Embed Size (px)
Citation preview

Về tác giả: 1. Họ và tên: Lê Đức Ngoan 2. Năm sinh: 05-01-1953 3. Học vị, học hàm: PGS.TS 4. Cơ quan công tác: khoa CNTY, trường đại học Nông Lâm, Đại học Huế 5. E-mail: [email protected]
Phạm vi và đối tượng sử dụng giáo trình: 1. Giáo trình cho các ngành đào tạo ở bậc đại học, cao đẵng các ngành: thủy sản, ngư
y.. 2. Loại hình trường sử dụng: Nông nghiệp, Thủy sản 3. Từ khóa: Dinh dưỡng thủy sản; Năng lượng; Protein và axit amin; Lipit và axit béo
thiết yếu; Carbohydrate và xơ; Nhu cầu dinh dưỡng; Lượng ăn vào; Khoáng thiết yếu; Khoáng vi lượng và đa lượng; Vitamin tan trong nước; Vitamin tan trong dầu; Tiêu hóa và hấp thu;
4. Yêu cầu kiến thức: phải học xong môn Sinh hóa động vật thủy sản, Sinh lý động vật thủy sản
5. Giáo trình đã được xuất bản 2008 tại Nhà xuất bản Nông nghiệp Hà Nội; giấy phép xuất bản 1079-2008/CXB/275-90/NN do Cục xuất bản cấp 02/12/2008. Nộp lưu chiểu 12/2008.

2
LÊ ĐỨC NGOAN, VŨ DUY GIẢNG, NGÔ HỮU TOÀN
GIÁO TRÌNH
DINH DƯỠNG VÀ THỨC ĂN THUỶ SẢN
Giáo trình điện tử (version 1)
NHÀ XUẤT BẢN NÔNG NGHIỆP

3
LỜI NÓI ĐẦU Đào tạo đại học ngành nuôi trồng thủy sản của trường Đại học Nông Lâm, Đại học
Huế bắt đầu từ năm 1995. Những năm gần đây, đào tạo kỹ sư nuôi trồng thủy sản của trường nói riêng và cả nước nói chung đã và đang được phát triển đáng kể về số lượng lẫn chất lượng. Để phục vụ việc nâng cao chất lượng đào tạo, nhà trường luôn khuyến khích các giảng viên, cán bộ khoa học tham gia biên soạn giáo trình, bài giảng và tài liệu tham khảo.
„Giáo trình dinh dưỡng và thức ăn thủy sản" được GS.TS. Vũ Duy Giảng (trường Đại học Nông nghiệp Hà Nội), PGS.TS. Lê Đức Ngoan và TS. Ngô Hữu Toàn (trường Đại học Nông Lâm, Đại học Huế) biên soạn nhằm góp phần vào công tác đào tạo và nghiên cứu của nhà trường. Hy vọng quyển sách này là tài liệu tham khảo tốt cho các trường đại học, cao đẳng khác trong cả nước. Giáo trình được nhóm tác giả biên soạn công phu, tham khảo nhiều tài liệu trong, ngoài nước và thông tin cập nhật. Giáo trình bao gồm 14 chương và 5 nhóm phụ lục, đã được TS. Lại Văn Hùng – trưởng khoa Nuôi ttrồng thủy sản, Đại học Nha Trang- góp ý và phản biện.
Trong khuôn khổ thời lượng của một môn học thuộc chương trình đào tạo ngành nuôi trồng thủy sản với 3 tín chỉ, chắc chắn nội dung và thông tin trong giáo trình này chưa thể bao trùm những vấn đề chuyên sâu về lĩnh vực dinh dưỡng và thức ăn cho tất cả các đối tượng nuôi trồng thủy sản. Chúng tôi mong muốn nhận được sự đóng góp ý kiến xây dựng từ quý thầy cô giáo, các nhà nghiên cứu, các sinh viên và đọc giả để tài liệu có thể hoàn chỉnh hơn trong lần tái bản sau.
Chúng tôi xin trân trọng cám ơn TS. Lại Văn Hùng về những góp ý có giá trị khoa học và dự án Nghiên cứu phát triển nông thôn bền vững (RDVIET) do Sida/SAREC tài trợ đã trợ giúp in ấn giáo trình này.
Giáo trình được biên soạn lại ở dạng điện tử, có bổ sung và sửa chữa. PGS. TS. Trần Văn Minh Hiệu trưởng, Chủ tịch Hội đồng Khoa học - Giáo dục

4
MỤC LỤC
MỞ ĐẦU.................................................................................................................10
ĐẶC ĐIỂM DINH DƯỠNG ĐỘNG VẬT THỦY SẢN .....................................10 I. MỘT SỐ KHÁI NIỆM .....................................................................................10
1.1. Thức ăn .....................................................................................................10 1.2. Dinh dưỡng ...............................................................................................10 1.3. Lịch sử phát triển dinh dưỡng học động vật thủy sản ..............................11
II. NHỮNG ĐẶC ĐIỂM DINH DƯỠNG ĐỘNG VẬT THỦY SẢN................11 III. QUAN HỆ GIỮA THỨC ĂN VỚI CÁC HÌNH THỨC NUÔI TRỒNG
THỦY SẢN .....................................................................................................................12
CHƯƠNG I.............................................................................................................14
SINH LÝ TIÊU HOÁ CỦA CÁ............................................................................14 1.1. CẤU TẠO GIẢI PHẪU BỘ MÁY TIÊU HOÁ CỦA CÁ.............................14 1.2. ĐẶC ĐIỂM CÁC LOẠI DỊCH TRONG ỐNG TIÊU HÓA CỦA CÁ..........14
1.2.1. Dịch vị (dạ dày - gastric secretion) .......................................................14 1.2.2. Dịch tuỵ (pancreatic secretion) .............................................................15 1.2.3. Dịch mật (bile secretion) .......................................................................15 1.2.4. Dịch ruột (intestial secretion)................................................................15
1.3. SỰ TIÊU HÓA CÁC CHẤT DINH DƯỠNG ...............................................16 1.3.1. Sự tiêu hóa do các enzyme .....................................................................16 1.3.2. Sự tiêu hoá do vi sinh vật .......................................................................18
1.4. SỰ HẤP THU CÁC CHẤT DINH DƯỠNG.................................................18 1.5. TỶ LỆ TIÊU HOÁ THỨC ĂN......................................................................19
CHƯƠNG II ...........................................................................................................22
CHUYỂN HÓA VÀ TÍCH LŨY CHẤT DINH DƯỠNG Ở CÁ........................22 2.1. MỞ ĐẦU.......................................................................................................22 2.2. CHUYỂN HOÁ CARBOHYDRATE...........................................................22
2.2.1. Sự thuỷ phân glucose .............................................................................23 2.2.2. Tổng hợp carbohydrate..........................................................................24 2.2.3. Con đường chuyển hóa pentose phosphate............................................24 2.2.4. Thức ăn và sự chuyển hoá carbohydrate ...............................................24
2.3. CHUYỂN HÓA LIPID..................................................................................24 2.4. CHUYỂN HOÁ AMINO ACID ...................................................................26 2.5. CÁC YẾU TỐ ẢNH HƯỞNG ĐẾN QUÁ TRÌNH TRAO ĐỔI CHẤT........28
2.5.1. Ảnh hưởng của khẩu phần đến sự trao đổi trung gian ..........................28 2.5.2. Lượng ăn vào và sự trao đổi chất ..........................................................28 2.5.3. Thành thục giới tính và trao đổi chất ....................................................29
CHƯƠNG III..........................................................................................................31
NĂNG LƯỢNG VÀ NHU CẦU NĂNG LƯỢNG...............................................31 3.1. TRAO ĐỔI NĂNG LƯỢNG........................................................................31

5
3.1.1. Khái niệm chung ....................................................................................31 3.1.2. Chuyển hóa năng lượng của thức ăn .....................................................31
3.2. NHU CẦU NĂNG LƯỢNG .........................................................................34 3.2.1. Nhu cầu năng lượng duy trì ...................................................................34 3.2.2. Nhu cầu năng lượng cho sự tăng trưởng ...............................................35
CHƯƠNG IV..........................................................................................................36
DINH DƯỠNG PROTEIN VÀ AMINO ACID ..................................................36 4.1. PROTEIN......................................................................................................36
4.1.1. Phân loại ................................................................................................36 4.1.2. Vai trò của protein .................................................................................36 4.1.3. Nhu cầu protein của cá ..........................................................................37 4.1.4. Tỷ lệ năng lượng/protein........................................................................37 4.1.5. Đánh giá chất lượng protein thức ăn.....................................................38
4.2. AMINO ACID...............................................................................................40 4.2.1. Các amino acid thiết yếu........................................................................40 4.2.2. Nhu cầu amino acid ...............................................................................41 4.2.3. Vấn đề bổ sung amino acid công nghiệp vào khẩu phần.......................42
CHƯƠNG V ...........................................................................................................43
DINH DƯỠNG LIPID...........................................................................................43 5.1. KHÁI NIỆM, PHÂN LOẠI VÀ CHỨC NĂNG ..........................................43
5.1.1. Khái niệm...............................................................................................43 5.1.2. Phân loại ................................................................................................43 4.1.3. Chức năng..............................................................................................48
5.2. VAI TRÒ DINH DƯỠNG CỦA AXIT BÉO ................................................49 5.2.1. Sinh tổng hợp các axit béo của động vật thuỷ sản.................................49 5.2.2. Các yếu tố ảnh hưởng đến thành phần axit béo trong động vật thuỷ sản
......................................................................................................................................50 5.2.3. Vai trò và nhu cầu axit béo thiết yếu .....................................................50
CHƯƠNG VI..........................................................................................................53
CARBOHYDRATE VÀ NHU CẦU ĐỐI VỚI CÁ.............................................53 6.1. KHÁI NIỆM..................................................................................................53 6.2. PHÂN LOẠI CARBOHYDRATE ...............................................................53
6.2.1. Monosaccharide.....................................................................................55 6.2.2. Oligosaccharide.....................................................................................56
6.3. SỰ CHUYỂN HOÁ ĐƯỜNG Ở CÁ .............................................................60 6.4. SỬ DỤNG TINH BỘT VÀ CHẤT XƠ Ở CÁ ...............................................60
6.4.1. Tinh bột ..................................................................................................60 6.4.2. Chất xơ ...................................................................................................61
CHƯƠNG VII ........................................................................................................62
DINH DƯỠNG VÀ NHU CẦU VITAMIN .........................................................62

6
7.1. VITAMIN A..................................................................................................62 7.1.1. Công thức cấu tạo..................................................................................62 7.1.2. Vai trò sinh học......................................................................................63
7.2. VITAMIN D..................................................................................................63 7.2.1. Công thức...............................................................................................63 7.2.2. Vai trò sinh học......................................................................................64
7.3. VITAMIN E ..................................................................................................65 7.3.1. Công thức...............................................................................................65 7.3.2. Vai trò sinh học......................................................................................65
7.4. VITAMIN K..................................................................................................66 7.4.1. Công thức...............................................................................................66 7.4.2. Chức năng..............................................................................................67
7.5. VITAMIN C (AXIT ASCORBIC) ................................................................67 7.6. VITAMIN NHÓM B.....................................................................................67 7.7. NHU CẦU VITAMIN CỦA CÁ ...................................................................68 7.8. SỬ DỤNG VITAMIN TRONG THỨC ĂN NUÔI CÁ.................................68
CHƯƠNG VIII.......................................................................................................70
DINH DƯỠNG VÀ NHU CẦU CHẤT KHOÁNG.............................................70 8.1. KHÁI NIỆM CHUNG ..................................................................................70 8.2. CANXI, PHOSPHO, MAGIE .......................................................................71
8.2.1. Canxi (Ca).............................................................................................71 8.2.2. Phospho (P) ...........................................................................................72 8.2.3. Magiê (Mg) ............................................................................................72
8.3. CÁC NGUYÊN TỐ KHOÁNG KHÁC ........................................................73
CHƯƠNG IX..........................................................................................................74
ĐẶC ĐIỂM THỨC ĂN TRONG NUÔI TRỒNG THUỶ SẢN.........................74 9.1. PHÂN LOẠI THỨC ĂN ..............................................................................74 9.2. ĐẶC ĐIỂM DINH DƯỠNG CỦA CÁC LOẠI THỨC ĂN ........................74
9.2.1. Thức ăn giàu năng lượng.......................................................................74 9.2.2. Thức ăn giàu protein..............................................................................74 9.2.3. Thức ăn bổ sung (feed additives) ...........................................................77 9.2.4. Thức ăn tự nhiên ....................................................................................78 9.2.5. Mùn bã hữu cơ và sinh khối vi khuẩn ....................................................78 9.2.6. Probiotic trong nuôi trồng thuỷ sản.......................................................78
CHƯƠNG X ...........................................................................................................83
THỨC ĂN TỰ NHIÊN..........................................................................................83 10.1. VAI TRÒ CỦA THỨC ĂN TỰ NHIÊN ĐỐI VỚI NUÔI TRỒNG THỦY
SẢN..................................................................................................................................83 10.1.1. Kích thước miệng ấu trùng ở pha nuôi dưỡng đầu..............................83 10.1.2. Ống tiêu hoá.........................................................................................84
10.2. VI TẢO (MICRO-ALGAE)..............................................................................84

7
10.2.1.Các loài tảo nuôi trồng chủ yếu 82
10.2.2. Giá trị dinh dưỡng của vi tảo...............................................................86 10.2.3. Vi tảo trong nuôi trồng thủy sản..........................................................89
10.3. LUÂN TRÙNG (ROTIFERS).........................................................................91 10.3.1. Hình thái ..............................................................................................91 10.3.2. Một vài đặc điểm sinh học quan trọng.................................................91 10.3.3. Giá trị dinh dưỡng của rotifer .............................................................92 10.3.4. Rotifer trong nuôi trồng thủy sản.........................................................92
10.4. ARTEMIA ..................................................................................................93 10.4.1. Một số đặc điểm sinh học quan trọng..................................................93 10.4.2. Giá trị dinh dưỡng ...............................................................................94
10.5. CÁC ZOOPLANKTON KHÁC..................................................................96 10.5.1. Copepod ...............................................................................................96 10.5.2. Daphnia và Moina ...............................................................................97 10.5.3. Nematode .............................................................................................98 10.5.4. Ấu trùng bánh xe (trochophora larvae) ...............................................99
10.6. QUẢN LÝ AO NUÔI ĐỂ PHÁT TRIỂN THỨC ĂN TỰ NHIÊN..............99 10.6.1. Yếu tố vật lý..........................................................................................99 10.6.2. Yếu tố sinh học .....................................................................................99 10.6.3. Yếu tố hoá học....................................................................................100
CHƯƠNG XI........................................................................................................102
CHẾ BIẾN THỨC ĂN VÀ THỨC ĂN CÔNG NGHIỆP ................................102 11.1. CHẾ BIẾN THỨC ĂN HẠT .....................................................................102
11.1.1. Tính chất vật lý, hóa học của tinh bột................................................102 11.1.2. Biến đổi vật lý, hóa học của tinh bột trong quá trình chế biến..........102 11.1.3. Kỹ thuật chế biến ...............................................................................103
11.2. THỨC ĂN HỖN HỢP VÀ CÔNG NGHỆ THỨC ĂN HỖN HỢP .........104 11.2.1. Phân loại thức ăn công nghiệp ..........................................................104 11.2.2. Tiêu chuẩn chất lượng của thức ăn hỗn hợp .....................................104 11.2.3. Các quy định pháp lý đối với thức ăn hỗn hợp ..................................105
11.2.4. Công nghệ thức ăn hỗn hợp ................................................................106 11.2.5. Những thiết bị cần thiết của một nhà máy thức ăn hỗn hợp.. ............108
CHƯƠNG XII ......................................................................................................109
TIÊU CHUẨN ĂN VÀ KHẨU PHẦN ...............................................................109 12.1. KHÁI NIỆM..............................................................................................109
12.1.1. Tiêu chuẩn ăn.....................................................................................109 12.1.2. Nội dung tiêu chuẩn ăn......................................................................110 12.1.3. Khẩu phần ăn.....................................................................................110
12.2. NGUYÊN TẮC PHỐI HỢP KHẨU PHẦN ĂN........................................110 12.2.1. Nguyên tắc khoa học..........................................................................110 12.2.2. Nguyên tắc kinh tế..............................................................................111
12.3. PHƯƠNG PHÁP PHỐI HỢP KHẨU PHẦN ĂN CHO TÔM, CÁ...........111 12.3.1. Phương pháp phối hợp thông thường................................................111

8
12.3.2. Phương pháp hình vuông Pearson.....................................................112 12.3.3. Phương pháp giải phương trình ........................................................112
CHƯƠNG XIII.....................................................................................................114
DINH DƯỠNG VÀ NUÔI DƯỠNG MỘT SỐ ĐỐI TƯỢNG TÔM, CÁ.......114 13.1. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ CHÉP (CIPRINUS CARPIO) .........114
13.1.1. Giới thiệu ...........................................................................................114 13.1.2. Nhu cầu dinh dưỡng...........................................................................115 13.1.3. Khẩu phần ăn.....................................................................................119 13.1.4. Nuôi dưỡng ........................................................................................121
13.2. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ TRÔI ẤN ĐỘ (INDIAN MAJOR CARP) ..............................................................................................................................123
13.2.1. Giới thiệu ...........................................................................................123 13.2.2. Nhu cầu dinh dưỡng...........................................................................123 13.2.3. Khẩu phần và nuôi dưỡng..................................................................126
13.3. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ RÔ PHI (OEROCHROMIS SPP.) ..129 13.3.1. Giới thiệu ...........................................................................................129 13.3.2. Nhu cầu dinh dưỡng...........................................................................129 13.3.3. Khẩu phần thức ăn.............................................................................138 13.3.4. Nuôi dưỡng ........................................................................................138
13.4. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ QUẢ VÀ CÁ DA TRƠN (PANGASIUS)...................................................................................................................140
13.4.1. Giới thiệu ...........................................................................................140 13.4.2. Nhu cầu dinh dưỡng...........................................................................140 13.4.3. Khẩu phần ăn.....................................................................................141 13.4.4. Nuôi dưỡng ........................................................................................142
13.5. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ CHÌNH (ANGUILLA SP.) ...........143 13.5.1. Giới thiệu ...........................................................................................143 13.5.2. Nhu cầu dinh dưỡng...........................................................................143 13.5.3. Khẩu phần ăn.....................................................................................146 13.5.4. Nuôi dưỡng ........................................................................................146
13.6. DINH DƯỠNG VÀ NUÔI DƯỠNG TÔM HE.........................................147 13.6.1. Giới thiệu ...........................................................................................147 13.6.2. Các hình thức nuôi.............................................................................148 13.6.3. Nhu cầu dinh dưỡng và nguồn cung cấp ...........................................148 13.6.4. Khẩu phần ăn và nuôi dưỡng.............................................................151
CHƯƠNG XIV.....................................................................................................155
ĐỘC TỐ TRONG THỨC ĂN THỦY SẢN.......................................................155 14.1. ĐÔC TỐ TỰ NHIÊN................................................................................155
14.1.1. Độc tố có nguồn gốc thực vật ............................................................155 14.1.2. Độc tố có nguồn gốc động vật ..............................................................159 14.1.3. Độc tố có nguồn gốc vi sinh vật ............................................................160
14.2. CÁC HỢP CHẤT KHÔNG TỰ NHIÊN VÀ PHỤ GIA TRONG KHẨU PHẦN.............................................................................................................................163
14.2.1. Hóa chất hữu cơ.................................................................................163

9
14.2.2. Kim loại nặng.....................................................................................164 PHỤ LỤC I.........................................................................................................166 HỖN HỢP THỨC, PREMIX KHOÁNG, VITAMIN CHO TÔM, CÁ .......................................166 PHỤ LỤC II .......................................................................................................172 BẢNG NHU CẦU DINH DƯỠNG CỦA MỘT SỐ LOÀI CÁ..................................................172 PHỤ LỤC III ......................................................................................................175 THÀNH PHẦN HOÁ HỌC CỦA MỘT SỐ THỨC ĂN CHỦ YẾU ......................................175 CHO CÁ Ở VIỆT NAM (%) ........................................................................................175 PHỤ LỤC IV......................................................................................................177 TIÊU CHUẨN NGÀNH MỘT SỐ LOẠI THỨC ĂN THỦY SẢN ..........................................177 PHỤ LỤC V .......................................................................................................192 TÊN KHOA HỌC CỦA MỘT SỐ LOẠI CÁ.....................................................................192 TÀI LIỆU THAM KHẢO CHÍNH.....................................................................193
Tiếng Việt .......................................................................................................193 Tiếng Anh.......................................................................................................193

10
MỞ ĐẦU ĐẶC ĐIỂM DINH DƯỠNG ĐỘNG VẬT THỦY SẢN
I. MỘT SỐ KHÁI NIỆM
1.1. Thức ăn Trong nuôi trồng thủy sản (NTTS), thức ăn đóng vai trò quan trọng vì chiếm tỷ lệ
cao trong chi phí (60-80% tổng chi phí). Tiết kiệm chi phí thức ăn làm tăng đáng kể lợi nhuận trong nuôi trồng. Về nguyên tắc, phương pháp tiết kiệm chi phí thức ăn bao gồm giảm đơn giá thức ăn và giảm lượng thức ăn tiêu thụ. Giảm giá thức ăn bao gồm chọn nguyên liệu đầu vào hợp lý về chất dinh dưỡng và giá. Giảm lượng thức ăn tiêu thụ cần phải hiểu biết rõ về nhu cầu dinh dưỡng để cân đối dinh dưỡng khẩu phần.
Thức ăn là vật chất chứa chất dinh dưỡng mà động vật có thể ăn, tiêu hóa và hấp thu để duy trì sự sống và tích lũy trong các mô cơ thể.
Trong tự nhiên, một loại vật chất có thể là thức ăn của loài cá này, giai đoạn phát triển cơ thể này nhưng chưa hẳn đã là thức ăn của loài cá khác, giai đoạn phát triển cơ thể khác. Sự khác biệt đó hoặc là do đặc điểm dinh dưỡng khác nhau theo loài, mà nguyên nhân chính là khả năng thu nhận và tiêu hóa các loại thức ăn khác nhau theo loài hoặc do sự khác biệt về mức độ hoàn thiện bộ máy tiêu hóa theo giai đoạn phát triển cơ thể. Đó cũng thể hiện đặc tính loài.
Thức ăn tự nhiên (live food, natural food): như các loài rong tảo và các sinh vật phù du động vật là những cơ thể sinh vật sống và phát triển trong hệ thống nuôi hoặc sinh vật sống được nuôi có thể dùng làm thức ăn cho động vật thuỷ sản.
Thức ăn nhân tạo (man-made food) còn được gọi là thức ăn công nghiệp (commercial food) hay thức ăn viên (pellet food). Trong thức ăn công nghiệp, nó còn được chia ra gồm thức ăn viên chìm (sinking food) sử dụng chủ yếu nuôi giáp xác và thức ăn nổi (floating food) sử dụng nuôi cá.
Thức ăn tươi sống (fresh food): là các loại động vật chưa qua chế biến, còn tươi.. dùng làm thức ăn cho tôm, cá như: tôm cá tạp, ốc, cua…
Thức ăn tự chế (home-made food): thức ăn do người nuôi tự phối chế chủ yếu từ các nguồn nguyên liệu sẵn có với qui trình đơn giản nhằm giảm giá thành và chủ động khi sử dụng. Loại thức ăn tự chế này thường ở dạng ẩm và sử dụng ngay sau khi chế biến. 1.2. Dinh dưỡng
Dinh dưỡng là các quá trình hoạt động sinh lý và hoá học để chuyển hóa những chất dinh dưỡng có trong thức ăn thành những chất dinh dưỡng cho cơ thể sử dụng.
Có 4 quá trình trong quá trình dinh dưỡng: thu nhận thức ăn, tiêu hoá hấp thu thức ăn, chuyển hoá và bài tiết các chất dinh dưỡng khỏi cơ thể. Môn học nghiên cứu các quá trình trên gọi là dinh dưỡng học.
Mục đích của dinh dưỡng học động vật thuỷ sản là nghiên cứu cơ sở khoa học và thực tiễn để cho quá trình chuyển những chất dinh dưỡng của thức ăn thành những chất dinh dưỡng của cơ thể hiệu quả nhất (con vật khoẻ mạnh, sinh trưởng phát triển tốt và có hiệu suất lợi dụng thức ăn cao nhất).
Chất dinh dưỡng là các nguyên tố hay hợp chất hóa học có trong khẩu phần làm thỏa mãn sự sinh sản, sinh trưởng hay duy trì quá trình sống bình thường. Sáu nhóm chất

11
dinh dưỡng đã được phân loại như sau: nước, protein và amino acid, carbohydrate, lipit, vitamin và các nguyên tố khoáng.
Nước Carbohydrate Lipit THỨC ĂN Hữu cơ Protein và axit nucleic
Axit hữu cơ
Vật chất khô Vitamin Lignin hỗn hợp; axit hữu cơ; các hợp chất tạo màu, mùi và vị; hormone...
Vô cơ: Thiết yếu: Ca, Cl, K, Mg, Na, P, S, Co, Cr, Cu, F, Fe, I, Mn, Mo, Ni,Se, Si, Sn, V, Zn.
Không thiết yếu: Ag, Al, Au, Bi, Ge, Hg, Pb, Rb,Sb, Ti.
Độc: As, Cd, F, Hg, Mo, Pb, Se, Si. Năng lượng mà tất cả động vật đều cần được lấy từ mỡ, carbohydrate và từ các sản
phẩm khử amin của các amino acid. Các chất dinh dưỡng cung cấp cho tế bào: nước, các vật liệu, các hợp chất cấu trúc (da, cơ, xương, thần kinh, mỡ) và chất điều chỉnh quá trình trao đổi chất trong cơ thể. Động vật cần hơn 40 chất dinh dưỡng khác nhau và được lấy từ khẩu phần thức ăn và có những chất bản thân cơ thể không tổng hợp được gọi là ”chất dinh dưỡng thiết yếu”, và một số chất bản thân có thể tổng hợp được gọi là “chất dinh dưỡng không thiết yếu”. Nhóm chất dinh dưỡng thiết yếu bao gồm: các amino acid thiết yếu, các axit béo thiết yếu và các khoáng thiết yếu (sẽ trình bày cụ thể trong các chương IV, V và VIII). 1.3. Lịch sử phát triển dinh dưỡng học động vật thủy sản
Dinh dưỡng học thuỷ sản chỉ mới phát triển gần đây. Những nghiên cứu đầu tiên về dinh dưỡng thủy sản được thực hiện tại Corland (Ohio, Mỹ) vào những năm 40 và phát triển nhanh sau những năm 60 của thế kỹ XX. Thức ăn nhân tạo cho động vật thuỷ sản bắt đầu áp dụng từ thập niên 50 và cuối thập niên của thế kỷ trước, thức ăn viên được dùng phổ biến tại Mỹ và Châu Âu.
Động vật thuỷ sản chủ yếu bao gồm các loài cá có xương (finfish), giáp xác (crustacean) và nhuyễn thể (mollusca). Chúng có những đặc điểm dinh dưỡng khác với các động vật trên cạn. Số lượng các loài cá rất phong phú, nhưng hiện chỉ có khoảng 20 loài được nghiên cứu về dinh dưỡng và đại bộ phận tập trung vào những loài cá ôn đới.
II. NHỮNG ĐẶC ĐIỂM DINH DƯỠNG ĐỘNG VẬT THỦY SẢN
Cá có cấu trúc ống tiêu hoá và chức năng tiêu hoá rất khác nhau và đa số động vật thuỷ sản đều trãi qua giai đoạn ấu trùng. Ở giai đoạn này nhu cầu dinh dưỡng của chúng biến đổi rất lớn, do vậy nghiên cứu về dinh dưỡng của động vật thủy sản khó hơn so với động vật trên cạn.
Cá là động vật biến nhiệt (poikilotherms) nên có nhu cầu năng lượng thấp hơn động vật máu nóng vì không tiêu tốn năng lượng vào việc điều tiết thân nhiệt. Tuy nhiên, cá lại nhạy cảm với stress của môi trường, đặc biệt là nhiệt độ nước. Do vậy, nhu cầu dinh dưỡng thường được xác định ở khoảng nhiệt độ nước thích hợp nhất định, gọi là nhiệt độ môi trường tiêu chuẩn (SET: Standard Environmental Temperatures).

12
Ví dụ: theo NRC thì SET của một số loại cá như sau: Cá hồi (chinook salmon): 59o F (15o C) Cá hồi vân (rainbow trout): 50oF (10oC) Cá da trơn Mỹ (channel catfish): 86oF (30oC) Nhu cầu năng lượng của động vật thuỷ sản thấp hơn động vật trên cạn (vì không
mất năng lượng để điều hoà thân nhiệt, không tốn nhiều năng lượng để vận động, không mất nhiều năng lượng trong chuyển hoá protein (vì vậy, cá được xếp vào nhóm ammoniotelic- bài tiết amoniac).
Nhu cầu vitamin cũng cao hơn, đặc biệt vitamin C, do cá không tự tổng hợp được trong cơ thể, vì vậy nhu cầu vitamin phụ thuộc nhiều vào thức ăn.
Nhu cầu chất khoáng thấp hơn vì cá có thể lấy chất khoáng từ môi trường nước. Hầu hết các loài cá có nhu cầu về axit béo nhóm Ω-3 (hay n-3) và các nhóm động vật thuỷ sản khác nhau thì có nhu cầu axit béo này khác nhau.
Về hiệu suất sử dụng thức ăn. Hiệu suất sử dụng (HSSD) thức ăn của cá cao hơn động vật trên cạn. HSSD của cá trong khoảng 1,2 - 1,7/1, trong khi đó HSSD của lợn là 3/1 và của gà là 2/1).
Về phương thức lấy thức ăn của cá. Có nhiều phương thức như bắt mồi (cá hồi), gặm (như cá đối), lọc (như cá mòi, có thể lọc 6 gallons nước/phút qua mang), hút, ký sinh (như cá mút đá ...). Do đó, thức ăn phải được chế biến và cho ăn theo phương thức lấy thức ăn của cá.
III. QUAN HỆ GIỮA THỨC ĂN VỚI CÁC HÌNH THỨC NUÔI TRỒNG THỦY SẢN
Trong nuôi trồng thuỷ sản, tuỳ phương thức sản xuất, thức ăn tự nhiên và thức ăn nhân tạo chiếm vị trí khác nhau. Trong nuôi quảng canh, thức ăn tự nhiên là quan trọng, khi phương thức quảng canh được thay dần bằng bán thâm canh hoặc thâm canh thì thức ăn nhân tạo thay thế dần cho thức ăn tự nhiên. Trình độ thâm canh càng cao thì thức ăn nhân tạo càng giữ vai trò quan trọng (sơ đồ 1). Hai tam giác ngược chiều nhau (thức ăn nhân tạo và thức ăn tự nhiên) chỉ rõ mức độ đóng góp của các nguồn thức ăn khi thay đổi các phương thức nuôi trồng thủy sản.
Sơ đồ 1. Mối quan hệ giữa thức ăn tự nhiên và thức ăn nhân tạo trong nuôi trồng thuỷ sản
Thức ăn
nhân tạo
Thức ăn tự
nhiên
Nuôi quảng canh
Nuôi bán thâm canh
Nuôi thâm canh

13
CÂU HỎI 1. Thế nào là thức ăn và dinh dưỡng? 2. Nêu những đặc điểm dinh dưỡng của động vật thủy sản? 3. Quan hệ giữa thức ăn với các hình thức nuôi trồng thủy sản?
TÀI LIỆU THAM KHẢO
Tiếng Việt Vũ Duy Giảng, Nguyễn Thị Lương Hồng, Tôn Thất Sơn (1997). Dinh dưỡng và thức ăn
gia súc. Nhà XBNN, Hà Nội. Lại Văn Hùng (2004). Dinh dưỡng và thức ăn trong nuôi trồng thủy sản. Nhà XBNN, Tp
Hồ Chí Minh. Lê Đức Ngoan (2002). Dinh dưỡng gia súc. Nhà XBNN, Hà Nội.
Tiếng Anh Michael B. New (1987). Feed and Feeding of Fish and Shrimp (A manual on the
preparation and presentation of compound feed for shrimp and fish in aquacultue), UNDP, FAO, Rome.
Webster, C.D. and Lim C. (eds) (2002). Nutritient Requirement and Feeding of Finfish for Aquaculture. CAB international.

14
CHƯƠNG I SINH LÝ TIÊU HOÁ CỦA CÁ
1.1. CẤU TẠO GIẢI PHẪU BỘ MÁY TIÊU HOÁ CỦA CÁ Ống tiêu hoá cá có 4 phần: phần đầu, ruột trước, ruột giữa và ruột sau (sơ đồ 1.1 và
1.2). Phần đầu gồm xoang miệng và mang. Ruột trước gồm thực quản, dạ dày, hạ vị. Một số loài cá không có dạ dày (khoảng 15% loài cá không có dạ dày) thì ruột trước chỉ có thực quản và một đoạn ruột bắt đầu từ cuối ống thực quản kéo đến cửa ống dẫn mật. Ruột giữa là đoạn ruột từ sau van hạ vị đến đầu đoạn ruột sau. Gần van hạ vị có túi mù hạ vị (pylorus caecae), cá chó có 5-8 túi , cá hồi nước ngọt có 35-100 túi. Ruột sau gồm kết tràng và lỗ thải phân.
Niêm mạc ruột là các lông nhung, kích cỡ lông nhung biến đổi theo thời tiết và thức ăn (cá sống môi trường lạnh có lông nhung dài và dày hơn so với cá sống môi trờng nóng, tuy nhiên tổng số lông nhung thì không biến đổi).
Đặc điểm chung về ống tiêu hoá tất cả các loài cá là: cấu tạo giải phẫu biến đổi theo tập tính ăn. Ruột của loài ăn thực vật (herbivores) dài hơn loài ăn động vật (carnivores). Chiều dài ruột/dài thân của carnivores, omnivores và herbovores lần lượt là 0,2-0,5/1; 0,6-8,0/1 và 0,8-15/1.
1.2. ĐẶC ĐIỂM CÁC LOẠI DỊCH TRONG ỐNG TIÊU HÓA CỦA CÁ Ống tiêu hoá của cá chứa nhiều loại dịch như dịch vị, dịch tuỵ và dịch ruột, trong
đó có chứa các enzyme giữ vai trò quan trọng trong quá trình tiêu hoá các chất dinh dưỡng. 1.2.1. Dịch vị (dạ dày - gastric secretion)
Dịch vị có tính axit có ở hầu hết các loài cá, trừ cá không có dạ dày. Thành phần dịch vị gồm: axit
hydrochloric,

15
tiết ra từ dạ dày khi có thức ăn, nhờ đó pH dịch dạ dày có thể đạt tới 2 sau khi ăn vài giờ; enzyme: pepsin được hình thành từ pepsinogen trong môi trường axit. Pepsin phân cắt các peptide thành những mạch ngắn hơn, nó phân giải được hầu hết protein nhưng không phân giải được mucins, spongin, conchiolin, keratin hay những peptide phân tử lượng thấp.
Dich vị cũng chứa một số enzyme không phân giải protein, đó là các enzyme: amylase (ở cá trích), lipase (cá rô phi), esterase (pH = 5,3 - 8,0), chitinase (Coryphaenoides sp), hyaluronidase (Scomberjaponicus), cellulase (trong một vài loài cá cửa sông và cá nước ngọt) - enzyme này có nguồn gốc vi sinh vật chứ không phải của cá. 1.2.2. Dịch tuỵ (pancreatic secretion)
Dịch tụy do tuyến tụy ngoại tiết tiết ra, bao gồm: bicarbonate tiết ra để trung hoà HCl tiết ra từ dạ dày và các nhóm enzyme khác nhau.
Nhóm protease Trypsin: hình thành do thuỷ phân trypsinogen, phân giải liên kết peptide có nhóm
carboxyl đến từ arginine hay lysine. Hoạt động tối ưu ở pH=7. Chymotrypsin: hình thành do trypsin tác động vào chimotrypsinogen, phân giải liên
kết peptide có chứa nhóm carboxyl của axit amin mạch nhánh (tyrosine, tryptophan, phenylalanine).
Elastase được hình thành khi pro-elastase được hoạt hoá bởi trypsin, nó phân giải dây nối peptide của elastin.
Carboxypeptidase hình thành từ procarboxypeptidase sau khi được trypsin hoạt hoá, nó thuỷ phân các mạch peptide cuối cùng của cơ chất.
Amylase: tuyến tuỵ là nguồn chủ yếu của amylase của cá, pH tối ưu cho hoạt động của nó là 6,7.
Chitinase: có ở nhiều loài cá, đặc biệt các loài cá ăn côn trùng và giáp xác. Enzyme này hoặc sinh ra từ tuỵ (pH cho hoạt động tối ưu là 8-10) hoặc từ dạ dày (pH cho hoạt động tối ưu là 1,25-3,5).
Lipase: thuỷ phân mỡ triglyceride, phospholipide và sáp. Carbonic anhydrase: thấy ở ruột cá ở rạn san hô, người ta cho rằng enzyme này
dùng để phân giải calcium carbonate. 1.2.3. Dịch mật (bile secretion)
Về cơ bản, mật cá giống mật động vật có vú, nhưng vì mô gan và mô tuỵ của một vài loài cá trộn lẫn nhau cho nên dịch mật có chứa enzyme của tuỵ. Dịch mật có tính kiềm yếu, chứa muối mật, cholesterol, phospholipides, sắc chất mật, anion hữu cơ, glycoproteins và ion vô cơ. Dịch mật là tác nhân nhũ hoá mỡ trong quá trình tiêu hoá mỡ. 1.2.4. Dịch ruột (intestial secretion)
Dịch ruột chứa nhiều loại enzyme như amino-di-tripeptidase; alkali và axit nucleosidase (phân chia nucleoside); polynucleotidase (phân chia axit nucleic); lecithinase (phân chia phospholipid); lipase và những esterase khác (phân chia lipid); amylase, maltase, isomaltase, sucrase, lactase, trehalase và laminarinase (tiêu hoá carbohydrate). Hoạt tính amylase ruột cá chép cao hơn ở cá hồi vân. Laminarinase trong ruột cá rô phi (Tilapia macrochira) nuôi bằng vi sinh vật phù du. Laminarinase phân giải laminarin (β-1,3-glucan), có nhiều trong nhóm tảo Laminariaceae.

16
1.3. SỰ TIÊU HÓA CÁC CHẤT DINH DƯỠNG
1.3.1. Sự tiêu hóa do các enzyme
Nhóm enzyme protease và peptidase Ở động vật dạ dày đơn, tiêu hóa protein xảy ra trong ống tiêu hoá, bắt đầu ở dạ dày
trong những loài cá có dạ dày. Protein bị phân cắt thành những mảnh polypeptide sau đó thành dipeptide và amin acid (sơ đồ 1.3). Các poly, di-peptide tiếp tục được tiêu hoá ở ruột. Dưới tác động của nhóm enzyme protease từ dịch vị, dịch tụy và dịch ruột. Protein bị phân giải theo sơ đồ:
Protein pepton, polypeptide peptide đơn giản
amino acid Như đã biết, protein có
nhiều mức cấu trúc nên cần có lượng lớn enzyme protease và peptide và mỗi loại enzyme có chức năng chuyên biệt ứng với cấu trúc protein. Protease có vai trò như protease nội thì cắt giữa chuổi peptide, nếu là protease ngoại thì cắt đầu các chuổi peptide tự do. Ví dụ, aminopeptidase là một peptidase ngoại cắt các amino acid từ đầu cuối có nhóm amin của chuổi peptide từng lần một.
Protease là nhóm enzyme quan trọng để hoạt hóa các tiền enzyme của nhiều enzyme tiêu hóa thành enzyme hoạt động. Một ví dụ kinh điển là trypsinogen được hoạt hóa bởi enterokinase. Trypsin lại hoạt hóa các tiền enzyme tiêu hóa khác như chymotrypsin, elastase, collagenase và lipase.
Hoạt tính enzyme tiêu hoá protein của cá phụ thuộc yếu tố sau: - Loài: hoạt tính proteolytic của loài ăn động vật lớn hơn loài ăn thực vật. - Tuổi: hoạt tính enzyme peptic và tryptic tăng mạnh trong 20 ngày tuổi đầu, sau đó
hoạt tính tryptic tăng mạnh hơn peptic (40 ngày tuổi hoạt tính tryptic tăng 10 lần còn hoạt tính peptic tăng 4 lần).
- Thành phần thức ăn trong khẩu phần: khẩu phần chứa nhiều tinh bột và xơ làm giảm hoạt tính proteolytic.
- Nhiệt độ nước: enzyme proteolytic tiết nhiều và có hoạt lực cao ở nhiệt độ cao (40-50oC), ở nhiệt độ từ 5oC đến 20oC, hoạt lực proteolytic giảm 30-40% giá trị ban đầu.
- pH: đối với nhóm ăn lọc, pH tối ưu cho pepsin dạ dày là 3, cho trypsin và chimotrysin là 8,2 và 7,8; đối với nhóm ăn tạp những con số tương ứng là 2,5-3,3 đối với pepsin (nhiệt độ 40-50oC) và là 7,6 đối với trypsin (nhiệt độ 46oC).
- Thời gian sau khi ăn: hoạt tính protease dịch ruột cá chép đạt tối đa sau khi ăn 5 giờ, hoạt tính amylase giảm sau khi ăn 1 giờ , nhưng sau 5-6 giờ lại tăng lên.

17
Nhóm enzyme lipase Nhìn chung, lipid được thủy
phân ngoài tế bào (Higg và Dong, 2000) ở ruột non và manh tràng (sơ đồ 1.4) bởi nhiều nhóm enzyme lipase và colipase (Sargen et al., 1989). Hiểu chung nhất, các acid béo mạch ngắn (2-10 carbon) và glycerol được hấp thu trưc tiếp qua lông nhung của niêm mạc ruột. Các acid béo mạch dài (trên 12 carbon) được cắt ngắn bỏi lipase và nhủ hóa do muối mật thành các micelle. Micelle được chuyển vận từ ống tiêu hóa vào lông nhung, ở đó tách khỏi muối mật và acid béo khuếch tán qua màng biểu mô. Các acid béo tái este và nhóm với protein thành chylomicron.
Ở cá, lipid quan trọng nhất là triaglycerole, cholesterol và phospholipid (Sargen et al., 1989). Các chất này bị thủy phân thành các acid béo, glycerol, 2-monoacylglycerol và lysophospholipid. Các hợp chất này được hấp thu qua màng ruột (Higg và Dong, 2000). Lipid cũng được hấp thu ở dạng hạt nhỏ (Smith, 1989).
Nhóm enzyme carbohydrase Enzyme tiêu hóa carbohydrate có rất nhiều nhóm và được tiết ra ở các đoạn khác
nhau của đường tiêu hóa. Amylase và maltase tiết ra chủ yếu ở đoạn ruột giữa, saccharase tiết chủ yếu ở đoạn ruột sau. Tuy nhiên, ở cá chép amylase tiết ra chủ yếu ở tuỵ và hầu như không tiết ra ở ruột. Cá con (6,5 g) có hoạt tính amylase và maltase cao hơn cá lớn (400 g); khẩu phần giàu tinh bột làm tăng hoạt tính của amylase và maltase; nhiệt độ thích hợp cho carbohydrase hoạt động thì tương đối rộng (20-40oC).
Nói chung, carbohydrate bị thủy phân ngoài tế bào ở dạ dày, ruột và manh tràng (Divakaran et al., 1999). Sản phẩm của thủy phân là polysaccharide và monosaccharide. Ở động vật có vú, monosaccharide được hấp thu chủ động qua tế bào lông nhung ruột non (Lentner, 1981). Đối với cá, cơ chế này chưa rõ ràng.
Khả năng tiêu hóa và sử dụng carbohydrate khác nhau rất lớn ở các loài (Chan và Horn, 1999; Divakaran et al., 1999). Sự sai khác về tỷ lệ tiêu hóa do khác nhau về số lượng và chủng loại carbohydrate tìm thấy ở các loài đó (Chan và Horn, 1999). Hầu hết các loài sử dụng tinh bột sống (chưa nấu) rất kém (Hemre et al., 2000). Ngược lại, tinh bột nấu chín và polysaccharide đơn giản được tiêu hóa rất tốt ở cá. Nhiều nghiên cứu cho thấy có mối tương quan dương giữa độ chính với tỷ lệ tiêu hóa tinh bột (Jeong et al., 1992; Podoskina et al., 1997). Mặt khác, carbohydrate có hoặc không thể được đồng hóa tốt ở các loài cá. Cá hồi (Oncorhynchus mykiss) sử dụng carbohydrate rất kém.

18
Chitin bị phân giải thành đường và N-acetylamin nhờ enzyme chitinase. Moe và Place (1999) cho thấy tương quan giữa hoạt tính chitinase và loại khẩu phần ở 8 loài cá biển đánh bắt. Tuy nhiên, Lindsay không tìm thấy tương quan nào ở 29 loài cá. Hoạt tính của enzyme này chỉ phát hiện ở dạ dày (pH thấp) và ruột và manh tràng (pH trung tính), điều này chỉ ra rằng có it nhất hai nhóm chitinase, một tiết ra từ dạ dày và một tiết ra từ tụy (Smith, 1999). Chitinase có thể không bị tiêu hóa do vi sinh vật và ký sinh trùng ở các nơi khác (Lundblad et al., 1979; Smith, 1999). 1.3.2. Sự tiêu hoá do vi sinh vật
Vi sinh vật trong đường tiêu hóa của cá gồm: vi khuẩn và protozoa, có ở phần cuối đoạn ruột nơi tiếp giáp trực tràng, chúng tiết ra các enzyme proteolytic, amylolytic, chitinase, lecithinase và cellulase. Tuy nhiên, vi khuẩn chỉ đóng một vai trò nhỏ trong quá trình tiêu hoá chitin và cellulose.
1.4. SỰ HẤP THU CÁC CHẤT DINH DƯỠNG Sự vận chuyển các chất dinh dưỡng từ ống ruột non vào tế bào bởi nhiều con
đường khác nhau, như thực bào, thẩm thấu, khuếch tán, hấp thu chủ động, trao đổi ion.. (Smith, 1989). Thực bào (pinocytosis) có thể vận chuyển lượng lớn các hợp chất sẽ được tiêu hóa nội bào hoặc sử dụng cho hệ miễn dịch hay các dịch tiết tiêu hóa (enzyme, muối mật..). Thẩm thấu là do chênh lệch nồng độ trong và ngoài tế bào. Hấp thu chủ động (active transport) là sự lựa chọn và cần gradient Na để bơm chất dinh dưỡng (ví dụ, amino acid) qua màng tế bào lông nhung (Storelli và Verri, 1993). Hấp thu chủ động cần năng lượng và quan trọng hơn thẩm thấu cũng như thực bào, đặc biệt đối với các amino acid thiết yếu (Verri, 1993).
Ít nhât có hai nhóm enzyme quan trọng ở cá và động vật có xương sống tham gia hấp thu chủ động là alkaline phosphatase và gama-glutamyltransferase. Phosphatase khử phospho các chất dinh dưỡng và có vai trò trong hấp thu lipd và protein như là chất xúc tác cho phản ứng chuyển phospho (Villanueva et al., 1997). Gama-Glutamyltransferase xúc tác thủy phân các liên kết peptide gama-glutamyl và tham gia vận chuyển protein qua màng (Baumrucker et al., 1989).
Bảng 1.1. Các hệ thống vận chuyển các chất dinh dưỡng
Hệ thống vận chuyển
Cơ chất, chất mang Vị trí Nguồn
Amino acid Alanine (A) Hầu hết amino acid, chất
mang Na Thành ruột Baumrucker et
al. 1989 Leucine (l) Chuổi nhánh lớn hay chuổi
chất thơm nt nt
LNBB Hầu hết amino acid trung tính, chất mang Na
nt Mercer et al., 1989
Phenylalanine Phe, Met; chất mang Na nt Baumrucker et al. 1989
Cation Amino acid cation (Arg, Lys, Orn)
nt nt
Anion Amino acid anion (Asp, Glu) nt nt Imino Proline; chất mang Na nt nt

19
Khoáng Ca Vit D có vai trò quan trọng
trong hấp thu ở ruột. Hấp thu giảm do xơ, Zn, Mg và phytate
Mang, biểu mô; Ruột
Lall, 1989
P Phosphate hòa tan; trao đổi P bị ức chế bởi Fe và Mn
Ruôt, mang và da
nt
Na/K/Clo Bơm năng lượng chuyển các Ion quan màng
Mang, da nt
Fe Tiến trình phức tạp, Fe được hấp thu qua hệ thống protein và chuyển vào máu qua protein khác
Ruột, mang nt
Ở động vật có vú có bảy hệ thống vận chuyển chủ động amino acid trong lông
nhung chất nhầy ruột (bảng 1.1). Một số tác giả cho rằng các hệ thống này ở cá tương tự với động vật có vú nhưng đến nay chúng vẫn chưa được nghiên cứu chi tiết trên cá. Mặc dù vậy, có thể tóm tắt sự hấp thu ba nhóm chất dinh dưỡng như sau:
Protein. Amino acid được hấp thu theo gradient nồng độ sau khi kết hợp với ion vô cơ, những peptide đơn giản được hấp thu bằng cơ chế thực bào (pinocytosis).
Lipid. Những sản phẩm hoà tan của tiêu hoá lipid được hấp thu chủ yếu ở niêm mạc ruột trước và cả ở các túi mù hạ vị.
Carbohydrate. Glucose được hấp thu ở niêm mạc ruột theo cơ chế hấp thu tích cực và theo gradient nồng độ.
1.5. TỶ LỆ TIÊU HOÁ THỨC ĂN Để xác định khả năng tiêu hóa thức ăn, người ta sử dụng khái niệm tỷ lệ tiêu hóa.
Tỷ lệ tiêu hoá (TLTH) thức ăn được xác định bằng tỷ lệ của chất dinh dưỡng tiêu hoá so với chất dinh dưỡng ăn vào. TLTH có thể biểu thị bởi hệ số hay phần trăm.
TLTH = Chất dinh dưỡng ăn vào - Chất dinh dưỡng trong phân
Chất dinh dưỡng ăn vào
Chất dinh dưỡng được định nghĩa như là năng lượng, protein, axít amin, carbon
hydrate, lipid. Riêng đối với chất khoáng không áp dụng công thức trên vì lượng khoáng trao đổi thải qua phân khá lớn.
Ví dụ, trong một ngày, một cá trắm cỏ ăn hết 400 g cỏ tươi (chứa 10% vật chất khô, tương ứng 40 g chất khô ăn vào) và thải ra 50 g phân (chứa 20% vật chất khô, tương ứng 10 g chất khô thải ở phân), thì TLTH vật chất khô sẽ là 0,75 (hay là 75%).
Ý nghĩa: Tỷ lệ tiêu hóa nói lên khả năng tiêu hóa của vật nuôi với một loại thức ăn, khẩu phần hay một chất dinh dưỡng nào đó. Giá trị 75% có nghĩa là nếu cho cá ăn 1 kg thức ăn thì chỉ có 750 g được tiêu hóa để hấp thu, còn 250 g thải qua phân.
Tỷ lệ tiêu hóa biểu kiến (Apparent Digestibility) và tỷ lệ tiêu hóa thực (True Digestibility). Chất thải trong phân không chỉ có thức ăn không tiêu hóa mà còn có các chất dinh dưỡng (ngoại trừ xơ) thải qua phân có nguồn gốc nội sinh. Những chất dinh

20
dưỡng này là những chất có trong tế bào ruột và các chất tiết của đường tiêu hóa.có nguồn gốc từ cơ thể vật chủ. Các chất nội sinh này không thể phân biệt với chất không tiêu hóa trong thức ăn được. Vì vậy người ta đã đưa ra khái niệm TLTH biểu kiến (bao gồm cả phần nội sinh) để phân biệt với TLTH thực (không bao gồm các chất nội sinh). Tính tỷ lệ tiêu hóa biểu kiến (theo công thức trên) sẽ luôn luôn cho giá trị thấp hơn tỷ lệ tiêu hóa thực của thức ăn. TLTH biểu kiến đối với các chất khoáng gần như không có ý nghĩa.
Tỷ lệ tiêu hóa thực (TLTHt) là tỷ lệ chất ăn vào được hấp thu từ đường tiêu hóa, không tính đến lượng nội sinh so với lượng ăn vào. Công thức tính như sau:
TLTHt = Chất dinh dưỡng ăn vào - (Chất dinh dưỡng trong phân + Chất nội sinh)
Chất dinh dưỡng ăn vào
Chất nội sinh của cơ thể thải qua phân chủ yếu là Nitơ - được gọi là N trao đổi
trong phân. Một trong các phương pháp để xác định N trao đổi là nuôi cá với khẩu phần không chứa N.
Hiện nay, TLTH biểu kiến và TLTH thực đang được sử dụng rộng rãi trong các tài liệu. Trong thực tế, TLTH thực được xác định rất khó khăn ví khó để tách biệt các chất có nguồn gốc thức ăn và nguồn gốc từ cơ thể, cho nên trong dinh dưỡng động vật thuỷ sản người ta thường chỉ sử dụng TLTH biểu kiến.
Để xác định tỷ lệ tiêu hoá một chất dinh dưỡng nào đó trong thức ăn, có nhiều phương pháp khác nhau, tuy nhiên có một phương pháp phổ biến trong nghiên cứu dinh dưỡng cá là phương pháp dùng chất đánh dấu.
Chất đánh dấu thường dùng là oxit crom (Cr2O3), chất này hầu như không bị tiêu hoá hấp thu trong đường tiêu hoá. Oxit crom được trộn vào thức ăn theo tỷ lệ 1-2%. Sau khi cho ăn một thời gian, người ta lấy mẫu phân của cá. Cùng với việc định lượng tỷ lệ của các chất dinh dưỡng trong thức ăn, người ta cũng xác định tỷ lệ của oxit crom trong phân. Sau đó, áp dụng công thức sau để tính TLTH biểu kiến:
TLTH = 100 - [ 100 A x b ] B a Trong đó, A: % chất đánh dấu có trong thức ăn (theo khối lượng chất khô) B: % chất đánh dấu có trong phân (theo khối lượng chất khô) a: % chất dinh dưỡng có trong thức ăn (theo khối lượng chất khô) b: % chất dinh dưỡng có trong phân (theo khối lượng chất khô)
CÂU HỎI 1. Sự giống và khác nhau giữa thức ăn và chất dinh dưỡng ? 2. Những đặc điểm cấu tạo ống tiêu hoá của cá ? 3. Những enzyme tiêu hoá protein, lipid và carbohydrate và kết quả tác động của
những enzyme này trong quá trình tiêu hoá thức ăn ? 4. Hấp thu protein, lipid và carbohydrate của cá ? 5. Tỷ lệ tiêu hóa và cách xác định ?

21
TÀI LIỆU THAM KHẢO
Tiếng Việt Lại Văn Hùng (2004). Dinh dưỡng và thức ăn trong nuôi trồng thủy sản. Nhà XBNN, TP
Hồ Chí Minh. Tiếng Anh Halver, J.E. and Hardy, R.W. (2002). Fish Nutrition. 3rd Ed. Academic Press, Imprint of
Elsevier Science. Steffens, W. (1989). Principles of Fish Nutrition. Ellis Horwood Limited, England.

22
CHƯƠNG II CHUYỂN HÓA VÀ TÍCH LŨY CHẤT DINH DƯỠNG Ở CÁ
2.1. MỞ ĐẦU Sự chuyển hoá các chất dinh dưỡng của cá là tổng hợp các phản ứng hoá sinh trong
suốt quá trình sống. Những nét cơ bản về con đường chuyển hoá dinh dưỡng của các loài động vật sống trên cạn đã được nêu ra trong những nghiên cứu cổ điển về sinh hoá của Lehninger (1979), West et al. (1966) và Kaneko (1989),
nó cũng phù hợp với các loài động vật biến nhiệt như cá. Các loài cá khác nhau từ loài ăn thịt đến ăn tạp và ăn thực vật sống trong giới hạn nhiệt độ nước khá rộng từ 00C đến 400C, thích ứng với khả năng sử dụng nguồn carbohydrate, protein và lipid để tạo năng lượng và các chất trao đổi cho quá trình đồng hoá xảy ra ở mô cơ. Sự khác nhau chính về dinh dưỡng giữa động vật trên cạn và động vật dưới nước là khả năng sử dụng các loài thực vật làm thức ăn. Động vật trên cạn lợi dụng khả năng tự dưỡng của thực vật, biến đổi năng lượng ánh sáng mặt trời vào trong lá và hạt một cách trực tiếp hoặc gián tiếp trong chuỗi thức ăn. Một vài loài cá sử dụng trực tiếp thực vật nước nhưng phần lớn là các sinh vật phù du (plankton) bao gồm cả thực vật phù du (phytoplankton) và động vật phù du (zooplankton) cũng trực tiếp hoặc gián tiếp trong chuỗi thức ăn. Không có loài cá nào giống với động vật nhai lại. Chương này sẽ giới thiệu các con đường dinh dưỡng chính và nhấn mạnh sự khác nhau giữa các loài cá và giữa cá và động vật trên cạn.
Phần lớn cá sống trong vùng nóng và tốc độ trao đổi chất của chúng bị chi phối bởi nhiệt độ nước. Vì vậy, hầu hết cá có hệ thống enzyme thích nghi với môi trường nhiệt và “isozymes di truyền” của chúng. Những nhóm cá khác nhau có khả năng tiêu hoá và sử dụng polysaccharide phức hợp làm nguồn năng lượng là khác nhau, nhưng cũng thích ứng tốt với việc sử dụng protein và lipid như nguồn năng lượng hoàn hảo. Cá nước ngọt ở vùng nước ấm thường sử dụng nhiều carbohydrate trong khẩu phần và cá ăn thực vật phát triển tốt trong điều kiện sử dụng thức ăn thực vật. Tất cả các loài cá chỉ cần chuyển hoá protein thành amino acid cho đồng hoá để sinh trưởng. Như vậy, tất cả các loài cá được nghiên cứu có hệ thống enzyme phân giải protein phát triển tốt và một vài loài có cả enzyme tripsin có tính acid và base, trái ngược với động vật trên cạn. Nét đặc trưng khác nữa là sự có mặt của enzyme chitinase hoạt động trong nhiều loài với khả năng tiêu hoá và sử dụng 2-aminoglucose polymer có mặt trong vỏ của côn trùng và giáp xác. Phần lớn các loài cá đều sử dụng tốt lipid mà không phát sinh độc ở gan và béo phì, một số loài thuộc họ cá chép hạn chế sử dụng khẩu phần chứa lipid cao. Cá sống trong môi trường nước, rất thuận lợi để tiếp xúc trực tiếp với dung môi tốt cho sự hấp thu chất bài tiết. Vì thế, cá bài tiết chủ yếu nitơ là kết quả của quá trình dị hoá amino acid và chuyển hoá các hợp chất chứa nitơ khác vào nước như amoniac qua mang. Đó là một phương thức tiết kiệm năng lượng được gia tăng bởi giá trị năng lượng chuyển hoá mà cá nhận từ chuyển hoá protein. Như vậy, cá sử dụng nguồn protein có hiệu quả hơn động vật ổn nhiệt trên cạn, chúng phải bài tiết các hợp chất chứa nitơ ở dạng urea hoặc acid uric.
2.2. CHUYỂN HOÁ CARBOHYDRATE Các loài cá ăn thịt tiêu hoá polysaccharide không tốt lắm. Cá hồi và phần lớn các
loài cá nước mặn sử dụng protein và lipid như một nguồn năng lượng chính và các sản phẩm trung gian sử dụng cho sinh trưởng. Cowey (1989) đã tóm tắt hoạt động của

23
amylase, pepsin và trypsin trong cá chép và cá tráp cao gấp 10 lần hoạt động của amylase trong cá chép ở vùng nước ấm so sánh với cá tráp biển.
Glycogen chỉ là dạng carbohydrate dự trữ chính trong gan và cơ của cá, ngay cả trong suốt thời kỳ dài nhịn đói và di trú, glycogen trong gan hoặc cơ của cá hồi cũng chỉ mất rất ít (French et al., 1983). Đó là bằng chứng cho thấy sự hình thành glucose trong cơ thể động vật từ sự dị hoá các amino acid trong suốt thời kỳ nhịn đói. 2.2.1. Sự thuỷ phân glucose
Con đường của Emden - Meyerh cổ điển về sự chuyển hoá glucose tồn tại trong cá như các động vật trên cạn, bắt đầu từ sự hình thành glucose 1-phosphate. Theo đó, glucose 6-phosphate được hình thành, nó được biến đổi thành fructose 1,6-diphosphate, được phân chia thành hai phân tử glyceraldehydes 3 - phosphate và sau đó hình thành hai phân tử pyruvate có thể đi vào chu trình tricaboxylic acid (TCA, sơ đồ: 2.1) như acetyl - coenzyme A (acetyl - CoA). Tất cả các enzyme của chu trình được tìm thấy trong các cơ quan của cá (Tarr, 1972).
Con đường chuyển hóa glucose thành acetyl - CoA kèm theo sự hình thành năng lượng liên kết cao năng adenosine triphosphate (ATP) từ adenosine diphosphate (ADP). Kết quả là từ 1 mol glucose tạo thành 36 mol ATP. Mỗi mol (phân tử) ATP thuỷ phân trong tế bào mang lại 7 kcal năng lượng cho các phản ứng đồng hoá và dị hoá. Hiệu suất sinh học của chuyển hoá hiếu khí của glucose có thể được tính từ các năng lượng liên kết liên quan như sau:
C6H12O3 + 6O2 6CO2 + 6 H2O + 686 kcal Trong khi, sự thuỷ phân glucose bởi enzyme cho 36 ATP x 7 kcal thành 252 kcal,
vì vậy, hiệu suất sinh học theo lý thuyết là 38% (252/686).

