Embed Size (px)
DESCRIPTION
Textbook Mikrobiologi13
Citation preview

BAB 13. FERMENTASI
Organisme membentuk energi dari nutrien melalui 2 cara, yaitu transfer elektron
di membran sel dan transfer elektron di sitoplasma. Sumber elektron pada transfer
elektron di sitoplasma adalah senyawa organik (heterotrof). Sumber elektron pada
transfer elektron di membran sel ada 4, yaitu pigmen penangkap cahaya (fototrof),
senyawa anorganik (autolitotrof), senyawa C1 (kemoautotrof), dan senyawa organik
(heterotrof)
Fotosintesis adalah pemanfaatan cahaya untuk pembentukan energi melalui
transfer elektron di membran. Respirasi adalah pemanfaatan senyawa kimia (baik
organik maupun anorganik) untuk pembentukan energi melalui transfer elektron di
membran. Fermentasi adalah pemanfaatan senyawa organik untuk pembentukan energi
melalui transfer elektron di sitoplasma. Pembentukan energi melalui transfer elektron di
membran disebut juga fosforilasi oksidatif. Dengan demikian respirasi didefinisikan
pembentukan energi melalui fosforilasi oksidatif.
Pembentukan energi melalui transfer elektron di sitoplasma disebut fosforilasi
tingkat substrat. Pembentukan energi dengan cara demikian ditemukan pada organisme
fermentatif. Oleh karena itu, fermentasi didefinisikan sebagai pembentukan energi
melalui fosforilasi tingkat substrat. Fermentasi sebenarnya proses yang tidak
memerlukan oksigen. Fungi mampu melakukan fermentasi dalam kondisi aerob. Jadi
fermentasi sebenarnya adalah metabolisme tanpa melibatkan oksigen, tetapi organisme
fermentatif terkadang memerlukan oksigen untuk proses metabolisme lainnya maupun
pertumbuhannya.
Prokariota mampu melakukan fermentasi dan respirasi untuk menghasilkan
energi. Respirasi aerob dilakukan, jika terdapat O2 sebagai akseptor elektron. Akan
tetapi, jika tidak ada O2, maka prokariota melakukan respirasi anaerob (jika ada akseptor
elektron selain oksigen) atau fermentasi (jika tidak ada akseptor elektron).
Sebagian besar prokariota obligat anaerob terbunuh, jika ada oksigen di
lingkungannya. Oksigen di dalam sel terakumulasi menjadi produk toksik, yaitu radikal
hidroksil, radikal superoksida, dan hidrogen peroksida. Pada eukariota produk toksik ini
dapat dinetralisir oleh peroksida dismutase atau katalase, tetapi prokariota obligat
anaerob tidak memiliki enzim-enzim tersebut.
Sebagian besar prokariota fermentatif menghasilkan semua ATP melalui
fosforilasi tingkat substrat, kemudian ATP dihidrolisis oleh ATPsintase, sehingga dapat
menghasilkan p yang dipakai untuk aktivitas membran.

Pada lingkungan anaerob dapat terjadi rantai makanan. Karbohidrat, protein,
lemak, dan senyawa organik lainnya difermentasi menjadi alkohol atau asam organik
oleh organisme anaerob A. Asam organik atau etanol difermentasi menjadi CO2 oleh
organisme anaerob B. CO2 ditambat oleh organisme anaerob C menjadi karbohidrat,
protein, lemak, dan lainnya. Proses rantai makanan di lingkungan anaerob disebut rantai
makanan anaerob.
Pada organisme fermentatif permasalahan besar adalah mengoksidasi NADH
yang dihasilkan dari proses glikolisis. Pada organisme respiratif masalah tersebut dapat
diatasi, karena NADH langsung dioksidasi melalui serial transfer elektron sampai ke
akseptor elektron. Fermentasi sebagian besar terjadi, karena ketiadaan akseptor
elektron respiratif. Oleh karena itu, organisme fermentatif harus menyediakan akseptor
elektron alternatif untuk menerima elektron dari NADH.
Proses transfer elektron yang terjadi pada fermentasi disebut pembuangan
elektron. Disebut demikian karena elektron bukan melewati serial transfer elektron,
tetapi hanya sekali transfer elektron ke akseptor elektron. Biasanya produk yang
dihasilkan dari pembuangan elektron disekresi ke luar sel (kalau produk hasil respirasi
masih dapat dimanfaatkan lagi oleh sel), karena dapat meracuni sel prokariota, baik
secara langsung, maupun tidak langsung. Etanol dapat membunuh prokariota, karena
sebagian besar prokariota mati oleh akumulasi etanol dalam media. Asam organik dapat
membunuh prokariota secara tidak langsung, karena asam organik akan menurunkan
nilai pH, sehingga sebagian besar protein terdenaturasi. Beberapa hal yang perlu
diperhatikan dalam meningkatkan produk fermentasi adalah meminimalisir pertambahan
biomassa, menggunakan strain homofermentatif, dan mempertahankan lingkungan
pada kondisi ideal.
KESETIMBANGAN FERMENTASI
Pada fermentasi sering dihasilkan lebih dari satu senyawa (multi-produk). Oleh
karena itu, kita harus dapat menghitung apakah telah terjadi fermentasi sempurna atau
tidak. Jika tidak, maka pasti terdapat produk lain yang tidak terukur.
Kesetimbangan fermentasi dapat dihitung dengan 3 cara, yaitu metode O/R (O/R
method), metode ketersediaan hidrogen (H available method), metode rekoveri karbon
(C recovery method). Ketiga metode ini untuk menghitung keseimbangan konsumsi
substrat dan produk yang dihasilkan oleh mikroba (Tabel 13.1). Nilai ideal dari
kesetimbangan fermentasi adalah 1. Hal ini berarti semua substrat dikonversi menjadi
produk fermentasi.

