体を認識しているのかを確かめるため,GFP-TDP25
と HSP70-RFP (RFP は赤色蛍光タンパク質)を共発現させた Neuro2A 細胞を共焦点蛍光顕微
鏡で観察した.細胞質に形成された GFP-TDP25の封入体に,HSP70-RFP は多く共局在して
いた.一方,GFP-NLS-TDP25 が集積した核小体における HSP70-RFP の共局在性を同様に調
べたところ,TDP25 の細胞質封入体ほど多くの HSP70 では無かったが,弱く共局在が見ら
れた.従って,核小体に集積した NLS-TDP25は,細胞質の TDP25 封入体とは異なる構造を
取っており,細胞質封入体はより重
度の変性した構造を取る TDP25が集

18
積していることが示唆された(図6).
4-2.TDP25 の凝集性を規定する領域の同定
4-2-1.TDP25 欠損変異体の凝集形成
細胞質封入体は凝集したタンパク質が集積してできる構造であると考えられる.従って,
以後,細胞質封入体の形成率を測ることで,解析対象のタンパク質の凝集性を調べることに
した.
TDP25は凝集性が高い TDP-43のカルボキシル末端断片であるが,この凝集体がどのよう
にして形成されるのか明らかになっていない.TDP25は,N 末を一部欠損した RNA/DNA結
合 RRM2ドメイン (220-262番目)と,その C 末にグリシンに富んだ領域 (263-414番目)を含
む.一方,グリシンに富んだ領域は天然変性領域であるものの,アミロイド様の凝集体に転
移し,その構造が分子間で伝播するプリオン様ドメインとしても知られている.我々の先行
研究から,この領域に GFP を融合し,Neuro2A に発現させたところ,細胞質封入体は形成
しなかった.また,同タンパク質を発現する Neuro2A細胞抽出液を得て,0.1% SDS 可溶性
画分と 0.1% SDS 不溶性・1 M尿素可溶性画分に含まれる量比を求めたところ,ほぼすべて
0.1% SDS 可溶性として存在することがわかった(Kitamura et al 2016).すなわち,263-414番
の領域は凝集体に転移する性質は有しているものの,通常条件では凝集体を形成しにくい
と考えられる.しかしながら一方で,220-414番目の領域断片である TDP25は高い凝集性を
持つ.これらのことから,220-262番目のどこかに凝集への構造変化を規定する配列が存在
するのではないかと考えられた.
次に,凝集体形成の原因となる疎水性相互作用を形成するアミノ酸について調べるため,
TDP25 配列の疎水度プロットおよび二次構造予測を行った.この予測の結果,RRM2 には

19
TDP-43全長の中で相対的に疎水性の高い領域が存在していた. これらのことから,N 末側
が欠損した RRM2 では,疎水性の配列が露出することで凝集形成の契機となるが,この疎
水性配列が N 末に来なければ,凝集形成しにくいのではないかと考えられる.
4-2-2.TDP25 凝集形成における欠損 RRM2 領域の役割
TDP25 は RRM2 の N 末 18 残基が欠損している.そのため,この欠損により,残された
RRM2 の配列が正しくフォールディングできず,この構造不安定性が凝集の契機になるの
ではないかと考えた.そこで,残された RRM2 である 220-262 番目のペプチド領域 (TDP-
43220-262)が細胞質封入体を形成するか調べた.GFP 融合型 TDP-43 (GFP-TDP-43220-262)を
Neuro2A細胞に発現したところ,11%の細胞で細胞質封入体を形成した.また同条件におけ
る GFP-TDP25の細胞質封入体形成率は 23%であった.このことから,残された RRM2領域
(220-262 番目) はそれ自身,構造が不安定で正しくフォールディングされた構造を取れない
ことが示唆された.さらに,C 末端のグリシンに富んだ領域がこの 220-262 番目の領域と融
合すると,封入体形成率が増加することから,220-262番目の領域とグリシンに富んだ領域
が両者存在することで,高い細胞質封入体形成性を示すと考えられる.
次に,220-262番目の領域とグリシンに富んだ領域がどのようにして封入体形成に至るの
かを考えるために,構成するアミノ酸の疎水性に着目した.グリシンに富んだ領域の 310-
330 領域付近には,疎水性が高いアミノ酸が保存された領域が存在する.前項 4-2-1 で述べ
た TDP43の 220-262 領域にも疎水性の高いアミノ酸が保存されていることから,220-263領
域に存在する,元来 RRM2 の折り畳み構造を安定化するための疎水性残基が,一部配列の
欠損により露出するようになり,310-330領域の疎水性アミノ酸残基と疎水性相互作用を引
き起こして,凝集に至るのではないかと考えられる.

