Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DA BAHIA
ESCOLA DE MEDICINA VETERINÁRIA
CIÊNCIA ANIMAL NOS TRÓPICOS
AVALIAÇÃO DO HEMOGRAMA, BIOQUÍMICA SÉRICA E
PERFIL ELETROFORÉTICO DE PROTEÍNAS DE TARTARUGAS
MARINHAS Caretta caretta (LINNAEUS, 1758) DE VIDA LIVRE E
EM CATIVEIRO.
THAÍS TORRES PIRES
Salvador – Bahia 2007

Universidade Federal da Bahia
Escola de Medicina Veterinária
Mestrado em Ciência Animal nos Trópicos
AVALIAÇÃO DO HEMOGRAMA, BIOQUÍMICA SÉRICA E
PERFIL ELETROFORÉTICO DE PROTEÍNAS DE TARTARUGAS
MARINHAS Caretta caretta (LINNAEUS, 1758) DE VIDA LIVRE E
EM CATIVEIRO.
THAÍS TORRES PIRES
Salvador – Bahia 2007

THAÍS TORRES PIRES
AVALIAÇÃO DO HEMOGRAMA, BIOQUÍMICA SÉRICA E PERFIL
ELETROFORÉTICO DE PROTEÍNAS DE TARTARUGAS MARINHAS
Caretta caretta (LINNAEUS, 1758) DE VIDA LIVRE E EM CATIVEIRO.
Dissertação apresentada à Escola de Medicina Veterinária da Universidade Federal da Bahia, como requisito para a obtenção do título de Mestre em Ciência Animal nos Trópicos, na área de Saúde Animal.
Orientador: Prof. Dr. José Eugênio Guimarães
Salvador – Bahia 2007

AVALIAÇÃO DO HEMOGRAMA, BIOQUÍMICA SÉRICA E PERFIL
ELETROFORÉTICO DE PROTEÍNAS DE TARTARUGAS MARINHAS
Caretta caretta (LINNAEUS,1758) DE VIDA LIVRE E EM CATIVEIRO.
THAÍS TORRES PIRES
Dissertação defendida e aprovada para obtenção do grau de Mestre em Ciência Animal
nos Trópicos.
Salvador, 29 de junho de 2007.
Comissão Examinadora:
______________________________________ Prof. Dr. José Eugênio Guimarães – UFBA
Orientador
______________________________________ Profa. Dra. Eliana Reiko Matushima – USP
______________________________________ Prof. Dr. Luis Fernando Pita Gondim – UFBA

Ao meu marido, John, por seu amor incondicional.

AGRADECIMENTOS
A Deus por ter iluminado o meu trajeto para alcançar esta vitória. Aos meus pais por toda dedicação e carinho. Ao meu marido, John, pela colaboração e companheirismo. Aos meus irmãos, Dalmar e Danilo, pelo apoio para que eu pudesse alcançar meus objetivos. Ao Professor José Eugênio Guimarães pelo exemplo de profissionalismo, orientação, colaboração e incentivo durante a realização desta pesquisa. À Professora Maria Ângela Ornelas de Almeida, pela orientação e esclarecimentos a respeito da eletroforese. Ao Projeto Tamar-Ibama, em especial a Gonzalo Rostan, Cecília Baptistotte, Guy Marcovaldi e Neca Marcovaldi pela receptividade e apoio para realização deste trabalho. À base de Arembepe, em especial a Thiago, Mari, Eduardo e Michele, pelas madrugadas na praia e apoio, sem o qual seria impossível a realização deste estudo. À Professora Thereza Cristina Calmon Bittencourt, pela ajuda com a análise estatística. A todos aqueles que de alguma forma colaboraram com este trabalho, em especial aos amigos Bruno Bastos, Elitiere Neto, Sue Yohii, Uiara Burghgrave, Gustavo Rodamilans, Jaciara Moreira e Marcelo Renan. Às companheiras de laboratório Romana, Íris e Rose. Aos maravilhosos amigos Beta, Nando, Manu, Bebel, Sam, Wi, Erica, Pedro, Bel, Driks, Marquinhos, Leo, Mari e Fred pelo incentivo constante. À CAPES pela concessão de bolsas e FAPESB pelo apoio financeiro ao projeto de mestrado. Aos animais que tanto amo, e em especial às tartarugas marinhas.

ÍNDICE
LISTA DE TABELAS ................................................................................................... vii
LISTA DE FIGURAS ...................................................................................................... x
LISTA DE ABREVIATURAS ....................................................................................... xi
RESUMO ....................................................................................................................... xii
SUMMARY .................................................................................................................. xiii
1 INTRODUÇÃO GERAL .............................................................................................. 1
2 REVISÃO DE LITERATURA .................................................................................... 7
2.1 Exames complementares em tartarugas marinhas ..................................................... 7
2.1.1 Importância dos exames complementares em tartarugas marinhas ........................ 7
2.1.2 Colheita sangüínea .................................................................................................. 7
2.1.3 Anticoagulantes ...................................................................................................... 9
2.1.4 Eritrograma ............................................................................................................. 9
2.1.4.1 Volume Globular ........................................................................................ 9 2.1.4.2 Morfologia e Contagem de Hemácias ...................................................... 10 2.1.4.3 Concentração de Hemoglobina ................................................................. 11 2.1.4.4 Índices Hematimétricos ............................................................................ 11
2.1.5 Leucograma .......................................................................................................... 11
2.1.5.1 Contagem de Leucócitos .......................................................................... 12 2.1.5.2 Contagem Diferencial de Leucócitos ....................................................... 12
2.1.6 Contagem de Trombócitos .................................................................................... 15
2.1.7 Parâmetros Bioquímicos ....................................................................................... 16
2.1.7.1 Proteína Total, Albumina e Globulina ...................................................... 16 2.1.7.2 Relação Albumina: Globulina (A:G) ........................................................ 18 2.1.7.3 Glicose ...................................................................................................... 18 2.1.7.4 Ácido úrico ............................................................................................... 19 2.1.7.5 Creatinina ................................................................................................. 20 2.1.7.6 Colesterol .................................................................................................. 20 2.1.7.7 Triglicerídeos ............................................................................................ 21 2.1.7.8 Fosfatase Alcalina ..................................................................................... 21 2.1.7.9 Aspatato Aminotransferase ....................................................................... 22
2.1.8 Eletroforese das Proteínas Séricas ........................................................................ 23

3 ARTIGOS CIENTÍFICOS ......................................................................................... 25
3.1 Artigo I: Hemograma e bioquímica sérica de tartarugas cabeçudas (Caretta caretta) de vida livre e mantidas em cativeiro, no Litoral Norte da Bahia ........................... 25
3.1.1 Introdução .....................................................................................................26 3.1.2 Materiais de Métodos ...................................................................................27 3.1.3 Resultado .....................................................................................................29 s3.1.4 Discussão .....................................................................................................31 .3.1.5 Conclusão .....................................................................................................36 3.1.6 Referências Bibliográficas ............................................................................37
3.2 Artigo II: Eletroforese de proteínas séricas de tartarugas cabeçudas (Caretta caretta) de vida livre e mantidas em cativeiro no Litoral Norte da Bahia .............................40
3.2.1 Introdução .....................................................................................................41 3.2.2 Materiais de Métodos ...................................................................................42 3.2.3 Resultados .....................................................................................................43 3.2.4 Discussão .....................................................................................................45 .3.2.5 Conclusão .....................................................................................................47 3.2.6 Referências Bibliográficas ............................................................................48
4 CONSIDERAÇÕES GERAIS .................................................................................... 50
5 REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................... 51
ANEXOS ....................................................................................................................... 57

LISTA DE TABELAS
ARTIGO CIENTÍFICO I
Tabela 01 - Médias, desvios-padrão, valores máximo e mínimo das variáveis do eritrograma e contagem de trombócitos de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e de cativeiro (n = 5). Bahia - 2006 .........
29
Tabela 02 - Médias, desvios-padrão, valores máximo e mínimo das variáveis do leucograma de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e de cativeiro (n = 5). Bahia - 2006 ...................................................
30
Tabela 03 - Médias, desvios-padrão, valores máximo e mínimo dos parâmetros da bioquímica sérica de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e em cativeiro (n = 5). Bahia - 2006 ..........................................
30
ARTIGO CIENTÍFICO II
Tabela 01 - Médias, desvios-padrão, valores máximo e mínimo de proteína total, das frações albumina, alfa-1 e alfa-2 globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=20), de cativeiro (n=5) e de vida livre e cativeiro juntas (n=25). Bahia - 2006.............................................................................................
45
ANEXOS
Tabela 01 - Valores individuais do comprimento curvilíneo de carapaça (CCC), variáveis do eritrograma e contagem de trombócitos de tartarugas marinhas (Caretta caretta) de vida livre (n = 22). Bahia - 2006 ............
59
Tabela 02 - Valores individuais do comprimento curvilíneo de carapaça (CCC), variáveis do eritrograma e contagem de trombócitos de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia - 2006 ..............
60
Tabela 03 - Valores individuais das variáveis do leucograma de tartarugas marinhas (Caretta caretta) de vida livre (n = 22). Bahia - 2006 ............
61
Tabela 04 - Valores individuais das variáveis do leucograma de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia - 2006 ..............
62

Tabela 05 - Valores individuais de parâmetros da bioquímica sérica de tartarugas
marinhas (Caretta caretta) de vida livre (n = 22). Bahia - 2006 ............
63
Tabela 06 - Valores individuais de parâmetros da bioquímica sérica de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia - 2006 ..............
64
Tabela 07 - Valores individuais de proteína total, das frações albumina, alfa-1 e alfa-2 globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=18). Bahia - 2006 ........................................................................................................
65
Tabela 08 - Valores individuais de proteína total, das frações albumina, alfa globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta), de vida livre (n=2). Bahia - 2006 ...............
66
Tabela 09 - Valores individuais de proteína total, das frações albumina, alfa-1 e alfa-2 globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia - 2006 ........................................................................................................
66

LISTA DE FIGURAS
ARTIGO CIENTÍFICO II
Figura 01 - Representação do eletroforetograma das proteínas séricas de tartarugas cabeçudas (Caretta caretta). A – Albumina, B – Alfa-1 globulina, C – Alfa- 2 globulina, D – Beta globulina, e E – Gama globulina. ................................................................................................
44
ANEXOS
Figura 01 - Exemplar de tartaruga cabeçuda (Caretta caretta) ................................
57
Figura 02 - Coleta de sangue no seio cervical dorsal de tartaruga marinha ..............
58

LISTA DE ABREVIATURAS
AST – Aspartato Aminotransferase
CCC – Comprimento Curvilíneo de Carapaça
CHGM – Concentração de Hemoglobina Globular Média
EDTA – Ácido etileno diamino tetracético
EMV – Escola de Medicina Veterinária
IBAMA – Instituto Brasileiro de Meio Ambiente
IUCN – World Conservation Union
Relação A:G – Relação Albumina:Globulina
SPSS – Statistical Package for the Social Sciences
UFBA – Universidade Federal da Bahia
VG – Volume Globular
VGM – Volume Globular Médio

PIRES, T.T. Avaliação do hemograma, bioquímica sérica e perfil eletroforético de
proteínas de tartarugas marinhas Caretta caretta (Linnaeus,1758) de vida livre e
em cativeiro. Salvador, Bahia, 2007. 66p. Dissertação (Mestrado em Ciência
Animal nos Trópicos) – Escola de Medicina Veterinária, Universidade Federal da
Bahia, 2007.
Resumo A obtenção de valores hematológicos é de grande importância na clínica médica para o
diagnóstico e tratamento das enfermidades que acometem tartarugas marinhas, por se
tratar de espécies vulneráveis à extinção. Com o objetivo de obter dados que auxiliem
na rotina clínica destes animais, avaliou-se o hemograma, alguns parâmetros
bioquímicos e eletroforese de proteínas de tartarugas cabeçudas (Caretta caretta),
fêmeas, adultas e clinicamente sadias. Os indivíduos foram divididos em dois grupos:
G1 de vida livre (n=22), que desovam no litoral norte da Bahia, e G2 de cativeiro (n=5)
mantidos no Centro de Visitantes do Projeto Tamar-Ibama, na Praia do Forte, município
de Mata de São João, Bahia. Os achados hematológicos incluíram a contagem de
eritrócitos, dosagem da concentração de hemoglobina, determinação do volume
globular, contagens total e diferencial de leucócitos, índices hematimétricos (VGM e
CHGM) e avaliação microscópica de esfregaços sangüíneos. Para a avaliação da
bioquímica sérica determinou-se a proteína total, albumina, glicose, colesterol,
triglicerídeos, ácido úrico, creatinina, aspartado aminotransferase e fosfatase alcalina,
por meio de kits comerciais, e corrida eletroforética de proteínas séricas em gel de
agarose, com estabelecimento da relação A:G. Os valores obtidos para os dois grupos
foram comparados, sendo que a análise estatística revelou diferenças significativas ao
nível de 5% para as contagens relativas de eosinófilos e monócitos, contagem absoluta
de eosinófilo, proteína total, globulina, glicose, colesterol, e aspartato aminotransferase
e fração gama globulina na eletroforese. Conclui-se no geral que não houve diferença
entre os grupos estudados, apesar de alguns parâmetros mostrarem alterações,
possivelmente pelo número reduzido de animais, condições ambientais, nutricionais
e/ou reprodutivos, a que estes animais estavam submetidos.
Palavras-chave: hemograma, bioquímica sérica, eletroforese, tartaruga cabeçuda.

PIRES, T.T. Evaluation of hemogram, serum biochemistry and protein
electrophoretic profile for captive and free-ranging sea turtles Caretta caretta
(Linnaeus,1758). Salvador, Bahia, 2007. 66p. Dissertation (Master of Science in
Tropical Veterinary Medicine) – School of Veterinary Medicine, Federal
University of Bahia, 2007.
SUMMARY
The study of hematological values has been very important in the clinical work for the
diagnostic and treatment of diseases that affect sea turtles, especially because these
species are endangered of extinction. The objective of this study was to obtain
information to be used in the clinical evaluation of sea turtles. It was analyzed the
hemogram, biochemical and protein electrophoresis parameters for loggerhead sea
turtles (Caretta caretta), female, adult and clinically healthy. The animals were
separated in two groups: G1 the free-ranging (n=22), which nest in the north coast of
Bahia, and G2 from captivity (n=5) which are kept in the visitor center of the Projeto
Tamar-Ibama, in Praia do Forte, municipality of Mata de São João, Bahia, Brazil. The
hematological findings included the red blood cell count, hemoglobin concentration,
packed cell volume, white blood cell count and differential count, red blood cell indices
(VGM and CHGM) and blood smear microscopic analysis. The biochemical evaluation
was based on the total protein, albumin, glucose, cholesterol, triglycerides, uric acid,
creatinine, aspatate aminotransferase and alkaline phosphatase, using commercial kits,
and serum protein electrophoresis in agarosis gel, with the determination of the A:G
ratio. Statistical comparisons between the two groups exhibit significant differences at
5% level for the relative eosinophil and monocyte count, absolute eosinophil count,
total protein, globulin, glucose, cholesterol, aspatate aminotransferase and gamma
globulin fraction from the electrophoresis analysis. Although it had been observed
variation in a few parameters, probably related to the reduced number of animals or in
the environmental, nutritional and reproductive conditions, there was no difference
between the two groups.
Keywords: hemogram, serum biochemistry, electrophoresis, loggerhead sea turtles

FICHA CATALOGRÁFICA
PIRES, Thaís Torres Avaliação do hemograma, bioquímica sérica e perfil eletroforético de proteínas de tartarugas marinhas Caretta caretta (Linnaeus,1758) de vida livre e em cativeiro. Thaís Torres Pires – Salvador, 2007. 66p. Dissertação (Mestrado em Ciência Animal nos Trópicos) – Escola de Medicina Veterinária da Universidade Federal da Bahia, 2007.
Professor Orientador: José Eugênio Guimarães Palavras-chaves: hemograma, bioquímica sérica, eletroforese, tartaruga cabeçuda.
1. Autor 2. Assunto-Dissertação 3. Assunto-Dissertação I. Título

1 INTRODUÇÃO GERAL
As tartarugas marinhas existem há mais de 150 milhões de anos e conseguiram
sobreviver a todas as mudanças do planeta. Sua origem foi na terra e durante a evolução
desenvolveram características de adaptação ao ambiente marinho, como a diminuição
do número de vértebras e a fusão das que restaram às costelas, formando uma carapaça
resistente, embora leve. Perderam os dentes, ganharam um tipo de bico córneo e suas
patas se transformaram em nadadeiras.
Existem sete espécies de tartarugas marinhas, agrupadas em duas famílias
distintas: Dermochelyidae, com uma única espécie, Dermochelys coriacea (tartaruga de
couro), e Cheloniidae, com cinco gêneros e seis espécies: Chelonia mydas (tartaruga
verde), Caretta caretta (tartaruga cabeçuda), Eretmochelys imbricata (tartaruga de
pente), Lepidochelys olivacea (tartaruga oliva), Lepidochelys kempii (kemps ridley) e
Natator depressa (Australian flatback) (DAVENPORT, 1997; SOSLAU et al., 2005).
No Brasil ocorrem cinco espécies de tartaruga: Caretta caretta, Eretmochelys
imbricata, Lepidochelys olivacea, Chelonia mydas e Dermochelys coriacea, que desde
1986 são protegidas pela legislação brasileira (MARCOVALDI & MARCOVALDI,
1999; BAPTISTOTTE et al., 2003). Essas espécies são classificadas pelo IUCN (World
Conservation Union) como ameaçadas ou criticamente ameaçadas de extinção. Durante
muito tempo o consumo da carne e ovos de tartarugas marinhas, e a utilização de sua
carapaça na produção de adornos, contribuíram para o declínio de suas populações,
além de fatores como a degradação de seu habitat e a interação com a atividade
pesqueira (TROËNG & DREWS, 2004).
Devido à pressão internacional para criação de um programa de conservação
para as tartarugas marinhas no Brasil, já que são animais que realizam grandes
migrações, visitando águas de diferentes países, em 1980 o governo brasileiro
estabeleceu o Programa Nacional de Conservação das Tartarugas Marinhas (Projeto
Tamar), vinculado ao Instituto Brasileiro de Meio Ambiente (IBAMA). Atualmente, o
Projeto Tamar possui 22 bases de conservação e pesquisa, em praias do continente e

ilhas oceânicas, monitorando cerca de 1100 km de praias (MARCOVALDI &
MARCOVALDI, 1999; BAPTISTOTTE et al., 2003).
O litoral norte da Bahia caracteriza-se por ser uma das principais áreas
remanescentes de desova de Caretta caretta e Eretmochelys imbricata (D’AMATO &
MARCOVALDI, 1997), possuindo grande importância biológica para as tartarugas
marinhas, pois grande parte das desovas registradas na costa brasileira ocorre em praias
baianas. O projeto Tamar possui quatro bases e uma sub-base nesta área, protegendo
cerca de 200km de praia (MARCOVALDI et al., 1999), destacando-se as bases de Praia
do Forte e de Arembepe.
A base de pesquisa e conservação da Praia do Forte, município de Mata de São
João na Bahia, foi uma das três primeiras fundadas em 1982 (Comboios, no Espírito
Santo; e Pirambu, em Sergipe, foram as outras bases) e é a praia continental com maior
densidade de desova de tartaruga marinha do Brasil (MARCOVALDI & LAURENT,
1996). A área monitorada por esta base é visitada principalmente pela tartaruga
cabeçuda (Caretta caretta), ocorrendo também desovas de tartaruga de pente
(Eretmochelys imbricata), oliva (Lepidochelys olivacea) e verde (Chelonia mydas) em
ordem decrescente de ocorrência (SANTOS et al., 2000). A Praia do Forte conta com
um centro de visitação, onde a comunidade local e turistas podem conhecer o trabalho
de conservação destas espécies (MARCOVALDI & LAURENT, 1996). Nestes centros
de visitação são mantidas tartarugas marinhas em diferentes estágios de maturidade, em
condições saudáveis, com a finalidade de educação ambiental (MARCOVALDI &
MARCOVALDI, 1999).
A base de Arembepe atualmente monitora uma área de 47 km de praia, inserida
em três municípios: Salvador, Lauro de Freitas e Camaçari, e possui grande importância
por abranger áreas com grande aglomerado urbano e fluxo turístico intenso. Localizada
à 30Km da Base de Praia do Forte, conta com um Centro de Visitantes, tendo a
tartaruga cabeçuda como a espécie que mais desova nesta área.
A tartaruga cabeçuda pertence à família Cheloniidae e é diferenciada das outras
espécies de tartarugas marinhas por possuir a cabeça proporcionalmente larga, com dois
pares de escudos prefrontais, e o bico muito forte. Sua carapaça possui cinco pares de

escudos laterais (MARQUEZ, 1990). Esta espécie apresenta maior índice de desova nas
praias do continente, no Brasil (MARCOVALDI & MARCOVALDI, 1999;
BAPTISTOTTE et al., 2003), contribuindo com 72 a 89% dos ninhos durante cada
estação reprodutiva na Praia do Forte, sendo que o pico de desova da tartaruga cabeçuda
ocorre entre outubro e dezembro (MARCOVALDI & LAURENT, 1996).
Cada espécie de tartaruga marinha possui um regime alimentar, sendo esses
animais bastante eficientes no aproveitamento de sua dieta especializada, a qual se
encontra diretamente relacionada à flora bacteriana de seu sistema digestório. A forma
do bico indica quais os itens da dieta de cada espécie, e na Caretta caretta ele é
extremamente forte servindo para triturar conchas, carapaças de moluscos e crustáceos;
sua alimentação ainda inclui peixes, camarões, caramujos e algas (FRAIZER, 2001).
Como resultado do trabalho que está sendo desenvolvido no Projeto Tamar, a
matança de fêmeas e a coleta de ovos de tartarugas marinhas foram drasticamente
reduzidas (MARCOVALDI & MARCOVALDI,1999), mas a captura incidental, por
diversas artes de pesca, juntamente com o crescimento urbano desordenado, constituem
ameaças atuais às populações destas espécies.
Recentemente as tartarugas marinhas têm adquirido uma importância para fins
de não consumo, como objeto para o turismo, atividades educacionais e pesquisas
científicas, gerando oportunidades de empregos, assim como outros ganhos
econômicos. Outra consideração menos aparente, mas de valor insubstituível, é como
recurso ecológico. Estes répteis são componentes singulares de sistemas ecológicos
complexos, cuja vitalidade se inter-relaciona com outros recursos exploráveis, como
peixes, moluscos e áreas de mangue. Devido às migrações que realizam de milhares de
quilômetros e da maturidade sexual tardia, as tartarugas servem como importantes
indicadores da saúde dos ambientes costeiros e marinhos, tanto em escala local como
mundial (FRAZIER, 1999).
É fundamental compreender que a condição ambiental está intimamente
relacionada com a qualidade de vida das comunidades humanas, e em muitos casos, a
conservação com base na comunidade se considera parte do processo de
desenvolvimento desta mesma comunidade, sendo indispensável à conservação de