Kesetimbangan fermentasi dapat dipakai untuk mengukur efisiensi fermentasi.
Efisiensi fermentasi dapat diukur berdasarkan produk secara keseluruhan atau
berdasarkan produk yang diinginkan. Pada kondisi aplikatif efisiensi fermentasi biasanya
dihitung berdasarkan produk yang diinginkan.
Pada fermentasi etanol efisiensi produksi etanol dihitung berdasarkan
perbandingan berat etanol dengan konsumsi substrat. Misalnya 1 kg molase
difermentasi menjadi 550 g etanol, efisiensi fermentasi adalah
550/1000 x 100% = 55%
Hal ini berarti hanya 55% molase yang diubah menjadi etanol.
Tabel 13.1 Kesetimbangan fermentasi glukosa oleh Lactobacillus pentoaceticus.Senyawa mmol Mmol
karbonNilai O/R
Produk teroksidasi
Produk tereduksi
Hidrogen tersedia
Mmol hidrogen
Glukosa 100 600 0 24 2400LaktatGliserolEtanolAsetatCO2
1007
807
85
30021
1601485
0-1-20
+2 170
7160
12141280
120098
96056
Total 580 170 167 2314Kesetimbangan fermentasi berdasarkan metode O/R adalah 167/170=0,98
metode ketersediaan H adalah 2314/2400=0,96 metode rekoveri karbon= 580/600=0,97
FERMENTASI ETANOL
Fermentasi etanol dilakukan baik oleh eukariota dan prokariota. Eukariota yang
paling sering dipakai sebagai fermentor etanol adalah khamir (Saccharomeces spp).
Prokariota yang sering digunakan dalam fermentasi etanol adalah Pseudomonas dan
Zymomonas mobilis.
Fermentasi Etanol oleh Khamir (Saccharomeces cerevisiae)
Khamir Saccharomeces cerevisiae menggunakan jalur EMP dalam
memfermentasi glukosa menjadi etanol pada kondisi netral atau sedikit asam dan
anaerob. Pada kondisi mikroaerofil S. cerevisiae melakukan respirasi. Pada kondisi
tersebut 10% glukosa biasanya direspirasi menjadi CO2. Fermentasi etanol oleh S.
cerevisiae menghasilkan etanol kurang dari 50%. Pada kondisi aerob khamir melakukan
respirasi.
Terdapat perubahan produk pada fermentasi etanol akibat perubahan kondisi
media. Jika pada media terkandung natrium sulfit, maka menghasilkan gliserol sebagai
produk yang dominan. Hal ini karena asetaldehid terjerat oleh sulfit, sehingga menjadi

bisulfid. Pada kondisi ini asetaldehid tidak dapat menjadi akseptor elektron bagi NADH,
sehingga gliserol fosfat berperan sebagai akseptor elektron bagi NADH dan gliserol
fosfat diubah menjadi gliserol. Reaksi keseluruhan adalah sebagai berikut.
C6H12O6 + HSO3- C3H8O3 + CO2 + C2H4O-HSO3
- glukosa sulfit gliserol asetaldehidbisulfid
Jika media dalam kondisi alkali, terdapat perubahan komposisi produk. Pada
kondisi alkali glukosa akan diubah menjadi gliserol, etanol, asetat, dan CO2. Pada
kondisi alkali maka asetaldehid akan dioksidasi menjadi asetat dan NADH. NADH
dipakai untuk mereduksi asetaldehid lainnya menjadi etanol. NADH hasil oksidasi
glukosa menjadi 2 asetaldehid digunakan untuk mereduksi dihidroksiaseton fosfat
menjadi gliserol fosfat, kemudian menjadi gliserol.
glukosa + 2 NAD+ 2 asetaldehid + 2 NADH + 2 H+ + 2 CO2 glukosa + 2 ATP 2 dihidroksiaseton fosfat + 2 ADPasetaldehid + NAD+ + H2O asetat + NADH + H+
asetaldehid + NADH + H+ NAD+ + etanol2 dihidroksiaseton fosfat + 2 NADH + 2 H+ 2 gliserol fosfat + 2 NAD+
2 gliserol fosfat 2 gliserol + 2 Pi +. 2 C6H12O6 + 2 ATP + H2O 2 C3H8O3 + 2 CO2 + C2H6O + C2H4O2 +2(ADP+Pi) glukosa gliserol etanol asetat
Gambar 13.1 Perbandingan jalur fermentasi etanol oleh S. cerevisiae (kiri) dan Z. mobilis (kanan).

Fermentasi Etanol oleh Zymomonas mobilis
Zymomonas mobilis merupakan prokariota obligat fermentatif dan mampu
memfermentasi glukosa menjadi etanol dan karbon dioksida (Gambar 13.1). Glikolisis
glukosa menjadi piruvat melalui jalur Entner Doudoroff. Etanol diperoleh dari
dekarboksilasi piruvat menjadi asetaldehid dan reduksi asetaldehid menjadi etanol.
Pada fermentasi etanol dihasilkan produk sampingan, yaitu karbon dioksida.
Karena berat molekul etanol dan karbondioksida relatif sama, maka efisiensi produksi
etanol dari glukosa adalah 50%.
Beberapa keuntungan fermentasi etanol dengan menggunakan Z. mobilis
dibandingkan S. cerevisiae. Keuntungan itu adalah sebagai berikut. Efisiensi produksi
etanol dapat meningkat sampai mendekati 50%. Produksi etanol relatif cepat, karena Z.
mobilis mempunyai piruvat dekarboksilase lebih banyak, sehingga dengan cepat piruvat
didekarboksilasi menjadi asetaldehid, kemudian menjadi etanol. Produk samping relatif
sedikit, itupun didominasi CO2. Hal ini karena Z. mobilis merupakan bakteri obligat
fermentatif. Produksi etanol dalam media mampu mencapai 40%. Hal ini karena
toleransi Z. mobilis yang tinggi terhadap etanol.
FERMENTASI ASAM LAKTAT
Bakteri asam laktat mampu mengubah glukosa menjadi asam laktat. Bakteri
tersebut adalah Lactobacillus, Streptococcus, Leuconostoc, Pediococcus, dan
Bifidobacterium. Ada 2 kelompok fermentasi asam laktat, yaitu homofermentatif dan
heterofermentatif. Yang disebut lebih dulu menggunakan glikolisis melalui jalur EMP dan
yang satunya menggunakan glikolisis melalui jalur HMP.
Fermentasi Asam Laktat Homofermentatif
Bakteri asam laktat homofermentatif menghasilkan mayoritas asam laktat
dengan sedikit produk samping, yaitu gliserol, etanol, asetat, format dan CO2 (Gambar
13.2). Bakteri asam laktat homofermentatif mengoksidasi glukosa menjadi 2 piruvat
melalui jalur EMP. Pada jalur ini menghasilkan 2 ATP. NADH yang dihasilkan pada jalur
ini dipakai untuk mereduksi piruvat menjadi asam laktat. Reaksi keseluruhan adalah
Glukosa + 2ADP + 2Pi 2 Laktat + 2 ATP
Produk samping diperoleh, karena bakteri asam laktat homofermentatif
mempunyai berbagai enzim yang dapat mengubah piruvat menjadi etanol dan CO2,
asetat dan format, dan laktat. Jika piruvat tidak segera diubah menjadi produk di atas,
NADH dipakai untuk mereduksi dihidroksi aseton fosfat menjadi gliserol.