24
Trong điều kiện kỵ khí ở cơ cá, glucose được chuyển hóa thành lactate và chỉ 2 mol ATP. Ngoài ra, lactate cần thời gian đáng kể để bài tiết từ cơ với đủ lượng oxy ra môi trường nước. Tarr (1972) đã liệt kê danh sách các enzyme liên quan đến sự thuỷ phân glycogen trong mô cá. Cowey và Walton (1989) đã thảo luận về những phản ứng không thuận nghịch và các nhân tố điều chỉnh có thể có trong sự thuỷ phân và sự hình thành glucose trong cơ thể động vật và ngoài phạm vi của enzyme glycolytic trong cá hồi (Walton và Cowey, 1982). 2.2.2. Tổng hợp carbohydrate
Tổng hợp carbohydrate là một quá trình rất tốn năng lượng trong mô cá. Để tổng hợp 1 mol glucose cần 2 mol pyruvate và sử dụng 6 liên kết phosphate cao năng, nhưng sự chuyển đổi kỵ khí của glucose thành lượng pyruvate chỉ cần 2 mol ATP. Sự hình thành glucose trong cơ thể động vật xảy ra như ở cá, đặc biệt trong suốt thời kỳ nhịn đói và di trú kéo dài, glycogen ở cơ và gan phải cung cấp cho glucose máu cần thiết cho quá trình sống. 2.2.3. Con đường chuyển hóa pentose phosphate
Con đường chuyển hóa pentose phosphate là một phần nhỏ của tổng hợp phân tử ribose và niacin adenine diphosphate (NADP) đã được Walton và Cowey (1982) mô tả ở cá như sau:
D- Glucose 6-phosphate + 2 NADP + H2O → D-ribose 5-phosphate + CO2 + + 2 NADPH + 2 H+ Các chất sau phản ứng khử này có thể được dùng để tổng hợp lipid. Khẩu phần
chứa carbohydrate cao đối với cá da trơn làm tăng mức enzyme glucose-6-phosphate dehydrogenase ở trong gan (Likimandi và Wilson, 1982). 2.2.4. Thức ăn và sự chuyển hoá carbohydrate
Sự cân bằng trong khẩu phần ảnh hưởng đến sự chuyển hoá carbohydrate. Kết quả thí nghiệm của Cowey et al. (1977) cho thấy, cá hồi sử dụng 60% protein và không có carbohydrate dễ tiêu trong khẩu phần có lượng thuỷ phân glucogen cao hơn đáng kể so với cá hồi nuôi khẩu phần thấp protein và với 50% carbohydrate. Ngoài ra, ở khẩu phần có chứa protein cao cá nhịn ăn có tốc độ thủy phân glycogen cao. Hai enzyme glycolytic cho thấy mức độ hoạt động cao, tương tự như cá ăn khẩu phần có lượng protein cao. Buhler và Halver (1961) cho rằng cá hồi sử dụng hexoses đơn dễ dàng và disaccharide tốt nhưng sử dụng polysaccharide kém trong nước lạnh nơi mà cá hồi cư ngụ. Giá trị năng lượng tiêu hoá được tính theo các hệ số sau:
Tinh bột thô: 1.5 - 2 kcal/g Tinh bột nấu chín: 2.5 - 3 kcal Dextrin: 3 - 3.5 kcal Glucose: 4.0 kcal Saccharose: 4.0 kcal Cellulose: 0.1 kcal Dưới đây là giá trị năng lượng tiêu hoá của protein và lipid tiêu hóa (Smith,1989): Protein tiêu hoá: 5.0 kcal/g Lipid tiêu hoá: 8.0 kcal/g
2.3. CHUYỂN HÓA LIPID Những chất béo đơn giản được tiêu hoá bởi enzyme lipase, giải phóng ra acid béo
và glycerol. Những acid béo bị thủy phân thành 2 đơn vị carbon tại một thời điểm trong

25
bước oxy hoá ở vị trí β-cổ điển, acetyl-CoA sinh ra chu chuyển vào chu trình TCA và sinh ra các liên kết phosphate cao năng trong ATP (sơ đồ 2.2). Chuỗi oxy hoá chính được thể hiện ở công thức sau:
Acid béo + ATP và S-CoA → Acid béo - S-CoA
Khử nước để tạo thành β-dehydro acid béo - CoA
Hydrat hoá tạo thành β-hydroxy acid béo - CoA
Oxy hoá tạo thành β-keto acid béo - CoA
Sự tiêu (lysis) bằng CoA-SH tạo thành acid béo-CoA mới và acetyl-CoA
Sau đó, quá trình β-oxy hoá lặp lại các bước trên để sinh ra acetyl-CoA nhiều hơn. Mỗi vòng như vậy làm giảm 2 cacbon của cả chuổi cacbon của acid béo trong thức ăn.
Ví dụ, acid palmitic (C16) tạo thành palmityl-CoA và sinh ra 8 phân tử acetyl-CoA. Trong chu trình đó, 7 flavin adenine dinucleotie (FADH) và 7 diphosphopyridine nucleotide (DPNH) được sinh ra ở dạng khử.
Năng lượng thuần thu được như sau: Acid Palmitic tạo ra palmityl-CoA→ 2ATP 7 Palmityl-CA sinh ra 8 acetyl-CoA 7 FADH tạo ra 7 FP (7x2) → + 14 ATP 7 DPNH tạo ra 7 DPN (7x3) → + 21 ATP
8 Acetyl-CoA tạo ra 16 CO2 + 8 HOH + 8 CoASH (8x12) → + 96 ATP Ước tính năng suất sinh học: 129 ATP (96 + 14 + 21 – 2) x 7 kcal = 903 kcal Nhưng, quá trình oxy hoá hoàn toàn của acid palmitic tạo ra CO2 và H2O sinh ra
2340 kcal. Do đó, hiệu suất sinh học là 39% (= 903/2340). Hiệu suất sinh học của quá trình oxy hoá tế bào acid béo xấp xỉ như carbohydrate hoặc lipid hoặc protein tiêu hoá.
Một vài vitamin tham gia trong các bước khác nhau của quá trình oxy hoá: acid pantothenic trong CoA ở bước 1; riboflavin như flavin adenine diphosphate trong bước 2; biotin và acid ascorbic trong bước 3; riboflavin, niacin và pyridoxine trong bước 4; acid pantothenic lần nữa trong bước 5 và acid lipoic trong suốt chu trình TCA để sử dụng “acetate hoạt hóa” sinh ra ATP đảm bảo nhu cầu năng lượng chuyển hoá trong tế bào (sơ đồ 2.2. Chu trình dinh dưỡng và ATP).
Quá trình oxy hoá acid béo mạch dài có số carbon lẻ Acid béo mạch dài có số carbon lẻ có một lượng nhỏ trong chất nhớt ở trên da cá
và trên bề mặt của lớp vỏ ở côn trùng và giáp xác. Các chất này giống như tác nhân tiêu diệt vi khuẩn khi bị phân giải bởi quá trình β-oxy hoá tạo ra acid propionic 3 cacbon. Chất

26
này không bị phân huỷ bởi phần lớn các vi khuẩn. Quá trình trao đổi các acid béo liên quan đến loại 1 nguyên tử carbon khỏi acid béo và do vậy sinh ra một acid béo mạch dài có số carbon chẵn có thể sau đó β-oxy hoá sản sinh năng lượng. Những enzyme đặc hiệu cho quá trình khử methyl ở cá tương tự như đã tìm thấy ở động vật trên cạn (Nicolaides và Laves, 1958; Gurr và Harwood, 1991).
Chuyển bậc electron Trong phản ứng oxy hoá - khử, các electron được chuyển từ các chất cho electron
đến một chất oxy hoá hoặc chất nhận electron. Có 4 nhóm enzyme chính: Pyridine dehydrogenase xúc tác di chuyển thuận nghịch của electron từ cơ chất đến
niacin adenine dinucleotide (NAD) hoặc phosphate NAD tạo thành NADH hoặc NADPH. Flavin dehydrogenase chứa flavin mononucleotide (FMN) hoặc dinucleotide
(FDA). Các flavin oxidase quan trọng nhất là succinic dehydrogenase vàác Các protein phức sắt - lưu huỳnh có từ 2 đến 8 nguyên tử sắt và số các nguyên tử
sulfur acid không bền tương ứng, khi mà nguyên tử sắt Fe2+ đổi thành Fe3+ như vậy là một electron đã bị chuyển đi.
4. Các cytochrome vận chuyển electron từ flavoprotein đến oxygen. Những hợp chất này chứa phức sắt - porphyrin và có thể thực hiện phản ứng thuận nghịch chuyển Fe2+ thành Fe3+ và ngược lại. Sự vận chuyển electron từ NADH đến oxygen gắn với việc giảm năng lượng tự do. Như đã biết, năng lượng tự do có thể chuyển đổi bởi quá trình phosphoryl hóa (phosphorylation) ADP thành ATP để sinh ra liên kết cao năng, dễ dàng sử dụng như một nguồn năng lượng cho đồng hoá và dị hoá ở trong mô. Cytochrome chứa đồng cũng tham gia vào con đường chuyển hóa này, đồng có thể được dùng cho vận chuyển electron từ trạng thái Cu+ đến Cu+2. Oxy phân tử có thể được sử dụng trong hệ thống có hợp chất hữu cơ hydroxyl hoá. Dioxygenase chèn hai nguyên tử oxy vào cơ chất, trong khi monooxygenase chỉ chèn một nguyên tử oxy. Cytochrome P450 là một đồng tố hoạt động trong quá trình hydroxy hoá bằng enzyme (enzymatic hydroxations).
2.4. CHUYỂN HOÁ AMINO ACID Cá và tất cả các động vật cần một nguồn ổn định các amino acid kết hợp để tổng
hợp protein mô cơ và cùng với quá trình trao đổi chất để tổng hợp protein ở các mô và tổng hợp các hợp chất liên quan đến quá trình trao đối chất như hormone, các hợp chất purine và các enzyme chuyển hoá. Ngoài ra, các amino acid còn để cung cấp năng lượng trao đổi cho cơ thể. Các amino acid dùng cho những mục đích trên được cung cấp từ thức ăn và các amino acid trong protein ở mô cơ. Mức amino acid tự do trong mô thường thấp; phần lớn các amino acid trong cơ thể là những hợp chất của protein ở mô cơ. Như vậy, cần cung cấp thường xuyên các amino acid cho chuyển hoá cần thiết để bảo tồn sự sống của cá. So với chuột, cá phụ thuộc nhiều hơn vào amino acid thức ăn để đáp ứng cho quá trình chuyển hoá. Cowey và Luquet (1983) ước tính dị hoá protein ở mô cá cung cấp khoảng 50% amino acid để sử dụng cho quá trình trao đổi chất; trong khi với chuột con số này khoảng 70% (Millward et al., 1976).
Như đã đề cập ở trên, cá bài tiết nitơ từ quá trình dị hoá amino acid tạo nên chủ yếu là amoniac, trong khi ở động vật có vú hoặc chim là ure hoặc acid uric. Tổng hợp ure và acid uric từ amoniac tiêu tốn năng lượng, do vậy, động vật có vú và chim nhận ít năng lượng chuyển hoá từ sự dị hoá amino acid hơn cá. Điều đặc biệt có thể giải thích tại sao trong điều kiện thí nghiệm, cá không thể tích lũy hơn 55% N khẩu phần, trong khi các động vật có vú tích lũy N cao hơn trong điều kiện lý tưởng (NRC, 1974). Amoniac chiếm khoảng 70-90% tổng lượng chất thải có chứa nitơ ở cá (Mommsen và Walsh, 1992), với 5-

27
15% bài tiết dạng ure, phụ thuộc vào loài cá (Dosdat et al., 1996). Khoảng 80-90% nitơ bài tiết qua mang.
Quá trình dị hoá amino acid xảy ra chủ yếu trong gan do phản ứng khử amin, tức là các phản ứng khử amin có sự tham gia của các enzyme aminotranferase xảy ra ở cytosol và mitochondria. Mỗi một amino acid có con đường dị hóa riêng với sự tham gia của enzyme đặc hiệu và cho sản phẩm cuôi cùng riêng biệt. Vấn đề này chưa được nghiên cứu nhiều ở cá, nó được xem như là những con đường và các phản ứng đã tìm thấy ở động vật cũng tồn tại ở cá. Cowey và Walton (1989) đã liệt kê các enzyme và con đường dị hoá cho mỗi amino acid, và những thông tin này vẫn còn sử dụng đến ngày nay. Bước đầu tiên trong sự dị hoá amino acid là việc di chuyển của nhóm amino (transdeamination) tạo ra hợp chất trung gian có thể xúc tiến sự chuyển hoá trong chu trình TCA để sinh ra năng lượng hoặc sử dụng như cơ chất cho tổng hợp các hợp chất khác. Nhóm amino, chứa nitơ, được chuyển thành α-ketoglutarate tạo thành acid glutamic. Đây là phản ứng liên quan đến các enzyme transaminase trong tế bào chất. Sản phẩm đã loại nhóm amin (deaminated) có thể sau đó được chuyển hoá thông qua con đường oxy hoá để sản sinh năng lượng. Hiệu suất sinh học thuần thu được của chuyển đổi protein thành năng lượng là khoảng 40%.
Acid glutamic được vận chuyển vào mitochondria và bi khử amin bởi glutamate dehydrogenase và tạo ra NH4
+ thoat ra khỏi mitochondria (Walsh, 1997). Giả thuyết đó cho rằng, phần NH4
+ trong “con thoi” amoniac do enzyme glutamine tổng hợp tế bào chất để hình thành glutamine. Glutamine được chuyển vào mitochondria, ở đó nó được chuyển đổi thành NH3. Cơ chế này tồn tại trong thận của bộ cá nhám và giúp thiết lập gradient chemiosmotic cần cho sự hình thành ATP.
Ở cá, hầu như toàn bộ nitơ bài tiết qua mang. Ở gan, acid glutamic loại bỏ nhóm amino (NH2) hình thành α-ketoglutarate và NH3, và sau đó luân chuyển qua hệ thống tuần hoàn của máu đến mang. Chúng được bài tiết bằng khuếch tán trực tiếp từ máu đến nước, khuếch tán trực tiếp NH3 và/hoặc sự trao đổi Na+/NH4
+ chức năng. Cơ chế nào chiếm ưu thế còn phụ thuộc vào môi trường sống của cá ở nước mặn hay nước ngọt. Ở những loài nước ngọt, sự khuyếch tán NH3 là cơ chế chính để bài tiết. Phần lớn cá có xương bài tiết amoniac, một phần sản phẩm bài tiết nitơ của chúng dưới dạng ure. Bộ cá nhám, cá vây tay và một số loài cá xương khác sống trong môi trường đặc biệt là bài tiết ure, và sản phẩm bài tiết nitơ chủ yếu của chúng là ure (Walsh, 1997). Bộ cá nhám và cá vây tay là cá thẩm thấu ure và có thể duy trì sự điều chỉnh thẩm thấu bằng cách duy trì mức ure cao trong mô cá. Ở cá nhám góc có gai (Squalus acanthias), hơn 90% sản phẩm bài tiết nitơ là ure được bài tiết qua mang (Wood et al., 1995). Ure là chất ít độc hơn amoniac, nhưng ở mức tìm thấy ở các loài cá thẩm thấu ure, ure có thể làm kết tủa protein. Bản thân cá tự làm mất tác dụng này bởi trimethylamine oxide (TMAO) tồn tại với hàm lượng cao và trung hoà ảnh hưởng của ure và làm ổn định protein (Somero, 1986).
Bởi vì amoniac là chất độc, nó phải được bài tiết một cách nhanh chóng để ngăn chặn sự tích luỹ lại ở mô. Nồng độ amoniac huyết tương có liên quan đến protein ăn vào (Kaushik, 1980; Jobling, 1981; Kikuchi và cộng sự, 1991) và bắt đầu tăng 3 - 8 giờ sau bữa ăn, phụ thuộc vào loài cá, protein ăn vào, và chất lượng của protein trong thức ăn (Dosdat và cộng sự, 1996). Những loài nước ngọt có mức nitơ trong huyết tương cao sau bữa ăn (6,5 mg/l) hơn những loài nước mặn (cá vược, cá tráp, cá bơn) đều thấp giống nhau (3,5mg/l). Ngược lại, nồng độ ure trong huyết tương ở những loài nước mặn không liên quan đến protein ăn vào và ở loài nước mặn cao hơn 7 đến 8 lần so với loài nước ngọt (44-59 so với 6,5 – 7 mg/l). Sản phẩm bài tiết nitơ cao nhất xuất hiện 3-5 giờ sau bữa ăn đối

28
với cá nhỏ (10g), đối với cá lớn (100g) là 5-8 giờ sau bữa ăn. Khoảng 30 - 38% lượng nitơ ăn vào được bài tiết trong 24 giờ ở cá hồi, cá vược và cá tráp nhưng đối với cá bơn giá trị này chỉ là 21% (Dosdat và cộng sự, 1996).
2.5. CÁC YẾU TỐ ẢNH HƯỞNG ĐẾN QUÁ TRÌNH TRAO ĐỔI CHẤT
2.5.1. Ảnh hưởng của khẩu phần đến sự trao đổi trung gian Lysine trong khẩu phần ăn ảnh hưởng đến tốc độ (tỷ lệ) tổng hợp và thoái biến
protein trong gan và cơ của cá hồi (Garzon và cộng sự, 1994). Cá ăn khẩu phần thiếu lysine tăng tốc độ của sự thoái biến protein, không có thay đổi đáng kể tỷ lệ tổng hợp protein trong gan, trong khi ở trong cơ, thiếu lysine làm tăng đáng kể tỷ lệ thoái biến protein và tốc độ tổng hợp protein. Một số nghiên cứu khác cũng đã xác nhận các enzyme amino acid transferase khác nhau có hoạt tính cao hơn ở cá ăn khẩu phần chứa protein cao so với cá ăn khẩu phần chứa protein thấp, nhưng ảnh hưởng này không đồng nhất (Cowey và Walton, 1989). Protein thức ăn ăn vào ít ảnh hưởng đến hoạt tính của các enzyme dị hoá amino acid.
Cách xác định sự tích lũy và thoái biến của protein Các phương pháp mới và nhạy đã được phát triển để đo tốc độ tổng hợp và thoái
biến protein trong mô sử dụng liều flooding [3H] phenylalanine, bằng cách tiêm (Garlick et al., 1980). Sử dụng phương pháp này, các nhà nghiên cứu đã hiểu bản chất của quá trình chuyển hoá liên quan tích lũy protein trong mô cơ. Houlihan (1991) đã tóm tắt như sau: “Một điều rõ ràng rằng, lượng protein tổng hợp vượt quá lượng tích lũy cho sinh trưởng”. Sự biến đổi giá trị này phụ thuộc protein thức ăn ăn vào, kích cỡ cá và loài, ngoài ra còn có sự khác nhau của các mô cơ. Carter và cộng sự (1993) cho thấy có một mối tương quan giữa protein ăn vào, tổng hợp protein và hiệu suất tích lũy protein (tỷ lệ protein tích lũy/protein ăn vào) trong cá trắm cỏ, nhưng hiệu suất protein sinh trưởng lại được xác định bằng tốc độ của sự thoái biến protein. Cá sinh trưởng nhanh hơn khi có tỷ lệ thoái biến protein thấp, tích lũy protein được tổng hợp cao hơn, hoạt tính RNA cao hơn, năng lực tổng hợp và tốc độ tổng hợp protein thấp hơn. Houlihan và cộng sự (1988) cho rằng tốc độ tổng hợp protein khác nhau giữa các mô cơ: gan > mang > ruột > lá lách > tâm thất > dạ dày > tuyến sinh dục > cơ trắng ở cá tuyết. Cơ trắng ở cá tuyết có hiệu suất sử dụng protein cao nhất và tính được khoảng 40% của tổng protein tích lũy trong cơ thể mỗi ngày, ngoài ra Facuconneau và cộng sự (1995) cũng quan sát thấy đối với các cá “rainbow trout” (loại cá hồi có đốm đen và hai vệt hơi đỏ kéo từ mõm đến đuôi). Houlihan và cộng sự (1986) đã xếp hạng tốc độ tổng hợp và thoái biến protein ở các mô cơ cá “rainbow trout” như sau: mang > tâm thất > cơ đỏ > cơ trắng. Tuy nhiên, việc sắp xếp dựa trên tổng lượng protein tổng hợp mỗi ngày như sau: cơ trắng = mang > cơ đỏ > tâm thất. Nhiều nghiên cứu cho rằng, 76% protein tổng hợp trong cơ trắng cho sinh trưởng, còn ở mang thì chỉ có 4%. Gần đây, Overturf và Hardy (2001) giới thiệu phương pháp đánh giá tốc độ tổng hợp protein (cơ) bằng cách đo hàm lượng mRNA ở mô liên quan với tổng hợp myosin. Nếu có một phương pháp tương tự để đo tốc độ thoái biến protein, thì có thể hiểu sâu hơn về các nhân tố điều chỉnh hiệu suất tích lũy protein và cung cấp công cụ để thiết lập khẩu phần và chọn lọc di truyền của giống cá sinh trưởng nhanh và kinh tế, và các ứng dụng có lợi khác. 2.5.2. Lượng ăn vào và sự trao đổi chất
Lượng ăn vào xác định tốc độ tổng hợp protein ở cá ngừ Đại Tây Dương (Houlihan và cộng sự, 1995), Carter và cộng sự (1993). Các tác giả đã kết luận rằng, protein và acid amin sử dụng cho tái sử dụng và tổng hợp trong cơ thể (cho sinh trưởng) lớn hơn nhiều so

29
với protein bị oxy hoá và thải ra ngoài, làm cho một số cá hồi tích lũy protein một cách hiệu quả hơn. Ở cá bơn, sự khác nhau giữa các cá thể về tốc độ sinh trưởng liên quan đến protein thoái biến, cá lớn nhanh có tốc độ tổng hợp và thoái biến protein thấp (Carter và cộng sự, 1998). 2.5.3. Thành thục giới tính và trao đổi chất
Trao đổi protein trong thời kỳ thành thục khác với các giai đoạn phát triển khác trong vòng đời của cá. Martin và cộng sự (1993) cho rằng có sự quay vòng protein đáng kể và sự phân phối lại
các amino acid trong suốt giai đoạn nhịn đói và thành thục ở cá hồi Đại Tây Dương. Rõ ràng, buồng trứng có nhu cầu lớn nhất về năng lượng và amino acid của cá trong suốt thời kỳ này. Phần lớn các amino acid cần thiết cho sự thành thục của buồng trứng bắt nguồn từ cơ trắng và được hình thành như là một kết quả của sự thoái biến protein. Gần đây, kỹ thuật sử dụng các chất đồng vị, chính là 15N, sử dụng kỹ thuật liều chảy tràn với chất đồng vị phóng xạ của amino acid dùng để xác định những thông tin nói ở trên đã được sử dụng (Owen và cộng sự, 1999). Lợi ích của phương pháp tiếp cận này có thể sử dụng trong những lĩnh vực hoặc tình huống sử dụng chất đồng vị phóng xạ có giới hạn. Một vài nét chính của dòng dinh dưỡng, sử dụng và chuyển hoá ở cá được chỉ ra ở sơ đồ 2.3.
Nâng cao hiệu suất tích lũy protein (protein retention efficiency) Như đã thảo luận ở trên, hiệu suất tích lũy protein bị ảnh hưởng bởi một số nhân tố
nội sinh và ngoại sinh, bao gồm lượng thức ăn ăn vào, mức protein và năng lượng thức ăn, mức amino acid, giá trị sinh học của amino acid, giai đoạn sinh trưởng và tốc độ điều chỉnh về mặt di truyền của protein thoái biến. Hơn một thập kỷ qua, sử dụng protein (nitơ) ở các loài cá nuôi được nâng cao mà không ảnh hưởng đến mức lipid cao trong khẩu phần

30
như cá hồi Đại Tây Dương. Cuối năm 1980, mức protein trung bình trong khẩu phần cá hồi Đại Tây Dương nuôi là 22-25%, với hơn 75% lượng nitơ ăn vào bị bài tiết. Năm 2000, mức protein được tăng lên trên 45% do sự thay đổi mức năng lượng trong khẩu phần (với 35% lipid) và sự cải thiện chất lượng protein. Đối với cá hồi Đại Tây Dương, việc tăng protein thêm nữa là có thể, nhưng điều đó bị giới hạn bởi chuyển hoá thuần của protein. Ở một số loài cá nuôi khác, cơ hội để nâng cao hiệu suất sử dụng protein có thể liên quan đến lipid cao trong khẩu phần.
Nâng cao hiệu suất tích lũy protein ở các loài cá nuôi rất cần thiết để giảm ảnh hưởng đến môi trường nuôi trồng, và ngoài ra làm tăng hiệu quả sử dụng protein trong khẩu phần, thành phần đắt tiền nhất trong công thức thức ăn cho cá. Có thể, tiềm năng cho việc nâng cao tích lũy protein đó là lựa chọn cặp bố mẹ có tốc độ tích lũy protein cao do tốc độ thoái biến protein thấp, nhưng tiềm năng tốt hơn cả là cải thiện việc tích lũy protein thông qua việc lập khẩu phần “protein lý tưởng” bởi tăng mức năng lượng khẩu phần.
Câu hỏi: 1. Trình bày sự chuyển hóa carbohydrate? 2. Trình bày sự chuyển hóa lipid? 3. Trình bày sự chuyển hóa amino acid? 4. Các yếu tố ảnh hưởng đến sự trao đổi các chất dinh dưỡng?

31
CHƯƠNG III NĂNG LƯỢNG VÀ NHU CẦU NĂNG LƯỢNG
Năng lượng mặc dù không phải là chất dinh dưỡng nhưng là yếu tố quan trọng trong khẩu phần thức ăn của động vật thủy sản (ĐVTS). Tất cả khẩu phần thức ăn cho ĐVTS đều phải được phối trộn không những đáp ứng đủ các nhu cầu về dinh dưỡng mà còn thỏa mãn được nhu cầu về năng lượng cho từng giai đoạn phát triển của động vật thủy sản. Nguồn năng lượng này có nguồn gốc từ các chất dinh dưỡng của các loại thức ăn phối trộn trong khẩu phần. Vì vậy, cần phải xác định giá trị năng lượng đối với chúng. Chương này đề cập đến sự chuyển hóa năng lượng của thức ăn trong cơ thể và các phương pháp ước tính giá trị năng lượng của thức ăn, và nhu cầu năng lượng của cá.
3.1. TRAO ĐỔI NĂNG LƯỢNG
3.1.1. Khái niệm chung Năng lượng là đại lượng vật lý đặc trưng cho khả năng sinh ra công của vật chất
(Từ điển tiếng Việt, 1998). Thuật ngữ này không áp dụng trực tiếp trong dinh dưỡng động vật. Đối với dinh dưỡng động vật, năng lượng chính là nhiệt lượng sản sinh ra trong quá trình đốt cháy các hợp chất hữu cơ và biểu thị bằng calori. Calori (cal) sử dụng trong dinh dưỡng là calori 150C (tương đương lượng nhiệt cần thiết để làm nóng 1 g nước từ 14,5 đến 15,50C). Calori có các bội số là kilocalori (kcal = 1.000 cal) và megacalori (Mcal = 1.000 kcal). Joule (J) cũng là đơn vị biểu thị năng lượng và hiện nay đang được nhiều nước sử dụng (1 Joule là 1 kg-m2/s2). Có thể chuyển đổi calori sang joule (J), 1 cal = 4,184 J hay 1 J = 0,239 cal. Joule cũng có các bội số tương ứng là kJ (1.000 J) và MJ (1.000 kJ). 3.1.2. Chuyển hóa năng lượng của thức ăn
Năng lượng các chất hữu cơ của thức ăn được chuyển hóa trong cơ thể cá theo sơ đồ 3.1 như sau:
Năng lượng thức ăn (Năng lượng thô - GE) Năng lượng tiêu hóa (DE) Năng lượng phân (FE) Năng lượng trao đổi (ME) Năng lượng nước tiểu (UE) Năng lượng thải qua mang (GEE) (Metabolisable energy) (Urine Energy) (Gill Excretion Energy) Năng lượng thuần (NE) Sinh nhiệt (HE= Heat Energy) (Tiêu hóa, hấp thu; tạo SP tiêu hóa; tạo chất thải & bài tiết) Năng lượng tích lũy Năng lượng cho duy trì (NE for Maintenence = NEm) (NE for Production = NEp) (Trao đổi cơ bản, hoạt động bắt buộc, điều chỉnh thân nhiệt) Sơ đồ 3.1. Chuyển hóa năng lượng của thức ăn trong cơ thể động vật (Webster, Lim, 2002)

32
Năng lượng thô (Gross Energy - GE) Năng lượng hóa học có trong thức ăn chuyển đổi thành nhiệt năng nhờ đốt cháy bởi
có mặt ôxy. Nhiệt lượng sản sinh ra do đốt cháy hoàn toàn một đơn vị khối lượng thức ăn gọi là năng lượng thô.
Năng lượng thô được xác định bằng máy đo năng lượng (Bomb calorimeter). Giá trị năng lượng thô của một số chất dinh dưỡng và thức ăn như sau (MJ/kg chất khô):
Các tinh chất: Glucose 15,6 Tinh bột 17,7 Xelulose 17,5 Casein 24,5 Mỡ 38,5 Dầu 39,0
Sản phẩm lên men: Axit axetic 14,6 Propionic 20,8 Butyric 24,9 Lactic 15,2 Mêtan 55,0
Mô cơ thể: Nạc 23,6 Mỡ 39,3
Thức ăn: Hạt ngô 18,5 Rơm 18,5 Cỏ khô 18,9 Khô dầu ôliu 21,4 Sữa (4% mỡ) 24,9
Xác định năng lượng thô của các chất hữu cơ thông qua khả năng oxy hóa của chúng và biểu thị quan hệ giữa tỷ lệ cacbon + hydro so với ôxy. Tất cả carbohydrate có tỷ lệ này như nhau nên giá trị năng lượng thô xấp xỉ 17,5 MJ/kg. Mỡ trung tính có hàm lượng ôxy thấp rất nhiều so với cacbon và hydro nên giá trị năng lượng thô cao hơn nhiều (39 MJ/kg) so với carbohydrate. Năng lượng thô của từng axit béo khác nhau do số chuỗi cacbon; chuỗi cacbon càng ngắn (các axit béo bay hơi) thì năng lượng thô càng thấp. Protein có giá trị năng lượng thô cao hơn carbohydrate vì có chứa yếu tố ôxy hóa, N và S. Mêtan có giá trị năng lượng thô cao vì chỉ có cacbon và hydro.
Như vậy, thức ăn chứa nhiều mỡ và dầu thực vật thì năng lượng thô cao còn thức ăn chứa nhiều tinh bột thì thấp năng lượng. Hầu hết các loại thức ăn thông thường có giá trị năng lượng thô khoảng 18,5 MJ/kg (4.400 kcal).
Giống như tất cả các động vật khác, cá cần năng lượng để sống. Chu trình Krebs là nơi sản xuất ATP khởi đầu và thủy phân ATP cho các tế bào có năng lượng. Oxy hóa hoàn toàn 1 mol glucose tạo ra 686 kcal và thủy phân 1 mol ATP cho 8 kcal. Về lý thuyết, thủy phân 1 mol glucose cho ra 85 mol ATP (686/8); tuy nhiên, thực tế chỉ có 39 mol ATP mà thôi, phần còn lại mất qua nhiệt vào môi trường nước.
Năng lượng tiêu hóa (DE- digestible Energy) và trao đổi (ME - Metabolisable Energy)
Năng lượng tiêu hóa (DE = GE – FE) phụ thuộc khả năng tiêu hóa thức ăn của cá. Trong khi đó, năng lượng trao đổi chỉ phụ thuộc năng lượng của N bài tiết qua mang ở dạng NH3 (chứ không phải ure). Vì mất ít năng lượng thải qua mang nên chênh lệch giữa DE và ME ở cá nhỏ hơn ở động vật có vú (bảng 3.1). Giá trị DE và ME của một số loại

33
thức ăn trên một số đối tượng nuôi có thể tham khảo ở các bảng 3.2, 3.3 và 3.4. Tuy nhiên, việc nghiên cứu về năng lượng trên cá chưa được đề cập nhiều ở Việt Nam. Bảng 3.1. Sự sai khác giữa giá trị năng lượng tiêu hóa (DE) và năng lượng trao đổi (ME) ở cá hồi Rainbow trout
Chất dinh dưỡng DE (kJ/g) ME (kJ/g) ± % Glucose Tinh bột chín Bột sống
15,6 10,6 4,8
13,1 9,0 3,0
83,9 84,9 62,5
Bảng 3.2. Giá trị GE và DE của một số chất dinh dưỡng
DE (kJ/g) Chất dinh dưỡng GE (kJ/g) Cá chình Rô phi Cá chép
Protein Mỡ Cacbohydrate
23,9 39,8 17,6
22,2 33,3 6,8
18,9 37,7 16,8
16,8 33,5 14,7
Bảng 3.3. Giá trị DE và ME của một số loại thức ăn cá
DE (MJ/kg) Nguyên liệu Cá da trơn Rô phi Cá hồi
ME (MJ/kg) (Cá hồi)
Ngô (extruded) 30% khẩu phần 60% khẩu phần Bột ngô Lúa mì Bột đỗ tương Khô dầu bông Bột cá Bột cỏ Dầu động vật Bột phụ phẩm gia cầm
4,6 8,5 -
10,7 10,7 11,2 17,2 2,5 - - -
- -
13,0 -
11,2 -
16,1 -
36,4 15,2 11,2
- - - -
12,5-14,8 11,3
14,6-19,8 8,1 -
11,5 7,1-10.2
- - - -
10,8-137 9,5-10,3 12,5-17,3
5,8 - -
5,2-9,4 Trong sản xuất, để dễ ước tính giá trị năng lượng tiêu hoá (DE) của thức ăn, ADCP
(1983) đề nghị sử dụng những giá trị DE sau đây cho các chất dinh dưỡng (bảng 6.4). Giá trị năng lượng tiêu hoá của một số loại thức ăn tính toán trên cơ sở các số liệu ở bảng 6.4 được ghi ở phụ lục 3 “Thành phần hoá học một số loại thức ăn tôm - cá’’.
Bảng 3.4. Giá trị DE của một số chất dinh dưỡng dùng để ước tính DE của thức ăn thuỷ sản (ADCP, 1983)
Chất dinh dưỡng GE (kcal/g) DE (kcal/g) Carbohydrate (không phải rau cỏ) Carbohydrate (rau cỏ) Protein (động vật) Protein (thực vật) Chất béo
4,1 -
5,5 -
9,1
3,00 2,00 4,25 3,80 8,00

34
3.2. NHU CẦU NĂNG LƯỢNG Trong thực tế, ĐVTS có nhu cầu cao về protein hơn các động vật khác và nhu cầu
năng lượng thì lại thấp hơn. Nguyên nhân thứ nhất là ĐVTS không cần năng lượng để duy trì nhiệt độ cơ thể như vật nuôi. Thứ hai, ĐVTS sống trong nước nên hạn chế tối thiểu năng lượng mất đi để duy trì thăng bằng trong khoảng không. Động vật trên cạn phải mất năng lượng để giữ vị trí cơ thể khỏi tác động với trọng lực. Nhiều loài cá có bong bóng nhằm duy trì vị trí cơ thể trong các tầng nước nên cơ của chúng ít hoạt động để giữ yên vị trí của chúng. Nguyên nhân thứ ba là cá thải khoảng 85% chất thải trao đổi dưới dạng NH3 trực tiếp qua mang vào môi trường nước và cần rất ít năng lượng. Mặt khác, động vật có vú phải dùng năng lượng để tạo urea và gia cầm thì tạo acid uric.
Nhiều yếu tố ảnh hưởng đến nhu cầu năng lượng của cá như: hoạt động cơ học, nhiệt độ, kích cỡ, tốc độ sinh trưởng, giống loài và thức ăn tiêu thụ. Hoạt động cơ học bao gồm bơi, lẫn trốn khỏi kẻ thù hay stress. Nhiệt độ nước là yếu tố ảnh hưởng lớn nhất đến nhu cầu năng lượng của cá. Như đã biết, nhiệt độ cá gần giống nhiệt độ môi trường nước xung quanh, khi nhiệt độ nước gần nhiệt độ tối ưu cho từng loại cá thì tăng trao đổi chất, độ ngon miệng, tốc độ sinh trưởng và hoạt động. Ngược lại, khi nhiệt độ nước giảm dưới ngưỡng tối ưu thì tốc độ trao đổi chất giảm. Kích cỡ cá có ảnh hưởng đến nhu cầu năng lượng của chúng, cá nhỏ nhu cầu năng lượng trên đơn vị thể trọng cao hơn cá lớn. Tương tự, cá lớn nhanh cần nhiều năng lượng hơn cá chậm lớn.
Đối với hầu hết động vật trên cạn, carbohydrate là nguồn năng lượng chủ yếu, tuy nhiên, cá không dùng nguồn này nhiều như động vật trên cạn. Lợn có thể tiêu hóa 90% carbohydrate trong khi đó cá trắm cỏ chỉ tiêu hóa 55-60% mà thôi. Carbohydrate chứa 4,1 kcal GE/gram và là nguồn năng lượng rẻ nhất, vì vậy nó là nguồn thức ăn chủ yếu cho các loài cá ăn cỏ và ăn tạp. Protein chứa 5,6 kcal GE/gram, cá sử dụng rất mạnh. Tuy nhiên, protein là nguồn đắt nhất trong khẩu phần thức ăn. vì vậy, protein không phải là nguồn năng lượng chính trong khẩu phần. Lipid chứa khoảng 9.4 kcal GE/gram, là nguồn năng lượng phụ thêm trong thức ăn thủy sản. Vì vậy, lipid và carbohydrate là nguồn năng lượng làm tăng mật độ năng lượng khẩu phần. 3.2.1. Nhu cầu năng lượng duy trì
Nhu cầu năng lượng duy trì là nhu cầu năng lượng chỉ đủ để cho cá không thay đổi thể trọng trong thời gian thí nghiệm. Để xác định nhu cầu năng lượng, người ta tiến hành phương pháp cân bằng cacbon hay cân bằng năng lượng hoặc bằng phương pháp nuôi dưỡng.
Bảng 3.5. Nhu cầu NL duy trì của ba nhóm cá (Guillaume, 1999 theo Lê Thanh Hùng, 2000)
Nhóm cá Khối lượng cá (g) Nhiệt độ (0C) Duy trì (kJ/kg cá/ngày)
Cá chép Nhóm cá da trơn Nhóm cá hồi
80 80
10-20 100 150 300
10 20 25 25 18 15
28 67 84 72
85-100 60

35
Lượng năng lượng cho duy trì chính là phần mất mát tối thiểu qua nhiệt. Nhu cầu năng lượng duy trì của cá thấp hơn động vật trên cạn vì cá tiêu hao ít năng lượng cho sự vận động và giữ thăng bằng cơ thể, cá không có cơ chế điều tiết thân nhiệt, cá bài tiết amonia mà không bài tiết ure hay axit uric.Tuy nhiên, phương pháp nghiên cứu này tiến hành rất khó khăn.
Nhu cầu năng lượng duy trì cho cá bình quân 70 kJ/kg thể trọng hay 50 kJ/kg W0.75 (khi nhiệt độ nước 20-240C). Nhu cầu năng lượng duy trì so với tổng nhu cầu năng lượng hàng ngày chiếm tỷ lệ 14-17% ở cá chép, 17-24% ở cá hồi, còn ở động vật có vú tỷ lệ này là 30-59%. Bảng 6.5 cho biết nhu cầu năng lượng của một số nhóm cá. 3.2.2. Nhu cầu năng lượng cho sự tăng trưởng
Khẩu phần đủ protein, tăng năng lượng thì tăng sinh trưởng, ví dụ:
GE (MJ/kg thức ăn khô) 13,8 16,8 18,6 20,9-18,2 20,5 22,8 24,9 Tăng (% so với BW đầu) 148 257 392 380 - 150 218 283 320
Ở một mức năng lượng, tăng tỷ lệ protein có thể không làm làm tăng tốc độ sinh
trưởng (bảng 3.6)
Bảng 3.6. Ảnh hưởng của năng lượng và protein khẩu phần đến tốc độ sinh trưởng của cá (cá chép W=170g, cung cấp thức ăn ở mức 2% khối lượng cơ thể, t0: 240C)
Protein (% vật chất khô) DE (MJ/kg thức ăn khô) 41,3 46,5 51,4
18,3 20,1
2,01 2,15
1,99 2,17
2,01 2,14
Các kết quả trên cho thấy sự quan trọng của việc duy trì tỷ lệ năng lượng/protein trong khẩu phần của cá.
Câu hỏi 1. Các dạng năng lượng của thức ăn, công thức tính? 2. Nhu cầu năng lượng cho duy trì, sinh trưởng của cá, những yếu tố chi phối nhu
cầu năng lượng cho sinh trưởng? 3. Công thức P/E và cho một số chỉ tiêu P/E thích hợp của một số loài cá?

36
CHƯƠNG IV DINH DƯỠNG PROTEIN VÀ AMINO ACID
4.1. PROTEIN
4.1.1. Phân loại Về khía cạnh chức năng người ta chia protein thành nhiều loại sau: Protein đơn giản là protein chỉ cho axit amin khi thủy phân, bao gồm hai nhóm là
protein sợi và protein cầu. Protein sợi (fibrous protein): giữ vai trò cấu tạo các mô liên kết như collagen,
elastin và keratin. Protein này không hòa tan và bền với các enzyme tiêu hóa, collagen có hydroxyproline, là một axit amin quan trọng của protein này. Elastin cấu tạo gân và mạch máu, chuỗi polipeptid của elastin giàu alanine và glycine. Keratin có hai loại là α- keratin là protein của lông và tóc và β - keratin là protein của lông vũ, da…, keratin rất giàu axit amin chứa lưu huỳnh, ví dụ protein lông chứa tới 4 % lưu huỳnh.
Protein hình cầu (globular protein): là các enzyme, antigen và hormon. Các protein hình cầu chính, gồm:
Albumin: có ở sữa, máu, trứng, thực vật, đặc điểm: hòa tan trong nước, ngưng tụ bởi nhiệt
Histone: có ở nhân tế bào, ở đây nó gắn với deoxyribonucleic. Đặc điểm: hòa tan trong dung dịch muối và không bị ngưng tụ bởi nhiệt, khi thủy phân cho ra nhiều arginine và lysine.
Protamin: là protein kiềm gắn với axit nucleic có nhiều trong tinh trùng của động vật có vú, protamin giàu arginine nhưng không có tyrosine, tryptophan hay axit amin chứa lưu huỳnh.
Globulin protein có trong sữa, trứng, máu. Protein phức tạp loại protein này ngoài các axit amin còn có nhóm không phải
protein như glycoprotein, lipoprotein, photphoprotein và chromoprotein. Glycoprotein là thành phần của niêm dịch có tác dụng bôi trơn, cũng có trong lòng trắng trứng, ovalbumin. Lipoprotein là thành phần chính của màng tế bào. Photphoprotein như cazein của sữa và photphovitin của lòng đỏ. Chromoprotein như hemoglobin, cytochrome hoặc flavoprotein.
Về khía cạnh dinh dưỡng, người ta chia protein thành hai loại: Protein thô gồm cả protein và các hợp chất chứa nitơ không phải protein. Theo quy
ước của ngành thức ăn chăn nuôi và cá, protein thô được tính bằng công thức: Protein thô = N x 6,25 (N: hàm lượng N)
Hợp chất N phi protein là những hợp chất chứa N nhưng không có cấu trúc protein như: axit amin tự do, amin (putresine, histamine, cadaverine..), amid (urea, asparagine, glutamine..), nitrate, alkaloid (nicotine, cocaine, strichnine, morphine..).
Thức ăn thực vật non chứa nhiều hợp chất nitơ phi protein hơn thực vật trưởng thành ( 25-30% nitơ tổng số), thức ăn ủ xanh chứa tối đa 50-60% nitơ phi protein, thức ăn hạt chứa 10% nitơ phi protein. 4.1.2. Vai trò của protein
Protein ngoài vai trò cấu trúc (nguyên liệu tạo các mô và các sản phẩm) còn có những vai trò quan trọng sau:
Tạo các chất xúc tác (enzyme), hormone

37
Thực hiện chức năng vận chuyển như hemoglobin Tham gia chức năng cơ giới như colagen Chức năng bảo vệ như kháng thể Chức năng thông tin như protein thị giác Protein còn có vai trò tạo năng lượng, 1g protein sản sinh ra 4,5 kcal năng lượng (ở
cá). Cá là loại Aminotelic (thải amoniac) khác với động vật có vú là loại Ureotelic và chim là Uricotelic, đối với các loại này 1g protein chỉ cho 4 kcal năng lượng. 4.1.3. Nhu cầu protein của cá
Người ta chia nhu cầu làm hai loại: nhu cầu duy trì và nhu cầu sản xuất: - Nhu cầu protein cho duy trì ở cá cao hơn ở động vật có vú. Ví dụ: cá hồi vân
(Oncorhynchus mykiss) nặng 100g có nhu cầu protein duy trì hàng ngày là 52,1; 69,3 và 97,7 mg/ngày, tương ứng với nhiệt độ môi trường là 100C, 150C và 200C.
- Nhu cầu protein cho sản xuất (cho tăng trưởng) cũng cao hơn động vật có vú 4 lần, gà 2 lần và phụ thuộc vào:
+ Loài cá: ví dụ cá rô phi lớn nhanh hơn hai lần so với cá mè hoa. + Tính biệt: ví dụ cá chép cái lớn nhanh hơn cá chép đực + Tuổi và khối lượng cơ thể: nhu cầu protein tính cho một đơn vị khối lượng cơ
thể ở con vật non cao hơn con vật trưởng thành. Thí nghiệm nuôi dưỡng cá giai đoạn cá bột, cá hương và cá thương phẩm thấy rằng nhu cầu protein cao nhất ở giai đoạn cá bột, sau đó giảm dần, ở giai đoạn cá bột, protein khẩu phần phải đạt 50%, lúc 6 – 8 tuần giảm còn 40% đối với salmon và trout và 35% đối với salmonid ở giai đoạn nuôi thương phẩm.
+ Mật độ đàn: số lượng cá trên đơn vị diện tich mặt nước. + Mức độ hoạt động. + Yếu tố môi trường: nhiệt độ, ánh sáng, độ mặn, nồng độ O2, chất độc hoặc chất
chuyển hóa (như NH3 hay nitrite). Ví dụ: cá hồi (O. tshawytscha) yêu cầu khẩu phần chứa 40% protein khi nhiệt độ
nước là 80C, nhưng ở nhiệt độ nước 140C nhu cầu protein sẽ là 55% tính trên cơ sở khẩu phần vật chất khô.
+ Độ mặn cao thì yêu cầu về protein cũng cao. Ví dụ cá hồi (O. mykiss) yêu cầu protein là 40 và 43,5% khi độ mặn lần lượt là 100/00 và 200/00.
+ Chất lượng protein khẩu phần và cân đối năng lượng: Kanko (1968) đã thấy khẩu phần cá hồi chứa 40% protein sẽ cho tốc độ sinh trưởng tối ưu khi bột cá trắng là nguồn protein chính, nhưng với những khẩu phần giầu năng lượng protein chỉ cần 30% (chú ý cá hồi sử dụng mỡ tốt hơn carbohydrate). Protein có axit amin cân đối và có tỷ lệ tiêu hoá cao sẽ tạo cho nhu cầu protein thấp hơn so với loại protein không cân đối axit amin.
Do bị những yếu tố trên chi phối cho nên khó có được một hướng dẫn chung về protein cho cả kỳ sinh trưởng của cá. Bảng 3.1 và 3.2 sau đây cho biết những kết quả nghiên cứu về nhu cầu protein của cá (các thí nghiệm xác định nhu cầu protein của cá thường làm trên cá giống có khối lượng từ 5 - 50g). 4.1.4. Tỷ lệ năng lượng/protein
Có hai công thức, hoặc là tỷ lệ năng lượng/protein (E/P), hoặc là tỷ lệ protein/năng lượng (P/E):
Năng lượng của khẩu phần (KJ hoặc MJ/kg) E/P = -------------------------------------------------- Protein thô của khẩu phàn (%)

38
Số mg (hoặc g) protein Tỷ lệ P/E = ------------------------------------------------ Năng lượng khẩu phần KJ (MJ) Tỷ lệ E/P tối ưu cho cá chép 450-500 (tính theo DE). Như vậy thức ăn với mức 2%
thể trọng, DE khẩu phần là 16,9 - 20,1 MJ/kg và protein thô của khẩu phần là 30-42%. Khuyến cáo của NRC về nhu cầu protein của một số loài cá ghi ở bảng 4.1.
Bảng 4.1. Nhu cầu protein một số loài cá (NRC, 1993)
Loài Tỷ lệ protein trong khẩu phần (giai đoạn juvenile) Cá hồi Atlantic 45 Cá da trơn 32-36 Cá chép 31-38 Cá trắm cỏ 41-43 Cá hồi vân (Rainbow trout) 40 Cá lóc 52 Cá rô phi 30-34 Cá chình Nhật 44,5
Bảng 4.2. Tỷ lệ P/E hoặc E/P cho tăng trưởng tối ưu của một số loài cá
Giống loài Khối lượng
(g)
Protein tiêu hóa
(%)
DE (kJ/g)
DP/DE (mg/kJ)
Tác giả
Cá trơn Mỹ Cá rô phi Đài Loan Cá chép Cá hồi Salmo gairdneri Cá trê phi Cá tra Cá basa
526 34 10 266 600 50 20 90 94
15 20
22,2 28,8 27,0 27,0 24,4 30
31,5 33 42 40 32 28
9,7 12,8 11,6 13,1 12,8 12,3 12,3 15,1 17,2 18,6*
22,7 22,5 23,2 20,5 19,3 24,6 25,8 22,0 25,1
21,5** 18,6** 14,4**
Page&andrews,1973 Garling&Wilson, 1976 Mangalik, 1986 Mangalik, 1986 Li&Lovell, 1992 El Sayed, 1987 Takeuchi et al., 1979 Cho&Kaushik, 1985 Cho&Woodward, 1989 Machiels&Henken, 1985 Hung L.T, 1999 Hung L.T, 1999
* DE: năng lượng tiêu hoá; GE: năng lượng thô 4.1.5. Đánh giá chất lượng protein thức ăn
Protein của các loại thức ăn khác nhau có chất lượng khác nhau, người ta đo chất lượng protein theo các chỉ tiêu sau:
Protein tích lũy Năn vào–( Nphân+Nnước tiểu) Giá trị SVH (BV) = Protein tiêu hóa
= Năn vào - Nphân
x 100
Năn vào–( Nphân-Ntrao đổi)–( Nnước tiểu- Nnội sinh) = Năn vào – ( Nphân – Ntrao đổi)
x 100

39
Số liệu ở bảng 4.3 giới thiệu giá trị SVH (BV) của các protein khác nhau thí nghiệm trên cá chép thể trọng từ 50 – 100g với khẩu phần chứa 10% protein.
Bảng 4.3. Giá trị BV một số protein thức ăn cá (Ogino và Chen, 1973)
Loại thức ăn
BV Nhận xét
Lòng đỏ trứng khô Cazein Bột cá trắng Gelatin Bột gluten ngô Bột đỗ tương Nấm men từ dầu hỏa
89 80 76 23 55 74
73 - 79
Xử lý Ethanol và Ete không có vitamin C Khử mỡ
BV phụ thuộc vào loài, kỹ thuật chế biến. Ví dụ, bột cá hấp 1270C trong 3,5 giờ có
BV giảm 10 – 20% so với bột cá hấp trong 25 phút. Hiệu quả protein (PER: protein efficiency ratio):
Tăng trọng(g) PER = Protein tiêu thụ(g)
Hoặc một chỉ tiêu tương tự có tên là Giá trị protein sản xuất (PPV:productive
protein value).
Lượng protein của mô đã tăng(g) PPV = Protein tiêu thụ(g) x 100
Ví dụ: Sự liên quan giữa lượng thức ăn cung cấp ( tính theo % thể trọng cá), tăng
trọng trong 4 tuần ( tính theo % thể trọng ban đầu của cá), PER, PPV khi nuôi cá chép con ( khối lượng ban đầu là 40g) được trình bày ở bảng 4.4.
Bảng 4.4. Hiệu quả chuyển hóa thức ăn và hiệu quả lợi dụng protein theo các khẩu phần
Khẩu phần thức ăn Tăng trọng, % FCR PER PPV
1 3 5 7 9
19 81 117 128 101
1,41 1,14 1,41 1,87 2,76
1,46 1,80 1,46 1,10 0,74
28,2 30,3 22,8 18,1 12,1
Thang giá trị hóa học (CS: chemical score) Để xác định CS của một thức ăn nào đó cần biết hàm lượng các axit amin của nó,
đem so sánh hàm lượng từng axit amin của thức ăn với hàm lượng axit amin tương ứng của trứng gà từ đó tính CS.
Ví dụ: Tỷ lệ của lysine lúa mì so với lysine của trứng gà: (2,7/7,2) x 100 = 37,5%

40
CS của lysine lúa mì: 37,5 – 100 = - 62,5
Bảng 4.5. Thang giá trị hóa học của lúa mì
Axit amin Lúa mì (%) Trứng (%) CS Arg His Lys Try Phe Met Met + cys Thr Leu Iso Val
4,2 2,1 2,7 1,2 5,7 2,5 4,3 3,3 6,8 3,6 4,5
6,7 2,7 6,8 1,9 5,4 3,3 5,5 5,5 8,5 7,0 8,2
- 37,31 - 22,22 - 60,29 - 36,84 5,55 - 24,24 - 21,81 - 40,00 - 20,00 - 48,57 - 45,12
Chỉ số amino acid thiết yếu (EAAI = Essential Amino Acid Index)
EAAI = n
n
n
2
2
1
1
aea100
........xae
a.100x
aea.100
a1…an: amino acid của protein thức ăn( g/100g protein) ae1…aen: amino acid của protein trứng gà(g/100g protein)
4.2. AMINO ACID Amino acid là thành phần của protein. Protein tự nhiên có khoảng 23 axit amin. Có
hai loại amino acid là amino acid thiết yếu và không thiết yếu. Đối với tôm và cá có 10 loại amino acid được coi là amino acid thiết yếu. 4.2.1. Các axit amin thiết yếu
Bảng 4.6. Các axit amin thiết yếu của tôm và cá
Amino acid thiết yếu (Essential amino acids)
Amino acid không thiết yếu (Non-essential amino acids)
Arginine (Arg) Histidine (His) Isoleucine (Iso) Leucine (Leu) Lysine (Lys) Methionine (Met) Phenylalanine (Phe) Threonine (Thr) Tryptophan (Try) Valine (Val)
Alanine (Ala) Acid aspartic (Asp) Acid glutamic (Glu) Glycine (Gly) Proline (Pro), Hydroxyproline Cystine, Cysteine (Cys) Tyrosine (Tyr) Serine (Ser) Ornitine (Orn)
Chú ý:Phenylalanine có thể được thay thế một phần bằng tyrosine (thay thế được 5% phenylalanine ở cá da trơn).
Cystine có thể thay thế một phần (60%) methionine, thí nghiệm ở cá da trơn.

41
4.2.2. Nhu cầu amino acid Nhu cầu axit amin của cá ghi ở bảng 4.7, số liệu trong bảng cho thấy:
Bảng 4.7. Nhu cầu amino acid một số loài cá (% vật chất khô)*
Axit amin Cá chép Cá rô phi Cá chình Nhật bản Cá da trơn Cá hồi
Arginine Histidine Isoleucine Leucine Lysine Methionine Phenylalanine Threonine Tryptophan Valine
1,6 0,8 0,9 1,3 2,2 1,2a 2,5b
1,5 0,3 1,4
1,18 0,48 0,87 0,95 1,43 0,75c 1,05d 1,05 0,28 0,78
1,7 0,8 1,5 2,0 2,0 1,2a 2,2b 1,5 0,4 1,5
1,0 0,4 0,6 0,8 1,2 0,6 1,2e 0,5 0,12 0,71
1,4 0,64 0,96 1,76 2,12 0,72f 1,24 1,36 0,3 1,24
* a: không có cystine; b: không có tyrosine, nếu có, tyrosine chiếm 1% nhu cầu phenyalanine; c: có 0,15% cystine; d: có 0,5% tyrosine; e: có 0,3% tyrosine; f: có cystine
Nhu cầu arginine của nhóm cá hồi giai đoạn hương gần giống như gà nhưng lớn hơn của lợn và chuột rất nhiều. Ở động vật có vú và ở gia cầm, cơ thể có thể tự đáp ứng được một phần arginine nhưng cá thì không. Nhu cầu arginine của cá nước mặn thấp hơn cá nước ngọt. Không có sự đối kháng arginine - lysine ở cá.
Nhu cầu histidine cho sinh trưởng của hồi giai đoạn hương là 0,7% tính theo chất khô khẩu phần (1,6g/16gN). Histidine được dùng để hình thành dipeptit carnosine và ansezine. Khử cacboxyl histidine thành histamine. Khẩu phần ăn cá hồi (O. mykiss) có nhiều histamine làm cho thành dạ dày bị loét giống ở gia cầm.
Nhu cầu isoleucine ở nhóm cá hồi giai đoạn cá bột phụ thuộc vào hàm lượng leucine trong khẩu phần, tăng isoleucine thì cũng tăng nhu cầu leucine. Mối tương tác giữa isoleucine và leucine cũng thấy ở cá da trơn.
Đối với cá hồi giai đoạn cá hương, 1,9% lysine/CK khẩu phần (4,8g/16gN) sẽ cho sinh trưởng tốt, nhưng để có sinh trưởng và hiệu quả lợi dụng thức ăn cao thì lysine cần 2,5%. Nhu cầu lysine của cá hồi (O. mykiss) giai đoạn hương thì cao hơn (6,1g/16gN hay 9 g/kg khẩu phần), các giai đoạn sau thì mức lysine tối thiểu là 25 - 30g/kg khẩu phần. Cung cấp lysine thiếu sẽ dẫn đến sinh trưởng kém, thối loét vây và chết.
Axit amin chứa S rất quan trọng đối với dinh dưỡng cá, methionine có nhiều trong protein động vật nên những khẩu phần có quá ít protein động vật, sinh trưởng của cá giảm mạnh. Đối với cá hồi (O. mykiss) sinh trưởng sẽ thoả mãn khi khẩu phần chứa methionine + cystine ít nhất là 13 - 15g/kg VCK khẩu phần (0,85 - 1,05% khẩu phần).
Thiếu methionine (không thiếu cystine) dẫn đến viêm cata thuỷ tinh thể mắt ngoài hiện tượng giảm sinh trưởng. Thuỷ tinh thể bị đục sau 2 - 3 tháng tuỳ theo mức độ thiếu. Tuy nhiên, thừa methionine cũng dẫn đến ức chế sinh trưởng.
Threonine: nhu cầu của cá hồi (O. keta) đối với axit amin này là 1,2%/VCK khẩu phần (3g/16gN), của cá hồi (O. mykiss) là 2,2 - 3,2g/16gN (8g/kg VCK khẩu phần), hàm lượng cao hơn làm giảm sinh trưởng.

42
Tryptophan: thiếu làm tăng chứng vẹo xương (scoliosis) ở nhóm cá hồi (vẹo xương là bệnh biến dạng cột sống do thiếu serotonin). Vẹo xương có thể liên quan đến mức serotonin ở não. Cho uống serotonin - creatinsulphate (0,25 - 3g/kg khẩu phần) giảm sự biến dạng cột sống của cá hồi (O. keta) giai đoạn hương, nhưng không ngăn ngừa được bệnh (chú ý ở cá da trơn không thấy có sự quan hệ giữa scoliosis với tryptophan). Ở nhóm cá hồi, thiếu tryptophan gây viêm cata thuỷ tinh thể mắt. Hiện tượng này xảy ra nghiêm trọng khi nhiệt độ giảm thấp (từ 20oC xuống 10oC). 4.2.3. Vấn đề bổ sung axit amin công nghiệp vào khẩu phần
Cá chép (Cyprius carpio) giai đoạn chưa trưởng thành không thể sinh trưởng được khi cho ăn khẩu phần thức ăn mà protein được thay thế bằng hỗn hợp axit amin công nghiệp có thành phần tương tự.
Cá da trơn cũng không lợi dụng được axit amin tự do bổ sung vào khẩu phần ăn của chúng. Hỗn hợp thức ăn có nguồn protein chủ yếu từ bột cá được thay bằng đỗ tương và bổ sung thêm methionine, cystine hay lysine và hầu hết những axit amin giới hạn đều không cải thiện được tăng trọng.
Tăng arginine vào khẩu phần gelatin của cá da trơn từ 11g lên 17g/kg làm tăng trọng tăng lên rõ rệt, nhưng thêm arginine, cystine, tryptophan hay methionine vào khẩu phần chứa cazein không làm thay đổi tăng trọng và hiệu suất sử dụng thức ăn.
Tuy nhiên họ cá hồi (salmonids) lại có thể sử dụng được axit amin tự do cho sinh trưởng. Bổ sung lysine, tryptophan vào khẩu phần cazein – gelatin, cho thấy sinh trưởng tăng rõ rệt so với khẩu phần này không bổ sung 2 axit amin trên.
Câu hỏi: 1. Vai trò protein trong dinh dưỡng thủy sản? 2. Nhu cầu protein phụ thuộc vào những yếu tố nào? Cho biết nhu cầu protein của
một số loài cá? 3. Cách đánh giá giá trị protein (BV, PER, CS, EAAI) và ứng dụng? 4. Kể tên 10 axit amin thiết yếu đối với tôm và cá, nhu cầu các axit amin này, vai
trò axit amin thiết yếu đối với cá?

43
CHƯƠNG V DINH DƯỠNG LIPID
Lipid đóng vai trò quan trọng trong dinh dưỡng động vật. Chúng có bốn chức năng chính sau đây:
- Cung cấp năng lượng cho các chức năng duy trì và sản xuất, và nguồn dự trữ, khi bị oxy hóa sinh ra năng lượng cao gấp 2,25 lần so với carbohydrate và protein
- Là nguồn cung cấp axit béo thiết yếu - Là chất vận chuyển các vitamin tan trong dầu và vận chuyển điện tử và các cơ chất
trong các phản ứng của enzyme - Thành phần quan trọng của màng tế bào. Trong chương này, chúng tôi xin tóm tắt
một số nội dung liên quan đến khía cạnh dinh dưỡng hơn là sinh lý và sinh hóa của lipit
5.1. KHÁI NIỆM, PHÂN LOẠI VÀ CHỨC NĂNG
5.1.1. Khái niệm Lipid hay chất béo (ether extract -EE) là các hợp chất hữu cơ không tan trong nước
nhưng tan trong nhiều dung môi hữu cơ như benzen, ete, cloroform.., và có các chức năng sinh lý, sinh hóa quan trọng trong cơ thể thực vật và động vật. 5.1.2. Phân loại
Căn cứ cấu tạo và chức năng mà lipit được phân thành các loại ở bảng 5.1.
Bảng 5.1. Phân loại lipid (MacDonald et al., 1988)
Lipid
Có glycerol Không có glycerol
Đơn giản Phức tạp
Sphingomyelin, Glycolipit Phosphoglycerit cerebrosit, sáp,
steroit, terpen
Dầu, mỡ Glycolipit Glactolipit Lecithin Cephalins
Dầu và mỡ Mỡ và dầu có trong thành phần cơ thể của động vật và thực vật, có cùng cấu trúc và
tính chất hóa học nhưng khác nhau về cấu trúc vật lý. Điểm nóng chảy của dầu thấp, ở điều kiện nhiệt độ bình thường chúng ở dạng lỏng, còn mỡ thì ngược lại. Đó là đặc điểm cơ bản để nhận biết dầu và mỡ. Thuật ngữ chất lipit hay chất béo (fat) dùng để chỉ cả hai nhóm.
Cấu tạo dầu và mỡ Dầu, mỡ là este của axit béo với glycerol-là một rượu no ba chức. Khi tất cả ba
nhóm rược được este với axit béo thì tạo thành một hợp chất gọi là triglyxerit hay triacylglycerol. Quá trình phản ứng như sau:

44
CH2- OH CH2- O- CO - R CH- OH + 3R-COOH => CH- O- CO - R + 3H2O CH2- OH CH2- O- CO - R Glycerol Axit béo Triglycerit Triacylglycerol (Triglycerit) khác nhau do đặc tính và vị trí của các axit béo. Vị trí
các axit béo quyết định đặc tính vật lý của các triacylglycerol. Các triacylglycerol chứa nhiều axit béo mạch ngắn và axit béo không no thì có nhiệt độ nóng chảy thấp. Nếu các axit béo khác nhau được nối vào các gốc rượu của glycerol thì công thức chung của triacylglycerol sẽ là:
CH2- O- CO - R1 CH - O- CO - R2 CH2- O- CO - R3 Nhiều bằng chứng cho thấy định dạng của triacylglycerol của dầu mỡ có thể ảnh
hưởng đến khả năng tiêu hóa. Ví dụ như, palmitat (hexadecanoat) phân bố ngẫu nhiên ở các vị trí 1, 2 và 3 thì tỷ lệ tiêu hóa thấp hơn phân bố ở vị trí 2- đây là vị trí thích hợp cho tác động của enzyme pancreatic lipase.
Phần lớn các axit béo có trong tự nhiên có một nhóm carboxyl và một chuỗi cacbon không phân nhánh, chúng là các axit béo no hoặc chưa no. Một vài axit béo quan trọng có trong tự nhiên được trình bày ở bảng 5.2.
Bảng 5.2. Một số axit béo thường gặp trong các dầu mỡ tự nhiên
Tên axit Công thức Nhiệt độ nóng chảy (0C)
Axit béo no (-anoic) (Saturated Fatty Acids – SFA) : Butyric (butanoic) Caproic (hexanoic) Caprylic (octanoic) Capric (decanoic) Lauric (dodecanoic) Myristic (tetradecanoic) Palmitic (hexadecanoic) Stearic (octadecanoic) Arachidic (eiocosanoic) Axit béo chưa no (-enoic)(Unsaturated Fatty Acids - USFA) Palmitoleic (9-hexadecenoic): 16:1 9 hay n-7-16:1* Oleic (octadecenoic): 18:1 9 hay n-9-18:1 Linoleic (octadecadienoic): 18:2 9,12 hay n-6,9-18:2 Linolenic (octadectienoic): 18:3 9,12,15 hay n-3,6,9-18:3 Arachidonic (eicosatetraenoic): 20:4 5,8,11,14 hay n-6,9,12,15-20:4 Eicosapentanoic: 20:5 5,8,11,14, 17 (n-3,6,9,12,15) Docosahexaenoic: 22:6 4,7,10,13,16,19 (n-3,6,9,12,15,18)
C3H7.COOH C5H11.COOH C7H15.COOH C9H19.COOH C11H23.COOH C13H27.COOH C15H31.COOH C17H35.COOH C19H39.COOH C15H29.COOH C17H33.COOH C17H31.COOH C17H29.COOH C19H31.COOH C20H33.COOH C22H35.COOH
-7,9 -3,2 16,3 31,2 43,9 54,1 62,7 69,6 76,3
0 13 -5
-14,5
-49,5
* Công thức viết tắt theo 2 cách khác nhau, chỉ điểm có liên kết đôi Axit béo thiết yếu (Essential Fatty Acid) Năm 1930, axit linoleic được phát hiện là có hiệu quả ngừa được nhiều chứng bệnh
của chuột cho ăn khẩu phần thiếu mỡ như da có vẩy, chậm tăng trưởng, sinh sản và có thể

45
chết. Các triệu chứng trên cũng được phát hiện ở nhiều loại động vật và cả người. Sau đó, axit arachidonic được chứng minh có vai trò như axit linoleic và có thể mạnh hơn các axit linoleic, và γ-linolenic có hoạt tính cao hơn 1,5 lần so với axit linoleic. Các axit arachidonic và γ-linolenic đều được tổng hợp từ axit linoleic trong cơ thể và không hoàn toàn là axit béo thiết yếu. Tuy nhiên, một trong số các bước tổng hợp, δ-6 desaturation, xảy ra rất hạn chế nên hàm lượng các axit trên có thể thấp và cần phải bổ sung từ bên ngoài. Axit α-Linolenic có hoạt lực thấp hơn linoleic nhưng không thể tổng hợp trong cơ thể và được coi là một axit béo thiết yếu khác ngoài axit linoleic.
Cơ chế chính xác về hoạt động duy trì chức năng bình thường của cơ thể của các axit béo thiết yếu chưa được biết rõ ràng, nhưng có thể tập trung vào hai lĩnh vực: (1) thành phần quyết định của cấu trúc lipit-protein của màng tế bào và (2) thành phần quan trọng của nhiều hợp chất gọi là eicosanoit có vai trò điều chỉnh việc tiết các hormon tuyến yên và tuyến dưới đồi.
Các axit béo này được xem là axit béo thiết yếu (EFA) cần thiết cho gà, lợn, cừu và dê. Giống như các axit béo chưa no khác, chúng là thành phần dùng để tổng hợp prostaglandin và thromboxan, là một chất giống như hormon điều hòa nhiều chức năng tế bào bao gồm trong việc đông máu, điều hòa huyết áp và đáp ứng miễn dịch. Động vật hấp thu axit linoleic vào cơ thể có thể chuyển hóa thành axit arachidonic.
Glycolipid Trong cấu trúc phân tử, hai nhóm rượu của glyxerol được kết nối với axit béo còn
một nhóm nối với đường. Lipit của hòa thảo và họ đậu chứa chủ yếu galactolipit (khoảng 60%), đường ở đây là galactose. Axit béo của galactolipit của cỏ hầu hết (95%) là axit α-linolenic và một ít linoleic (2-3%). Vi sinh vật dạ cỏ có khả năng phân giải galactolipit thành galactose, axit béo và glycerol. Ở động vật, glycolipid có chủ yếu trong não và thần kinh. Glycerol được thay bằng sphingosin bazơ chứa N:
CH3.(CH2)12.CH:CH.CH.CH.CH2.OH OH NH2 Cấu trúc thông thường của glycolipit của động vật là: CH3.(CH2)12.CH:CH.CH.CH.CH2-O- OH NH-CO.R Photpholipid Photpholipid là thành phần ban đầu cấu tạo nên phức hợp lipoprotein của các màng
sinh học. Chúng có trong các cơ tim, thận và mô thần kinh. Ví dụ, myelin của axon thần kinh chứa gần 55% photpholipit. Trứng và đậu nành chứa rất nhiều photpholipid. Photpholipid là hợp chất có chứa axit béo, glycerol, axit phosphoric và base nitơ. Lecithine và cephalin là hai đại diện của nhóm lipit này.
Glycerolphospholipid gồm phosphatidyl cholin (PA, còn có tên là lecithin), phosphatidyl ethanolamine (PE, còn có tên là cephalin), phosphatidyl inositol (PI), phosphotidyl serine (PS), phosphatidyl glycerol (PG). Sphingolipit phổ biến nhất là sphingomyelin.