recursos uma visão realista e de longo prazo (FRAZIER, 1999). O uso de não consumo,
como o turismo de observação de tartarugas marinhas, vem ganhando popularidade
mundial. Estudos sugerem que este tipo de exploração pode render muito mais, tendo ao
mesmo tempo menos impacto nas populações destes indivíduos que o uso de consumo,
caracterizado pela utilização da carne, ovos e carapaças (TROËNG & DREWS, 2004).
Animais mantidos em cativeiro proporcionam uma oportunidade única para o
estudo de alguns aspectos da biologia das tartarugas marinhas, permitindo uma
manipulação e experimentação que não é possível no habitat natural desses indivíduos.
Entretanto, estão sujeitas a uma ampla variedade de patógenos e parasitas, que em
condições naturais e com densidades populacionais silvestres, podem ter conseqüências
imperceptíveis, mas em cativeiro, onde estes animais encontram-se confinados, as
enfermidades podem causar grandes danos (ROSS, 2000). Assim como em aves e
mamíferos, a avaliação do estado de saúde e diagnóstico de doenças em tartarugas
marinhas requer amostras sangüíneas para investigação laboratorial (JACOBSON,
1998).
A avaliação hematológica é de grande valor na investigação de doenças em
répteis, assim como possivelmente pode mostrar alterações fisiológicas entre animais de
vida livre e em adaptação ao cativeiro (DIVERS et al., 1996). Contudo, são poucas as
informações de valores sanguíneos para as tartarugas marinhas. A maior parte dos
estudos apresenta um número reduzido de parâmetros bioquímicos baseados em uma
pequena quantidade de amostras de animais em cativeiro ou de histórico desconhecido.
Tartarugas mantidas em cativeiro podem apresentar diferenças significativas em seus
valores hematológicos, resultantes principalmente da dieta artificial ou de condições de
estresse, não sendo desta forma, recomendada a utilização destes valores para tartarugas
de vida livre (BOLTEN & BJORNDAL, 1992).
O estabelecimento do perfil bioquímico sangüíneo para populações selvagens,
saudáveis, de espécies de tartarugas marinhas ameaçadas de extinção é de grande
importância. A avaliação do estado fisiológico destas populações se faz necessário para
o desenvolvimento apropriado de planos de manejo e conservação. Para auxiliar na
avaliação de causas de doenças e estimar populações que estão potencialmente em risco,
são necessários estudos de grupos naturais de tartarugas marinhas que não sofram

impactos negativos por poluição ou degradação de habitat (BOLTEN & BJORNDAL,
1992).
Para qualquer animal o conhecimento do padrão sangüíneo normal é importante,
já que as mudanças na química sangüínea podem estar relacionadas com o estado
fisiológico, podendo ainda ser usado na identificação de condições patológicas. Com
exceção da tartaruga marinha, existem muitos estudos e revisões sobre as mudanças
sazonais na química sangüínea de répteis (LUTZ & DUNBAR-COOPER, 1987). Além
da influência da sazonalidade, dados avaliados indicam que fatores como idade,
tamanho, sexo, estação do ano, saúde, habitat e dieta podem afetar os parâmetros
hematológicos (WOOD & EBANKS, 1984). Embora alguns trabalhos sejam publicados
sobre valores hematológicos de tartarugas, é difícil o uso dessas informações em
estudos comparativos, por causa dos métodos de coleta das amostras, manipulação,
processamento e análise bioquímica, que variam muito entre esses trabalhos (BOLTEN
et al., 1992). Outra dificuldade bastante comum diz respeito à identificação e
descrições das características morfológicas de células sangüíneas de quelônios marinhos
(WORK et al., 1998).
A eletroforese de proteínas plasmáticas pode ser utilizada como uma importante
ferramenta de avaliação da saúde de tartarugas marinhas. Entretanto, é necessário o
estabelecimento de intervalos de referência dos valores normais (GICKING et al.,
2004). Mudanças nas frações protéicas são evidentes em diversas doenças e podem
auxiliar no diagnóstico quando outros testes não são conclusivos. Esta técnica
proporciona informações importantes sobre vários componentes de proteínas do soro ou
plasma, os quais auxiliam na redução do número de possibilidade de diagnóstico
diferencial na maioria dos casos. Aliado a isso, a eletroforese de proteínas requer
pequena quantidade de sangue do espécime, e é um teste diagnóstico que ainda precisa
ser explorado na clínica de répteis e anfíbios (ZAIS & CRAY, 2002).
Este trabalho propõe a realização do hemograma, bioquímica sérica e a
determinação do perfil eletroforético das proteínas séricas em tartarugas marinhas
cabeçudas de vida livre e em cativeiro, observando as possíveis diferenças entre esses
grupos, permitindo assim que os valores obtidos possam ser utilizados como método
auxiliar para o diagnóstico clínico de enfermidades que acometem esses animais,

avaliação da terapia e monitoramento do processo que o acomete. Ressalta-se aqui a
importância desses parâmetros no tratamento de animais de vida livre, por se tratar de
uma das espécies de tartarugas marinhas vulnerável à extinção, a reintrodução na
natureza desses indivíduos comprovadamente saudáveis, através dessas avaliações, é de
grande necessidade para os projetos de conservação. Portanto, torna-se imprescindível a
realização de trabalhos que visem a determinação do padrão hematológico dessa espécie
no Brasil, especificamente na Bahia, considerando todos os fatores que os influenciam.

2 REVISÃO DE LITERATURA
2.1 Exames complementares em tartarugas marinhas
2.1.1 Importância dos exames
Algumas características anatômicas e fisiológicas das tartarugas marinhas
dificultam o exame físico. A carapaça e plastrão ósseos limitam a auscultação, palpação
e a realização de exames como ultra-sonografia e radiografias. Por serem animais
pecilotérmicos necessitam de temperaturas ambiente confortáveis para manutenção das
funções fisiológicas normais (PAGE & MAUTINO, 1990; WHITAKER & KRUM,
1999). Os exames hematológicos e bioquímicos podem ser realizados através de
simples colheitas de amostras e permitem a avaliação de informações importantes sobre
o estado de saúde das tartarugas marinhas (HERBST & JACOBSON, 2003).
2.1.2 Colheita sangüínea
O volume sangüíneo total varia entre as espécies, geralmente corresponde a
cerca de 5 a 8% do peso corporal. Assim, uma tartaruga com 100g tem um volume
sangüíneo estimado em 5 a 8ml e, estando clinicamente saudável pode-se coletar 10%
deste volume sem conseqüências prejudiciais a sua saúde (JACOBSON, 1998).
Os locais de escolha incluem o coração, veia jugular, veia braquial, veia ventral
coccígena, seio orbital, borda da unha e seio cervical dorsal (OWENS & RUIZ, 1980;
ROSSKOPF, 1982; JACOBSON, 1998) e, com base na literatura, a punção cardíaca e
venipunção do seio cervical dorsal são os métodos mais utilizados (WOOD &
EBANKS, 1984; LUTZ & DUNBARCOOPER, 1987; WORK et al., 1998).
O método de coleta do seio cervical dorsal é considerado superior aos outros
procedimentos para colheita de sangue em tartarugas marinhas, já que reduz o estresse e
o risco de danos permanentes, requer o mínimo de equipamento, é de fácil realização e

permite a obtenção de amostras sem contaminações. O seu sucesso depende de uma
adequada contenção, que consiste em manter o animal em posição inclinada, com a
região anterior mais baixa e o pescoço estirado (OWENS & RUIZ, 1980).
Diferenças significativas podem ser encontradas quando se compara os valores
hematológicos e bioquímicos obtidos de amostras sangüíneas de jabutis do deserto
(Gopherus agassizii) através da punção do plexo venoso postocciptal e veia jugular.
Essa diferença é justificada pela contaminação por fluido extracelular e pela linfa,
levando a valores inferiores de hematócrito, contagens de eritrócitos e leucócitos, e
concentração de hemoglobina. Os répteis, incluindo os quelônios, possuem um sistema
linfático amplamente desenvolvido, com vasos adjacentes ao sistema venoso. A
contaminação também afeta os valores da bioquímica sangüínea, isto é, a atividade
enzimática e a concentração de proteínas totais, albumina e globulina. Assim, a
hemodiluição com linfa promove valores mais baixos dessas proteínas, fosfatase
alcalina, aspartato aminotransferase e alanina aminotransferase (GOTTDENKER &
JACOBSON, 1995).
MARKS & CITINO (1990) ao trabalharem com tartarugas da espécie Testudo
radiata também observaram diferenças nos valores hematológicos relacionados ao local
da coleta sangüínea, onde amostras obtidas da veia coccígena apresentaram
hemodiluição. Entretanto, WOOD & EBANKS (1984) não encontraram diferença
significativa associada ao método de coleta das amostras em trabalho com tartarugas
verdes (Chelonia mydas), onde o sangue foi coletado através de punção cardíaca, do
seio cervical dorsal e do fluxo sangüíneo livre após o corte do pescoço dos animais
sacrificados, sendo que o tamanho do animal determinou o método de coleta de sangue.
SAMOUR e colaboradores (1998) relataram que em algumas colheitas sangüíneas em
tartarugas verdes realizadas através da venipunção da jugular, observou-se um fluido
transparente na seringa, porém quando a colheita foi realizada no seio cervical dorsal
esta contaminação não ocorreu.

2.1.3 Anticoagulantes
A conservação adequada das amostras sangüíneas é de grande importância para
se obter resultados confiáveis. Em avaliações hematológicas de quelônios a heparina é o
anticoagulante de escolha, já que o EDTA quela o cálcio e outros íons metálicos,
causando hemólise extensa em sangue de répteis (PAGE & MAUTINO, 1990;
BOLTEN et al., 1992; KNOTKOVÁ et al., 2002). A heparina impede a coagulação
sangüínea por interferir na conversão da protombina em trombina, sendo eficaz por um
período de 10 a 12 horas (COLES, 1984).
2.1.4 Eritrograma
2.1.4.1 Volume Globular
A dieta, estado de saúde, exercício, hormônio circulante, temperatura e estresse
podem afetar o volume globular (VG) de tartarugas mantidas em cativeiro, além de
algumas doenças como infecções do plastrão, hemorragias prolongadas, deficiências
nutricionais ou inanição, podem ser relacionadas à sua diminuição, atingindo nestes
casos, valores abaixo de 20% (FRAIR, 1977a).
FRAIR (1977b) mostrou haver uma correlação significativa entre o tamanho
corporal e o valor do hematócrito em tartarugas marinhas. Contudo, BOLTEN &
BJORNDAL (1992) relatam que o VG de tartarugas verdes não apresentou relação
significativa entre sexo ou tamanho, sendo obtido valor médio de 35,2% (± 3,2). Tal
fato também foi constatado por WOOD & EBANKS (1984) que trabalharam com a
mesma espécie, verificando que não houve diferença significativa entre sexos para
valores desta variável, e que os mesmos parecem aumentar com a idade. A média
encontrada por estes autores foi de 45% (± 7) para 13 animais adultos.
LUTZ & DUNBARCOOPER (1987) em estudo com tartarugas marinhas
cabeçudas, encontraram valores constantes de hematócrito, os quais não sofreram
influência da estação do ano. A variação para este parâmetro foi de 28 a 48%, tendo
como média 35,4%. Entretanto, SOUTHWOOD e colaboradores (2003) observaram

valores mais elevados no VG de Chelonia mydas durante condições de baixas
temperaturas, obtendo valores de 33,6% (± 0,5) a 26ºC, e 40,0% (± 1,3) a 16ºC,
justificando que nestes períodos há uma diminuição do metabolismo, assim um VG
mais alto poderia ser uma maneira de manter a capacidade de transporte de oxigênio
para os tecidos durante os períodos de bradicardia.
Alguns autores obtiveram valores do hematócrito de tartarugas cabeçudas
mantidas em cativeiro: BALDASSIN (2003) encontrou valor de 22% (± 2,9), PIRES e
colaboradores (2006) 33,12% (± 2,35) e CUBAS & BAPTISTOTTE (2006) de 29,5%
(± 7,3). Para 48 juvenis de vida livre desta espécie, o valor encontrado por KELLER e
colaboradores (2004) foi de 31,5% (± 4,3).
2.1.4.2 Morfologia e Contagem de Hemácias
A morfologia dos eritrócitos de tartaruga marinha é similar às de outros
quelônios. São células ovais, com citoplasma homogêneo circundando completamente o
núcleo, que se apresenta em posição central e é fortemente basofílico (WOOD &
EBANKS, 1984; ALLEMAN et al.,1992; CASAL & ORÓS, 2007).
O número de hemácias foi alvo de estudo por parte de WOOD & EBANKS,
(1984) que trabalharam com a espécie Chelonia mydas, obtendo um valor médio para
esta variável de 341.000/µL de sangue. Já, WHITAKER & KRUM (1999) trabalhando
com a espécie Caretta caretta, com menos de 2 Kg, e mantidas em cativeiro, obtiveram
valores médios de 390.000/µL, enquanto que a média, encontrada por PIRES e
colaboradores (2006), para oito indivíduos desta espécie, foi de 275.000/µL. Valores
semelhantes a estes foram apresentados por BALDASSIN (2003) e CUBAS &
BAPTISTOTTE (2006) que obtiveram valores de 254.000 (n=7) e 288.000 (n=13) de
hemácias por microlitro de sangue, respectivamente, trabalhando com tartarugas da
mesma espécie em cativeiro.
Com relação à análise morfológica do esfregaço sangüíneo de tartarugas,
alterações como anisocitose, poiquilocitose e hipocromia podem ser observadas em
anemias regenerativas e por deficiência de ferro ou perda crônica de sangue

(CAMPBELL,1996), enquanto que a policromasia foi observada em tartarugas
cabeçudas expostas a petróleo bruto (LUTCAVAGE et al., 1995).
2.1.4.3 Concentração de Hemoglobina
A concentração de hemoglobina aparentemente aumenta com a idade e/ou
tamanho do animal, mas não apresenta diferença significativa entre sexos (WOOD &
EBANKS,1984), porém segundo ALKYNDI & MAHMOUD (2002) permanece estável
durante a temporada reprodutiva, apesar do exaustivo esforço físico realizado pelas
tartarugas durante as diferentes fases reprodutivas.
AGUIRRE e colaboradores (1995), ao trabalharem com cinco tartarugas verdes,
encontraram valor médio de 8,54g/dL (± 1,44) para a concentração de hemoglobina. Em
tartarugas cabeçudas ALKINDI & MAHMOUD (2002), BALDASSIN (2003), PIRES e
colaboradores (2006) e CUBAS & BAPTISTOTTE (2006), obtiveram valores médios
de 8,87 g/dl (± 1,41), 6,10 g/dl (± 2,40), 8,65 g/dl (± 0,80) e 8,60 g/dl (± 2,00) para esta
variável, respectivamente.
2.1.4.4 Índices Hematimétricos
AGUIRRE e colaboradores (1995) obtiveram valores médios para volume
globular médio (VGM), hemoglobina globular média (HGM) e concentração de
hemoglobina globular média (CHGM) de 725,00fL (± 312,35), 198,18pg (± 83,52) e
27,50% (± 1,25) respectivamente, em estudo com Chelonia mydas. Em trabalhos com
tartarugas Caretta caretta, PIRES e colaboradores (2006) apresentaram valores de
VGM de 1.214,36fL (± 131,99), HGM de 317,12pg (± 38,66) e CHGM de 26,10% (±
1,21), enquanto que para CUBAS & BAPTISTOTTE (2006) esses valores foram
1.106,20fL (± 413,10), 315,90pg (± 93,40) e 29,5% (± 4,90), respectivamente.

2.1.5 Leucograma
2.1.5.1. Contagem de Leucócitos
Assim como os mamíferos, os glóbulos brancos dos répteis são classificados em
granulócitos e agranulócitos, entretanto descrições morfológicas das características
destas células em quelônios marinhos são limitadas. Mesmo quando métodos
consistentes são utilizados, a contagem total de leucócitos em répteis pode ser altamente
variável, mas são bastante úteis em estudos comparativos ou quando se monitora a
evolução do quadro clínico (WORK et al., 1998).
O número total de leucócitos foi objeto de estudo para tartarugas juvenis da
espécie Chelonia mydas por AGUIRRE e colaboradores (1995), que obtiveram o valor
de 9.340/µL, enquanto que WORK e colaboradores (1998) e WORK & BALAZS
(1999) encontraram médias de 13.000/µL (n=120) e 11.890/µL (n=28),
respectivamente, e KELLER e colaboradores (2004) trabalhando com a espécie Caretta
caretta obtiveram valor de 14.800/µL. Para adultos desta espécie, PIRES e
colaboradores (2006) encontraram 3.656 leucócitos por microlitro de sangue, valor
próximo ao encontrado por CUBAS & BAPTISTOTTE (2006), de 4.249/µL.
Condições de cativeiro podem causar elevação nos valores da contagem total de
leucócitos, devido ao estado nutricional ou de estresse dos animais (WOOD &
EBANKS, 1984). Tartarugas marinhas expostas à contaminação por petróleo
apresentaram um aumento de até quatro vezes da contagem destas células
(LUTCAVAGE et al., 1995).
2.1.5.2 Contagem Diferencial de Leucócitos
WORK e colaboradores (1998) e CASAL & ORÓS (2007), utilizando-se de
técnicas citoquímicas, identificaram na corrente sangüínea de tartarugas verdes e
cabeçudas os seguintes leucócitos: linfócitos, monócitos, heterófilos, eosinófilos e
basófilos. As células mononucleares circulantes como linfócitos e monócitos são
similares entre os vertebrados superiores, mas é considerável a heterogeneidade da série

granulocítica. Diferentemente dos animais domésticos, o heterófilo é um granulócito
encontrado em répteis e aves e corresponde ao neutrófilo dos mamíferos. Outra
terminologia usada é acidófilo que pode ser aplicado para os heterófilos e eosinófilos, já
que estas células apresentam em seu citoplasma grânulos eosinofílicos (MONTALI,
1988).
Alguns pesquisadores ainda divergem sobre a terminologia e critérios para
distinguir os granulócitos acidofílicos (HARMS et al., 2000). Os neutrófilos são raros
em répteis, mas têm sido descritos em estudos com tartarugas cabeçudas (GEORGE,
1997) e verdes (WOOD & EBANKS, 1984; AGUIRRE et al. 1995). Entretanto, alguns
destes autores não utilizaram a citoquímica ou outra técnica mais específica para
confirmar e identificar os diferentes tipos de leucócitos (CASAL & ORÓS, 2007). A
descrição de azurófilos é rara em tartarugas marinhas, mas tem-se relatado este tipo
celular em poucos trabalhos (SAMOUR et al., 1998; KELLER et al, 2004), embora
outros não tenham identificado estas células (WOOD & EBANKS, 1984; CANNON,
1992; AGUIRRE et al., 1995; WORK et al., 1998; WORK & BALAZS, 1999; HARMS
et al., 2000; CASAL & ORÓS, 2007).
Os linfócitos são células que apresentam grande variação quanto à forma e ao
tamanho, o seu citoplasma é escasso (WORK et al., 1998; SAMOUR et al., 1998) e o
núcleo segue em geral a forma da célula, apresentando cromatina nuclear fortemente
condensada quando maduros (WOOD & EBANKS, 1984; ALLEMAN et al.,1992;
CASAL & ORÓS, 2007). Alterações no número de linfócitos podem ser causadas por
estresse, neoplasia ou resposta inflamatória não específica (CRAY et al., 2001), ao
passo que o estresse e a imunossupressão em tartarugas marinhas com
fibropapilomatose podem provocar linfopenia (AGUIRRE et al., 1995; WORK &
BALAZS, 1999).
Os monócitos são as maiores células encontradas no esfregaço sangüíneo
(WORK et al., 1998; SAMOUR et al., 1998), são arredondadas ou irregulares, com
citoplasma não muito abundante, podendo apresentar vacúolos. O núcleo é excêntrico
com contorno irregular, e sua cromatina menos densa que a do linfócito (WOOD &
EBANKS, 1984; CASAL & ORÓS, 2007). A presença de monocitose sugere um
processo infeccioso e/ou inflamatório crônico, sendo a proliferação monocítica

associada à granulomas, enquanto que a ausência de monócitos, na contagem
diferencial, é considerada normal (AGUIRRE et al., 1995; WORK & BALAZS, 1999).
Os heterófilos são células com funções análogas às dos neutrófilos dos
mamíferos. É uma célula grande, arredondada e de formato irregular (WORK et al.,
1998). O núcleo do heterófilo maduro é esférico, não lobulado, e de localização
excêntrica no citoplasma; este por sua vez, é preenchido com numerosos grânulos
fusiformes eosinofílicos (MONTALI, 1988; ALLEMAN et al., 1992; SAMOUR et al.,
1998; CASAL & ORÓS, 2007). Heterofilias são observadas em tartarugas marinhas
com fibropapilomatose, condições de estresse (AGUIRRE et al., 1995; WORK &
BALAZS, 1999) e em inflamações agudas (ROSSKOPF, 1982).
Os eosinófilos são células esféricas, com quantidade abundante de citoplasma
contendo grânulos arredondados eosinofílicos, que coram-se mais fracamente que os
dos heterófilos (MONTALI, 1988; CANNON, 1992) O núcleo tem forma arredondada a
irregular, em posição excêntrica, (ALLEMAN et al.,1992; WORK et al., 1998;
SAMOUR et al., 1998; CASAL & ORÓS, 2007). Uma eosinofilia é esperada em casos
de estímulo parasitário (ROSSKOPF, 1982; CAMPBELL, 1996), enquanto que
eosinopenia pode ocorrer em situações de estresse (AGUIRRE et al., 1995). WORK &
BALAZS (1999) observaram eosinopenia em casos de infecções por trematódeos
vasculares em tartarugas verdes, o que pode estar associado ao seqüestro de eosinófilos
nos tecidos como uma resposta inflamatória a estes parasitas. Existe uma correlação
entre esta célula e a idade do animal, com diminuição do seu número e aumento de
heterófilos com o envelhecimento (WOOD & EBANKS, 1984).
O basófilo é uma célula pequena que apresenta núcleo denso, de posição central,
estando freqüentemente encoberto por grandes grânulos citoplasmáticos arredondados e
fortemente basofílicos (WOOD & EBANKS, 1984; ALLEMAN et al., 1992; WORK et
al., 1998; SAMOUR et al., 1998). Contudo, sua presença na circulação de animais
hígidos é rara, como relatam WOOD & EBANKS (1984), CANNON (1992) e WORK e
colaboradores (1998) que trabalharam com as espécies Lepidochelys kempi e Chelonia
mydas, respectivamente. A basofilia pode ocorrer em resposta à inflamações crônicas
(ROSSKOPF, 1982).

No que diz respeito aos valores relativos de leucócitos obtidos na contagem
diferencial, CUBAS & BAPTISTOTTE (2006) obtiveram para tartarugas cabeçudas
mantidas em cativeiro, os seguintes resultados: heterófilos 54% (± 13,1); linfócitos
17,5% (± 8,4); eosinófilos 13,6% (± 7,7); monócitos 12,5% (± 7,2); e basófilos 0,1% (±
0,3), correspondendo a valores absolutos de: heterófilos 2.170,3/µL (± 738,5); linfócitos
800,8/µL (± 602,6); eosinófilos 594,7/µL (± 477,6); monócitos 560,5/µL (± 422,9); e
basófilos 4,2/µL (± 15,2). PIRES e colaboradores (2006), também trabalhando com a
espécie Caretta caretta em cativeiro, apresentaram os seguintes valores relativos e
absolutos, respectivamente: heterófilos 59,4% (± 16,3) e 2.156,9/µL (± 703,5);
linfócitos 29,2% (± 17,1) e 1.110,9/µL (± 783,6); eosinófilos 10,4% (± 6,3) e 366,9/µL
(± 216,4); monócitos 0,9% (± 2,1) e 19,1/µL (± 42,34); e basófilos 0,1% (± 0,3) e
2,5/µL (± 7,1). Valores relativos ligeiramente diferentes de heterófilos 75,8% (± 9,1);
linfócitos 18,4% (± 7,1); eosinófilos 4,5% (± 6,3); monócitos 1,2% (± 1,4); e basófilos
0% (± 0) foram obtidos por CASAL & ORÓS (2007) para 35 tartarugas desta mesma
espécie.
Heterofilia, linfocitopenia e eosinopenia foram observadas em tartarugas verdes
24 horas após a captura. Essa mudança no leucograma foi atribuída ao estresse agudo de
captura, similar ao que ocorre em outros répteis. Já a linfocitopenia severa em tartarugas
marinhas com fibropapilomatose podem indicar uma supressão do sistema imunológico
destes animais (AGUIRRE et al., 1995; WORK & BALAZS, 1999).
2.1.6 Contagem de Trombócitos
Trombócitos são células nucleadas com função hemostática análoga às plaquetas
dos mamíferos (SOSLAU et al., 2005). São células elípticas ou arredondadas, possuem
núcleo oval, central e fortemente basofílico (WORK et al., 1998). Essas células
geralmente são encontradas aglutinadas em esfregaços sangüíneos (WOOD &
EBANKS, 1984; HARMS et al., 2000; CASAL & ORÓS, 2007), podendo ser
confundidas com pequenos linfócitos, sendo diferenciadas destes por apresentar
citoplasma incolor, e uma relação núcleo/citoplasma menor (ALLEMAN et al.,1992).