Gambar 13.2 Fermentasi asam laktat homofermentatif.
Perubahan nilai pH pada media dapat mengubah komposisi produk fermentasi
asam laktat homofermentatif Lactobacillus casei (Tabel 13.2). Fermentasi asam laktat
idealnya dilakukan pada kondisi asam. Ketika kondisi diubah menjadi netral, sebagian
piruvat dioksidasi menjadi asetil KoA dan format. Asetil KoA kemudian tereduksi menjadi
asetat dan etanol.
Tabel 13.2 Efek pH pada fermentasi laktat oleh Lactobacillus caseiProduk pH 5 pH 7 pH 8
LaktatAsetatFormatEtanol
176,39,5
13,15,0
134,319,226,612,7
102,031,547,318,7
Efisiensi fermentasi (%) 91,3 87,7 84,0
Pada kondisi aerob S. faecalis mampu memfermentasi gliserol menjadi laktat.
Hal ini karena oksigen digunakan untuk mereduksi koenzim Fp (flavoprotein) menjadi Fp
dan hidrogen peroksida. Reaksi keseluruhan adalah sebagi berikut.
Gliserol + ADP + Pi + O2 laktat + H2O2 + ATP
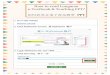
Fermentasi Asam Laktat Heterofermentatif
Bakteri asam laktat heterofermentatif menghasilkan asam laktat dan produk
fermentasi lainnya (kebanyakan etanol) dengan rasio yang seimbang (Gambar 13.3).

Hal ini karena mereka mengoksidasi glukosa menjadi piruvat dan asetil fosfat melalui
jalur HMP. Piruvat kemudian direduksi menjadi asam laktat, sedangkan asetil fosfat
kemudian direduksi menjadi etanol. Pada jalur ini menghasilkan 1 ATP. Reaksi
keseluruhan adalah.
Glukosa + ADP + Pi Laktat + etanol + CO2 + ATP
Gambar 13.3 Fermentasi asam laktat heterofermentatif.
Bakteri Streptococcus mutans mempunyai kemampuan dalam memfermentasi
glukosa menjadi laktat (heterofermentatif) dalam suasana aerob. Pada kondisi aerob
NADH dioksidasi menjadi NAD+ dengan bantuan oksigen dan NADH oksidase. Oleh
karena itu, terdapat perubahan produk, di mana etanol diubah menjadi asetil KoA dan

kemudian menjadi asetat. Perubahan asetil KoA menjadi asetat menghasilkan ATP.
Jamur Rhizopus oryzae juga mempunyai kemampuan memfermentasi karbohidrat (pati
dan glukosa) menjadi etanol dan asam laktat secara aerob.
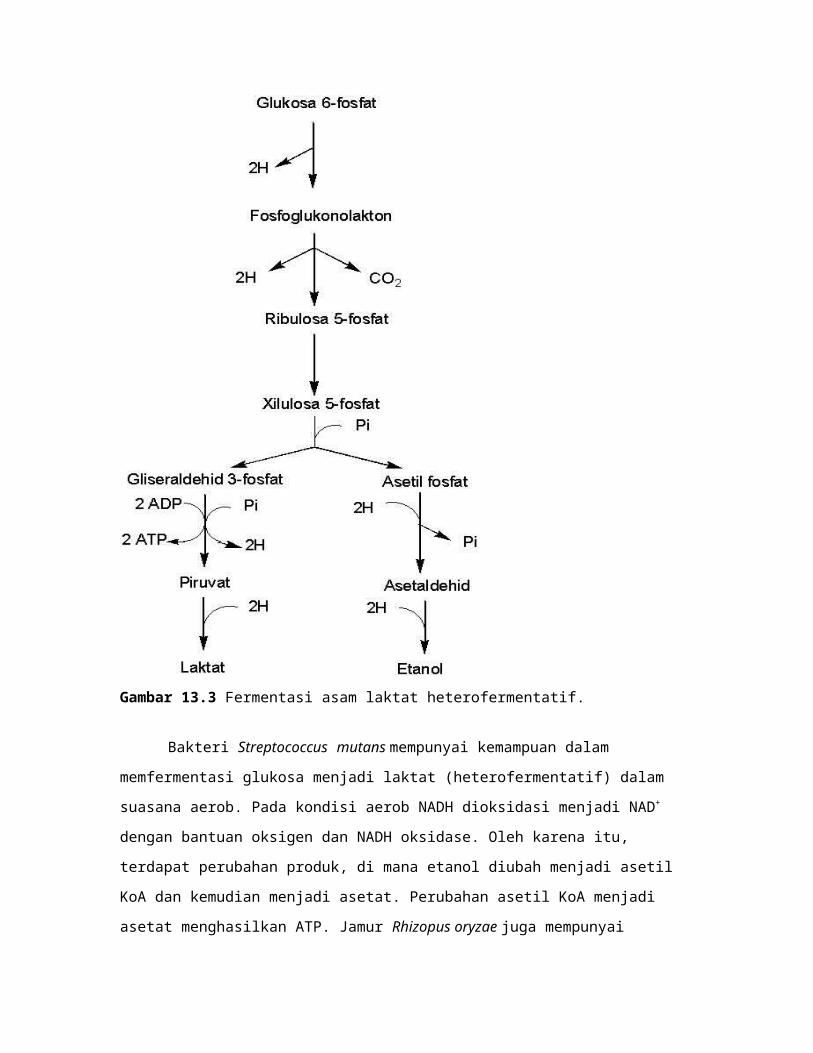
Gambar 13.4 Fermentasi asam laktat melalui jalur bifidium.
Fermentasi Asam Laktat Melalui Jalur Bifidium
Dinamakan jalur bifidium, karena ditemukan pada Bifidobacterium bifidium.
Secara keseluruhan fermentasi asam laktat melalui jalur bifidium adalah
2 Glukosa + 5 ADP + 5 Pi 3 asetat + 2 laktat + 5 ATP