46
CH2.O.CO.R1 CH2.O.CO.C15H31
CH.O.CO.R2 CH.O.CO.C17H33
O CH2.O.PO3.CH2.CH2.N+(CH3)3
CH2.O.P.OH
OH Phosphatidylcholine
Axit phosphatidic
Phospholipid có một vai trò rất quan trọng trong dinh dưỡng vì nó tham gia vào cấu trúc của tất cả các màng cơ bản và gữi vai trò quan trọng trong sự vận chuyển và hấp thụ lipid và tham gia vào các quá trình biến dưỡng trung gian trong cơ thể sinh vật. Phospholipid có vai trò quan trọng trong dinh dưỡng cho động vật thủy sản, đặc biệt là giáp xác. Nó tham gia vào cấu trúc của tất cả các màng tế bào cơ bản, giữ vai trò quan trọng trong sự vận chuyển và hấp thu lipid, tham gia vào quá trình biến dưỡng trung gian trong cơ thể sinh vật. Thường đối với thủy sản, nguồn cung cấp phospholipid chủ yếu là lecithin từ dầu đậu nành. Tất cả các loài giáp xác đều cần lecithin trong suốt quá trình phát triển đặc biệt là giai đoạn ấu trùng (Harrison, 1990).
Ấu trùng tôm biển giai đoạn mysis chết 100% nếu cho ăn khẩu phần thức ăn không có lecithin (Kazanawa và ctv, 1985). Teshima và Kavazawa (1986) nghiên cứu ảnh hưởng của hàm lượng lecithin lên sinh trưởng tôm he Nhật bản đã kết luận rằng: tốc độ tăng trưởng và hiệu quả sử dụng thức ăn sẽ giảm khi chỉ bổ sung lecithin dưới 3%. Đối với tôm hùm triệu chứng của việc thiếu hụt phospholipid là tôm không có khả năng lột xác hoàn toàn (bệnh "molt death: bẩy lột xác), bệnh này kéo dài đến 90 ngày tuổi làm giảm tỷ lệ sống của ấu trùng (Coklin và ctv, 1980). Lecithin lấy từ đậu nành và phosphostidylcholine (PC) đã được chứng minh là cần thiết cho sự sinh trưởng của tôm P. penicillatus (Jenn, 1989), tôm P. monodon (Piedad-Pascula, 1985) và P. chinensis (Kazanawa, 1993). Piedad-Pascula (1985) cho biết tăng trọng của tôm sú giống tăng khi bổ sung 2% lecithin kết hợp với dầu cá Tuyết trong khẩu phần ăn của chúng.
Bảng 5.3. Nhu cầu lecithin của một số đối tượng giáp xác
Loại tôm Nguồn phospholipid Mức tối ưu (%)
Tác giả
Tôm hùm (giống)
Lecithin đậu nành (Hỗn hợp dầu đậu nành, PC, PE và một số loại khác)*
7.5 Conklin và ctv (1980)
He Nhật Bản (P. japonicus)
Lecithin đậu nành (hỗn hợp PC (24%), PE (30%) và một số loại khác)
3 Teshima và ctv (1983)
Tôm sú (P. monodon)
Lecithin đậu nành 2% Piedad-Pascula, 1985
P. stylirostris Lecithin đậu nành 1.5 Bray và ctv (1990) * PC: phosphostidylcholine, PE: Phosphatidylethanolamine Steroit Steroit là nhóm chất gồm sterol, các axit mật, hormôn thượng thận và hormone sinh
dục. Chúng có đơn vị cấu trúc cơ bản chung là nhân phenanthren nối với vòng cyclopentan. Các hợp chất đơn lẻ chỉ khác nhau số lượng và vị trí nối đôi cũng như đặc tính của chuỗi nối với cacbon vị trí 17.

47
Đại diện của nhóm sterol là cholesterol, 7-dehydrocholesterol và ergosterol. Nó cũng là một thành phần chính của màng tế bào động vật, có vai trò quan trọng trong việc điều khiển độ nhớt. Cholesterol cũng là tiền chất của các steroids khác như hormone sinh dục, hormone của tuyến vỏ thượng thận (estrogen, androgen, progesterol, aldosterone, corticosterone) và axit mật. Nồng độ bình thường của cholesterol trong máu là 1,3 - 2,6 g/lit. Vì cholesterol có độ hoà tan rất thấp, khi có nhiều trong máu trong thời gian dài chúng sẽ tích tụ trên vách thành mạch, dần dần cứng lại tạo thành những plaque atherosclerotic. Đây chính là nguyên nhân của của nhồi máu cơ tim và những cơn đau tim.
Giáp xác phải lấy sterol từ thức ăn, mà duy nhất là từ lipid của thức ăn (Kanazawa và ctv, 1971; Castell và ctv, 1975). Một vài dạng của sterol là Cholesterol, phytosterol, isofucosterol, stigmasterol... Trong đó cholesterol được xem như là loại có ảnh hưởng lớn nhất đến sinh trưởng và tỷ lệ sống của nhiều loại giáp xác (Teshima và Kanazawa, 1983). Một số loài giáp xác như cua và tôm hùm sẽ chuyển hoá cholesterol ngoại sinh thành cholesteryl esters, corticoids, kích thích tố sinh dục, và kích thích tố lột xác (Kanazawa, 1985; Teshima và Kanazawa, 1971). Thí nghiệm của Teshima và ctv (1982) trên tôm P. japonicus cho thấy, khi bổ sung 1% cholesterol vào thức ăn cho loài tôm này đã làm gia tăng tỷ lệ sống cũng như tốc độ tăng trưởng. Nhu cầu cholesterol của tôm sú P. monodon được đề nghị bởi Wu (1986) là 0,5%.
Các axit mật (axit glycocholic) khác với nhóm trên ở chuỗi nối gồm 5 cacbon và cuối là nhóm cacboxyl, và được tổng hợp từ cholesterol. Nhóm hormone gồm oestrogen, androgen, progesteron, cortisol, aldosteron và corticosteron. Nhóm hormon thượng thận có vai trò quan trọng trong việc điều chỉnh trao đổi glucose và mỡ.
Eicosanoit Eicosanoit là nhóm hợp chất gồm prostaglandin, throboxan và prostacyclin bắt
nguồn từ axit béo C20. Các prostaglandin và các chất trao đổi của chúng ảnh hưởng đến sự co cơ, dung nạp của tiểu cầu, áp lực thành động mạch và áp suất máu. Chúng kìm chế tiết dịch vị và sản sinh các axit béo từ các mô mỡ và là chất gây viêm nhiễm. Prostaglandin thường dưới dạng PGF2 được sử dụng trong kích thích động dục hàng loạt ở cừu và bò và điều khiển thời gian đẻ của lợn nái.
Nhóm hợp chất eicosanoit liên quan tới các axit béo thiết yếu theo sơ đồ 5.1 sau: KHẨU PHẦN Axit linoleic Axit arachidonic Axit linolenic γ-linoleic Axit dihomo- Axit gammalinoleic Eicosapentaenoic PGE1 PGE2 TXA2 PGE2 PGE3 TXA3 PGI3 PGF1a PGF2a PGF3a
Ghi chú: PGE và PGF là các prostaglandin; PGI là prostacyclin; TXA là thromboxan. Sơ đồ 5.1. Mối liên hệ giữa các axit béo và nhóm eicosanoit

48
Sáp Sáp là este của một axit chuỗi dài và một gốc rượu chuỗi dài. Ơ một số loài cá như
cá sụn, sáp là một thành phần đáng kể của lipid và những loài cá nhỏ thường có khả năng oxy hoá sáp như là nguồn năng lượng.
Terpenes Terpenes được tạo nên từ những đơn vị isoprene liên kết với nhau thành chuỗi thẳng
hay vòng. Isoprene là hợp chất 5 cacbon có công thức: CH2 :C.CH :CH2 CH3 Nhiều isoprene thấy trong thực vật có mùi vị rất mạnh, chúng là thành phần của dầu
lemon và camphor; ở động vật isoprene có trong coenzyme như coenzyme nhóm Q. 4.1.3. Chức năng
Ở thực vật lipit ở hai dạng cấu trúc và dự trữ. Ở dạng cấu trúc, lipit có trong thành phần của màng tế bào và bảo vệ lớp bề mặt của tế bào và chiếm đến 7% lá của thực vật bậc cao. Lipit trên bề mặt tế bào chủ yếu là chất sáp, axit béo và cutin. Lipit còn là thành phần màng của ty thể, màng nguyên sinh chất....ở dạng glycolipit (40-50%) và photpholipit. Lipit dự trữ ở thực vật chủ yếu trong quả và hạt ở dạng dầu.
Ở động vật, lipit là nguồn dự trữ năng lượng chủ yếu dưới dạng mỡ và có thể lên đến 97% trong mô mỡ của gia súc béo phì. Năng lượng từ mỡ cao hơn nhiều so với carbohydrate, 1 kg mỡ ôxy hóa hoàn toàn cho 39 MJ trong khi đó glycogen cho 17 MJ mà thôi. Lipit cấu trúc trong mô của động vật chủ yếu là phospholipit (0,5-1% trong mô cơ và mô dự trữ) và 2-3% trong gan.
Trong khẩu phần, lipit có các tác dụng như cải thiện tính chất lý học của thức ăn như làm bớt độ bụi, tạo mùi vị thơm ngon nên gia súc dễ ăn hơn. Đây là một vai trò khá quan trọng của chất béo trong khẩu phần. Lipit có ảnh hưởng đến các chất dinh dưỡng khác như làm giảm sự sử dụng carbohydrate và protein.
Khẩu phần cá vùng nước lạnh cần nhiều lipid hơn cá vùng nước ấm vì năng lực sử dụng carbohydrate để lấy năng lượng kém hơn.
Takeuchi et.al. (1978) cho biết sinh trưởng của cá hồi vân (rainbow trout) không bị ảnh hưởng khi protein khẩu phần giảm từ 48% xuống 35% nếu lipid tăng từ 15% lên 20%.
Như vậy, khi xây dựng khẩu phần cho tôm và cá không chỉ đảm bảo cân đối tỷ lệ P/E mà còn cần có một tỷ lệ lipid nhất định (đối với nhiều loài cá tỷ lệ này là từ 20% trở lên). Tuy nhiên, quá nhiều lipid có thể làm mất cân bằng E/P và thừa mỡ tích luỹ ở mô và phủ tạng. Steffens et.al. khảo sát ảnh hưởng của việc bổ sung thêm dầu vào khẩu phần cá hồi vân đã thấy sinh trưởng và chuyển hoá thức ăn của cá tăng lên khi lipid khẩu phần tăng từ 4,7% lên 9%, các loại dầu khác nhau cũng cho kết quả khác nhau (bảng 5.3).
Bảng 5.3. Ảnh hưởng của bổ sung dầu đến tăng trọng và FCR của cá
Không thêm dầu
Dầu hướng dương
Dầu gan cá thu Dầu cá
Khẩu phần: - Mỡ % - Protein %
4,7 40,1
9,0 38,2
8,9 38,3
8,2 38,6
Thể trọng ban đầu (g) 35,5 35,2 39,6 34,2 Thể trọng cuối (g) 127,7 169,6 169,1 141,1 Tăng % 261 382 324 313 FCR (kg/kg tăng trọng) 1,98 1,28 1,46 1,57

49
Mức sử dụng tối đa lipid trong thức ăn trên cá nươc ngọt thường thấp hơn cá biển, mức này đối với cá chép là 12-15%, rô phi <10%, trê phi và cá trơn Mỹ 7-10%; cá hồi 18-20%, cá chẽm 13-18%, cá mú 13-14%, cá vền biển 12-15%.
5.2. VAI TRÒ DINH DƯỠNG CỦA AXIT BÉO
5.2.1. Sinh tổng hợp các axit béo của động vật thuỷ sản Tất cả động vật đều có thể tổng hợp được các axit béo no chuỗi dài từ axetat: n CH3COO- → CH3CH2CH2COO- → CH3CH2CH2CH2CH2CH2… Tất cả các loài động vật cũng tổng hợp được các axit béo chưa no bằng cách thêm
những nối đôi vào phía đầu chuỗi chứa nhóm cacboxyl nhưng không có thể thêm những nối đôi vào phía đầu chuỗi chứa nhóm methyl (trừ thực vật).
Sơ đồ sinh tổng hợp các axit béo trên cá và động vật thuỷ sản như sau: Acetate 14:0 (myristic acid) 14:1n5 16:1n5 16:0 (palmitic acid) 16:1n7 18:1n7 18:0 (stearic acid) 18:1n9 20:1n9 20:0 (arachidic acid) 20:1n11 22:1n11
Oleic acid Linoleic acid Linolenic acid 18:1n9 18:2n6 18:3n3 20:1n9 18:2n9 20:2n6 18:3n6 20:3n3 18:4n3 20:2n9 20:3n6 20:4n3
20:3n9 22:3n6 20:4n6 22:4n3 20:5n3
22:4n6 22:5n3
22:5n6 22:6n3
Sơ đồ 5.2. Sinh tổng hợp các axit béo trên cá và động vật thuỷ sản Như vậy, các axit béo họ n5, n7, n9 có thể được cá sinh tổng hợp từ các tiền chất là
các axit béo no, các họ n3 và n6 sinh ra từ tiền chất là axit linolenic (18:3n3) và axit linoleic (18:2n6), các tiền chất này không có trong cơ thể mà hoàn toàn phải lấy từ thức ăn.
Như vậy, hai axit béo linolenic và linoleic là hai axit béo thiết yếu. Xem thêm sơ đồ 4.3 biểu thị những con đường tạo axit béo chưa no và kéo dài chuỗi carbon từ tiền chất là axit linolenic và axit linoleic của Dave A.Higgs và Faye M.Dong (2000) cuối chương. Các axit linolenic và linoleic được gọi là HUFA (highly unsaturated fatty acid) và những axit béo trong hai họ trên có chuỗi carbon dài trên 20 như 20:3n3; 22:4n3; 20:2n6; 22:3n6 được gọi là PUFA (Polyunsaturated fatty acid).

50
5.2.2. Các yếu tố ảnh hưởng đến thành phần axit béo trong động vật thuỷ sản Độ mặn. Cá nước ngọt chứa nhiều axit béo C16 và C18, trong khi cá biển chứa
nhiều axit béo có chuỗi carbon dài hơn như C20 và C22. Ngoài ra, cá biển chứa một tỷ lệ cao các họ axit béo n-3 hơn họ n-6 so với cá nước ngọt.
Tỷ lệ n-6/n-3 thay đổi từ 0,34 và 0,15 lần lượt đối với cá nước ngọt và cá nước biển. Tỷ lệ n-6/n-3 cũng thấy khác nhau đối với loài cá di cư từ biển vào sông hay ngược
lại (bảng 5.4).
Bảng 5.4. Thành phần của axit béo thay đổi khi cá di cư
Plecoglosus altivelis Onchorhynchus masu Biển Nước ngọt Nước ngọt Biển
Axit béo
TG PL TG PL TG PL TG PL No Mono n-6 n-3 n-6/n-3
34,9 27,4 4,4 31,7 0,14
31,8 16,1 2,2 49,4 0,04
35,1 32,0 7,2 23,9 0,30
53,8 35,9 3,2 6,9 0,46
31,9 18,6 4,0 39,8 0,10
37,5 18,6 4,0 39,8 0,10
31,0 43,1 23,0 23,2 0,10
36,0 19,2 1,5 43,1 0,03
Nhiệt độ. Cá vùng ôn đới thường chứa nhiều PUFA trong thành phần axit béo hơn
cá vùng nhiệt đới, tỷ số n-6/n-3 giảm theo sự giảm nhiệt độ. Thức ăn. Tỷ lệ axit béo n-6/n-3 thay đổi rất lớn theo tỷ lệ n-6/n-3 của thức ăn. Khi
cho cá ăn thức ăn chứa nhiều n-6 như mỡ bò, dầu thực vật, cá có khuynh hướng thay đổi tỷ lệ n-6/n-3 trong cơ thể bằng cách tăng tỷ lệ n-6/n-3 và ngược lại khi cho cá ăn thức ăn giàu axit béo n-3.
Cá có khả năng điều hoà số lượng axit béo trong cơ thể, tuy nhiên người ta thấy rằng một khi có lượng axit béo dư thừa nó có thể ức chế sự hấp thu và tích luỹ các axit béo khác. Axit béo 18:2 có thể ngăn cản sự tích luỹ và sử dụng axit béo 16:1 và 18:1. Như vậy, thành phần axit béo trong cơ thể là kết quả của sự điều chỉnh cân bằng giữa axit béo thức ăn và axit béo tồng hợp từ các nguồn chất trong cơ thể.
Mùa vụ. Thành phần axit béo trong cá thay đổi theo mùa. Lượng lipit tổng số và chỉ số iốt của đầu cá mòi có chỉ số iốt hạ thấp nhất vào tháng giêng và tăng cao vào tháng sáu hàng năm. 5.2.3. Vai trò và nhu cầu axit béo thiết yếu
Vai trò dinh dưỡng. Thiếu các axit béo thiết yếu (EFA) có thể gây những rối loạn sau: thối loét vẩy và vây, tăng tỷ lệ chết, viêm cơ tim, giảm khả năng sinh sản (cá chép, cá hồi, cá tráp), giảm sinh trưởng…
Trong quá trình phát triển của trứng và ấu trùng cá, triglyxerite và phospholypide là nguồn năng lượng chính và axit béo họ n-3 (n3-HUFA) cũng giữ vai trò quan trọng, khẩu phần thiếu họ axit béo này, tỷ lệ chết tăng cao trong vòng 19 ngày (thí nghiệm trên cá tráp). Cũng trên loài cá này, Fernandez Palacios et al. (1995) báo cáo rằng, khẩu phần chứa một nồng độ tối ưu n-3-HUFA trong 3 tuần sẽ làm cho khả năng sinh sản, bao gồm tỷ lệ đẻ, tỷ lệ nở và chất lượng ấu trùng được cải thiện rõ rệt.
Tuy nhiên, cần chú ý rằng khẩu phần thừa n3-HUFA hay tỷ lệ EPA (eicosapentaenoic acid chưa este hoá- 20:5n3) và DHA và arachidonic acid không thích hợp có ảnh hưởng xấu đến tất cả khả năng sinh sản của tôm và cá.
Thành phần axit béo khẩu phần cũng có ảnh hưởng đến khả năng miễn dịch của cơ thể. Trong một nghiên cứu trên cá hồi, Thomson et al., (1996) đã thấy rằng khẩu phần đầy

51
đủ axit béo n-3 nhưng tỷ lệ n-3/n-6 thấp thì sức đề kháng với bệnh Aeromonas salmonicida và Vibrio anguillarum kém hơn khẩu phần có tỷ lệ n-3/n-6 cao.
Nhu cầu EFA của cá. Nhu cầu EFA của cá khác nhau theo loài và cho đến nay cũng chưa được hiểu biết một cách đầy đủ, bảng 5.5 giới thiệu nhu cầu axit béo thiết yếu của một số loài cá.
Bảng 5.5. Nhu cầu các axit béo quan trọng (EFA) của cá
Loài Nhu cầu EFA Nguồn tài liệu
Cá nước ngọt Ayu Cá da trơn Cá hồi Chum Cá hồi Coho Cá chép Cá chình Nhật Bản Cá hồi (Rainbow trout) Rô phi Nile Rô phi Zillii Cá vược Cá biển Cá tráp hồng Cá pecca (cárô biển) Cá sọc vằn Cá bơn Cá cam
1% linoleic acid hay 1% EPA 1-2% linoleic acid hay 0,5-0,75 EPA và DHA 1% linoleic acid và 1% linolenic acid 1-2,5%linolenic acid 1% linoleic acid và 1linolenic acid 0,5 linoleic acid và 0,5% linolenic acid 1% linolenic acid 0,8% linolenic acid 20% lipid dưới dạng linolenic acid hoặc 10% lipid dưới dạng EPA và DHA 0,5% linoleic acid 1% linoleic acid hay 1% arachidonic acid 0,5% EPA và DHA 0,5% EPA và DHA hay 0,5% EPA 1% EPA và DHA 1,7% EPA và DHA hay 1,7% DHA 0,8% EPA và DHA 2% EPA và DHA
Kanazaw et al. (1982) Satoh et al. (1989) Takeuchi &Watanabe (1982)Yu & Sinnhuber (1979) Watanabe et al. (1975), Takeuchi & Watanabe (1977) Takeuchi et al. (1980) Gastell et al. (1972) Watanabe et al. (1974) Takeuchi&Watanabe(1977) Takeuchi et al. (1983) Kanazawa et al. (1980) Webster & Lovell (1990) Yone et al. (1971) Buranapanidgit et al(1989) Watanabe et al. (1989) Gatesoupe et al. (1977) Deshimaru and Kuroki (1983)
Nguồn thức ăn EFA. Nói chung, dầu thực vật (dầu lạc, dầu bông, dầu lanh, dầu dừa,
dầu ngô, dầu cải dầu), mỡ lợn, mỡ bò, cừu nhiều axit béo họ ω6, còn dầu cá biển (cá hồi, cá thu, cá sardin) nhiều axit béo họ ω-3.

52
18: 2n-6 (Linoleic acid) 18: 3n-3 (Linolenic acid) Desaturase 18: 3n-6 (γ linolenic acid) 18: 4n-3 (Octadecatetraeinoic acid) Elongase 20: 3n-6 (Diomo γ linolenic acid) 20: 4n-3 (Eicosatetraeinoic acid) Desaturase 1-series prostanoic 20: 4n-6 (Arachidonic acid, AA 20: 5n-3(Eicosapentanenoic acid, EPA
nonesterified) non-esterified) Cyclooxygenase Lipoxygenase 2-series prostanoid 3-series prostanoid 4-series leukotrienes 5-series leucotrienes Elongase & lipoxins & lipoxins 22: 4n-6 (Docosatetraeinoic acid) 22: 5n-3 (Docosapentaeinoic acid) Elongase Eicosanoids 24:4n-6 22: 5n-6 24:5n-3 22: 6n-3
(Docosahexaenoic acid, DHA)
Desaturase 24: 5n-6 β-oxidation 24: 6n-3 β –oxidation
Sơ đồ 5.3. Con đường chuyển hoá axit béo họ n6 và n3 (Dave A.Higgs và Faye M.Dong, 2000)
Câu hỏi: 1. Vai trò lipid đối với cá. Cho một số con số về nhu cầu lipid của một số loài cá? 2. Phân loại, cách gọi tên và ký hiệu của axit béo no và không no? 3. Đặc điểm chuyển hoá axit béo của cá. Từ axit linoleic và axit linolenic sẽ cho
những axit béo nào trong họ n6 và n3? 4. Vai trò và nhu axit béo chưa no n3 và n6 của một số loài cá?

53
CHƯƠNG VI CARBOHYDRATE VÀ NHU CẦU ĐỐI VỚI CÁ
6.1. KHÁI NIỆM Tên carbohydrate bắt nguồn từ tiếng Pháp hydrate de carbone, là hợp chất trung tính có
chứa carbon, hydro và oxy, và tỷ lệ hydro và oxy giống như cấu tạo của phân tử nước. Phần lớn carbohydrat có công thức cấu tạo chung là (CH2O)n trong đó n từ 3 trở lên.
Định nghĩa trên không thật chính xác khi phát hiện ra những carbohydrate có chứa không những C, H, O mà còn có phospho, nitơ và hưu huỳnh. Hơn nữa, một số hợp chất như là deoxyribose (C5H10O4) không có tỷ lệ hydro và oxy như trong phân tử nước.
Quan điểm hiện tại để định nghĩa carbohydrate là những polyhydroxy aldehyt, xeton, rượu hoặc axit hay những dẫn xuất đơn giản của các hợp chất kể trước đó và bất cứ hợp chất nào mà đều có thể bị thủy phân cho ra chúng.
Trong thức ăn thực vật, carbohydrate chiếm tỷ lệ lớn hơn bất cứ chất dinh dưỡng nào khác. Carbohydrate là nguồn cung cấp năng lượng chủ yếu và là nguồn năng lượng ban đầu cho các hoạt động của cơ thể. Khác với thức ăn thực vật, thức ăn động vật có hàm lượng carbohydrate thấp, chiếm khoảng 1-1,5%, tuy vậy, chúng giữ vai trò không kém phần quan trọng. Glycogen được hình thành từ nhiều phân tử glucose làm nhiệm vụ dự trữ năng lượng trong cơ thể.
6.2. PHÂN LOẠI CARBOHYDRATE Carbohydrate có thể phân chia theo bản chất hóa học thành 2 nhóm chính: nhóm các loại
đường và nhóm không chứa đường (Bảng 6.1). Nhóm đường đơn giản nhất bao gồm các monosaccharide chia làm các nhóm phụ triose, tetrose, pentose, hexo và heptose, phụ thuộc vào số nguyên tử cacbon trong phân tử. Monosaccharide liên kết với nhau để tạo thành di, tri hoặc tetra-polysaccharide.
Thuật ngữ “đường” giới hạn với carbohydrat chứa ít hơn 10 đơn vị monosaccharide, đồng thời thuật ngữ oligosaccharide (oligos tiếng Ai Cập là một vài) dùng để chỉ các loại đường trừ monosaccharide.
Polysaccharide, còn gọi là glycan là các polyme của các đơn vị monosaccharide. Chúng được phân thành 2 nhóm: nhóm homoglycan chứa một đơn vị monosaccharide và nhóm heteroglycan khi thủy phân cho ra nhiều đơn vị monosaccharide. Khối lượng phân tử của polysaccharide rất khác nhau từ 8.000 trong một vài loại fructan của thực vật đến 100 triệu trong amylopectin của tinh bột. Quá trình thủy phân polysaccharide thành đường chịu ảnh hưởng bởi hoạt động của các enzyme đặc hiệu hoặc axit.
Carbohydrate phức tạp chứa carbohydrate kết hợp với phân tử không chứa carbohydrate, bao gồm glycolipit và glycoprotein.

54
Bảng 6.1. Phân loại carbohydrate Triose
C3H6O3 Glyceraldehyte
Dihydroxyacetone Treose
(C4H8O4) Erythrose
Monosaccharide Pentose (C5H10O5)
Arabinose Xylose Xylulose Ribose Ribulose
Đường
Hexose (C6H12O6)
Glucose Galactose Mannose Fructose
Heptose (C7H14O7)
Sedoheptulose
Disaccharide
Succarose Lactose Maltose Cellobiose
Oligosacharide Triasaccharide
Raffinose Kestose
Tetrasaccharide Arabinan Xylan
Stachynose
Homoglycan
Glucan
Tinh bột Dextrin Glycogen Cellulose Callose
Polysaccharide
Fructan Galactan Mannan Glucosamine
Inulin Levan
Hợp chất không chứa đường
Heteroglycan
Hợp chất pectic Hemicellulose Gum Các chất nhầy axit Axit hyaluronic Chondroitin
Carbohydrate Cacbon phức tạp
Glycolipit Glycoprotein

55
1CHO CHO
HO3CH HCOH
H2COH HOCH
H4COH HOCH
H5COH HOCH
6CH2OH CH2OH
D-Glucoz L-Glucoz
6.2.1. Monosaccharide Cấu trúc. Công thức của glucose có thể viết ở dạng thẳng như sau: Rõ ràng là phân tử monosaccharide có chứa nhóm aldehyte (CHO) và đường chứa nhóm
này thuộc nhóm aldose. Do sự có mặt của 4 nguyên tử cacbon trong aldohexose nên có 16 đồng phân lập thể, 8 dạng D và 8 dạng L. Công thức chuỗi của hexose có thể chứa một nhóm xêton (CO) thay vì nhóm aldehyte, ví dụ trường hợp của fructose.
Monosaccharide tồn tại ở dạng tự do, chỉ có ở một số thực vật và mô cơ động vật. Trong thực vật, monosaccharide là tiền thân của polysaccharide (đường đơn và đường đa). Đặc tính chung của monosaccharide là hoạt động như những chất khử do sự có mặt các nhóm aldehyt hay xeton. Đặc tính khử của đường thể hiện ở chỗ: khử ion kim loại như Zn hoặc Cu trong môi trường kiềm. Nhóm aldehyt hay xeton có thể bị khử bởi hóa học hoặc bởi enzyme để hình thành rượu của đường tương ứng.
Các nhóm monosaccharide quan trọng Hexose. Đường quan trọng nhất của hydrat cacbon trong thức ăn và trong cơ thể động
vật là đường 6 cacbon gọi là hexose. Đại diện của đường 6 cacbon có: glucose, galactose, fructose và mantose. Đường hexose tồn tại ở dạng tự do rất ít (ví dụ, fructose có trong nước hoa quả, glucose ở trong máu).
Pentose. Đường pentose (đường đơn có 5 nguyên tử cacbon trong phân tử có arabiose và xylose. Chúng là những đơn vị cơ bản để cấu trúc nên đường pentose loại đường này có nhiều ở vách tế bào thực vật; hoặc đường ribose và deroxyribose là đơn vị cơ bản của axit nucleotic và các enzym. Dẫn xuất quan trọng nhất của đường hexose là đường amino (thành phần của mucopolysaccharide và glycoside).
Heptose. D-sedohetulose là ví dụ quan trọng của monosaccharide chứa 7 cacbon. Đường heptose xuất hiện dạng photphat hay chất trung gian trong con đường chuyển hóa photphat pentose.
Dẫn suất của monosaccharide Este axit photphoric. Este của axit photphoric với đường có vai trò quan trọng trong các
phản ứng trong cơ thể sống. Este quan trọng nhất là với glucose ở vị trí nguyên tử cácbon 1 hoặc 6 hoặc cả hai.
Đường có nhóm amino. Nếu nhóm hydro ở nguyên tử cacbon số 2 của aldohexose được thay thế bởi nhóm amino (-NH2) thì hợp chất mới là một đường có nhóm amino. Trong tự nhiên tồn tại hai nhóm quan trọng là D-glucosamine là thành phần chính của chitin (trong vỏ tôm, châu chấu..) và D-galactosamine là thành phần polysaccharide của sụn.
1CH2OH CH2OH
HO3CH HCOH
2C=O O=C
H4COH HOCH
H5COH HOCH
6CH2OH CH2OH
D-Fructoz L-Fluctoz

56
Đường deoxy. Thay thế nhóm hydroxyl bởi hydro hình thành đường deoxy. Dẫn xuất của ribose, deoxyribose là thành phần cấu tạo của axit deoxyribonucleic (DNA). Tương tự như vậy, dẫn xuất của galactose là fucose và mannose là rhamnose, và chúng thành phần của heteropolysaccharide.
Axit từ đường. Các aldose có thể bị oxy hóa thành axit, trong đó có các axit quan trọng sau: axit aldonic, axit aldaric và axit uronic. Đối với glucose thì có các dẫn xuất axit sau: gluconic, glucanic và glucoronic.
Rượu từ đường. Các đường đơn giản có thể bị ôxy hóa thành rượu polyhydric, ví dụ, glucose thành sorbitol, galactose thành dulciol, còn mannose và fructose đều thành mannitol. Mannitol xuất hiện trong cỏ ủ silô do hoạt động của vi khuẩn phân giải fructose.
Glycoside. Nếu hydro của nhóm hydroxyl bất kỳ của glucose được thay bởi este hay alcohol hay phenol hình thành nên glucoside. Tương ứng, galactose hình thành galactoside và fructose thành fructosit. Oligosaccharide và polysaccharide thuộc nhóm glycoside khi thủy phân tạo thành đường hoặc dẫn suất của đường.
Glycoside cyanogenetic khi thủy phân giải phóng HCN-có tính độc nên những thực vật chứa nhóm glycoside trên đều độc đối với gia súc. Bản thân glycoside không gây độc và phải thủy phân chúng trước khi gây độc. Tuy nhiên, glycosit dễ dàng bị thủy phân bởi enzyme có mặt trong thực vật. Ví dụ, linamarin (còn gọi phaseolunatin) có trong sắn, đậu Java và hạt lanh khi bị thủy phân (hydrolysis) tạo thành HCN, glucose và acetone. Một số glycoside quan trọng có trong tự nhiên trình bày ở bảng 6.2.
Bảng 6.2. Một số glycoside cyanogenetic tự nhiên quan trọng
Tên Nguồn Sản phẩm khi thủy phân ngoài glucose và HCN
Linamarin Đậu Java, sắn và hạt lanh Acetone Vicianin Vicia angustfolia Arabinose, benzaldehyt Amygdalin Mận, ruột quả đào, dứa và quả của
Rosaceae Benzaldehyte
Dhurrin Lá kê (Sorghum vulgare) p-hydroxy-benzaldehyt Lotaustralin Trefoil (Lotus australis), cỏ ba lá
trắng (Trifolium repens) Methylethyl xeton
6.2.2. Oligosaccharide Nhờ sự liên kết từ 2 đến 10 monosaccharide và giải phóng nước, xuất hiện
oligosaccharide. Oligosaccharide có ba nhóm: di-, tri- và tetrasaccharide. Disaccharide Nhóm này xuất hiện nhiều trong tự nhiên và có những đường quan trọng sau: saccarose,
maltose, lactose và cellobiose khi thủy phân cho hai phân tử hexose: C12H22O11 + H2O = 2C6H12O6 Đường saccharose có nhiều trong mía và củ cải đường, có cấu trúc phân tử: α-D-glucose
+ β-D-fructose. Mía chứa 20% đường saccharose, củ cải đường chứa 15-20%... Đường lactose còn gọi là đường sữa là sản phẩm tổng hợp từ tuyến vú. Sữa bò chứa 43-
48 g/kg. Lactose tan trong nước không nhanh và không ngọt như glucose. Lactose chứa 1 phân tử α-glucose liên kết với một phân tử β-galactose qua liên kết β-(1-4). Lactose bị lên men một cách nhanh chóng do vi sinh vật, chủ yếu là Streptococcus lactis. Nhóm vi sinh vật này làm

57
chua sữa do biến đường lactose thành axit lactic (CH3.CHOH.COOH). Ở nhiệt độ 1500C lactose chuyển thành màu vàng, 1750C thành màu nâu, gọi là lactocaramel.
Mantose (đường mạch nha) là sản phẩm phân giải tinh bột và glycogen bởi axit loãng hoặc enzyme. Matose thường có nhiều trong các loại hạt như đại mạch, mì, thóc... Đại mạch hoặc thóc khi lên mầm bị khống chế và đem sấy để làm mạch nha sử dụng cho sản xuất bia. Maltose dễ tan trong nước và không ngọt như saccaroz. Maltose do hai phân tử α-D-glucose liên kết với nhau qua vị trí α-1,4 và có một nhóm hoạt động.
Cellobiose không tồn tại trong tự nhiên như là đường tự do, là sản phẩm phân giải từ cellulose. Cellubiose được tạo thành từ hai β-D-glucose là sản phẩm của phân giải celulose liên kết thông qua mạch nối β-(1 4). Liên kết này không bị bẻ gãy bởi enzym tiêu hóa của động vật có vú, tuy nhiên, bị phân giải bởi enzyme vi sinh vật. Cellobiose cũng chỉ có một nhóm hoạt động.
Trisaccharide Raffinose và cestose là hai đại diện của nhóm trisaccharide. Chúng đều không phải là
chất khử và khi thủy phân cho ba phân tử đường hexose: C18H32O16 + H2O = 3C6H12O6 Raffinose là nhóm đường phổ biến nhất, có hầu hết trong thực vật như là saccharose. Nó
có lượng nhỏ trong củ cải đường và hình thành trong rỉ mật trong khi chế biến đường saccharose. Hạt bông chứa khoảng 80 g raffinose/kg. Khi thủy phân raffinose tạo thành glucose, fructose và galactose. Cestose cũng như dẫn suất của nó có trong từng phần của rau và hạt cỏ hòa thảo.
Tetrasaccharide Nhóm này được hình thành từ bốn monosaccharide. Stachyose thuộc nhóm này, có mặt
hầu hết trong thực vật bậc cao. Nó không phải là đường khử và thủy phân tạo thành hai phân tử galactose, một phân tử glucose và một phân tử fructose:
C24H42O21 + 3H2O = 4C6H12O6 2.3. Polysaccharide (Glycan) Polysaccharide được cấu tạo từ các monosaccharide, gồm có homoglycan, heteroglycan
và lignin. Homoglycan Nhóm carbohydrate này rất khác với nhóm đường. Khác cơ bản là trọng lượng phân tử
cao, hình thành từ nhiều pentose và hexose. Homoglycan không mang đặc tính phản ứng đường của aldose và cestose. Nhiều loại homoglycan có trong thực vật như tinh bột (nguyên liệu dự trữ) và cellulose (chất liệu cấu tạo).
Arabinan và xylan Arabinan và xylan là polyme của arabinose và xylose. Chúng được tìm thấy trong sự kết
hợp với các loại đường khác như là thành phần của heterpglycan. Glucan Tinh bột là một glucan có nhiều trong thực vật như là nguồn dự trữ cacbon hydrat. Nó có
rất nhiều trong hạt, quả, củ và rễ. Tinh bột có trong tự nhiên ở dạng hạt với kích cỡ và hình thù khác nhau.Tinh bột là hỗn hợp của hai polysaccharide khác nhau: amylose và amylopectin, tuy nhiên rất khác nhau về cấu trúc hóa học trừ một số trường hợp. Tỷ lệ của các nhóm trong tinh bột phụ thuôc vào nguồn nhưng amylopectin là thành phần chủ yếu chiếm khoảng 70-80%. Người ta có thể kiểm tra nhanh chất lượng tinh bột thông qua phản ứng với iôt: Amylose cho màu xanh đậm và amylopectin cho màu violet hoặc màu đỏ tía.

58
Amylose có cấu trúc thẳng, đó là liên kết giữa α-D-glucose ở vị trí cácbon 1 và cácbon 4 của phân tử glucose khác. Liên kết α-(1→6) chiếm tỷ lệ rất nhỏ trong amylose. Amylopectin có cấu trúc phức tạp hơn bao gồm liên kết α-(1→4) là chính và cả liên kết α-(1→6).
Hạt tinh bột không tan trong nước lạnh nhưng khi khuấy đều và đun nóng thì bị trương phồng và dẻo. Hạt tinh bột khoai tây trương phồng to và vỡ ra, còn tinh bột hạt cốc thì chỉ trương phồng. Trong thực tế chăn nuôi, gia súc được cho ăn một khối lượng lớn tinh bột của hạt cốc và sản phẩm phụ của hạt cốc.
Glycogen Glycogen là thuật ngữ để chỉ nhóm polysaccarrit mạch nhánh được tách ra từ động vật
hay vi sinh vật. Glycogen có nhiều trong gan, cơ và các mô khác của động vật. Glycogen là những glucan có cấu trúc tương tự amylopectin vì vậy có thuật ngữ riêng cho glycogen là "tinh bột động vật". Glycogen là cacbon hydrat dự trữ ở động vật và đóng vai trò quan trọng trong trao đổi năng lượng.
Khối lượng phân tử glycogen rất khác nhau tùy theo loại động vật, dạng mô cơ và trạng thái sinh lý. Ví dụ, glycogen của gan chuột có khối lượng phân tử 1 - 5 x 108, trong khi đó glycogen cơ là 5 x 106.
Dextrin Dextrin là sản phẩm trung gian của quá trình thủy phân tinh bột và glycogen: Tinh bột dextrin → maltose → glucose Glycogen Dextrin tan trong nước và tạo thành dung dịch giống như gôm (dính). Sự có mặt dextrin
trong thức ăn ngũ cốc hình thành những tính chất như cứng, nướng được và cháy từng phần. Cellulose Cellulose là polyme đơn phổ biến trong thực vật, là cấu trúc vững chắc cho màng tế bào.
Dạng thuần của cellulose được tìm thấy trong sợi bông. Cellulose thuần là một homoglycan có khối lượng phân tử cao được hình thành từ các đơn vị cellobiose (hình 6.1).
Trong thực vật, chuỗi cellulose được hình thành theo trật tự nhất định thông qua cầu nối hydro cả trong và ngoài phân tử. Trong các tế bào thực vật, cellulose liên quan chặt chẽ về cả lý tính và hóa tính với các thành phần khác, đặc biệt là hemicellulose và lignin.
Callose. Callose là thuật ngữ chỉ nhóm polysaccharide chứa liên kết β-(1,3) và β-(1,4). Các β-glucan này có mặt trong thực vật bậc cao như là thành phần của màng tế bào chỉ xuất hiện trong một số giai đoạn phát triển nhất định.
Hình 6.1. Cấu trúc cellulose và chitin

59
Fructan Fructan là nguyên liệu dự trữ trong rễ, thân, lá và hạt của nhiều loài thực vật, nhưng chủ
yếu ở hai họ Compositae và Gramineae. Trong họ Gramineae, fructan chỉ có trong các loài cây ôn đới.
Fructan tan trong nước lạnh và có khối lượng phân tử thấp. Toàn bộ fructan đều chứa các mảnh β-D-frutose liên kết với nhau qua cầu nối 2,6 hoặc 2,1. Fructan có thể chia làm ba nhóm: nhóm levan với đặc trưng liên kết 2,6; nhóm inulin chứa liên kết 2,1; và nhóm fructan có nhánh tìm thấy trong cỏ Agropyron repens. Nhóm này chứa cả hai loại liên kết trên.
Galactan và mannan Galactan và mannan là polyme của galactose và mannose có mặt trong thành tế bào thực
vật. Mannan là thành phần chính của thành tế bào hạt cây cọ và dừa. Trong khi đó, hạt của nhiều cây bộ đậu như cỏ clover, trefoil và medicago chứa galactan.
Bảng 6.3. Khối lượng và số lượng phân tử glucose trong cấu trúc polysaccharide khác nhau
Polysaccharide Khối lượng phân tử Số lượng phân tử glucose Amylose 16.200 - 97.200 100 - 600 Amylopectin 48.600 - 972.000 300 - 6.000 Glycogen 972.000 - 4.050.000 6.000 - 25.000 Cellulose 972.000 - 1.296.000 6.000 - 8.000
Glucosamine Chitin là điển hình của homoglucan chứa glucosamine. Chitin phân bố nhiều trong động
vật bậc thấp và đặc biệt có nhiều trong Crustacae (giáp xác), trong nấm và vài loại tảo xanh. Chitin có cấu trúc giống với cellulose (hình 6.1) và là polysaccharide phổ biến trong tự nhiên sau cellulose.
Heteroglycan Heteroglycan là một loại polysaccharide, trong đó gồm nhiều loại hương vị một số chất
khác. Những heteroglycan có ý nghĩa trong dinh dưỡng bao gồm: hemicellulose, pectin, mucopolysaccharide và glycopotein.
Hemicellulose Hemicellulose là polysaccharide trong màng tế bào tan trong dung dịch kiềm và có liên
kết chặt chẽ với cellulose. Về cấu trúc, hemicellulose có thành phần chính là D-glucose, D-galactose, D-mannose, D-xylose và L-arabinose liên kết với các thành phần khác và nằm trong liên kết glucoside. Hemicellulose cũng có thể chứa axit uronic.
Lignin Lignin không phải là carbohydrate nhưng có liên kết chặt chẽ với nhóm này để cấu tạo
màng tế bào và làm cho cây cứng cáp. Lignin là một polyme bắt nguồn từ ba dẫn xuất của phenylpropal: rượu coumaryl, coniferil và sinapyl. Một phân tử lignin bao gồm nhiều đơn vị phenylpropanoit đan chéo nhau thành liên kết phức hợp.
Fructan (chuäøi saccharose)

60
Rượu coumaryl: R = R1 = H Rượu coniferil: R = H; R1 = OCH3 Rượu sinapyl: R = R1 = OCH3
Trong dinh dưỡng động vật, lignin rất đáng quan tâm vì nó không bị tiêu hóa bởi enzym của cơ thể vật chủ. Lignin còn liên kết với nhiều polysaccharide và protein màng tế bào ngăn trở quá trình tiêu
hóa các hợp chất đó. Gỗ, cỏ khô và rơm rất giàu lignin nên tỷ lệ tiêu hóa thấp trừ khi được xử lý hóa học để bẻ gãy liên kết giữa lignin với các hydrat cacbon khác.
Thực vật càng già, lượng lignin tích tụ càng lớn. Lignin chỉ bị phân giải bởi enzyme của mối, mọt... Lignin chiếm tới khoảng 13-15% trong chất khô của cây cỏ hòa thảo.
6.3. SỰ CHUYỂN HOÁ ĐƯỜNG Ở CÁ Tinh bột Dextrin + Maltose + Glucose
Dextrin Glucose + Maltose
Maltose Glucose + Glucose
Lactose Glucose + Galactose
Sucrose Glucose + Fructose
Cá có khả năng hấp thu tốt glucose, nhưng khả năng sử dụng glucose thì kém hơn động vật trên cạn. Theo dõi trên cá ăn khẩu phần giàu tinh bột hay glucose, thấy rằng lượng đường trong máu tăng lên rất cao và kéo dài, khác với động vật có vú có hàm lượng đường trong máu rất thấp và ổn định. Thực ra, cá có khả năng tiết ra insulin (sau bữa ăn hàm lượng insulin tăng trong khoảng 5-48mg/ml plasma, nhưng khi nhịn đói chỉ có 1-3mg/ml). Insulin có tác dụng biến glucose thành glycogen dự trữ ở gan, cơ và não và từ đó làm giảm lượng đường trong máu. Tuy nhiên, quá trình chuyển glucose thành glycogen còn phụ thuộc vào các “chất nhận” (receptor) có trong tế bào và mối tương quan với glucagon. Các nghiên cứu gần đây cho thấy, cá rô phi thiếu “chất nhận” glucose do insulin đem đến và đã có phản ứng chậm và không hiệu quả đối với insulin. Tuy nhiên, nếu tăng số lần cho ăn thì thấy tăng khả năng sử dụng glucose. Điều này cho kết luận là khả năng chuyển hoá glucose của cá chậm hơn động vật trên cạn, thêm một lượng lớn thức ăn chứa glucose sẽ dẫn đến sự gia tăng đột ngột và lâu dài glucose plasma của cá.
Ở cá con đường sản sinh glucose (glucogenesis) từ protein và lipid là con đường quan trọng. Các axit amin đều có thể được chuyển hoá tạo ra glucose, trong đó alanine, serine và glycine có ưu thế hơn axit glutamic và axit aspartic.
Đối với triglyceride thì sau khi thuỷ phân, glycerol sẽ được phosphoryl hoá tạo ra dihydroxyacetone phosphat rồi hình thành pyruvat và đi vào chu trình Krebs để tạo năng lượng. Còn các axit béo sẽ theo con đường oxy hoá tạo nên acetyl-CoA đi vào chu trình Krebs.
6.4. SỬ DỤNG TINH BỘT VÀ CHẤT XƠ Ở CÁ 6.4.1. Tinh bột
Tinh bột là nguồn năng lượng rẻ tiền hơn protein và lipid và được các nhà sản xuất đưa vào khẩu phần với những tỷ lệ khác nhau tuỳ theo loài cá. Nhóm cá hồi ăn động vật, carbohydrate làm giảm sinh trưởng. Tuy nhiên, ở cá bơn (Pluronectus platessa) và cá chép thì bổ sung tinh bột làm tăng tốc độ sinh trưởng. Tỷ lệ tiêu hoá tinh bột của cá chép trong khoảng 40-80% phụ thuộc vào nguồn tinh bột (bảng 6.4).
α
α
α Lactase
sucrase
R R1OH
CH CH.CH2OH

61
Bảng 6.4. Tỷ lệ tiêu hoá các carbohydrate khác nhau của cá chép 2 năm tuổi (Scerbina, 1973)
Loại thức ăn Hàm lượng carbohydrate (%) Tỷ lệ tiêu hoá Đại mạch (barley) 55,0 74 Yến mạch (oats) 37,3 75 Mạch đen (rye) 46,8 84 Lúa mì (wheat) 43,6 58 Đậu peas 34,1 45 Đậu lupins 22,8 56 Khô lạc (groundnut meal) 15,0 65 Khô đậu tương (soyabean meal) 25,4 51 Thức ăn hỗn hợp 14,8 – 30,5 46 - 75
Lượng tinh bột có thể sử dụng tối đa trong khẩu phần của một số loài cá ghi ở bảng 6.5. Cần chú ý rằng những nhóm cá sử dụng được tinh bột nếu tăng tinh bột trong khẩu phần thì làm tăng hàm lượng lipit cơ thể.
Bảng 6.5. Tỷ lệ tinh bột có thể sử dụng tối đa trong thức ăn một số loài cá
Cá nước ngọt % tinh bột Cá biển % tinh bột Chép Cá da trơn Mỹ Cá hồi Cá rô phi Cá chình
40-45 30-35 25-30 35-40 25-30
Cá măng biển Cá chẽm
Cá bơn Atlantic
35-40 20-25 15-20
(Nguồn: Halver và Hardy, 2002) Để tăng hiệu quả sử dụng tinh bột trong thức ăn thuỷ sản nên áp dụng các biện pháp như
sau: hồ hoá tinh bột qua biện pháp nấu chín, ép viên hay ép đùn để tăng tỷ lệ tiêu hoá tinh bột; và tăng số lần cho ăn để tránh glucose tăng đột ngột sau bữa ăn. 6.4.2. Chất xơ
Hoạt tính enzyme cellulase rất yếu trong đường tiêu hoá của cá. Xơ trong khẩu phần làm tăng sản xuất phân, giảm tỷ lệ tiêu hoá, tăng khối lượng ống tiêu hoá (bảng 6.6).
Tỷ lệ xơ trong khẩu phần cá thường được khuyến cáo từ 8-10%, đối với tôm thì không quá 5%. Nếu xơ không ảnh hưởng đến tỷ lệ tiêu hoá và độ lợi dụng của các chất dinh dưỡng khác, có thể sử dụng xơ như chất pha loãng và để điều chỉnh tỷ lệ P:E khi phối hợp khẩu phần.
Bảng 6.6. Ảnh hưởng của xơ thô đến tỷ lệ tiêu hoá VCK khẩu phần
Xơ thô (%/CK) 0 10 20 Tỷ lệ tiêu hoá VCK (%) Thời gian rỗng dạ dày (phút) Tỷ lệ khối lượng dạ dày/WB
71 782 1,4
66 379 1,8
59 412 1,9
(Nguồn: Werner Steffens, 1985, thí nghiệm trên cá hồi) Câu hỏi 1. Phân loại carbohydrate và ý nghĩa của nó? 2. Vai trò của tinh bột đối với cá và sự lợi dụng tinh bột của cá? 3. Bản chất của chất xơ, cá có sử dụng được chất xơ không?

62
CHƯƠNG VII DINH DƯỠNG VÀ NHU CẦU VITAMIN
Cá được nuôi hàng nghìn năm nay nhưng những biểu hiện thiếu vitamin chỉ mới phát hiện gần đây khi cá được nuôi thâm canh bằng thức ăn công nghiệp. Bổ sung vitamin cho cá trong điều kiện nuôi thâm canh không những thúc đẩy được tăng trưởng của cá mà còn ngăn chăn được những rối loạn bệnh lý do thiếu vitamin. Thông thường, vitamin bổ sung trong thức ăn chỉ chiếm 1-2%, nhưng chi phí lại chiếm tới 15% tổng giá tiền thức ăn.
Có hai nhóm vitamin là vitamin hoà tan trong mỡ (gồm vitamin A, D, E, K) và vitamin hoà tan trong nước (gồm vitamin B1, B2, PP, B5, B6, B12, biotin, axit folic, cholin, vitamin C...).
Dưới đây trình bày vai trò dinh dưỡng, nhu cầu và nguồn cung cấp một số vitamin quan trọng đối với cá, những vitamin khác thì được ghi trong bảng tóm tắt.
7.1. VITAMIN A
7.1.1. Công thức cấu tạo
CH2-OH
CH3 CH3 CH3 CH3
CH3 Các dẫn xuất của Vitamin A. Vitamin A có các dẫn xuất sau: retinol, retinaldehyd,
retinoic, retinilacetat, retinilpropionat, retinilpalmitat. Đơn vị tính là đơn vị quốc tế 1UI = 0,300 microgram retinol = 0,344 microgram retinilacetate = 0,440 microgram retinilpalmitate Các chất tiền vitamin A. Vitamin A không có ở thực vật, nhưng có mặt tiền vitamin ở
dạng các caroteneoit, sẽ chuyển đổi thành vitamin A trong cơ thể động vật. Có ít nhất 80 tiền vitamin được biết bao gồm các α-, β-, γ- caroten, cryptoxanthin có mặt ở thực vật bậc cao và myxoxanthin có mặt trong tảo lục và tảo lam. Không phải tất cả các caroteneoit là tiền vitamin
A. Ví dụ, xantophyl là một sắc tố đáp ứng chủ yếu cho lòng đỏ trứng. Trong các tiền vitamin thì β-caroten có phân bố rộng rãi và hoạt động nhất. Tinh thể β-caroten có màu vàng đỏ và dung dịch có màu vàng cam. Tất cả tiền vitamin đều không tan trong trong nước nhưng tan trong dầu và dung môi hữu cơ. β-caroten rất dễ bị oxy hoá, đặc biệt là ở nhiệt độ cao của không khí. Thức ăn phơi dưới ánh sáng mặt trời hàm lượng β-caroten mất rất nhiều.

63
7.1.2. Vai trò sinh học Vitamin A đóng vai trò quan trọng trong thị giác. Trên tế bào võng mạc mắt có một
quang chất tên là rhodopsin, khi có ánh sáng, rhodopsin bị phân giải thành retinol và opsin, trong tối thì có quá trình tái tổng hợp ngược lại. Rhodopsin tạo nên kích thích thần kinh và gây phản xạ nhìn. Khi khẩu phần thiếu vitamin A động vật bị bệnh quáng gà (hình 7.1).
Vai trò với niêm mạc thượng bì. Tế bào thượng bì do tế bào gốc biệt hoá mà thành. Khi khẩu phần có đầy đủ vitamin A, tế bào gốc biệt hoá hình thành tế bào tiết niêm dịch (đó là các tế bào cuboidal, columna và tế bào goblet), còn nếu khẩu phần thiếu vitamin A, tế bào gốc chủ yếu biệt hoá hình thành tế bào vẩy cá, loại tế bào này tiết ít niêm dịch, lớp thượng bì, niêm mạc sẽ khô, sừng hoá, khả năng ngăn cản sự xâm nhập của vi khuẩn bị giảm (hình 7.2). Tóm tắt như sau:
+ VitaminA → Tế bào Cuboidal, columna và goblet Basal cells (tế bào gốc) - VitaminA → Tế bào Squamous (giống vẩy cá)
Vai trò liên quan đến sức đề kháng của cơ thể.
Thiếu vitamin A, sự sản sinh kháng thể bị giảm thấp. Như vậy cùng với hiện tượng sừng hoá, giảm hoạt động của hệ thống kháng thể đã làm cho sức chống bệnh của cơ thể bị suy giảm. Biểu hiện chung của sự thiếu vitamin A ở cá là: xuất huyết hố mắt, gốc mang cá, nắp mang bị xoắn lại. Cá trơn của Mỹ nuôi bằng khẩu phần có 0,4mg caroten/kg thức ăn, có hiện tượng cá chậm tăng trưởng, mắt lồi, thận xuất huyết. Cá chép thiếu vitamin A sẽ có màu nhợt nhạt, xuất huyết da và vây, biến dạng nắp mang. Tuy nhiên, quá nhiều vitamin A (2,2 triệu UI/kg dưới dạng retinyl palmitat) làm cho cá chậm tăng trưởng, thiếu máu, biến dạng cuống đuôi.
7.2. VITAMIN D 7.2.1. Công thức
Trong tự nhiên có hai vitamin D phổ biến là vitamin D2 và D3 (còn có tên là ergocalciferol và cholecalciferol), tiền của vitamin D2 là ergosterol và tiền vitamin D3 là 7-dehydrocholesterol. Dưới tác động của tia tử ngoại, tiền vitamin D biến thành vitamin D. Dưới đây là công thức của tiền vitamin D3, Vitamin D2 và vitamin D3:
OH
CH3
CH3
CH3
CH3
CH3
CH3
7-dehydro cholesterol

64
OH
CH3
CH3 CH3
CH3
CH3
CH2
Vitamin D3
OH
CH3
CH3
CH3 CH3
CH3
CH3
Ergosterol
OH
CH2
CH3
CH3 CH3
CH3
CH3
Vitamin D2 - 1UI = 0,025 microgram vitamin D3 tinh thể.
7.2.2. Vai trò sinh học Vitamin D3 (cholecalciferol) hấp thu vào máu đến gan, ở gan được thuỷ phân thành 25-
hydroxy cholecalciferol (viết tắt 25 (OH)-vitamin D3), khi đến thận nó lại bị thuỷ phân tiếp để biến thành 1,25(OH)2-vitamin D3 hoặc 24,25(OH)2-vitamin D3. Sản phẩm thuỷ phân 1,25(OH)2-vitamin D3 có hoạt tính mạnh nhất, nó kích thích thành ruột tiết một protein vận chuyển (BP = binding protein), nhờ protein này ion Ca được hấp thu vào máu cũng như vận chuyển Ca vào xương và các sản phẩm khác cùng với phospho (sơ đồ 7.3). Hoạt tính sinh học của D3 trên các loài cá hồi và cá trơn Mỹ gấp 3 lần vitamin D2.
Xương
Ca++ + BP CaBP Ca+ + + BP CaBP Ca++ Ruột BP BP Sữa Máu Trứng Tổ chức
Sơ đồ 7.3. Vai trò vitamin D3 trong hấp thu Ca

65
Chức năng sinh hoá của vitamin D là thúc đẩy sự hấp thu Ca (và cả P) ở ruột để duy trì sự khoáng hoá bình thường của xương. Thiếu vitamin D3: sinh trưởng kém, gan nhiều mỡ, cơ chế homeostasis đối với Ca bị cản trở (biểu hiện co cứng cơ xương).
Hiện tại người ta vẫn chưa hiểu rõ hoàn toàn nhu cầu vitamin D của cá. Ở nhóm cá hồi, người ta thấy nhu cầu vitamin D rất nhỏ, thậm chí khẩu phần không chứa calciferol thì rainbow trout cũng không biểu hiện một triệu chứng nào cả.
Thông thường người ta bổ sung dầu thực vật thì cũng có đủ vitamin D, tuy nhiên với thức ăn viên người ta thường đưa vào 2000-3000 IU vitamin D3/kg thức ăn. Cá trao đổi Ca trực tiếp với nước qua mang, cho nên vùng nước nào nghèo Ca thì mới phải bổ sung Ca cùng với vitamin D.
7.3. VITAMIN E
7.3.1. Công thức Vitamin E có nhiều đồng phân như α-tocopherol, β-tocopherol, γ-tocopherol và δ-
tocopherol, nếu hoạt tính của α-tocopherol là 100, thì các tocopherol β , γ và δ lần lượt là 30-40, 10 và 1.
HO
H3C -CH2-CH2-CH2-CH-CH2-CH2-CH2-CH-CH2-CH2-CH2-CH
CH3 CH3 CH3 CH3
CH3
CH3
CH3
O
α -Tocopherol
7.3.2. Vai trò sinh học Vai trò sinh học chính của vitamin E là chất chống oxy hoá sinh học, ngăn cản sự oxy
hoá các axit béo không no PUFA và HUFA có trong màng tế bào. Thiếu vitamin E thường dẫn đến tổn thương gan, cơ thoái hoá, và cơ quan sinh dục bị ảnh hưởng. Trên cá chép, người ta ghi nhận vitamin E làm tăng khả năng sinh sản. Cá ăn khẩu phần bổ sung vitamin E có hệ số thành thục là 14,1% thay vì 3,3% trên khẩu phần không bổ sung vitamin E. Ngoài ra, vitamin E còn giúp nâng cao tỷ lệ nở của trứng.
Bảng 7.1. Tác dụng của vitamin E và Se bổ sung vào thức ăn cá (Bell et al. 1985; dẫn theo W.Steffens, 1989)
Vitamin E (mg/kg) Se (mg/kg)
2,0 0,06
41 0,06
2,0 0,9
41 0,9
Tăng trọng (%) FCR (kg/kg TT) Vitamin E:
- Máu (microg/ml) - Gan (microg/g)
Tỷ lệ hồng cầu vỡ (%)
2322 1,89 1,7 2,3 51,5
3125 1,62 16,0 36,8 30,9
2976 1,63 2,8 3,4 21,6
3137 1,53 15,9 35,6 20,1

66
Vitamin E và Se có quan hệ hỗ trợ nhau trong việc ngăn trở sự oxy hoá những axit béo không no. Vitamin E có vai trò ngăn cản sự hình thành peroxit, còn Se tham gia vào một enzyme có tên là glutathion peroxidase (GSH-Px), có tác vai trò xúc tác sự phân giải peroxit thành nước:
GSH-Px 2 GSH + H2O2 GSSH + H2O Bổ sung vitamin E và Se vào thức ăn cá có tác dụng làm tăng tốc độ sinh trưởng, FCR và
độ bền của huyết cầu (bảng 7.1).
7.4. VITAMIN K
7.4.1. Công thức Cho đến nay người ta biết vitamin K có 3 dạng hoá học như sau: Vitamin K1 có trong
thực vật có tên là phytokinon, vitamin K2 do vi sinh vật tạo ra có tên là menakinon và vitamin K3 tổng hợp bằng con đường hoá học có tên là menadinon. Hoạt tính của vitamin K3 lớn hơn 2 lần K1 hoặc K2.
O
O CH2-CH=C-CH2-(CH2-CH2-CH-CH2)3H
CH3
CH3 CH3
Vitamin K1
CH3
CH3
(CH2-CH=C-CH2)nH O
O
Vitamin K2
O
O H
CH3
Vitamin K3

67
7.4.2. Chức năng Vitamin K tham gia vào một enzyme hoạt hoá protrombin, cần cho sự đông máu của
động vật trên cạn và cá. Lượng vitamin K 0,5 - 1 mg/kg trong thức ăn đủ để duy trì sự đông máu bình thường trên các loài cá hồi.
Thức ăn động vật như bột cá là nguồn cung cấp quan trọng của vitamin K2. Vitamin K3 bền khi không trộn vào thức ăn hỗn hợp hoặc trong premix (vì cholin chlorid và các ion kim loại xúc tác phân huỷ chúng).
7.5. VITAMIN C (AXIT ASCORBIC) Vitamin C còn gọi là axit ascorbic, hầu hết các loài cá không tự tổng hợp được vitamin
này trong cơ thể (người, khỉ, chuột biển cũng không tổng hợp được vitamin C trong cơ thể). Axit ascorbic là một đồng yếu tố tham gia vào quá trình hydroxin hóa prolin và lysine để
hình thành hydroxyprolin và hydroxylysine trong procollagen (tiền collagen). Như vậy, axit ascorbic cần cho việc hình thành mô liên kết, mô sẹo và khung protein xương.
Vitamin C giúp cho sắt được hấp thu tốt do đó ngăn ngừa được hiện tượng thiếu máu hay gặp ở cá do thiếu vitamin C. Ngoài ra, vitamin C cùng với vitamin E tham gia vào quá trình hạn chế sự hình thành peroxit ở lipit trong mô cá.
Thiếu vitamin C ở cá hồi và trout có biểu hiện biến dạng cấu trúc (vẹo xương-scoliosis, ưỡn lưng-lordosis, sụn mắt, mang và vây bất thường), xuất huyết nội. Những dấu hiệu này xảy ra trước cả những dấu hiệu không đặc trưng như giảm ăn và yếu ớt, kém linh hoạt (anorexia và lethargy). Biến dạng cấu trúc cũng thấy ở cá da trơn, chép, rôphi...
Gần đây, người ta cũng thấy bổ sung vitamin C cho cá da trơn và rainbow trout đã có tác dụng tăng đáp ứng miễn dịch (tăng hoạt tính thực bào của tế bào hệ thống miễn dịch).
Môi trường ô nhiễm kim loại nặng (Yamamoto et.al. 1985), thuốc diệt côn trùng chứa hydrocacbon chlorinate (Mayer et.al. 1978) làm tăng nhu cầu vitamin C của cá.
Vitamin C rất dễ bị phá hủy trong quá trình dự trữ và chế biến, do vậy người ta phải bảo vệ nó trước khi bổ sung vào thức ăn cá.
Vitamin C khi sử dụng cho cá thường ở dạng bọc với ethylcellulose hay bọc với mỡ, dạng phosphorylated ascorbic là dạng khá bền nhưng đắt tiền cho nên cũng it được dùng.
7.6. VITAMIN NHÓM B Vitamin nhóm B thuộc nhóm vitamin tan trong nước gồm có các loại vitamin B1, B2, B6,
B12... Chúng đóng vai trò quan trọng trong quá trình sống của động vật thủy sản. Khi thiếu vitamin nhóm B, chúng thường xuất hiện những triệu chứng bệnh lý ảnh hưởng đến sức khỏe của vật nuôi và thiệt hại đáng kể đến hiệu quả sản xuất. Bảng 7.2. trình bày nhu cầu của cá đối với vitamin nhóm B và các triệu chứng khi thiếu nhóm vitamin này
Bảng 7.2. Tóm tắt những triệu chứng thiếu vitamin nhóm B của cá
Vitamin Nhu cầu (mg/kg)
Triệu chứng thiếu
Thiamin (B1
0,5 10-20*
Chảy máu vây, hiện tượng thần kinh, nhạt màu, kém ăn, chậm lớn. Sử dụng cá sống làm thức ăn sẽ thiếu B1 vì trong thịt cá sống có thiaminase gây vô hoạt thiamin
Riboflavin (B2 4-7 15-20*
Kém ăn, chậm lớn, tỷ lệ chết cao, chảy máu ở da và vây, hiện tượng thần kinh, sợ ánh sáng

68
Pyridoxin 4-5 8-12*
Kém ăn, chậm lớn, rối loạn thần kinh
Pentothenic axit 30-50 40-50*
Kém ăn, chậm lớn, lờ đờ, chậm chạp, thiếu máu, chảy máu da, lồi mắt
Nicotinic axit 28 80-120*
Chảy máu da, tỷ lệ chết cao
Biotin 1-2,5 0,5-1*
Chậm lớn, giảm hoạt động
Folic axit Vitamin B12
NR NR
Inositol 440 100-150
Chậm lớn, chảy máu da và vây, mất niêm mạc da.
Cholin 4000 800-1200
Chậm lớn, gan nhiễm mỡ.
Vitamin C NR 300-500
Chậm lớn, biến dạng cột sống, xuất huyết vây, đầu và da.
NR: Không có nhu cầu (dưới điều kiện thí nghiệm); * Tăng 30 giai đoạn hương và 50% giai đoạn giống
7.7. NHU CẦU VITAMIN CỦA CÁ Những nghiên cứu về nhu cầu vitamin hầu hết thực hiện trên cá hồi và những kết quả
nghiên cứu này được chấp nhận cho những loài cá khác (bảng 7.3).
Bảng 7.3. Nhu cầu vitamin của nhóm cá hồi (mg/kg thức ăn)
Vitamin NRC (1993) Mức thêm vào thức ăn Vitamin A (IU) Vitamin D3 (IU) VitaminE Vitamin K3 Thiamin (B1) Riboflavin (B2) Pyridoxine Pantothenic acid Niacin Biotin Folic acid Vitamin B12 Inositol Cholin Vitamin C
2500 2400 50 R* 1
4-7 3-6 20 10
0,15 1
0,01 300 1000 50
6000 2000
300-500 10 15 25 15 50 180 0,6 8
0,03 130 1000 150**
R* : có nhu cầu nhưng không xác định được số lượng; ** : dùng loại vitamin C bền
7.8. SỬ DỤNG VITAMIN TRONG THỨC ĂN NUÔI CÁ Độ bền của vitamin D3 trong điều kiện bảo quản như vitamin A bằng 75 - 80%. Các
dạng vitamin khác và độ bền của nó trong thức ăn viên (ép đùn) và trong premix ghi ở bảng 6.7. Tuy nhiên, cần lưu ý đến độ bền của vitamin C. Tinh thể axit ascorbic cực kỳ nhạy cảm với sự

69
oxy hoá. Trong 3 ngày dự trữ ở nhiệt độ thường, toàn bộ hoạt tính vitamin mất hết, trong viên hoạt tính vitamin C chỉ còn lại 20% sau khi sử lý nhiệt và dự trữ. Gần đây, người ta sử dụng asorbate-2-monophosphate (sản phẩm này có ascorbate-2-monophosphate và một lượng nhỏ ascorbate-2-polyphosphate, hoạt tính acid ascorbic là 33% và 35 % lần lượt). Dạng vitamin này chỉ mất 15 % hoạt tính trong viên ép đùn và dự trữ 3 tháng ở nhiệt độ trong phòng, trong khi viên vitamin C bọc mỡ hay ethylcellulose mất tới 70-90% hoạt tính trong cùng điều kiện.
Hầu hết, vitamin bổ sung vào thức ăn cá được sản xuất bằng con đường hoá học hoặc vi sinh vật hoặc kết hợp cả hai chứ không phải chiết từ thức ăn tự nhiên, vì các vitamin chiết từ nguồn tự nhiên rất đắt và hiệu quả thấp. Các vitamin tổng hợp được sản xuất ra dưới dạng khác nhau và được bảo vệ để chống lại sự phân huỷ trong quá trình chế biến và dự trữ. Khi sử dụng vitamin để trộn vào thức ăn phải chú ý đến độ bền của vitamin. Các dạng vitamin khác nhau và cách bảo vệ khác nhau thì có độ bền khác nhau.
Ví dụ: Vitamin A dưới dạng vitamin A acetate chứa trong viên nang, trong nang chứa một cái khung bằng gelatin có cấu trúc liên kết chéo, vitamin phân tán khắp trong cái khung này cùng với chất chống oxy hoá và được bọc một lớp vỏ bảo vệ bằng tinh bột ngô. Thường trong viên gelatin người ta thêm cả vitamin D3.
Bảng 7.4. Độ bền của vitamin trong premix và trong viên ép đùn sau 3 tháng dự trữ ở nhiệt độ trong phòng (F.Hoffmann-La Roche, 1988)
Hoạt tính còn sau 3 tháng dự trữ ở nhiệt độ trong phòng (%)
Vitamin
Dạng sử dụng
Trong premix Trong viên ép đùn
Vitamin A Vitamin D Vitamin E Vitamin K Vitamin B1 Vitamin B2 Pyridoxine Pantothenic acid Niacin Biotin Folic acid Vitamin B12 Cholin Inositol Ascorbic acid
Vitamin A acetate Cholecalcalciferol dl-a tocoferol acetate Muối menadione (K3) Thiamin mononitrate Tinh thể Pyridoxine hydrochoride Calcium d-pantothenate Niacinamide và nicotinic acid D-Biotin Tinh thể Dung dịch 1% Muối chloride Ascorbate-2-polyphosphate Tinh thể
70 - 90 80 - 100 90 - 100 65 - 85 70 - 80 90 - 100 80 - 90 80 - 100 90 - 100 80 - 100 50 - 70 50 - 80
không thêm 100 90
30 - 70
70 - 90 75 - 100 90 - 100 40 - 70 60 - 80 90 - 100 80 - 90 80 - 100 90 - 100 70 - 90 50 - 65 40 - 80
100 100 90
10 - 30 Câu hỏi 1. Công thức vitamin A, vai trò và nhu cầu đối với cá? 2. Công thức vitamin D vai trò và nhu cầu đối với cá? 3. Công thức vitamin E vai trò và nhu cầu đối với cá? 4. Vai trò vitamin C đối với cá và những chú ý khi bổ sung vitamin C trong thức ăn cá. Những chú ý khi sử dụng vitamin trong thức ăn tôm và cá?