O valor obtido na contagem de trombócitos de tartarugas cabeçudas mantidas em
cativeiro, por PIRES e colaboradores (2006) foi de 10.968,1/µL, enquanto que CUBAS
& BAPTISTOTTE (2006) apresentaram uma média de 21.932,1/µL para a mesma
espécie.
2.1.7 Parâmetros Bioquímicos
Mudanças nos valores de parâmetros bioquímicos em tartarugas marinhas
podem indicar seu estado fisiológico ou condições patológicas. Esses valores podem ser
afetados por muitos fatores intrínsecos como: espécie, sexo e idade, e extrínsecos como
sazonalidade, temperatura, habitat e dieta (LUTZ & DUNBAR-COOPER, 1987;
AGUIRRE et al., 1995). Além de ser utilizado como meio diagnóstico de um exemplar
de determinada espécie, os dados bioquímicos podem auxiliar na avaliação do estado de
sanidade de populações (SWIMMER, 2000). Sendo assim, o estabelecimento do perfil
da bioquímica sangüínea de tartarugas marinhas de vida livre saudáveis é uma das
prioridades na tentativa de conservação destas espécies (AGUIRRE & BALAZS, 2000).
Entretanto ainda não se tem estudos aprofundados relacionando os parâmetros
bioquímicos as enfermidades em tartarugas marinhas, destacando-se somente a
fibropapilomatose. Esta é uma doença que apresenta alta prevalência entre as
populações de tartarugas verdes, e por este motivo, vem sendo alvo de investigações
quanto as alterações bioquímicas e hematológicas que produz nos indivíduos afetados
(WORK & BALAZS, 1999; SWIMMER, 2000; WORK et al., 2001; CRAY et al.,
2001).
2.1.7.1 Proteína Total, Albumina e Globulina
Há pelo menos uma centena de proteínas plasmáticas descritas e identificadas,
no homem e nos animais, com inúmeras funções no organismo. As principais e mais
conhecidas proteínas são a albumina, globulina e suas frações, e o fibrinogênio, cujos
valores normais variam de acordo com a idade, espécie animal e condições fisiológicas
(KANEKO, 1997). Em répteis os valores normais de proteína plasmática total

geralmente variam de 3 a 8 g/dL (CAMPBELL, 1996), e em estudos realizados com
tartarugas cabeçudas BOLTEN e colaboradores (1992 e 1994) obtiveram uma média de
4,1g/dL (± 1,2), enquanto que em Caretta caretta juvenis valor de 4,0g/dL (± 0,8) foi
obtido por KELLER e colaboradores (2004).
Em situações de cativeiro as tartarugas podem apresentar níveis protéicos mais
elevados como resultado da dieta rica em proteína oferecida (SWIMMER, 2000). Para
exemplares de tartarugas cabeçudas, mantidos nesta condição, a média encontrada por
CUBAS & BAPTISTOTTE (2006) foi de 4,2g/dL (± 0,4). Contudo, em tartarugas
verdes na Austrália, submetidas à baixas temperatura (17oC/16 semanas), simulando
condições de inverno, pode-se obter valores mais baixos de proteína total, o que se
justifica pelo menor consumo de alimento nestas condições, onde a taxa metabólica
encontra-se diminuída (SOUTHWOOD et al., 2003).
Para a albumina, em pesquisa realizada com tartarugas marinhas cabeçudas,
foram obtidos os seguintes valores: 0,9g/dL (± 0,3) em 23 amostras de plasma e 0,7g/dL
(± 0,2) em 168 amostras séricas (BOLTEN et al., 1992 e 1994), enquanto que 1,1g/dL
(± 0,2) e 0,7g/dL (± 0,2) foram apresentados por KELLER e colaboradores (2004) e
CUBAS & BAPTISTOTTE (2006) ao avaliarem plasma.
Quanto a globulina, valores de 3,2g/dL (± 0,9) e 2,9g/dL (± 0,7) foram obtidos
para tartarugas marinhas cabeçudas por BOLTEN e colaboradores (1994) e KELLER e
colaboradores (2004), respectivamente.
Com relação a alterações de valores sangüíneos, a hipo ou hiperproteinemia são
atribuídas principalmente a diminuição de albumina e/ou aumento de globulina. As
hipoalbuminemias em répteis estão associadas freqüentemente à má nutrição, podendo-
se incluir também má absorção e má digestão, geralmente associadas a parasitismo
intestinal, enteropatias, severa perda de sangue, e doenças hepáticas ou renais crônicas
(CAMPBELL, 1996). Em tartarugas marinhas Chelonia mydas com fibropapilomatose
relatou-se hipoproteinemia-hipoalbuminemia (NORTON et al., 1990; AGUIRRE et al.,
1995; AGUIRRE E BALAZS, 2000; CRAY et al., 2001). As hiperproteinemias
ocorrem com a hemoconcentração (desidratação) ou elevação das globulinas em
doenças inflamatórias crônicas (CAMPBELL, 1996).

2.1.7.2 Relação Albumina: Globulina (A:G)
A relação entre a albumina e globulina é de grande valor na clínica veterinária,
servindo de base na classificação do perfil eletroforético das proteínas, bem como na
natureza deste perfil. A relação A:G obtida em estudo com tartarugas cabeçudas de vida
livre foi de 0,3 (± 0,1) (BOLTEN et al., 1994), valor próximo ao encontrado por
KELLER e colaboradores (2004) de 0,4 (± 0,08), em trabalho com a mesma espécie.
Em trabalho realizado com a espécie Caretta caretta, KELLER e colaboradores
(2004) observaram uma diminuição da relação A:G, associada a concentrações de
contaminantes organoclorados no sangue e gordura, e CRAY e colaboradores (2001)
relatam também um decréscimo nessa relação em um grupo de Chelonia mydas com
fibropapilomatose, devido a uma diminuição na concentração de albumina e aumento na
de gamaglobulinas. Em trabalho com tartarugas verdes, severamente acometidas por
fibropapilomatose, constatou-se também diminuição da albumina e globulinas,
entretanto não houve diferença entre a relação A:G, quando comparada com animais
sem tumores ou aqueles levemente acometidos pela doença (WORK et al., 2001).
2.1.7.3 Glicose
A concentração sangüínea de glicose em répteis varia de acordo com a espécie
animal, o estado nutricional e condições ambientais, podendo ainda apresentar variação
sazonal em algumas espécies (CAMPBELL, 1996). Em 168 amostras plasmáticas de
tartarugas marinhas, um valor médio de 97mg/dL (± 21) foi obtido por BOLTEN e
colaboradores (1994), enquanto que valores de 109mg/dL (± 18) e 102,8mg/dL (± 17)
foram obtidos por KELLER e colaboradores (2004) e CUBAS & BAPTISTOTTE
(2006), respectivamente. Em outro estudo utilizando soro a média encontrada foi de
80,1mg/dL (± 22,3) (BOLTEN et al., 1992).
A hipoglicemia nestes animais pode ser resultado de má nutrição, dietas com
altos níveis protéicos, hepatopatias severas e endocrinopatias, enquanto que a
hiperglicemia (valores de glicose acima de 200mg/dL) raramente é documentada nestas
espécies, sendo forte indício de diabetes mellitus (CAMPBELL, 1996). Estudos relatam

hiperglicemia em resposta ao estresse agudo em tartarugas marinhas, assim como nos
mamíferos (AGUIRRE et al., 1995; GREGORY & SCHMID, 2001).
2.1.7.4 Ácido úrico
O ácido úrico é o produto final do catabolismo primário das proteínas, nitrogênio
não protéico e purinas em répteis, sendo grande parte do nitrogênio excretado desta
forma. A dieta pode influenciar os valores do ácido úrico sangüíneo, especialmente
aquelas ricas em proteínas e uréia (CAMPBELL, 1996). Alguns trabalhos apresentam
valores deste parâmetro para tartarugas cabeçudas de vida livre, BOLTEN e
colaboradores (1992) obtiveram uma média de 0,8mg/dL (±0,2), assim como KELLER
e colaboradores (2004) 0,8mg/dL (±0,7), enquanto que para indivíduos desta espécie
mantidos em cativeiro CUBAS & BAPTISTOTTE (2006) relatam valores de 1,0mg/dL
(±0,4).
Em estudo com juvenis de tartarugas verdes, BOLTEN & BJORNDAL (1992)
observaram diferença entre os valores de ácido úrico obtidos para machos e fêmeas,
onde os machos (n=38) apresentaram valores mais elevados, de 1,8mg/dL (±0,6) que as
fêmeas (n=60), de 1,4mg/dL (±0,5).
Tartarugas verdes mantidas em cativeiro apresentaram valores
significativamente mais elevados que animais de vida livre, devido a uma alimentação
exclusivamente protéica, entretanto animais cativos que recebem alimentação
balanceada apresentam níveis de ácido úrico similares àqueles encontrados em animais
de vida livre (SWIMMER, 2000).
Hiperuricemia pode ser associada à doença renal severa e gota. O uso de drogas
nefrotóxicas, como aminoglicosídeos e sulfa, pode resultar em necrose tubular renal,
causando aumento do ácido úrico plasmático (CAMPBELL, 1996).

2.1.7.5 Creatinina
A determinação da concentração de creatinina sérica pode ser pouco conclusiva
em doenças pré-renais e renais em répteis, uma vez que nestes animais o nitrogênio é
eliminado como ácido úrico, razão pela qual as tartarugas são considerados indivíduos
uricotélicos. Nestas espécies os valores normais de creatinina sérica são geralmente
muito baixos, inferiores a 1 mg/dL, e espera-se valores elevados na desidratação severa
e doenças renais (CAMPBELL, 1996)
BOLTEN e colaboradores (1994) encontraram uma média de 0,3mg/dL (± 0,1)
de creatinina no plasma de tartarugas Caretta caretta. Valor idêntico foi obtido em
estudo com amostras de soro desta espécie (BOLTEN et al., 1992). A determinação da
concentração de creatinina também foi objeto de estudos de KELLER e colaboradores
(2004) que apresentaram valor inferior a 0,1mg/dL, e de CUBAS & BAPTISTOTTE
(2006) que obtiveram média de 0,2mg/dL (± 0,1), ainda para a espécie cabeçuda.
2.1.7.6 Colesterol
Distúrbios do metabolismo lipídico podem ser mensurados através das alterações
no colesterol sérico. Algumas causas do aumento dos seus níveis incluem dietas ricas
em gorduras, lesões hepáticas e renais (MEYER et al., 1995). Tartarugas marinhas
mantidas em cativeiro podem apresentar níveis mais elevados de colesterol devido à
dietas ricas em gorduras (SWIMMER, 2000).
BOLTEN & BJORNAL (1992) relataram correlação positiva entre os níveis de
colesterol e o peso corporal, assim como concentrações mais elevadas em fêmeas que
em macho de tartaruga verde.
Para a espécie Caretta caretta de vida livre, BOLTEN e colaboradores (1994)
obtiveram uma média de 170mg/dL (± 89) de colesterol total em amostras de plasma,
enquanto que no soro de tartarugas cabeçudas este valor foi de 106mg/dL (± 57)
(BOLTEN et al., 1992). Por outro lado, em tartarugas cabeçudas cativas a média

apresentada foi de 107mg/dL (± 26,1) para esta mesma variável (CUBAS &
BAPTISTOTTE, 2006).
Valores baixos de colesterol sérico foram relatados associados a
fibropapilomatose severa em tartarugas verdes, devido a condição crônica debilitante da
enfermidade (AGUIRRE et al., 1995; AGUIRRE & BALAZS, 2000).
2.1.7.7 Triglicerídeos
Os valores plasmáticos médios de triglicerídeos apresentados em estudos com
tartarugas cabeçudas foram de 179mg/dL (± 68,5) (CUBAS & BAPTISTOTTE, 2006),
e 212mg/dL (± 334) (BOLTEN et al., 1994), enquanto que BOLTEN e colaboradores
(1992) ao avaliarem o soro de exemplares desta espécie obtiveram o valor de 81mg/dL
(± 204) e BALDASSIN (2003) de 587,2mg/dL (± 533,3) para animais cativos.
Em répteis mantidos em cativeiro, a causa mais comum de aumento nos níveis
de triglicerídeos é um defeito metabólico no transporte dos lipídios ou síntese de
proteínas, resultando em obesidade, causado por superalimentação. Assim, valores
elevados encontrados em tartarugas marinhas cativas são devido à dieta rica em
gorduras, exercício insuficiente ou ambos (SWIMMER, 2000).
Diferenças significativas nos níveis de triglicerídeos foram relatados, ao se
comparar populações de tartarugas verdes juvenis, clinicamente saudáveis e acometidas
por fibropapilomatose em ilhas do Hawaii (AGUIRRE & BALAZS, 2000), sendo que
este fato já tem sido constatado e com valores baixos em tartarugas marinhas com esta
alteração severa (AGUIRRE et al., 1995).
2.1.7.8 Fosfatase Alcalina
Poucos estudos são realizados a respeito das alterações enzimáticas em répteis.
Geralmente os valores obtidos são interpretados da mesma forma que para mamíferos e
aves domésticos. A fosfatase alcalina está presente em vários tecidos e a sua

determinação não é considerada um teste específico para determinado órgão. Esta
enzima pode estar aumentada na atividade osteoblástica ou doença hepatobiliar em
algumas espécies; animais mais jovens podem apresentar valores mais elevados que os
adultos (CAMPBELL, 1996). São reportados ainda elevação da fosfatase alcalina no
hiperparatireoidismo, doenças ósseas e insuficiência renal (SWIMMER, 2000).
BOLTEN e colaboradores (1994) ao avaliarem o plasma de tartarugas marinhas
cabeçudas, obtiveram média de 15U/L (± 9) para fosfatase alcalina, enquanto que em
amostras de soro desta mesma espécie o valor encontrado foi de 13U/L (± 4) (BOLTEN
et al., 1992). Por outro lado, valores mais elevados de 23U/L (± 14) e 36,3UI/L (± 16,3)
foram obtidos para esta variável em plasma de juvenis de tartarugas marinhas
cabeçudas, e animais em cativeiro por KELLER e colaboradores (2004) CUBAS &
BAPTISTOTTE (2006), respectivamente.
2.1.7.9 Aspartato Aminotransferase
A Aspartato Aminotransferase (AST) está presente em todos os tecidos dos
répteis. Embora a sua análise não seja considerada um teste específico, altas
concentrações são encontradas em tecidos muscular e hepático. Assim, elevações do seu
nível sérico relacionam-se a danos destes tecidos. Geralmente, os valores considerados
normais são inferiores a 250 U/L nestas espécies e, valores superiores aqueles são
sugestivos de danos hepático, na musculatura esquelética ou no miocárdio. Em doenças
generalizadas o aumento da AST está relacionado com necrose celular de diferentes
tecidos (CAMPBELL, 1996), enquanto que em tartarugas verdes esta elevação foi
observada em animais com fibropapilomatose (AGUIRRE et al., 1995; NORTON et al.,
1990).
A AST foi avaliada no soro de tartarugas marinhas cabeçudas de vida livre,
apresentando um valor médio de 180U/L (± 84) (BOLTEN et al., 1992). Amostras de
plasma desta espécie apresentaram um valor médio de 204U/L (± 90) para esta variável
(BOLTEN et al., 1994), e 229U/L (± 54) em juvenis (KELLER et al., 2004) e para
Caretta caretta mantidas em cativeiro o valor foi de 323,2UI/L (± 178,4) (CUBAS &
BAPTISTOTTE, 2006).

2.1.8 Eletroforese das Proteínas Séricas
A eletroforese de proteínas pode ser utilizada tanto para avaliar o estado de
saúde, quanto para auxiliar no diagnóstico de doenças em tartarugas marinhas
(GICKING et al., 2004). As técnicas atuais permitem uma alta qualidade na separação
das frações protéicas, com custo acessível e pequeno volume de amostra, sendo ainda
considerado como método acurado para mensurar albumina e frações de globulinas,
constituindo em um importante complemento na rotina dos exames hematológicos e
bioquímicos (CRAY & TATUM, 1998).
A albumina é considerada a mais abundante das proteínas séricas na
eletroforese, constituindo de 35 a 50% do seu total. E, sua magnitude auxilia, não só em
termos fisiológicos, mas também na interpretação do gráfico obtido por esta técnica,
sendo assim um guia útil para diferenciar os picos monoclonais de globulina
(KANEKO, 1997).
A determinação das proteínas séricas através da eletroforese pode auxiliar
também no diagnóstico laboratorial de répteis com hiperglobulinemia; as frações α, β e
γ podem estar aumentadas em doenças infecciosas e, geralmente as α-globulinas se
elevam em necrose tecidual estando diminuídas em doenças hepáticas severas, má
nutrição e má absorção. Além disso, importantes proteínas de fase aguda compõem esta
fração, como a seromucoide, ceruloplasmina e haptoglobina, sendo consideradas
marcadores de doença inflamatória aguda. A fração β-globulina é formada por proteínas
de grande valor diagnóstico, entre elas o fibrinogênio e a proteína C-reativa, que estão
também aumentadas em doenças inflamatórias agudas. As alterações das proteínas de
fase aguda geralmente ocorrem antes das mudanças dos valores bioquímicos e
hematológicos. A gama globulina, na maioria dos animais, é observada como duas
frações: gama 1 e gama 2, onde as imunoglobulinas (anticorpos) migram nestas duas
regiões (CAMPBELL, 1996; KANEKO, 1997; CRAY & TATUM, 1998).
FRAIR (1982) utilizou eletroforese de proteínas séricas em estudo taxonômico
de diferentes espécies de tartarugas marinhas e concluiu que aparentemente todas têm
linhas de migração similares, e também tem padrões relativamente curtos quando

comparados com humanos. No entanto, alguma variabilidade intraespecífica ocorreu,
sendo mais quantitativa (altura das linhas) do que qualitativa (posição das linhas).
CRAY e colaboradores (2001) detectaram bandas fracionadas, em estudo com
16 tartarugas verdes, equivalentes àquelas observadas em mamíferos: albumina, α1-
globulina, α2-globulina, β-globulina e γ-globulina. Ao realizar a eletroforese de
proteínas plasmáticas de 29 tartarugas cabeçudas, GICKING e colaboradores (2004)
encontraram uma concentração média de albumina de 1,0g/dL (±0,17), para a α-
globulina a média encontrada foi de 0,48g/dL (±0,1), enquanto que para β-globulina o
valor obtido foi de 0,80g/dL (±0,2) e para concentração de γ-globulina a média foi de
1,94g/dL (±0,62), valor considerado alto quando os autores compararam com espécies
de aves e mamíferos.
Poucos são os estudos com eletroforese em tartarugas marinhas, entretanto já há
um interesse em utilizar este meio diagnóstico nestas espécies, principalmente em
animais acometidos pela fibropapilomatose, que é uma doença debilitante, podendo
causar a morte do animal e, que tem crescido bastante entre a espécie Chelonia mydas,
alcançando altas incidências em algumas populações, tornando-as ameaçadas
(SWIMMER, 2000; AGUIRRE & BALAZS, 2000; CRAY et al., 2001). Tartarugas
verdes com fibropapilomatose severa apresentam diminuição dos valores de α1, α2 e γ-
globulinas (WORK, et al., 2001), sendo que valores baixos da fração gama são
indicativos de imunossupressão (KANEKO, 1997; WORK et al., 2001).

3 ARTIGOS CIENTÍFICOS
3.1 Artigo Científico I:
Hemograma e bioquímica sérica de tartarugas cabeçudas (Caretta caretta) de vida
livre e mantidas em cativeiro, no Litoral Norte da Bahia.
Hemogram and serum biochemistry for loggerhead sea turtles (Caretta caretta), free-ranging and captive in north coast of Bahia, Brazil.
Thaís Torres Pires1,3, Gonzalo Rostan3, Thereza Cristina Calmon de Bittencourt2 e José
Eugênio Guimarães1
1- Departamento de Patologia e Clínicas – Hospital de Medicina Veterinária - EMV/UFBA – Salvador/BA - Brasil
2- Departamento de Produção Animal – EMV/UFBA – Salvador/BA - Brasil 3- Projeto Tamar-Ibama – Mata de São João/BA - Brasil
RESUMO
Com o objetivo de se obter dados que auxiliem na rotina clínica de tartarugas marinhas, realizou-se o hemograma e a determinação de alguns parâmetros bioquímicos séricos de fêmeas de tartarugas cabeçudas (Caretta caretta) de vida livre (n = 22), animais que desovam no Litoral Norte da Bahia, e em cativeiro (n = 5), animais mantidos no Centro de Visitantes do Projeto Tamar-Ibama, na Praia do Forte. Os valores obtidos para os dois grupos foram comparados, onde observou-se diferença estatística significativa (p<0,05) para as contagens relativas e absolutas de eosinófilos e contagem relativa de monócitos dentre as variáveis do hemograma e, para a proteína total, globulina, glicose, colesterol e aspartato aminotransferase. Possivelmente, diferenças nas condições ambientais, nutricionais e reprodutivas, as quais estes animais foram submetidos, influenciaram nos resultados.
Palavras-chave: hemograma, bioquímica sérica, tartaruga marinha cabeçuda.

SUMMARY The aim of this study was to improve the clinical evaluation in sea turtles by analyzing the hemogram and serum biochemistry in loggerhead sea turtles (Caretta caretta), female, free-ranging (n=22) nesting in the north coast of Bahia and in captivity (n=5) in the visitor center of the Projeto Tamar-Ibama, in Praia do Forte, municipality of Mata de São João, Bahia, Brazil. The values obtained were compared between the two groups and it was observed that there was statistically significant difference (p<0,05) for the relative and absolute eosinophil count, relative monocyte count, total serum protein, globulin, glucose, cholesterol and aspartate aminotransferase. The results variation is probably related to the differences in environmental, nutritional and reproductive conditions. Keywords: hemogram, serum biochemistry, loggerhead sea turtles.
3.1.1 Introdução
Na investigação clínica de répteis, amostras sangüíneas podem ser facilmente
obtidas e são de grande valor diagnóstico. Entretanto, a patologia clínica é uma área da
medicina herpetológica que ainda necessita de vários estudos (DIVERS et al., 1996). A
bioquímica sérica representa uma importante ferramenta de monitoramento da saúde e
estado fisiológico de tartarugas marinhas. Pesquisas sobre este tema podem fornecer
informações fundamentais para a manutenção e conservação das populações selvagens
destas espécies (NORTON et al., 1990; BOLTEN & BJORDAL, 1992; AGUIRRE et
al., 1995; SAMOUR et al., 1998). Alguns trabalhos foram realizados, porém a
comparação dos dados obtidos é limitada devido as possíveis diferenças entre as
populações, bem como as variações de técnicas utilizadas (BOLTEN et al., 1992;
SWIMMER, 2000). Fatores como idade, tamanho, sexo, sazonalidade, saúde, habitat e
dieta podem afetar os parâmetros hematológicos, tanto em tartarugas marinhas como em
répteis de maneira geral (WOOD & EBANKS, 1984; GOTTDENKER & JACOBSON,
1995). Além disso, as descrições de características morfológicas de células sangüíneas
de quelônios marinhos são limitadas (CANNON, 1992; WORK et al., 1998; CASAL &
ORÓS, 2007).