Fermentasi asam laktat pada jalur ini melalui glikolisis melalui modifikasi jalur
Pentosa Fosfat (Gambar 13.4). Dua molekul glukosa difosforilasi menjadi 2 molekul
fruktosa 6-fosfat (perlu 2 ATP). Satu molekul fruktosa 6-fosfat dipecah menjadi eritrosa
4-fosfat dan asetil fosfat. Eritrosa 4-fosfat kemudian bereaksi dengan satu molekul
fruktosa 6-fosfat lainnya menghasilkan sedoheptulosa 7-fosfat dan fosfogiseraldehid.
Sedoheptulosa 7-fosfat bereaksi lagi dengan fosfogiseraldehid menghasilkan xilulosa 5-
fosfat dan ribulosa 5-fosfat. Ribulosa 5-fosfat berisomerasi menjadi xilulosa 5-fosfat.
Dua molekul xilulosa 5-fosfat dipecah menjadi 2 fosfogliseraldehid dan 2 asetil
fosfat. Dua molekul fosfogliseraldehid dioksidasi menjadi 2 piruvat kemudian direduksi
menjadi 2 laktat (menghasilkan 4 ATP). Dua molekul asetil fosfat diubah menjadi 2
asetat (menghasilkan 2 ATP).
FERMENTASI ASETAT
Clostridium thermoaceticum mampu memfermentasi piruvat menjadi asetat
melalui jalur asetil KoA. Karena produknya asetat, maka disebut juga asetogenesis.
Selain C. thermoaceticum, bakteri pereduksi sulfat Desulfotomaculum thermobenzoicum
juga mampu memfermentasi piruvat menjadi asetat (Gambar 13.5) di media tanpa
sulfat. Kedua prokariota ini melakukan fermentasi piruvat menjadi asetat dengan model
dan enzim yang sama. Kedua prokariota juga mampu menambat CO2 menjadi asetat
juga dengan model yang sama. Yang membedakan adalah protein pembawa karbon
(lihat penambatan CO2 pada bab sebelumnya).
Piruvat diperoleh dari hasil glikolisis glukosa. Empat molekul piruvat dioksidasi
oleh piruvat dehidrogense menjadi 4 molekul asetil KoA. Pergantian gugus KoA dengan
fosfat oleh fosfotransasetilase membuat asetil KoA berubah menjadi asetil fosfat.
Defosforilasi (dikopling dengan sintesis ATP) asetil fosfat oleh asetat kinase
menghasilkan asetat.
Pada proses fermentasi piruvat menjadi asetat menghasilkan 8 molekul NADH.
NADH dipakai untuk menambat CO2 menjadi asetat. Enam elektron dipakai untuk
menambat CO2 menjadi gugus metil [CH3] melalui jalur asetil KoA pada metanogen. Dua
elektron dipakai untuk menambat CO2 menjadi kompleks karbonmonoksida
karbonmonoksida dehidrogenase (CO-CODH). Selanjutnya, gugus metil diinkorporasi ke
kompleks CO-CODH menjadi CH3-CO-CODH. Gugus CH3-CO- ditransfer ke KoA juga
oleh karbonmonoksida dehidrogenase, sehingga menjadi asetil KoA. Selanjutnya asetil
KoA diubah menjadi asetat sama seperti pada fermentasi piruvat menjadi asetat. Reaksi