20
一方,グリシンに富んだ領域である 263-414 番目の領域は天然変性領域であると共に,プ
リオン様領域とも呼ばれ,構造変化を伴いアミロイド様の凝集体を形成する領域である.こ
れらのことから,グリシンに富んだ領域がそれだけで凝集体を形成するものではなく,
TDP25 の 220-262 領域に含まれる疎水性アミノ酸が露出し,グリシンに富んだ領域の疎水
性アミノ酸と相互作用することで,凝集形成に至る可能性が考えられる.この凝集過程をさ
らに詳細に検証するために,疎水性アミノ酸を親水性アミノ酸に置換することで,細胞質封
入体の形成率から凝集形成過程を考察していく必要があると考えられる.
4-3.TDP-43の RNAi ノックダウンによる RNAに与える影響
4-3-1.TDP-43 の RNAi ノックダウンにより RNAの量が減少する
前項までは,TDP-43 のカルボキシル末端断片の凝集体または凝集性のオリゴマーが細胞
死を引き起こすという毒性獲得モデルを念頭に解析を行ってきたが,ALS では TDP-43の機
能損失が原因となり得るという仮説も存在する.そこで,siRNA による RNAi 法を用いて,
Neuro2A細胞内在性の TDP-43をノックダウンし,機能解析を行うことにした.
TDP-43は mRNA のスプライシングや,microRNA のプロセッシングに関与する(Buratti &
Baralle 2010, Kawahara & Mieda-Sato 2012).TDP-43が関与する経路と関与しない経路があり,
その詳細な役割は未知である.このような経緯を踏まえ,我々は TDP-43 依存的に量変化す
る micro RNA を探索するために,まず,TDP-43 をノックダウンした細胞から得た RNA に
対し,低分子 RNA を対象とした逆転写反応を行った後,定量的 PCR (qPCR)を用いて増幅
量を比較した.この結果,TDP-43 をノックダウンした Neuro2A 細胞ではある RNA X の量
が減少することが再現性を持って得られた.この時,ハウスキーピング遺伝子由来の転写産
物 18S rRNA,7SL RNA, snoRNA 202 に着目し,これらの PCR 増幅量を比較したところ,ノ

21
ックダウン対象を持たない siRNA を導入した Neuro2A細胞由来の試料(ネガティブコント
ロール)由来の増幅量に対して,18S rRNAと 7SL RNAは変化しなかった.これらの結果に
より,18S rRNA と 7SL RNA は,TDP-43 のノックダウンによって量が変化しない RNA で
あり,内部標準に適切であることがわかった.さらに 7SL RNAを内部標準として RNA Xの
増幅量を調べたところ,TDP-43ノックダウン細胞で有意に減少することがわかった.
4-3-2.RNA Xの発現は TDP-43ノックダウンによる起こる細胞死を抑制す
る
TDP-43 をノックダウンする
と,細胞死が起こることが報告
されている(Iguchi et al 2009).実
際,我々が TDP-43 を siRNA に
よりノックダウンしたときにも,
siRNA 導入後 120 時間で細胞死
の割合がコントロール siRNA導
入細胞に対して増加した.そこ
で次に,RNA X の発現量とこの細胞死が関与するのか調べるために,TDP-43をノックダウ
ンした細胞に対し,RNA X を一過的に発現させることで,細胞死を回避できるかを調べた.
TDP-43をノックダウンした細胞で RNA X を一過的に発現させたところ,120時間経過した
時点での細胞死の割合は,空ベクターを導入した細胞に比べ減少した.コントロール siRNA
を導入した細胞に RNA X発現ベクターまたは空ベクターを導入しても,細胞死の割合に変
化は無かった(図7).このことから,RNA X は TDP-43の機能阻害により引き起こされる