Com a necessidade crescente de avaliação do estado de saúde em tartarugas
marinhas, para uma possível manutenção de animais saudáveis em cativeiro e
reabilitação de indivíduos de vida livre, se faz necessário um maior empenho em
desenvolver estudos relacionados aos meios auxiliares de diagnóstico clínico, onde a
avaliação hematológica possui grande destaque (PIRES et al., 2006).
Este trabalho propõe a determinação e comparação dos valores do hemograma e
de alguns parâmetros da bioquímica sérica de tartarugas cabeçudas (Caretta caretta)
selvagens que visitam o litoral norte da Bahia e de exemplares desta espécie criadas em
cativeiro, obtendo-se assim valores que possam auxiliar no diagnóstico clínico de
enfermidades que acometem estes animais.
3.1.2 Material e Métodos
Foram analisados dois grupos distintos de tartarugas marinhas cabeçudas
(Caretta caretta): animais de vida livre e animais em cativeiro. As amostras de sangue
dos animais de vida livre foram coletadas de 22 exemplares fêmeas que desovam no
Litoral Norte da Bahia, em áreas protegidas e monitoradas pelas bases do Projeto
Tamar-Ibama da Praia do Forte (12º34’S 38º00’W) e Arembepe (12º45'S 38º10'W),
logo após a desova, durante as temporadas reprodutivas, setembro a março, de
2004/2005 e 2005/2006.
Para os animais em cativeiro, colheu-se amostras sangüíneas de cinco fêmeas
que estão no Centro de visitantes do Projeto Tamar-Ibama da base de Praia do Forte –
Bahia, desde o nascimento, e com idades de 18 a 20 anos, no mês de março de 2006. Os
mesmos são alimentados diariamente com três tipos de peixe: corvina, galo e sardinha, e
mantidos em tanques com volume de 102 e 30 m3, recebendo água diretamente captada
do mar, em sistema aberto, que passa por filtros de areia.
As tartarugas foram mensuradas, e os valores médios do comprimento curvilíneo
de carapaça (CCC) foram de 1,00 m (±0,05) para os animais de vida livre e 0,96 m
(±0,03) para aquelas mantidas em cativeiro, caracterizando-os assim animais adultos.
Todos os animais foram avaliados clinicamente, onde se observou condição corporal,

presença de ectoparasitas, tumores e lesões cutâneas, sendo coletadas amostras apenas
dos animais considerados clinicamente saudáveis.
Após a contenção dos animais, segundo recomendações de OWENS & RUIZ
(1980), cinco mililitros de sangue foram coletados do seio cervical dorsal de cada
tartaruga. As amostras foram imediatamente armazenadas em tubos com heparina de
sódio para a realização do hemograma, e em tubos sem anticoagulante para obtenção de
soro, que foi congelado a -20ºC até o momento das análises bioquímicas. Em seguida
foram enviadas e processadas no Laboratório de Patologia Clínica Veterinária da Escola
de Medicina Veterinária da UFBA.
O volume globular (VG) foi determinado através da técnica de microhematócrito
(JAIN, 1986), enquanto que a concentração de hemoglobina, pelo método da
cianometahemoglobina, através de kit comercial (Doles). As contagens de eritrócitos e
leucócitos foram realizadas em Câmara hematimétrica de Neubauer, tendo como
diluente a solução de NATT & HERRICK (1952). As contagens diferencial de
leucócitos e de trombócitos foram estabelecidas em esfregaço sangüíneo corado
segundo a técnica de Rosenfeld descrita por BIRGEL e colaboradores (1983). A partir
das variáveis do eritrograma estabeleceu-se matematicamente os índices
hematimétricos: Volume Globular Médio (VGM) e Concentração de Hemoglobina
Globular Média (CHGM).
Para a determinação dos parâmetros da bioquímica sérica foram utilizados kits
comerciais (Labtest), onde a proteína sérica total foi determinada pelo método do
biureto modificado, com leitura espectrofotométrica em 550nm, e a albumina pelo
método Verde de bromocresol modificado com leitura espectrofotométrica em 630nm.
Através da diferença dos valores obtidos para essas variáveis, obteve-se o valor total de
globulina. A relação Albumina:Globulina (A:G) foi calculada a partir destes resultados.
As concentrações de fosfatase alcalina e aspartato amino transferanse foram obtidas
pelos métodos de Bowers & McComb modificado e cinética UV-IFCC, com leitura
espectrométrica em 405nm e 340nm, respectivamente, enquanto que para a creatinina
utilizou-se o método Labtest em comprimento de 510nm. O ácido úrico, o colesterol e
os triglicérideos foram obtidos através do método enzimático-Trinder com leitura em
espectrofotômetro utilizando o comprimento de onda de 520, 500 e 505nm,
respectivamente. E, para a concentração da glicose a metodologia utilizada foi a de
GOD-Trinder com leitura espectrofotométrica em 505nm.
Considerando-se o número reduzido de animais entre os grupos avaliados,
utilizou-se para efeito de estudo o teste não paramétrico de Mann & Whitney (1947),
por meio do programa SPSS, versão 11.0, comparando-se as médias dos grupos em
questão. O nível de significância usado para o teste foi de 5%.
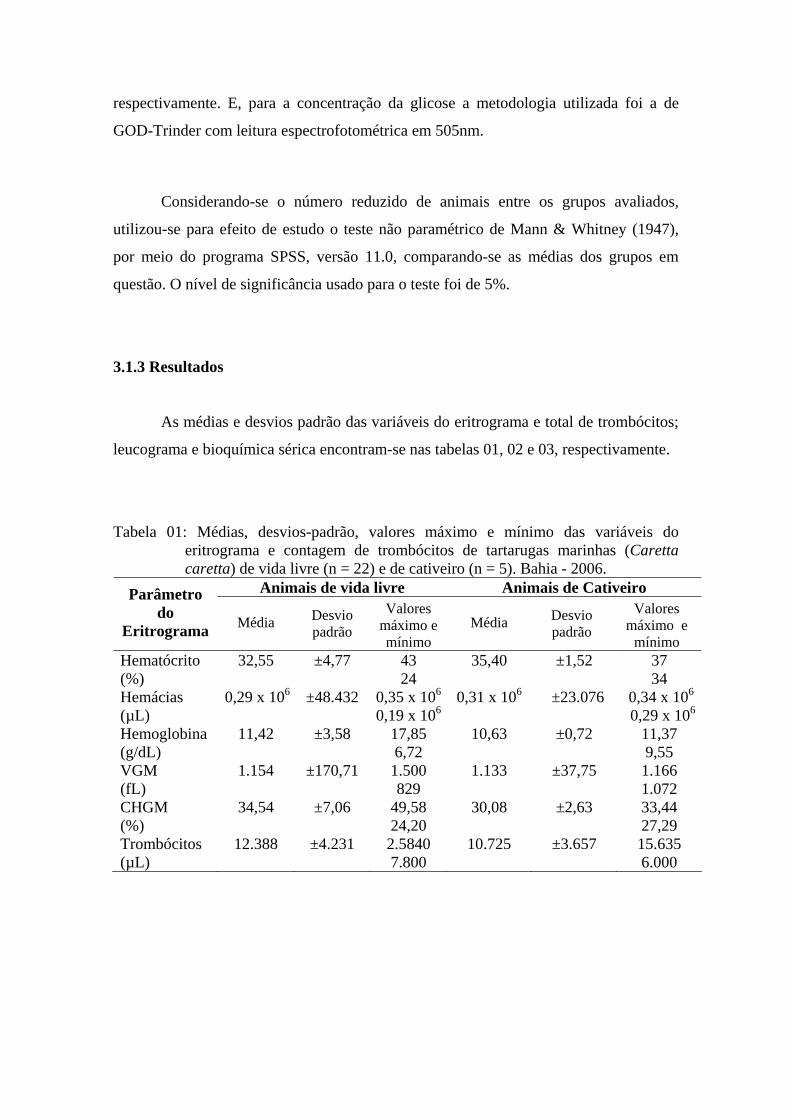
3.1.3 Resultados
As médias e desvios padrão das variáveis do eritrograma e total de trombócitos;
leucograma e bioquímica sérica encontram-se nas tabelas 01, 02 e 03, respectivamente.
Tabela 01: Médias, desvios-padrão, valores máximo e mínimo das variáveis do eritrograma e contagem de trombócitos de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e de cativeiro (n = 5). Bahia - 2006.
Animais de vida livre Animais de Cativeiro Parâmetro do
Eritrograma
Média Desvio padrão
Valores máximo e mínimo
Média Desvio
padrão Valores
máximo e mínimo
Hematócrito (%)
32,55 ±4,77 43 24
35,40 ±1,52 37 34
Hemácias (µL)
0,29 x 106 ±48.432 0,35 x 106
0,19 x 1060,31 x 106 ±23.076 0,34 x 106
0,29 x 106
Hemoglobina (g/dL)
11,42 ±3,58 17,85 6,72
10,63 ±0,72 11,37 9,55
VGM (fL)
1.154 ±170,71 1.500 829
1.133 ±37,75 1.166 1.072
CHGM (%)
34,54 ±7,06 49,58 24,20
30,08 ±2,63 33,44 27,29
Trombócitos (µL)
12.388 ±4.231 2.5840 7.800
10.725 ±3.657 15.635 6.000
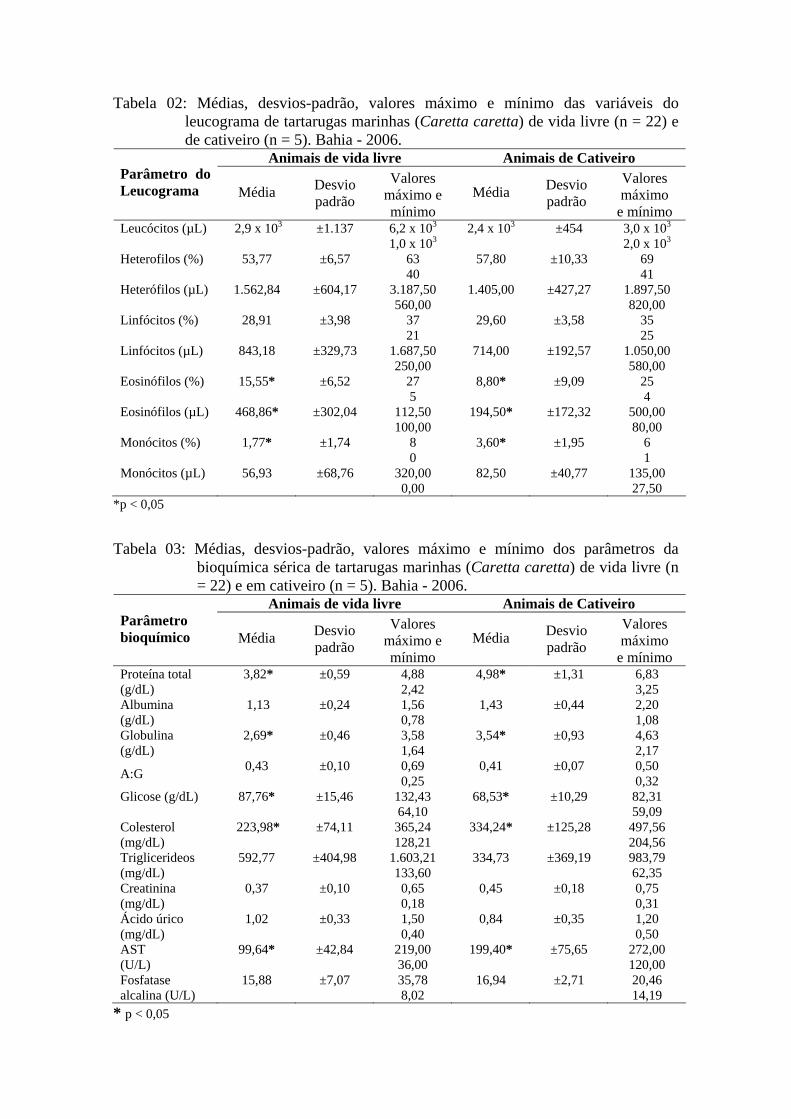
Tabela 02: Médias, desvios-padrão, valores máximo e mínimo das variáveis do leucograma de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e de cativeiro (n = 5). Bahia - 2006.
Animais de vida livre Animais de Cativeiro Parâmetro do Leucograma
Média Desvio
padrão Valores
máximo e mínimo
Média Desvio
padrão Valores máximo
e mínimo Leucócitos (µL) 2,9 x 103 ±1.137 6,2 x 103
1,0 x 1032,4 x 103 ±454 3,0 x 103
2,0 x 103
Heterofilos (%) 53,77 ±6,57 63 40
57,80 ±10,33 69 41
Heterófilos (µL) 1.562,84 ±604,17 3.187,50 560,00
1.405,00 ±427,27 1.897,50 820,00
Linfócitos (%) 28,91 ±3,98 37 21
29,60 ±3,58 35 25
Linfócitos (µL) 843,18 ±329,73 1.687,50 250,00
714,00 ±192,57 1.050,00 580,00
Eosinófilos (%) 15,55* ±6,52 27 5
8,80*
±9,09 25 4
Eosinófilos (µL) 468,86* ±302,04 112,50 100,00
194,50* ±172,32 500,00 80,00
Monócitos (%) 1,77* ±1,74 8 0
3,60*
±1,95 6 1
Monócitos (µL) 56,93 ±68,76 320,00 0,00
82,50 ±40,77 135,00 27,50
*p < 0,05
Tabela 03: Médias, desvios-padrão, valores máximo e mínimo dos parâmetros da
bioquímica sérica de tartarugas marinhas (Caretta caretta) de vida livre (n = 22) e em cativeiro (n = 5). Bahia - 2006.
Animais de vida livre Animais de Cativeiro Parâmetro bioquímico
Média Desvio
padrão Valores
máximo e mínimo
Média Desvio
padrão Valores máximo
e mínimo Proteína total (g/dL)
3,82* ±0,59 4,88 2,42
4,98*
±1,31 6,83 3,25
Albumina (g/dL)
1,13 ±0,24 1,56 0,78
1,43
±0,44 2,20 1,08
Globulina (g/dL)
2,69* ±0,46 3,58 1,64
3,54*
±0,93 4,63 2,17
A:G 0,43 ±0,10 0,69 0,25
0,41
±0,07 0,50 0,32
Glicose (g/dL) 87,76* ±15,46 132,43 64,10
68,53* ±10,29 82,31 59,09
Colesterol (mg/dL)
223,98* ±74,11 365,24 128,21
334,24* ±125,28 497,56 204,56
Triglicerideos (mg/dL)
592,77 ±404,98 1.603,21 133,60
334,73 ±369,19 983,79 62,35
Creatinina (mg/dL)
0,37 ±0,10 0,65 0,18
0,45
±0,18 0,75 0,31
Ácido úrico (mg/dL)
1,02
±0,33 1,50 0,40
0,84
±0,35 1,20 0,50
AST (U/L)
99,64* ±42,84 219,00 36,00
199,40*
±75,65 272,00 120,00
Fosfatase alcalina (U/L)
15,88 ±7,07 35,78 8,02
16,94 ±2,71 20,46 14,19
* p < 0,05

3.1.4 Discussão
Os valores obtidos para as variáveis do eritrograma não apresentaram diferença
estatística entre os grupos de vida livre e de cativeiro. As médias encontradas para o
volume globular para os dois grupos, foram próximas àquelas relatadas em estudos
realizados por BOLTEN & BJORNDAL (1992), AGUIRRE e colaboradores (1995) e
SAMOUR e colaboradores (1998) com tartarugas verdes, que apresentaram valores de
31 a 35%, enquanto que para tartarugas cabeçudas os valores obtidos variaram de 28 a
33% (ALKINDI & MAHMOUD, 2002; KELLER et al., 2004; PIRES et al., 2006;
CASAL & ORÓS, 2007).
Para a contagem total de eritrócitos as médias obtidas para tartarugas de vida
livre e para as cativas foram inferiores àquelas apresentadas por AGUIRRE e
colaboradores (1995) e SAMOUR e colaboradores (1998) para tartarugas verdes, de
400.000/µL (±95.000) para animais juvenis e 480.000/µL (±160.000) para fêmeas
adultas, e por KELLER e colaboradores (2004) para tartarugas cabeçudas juvenis de
410.000/µL (±98.000). Esta diferença pode ser atribuída a espécie e/ou idade dos
animais. Contudo, PIRES e colaboradores (2006) e CUBAS & BAPTISTOTTE (2006)
apresentam valores semelhantes aos encontrados neste trabalho, para exemplares de
cativeiro da espécie Caretta caretta, de 275.000/µL (±28.030) e 288.000/µL (±86.000),
respectivamente.
Ao avaliar as concentrações de hemoglobina, observa-se que KELLER e
colaboradores (2004) relatam valores semelhantes para esta variável, de 9,82g/dL
(±1,46) para tartarugas cabeçudas. Entretanto, AGUIRRE e colaboradores (1995),
ALKINDI & MAHMOUD (2002) e PIRES e colaboradores (2006) apresentaram
valores próximos, de 8,54g/dL (±1,44) para juvenis de tartaruga verde, e 8,87g/dL (±
1,41) e 8,64g/dL (±0,80), para tartaruga cabeçuda adultas, respectivamente.
Os valores do índice hematimétrico VGM (Volume Globular Médio)
encontrados neste trabalho foram discrepantes daqueles obtidos por AGUIRRE e
colaboradores (1995), em estudo com juvenis de tartarugas verdes de vida livre de
725,00fL (± 312,35), o que era esperado, já que estes autores apresentaram valores mais
elevados para a contagem de eritrócitos que é utilizado para o cálculo matemático deste

índice, porém próximos aos valores apresentados por PIRES e colaboradores (2006) e
CUBAS & BAPTISTOTTE (2006) de 1.214,36 fL (±131,99) e 1.106,2 fL (±413,1) em
tartarugas cabeçudas.
Com relação à Concentração de Hemoglobina Globular Média (CHGM), a
média obtida para tartarugas selvagens e de cativeiro foram próximas aos valores
apresentados pela literatura, que variou de 26,10% (PIRES et al., 2006) a 29,5%
(CUBAS & BAPTISTOTTE, 2006).
Os valores obtidos na contagem de trombócitos de 12.388/µL (±4.231) para o
grupo de vida livre e de 10.725/µL (±3.657) cativeiro foram similares aos apresentados
por WOOD & EBANKS (1984) de 11.520/µL, para tartarugas verdes e por PIRES e
colaboradores (2006) de 10.968/µL (±3.109), para tartarugas cabeçudas, enquanto que
CUBAS & BAPTISTOTTE (2006) apresentaram uma média superior de 21.932/µL
(±14.859) para a mesma espécie. Vale salientar que a presença de aglutinados de
trombócitos nos esfregaços é comum (WOOD & EBANKS, 1984; HARMS et al., 2000;
CASAL & ORÓS, 2007), podendo interferir nos valores da contagem deste tipo celular,
levando a resultados subestimados. Além disso, possivelmente a utilização de técnicas
diferentes, a exemplo da contagem em câmara de Neubauer, pode apresentar valores
superestimados, pois a presença de núcleos das hemácias lisadas dificulta esta
contagem.
As contagens de leucócitos totais não apresentaram diferença significativa entre
os grupos estudados e foram inferiores às relatadas por KELLER e colaboradores
(2004) de 13.300/µL (±5.300) para juvenis desta espécie, e ligeiramente inferiores aos
valores apresentados por CUBAS & BAPTISTOTTE (2006) de 4.249/µL (±1586.7) e
PIRES e colaboradores (2006) de 3.656/µL (±963,04) para tartarugas cabeçudas cativas.
Esta variação pode ser explicada pelo período da coleta das amostras de fêmeas que
ocorreu na estação reprodutiva, onde devido a presença de hormônios, como a
gonodotrofina e esteróides sexuais, normalmente as fêmeas apresentam valores
inferiores às dos machos (WOOD & EBANKS, 1984).
Ao avaliar os valores obtidos no diferencial de leucócitos para os grupos,
observou-se que as contagens relativa e absoluta dos eosinófilos apresentaram

diferenças significativas, onde animais de vida livre apresentaram um número mais
elevado desta célula. Como as tartarugas cativas são mantidas em ambiente controlado e
passaram por procedimento de vermifugação, quando necessário, entende-se que os
animais de vida livre possuem um estímulo parasitário maior para a produção de
eosinófilos. O aumento deste tipo celular pode ser relacionado a casos de parasitismo
(ROSSKOPF, 1982), já que uma das funções do eosinófilo é participar da defesa contra
infecções parasitárias (AZEVEDO & LUNARDI, 2003).
Houve diferença significativa entre os valores da contagem relativa de
monócitos entre os dois grupos, entretanto esta diferença não está presente na avaliação
dos valores da contagem absoluta. Não foram observados basófilos nos grupos
estudados, contudo, a presença deste tipo celular na circulação de tartarugas marinhas
hígidas é rara, como relatam WOOD & EBANKS (1984), WORK e colaboradores
(1998) e CANNON (1992). Para a contagem diferencial de leucócitos a literatura
apresenta valores discrepantes, possivelmente devido à grande variação entre indivíduos
da mesma faixa etária (WOOD & EBANKS, 1984) e à dificuldade de classificação dos
leucócitos, que possuem morfologia variável entre os répteis em geral, não tendo ainda
critérios bem estabelecidos para a descrição das células sangüíneas das tartarugas
marinhas, assim como as diferenças geográficas (KNOTKAVÁ et al., 2002; CASAL &
ORÓS, 2007).
Entre os parâmetros bioquímicos analisados, a proteína total apresentou
diferenças estatísticas significativas ao se comparar os grupos estudados, onde animais
de cativeiro apresentaram valores mais elevados, o que pode ser resultado de uma dieta
rica em proteína oferecida a estes (SWIMMER, 2000). Os valores obtidos para proteína
total de 3,82g/dL (±0,59) para o grupo de animais de vida livre e 4,98 (±1,31) para as
tartarugas de cativeiro foram semelhantes aos descritos para esta espécie por BOLTEN
e colaboradores (1992 e 1994) de 4,1g/dL (± 1,2), KELLER e colaboradores (2004), de
4,0g/dL (± 0,8) e por CUBAS & BAPTISTOTTE (2006) de 4,2g/dL (± 0,4).
Para a albumina as médias encontradas neste trabalho foram 1,13g/dL (±0,24) e
1,43g/dL (±0,44) para animais de vida livre e cativos, respectivamente. Alguns
trabalhos apresentam valores para esta variável: BOLTEN e colaboradores (1992 e
1994) obtiveram uma média de 1,1g/dL (± 0,2) e 0,9g/dL (± 0,3), KELLER e

colaboradores (2004) de 1,1g/dL (± 0,2), enquanto que para CUBAS & BAPTISTOTTE
(2006) esse valor foi de 0,7g/dL (± 0,2), Esta diferença entre os valores pode ser
atribuída às diferentes técnicas utilizadas nos estudos, sendo que BOLTEN e
colaboradores (1992) ao testarem duas técnicas distintas apresentaram resultados
significativamente diferentes para alguns parâmetros bioquímicos, inclusive para a
albumina.
Em relação às globulinas, os resultados encontrados neste estudo de 2,69g/dL
(±0,46) para animais selvagens e 3,54 (±0,93) para os cativos, diferiram
estatisticamente, sendo que os valores inferiores para o grupo de vida livre,
possivelmente possa ser atribuído ao efeito do estresse reprodutivo sobre a função
imunológica destas fêmeas. Entretanto, os valores obtidos foram próximos aos valores
relatados por BOLTEN e colaboradores (1994), de 3,2g/dL (± 0,9), e KELLER e
colaboradores (2004) de 2,9g/dL (± 0,7) para tartarugas cabeçudas.
A média da relação A:G, de 0,43 para os animais de vida livre e 0,41 para os
animais de cativeiro foram bastante próximas as obtidas em outros trabalhos com a
espécie em questão, de 0,3 (± 0,1) (BOLTEN et al., 1994), e 0,4 (± 0,08) (KELLER et
al., 2004).
Para a variável glicose, a média do grupo de vida livre de 87,76mg/dL (± 15,46)
foi superior e estatisticamente diferente daquela encontrada para o grupo de cativeiro de
68,53mg/dL (± 10,29). Este resultado pode ter sido causado pelo estresse reprodutivo, já
que estudos relatam hiperglicemia em resposta ao estresse agudo em tartarugas
marinhas, assim como nos mamíferos (AGUIRRE et al., 1995; GREGORY &
SCHMID, 2001). Os valores obtidos foram ligeiramente inferiores aos encontrados por
outros autores, onde BOLTEN e colaboradores (1994) relatam uma média de 97mg/dL
(± 21), KELLER e colaboradores (2004) de 109mg/dL (± 18) e CUBAS &
BAPTISTOTTE (2006) de 102,8mg/dL (± 17), ao avaliarem o plasma de tartarugas
cabeçudas. Em estudo que utilizou o soro desta espécie, a média encontrada foi de
80,1mg/dL (± 22,3) (BOLTEN et al., 1992).
Houve diferença significativa entre as médias obtidas para o colesterol, onde
para animais de vida livre o valor foi de 223,98mg/dL (± 74,11) e para os cativos de