keseluruhan asetogenesis Desulfotomaculum thermobenzoicum adalah 4 Piruvat + 4
ADP + 4 Pi 5 asetat + 2 CO2 + 4 ATP
Gambar 13.5 Asetogenesis dari piruvat pada Desulfotomaculum thermobenzoicum.
Pada bakteri asam asetat lainnya, misalnya Acetobacter dan Gluconobacter
mampu memfermentasi glukosa dan etanol menjadi asetat. Gluconobacter suboxydans
tidak mempunyai enzim-enzim pada siklus asam sitrat, kecuali suksinat dehidrogenase.
Modifikasi siklus pentosa fosfat digunakan untuk memfermentasi glukosa
menjadi asetat (Gambar 13.6). Glukosa difosforilasi menjadi glukosa 6-fosfat, kemudian
diisomerisasi menjadi 2 molekul fruktosa 6-fosfat. Fruktosa 6-fosfat dipecah menjadi
asetil fosfat dan eritrosa 4-fosfat. Asetil fosfat dideforforilasi menjadi asetat. Eritrosa 4-
fosfat bereaksi dengan fruktosa 6-fosfat menjadi sedoheptulosa 7-fosfat dan
gliseraldehid 3-fosfat. Reaksi ini dikatalisis transaldolase. Sedoheputosa 7-fosfat dan
gliseraldehid 3-fosfat bereaksi menjadi ribulosa 5-fosfat dan xilulusa 5-fosfat. Reaksi ini
dikatalisis transketolase. Xilulosa 5-fosfat diisomerisasi menjadi ribulosa 5-fosfat. Dua
molekul Ribulosa 5-fosfat dipecah menjadi 2 molekul gliseraldehid 3-fosfat dan 2
molekul asetil fosfat. Dua molekul asetil KoA didefosforilasi menjadi asetat. Dua molekul
gliseraldehid 3-fosfat berkondensasi menjadi fruktosa 1,6-bisfosfat. Fruktosa 1,6-
bisfosfat terdefosforilasi menjadi fruktosa 6-fosfat. Fruktosa 6-fosfat menggantikan
fruktosa 6-fosfat yang bereaksi dengan eritrosa 4-fosfat.
Gambar 13.6 Fermentasi asetat dari glukosa oleh Gluconobacter.
Karakteristik Asetobacter dan Gluconobacter adalah kemampuannya
mengoksidasi etanol menjadi asetat. Dua dehidrogenase membran sel terlibat dalam
oksidasi etanol menjadi asetat, yaitu etanol dehidrogenase dan asetaldehid
dehidrogenase. Reaksi keseluruhan oksidasi fermentasi etanol menjadi asetat adalah
sebagai berikut.
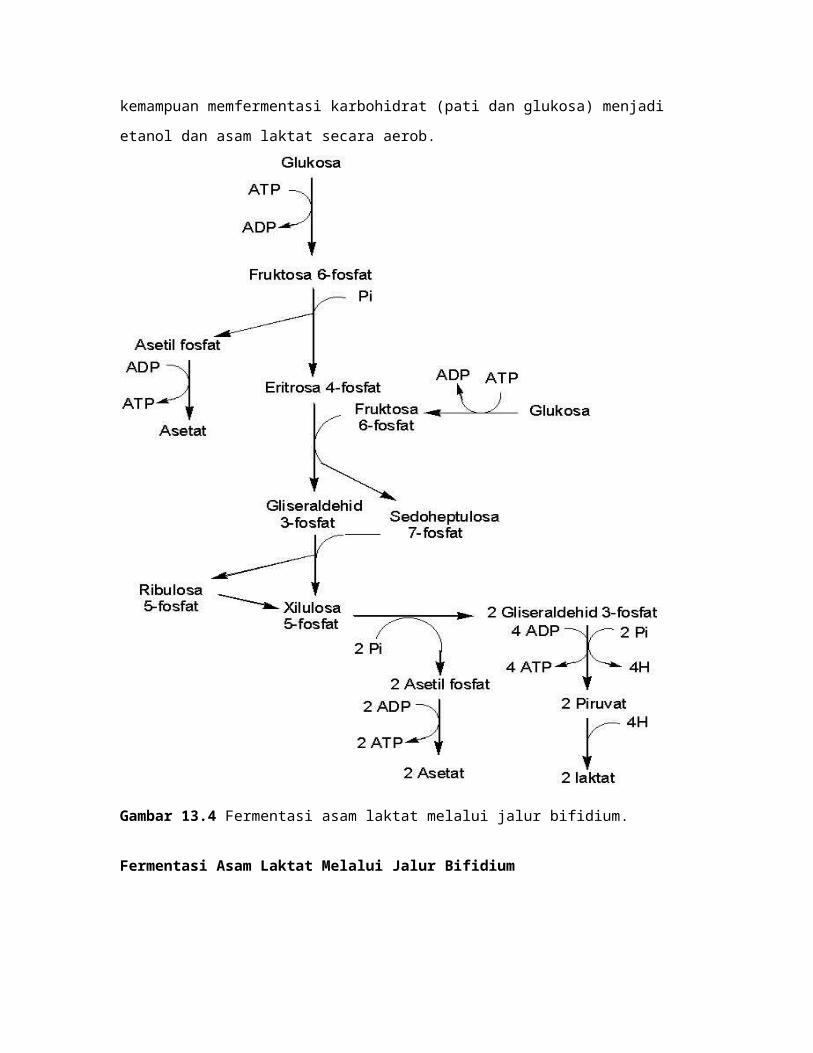
CH3CH2OH + ½ O2 CH3COOH + 2H etanol asetat
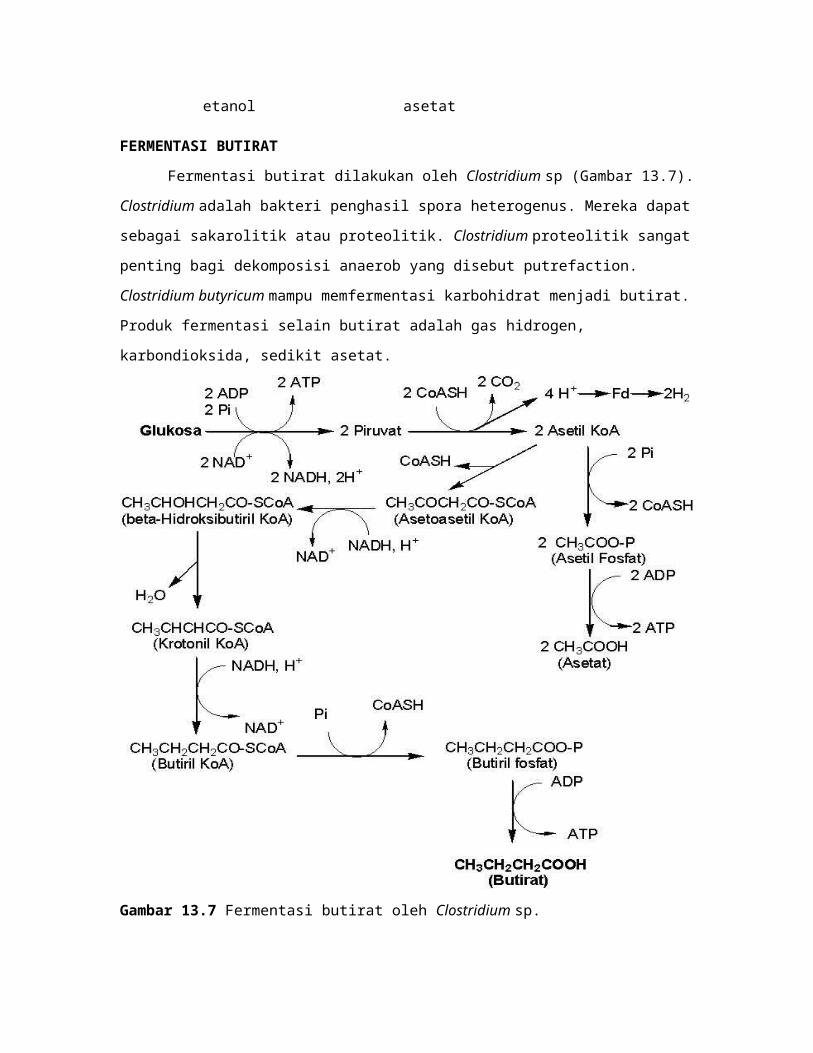
FERMENTASI BUTIRAT
Fermentasi butirat dilakukan oleh Clostridium sp (Gambar 13.7). Clostridium
adalah bakteri penghasil spora heterogenus. Mereka dapat sebagai sakarolitik atau
proteolitik. Clostridium proteolitik sangat penting bagi dekomposisi anaerob yang disebut
putrefaction. Clostridium butyricum mampu memfermentasi karbohidrat menjadi butirat.
Produk fermentasi selain butirat adalah gas hidrogen, karbondioksida, sedikit asetat.
Gambar 13.7 Fermentasi butirat oleh Clostridium sp.
Glukosa dipecah menjadi piruvat melalui jalur EMP (menghasilkan 4 elektron dan
2 ATP). Piruvat didekarboksilasi oleh piruvatferedoksi eksidoreduktase menjadi asetil
KoA, CO2. H2 diperoleh dari aktivitas oksidasi hidrogenase terhadap feredoksin. Dua
molekul asetil KoA berkondensasi menghasilkan asetoasetil KoA dengan bantuan asetil