22
細胞死経路の重要な標的分子であることが初めて明らかになった.
5.考察
TDP25 は凝集性が高く細胞質封入体を形成しやすい性質を持つ.TDP25 が核及び細胞質
に存在すると細胞死が起こるが,核局在化シグナル配列を付加して TDP25 を核のみに局在
させると,細胞死が低減できることを我々のグループは報告済みである(Kitamura et al 2016).
また,我々の先行研究から NLS-TDP25は核小体において秩序正しく分子が配向した集積構
造を取るのに対し,細胞質封入体における TDP25 はそのような秩序正しい分子配向は取ら
ないことが FRET の解析から示唆された (Kitamura et al 2017).このような集積に至る過程
で,核質または細胞質で拡散している状態の TDP25 または NLS-TDP25 は,細胞内の内在性
タンパク質と結合したオリゴマー状態になっていると考えられる.
TDP25 に結合する内在性タンパク質の一つとして,分子シャペロン HSP70 を我々は同定
しており,この結果は他のグループから出された報告とも一致する(Zhang et al 2010).HSP70
が TDP25 に結合する生理的理由であるが,HSP70がタンパク質の凝集形成を抑制または脱
凝集を行うものであるため,TDP25 に対してもそのように働いて結合していると考えられ
る.ただし,単に凝集を抑制しているのか,あるいは積極的に脱凝集を行っているのかにつ
いては明かではない.我々が HSP70を TDP25の結合相手として同定した免疫沈降-マスス
ペクトロスコピーの結果では,脱凝集活性を HSP70 と共に発揮するために必要な HSP40な
らびに HSP110 の共沈降は同定されなかった(未発表データ).ただし,HSP40 の基質結合
力は HSP70と比べて相対的に弱いため,免疫沈降途中において TDP25から解離しているの
かもしれない.この点を明らかにするためには,in vitro で生化学的な脱凝集反応を検出す
る系を用いるか,加えて HSP40 や HSP110 の RNAi ノックダウンを行うことで,TDP25 の

23
脱凝集に対する効果を調べていくことが必要であると考えられる.
さらに,免疫共沈降される HSP70 の量は,TDP25 と NLS-TDP25 で差は見られなかった
ものの,TDP25の細胞質封入体に HSP70は効率的に共局在するのに対し,NLS-TDP25が局
在する核小体にはあまり効率的に局在しないことから,HSP70は TDP25 の構造を見極め結
合していることが示唆される.従って,FRET による分子間配向の結果とも合わせると,NLS-
TDP25は TDP25とは異なるオリゴマー構造を形成しており,このオリゴマー構造の違いが
細胞死を引き起こす毒性の違いに関係しているのでは無いかと考えられる.
我々は,TDP25 が凝集を形成する機構として,欠損 RRM2 領域内にある疎水性アミノ酸
が露出し,C末のグリシンにある領域に存在する疎水性領域と疎水性相互作用を介して起こ
るのではないかと考えている.これをさらに詳細に明らかにするためには,疎水性領域に変
異を入れるなどの実験が必要であるが,疎水性領域は複数のアミノ酸から構成されている
ことから,単純な数個のアミノ酸置換では解析できない.そのため,実際に変異を導入しな
い TDP25 を元にして構造変化が起こることを直接解析することが重要と考えている.その
ための方法として,低分子の蛍光分子をラベルした TDP25の一分子構造変化について FRET
を用いて解析することに加え,NMRの利用も検討中である.ただし過去の我々の検討から,
TDP25 の大腸菌リコンビナント体は極めて発現量が低くく,その凝集性もあることから回
収が難しかった(未発表データ).この領域を詳細に検討することで,大腸菌リコンビナン
ト体発現を検討することで,回収量を増やせる可能性もある.今後は,TDP25が凝集する過
程の構造変化を解析していきたいと考えている.
ALS 病態の原因として,TDP-43 の機能損失によるものと,TDP25 のような凝集体が毒性
を獲得することに寄るものか未だ明らかになっていない.この点を解明することは,ALSの
病因解明において極めて重要な課題であり,この点無くして根本的治療法は確立できない

24
と考えられる.我々はTDP-43の機能損失と,凝集性獲得という両者の観点から解析を行い,
TDP-43の機能損失があるとき,RNA X が減少し,この発現量を維持することで細胞死を抑
制できることを発見した.ただし,RNA X には TDP-43が結合する普遍的配列が見られない
ことから,TDP-43 は RNA X の安定性または分解に寄与する他の因子の発現バランスを乱
す可能性もある.今後は,TDP-43がどのような機構で RNA Xの発現量維持に働いているの
か,その点について明らかにする必要があると考えられる.
また,我々の先行研究から,TDP25 が凝集すると全長の TDP-43を巻き込み封入体を形成
することが示唆されている(Kitamura et al 2016).このようにして封入体に巻き込まれた TDP-
43 は機能損失の状態にあると考えられる.細胞に存在する TDP-43 の多数が封入体に巻き
込まれると,TDP-43 そのものをノックダウンしたときと同様の状態が起こり,RNA Xの現
象と共に細胞死が引き起こされるのではないかと考えられる.またこの時,TDP25 の凝集
体がもつ細胞毒性が神経細胞死を増強させてしまうことも考えられる(図8).今後はこれ
らのつながりについて詳細に明らかにしていかねばならないと考えられる.
6.謝辞
このような研究の機会を頂いた貴協会に感謝いたします.貴協会からの研究助成金は,上