334,24mg/dL (± 125,28). Tartarugas marinhas mantidas em cativeiro podem apresentar
níveis mais elevados de colesterol devido à dietas ricas em gorduras (SWIMMER,
2000) o que justifica os valores encontrados neste estudo. Ao comparar os dados deste
trabalho constata-se também que estes foram superiores aos relatados pela bibliografia
para a espécie Caretta caretta, onde BOLTEN e colaboradores (1992 e 1994) obtiveram
uma média de 106mg/dL (± 57) e 170mg/dL (± 89) para esta variável, enquanto que
CUBAS & BAPTISTOTTE (2006) encontraram uma média de 107mg/dL (± 26,1) para
o colesterol. Esta diferença pode ser atribuída ao sexo como afirmam BOLTEN &
BJORNDAL (1992).
As médias encontradas para os triglicerídeos, para animais de vida livre e de
cativeiro, não apresentaram diferenças estatísticas significativas e foram superiores às
relatadas por outros autores que trabalharam com tartarugas cabeçudas, de 81mg/dL (±
204) (BOLTEN et al., 1992), 212mg/dL (± 334) (BOLTEN et al., 1994) e 179mg/dL (±
68,5) (CUBAS & BAPTISTOTTE, 2006). Entretanto, SWIMMER (2000) obteve um
valor médio de 486,7mg/dL (± 31,66) para tartarugas verdes de cativeiro, afirmando que
a exemplo do colesterol a dieta rica em gorduras ofertadas as tartarugas marinhas
cativas favorecem níveis elevados deste parâmetro Além disso, cita ainda o exercício
insuficiente como outro fator para o aumento de triglicerídeos, o que justifica os valores
obtidos para o grupo de cativeiro deste trabalho. Para o grupo de vida livre observou-se
uma grande variação dos valores, que se reflete no desvio padrão elevado, podendo ser
causado por coletas sangüíneas em períodos de lipemia pós prandial, já que não é
possível controlar a alimentação dos animais selvagens.
Os valores obtidos para a creatinina não apresentaram diferenças estatísticas
significativas entre os grupos estudados e foram próximos aos encontrados por
BOLTEN e colaboradores (1994) de 0,3mg/dL (± 0,1) e CUBAS & BAPTISTOTTE
(2006) de 0,2mg/dL (± 0,1) para a espécie em questão.
Não houve diferença entre as médias obtidas para o ácido úrico entre os grupos
de cativeiro e vida livre, estando os valores relatados neste estudo de acordo com a
literatura, que apresentam valores para a espécie cabeçuda de 0,8mg/dL (BOLTEN et
al., 1992; KELLER et al., 2004) e 1,0mg/dL (± 0,4) (CUBAS & BAPTISTOTTE,
2006).

Para a aspartato aminotransferase (AST) houve diferença significativa entre os
grupos estudados, o que pode ser atribuído ao confinamento e dietas não naturais como
sugere SWIMMER (2000). Contudo, ao se analisar os valores da literatura constata-se
que a média encontrada neste trabalho para as tartarugas de vida livre, de 99,64U/L (±
42,84), foi inferior àquelas relatadas por BOLTEN e colaboradores (1992 e 1994), e
KELLER e colaboradores (2004) de 180U/L (± 84), 204U/L (± 90) e 229U/L (± 54),
respectivamente, porém é próxima a obtida para o grupo de cativeiro de 199,40U/L (±
75,65). Vale ressaltar que CAMPBELL (1996) sugere como referência para esta
variável um intervalo de 100 a 350U/L para tartarugas marinhas, estando os valores
deste trabalho compatíveis com este autor.
Outro parâmetro avaliado nesta pesquisa foi a fosfatase alcalina onde os valores
obtidos não apresentaram diferença significativa entre os grupos estudados e foram
semelhantes aos relatados na literatura para a espécie, que variam de 15 a 36U/L
(BOLTEN et al., 1994; KELLER et al., 1994; CUBAS & BAPTISTOTTE, 2006).
3.1.5 Conclusão
As condições ambientais, nutricionais e reprodutivas, as quais estes animais
foram submetidos, influenciaram os resultados, observando-se valores distintos para as
contagens relativas de eosinófilos e monócitos e absoluta de eosinófilos, concentração
de glicose, colesterol, proteína total, globulina e aspartato aminotransferase.

3.1.6 Referências Bibliográficas
AGUIRRE, A. A.; BALAZS, G. H.; SPRAKOR, T. R.; GROSS, T. S. Adrenal and hematological responses to stress in juvenile green turtles, Chelonia mydas, with and without fibropapillomas. Phisiologycal Zoology, Chicago, v. 68, n. 5, p. 831 - 854, 1995. ALKINDI, A.Y.A.; MAHMOUD, I.Y. Hematological survey in two species of sea turtles in the Arabian sea during nesting season. Pakistan Journal of Biological Sciences, v. 5, n. 3, p. 359-361, 2002. AZEVEDO, A.; LUNARDI, L. O. Cytochemical characterization of eosinophilic leukocytes circulating in blood of the turtles (Chrysemys dorbignih). Acta Histochemica, Jena, v. 105, n. 1, p. 99-105, 2003.
BIRGEL, E. H.; LARSSON, M. H. A.; HAGIWARA, M. K.; VASCONSELOS, S. A.; LARSSON, C. E.; OGASSAWARA, S. BENESI, F. J. Patologia clínica veterinária. São Paulo, 1983. 260p.
BOLTEN, A. B.; BJORNDAL, K. A. Blood profiles for a wild population of green turtles (Chelonia mydas) in the southern Bahamas: size-specific and sex-specific relationships. Journal of Wildlife Diseases, Ames, v. 28, n. 3, p. 407 - 413, 1992. BOLTEN, A. B.; BJORNDAL, K. A.; ELIAZAR, P.J.; GREGORY, L.F. Seasonal abundance, size distribution, and blood biochemical values of loggerheads (Caretta caretta) in Port Canaveral Ship Channel, Florida. Miami: NOAA, 1994. 39p. (NOAA Technical Memorandum NMFS-SEFSC, 353). BOLTEN, A. B.; JACOBSON, E. R.; BJORNDAL, K. A. Effects of anticoagulant and auto analyzer on blood biochemical values of loggerhead sea turtle (Caretta caretta). American Journal of Veterinary Research, Schaumburg , v. 53, n. 12, p. 2224 - 2227, 1992. CAMPBELL, T. W. Clinical pathology. In: MADER, D. R. Reptile Medicine and Surgery. Philadelphia: W. B. Saunders Company, 1996. p. 248 - 257. CANNON, M. S. The morphology and cytochemistry of blood leukocytes of Kemp’s sea turtles (Lepidochelys kempi). Can. J. Zoo., v. 70, n. 7, p. 1336 - 1340, 1992. CASAL, A.B.; ORÓS, J. Morphologic and cytochemical characteristic of blood cells of juvenile loggerhead sea turtles (Caretta caretta). Research in Veterinary Science, London, v. 82, n. 2, p. 158-165, 2007. CUBAS, P.H.; BAPTISTOTTE, C. Chelonia (tartaruga, cágado e jabuti), In: CUBAS, Z.S.; SILVA, J.C.R.; CATÃO-DIAS, J.L. Tratado de Animais Selvagens: Medicina Veterinária. São Paulo: Roca, 2006. p. 86-119. DIVERS, S. J.; REDMAYNE, G.; AVES, K. Haematological and biochemical of 10 green iguanas (Iguana iguana). The Veterinary Record, London, v. 138, n. 3, p. 203 - 205, 1996.

GOTTDENKER, N. L.; JACOBSON, E. R. Effect of venipuncture sites on hematologic and clinical biochemical values in desert tortoises (Gopherus agassizii). American Journal of Veterinary Research, Schaumburg, v.56, n. 1, p. 19 - 21, 1995. GREGORY, L.F.; SCHMID, J.R. Stress responses and sexing of wild kemp’s ridley sea turtles (Lepidochelys kempii) in the northeastern Gulf of Mexico. General and Comparative Endocrinology, v. 124, n. 1, p. 66-74, 2001. HARMS, C.A.; KELLER, J.M.; KENNEDY-STOSKOPF, S. Use os a two-step percoll® gradient for separation os loggerhead sea turtle peripheral blood mononuclear cells. Journal of Wildlife Diseases, Ames, v. 36, n. 3, p. 535-540, 2000. JAIN, N.C. Schalm’s veterinary hematology. 4 ed. Philadelphia: Lea & Febiger, 1986. 1221p. KELLER, J. M., KUCKLICK, J. R., STAMPER, M. A., HARMS, C. A., MCCLELLAN-GREEN, P. D. Associations between organochlorine contaminant concentrations and clinical health parameters in loggerhead sea turtles from North Carolina, USA. Environmental Health Perspectives, v. 112, n. 10, p. 1074-1079, 2004. KNOTKOVÁ, Z.; DOUBEK, J.; KNOTEK, Z.; HÁJKOVÁ P. Blood cell morphology and plasma biochemistry in Russian tortoises (Agrionemys horsfieldi). Acta Veterinaria Brno, Brno, v. 71, n. 2, p. 191-198. 2002. MANN, H.B.; WHITNEY, D.R. On a test of whether one of two random variables is statistically larger than the other. Ann. Math. Stat., v. 18, p.50-60, 1947. NORTON, T.M., JACOBSON, E.R., SUNDBERG, J.P. Cutaneous fibropapillomas and renal mixofibroma in a Green turtles, Chelonia mydas. Journal of wildlife diseases, Ames, v. 26, n. 2, p. 265-270, 1990. NATT, M. P.; HERRICK, G. A. A new blood diluent for counting erythrocytes and leukocytes of the chicken. Poultry Science, v. 31, p. 735 - 738, 1952. OWENS, D. W.; RUIZ G. J. New methods of obtaining blood and cerebrospinal fluid from marine turtles. Herpetologica, Johnson City, v. 36, n.1, p.17 - 20, 1980. PIRES, T.T.; ROSTAN, G.; GUIMARÃES, J.E. Hemograma e determinação da proteína plasmática total de tartarugas marinhas da espécie Caretta caretta (Linnaeus, 1758), criada em cativeiro, Praia do Forte, Município de Mata de São João – Bahia. Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 43, n. 3, p. 348-353, 2006. ROSSKOPF, W. J. Normal hemogram and blood chemistry values for California desert tortoises. Veterinary Medicine/ Small Animal Clinician, Book Stacks, v.77, n. 1, p. 85 - 87, 1982.

SAMOUR, H. J.; HOWLETT, J.C.; SILVANOSE, C.; HASBUN, C.R.; AL-GHAIS, S.M. Normal haematology of free-living green turtles (Chelonia mydas) from the United Arab Emirates. Comparative Haematology International, London, v. 8, n. 2, p. 102-107, 1998.
SWIMMER, J.Y. Biochemical responses to fibropapilloma and captivity in the green turtles. Journal of Wildlife Diseases, Ames, v. 36, n. 1, p. 102-110, 2000.
WOOD, F. E.; EBANKS, G. Blood cytology and hematology of the green sea turtle, Chelonia mydas.Herpetologica, Johnson City, v. 40, n.3, p.331 - 336, 1984. WORK, T. M.; RASKIN R. E.; BALAZS, G. H.; WHITTAKER, S. D. Morphologic and cytochemical characteristics of blood cells from Hawaiian green turtles. American Journal of Veterinary Research, Schaumburg, v.59, n. 10, p. 1252 - 1257, 1998.

3.2 Artigo Científico II
Eletroforese de proteínas séricas de tartarugas cabeçudas (Caretta caretta) de vida
livre e mantidas em cativeiro no Litoral Norte da Bahia.
Serum protein electrophoresis for loggerhead sea turtles (Caretta caretta), free-ranging and captive in north coast of Bahia, Brazil.
Thaís Torres Pires1,3, Gonzalo Rostan3, Thereza Cristina Calmon de Bittencourt2, Maria Ângela Ornelas de Almeida1 e José Eugênio Guimarães1
1- Departamento de Patologia e Clínicas – Hospital de Medicina Veterinária - EMV/UFBA – Salvador/BA - Brasil
2- Departamento de Produção Animal – EMV/UFBA – Salvador/BA - Brasil 3- Projeto Tamar-Ibama – Mata de São João/BA - Brasil
RESUMO
Este trabalho objetivou a determinação e comparação do perfil eletroforético das proteínas séricas de tartarugas cabeçudas (Caretta caretta), fêmeas, de vida livre (n=20) que desovam no Litoral Norte da Bahia e daquelas mantidas em cativeiro (n=5) no Centro de Visitantes do Projeto Tamar-Ibama da Praia do Forte. Os valores obtidos para os animais de vida livre para as variáveis foram: proteína total de 3,84±0,56g/dL, albumina 1,39±0,30g/dL, alfa-1 e alfa-2 globulina 0,34±0,09g/dL e 0,42±0,22g/dL, beta globulina 0,57±0,26g/dL e gama globulina 1,16±0,30g/dL, e relação A:G 0,58±0,16. Para os animais cativos estes valores foram: proteina total 4,98±1,31g/dL, albumina 1,64±0,55g/dL, enquanto que para as frações alfa-1, alfa-2, beta e gama globulinas 0,39±0,24g/dL, 0,68±0,44g/dL, 0,68±0,13g/dL e 1,59±0,31g/dL, respectivamente; para a relação A:G de 0,49±0,08. Avaliando os resultados observou-se diferenças estatísticas significativas (p<0,05) para os valores de proteína total onde os animais de cativeiro apresentaram níveis mais elevados, o que pode ser atribuído a alimentação rica em proteína oferecida, e para a gama globulina onde o grupo de vida livre obteve valores inferiores, possivelmente devido a uma imunodepressão relacionada ao estresse reprodutivo. Apesar destas diferenças o padrão eletroforético foi semelhante entre os grupos estudados.
Palavras-chave: eletroforese, frações protéicas, tartarugas cabeçudas.

SUMMARY The purpose of this study was to determine and to compare the serum protein electrophoresis profile in loggerhead sea turtles (Caretta caretta), female, free-ranging (n=20) nesting in the north coast of Bahia and the animals kept in captivity (n=5) in the visitor center of the Projeto Tamar-Ibama, in Praia do Forte, municipality of Mata de São João, Bahia, Brazil. The values obtained for the free-ranging turtles were: total protein 3,84±0,56g/dL, albumin 1,39±0,30g/dL, alfa-1 and alfa-2 globulin 0,34±0,09g/dL and 0,42±0,22g/dL, beta globulin 0,57±0,26g/dL, and gama globulin 1,16±0,30g/dL, and A:G ratio 0,58±0,16, while for the captives ones these values were: total protein 4,98±1,31g/dL, albumin 1,64±0,55g/dL, alfa-1, alfa-2, beta and gama globulin 0,39±0,24g/dL, 0,68±0,44g/dL, 0,68±0,13g/dL and 1,59±0,31g/dL, respectively; A:G ratio 0,49±0,08. The total protein values were significantly higher (p<0,05) for the captivities turtles; this fact may be related with the high protein feeding. Differences were also identified for the gamma globulin for the free-ranging, that presented lower values, probably due to reproductive stress and immunosuppression. Despite these considerations, the electrophoresis profile for the two loggerhead turtles groups were similar in general. Keywords: electrophoresis, protein fractions, loggerhead sea turtles. 3.2.1 Introdução
O litoral norte da Bahia é uma das principais áreas de desova de tartarugas
marinhas no Brasil, principalmente das tartarugas cabeçudas (Caretta caretta) e
tartarugas de pente (Eretmochelys imbricata) (D’AMATO & MARCOVALDI, 1997).
Outras espécies como a oliva (Lepidochelys olivacea) e verde (Chelonia mydas)
também freqüentam este litoral, porém em menor número. Assim, esta área apresenta
grande importância biológica para estas espécies, pois grande parte das desovas
registradas na costa brasileira ocorre em praias baianas, mais especificamente na Praia
do Forte, município de Mata de São João e Arembepe, município de Camaçari.
Sendo a tartaruga cabeçuda a espécie com maior índice de desova nas praias do
continente, no Brasil (MARCOVALDI & MARCOVALDI, 1999; BAPTISTOTTE et
al., 2003), ela contribui com cerca de 80% dos ninhos durante cada estação reprodutiva.
Todas as espécies de tartarugas marinhas são listadas como vulneráveis ou ameaçadas
de extinção pela World Conservation Union (IUCN, 2006) e, o Projeto Tamar-Ibama é

responsável pela pesquisa e conservação das tartarugas marinhas no Brasil, desde 1980
(MARCOVALDI & MARCOVALDI, 1999).
Os esforços para conservar as espécies de tartarugas marinhas, os avanços na
medicina veterinária, com estudos dos parâmetros fisiológicos, têm crescido
recentemente (CASAL & OROS, 2007). Entretanto, ainda são escassas as informações
de valores de parâmetros sangüíneos para estes animais. A avaliação do estado
fisiológico de populações de espécies ameaçadas se faz necessário para o
desenvolvimento apropriado de planos de manejo e conservação (BOLTEN &
BJORNDAL, 1992).
A eletroforese de proteínas é de grande valor como método auxiliar para o
diagnóstico de doenças e compreensão de alterações fisiológicas (CRAY & TATUM,
1998; KANEKO, 1997), pois mudanças dramáticas nas frações protéicas são evidentes
em diversas enfermidades e podem auxiliar na procura de um real diagnóstico, quando
outros testes são pouco conclusivos. Esta técnica proporciona informações importantes
sobre vários componentes de proteínas do soro ou plasma, os quais podem ajudar no
diagnóstico diferencial em muitos casos, além de requerer pequena quantidade de
amostra para o seu processamento. Ela tem se mostrado um teste diagnóstico auxiliar
muito útil em mamíferos e aves, que ainda precisa ser explorado na clínica de répteis e
anfíbios (ZAIAS & CRAY, 2002). Assim sendo, a eletroforese de proteínas plasmáticas
pode ser utilizada como uma importante ferramenta na avaliação da saúde de tartarugas
marinhas, e a obtenção de valores de referência é necessária, respeitando-se as
condições ambientais, nutricionais e fatores fisiológicos que possam interferir ou
contribuir para aqueles dados (GICKING et al., 2004).
Este trabalho teve como objetivo estabelecer e comparar o perfil eletroforético
de tartarugas marinhas cabeçudas (Caretta caretta), fêmeas, clinicamente saudáveis,
criadas em cativeiro e em vida livre, de maneira que posteriormente possa auxiliar na
avaliação clínica desta espécie.