KoA asetiltransferase. Asetoasetil KoA direduksi menjadi -hidroksibutiril KoA oleh
dehidrogenase. -Hidroksibutiril KoA didehidrasi menjadi krotonil KoA oleh krotonase.
Krotonil KoA direduksi menjadi butiril KoA oleh butiril KoA dehidrogenase. Pengantian
gugus KoA oleh fosfat mengakibatkan butiril KoA menjadi butiril fosfat. Reaksi ini
dikatalisis fosfotransbutirilase. Butiril fosfat didefosforilasi (dikopling dengan sintesis
ATP) menjadi butirat oleh butirat kinase.
C. tyrobutyricum mampu memproduksi butirat dan asetat dari glukosa (Gambar
13.7). Untuk menurunkan produksi asetat, gen pta yang mengkode asetat kinase dapat
dihilangkan. Meskipun gen pta dihilangkan, tetapi C. tyrobutyricum masih mampu
menghasilkan asetat. Hal ini menunjukkan bahwa terdapat jalur atau enzim lain yang
mampu menghasilkan asetat.
Bakteri rumen Butyrivibrio fibrisolvens mampu memfermentasi glukosa menjadi
butirat. Fermentasi glukosa menjadi butirat oleh Butyrivibrio fibrisolvens melalui rute/jalur
yang sama dengan Clostridium. Pada kondisi pertumbuhan tinggi (pada fase
eksponensial) glukosa difermentasi menjadi butirat, asetat, H2 dan CO2. Asetat
merupakan produk samping dan diperoleh dari forforilasi asetil KoA menjadi asetil fosfat
oleh fosfotransasetilase. Asetil fosfat kemudian didefosforilasi menjadi asetat oleh asetat
kinase.
Tabel 13.3 Perbandingan kadar produk pada fermentasi butirat dan Butanol-aseton (mmol/100 mmol glukosa)Produk C.
saccharo-butyricum
C. aceto-butylicum
C. butylicum
C. thermac-charolyticum
B. aceto- ethylicum
B. polymyxa
CO2
H2
AsetatButiratFormatLaktatEtanolButanolAsetonIsopropanolAsetoin2,3-Butandiol
195,5233,0
42,675,3
Sedikit
220,0165,9
24,87,1
4,947,422,3
5,7
207,0111,1
20,314,5
50,2
18,0
174,0229,4
48,559,5
25,7
215,0137,0
16,0
10,0
122,0
28,0
12,0
195,054,0
5,0
95,04,56,0
39,0Rekoveri C % 97,0 98,0 93,5 98,0 105,0 100,0Sumber: Moat A.G. & J.W. Foster. 1995. Microbial Physiology. Wiley-Lyss, New York

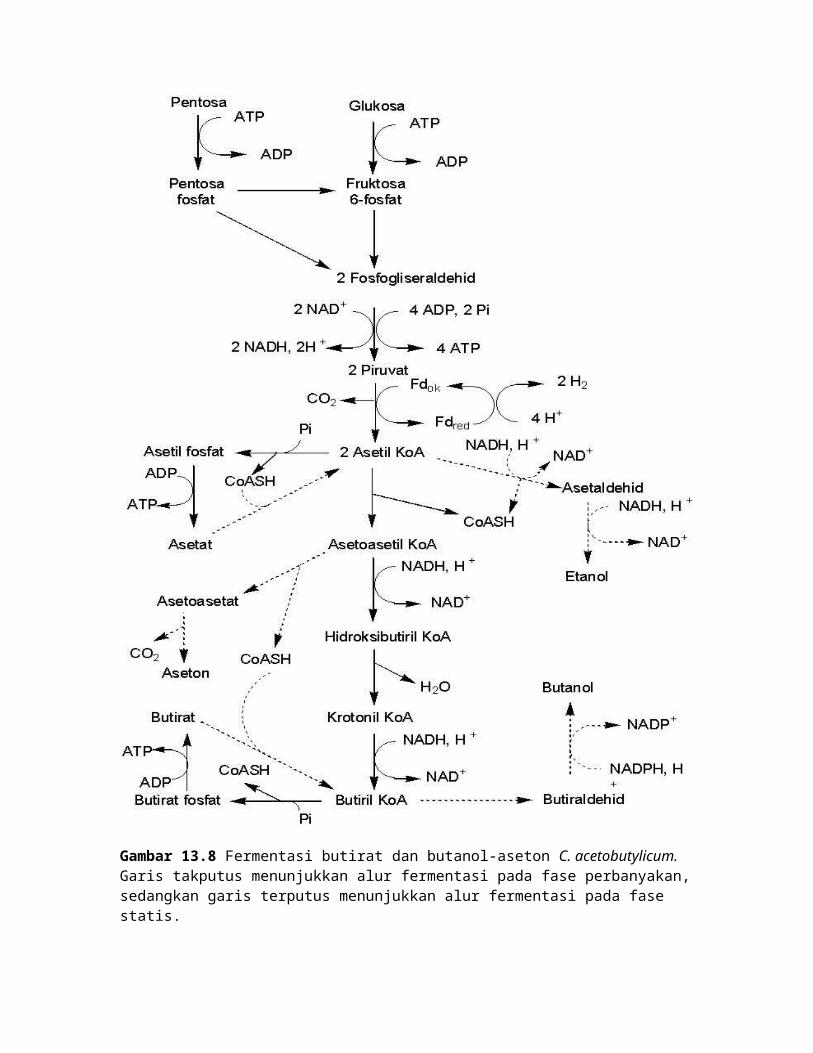
Gambar 13.8 Fermentasi butirat dan butanol-aseton C. acetobutylicum. Garis takputus menunjukkan alur fermentasi pada fase perbanyakan, sedangkan garis terputus menunjukkan alur fermentasi pada fase statis.