70
CHƯƠNG VIII DINH DƯỠNG VÀ NHU CẦU CHẤT KHOÁNG
Trong tự nhiên có ít nhất 22 chất khoáng mà cơ thể động vật cần. Khoáng được chia làm hai nhóm căn cứ vào nhu cầu của động vật: nhóm khoáng đa lượng và khoáng vi lượng. Trong chương này chúng tôi đề cập đến vai trò sinh học của một số khoáng đa lượng và vi lượng là những nguyên tố hết sức quan trọng đối với cá
8.1. KHÁI NIỆM CHUNG Mặc dù hầu hết các chất khoáng tìm thấy trong tự nhiên đều có mặt trong các mô của
động vật vì chúng có trong thức ăn nhưng không phải chất khoáng nào cũng có vai trò trong trao đổi chất của cơ thể. Một số chất khoáng với hàm lượng rất thấp có thể còn gây độc cho cơ thể. Ngay cả một số chất khoáng cần thiết đối với động vật thủy sản nhưng được cung cấp với lượng vượt mức nhu cầu cũng gây độc cho cơ thể.
Trong cơ thể người, động vật và cá chất khoáng chiếm tỷ lệ rất thấp so với các chất hữu cơ khác (bảng 8.1; bảng 8.2).
Thuật ngữ khoáng thiết yếu (Essential mineral element) dùng để diễn tả những chất tham gia vào quá trình trao đổi chất của cơ thể. Để nhận biết một chất khoáng là thiết yếu hay không thì khi con vật ăn một phần không có chứa chất khoáng ấy và gây ra những triệu chứng bệnh lý chỉ có thể điều trị hoặc phòng ngừa bằng chính chất đó. Phần lớn các nghiên cứu về dinh dưỡng khoáng đều sử dụng phương pháp trên. Tuy nhiên, đối với những chất khoáng mà cơ thể cần với số lượng rất nhỏ thì không thể kiểm soát được sự có mặt của chúng trong nước uống, máng ăn, chuồng trại, các dụng cụ vệ sinh, bụi trong không khí.
Bảng 8.1. Hàm lượng một số nguyên tố khoáng trong cơ thể động vật
Đa khoáng g/kg thể trọng Vi khoáng mg/kg thể trọng Ca P K Na Clo S Mg
15 10 2
1,6 1,1 1,5 0,4
Fe Zn Cu Mo Se I
Mn Co
20-80 10-50 1-5 1-4 1-2
0,3-0,6 0,2-0,5 0,02-0,1
Đến năm 1950, 13 chất được coi là khoáng thiết yếu, bao gồm Ca, P, K, Na, Cl, S, Mg,
Fe, I, Cu, Mn, Zn và Co. Đến năm 1970, người ta bổ sung thêm Mo, Se, Cr, Fl, As, Bo, Pb, Li, Ni, Si, Sn và Va. Có khoảng trên dưới 40 chất khoáng tham gia vào quá trình trao đổi chất trong cơ thể động vật.
Ngoài ra, chất khoáng thường xếp vào hai nhóm tùy theo nồng độ là khoáng đa lượng và khoáng vi lượng. Thông thường những chất khoáng được gọi là vi lượng khi chúng có mặt trong cơ thể động vật không lớn hơn 50 mg/kg. Một số tài liệu khác cho rằng, khoáng đa lượng là những chất mà động vật có nhu cầu lớn hơn 100mg/kg khẩu phần như Ca, P, Mg, K, Na, Cl, và S; khoáng vi lượng: nhu cầu nhỏ hơn 100mg/kg khẩu phần như Fe, Cu, Mn, Zn, Co, Mo, Cr, Se, F, I, và Ni.

71
Bảng 8.2. Thành phần chất khoáng trong cơ thể cá (Shearer, 1984, Kirchgessmer và Shwarz, 1986)
Các loại khoáng phổ biến Cá hồi (10-1800g)
Cá chép (170-1150g)
Khoáng đa lượng (g/kg WB) Ca P Mg K Na
5,2 4,8 0,33 3,2 1,3
6,1 5,0 0,25 2,1 0,85
Khoáng vi lượng (mg/kg WB) Fe Cu Mn Zn
12 1,2 1,8 25
20 1,1 0,7 63
Về cơ bản người ta chấp nhận rằng nhu cầu khoáng của cá tương ứng với động vật bậc
cao. Tuy nhiên, môi trường xung quanh (nước) là nguồn cung cấp khoáng quan trọng, ngoài thức ăn. Sự trao đổi khoáng của cá thể hiện ở sơ đồ 8.1.
Nước
Máu Mô (cơ thể)
Khẩu Phân, phần Nước tiểu
Sơ đồ 8.1. Trao đổi khoáng ở cá
8.2. CANXI, PHOSPHO, MAGIE
8.2.1. Canxi (Ca) Ca của cơ thể cá phân bố tập trung ở xương, vây. Hàm lượng Ca của cá chép (khối lượng
- 340-3300mg) như sau: Cột sống: 80g/kg (69-96g/kg) Cơ : 124mg/kg (57-410mg/kg) Gan : 38mg/kg (21-155mg/kg) Tỷ lệ Ca/P của xương và vây là 1,5 - 2:1 và của toàn bộ cơ thể là 0,7-1,6. Nồng độ Ca của nước là 200mg/l đáp ứng đủ nhu cầu Ca cho cá hồi. Nếu lượng Ca của
nước thấp (5mg/l) thì cá phải lợi dụng Ca của khẩu phần.

72
Như vậy, sinh trưởng của cá phụ thuộc vào nồng độ Ca và pH của nước. Người ta cũng thấy hàm lượng nhôm trong máu cao làm giảm sự hấp thu Ca. Những hồ nước axit, pH thấp, Ca thấp và Al cao làm giảm tỷ lệ sống của cá rõ rệt. Những hồ nào có hàm lượng Ca nhỏ hơn 0,8mg/l; pH nhỏ 4,5 thường không có cá (Howell et al., 1983).
Vậy, nhu cầu của cá trong khẩu phần là bao nhiêu? Nói chung, khó xác định được nhu cầu Ca của cá. Cá hồi có thể trọng ban đầu là 1,2 g không thấy biểu hiện sinh trưởng khác nhau khi khẩu phần chứa 0,3-3,4g Ca/kg và hàm lượng Ca nước là 20-30mg/l.
Cá da trơn có thể trọng 6-24g sống trong nước có hàm lượng Ca là 56mg/l đáp ứng sinh trưởng tối ưu khi khẩu phần chứa 8g Ca/kg (+8g photpho hữu dụng) nhưng sinh trưởng giảm khi Ca khẩu phần là 20g/kg. Cá hồi châu Âu, cá chép thích hợp với khẩu phần 18-22g/kg Ca. Cá hồi O. aureus thích hợp khẩu phần 8g Ca/kg trong điều kiện nước không có Ca. 8.2.2. Phospho (P)
Cá lấy P từ nước kém hơn Ca. Ví dụ, cá hồi (cá giống) hấp thụ P từ nước chỉ bằng 1/400 so với Ca từ nước. Hấp thu P từ nước cũng phụ thuộc vào nhiệt độ nước và hàm lượng Ca nước. Hấp thu P từ nước tăng khi nhiệt độ tăng và nồng độ Ca nước giảm.
Như vậy, nguồn P khẩu phần đối với cá quan trọng hơn là nguồn P từ nước. Nhu cầu P khẩu phần của cá nằm trong phạm vi 0,4-0,7% khẩu phần phụ thuộc vào:
+ Cấu tạo ống tiêu hoá: loài cá có dạ dày hấp thu P tốt hơn cá không có dạ dày, ngay cả khi nguồn P có độ lợi dụng kém.
+ Nguồn Photpho: Phospho phytic không lợi dụng được vì cá không có enzyme phytase. Giống như động vật trên cạn, photpho monocanxi có độ lợi dụng cao nhất, đi và tri-canxi thì kém hơn, nhất là cá chép (bảng 8.3).
Bảng 8.3. Sinh trưởng của cá hồi và cá chép (g) theo độ lợi dụng của photpho khẩu phần
Cá hồi (11 tuần) Cá chép (4 tuần) Monocanxi photphat Dicanxi photphat Tricanxi photphat
640-710 610 494
270-287 150 112
P trong bột cá, cazein, nấm men đều được rainbow trout lợi dụng tốt, cá chép lợi dụng
tốt P trong nấm men và cazein. 8.2.3. Magiê (Mg)
Mg giữ vai trò quan trọng trong phản ứng photphoryl hoá và một vài enzyme. Mức Mg trong nước ngọt không đáp ứng đủ nhu cầu Mg của cá, phải bổ sung một lượng thích đáng vào khẩu phần. Nước biển chứa 1,3g Mg/lít thì đáp ứng đủ nhu cầu Mg cho cá biển.
Cowey et. al. (1977) làm thí nghiệm với cá hồi nặng khoảng 30g thấy rằng tính ham ăn, tăng trọng và FCR tốt khi khẩu phần chứa 1000mg/kg so với khẩu phần 26-63mg/kg. Thí nghiệm ở cá hồi non thấy 200-300mg/kg thì đủ cho sinh trưởng nếu nước chứa 1,7mg/lít.
Thiếu Mg gắn với mức Ca 26-40g/kg khẩu phần đã làm tăng nephrocalcinosis (nephocalcinosis: Ca lắng đọng ở thận), natri trong cơ cũng tăng, làm cho thịt nát..vì cơ thịt chứa nhiều nước.
Ơ cá chép thiếu Mg làm giảm thu nhận thức ăn, nghèo sinh trưởng và inertia. Trong một thí nghiệm người ta thấy mức Mg là 52mg/kg đã làm tăng tỷ lệ tử vong từ đó người ta thấy nhu cầu tối thiểu Mg phải là là 400-700mg/kg khẩu phần.

73
8.3. CÁC NGUYÊN TỐ KHOÁNG KHÁC Bảng 8.4 sau đây tóm tắt chức năng của các nguyên tố vi khoáng.
Bảng 8.4. Tóm tắt vai trò dinh dưỡng của một số nguyên tố vi lượng
Nguyên tố Chức năng Biểu hiện thiếu và nhu cầu Fe Cấu tạo Hb, myoglobin, cytocrome và
nhiều enzyme khác Chậm lớn, thấp Hb và hematocrit. 200mg/kg thức ăn của cá chép, 30mg/kg thức ăn của cá da trơn.
Cu Tham gia vào các enzym có đồng như cytocrome oxidase, feroxydase, tyrosinase, superoxide dismutase
Chậm lớn, viêm cata, tim yếu. 3mg/kg thức ăn của cá chép. 5mg/kg thức ăn của cá da trơn
Mn Coenzym của một số enzyme tổng hợp ure, trao đổi axit amin, axit béo và oxy hoá glucose.
Chậm lớn, cột sống ngắn. Viêm cata, tỷ lệ tử vong cao, 2,4mg/kg thức ăn của cá da trơn , 13mg/kg thức ăn của cá chép
Zn Cofactor của một số enzyme và thành phần của nhiều metaloenzyme như cacbonic anhydrase, carboxypeptidase, malic dehydrogenase, alkali photphatase, superoxid dismutase, ribonuclease và DNApolymerase.
Chậm lớn, kém ăn, viêm cata, thối vây và da, tử vong cao. 20mg/kg thức ăn của cá da trơn. 15-30mg/kg thức ăn của cá chép
Co Thành phần của vitamin B12 Chậm lớn, số lượng hồng cầu giảm, mứckhuyến cáo 1 - 6mg/kg thức ăn.
Se Thành phần của glutathion peroxidase, phân giải peroxid sinh ra trong quá trình oxy hoá mỡ
Chậm lớn, thiếu máu, viêm cata. Mức khuyến cáo 0,25mg/kg thức ăn của cá da trơn
I Thành phần hormon thyroxin 0,6-2,8mg/kg thức ăn của cá hồi
Giới thiệu hỗn hợp khoáng dùng cho cá (premix):
2,1% CaCO3 0,034% Cu (OH)2.2CuCO3 73,5% CaHPO4.2H2O 0,081% ZnCO3 8,1% K2HPO4 0,001% KI 6,8% K2SO4 0,002% NaF 3,1% NaCl 0,020% CoCl2 2,5% MgO 0,0686% axit citric 0,558% MnCO3
Câu hỏi
1. Sơ đồ chuyển hoá khoáng của cá? 2. Vai trò của Ca, P, Mg, và nhu cầu của cá. Tại sao khả năng lợi dụng P dưới dạng axit
phytic ở cá rất thấp, biện pháp khăc phục? 3. Cho ví dụ về một công thức khoáng hỗn hợp của cá và cho ý kiến nhận xét về công
thức khoáng này?

74
CHƯƠNG IX ĐẶC ĐIỂM THỨC ĂN TRONG NUÔI TRỒNG THUỶ SẢN
9.1. PHÂN LOẠI THỨC ĂN Người ta xếp thức ăn cho động vật thuỷ sản thành 5 nhóm dựa vào thành phần dinh
dưỡng và cách sử dụng: + Thức ăn thô xanh: bao gồm thức ăn xanh như rau cỏ xanh, thức ăn thô khô như cỏ khô,
rơm, thân cây ngô … Tỷ lệ xơ trong thức ăn thường lớn hơn 18%. + Thức ăn giàu năng lượng: nhóm thức ăn có hàm lượng protein nhỏ hơn 20% và xơ nhỏ
hơn 18%. + Thức ăn giàu protein: nhóm thức ăn có hàm lượng protein lớn hơn hoặc bằng 20%, đó
là protein nguồn gốc động vật như bột thịt, bột cá, bột lông vũ thuỷ phân... và protein nguồn gốc thực vật như khô dầu đỗ tương, khô dầu bông, gluten ngô …
+ Thức ăn giàu khoáng như bột đá, monocanxiphosphat, dicanxiphosphat… + Thức ăn bổ sung: gồm thức ăn bổ sung dinh dưỡng như vitamin, chất khoáng, axit
amin, và thức ăn bổ sung phi dinh dưỡng (feed additives) như chất chống oxy hoá, sắc chất, các thuốc phòng bệnh...
9.2. ĐẶC ĐIỂM DINH DƯỠNG CỦA CÁC LOẠI THỨC ĂN
9.2.1. Thức ăn giàu năng lượng Các loại hạt ngũ cốc và phụ phẩm của ngũ cốc, các loại bột củ và phụ phẩm nông nghiệp
như cám gạo, cám mì… nằm trong nhóm thức ăn này. Thành phần hoá học: tinh bột chiếm 2/3 khối lượng hạt, protein 9 – 12%, mỡ 2 – 4%, xơ trung bình 6%, nhưng khác nhau nhiều giữa các loại hạt và phụ phẩm của hạt.
Bột sắn là nguồn thức ăn giầu tinh bột (60-70% trong chất khô), nhưng protein lại rất thấp (0,9-2,1%).
Đặc điểm chung của nhóm thức ăn giàu năng lượng là tỷ lệ protein thấp, acid amin thiếu và không cân đối; đa số có hàm lượng Lipid thấp (trừ cám gạo có 10-15% lipid); tỷ lệ xơ cao; hàm lượng khoáng thấp và không thích hợp cho động vật thủy sản.
Tùy vào đối tượng thủy sản nuôi cụ thể mà ta có thể sử dụng các nguồn thức ăn này khác nhau. Đối với nhóm đối tượng ăn thiên về động vật, lượng tinh bột không nên sử dụng quá 20%. Bột sắn là nguồn thức ăn cung cấp năng lượng tốt nhất. Cám gạo có hàm lượng xơ từ 11- 20% (tùy loại cám) do đó ít được sử dụng làm thức ăn cho tôm. 9.2.2. Thức ăn giàu protein
Có hai nhóm là thức ăn protein có nguồn gốc thực vật và nguồn gốc động vật. Nhóm thức ăn protein nguồn gốc thực vật có hai nhóm nhỏ: - Nhóm thức ăn có 20 – 30% protein thô, trong nhóm này có bã rượu, bã bia, bã mạch
nha.. hàm lượng protein 25 – 27%, chất lượng protein thấp (thiếu lysine), xơ tương đối cao (12 – 15%).
- Nhóm thức ăn chứa 30 – 45% protein, trong nhóm này có các loại khô dầu lạc, khô dầu bông, khô dầu lang, khô đỗ tương, khô hướng hướng dương, khô cải dầu, … Hàm lượng protein của các loại khô dầu này từ 42 – 46%. Chất lượng protein cao hơn nhóm thức ăn trên. Tuy nhiên, khô dầu lanh và khô dầu bông nghèo lysine, khô dầu lạc nghèo axit amin chứa S, chỉ có khô đỗ tương là có chất lượng khá hoàn toàn. Tỷ lệ xơ của nhóm thức ăn này thấp hơn nhóm

75
trên (9 – 11%), riêng khô đỗ tương rất thấp (5%). Chất béo của nhóm khô dầu phụ thuộc vào cách lấy dầu, nếu lấy dầu bằng cách ép thì chất béo của sản phẩm còn 6- 8%, còn chiết dầu bằng dung môi hữu cơ thì chất béo chỉ còn 1 – 2%.
Thành phần dinh dưỡng của một số loại thức ăn protein thực vật trình bày ở bảng 9.1.
Bảng 9.1. Thành phần dinh dưỡng của một số loại thức ăn giàu protein có nguồn gốc thực vật
Thành phần Khô đậu tương
Khô dầu bông
Khô dầu dừa
Khô dầu lạc
Vật chất khô % 88 91 90 89 Protein % 45-48 41 21.5 45-48 Lipid % 1.9 1.4 1.6 1.1 Dẫn xuất không đạm % 28.5 29.1 43.9 - Khoáng % 6.2 6.5 7.0 4.5 Năng lượng thô (MJ/kg) 17.5 17.9 16.1 - Năng lượng tiêu hóa (MJ/kg) 13.5 9.1 - -
Đặc điểm dinh dưỡng của một số loại khô dầu Khô đậu tương. 42 – 48% protein, giàu lysine (2,45%) nhưng hạn chế methionine và
cystine, Ca, P, và vitamin B. Khẩu phần nuôi cá O. niloticus có thể thay hoàn toàn bột cá bằng khô đỗ tương nếu bổ sung thêm 0,25% methionine.
Khô dầu bông: 40 – 45% protein, nghèo methionine, cystine, lysine, Ca, P, giầu vitamin B1, chứa 0,003 – 0,2% gossypol là chất độc gây ức chế enzym tiêu hoá và làm giảm độ ngon của khô dầu bông.
Khô dầu lạc. 45 – 50% protein, nghèo lysine, methionine, cystine, dự trữ trong điều kiện nóng ẩm sẽ sản sinh mycotoxin, đặc biệt là aflatoxin. Cá rất nhạy cảm với độc tố aflatoxin, liều gây độc ở cá là 1 ppb trong khi ở gà con là > 50 ppb, lợn con ≥ 50 ppb, bê > 200 ppb.
Khô dầu cải dầu. Protein giống khô dầu đỗ tương, chứa glucozit làm giảm sinh trưởng của cá chép. Glucoside bền đối với nhiệt.
Khô dầu hướng dương: 35 – 40% protein, không thấy có chất độc, xơ cao ( 16%). Khô dầu vừng. 40% protein, giàu methionine, arginine và leucine, nhưng thiếu lysine.
Chứa axit phytic dễ kết hợp với chất khác như axit amin, vi khoáng.. tạo thành phytat không hoà tan, không hấp thu được, làm mất cân đối axit amin và vi khoáng khẩu phần.
Nhóm thức ăn giàu protein nguồn gốc động vật gồm có bột thịt, bột thịt xương, bột máu, bột cá, phụ phẩm sữa,… Thành phần dinh dưỡng của một số loại thức ăn này ghi ở bảng 9.2.
Đặc điểm một số thức ăn giàu protein nguồn động vật Bột cá: Bột cá là nguồn cung cấp protein tốt nhất cho các đối tượng nuôi thủy sản, hàm
lượng protein thô biến động từ 45% đến 70% phụ thuộc vào nguồn cá, cách chế biến. Bột cá chứa đầy đủ các acid amin cần thiết cho động vật thủy sản, giàu lysine (7,8% so với CP), methionine (3,5% so với CP), methionine+cystine (4,7% so với CP), tryptophan (1,3% so với CP), threonine (4,9% so với CP). Đặc biệt trong thành phần lipid của bột cá có nhiều acid béo cao phân tử không no (HUFA). Trong bột cá có hàm lượng vitamin A và D cao và thích hợp cho việc bổ sung vitamin A trong thức ăn. Bột cá làm cho thức ăn trở nên có mùi hấp dẫn và tăng tính ngon miệng của thức ăn. Hàm lượng khoáng trong bột cá luôn lớn hơn 16% và là nguồn khoáng được động vật thủy sản sử dụng hiệu quả. Năng lượng thô của bột cá từ 4100 - 4200 kcal/kg. Ngoài ra, một số nghiên cứu cho thấy trong bột cá có chứa chất kích thích sinh trưởng,

76
đây là nguyên nhân chính khi thay thế bột cá bằng các nguồn protein động vật khác kết quả không hoàn toàn đạt được như sử dụng bột cá.
Bột cá được chia làm hai loại: bột cá nhạt và bột cá mặn. Trong chế biến thức ăn cho động vật thủy sản chỉ sử dụng bột cá nhạt (độ muối <5%). Bột cá thường được làm từ cá trích, cá mòi và cá cơm. Chất lượng bột cá phụ thuộc vào loài, độ tươi của nguyên liệu tươi, phương thức chế biến và bảo quản. Có hai phương pháp chế biến bột cá:
- Chế biến ẩm: Cá tươi đem hầm hơi sau đó ép để tách nước và dầu, bã được sấy khô nghiền thành bột, làm bền bột bằng các chất chống oxy hoá. Dịch ép được chiết mỡ, cô đặc rồi cho thêm vào bột để bổ sung vitamin và protein hoà tan.
- Chế biến khô: Cá sau khi nấu chín (hoặc hầm chín) rồi làm khô không tách mỡ. Bột cá cũng có thể chế biến theo phương pháp chiết bằng dung môi. Bột cá chế biến theo phương pháp này rất giầu protein (80%) và ít mỡ.
Bảng 9.2. Thành phần dinh dưỡng của nhóm thức ăn protein nguồn gốc động vật
Khoáng (%) Thức ăn
Protein%
Béo% Na P
8,0 10,0 0,3 - - -
1,2 0,9
4,03 5,07 0,22
- - -
1,0 0,8
Bột thịt Bột thịt xương Bột máu Bột cá 50% protein 70% protein 65% protein Sữa : Bột sữa khử mỡ Váng sữa ( whey) Bột tôm Phụ phẩm gia cầm Bột lông vũ thuỷ phân
53 51 80 53 74 68 34 14
49-74 60-65
80-85
10 10 2 4 1 1 1 1 -
15-20 -
Nghèo lysine hơn bột cá da trơn, có thể dùng 15% bột lông vũ thuỷ phân
Bột tôm. Bột tôm là phụ phẩm của nhà máy chế biến tôm đông lạnh bao gồm đầu tôm và
vỏ tôm. Hàm lượng protein 30-40%, chứa đầy đủ các axit amin thiết yếu, tuy nhiên, methionine axit amin là giới hạn một. Bột đầu tôm có thể thay 20-30% bột cá và không nên bổ sung quá 15% trong khẩu phần ăn của tôm. Bột đầu tôm là nguồn cung cấp axit béo n3, cholesterol, asthaxanthin, khoáng và chứa cả chất dẫn dụ. Ngoài ra, bột đầu tôm giàu chitin (15-23%) là chất cần thiết cho quá trình hình thành vỏ của tôm. Mục đích bổ sung bột đầu tôm vào thức ăn nhằm cải thiện mùi vị hấp dẫn của thức ăn. Chất lượng của bột đầu tôm rất biến động phụ thuộc vào nguyên liệu, phương thức chế biến và bảo quản.
Cá ủ ướp (fish silage). Đây là cách bảo quản bằng phương pháp lên men sinh học trong môi trường axit. Cá hay phụ phẩm cá được chặt nhỏ, trộn với bột sắn và rỉ mật theo tỷ lệ 5 kg cá + 3 kg bột sắn + 2 kg rỉ mật. Có thể thay bột sắn bằng bột gạo, bột ngô, bột các loại khoai củ hoặc chỉ cần ủ với rỉ mật. Có điều kiện thêm axit hữu cơ như axit formic hay chế phẩm lên men lactobacillus.
Cá đã băm nhỏ đặt vào các thùng nhựa ủ kín, thỉnh thoảng khuấy đều. Trong quá trình ủ, axit lactic được hình thành, pH xuống dưới 4,5 nhờ vậy, sản phẩm được bảo quản vài tháng không hư hỏng.

77
Cá ủ ướp có hàm lượng protein từ 30 – 50% so với vật chất khô, tuỳ theo nguyên liệu đem ủ và có nhiều axit amin quý như lysine, methionine.
Bột lông vũ thuỷ phân. Hàm lượng protein cao tới 80-85% nhưng không cân đối axit amin (nghèo lysine, histidine và tryptophan). Thành phần protein chủ yếu là keratin nên tỷ lệ tiêu hóa rất thấp (khoảng 50%). Bột lông vũ thủy phân có thể thay 30% bột cá trong khẩu phần cá rô phi và cá trê.
Bột thịt. Bột thịt bao gồm phế phụ phẩm lò mổ gồm tất cả những phần không dùng làm thức ăn cho người như: ruột già, gân, móng, thức ăn trong dạ dày, móng và lông. Hàm lượng dinh dưỡng biến đổi nhiều tuỳ theo nguồn phụ phẩm, protein từ 30-60%, lipit 8-11%, Ca 4,5-5% và P 8-10%. Nhìn chung giá trị protein của cả hai loại bột này đều không cao, hàm lượng methionin thấp nên hiệu quả sử dụng không cao khi làm thức ăn cho động vật thuỷ sản. Hàm lượng bột thịt xương được đề nghị sử dụng trong thức ăn cho tôm không quá 15%.
Bột máu. Bột máu có hàm lượng protein cao tới 85-90%, rất giàu lysine (9-11%), tuy nhiên thiếu Isoleusine và Methionin. Khả năng tiêu hóa bột máu của động vật thuỷ sản thấp, tỷ lệ tiêu hoá protein biến động từ 40-80% tuỳ theo phương pháp sấy. Protein và acid amin trong bột máu dễ bị phân hủy trong quá trình sấy. Bột máu rất dễ bị hư trong quá trình tồn trữ. Hàm lượng bột máu phối trộn trong khẩu phần ăn cho tôm không nên quá 10%. 9.2.3. Thức ăn bổ sung (feed additives)
Có hai nhóm thức ăn bổ sung, đó là thức ăn bổ sung dinh dưỡng và thức ăn bổ sung phi dinh dưỡng:
Thức ăn bổ sung dinh dưỡng bao gồm thức ăn bổ sung vitamin, vi khoáng, axit amin tổng hợp... Thông thường, thức ăn bổ sung loại này được sản xuất dưới dạng premix.
Premix là một hỗn hợp chứa hoạt chất và chất mang. Hoạt chất có thể là axit amin, vitamin, nguyên tố vi lượng, còn chất mang thì tuỳ loại hoạt chất mà có thể khác nhau. Ví dụ, đối với hoạt chất là vitamin, axit amin.. thì hoạt chất là trấu hay cám mì nghiền mịn, đối với hoạt chất là vi khoáng thì chất mang là bột đá. Để cho các hoạt chất trộn đều vào chất mang thì khối lượng riêng và kích thước của hoạt chất- chất mang phải tương đương nhau, thời gian bảo quản tương đồng và không phá hoại nhau.
Thức ăn bổ sung phi dinh dưỡng bao gồm chất chống oxy hoá, chất tạo mầu, tạo mùi, chất hấp phụ mycotoxin, enzyme, chất dẫn dụ, chất kết dính...
- Chất chống oxy hoá: trong công nghiệp thức ăn thuỷ sản, người ta thường sử dụng các chất chống oxy hoá là Ethoxyquin (1,2 dihydro-6 cthoxy-2,2,4 trimethyl quinoline) với liều 150ppm, BHT (Butylated hydroxy toluene) với liều 200ppm, BHT (Butylaled hydroxy anisole) với liều 200ppm.
- Chất chống nấm: có các loại như potassium sorbate, polypropilene glycol, aluminiumsilicat, Mycofix-plus.. ngăn ngừa sự phát triển của một số nấm mốc sản sinh mycotoxin.
- Sắc chất: cá không thể sinh tổng hợp được các sắc tố nên phải được cung cấp từ thức ăn. Sắc tố có nhiều trong thức ăn tự nhiên như rong, tảo, rau cỏ xanh, các sắc tố tổng hợp thường chiết rút từ các nguồn thực vật trên.
Hai loại sắc tố sử dụng trong thức ăn của cá hồi (làm đỏ da, cơ và vây) là asthaxanthin và cathaxanthin. Asthaxanthin cũng dùng để tạo màu cho tôm hùm. Các sắc tố trên cũng dùng cho cá cảnh.
Đối với cá tra hay basa người tiêu dùng ưa thích thịt cá trắng không vàng, như vậy, đối với loaị cá này người ta không dùng sắc tố hay những loại thức ăn tự nhiên chứa sắc tố như ngô vàng hay rau xanh.

78
- Chất dẫn dụ: Các axit amin tự do như glycine, alanine, glutamate; một số peptid như betain có tác dụng hấp dẫn tôm, cá. Các chất này hiện diện trong bột tôm, bột cá, bột mực, bột các loài nhuyễn thể hay trong các sản phẩm thuỷ phân của những thức ăn này, các loại này có thể trộn vào thức ăn ở mức 1-5%.
- Chất kết dính: Chất kết dính làm cho thức ăn bền trong nước. Chất kết dính thường dùng là tinh bột ngũ cốc, tinh bột sắn có thể dùng 5% trong hỗn hợp thức ăn viên. Các chất kết dính khác được dùng nhiều trong công nghiệp thức ăn thuỷ sản là alginate, gelatin, chitosan, bentonite, CMC (carboxyl methyl cellulose). 9.2.4. Thức ăn tự nhiên
Thức ăn tự nhiên đóng vai trò rất quan trọng, quyết định sự thành công trong ương nuôi nhiều loài động vật thủy sản, đặc biệt là ở giai đoạn ấu trùng. Nghiên cứu đặc điểm sinh học, kỹ thuật nuôi một số loại thức ăn tươi sống cho động vật thủy sản đã được nhiều nhà nghiên cứu quan tâm. Các đối tượng chủ yếu đang được quan tâm nghiên cứu như: vi tảo, luân trùng, giáp xác râu ngành, Artemia, trùng chỉ.. vì chúng có giá trị dinh dưỡng cao (bảng 9.3)
Bảng 9.3. Thành phần dinh dưỡng của một số zooplankton nước biển và nước ngọt (Hepher, 1988)
Thành phần dinh dưỡng (% vật chất khô) Zooplankton VCK CP EE Tro GE kcal/kg
Rotifier Artemia Cladocera Malacostraca Ostracoda Copepoda
11,2 11,0 9,8 24,6 35,0 10,3
64,3 61,3 56,5 49,9 41,5 52,3
20,3 19,5 19,3 20,3
- 7,1
6,2 10,1 7,7 19,6
- 1,7
4866 5835 4800 5537 5683 5445
Nghề nuôi giáp xác, cá biển và nhuyễn thể đang ngày càng phát triển mạnh, vì thế, nhu cầu con giống ngày càng gia tăng. Trong sản xuất giống ấu trùng, thức ăn và kỹ thuật cho ăn là vấn đề rất quan trọng. Ngày nay, mặc dù có nhiều kỹ thuật tiên tiến trong sản xuất thức ăn nhân tạo cho ấu trùng, nhưng những thức ăn tươi sống như tảo, luân trùng, giáp xác râu ngành, Artemia.. vẫn được xem là thức ăn vô cùng quan trọng và có tiềm năng rất lớn trong sản xuất giống. Việc nuôi và sử dụng các sinh vật làm thức ăn này đã có một lịch sử lâu đời ở nhiều nước và ngày nay đang được áp dụng rộng rãi tên toàn thế giới.
Chi tiết về thức ăn tự nhiên được trình bày ở chương tiếp theo. 9.2.5. Mùn bã hữu cơ và sinh khối vi khuẩn
Mùn bã hữu cơ là sản phẩm phân giải chất hữu cơ có nguồn gốc từ động vật hoặc thực vật. Mùn bã hữu cơ có giá trị dinh dưỡng thấp nhưng gắn trên mùn bã hữu cơ là những tập đoàn nấm, vi khuẩn và các protozoa làm nhiệm vụ phân huỷ chất hữu cơ. Lượng protein trong mùn bã hữu cơ chỉ khoảng 10%, nhưng nếu kể chung với tập đoàn sinh vật công sinh thì giá trị dinh dưỡng của mùn bã hữu cơ tăng lên nhiều lần, đặc biệt sinh khối vi khuẩn là nguồn cung cấp axit amin thiết yếu, nhất là nhóm axit amin chứa lưu huỳnh. 9.2.6. Probiotic trong nuôi trồng thuỷ sản
Do yêu cầu về vệ sinh an toàn thực phẩm ngày càng cao, kháng sinh và các hoá chất dùng làm thức ăn bổ sung (feed additives) trong thức ăn cho động vật nuôi trên cạn và động vật nuôi dưới nước được kiểm soát ngày càng chặt chẽ (xem phụ lục: Kháng sinh cấm sử dụng).

79
Khoảng 20 năm trước đây, kháng sinh dùng với một lượng rất lớn, hàng năm lượng kháng sinh sản xuất ra lên tới 27 ngàn tấn, 90% số lượng này bổ sung vào thức ăn chăn nuôi (International Poultry Production, 1998). Kháng sinh đã tạo ra những dòng vi kháng sinh, loài người đã tốn hàng tỷ đô la để nghiên cứu và tìm ra những loại kháng sinh mới thay thế kháng sinh cũ đã mất tác dụng diệt khuẩn.
Ngày nay, kháng sinh đã bị hạn chế sử dụng làm chất kích thích sinh trưởng và phòng bệnh trong chăn nuôi và nuôi trồng thuỷ sản. Bắt đầu từ năm 2006, Cộng đồng Châu Âu đã cấm sử dụng tất cả các loại kháng sinh làm thức ăn bổ sung trong chăn nuôi và nuôi trồng thuỷ sản.
Nhiều biện pháp đã được áp dụng để thay thế kháng sinh, trong số các biện pháp này probiotic đã có tác dụng lớn và có nhiều triển vọng. Na Uy giảm lượng kháng sinh trong NTTS từ 50 tấn vào năm 1987 xuống còn 746,5kg năm 1997, nhưng sản lượng cá vẫn tăng từ 50 ngàn tấn lên 350 ngàn tấn.
Sử dụng probiotic trong NTTS còn rất mới. Yasuda và Taga, lần đầu tiên sử dụng vi khuẩn như một nguồn thực phẩm và như một tác nhân sinh học để khống chế bệnh của cá vào năm 1980. Vi khuẩn Vibrio alginolyticus được dùng như một probiotic cho tôm ở Ecuador vào năm 1992, nhờ vậy sản lượng tôm đã tăng 35% trong khi toàn bộ lượng dùng kháng sinh giảm 94% trong thời gian từ 1991 đến 1995. Ngày nay, việc sử dụng probiotic trong nuôi tôm ở Mexico cũng trở phổ biến. FAO cũng coi việc nghiên cứu probiotic cùng với các chất kích thích miễn dịch (immunostimulant), các chất nâng cao khả năng miễn dịch (immune enhancers) như một trong các biện pháp chủ yếu để cải thiện chất lượng môi trường nuôi thuỷ sản.
Probiotic là gì? Theo Laurent Verschuere và ctv (2000), probiotic được định nghĩa như sau: “Probiotic là những vi khuẩn sống có ảnh hưởng tốt cho vật chủ nhờ vào sự biến đổi hệ vi khuẩn gắn với vật chủ hay xung quanh con vật chủ, từ đó cải thiện khả năng sử dụng thức ăn, nâng cao khả năng chống bệnh của vật chủ và cải thiện chất lượng môi trường xung quanh”
Dựa trên định nghĩa này chúng ta thấy rằng, probiotic có thể bao gồm những vi khuẩn ngăn ngừa bệnh sinh (pathogen) phát triển trong ống tiêu hoá, trên cấu trúc bề mặt động vật chủ và trong môi trường nuôi. Những vi khuẩn cung cấp chất dinh dưỡng không hỗ trợ cho con vật chủ hay không có mối quan hệ tương tác với vi khuẩn khác, với môi trường sống của con vật chủ thì không bao gồm trong định nghĩa này.
Probiotic còn có các tên khác là: Probiont, Probiotic Bacteria, Beneficial Bacteria. Hầu hết probiotic được dùng như một tác nhân kiểm soát sinh học trong nuôi trồng thuỷ sản thuộc về vi khuẩn lactic (Lactobacillus, Carnobacterium...), Vibrio (Vibrio alginolyticus...), Bacillus và Pseudomonas.
Kiểu tác động của probiotic. Có nhiều nghiên cứu về cơ chế tác động của probiotic, tuy nhiên cũng chưa có những lý giải được chấp nhận hoàn toàn. Sau đây là tóm tắt những kiểu tác của probiotic (Verschuere và ctv., 2000 và Jenn-Kan Lu, 2003):
Probiotic sản sinh các chất ức chế Vi khuẩn probiotic có thể sản sinh một số hoá chất có tác dụng diệt khuẩn hay ức chế
hoạt động của vi khuẩn bệnh dưới dạng đơn hay hỗn hợp như kháng sinh, bactericins, siderophores, lysozymes, proteases, hydrogen peroxid, axit hữu cơ hay diacetyl, nhờ đó ngăn chặn bệnh và hạn chế sự cạnh tranh các chất dinh dưỡng và năng lượng của những vi khuẩn có hại.
Tác động này có thể thực hiện trên bề mặt (trên da) và trong ruột động vật chủ hay trong môi trường nuôi cấy vi khuẩn.
Các nhà vi sinh học đã đưa ra các bằng chứng sau: Lactobacillus sp. có thể sản sinh bactericins là chất ưc chế trưởng của vi khác (chủ yếu là vi khuẩn gram (+). Các vi khuẩn biển

80
sản sinh enzyme phân giải vi khuẩn chống lại Vibrio parahaemolyticus (V. parahaemolyticus là vi khuẩn hiếu khí gram (-), thường nhiễm trong hải sản và gây độc cấp tính cho người).
Alteromonas sp.dòng B-10-31, phân lập từ nước biển gần bờ của Nhật sản sinh monastatin có tác dụng ức chế hoạt tính protease của Aermonas hydrophila và V. anguillarum (hai vi khuẩn này thuộc vi khuẩn gram (-) thường nhiễm trong nước, trong đất và thực phẩm).
Tranh dành dinh dưỡng và năng lượng với những vi khuẩn khác Tất cả các vi khuẩn đều yêu cầu Fe để tăng trưởng. Siderophores là chelat sắt ba khối
lượng phân tử thấp (<1500) do vi khuẩn probiotic sinh ra và có thể hoà tan sắt kết tủa để cho vi khuẩn sử dụng, các khuẩn hại không có khả năng này cho nên thiếu sắt cho tăng trưởng của chúng.
Bản thân vi khuẩn có hại bị loại bỏ thì cũng có nghĩa là loại bỏ được đối thủ cạnh tranh các chất dinh dưỡng và năng lượng dùng cho vi khuẩn probiotic và cho con vật chủ. Cơ chế này xãy ra ở mô và được thực nghiệm với V. anguillarum (dòng VL4335).
Tranh giành vị trí bám dính với vi khuẩn có hại Vi khuẩn probiotic có thể ngăn trở sự khu trú của bệnh sinh theo cơ chế tranh giành trí
bám dính trên vách ruột hay trên bề mặt các mô khác. Vi khuẩn bám dính trên niêm mạc ruột nhờ cơ chế đặc trưng (dựa vào chất bám dính của vi khuẩn và các phân tử receptor của thượng bì ruột) và cơ chế không đặc trưng (dựa vào những yếu tố hoá vật lý học).
Năng lực cạnh tranh bám dính và sinh trưởng trong ruột hay niêm dịch đã được chứng minh in vitro trên vi khuẩn probiotic Carnobacterium dòng K1 đối với V. anguillarum và A. hydrophila.
Nâng cao đáp ứng miễn dịch của ruột Kháng nguyên của probiotic kích thích tế bào niêm ruột sản sinh kháng thể chống lại
bệnh. Chất ức chế của probiotic tiếp tục nâng cao hiệu quả của kháng thể của con vật chủ. Trong thực nghiệm người ta thấy động vật thí nghiệm được bổ sung vi khuẩn lactic đã tăng khả năng chống lại bệnh truyền nhiễm đường ruột khá rõ rệt.
Cải thiện chất lượng nước Xử lý nước nuôi thuỷ sản bằng probiotic đã thấy giảm được chất hữu cơ trong nước của
ao nuôi, giảm đuợc BOD (Biochemical Oxygen Demand: nhu cầu oxy sinh hoá) và giảm độc amonia, nitrit và hydrogen sulfide, khống chế được vi khuẩn bệnh.
Ngoài các kiểu tác động trên người ta cũng thấy rằng vi khuẩn probiotic còn góp thêm enzym cho con vật chủ cũng như cung cấp các nguyên tố đa vi lượng cho con vật chủ, giúp con vật chủ sử dụng thức ăn hiệu quả hơn và tăng trưởng tốt hơn.
Tuy nhiên, cũng cần lưu ý rằng trong mối tương tác với vi tảo thì vi khuẩn probiotic lại ức chế sự phát triển của vi tảo, đặc biệt tảo đỏ. Trong số 41 dòng vi khuẩn thử nghiệm có tới 23 dòng ức chế sinh trưởng của tảo đơn bào Pavlova lutheri ở những mức độ khác nhau. Vi khuẩn probiotic hạn chế sự phát triển của tảo là điều không mong muốn trong nuôi ấu trùng bằng kỹ thuật nước xanh (green-water technique), tức là kỹ thuật bổ sung thêm tảo đơn bào cho ấu trùng.
Ứng dụng probiotics trong nuôi trồng thuỷ sản. Có 3 cách sử dụng probiotic trong nuôi trồng thuỷ sản: Đưa trực tiếp vào nước để vi khuẩn probiotic khu trú trước trong nước, trộn probiotics vào thức ăn, ương nuôi thức ăn tự nhiên (rotifier, artemia) với probiotic.
Kết quả thực nghiệm trên cá Cá hồi Đại Tây dương (Atlantic salmon) và cá hồi nước ngọt (O. mykiss) nuôi bằng thức
bổ sung vi khuẩn probiotic là Camobacterium, vi khuẩn bệnh được dùng trong thực nghiệm là Aeromonas salmonicida. Số lượng A.salmonicida trong phân từ 105 CFU/g giảm còn 102 CFU/g phân sau khi dùng probiotic trong 3 ngày.

81
Vi khuẩn Pseudomonas fluorescens cũng có tác dụng hạn chế A. salmonicida do cơ chế tranh giành sắt. Vi khuẩn P. fluorescens tác động bên ngoài cơ thể cá.
Kết quả thực nghiệm trên các loài giáp xác (crustaceans) Kết quả thực nghiệm trên tôm. Artemia ương nuôi bằng môi trường bổ sung Bacillus sp.
đã cải thiện tỷ lệ nuôi sống của tôm Penaeus monodon khi cho nhiễm Vibrio harveyi. Sau 10 ngày nuôi nhóm tôm thí nghiệm có tỷ lệ nuôi sống là 100% còn nhóm đối chứng là 26%.
Vi khuẩn proboiotic là V. alginolyticus hàng ngày đưa vào bể nuôi ấu trùng tôm (bể 25-60 tấn) đã làm tỷ lệ nuôi sống và khối luợng của tôm cao hơn tôm đối chứng hay tôm dùng oxytetracycline.
Vi khuẩn có hại V. paraheamolyticus không phát hiện thấy trên tôm dùng probiotic, trong khi đó phát hiện thấy 10% mẫu tôm có vi khuẩn gây bệnh này trên tôm nhận kháng sinh hay trên tôm đối chứng.
Ở Indonesia người ta dùng Bacillus cho ao nuôi tôm đã giúp tôm phát triển 160 ngày không có vấn đề gì, trong khi đó tất cả những ao nuôi không dùng Bacillus đều thất bại do Vibrio đã giết chết tôm trước 80 ngày nuôi.
Kết quả thực nghiệm trên cua. Diatom và rotifier ủ với dòng vi khuẩn PM-4 rồi đưa vào bể nuôi ấu trùng cua (Portunus trituberculatus) trong 7 ngày, dung tích bể 200m3, bể cũng đã nuôi diatom và rotifier. Có mối tương quan âm giữa sự có mặt của PM-4 với mật độ của Vibrio spp. Trong 7 thí nghiệm, tỷ lệ sống của ấu trùng cua bổ sung PM-4 là 27,2%; trong 6/9 thí nghiệm không có PM-4, ấu trùng không phát triển, tỷ lệ sống chỉ đạt 6,8%.
Probiotics dùng trong xử lý nước. Hợp chất hữu cơ trong nước, dưới tác dụng của hầu hết vi khuẩn, bị phân giải thành amoniac, amoniac tham gia vào quá trình nitrit hoá biến thành nitrit (NO2
-) rồi thành nitrat (NO3-) nhờ nhóm vi khuẩn tự dưỡng mà chủ yếu là Nitrosomonas
và Nitrobacter theo phản ứng tóm tắt như sau: Nitrosomonas 2NH3-N + 3O2 2NO2
--N +2H2 + 2H- Nitrobacter 2NO2
- -N + O2 2NO3--N
Nitrate lại được thực vật và tảo biến thành nitrit rồi được chloroplast chuyển thành NH4
+ để sinh tổng hợp protein. Protein lại bị vi khuẩn phân giải thành NH3. Quá trình phân giải hình thành NH3 nhanh hơn quá trình lấy NH3 để tổng hợp protein, nồng độ NH3 trong nước sẽ cao và có hại cho sự sống của động vật nước.
Thông thường, người ta dùng chất hấp phụ (như zeolite hay De-Odorase chiết từ cây Yucca) để giảm lượng NH3 trong môi trường nuôi. Vi khuẩn probiotic (chủ yếu là Nitrobacter) có vai trò thúc đẩy quá trình nitrit hoá nhờ đó làm giảm bớt nồng độ NH3 trong nước.
Nhóm vi khuẩn Bacillus sp. cũng thường được sử dụng để xử lý nước. Ngày nay probiotic đã được dùng khá phổ biến trong chăn nuôi và nuôi trồng thuỷ sản.
Thông thường người ta phối hợp probiotic với prebiotic (prebiotic là những chất hỗ trợ vi khuẩn có lợi phát triển và loại bỏ những vi khuẩn có hại như manan oligosacharide và glucan oligosacharide), với vi khoáng và với chất hấp phụ amoniac. Ví dụ, Shrimp-604TM của hãng Alltech (Mỹ) là sản phẩm phối hợp của manan oligosacharide, crôm và selen hữu cơ cùng với chất hấp phụ amoniac chiết rút từ cây yucca dùng cho tôm rất hiệu quả (năng suất và tỷ lệ nuôi sống tăng 10%, hiệu suất chuyển hoá thức ăn tăng 20%, tốc độ tăng trưởng tăng 30%).

82
Câu hỏi 1. Đặc điểm dinh dưỡng của nhóm thức ăn giầu năng lượng và protein 2. Kể tên và nêu vai trò của những thức ăn bổ sung dinh dưỡng và phi dinh dưỡng. 3. Vì sao người ta đánh giá cao giá trị dinh dưỡng của thức ăn tự nhiên, ấu trùng tôm cá
thích loại thức ăn nào? 4. Các chất độc nguồn gốc hóa học, sinh học và các chất kháng dinh dưỡng trong thức ăn
của cá và tác hại của chúng.

83
CHƯƠNG X THỨC ĂN TỰ NHIÊN
10.1. VAI TRÒ CỦA THỨC ĂN TỰ NHIÊN ĐỐI VỚI NUÔI TRỒNG THỦY SẢN Thức ăn tự nhiên hay thức ăn sống (natural food, live food ) là các phiêu sinh (plankton).
Phiêu sinh chính là chuỗi thức ăn sơ cấp và thứ cấp cho hầu hết động vật nước. Phiêu sinh có hai nhóm, đó là phiêu sinh thực vật (phytoplankton) và phiêu sinh động vật (zooplankton). Phytoplankton được coi là sinh vật tự dưỡng, bởi vì chúng có thể sử dụng nguồn cacbon, nitơ đơn giản và ánh sáng mặt trời để sinh trưởng và phát triển; chúng là điểm xuất phát của chuỗi thức ăn. Zooplankton được coi là sinh vật dị dưỡng, chúng ăn những sinh vật tự dưỡng và các sinh vật dị dưỡng khác. Zooplankton là nguồn thức ăn quan trọng của ấu trùng tôm và cá trong tự nhiên hay nuôi trồng.
Trong những năm 70, sản xuất của các trại cá và tôm hầu như dựa chủ yếu vào việc đánh bắt những cá giống (giai đoạn cá bột) sống trong tự nhiên. Tuy nhiên, từ sau khi kỹ thuật sản xuất ấu trùng từ đàn bố mẹ trở nên phổ biến thì hàng tỷ ấu trùng cá, ấu trùng thân mềm và giáp xác đã được sản xuất trong các trại giống trên toàn thế giới.
Ấu trùng là những sinh vật còn rất nhỏ, yếu đuối và chưa phát triển đầy đủ về mặt sinh lý như kích thước miệng còn nhỏ, giác quan và hệ thống tiêu hoá chưa hoàn thiện. Những yếu tố này hạn chế việc lựa chọn và sử dụng thức ăn thích hợp trong những pha nuôi dưỡng đầu tiên của ấu trùng. 10.1.1. Kích thước miệng ấu trùng ở pha nuôi dưỡng đầu
Kích thước miệng của ấu trùng pha đầu giới hạn kích thước của các tiểu phần thức ăn mà ấu trùng có thể ăn. Nói chung, kích thước miệng có liên quan với kích thước cơ thể và kích thước cơ thể lại bị chi phối bởi đường kính của trứng và thời kỳ nuôi dưỡng nội sinh (tức là thời kỳ tiêu thụ túi lòng đỏ). Ví dụ: trứng cá hồi Atlantic thường lớn hơn trứng Gilhead seabream ít nhất 4 lần (bảng 10.1). Kết quả là ở lúc nở, ấu trùng cá hồi khá lớn và túi lòng đỏ cũng lớn đủ cung cấp thức ăn nội sinh trong 3 tuần phát triển đầu tiên, còn như ấu trùng cá Gilhead seabream thì rất nhỏ, túi lòng đỏ cũng nhỏ và cung cấp thức ăn nội sinh chỉ đủ trong khoảng 3 ngày. Ở pha đầu, ấu trùng cá hồi có thể ăn được những tiểu phần thức ăn có kích thước 1mm, còn như ấu trùng Gilthead seabream chỉ ăn được những tiểu phần có kích thước 0,1mm.
Bảng 10.1. Kích thước trứng và chiều dài ấu trùng lúc nở của một số loài cá*
Loài Đường kính trứng (mm) Chiều dài ấu trùng (mm) Cá hồi Atlantic (Salmo salar) Cá hồi vân (Onchorhyncus mykiss) Cá chép (Cyprinus caprio) Seabass châu Âu (Dicentrarchus labrax) Gilthead seabream (Sparus aurata) Turbo (Scophthalmus maximus) Sole (Solea solea) Milkfish (Chanos chanos) Grey mullet (Mugil cephalus) Greasy grouper (Epinephelus tauvina) Bream (Acanthopagrus cuvieri)
5,0 - 6,0 4,0 0,9 - 1,6 1,2 - 1,4 0,9 - 1,2 0,9 - 1,2 1,0 - 1,4 1,1 - 1,25 0,9 - 1,0 0,77 - 0,90 0,78 - 0,84
15,0 - 25,0 12,0 - 20,0 4,8 - 6,2 7,0 - 8,0 3,5 - 4,0 2,7 - 3,0 3,2 - 3,7 3,2 - 3,4 1,4 - 2,4 1,4 - 2,4 1,8 - 2,0
* Nguồn: Jones và Houde, 1981 (dẫn theo Patric Lavens và Patric Sorgelooos,1996)

84
10.1.2. Ống tiêu hoá Tình trạng phát triển của hệ thống tiêu hoá của ấu trùng pha đầu cũng cho biết ấu trùng
có thể tiêu hoá được thức ăn ăn vào hay không. Ví dụ: ấu trùng cá hồi pha đầu đã có ống tiêu hoá phát triển tốt với hệ thống enzyme cho phép tiêu hoá được thức ăn đập vụn (feed crumble). Ngược lại, ấu trùng cá Gilhead seabream (giống như ấu trùng nhiều loài cá khác) không có dạ dày hoàn chỉnh, ống tiêu hoá thì ngắn, chỉ có một số enzyme hoạt động ở đầu pha nuôi dưỡng. Từ đó, thấy rằng ấu trùng của những cá này chỉ có thể sử dụng được những nguồn thức ăn (i) dễ tiêu hoá (như thức ăn chứa một số lớn axit amin tự do và oligopeptide thay cho những phân tử protein phức hợp), (ii) chứa enzyme cho phép thức ăn tự phân giải, và (iii) cung cấp đầy đủ các chất dinh dưỡng quan trọng mà ấu trùng yêu cầu.
Nói chung, thức ăn phối chế không đáp ứng được tất cả các yêu cầu cho ấu trùng các loài cá nhỏ như Gilhead seabream và kết quả là ấu trùng nghèo sinh trưởng và có tỷ lệ chết cao. Trái lại, thức ăn sống thì có thể đáp ứng được tất cả các tiêu chuẩn cần thiết của ấu trùng các loài cá nhỏ. Ngoài ra, thức ăn sống còn có tác dụng kích thích sự phát triển các giác quan của ấu trùng. Các giác quan của ấu trùng như thị giác, khứu giác, vị giác và đường bên rất quan trọng đối với việc phát hiện thức ăn. Tuy vậy, các cơ quan này lại phát triển chưa hoàn thiện khi ấu trùng còn non. Ví dụ: võng mạc mắt của ấu trùng cá chỉ chứa tế bào hình nón, khiến cho năng lực thị giác kém, nhưng võng mạc mắt của cá giai đoạn lớn hơn (giai đoạn cá hương), ngoaì tế bào hình nón còn có tế bào hình gậy với nhiều sắc chất thị giác trên võng mạc, nhờ vậy năng lực thị giác tốt hơn. Thức ăn sống thường có độ tương phản tốt hơn thức ăn nhân tạo và nhờ vận động liên tục mà có hiệu quả kích thích, giúp cho giác quan ấu trùng phát triển. Ngoài ra, nhờ khả năng bơi của thức ăn sống mà thức ăn được phân bố đều trong cột nước, giúp cho ấu trùng có nhiều cơ hội gặp được thức ăn.
Khẩu phần tự nhiên của hầu hết các loài cá nuôi gồm những loài phytoplankton (diatoms, flagellates vv..) và zooplankton khác nhau (copepods, cladocerans, ấu trùng decapod, rotifers, cilliates vv..). Sự phong phú và đa dạng của thức ăn sống với những kích thước và thành phần dinh dưỡng khác nhau đã cho ấu trùng cá những cơ may tuyệt vời để tăng trưởng và phát triển.
Ngày nay, ba nhóm thức ăn sống dùng phổ biến để nuôi ấu trùng ở quy mô công nghiệp (industrial larviculture) là:
+ Những loài vi tảo có kích thước 2 đến 20μm sử dụng cho hai mảnh vỏ (bivalves), tôm he (penaeid shrimp), rotifer, copepods.., cá.
+ Brachionus plicatilis sử dụng cho giáp xác (crustaceans), cá biển + Artemia spp. (meta-)nauplii cho giáp xác, cá Ngoài các nhóm trên, một số loại thức ăn sống khác cũng được dùng với mức độ hạn chế
hơn, bao gồm Brachionus rubens, Moina spp., daphnids, trứng Artemia khử vỏ cho cá nước ngọt và ấu trùng tôm và sinh khối Artemia cho ấu trùng tôm hùm, postlarrvae tôm và tôm bố mẹ, cá biển giai đoạn cá hương. Trong vài năm gần đây người ta đạt được kết quả rất tốt trong việc nuôi tôm bằng một số sản phẩm bổ sung và thay thế thức ăn sống. Tuy nhiên, đối với việc nuôi cá biển giai đoạn đầu bằng loại thức ăn này thì kết quả còn rất hạn chế.
10.2. VI TẢO (MICRO-ALGAE) Vi tảo là nguồn thức ăn cho tất cả các giai đoạn sinh trưởng của động vật hai mảnh vỏ,
thức ăn cho ấu trùng của một số loài giáp xác và thức ăn cho một số loài cá ở giai đoạn sinh trưởng đầu. Mặt khác tảo lại là nguồn thức ăn của zooplankton và chính zooplankton lại là nguồn thức ăn cho ấu trùng của cá và giáp xác giai đoạn ấu trùng và giai đoạn cá hương. Bên

85
cạnh đó, việc nuôi ấu trùng cá biển theo kỹ thuật nước xanh (green water technique), tảo được dùng trực tiếp trong các tank ấu trùng và chúng đóng vai trò làm ổn định chất lượng nước, cung cấp dinh dưỡng cho ấu trùng và khống chế vi khuẩn (hình 10.1).
10.2.1.Các loài tảo nuôi trồng chủ yếu Ngày nay 40 loài vi tảo
khác nhau được phân lập từ khắp nơi trên thế giới được nuôi trồng theo phương thức thâm canh.
Bảng 10.2 cho biết 8 lớp chính và 32 giống tảo nuôi trồng đang được dùng để nuôi dưỡng những nhóm động vật nước quan trọng.
Bảng kê các loài tảo trong bảng bao gồm các loài diatoms, tảo xanh chlorococcalean và tảo xanh hình roi (flagellated), tảo xanh hình sợi có kích thước từ vài micromet đến hơn 100μm. Hầu hết những loài tảo dùng thường xuyên trong nuôi biển là diatoms (Skeletonema costatum, Thalassiosira pseudonana, Cheatoceros gracilis, C. Calcitrans), tảo roi (Isochrysis galbana, Tetraselmis suecica, Monochrysis lutheri và Chorella spp.) (hình 10.2).
Bảng 10.2. Các lớp và giống vi tảo chủ yếu được nuôi trồng (De Pauw và Persoone, 1988)
Lớp Giống Ứng dụng Skeletonema PL, BL, BP Thalassiosira PL, BL, BP Phaeodactylum PL, BL, BP, ML, BS Chaetoceros PL, BL, BP, BS Cylindrotheca PL Bellerochea BP Actinocyclus BP Nitzchia BS
Bacillariophyceae
Cyclotella BS Isochrysis PL, BL, BP, ML, BS Pseudoisochrysis BL, BP, ML
Haptophyceae
Dicrateria BP Chrysophyceae Monochrysis (Pavlova) BL, BP, BS, MR
Tetraselmis (Platymonas) PL, BL, BP, AL, BS, MR Prasinophyceae Pyramimonas BL, BP

86
Micromonas BP Chroomonas BP Cryptomonas BP
Cryptophyceae
Rhodomonas BL, BP Cryptophyceae Chlamydomonas
Chlorococcum BL, BP, FZ, MR, BS BP
Xanthophyceae Olisthodiscus BP Carteria BP Chlorophyceae Dunaliella BP, BS, MR
Cyanophyceae Spirulina PL, BP, BS, MR Ghi chú: PL: ấu trùng tôm he BL: ấu trùng nhuyễn thể hai mảnh vỏ ML: ấu trùng tôm nước ngọt BP: hậu ấu trùng nhuyễn thể hai mảnh vỏ
AL: ấu trùng bào ngư MR: rotifers nước mặn (Brachionus) BS: Tôm nước mặn (Artemia) SC: copepods nước mặn FZ: zooplankton nước ngọt
10.2.2. Giá trị dinh dưỡng của vi tảo
Giá trị dinh dưỡng của vi tảo phụ thuộc vào kích thước tế bào, tỷ lệ tiêu hoá, chất độc và thành phần sinh hoá (bảng 10.3). Điều kiện nuôi trồng cũng ảnh hưởng nhiều đến giá trị dinh dưỡng của vi tảo. Tuy biến động khá rộng tuỳ theo các lớp và các loài, nhưng hàm lượng protein, lipid và carbohydrate biểu thị bằng % chất khô nằm trong phạm vi 12-35, 7,2-23 và 4,6-23, lần lượt.
Hình 10.2. Một vài loại tảo biển dùng làm thức ăn trong nuôi trồng thuỷ sản: (a) Tetraselmis spp., (b) Dunaniella spp., (c) Chaetoceros spp. (Laing, 1991)
Các acid béo chưa no HUFA, đặc biệt eicosapentaenoic acid (EPA, 20:5n-3),
arachidonic acid (ARA, 20:4n-6) và docosahexaenoic acid (DHA, 22:6n-3) giữ vai trò quan trọng trong việc đánh giá giá trị dinh dưỡng đối với với một loài tảo dùng để nuôi động vật biển. EPA có nhiều trong các loài diatom (Chaetoceros calcitrans, C. gracilis, S. costatum, T. pseudomonas) và prymnesiophyte Platymonas lutheri, còn DHA thì có nhiều trong prymnesiophytes (P. lutheri, Isochrysis sp.) và Chroomonas salina.
Thành phần acid béo của 10 loài vi tảo phát triển dưới những điều kiện xác định và thu hoạch ở pha log được trình bày ở đồ thị 10.3.
Vi tảo cũng là một nguồn giầu vitamin C (0,11 - 1,62 %/chất khô).
Bảng 10.3. Hàm lượng chlorophyl a (Chl a), protein, carbohydrtae (CHO) và lipid của 16 giống vi tảo dùng phổ biến trong nuôi thuỷ sản (% theo chất khô)