3.2.2 Materiais e Métodos
Foram analisados dois grupos distintos de tartarugas marinhas cabeçudas
(Caretta caretta): animais de vida livre (n=20) e em cativeiro (n=5). As amostras de
sangue dos animais de vida livre foram coletadas das fêmeas, durante as temporadas
reprodutivas, de setembro a março de 2004/2005 e 2005/2006, após desova no Litoral
Norte da Bahia, em áreas protegidas pelas bases do Projeto Tamar-Ibama da Praia do
Forte (12º34’S,38º00’W) e Arembepe (12º45'S 38º10'W). Para os animais em cativeiro,
foram coletadas amostras de fêmeas existentes no Projeto Tamar-Ibama, Centro de
visitantes da base de Praia do Forte – Bahia, em março de 2006. Os animais foram
mensurados, os valores médios do comprimento curvilíneo de carapaça (CCC) foram de
1,00 m (±0,05) para os animais de vida livre e 0,97 m (±0,03) para as tartarugas
mantidas em cativeiro. Todos os animais foram avaliados clinicamente, onde observou-
se condição corporal, presença de ectoparasitas, tumores e lesões cutâneas. Foram
coletadas amostras apenas dos animais considerados clinicamente saudáveis.
As amostras de sangue foram coletadas do seio cervical dorsal (OWENS &
RUIZ, 1980), imediatamente armazenadas em tubos sem anticoagulante, e em seguida
enviadas ao Laboratório de Patologia Clínica do Hospital Veterinário da Escola de
Medicina Veterinária da UFBA, onde foram centrifugadas para obtenção do soro, que
foi congelado a -20ºC.
A separação eletroforética das proteínas foi realizada em gel de agarose
utilizando-se kit comercial (Celm Gel). As fitas de agarose foram coradas com o Amido
Black e lidas em densitômetro (CELM DS-35) com comprimento de onda de 520nm. A
proteína sérica total foi determinada, pelo método do biureto modificado através de kit
comercial (Labtest), com leitura espectrofotométrica em 550nm.
Considerando-se o número reduzido de animais entre os grupos avaliados,
utilizou-se para efeito de estudo o teste não paramétrico de Mann & Whitney (1947),
por meio do programa SPSS, versão 11.0, comparando-se as médias dos grupos em
questão. O nível de significância usado para o teste foi de 5%.
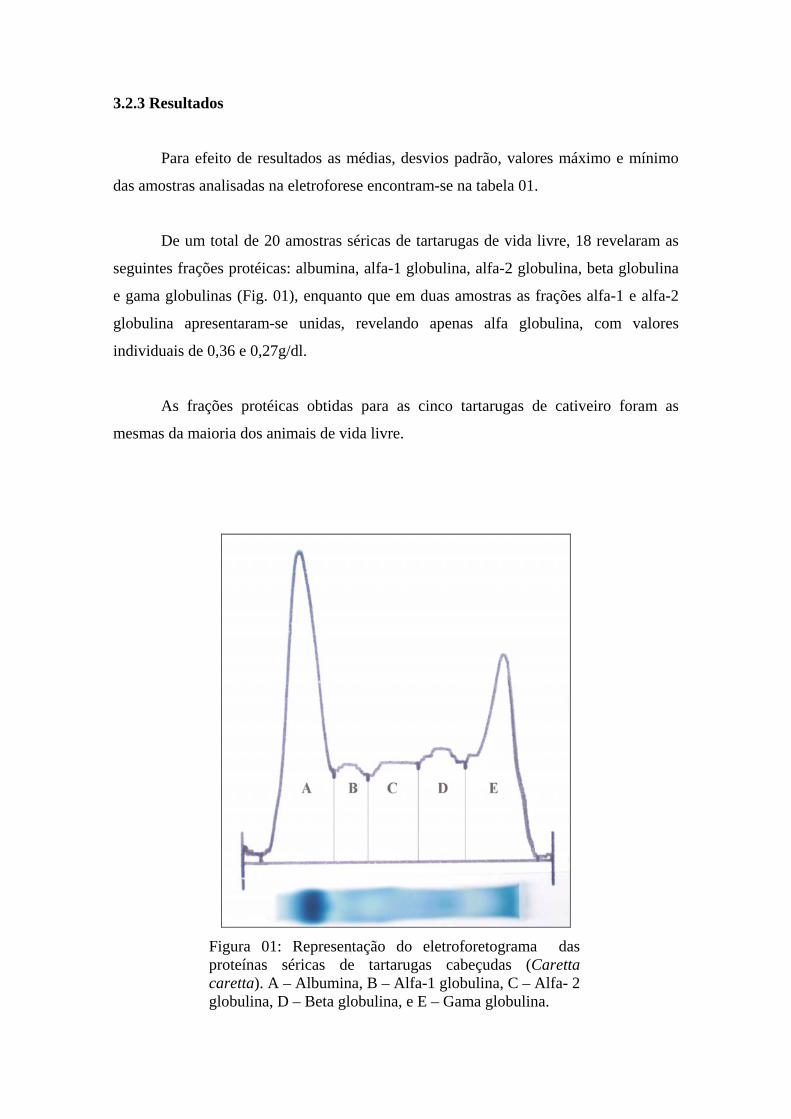
3.2.3 Resultados
Para efeito de resultados as médias, desvios padrão, valores máximo e mínimo
das amostras analisadas na eletroforese encontram-se na tabela 01.
De um total de 20 amostras séricas de tartarugas de vida livre, 18 revelaram as
seguintes frações protéicas: albumina, alfa-1 globulina, alfa-2 globulina, beta globulina
e gama globulinas (Fig. 01), enquanto que em duas amostras as frações alfa-1 e alfa-2
globulina apresentaram-se unidas, revelando apenas alfa globulina, com valores
individuais de 0,36 e 0,27g/dl.
As frações protéicas obtidas para as cinco tartarugas de cativeiro foram as
mesmas da maioria dos animais de vida livre.
Figura 01: Representação do eletroforetograma das proteínas séricas de tartarugas cabeçudas (Caretta caretta). A – Albumina, B – Alfa-1 globulina, C – Alfa- 2 globulina, D – Beta globulina, e E – Gama globulina.
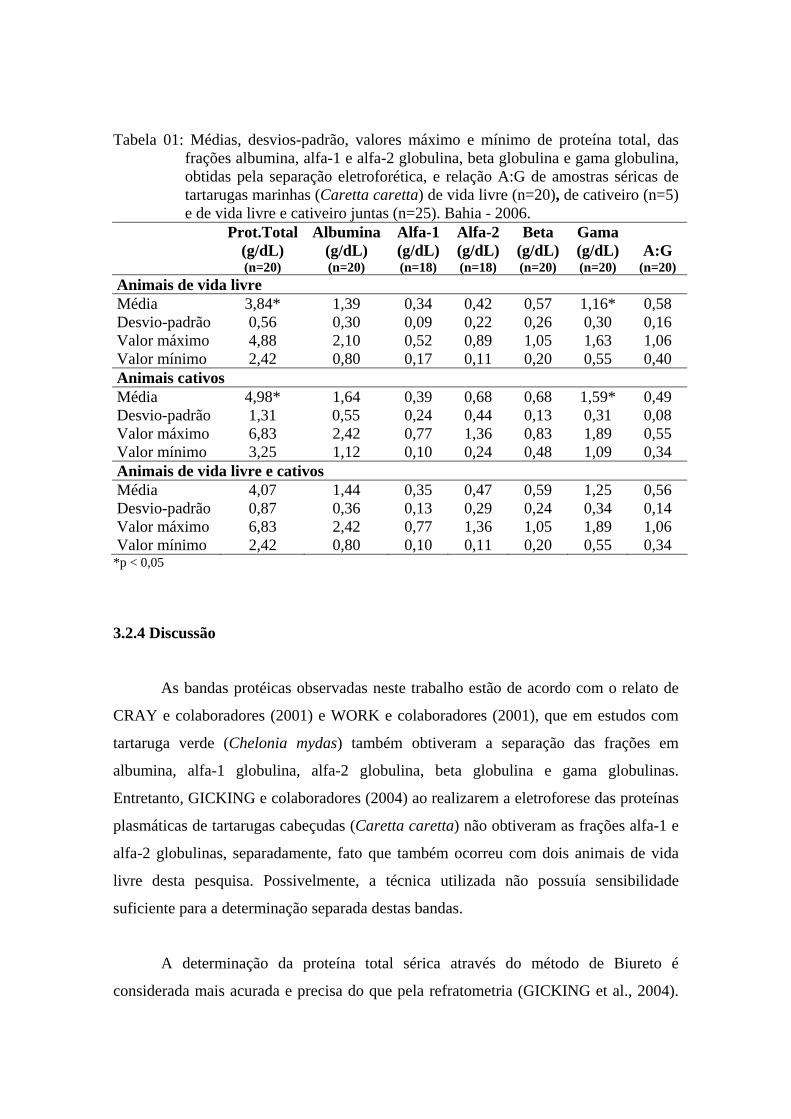
Tabela 01: Médias, desvios-padrão, valores máximo e mínimo de proteína total, das
frações albumina, alfa-1 e alfa-2 globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=20), de cativeiro (n=5) e de vida livre e cativeiro juntas (n=25). Bahia - 2006.
Prot.Total (g/dL) (n=20)
Albumina (g/dL) (n=20)
Alfa-1 (g/dL) (n=18)
Alfa-2 (g/dL) (n=18)
Beta (g/dL) (n=20)
Gama (g/dL) (n=20)
A:G (n=20)
Animais de vida livre Média 3,84* 1,39 0,34 0,42 0,57 1,16* 0,58 Desvio-padrão 0,56 0,30 0,09 0,22 0,26 0,30 0,16 Valor máximo 4,88 2,10 0,52 0,89 1,05 1,63 1,06 Valor mínimo 2,42 0,80 0,17 0,11 0,20 0,55 0,40 Animais cativos Média 4,98* 1,64 0,39 0,68 0,68 1,59* 0,49 Desvio-padrão 1,31 0,55 0,24 0,44 0,13 0,31 0,08 Valor máximo 6,83 2,42 0,77 1,36 0,83 1,89 0,55 Valor mínimo 3,25 1,12 0,10 0,24 0,48 1,09 0,34 Animais de vida livre e cativos Média 4,07 1,44 0,35 0,47 0,59 1,25 0,56 Desvio-padrão 0,87 0,36 0,13 0,29 0,24 0,34 0,14 Valor máximo 6,83 2,42 0,77 1,36 1,05 1,89 1,06 Valor mínimo 2,42 0,80 0,10 0,11 0,20 0,55 0,34
*p < 0,05
3.2.4 Discussão
As bandas protéicas observadas neste trabalho estão de acordo com o relato de
CRAY e colaboradores (2001) e WORK e colaboradores (2001), que em estudos com
tartaruga verde (Chelonia mydas) também obtiveram a separação das frações em
albumina, alfa-1 globulina, alfa-2 globulina, beta globulina e gama globulinas.
Entretanto, GICKING e colaboradores (2004) ao realizarem a eletroforese das proteínas
plasmáticas de tartarugas cabeçudas (Caretta caretta) não obtiveram as frações alfa-1 e
alfa-2 globulinas, separadamente, fato que também ocorreu com dois animais de vida
livre desta pesquisa. Possivelmente, a técnica utilizada não possuía sensibilidade
suficiente para a determinação separada destas bandas.
A determinação da proteína total sérica através do método de Biureto é
considerada mais acurada e precisa do que pela refratometria (GICKING et al., 2004).

Os valores obtidos neste estudo, de 3,84g/dL para as tartarugas de vida livre e 4,98g/dL
para as cativas, foram próximos àqueles publicados para tartaruga cabeçuda, de 4,1 ±
1,2 g/dL (BOLTEN et al., 1992), 4,1 ± 1,0 g/dL (BOLTEN et al., 1994), 4,4 ± 0,75 g/dL
(GICKING et al., 2004), 4,0 ± 0,8 g/dL (KELLER et al., 2004) e 4,2 ± 0,4 g/dL
(CUBAS & BAPTISTOTTE, 2006). Foi observada diferença significativa entre os
valores de proteína sérica total dos grupos estudados, onde os animais cativos
apresentaram valores mais elevados (4,98 ± 1,31 g/dL) que os animais de vida livre
(3,82 ± 0,57 g/dL). Muito possivelmente, esta diferença pode ser atribuída ao manejo
nutricional, pois sabe-se que tartarugas mantidas em cativeiro podem apresentar níveis
protéicos mais elevados, resultado da dieta rica em proteína oferecida (SWIMMER,
2000). Os animais mantidos em cativeiro no Centro de Visitantes do Projeto Tamar-
Ibama são alimentados diariamente com peixes (sardinha, galo e corvina), na proporção
de 1% do seu peso corporal.
Os valores encontrados para albumina, alfa-1 globulina, alfa-2 globulina e beta
globulina (Tab. 01) não apresentaram diferença significativa entre os grupos, entretanto
a fração gama globulinas, de 1,16g/dL para os animais de vida livre e 1,59g/dL para as
tartarugas de cativeiro, apresentou diferença significativa o que pode ser atribuído ao
estresse reprodutivo provocando uma leve imunodeficiência, já que apesar de mais
elevados, os valores obtidos para o grupo de animais de cativeiro são próximos aqueles
relatados pela literatura.
Os valores das frações protéicas encontrados no presente trabalho foram
inferiores aos obtidos por CRAY e colaboradores (2001), que em trabalho com 16
tartarugas verdes de cativeiro, apresentaram os seguintes valores: albumina 3,5 ± 0,7
g/dL, alfa-1 globulinas 0,4 ± 0,2 g/dL, alfa-2 globulinas 1,2 ± 0,3 g/dL, beta globulinas
1,1 ± 0,2 g/dL e gama globulinas 1,4 ± 0,3 g/dL, esta diferença pode ser devido ao uso
de outro método na determinação da proteína total, onde estes autores utilizaram a
refratometria. Ao comparar os valores encontrados neste estudo com aqueles obtidos
por GICKING e colaboradores (2004) para sete fêmeas adultas de tartarugas cabeçudas,
pode-se observar que os valores apresentados por estes autores foram levemente
inferiores para a albumina (0,97 ± 0,13 g/dL) e alfa globulinas (0,49 ± 0,05 g/dL),
enquanto que para beta globulinas (0,81 ± 0,14 g/dL) e gama globulinas (2,1 ± 0,64
g/dL) foram mais elevados. Vale salientar que estes autores utilizaram plasma em suas

análises, onde a presença do fibrinogênio, uma das proteínas de fase aguda que compõe
a fração beta globulina (KANEKO, 1997), pode interferir nos resultados.
A relação A:G obtida não apresentou diferença significativa entre os dois grupos
e foi maior que aquelas relatadas em estudos com tartarugas cabeçudas de 0,30 ± 0,62,
para fêmeas adultas (GICKING et al., 2004), 0,3 ± 0,1 (BOLTEN et al., 1994), em
estudo com animais em diferentes fases de vida, e 0,4 ± 0,08 (KELLER et al., 2004)
para juvenis desta espécie. Estas diferenças podem ser devido a diferenças nas técnicas
e pela utilização de plasma, enquanto que neste trabalho utilizou-se soro.
3.2.5 Conclusão
De acordo com o protocolo experimental utilizado nesta pesquisa e com base
nos resultados obtidos, pode-se concluir que o padrão eletroforético de proteínas de
tartarugas Caretta caretta de vida livre e de cativeiro são semelhantes.

3.2.6 Referências Bibliográficas
BAPTISTOTTE, C.; THOMÉ, J.C.A.; BJORNDAL, K.A. Reproductive biology and conservation status of the loggerhead sea turtle (Caretta caretta) in Espírito Santo State, Brazil. Chelonian Conservation and Biology, Washington, v. 4, n. 3, p. 523-529, 2003. BOLTEN, A. B.; BJORNDAL, K. A. Blood profiles for a wild population of green turtles (Chelonia mydas) in the southern Bahamas: size-espcific and sex-specific relationships. Journal of Wildlife Diseases, Ames, v. 28, n. 3, p. 407 - 413, 1992. BOLTEN, A. B.; BJORNDAL, K. A.; ELIAZAR, P.J.; GREGORY, L.F. Seasonal abundance, size distribution, and blood biochemical values of loggerheads (Caretta caretta) in Port Canaveral Ship Channel, Florida. Miami: NOAA, 1994. 39p. (NOAA Technical Memorandum NMFS-SEFSC, 353). BOLTEN, A. B.; JACOBSON, E. R.; BJORNDAL, K. A. Effects of anticoagulant and autoanalyzer on blood biochemical values of loggerhead sea turtle (Caretta caretta). American Journal of Veterinary Research, Schaumburg , v. 53, n. 12, p. 2224 - 2227, 1992. CASAL, A.B.; ORÓS, J. Morphologic and cytochemical characteristic of blood cells of juvenile loggerhead sea turtles (Caretta caretta). Research in Veterinary Science, London, v. 82, n. 2, p. 158-165, 2007. CRAY, C.; TATUM, L.M. Applications of protein electrophoresis in avian diagnostics. Journal of Avian Medicine and Surgery, Bedford, v. 12, n. 1, p. 4-10, 1998. CRAY, C.; VARELLA, R.; BOSSART, G. D.; LUTZ, P. Altered in vitro immune responses in green turtles (Chelonia mydas) with fibropapillomatosis. Journal of Zoo and Wildlife Medicine, Lawrence, v. 32, n. 4, p. 436-440, 2001. CUBAS, P.H.; BAPTISTOTTE, C. Chelonia (tartaruga, cágado e jabuti), In: CUBAS, Z.S.; SILVA, J.C.R.; CATÃO-DIAS, J.L. Tratado de Animais Selvagens: Medicina Veterinária. São Paulo: Roca, 2006. p. 86-119. D’AMATO, A.F.; MARCOVALDI, A.M. Aspectos da biologia de tartarugas amerinhas (Testudenes: Chelonidae) na região de Praia do Forte Bahia. XI Encontro de Zoologia do Nordeste, 1997, Fortaleza. Resumos do XI Encontro de Zoologia do Nordeste, 1997. v. 1. p. 54-55. GICKING, J.C.; FOLEY, A.M.; HARR, K.E.; RASKIN, R.E.; JACOBSON, E. Plasma protein electrophoresis of the Atlantic loggerhead sea turtles, Caretta caretta. Journal of Herpetological Medicine and Surgery, v. 14, n. 3, 2004. IUCN. 2006 IUCN Red List of Threatened Species. Disponível em: <http://www.iucnredlist.org>. Acesso em 12 April 2007.

KANEKO, J.J. Serum proteins and dysproteinemias. In: KANEKO, J.J.; HARVEY, J.W.; BRUSS, M.L. Clinical Biochemistry of Domestic Animals. 5 ed. San Diego: Academic Press, 1997. p. 117-138. KELLER, J. M., KUCKLICK, J. R., STAMPER, M. A., HARMS, C. A., MCCLELLAN-GREEN, P. D. Associations between organochlorine contaminant concentrations and clinical health parameters in loggerhead sea turtles from North Carolina, USA. Environmental Health Perspectives, v. 112, n. 10, p. 1074-1079, 2004. MANN, H.B.; WHITNEY, D.R. On a test of whether one of two random variables is statistically larger than the other. Ann. Math. Stat., v. 18, p.50-60, 1947. MARCOVALDI, M.A.; MARCOVALDI, G.G. Marine turtles of Brazil: the history and structure of projeto Tamar-Ibama. Biological conservation, Essex, n. 91, p.35-41, 1999. OWENS, D. W.; RUIZ G. J. New methods of obtaining blood and cerebrospinal fluid from marine turtles. Herpetologica, Johnson City, v. 36, n.1, p.17 - 20, 1980. SWIMMER, J.Y. Biochemical responses to fibropapilloma and captivity in the green turtles. Journal of Wildlife Diseases, Ames, v. 36, n. 1, p. 102-110, 2000. WORK, T.M.; RAMEYER, R.A.; BALAZS, G.H.; CRAY, C.; CHANG, S.P. Immune status of free-ranging green turtles with fibropapillomatosis from Hawaii. Journal of Wildlife Diseases, Ames, v. 37, n. 3, p. 574-581, 2001. ZAIAS, J.; CRAY, C. Protein Electrophoresis: A Tool for the Reptilian and Amphibian Practitioner Journal of Herpetological Medicine and Surgery, v. 12, n. 1, p.30-32. 2002.

4 CONSIDERAÇÕES GERAIS
Os resultados deste estudo, nas condições em que foi realizado, permitem as seguintes
considerações:
Não houve diferença entre os grupos estudados quanto à maioria dos parâmetros
avaliados.
Os dados obtidos para tartarugas cabeçudas fêmeas de vida livre em estágio
reprodutivo constituem valores de normalidade para esta espécie nesta fase,
constituindo-se em uma importante ferramenta para auxiliar na avaliação clínica e
reabilitação de tartarugas selvagens que venham a encalhar em nosso litoral.
Os valores obtidos para o hemograma, alguns parâmetros bioquímicos séricos e
eletroforese, para as tartarugas cabeçudas mantidas no Centro de Visitantes do Projeto
Tamar-Ibama na Praia do Forte, podem ser considerados de normalidade, auxiliando no
monitoramento da saúde das tartarugas marinhas mantidas em cativeiro no litoral do
estado da Bahia, e outras instituições que mantenham está espécie em seus plantéis.
Outros estudos sobre o tema são necessários para que se possa determinar
intervalos de referência para estas variáveis hematológicas, em diferentes fases de vida
e condições ambientais, fortalecendo assim a avaliação sangüínea como meio
diagnóstico para as tartarugas cabeçudas.

5 REFERÊNCIAS BIBLIOGRÁFICAS
AGUIRRE, A, A; BALAZS, G.H. Blood biochemistry values of green turtles, Chelonia mydas, with and without fibropapillomatosis. Comparative Haematology International, London, v. 10, p. 132-137, 2000. AGUIRRE, A. A.; BALAZS, G. H.; SPRAKOR, T. R.; GROSS, T. S. Adrenal and hematological responses to stress in juvenile green turtles, Chelonia mydas, with and without fibropapillomas. Physiological Zoology, Chicago, v. 68, n. 5, p. 831 - 854, 1995. ALKINDI, A.Y.A.; MAHMOUD, I.Y. Hematological survey in two species of sea turtles in the Arabian sea during nesting season. Pakistan Journal of Biological Sciences, v. 5, n. 3, p. 359-361, 2002. ALLEMAN, A. R.; JACOBSON, E. R.; RASKIN, R. E. Morphologic and cytochemical characteristics of blood cells from desert tortoise (Gopherus agassizii). American Journal of Veterinary Research, Schaumburg, v.53, n. 9, p. 1645 - 1651, 1992. AZEVEDO, A.; LUNARDI, L. O. Cytochemical characterization of eosinophilic leukocytes circulating in blood of the turtles (Chrysemys dorbignih). Acta Histochemica, Jena, v. 105, n. 1, p. 99-105, 2003. BALDASSIN, J.P. Perfil hematológico e bioquímico das tartarugas marinhas mantidas em cativeiro na base do Projeto Tamar em Ubatuba. 2003. 58f. Monografia (Graduação em Medicina Veterinária)-Universidade Paulista. Instituto de Ciências da Saúde, São Paulo, 2003. BAPTISTOTTE, C.; THOMÉ, J.C.A.; BJORNDAL, K.A. Reproductive biology and conservation status of the loggerhead sea turtle (Caretta caretta) in Espírito Santo State, Brazil. Chelonian Conservation and Biology, Washington, v. 4, n. 3, p. 523-529, 2003.
BIRGEL, E. H.; LARSSON, M. H. A.; HAGIWARA, M. K.; VASCONSELOS, S. A.; LARSSON, C. E.; OGASSAWARA, S. BENESI, F. J. Patologia Clínica Veterinária. São Paulo, 1983. 260p. BOLTEN, A. B.; BJORNDAL, K. A. Blood profiles for a wild population of green turtles (Chelonia mydas) in the southern Bahamas: size-specific and sex-specific relationships. Journal of Wildlife Diseases, Ames, v. 28, n. 3, p. 407 - 413, 1992. BOLTEN, A. B.; BJORNDAL, K. A.; ELIAZAR, P.J.; GREGORY, L.F. Seasonal abundance, size distribution, and blood biochemical values of loggerheads (Caretta caretta) in Port Canaveral Ship Channel, Florida. Miami: NOAA, 1994. 39p. (NOAA Technical Memorandum NMFS-SEFSC, 353).

BOLTEN, A. B.; JACOBSON, E. R.; BJORNDAL, K. A. Effects of anticoagulant and autoanalyzer on blood biochemical values of loggerhead sea turtle (Caretta caretta). American Journal of Veterinary Research, Schaumburg , v. 53, n. 12, p. 2224 - 2227, 1992. CAMPBELL, T. W. Clinical pathology. In: MADER, D. R. Reptile Medicine and Surgery. Philadelphia: W. B. Saunders Company, 1996. p. 248 - 257. CANNON, M. S. The morphogy and cytochemistry of blood leukocytes of Kemp’s sea turtles (Lepidochelys kempi). Can. J. Zoo., v. 70, n. 7, p. 1336 - 1340, 1992. CASAL, A.B.; ORÓS, J. Morphologic and cytochemical characteristic of blood cells of juvenile loggerhead sea turtles (Caretta caretta). Research in Veterinary Science, London, v. 82, n. 2, p. 158-165, 2007. COLES, E. H. Patologia Clínica Veterinária. 3. ed. São Paulo: Manole, 1984. 566p. CRAY, C.; TATUM, L.M. Applications of protein electrophoresis in avian diagnostics. Journal of Avian Medicine and Surgery, Bedford, v. 12, n. 1, p. 4-10, 1998. CRAY, C.; VARELLA, R.; BOSSART, G. D.; LUTZ, P. Altered in vitro immune responses in green turtles (Chelonia mydas) with fibropapillomatosis. Journal of Zoo and Wildlife Medicine, Lawrence, v. 32, n. 4, p. 436-440, 2001. CUBAS, P.H.; BAPTISTOTTE, C. Chelonia (tartaruga, cágado e jabuti), In: CUBAS, Z.S.; SILVA, J.C.R.; CATÃO-DIAS, J.L. Tratado de Animais Selvagens: Medicina Veterinária. São Paulo: Roca, 2006. p. 86-119. D’AMATO, A.F.; MARCOVALDI, A.M. Aspectos da biologia de tartarugas marinhas (Testudenes: Chelonidae) na região de Praia do Forte Bahia. XI Encontro de Zoologia do Nordeste, 1997, Fortaleza. Resumos do XI Encontro de Zoologia do Nordeste, 1997. v. 1. p. 54-55. DAVENPORT, J. Temperature and the life-history strategies of sea turtles. Journal of Thermal Biology. v.22, n. 6, pp.479-488, 1997. DIVERS, S. J.; REDMAYNE, G.; AVES, K. Haematological and biochemical of 10 green iguanas (Iguana iguana). The Veterinary Record, London, v. 138, n. 3, p. 203 - 205, 1996. FRAIR, W. Serum electrophoresis and sea turtle classification. Comparative Biochemistry and Physiology, New York, v. 72B, p. 1 -5,1982. _____. Turtles red blood cell packed volumes, sizes, and numbers. Herpetologica, Johnson City, v. 33, n.2, p. 167 - 190, 1977a. _____. Sea turtle red blood cell parameters correlated with carapace lengths. Comparative Biochemistry and Physiology, Oxford, v. 56A, n. 4, p. 467 - 472, 1977b.