Ketika sel masuk ke fase statis dan kandungan butirat tinggi, terjadi fermentasi
glukosa dan pentosa menjadi aseton. Selain itu, terjadi konsumsi butirat asetat menjadi
butanol dan etanol (Gambar 13.8 dan Tabel 13.3).
Glukosa dan pentosa diglikolisis menjadi piruvat. Piruvat didekarboksilasi
menjadi asetil KoA oleh piruvatferedoksin oksidoreduktase. Kondensasi 2 molekul asetil
KoA menjadi asetoasetil KoA oleh asetil KoA transasetilase. Asetoasetat dipecah
menjadi aseton dan CO2 oleh asetoasetat dekarboksilase.Gugus KoA dari asetoasetil
KoA ditransfer ke butirat atau asetat, sehingga menjadi butiril KoA atau asetil KoA .
Reaksi ini dikatalisis asetoasetil KoA-butirat atau asetat-KoA transferase. Butiril KoA
direduksi menjadi butiraldehid oleh butiraldehid dehidrogenase, kemudian direduksi
menjadi butanol oleh butanol dehidrogenase. Sedangkan asetil KoA direduksi menjadi
asetaldehid oleh asetaldehid dehidrogenase, kemudian direduksi menjadi etanol oleh
etanol dehidrogenase.
FERMENTASI PROPIONAT
Propionat, asetat, dan karbon dioksida merupakan produk utama dari fermentasi
laktat, glukosa, dan gliserol oleh Propiniobacterium, Veillonella, Bacteroides dan
beberapa Clostridium spp. Hipotesis awal menyatakan bahwa langkah awal fermentasi
propionat adalah dehidrasi laktat menjadi akrilat. Akrilat kemudian direduksi menjadi
propionat. Rute tersebut teramati pada Clostridium propionicum, Bacteroides rumicola,
dan Peptostreptoccus. Pada Propionibacterium dan Veillonella pembentukan propionat
melalui rute yang lebih kompleks.
Fermentasi Propionat oleh Clostridium propionicum
C. propionicum mampu memfermentasi asam laktat menjadi asetat (melalui jalur
asetil KoA) dan propionat (melalui jalur akrilil KoA) dan menghasilkan 1 ATP (Gambar
13.9). Satu molekul laktat didehidrogenasi menjadi piruvat oleh laktat dehidrogenase.
Piruvat didehidrogenasi dan dekarboksilasi menjadi asetil KoA oleh piruvatferedoksin
oksidoreduktase. Gugus fosfat menggantikan gugus KoA oleh fosfotransasetilase,
sehingga Asetil KoA diubah menjadi asetil fosfat. Asetil fosfat didefosforilasi (dikopling
dengan sintesis ATP) menjadi asetat oleh asetat kinase. Pada jalur ini menghasilkan 1
ATP, 1 CO2, dan 4 elektron. Empat elektron dipakai untuk mereduksi 2 molekul laktat
menjadi 2 molekul propionat.
Gugus KoA (berasal dari propionil KoA) ditransfer ke laktat oleh KoA transferase,
sehingga menjadi laktil KoA. laktil KoA terdehidrasi menjadi akrilil KoA. Reaksi ini
dikatalisis akrililase. Akrilil KoA direduksi menjadi propionil KoA oleh propionil KoA

dehidrogenase. Propionil KoA diubah menjadi propionat. Reaksi keseluruhan adalah
sebagai berikut.
3 Laktat + ADP + Pi 2 propionat + asetat + CO2 + ATP
Gambar 13.9 Fermentasi propionat melalui jalur akrilat oleh C. propionicum.
Fermentasi Propionat oleh Propionibacterium
Propionibacterium memfermentasi laktat, triosa, dan heksosa menjadi propionat
(jalur suksinat-propionat), asetat (jalur asetil KoA), dan karbon dioksida (Gambar 13.10
dan Tabel 13.4). Tiga molekul laktat diubah menjadi tiga molekul piruvat oleh laktat
dehidrogenase. Satu molekul piruvat diubah menjadi satu molekul asetat sama seperti
pada C. propionicum. Fermentasi laktat menjadi asetat menghasilkan 2 elektron dan
perubahan 2 molekul laktat menjadi 2 molekul piruvat menghasilkan 6 elektron. Delapan
elektron ini dipakai untuk mereduksi piruvat menjadi propionat.
Piruvat dikarboksilasi (berasal dari dekarboksilasi metilmalonil KoA) menjadi
oksaloasetat oleh transkarboksilase. Reduksi oksaloasetat menjadi malat oleh malat
dehidrogenase. Malat dihidrasi menjadi fumarat oleh fumarase. Fumarat direduksi
menjadi suksinat oleh fumarat reduktase. Transfer gugus KoA (berasal dari propionil
KoA) ke suksinat, sehingga menjadi suksinil KoA. Reaksi ini dikatalisis suksinil KoA
transferase. Re-aransemen suksinil KoA menjadi metil malonil KoA oleh metil malonil
KoA rasemase. Dekarboksilasi metil malonil KoA oleh transkarboksilase, sehingga
menjadi propionil KoA. Propionil KoA diubah menjadi propionat. Secara teoritis rasio
propionat dan asetat adalah 2, tetapi tidak jarang rasionya lebih dari 2 (Tabel 13.4).
Reaksi keseluruhan adalah sebagai berikut.
3 Laktat + 2 ADP + 2 Pi 2 propionat + asetat + CO2 + 2 ATP
Gambar 13.10 Fermentasi propionat oleh Propionibacterium.
Tampak bahwa reduksi piruvat menjadi suksinil KoA merupakan rute pada jalur
reduktif-asam sitrat, Jadi boleh dikatakan bahwa fermentasi propionat pada
Propionibacterium melalui jalur reduktif-asam sitrat. Produksi propionat dan asetat dapat
ditingkatkan, jika tekanan gas CO2 diturunkan.
FERMENTASI ASAM CAMPURAN
Pada kondisi anaerob dan ketiadaan akseptor elektron, anggota
Enterobacteriaceae (Escherichia, Enterobacter, Salmonella, Klebsiella, dan Shigella)
memfermentasi glukosa menjadi campuran asetat, format, suksinat, etanol, CO2 dan H2
(Tabel 13.5). Semua produk diperoleh dari fosfoenol piruvat (PEP) (Gambar 13.11).
Atau lebih tepatnya suksinat dari PEP. Lainnya dari piruvat (piruvat diperoleh dari PEP).
Tabel 13.4 Fermentasi propionat oleh Propionibacterium (mmol/100 mmol glukosa)Bakteri Substrat Propionat Asetat CO2 Suksinat Rasio
P/A
P. freudenreichii
P. freudenreichiiP. freudenreichiiP. shermaniiP. shermaniiP. shermaniiP. peterssoniiP. arabinosum
GlukosaLaktat
GliserolGlukosaLaktat
GliserolGlukosaGlukosa
134,063,5
100,0140,0
62,5102,0114,0148,0
52,635,3
9,956,836,510,654,010,0
46,235,8
6,956,437,0
7,351,063,6
12,637,811,712,0
9,311,211,1
7,9
2,561,8
10,02,46
1,710,0
2,114,8
Sumber: Moat A.G. & J.W. Foster. 1995. Microbial Physiology. Wiley-Lyss, New York
Gambar 13.11 Fermentasi asam campuran.