87
Lớp và giống vi tảo Chất khô (pg.cell-1)
Chl a Protein (pg.cell-1)
CHO Lipid
Bacillariophyceae Chaetoceros calcitrans 11.3 0.34 3.8 0.68 1.8 Chaetoceros gracilis 74.8 0.78 9.0 2.0 5.2 Nitzchia closterium - - - - - Phaeodactylum tricornutum 76.7 0.41 23.0 6.4 10.7 Skeletonema costatum 52.2 0.63 13.1 2.4 5.0 Thalassiosira pseudonana 28.4 0.27 9.7 2.5 5.5 Chlorophyceae Dunaliella tertiolecta 99.9 1.73 20.0 12.2 15.0 Nannochloris atomus 21.4 0.080 6.4 5.0 4.5 Cryptophyceae Chroomonas salina 122.5 0.98 35.5 11.0 14.5 Eustigmatophyceae Nannochloropsis oculata 6.1 0.054 2.1 0.48 1.1 Prasinophyceae Tetraselmis chui 269.0 3.83 83.4 32.5 45.7 Tetraselmis suecica 168.2 1.63 52.1 20.2 16.8 Prymnesiophyceae Isochrysis galbana 30.5 0.30 8.8 3.9 7.0 Isochrysis aff. Galbana (T-iso) 29.7 0.29 6.8 1.8 5.9 Pavlova lutheri 102.3 0.86 29.7 9.1 12.3 Pavlova salina 93.1 0.34 24.2 6.9 11.2 % theo vật chất khô Bacillariophyceae Chaetoceros calcitrans 11.3 3.01 34 6.0 16 Chaetoceros gracilis 74.8 1.04 12 4.7 7.2 Nitzchia closterium - - 26 9.8 13 Phaeodactylumtricornutum 76.7 0.53 30 8.4 14 Skeletonema costatum 52.2 1.21 25 4.6 10 Thalassiosira pseudonana 28.4 0.95 34 8.8 19 Chlorophyceae Dunaliella tertiolecta 99.9 1.73 20 12.2 15 Nannochloris atomus 21.4 0.37 30 23.0 21 Cryptophyceae Chroomonas salina 122.5 0.80 29 9.1 12 Eustigmatophyceae

88
Nannochloropsis oculata 6.1 0.89 35 7.8 18 Prasinophyceae Tetraselmis chui 269.0 1.42 31 12.1 17 Tetraselmis suecica 168.2 0.97 31 12.0 10 Prymnesiophyceae Isochrysis galbana 30.5 0.98 29 12.9 23 Isochrysis aff. Galbana (T-iso) 29.7 0.98 23 6.0 20 Pavlova lutheri 102.3 0.84 29 9.0 12 Pavlova salina 93.1 0.98 26 7.4 12
Tảo Chlorella Chlorella được biết đến nhiều bởi vai trò quan trọng của nó về dinh dưỡng cũng như là
nhân tố môi trường trong nuôi trồng thủy sản. Việc dùng Chlorella vào trong sản xuất đó là phương pháp nước xanh được áp dụng rộng rãi trong sản xuất giống tôm càng, một số loài cá và hai mảnh vỏ.
Khi ương ấu trùng tôm càng xanh, Cohen (1976) thấy rằng sự hiện diện của thực vật phiêu sinh có thể thúc đẩy sự tăng trưởng của ấu trùng tôm thông qua việc loại bỏ NH3 và một số chất độc khác. Joshep (1977) cũng ghi nhận sự bổ sung tảo sẽ làm cho môi trường nước trở nên giàu dinh dưỡng, cung cấp những hợp chất vi lượng mà thức ăn ban đầu và thức ăn bổ sung không có.
Chlorella cũng được chú ý nhiều trong sản xuất giống cua Scylla serrata. Ở Đài Loan, Nhật Bản, ấu trùng cua được ương trong môi trường có bổ sung Chlorella. Thêm Chlorella vào môi trường ương, tỉ lệ sống của cua tăng cao.
Ngoài ra, Chlorella còn là thức ăn rất quan trọng trong ương nuôi luân trùng và động vật phiêu sinh khác. Bên cạnh những loài Chlorella biển, loài Chlorella nước ngọt, Chlorella vulgairs, cũng được thử nghiệm thành công làm thức ăn cho luân trùng. Theo một số báo cáo, tốc độ tăng trưởng và sức sinh sản trung bình của luân trùng cao nhất khi cho ăn Chlorella, tiếp theo là loài Isochrysis galbana (Nagata và Whyte, 1992). Với hàm lượng HUFA cao, Chlorella không chỉ là thức ăn quan trọng của luân trùng mà còn được dùng để làm giàu acid béo cho luân trùng và một số động vật phù du khác trước khi dùng chúng làm thức ăn cho cá và các loài nuôi thủy sản khác.
Tảo Dunaliella Tảo Dunaliella có chứa hàm lượng glycerol và β-caroten cao nên được xem là đối
tượng nuôi đầy triển vọng, dùng làm thức ăn không chỉ trong nghề nuôi thủy sản mà còn nhiều lĩnh vực khác. Trong các loài thuộc giống Dunaliella, loài D. salina có hàm lượng β-caroten cao nhất (Borowithzka, 1990), chiếm 20% trọng lượng khô (Kranzfelder, 1991). Trong nuôi thủy sản, Dunaliella đóng vai trò trong chế độ dinh dưỡng của nhuyễn thể nhưng với mức độ khác nhau tùy theo loài nhuyễn thể.
Quan sát quá trình tiêu hóa của ấu trùng Strombus gigas với 8 loại tảo khác nhau (Isochrysis galbana, Tetraselmis chuii, T. seucica, Dunaliella tertiolecta, Chlamidomonas cocoides, Thalassiosira fluviatilis, Chlorella sp., và Chaetoceros sp.), Aranda (1994) thấy rằng quá trình tiêu hóa tảo Tetraselmis chuii, Chaetoceros sp. và Chlorella sp. nhanh hơn so với 5 loài tảo còn lại. Sự tiêu hóa tảo Chaetoceros sp. và Chlorella sp. nhanh hơn so với tảo D. tertiolecta...Khi ương ấu trùng Mytilus galloprovincialis với các loài tảo Dunaliella tertiolecta, T. seucica, I. Galbana, P. tricornutum, dạng đơn lẻ hay hỗn hợp, kết quả cho thấy tỉ lệ sống và

89
sức tăng trưởng của ấu trùng thấp nhất ở nghiệm thức cho ăn hỗn hợp T. seucica và I. Galbana (Moskera và ctv., 1989). Dunaliella không chỉ có vai trò quan trọng trong ương nuôi nhuyễn thể mà chúng còn dùng làm thức ăn cho một số loài cá biển, cho Artemia và động vật phiêu sinh khác.
Đồ thị 10.3. Thành phần acid béo của 10 loài vi tảo (Volkman et al., 1989).
10.2.3. Vi tảo trong nuôi trồng thủy sản
Tảo Spirulina Cùng với Chlorella và Dunaliella,
Spirulina cũng là loài tảo rất giàu protein, acid amin thiết yếu, acid béo, khoáng, vitamin và các hợp chất carotenoid nên chúng được xem là nguồn dinh dưỡng rất tốt trong nuôi thủy sản. Mustafa và ctv. (1994) thông báo Spirulina được thêm vào làm thức ăn bổ sung cho Pagrus major với tỉ lệ 5% đã làm tăng tốc độ tăng trưởng của cá, hiệu quả chuyển đổi thức ăn và hiệu suất sử dụng protein; thành phần protein có trong thịt cá không bị ảnh hưởng xấu. So với những loại tảo có kích thước lớn được thí nghiệm trước đó thì loài Spirulina ảnh hưởng tốt nhất đến sự tăng trưởng và sử dụng thức ăn của cá Red sea beam. El (1994) cũng cho biết tốc độ tăng trưởng và hiệu quả sử dụng thức ăn của cá Silver sea beam khác nhau không có ý nghĩa giữa
nghiệm thức có bổ sung 50% Spirulina trong khẩu phần ăn với nghiệm thức đối chứng 100% bột cá. Tuy nhiên, thay 75% Spirulina thì có ảnh hưởng bất lợi. Spirulina cũng được đề nghị thay thế một phần bột cá trong chế độ ăn của cá rô phi O. mossambicus.
Boonyarapalin và ctv (1989) nghiên cứu về sự thay đổi màu sắc của cá rô phi đỏ Oreoromic niloticus với các nguồn bổ sung sắc tố khác nhau, gồm: Spirulina marigold, Pepal meal, bột đầu tôm, bột nghệ và thức ăn đối chứng. Ông thấy rằng tương ứng với các loại thức ăn trên, màu sắc của cá rô phi thay đổi theo thứ tự nâu đỏ, vàng lam, lam, cam và hơi cam. Một
Ghi chú: (a) C16- & C18-polyunsaturated fatty acids (PUFA); (b) 20:5n-3 and 22:6n-3; (c) (n-3) and (n-6) PUFA. Các loài tảo (viết tắt): C. CAL: Chaetoceros calcitrans; C.GRA: C. gracilis; SKEL: Skeletonema costatum; THAL: Thalassiosira pseudonana; ISO: Isochrysis sp. (Tahitian); PAV: Pavlova lutheri; DUN: Dunaliella tertiolecta; NAN: Nannochloris atomus; TET: Tetraselmis suecica; CHRO: Chroomonas salina.

90
nghiên cứu khác cũng cho biết sắc tố của cá chép trở nên đậm hơn khi cho cá ăn khẩu phần có bổ sung 10% Spirulina. Nguồn cung cấp sắc tố khác là Marigold petal chỉ cho sự biến đổi màu nhẹ (Wutiporn-Phromkunthong, 1984). Sự biến đổi màu sắc của tôm sú nuôi cũng được Okada và ctv nghiên cứu. Các tác giả nghiên cứu sử dụng các nguồn bổ sung carotenoid khác nhau trong khẩu phần ăn của tôm từ β-caroten, Spirulina, Phaffia và krill oil. Họ nhận thấy Spirulina cho kết quả tốt nhất về sự gia tăng hàm lượng caroten trong vỏ của tôm. Nên cho tôm ăn với khẩu phần chứa 3% Spirulina trong 1 tháng trước khi thu hoạch.
Nghiên cứu về ảnh hưởng của các nguồn protein khác nhau trong khẩu phần ăn của tôm thẻ (Penaeus indicus), Ali (1992) phát hiện Spirulina và đậu phụng cho sức tăng trưởng của tôm tốt hơn so với bánh dầu dừa; hiệu quả sử dụng protein thô và giá trị sinh học của Spirulina cao hơn so với đậu phụng. Khi ương ấu trùng tôm he (Penaeus) từ gia đoạn Zoea 1 đến Mysis 2, Gu và ctv (1989) thấy rằng ấu trùng được cho ăn Spirulina apletensis và S. platensis cộng với bột đậu nành, đạt kích cỡ 663-757 µm, dài hơn có ý nghĩa so với thức ăn đối chứng chỉ dùng bột đậu nành. Tỉ lệ tôm sống của nghiệm thức trên đạt 48-53%, cao hơn rất nhiều so với đối chứng (11%).
Tảo khuê Trong lớp tảo khuê, loài Skeletonema costatum được phân lập lần đầu tiên bởi Masue
(1941) đã được dùng rộng rãi và là thức ăn rất quan trọng của âu trùng tôm biển. Hudinaga đã đạt được thành công đầu tiên trong việc sử dụng tảo này làm thức ăn cho ấu trùng tôm, tỉ lệ sống ở giai đoạn Mysis đạt 30%, cao hơn rất nhiều so với các kết quả trước đây, chỉ đạt 1% (Liao, 1983). Từ kết quả đó, nhiều loài tảo khuê khác như Chaetoceros sp., Thlasiosira, Isochrysis,... cũng được nghiên cứu làm nguồn thức ăn cho ấu trùng tôm. Tùy theo từng loài tảo và đặc điểm của chúng mà mỗi loài đều có những ưu điểm và nhược điểm riêng đối với ấu trùng tôm.
Trong quá trình phát triển của lĩnh vực sản xuất tôm giống, người ta đã chế biến ra nhiều loại thức ăn nhân tạo để thay thế một phần hoặc toàn bộ tảo khuê. Tuy nhiên, đến nay tảo khuê vẫn được xem là thức ăn tươi sống rất quan trọng của ấu trùng tôm. Trong thí nghiệm so sánh về sự ảnh hưởng của 9 loại thức ăn nhân tạo dùng thay cho tảo khuê (Chaetoceros) làm thức ăn cho ấu trùng tôm, Utama và ctv (1992) nhận thấy giảm mật độ tảo từ 50.000 xuống còn 5.000 tế bào/ml do việc thay thế tảo bằng thức ăn nhân tạo vẫn cho kết quả tốt, tỉ lệ sống của ấu trùng tôm giữa các nghiệm thức vẫn không khác nhau, nhưng mật độ tảo không thể thấp hơn 5.000 tế bào/ml. Hơn nữa, Chu (1991) cũng nhận thấy ấu trùng tôm Metapenaeus ensis và Penaeus chinese cho ăn thức ăn nhân tạo bị chậm lớn và tỉ lệ sống luôn thấp hơn so với tôm cho ăn Chaetoceros garcilis và Artemia. Kết quả thí nghiệm của Chu (1989) cho thấy chỉ dùng một loài tảo Chaetoceros gracilis có thể cung cấp chế độ ăn đầy đủ dinh dưỡng cho ấu trùng tôm Metapenaeus ensis từ giai đoạn Zoea đến PL6 với tỉ lệ sống đạt 35-63%. Việc bổ sung Artemia không làm cải thiện được tỉ lệ sống của ấu trùng tôm.
Bên cạnh đó, tảo khuê còn đóng vai trò quan trọng trong nuôi nhuyễn thể. Okauchi (1990) thí nghiệm tìm hiểu về vai trò của tảo đối với spat của trai (Pintctada fucata) và thấy sức tăng trưởng của spat cho ăn chỉ có tảo Isochrysis aff galbana thấp hơn so với spat cho ăn kết hợp Isochrysis galbana và Chaetoceros garcilis. Laing và ctv (1990) nghiên cứu về giá trị dinh dưỡng của tảo khô loài Nannochloris sp. và Tetraselmis seucica so với lô đối chứng gồm hỗn hợp tảo Chaetoceros calcitral và T-ISO dùng làm thức ăn cho ấu trùng nghêu Manila (Tapes philipinarum), thấy rằng ấu trùng nghêu cho ăn tảo khô có sức tăng trưởng bằng hoặc cao hơn so với cho ăn dạng tươi sống, nhưng thấp hơn so với lô đối chứng.

91
10.3. LUÂN TRÙNG (ROTIFERS)
10.3.1. Hình thái Rotifer thuộc về lớp động vật đa bào nhỏ nhất. Trong số hơn 1000 loài đã được nhận
biết, có tới 90% loài sống ở môi trường nước ngọt. Chiều dài cơ thể của rotifer khoảng 2mm, con đực nhỏ hơn và kém phát triển hơn con cái. Cơ thể của tất cả các loài gồm một số tế bào cố định (ví dụ các loài Brachionus chứa khoảng 1000 tế bào) (xem hình 10.6 mô tả sơ lược một số loài rotifer nước mặn và nước ngọt).
Rotifer có một lớp vỏ keratin được gọi lorica, hình thái và các loại gai cho phép phân biệt sự khác nhau giữa các loài.
Cơ thể rotifer có 3 phần là đầu, thân và chân. Đầu mang cơ quan vận động, đó là các lông hình khuyên, giúp con vật vận động xoay tròn (vì thế rotifer có tên gốc là Rotatoria). Thân chứa ống tiêu hóa, cơ quan bài tiết và sinh sản. Cơ quan đặc trưng của rotifer là mastax, một cơ quan đã được canxi hóa ở vùng miệng, có vai trò nghiền những tiểu phần thức ăn đã ăn vào (xem hình 10.4). 10.3.2. Một vài đặc điểm sinh học quan trọng
Rotifer có hai nhóm loài, nhóm loài nước mặn và nhóm loài nước ngọt.
Trong nhóm loài nước mặn, rotifer dùng trong nuôi ấu trùng biển thường là rotifer kiểu nhỏ (có tên là Brachionus rotundiformis) và kiểu lớn (có tên là Brachionus plicatilis). Chiều dài lorica của rotifer kiểu lớn trung bình là 239m/m (130-340m/m), còn của kiểu nhỏ là 160m/m ( 100-21-m/m).
Rotifer nước ngọt thường dùng trong nuôi ấu trùng nước ngọt là các loài Brachionus calyciflorus và Brachionus rubens.
Đời sống của rotifer nước mặn kéo dài khoảng 3,4 - 4,4 ngày ở 25oC. Nói chung, ấu trùng trưởng thành sau 0,5 - 1,5 ngày và sau đó thì con cái đẻ trứng cứ 4 giờ một lần. Người ta cho rằng con cái có thể sinh ra 10 thế hệ trước khi chết. Năng lực sinh sản của Brachionus phụ thuộc vào nhiệt độ môi trường, trong phạm vi từ 15 đến 25oC tuổi con cái đẻ lần đầu, khoảng cách giữa hai lần đẻ và tuổi thọ càng ngắn khi nhiệt độ càng cao và số trứng đẻ được trong cả đời cũng nhỏ hơn khi nhiệt độ tăng.
Trong vòng đời của Brachianus plicatilis thường có hai kiểu sinh sản, sinh sản vô tính (asexual) và hữu tính (sexual). Trong sinh sản đơn tính, con cái sản sinh trứng amictic (diploid, 2 nhiễm sắc thể), trứng sẽ nở thành con cái amictic. Trong điều điều kiện môi trường nào đó, con cái sinh sản

92
hữu tính, sinh ra con cái mictic và amictic. Tuy không phân biệt được về hình thái, nhưng con cái mictic thì sản sinh ra trứng haploid (n nhiễm sắc thể). Ấu trùng nở ra từ trứng mictic không thụ tinh thì phát triển thành con đực haploid, có kích thước nhỏ chỉ bằng khoảng 1/4 kích thước của con cái, không có ống tiêu hóa và bong bóng nhưng có một dịch hoàn chứa đầy tinh trùng. Trứng mictic thụ tinh thì có kích thước lớn hơn và có một lớp vỏ hạt dày bọc ngoài. Đây là những trứng nghỉ mà sẽ chỉ phát triển và nở ra những con cái amictic sau khi tiếp xúc với môi trường thuận lợi (xem hình 10.5).
Điều kiện sống của rotifer phụ thuộc vào độ mặn, nhiệt độ, oxy hòa tan, pH, hàm lượng NH3, vi khuẩn.
Brachianus plicatilis có thể sống trong môi trường có độ mặn từ 1 đến 97 ppt, tuy nhiên khả năng sinh sản tốt nhất khi độ mặn dưới 35 ppt (Lubzens, 1987). Hầu hết các cơ sở ương nuôi rotifer sử dụng độ mặn 10-20 ppt. Độ mặn thay đổi 5 ppt có thể ức chế khả năng bơi và gây chết, như vậy việc thay đổi độ mặn phải thực hiện từ từ và thận trọng.
Nhiệt độ tối ưu cho rotifer kiểu lớn thấp hơn kiểu nhỏ, nói chung tăng nhiệt độ trong phạm vi thích hợp thì tăng khả năng sinh sản. Nuôi rotifer dưới nhiệt độ tối ưu thì làm chậm sự tăng trưởng của quần thể. Nhiệt độ tối ưu thích hợp với hầu hết các nhóm rotifier là 28-32oC. 10.3.3. Giá trị dinh dưỡng của rotifer
Rotifer rất giàu protein (52-59% CP) và chất béo (13% EE), đặc biệt là các HUAF (3,1% n-3 HUFA). Rotifer là thức ăn lý tưởng của ấu trùng tôm và cá vì chúng có kích thước nhỏ, bơi chậm, sinh trưởng với mật độ cao và sinh sản nhanh.
Giá trị dinh dưỡng của rotifer phụ thuộc vào môi trường. Người ta có thể tăng giá trị dinh dưỡng của rotifer bằng cách nuôi rotifer trong các môi trường được làm giầu các chất dinh dưỡng như acid béo omega-3, vitamin C, protein ...
Ví dụ, hàm lượng HUFA của rotifer nuôi trong môi trường tảo Chlorella sp., men bánh mì hay các môi trường nhân tạo khác của hãng Selco đã thay đổi rất nhiều theo với các môi truờng khác nhau, HUFA của rotifer nuôi trong môi trường của hãng Selco có thể tăng 5-10 lần
so với môi trường tảo Chlorella hay tảo kết hợp men bánh mì (đồ thị 10.4).
Đồ thị 10.4. Hàm lượng HUFA của rotifer nuôi trong các môi trường khác nhau (CHL: Chlorella sp., Men bánh mì: BY, Protein Selco: PS, môi trường Selco: CS và Super Selco: SS)
10.3.4. Rotifer trong nuôi trồng thủy sản
Wendy (1991) từ kết quả nghiên cứu của Nagata
(1989) cho biết Brachionus plicatilis được sử dụng rất rộng rãi trên thế giới trong ương nuôi ấu trùng của trên 60 loài cá biển và 18 loài giáp xác. Zheng và ctv. (1994) nghiên cứu sử dụng rotifer cho tôm he cũng khẳng định Brachionus plicatilis là một trong những loại thức ăn thích

93
hợp nhất cho ấu trùng tôm he ở giai đoạn Mysis 3. Brachionus plicatilis còn là nguồn thức ăn tốt cho ương ấu trùng tôm Penaeus monodon, P. indicus và P. Merguiensis
Cruz và ctv (1989) báo cáo cá bột rô phi cho ăn kết hợp thức ăn chế biến với rotifer đạt được trọng lượng cuối, tốc độ tăng trưởng hàng ngày và năng suất cao hơn so với cá bột chỉ cho ăn một loại thức ăn, hoặc rotifer hoặc thức ăn chế biến. Việc sử dụng rotifer làm thức ăn cho cá bột mặc dù không làm tăng tỉ lệ sống của cá, nhưng lại làm tăng tốc độ tăng trưởng và năng suất cá bột.
10.4. ARTEMIA Trong các khẩu phần nuôi ấu trùng
tôm và cá, nauplii của Artemia được dùng nhiều nhất. Hàng năm trên 2000 tấn cyst khô Artemia được bán trên thị trường. Tính chất
duy nhất của Artemia là hình thành những phôi ngủ, gọi là “cyst”, đó là nguồn thức ăn tuyệt vời của ấu trùng tôm cá. Cyst có quanh năm với số lượng lớn dọc theo bờ các hồ nước mặn, đầm phá ven biển hay những ruộng muối trên khắp thế giới. Sau 24 giờ ấp trong nước biển, cyst nở ra những nauplii bơi tự do và có thể trực tiếp đem nuôi các loại ấu trùng động vật biển hay nước ngọt.
10.4.1. Một số đặc điểm sinh học quan trọng
Artemia trưởng thành có kích thước trên dưới 1 cm, có hai mắt kép, ống tiêu hóa thẳng, có râu anten và 11 cặp đốt ngực. Con đực có cơ quan giao cấu (penis) ở phần sau của vùng thân. Con cái có tử cung ngay sau cặp đốt ngực số 11. Trứng phát triển trong hai buồng trứng ở xoang bụng. Khi chín trứng trở nên tròn và đi vào tử cung theo ống dẫn trứng (hình 10.7).
Trong môi trường tự nhiên, Artemia sản sinh cyst, các cyst này nổi trên mặt nước. Cyst có một lớp vỏ cứng, màu nâu đậm, bảo vệ rất hiệu quả phôi sống ở bên trong và chống lại được các điều kiện bất lợi như khô, nhiệt độ chênh lệch từ 0 - 100oC, bức xạ năng lượng cao và một số dung môi hữu cơ. Cyst khô có thể dự trữ hàng tháng hay hàng năm mà không ảnh hưởng đến độ nở.
Khi ngâm vào nước biển, từ hình tròn lõm hai mặt, cyst biến thành hình cầu bên trong chứa phôi. Sau 20 giờ vỏ ngoài của cyst vỡ ra, phôi xuất hiện với màng bọc bên ngoài, đó là nauplii. Nauplii phát triển qua nhiều giai đoạn, khi bước vào giai đoạn bung dù (phôi treo dưới vỏ rỗng) thì có nghĩa là nauplii đã phát triển hoàn toàn. Sau khi màng ngoài của nauplii vỡ ra, nauplii bước vào giai đoạn bơi tự do (hình 10.8).

94
Thực ra phôi phát triển qua nhiều giai đoạn, giai đoạn đầu gọi là instar I (kích thước 400-500 microns, instar I chỉ sống bằng chất dự trữ trong túi lòng đỏ), sau 8 giờ thì vào giai đoạn instar II, ở giai đoạn này nhận được thức ăn có kích thước 1-50 microns như tảo đơn bào, vi khuẩn... Sau 15 lần lột xác (từ instar I đến instar XV), ấu trùng trở thành Artemia thành thục. Bình thường, 200.000- 300.000 nauplii nở ra từ 1 g cyst chất lượng cao.
Điêù kiện tối ưu cho cyst nở: nhiệt độ > 250C (tốt nhất 280C), độ mặn 5 ppt, nước sục khí và tuần hoàn tốt, ánh sáng đầy đủ, pH = 8, mật độ không quá 5 g cyst/lit nước. 10.4.2. Giá trị dinh dưỡng
Căn cứ thành phần axit béo, Artemia được chia làm hai nhóm: nhóm chứa nhiều EFA, phù hợp với ấu trùng cá nước ngọt (có ở Nam Mỹ); nhóm chứa nhiều PUFA (20: 5ω3) và DHA (22: 6ω3), phù hợp với ấu trùng cá nước mặn (có ở hồ Saskachewan Canada, Greatlake, Mỹ và Tien-tsin, Trung quốc).
Cyst của Artemia giầu protein (50%), chất béo (14%), nauplii có hàm lượng protein thấp hơn một chút nhưng hàm lượng chất béo lại cao hơn (bảng 10.4). Tỷ lệ acid amin tự do so với protein tổng số của nauplii cao hơn cyst; hàm lượng acid amin của nauplii khác nhau nhiều theo vùng địa lý (bảng 10.5). Phổ acid béo của cyst và nauplii giống nhau nhưng khác nhau về mứcViatmin C trong cyst ở dạng ascorbic acid 2-sulfate (AAS), là dạng rất bền nhưng có độ lợi dụng sinh học kém. Trong quá trình cyst nở thành nauplii, vitamin C được thủy phân thành dạng tự do, có độ bền thấp nhưng có độ lợi dụng sinh học cao đối với ấu trùng tôm cá.
Bảng 10.4. Thành phần dinh dưỡng của cyst Artemia bỏ vỏ và nauplii instar I (% chất khô)
Hồ Nước Lớn Vịnh San Francisco cysts nauplii Cysts nauplii
Protein ± 50 41-47 ± 57 47-59 Lipid ± 14 21-23 ± 13 16-27 Carbohydrate - 11 - 11 Tro ± 9 10 ± 5 6-14
Bảng 10.5. Thành phần acid amin của nauplii Artemia (mg.g-1 protein) (Seidel et al., 1980)
Macau, Brazil Great Salt Lake, UT-USA San Pablo Bay, CA-USA Aspartic acid 110 113 141 Threonine 52 48 60 Serine 45 54 77
95
Glutamic acid 131 135 102 Proline 57 59 49 Glycine 60 60 74 Alanine 46 49 42 Valine 53 52 55 Methionine 22 37 26 Isoleucine 56 68 54 Leucine 89 100 84 Tyrosine 105 66 77 Phenylalanine 51 85 104 Pistidine 49 27 35 Lysine 117 93 87 Arginine 115 97 98
Bảng 10.6. Hàm lượng axit béo của cyst Artemia (vùng Hồ lớn, San Francisco)
C18:0 Stearic 9.0 mg/g C18:1 Oleic 27.2 C18:3 Linoleic 9.1 C18:4 Octadecatetra 11.2 C20:0 Arachidic 1.1 C20:4 Arachidonic 4.5 C20:3 Eicosatrienoic 2.7 C20:5 Eicosapentaenoic (EPA) 4.1 C22:6 Docosahexaenoic (DHA) 0.1 Tổng Omega-3 Fatty Acids 4.0
Artemia được dùng để nuôi ấu trùng tôm penaeid giai đoạn ấu trùng sau và postlarval; nauplii mới nở thường cung cấp cho tôm ở lúc khởi phát giai đoạn mysis đầu. Tuy nhiên, penaeid thường ăn tảo trước khi ăn Artemia, thêm Artemia quá sớm trong chu kỳ sống của tôm sẽ xẩy ra hiện tượng cạnh tranh tảo giữa Artemia (Artemia không được tôm ăn) với tôm. Giải pháp thuận tiện là dùng nauplii đã làm chết (bằng cách ngâm nhanh nauplii vào nước nóng 80oC hay làm lạnh ở -100C) hay sử dụng cyst Artemia khử vỏ. Tuy nhiên ấu trùng tôm nước ngọt lại có thể sử dụng tốt nauplii của Artemia, ấu trùng Macrobrachium có thể cho ăn nauplii mới nở ngay từ giai đoạn đầu.
Ấu trùng của nhiều loài cá biển như cá bơn (turbot), cá mú (grouper) chỉ có thể sử dụng Artemia sau thời kỳ ăn những mồi nhỏ như rotifer. Tuy vậy, thời kỳ ăn Artemia của ấu trùng cá biển tương đối dài (20-40 ngày) cho nên cyst Artemia vẫn là loại thức ăn sống được tiêu thụ nhiều nhất của ấu trùng cá biển (để sản xuất 1000 fry cần 200-500 g cyst Artemia).
Artemia làm thức ăn cho tôm biển: Artemia là thức ăn rất quan trọng trong ương nuôi ấu trùng và hậu ấu trùng tôm biển. Artemia bắt đầu cho ăn khi ấu trùng đạt đến giai đoaün Mysis1 hay ngay cả Zoa E2-3. Nhiều nghiên cứu đã cho thấy rằng, thành phần dinh dưỡng của những nguồn Artemia khác nhau và ngay cả ở những đợt sản xuất khác nhau là khác nhau. Amat

96
nghiên cứu ảnh hưởng của các nguồn Artemia khác nhau lên ấu trùng một số loài tôm biển đã cho thấy rằng kết quả tốt nhất ở tôm ăn Artemia có thành phần HUFA cao nhất.
Việc sản xuất giống tôm càng xanh cũng phụ thuộc căn bản vào nguồn Artemia. Không như tôm biển, ấu tôm càng xanh có thể bắt đầu cho ăn từ giai đoạn đầu với Artemia mới nở. Tuy nhiên, cũng giống như tôm biển, trong ương nuôi ấu trùng tôm càng xanh, ngoài Artremia cần bổ sung thêm các nguồn thức ăn nhân tạo khác để tăng cường dinh dưỡng và nâng cao tỷ lệ sống của ấu trùng.
Đối với cá biển, trong sản xuất giống, Artemia thường được bắt đầu cho ăn một tuần sau khi cho ăn bằng rotifer với kích cỡ nhỏ. Lượng Artemia cần cho sản xuất giống cá thường tốn hao nhiều hơn so với sản xuất giống tôm biển và tôm càng xanh. Kết quả sản xuất giống cá biển được nâng cao rất có ý nghĩa khi Artemia được làm giàu hóa HUFA.
Đối với cá nước ngọt, rất nhiều loài cá được ương nuôi trực tiếp ở ao sau khi hết giai đoạn noãn hoàng. Ấu trùng cá sẽ sử dụng nguồn thức ăn tự nhiên trong ao như tảo, động vật phù du được gây nuôi bằng cách bón phân. Tuy nhiên, cũng có nhiều loài cá cần được ương nuôi trên bể như cá Basa, cá tra, cá lăng .., và vì thế Artemia là thức ăn rất quan trọng cho sản xuất giống những loài cá này.
Ngoài ra, trong nghề nuôi nuôi cá cảnh, cả ấu trùng Artemia và Artemia trưởng thành đều được sử dụng cho mục đích sản xuất giống hay nuôi.
Mặc dù Artemia có giá trị dinh dưỡng rất cao và có một số loài có thể dựa vào nguồn thức ăn là Atermia đơn độc, song, trong sản xuất giống, việc bổ sung các nguồn thức ăn nhân tạo hay làm giàu Artemia là điều rất cần thiết và rất phổ biến.
10.5. CÁC ZOOPLANKTON KHÁC
10.5.1. Copepod Copepod có hai nhóm, một là calcanoid (Acartia tonsa, Eurytemora affinis, Calcanus
finmarchius & C. helgolandicus, Pseudocalcanus elongatus) và hai là harpacticoid (Tsibe holothuriae, Tigriopus japonicus, Tisbenta elongata, Schizopera elatensis). Calcanoid có râu anten thứ nhất rất dài (khoảng 16-26 đốt), còn harpacticoid thì râu anten ngắn hơn (khoảng 10 đốt).
Hầu hết copepod trưởng thành có chiều dài trong khoảng 1-5m/m, cơ thể hình ống, phần trước phình to hơn phần sau. Thân gồm hai phần rõ rệt, đó là ngực và bụng, đầu lẫn vào đốt thứ nhất của phần ngực có. Đầu có mắt và râu anten khá dài. Con đực nhỏ hơn con cái, khi giao phối con đực ôm lấy con cái bằng râu anten rồi phóng tinh vào cửa của lỗ tiếp nhận tinh của con cái, sau đó cửa được bịt lại bởi một loại chất gắn đặc biệt. Trứng thường được bao bằng một túi trứng và gắn vào đốt bụng đầu tiên của con cái.
Calcanoid đẻ trứng vào nước, trứng nở thành nauplii rồi sau 5-6 lần lột xác ấu trùng trở thành copepodite. Sau 5 lần lột xác nữa copepodite bước vào giai đoạn trưởng thành. Sự phát triển từ trứng đến khi trưởng thành có thể kéo dài một tuần đến một năm. Tuổi thọ của copepod kéo dài từ 6 tháng đến một năm. Dưới những điều kiện không thuận lợi, copepode có thể sản sinh trứng nghỉ (cyst).
Kích thước của copepod phụ thuộc vàp loài và sự phát triển của cá thể. Nhờ kích thước khác nhau mà đảm bảo được thức ăn cho ấu trùng tôm cá trong tất cả các giai đoạn phát triển.
Harpacticoid Tsibe holothuriae ở giai đoạn nauplii chỉ có kích thước 55 micron đến khi trưởng thành thì có kích thước 180 micron. Schizopera elatensis có kích thước 50-500 micron, Tisbenta elongata có kích thước từ 150 đến hơn 750 micron.

97
Trong nuôi trồng copepod, người ta thấy harpacticoid ít nhậy cảm và dung nạp tốt đối với điều kiện môi trường hơn calcanoid (độ mặn: 15-70 g/lit nhiệt độ 17-30oC) và như vậy harpacticoid dễ nuôi thâm canh hơn.
Nhiều loài copepod ôn đới có thể sản sinh cyst giống như Artemia. Cyst có thể dung nạp được điều kiện làm khô ở 25oC và đông lạnh ở -25oC, chúng có thể chịu được nhiệt độ thấp (3-5oC) dài tới 9-15 tháng.
Về mặt thức ăn và chất lượng dinh dưỡng, copepod có một số ưu điểm trong nuôi ấu trùng cá biển như sau:
- Copepod có thể đem nuôi ấu trùng cá ở các giai đoạn khác nhau như nauplii, copepodite…
- Chuyển động zic-zac của copepod có tác dụng kích thích thị giác của nhiều loài cá. - Chất lượng dinh dưỡng của Copepode được cho là cao hơn Artemia, protein trong
phạm vi 44-52%, tỷ lệ acid amin cân đối trừ methionine và histidine. Thành phần acid béo khác nhau nhiều theo điều kiện nuôi trồng. Ví dụ: (n-3) HUFA của Tisbe trưởng thành nuôi bằng tảo Dunaliella (thấp HUFA) hay tảo Rhodomonas (cao HUFA) là 0,8% và 1,3% tính theo chất khô, lần lượt. Hàm lượng EPA và DHA lần lượt là 6% và 17% của Tisbe trưởng thành cho ăn tảo Dunaliella và lần lượt là 18% và 32% khi nuôi bằng tảo Rhodomonas. (n-3) HUFA của nauplii thì cao hơn Tisbe trưởng thành.
- Copepod được cho là có hàm lượng enzyme tiêu hóa cao hơn Artemia (Pedeson, 1984, khảo sát trên ấu trùng cá herring đã thấy rằng copepod được tiêu hóa tốt hơn Artemia). 10.5.2. Daphnia và Moina
Daphnia Daphnia thuộc về phân bộ Cladocera sống hầu như duy nhất ở nước ngọt. Vỏ giáp bọc
toàn thân, trừ đầu và gai đuôi (apical spine) (hình 10.9). Hiện tại có 50 loài được báo cáo trên toàn thế giới, trong đó chỉ có 6 loài có ở vùng đất
thấp nhiệt đới. Kích thước của Daphnia trưởng thành khác nhau nhiều tùy thuộc vào thức ăn; khi thức ăn phong phú, con trưởng thành có vỏ giáp (carapace) dài gấp hai lần các cá thể mới
thành thục. Không những vậy, kích thước của đầu cũng biến đổi từ tròn chuyển sang bầu dục vào mùa xuân và giữa hè, nhưng từ giữa hè trở đi thì đầu lại trở về hình tròn. Sự biến dạng này được gọi là biến hình theo chu kỳ (cyclomorph), có thể xẩy ra do những yếu tố nội tại hay do mối tương tác giữa các yếu tố di truyền và môi trường.
Bình thường, Daphnia sinh trưởng từ nauplii tới khi thành thục phải trải qua 4-5 lần lột xác, thời gian lột xác của mỗi thời kỳ phụ thuộc chủ yếu vào nhiệt độ (11 ngày ở 10oC hay 2 ngày ở 25oC) và vào thức ăn. Các loài Daphnia sinh sản đơn tính hầu như chỉ sinh ra con đực. Trứng được sản sinh trong túi trứng với 200 tới vài trăm trứng. Một con cái có thể sản xuất vài ổ trứng. Trứng đơn tính nếu được sản sinh theo kiểu không giảm phân thì kết quả cho
98
ra những con cái, nhưng đôi khi cũng xuất hiện con đực. Sự phát triển của phôi cladoceran xẩy ra trong túi ấp (broodpouch) và ấu trùng là phiên bản nhỏ của con trưởng thành. Trong điều kiện không thuận lợi trứng đã thụ tinh hình thành lớp vỏ bảo vệ phôi bên trong, đó là các trứng nghỉ (cyst).
Giá trị dinh dưỡng của Daphnia phụ thuộc nhiều vào thành phần hóa học của nguồn thức ăn. Nói chung Daphnia nghèo acid béo quan trọng, tuy nhiên Daphnia chứa một phổ rộng enzyme tiêu hóa như protease, peptidase, lipase và cellulase, là nguồn enzyme ngoại sinh tốt trong ruột của ấu trùng cá.
Moina Moina cũng thuộc phân bộ Cladocera, nhiều đặc điểm sinh học giống như Daphnia.
Moina phát triển mạnh ở ao và những hồ nước. Thời kỳ để đạt thành thục sinh sản cần 5 ngày ở nhiệt độ 26oC. Con đực nhỏ hơn con cái, con cái thành thục sinh dục chỉ mang hai trứng nằm trong ephippium. Moina nhỏ hơn Daphnia nhưng có hàm lượng protein cao hơn. Sinh khối của moina thường dùng để nuôi ấu trùng cá hồi vân, cá hồi và các ấu trùng cá nước ngọt hay nước mặn khác. Moina micrura cũng được báo cáo là có tác dụng tốt khi thay thế một phần Artemia trong nuôi ấu trùng tôm Macrobrachium rosenbergii. 10.5.3. Nematode
Nematode, Panagrellus redivivus được chứng minh là thức ăn tốt để nuôi thành công ấu trìng cá chép, cá silver carp và tôm Penaeus blebejus.
Panagrellus redivivus là loại thức ăn sống tốt vì nó có kích thước nhỏ (đuờng kính 50 micron), profile acid amin tương tự như của Artemia (bảng 10.7), nhưng EPA DHA thì cao hơn của Artmia. Mặt khác P. redivivus lại có thể nuôi trồng rất đơn giản trong các khay chứa 70 g bột mì (loại 10,8% protein)/100cm2, sau đó giữ ẩm bằng cách phun nước. Môi trường nuôi cấy hàng tuần được bổ sung 0,5 g men/100cm2. Khay nuôi cần giữ nơi thoáng khí có nhiệt độ 20-23oC, khay cần che đậy bằng vải mỏng để tránh côn trùng. Nematode có thể thu họach hàng ngày trong khoảng 53 ngày trên cùng một môi trường. Sản lượng tối đa có thể đạt 75-100 mg/100cm2 sau 3 tuần nuôi.
Bảng 10.7. So sánh hàm lượng protein và amino acid của P. redivivus and Artemia (% tổng số amino acid) (Watanabe & Kiron, 1994)
P. redivivus Artemia P. redivivus Artemia Protein 48.3 61.6 Amino acids Amino acids ILE 5.1 3.8 ARG 6.6 7.3 LEU 7.7 8.9 HIS 2.9 1.9 MET 2.2 1.3 ALA 8.8 6.0 PHE 4.7 4.9 ASP 11.0 11.2TYR 3.2 5.4 GLU 12.8 12.9THR 4.7 2.5 GLY 6.4 5.0 TRY 1.5 PRO 5.4 6.9 VAL 6.4 4.7 SER 3.7 6.7 LYS 7.9 8.9

99
10.5.4. Ấu trùng bánh xe (Trochophora larvae) Một số loài cá biển như cá bơn, cá hanh… thì
rotifer thì quá lớn cho pha nuôi dưỡng đầu của ấu trùng, ấu trùng bánh xe có thể thay thế rotifer. Ấu trùng bánh xe có kích thước khoảng 50 micron (xem H. 10.10), bơi chậm và có giá trị dinh dưỡng cao đối với ấu trùng cá biển (ấu trùng trocophora chứa 15% EPA và DHA so với tổng acid béo.
10.6. QUẢN LÝ AO NUÔI ĐỂ PHÁT TRIỂN THỨC ĂN TỰ NHIÊN
Chất lượng nước của ao nuôi cá nước ngọt phụ thuộc vào các yếu tố: vật lý, hoá học và sinh học. 10.6.1. Yếu tố vật lý
Khác nhau theo các vùng trong ao nuôi (hình 10.11). Ở tầng mặt (littoral zone), cây thuỷ sinh ảnh hưởng đến lượng oxy hoà tan (DO) của nước trong ao. Ở tầng đáy (benthic zone), tính chất đất có ảnh hưởng quan trọng đến chất lượng nước. Ở tầng giữa (limnetic zone), sinh vật nước sống ở vùng này và các số đo về chất lượng nước thực hiện tại đây.
Hình 10.11. Sơ
đồ lát cắt các vùng ao nuôi
10.6.2. Yếu tố sinh học
Yếu tố sinh học liên quan đến các sinh vật sống trong ao thông qua chu trình sau:
Vi khuẩn → Phytoplankton → Zooplankton → Cá & Tôm → Chất hữu cơ lắng cặn → Vi khuẩn → Phytoplankton …
Các pha phát triển sinh vật như ở sơ đồ cho thấy, pha trước có ảnh hưởng đến pha sau. Phân vô cơ và hữu cơ và chất lượng nước ao nuôi có ảnh hưởng đến chu trình phát triển hệ sinh vật trong ao (hình 10.12).
Hình 10.12. Sơ đồ Chu trình phát triển hệ vi sinh vật trong ao nuôi (Boyd, 1998)
Giải pháp phát triển zooplankton bằng việc bón phân hữu cơ, phân vô cơ hoặc phân hữu cơ kết hợp với phân vô cơ. Ưu điểm của phân vô cơ đó là thúc đẩy sinh vật tự dưỡng (P là nguồn thức ăn rất cơ bản của phytoplankton), thành phần tương đối ổn định, sẵn có, dễ mua, và

100
tốn ít công và dễ dự trữ. Trong khi, phân hữu cơ (phân gia súc, khô dầu bông, khô đỗ tương, bột cỏ alfalfa..) làm thúc đẩy phát triển sinh vật tự dưỡng (autotrophic organism) và dị dưỡng (heterotrophic organism), giải phóng CO2 cung cấp cho thực vật, giúp làm giảm độ đục, và có ảnh hưởng lâu bền.
Ngoài ra, việc bón vôi cho môi trường ao nuôi cũng có các tác dụng như: khử chua, tăng pH, tăng độ hoà tan của phosphor, và thúc đẩy sự phân giải của chất hữu cơ. 10.6.3. Yếu tố hoá học
Các yếu tố hoá học là: oxy hoà tan, nhiệt độ, pH, độ mặn, độ đục, độ kiềm và độ cứng, amoniac, chất dinh dưỡng.. đều ảnh hưởng đến phát triển của hệ sinh vật trong ao nuôi.
Boyd, 1998 đưa ra hàm lượng các chất vô cơ hoà tan thích hợp trong ao nuôi được trình bày ở bảng 10.8.
Bảng 10.8. Hàm lượng các chất vô cơ hoà tan thích hợp trong ao nuôi
Nguyên tố Dạng tồn tại trong nước
Hàm lượng thích hợp
Nguyên tố Dạng tồn tại trong nước
Hàm lượng thích hợp
Oxy O2 5-15 mg/l Hydro H+ pH 7-9 Nitơ N2 Bảo hòa Nitơ NH4 0,2-2 mg/l Nitơ NH3 <0,1 mg/l Nitơ NO3 0,2-10 mg/l Nitơ NO2 <0,3 mg/l Sulfur H2S; SO4 5-100 mg/l
Carbon CO2 1-10 mg/l Ca Ca2+ 5-100 mg/l Mg Mg2+ 5-100 mg/l
Quản lý ao nuôi trước và sau khi thả cá có ảnh hưởng rất quan trọng đến sự phát triển
nguồn thức ăn tự nhiên, từ đó có ảnh hưởng đến năng suất và hiệu quả nuôi cá. Sau đây là một điểm cần chú ý:
- Dọn tẩy ao trước khi thả cá. Nếu là ao mới đào thì phải dẫn nước vào – ra vài lần để rửa ao. Tẩy vôi với 10-12kg/100m2 đáy ao. Cày bừa đáy ao và phơi nắng 5-7 ngày. Bón lót phân chuồng (50kg/100m2 đáy ao). Nếu là ao cũ đã nuôi cá, phải tháo cạn để bắt hết cá, vét bớt bùn thối, phát quang bụi rậm quanh ao, lấp kín các hang hốc, đắp bờ, sửa cống cấp thoát nước. Tẩy vôi với 8-10kg/100m2 đáy ao và phơi nắng 5-7 ngày. Bón lót bằng phân chuồng và phân xanh mỗi thứ 30-40 kg/100m2. Bừa san phẳng đáy ao 1-2 lượt.
- Sau khi bón lót, cho nước vào ao, lọc nước qua đăng chắn. Mức nước lấy vào chỉ cần 0,5-0,7 m, ngâm ao 2-3 ngày, nước ao sẽ có mầu xanh nõn chuối. Trước khi thả cá phải lấy đủ nước, đảm bảo mức nước sâu từ 1 – 1,5m.
- Sau khi thả cá phải chú ý đến việc bón phân cho ao, có thể dùng phân hữu cơ như phân chuồng, phân xanh và phân vô cơ như đạm, lân, vôi... Nuôi cá kết hợp với chăn nuôi (hệ thông VAC) vừa có nguồn phân hữu cơ cho cá, vừa kiểm soát được ô nhiễm môi trường do chăn nuôi.
- Thêm nước mới vào ao nuôi cá mỗi tuần một lần (dâng cao khoảng 0,3m nước), sau 2 tuần tháo nước đáy và thêm nước mới vào tầng mặt. Những ngày có mưa giông phải kiểm tra bờ ao, cống ...Những ngày trời oi bức, thời tiết thay đổi phải theo dõi hiện tượng cá nổi đầu, nếu cá bị thiếu oxy thì phải xử lý kịp thời.
- Sau khi thu hoạch cá lại thực hiện việc xử lý ao như trình bày ở trên: vét bùn, dọn tẩy ao, bón phân... để bước vào một vụ nuôi cá mới.

101
Câu hỏi 1. Vai trò của thức ăn sống đối với nuôi trồng thủy sản? 2. Các loài tảo thường sử dụng trong NTTS? 3. Sử dụng Rotifer và Atermia trong NTTS? 4. Giải pháp phát triển nguồn thức ăn tự nhiên?

102
CHƯƠNG XI CHẾ BIẾN THỨC ĂN VÀ THỨC ĂN CÔNG NGHIỆP
11.1. CHẾ BIẾN THỨC ĂN HẠT
11.1.1. Tính chất vật lý, hóa học của tinh bột Tinh bột là polysaccharide dự trữ trong hạt, củ, quả. Đại phân tử tinh bột có hai cấu tử là
amylose và amylopectin. Amylose là một polyme mạch thẳng do các phân tử α-glucose liên kết với nhau theo kiểu 1,4-α-glucoside. Thông thường, tinh bột hạt ngũ cốc chứa 20 – 30% amylose (gạo ngon chứa 25 – 30% amylose, tỷ lệ này cao hơn hay thấp hơn đều làm thay đổi độ ngon). Amylopectin là một polyme mạch nhánh do các glucose liên kết với nhau qua mạch 1,4 và 1,6-α-glucoside. Trong lượng phân tử của amylopectin là hàng chục triệu, trong khi đó amylose chỉ khoảng 106.
Cấu trúc hạt tinh bột khá đặc biệt, mỗi hạt có một rốn hạt (hilum), xung quanh rốn hạt là các vòng đồng tâm (còn gọi là vòng sinh trưởng). Các hạt tinh bột ngũ cốc thường có các vết nứt hình thành do sự mất nước nhanh của ngũ cốc trong quá trình thành thục. Chính nhờ những vết nứt này mà enzym tiêu hóa dễ thâm nhập tạo điều kiện dễ dàng cho sự phân giải. Các hạt tinh bột các loại củ không có vết nứt này cho nên khó tiêu hóa hơn. Tinh bột có cấu trúc tinh thể, dưới tác dụng của nhiệt hay axit, cấu trúc tinh thể bị phá vỡ 11.1.2. Biến đổi vật lý, hóa học của tinh bột trong quá trình chế biến
Sự gelatin hóa. Dưới tác dụng của cơ, nhiệt hoặc hóa chất, liên kết hydro giữa các đại phân tử amylose và amylopectin bị phá vỡ, cấu trúc tự nhiên của hạt bị biến đổi. Khi tinh bột được ngâm trong nước và nhiệt độ nước tăng dần lên tới 550C, các hạt tinh bột hút nước và trương phồng lên. Sự trương phồng này là quá trình thuận nghịch, sau khi làm lạnh và khô, hạt trở lại bình thường. Tuy nhiên, nếu ngâm nước và đun nóng ở nhiệt độ cao hơn (60 - 800C), các hạt tinh bột mất đi cấu trúc tinh thể. Nhiệt độ đun nóng càng cao, càng kéo dài và có sự rung động mạnh thì cấu trúc tinh thể cũng bị phá vỡ nhiều.
Đây là quá trình gelatin hóa. Nhiệt độ gelatin hóa khác nhau phụ thuộc vào nguồn gốc tinh bột: tinh bột đại mạch 59 - 640C, tinh bột ngô 62 - 720C, tinh bột lúa mỳ 65 - 670C, tinh bột cao lương 67 - 770C. Gelatin hóa có thể xuất hiện khi nghiền hoặc cán mỏng các hạt ngũ cốc, một số dung dịch kiềm hay axit cũng có tác dụng thúc đẩy quá trình gelatin hóa.
Sự rắn đanh (retrogradation) và dextrin hóa. Rắn đanh là quá trình trong đó các hạt tinh bột từ trạng thái trương phồng hoặc gelatin hóa trở về trạng thái quần tụ thành từng đám và không hòa tan. Kết quả của quá trình này là liên kết hydro giữa amylopectin được phục hồi.
Mức độ rắn đanh phụ thuộc vào bản chất, vào hàm lượng nước tự do, nhiệt độ. Sự rắn đanh của tinh bột làm giảm hiệu quả tác động của enzyme, do đó là giảm tỷ lệ tiêu hóa.
Dextrin hóa là quá trình cắt phân đoạn các cấu tử amylose và amylopectin. Dextrin hóa có thể thực hiện bằng phương pháp xử lý tinh bột bằng nhiệt độ và độ ẩm, bằng dung dịch axit hoặc muối. Xử lý bằng tia hồng ngoại (micronizzing), nổ bỏng (poping) là các phương pháp dextrin hóa điển hình, tỷ lệ tiêu hóa tinh bột tăng lên rõ rệt.
Tác động của enzyme amylase lên tinh bột. Các α-amylase có thể gắn vào một điểm bất kỳ bên trong của chuỗi tinh bột mạch thẳng và bẻ gẫy chúng thành các đoạn có kích thước nhỏ hơn, chuyển chúng thành đường maltose và dextrin mạch thẳng hoặc mạch nhánh.
Các β-amylase gắn vào các mạch α.1,4 và α.1,6-glucoside ở phần ngoại vi của đại phân tử tinh bột, thủy phân tinh bột thành maltose và glucose. Đối với các cấu tử amylopectin, α-

103
amylase đầu tiên tác động vào các liên kết gần các điểm phân nhánh và bẻ gẫy chúng thành các oligosaccharide có liên kết phân nhánh. Về đại thể các amylase tác động lên bề mặt hạt tinh bột, trước hết ở những chỗ gãy hay những vùng không hoàn hảo về cấu trúc, sau đó lan tỏa ra các vùng xung quanh, tạo thành các hốc hình nón, xói mòn các hạt và làm cho chúng hòa tan hoàn toàn. 11.1.3. Kỹ thuật chế biến
Thức ăn hạt thường được chế biến theo các kỹ thuật sau (bảng 11.1).
Bảng 11.1. Tóm tắt các phương pháp chế biến thức ăn hạt
Chế biến khô Chế biến ướt Khô lạnh Khô nóng Ướt lạnh Ướt nóng
Nghiền búa Nghiền trục lăn
Nổ bỏng (Poping) Xử lý tia hồng ngoại (Micronizing) Rang chín (Roasting) Ép đùn (Dry extruding)
Ngâm ướt (Soaking) ủ hạt ướt Xử lý kiềm (Alkali treatment) Xử lý axit (Acid treatment)
Hấp cám (Steam rollring) Hấp và làm vỡ (Steam flaking) Làm giãn nở (Expanding) Nấu chín (Cooking)
Các phương pháp chế biến khô. Nghiền bằng búa: trong máy nghiền, hạt được đập vỡ
bằng hệ thống búa đập. Độ nhỏ của hạt phụ thuộc vào loại hạt, độ ẩm của hạt, kích cỡ mặt sàng, tốc độ dòng hạt lưu chuyển.
Nghiền bằng trục lăn: hạt được làm vỡ, bị cán mỏng và nghiền nhỏ bởi các trục lăn trong máy nghiền. Độ nhỏ của hạt phụ thuộc vào kích cỡ và cấu trúc, tốc độ của vòng quay của con lăn và các yếu tố khác như loại hạt, độ ẩm của hạt.
Nổ bỏng (popping): đây là phương pháp làm giãn nở và phá vỡ các hạt bằng nhiệt độ và áp suất cao. Ngô, gạo, cao lương, lúa mì có thể áp dụng nổ bỏng nhưng đại mạch, yến mạch không thực hiện được. Nhiệt độ nổ bỏng thường là 150oC, nhưng mức độ bung nổ khác nhau nhiều tùy theo loại hạt và độ ẩm của hạt.
Phương pháp dùng sóng cực ngắn (micronizing): nhờ tác động của vi sóng, nhiệt độ của hạt tăng nhanh trong khoảng 140-180oC với thời gian vài chục giây thùy theo loại hạt, tinh bột hạt được được gelatin hóa, vitamin được bảo toàn.
Phương pháp rang chín: hạt được quay trong một khoang kim loại chuyển động theo chu kỳ. Nhiệt độ của hạt trong quá trình rang đạt khoảng 150oC.
Ép đùn (extruding): hạt được ép qua một syranh trơn, bên trong là một trục có rãnh xoắn. Lực ma sát tạo ra nhiệt độ khoảng 95oC. Tinh bột được gelatin hóa và chất kháng dinh dưỡng cũng bị phá hủy, các chất dinh dưỡng được bảo toàn.
Các phương pháp chế biến ướt. Ngâm nước: hạt được ngâm nước trong khoảng thời gian 12-24 giờ, hạt ngấm một lượng nước.
Phương pháp ủ hạt ướt: hạt được ngâm nước đạt hàm lượng nước 25-30% sau đó đem ủ yếm khí 20 ngày. Trong quá trình ủ, các enzym có sẵn trong hạt sẽ tác động đến tinh bột. Chất lượng của sản phẩm chế biến sẽ phụ thuộc vào loại hạt, nhiệt độ môi trường và tỷ lệ nước trong hạt.

104
Xử lý kiềm (alkali treatment): hạt được ngâm hoặc phun bằng dung dịch xút nồng độ 2,5-4% phụ thuộc vào loại hạt. Cũng có thể sử dụng dung dịch amoniac, tuy nhiên phương pháp này đóng vai trò bảo quản nhiều hơn là làm thay đổi tính chất vật lý, hóa học của hạt.
Xử lý axit (acid treatment): xử lý axit thường áp dụng cho hạt cốc tươi, axit được dùng là axetic, propionic, izobutyric, formic, benzoic nhưng phổ biến là axit axetic hoặc propionic hoặc hỗn hợp của hai axit này. Tùy theo độ ẩm của hạt mà tỷ lệ axit được dùng từ 0,5-3% tính theo khối lượng hạt, độ ẩm càng cao lượng axit càng nhiều. Ưu điểm của phương pháp xử lý axit là không cần có hầm ủ kín ( hạt đã ngấm axit có thể bảo quản trong túi polyetylen, trong thùng gỗ ), thời gian bảo quản có thể kéo dài tới một năm. Tác dụng bảo quản của hạt vẫn còn duy trì khi đưa hạt ra khỏi nơi bảo quản.
Phương pháp hấp cán (steam rolling): trước hết hạt phải chịu tác động bởi hơi nước nóng trong khoảng thời gian từ 3-5 phút, sau đó hạt được nghiền bằng trục lăn.
Hấp và làm vỡ (steam flaking): theo phương pháp này hạt được phun một lượng hơi nước nóng trong khoảng thời gian sao cho độ ẩm của hạt nâng lên 18% (với ngô mất 12 phút, với hạt cao lượng mất 25 phút). Nhiệt độ hạt đạt được khoảng 100oC khi đưa vào trục lăn.
Hấp chín áp suất cao (pressure cooking): hạt được ấp chín ở nhiệt độ 143oC và áp suất 3kg/cm2. Sau khi hấp chín nguyên liệu được làm mát cho đến khi nhiệt độ còn 90oC và hàm ẩm giảm còn 20% trước khi cán và nghiền bằng trục lăn.
Phương pháp làm giãn nở (expanding): theo phương pháp này hạt được hấp chín trong điều kiện có hơi nước ở áp suất và nhiệt độ cao (15kg/cm2, 200oC trong 20 giây). Dưới áp suất và nhiệt độ cao, hạt bị trương phồng sau đó giãn nở đến mức tối đa.
Ép viên (pelleting): thông thường người ta đưa hơi nước nóng vào khối nguyên liệu để đưa nhiệt độ lên khoảng 60-94oC, ở nhiệt độ này một phần tinh bột được gelatin hóa. Nhờ sức ép của trục lăn trên syranh, viên thức ăn được hình thành khi chui qua các rãnh của thành syranh.
11.2. THỨC ĂN HỖN HỢP VÀ CÔNG NGHỆ THỨC ĂN HỖN HỢP
11.2.1. Phân loại thức ăn công nghiệp Định nghĩa: Thức ăn công nghiệp là một hỗn hợp thức ăn bao gồm một số nguyên liệu
đã qua chế biến và phối hợp theo công thức của nhà chế tạo, đáp ứng nhu cầu dinh dưỡng của tôm và cá.
Phân loại thức ăn hỗn hợp: thức ăn hỗn hợp có 2 loại. Hỗn hợp hoàn chỉnh: đầy đủ các chất dinh dưỡng, khi cho ăn không phải bổ sung thêm
bất cứ một chất dinh dưỡng nào khác (dùng trong hệ thống nuôi công nghiệp). Hỗn hợp bổ sung: bổ sung thêm một số chất dinh dưỡng cần thiết như vitamin, chất
khoáng… Các dạng thức ăn hỗn hợp: Có nhiều dạng thức ăn công nghiệp cho tôm và cá như dạng khô, dạng ướt; trong đó lại
chia làm dạng bột hay viên, dạng mảnh hay hạt.., mỗi dạng đòi hỏi một công nghệ sản xuất riêng (sơ đồ 11.1). 11.2.2. Tiêu chuẩn chất lượng của thức ăn hỗn hợp
Độ ẩm. Hàm lượng ẩm của TACN (dạng khô) rất quan trọng vì độ ẩm của thức ăn có quan hệ đến sự phát triền của vi sinh vật từ đó ảnh hưởng đến thời hạn sử dụng. Bảng 11.2 sau đây cho biết mối quan hệ giữa hàm ẩm của thức ăn đến sự phát triển của vi sinh vật và côn trùng.

105
Bảng 11.2. Mối quan hệ giữa hàm ẩm của thức ăn với sự phát triển của côn trùng và vi sinh vật
Độ thức ăn (%)
Độ ẩm không khí (RH%)
Hậu quả
0 – 8 0 – 30 Không có hoạt động sinh học 9 -14 30 –70 Nhiễm côn trùng khi RH>60% 14 – 20 70 -90 Nhiễm côn trùng và mốc 20 – 25 90 – 95 Mốc và vi khuẩn phát triển >25 >95 Vi khuẩn tăng và nẩy mầm Kích thước. Kích thước tùy thuộc vào loài và giai đoạn sinh trưởng của tôm cá (bảng 11.3)
Bảng 11.3. Kích thước viên của thức ăn công nghiệp cho cá
Cá Rô phi Cá da trơn Kich cỡ/tuổi Kích cỡ viên Kích cỡ (g) Kích cỡ viên (mm)
Hương 1-24 giờ Hương 2-10 ngày Hương 10-30 ngày Juvenille 1 –30 g 20 – 120 g 125 – 25 g >250 g
liquify 500 microns 500 –1000 microns 500 –1500 microns 1 – 2mm 2 mm 3 mm 4 mm
0,02 – 0,25 0,25 – 1,50 1,50 – 5,00 5,0 – 20,0
0,4 – 0,8 0,8 – 1,4 1,4 – 2,8 2,8 – 4,0
Tiêu chuẩn cảm quan. Thức ăn phải có mầu đặc trưng của nguyên liệu trong hỗn hợp có
mùi thơm, không có mùi mốc hoặc hôi thối… Tiêu chuẩn dinh dưỡng. Tiêu chuẩn này tùy thuộc vào loài và giai đoạn sinh trưởng của
tôm cá. Các tiêu chuẩn thường được quy định là năng lượng (DE hoặc ME tính theo MJ hay Mcal/kg hỗn hợp), tỷ lệ P/E, protein và một số axit amin, chất béo và axit béo n-3, một số chất khoáng (Ca, P…), một số vitamin ( các chất này được tính theo % hay g, mg/kg hỗn hợp). 11.2.3. Các quy định pháp lý đối với thức ăn hỗn hợp
Thức hỗn hợp là một loại hàng hóa cho nên phải tuân thủ những quy định pháp luật của loại hàng hóa này, những quy định này phải được thể hiện trên nhãn hàng.
Những quy định này gồm: - Hàm lượng ẩm: đối với hỗn hợp khô, hàm ẩm được quy định là <14%. - Hàm lượng tối thiểu chất dinh dưỡng (đối với cá chất dinh dưỡng quý như protein, axit
amin, chất béo, Ca, P…) và hàm lượng tối đa các chất dinh dưỡng (đối với các chất dinh dưỡng gây ảnh hưởng xấu đến sinh trưởng và hiệu suất sử dụng thức ăn của động vật như tro, cát sạn (khoáng không hòa tan trong axit HCl), chất xơ.
- Các chỉ tiêu vệ sinh an toàn: bao gồm các chỉ tiêu vi sinh vật gây bệnh và nấm mốc độc hại (như aflatoxin…), các chất hóa chất độc hại (như kim loại nặng Pb, Hg, Cd…), các thuốc (như kháng sinh…).
- Các quy định về nhãn hàng: nhãn hàng phải ghi tên thương phẩm (đối với những hỗn hợp có bổ sung thuốc, một số nước bắt buộc phải ghi trên nhãn hàng chữ “có sử dụng thuốc” (MEDIATED), đối tượng sử dụng, các phân tích đảm bảo (guaranteed analysis), các nguyên liệu trong hỗn hợp (chỉ ghi tên nguyên liệu không cần khối lượng hay tỷ lệ), hướng dẫn sử dụng, thời hạn sử dụng, tên và địa chỉ nhà sản xuất, khối lượng tịnh.