25
記のような研究結果を得ることにつながりました.これらの結果の中で,項目 4-1で説明し
た内容の一部の結果に加え,過去の研究結果を用いて構成した論文は,日本分子生物学会の
国際英文専門誌である Genes to Cells誌に 2017 年 3月 27日付けで掲載受理済(助成金受領
者が筆頭著者.全著者四名)となっております.また,項目 4-2で説明した内容は,私のア
イデア,指導を元に,北海道大学理学部生物科学科・高分子機能学専修 四年生の森谷香南
さんが主たる実験を遂行してくれました.当該内容は,平成 28年度北海道大学細胞生物研
究集会にて口頭発表を行いました.今後も引き続き解析を続ける予定です.項目 4-3で説明
した内容は,北海道大学大学院生命科学院生命融合科学コース博士課程三年の矢原真郎さ
んが私のアイデアを元に,自身のアイデアも加えて遂行した内容の一部です.当該成果は現
在投稿準備中であり,すでに投稿に向け論文が完成に近い状態です(助成金受領者が第二著
者.全著者三名).ここに謝意を表します.
7.引用文献
Arai T, Hasegawa M, Akiyama H, Ikeda K, Nonaka T, et al. 2006. TDP-43 is a component of ubiquitin-
positive tau-negative inclusions in frontotemporal lobar degeneration and amyotrophic lateral
sclerosis. Biochemical and biophysical research communications 351: 602-11
Arnold ES, Ling SC, Huelga SC, Lagier-Tourenne C, Polymenidou M, et al. 2013. ALS-linked TDP-
43 mutations produce aberrant RNA splicing and adult-onset motor neuron disease without
aggregation or loss of nuclear TDP-43. Proceedings of the National Academy of Sciences of
the United States of America 110: E736-45
Bunina TL. 1962. [On intracellular inclusions in familial amyotrophic lateral sclerosis]. Zh Nevropatol
Psikhiatr Im S S Korsakova 62: 1293-9

26
Buratti E, Baralle FE. 2010. The multiple roles of TDP-43 in pre-mRNA processing and gene
expression regulation. RNA Biol 7: 420-9
Charcot JM, Joffroy A. 1869. Deux cas d’atrophie musculaire progressive avec lesion de la substance
grise et des faisceaux antéro-latéraux de la moelle épinière. Arch. Physiol. Neurol. Path. 2:
744-754.
Gurney ME, Pu H, Chiu AY, Dal Canto MC, Polchow CY, et al. 1994. Motor neuron degeneration in
mice that express a human Cu,Zn superoxide dismutase mutation. Science 264: 1772-5
Hartl FU, Bracher A, Hayer-Hartl M. 2011. Molecular chaperones in protein folding and proteostasis.
Nature 475: 324-32
Hirano Y, Matsuda A, Hiraoka Y. 2015. Recent advancements in structured-illumination microscopy
toward live-cell imaging. Microscopy 64: 237-49
Iguchi Y, Katsuno M, Niwa J, Yamada S, Sone J, et al. 2009. TDP-43 depletion induces neuronal cell
damage through dysregulation of Rho family GTPases. The Journal of biological chemistry
284: 22059-66
Kampinga HH, Craig EA. 2010. The HSP70 chaperone machinery: J proteins as drivers of functional
specificity. Nature reviews. Molecular cell biology 11: 579-92
Kawahara Y, Mieda-Sato A. 2012. TDP-43 promotes microRNA biogenesis as a component of the
Drosha and Dicer complexes. Proceedings of the National Academy of Sciences of the United
States of America 109: 3347-52
Kitamura A, Inada N, Kubota H, Matsumoto G, Kinjo M, et al. 2014. Dysregulation of the proteasome
increases the toxicity of ALS-linked mutant SOD1. Genes to cells : devoted to molecular &
cellular mechanisms 19: 209-24