FRAIZER, J. Generalidades de la historia de vida de las tortugas marinas. In: CONSERVACIÓN DE TORTUGAS MARINAS EN LA REGIÓN DEL GRAN CARIBE – UN DIÁLOGO PARA EL MANEJO REGINAL EFECTIVO, 1999, Santo Domingo. Memorias...Santo Domingo:[s.n], 2001. v. 1. p. 3 - 18. FRAZIER, J. G. Community-based conservation. In: ECKERT, K.L.; BJORNDAL, K. A.; ABREU-GROBOIS, F.A.; DONNELLY, M. Research and Management Techniques for the Conservation of Sea Turtles. Gland : IUCN, 1999, p. 15-18 (IUCN/SSC Marine Turtles Specialist Group Publication, n. 4). GEORGE, R.H. Health problems and diseases of sea turtles. In: LUTZ, P.L.; MUSICK, J. A. The Biology of Sea Turtles. Boca Raton: CRC Press, 1997. p. 363-385. GICKING, J.C.; FOLEY, A.M.; HARR, K.E.; RASKIN, R.E.; JACOBSON, E. Plasma protein electrophoresis of the Atlantic loggerhead sea turtles, Caretta caretta. Journal of Herpetological Medicine and Surgery, v. 14, n. 3, 2004. GOTTDENKER, N. L.; JACOBSON, E. R. Effect of venipuncture sites on hematologic and clinical biochemical values in desert tortoises (Gopherus agassizii). American Journal of Veterinary Research, Schaumburg, v.56, n. 1, p. 19 - 21, 1995. GREGORY, L.F.; SCHMID, J.R. Stress responses and sexing of wild kemp’s ridley sea turtles (Lepidochelys kempii) in the northeastern Gulf of Mexico. General and Comparative Endocrinology, v. 124, n. 1, p. 66-74, 2001. HARMS, C.A.; KELLER, J.M.; KENNEDY-STOSKOPF, S. Use os a two-step percoll® gradient for separation os loggerhead sea turtle peripheral blood mononuclear cells. Journal of Wildlife Diseases, Ames, v. 36, n. 3, p. 535-540, 2000. HERBST, L.H.; JACOBSON, E.R. Practical approaches for studying sea turtle health and disease. In: LUTZ, P.L.; MUSICK, J.A.; WYNEKEN, J. The Biology of Sea Turtles. Boca Raton: CRC Press, 2003. v. 2, p. 384-410. IUCN. 2006 IUCN Red List of Threatened Species. Disponível em: <http://www.iucnredlist.org>. Acesso em 12 April 2007. JACOBSON, E. R. Collecting and processing blood from sea turtles for hematologic and plasma biochemical determinations. In: REPORT OF THE SEA TURTLE HEALTH ASSESSMENT WORKSHOP, 1998, Charleston, Memorandum... Charleston:[s.n.], 1998. p. 24 - 31. JAIN, N.C. Schalm’s veterinary hematology. 4 ed. Philadelphia: Lea & Febiger, 1986. 1221p. KANEKO, J.J. Serum proteins and dysproteinemias. In: KANEKO, J.J.; HARVEY, J.W.; BRUSS, M.L. Clinical Biochemistry of Domestic Animals. 5 ed. San Diego: Academic Press, 1997. p. 117-138.

KELLER, J. M., KUCKLICK, J. R., STAMPER, M. A., HARMS, C. A., MCCLELLAN-GREEN, P. D. Associations between organochlorine contaminant concentrations and clinical health parameters in loggerhead sea turtles from North Carolina, USA. Environmental Health Perspectives, v. 112, n. 10, p. 1074-1079, 2004. KNOTKOVÁ, Z.; DOUBEK, J.; KNOTEK, Z.; HÁJKOVÁ P. Blood cell morphology and plasma biochemistry in Russian tortoises (Agrionemys horsfieldi). Acta Veterinaria Brno, Brno, v. 71, n. 2, p. 191-198. 2002. LUTCAVAGE, M.E.; LUTZ, P.L.; BOSSART, G.D.; HUDSON, D.M. Physiologic and clinicopathologic effects of crude oil on loggerhead sea turtles. Archives of Environmental Contamination and Toxicology, New York, v. 28, n. 4, p. 417-422, 1995. LUTZ, P. L.; DUNBAR-COOPER, A. Variations in the blood chemistry of the loggerhead sea turtle, Caretta caretta. Fishery Bulletin, Seattle, v. 85, n. 1, p. 37 - 43, 1987. MANN, H.B.; WHITNEY, D.R. On a test of whether one of two random variables is statistically larger than the other. Ann. Math. Stat., v. 18, p.50-60, 1947. MARCOVALDI, M. A.; LAURENT, A. A six season study of marine turtle nesting at Praia do Forte, Bahia, Brazil, with implications for conservation and management. Chelonian Conservation and Biology, v. 2, n. 1, p. 55-59, 1996. MARCOVALDI, M.A.; MARCOVALDI, G.G. Marine turtles of Brazil: the history and structure of Projeto Tamar-Ibama. Biological conservation, Essex, n. 91, p.35-41, 1999. MARCOVALDI, M.A.; VIEITAS, C. F.; GODFREY, M. H. Nesting and conservation management of hawksbill turtles (Eretmochelys imbricata) in northern Bahia, Brazil. Chelonian Conservation and Biology, v. 3, n. 2, p. 301-307, 1999. MARKS, S. K.; CITINO, S. B. Hematology and serum chemistry of the radiated tortoise (Testudo radiata). Journal of Zoo and Wildlife Medicine, Lawrence, v. 21, p. 342 - 344, 1990. MARQUEZ, R. Sea turtles of the world. FAO Species Catologue: Sea turtles of the world. Rome, p.69, 1990. (FAO Fisheries Synopsis (FAO), n.125, v.11). MEYER, D.J.; COLES, E.H; RICH L.J. Medicina de Laboratório Veterinária: Interpretação e diagnóstico. São Paulo: Roca, 1995. 308p. MONTALI, R.J. Comparative pathology of inflammation in the higher vertebrates (reptiles, birds, mammals). Journal of Comparative Pathology, v. 99, p. 1 - 21, 1988. NATT, M. P.; HERRICK, G. A. A new blood diluent for counting erythrocytes and leukocytes of the chicken. Poultry Science, v. 31, p. 735 - 738, 1952.

NORTON, T.M., JACOBSON, E.R., SUNDBERG, J.P. Cutaneous fibropapillomas and renal mixofibroma in a Green turtles, Chelonia mydas. Journal of wildlife diseases, Ames, v. 26, n. 2, p. 265-270, 1990. OWENS, D. W.; RUIZ G. J. New methods of obtaining blood and cerebrospinal fluid from marine turtles. Herpetologica, Johnson City, v. 36, n.1, p.17 - 20, 1980. PAGE, C. D.; MAUTINO, M. Clinical management of tortoise. Compendium on Continuing Education for the Practicing Veterinarian, Montreal, v. 12, n. 2, p. 221 - 228, 1990. PIRES, T.T.; ROSTAN, G.; GUIMARÃES, J.E. Hemograma e determinação da proteína plasmática total de tartarugas marinhas da espécie Caretta caretta (Linnaeus, 1758), criada em cativeiro, Praia do Forte, Município de Mata de São João – Bahia. Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 43, n. 3, p. 348-353, 2006. ROSS, J. P. La crianza y reproducción en cautiverio de tortugas marinas: Una evaluación de su uso como estrategia de conservación. In: ECKERT, K.A. et al. Técnicas de Investigación y Manejo para la Conservación de las Tortugas Marinas. Washington, D.C.: SSC/IUCN Marine Turtle Specialist Group, 2000. 278 p. (IUCN/SSC Marine Turtle Specialist Group, 4). ROSSKOPF, W. J. Normal hemogram and blood chemistry values for California desert tortoises. Veterinary Medicine/ Small Animal Clinician, Book Stacks, v.77, n. 1, p. 85 - 87, 1982. SAMOUR, H. J.; HOWLETT, J.C.; SILVANOSE, C.; HASBUN, C.R.; AL-GHAIS, S.M. Normal haematology of free-living green turtles (Chelonia mydas) from the United Arab Emirates. Comparative Haematology International, London, v. 8, n. 2, p. 102-107, 1998. SANTOS, A. S.; MARCOVALDI, M. A.; GODFREY, M.H. Update on the nesting population of loggerhead sea turtles monitored by Projeto Tamar-Ibama, Praia do Forte, Bahia, Brazil. Marine Turtles Newsletter, n. 89, p. 8-11, 2000. SOSLAU, G.; PREST, P. J.; CLASS, R.; GEORGE, R.; PALADINO, F.; VIOLETTA, G. Comparison of sea turtles thrombocyte aggregation to human platelet aggregation in whole blood. Comparative Biochemistry and physiology, Part B, New York, v. 142, p. 353-360, 2005. SOUTHWOOD, A.L.; DARVEAU, C.A.; JONES, D.R. Metabolic and cardiovascular adjustments of juvenile green turtles to seasonal changes in temperature and photoperiod. The Journal Experimental Biology, Cambridge, v. 206, n. 24, p. 4521-4531, 2003. SWIMMER, J.Y. Biochemical responses to fibropapilloma and captivity in the green turtles. Journal of Wildlife Diseases, Ames, v. 36, n. 1, p. 102-110, 2000.

TROËNG, S.; DREWS, C. Moneys Talks: Economic aspects of marine turtles use and conservation. WWF-International: Glande, 2004. 62 p. Disponível em: <http://www.panda.org>. Acesso em 25 de março de 2006. WOOD, F. E.; EBANKS, G. Blood cytology and hematology of the green sea turtle, Chelonia mydas.Herpetologica, Johnson City, v. 40, n.3, p.331 - 336, 1984. WORK, T.M.; BALAZS, G.H. Relating tumor score to hematology in green turtles with fibropapillomatosis in Hawaii. Journal of Wildlife Diseases, Ames, v. 35, n. 4, p. 804-807, 1999. WORK, T.M.; RAMEYER, R.A.; BALAZS, G.H.; CRAY, C.; CHANG, S.P. Immune status of free-ranging green turtles with fibropapillomatosis from Hawaii. Journal of Wildlife Diseases, Ames, v. 37, n. 3, p. 574-581, 2001. WORK, T. M.; RASKIN R. E.; BALAZS, G. H.; WHITTAKER, S. D. Morphologic and cytochemical characteristics of blood cells from Hawaiian green turtles. American Journal of Veterinary Research, Schaumburg, v.59, n. 10, p. 1252 - 1257, 1998. WHITAKER, B. R.; KRUM, H. Medical management of sea turtles in aquaria. In: FOWLER, M. E.; MILLER, R. E. Zoo & Wild animal medicine. v.4. Philadelphia: W. B. Saundes Company, 1999. p. 217 - 231. ZAIAS, J.; CRAY, C. Protein Electrophoresis: A Tool for the Reptilian and Amphibian Practitioner Journal of Herpetological Medicine and Surgery, v. 12, n. 1, p.30-32. 2002.

ANEXOS
Figura 01 - Exemplar de tartaruga cabeçuda (Caretta caretta)