106
11.2.4. Công nghệ thức ăn hỗn hợp Công nghệ chế biến thức
ăn hỗn hợp xuất hiện từ những năm 50, ngày nay công nghệ này ngày càng hiện đại. ở giai đoạn đầu, thức ăn hỗn hợp chỉ được trộn bằng tay trên sàn kho, rồi tiến tới trộn cơ khí, trộn liên tục và bây giờ máy tính đã kiểm soát việc trộn và viên. Các công đoạn của chế biến thức ăn hỗn hợp gồm : nhập kho, cân, nghiền, trộn, viên, làm nguội, bao gói, bảo quản (sơ đồ 11.1).
Không đi sâu vào các công đoạn trên, ở đây chỉ nhấn mạnh những điểm sau:
(1). Cân nguyên liệu: Để phối hợp đúng công thức, việc cân chính xác nguyên liệu là rất quan trọng, có một số nguyên liệu cần phải nghiền trước khi cân, cân thức ăn bổ sung phải chú ý vì khối lượng nhỏ và đắt tiền.
(2).Nghiền (hình 11.1): Đây là công đoạn tốn nhiều năng lượng nhất. Chi phí năng lượng cho việc nghiền thức ăn thuỷ sản thường gấp 5-6 lần so chi phí năng lượng cho việc nghiền nguyên liệu làm thức ăn cho động vật trên cạn. (Ví dụ: máy nghiền công suất 22 KW/giờ, nếu ngô đem nghiền mịn, mỗi giờ nghiền được 200kg, còn nghiền thô thì được tới 2000kg).
Kích thước nghiền phụ thuộc vào công thức ăn cho từng loại cá, nghiền nhỏ giúp dễ trộn và tăng tỷ lệ tiêu hoá. Tuy nhiên nghiền quá nhỏ thì không tốt vì bề mặt tiếp xúc tăng, khó bảo quản, tỷ lệ tiêu hoá có thể giảm. Nghiền làm cho nhiệt độ nguyên liệu tăng lên ( 10 – 200C) do vậy máy nghiền phải có thiết bị làm mát (quạt, …). Thường trước khi nghiền người ta phải loại bỏ kim loại và những tạp chất khác.
(3). Trộn: Trộn là một “ nghệ thuật”, chứ không phải “kỹ thuật”. Mục đích của trộn là làm cho hỗn hợp đồng nhất. Đối với thức ăn cá, việc trộn đều càng cần thiết vì hàng ngày cá ăn ít, trộn không đều làm cho lượng thức ăn ăn vào biến động. Đối với quy mô nhỏ, để trộn đều người ta đưa thêm chất màu vào để làm chất chỉ thị. Thức ăn
bổ sung khi đưa vào hỗn hợp phải trộn trước ( ở dạng premix) và đảm bảo khối lượng không dưới 1 kg tính cho 100 kg hỗn hợp.
Máy trộn có hai loại (hình 11.2):

107
+ Máy trộn đứng (vertical mixers) : Trộn bằng vít xoắn ở tốc độ 100 – 200 vòng/phút, thời gian trộn kéo dài 10 – 15 phút. Máy trộn đứng khi phải trộn thức ăn cùng với chất lỏng thì ít hiệu quả vì chất lỏng có khuynh hướng tạo thành những cục nhỏ chứ không bám đều vào
nguyên liệu khô. + Máy trộn nằm ngang
(horizontal mixers) : Thời gian trộn ngắn hơn ( 3 – 6 phút), máy này cũng thích hợp với việc trộn 8% chất lỏng trong hỗn hợp, máy này đắt hơn máy trộn đứng.
(4). Viên (hình 11.3): Viên nguội thức ăn đã trộn, đưa vào máy dập viên ở nhiệt độ trong phòng. Đưa độ ẩm của khối thức ăn lên 15 – 16%. Viên thực hiện bằng cách ép đùn cho nên trong quá trình viên nhiệt độ tăng lên 60 – 700C, sau đó viên được làm khô và làm nguội đến nhiệt độ trong phòng.
Viên nóng (conditioner pelleting) : Hỗn hợp thức ăn được làm nóng bằng hơi nước khô nhờ một bộ phận tạo hơi nước nóng. Hỗn hợp thức ăn được ép bằng máy dập khuôn để tạo viên, sau đó viên được làm khô và làm nguội.
Trong công nghiệp thức ăn hỗn hợp cần chú ý đến chất lượng viên thức ăn. Chất lượng viên (chỉ xét về khía cạnh vật lý) thể hiện ở độ cứng, độ bóng, độ gelatin hoá, độ chín và độ bền với nước khi sử dụng.
Chất lượng viên phụ thuộc vào nguyên liệu đưa vào ép viên, thiết bị và công nghệ viên.
Về nguyên liệu. - Khối lượng riêng, kết cấu và thành phần hoá học nguyên liệu. Hỗn hợp nguyên liệu có
khối lượng riêng (klr) 0,4 g/cc khi được ép thành viên có klr 0,5-0,6 g/cc (với khuôn vòng, áp xuất viên là 75-600 kg/cm2).
- Thức ăn nhiều xơ viên cứng hơn thức ăn ít xơ. - Thức ăn nhiều mỡ (>8-10%), nhiều nước (>15%) làm giảm chất lượng viên (giảm độ
cứng). Về công nghệ. Quan hệ giữa đường kính x độ dài khuôn viên; tốc độ quay của khuôn, tốc
độ thức ăn đa vào buồng nguyên liệu; lượng hơi nước vào khuôn; độ ẩm không khí đều có ảnh hưởng đến chất lượng viên thức ăn.
(5).Đóng bao: Thức ăn hỗn hợp rời hay viên được bao gói bằng túi polyetylen, bên ngoài bằng bao dứa. Dự trữ nơi khô ráo, thoáng mát, tránh chuột bọ xâm hại.

108
11.2.5. Những thiết bị cần thiết của một nhà máy thức ăn hỗn hợp cho tôm và cá Giới thiệu những thiết bị cần thiết để sản xuất các loại thức ăn hỗn hợp khác nhau cho
tôm và cá (bảng 11.4 và sơ đồ 11.1. và 11.2).
Bảng 11.4. Các thiết bị cần thiết để sản xuất thức ăn hỗn hợp khô cho tôm và cá
Bột
Viên nổi Viên chìm Hạt
Máy nghiền (grinder) Máy trộn khô (dry mixer) Máy nâng (elevator) Băng tải (conveyor) Máy ép viên (pelleter/dies) Nồi nấu hay máy ép đùn (cooker/extruder) Surge bins Máy làm nguội/làm khô (cooler/dryer) Máy phun mỡ (fat sprayer) Nồi hơi (steam boiler) Máy đập mảnh (crumbler) Máy nâng (sifter) Máy khâu bao (bag sewer) Cân (scales)
+(*) + ? ? - - - - - - - - + +
+ + + + - +
+ + ? + - + + +
+ + ? ? + - + + -
+ - + +
+
+ + ? ? + - + + ?
+
+ + + +
*(+): cần; (-): không cần; (?): có thể cần
Câu hỏi: 1. Những biến đổi vật lý, hoá học của tinh bột khi chế biến? 2. Kỹ thuật chế biến thức ăn hạt? 3. Cho biết đặc điểm các loại và các dạng thức ăn hỗn hợp cho tôm và cá? 4. Các khâu chính trong công nghệ sản xuất thức ăn hỗn hợp cho tôm và cá?

109
CHƯƠNG XII TIÊU CHUẨN ĂN VÀ KHẨU PHẦN
Mỗi loài động vật khác nhau yêu cầu khác nhau về các chất dinh dưỡng, tuỳ theo đặc điểm riêng của từng loài, giống và khả năng sản xuất của chúng. Trong từng loài, giống tuỳ theo giai đoạn phát triển mà nhu cầu dinh dưỡng khác nhau. Cân đối các chất dinh dưỡng cần thiết trong khẩu phần so với nhu cầu của động vật là biện pháp rất quan trọng nhằm tăng năng suất và hiệu quả trong nuôi trồng thủy sản.
12.1. KHÁI NIỆM
12.1.1. Tiêu chuẩn ăn Tiêu chuẩn ăn được xác định dựa trên nhu cầu các chất dinh dưỡng (phần này đã đề cập
trong các chương trước). Như đã biết, nhu cầu dinh dưỡng là khối lượng chất dinh dưỡng mà tôm, cá cần để duy trì hoạt động sống và tạo sản phẩm (tăng trọng, đẻ trứng..) trong ngày đêm.
Tiêu chuẩn được xây dựng trên cơ sở nhu cầu. Vì vậy, có thể khái niệm tiêu chuẩn ăn là khối lượng các chất dinh dưỡng (được tính bằng đơn vị khối lượng hoặc tính bằng phần trăm trong thức ăn hỗn hợp) mà tôm, cá yêu cầu trong một ngày đêm. Tiêu chuẩn ăn có thể hiểu như sau:
Tiêu chuẩn ăn = Nhu cầu + Số dư an toàn Số dư an toàn là số lượng
chất dinh dưỡng cần thêm vào ngoài nhu cầu của tôm, cá được xác định thông qua các thực nghiệm. Trong thực tế, xác định nhu cầu dinh dưỡng được tiến hành trong phòng thí nghiệm (on-station) với nhiều cá thể và giá trị thu được là trung bình số học của các quan sát. Giá trị về nhu cầu dinh dưỡng (ví dụ: 10 MJ ME) là giá trị trung bình của các giá trị thu được trên hoặc dưới giá trị trung bình nói trên (có thể 9-11 MJ ME). Có nghĩa, nếu áp dụng giá trị trung bình trên để xác định nhu cầu thì một số cá thể không đáp ứng nhu cầu chất dinh dưỡng (những quan sát trên 10 MJ ME). Do đó, người ta mới sử dụng khái niệm số dư an toàn.
Tiêu chuẩn ăn được qui định bởi một số chỉ tiêu cơ bản, những chỉ tiêu này phụ thuộc vào sự phát triển nuôi trồng thủy sản của mỗi nước.
Ví dụ 1: Nhu cầu dinh dưỡng của cá chép:
Lipit: 18% trở lên Protein: 25-38% Lisine: 5,7% của thức ăn Methionine: 3,1% của protein (không có cystine) Photpho hữu dụng: 0,6-0,7%. DE kcal/kg: 2700-3100.
Nhu cầu của cá rôphi (Jaucey, Ross, 1982): Lipit: 10% (từ hương đến 0,5g)
8% (từ 0,5 đến 35g) 6% (từ 35g đến bán)
Protein: 50% (từ hương đến 0,5g) 35% (từ 0,5 đến 35g)
30% (từ 35 đến bán) Lysine: 4,1% protein thức ăn Methionine + 50% Cystine: 1,7% protein thức ăn Carbohydrate tiêu hoá: 25% Xơ thô: 8 – 10% DE (kcal/kg): 2500 – 3400

110
12.1.2. Nội dung tiêu chuẩn ăn - Nhu cầu năng lượng (ví dụ 1): Biểu thị bằng kcal (Mcal) hay kJ (MJ) của DE, ME tính
cho một ngày đêm hay tính cho 1 kg thức ăn. Khi nhu cầu năng lượng tính trên 1 kg thì gọi là mật độ năng lượng hay mức năng lượng.
- Nhu cầu protein và axít amin (ví dụ 1): Nhu cầu protein có thể thể hiện bằng khối lượng (g; kg) cho một ngày đêm hay tỷ lệ (%) protein thô hay protein tiêu hóa trong khẩu phần. Axit amin cũng được tính theo khối lượng (g) cho một ngày đêm hay tỷ lệ (%) so với vật chất khô hoặc tỷ lệ (%) so với protein.
- Nhu cầu mỡ và axit béo (ví dụ 1): Nhiều nước đã sử dụng các axit béo thiết yếu trong tiêu chuẩn ăn của vật nuôi (Anh, Mỹ, Australia..).
- Nhu cầu các chất khoáng (ví dụ 1): + Khoáng đa lượng: Ca, P, Mg, Na, Cl ,K, S (g/con ngày hoặc % thức ăn). + Khoáng vi lượng: Fe, Cu, Co, Mn, Zn.. (mg/con ngày). - Nhu cầu vitamin: A, D, E (UI), VTM nhóm B, C, K (mg); B12 (µg).
12.1.3. Khẩu phần ăn Để hiển thị tiêu chuẩn ăn bằng các loại thức ăn cụ thể thì người ta sử dụng khái niệm
“khẩu phần ăn”. Khẩu phần ăn là khối lượng các loại thức ăn cung cấp cho con vật để thoả mãn tiêu chuẩn ăn. Khẩu phần ăn được tính bằng khối lượng trong một ngày đêm hoặc tỷ lệ phần trăm trong thức ăn hỗn hợp.
Ví dụ, để đảm bảo tiêu chuẩn ăn cho cá da trơn với yêu cầu là protein thô 34,5; lipit 10,8; khoáng 11,9; Ca 1,0 và P 1,2% thì khẩu phần như ở ví dụ 2.
12.2. NGUYÊN TẮC PHỐI HỢP KHẨU PHẦN ĂN
Tối ưu hoá khẩu phần ăn hay còn gọi là lập khẩu phần ăn để thoả mãn nhu cầu dinh dưỡng của gia súc gia cầm với giá thành thấp nhất là rất quan trọng để nâng cao hiệu quả kinh tế trong chăn nuôi. Có hai nguyên tắc để lập khẩu phần ăn là nguyên tắc khoa học và nguyên tắc kinh tế. 12.2.1. Nguyên tắc khoa học
Khẩu phần ăn phải đáp ứng đầy đủ nhu cầu dinh dưỡng, thoã mãn được tiêu chuẩn ăn. Đảm bảo được sự cân bằng các chất dinh dưỡng: axit amin, khoáng, vitamin...
Khối lượng khẩu phần ăn phải thích hợp với sức chứa của bộ máy tiêu hoá. Để khống chế khối lượng khẩu phần ăn người ta dùng lượng thức ăn (% vật chất khô) có
thể thu nhận tính theo tỷ lệ khối lượng cơ thể. Ví dụ, khối lượng thức ăn nuôi tôm khoảng 2-3% khối lượng tôm.
Ví dụ 2. Khẩu phần cho cá da trơn kg
Bột cá thương phẩm Khô đỗ tương Khô lạc Bột mì Mỡ lợn Dicanxiphotphat Muối ăn Chất kết dính (Cacboxymethyl celulose) Premix vitamin Nước Tổng cộng
15,17 8,50 8,50 18,70 5,11 1,04 2,00 2,00
0,92 38,56 100,00

111
12.2.2. Nguyên tắc kinh tế Khẩu phần thức ăn phải có giá cả hợp lý và rẻ. Để khẩu phần thức ăn vừa đảm bảo nhu
cầu dinh dưỡng cho tôm, cá và vừa tối ưu về mặt kinh tế cho người nuôi, khi lập khẩu phần ăn phải chú ý các vấn đề sau đây:
+ Tính sẵn có, chất lượng và giá cả của nguồn nguyên liệu thức ăn + Đặc tính sinh học của tôm cá và cách cho ăn + Mục tiêu nuôi (thịt, làm giống...) + Đặc điểm cơ bản của hệ thống nuôi dưỡng, ăn tự do hay hạn chế + Nhiệt độ, độ ẩm của môi trường...
12.3. PHƯƠNG PHÁP PHỐI HỢP KHẨU PHẦN ĂN CHO TÔM, CÁ Muốn phối hợp khẩu phần ăn cho tôm, cá một cách khoa học và hợp lý chúng ta cần
biết: - Tiêu chuẩn ăn đối với các chất dinh dưỡng như: năng lượng, protein, axit amin, hàm
lượng lipit, vitamin, khoáng... - Thành phần hoá học và giá trị dinh dưỡng, giá cả của các loại thức ăn dự kiến sẽ sử
dụng trong khẩu phần ăn (chú ý giới hạn tốt đa % của từng loại nguyên liệu). - Giá cả của các nguyên liệu làm thức ăn có thể tính cho 1 kg hay cho 1.000 kcal năng
lượng (tiêu hoá hay trao đổi) và 100 gam protein thô trong thức ăn.
Bảng 12.1. Tính toán giá trị năng lượng tiêu hoá (DE) cho cá
Chất dinh dưỡng Năng lượng thô (GE, kcal/g)
Năng lượng tiêu hoá (DE, kcal/g)
Carbohydrate (không phải cây bộ đậu) Carbohydrate (cây bộ đậu) Protein (động vật) Protein (thực vật) Chất béo
4,1 4,1 5,5 5,5 9,1
3,0 2,0 4,25 3,8 8,0
Có thể sử dụng bảng “Thành phần dinh dưỡng thức ăn cho tôm, cá” của ADCP
(Aquaculture Developpement and Coordination Programe - FAO) ở phần phụ lục. Trong bảng có ghi thành phần protein thô, chất béo, xơ thô, khoáng, chất chiết không N, Ca, P, Met + Cys, lysine (tính trên cơ sở chất khô của thức ăn) và năng lượng tiêu hoá (DE, kcal/kg). DE của thức ăn cho tôm, cá tính trên cơ sở giá trị DE của 1g các chất dinh dưỡng trong thức ăn theo khuyến cáo của Michael B. New (1987, bảng12.1). 12.3.1. Phương pháp phối hợp thông thường
Phương pháp này áp dụng tính toán cho các khẩu phần thức ăn chỉ bao gồm một vài nguyên liệu và yêu cầu tính một vài chất dinh dưỡng chủ yếu trong khẩu phần. Các phương pháp kinh điển được sử dụng để xây dựng khẩu phần như: Phương pháp thử - sai (trial - error), phương pháp hình vuông Pearson, phương pháp lập phương trình đại số…
Nhìn chung, các phương pháp có các bước như sau: Bước 1: Xác định nhu cầu dinh dưỡng, tiêu chuẩn cho tôm, cá. Nhu cầu dinh dưỡng theo tiêu
chuẩn Việt nam (TCVN) hoặc tham khảo tiêu chuẩn NRC (Mỹ), ARC (Anh), tiêu chuẩn của Nhật Bản, Hà Lan, Ân Độ.. phù hợp với khí hậu và các vùng sinh thái khác nhau, và môi trường nuôi trồng thủy sản.

112
Bước 2: Chọn lựa các nguyên liệu thức ăn để lập khẩu phần ăn. Phải biết giá trị dinh dưỡng và
giá thành các nguyên liệu thức ăn đó. Nguyên liệu thức ăn phải bảo đảm chất lượng tốt và phải phù hợp với tôm, cá. Nói chung, nguồn thức ăn cho thủy sản phải đảm bảo hàm lượng protein cao.
Bước 3: Tiến hành lập khẩu phần. Phương pháp này thường theo các bước chính sau đây: - Xác định khối lượng các loại thức ăn bổ sung như khoáng vi lượng , premix vitamin...
Các loại thức ăn này thường chiếm tỷ lệ thấp trong khẩu phần (bảng 10…). - Ấn định khối lượng một số loại thức ăn giàu protein hoặc thức ăn giàu năng lượng
(tham khảo khuyến cáo ở trên). - Trên cơ sở thức ăn đã ấn định tính toán khối lượng các loại thức ăn còn lại. Ta có thể
xác định khối lượng của từng loại thức ăn này bằng 2 phương pháp: phương pháp Pearson hoặc phương pháp dùng phương trình đại số.
- Tính toán giá trị dinh dưỡng của khẩu phần dự kiến. - Điều chỉnh và bổ sung. Dựa vào tiêu chuẩn ăn để điều chỉnh và bổ sung các chất dinh
dưỡng cho phù hợp với nhu cầu của tôm, cá. 12.3.2. Phương pháp hình vuông Pearson
Ví dụ 1: hãy phối hợp 1 hỗn hợp có 25% protein cho cá lóc, gồm khô dầu dừa (chứa 20% protein) và bột cá (chứa 60% protein).
Khô dầu dừa 20 35 (35/40) x 100 = 87,5/100
25
Bột cá 60 5 (5/40) x 100 = 12,5/100
Như vậy, cần phối hợp 12,5 kg bột cá và 87,5 kg khô dầu dừa, chúng ta sẽ có một hỗn hợp chứa khoảng 25% protein thô. 12.3.3. Phương pháp giải phương trình
Ví dụ 2: Lấy lại thí dụ 1, gọi x là kg khô dầu dừa và y là kg bột cá trong 100 kg thức ăn, ta có phương trình 2 ẩn số:
0,20x + 0,60y = 25 (1) x + y = 100 (2) Giải phương trình trên ta được x = 87,5 kg và y = 12,5 kg. Ví dụ 3: Phối hợp 1 khẩu phần có 35% protein thô (CP) bằng các nguyên liệu sau cho cá
rô phi: Bột cá 60%CP, cám gạo 10%CP, khô dầu đỗ tương 45%CP. Hỗn hợp cuối cùng chứa
protein động vật không quá 1/3 lượng protein khẩu phần. Hỗn hợp chứa 4% premix vitamin - khoáng, 4% mỡ và 2% chất kết dính.
Giải: Gọi x: kg bột cá trong 100kg thức ăn hỗn hợp y: kg cám trong 100kg thức ăn hỗn hợp z : kg đỗ tương trong 100kg thức ăn hỗn hợp Các chất bổ sung đã có trong hỗn hợp: 4 premix + 4% mỡ + 2% chất kết dính = 10% Giới hạn protein động vật: 0,60x = 35/3 = 11,67

113
Ta lập được phương trình: 0,6 x + 0,1y + 0,45z = 35 (1)
x + y + z = (100 - 10) (2)
0,6 x = 11,67 (3)
Từ (3) ta có x = 11,67/0,6 = 19,45
Thay giá trị của x vào (1) và (2), ta có phương trình: 0,1 y + 0,45 z = 23,43 (4)
y + z = 70,56 (5)
Giải phương trình ta được: x = 19,45 y = 42,03 z = 38,42 Tóm lại khẩu phần ăn chứa: Bột cá 19,45 kg Cám 42,03 kg Khô đỗ tương 38,42 kg Premix 4,00 kg Mỡ 4,00 kg Chất kết dính 2,00 kg 100,00 kg Ngoài các phương pháp trên, phương pháp phối hợp khẩu phần bằng các chương trình
phần mềm trên máy vi tính cũng đã được áp dụng nhiều. Tùy theo khả năng mà có thể sử dụng các phầm mềm khác nhau. phần mềm đơn giản nhất sử dụng chương trình trên phần mềm Excel để thiết lập bảng tính với những công thức thích hợp hoặc các phần mềm chuyên biệt như: UFFDA, Feedlive, Feedmania… Các phần mềm chuyên biệt ngoài việc tính toán công thức thức ăn còn có thể có chức năng so sánh giá trị dinh dưỡng của các nguồn nguyên liệu, tồn trữ các công thức, dữ liệu để giảm bớt thời gian tìm kiếm, tính toán
Câu hỏi 1. Tiêu chuẩn, khẩu phần ăn là gì? Cho ví dụ? 2. Cho biết nhu cầu dinh dướng của một số loài tôm và cá? 3. Cho biết các nguyên tắc và phương pháp phối hợp khẩu phần, cho ví dụ?

114
CHƯƠNG XIII DINH DƯỠNG VÀ NUÔI DƯỠNG MỘT SỐ ĐỐI TƯỢNG TÔM, CÁ
13.1. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ CHÉP (CIPRINUS CARPIO)
13.1.1. Giới thiệu Cá chép (Cyprinus carpio) thuộc họ Cyprinidae. Trong tự nhiên, cá chép sống ở vùng
trung và hạ lưu các con sông có dòng chảy chậm hoặc trong các đầm lầy. Môi trường sống của chúng thường là vùng nước nhiều cỏ dại và đáy bùn. Thức ăn của cá hương là động vật phù du như rotifer và copepods, nhưng khi trưởng thành thì chúng là những đối tượng ăn đáy, thức ăn của chúng là động vật và các loại sinh vật khác.
Những loài trong họ cá chép đã được nuôi từ rất lâu đời cho đến nay, đây là họ cá có xương quan trọng nhất của nghề nuôi cá nước ngọt trên toàn thế giới. Sản lượng hiện nay của cá chép đạt đến hơn 13 triệu tấn mỗi năm. Cá chép là một đối tượng nuôi quan trọng của họ cyprinids, chỉ sau cá mè trắng và cá trắm cỏ, sản lượng của nó đã tăng gấp đôi so với 1 thập niên trước đây, đạt khoảng 2,5 triệu tấn vào năm 1998, trị giá 2,8 tỷ USD. Một tỷ lệ lớn từ nguồn cá chép nuôi ở châu Á, đặc biệt là ở Trung Quốc. Trong khi hầu hết sản lượng cá chép từ hình thức nuôi tự nhiên, thì chỉ khoảng
Hình 13.1. Cá Chép (Cyprinus carpio) 3% trong họ cá chép được nuôi theo các hệ thống chuyên canh (Tacon, 1993).
Cá chép được nuôi truyền thống trong ao hoặc trong ruộng lúa, hoặc theo hình thức nuôi hiện đại trong ao chủ động nước, hệ thống nuôi nước chảy và nuôi lồng lưới trong hồ. Nhiều kỹ thuật nuôi đã được áp dụng trong nuôi vỗ cá bố mẹ, cá hương, cá giống và cá thương phẩm. Các giai đoạn, hình thức nuôi và cách cho ăn được trình bày bảng 13.1. Khẩu phần với giá thành thấp được sử dụng nhằm cung cấp các chất dinh dưỡng thích hợp từ các loại thức ăn sẵn có.
Bảng 13.1. Hình thức nuôi và khẩu phần thức ăn trong những giai đoạn phát triển khác nhau của cá chép (Cyprinus carpio) (Kafuku, 1992)
Giai đoạn Hình thức nuôi Thức ăn/cho ăn Cá bố mẹ Cá hương Cá giống Cá thương phẩm
Ao nuôi vỗ Ao không lưu thông nước Ao không lưu thông nước Ao nước chảy một phần Ao ruộng Ao ruộng, ao nước chảy, Ao lấy nước suối Ao tuần hoàn Lồng lưới
70% thức ăn xanh và 30% thức ăn động vật đã được làm giàu vitamin và khoáng Daphnia và động vật phù du khác Thức ăn tự chế biến Thức ăn viên, ăn hơn 5 lần/ngày Thức ăn tự chế biến, nhộng tằm Rau cỏ, lúa mì nấu chín

115
13.1.2. Nhu cầu dinh dưỡng Nhu cầu dinh dưỡng của cá chép trong khẩu phần về protein, amino acids, lipid, axit
béo, carbohydrates, vitamin, khoáng, năng lượng và protein/năng lượng đã được kiểm nghiệm bởi nhiều nhà khoa học và các nhà nghiên cứu (Satoh, 1991; Oma, 1992; Song, 1994; De Silva và Anderson, 1995; Kaushik, 1995; Takeuchi, 1999).
Protein và amino acid. Một số nghiên cứu cho biết nhu cầu protein hàng ngày của cá chép khoảng 1g/kg khối lượng cơ thể cho duy trì và 12g/kg khối lượng cơ thể cho tích lũy protein tối đa (Ogino và Chen, 1973; Ogino, 1980). Tuy nhiên, hiệu quả sử dụng N cho quá trình tăng trưởng cao nhất khi lượng protein ăn vào 7-8 g/kg khối lượng cơ thể/ ngày.
Nhu cầu tối ưu của cá chép về protein thô là: 30-38% (Jauncey, 1982; Wantanabe, 1988). Nhìn chung, mức protein thô này được xác định bởi việc sử dụng khẩu phần gồm một nguồn đơn protein có chất lượng cao, như casein, protein trứng gà hoặc bột cá. Nếu đủ năng lượng tiêu hóa có trong khẩu phần, mức protein tối ưu có thể giữ 30-35 % (Watanabe, 1982).
Bảng 13.2. Nhu cầu dinh dưỡng của cá chép (Ciprinus carpio)
Thành phần dinh dưỡng Nhu cầu Protein Lipit Acid béo no thiết yếu Linoleate Linolenate Năng lượng tiêu hóa Carbohydrate (như tinh bột)
30–35 g /100g 5-15 g /100g (tùy theo mức năng lượng) 1 g/100g 1 g/100 g 13-15 MJ/ kg (310-360 kcal) 30-40 g/100g
Amino acid cấu trúc cơ thể cá chép không bị ảnh hưởng bởi sự biến động các thành phần
khác nhau trong khẩu phần hoặc bởi tuổi của cá (Schwarz và Kirchgessner, 1988). Mười amino acid thiết yếu (EAA) tương tự cho hầu hết các loại cá cũng đều cần thiết cho quá trình sinh trưởng của cá chép. Nhu cầu về số lượng các amino acid được thiết lập thông qua các nghiên cứu khác nhau và được chỉ ra ở bảng 13.3. Cần chú ý là có những thay đổi nhỏ trong nhu cầu đối với từng amino acid tùy theo từng giai đoạn sinh trưởng (Baloguma, 1995). Nhu cầu lysine ở giai đoạn cá giống là 2,25% của khẩu phần (6% của protein) và giảm xuống 1,75% (5,4% của protein) ở giai đoạn thịt, còn đối với methionine thì không có sự thay đổi. Điều này cũng được nhận thấy ở các đối tượng khác, cysteine và tyrosine có thể dư hoặc lần lượt thay thế một phần của methionine và phenylalanine khẩu phần. Những nghiên cứu hiện nay về amino acid có chứa lưu huỳnh cho thấy rằng hoạt lực enzyme cysteinsulphinate decarboxylase (CSD) ở gan của cá chép khá yếu so với cá hồi nước ngọt (Oncorhynchus mykiss) và cá rô phi (Oreochromis niloticus) đến 50 lần (Yokoyama và cộng sự, 2001). Ta biết rằng cơ của cá chép có chứa một lượng lớn Taurine. CSD là một enzyme tham gia vào quá trình sinh tổng hợp Taurine từ cysteine và, nếu như hoạt động của nó yếu, có nghĩa là nhu cầu taurine của cá chép phải được cung cấp thêm trong khẩu phần.
Nhu cầu amino acid có thể ước tính từ số liệu về thành phần amino acid và nguồn protein tích lũy hàng ngày của cơ thể (Ogino, 1980). Nếu khẩu phần chứa 35% protein, tỷ lệ tiêu hóa (TLTH) protein là 80%, định mức cho ăn 3% khối lượng cơ thể, giả thiết rằng cá có thể tích lũy được 0,58g protein/100g khối lượng cơ thể mỗi ngày. Tuy nhiên, mức tích lũy này không
116
tính đến các đường chuyển hóa của amino acid không dẫn đến quá trình tổng hợp protein. Nhu cầu về acid amin của cá chép được trình bày ở bảng 13.3.
Bảng 13.3. Nhu cầu về amino acid của cá chép (Cyprinus carpio)
Nose (1979) Các amino acid % protein trong
khẩu phần % khẩu phần
Ogino (1980) % protein trong
khẩu phần
Dabrowski (1983) mg/kg/ngày
Arginine 4,3 1,6 4,4 506 Histidine 2,1 0,8 1,5 145 Isoleucine 2,5 0,9 2,6 255 Leucine 3,3 1,3 4,8 429 Lysine 5,7 2,2 6,0 458 Methionine 2,1 0,8 1,8 105 Cystein 5,2 2,0 0,9 - Phenylalanine 3,4 1,4 3,4 254 Tyrozine 2,6 1,0 2,3 190 Threonine 3,9 1,5 3,8 213 Valine 3,6 1,4 3,4 305 Tryptophan 0,8 0,3 0,8 -
Tỷ lệ hấp thu các amino acid rất khác nhau phụ thuộc vào nguồn protein và khoảng thời gian sau khi ăn (Dabrowshi, 1983, 1986). Gần đây, Akiyama và cộng sự (1997) đã so sánh tỷ lệ amino acid/năng lượng (A/E; liên quan đến tỷ lệ của từng EAA/tổng của EAA) giữa các loài cá, và giữa cá chép với cá catla (Catla catla), cả hai đều thuộc họ Cyprinidae.
Năng lượng. Nhu cầu năng lượng của cá chép rất ít thông tin so với lượng thông tin về các chỉ tiêu dinh dưỡng khác. Như đã mô tả ở các loài cá xương khác, cả tỷ lệ chuyển hóa năng lượng và nhu cầu năng lượng duy trì đều bị ảnh hưởng bởi nhiệt độ nước. Tỷ lệ chuyển hóa kém tích cực ở nhiệt độ dưới 170C là khá thấp (Kaushik, 1995). Tương quan tuyến tính giữa nitrogen (N) ăn vào và sự gia tăng nhiệt độ thức ăn cũng đã được đề cập. Kết quả nghiên cứu cho thấy tỷ lệ năng lượng vào khoảng 40 kJ/g N ăn vào (Chakraborti và cộng sự, 1992; Kaushik, 1995).
Nhu cầu protein và lipit liên quan đến năng lượng tiêu hóa. Giá trị tối ưu của năng lượng tiêu hóa/protein cho sinh trưởng tối đa là 97-116 (Takeuchi và cộng sự, 1979). Nghiên cứu chuyển hóa năng lượng của khẩu phần thực tế bao gồm 25% bột cá, 4% bột thịt, 10% bột đậu nành và 8% bột ngô, Ohta và Wantanabe (1996) cho thấy, 29,9% năng lượng mất qua phân, 1,5% mất qua phần thải khác, 31,9% mất do sinh nhiệt và 36,7% năng lượng thuần (bao gồm 12,6% cho duy trì và hoạt động và 24,1% cho sản xuất). Các tác giả cũng khuyến cáo rằng, nhu cầu năng lượng tiêu hóa cho sinh trưởng tôí đa là 285; 548 và 721 kJ/kg khối lượng cơ thể tương ứng với mức cho ăn 1,83; 3,60 và 5,17% của khối lượng cơ thể/ngày, chịu ảnh hưởng bởi cả khẩu phần và kích cỡ cá.
Lipid và acid béo. Cá chép là loài cá ăn tạp và có thể sử dụng hiệu quả cả lipid và carbohydrates như nguồn năng lượng từ khẩu phần, và vì vậy năng lượng tiêu hóa quan trọng hơn so với lipid trong khẩu phần. Điều đó có nghĩa là việc làm giàu thêm thành phần tạo năng lượng tiêu hóa từ 13 đến 15 MJ/ kg khẩu phần bởi thêm vào 5-15% lipid không làm cải thiện tốc độ sinh trưởng và sử dụng protein thuần (Takeuchi và cộng sự, 1979). Hơn nữa, một mặt tiêu cực của việc tăng tỷ lệ lipid trong khẩu phần là có thể làm tăng tích lũy mỡ trong cơ thể, đặt biệt là mỡ ruột (Zeitler và cộng sự, 1983; Murai và cộng sự, 1985).

117
Cá rất cần các acid béo thiết yếu. Cá chép và cá trắm (Ctenopharygodon adella) đòi hỏi cả acid béo n-6 và n-3. Ước tính cung cấp 1% mỗi acid béo này đảm bảo tốt nhất cho quá trình sinh trưởng và hiệu quả cho ăn đối với cá chép giống (Takeuchi và Watanabe, 1977). Tuy nhiên, cũng có một báo cáo khác thấp hơn nhu cầu này (Kaushik, 1995). Dù vậy, triệu chứng thiếu hụt liên quan đến EFA khó có thể nhận thấy ở cá chép như là chậm lớn, tỷ lệ chết cao và mất sắc tố da. Mặt khác, ở cá trắm cỏ, một loài gần gũi, sự thiếu hụt thể hiện rõ như hiện tượng cong thân, giống như “bệnh Sekoke”, nhưng những dấu hiệu của nó biểu hiện rõ ở sự loạn dưỡng mô cơ (Takeuchi và cộng sự, 1992).
Vai trò của chuỗi triglycerides (MCT) mạch trung bình trong dinh dưỡng ấu trùng cá chép vừa mới được nghiên cứu gần đây (Fontagné và cộng sự, 1999, 2000). Trong lúc tricaproin (C6:0), tricaprin (C10:0), trilaurin (C12:0) và triolein (C18:1) cho thấy có ảnh hưởng một cách
hiệu quả về sinh trưởng và tỷ lệ sống, còn tricaprylin (C8:0) thì không. Điểm riêng của tricaprylin trong các MCT khác, dường như được sử dụng tốt trên 30g/100g của tổng acid béo khẩu phần, liên quan đến sự tổn thương tổ chức của hệ thống enzyme phân giải acid béo. Hình 13.2. Cá Trắm cỏ Phosphilipids (PL) có một số chức năngtrong thức ăn của ấu trùng bao gồm
ảnh hưởng bề mặt, chống lại sự oxi hóa và giữ ổn định nước trong các mẫu thức ăn (Coutteau và cộng sự, 1995). Hậu quả của sự thiếu hụt PL khẩu phần là làm tích lũy các giọt mỡ trong biểu bì ruột trước, làm tăng chất nhầy niêm mạc và giảm thể tích tế bào gan. Tuy nhiên, việc bổ sung phosphatidylcholine (PC) từ lòng đỏ trứng gà hoặc từ đậu tương ngăn chặn được thoái hóa ruột và tăng thể tích tế bào gan (Fontagné và cộng sự, 1998). Những nghiên cứu đã đề cập ở trên cho thấy rằng, PL cần thiết cho việc hấp thu lipid trung tính mặc dù hiệu quả phụ thuộc vào đặc tính nhũ tương hóa của nó. Dựa trên so sánh một vài nguồn PL tạo thành từ lecithin đậu tương, Geurden và cộng sự (1998) thấy rằng PC quan trọng đối với sự hấp thu cao đầu tiên của quá trình sinh trưởng, phosphatidylinositol liên quan chặt chẽ tới sự phát triển của ấu trùng cá chép. Họ đề nghị rằng thêm PL vào khẩu phần bằng phối trộn như trong chất tổng hợp của cơ thể ấu trùng. Trong một nghiên cứu sau này Geurden và cộng sự (1999) nhấn mạnh phân bố bảo tồn của PL sắp sếp theo kích cỡ cá nhất định, không phụ thuộc vào khẩu phần, điều đó giải thích rằng chỉ có sự tái tổ chức có giới hạn là cần thiết.
Carbohydrate. Một vài nghiên cứu đã cho biết cách sử dụng carbohydrate của cá chép. Hoạt động của enzyme amylase trong ống tiêu hóa và tỷ lệ tiêu hóa tinh bột ở cá thường thấp hơn so với sinh vật ở cạn. Trong các loài cá, hoạt động của enzyme amylase ở ruột cao hơn ở các đối tượng ăn tạp, bao gồm cá chép, hơn là các đối tượng ăn động vật. Điều đó cho thấy tỷ lệ chiều dài ruột với chiều dài cơ thể cá chép là 1,8-2,0, giá trị này lớn hơn 4 lần so với cá hồi và cá chình Nhật (Anguilla japonicus), điều này giải thích việc sử dụng tinh bột tốt hơn ở cá chép. Murai và cộng sự (1983) đã kiểm tra ảnh hưởng của các khẩu phần tinh bột khác nhau và việc thường xuyên cho ăn các mẫu thức ăn sử dụng cho cá chép. Sử dụng tinh bột trong khẩu phần hiệu quả nhất khi cho ăn 2 lần mỗi ngày, còn glucose và maltose ít nhất 4 lần. Điều đó chỉ ra

118
rằng, có sự giảm sút trong hiệu quả hấp thu của glucose và maltose khi sử dụng số lượng lớn cùng một thời điểm.
Ogino và cộng sự (1976) nhận thấy rằng, cá trắm cỏ sử dụng tinh bột một cách hiệu quả như là một nguồn cung cấp năng lượng. Sau đó, Takeuchi và cộng sự (1979) cũng xác định giá trị khẩu phần của carbohydrates như là nguồn cung cấp năng lượng. Kết quả của nhiều nghiên cứu cho thấy, lượng tối ưu của carbohydrate khẩu phần dao động 30-40% khối lượng thức ăn đối với cá chép.
Vitamin và muối khoáng. Nhu cầu về chất lượng và số lượng Vitamin của cá chép đã được xác định; các biểu hiện do thiếu Vitamin được trình bày ở bảng 13.4. Nhu cầu trong khẩu phần đối với acid folic và vitamin B12, D và K chưa được nghiên cứu, nhưng một số vitamin này có thể được tổng hợp bởi các vi khuẩn đường ruột đối với cá chép và một số loại cá nước ngọt khác (Lovell và Limsuwan, 1982). Nhu cầu vitamin ở cá chép có thể bị tác động bởi nhiều yếu tố, như kích cỡ cá, nhiệt độ nước và thành phần thức ăn. Ví dụ, cá giống và cá chép trưởng thành không có nhu cầu về vitamin C bởi vì bản thân chúng có thể tổng hợp vitamin C từ D-glucose. Tuy nhiên, ấu trùng cá chép có những biểu hiện do thiếu vitamin C như mòn vây đuôi, biến dị xương cung mang (Dabrowski và CS, 1098). Những nghiên cứu về dinh dưỡng giai đoạn đầu của ấu trùng cho thấy rằng nhu cầu tích lũy tối đa trong mô (270mg vitamin C/1kg) là cao hơn nhu cầu cho duy trì và sinh trưởng (45mg vitamin C/kg) (Goullou-Coustans và CS, 1988).
Bảng 13.4. Nhu cầu vitamin ở cá chép và những triệu chứng thiếu (Satoh, 1991; NRC, 1993)
Vitamin Nhu cầu (mg/kg)
Dấu hiệu thiếu hụt
Thiamine 0,5 Tăng trưởng kém, hốt hoảng, mất sắc tố da, xuất huyết dưới da Riboflavin 7 Biếng ăn, tăng trưởng kém, xuất huyết ở gan, da và vây, gầy
mòn, sợ ánh sáng, hốt hoảng, hoại tử thận sớm, Pantothenate 30 Biếng ăn, chậm lớn, cáu kỉnh, xuất huyết ở da, đờ đẫn, mắt lồi Niacin 28 Biếng ăn, chậm lớn, sống yếu, xuất huyết da, tỷ lệ chết cao Biotin 1 Chậm lớn, hồng cầu dễ vỡ ra từng mảnh, hôn mê, tăng tế bào
nhầy da Choline 500 Chậm lớn, gan nhiễm mỡ, rỗng hóa tế bào gan Vitamin A 4000 IU Biếng ăn, chậm lớn, mắt lồi, xuất huyết mang, da, mang xoắn,
mất sắc tố da Vitamin E 100 Loạn dưỡng cơ, mắt lồi, lưng cong, thoái hóa gan, thận. Vitamin C Đã thỏa mãn Mòn vây đuôi, biến dị xương cung mang ở giai đoạn ấu trùng,
sinh trưởng kém Liên quan đến vitamin E, nhu cầu có thể tăng lên phụ thuộc vào mức độ của axit béo
không có khả năng sinh cholesterol trong khẩu phần. Hiện nay kỹ thuật đùn thức ăn đã được áp dụng nên có thể làm thay đổi khẩu phần và một vài axit amin có thể bị mất tác dụng trong quá trình chế biến.
Mức độ bổ sung vitamin vào khẩu phần thức ăn cao gấp 2-5 lần so với nhu cầu muối khoáng và những biểu hiện do thiếu khoáng được tóm tắt ở bảng 13.5 (cá chép có nhu cầu về Co, Cu, Mg, Mn, P, Zn). Cá chép thiếu 1 acid thiết yếu tiết ra từ dạ dày cho tiêu hóa và hòa tan các hợp chất chứa cả Ca và P; như vậy P tồn tại phụ thuộc vào muối và thành phần khác hòa tan trong nước (Satoh và cộng sự, 1992,1997). Hàm lượng P từ tricalcium phosphate của bột cá (FM) ít hơn so với nhiều mono và dicalcium phosphate hòa tan.

119
Bảng 13.5. Nhu cầu muối khoáng của cá chép và những triệu chứng do thiếu muối khoáng (Satoh, 1991; NRC, 1993; Kim và cộng sự,1998)
Muôi khoáng
Nhu cầu Dấu hiệu thiếu hụt
Phốt pho 6 – 8 g/kg Chậm lớn, xương không bình thường, hiệu quả thức ăn kém, lượng khoáng thấp toàn cơ thể và cột sống, tăng mỡ nội tạng
Magiê 0,4 - 0,5 g/kg Chậm lớn, biếng ăn, tỷ lệ chết cao, lờ đờ và co giật, lượng canxi cao, magiê giảm trong xương, đục nhân mắt.
Sắt 150 mg/kg Mất trọng lực, Hb, huyết tương không bình thường Kẽm 15-30 mg/kg Chậm lớn, tỷ lệ chết cao, mòn mang và da, kẽm thấp trong xương Mangan 13 mg/kg Chậm lớn, còi cọc, xương không bình thường, tỷ lệ chết cao,
Canxi, Mg, P, kẽm và Mn thấp trong xương Đồng 3 mg/kg Chậm lớn Coban 0,1 mg/kg Chậm lớn
Việc bổ sung mono phosphate vào khẩu phần chứa bột cá là nguyên liệu cơ bản đã mang lại kết quả tốt về tăng trưởng của cá chép. Nó cũng cho thấy rằng, việc bổ sung các chất từ bên ngoài như Cu, Mn, Mg và Zn rất cần thiết cho khẩu phần thức ăn của cá chép. Tuy nhiên, ở cá chép phải chú ý rằng một lượng giới hạn của tricalcium phosphate có thể ngăn cản sự cung cấp của một số các yếu tố, như Zn và Mn, mặc dù ít hơn nhiều so với cá hồi (Satoh và cộng sự, 1989). Trong một nghiên cứu xem xét sự tương tác giữa thiếu Zn và lipid, người ta thấy các chất dinh dưỡng không được hấp thu và liên quan đến sự tích tụ lipid ở ruột (Taneja và Arya, 1994). 13.1.3. Khẩu phần ăn
Thay đổi theo thời gian, phương thức cho ăn truyền thống với thức ăn thô được chế biến bằng những nguyên liệu sẵn có ở địa phương chuyển dần sang cách cho ăn thức ăn công nghiệp. Ở những nước đang phát triển như ở Trung Quốc, người dân vẫn còn áp dụng hình thức nuôi quảng canh dựa vào thức ăn tự nhiên, thức ăn tươi sống và những thức ăn khô chế biến đơn giản (Song, 1994). Khẩu phần ăn đơn giản bao gồm khô dầu dừa, cám, cỏ và thức ăn xanh được sử dụng trong nuôi quảng canh, trong lúc khẩu phần bao gồm bột cá, thịt và bột xương, nhộng tằm và bột máu được áp dụng cho hình thức nuôi thâm canh.
Khẩu phần thức ăn cho cá chép lấy bột cá làm nguyên liệu cơ bản. Các nghiên cứu nhằm vào mục đích giảm giá thành và tạo nên một khẩu phần phù hợp hơn cho môi trường đã được tiến hành. Sử dụng thức ăn đơn lẻ nhanh chóng được áp dụng đối với nhiều đối tượng nuôi, trong đó bao gồm cả cá chép. Thức ăn đơn lẻ có những đặc điểm giống viên thức ăn đùn, viên thức ăn khô và đồng thời có chi phí thấp.
Khả năng tiêu hóa các thành phần thức ăn. Chúng ta biết rõ rằng, tiêu hóa protein biểu kiến (APD) và năng lượng ở một số thành phần của khẩu phần phụ thuộc vào mỗi loài và nhiệt độ. Đặc biệt là đối với thức ăn xanh. Bảng 13.6 chỉ ra tỷ lệ tiêu hóa biểu kiến protein và năng lượng của một số loại thức ăn ở nhiệt độ nước khác nhau của cá chép (Takeuchi, 1991). Giá trị tiêu hóa không bị ảnh hưởng bởi kích thước cá có khối lượng dao động từ 3-295 g.
Trong thiết lập khẩu phần ăn cho cá chép, người ta đang cố gắng để thay thế bột cá bằng nguồn protein từ thực vật mà vẫn cho hiệu quả tốt. Takeuchi và cộng sự (1990) cho rằng, carbohydrate từ các nguồn như bột khoai tây, bột ngô, lúa mạch, bột mì đã được cải thiện nhờ xử lý đùn, làm tăng mức gelatin hóa cũng như năng lượng. Jeong và cộng sự (1992) đã điều tra về ảnh hưởng của tỉ lệ carbohydrate được gelatin hóa trong khẩu phần với năng lượng tiêu hóa

120
của cá chép con nuôi bằng khẩu phần chứa khoai tây và khoai tây đùn. Sinh trưởng đạt tốt nhất khi mức gelatin hóa là 40% trong khẩu phần có năng lượng thấp (19,7 MJ/kg) và 20% trong khẩu phần có năng lượng cao (20,9 MJ/kg).
Bảng 13.6. Tỷ lệ tiêu hóa protein biểu kiến và năng lượng của các loại thức ăn khác nhau trong khẩu phần ăn của cá chép ở các mức nhiệt độ khác nhau
Thành phần TLTH biểu kiến (%) Protein Năng lượng
Thành phần thức ăn Ẩm
độ, % Protein %
GE, MJ/kg 15oC 20oC 25oC 15oC 20oC 25oC
Bột cá trắng 8.8 63.8 0.17 94 90 92 86 89 88 Bột cá địa phương 10.8 63.5 0.18 90 90 93 87 87 86 Bột thịt 11.8 69.0 0.22 91 91 95 76 79 82 Bột nhộng tằm 7.8 52.6 0.24 87 86 88 80 79 82 Bột Gluten ngô 8.8 64.3 0.22 82 88 93 73 80 85 Bột đỗ tương khử mỡ
10.7 45.3 0.17 94 96 95 76 79 80
Mầm lúa mỳ khử mỡ 10.6 31.3 0.17 92 93 94 70 74 77 Cám gạo khử mỡ 11.3 15.8 0.16 60 85 88 48 71 76 Bột mỳ 13.1 18.8 0.16 78 79 80 49 73 79
Loại thức ăn từ protein đậu nành cô đặc (SPC) có hệ số tiêu hóa biểu kiến (TLTH) protein và lipit ở 180C cao hơn bột cá trích (HM) (Kim và cộng sự, 1988). Tuy nhiên, TLTH năng lượng không có sự khác nhau giữa SPC và HM. Trong nghiên cứu khác, sử dụng khô dầu được làm từ vừng, hạt cải, hạt lanh và dừa khô các mức từ 25%-75% so tổng số protein trong khẩu phần, cho thấy rằng TLTH protein không sai khác giữa các khẩu phần và trong khoảng 78%-90% (Hasan và cộng sự ,1997).
Protein trong bột mạch nha (MPF), bã bia được sử dụng như 1 nguồn thức ăn giàu protein. MPF có thể thay thế 40% hoặc 60% bột cá trắng (WFM) trong khẩu phần cá con. Nếu thay thế 60% thì cần bổ sung EAA (axit amin thiết yếu) tinh khiết (Yamamoto và cộng sự, 1996). Ngoài ra, thay thế 60% MPF cho bột đậu nành (SBM) trong khẩu phần cho tăng trọng tốt hơn khẩu phần chỉ có WFM. Vì vậy, MPF cũng là một lựa chọn tốt để làm nguồn thức ăn giàu protein.
Trong thiết lập khẩu phần sử dụng nhiều nguồn protein, các dữ liệu về TLTH protein cũng như các amino acid là rất cần thiết. Phương thức chế biến thức ăn cũng có thể làm mất phần lớn hàm lượng các chất dinh dưỡng trong khẩu phần. Thức ăn chế biến đùn hoặc gelatin hóa carbohydrate có thể làm mất các amino acid bởi sự oxi hoá và bị giảm hàm lượng một vài acid amin (như lysine) thông qua phản ứng Maillard. Yamamoto và cộng sự (1998) đã so sánh TLTH biểu kiến của WFM, SBM, đùn SBM (ExSBM ), MPF, ExMPF và bột ngô (CGM), kết quả cho thấy TLTH biểu kiến của chúng thấp hơn xấp xỉ 1,9% tỷ lệ tiêu hóa protein thực (TLTHT). Sự khác nhau giữa tỷ lệ tiêu hóa acid amin thực và acid amin biểu kiến cũng giống như sự khác nhau giữa TLTH protein thực và TLTH protein biểu kiến. Xác định được TLTH của các acid amin giúp ta đánh giá chính xác hơn về số lượng chất dinh dưỡng của các nguyên liệu phối trộn trong khẩu phần của thức ăn chế biến cũng như thức ăn công nghiệp.
Dầu cá ngừ là nguồn lipit trong khẩu phần thức ăn cá chép (Applfold và Anderson, 1997). Dầu cá có TLTH khác nhau tại các điểm nóng chảy khác nhau, Takeuchi và cộng sự (1979) nhấn mạnh rằng việc tiêu hóa dầu cá tại điểm nóng chảy cao (530C) là thấp, đặc biệt là

121
trong tăng trưởng của với cá chép dưới 10 g. Khi điểm nóng chảy của dầu cá thấp hơn (380C) thì nó được sử dụng hiệu quả (hơn 70%) không tính đến kích cỡ cá và nhiệt độ nước. Vì vậy, lipid trong nguyên liệu kém chất lượng có thể dùng như một nguồn cung cấp năng lượng mà không có tác dụng phụ nào.
Trong số tất cả các giải pháp nhằm thay thế bột cá trong khẩu phần đã được thử nghiệm, bột đậu tương (SBM) đã được sử dụng rộng rãi nhất. Một nghiên cứu dài hạn bao gồm cả trong phòng thí nghiệm và ngoài trời đã chứng minh hiệu quả của bột đậu nành là nguyên liệu chính trong khẩu phần (Noble và cộng sự, 1998).
Việc thay thế hòan toàn bột cá nâu (BFM) bằng sự phối hợp của SBM, CGM và bột thịt (MM) đã được bổ sung các amino acid tổng hợp như lysine, methionine và threonine đã không cho kết quả tương đương về tăng trưởng và khả năng sử dụng thức ăn, mặc dù hương vị của nó cũng hấp dẫn như khẩu phẩn bột cá (Pongmaneerat và cộng sự, 1993). Pongmaneerat và cộng sự (1993) đề nghị thay thế 56% BFM trong khẩu phần của cá chép bằng 25% SBM và 10% CGM trong khẩu phần chứa 38% protein. Điều này trái ngược với mức thay thế khoảng 90% ở mức 44% protein khẩu phần cho cá hồi (Pongmaneerat, 1993). Mặc dù xét một cách tổng quát, cá chép có thể sử dụng protein thực vật hiệu quả hơn so với cá hồi, các mức thấp hơn đã đạt được ở cá chép có thể là do sử dụng thấp hơn các phân tử amino acid được cung cấp. Cũng nên kiểm tra Taurine nếu như Taurine là một amino acid thiết yếu như đã đề cập trên đây.
Những thí nghiệm thay thế bột cá khẩu phần cũng đã chứng minh rằng photspho trong môi trường có thể giảm xuống tương ứng với những tỷ lệ bột cá khác nhau. Căn cứ trên photspho tổng số và nitrogen, Jahan và cộng sự (2000) cho rằng tỉ lệ FM phù hợp trong khẩu phần thức ăn của cá chép là từ 20-25%. 13.1.4. Nuôi dưỡng
Việc nuôi thâm canh cá chép không chỉ phải chú ý đến tỉ lệ phối trộn các loại thức ăn để đạt sinh trưởng tối ưu mà còn phải kiểm soát được chất thải. Do vậy, nên sử dụng các công thức phối chế khẩu phần và cách cho ăn thích hợp, chú ý đến vấn đề tiêu hoá – trao đổi và chất thải. Nhiệt độ và kích cỡ cá ảnh hưởng lớn đến tỷ lệ cho ăn hàng ngày đối với cá chép (bảng 13.7 và 13.8). Nhiệt độ thích hợp nhất cho quá trình sinh trưởng của cá là 30-320C, khi nhiệt độ giảm xuống trên dưới 100C thì sinh trưởng sẽ chậm lại.
Bảng 13.7. Tỷ lệ cho ăn hàng ngày thay đổi theo nhiệt độ và kích cỡ cá chép nuôi bằng thức ăn công nghiệp (Miyatke, 1997)
Khối lượng cá (g) To nước (oC) 2-5 5-10 10 -
20 20 -30
30 -40
40 -50
50 -100
100-200
200-400
400-600
600-800
800-1000
15 4.5 3.7 3.2 2.8 2.5 2.3 2.0 1.7 1.5 1.3 1.1 0.9 16 4.8 4.0 3.4 3.0 2.7 2.4 2.1 1.8 1.6 1.4 1.1 0.9 17 5.2 4.3 3.7 3.2 2.9 2.6 2.2 1.9 1.7 1.5 1.2 1.0 18 5.6 4.7 4.0 3.4 3.1 2.8 2.3 2.0 1.8 1.6 1.3 1.0 19 6.0 5.1 4.3 3.7 3.4 3.0 2.5 2.2 1.9 1.7 1.3 1.1 20 6.5 5.5 4.6 4.0 3.7 3.2 2.7 2.4 2.2 1.8 1.4 1.2 21 7.0 6.0 4.9 4.3 4.0 3.4 2.9 2.6 2.1 1.9 1.5 1.3 22 7.6 6.5 5.3 4.6 4.3 3.7 3.1 2.8 2.3 2.0 1.6 1.4 23 8.2 7.0 5.7 4.9 4.6 4.0 3.4 3.0 2.5 2.1 1.7 1.5 24 8.8 7.5 6.1 5.3 4.9 4.3 3.7 3.2 2.7 2.3 1.8 1.6 25 9.5 8.0 6.5 5.7 5.2 4.6 4.0 3.4 2.9 2.5 2.0 1.7

122
26 10.2 8.5 6.9 6.1 5.5 4.9 4.3 3.7 3.1 2.7 2.2 1.8 27 10.9 9.0 7.4 6.5 5.8 5.2 4.6 4.0 3.3 2.9 2.4 1.9 28 11.6 9.5 7.9 6.9 6.2 5.6 4.9 4.3 3.6 3.1 2.6 2.0 29 12.3 10.1 8.4 7.4 6.6 6.0 5.2 4.6 3.9 3.3 2.8 2.1 30 13.1 10.7 8.9 7.9 7.0 6.4 5.7 4.9 4.2 3.5 3.0 2.2
Bảng 13.8. Tỷ lệ cho ăn hàng ngày thay đổi theo nhiệt độ và kích cỡ cá chép nuôi bằng khẩu phần nghèo protein nhưng giàu năng lượng do Kasumigaura điều chỉnh (Miyatke, 1997)
Khối lượng cá (g) To nước (oC) 1-5 5-10 10-50 50 -100 100-200 200-400 400-600 600-800 800-1000
15 4.65 3.42 2.16 1.59 1.26 0.95 0.84 0.76 0.69 16 5.15 3.80 2.39 1.76 1.40 1.05 0.94 0.84 0.77 17 5.17 4.21 2.65 1.95 1.55 1.17 1.04 0.93 0.85 18 6.33 4.66 2.94 2.17 1.72 1.30 1.15 1.03 0.95 19 7.02 5.17 3.26 2.40 1.90 1.44 1.28 1.14 1.05 20 7.78 5.73 3.61 2.66 2.11 1.59 1.41 1.26 1.16 21 8.62 6.35 4.00 2.95 2.34 1.76 1.57 1.40 1.29 22 9.65 7.04 4.44 3.27 2.59 1.96 1.74 1.55 1.43 23 10.60 7.81 4.92 3.62 2.88 2.17 1.93 1.72 1.58 24 11.75 8.65 5.45 4.02 3.19 2.40 2.13 1.91 1.75 >=25 13.02 9.60 6.04 4.45 3.53 2.66 2.37 2.11 1.94

123
13.2. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ TRÔI ẤN ĐỘ (INDIAN MAJOR CARP)
13.2.1. Giới thiệu Cá trôi Ấn Độ là một đối tượng nuôi kinh tế ở Ấn Độ và các tiểu lục Ấn Độ. Catla
(Catla catla), Rohu (Labeo rohita) và Mrigal (Cirrhinus mrigala) là 3 loài chủ yếu trong nhóm cá trôi Ấn Độ. Một số cá trôi với kích cỡ nhỏ và trung bình phát triển tốt trong các ao nuôi nước ngọt. Phần lớn cá trôi Ấn Độ được nuôi ghép. Ở hình thức nuôi này sử dụng chủ yếu nguồn thức ăn có sẵn trong tự nhiên ở những mức khác nhau để đạt được năng suất tối đa trên mỗi đơn vị diện tích (Jhingran, 1991).
Cá trôi không chỉ được nuôi ở Ấn Độ mà còn được nuôi ở các nước lân cận như Pakistan, Bangladesh, Nepal, Sri Lanka và các nước lân cận. Cá trôi nuôi ở Ấn Độ đã từ vài thế kỷ trước, hầu hết cá trôi nuôi được tiêu thụ nội địa. Trong những năm gần đây, thịt fillet cá trôi và các sản phẩm khác của nó được xuất khẩu đến các nước Trung Đông.
Sản phẩm Cá Trôi từ Ấn Độ đứng hàng thứ hai trên Hình 13.3. Cá Trôi Ấn Độ thế giới sau Trung Quốc. Trong 2000 tấn thuỷ sản của Án Độ, phần lớn là các sản phẩm cá trôi nuôi. Cho đến nay, trên thế giới đã có nhiều công trình phối hợp nghiên cứu về dinh dưỡng thức ăn và nuôi dưỡng trên đối tượng cá trôi Ấn Độ. 13.2.2. Nhu cầu dinh dưỡng
Protein và amino acid. Nhu cầu protein khẩu phần tối ưu ảnh hưởng bởi giá trị dinh dưỡng của protein thức ăn và năng lương từ nguồn khác protein trong khẩu phần. Khi nguồn năng lượng như lipid và cacbohydrate được cung cấp đầy đủ trong khẩu phần, hầu hết protein tiêu hoá được sử dụng cho quá trình tổng hợp protein. Cá trôi Ấn Độ trưởng thành cần 30% protein khẩu phần cho nhu cầu duy trì và sinh trưởng. Nhu cầu của cá giống và cá hương là 35% và 40% protein khẩu phần cho sinh trưởng tốt (Sen và cộng sự, 1978; Renukaradhya và Varghese, 1986). Theo báo cáo của Mondal và cộng sự (2000), nhu cầu protein của cá hương cho duy trì và sinh trưởng tối ưu là 38,5% khẩu phần.
Cá trôi Ấn Độ trưởng thành cũng giống như những động vật khác, không có nhu cầu về protein nguyên chất nhưng lại đòi hỏi hỗn hợp thức ăn cân đối về các amino acid thiết yếu và không thiết yếu (Murthy và Varghese, 1998). Chất lượng và số lượng về nhu cầu amino acid của cá trôi Ấn Độ được xác định bởi những nghiên cứu về sinh trưởng thông qua phương pháp nuôi dưỡng. Nhu cầu về chất lượng amino acid của cá trôi cũng được thiết lập bởi đồng vị C14. Thành phần các amino acid của cơ thể chỉ giúp ta ước tính một cách sơ lược về nhu cầu, nên chỉ sử dụng trong trường hợp không có thông tin về nhu cầu amino acid khẩu phần. Những nghiên cứu về nhu cầu khẩu phần ăn của cá trôi đã cho

124
thấy, chúng cần tất cả 10 amino acid thiết yếu (EAA). Nhu cầu về số lượng amino acid khẩu phần của cá Catla, Rohu và Mrigan đối với 10 amino acid được trình bày trong bảng 13.9, 13.10 và 13.11. Cysteine có giá trị thay thế khoảng 50% đối với cá trôi Ấn Độ.
Bảng 13.9. Nhu cầu amino acid khẩu phần của Catla catla (% protein khẩu phần)
Amino acid Cá bột Cá hương/trưởng thành Arginine 4,80 5,63 Histidine 2,45 2,38 Isoleucine 2,35 2,75 Leucine 3,70 4,38 Lysine 6,23 6,86 Methionine 3,55 3,00 Phenylalanine 3,70 4,50 Threonine 4,95 4,50 Tryptophan 0,95 1,03 Valine 3,55 3,60
Bảng 13.10. Nhu cầu amino acid khẩu phần của Labeo rohita
Amino acid % khẩu phần % protein KP Nguồn Arginine 2,30 5,75 Murthy và Varghese (1995) Histidine 0,90 2,25 Murthy và Varghese (1995) Isoleucine 1,20 3,00 Murthy và Varghese (1996a) Leucine 1,85 4,63 Murthy và Varghese (1997a) Lysine 2,27 5,58 Murthy và Varghese (1997) Methionine* 1,15 2,88 Murthy và Varghese (1998) Phenylalanine** 1,60 4,00 Murthy và Varghese (1996) Threonine 1,71 4,28 Murthy và Varghese (1996) Tryptophan 0,45 1,13 Murthy và Varghese (1997) Valine 1,50 3,75 Murthy và Varghese (1997) * Tổng số amino acid gốc Sulfua (Methionine + Cysteine) là 1,42% (3,55% protein khẩu phần)
** Tổng số amino acid thơm (Phe + Tyrosine) là 2,31% (5,77% protein khẩu phần)
Bảng 13.11. Nhu cầu amino acid khẩu phần của Cirrhinus mrigala
Amino acid % khẩu phần % protein KP Arginine 2,10 5,25 Histidine 0,85 2,13 Isoleucine 1,10 2,75 Leucine 1,70 4,25 Lysine 2,35 5,88 Methionine 1,27 3,18 Phenylalanine 1,60 4,00 Threonine 1,65 4,13 Tryptophan 0,43 1,08 Valine 1,40 3,50