Suksinat diperoleh dari karboksilasi PEP melalui jalur reduktif-asam sitrat (jalur
suksinat). PEP diubah menjadi oksaloasetat oleh PEP karboksilase. Perubahan
oksaloasetat menjadi suksinat melalui rute dan melibatkan enzim yang sama seperti
pada perubahan oksaloasetat menjadi suksinat pada fermentasi propionat pada bakteri
Propionibacterium. Laktat diperoleh langsung dari reduksi piruvat oleh laktat
dehidrogenase. Format diperoleh dari pemecahan piruvat (hasil lain adalah asetil KoA),
kemudian dapat diubah CO2 dan H2. Asetil KoA dapat diubah menjadi etanol maupun
asetat.
Lactobaciilus helveticus memfermentasi sitrat dan laktosa menjadi laktat. Akan
tetapi, jika laktosa ditiadakan, terjadi perubahan produk fermentasi, yaitu menghasilkan
asetat dan suksinat, bukan laktat. Asetoin dan diasetil tidak terdeteksi pada produk
fermentasi L. helveticus. Tampaknya produksi asetat dari piruvat (hasil konversi sitrat)
diperantarai NADH okdidase, bukan asetat kinase.
Tabel 13.5 Produk hasil fermentasi asam campuran (mmol/100 mmol glukosa)Produk Escherichia
coliEnterobacter
aerogenesSalmonella
typhiiLaktatEtanolAsetatFormat CO2
H2
Suksinat 2,3-butandiol
108,841,332,0
1,654,045,218,0
0
53,459,410,1
5,5126,9
44,26,0
34,6
121,725,425,639,3
00
10,80
Rekoveri C (%) 100,0 99,5 93,3Sumber: Moat A.G. & J.W. Foster. 1995. Microbial Physiology. Wiley-Lyss, New York
FERMENTASI BUTANDIOL
Fermentasi butandiol ditandai dengan produksi 2,3-butandiol dan asetoin (juga
laktat, etanol, dan format; Gambar 13.12).Glukosa dioksidasi melalui glikolisis jalur EMP
menjadi piruvat. Terdapat 3 arah metabolisme piruvat pada fermentasi butandiol. Piruvat
direduksi menjadi laktat oleh laktat dehidrogenase. Piruvat juga dapat dipecah menjadi
asetil KoA dan format oleh piruvat format liase. Asetil KoA terduksi menjadi asetaldehid
oleh asetaldehid dehidrogenase, kemudian direduksi menjadi etanol oleh etanol
dehidrogenase. Piruvat juga mengalami dekarboksilasi menjadi asetaldehid aktif,
kemudian diubah menjadi asetolaktat. Kedua reaksi ini dikatalisis asetolaktat sintase.
Dekarboksilasi asetolaktat menjadi asetoin oleh asetolaktat dehidrogenase. Reduksi
Asetoin menjadi 2,3-butandiol oleh 2,3-butandiol dehidrogenase.
Produksi butandiol dipengaruhi oleh nilai pH media. Fermentasi glukosa menjadi
butandiol mudah terjadi pada media dengan nilai pH sedikit asam, tetapi, jika nilai pH
media menjadi asam, glukosa cenderung diubah menjadi asam campuran.
Gambar 13.12 Fermentasi butandiol.
FERMENTASI BAKTERI RUMEN
Bakteri rumen Ruminococcus albus mampu memfermentasi glukosa menjadi
etanol, asetat, hidrogen, dan karbondioksida melalui glikolisis jalur EMP (Gambar
13.13). Glukosa lebih dulu dioksidasi menjadi piruvat oleh enzim-enzim pada glikolisis
jalur EMP. Piruvat kemudian didekarboksilasi oleh piruvat feredoksin oksidoreduktase
menjadi asetil KoA dan gas hidrogen. Asetil KoA direduksi menjadi asetaldehid oleh
asetaldehid dehidrogenase, kemudian direduksi menjadi etanol oleh etanol
dehidrogenase. Asetil KoA dapat juga mengalami fosforilasi menjadi asetil fosfat oleh
fosfotransasetilase. Asetil fosfat mengalami defosforilasi menjadi asetat oleh asetat
kinase. Laktat pada fermentasi rumen dihasilkan oleh bakteri laktat rumen, tetapi butirat
pada fermentasi rumen dapat dihasilkan oleh bakteri maupun protozoa rumen.
Ketika R. albus ditumbuhkan bersama (kultur campuran) dengan bakteri
metanogen terdapat perubahan produksi hasil fermentasi (Gambar 13.13). Hidrogen
yang dihasilkan melalui glikolisis jalur EMP dan dekarboksilasi piruvat dimanfaatkan oleh
bakteri metanogen untuk penambatan karbondioksida menjadi metana. Karena
ketiadaan hidrogen, maka sedikit NADH yang diproduksi R. albus. Hal ini
mengakibatkan asetil KoA bukan diubah menjadi etanol, tetapi diubah menjadi asetat.
Perubahan asetil KoA menjadi asetat tidak memerlukan NADH.
Gambar 13.13 Fermentasi etanol-asetat oleh bakteri rumen R. albus.