Figura 02 - Coleta de sangue no seio cervical dorsal de tartaruga marinha
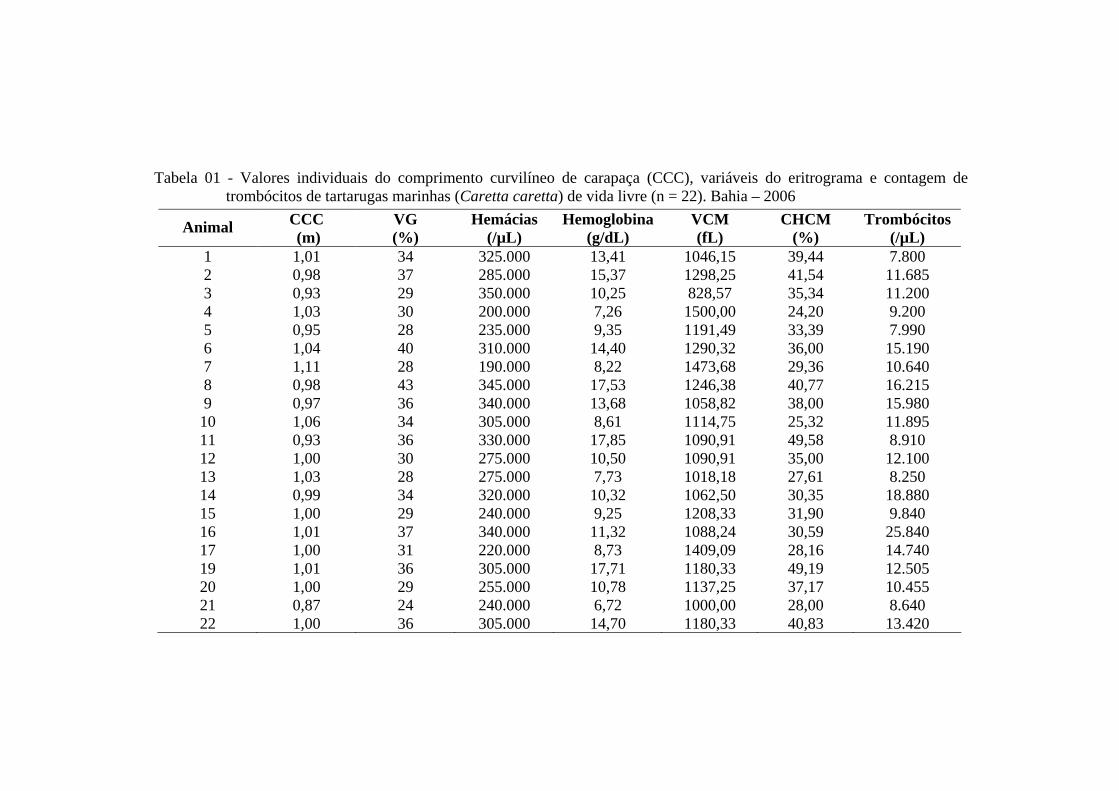
Tabela 01 - Valores individuais do comprimento curvilíneo de carapaça (CCC), variáveis do eritrograma e contagem de
trombócitos de tartarugas marinhas (Caretta caretta) de vida livre (n = 22). Bahia – 2006 CCC
(m) VG (%)
Hemácias (/µL)
Hemoglobina (g/dL)
VCM (fL)
CHCM (%)
Trombócitos (/µL) Animal
1 1,01 34 325.000 13,41 1046,15 39,44 7.800
2 0,98 37 285.000 15,37 1298,25 41,54 11.685
3 0,93 29 350.000 10,25 828,57 35,34 11.200
4 1,03 30 200.000 7,26 1500,00 24,20 9.200
5 0,95 28 235.000 9,35 1191,49 33,39 7.990
6 1,04 40 310.000 14,40 1290,32 36,00 15.190
7 1,11 28 190.000 8,22 1473,68 29,36 10.640
8 0,98 43 345.000 17,53 1246,38 40,77 16.215
9 0,97 36 340.000 13,68 1058,82 38,00 15.980
10 1,06 34 305.000 8,61 1114,75 25,32 11.895 11 0,93 36 330.000 17,85 1090,91 49,58 8.910 12 1,00 30 275.000 10,50 1090,91 35,00 12.100 13 1,03 28 275.000 7,73 1018,18 27,61 8.250 14 0,99 34 320.000 10,32 1062,50 30,35 18.880 15 1,00 29 240.000 9,25 1208,33 31,90 9.840 16 1,01 37 340.000 11,32 1088,24 30,59 25.840 17 1,00 31 220.000 8,73 1409,09 28,16 14.740 19 1,01 36 305.000 17,71 1180,33 49,19 12.505 20 1,00 29 255.000 10,78 1137,25 37,17 10.455
21 0,87 24 240.000 6,72 1000,00 28,00 8.640 22 1,00 36 305.000 14,70 1180,33 40,83
13.420
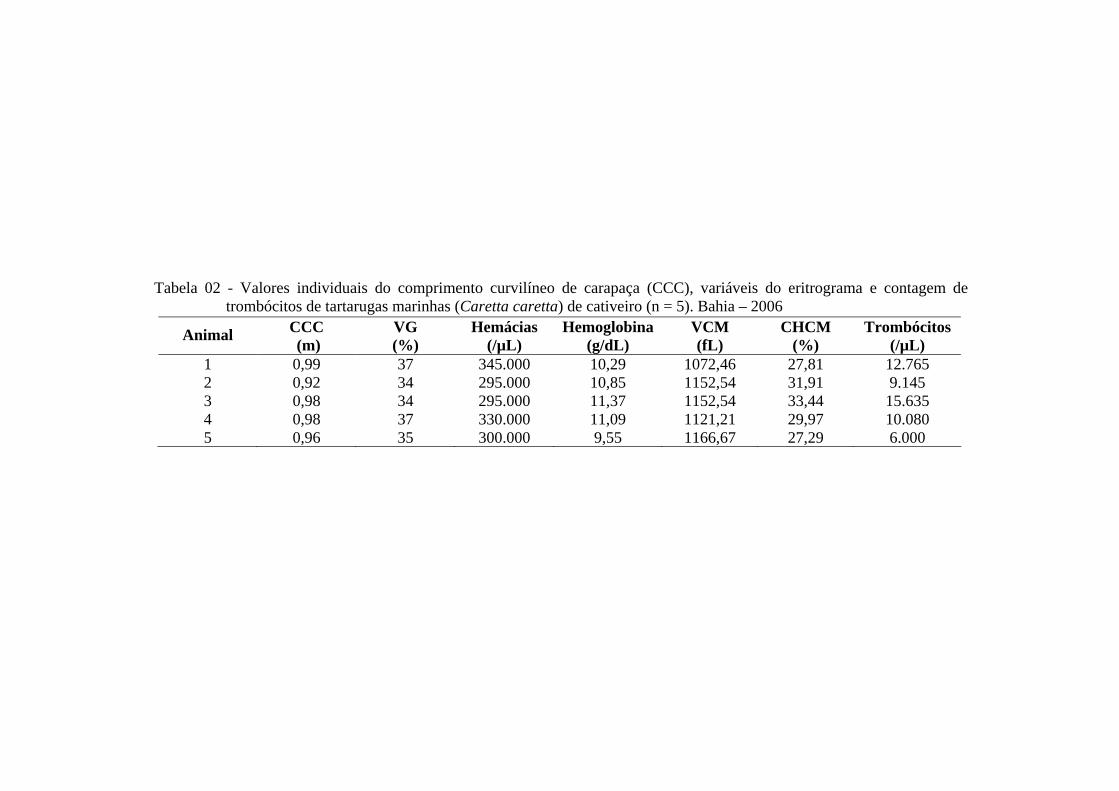
Tabela 02 - Valores individuais do comprimento curvilíneo de carapaça (CCC), variáveis do eritrograma e contagem de
trombócitos de tartarugas marinhas (Caretta caretta) de cativeiro (n = 5). Bahia – 2006 CCC
(m) VG (%)
Hemácias (/µL)
Hemoglobina (g/dL)
VCM (fL)
CHCM (%)
Trombócitos (/µL) Animal
1 0,99 37 345.000 10,29 1072,46 27,81 12.765
2 0,92 34 295.000 10,85 1152,54 31,91 9.145
3 0,98 34 295.000 11,37 1152,54 33,44 15.635
4 0,98 37 330.000 11,09 1121,21 29,97 10.080
5 0,96 35 300.000 9,55 1166,67 27,29 6.000
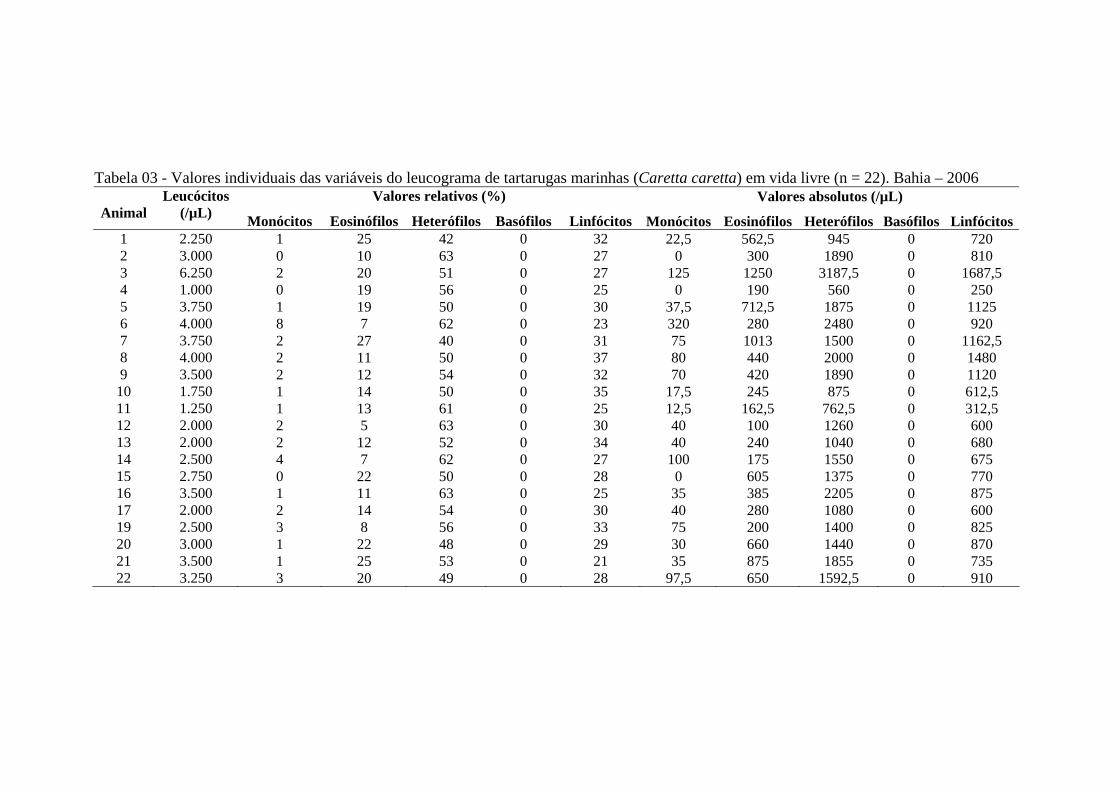
Tabela 03 - Valores individuais das variáveis do leucograma de tartarugas marinhas (Caretta caretta) em vida livre (n = 22). Bahia – 2006
Valores relativos (%) Valores absolutos (/µL) Animal
Leucócitos (/µL) Monócitos Eosinófilos Heterófilos Basófilos Linfócitos Monócitos Eosinófilos Heterófilos Basófilos Linfócitos
1 2.250 1 25 42 0 32 22,5 562,5 945 0 720 2 3.000 0 10 63 0 27 0 300 1890 0 810 3 6.250 2 20 51 0 27 125 1250 3187,5 0 1687,5 4 1.000 0 19 56 0 25 0 190 560 0 250 5 3.750 1 19 50 0 30 37,5 712,5 1875 0 1125 6 4.000 8 7 62 0 23 320 280 2480 0 920 7 3.750 2 27 40 0 31 75 1013 1500 0 1162,5 8 4.000 2 11 50 0 37 80 440 2000 0 1480 9 3.500 2 12 54 0 32 70 420 1890 0 1120
10 1.750 1 14 50 0 35 17,5 245 875 0 612,5 11 1.250 1 13 61 0 25 12,5 162,5 762,5 0 312,5 12 2.000 2 5 63 0 30 40 100 1260 0 600 13 2.000 2 12 52 0 34 40 240 1040 0 680 14 2.500 4 7 62 0 27 100 175 1550 0 675 15 2.750 0 22 50 0 28 0 605 1375 0 770 16 3.500 1 11 63 0 25 35 385 2205 0 875 17 2.000 2 14 54 0 30 40 280 1080 0 600 19 2.500 3 8 56 0 33 75 200 1400 0 825 20 3.000 1 22 48 0 29 30 660 1440 0 870 21 3.500 1 25 53 0 21 35 875 1855 0 735 22 3.250 3 20 49 0 28 97,5 650 1592,5 0 910
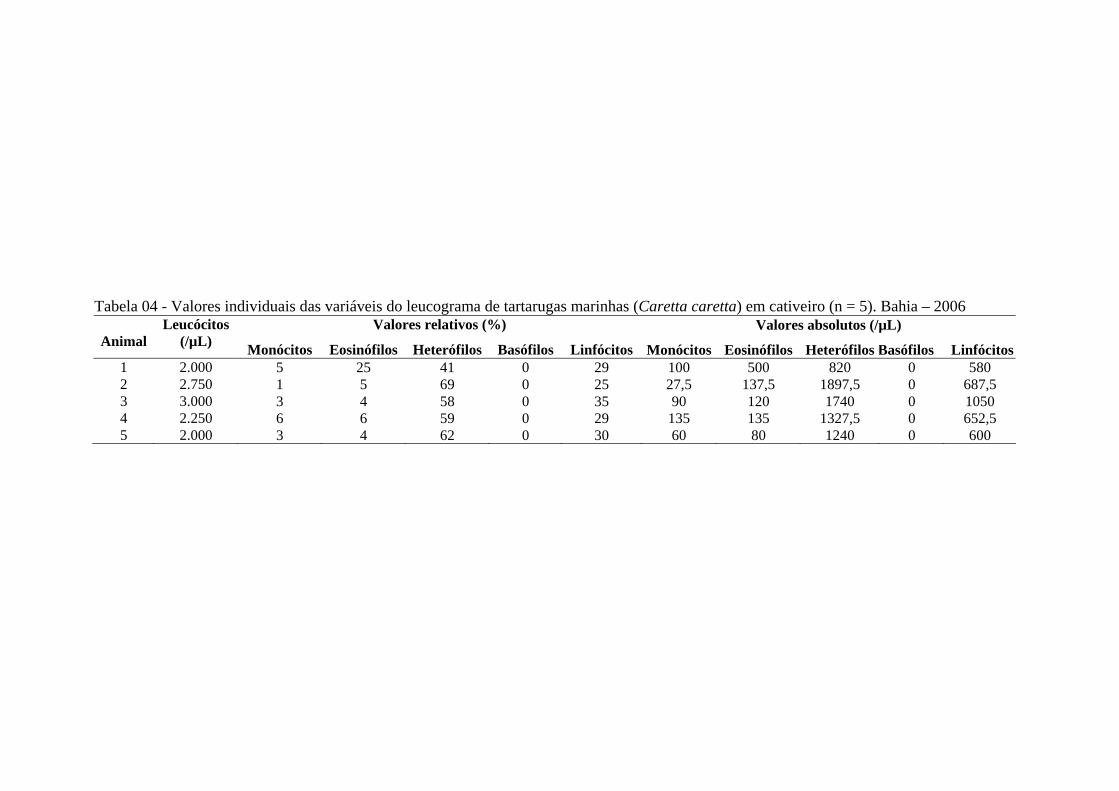
Tabela 04 - Valores individuais das variáveis do leucograma de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia – 2006 Valores relativos (%) Valores absolutos (/µL)
Animal Leucócitos
(/µL) Monócitos Eosinófilos Heterófilos Basófilos Linfócitos Monócitos Eosinófilos Heterófilos Basófilos Linfócitos 1 2.000 5 25 41 0 29 100 500 820 0 580 2 2.750 1 5 69 0 25 27,5 137,5 1897,5 0 687,5 3 3.000 3 4 58 0 35 90 120 1740 0 1050 4 2.250 6 6 59 0 29 135 135 1327,5 0 652,5 5 2.000 3 4 62 0 30 60 80 1240 0 600
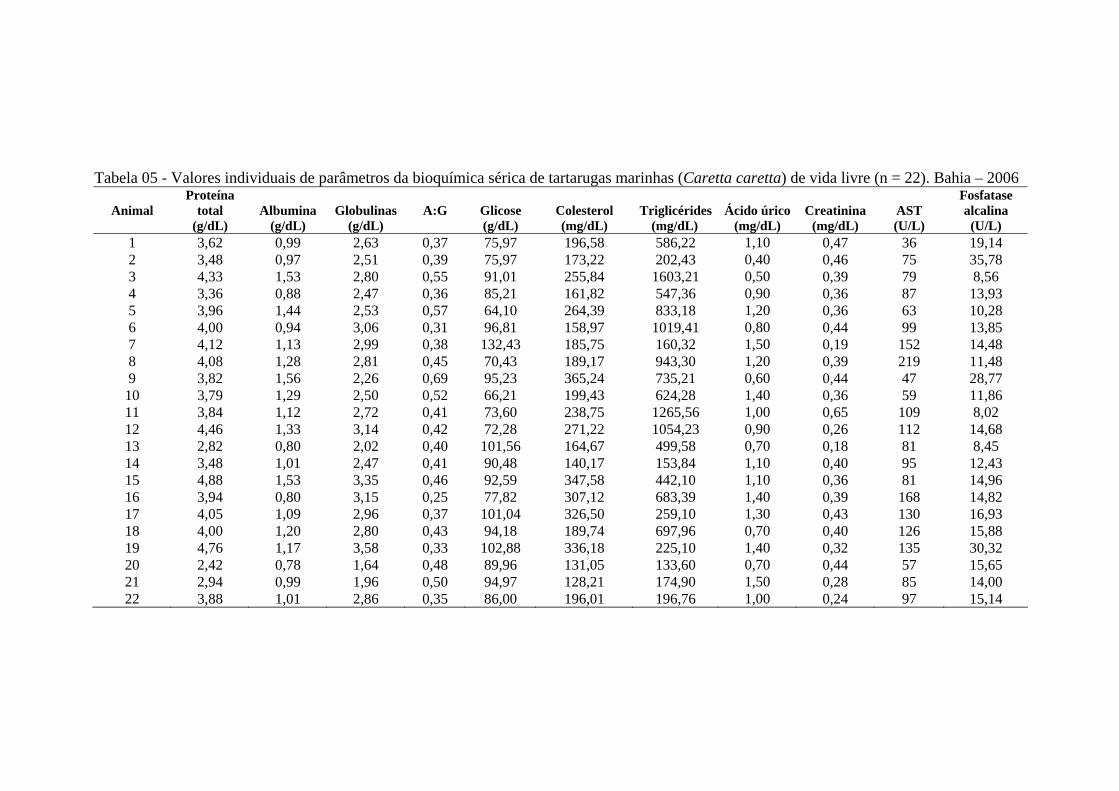
Tabela 05 - Valores individuais de parâmetros da bioquímica sérica de tartarugas marinhas (Caretta caretta) de vida livre (n = 22). Bahia – 2006
Animal
Proteína total
(g/dL) Albumina
(g/dL) Globulinas
(g/dL) A:G
Glicose (g/dL)
Colesterol (mg/dL)
Triglicérides (mg/dL)
Ácido úrico
(mg/dL) Creatinina
(mg/dL) AST (U/L)
Fosfatase alcalina (U/L)
1 3,62 0,99 2,63 0,37 75,97 196,58 586,22 1,10 0,47 36 19,14 2 3,48 0,97 2,51 0,39 75,97 173,22 202,43 0,40 0,46 75 35,78 3 4,33 1,53 2,80 0,55 91,01 255,84 1603,21 0,50 0,39 79 8,56 4 3,36 0,88 2,47 0,36 85,21 161,82 547,36 0,90 0,36 87 13,93 5 3,96 1,44 2,53 0,57 64,10 264,39 833,18 1,20 0,36 63 10,28 6 4,00 0,94 3,06 0,31 96,81 158,97 1019,41 0,80 0,44 99 13,85 7 4,12 1,13 2,99 0,38 132,43 185,75 160,32 1,50 0,19 152 14,48 8 4,08 1,28 2,81 0,45 70,43 189,17 943,30 1,20 0,39 219 11,48 9 3,82 1,56 2,26 0,69 95,23 365,24 735,21 0,60 0,44 47 28,77
10 3,79 1,29 2,50 0,52 66,21 199,43 624,28 1,40 0,36 59 11,86 11 3,84 1,12 2,72 0,41 73,60 238,75 1265,56 1,00 0,65 109 8,02 12 4,46 1,33 3,14 0,42 72,28 271,22 1054,23 0,90 0,26 112 14,68 13 2,82 0,80 2,02 0,40 101,56 164,67 499,58 0,70 0,18 81 8,45 14 3,48 1,01 2,47 0,41 90,48 140,17 153,84 1,10 0,40 95 12,43 15 4,88 1,53 3,35 0,46 92,59 347,58 442,10 1,10 0,36 81 14,96 16 3,94 0,80 3,15 0,25 77,82 307,12 683,39 1,40 0,39 168 14,82 17 4,05 1,09 2,96 0,37 101,04 326,50 259,10 1,30 0,43 130 16,93 18 4,00 1,20 2,80 0,43 94,18 189,74 697,96 0,70 0,40 126 15,88 19 4,76 1,17 3,58 0,33 102,88 336,18 225,10 1,40 0,32 135 30,32 20 2,42 0,78 1,64 0,48 89,96 131,05 133,60 0,70 0,44 57 15,65 21 2,94 0,99 1,96 0,50 94,97 128,21 174,90 1,50 0,28 85 14,00 22 3,88 1,01 2,86 0,35 86,00 196,01 196,76 1,00 0,24 97 15,14
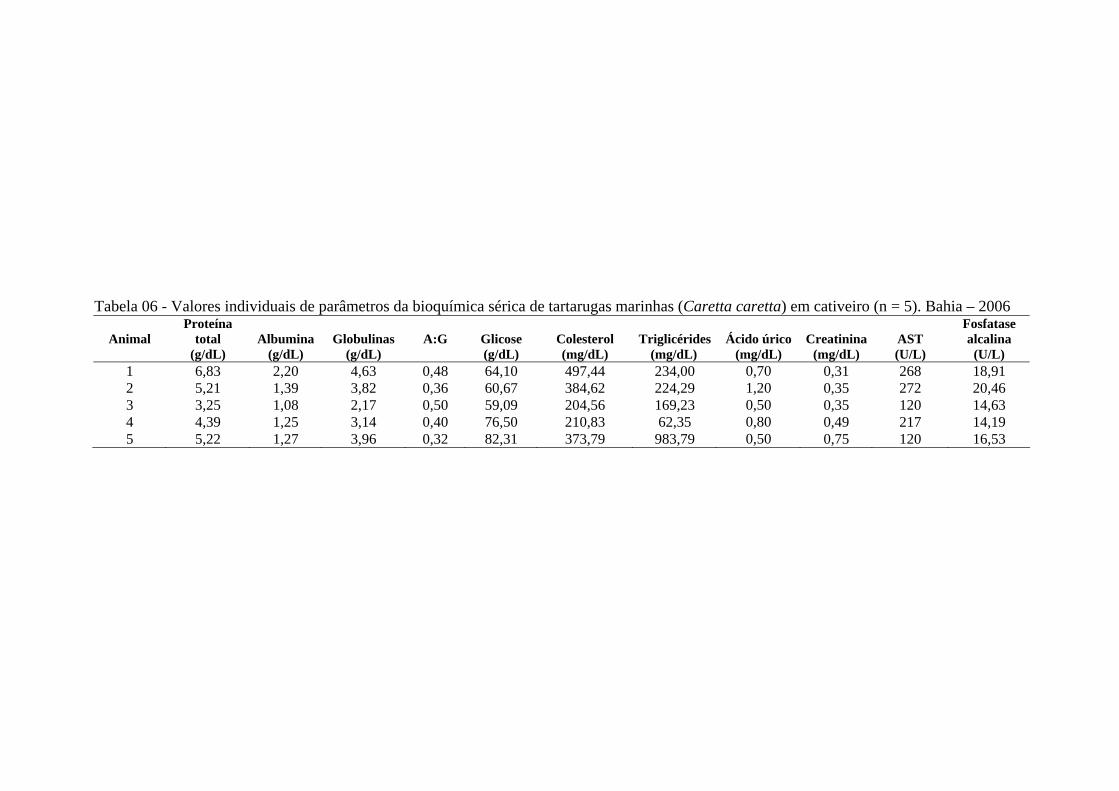
Tabela 06 - Valores individuais de parâmetros da bioquímica sérica de tartarugas marinhas (Caretta caretta) em cativeiro (n = 5). Bahia – 2006
Animal
Proteína total
(g/dL) Albumina
(g/dL) Globulinas
(g/dL) A:G
Glicose (g/dL)
Colesterol (mg/dL)
Triglicérides (mg/dL)
Ácido úrico
(mg/dL) Creatinina
(mg/dL) AST (U/L)
Fosfatase alcalina (U/L)
1 6,83 2,20 4,63 0,48 64,10 497,44 234,00 0,70 0,31 268 18,91 2 5,21 1,39 3,82 0,36 60,67 384,62 224,29 1,20 0,35 272 20,46 3 3,25 1,08 2,17 0,50 59,09 204,56 169,23 0,50 0,35 120 14,63 4 4,39 1,25 3,14 0,40 76,50 210,83 62,35 0,80 0,49 217 14,19 5 5,22 1,27 3,96 0,32 82,31 373,79 983,79 0,50 0,75 120 16,53
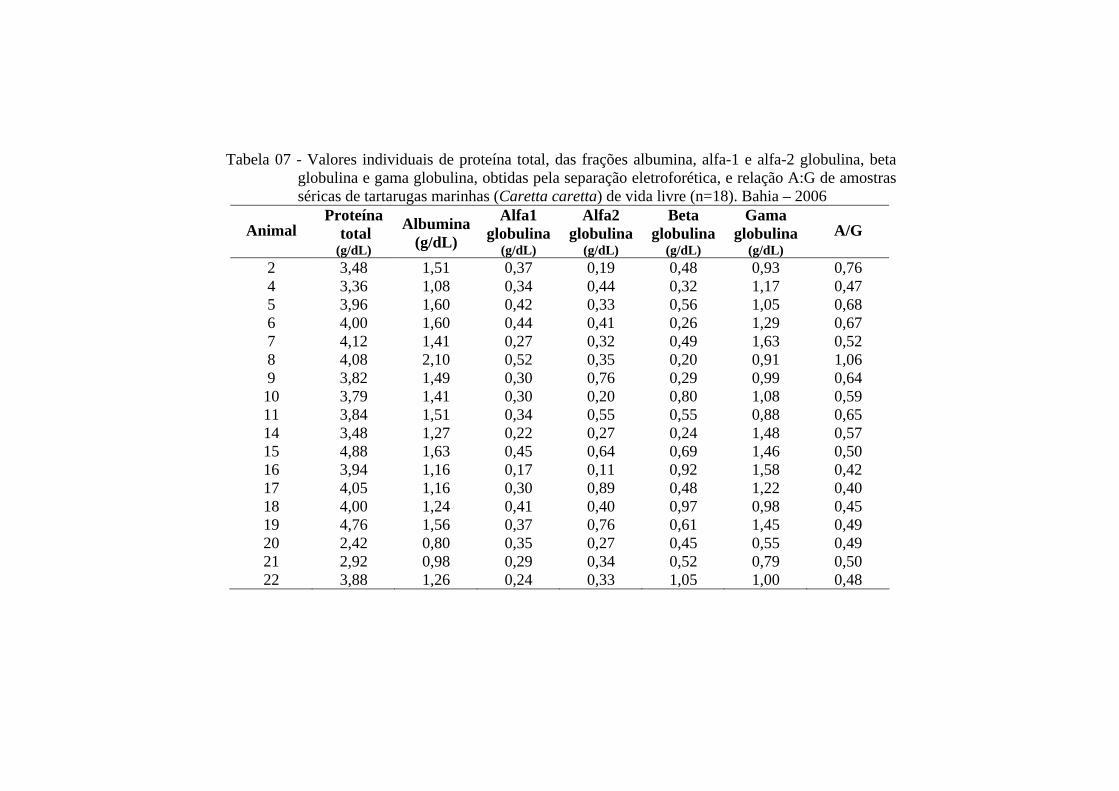
Tabela 07 - Valores individuais de proteína total, das frações albumina, alfa-1 e alfa-2 globulina, beta
globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=18). Bahia – 2006
Animal
Proteína total (g/dL)
Albumina (g/dL)
Alfa1 globulina
(g/dL)
Alfa2 globulina
(g/dL)
Beta globulina
(g/dL)
Gama globulina
(g/dL) A/G
2 3,48 1,51 0,37 0,19 0,48 0,93 0,76 4 3,36 1,08 0,34 0,44 0,32 1,17 0,47 5 3,96 1,60 0,42 0,33 0,56 1,05 0,68 6 4,00 1,60 0,44 0,41 0,26 1,29 0,67 7 4,12 1,41 0,27 0,32 0,49 1,63 0,52 8 4,08 2,10 0,52 0,35 0,20 0,91 1,06 9 3,82 1,49 0,30 0,76 0,29 0,99 0,64 10 3,79 1,41 0,30 0,20 0,80 1,08 0,59 11 3,84 1,51 0,34 0,55 0,55 0,88 0,65 14 3,48 1,27 0,22 0,27 0,24 1,48 0,57 15 4,88 1,63 0,45 0,64 0,69 1,46 0,50 16 3,94 1,16 0,17 0,11 0,92 1,58 0,42 17 4,05 1,16 0,30 0,89 0,48 1,22 0,40 18 4,00 1,24 0,41 0,40 0,97 0,98 0,45 19 4,76 1,56 0,37 0,76 0,61 1,45 0,49 20 2,42 0,80 0,35 0,27 0,45 0,55 0,49 21 2,92 0,98 0,29 0,34 0,52 0,79 0,50 22 3,88 1,26 0,24 0,33 1,05 1,00 0,48
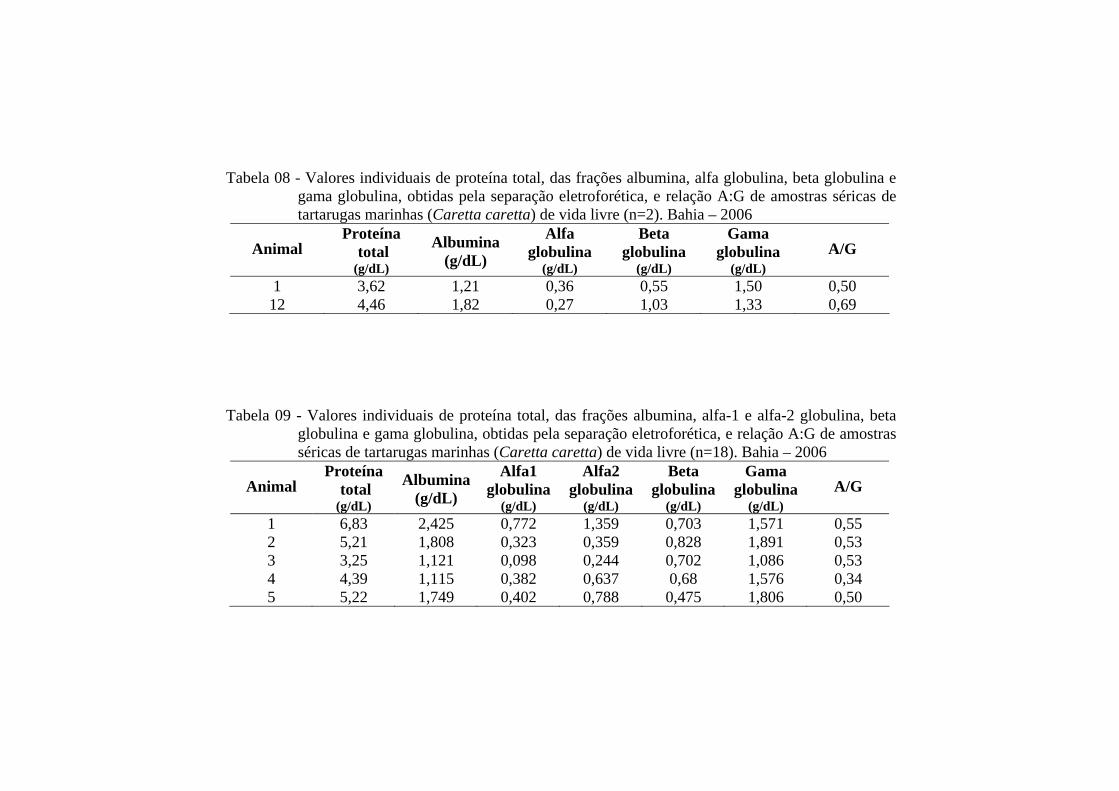
Tabela 08 - Valores individuais de proteína total, das frações albumina, alfa globulina, beta globulina e
gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=2). Bahia – 2006
Animal
Proteína total (g/dL)
Albumina (g/dL)
Alfa globulina
(g/dL)
Beta globulina
(g/dL)
Gama globulina
(g/dL) A/G
1 3,62 1,21 0,36 0,55 1,50 0,50 12 4,46 1,82 0,27 1,03 1,33 0,69
Tabela 09 - Valores individuais de proteína total, das frações albumina, alfa-1 e alfa-2 globulina, beta globulina e gama globulina, obtidas pela separação eletroforética, e relação A:G de amostras séricas de tartarugas marinhas (Caretta caretta) de vida livre (n=18). Bahia – 2006
Animal
Proteína total (g/dL)
Albumina (g/dL)
Alfa1 globulina
(g/dL)
Alfa2 globulina
(g/dL)
Beta globulina
(g/dL)
Gama globulina
(g/dL) A/G
1 6,83 2,425 0,772 1,359 0,703 1,571 0,55 2 5,21 1,808 0,323 0,359 0,828 1,891 0,53 3 3,25 1,121 0,098 0,244 0,702 1,086 0,53 4 4,39 1,115 0,382 0,637 0,68 1,576 0,34 5 5,22 1,749 0,402 0,788 0,475 1,806 0,50






