125
Lipid và acid béo. Dầu và mỡ (gọi chung là lipid) là nguồn cung cấp năng lượng và các acid béo thiết yếu. Hơn nữa, lipid còn đóng vai trò vận chuyển các vitamin hoà tan trong mỡ. Acid béo và phospholipid giúp duy trì cấu trúc màng tế bào. Nhu cầu lipid tổng số đối với cá trôi Ấn Độ trưởng thành là 7-8% của khẩu phần. Cá con có nhu cầu về lipid và protein cao hơn cá trưởng thành.
Cả 3 loài các trôi Ấn Độ trưởng thành đều sinh trưởng tốt khi khẩu phần chứa 1% acid béo n-3 và 1% acid béo n-6. Cơ thể cá trôi Ấn Độ trưởng thành chứa một tỷ lệ lớn acid béo n-3 cũng như acid béo n-6. Đã có những nghiên cứu sâu về nhu cầu acid béo khẩu phần của cá trôi Ấn Độ. Tuy nhiên vai trò của các acid béo mạch dài chưa bão hoà (HUFA), như là eicosapentaenoic acid (EPA) (20:5n-3) và docosahexaenoic acid (DHA) (22:6n-3) vẫn chưa được nghiên cứu trên cá trôi Ấn Độ.
Carbohydrates. Carbohydrate là loại thức ăn tinh và là nguồn cung cấp năng lượng rẻ tiền nhất đối với cá trôi. Là sinh vật ăn thực vật/ ăn tạp, cá trôi Ấn Độ trưởng thành dễ dàng tiêu hoá một lượng lớn tinh bột trong khẩu phần. Mức 22-30% tinh bột trong khẩu phần cho kết quả sinh trưởng tối ưu đối với cá trôi Ấn Độ. Khi tỷ lệ tinh bột vượt quá 35% của khẩu phần thì sẽ làm giảm sinh trưởng và hiệu quả sử dụng thức ăn thấp. Trong thực tế, cám, bột sắn, gạo là những nguồn tinh bột được sử dung trong phối hợp khẩu phần. Những nguyên liệu này cũng đóng vai trò là chất kết dính tự nhiên trong khẩu phần. Sự thiếu hụt tinh bột trong khẩu phần của cá trôi có thể dẫn đến việc huy động protein để sử dụng như một nguồn năng lượng.
Vitamin và muối khoáng. Thiamine là một trong những vitamin cần thiết cho quá trình sinh trưởng và tồn tại của một số loài cá nước ngọt. Chưa có một nghiên cứu nào về nhu cầu thiamine trong khẩu phần thức ăn của cá trôi Ấn Độ. Tuy nhiên, ước lượng từ 8-12mg/kg thức ăn đảm bảo đủ nhu cầu, phụ thuộc vào từng giai đoạn sống. Trong khẩu phần thức ăn công nghiệp, người ta thường cho vitamin này với một lượng cao để trừ bù tỷ lệ hao hụt qua quá trình chế biến và bảo quản.
Riboflavin cần thiết cho cá trôi đã trưởng thành, sự thiếu hụt Riboflavin dẫn đến hậu quả mòn cụt vây, bỏ ăn, mất sắc tố và đục mắt ở cá. Nhu cầu về Riboflavin ở cá trôi Ấn Độ vào khoảng 6-8mg/kg thức ăn. Tuy nhiên, trong thức ăn công nghiệp, bổ sung một lượng Riboflavin 10mg/kg thức ăn hoặc hơn nhằm loại trừ lượng vitamin mất khi hoà tan trong nước hoặc qua quá trình lọc.
Ước lượng, đối với 3 loài cá trôi Ấn Độ, khoảng 10-12mg niacin/kg thức ăn đảm bảo cho quá trình sinh trưởng bình thường.
Nhu cầu về pantothenic acid đối với cá trôi Ấn Độ là khoảng 9-11 mg/kg thức ăn. Sử dụng các dạng ít bị phân huỷ của vitamin C, như ascorbate 2-monophosphate
hoặc ascorbate 2-sulphate trong thức ăn cá trôi vì vitamin C dễ dàng bị phân hủy và mất qua lọc. Một vài nghiên cứu về nhu cầu ascorbic acid của cá trôi Ấn Độ trưởng thành đã được thực hiện. Tuy nhiên, mức 300mg ascorbic acid/kg trong khẩu phần thức ăn là đủ cho quá trình sinh trưởng và sống bình thường của ấu trùng và cá hương Rohu và Mrigan. Người ta đã làm các thí nghiệm tăng các mức vitamin C lần lượt trong khẩu phần ăn lên đến 1000mg/kg thức ăn cho cả cá Rohu và Mrigan. Khả năng đề kháng, đặc biệt là đối với vi khuẩn Aeromonas hydrophyla được tăng lên bởi việc bổ sung vitamin C và giảm thiểu tỷ lệ chết khi cá Rohu được bổ sung 600 mg vitamin C/ kg thức ăn và Mrigan là 1000 mg/kg thức ăn (Sobana và cộng sự, 1997). Chưa có báo cáo nào về ảnh hưởng của việc bổ sung Vitamin C đến tỷ lệ chết, các biểu hiện thiếu vitamin C hoặc các dấu hiệu sinh trưởng không bình thường so với việc không bổ sung vitamin C (Hasan và cộng sự, 1993). Nhìn

126
chung, bổ sung 100-150 mg ascorbic acid/kg thức ăn là phù hợp cho quá trình sinh trưởng bình thường của cá trôi Ấn Độ trưởng thành. Sự thiếu hụt ascorbic acid sẽ dẫn đến việc bỏ ăn, chậm lớn, tỷ lệ chết cao, hoại tử vây, mất sắc tố da, cong cột sống ở cá Mrigan (Agarwal và Mahajan, 1980; Mahajan và Agarwal, 1980), xuất huyết dưới da, thiếu máu ngoại vi và thiếu máu hypochronic cá trôi Ấn Độ (Sabana, 1997).
Các biểu hiện do thiếu Pyridoxine ở cá là bỏ ăn, hoại tử, rối loạn thần kinh, bấn loạn và ngột thở. Nhu cầu pyridoxine: 6-8mg pyridoxine/ kg thức ăn.
Nhu cầu về vitamine B12 vẫn chưa được xác định ở cá trôi Ấn Độ, nhưng khuyến cáo bổ sung 0,001-0,01 mg/kg.
Nhu cầu inositol dao động từ 300 đến 350 mg/kg thức ăn; biotin 5-8mg/kg thức ăn; acid folic là 0,5-1mg/kg thức ăn; choline 500-600 mg/kg thức ăn.
Sự thiếu hụt Vitamin A (retinoic acid) gây nên các hiện tượng như: thay đổi vị trí của thấu kính mắt, loãng dịch màng mắt, chậm lớn, lồi mắt, mất sắc tố trên da cá. Nhu cầu vitamin A của cá trôi Ấn Độ trưởng thành là 1.500UI.
Việc thiếu hụt vitamin D sẽ dẫn đến giảm quá trình sinh trưởng và teo cơ trong vây cá, kể cả cá trôi. Việc điều chỉnh các muối khoáng cũng đóng vai trò rất quan trọng, nhất là Ca và P. Không có nghiên cứu nào về nhu cầu vitamin D ở cá trôi Ấn Độ trưởng thành; tuy nhiên mức vitamin D từ 200-500 UI đã được khuyến cáo; vitamin E là 40 -50 g/ kg thức ăn; vitamin K dao động 5-10 mg/ kg thức ăn.
Về muối khoáng, nhu cầu canxi khẩu phần của cá trôi Ấn Độ đến nay vẫn chưa được nghiên cứu, nhưng mức 4000-5000 mg canxi/ kg được khuyến cáo bổ sung. Biểu hiện do thiếu canxi vẫn chưa được phát hiện ở cá trôi (Ogino và Takeda, 1976; Lall và cộng sự, 1985). Thiếu photspho dẫn đến hiệu suất sử dụng thức ăn kém, bỏ ăn, giảm sự khoáng hoá ở xương, cong cột sống, biến dạng sọ và những rối loạn khác ở cá trôi. Mức 5.000-6.000 mg/kg thức ăn đảm bảo an toàn cho chúng. Mức Magiê 500 mg/kg thức ăn là tốt nhất cho cá trôi, trong khi đồng là 3-4 mg/ kg thức ăn vẫn chưa đủ nhu cầu của nó. Khi bổ sung 0,1 mg cobalt/ kg thức ăn đối với cá hương trong ao ương, ở dạng cobalt chloride, tăng sức sống và tỷ lệ tăng trưởng (Alikunhi, 1987). Nhu cầu các muối khoáng khác trong khẩu phần của cá trôi Ấn Độ vẫn chưa được nghiên cứu. 13.2.3. Khẩu phần và nuôi dưỡng
Việc bổ sung dinh dưỡng là cần thiết để tăng sản lượng cá hương trong ao. Tỷ lệ sống trung bình của cá trôi Ấn Độ trưởng thành trong giai đoạn đầu khá thấp (khoảng 30% từ trứng lên cá hương) và xấp xỉ 50% từ cá hương lên cá giống. Khi cá có tỉ lệ chết cao cần cung cấp đầy đủ thức ăn cân đối dinh dưỡng và quản lý tốt.
Khẩu phần truyền thống là sử dụng cá vụn trộn với cám gạo và bánh dầu theo tỉ lệ bằng nhau để nuôi cá trôi Ấn Độ trưởng thành. Khẩu phần thức ăn có tỷ lệ 1:1 cám gạo và bánh khô dầu thường chứa 25-28% protein thô. Tuy nhiên, khẩu phần này không phải là một khẩu phần cân bằng dinh dưỡng. Thông thường, bánh khô dầu được ngâm trong nước sau vài giờ rồi trộn với cám gạo để tạo thành hỗn hợp thức ăn bột, dạng thức ăn chưa nấu chín, 3 lần mỗi ngày cho xuống ao ương, lượng cho ăn ứng với 10-20% khối lượng cá.
Khẩu phần cân bằng dinh dưỡng được sử dụng cho cá trôi giống và cá trôi trưởng thành do phối hợp nhiều nguyên liệu như trình bày ở bảng 13.12.
Nguồn bột cá, khô dầu, nhộng tằm, phụ phẩm chế biến tôm.. là nguồn protein có giá trị cao cho cá trôi (Varghese và cộng sự, 1976; Jeychandran và Paulraj, 1977; Jayaram và Shetty, 1980). Bột của bèo khô và bắp cải được phối hợp trong khẩu phần nhưng không cho kết quả như mong muốn (Devaraj và cộng sự, 1981). Kết hợp của bột nhộng tằm, bơ

127
thực vật, dầu cá mòi và bột cá Cassia tora trong công thức thức ăn làm tăng sức sống và tốc độ tăng trưởng cuả cá trôi Ấn Độ trưởng thành (Manissery và cộng sự 1988). Murthy và Devaraj (1990,1991) nghiên cứu sử dụng 3 loại thực vật thuỷ sinh dạng bột là Eichhornia, Pistia và Salvinia, cho thấy rằng, thức ăn chứa Pistia có hiệu quả hơn hẳn 2 khẩu phần kia.
Bảng 13.12. Một số loại thức ăn thường gặp trong khẩu phần của cá trôi ở Ấn Độ
Thành phần thức ăn Ẩm độ (%)
Protein thô %
Mỡ thô (%)
Xơ thô (%)
Khoáng (%)
DXKĐ (%)
Gạo đánh bóng 12,6 14,5 17,3 7,5 -* - Tấm gạo 10,1 12,6 11,3 19,3 10,2 36,5 Cám gạo khử mỡ 7,2 12,1 1,3 15,2 23,8 40,4 Cám mỳ 12,3 15,8 4,3 8,7 - - Tấm mỳ 9,0 11,5 1,9 4,0 0,2 73,4 Bột mỳ 12,6 14,5 3,7 2,7 2,3 64,2 Khô dầu lạc 10,0 42,0 7,3 13,0 2,5 25,2 Khô dầu hướng dương 8,0 31,0 2,1 18,4 1,5 39,0 Bột đậu nành 11,8 46,3 1,3 5,0 - - Bột ngô 13,5 9,5 4,0 4,0 1,5 67,5 Ngô hạt 10,4 4,6 7,8 3,5 1,0 72,7 Hạt cao lương 10,0 9,0 2,8 3,0 0,1 75,1 *: Không có số liệu
Cá hương của Catla, Rohu và Mrigal có kính thước từ 5-10mm sử dụng chủ yếu tảo
đơn bào, đến khi kích thước đạt 10-20mm thì ăn động và thực vật phù du có kích thước nhỏ. Để giảm giá thanh, nên sử dụng các nguyên liệu sẵn có ở địa phương như nhộng tằm, bột đậu nành (Chakraborthy và cộng sự 1973), cá và bột đầu tôm (Mahajan và Yadav 1974), protein thực vật và gạo (Chakraborthy và Kar, 1975), bột tảo trộn với bột cá (Singh và Bhanot, 1988) và bột của bèo Nymphoides và Spirodella (Patnaik và Das, 1979).
Nuôi cá bột và hương bằng thức ăn nổi được chế biến từ cám gạo, đậu lạc hoặc tép. Các nguyên liệu này được trộn đều, nấu chín và rây qua dụng cụ có kích cỡ mắt lưới là 300 -400µm và cho ăn vài lần trong 1 ngày sau khi trứng cá trôi nở (Alikunhi, 1987). Trong những nghiên cứu gần đây, bột lá của cây dâu tằm, cây keo dậu (Leucena leucocephala) và nhộng tằm sấy khô đã được đánh giá như là các nguyên liệu tốt cho thức ăn cá trôi hương (Vijayakumaraswamy và Devaraj, 1994). Khẩu phần thức ăn cho cá trôi hương được chế biến từ khô đậu lạc, cám gạo với bột cá (Swamy và cộng sự, 1988; Mohanty và cộng sự, 1990). Cá Catla hương cũng được nuôi các khẩu phần khác nhau từ bột đậu nành, bột cá và cám gạo (Mondal và cộng sự, 2000) có tỷ lệ sống cao hơn và FCR tốt hơn khi nuôi với thức ăn công nghiệp chứa 39% protein.
Công thức của hỗn hợp thức ăn được trình bày ở bảng 13.13. Hỗn hợp thức ăn brood-stock này chứa 25-30% protein thô được phối trộn từ
những nguyên liệu sẵn có tại địa phương, là những nguyên liệu ít đắt tiền và cho ăn 2% khối lượng cơ thể/ ngày. Khẩu phần này giúp cho cá trôi Ấn Độ thành thục nhanh và tăng khả năng sinh sản sau 1-2 tháng. Hơn nũa, khẩu phần này còn làm tăng sức sinh sản và chất lượng trứng tốt hơn.

128
Bảng 13.13. Tỷ lệ phối trộn các loại thức ăn trong hỗn hợp thức ăn brood-stock
Thành phần nguyên liệu Tỷ lệ phối trộn % Cám gạo 25 Khô dầu lạc 25 Bột cá 10 Ngô 10 Gạo vỡ 10 Đậu ngựa 10 Đậu đen 10 Vitamin và muối khoáng <1
Hormone thường được sử dụng trong kích thích sinh sản ở cá trôi. Một số hormone tương tự với mức sử dụng thấp trong khẩu phần lại nâng cao khả năng sinh trưởng của cá. Các loại hormon, như 17α-methyl-testosterone, khi phối hợp 2,5- 5 ppm trong khẩu phần cho cá chép (Basavaraja và cộng sự, 1989) và 1ppm cho cá Catla và Rohu (Deb và Varghese, 1988), tăng sinh trưởng và sức sống. Sử dụng những loại hormone khác, như 19-norethyl-stillboestrol ở cá chép (Nanjundappa và Vargadhara, 1989), hormone chorionic gonadotropic ở người với mức 5ppm cho cá trôi (Keshavanath và Matty, 1994) và 15 µg/g thức ăn cho cá Labeo fimbriatus (Jayaprakas và Sherly, 1996), làm tăng tốc độ sinh trưởng so với lô đối chứng. Tuy nhiên, hiện nay vấn đề sử dụng hormone cũng như kháng sinh trộn vào thức ăn nhằm tăng tốc độ sinh trưởng và phòng bệnh không được khuyến khích (một số loại bị cấm sử dụng) vì mục đích an toàn vệ sinh thưc phẩm và sức khỏe của con người
Phương pháp cho ăn phổ biến nhất trong nuôi cá trôi Ấn Độ là cho ăn bằng tay (cho ăn thủ công), đặc biệt là những trại nuôi cá quy mô nhỏ. Thức ăn được cho ăn mỗi ngày một lần với tỷ lệ dao động từ 2 -4% khối lượng thân. Cách cho ăn: trộn 1/2 độ ẩm với nhiều kích thước và hình dạng được vãi khắp bề mặt ao hay cho thức ăn vào túi treo. Thức ăn được cho vào trong một túi nhựa rỗng, có một vài lỗ (đường kính 1- 2 cm) ở đáy túi. Túi cho thức ăn đầy được treo vào một cọc tre, rồi được nhấn xuống nước, vì vậy mà phần đáy của túi nằm dưới mặt nước. Toàn bộ thức ăn sau khi đã được tiêu thụ hết trong 2-3 giờ. Sau khi hết thức ăn, túi được đưa lên, rửa sạch và phơi khô trước khi sử dụng cho ngày kế tiếp.

129
13.3. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ RÔ PHI (OEROCHROMIS SPP.)
13.3.1. Giới thiệu Cá rô phi là loài cá chủ yếu sống trong ao hồ và thích nghi tốt với môi trường nước
tù đọng. Đây là loài cá có tốc độ sinh trưởng cao, chống chịu tốt với bệnh tật và quá trình vận chuyển, dễ dàng tái sản xuất quần đàn ngay cả trong điều kiện nuôi nhốt và có thể chịu đựng những biến đổi lớn của điều kiện môi trường. Cá rô phi được nuôi rộng rãi ở vùng nhiệt đới và cận nhiệt đới và là nhóm cá nuôi đứng thứ 3 về số lượng trong các ao nuôi, với tỷ lệ tăng trưởng hàng năm vào khoảng 11,5% (El-Sayed, 1999). Sản phẩm mang tính chất toàn cầu của rô phi tăng lên hơn gấp 3 lần từ năm 1984, từ 186.544 đến 659.000 triệu tấn, chiếm 4.48% trong tổng số ao nuôi cá năm 1995 (Tacon, 1997).
Hầu hết cá rô phi nuôi đều thuộc 2 Hình 13.4. Cá rô phi giống (Trewavas, 1982): Tilapia và Oreochromis. Loài nuôi chính là Oreochromis niloticus, Oreochromis aureus, con lai O. niloticus O. aurenus, Oreochromis mossambicus và Tilapia zill. Đối với giống Oreochromis, sinh trưởng của cá rô phi cái chậm nên hình thức nuôi đơn rô phi đực được ưa chuộng hơn. Vì vậy, cần phải xác định đực cái trong nuôi cá rô phi và kiểm soát giới tính thông qua việc phân loại, chuyển đổi giới tính bằng xử lý hormone hoặc lai tạo.
Trong sự phát triển mạnh mẽ của công nghiệp và kỹ thuật công nghệ, hình thức nuôi rô phi truyền thống dần được thay thế bởi hình thức nuôi bán công nghiệp và công nghiệp. Các sinh vật tự nhiên trong ao là nguồn dinh dưỡng duy nhất cho sự phát triển của cá ở hình thức nuôi quảng canh. Ở hình thức nuôi bán thâm canh, thức ăn bổ sung bao gồm các nguyên liệu sẵn có ở địa phương, là các thực phẩm rẻ tiền, như cám gạo, bột ngô, bột cùi dừa, cà phê lững, bã bia và/ hoặc hỗn hợp của chúng, thường được sử dụng như thức ăn bổ sung cho thức ăn tự nhiên (Lim, 1989). Để tăng tỷ lệ đàn giống, thành phần thức ăn tự nhiên giảm và cần nhiều hơn thức ăn đầy đủ các thành phần dinh dưỡng. Trong hình thức nuôi bán thâm canh và nuôi thâm canh, thức ăn là thành phần chiếm tỷ lệ chi phí cao nhất, dao động từ 30% đến 60% trong toàn bộ chi phí. Vì vậy, việc sử dụng giá tối thiểu, đảm bảo cân bằng dinh dưỡng và quản lý tốt việc cho ăn là 2 yếu tố quan trọng nhât quyết định đến sự thành công của việc nuôi cá. 13.3.2. Nhu cầu dinh dưỡng
Protein và các amino acid. Protein là chất dinh dưỡng không thể thiếu trong cấu trúc và chức năng của tất cả các cơ thể sống, bao gồm cả cá rô phi. Bởi vì, protein luôn luôn cần để sử sụng cho duy trì, sinh trưởng và tái sản xuất, là nguồn cung cấp liên tục các amino acid. Cá không có nhu cầu chính xác về protein, nhưng nó cần hỗn hợp cân bằng các amino acid thiết yếu và không thiết yếu. Việc thiếu hụt protein khẩu phần có thể dẫn đến ngưng trệ hoặc làm giảm quá trình sinh trưởng của cá và đưa đến việc giải phóng dần dần protein tích luỹ ở các mô để duy trì các chức năng sống của chúng. Nếu protein được cung cấp quá nhiều, chỉ một phần sẽ được sử dụng cho các phản ứng sinh hoá tạo mô mới và phần còn lại chuyển hoá trở lại thành năng lượng (NRC, 1983).

130
Nhiều nhà nghiên cứu đã sử dụng thức ăn tinh chế và bán tinh chế để đánh giá mức protein khẩu phần tối ưu của cá rô phi. Mức protein khẩu phần tối ưu của cá rô phi phụ thuộc vào kích cỡ, tuổi và dao động từ 28% đến 50% (bảng 13.14). Đối với cá hương, protein khẩu phần dao động từ 36 đến 50% cho tăng trưởng tối đa (Davis và Stickney, 1978; Jauncey và Ros, 1982; Santiago và Laron, 1991; El-Sayed và Teshima, 1992); đối với cá giống, 29-40% protein khẩu phần cho tăng trưởng tối ưu (Cruz và Laudensia, 1977; Teshima và cộng sự, 1978; Mazid vac, 1979; Kubaryk, 1980; Jauncey, 1982; Jauncey và Ross, 1982; Siddiqui và cộng sự, 1988; Wee và Tuan, 1988; Twibell và Brown, 1998).
Bảng 13.14. Nhu cầu protein của cá rô phi nuôi ở nước ngọt (% khẩu phần)
Giống Kích cỡ cá (g) Nhu cầu Nguồn 1,0-2,5 29-38 Cruz và Laudencia (1977) Cá bột 50 0,5-1,0 40 6-30 30-35
Jauncey và Ross (1982) O. mossambicus
1,8 40 Jauncey (1982) 1,5-7,5 36 Kubaryk (1980) 3,2-3,7 30 Wang et all (1085) 0,838 40
40 30 Siddiqui et al (1988)
24 27,5-35 Wee và Tuan (1988)
O. niloticus
0,012 45 El-Sayed và Teshima (1992) 0,3-0,5 36 Davis và Stickney (1978) O. aureus 0,16 40 Santiago và Laron (1991)
1,7 35-40 Teshima et al (1978) Tilapia zillii 1,65 35 Mazid et al (1979) 0,6-1,1 32 Shiau và Peng (1993) O.niloticus x O.aureus 21 28 Twibell và Brown (1998)
Khẩu phần ăn của cá rô phi thường chứa 25-35% protein thô. Trong ao nuôi, tuy
nhiên, cá có thể ăn thức ăn tự nhiên giàu protein; vì vậy mức protein trong khẩu phần giảm xuống 20-25% đã đảm bào đủ nhu cầu (Newman vac, 1979; Lovell, 1980; Wannigama và cộng sự, 1985).
Độ mặn của nước cũng ảnh hưởng đến nhu cầu protein, nhu cầu này sẽ thấp đi nếu độ mặn càng cao (bảng 13.15).
Bảng 13.15. Nhu cầu protein của cá rô phi nuôi ở các độ mặn khác nhau
Giống Cỡ cá (g)
Độ mặn (ppt)
Nhu cầu (%)
Nguồn
0 30,4 5 30,4 10 28,0 O. niloticus 0,024
15 28,0
De Silva và Perera (1985)
O.niloticus x O.aureus 2,88 32-34 24,0 Shiau và Huang (1989) Rô phi đỏ Florida 10,6 37 20,0 Clark, A.E. et al (1990)

131
Cá rô phi cũng có nhu cầu về 10 amino acid thiết yếu (arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalamine, threonin, tryptophan và valine) như các loài cá khác và các động vật ở cạn. Số lượng nhu cầu các amino acid thiết yếu cho sự phát triển của cá rô phi Nile nhỏ (O. niloticus) được xác định bởi Santiago và Lovell (1988) qua bảng 13.16.
Bảng 13.16. Nhu cầu các amino acid thiết yếu của cá rô phi
Amino acid Nhu cầu (% protein khẩu phần) Arginine 4,20 Histidine 1,72 Isoleucine 3,11 Leucine 3,39 Lysine 5,12 Methionine* 2,68 Phenylalanine** 3,75 Threonine 3,75 Tryptophan 1,00 Valine 2,80 * Tổng số nhu cầu amino acid gốc Sulfua (Methionine + Cystein) là 3,21% protein khẩu phần ** Tổng số nhu cầu amino acid thơm (Phe + Tyrosine) là 5,54% protein khẩu phần
Các amino acid không thiết yếu có thể được cá tổng hợp nhưng sự có mặt của
chúng trong khẩu phần vẫn có ý nghĩa dinh dưỡng bởi vì giảm được nhu cầu tổng hợp chúng. Hai ví dụ điển hình dễ giải thích là chuyển đổi của methionine thành cysteine và phenylalanine thành tyrosine. Những AA không thiết yếu này chỉ có thể được tổng hợp từ những AA thiết yếu tiền thân (NRC, 1983). Cá rô phi thực ra vẫn có nhu cầu về các amino acid nhân sulfua nhưng có thể thỏa mãn hoặc chỉ cung cấp methionine hoặc cung cấp hỗn hợp thích hợp methionine và cystein. Cystein khẩu phần có thể thay thế hơn 50% tổng số nhu cầu amino acid chứa lưu huỳnh đối với O. mossambicus (Jauncey và Ross, 1982). Một quan hệ tương tự tồn tại giữa các amino acid thơm, sự hiện diện của Tyrosine trong khẩu phần sẽ làm giảm nhu cầu về phenylalanine (NRC, 1993).
Nhìn chung, các nguồn protein có chứa các AA thiết yếu với hàm lượng gần với nhu cầu AA thiết yếu là những thức ăn có giá trị dinh dưỡng cao. Bột cá được sử dụng như nguồn protein chính trong khẩu phần thức ăn thuỷ sản. Đã có những cố gắng nhằm thay thế từng phần hoặc toàn bộ bột cá với chi phí thấp nhất bằng nguồn protein có sẵn ở địa phương cho cá rô phi. Bột đậu nành đã được nghiên cứu rộng rãi và có nhiều mức độ thành công khác nhau. Các yếu tố, như kháng dinh dưỡng (Tacon, 1993), amino acid giới hạn (Jackson và cộng sự, 1982; Tacon và cộng sự, 1983; Viola và Arieli, 1983; Téhima và Kanazawa, 1988; Shiau và cộng sự,1989), muối khoáng (Viola và cộng sự, 1986, 1988), mức nhu cầu protein (Shiau và cộng sự, 1989; Viola và cộng sự, 1994) đã được nghiên cứu và thảo luận. Một đánh giá toàn diện về việc chọn lựa nguồn protein khẩu phần, bao gồm các sản phẩm phụ động vật, dầu thực vật, thực vật thuỷ sinh, protein đơn bào, hạt rau màu và protein thực vật, đối với cá rô phi cũng đã được thực hiện (El-Sayed, 1999).
Năng lượng. Năng lượng không phải là dinh dưỡng nhưng nó là thuộc tính của dinh dưỡng được tạo thành trong suốt quá trình oxy hoá protein, carbohydrate và lipid. Thông thường, protein dược coi là ưu tiên hàng đầu trong thiết lập khẩu phần cho cá, vì nó là

132
thành phần đắt nhất trong số các nguyên liệu phối hợp khẩu phần. Tuy nhiên, năng lượng là yếu tố dinh dưỡng được chọn lựa đầu tiên khi cá rô phi cũng như các loại cá khác ăn vào để thỏa mãn nhu cầu năng lượng của chúng. Nếu như luôn thiếu năng lượng phi protein thì một phần của protein sẽ được sử dụng như là nguồn cung cấp năng lượng. Tuy nhiên, thừa năng lượng thì sẽ tạo thành mỡ cá, làm giảm sự thu nhận khẩu phần ăn (giảm tổng lượng protein ăn vào) và hạn chế mức sử dụng thích hợp các loại thức ăn khác. Kubaryk (1980) quan sát thấy rằng, khi năng lượng tiêu hóa (DE) trong khẩu phần tăng thì lượng ăn vào của cá rô phi giảm, còn số lượng protein trong khẩu phần thì đã không ảnh hưởng đến lượng ăn vào.
Nhu cầu về năng lượng của cá rô phi được đưa ra dưới dạng năng lượng tổng số hay còn gọi là năng lượng thô (GE), năng lượng tiêu hóa (DE) hay năng lượng trao đổi (ME) trong sự tương quan với mức protein khẩu phần (bảng 13.17). Nhìn chung, tỷ lệ protein/năng lượng (P/E) được yêu cầu cho tăng trưởng tối đa của cá sẽ giảm khi khối lượng cơ thể cá càng tăng. Đối với cá bột (cho đến 0,5g) và cá hương (từ 0,5g đến 5g), tỷ lệ P/E là 95,3-123 mgP/Kcal DE, đối với cá trưởng thành (từ 5-50g) là 99,48 - 108 mg P/Kcal DE. Shiau và Huang (1990) kết luận rằng tăng trưởng của cá rô phi O.niloticus nuôi nước biển (độ mặn 32-34ppt) đạt tối đa khi protein khẩu phần là 21 và 24% với tỷ lệ P/ME là 67,74 và 104,35 mg P/Kcal ME.
Bảng 13.17. Tỷ lệ Protein/Năng lượng tối thích cho cá rô phi
Giống cá rô phi Cỡ cá (g) P/E Nguồn 1,80 116,6 mg P/Kcal ME Jauncey (1982) O. mossambicus 5,19 99,48 mg P/Kcal DE El-Dahhar và Lovell (1995) 0,012 110 mg P/Kcal GE El-Sayed và Teshima (1992) O. niloticus 1,70 120 mg P/Kcal DE Kubaryk (1980) 2,50 123 mg P/Kcal DE Winfree và Stickney (1981) O. aureus 7,50 108 mg P/Kcal DE Winfree và Stickney (1981) 1,65 95,3 mg P/Kcal DE Mazid et al (1979) Tilapia zillii 50,00 103 mg P/Kcal DE El-Sayed (1987)
O. niloticus x O. aureus
0,16 111 mg P/Kcal DE Santiago và Laron (1991)
Lipit và acid béo. Lipid là chất dinh dưỡng cần thiết được sử dụng để giảm chi phí
khẩu phần ăn và tối ưu hóa tích lũy nitơ. Lipid là nguồn axit béo thiết yếu cần cho sự tăng trưởng và phát triển bình thường của cá. Chúng cũng là chất mang quan trọng và hỗ trợ cho việc hấp thu các vitamin hòa tan trong mỡ. Lipid, đặc biệt là phospholipid, có vai trò quan trọng trong khả năng linh hoạt và tính thấm của màng và cấu trúc tế bào.
Tăng khẩu phần lên đến 15% đã làm cải thiện một cách có ý nghĩa tỷ lệ hiệu dụng protein (Protein Efficiency Ratio - PER) và giá trị sản xuất protein (Protein Production Values – PPV) của cá Tilapia zillii (Teshima, 1978, El-Sayed và Garling, 1988). Teshima (1985) cũng tìm thấy kết quả tương tự khi nghiên cứu trên đối tượng cá O. Nilotcus. Hanley (1991) cho rằng, cá rô phi có thể tích trữ một lượng lipid có ý nghĩa ở thân thịt và nội tạng của chúng nhưng không thể sử dụng nguồn năng lượng này cho tăng trưởng. Tuy nhiên, thí nghiệm này được tiến hành ‘ngoài trời’ nên năng suất tự nhiên trong hệ thống nuôi có thể dã ảnh hưởng tới kết quả nghiên cứu. Lipid khẩu phần có tác dụng là tiết kiệm việc sử dụng protein, mức protein khẩu phần của cá O. Niloticus có thể giảm từ 33,2%

133
xuống 25,7% bằng cách tăng lipid từ 5,7 đến 9,4% và bột đường từ 31,9% lên 36,9% (Li, 1991). Mức lipid khẩu phần vượt quá 12% sẽ kìm hãm tốc độ sinh trưởng của con lai O. aureus và O. noliticus (Jauncey và Ross, 1982). Sự phát triển của O. aureus có thể được cải thiện một cách chắc chắn khi bổ sung dầu cá mòi hoặc dầu cá 7,5-10% khẩu phần so với mức lipid khẩu phần thấp. Tuy nhiên, hiệu suất tốt nhất đạt được đối với dầu cá mòi là 10% khẩu phần (Stickney và Wurts, 1986). Mức lipid tối ưu trong khẩu phần cá rô phi được xác định bởi Chou và Shiau (1996). Các chất 5-isoenerdetic và isonitrogenous được tinh chế từ khẩu phần chứa 0-20% lipid (dầu ngô, dầu gan cá và mỡ lợn ở tỷ lệ 1 :1 :1) với 5% lượng phụ gia dùng để nuôi con lai giữa O. niloticus và O. aureus giai đoạn cá con. Kết quả nghiên cứu cho thấy rằng, 5% lipid khẩu phần đủ cho nhu cầu tối thiểu của các rô phi giai đoạn chưa trưởng thành, nhưng mức 12% mới cho kết quả sinh trưởng tối ưu.
Cá sống trong môi trường nước lạnh có nhu cầu về n-3 acid béo mạch dài không no (PUFA) cao hơn, trong khi cá ở vùng nước ấm hướng đến nhu cầu về n-6 acid béo. Một vài nghiên cứu cho thấy rằng nhu cầu acid đối với nhiều loài cá rô phi là khác nhau. Ranozawa và cộng sự (1980) đã chỉ ra rằng T.zilli có khả năng biến đổi nhiều 18:2n-6 thành 20:4n-6. Tuy nhiên, Kanazawa và cộng sự (1980) đề nghị rằng những loài gần nhau chắc chắn biến đổi được 18:2n-6 thành 20:4n-6. Stickney và McGeachin (1985) đã chỉ ra rằng sự phát triển của cá rô phi xanh, O. aureus, không bị ảnh hưởng với mức khẩu phần chứa 2% linoleic acid. Khi cá rô phi xanh ăn thức ăn chứa dầu đậu nành, sự phát triển của nó được cải thiện theo tỷ lệ tăng của linoleic acid (Stickney và cộng sự, 1985). Takeuchi và cộng sự (1983) đã nhận thấy rằng tốc độ sinh trưởng của cá rô phi O. aureus giảm đáng kể khi sử dụng thức ăn chưa dầu cá (pollock liver oil) so với dầu ngô hoặc dầu đậu nành. Ramachandran Nair và Gopakumar (1981) đã chứng minh có hàm lượng cao 22:6n-3 trong trứng cá O.mossambica, khuyến cáo một vài vai trò quan trọng của docosahexaenoic acid trong giai đoạn phát triển phôi. Tuy nhiên, Santiago và Reyes (1993) cũng chỉ ra rằng, mặc dù dầu cá (cod-liver oil) có 22:6n-3 cao thúc đẩy tăng trọng tối đa đối với cá rô phi sông Nile, nhưng khi sử dụng dầu tương tự với dầu cá này thì đã cho kết quả giảm chất lượng sinh sản. Cá rô phi xanh có tốc độ sinh trưởng tốt khi sử dụng thức ăn chứa 10% dầu đậu nành, với tỷ lệ 20:5n-3 và 22:6n-3 cao (Stickney và McGeachin, 1983). Tác giả Stickney và Hardy (1989), tuy nhiên, đã cho rằng nhu cầu của O. aureus có thể giảm khi có mặt n-3 acid béo. Stickney và Wurts (1986) khi so sánh khẩu phần chứa các mức dầu cá da trơn và dầu mendahen khác nhau đã thấy rằng, sự phát triển tốt nhất ở cá rô phi sử dụng 10% dầu menhaden khẩu phần. Những nghiên cứu này đã cho những kết quả trái ngược nhau về nhu cầu n-3 và n-6 PUFA đối với cá rô phi. Gần đây, Chou và Shiau (1999) đã chứng minh rằng cả n-3 và n-6 acid béo mạch dài chưa no đều đóng vai trò quan trọng trong phát triển tối đa của cá lai (O.niloticus ×O. aureus). Cho đến nay, việc tiếp tục nghiên cứu để xác định số lượng của acid béo thiết yếu cho nhu cầu của các loài cá rô phi và cá lai là cần thiết.
Carbohydrate. Carbohydrate là thành phần rẻ tiền nhất trong năng lượng khẩu phần thức ăn của con người và vật nuôi, nhưng việc sử dụng chúng có sự thay đổi đối với động vật thuỷ sản và còn cần phải nghiên cứu thêm. Cá nói chung sử dụng kém nguồn carbohydrate khẩu phần. Nhu cầu về carbohydrate ở cá vẫn chưa được chứng minh. Việc sử dụng tinh bột cho kết quả tăng trưởng cao hơn một cách có ý nghĩa so với sử dụng glucose đối với cá rô phi (Anderson,1984; Tung và Shiau, 1991; Shiau và Chen, 1993; Shiau và Lin, 1993). Báo cáo cũng chỉ ra rằng cá rô phi sử dụng disacharide tốt hơn gluco nhưng lại kém hơn tinh bột. Đối với disacharide thì mức độ hấp thu tốt nhất là đường

134
maltose tiếp đến là sucrose và lactose (Shiau và Chuang,1995). Báo cáo của Lin và Shiau (1995) cho rằng enzyme malic, glucose-6-phosphat dehydrogennase (G-6-PD) và phosphogluconate đehdrogenase (PGD) hoạt động cao hơn trong khẩu phần thức ăn tinh bột hơn là khẩu phần thức ăn chứa glucose. Việc thay đổi thức ăn từ tinh bột sang gluco sẽ làm tăng lượng enzyme malic, G-6-PD và PGD hoạt động, trong khi đó việc thay đổi thức ăn chứa glucose sang tinh bột sẽ làm tăng hoạt động của các enzym trong gan cá. Các tác giả này cho rằng hoạt động của enzyme lipogenic ở gan cá rô phi có thể hấp thu được carbohydrate khẩu phần.
Tung và Shiau (1991) đã nghiên cứu ảnh hưởng của việc sử dụng carbohydrate hằng ngày đối với cá rô phi. Khẩu phần chứa 44% glucose dextrin và tinh bột được sử dụng cho cá ăn 3 hoặc 6 lần/ngày. Tác giả nhận thấy rằng cá sử dụng nguồn carbohydrate 6 lần/ngày cho tăng trọng cao hơn, hiệu quả sử dụng protein khẩu phần, protein và năng lượng tích luỹ một cách có ý nghĩa hơn là nuôi 2 lần/ ngày. Nghiên cứu này cũng đã chứng minh rằng, nếu việc cho ăn được tăng từ 2 lần/ ngày lên 6 lần/ ngày, thì cá ăn tinh bột và dextrin cho tăng trưởng tốt hơn là ăn glucose, nhưng sự khác nhau này không có ý nghĩa thống kê. Nó cũng chỉ ra rằng tốc độ tăng trưởng của cá rô phi ăn thức ăn chứa glucose 6 lần/ngày cao hơn cá ăn tinh bột hoặc dextrin 2 lần / ngày. Hiệu quả của chiến lược cho cá rô phi ăn carbohydratee đã được Shiau và Lei (1999) xác nhận gần đây, khi họ đánh giá lại hiệu quả của việc cho ăn carbohydratee liên tục và 2 bữa/ ngày đối với cá rô phi. Kết quả này cho thấy việc cho ăn thường xuyên trong ngày tăng hiệu quả sử dụng carbohydrate.
Shiau và cộng sự (1988) đã nghiên cứu ảnh hưởng của carboxymethycellulose (CMC) lên tốc độ tăng trưởng, tiêu hóa và thời gian rỗng dạ dày của cá rô phi. Trong nghiên cứu này, CMC được bổ sung với mức 2, 6, 10 và 14% khẩu phần. Dextrin cũng được sử dụng như một nguồn carbohydrate. Cả việc tăng trọng lượng và tiêu tốn thức ăn đều bị giảm khi tăng mức CMC khẩu phần. Shiau và cộng sự (1989) đã nghiên cứu ảnh hưởng của 5 khẩu phần dạng sợi (guar gum, agar, carrageenan, CMC và cenllulose) đối với việc sử dụng dextrin và gluco ở cá rô phi. Mỗi dạng sợi được bổ sung 10% vào khẩu phần. Kết quả cho thấy rằng trọng lượng tăng lên chỉ có ít ý nghĩa đối với cá rô phi ăn thức ăn có thành phần dạng sợi hơn là thức ăn có thành phần dextrin hoặc glucose. Việc hấp thụ carbohydrate ở ruột thấp hơn so với cá ăn thức ăn có thành phần dạng sợi, dù là loại thức ăn dạng sợi nào. Gần đây, Shiau và Yu (1999) đã chứng minh rằng việc bổ sung chitin hoặc chitosan làm giảm tốc độ tăng trưởng cá rô phi dù là bổ sung ở mức độ nào (2 - 5 và 10%).
Niacin là tiền chất của một vài coenzyme cần cho quá trình chuyển hoá carbohydrate. Shiau và Suen (1992) đã chứng minh được số lượng niacin cần cho cá rô phi thay đổi theo nguồn carbohydrate trong khẩu phần. Họ chỉ ra rằng mức niacin thích hợp đảm bảo cho cá rô phi giống tăng trưởng tối đa là 26mg/kg thức ăn ở cá ăn thức ăn chứa gluco và 121mg/kg ở cá ăn thức ăn chứa dextrin, và cũng chỉ ra rằng nhu cầu niacin trong thức ăn của cá rô phi phụ thuộc vào dạng carbohydrate trong khẩu phần.
Shiau và Peng (1993) đã tiến hành nghiên cứu nhằm đánh giá hiệu quả về sử dụng tiết kiệm protein của carbohydrate ở cá rô phi. 3 khẩu phần protein với mức (32, 28, 24%) đã đạt được bằng việc thay thế 3 mức (33, 34, 41%) và 3 nguồn (glucose, dextrin, tinh bột) của carbohydrate khẩu phần. Những kết quả nghiên cứu cho thấy hiệu quả tiết kiệm sử dụng protein của carbohydrate (dextrin hoặc tinh bột) chỉ xảy ra khi mức protein khẩu phần dưới mức tối ưu. Shiau và Liang (1995) đã báo cáo rằng việc sử dụng carbohydrate của cá rô phi chịu ảnh hưởng của nguồn protein trong khẩu phần thức ăn.

135
Những chất dinh dưỡng khác, như chromium, cũng được báo cáo về tác động của việc sử dụng carbohydrate cho cá. Shiau và Lin (1993) lần đầu tiên chứng minh rằng việc bổ sung chromium trong công thức thức ăn của chromium chloride đã cải thiện có ý nghĩa về việc sử dụng gluco nhưng không cải thiện đối với việc sử dụng tinh bột ở cá rô phi. Shiau và Chen (1993) cũng có những báo cáo ngay sau đó về việc cải thiện việc sử dụng gluco bởi cá rô phi ăn thức ăn bổ sung chromic oxide trong việc sử dụng các khẩu phần chứa gluco thay đổi. Cá ăn gluco khẩu phần chứa 5g chromic oxide/ kg thức ăn thì tăng trọng tốt hơn cá sử dụng 20 g chromic oxide/ kg thức ăn. Họ nhận xét, việc sinh trưởng ở cá rô phi càng chậm khi ăn khẩu phần chứa lượng chromic oxide càng cao có thể là do ảnh hưởng của độc tố có trong chromium khẩu phần. Gần đây, báo cáo của Shiau và Shy (1998) cho thấy sự sinh trưởng tốt đa và việc sử dụng gluco ở cá rô phi lai sẽ đạt được với mức khẩu phần chứa 204,4 mg chromic/ kg thức ăn.
Lin và cộng sự (1995) đã chỉ ra rằng nồng độ isulin huyết tương ở cá rô phi đã ăn vào với glucose là cao hơn so với ăn vào với tinh bột và cơ cấu insulin huyết tương của cá rô phi tương tự như ở người. Tuy nhiên, sự chuyển hoá insulin trong việc sử dụng carbohydrate ở cá rô phi vẫn chưa được làm sáng tỏ.
Amonia thải ra và việc tiêu thụ oxy của cá rô phi chịu tác động bởi dạng carbohydrate ăn vào. Shiau và Cheng (1999) cho cá rô phi ăn khẩu phần chứa 3 mức carbohydrate (gluco và tinh bột) (33, 37 và 41%) trong 8 tuần ; cá ăn thức ăn chứa tinh bột thải ra ít amonia hơn ăn thức ăn chứa gluco ở cả ba mức đó. Amonia thải ra trong thức ăn chứa glucose giảm khi mức carbohydrate tổng số tăng. Ở cá ăn thức ăn tinh bột, amonia thải ra là thấp hơn ở mức 41% hơn là 33 và 37%. Tổng số oxy tiêu thụ là cao hơn ở cá ăn thức ăn chứa tinh bột hơn so với cá ăn thức ăn chứa gluco. Trong cả 2 trường hợp, lượng oxy tiêu thu cao hơn đã được quan sát ở mức 37% và 41% hơn là mức 33%.
Các yếu tố sinh học có thể ảnh hưởng tới việc sử dụng carbohydrate ở cá rô phi. Tung và Shiau (1993) đã so sánh việc sử dụng 2 loại carbohydrate (glucose, tinh bột) cho 2 kích cỡ cá rô phi khác nhau (4.55 và 0.46g). Tác giả đã nhận thấy rằng cá có kích cỡ lớn tăng trọng nhanh hơn cá có kích cỡ nhỏ khi ăn thức ăn có chứa gluco. Tuy nhiên, con số này lại tương đương ở cá lớn lẫn cá nhỏ hơn khi ăn tinh bột.
Vitamin và muối khoáng. Việc bổ sung vitamin thường không bao gồm thức ăn dùng trong ao nuôi cá giống có bón phân với mật độ vừa phải. Ở hệ thống nuôi thâm canh hạn chế hoặc không có nguồn thức ăn tự nhiên thì cần phải bổ sung vitamin.
Bảng 13.18 trình bày nhu cầu vitamin ở các đối tượng rô phi khác nhau. Nhu cầu vitamin hoà tan trong nước của cá rô phi đã được xác định như sau: Thiamine, riboflavin, pyridoxine, vitamin B12, niacin, biotin, choline, pantothenic acid và ascorbic acid. Việc thiếu hụt thiamin ở cá rô phi đỏ giống (O.mossambicus × O. niloticus) nuôi nước biển (32 p.p.t độ muối) đã làm giảm quá trình sinh trưởng và hiệu quả sử dụng thức ăn. Mức thiamine cần bổ sung trong khẩu phần là 2.5 mg/ kg thức ăn đảm bảo cho sự tăng trưởng tối đa và ngăn ngừa các biểu hiện do thiếu thiamine. (Lim và LeaMaster, 1991). Dấu hiệu đặc trưng do thiếu riboflavin ở cá rô phi ăn khẩu phần không có riboflavin là chán ăn, chậm lớn, tỷ lệ chết cao, mòn vây, màu sắc cơ thể khác thường, còi cọc, đục thể thuỷ tinh. Nhu cầu riboflavin trong khẩu phần là 6 mg/ kg thức ăn cho cá O. aureus chưa trưởng thành phát triển trong môi trường nước ngọt (Soliman và Wilson, 1992) và 5mg/kg thức ăn cho cá điêu hồng (O. mossambicus × O.niloticus) nuôi ở 32 p.p.t độ muối (Lim et al., 1993).

136
Bảng 13.18. Nhu cầu vitamin ở các đối tượng rô phi khác nhau (mg/kg thức ăn)
Vitamin O.niloticus x O. aureus
O. aureus O.niloticus O.mossambicus x O.niloticus
Thiamine - - - 2,5 Riboflavin - 6 - 5 Pyridoxin 1,7-9,5 (28%CP) - - 3 15-16,5 (36%CP) Vitamin B12 Không cần thiết - Không cần - Niacin 121 - - - Biotin 0,06 - - - Folic acid - - - - Inositol - - - - Choline 1000 Không cần - - Pantothenic acid - 6-10 - - Vitamin A - - - - Vitamin D 0,00937 Không cần - - (374,8 IU/kg) Vitamin E 42-44 (5%lipid) 10 (3%L) 50-100(5%L) - 60-66 (12%lipid) 25 (6%L) 500 (10-15%L) - Viatmin K - - - -
Cá ăn thức ăn không bổ sung pyrodoxine sẽ xuất hiện những dấu hiệu thần kinh bất bình thường, bỏ ăn, co giật, mòn vây, tổn thương miệng, chậm lớn và tỷ lệ chết cao. Tăng trọng và hoạt động của enzyme vận chuyển axit amin alanine gan có tương quan phi tuyến tính, chứng tỏ nhu cầu pyridoxine tối ưu trong khẩu phần của cá lai O.niloticus ×O. aureus giai đoạn chưa trưởng thành nuôi ở nước ngọt là 1.7-9.5 mg/ kg thức ăn và 15.0-16.6 thức ăn chứa 28% và 36% protein theo thứ tự (Shiau và Hsied, 1997). Mức pyroxidine trong khẩu phần là 3 mg/ kg thức ăn đã được xác định đối với con lai O.mossambicus × O. niloticus sử dụng thức ăn chứa 38% protein và nuôi trong môi trường nước biển. Cá rô phi có khả năng sản xuất ra vitamin B12 trong ống tiêu hoá thông qua quá trình chuyển hoá của vi khuẩn nên không cần cung cấp thêm vitamin này qua thức ăn (Lovell và Limsuwan, 1982; Sugita và cộng sự, 1990; Shiau và Lung, 1993).
Niacin là một thành phần quan trọng trong khẩu phần đối với cá lai O.niloticus ×O. aureus nhưng nhu cầu thay đổi phụ thuộc vào nguồn carbohydrate cung cấp. Mức tối ưu trong khẩu phần cho sinh trưởng tối đa đã được nghiên cứu là 26 mg/kg thức ăn cho cá ăn thức ăn chứa glucose và 121 mg/ kg thức ăn cho cá ăn thức ăn chứa dextrin. Cá thiếu niacine có hiện tượng xuất huyết, miệng biến dạng, mang hoại tử và da, vây, miệng tổn thương (Shiau và Suen, 1992). Nhu cầu biotin trong khẩu phần cho cá lai O.niloticus ×O. aureu sinh trưởng tối đa là 0,06 mg/kg thức ăn (Shiau và Chin, 1999). Trong khi nhu cầu về methionine có thể được đảm bảo một phần từ choline, nhu cầu choline khẩu phần tối ưu đối với cá lai O.niloticus ×O. aureu là 1000 mg/kg thức ăn.
Khi thiếu Pantothenic acid có thể gây ra hiện tượng chậm lớn, xuất huyết, hoạt động lờ đờ, tỷ lệ chết cao, thiếu máu và tăng sản tế bào biểu bì tơ ở mang cá O. aureus. Mức 10mg calcium-pantothenate/kg thức ăn đảm bảo ngăn ngừa được các dấu hiệu do thiếu pantothenic acid (Soliman và Wilson, 1992). Tuy nhiên, O. aureus có thể đảm bảo

137
được nhu cầu pantothenic acid khi nuôi bằng vi khuẩn trong hệ thống tái tuần hoàn (Roem và cộng sự, 1990).
Cá rô phi có những dấu hiệu đặc trưng do thiếu vitamin C khi ăn khẩu phần không có thức ăn tự nhiên. Nhu cầu vitamin C đảm bào sinh trưởng bình thường đối với cá lai (O.niloticus ×O. aureus) là 50mg ascorbic acid/kg thức ăn (Stick và cộng sự, 1984). Cá giống cần 79mg ascorbic acid/kg thức ăn để đảm bảo cho sinh trưởng tối đa (Shiau và Jan,1992). Nhu cầu về vitamin C đối với O.niloticus là 420 mg/kg thức ăn (Soliman và cộng sự, 1994). Trong tất cả các nghiên cứu, L-ascrobic acid được sử dụng như một nguồn vitamin C. L-ascrobic acid (C1) dễ bị phân huỷ, nhất là qua các khâu chế biến thức ăn và mất dần trong quá trình bảo quản. Khoảng 75% lượng C1 bổ sung ban đầu trong thức ăn thủy sản có thể mất qua quá trình chế biến và bảo quản ở nhiệt độ phòng (Shiau và Hsu, 1993). Bởi vì ở dạng nguyên thuỷ, C1 dễ phân huỷ, nên việc sử dụng dạng ascorbic acid khó phân huỷ hơn là cần thiết. Một số dẫn xuất của C1 có các hoạt động đối kháng với ascorbic đối với rô phi; L-ascorbyl-2-sunphate (C2S) có hoạt động đối kháng về ascorbic với L-ascorbyl-2-monophosphate-magnesium (C2MP-Mg) đối với cá lai O.niloticus ×O. aureus. Nhu cầu để đảm bào sinh trưởng tối ưu của Oreochromis spilurus trong nước biển đã được chứng minh là trong khoảng 100 đến 200 mg C2S/ kg thức ăn (Al-Amoundi và cộng sự, 1992).
Những thông tin về vitamin hoà tan trong dầu đối với cá rô phi chỉ bao gồm vitamin D và vitamin E. Nhu cầu tối ưu về vitamin D3 (cholecalciferol) cho tốc độ sinh trưởng tối đa của con lai O.niloticus ×O. aureus l à 374.8 UI/kg thức ăn (Shiau v à Hwang, 1993). Qua báo cáo của O’Connell và Gatlin (1994) cho thấy vitamin D3 không phải là vitamin thiết yếu đối với O. aureus . Sự thiếu hụt vitamin E ở O. aureus làm xuất các biểu hiện chán ăn, giảm tăng trọng và giảm hiệu quả sử dụng thức ăn, xuất huyết dưới da, sụt giảm lượng hồng cầu, thoái hoá cơ, ceroid ở gan, lá lách và da; màu da có màu bất thường. Nhu cầu về vitamin E tăng lên theo mức tăng lipit trong khẩu phần. Nhu cầu đối với O. aureus là 10-25 mg D.L-α-tocopheryl acetate/ kg thức ăn đối với thức ăn chứa 3- 6% lipit hoặc 3-4 mg α-tocopheryl acetate cho mỗi % khẩu phần của lipit (Roem và cộng sự, 1990). Nhu cầu về vitamin E của O.niloticus được xác định là 50-100 mg/kg thức ăn đối với thức ăn chứa 5% lipit, và tăng lên 500 mg/ kg thức ăn đối với thức ăn chứa 10-15% lipit (Satoh và cộng sự, 1987).
Gần đây, các nhà khoa học đã chỉ ra nhu cầu của cá rô phi về cả n-3 acid béo mạch dài chưa no và n-6 acid béo đều cần cho quá trình phát triển tối đa (Chou và Shiau,1999). Do đó, nhu cầu về vitamin E của cá rô phi đã được đánh giá lại là 42-44 mg/kg và 60-66 mg/kg trong khẩu phần chứa 5% đến 12% lipit (Shiau và Shiau, 2001). Chưa có nghiên cứu nào về nhu cầu của cá vitamin A và K đối với cá rô phi.
Có rất ít thông tin về nhu cầu muối khoáng của cá rô phi. 5 muối khoáng chính, Ca, P, Mg, Zn và K đã được xác định về nhu cầu của chúng đối với cá rô phi (bảng 13.19). Trong nước chứa ít Ca, việc bổ sung 7,0g (7%) Ca và 5,0g (5%) P/ kg thức ăn đáp ứng đủ nhu cầu cho quá trình sinh trưởng bình thường và khoáng hoá của xương của O. aureus (Robinson và cộng sự, 1984, 1987). Nhu cầu P cho quá trình sinh trưởng tối đa và khoáng hoá bình thường của xương đối với O.niloticus là dưới 0,9% khẩu phần (Wantanabe và cộng sự, 1980). Đối với aureus, mức magnesium 0,05% (Reigh và cộng sự, 1991) và 0,002% Zn (McClain và Gatlin, 1988) đáp ứng được cho quá trình sinh trưởng tối ưu, trong khi nhu cầu của Mg là 0,059-0,077% (Dabrowska và cộng sự, 1989) và 0,003% Zn (Eid và Ghonim, 1994) đã được báo cáo đối với O. niloticus. Nhu cầu về K đối với con lai

138
O.niloticus ×O. aureus là xấp xỉ 0,2-0,3% (Shiau và Hsieh, 2001). Mức khẩu phần của Mg là 0,0012% và của đồng là 0,0003-0,0004% là nhu cầu của O. niloticus (Wantanabe và cộng sự, 1988).
Bảng 13.19. Nhu cầu muối khoáng của cá rô phi (% khẩu phần)
Muối khoáng O. aureus O. niloticus O.niloticus xO.aureus
Ca 0,7 - - P 0,5 <0,9 -
Mg 0,05 0,059-0,077 - Zn 0,002 0,003 - K - - 0,21-0,33
13.3.3. Khẩu phần thức ăn Cá rô phi đã được nuôi rất thành công với công thức thức ăn dành cho các đối
tượng khác như cá chép và cá da trơn. Công thức thức ăn chế biến thường bao gồm chỉ 3 hoặc 4 loại thức ăn, chủ yếu là thực vật. Công thức thức ăn công nghiệp đối với cá rô phi thường chứa 24-28% protein. Thức ăn tự nhiên trong ao đóng góp một phần protein đáng kể, vì vậy mức protein này được công nhận là đủ cao (Lim, 1989). Tuy nhiên, có ít nghiên cứu so sánh những công thức thức ăn khác nhau cho mô hình nuôi cá rô phi thâm canh và bán thâm canh. Thức ăn viên chứa 25% protein phối hợp từ 15% bột cá, 20% đậu nành, 20% lúa mì và 45% cao lương đã được sử dụng rất hiệu quả ở Israel trong phương thức nuôi này (Viola và Arieli, 1983). Tầm quan trọng của các yếu tố vi lượng bổ sung trong khẩu phần đối với thức ăn cho cá rô phi nuôi ao vẫn chưa được biết đến nhiều. Vì sự biến động cực lớn trong nuôi trồng, công thức phối trộn khẩu phần bổ sung chất dinh dưỡng một cách hiệu quả đối với thức ăn tự nhiên là không thể áp dụng được. Ở hình thức nuôi thâm canh, như nuôi lồng bè hoặc nuôi nước chảy, cá rô phi hoàn toàn dựa vào thức ăn chế biến như là nguồn dinh dưỡng duy nhất. Vì vậy, cần phải có khẩu phần đầy đủ tất cả các chất cần thiết với nhu cầu của cá.
Cá rô phi có thể sử dụng nhiều khẩu phần khác nhau, ở dạng bột và dạng nhão, viên chìm hay nổi. Thành phần thức ăn thô, như cám gạo, ở dạng bột, trong khi các thành phần thức ăn hầu hết được chế biến dưới dạng viên. Các loại thức ăn thô có thể không kinh tế khi tạo viên cho cá rô phi nuôi ao; tuy nhiên, khẩu phần ăn có chất lượng cao nên được chế biến thành viên để hạn chế làm tách rời các thành phần, cho ăn từng thành phần riêng rẽ, cũng như hạn chế sự hao hụt đối với các chất dinh dưỡng hoà tan trong nước và chất thải.
Cá rô phi dường như ưa thích thức ăn dạng viên nhỏ hơn cá da trơn và cá hồi có cùng kích cỡ. Kích cỡ viên thức ăn đối với cá rô phi được đề nghị bởi Jauncey và Ross (1982). Cỡ phổ biến nhất là viên có đường kính 3-4mm và dài 6-10mm. Thức ăn bột hoặc bóp vụn được sử dụng cho cá bột và cá giống. 13.3.4. Nuôi dưỡng
Nhìn chung tỷ lệ cho ăn/khối lượng cơ thể đối với cá nhỏ lớn hơn cá lớn. Tỷ lệ cho ăn theo kích cỡ cá rô phi được thể hiện qua bảng 13.20. Nhiệt độ nước cũng ảnh hưởng đến tỷ lệ chuyển hoá và tiêu hao năng lượng, vì vậy mà có ảnh hưởng lớn đến tỷ lệ cho ăn. Cá rô phi sẽ ít sử dụng thức ăn khi thời tiết lạnh hơn thời tiết ấm. Luquet (1991) khuyến cáo không nên cho cá rô phi ăn khi nhiệt độ nước xuống dưới 160C. Trong hình thức nuôi bán

139
thâm canh, thức ăn tự nhiên cũng góp phần quan trọng đối với nhu cầu dinh dưỡng của cá. Dưới điều kiện này, số lượng thức ăn được sử dụng nên ít hơn với cá nuôi thâm canh.
Bảng 13.20. Tỷ lệ và số lần cho ăn với các kích cỡ khác nhau của cá rô phi ở 28oC
Kích cỡ cá Tỷ lệ cho ăn/ngày (%P thân) Số lần cho ăn trong ngày 2 ngày tuổi đến 1 g 30 – 10 8
1 – 5 g 10 – 6 6 5 – 20 g 6 – 4 4
20 – 100 g 4 – 3 3 – 4 > 100 g 3 - 2 2 - 3
Do có nhiều yếu tố ảnh hưởng đến việc tiêu thụ thức ăn của cá, việc cho ăn tự do
(cho ăn ở mức nhiều nhất mà cá có thể ăn) có thể tốt hơn so với cho ăn hạn chế theo tỷ lệ cho ăn / trọng lượng thân như đã trình bày. Clark, J.H và cộng sự (1990) đã đạt được sinh trưởng tối đa của cá điêu hồng Florida Oreochromis urolepis hornorum× O. mossambicus nuôi lồng trên biển với tỷ lệ cho ăn gần như thoả mãn (90%). Gía thành thức ăn có thể giảm nhưng đã không làm giảm một cách có ý nghĩa đến sự tăng trưởng khi cho ăn ở 70% mức tự do.
Có thể cho cá ăn bằng tay, máy thổi, cho ăn tự động hoặc máy ăn theo nhu cầu. Cho ăn bằng tay tuy tốn lao động nhưng có thuận lợi hơn các cách cho ăn khác. Nó cho phép người cho ăn quan sát hoạt động bắt mồi và các hành vi của cá và vì vậy điều chỉnh được lượng thức ăn nhằm ngăn ngừa việc cho ăn quá mức. Tuy nhiên, phương pháp cho ăn này khó khả thi trong những trang trại quy mô lớn. Phương pháp phổ biến nhất để cho ăn đối với ao có diện tích lớn là cho thức ăn trên bề mặt phát tán theo hướng gió bằng cách sử dụng các thiết bị quạt. Cách cho ăn tự động, được điều khiển bởi đồng hồ hẹn giờ hoặc thiết bị điện, cho phép người công nhân đặt trước số lượng thức ăn để cho ăn vào các thời gian khác nhau. Máy cho ăn theo nhu cầu gồm có phễu ở đầu trên để đưa thức ăn vào và một đáy mở như một cổng di chuyển để phân phối thức ăn. Kèm với cổng là một tay đòn có đầu chìm vào nước nơi mà nó có thể được khởi động bởi cá. Khi mà cá vẫn còn tiếp tục tác động vào ống, thì thức ăn sẽ tiếp tục được chảy xuống. Clark J.H và cộng sự (1990) đã chỉ ra rằng máy ăn theo nhu cầu là phương pháp tốt nhất để cho cá rô phi ăn trong nuôi lồng ở biển đạt được sự sinh trưởng và chuyển đổi thức ăn tốt với việc giảm chi phí lao động.
Thông thường số lần cho ăn phụ thuộc vào kích cỡ cá. Cá càng lớn thì số lần cho ăn trong ngày sẽ giảm. Cho ăn 3 đến 4 lần một ngày đối với cá giai đoạn nhỏ, 2-3 lần đối với giai đoạn trưởng thành. Vì thói quen ăn liên tục và khả năng chứa của dạ dày nhỏ, cá rô phi đáp ứng tốt với việc cho ăn nhiều lần hơn là cá da trơn và cá hồi. Cá Oreochromic niloticus giống lớn nhanh khi ăn 4 lần/ngày hơn 2 lần/ngày nhưng không lớn nhanh hơn khi cho ăn 8 lần/ngày (Kubaryk, 1980).