27
Kitamura A, Nakayama Y, Kinjo M. 2015. Efficient and dynamic nuclear localization of green
fluorescent protein via RNA binding. Biochemical and biophysical research communications
463: 401-6
Kitamura A, Nakayama Y, Shibasaki A, Taki A, Yuno S, et al. 2016. Interaction of RNA with a C-
terminal fragment of the amyotrophic lateral sclerosis-associated TDP43 reduces cytotoxicity.
Scientific reports 6: 19230
Kitamura A, Yuno S, Muto H, Kinjo M. accepted in 2017. Different aggregation states of a nuclear
localization signal-tagged 25 kDa C-terminal fragment of TAR RNA/DNA-binding protein
43 kDa. Genes to Cells.
Lowe J, Lennox G, Jefferson D, Morrell K, McQuire D, et al. 1988. A filamentous inclusion body
within anterior horn neurones in motor neurone disease defined by immunocytochemical
localisation of ubiquitin. Neuroscience letters 94: 203-10
Miller SB, Ho CT, Winkler J, Khokhrina M, Neuner A, et al. 2015. Compartment-specific aggregases
direct distinct nuclear and cytoplasmic aggregate deposition. The EMBO journal 34: 778-97
Neumann M, Sampathu DM, Kwong LK, Truax AC, Micsenyi MC, et al. 2006. Ubiquitinated TDP-
43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science 314: 130-
3
Okamoto K, Mizuno Y, Fujita Y. 2008. Bunina bodies in amyotrophic lateral sclerosis.
Neuropathology : official journal of the Japanese Society of Neuropathology 28: 109-15
Pack C, Saito K, Tamura M, Kinjo M. 2006. Microenvironment and effect of energy depletion in the
nucleus analyzed by mobility of multiple oligomeric EGFPs. Biophysical journal 91: 3921-
36

28
Polymenidou M, Lagier-Tourenne C, Hutt KR, Huelga SC, Moran J, et al. 2011. Long pre-mRNA
depletion and RNA missplicing contribute to neuronal vulnerability from loss of TDP-43.
Nature neuroscience 14: 459-68
Rigler R, Mets U, Widengren J, Kask P. 1993. Fluorescence Correlation Spectroscopy with High Count
Rate and Low-Background - Analysis of Translational Diffusion. Eur Biophys J Biophy 22:
169-75
Rosen DR. 1993. Mutations in Cu/Zn superoxide dismutase gene are associated with familial
amyotrophic lateral sclerosis. Nature 364: 362
Rowland LP, Shneider NA. 2001. Amyotrophic lateral sclerosis. The New England journal of medicine
344: 1688-700
Shiga A, Ishihara T, Miyashita A, Kuwabara M, Kato T, et al. 2012. Alteration of POLDIP3 splicing
associated with loss of function of TDP-43 in tissues affected with ALS. PloS one 7: e43120
Sreedharan J, Blair IP, Tripathi VB, Hu X, Vance C, et al. 2008. TDP-43 mutations in familial and
sporadic amyotrophic lateral sclerosis. Science 319: 1668-72
Tan CF, Eguchi H, Tagawa A, Onodera O, Iwasaki T, et al. 2007. TDP-43 immunoreactivity in
neuronal inclusions in familial amyotrophic lateral sclerosis with or without SOD1 gene
mutation. Acta neuropathologica 113: 535-42
Tollervey JR, Curk T, Rogelj B, Briese M, Cereda M, et al. 2011. Characterizing the RNA targets and
position-dependent splicing regulation by TDP-43. Nature neuroscience 14: 452-8
Xiao S, Sanelli T, Chiang H, Sun Y, Chakrabartty A, et al. 2015. Low molecular weight species of
TDP-43 generated by abnormal splicing form inclusions in amyotrophic lateral sclerosis and
result in motor neuron death. Acta neuropathologica 130: 49-61

29
Yamashita T, Hideyama T, Hachiga K, Teramoto S, Takano J, et al. 2012. A role for calpain-dependent
cleavage of TDP-43 in amyotrophic lateral sclerosis pathology. Nature communications 3:
1307
Zhang YJ, Gendron TF, Xu YF, Ko LW, Yen SH, Petrucelli L. 2010. Phosphorylation regulates
proteasomal-mediated degradation and solubility of TAR DNA binding protein-43 C-terminal
fragments. Molecular neurodegeneration 5: 33
Zhang YJ, Xu YF, Cook C, Gendron TF, Roettges P, et al. 2009. Aberrant cleavage of TDP-43 enhances
aggregation and cellular toxicity. Proceedings of the National Academy of Sciences of the
United States of America 106: 7607-12