140
13.4. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ QUẢ VÀ CÁ DA TRƠN (PANGASIUS)
13.4.1. Giới thiệu Cá quả Ophiocephalus và cá Pangasius phân bố ở Nam Phi, Ấn Độ, Burma,
Indonesia, Malaysia, Campuchia, Việt Nam và Thái Lan (Wee, 1982; Chuapohuk, 1994). Những loài cá này được nuôi phổ biến vì chúng có tốc độ sinh trưởng rất nhanh, đề kháng với dịch bệnh và có sức chịu đựng tốt khi chất lượng nước kém và bị nuôi nhốt. Ở Thái Lan, cá quả và cá Pangasius là hai loài trong số 27 loài nước ngọt được nuôi với số lượng lớn, từ thâm canh cho sản phẩm thương mại đến nuôi quảng canh cho sản phẩm tiêu thụ trong gia đình. Cả hai loài cá quả và cá Pangasius thường được nuôi trong lồng và hồ. Theo thống kê, sản lượng cá quả và cá Pangasius ước tính chiếm khoảng 15% tổng sản lượng cá nuôi nước ngọt ở Thái Lan với giá trị hàng năm 18 triệu USD (Jantrarotal, 1994).
Cá quả. Cá quả thuộc họ Channidae. Nó còn có tên gọi là cá chuối hoặc cá đầu rắn.
Cá quả có mùi vị thơm ngon và là loài cá phổ biến nhất ở Nam Á và Đông Nam Á (De Silva, 1989). Đây là loài cá có giá trị, chất lượng thịt được đánh giá là bồi bổ sức khỏe, đặc biệt là hồi phục sức khỏe cho người bị bệnh và người lớn tuổi. Cá quả bắt đầu được nuôi từ năm 1955 ở Thái Lan và mở rộng ra các nước khác bao gồm Hồng Kông, Ấn Độ, Campuchia, Đài Loan và Việt Nam. Loài nuôi phổ biến là Channa striatus. Tuy nhiên, các loài khác như Channa micropeltes (cá lóc bông) và
Hình 13.5. Cá lóc bông Channa marulius cũng đưa vào nuôi (De Silva, 1989).
Cá da trơn Pangasius. Có hai loài thuộc Pangasids được nuôi khá nhiều ở các nước Đông Nam Á: Pangasius sutchi (cá tra) và Pangasius lamaudii (black-ear catfish). Chúng được tìm thấy ở các dòng sông ở Ấn Độ, Burma, Indonesia, Malaysia, Campuchia và Thái Lan (Ling, 1977). Cá Pangasius là loài cá có tốc độ sinh trưởng nhanh và sức chịu đựng với điều kiện môi trường tốt hơn cá quả. Nó được biết nhiều như là một loài cá thể thao. Cá Pangasius có giá trị cho chế biến do thịt cá có lượng thịt cao hơn so với cá nheo Mỹ (Ictalurus punctatus). 13.4.2. Nhu cầu dinh dưỡng
Cá quả. Nhu cầu về dinh dưỡng của cá quả rất ít được biết đến. Cá quả là loài cá ăn thịt và thường ăn các loại thức ăn có nguồn gốc là động vật. Mức nhu cầu protein cao hơn so với các loài cá nước ngọt khác. Theo Boonyaratpalin (1980) thì cá quả bột ăn 3,1 kcal năng lượng tiêu hóa (DE)/g thức ăn với 43% protein, trong khi đó cá 1 tháng tuổi yêu cầu mức năng lượng như trên chứa trong thức ăn 36% protein. Nhu cầu lipid cho sinh trưởng và tồn tại ở cá hương là 6% (Boonyaratpalin, 1981). Các acid béo cần thiết cho cá chưa có thông tin nhiều. Các acid béo mạnh dài n-6 hoặc n-3 được làm giàu cho lipid và được cá quả sử dụng tốt. Chưa có nghiên cứu nào về nhu cầu đối với vitamin, ngoại trừ acid

141
pantotheic. Boonyaratpalin (1981) cho rằng khẩu phần có pantothenate là rất cần thiết cho sinh trưởng bình thường và tỷ lệ sống cao của cá quả.
Cá Pangasius. So với cá quả, nhu cầu dinh dưỡng của cá Pangasius ít được biết hơn nhiều. Cá Pangasius là một loài ăn tạp và có nhu cầu về protein thấp hơn cá quả. Jantrarotai et al (1992) cho rằng cá P.sutchi giai đoạn giống có nhu cầu 27 - 29% protein cho sinh trưởng bình thường. Chuapoehak (1994) đã tiến hành một nghiên cứu về thức ăn ở cá P.sutchi và thấy rằng lượng protein có thể giảm còn 18% nhưng không làm giảm tốc độ sinh trưởng và hệ số thức ăn nếu nguồn protein có chất lượng cao được sử dụng trong thức ăn và tỷ lệ năng lượng/protein là 13kcal/g. Trong những nghiên cứu gần đây, Chutjareyaves et al. (1998) đã kết luận hàm lượng protein 35% thích hợp cho P. lamaudii giai đoạn cá hương. Còn giai đoạn cá P.lamaudii giống hàm lượng protein là 20% (Chutjareyaves et al., 1999). Chưa có nghiên cứu nhu cầu về các chất dinh dưõng khác của cá Pangasius. 13.4.3. Khẩu phần ăn
Thức ăn tự chế biến và thức ăn công nghiệp thường được sử dụng ở Thái Lan và những nước lân cận. Đối với cá quả, người nuôi thường tự chế biến thức ăn. Đôi khi, người nuôi cho cá ăn bằng thức ăn công nghiệp sản xuất cho cá da trơn - thức ăn công nghiệp. Thành phần thức ăn người nuôi sử dụng cho cá quả được trình bày ở bảng 13.21
Bảng 13.21. Công thức thức ăn tự chế biến cho cá quả nuôi ở Thái Lan
Thành phần Hàm lượng (%) Công thức 1 (Somseub, 1994) Cá tạp 90 Cám gạo 10 Công thức 2 (Somseub, 1994) Cá tạp 80 Cám gạo 20 Công thức 3 (S.Koonsomboon cung cấp thông tin) Cá tạp 50 Bột cá 17,5 Bột đậu tương 7,5 Gạo tấm 7 Cám gạo 17 Vitamin và chất khoáng 1
Thức ăn công nghiệp cho cá da trơn sản xuất tại Thái Lan dùng cho cá bột, cá
giống, cá trưởng thành và chứa tương ứng 40%, 35%, 30% và 25% protein. Thành phần thức ăn sử dụng cũng tương tự như các loại thức ăn khác bao gồm bột cá, bột đậu tương, bột đậu lạc, cám gạo, bột ngô, gạo tấm, bột sắn mì, vitamin, các chất khoáng và chất kết dính.
Thức ăn tự chế biến cho cá Pangasius sp thường là hỗn hợp của vài loại sản phẩm có sẵn ở địa phương. Tuy nhiên, người dân cũng thỉnh thoảng sử dụng thức ăn công nghiệp khi các phụ phẩm ở địa phương không có sẵn. Bảng 13.22 đưa ra các công thức thức ăn cho cá Pangasius sp. Thức ăn công nghiệp cho cá Pangasius nuôi thương phẩm chứa hàm lượng protein từ 15% đến 26%.

142
Bảng 13.22. Công thức thức ăn cho cá Pangasius (P.Rimteerakul)
Thành phần % Cá giống (30% protein) Bột cá 30 Cám gạo 45 Bột đậu tương 24 Vitamin và chất khoáng 1 Cá trưởng thành (25% protein) Bột cá 15 Bột đậu tương 15 Đậu phụng 24 Cám gạo 30 Gạo tấm 15 Vitamin và chất khoáng 1
Thông tin về nhu cầu vitamin và chất khoáng của cá quả và cá Pangasius chưa có nhiều. Tuy nhiên, NRC (1993) đã công bố nhu cầu một số vitamin và chất khoáng cho các loài cá nước ngọt khác ở vùng nhiệt đới. 13.4.4. Nuôi dưỡng
Cá quả. Diện tích hồ nuôi cá quả thường từ 1600 - 3200 m2 và có độ sâu 2-3m. Mật độ nuôi 25 con cá giống/m2. Nuôi cá quả hoàn toàn phụ thuộc vào nguồn protein động vật đặc biệt là cá tạp. Ở những vùng cá tạp có sẵn, cá quả cho ăn hoàn toàn cá tạp băm nhỏ và lượng cho ăn thoả mãn nhu cầu. Giai đoạn cá bột (15g) lên cá giống (50g) cá được cho ăn 3 lần/ngày. Khi cá đạt kích cỡ cá giống, lượng cá tạp giảm xuống 80% và thêm vào 20% cám gạo hoặc gạo tấm được nấu chín. Hỗn hợp các thành phần được băm nhỏ bằng máy băm thịt và được đặt trên tấm gỗ dọc theo hồ (S.Koonsomboon - người cung cấp thông tin). Cá sau 6 - 7 tháng đạt kích cỡ khoảng 1 kg, là kích cỡ thương phẩm thông dụng nhất ở Thái Lan.
Hai loài cá quả C.micropeltes và C.striatus thường được nuôi trong lồng. Lồng nuôi cá quả có thể tích 15 - 200m3. Cá có kích cỡ 50g được nuôi với mật độ 400 - 500 con/m3. Cá tạp được băm nhỏ trước khi cho ăn. Sau 8 - 12 tháng nuôi cá đạt kích cỡ 1 kg. Tốc độ sinh trưởng phụ thuộc vào khẩu phần thức ăn và mật độ nuôi. Hệ số chuyển đổi thức ăn của cá quả nuôi trong hồ và lồng khoảng 4:1 (Nuov và Nandeesha, 1994).
Cá da trơn Pangasius. Loài Pangasius được nuôi cả trong lồng và hồ. P.sutchi thường được nuôi trong hồ, còn lồng thường dùng để nuôi P.sutchi và P.lamaudii. Diện tích hồ nuôi cá Pangasius khoảng từ 600 - 2000m2 và độ sâu 2 - 3m. Cá giống kích cỡ 50g được nuôi với mật độ 3 tạ/ha. Phần lớn các hồ có hệ thống chứa nước và được làm đầy bằng nước mưa hoặc bằng máy bơm từ hồ bên cạnh. Kế hoạch cho ăn dựa trên tính sẵn có của các nguyên liệu trong vùng hơn là sự quan tâm đến kỹ thuật cao. Trong 2 tháng đầu, cá được cho ăn cám gạo. Một số người nuôi dùng hỗn hợp cám gạo, gạo tấm và một lượng nhỏ cá tạp. Từ tháng nuôi thứ 3 trở đi, tuỳ theo từng vùng nuôi và dựa trên thành phần thức ăn có sẵn ở mỗi địa phương mà các khẩu phần thức ăn cho cá khác nhau. Ví dụ, rau cỏ, gạo tấm nấu chín, phế phụ phẩm khác từ các nhà máy chế biến trái cây (như phụ phẩm từ dứa, bao lá từ trái ngô…), cá tạp, cá rẻ tiền và một số thứ khác có sẵn ở Thái Lan, Campuchia và Burma (New et al, 1994). Lượng cho ăn hàng ngày khoảng 10% trọng lượng thân từ

143
giai đoạn cá bột đến cá hương và giảm xuống còn 5% ở giai đoạn cá giống và cá trưởng thành.
Cá được cho ăn 3 lần/ngày trong 8 - 12 tháng nuôi cho đến khi đạt 1-1,5kg. Sản lượng thu được ở Thái Lan là 25,0 -37,5 tấn/ha (theo P.Rimteerakul). Hệ số chuyển đổi thức ăn 4 - 6:1. Pangasius còn được nuôi kết hợp với các loài khác trong hồ (Somsueb, 1994). Mật độ nuôi kết hợp là 1250 gà hoặc vịt hoặc 30- 60 con lợn với 12.500 con cá/ha.
Tiến hành nuôi lồng cá Pangasius ở các hồ lớn hoặc sông. Lồng có kích cỡ từ 20 - 900m3. Thường nuôi với mật độ 5 - 10kg/m3 giai đoạn cá giống (75 - 100g). Cho ăn cá tạp và thỉnh thoảng bổ sung thêm thức ăn viên công nghiệp. Sau 8 - 12 tháng, khi cá đạt 35 -65 kg/m3 tiến hành thu hoạch.
13.5. DINH DƯỠNG VÀ NUÔI DƯỠNG CÁ CHÌNH (ANGUILLA SP.)
13.5.1. Giới thiệu Cá chình là một trong những đối tượng nuôi trồng thuỷ sản chính, sản lượng toàn
cầu trong năm 1997 ước tính đạt trên 200.000 tấn. Bốn loài chính được nuôi đó là Anguilla japonica ở Châu Á, Anguilla anguilla ở Châu Âu, Anguilla rostrata ở Bắc Mỹ và Anguilla
australis ở Úc và New Zealand. Trong các loài đó thì A. japonica là đối tượng chính và được nuôi nhiều ở Trung Quốc và Nhật Bản.
Arai (1991) đã nghiên cứu nhu cầu dinh dưỡng của cá chình. Kể từ đó, một số thông tin mới cũng được trình bày. Những nghiên cứu gần đây cũng đã tập trung vào đặc điểm dinh dưỡng cơ bản của cá chình.
Hình 13.6. Cá Chình 13.5.2. Nhu cầu dinh dưỡng
Protein và amino acid. Cá chình là loài ăn động vật và có nhu cầu về protein cao hơn các loài ăn thực vật và ăn tạp cùng kích cỡ khác. Nose và Arai (1972) cho rằng khi khẩu phần thức ăn sử dụng cazein tinh chế thì A. japonica giai đoạn chưa trưởng thành cần hơn 45% protein trong thức ăn. Gần đây, Tibbets và cộng sự (2000) ước tính mức protein tối ưu cho cá chình Châu Mỹ giai đoạn giống (A. rostrata) là 47% khi trong khẩu phần sử dụng thành phần chính là cá trích. De la Higuera và cộng sự (1989) cho rằng lượng protein ăn vào hàng ngày tốt nhất cho A.anguilla sinh trưởng cao nhất khoảng 1,4g/100g cá và cao hơn cá hồi (Oncorhynchus mykiss) và cá chép (Cyprinus carpio) là 1,16g/100g cá (Ognio, 1980). Nhu cầu protein của cá chình (45-47%) tương đối cao hơn những loài cá khác.
Arai và cộng sự (1972) đã xác định nhu cầu về chất lượng các amino acid của cá A. japonica và A. anguilla khi sử dụng thức ăn thiếu các amino acid cần thiết. Khi cá chình thiếu arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, trytophan và valine trong thức ăn, chúng sẽ sinh trưởng thấp hơn so thức ăn đối chứng có đầy đủ các amino acid. Tuy nhiên, khi bổ sung đầy đủ các amino acid thì cá hồi phục tăng trưởng. Như vậy, cũng như các loài cá khác, cá chình cũng yêu cầu có đầy đủ 10 amino acid thiết yếu.
Nose (1979) đã tiến hành nghiên cứu để xác định nhu cầu về số lượng mỗi amino acid đối với cá chình Nhật Bản (bảng 13.23).
Nhu cầu về arginine của cá chình thấp hơn cá hồi Sinuc (Oncorhynchus tshawytscha). Tuy nhiên, nhu cầu về isoleucine và leucine lại cao hơn cá chép, cá nheo

144
(Ictalurus punctatus) và cá rô phi Nile (Oreochromis niloticus). Hơn thế nữa, nhu cầu về trytophan cao hơn các loài khác ngoại trừ cá rô phi. Gần đây, Akiyama et al (1997) đã tiến hành so sánh nhu cầu các amino acid thiết yếu với tỷ lệ mỗi amino acid thiết yếu chứa trong tổng các amino acid (tỷ lệ A/E) giữa các loài cá và kết luận rằng nhu cầu các amino acid trong thức ăn của cá chình Nhật Bản tương tự cá măng sữa (Chanos chanos); nhưng ở một khía cạnh khác nó lại xếp gần với cá rô phi Nile.
Bảng 13.23. Nhu cầu amino acid của cá chình Nhật Bản giai đoạn giống (Nose, 1979)
Amino acid Tỷ lệ % trong khẩu phần protein*
Tỷ lệ % trong vật chất khô
Arginine 4,5 1,7 Histidine 2,1 0,8 Isoleucine 4,0 1,5 Leucine 5,3 2,0 Lysine 5,3 2,0 Methionine 3,2 1,2 Phenylalanine 5,8 2,2 Threonine 4,0 1,5 Tryptophan 1,1 0,4 Valine 4,0 1,5
* Thức ăn trong thí nghiệm chứa 38% protein Bột cá chất lượng cao được xem là thành phần chính trong khẩu phần của cá chình
nhằm kích thích tăng trưởng và tăng mùi vị thức ăn, trung bình hàm lượng bột cá chiếm 75% của khẩu phần (Gallagher và Degani, 1988). Các nguồn protein có thể sử dụng thay thế bột cá cho cá chình châu Âu: thịt gà tươi tốt hơn máu gà và thịt gà, và chất thải của gà (Degani và cộng sự, 1984). Gần đây, Lee và Bai (1997) cho thấy rằng, trong khẩu phần ăn của cá chình Nhật Bản có thể thay thế bột haemoglobin đến 50% mà không cần bổ sung 3 loại amino acid thiết yếu là methionine, isoleucine và arginine.
Schmitz và cộng sự (1984) đã nghiên cứu khả năng tiêu hoá protein có thành phần là cazein, gelatin, cá, protein vi khuẩn, đậu tương, dung dịch protein đậu tương với các kết quả theo thứ tự là 99, 94, 94, 89, 96 và 94%.
Lipid và các acid béo. Cũng như các loài cá hồi và cá chép, cá chình cũng cần các acid béo cần thiết. Arai và cộng sự (1971) đã nghiên cứu ảnh hưởng của việc bổ sung lipid lên tốc độ tăng trưởng của cá chình Nhật Bản giai đoạn giống sử dụng thức ăn có cazein-gelatin tinh chế, và tìm thấy một hỗn hợp dầu bắp và dầu gan cá với tỷ lệ 2:1 là thích hợp cho sinh trưởng cá chình. Sau đó, Takeuchi và cộng sự (1980) đã xác định nhu cầu các acid béo cần thiết cho cá chình Nhật Bản khi sử dụng thức ăn chứa các methyl esters của acid béo. Họ cũng cho rằng nhu cầu về hai acid linoleic (18:2n-6) và linolenic (18:3n-3) cũng như cá chép nhưng thấp hơn còn 0,5% hơn là 1% đối với mỗi loại.
Lipid có vai trò rất quan trọng, là một nguồn năng lượng trong khẩu phần của cá, đặc biệt đối với những loài ăn tạp như cá chình khi nguồn năng lượng carbohydrates bị hạn chế. Lượng lipids thêm vào có thể làm cho việc sử dụng thức ăn protein tốt hơn. Burgos và cộng sự 1989) đã nghiên cứu ảnh hưởng của việc thay thế từng phần protein bằng chất béo. Các tác giả cho rằng giảm lượng protein trong khẩu phần không làm giảm tăng trọng của cơ thể, nhưng khi tăng lượng chất béo thì trọng lượng thân chỉ tăng đến một mức nhất định, như vậy chất béo chỉ có thể thay thế protein trong một giới hạn nhất định. Theo

145
Degani và cộng sự (1987), cá chình (40-120g) sinh trưởng tốt khi thức ăn chứa tỷ lệ protein/năng lượng (P/E) trên 16,7g/MJ. Ngoài ra, Gallego et al. (1993) đã nghiên cứu ảnh hưởng dự trữ của lipid khi sử dụng protein. Kết quả sinh trưởng tốt nhất khi cá sử dụng khẩu phần chứa 30% protein và 20% lipid, cụ thể là tỷ lệ protein/năng lượng (P/E) là 16,1g/MJ năng lượng thô và năng lượng thô 19MJ/kg thức ăn. Tibbets et al (2000) ước tính tỷ lệ protein/năng lượng tối ưu cho cá chình Châu Mỹ là 22g/MJ.
Dầu cá là nguồn lipid chính trong thức ăn của cá chình. Trong nghiên cứu của Gallagher và Degani (1988) đã chứng minh điều đó. Họ đã thử dùng gia cầm và dầu gia cầm thay cho cá và dầu cá trong khẩu phần của cá chình và thấy rằng những loại thức ăn được bổ sung 10% dầu cá có sự tăng trọng cao hơn là những loại thêm 5% dầu cá hoặc 5-10% dầu gia cầm.
Carbohydrate. Như đã đề cập ở trên, cá chình là loài ăn thịt và khả năng sử dụng carbohydrate có giới hạn. Mặc dù những thông tin về điều này còn rất hạn chế, có báo cáo cho rằng khả năng tiêu hóa gelatin tinh chất là khá cao từ 78 - 98% khi trong thức ăn chứa 20 -60%. Gelatin tinh chất giữ vai trò quan trọng như là chất kết dính trong thức ăn của cá chình. Nó là thành phần không thể thiếu, ngay khi có sự hoài nghi về tỷ lệ sử dụng thấp đối với cá chình.
Degan (1987) đã đánh giá ảnh hưởng của các nguồn carbohydrate khác nhau lên hoạt động của enzyme và glucose chứa trong cơ và gan của cá chình. Họ cho rằng cá chình ăn 30% bột mì hoặc tinh bột ngô thì có mức glucose trong cơ và gan cao, và hoạt động của aldolase cao khi cá ăn hạt lúa mì, bột mì hoặc tinh bột. Garcia - Gallego và cộng sự (1991) đánh giá khả năng của mức carbohydrate cao trong thức ăn công nghiệp cho cá chình và đề nghị tỷ lệ cho phép sử dụng tốt nhất ở mức 40% và cao hơn.
Vitamin. Arai và cộng sự (1972) đã đưa ra nhu cầu đối với các vitamin hoà tan của cá chình A. japonica giai đoạn non và các triệu chứng khi thiếu vitamin (bảng 13.24).
Bảng 13.24. Dấu hiệu thiếu vitamin ở cá chình (Arai và cộng sự, 1972)
Vitamin Dấu hiệu thiếu vitamin Thiamine Cong thân, mất điều hoà, xuất huyết và sung huyết ở vây, màu sắc tối
sẫm, bơi lờ đờ Riboflavin Xuất huyết và sung huyết ở vây, viêm da, nổi tầng mặt (thích sáng)
và bơi lờ đờ Pyridoxine Rối loạn thần kinh, co giật, bơi lung tung (như động kinh) Pantothenic acid Bơi dị thường, mất điều hoà, chết, xuất huyết ở biểu bì và viêm da. Niacin Bơi dị thường, mất điều hoà, thiếu máu, xuất huyết ở biểu bì và viêm
da. Biotin Bơi dị thường, màu sắc tối sẫm Folic acid Màu sắc tối sẫm, chậm lớn Cyanocobalamin Chậm lớn Choline Ruột trắng xám Inositol Ruột trắng xám Ascorbic acid Xuất huyết ở vây, đầu và da, miệng dưới bị ăn mòn p-aminobenzoic Không α-tocopherol Xuất huyết ở vây và da và viêm da
Cá chình Nhật Bản cần 11 vitamin, tuy nhiên, sự thiếu hụt không rõ ràng trong thức ăn của cá chình ngoại trừ acid p-aminobenzoic. Trong các loại vitamin, sự thiếu hụt về

146
pyridoxine, acid pantothenic và choline đã gây ra ảnh hưởng sớm hơn các loại khác, sự biểu hiện các dấu hiệu của chúng có ý nghĩa quan trọng. Phần lớn những dấu hiệu sẽ biến mất sớm sau khi cho ăn đầy đủ các vitamin.
Yamakawa và cộng sự (1975) đã mô tả dấu hiệu của sự thiếu hụt vitamin E và xác định số lượng nhu cầu α-tocopherol dùng vitamin tự do cazein như là nguồn protein và acid béo methyl esters như là nguồn lipid. Dấu hiệu quan sát được là bỏ ăn và sinh trưởng kém, xuất huyết và sung huyết ở vây và viêm ở da. Nhu cầu tối thiểu đối với α-tocopherol cho sinh trưởng của cá chình con là 200mg/kg thức ăn.
Chất khoáng. Nose và Arai (1979) đã xác định được nhu cầu chất khoáng của cá chình Nhật Bản trong thức ăn tinh chế. Cá chình ăn thức ăn thiếu canxi và photpho thì dần dần chúng sẽ chán ăn sau một tuần cho ăn và tiếp đó sẽ giảm tăng trưởng. Tuy nhiên, khi thức ăn thiếu magiê và sắt, cá sẽ bỏ ăn sau 3-4 tuần. Cá chình ăn thiếu sắt, bị bệnh thiếu máu. Lượng tối ưu đối với canxi, magiê, photpho và sắt cho cá chình con được ước tính lần lượt là 2700, 400, 2500-3200 và 170 mg/kg thức ăn.
Theo Park và Shimizu (1989) mức bổ sung nhôm và kẽm tối ưu cho thức ăn cá lần lượt là 15 và 50 - 100mg/kg. 13.5.3. Khẩu phần ăn
Thức ăn công nghiệp dùng cho cá chình Nhật Bản đã được sản xuất gần 35 năm trước. Gần đây, sự đổi mới các dạng thức ăn (dạng viên, dạng dính, dạng nổi) đã tăng hiệu quả sử dụng thức ăn của cá. Một số thông tin về thành phần của thức ăn công nghiệp đại diện cho cá chình ở Nhật Bản trình bày ở bảng 13.25. Thức ăn công nghiệp loại B chứa 5-6% lipid có nguồn gốc từ cá. Thức ăn tốt cho sinh trưởng và hiệu quả thức ăn cao khi tăng lượng dầu trong thức ăn đến 15% (Tsuda, 1997).
Bảng 13.25. Thành phần (%) của thức ăn công nghiệp cho cá chình (Tsuda, 1997)
Loại thức ăn Kích cỡ cá chình (g) Độ ẩm Protein Lipid Loại dính (nhão) Cá chình con (sau nở): 0,2 – 0,5 82 14 0.8 Loại A Cá chình con: 0,4 – 1,0
Cá chình con: 0,5 - 10 7 7
65 54
5 5
Loại B Cá chình con 10 - 20 Cá giống đến trưởng thành 20 -200
7 7
52 49
6 5
Nổi Cá chình trưởng thành 7 49 16 13.5.4. Nuôi dưỡng
- Ngày thứ nhất đến ngày thứ hai cho ăn Cladocera; - Ngày thứ ba đến ngày thứ tư cho ăn hồng trần; - Ngày thứ năm cho ăn hồng trần nghiền vụn trộn với 10 - 30% thức ăn tổng hợp.
Sau đó mỗi ngày tăng thêm 10% thức ăn tổng hợp đến ngày thứ 10 thức ăn tổng hợp chiếm 80%. Từ ngày thứ 15 trở đi hoàn toàn dùng thức ăn tổng hợp. Lượng thức ăn tổng hợp được tính bằng 10 - 15% trọng lượng cá trong ao, ngày cho ăn 2 lần vào lúc 7 - 8 giờ sáng và 4 - 5 giờ chiều.
Giai đoạn cho ăn hồng trần, lượng hồng trần được tính bằng 30 - 35% trọng lượng cá trong ao và ngày cho ăn 3 lần vào sáng, chiều, tối. Nếu nhiệt độ dưới 15oC chỉ cho ăn 1 lần hoặc không cho ăn. Khi cho ăn không sục khí, tập dần cho cá chỉ ăn ban ngày và khu vực cho cá ăn không cần che tối.

147
Thức ăn phải mềm cá mới ăn được nhưng không quá mềm, dễ tan trong nước. Nên thêm dầu dinh dưỡng vào thức ăn và trộn đều rồi mới cho cá ăn. Tỷ lệ thức ăn, dầu dinh dưỡng và nước để trộn thức ăn có quan hệ mật thiết với nhiệt độ (bảng 13.26) và tỷ lệ cho ăn so với trọng lượng thân cá tùy theo các giai đoạn phát triển của chúng (bảng 13.27)
Bảng 13.26. Tỷ lệ phối trộn thức ăn hỗn hợp với dầu và nước (kg)
Nhiệt độ (oC) Thức ăn Dầu dinh dưỡng Nước
< 18 100 0 130
18 - 23 100 3 - 5 170
> 23 100 5 - 8 200
Bảng 13.27. Tỷ lệ thức ăn so với trọng lượng thân cá chình ở các giai đoạn
Cỡ cá Cá bột Cá hương Cá giống Cá cỡ nhỏ Cá thương phẩm
Trọng lượng cá (g) 0,2- 0,8 1-1,5 16-40 40-100 150-200
Thức ăn (%) 6-10 4-6 3-4 2,8-3 2-2,5
13.6. DINH DƯỠNG VÀ NUÔI DƯỠNG TÔM HE
13.6.1. Giới thiệu Hiện nay, sản lượng đánh bắt tôm xấp xỉ đạt ngưỡng tối đa và không thể tiên đoán
được bởi các hiện tượng tự nhiên xảy ra bất thường không thể lường được. Hơn nữa, sự ô nhiễm và các hoạt động khác của con người đã phá hủy vùng sinh thái cho tôm đẻ ở nhiều khu vực. Nhu cầu tiêu tốn năng lượng cho việc khai thác tôm biển cao đẩy giá thành tôm lên cao. Do vậy, việc thỏa mãn nhu cầu tiêu thụ tôm ngày càng tăng trên thế giới phụ thuộc vào việc đẩy mạnh hoạt động nuôi. Năm 1986, tôm nuôi ao cung cấp khoảng 6 – 8% nhu cầu tôm trên thế giới , đến 1996 theo FAO con số này đã lên đến 43% nhu cầu tiêu thụ trên thế giới.
Sự phát triển các hình thức nuôi tôm bắt đầu từ những năm 1930 khi người Nhật cho đẻ và ương nuôi thành công ấu trùng tôm Penaeus japonicus trong bể. Từ đó, công nghệ tăng nhanh sinh khối ấu trùng tôm biển phát triển mạnh mẽ. Trong vòng 20 năm qua, tôm bố mẹ Pennaeus japonicus, P. monodon, P. indicus, P. mergiensis, P. aztecus, P. setiferus,
P. schmitti, P. chinensis, P.
Hình 13.7. Tôm sú (P. monodon) penicillatus, P. stylirostris và P. vannamei đã được nuôi vỗ và cho đẻ, ấu trùng của chúng được ương nuôi trong bể.

148
Do các trang trại tôm ngày càng tăng, các hình thức nuôi thâm canh thay thế cho nuôi truyền thống dùng các ngư cụ để bẫy và giữ tôm tự nhiên đi vào ao ven bờ mỗi khi thủy triều dâng nhằm đạt năng suất 24 tấn/ha/vụ nuôi. Thức ăn tự nhiên đóng vai trò chủ lực trong nuôi quảng canh trong khi thức ăn nhân tạo đóng vai trò chủ yếu trong hình thức nuôi bán thâm canh và thâm canh. Trong hình thức nuôi bán thâm canh và thâm canh, thức ăn chiếm đến 50% tổng chi phí sản xuất. Vì vậy, việc sử dụng thức ăn cân bằng dinh dưỡng và giá rẻ cùng với việc cho ăn tốt là những yếu tố cơ bản cho việc nuôi tôm thành công. 13.6.2. Các hình thức nuôi
Nhìn chung, có 3 hình thức nuôi tôm he là hình thức nuôi quảng canh, bán thâm canh và thâm canh.Thông thường, với hình thức quảng canh, mật độ nuôi post hoặc giống (juvenile) thấp hơn 2,5 con/m2. Hình thức lấy giống tự nhiên vào ao lúc nước triều dâng vẫn còn áp dụng ở một số nước châu Á. Rất ít khi cho thêm thức ăn bổ sung mà chủ yếu dựa vào nguồn thức ăn tự nhiên có sẵn trong ao. Bón thêm phân vô cơ và hữu cơ để tăng số lượng nguồn thức ăn tự nhiên trong ao. Quản lý nguồn nước dựa vào sự biến động của thủy triều. Với hình thức nuôi này, năng suất đạt từ 150-500 kg/ha/vụ.
Hầu hết tôm nuôi cung cấp trên thế giới được nuôi ở ao đất bằng hình thức bán thâm canh. Với hình thức này, mật độ thả giống từ 3-15 con/m2 hoặc gấp đôi mật độ nếu thả post. Bổ sung thêm thức ăn công nghiệp cùng với thức ăn tự nhiên. Phân bón được sử dụng ngay từ ban đầu để tăng khả năng sinh trưởng của thức ăn tự nhiên trong ao. Nước được bơm thêm vào ao hàng ngày từ 2 – 10% khi cho ăn. Nhìn chung, năng suất đạt 600-2.000 kg/ha/vụ, 2-2,5 vụ/năm. Khối lượng trung bình của tôm khi thu hoạch biến động từ 16-36 con/kg tùy thuộc vào mật độ nuôi, môi trường và quản lý ao nuôi. Giá bán khác nhau phụ thuộc vào kích cỡ của tôm lớn hay nhỏ.
Ở những nơi giá đất xây dựng ao nuôi quá cao hoặc không có đất và khi giá tôm lên cao, hình thức nuôi thâm canh được áp dụng. Hình thức nuôi thâm canh đòi hỏi công nghệ quản lý phức tạp, tỷ lệ sục khí và trao đổi nước cao cùng với khẩu phần ăn đầy đủ dinh dưỡng. Tôm được nuôi trong bể hoặc ao với mật độ 20-40 con/m2. Thức ăn nhân tạo là nguồn thức ăn cung cấp dinh dưỡng chủ lực cho tôm. Năng suất biến động từ 2.000-9.000 kg/ha/vụ, nuôi thâm canh trong bể ở Nhật có thể đạt tới 24 tấn/ha/vụ. 13.6.3. Nhu cầu dinh dưỡng và nguồn cung cấp
Mức protein trong khẩu phần cho tăng trưởng tối ưu của tôm dao động từ 28 – 60% tùy vào giống, kích cỡ, chất lượng protein, mức năng lượng phi protein, chất lượng của dạng viên thức ăn, tính ngon miệng, tỷ lệ cho ăn, chất lượng nước và lượng thức ăn tự nhiên sẵn có trong ao. Hầu hết, nhu cầu về số lượng protein được tính cho tôm nhỏ nuôi trong thùng chứa hoặc bể không có thức ăn tự nhiên. Tôm tăng trưởng cho đến kích cỡ thu hoạch ở ao có nhu cầu ít protein hơn so với số cung cấp trong tài liệu.
Tỷ lệ protein trong thức ăn công nghiệp nuôi thâm canh khoảng 35% hoặc cao hơn. Protein thức ăn trong nuôi bán thâm canh biến động từ 25-35%. Nghiên cứu trong phòng thí nghiệm Enrique Ensenot ở Panama cho thấy thức ăn chứa 25% protein cho năng suất tôm tương đương với thức ăn chứa tỷ lệ protein cao hơn trong nuôi bán thâm canh có bón phân với mật độ 5 con/m2. Hợp tác nghiên cứu giữa trường Đại học Auburn và trang trại Granjas Marinas ở Honduras cũng cho kết quả tương tự.
Một số nghiên cứu đã được thực hiện để đánh giá giá trị dinh dưỡng của các nguồn protein khác nhau. Bột cá là nguồn thức ăn chất lượng cao cho cá nhưng dường như có giá

149
trị dinh dưỡng thấp hơn đối với tôm, đặc biệt khi cho ăn chỉ đơn độc nguồn protein này. Điều này đã được xác định ở nhiều giống. Shigueno (1975) cho rằng, có thể vì do thiếu phenylalanine và acid amin căn bản (arginine, histidine và lysine) trong một số loại bột cá. Vai trò quan trọng của bột cá trong khảu phần nuôi tôm là để tăng tính ngon miệng.
Bột đậu tương là nguồn protein thực vật được sử dụng phổ biến nhất cho cá, đây cũng là nguồn protein tốt cho tôm. Bột đậu tương cho tăng trưởng của tôm P. duorarum tốt hơn bột cá, bột tôm, casein, bột gluten (Sick và Andrews, 1973). Trong thực tế sản xuất, bột đậu tương với tỷ lệ 20 – 50% khẩu phần có thể thay thế phần lớn bột cá, bột tôm, bột mực hoặc kết hợp giữa các loại này với nhau để nuôi tôm. Tuy nhiên, Lim và Dominy (1990) cho rằng nếu tính ngon miệng và sự ổn định về nước được duy trì thì có thể sử dụng bột đậu tương cho con giống P. vannamie lên đến 56% trong khẩu phần.
Bột hạt bông có thể sử dụng được cho tôm. Tuy nhiên, bột hạt bông (chứa 0,41% gossyfol tự do) không nên bổ sung quá 26% (hoặc 1100 ppm gossyfol tự do) trong khẩu phần của tôm giống P. vannamie. Bột hạt bông chứa rất ít lysine, do vậy, không thể thay thế một cách ngang bằng cho bột đậu tương được.
Tôm cũng cần có đủ 10 acid amin thiết yếu giống như cá và các động vật trên cạn. Arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan và valine là các acid amin thiết yếu đối với tôm Panaeus japonicus, P. azteus và P. monondon. Tuy nhiên nhu cầu về số lượng cho tất cả các acid amin thiết yếu này chưa được xác định. Trong điều kiện thiếu thông tin về nhu cầu số lượng này thi có thể áp dụng nhu cầu acid amin thiết yếu như của gia súc.
Bảng 13.28 sau đây liệt kê hàm lượng acid amin thiết yếu trong cơ của tôm P. japonicus và P. vannamei, và của mực, nguồn protein chất lượng cao đối với tôm. Khẩu phần nuôi tôm với tỷ lệ acid amin thiết yếu như trong cơ của tôm, mực, sò sẽ tạo được sự tăng trưởng tốt của tôm nuôi.
Bảng 13.28. Hàm lượng các acid amin thiết yếu của protein trong cơ thịt tôm, sò, mực, casein và trứng toàn phần
Nguồn protein (%) Acid amin thiết yếu P.japonicus P.vannamei Sò Mực Casein Trứng
Arginine 7,46 8,54 4,50 5,40 3,30 5,45 Histidine 1,66 1,86 1,27 1,50 2,65 1,71 Isoleucine 2,89 3,40 2,00 2,88 4,50 3,46 Leucine 7,04 6,28 4,01 5,79 8,76 6,47 Lysine 7,24 6,97 4,68 5,52 7,34 5,45 Methionine 2,92 2,48 1,70 2,30 2,51 3,01 Phenylalanine 3,90 3,39 2,13 2,86 4,75 4,15 Threonine 3,62 2,69 2,81 3,28 3,77 3,73 Tryptophan 0,52 1,27 0,51 0,72 1,21 3,76 Valine 2,87 3,38 2,18 2,66 5,83 3,76
Những nổ lực bổ sung trong các khẩu phần thiếu acid amin bằng các acid amin
tổng hợp đã không thành công đối với tôm, trái ngược với kết quả thu được từ cá da trơn và gia súc. Deshimaru (1982) cho rằng tỷ lệ tạo thành arginine dạng tự do vào trong protein cơ ít hơn 1% so với sự tạo thành 90% arginine ở dạng protein. Mai và cộng sự (1988) cho rằng con giống P. orientalis đã không hấp thu đồng thời cùng lúc methionine

150
và lysine tự do với các acid amin ở dạng gắn với protein. Sự bất lực của tôm trong việc sử dụng các acid amin dạng tự do thay cho acid amin dạng protein là do sự hấp thu với tỷ lệ khác nhau giữa acid amin tự do và acid amin dạng protein.
Việc cung cấp năng lượng tối ưu trong khẩu phần rất quan trọng vì khi thiếu hụt năng lượng phi protein, một phần protein sẽ được sử dụng để cung cấp năng lượng cho cơ thể. Cung cấp đủ nhu cầu carbohydrate và lipid trong khẩu phần sẽ giảm được nhu cầu protein mà vẫn không làm giảm khả năng tăng trưởng của tôm nuôi. Tuy nhiên, thừa năng lượng trong khẩu phần có thể làm giảm sự tiêu thụ thức ăn, do đó giảm lượng protein và các chất dinh dưỡng khác ăn vào. Sedgwick (1979) cho rằng lượng thức ăn tôm P. merguiensis tiêu thụ bị khống chế bởi mức năng lượng khẩu phần bất chấp hàm lượng protein có trong khẩu phần.Có rất ít nghiên cứu liên quan đến nhu cầu năng lượng của tôm. Tuy nhiên, tỷ lệ năng lượng / protein tối ưu đối với tôm gần giống với cá là 9-11 kcal/g protein. P. merguiensis đạt tăng trưởng tối đa khi khẩu phần có chứa 39,5% protein có năng lượng tổng số là 4,42 kcal/g (Sedgwick, 1979). Tỷ lệ protein thô – năng lượng tổng số trong thí nghiệm này vào khoảng 90 mg/kcal. Khẩu phần phối hợp có chứa 25 – 35% protein và 4-5% lipid sẽ tạo được tỷ lệ năng lượng – protein phù hợp.
Tôm sử dụng nguồn năng lượng từ carbohydrate và lipid trong khẩu phẩn để tiết kiệm protein. Tuy nhiên, chúng cũng sử dụng protein như một nguồn năng lượng, các nghiên cứu đã chỉ ra rằng khẩu phần chứa quá cao tỷ lệ protein – năng lượng phi protein làm giảm tỷ lệ tăng trưởng. Lipid trong khẩu phần của tôm không chỉ cung cấp năng lượng mà còn cung cấp các acid béo thiết yếu, các vitamin hòa tan trong mỡ, sterol và phospholipid cần thiết cho sự phát triển bình thường của tôm.
Những nghiên cứu về dinh dưỡng chỉ ra rằng acid béo linoleic (n-6) và linolenic (n-3) là cần thiết cho tôm P.japonicus, P. indicus, Palaemon serratus, P. stylirostris và P. vannamei. Tôm không có khả năng phá hủy sự bão hòa và kéo dài chuỗi các acid béo 18:3 n-3 thành các acid béo 20:5 n-3 và 22:6 n-3. Mức tối ưu trong khẩu phần đối với các acid béo không no n-3, 20:5 n-3 và 22:6 n-3 cho tôm dao động từ 0,5-1%, mức tối ưu của acid béo n-6 khoảng 0,5%. Khẩu phần chứa 0,5% acid béo n-6 và 0,5% acid béo n-3 cho sự tăng trưởng tối đa của nhiều giống tôm khác nhau.
Mặc dầu lipid trong khẩu phần có ảnh hưởng đến sự sử dụng tiết kiệm protein, nhưng tôm không chịu được khẩu phần chứa lượng lipid cao như đối với cá hồi. Nhiều nghiên cứu sử dụng các nguồn lipid khác nhau đã cho biết vượt quá tỷ lệ 10% lipid trong khẩu phần có thể làm giảm tăng trưởng.
Loài giáp xác không tổng hợp sterol từ acetate hoặc mevalonate như cá, do vậy chúng cần được cung cấp từ khẩu phần ăn. Cholesterol là sterol chủ yếu được tìm thấy ở loài giáp xác và là một tiền hormon giới tính, hormon lột xác và là một thành phần của lớp dưới da ở loài giáp xác. Bên cạnh acid béo thiết yếu, sterol, tôm biển còn có nhu cầu về phospholipid, như lecithin. Mức phối hợp phospholipid tối ưu trong khẩu phần dao động từ 1-3% phụ thuộc vào giống, vòng đời, nguồn và đặc tính của phospholipid, và có lẽ là các chất dinh dưỡng khác. Lecithin từ đậu tương là một nguồn cung cấp thích hợp và rẻ tiền phospholipid cho tôm.
Trong số 15 vitamin đã được xác định là cần thiết cho cá thì 14 vitamin đã được thử nghiệm thông qua các nghiên cứu về xác định nhu cầu cho tôm. Tỷ lệ phối trộn các vitamin trong khẩu phần đối với tôm cao hơn so với cá. Cũng có thể do nhu cầu trao đổi của tôm cao hoặc cũng có thể là do mất đi một lượng hòa tan vào nước trong quá trình ăn mà chưa xác định được. Hơn nữa, thí nghiệm xác định nhu cầu vitamin đối với post và giai

151
đoạn đầu của juvenile được thiết lập trong phòng thí nghiệm với các yếu tố môi trường được tối ưu hóa và ổn định.
Bảng 13.29. Lượng vitamin đề nghị bổ sung và lượng tối đa trong khẩu phần ăn của tôm
Số lượng/kg khẩu phần Các vitamin Bổ sung Tối đa Vitamin A 2000 IU 4000 IU Vitamin D 1000 IU 2000 IU Vitamin E 50 mg 100 mg Vitamin K 10 mg 20 mg Thiamin 20 mg 50 mg Riboflavin 10 mg 30 mg Pyridoxine 30 mg 60 mg Pantothenic acid 30 mg 80 mg Niacin 30 mg 80 mg Biotin 0 2 mg Acid folic 2 mg 5 mg Vitamin B12 0,01 mg 0,05 mg Inositol 50 mg 200 mg Vitamin C 100 mg 200 mg Choline chloride 500 mg 1500 mg
Tôm có nhu cầu bổ sung thêm phospho, nhưng trong nước biển chúng có thể hấp
thu đủ magiê và canxi từ nước. Bổ sung vào khẩu phần cho tôm các khoáng vi lượng, sắt, đồng, kẽm, Se và Mn để cải thiện sự tăng trưởng (Deshimaru và Yone, 1978).
Bảng 13.30. Lượng khoáng cần thiết trong khẩu phần của tôm
Khoáng Số lượng/kg thức ăn khô
Khoáng Số lượng/kg thức ăn khô
Khoáng đa lượng (g) Khoáng vi lượng (mg) Canxi 10 Mangan 40 Photpho 10 Kẽm 33 hoặc 200 Kali 6 Sắt 60 Magiê 0,4 Đồng 32
Iod 5 Selen 0,4
Coban 0,4 13.6.4. Khẩu phần ăn và nuôi dưỡng
Thức ăn tự nhiên Tôm he trưởng thành là loại ăn tạp, thức ăn là các sinh vật và mùn bã đáy, chúng là
những bọn bắt mồi cơ hội. Ở giai đoạn Zoea và Mys, ấu trùng ăn vi sinh vật bơi tự do. Giai đoạn Post, sống tầng đáy, thức ăn là mùn bã hữu cơ. Thói quen của Juvenile bắt đầu là ăn tạp sau đó đổi thành ăn thịt và chúng bắt chủ yếu là những động vật không xương sống nhỏ di chuyển chậm chạp.

152
Khẩu phần cho năng suất cao trong nuôi thâm canh đã được thiết lập. Tuy nhiên, những khẩu phần ăn như thế có lẽ là không kinh tế lắm đối với phương thức nuôi tôm bán thâm canh. Trong điều kiện ao hồ, nguồn thức ăn tự nhiên cho tôm là lớp mỏng ở đáy ao nơi bao gồm tảo, thủy sinh vật sống và chết, loài giáp xác và động vật thân mềm nhỏ, vi khuẩn, mùn bã và các sinh vật đáy khác như giun đốt và giun tơ. Mặc dầu tôm đã ăn một số lượng có ý nghĩa các chất dinh dưỡng từ nguồn thức ăn tự nhiên này hàng ngày nhưng khó biết được số lượng hay tỷ lệ phần trăm lấy được là bao nhiêu so với nhu cầu. Năng suất tôm có thể đạt đến 500 kg/ha/vụ trong những ao không cho ăn. Bởi vì tôm có xu hướng tiêu thụ mạnh thức ăn tự nhiên và sự biến động lớn về khả năng sẵn có của thức ăn trong những điều kiện quản lý và môi trường nuôi khác nhau nên khó có thể phối trộn được các loại thức ăn giá rẻ cho tôm nuôi bán thâm canh.
Tập quán ăn Tôm lấy thức ăn chủ yếu bằng cơ chế nhận biết hơn là nhìn thấy. Những cơ quan
thụ quan tập trung ở các phần phụ phía trước và râu. Ngay khi phát hiện được mùi của con mồi, tôm di chuyển về phía con mồi và nhanh chóng vồ chúng với những càng kẹp. Con mồi bị nhai nhỏ ra cỡ thích hợp trước khi nuốt. Thức ăn vào bụng đi qua thực quản và vào túi trước của dạ dày (ruột trước), nơi nó biến thành dạng sền sệt bởi các enzyme tiêu hóa. Dịch thức ăn đi xuống phần sau của ruột và cuối cùng đến các ống nhỏ của gan tụy tiếp tục tiêu hóa và hấp thu. Những hạt không mịn, thô chảy trực tiếp đến ruột giữa, nơi có vài sự tiêu hóa và là nơi quan trọng để hấp thu các chất dinh dưỡng. Những phần thức ăn không được tiêu hóa đi vào ruột sau, nơi chủ yếu tạo thành khuôn phân.
Khẩu phần ăn Thức ăn chế biến công nghiệp là nguồn dinh dưỡng cho tôm ở giai đoạn ương, nuôi
thương phẩm hình thức thâm canh và bán thâm canh. Tuy nhiên các mô bào tươi sống như bã cá, sò, mực thường dùng để nuôi vỗ tôm bố mẹ, nơi thức ăn tươi sống (rotifer, Artemia, tảo) là nguồn thức ăn chính để nuôi ấu trùng tôm.
Mặc dù thông tin về nhu cầu các chất dinh dưỡng cơ bản của nhiều giống tôm là sẵn có nhưng thiếu số liệu nghiên cứu về đề xuất các loại thức ăn nuôi ao. Bởi vì môi trường nuôi có ảnh hưởng lớn đến nhu cầu dinh dưỡng của tôm, phân tích lợi nhuận - chi phí thức ăn cho các hình thức nuôi và quản lý khác nhau là khó tính toán. Tính cần thiết của việc bổ sung các loại thức ăn ao khác nhau với tất cả các chất dinh dưỡng như vitamin và các chất béo cần thiết chưa được hình thành. Hầu hết các loại thức ăn công nghiệp có chứa hỗn hợp vitamin – khoáng. Nhìn chung, khẩu phần chứa hàm lượng protein cao được sử dụng sớm trong suốt giai đoạn post và giai đoạn juvenile, mức protein khẩu phần giảm dần trong suốt giai đoạn nuôi thương phẩm.
Chế biến thức ăn Vì tôm có thói quen ăn đáy, thức ăn công nghiệp nên được chế biến thành các dạng
viên chìm. Kích cỡ của viên thức ăn biến động tùy thuộc vào cỡ tôm. Dạng vụn nát được sử dụng trong suốt giai đoạn post; dạng viên được sử dụng kể từ giai đoạn juvenile cho đến cỡ thương phẩm. Đường kính của viên thức ăn để phù hợp với các cỡ tôm khác nhau nên là 1-2 mm đối với tôm 0,5-2 g; 2 mm đối với tôm 2-5 g; 3 mm đối với tôm 5-10g và 4mm cho tôm 4-10g và lớn hơn nữa. Tôm là con vật có tính ăn chậm và chọn lựa, vì vây, viên thức ăn nên duy trì được trong nước một thời gian cho đến khi ăn hết.
Hai quy trình chế biến là viên ép đùn và viên ép hấp được sử dụng phổ biến trong sản xuất thức ăn cho tôm. Ép được sử dụng rộng rãi trong sản xuất thức ăn nổi cho cá, tuy nhiên, cũng có thể sản xuất thức ăn chìm với máy đùn ép có nấu bằng cách giảm tỷ lệ giãn

153
nở nên viên thức ăn nặng. Viên thức ăn đùn ép thường có tính ổn định trong nước cao vì có sự keo hóa tinh bột. Tuy nhiên, hầu hết các loại thức ăn cho tôm được chế biến với viên nén được thiết kế đặc biệt gắn với máy phun hơi và hệ thống điều hòa hơi cho phép gia tăng nấu hỗn hợp thức ăn.
Các chất khác nhau (có thể là tự nhiên, chế biến, tổng hợp) được sủ dụng làm chất kết dính cho thức ăn tôm dạng viên hấp hơi. Bột mì, bột ngũ cốc, bột các loại củ là những chất kết dính tự nhiên được sử dụng phổ biến nhất trong sản xuất viên thức ăn cho tôm.. Những chất kết dính công nghiệp như lignin và bentonite được dùng phổ biến cho các loại thức ăn dạng viên cho vật nuôi đã được xác định là ít ảnh hưởng đến thức ăn tôm đòi hỏi nhu cầu ổn định lâu trong nước. Các hydrocolloid hữu cơ như carboxymethyl cellulose, alginate, gum đã được thử nghiệm thành công trong phòng thí nghiệm nhưng còn hạn chế đưa ra thị trường để sản xuất thức ăn cho tôm dạng viên vì giá thành quá cao. Các chất kết dính tổng hợp như Aqua-firm 1A, Aqua-firm 2A được sử dụng trong thức ăn cho tôm.
Các chất dẫn dụ (chất hấp dẫn) thường được sử dụng trong thức ăn của tôm để tăng sự hấp dẫn và tiêu thụ thức ăn. Những chất khác nhau như acid amin, acid béo và các chất chiết của cá, tôm, mực, sò.. kích thích sự bắt mồi của tôm. Những nghiên cứu này đã thành công trong phòng thí nghiệm nhưng chưa được chứng minh trong thực tiễn. Bột cá, nhìn chung, được sử dụng như là thành phần gây sự hấp dẫn trong thức ăn cho tôm và cá.
Cho ăn Bởi vì tôm là loài ăn chậm nên thỏa mãn việc cho ăn là khó và có lẽ không thể đạt
được. Môt số nhà sản xuất cho ăn dựa theo kinh nghiệm của vụ nuôi trước. Để tăng hiệu quả việc cho ăn, một số nhà sản xuất cố gắng tính toán hoạt động bắt mồi trong ao. Họ đặt các khay cho ăn trong ao và số lượng thức ăn còn lại trong khay 1 giờ, 2 giờ sau khi cho ăn sẽ quyết định lượng thức ăn cho ăn vào ngày tiếp theo tăng hay giảm. Một số nhà sản xuất lấy một vốc bùn đáy ở khu vực cho ăn để tìm thức ăn sót lại hoặc mùi hôi thối của thức ăn thối rữa. Những nhà sản xuất kinh nghiệm có thể quan sát thói quen bơi của tôm dọc theo bờ 1 hoặc 2 giờ sau khi cho ăn để biết tôm đã ăn đủ hay chưa. Các bảng hướng dẫn cho ăn đã được các công ty thức ăn chuẩn bị. Đây có thể xem như những hướng dẫn cách cho ăn nếu như có thể ước tính được khối lượng tôm trong ao nuôi. Lượng cho ăn không nên vượt quá khả năng của hệ thống để tiêu hóa hết những sản phẩm thải ra và duy trì đủ lượng oxy hòa tan trong ao.
Do tôm ăn chậm và liên tục nên cho ăn nhiều lần trong ngày. Trong điều kiện thí nghiệm, tần suất cho ăn tối ưu đối với tôm P. monodon giống là 3 lần/ngày (Lim và Pascual, 1979). Có thể hạn chế tối đa sự tan rã thức ăn và việc mất các chất dinh dưỡng hòa tan trong nước bằng cách cho ăn nhiều lần trong ngày hơn là cho ăn 1 lần trong ngày. Không như cá, tôm thường khu trú cố định và không bơi đi xa để bắt mồi. Do vây, việc phân phối thức ăn khắp đều trên mặt ao hoặc nơi tôm khu trú là rất quan trọng. Đối với hồ nhỏ có thể cho ăn bằng tay, đối với các hồ lớn có thể cho ăn bằng thuyền, máy thổi.
13.6.5. Chế phẩm sinh học EM trong quản lý môi trường ao nuôi tôm Nghề nuôi tôm ở nước ta đã được hình thành từ lâu nhưng mãi đến năm 1990 mới được phát triển mạnh cả về số lượng lẫn chất lượng để đáp ứng nhu cầu ngày càng tăng sản phẩm tôm cho tiêu dùng và xuất khẩu. Tuy nhiên, do phát triển tự phát, không theo đúng quy hoạch nên chất lượng môi trường ngày càng giảm, đang có nguy cơ bị ô nhiễm nghiêm trọng, tính rủi ro của nghề nuôi tôm ngày càng cao. Nhiều kết quả nghiên cứu cho thấy, một trong những nguyên nhân quyết định làm tôm tăng trưởng chậm, giảm sức đề kháng, dễ bị bệnh tấn công là do môi trường từ lớp bùn cặn bã hữu cơ dơ bẩn tích tụ lâu ngày ở bề

154
mặt đấy ao, bắt nguồn từ thức ăn dư thừa, từ các chất mùn, vỏ tôm, các cặn bã hữu cơ co sẵn trong nguồn nước. Chính những lớp bùn dơ bẩn đó là nguồn chứa đủ mọi vi sinh vật gây bệnh và tạo ra các khí độc. Trong ao nuôi có càng nhiều chất cặn bã hữu cơ hay thực phẩm dư thừa, cung cấp và tạo điều kiện cho vi sinh vật gây bệnh như : Vibrio, Aeromonas, E coli... sinh sôi nảy nở càng nhanh và sớm giết chất tôm nuôi trong ao. Thực chất là bản thân các vi sinh vật có trong ao nuôi không giết chết tôm nuôi hàng loạt, nhưng chúng là nhân tố làm suy yếu hệ thống miễn dịch của cơ thể tôm, tạo điều kiện cho virus giết chết tôm trong ao. Mặt khác do lạm dụng việc sử dụng các loại thuốc và hóa chất ngăn ngừa mầm bệnh, rong tảo, đã dẫn đến việc tiêu diệt các vi sinh vật gây bệnh và cả vi sinh vật hữu ích.
Nhằm từng bước khắc phục và cải thiện dần môi trường phục vụ cho nghề nuôi tôm, với mục tiêu hướng đến một môi trường nuôi tôm bền vững, người ta đã ứng dụng chế phẩm sinh học nhằm quản lý môi trường ao nuôi tôm. Loại chế phẩm sinh học dược sử dụng có hiệu qủa nhất là Effective Micro-organism (gọi tắt là chế phẩm EM). Kết quả thử nghiệm cho thấy, chế phẩm sinh học EM có khả năng phân giải tốt các chất thải hữu cơ hòa tan và không hòa tan từ uế chất của tôm, từ thức ăn thừa tích tụ ở đáy ao nuôi; tạo được sự ổn định và duy trì chất lượng nước, màu nước trong ao nuôi trong qúa trình nuôi; ức chế khả năng phát triển của vi sinh vật gây hại như Vibrio, Aeromonas và những mầm bệnh vi khuẩn khác; điều hòa hàng loạt các yếu tố môi trường phát sinh trong quá trình nuôi như pH, ôxy hòa tan, cặn lắng, độ trong, H2S, NH3...
Những nhóm vi sinh vật hữu hiệu có trong chế phẩm như: Rhodopreudomonas. Lactobacillus, Strepptomyces, Sacchamyces, Aspergillus, Penicillium có vai trò "tiêu thụ" các chất hữu cơ phát sinh trong ao hồ theo qúa trình nuôi tôm. Chế phẩm EM không hoạt động ở môi trường khô, nó chỉ hoạt động mạnh trong môi trường nước. Khi gặp nước, các Enzym được kích hoạt và bắt đầu thực sự phân giải rất mạnh các chất hữu cơ trong ao. Chính sự phân giải này đã tạo ra cơ chất làm thức ăn cho các chủng vi sinh vật hữu hiệu, tăng nhanh sinh khối, phân hủy các chất thải, các chất lơ lửng rồi kết tụ lắng xuống đáy ao, giúp môi trường ao nuôi trong sạch. Qúa trình này diễn ra liên tục theo chiều hướng tích cực và có lợi cho môi trường nuôi tôm.
Căn cứ điều kiện thời tiết, yếu tố mùa vụ việc sử dụng EM giống có thay đổi nhất định. Nếu thời tiết tốt điều kiện môi trường ổn định, rãi EM 1 tuần/ lần, liều lượng bình quân 0,5 - 1 lít EM giống /sào (500m2). Nếu thời tiết thất thường, biến động liên tục, tốt nhất là rãi EM giống liên tục hàng ngày với liều lượng bình quân 0,2-0,4 lít/sào. Nên rãi đều trên mặt nước ao vào buổi sáng, tốt nhất là khi mặt trời vừa hé sáng.
Câu hỏi: 1. Đặc điểm về dinh dưỡng, khẩu phần ăn và nuôi dưỡng cá chép, cá rô phi, cá trôi
Ấn Độ, cá quả, cá chình, tôm he? 2. Các giải pháp dinh dưỡng và nuôi dưỡng nhằm nâng cao năng suất các đối tượng
nuôi kể trên?

155
CHƯƠNG XIV ĐỘC TỐ TRONG THỨC ĂN THỦY SẢN
14.1. ĐÔC TỐ TỰ NHIÊN Độc tố tự nhiên chỉ những chất và hợp chất độc hại có tự nhiên trong thức ăn. Độc
tố có thể có từ nguồn gốc thực vật, động vật và vi sinh vật.
14.1.1. Độc tố có nguồn gốc thực vật Thời xa xưa, phối hợp khẩu phần ăn cho cá chủ yếu dựa vào các thức ăn có nguồn
gốc từ động vật (Law et al., 1961). Ngày nay, do sự hiểu biết về dinh dưỡng ngày càng tăng, giá thức ăn có nguồn gốc động vật vừa tăng cao lại vừa không đủ cung ứng kịp thời nên việc sử dụng thức ăn có nguồn gốc thực vật phối hợp trong khẩu phần ăn cho tôm, cá tăng nhanh một cách rõ rệt. Những thức ăn có nguồn gốc thực vật thường được sử dụng phổ biến là hạt đỗ tương, lạc và các sản phẩm phụ từ công nghiệp chế biến như bã đậu nành, khô dầu lạc, khô dầu bông... Đây là những loại thức ăn được nhiều nhà khoa học trên thế giới và trong nước nghiên cứu nhằm thay thế với tỷ lệ thích hợp các loại thức ăn có nguồn gốc động vật trong khẩu phần ăn của ĐVTS. Tuy nhiên, một vấn đề cần đối mặt khi sử dụng nguồn thức ăn này là các độc tố làm cản trở đến khả năng tiêu hóa các chất dinh dưỡng có trong các loại thức ăn này.
Chất ức chế trypsin (Trypsin inhibitor - I-trypsin) trong đỗ tương Đỗ tương là một trong những nguồn protein thực vật quan trọng thay thế bột cá. Đỗ
tương thường được bổ sung ở dạng viên trong khẩu phần ăn của cá da trơn và các loại cá khác với một lượng hạn chế do có chứa nhiều chất ức chế (anti-metabolites) mà chủ yếu nhất là chất ức chế trypsin. Chất ức chế trypsin là một protein hình cầu có khối lượng phân tử 21.500 (Liener và Kakade, 1980). Nó làm chậm sinh trưởng của gà, chuột, cá hồi (rainbow trout), cá da trơn, cá chép và các loại khác, và gây ra triệu chứng phình to tụy ở gà và chuột do tụy tăng cường hoạt động quá mức để đáp ứng nhu cầu cao việc cung cấp enzyme trypsin cần thiết cho cơ thể (Sandholm et al. 1976). Nó không làm phình to tụy ở bê, lợn, chó (Liener và Kakade, 1980). Tuy nhiên, Robinson et al (1981) mổ khám tụy cá da trơn nuôi bằng khô đỗ tương ở các mức I-trypsin khác nhau không thấy ảnh hưởng gì. Bằng chứng đó cho thấy cơ chế giảm sinh trưởng do đỗ tương rất phức tạp không đơn giản chỉ có ức chế tiêu hóa protein. Xử lý nhiệt có thể giảm hoạt tính của I-trypsin. Điều kiện nhiệt và ẩm và thời gian khi xử lý đỗ tương phụ thuôc loại vật. Nhiệt độ không đủ không hoạt hóa được I-trypsin sẽ ức chế tiêu hóa protein (Dabrowska, Kozak, 1979), nhưng quá nhiệt làm giảm một số axit amin sẵn có trong đỗ tương, đặc biệt lysine (Riesen et al. 1947; Evans và Butts, 1951).
Viola et al. (1982) nghiên cứu trên cá chép, kết luận rằng khô đỗ tương có mức năng lượng và hàm lượng lysine thấp hơn 10 -15% so nhu cầu. Các tác giả đề nghị chỉ nên thay thế một phần bột cá bằng đỗ tương, nếu thay thế hoàn toàn sẽ tăng chi phí bổ sung axit amin và tăng mức lipid. Họ còn chỉ ra rằng bột cá chứa một số chất kích thích sinh trưởng chưa định rõ tên mà trong đỗ tương không có.
Andrew và Page (1974) phát hiện thấy khi thay thế bột cá bằng đỗ tương cùng mức N làm giảm đáng kể sinh trưởng và hiệu quả sử dụng thức ăn cá da trơn.
Robinson et al (1981) thử nghiệm ảnh hưởng xử lý nhiệt lên I-trypsin trong khẩu phần cá da trơn và cho thấy làm giảm sinh trưởng và hiệu quả sử dụng thức ăn khi khẩu phần chứa trên 1,9 TIU (trypsin inhibitor unit)/mg mẫu. Nhưng, cũng nhóm tác giả này

156
(1985), họ đã không tìm thấy ảnh hưởng của I-trypsin ở cá da trơn khi nuôi khẩu phần chứa 3,6 TIU trong một thí nghiệm khác.
Các nghiên cứu trên phát hiện nhiều đặc tính quan trọng về ảnh hưởng của I-trypsin trong nuôi trồng thủy sản. Đặc điểm thứ nhất là độ nhạy cảm với I-trypsin khác nhau ở các đối tượng thử nghiệm. Cá hồi (Rainbow trout) nhạy cảm hơn, cá da trơn có sức kháng hơn, trong khi cá chép nằm ở vị trí trung gian. Đặc điểm thứ hai là xử lý nhiệt khô đỗ tương thương mại không có quy trình chuẩn nên có mức I-trypsin khác nhau giữa các mẫu phân tích. Trên cơ sở kinh tế và ưu điểm sinh học, nên dùng nguyên hạt đỗ tương (không ép dầu) sau khi xử lý nhiệt để làm thức ăn, ít nhất cho loại cá hồi này (Smith 1977). Đặc điểm thứ ba là bổ sung amino acid và khoáng cho kết quả khả quan có thể do sai khác các giống, và chất lượng khô đỗ tương. Đặc điểm thứ tư là rất ít các nghiên cứu trên cá về ảnh hưởng của I-trypsin lên tụy.
Phytic acid Phytic acid - hexaphosphate của myo-inositol là thành
phần của hầu hết các sản phẩm hạt cốc và khô dầu như đỗ tương, bông, oliu… Ở chuột, phytate gây ảnh hưởng đáng kể đến sinh trưởng đặc biệt khi có mức Ca cao trong khẩu phần (Moris và Ellis 1980). Phytate có khả năng kết hợp với các cation như Ca, Mg và Zn, và nhất là P có trong thực vật thành hợp chất không thích hợp cho động vật (Smith 1977). Điều này có thể giải thích lợi ích của bổ sung khoáng dưới dạng khẩu phần chứa đỗ tương (Ketola 1975). Nhiều nghiên cứu chỉ ra rằng hàm lượng axit phytic cao kìm hãm sinh trưởng, chuyển hóa protein và chức năng tuyến giáp, và tăng tỷ lệ chết ở cá (Richardson et al. 1985; Satoh et al. 1983).
Glucosinolate (GL) Glucosinolate (thioglucoside) được hình thành tự nhiên trong các thực vật dạng
hình chữ nhật như bắp cải, súp lơ, cải dầu, cải xoăn… GL bao gồm 5-vinyloxazolidine-thione, nitrite hữu cơ, isothiocyanate và ino thiocyanate (R-N=C=S). Tất cả các sản phẩm thủy phân GL là những hợp chất antithyroid (Van Etten 1969; Tookey et al 1980). GL không tự gây độc nhưng khi thủy phân bởi enzyme myrosinase tạo các hợp chất làm suy yếu chức năng tuyến giáp và gây các ảnh hưởng có hại khác (Tookey et al 1980; Van Etten và Tookey 1983).
GL trở nên được chú ý đặc biệt khi sử dụng khô cải dầu - nguồn protein rẻ tiền - trong khẩu phần ăn của cá hồi. Hơn nữa, ngoài chất GL, khô cải dầu còn chứa các hợp chất độc khác như axit erucic, tannin và các phenolic khác, I-trypsin và axit phytic. GL từ
nguồn cải dầu gây cường năng tuyến giáp và giảm thyroxine (T4) huyết thanh ở cá hồi (Yurkowski et al 1978).
Gossypol Hạt bông là nguồn bổ sung protein cho vật nuôi và cá với lượng hạn chế vì chứa nhiều độc tố tư nhiên. Độc tố được biết nhiều nhất là gossypol trong tuyến sắc tố màu vàng. Các tuyến sắc tố chứa 2,4-4,8% của hạt và gossypol 39-50% khối lượng tuyến (Berardi và Goddblatt 1980). Xử lý nhiệt, ẩm hạt bông

157
chuyển gossypol tự do thành liên kết và không gây độc cho động vật. Phương pháp chế biến hiện nay có thể loại trừ 80-90% gossypol, như vậy sử dụng khô dầu cotton làm thức ăn cho vật nuôi khá an toàn ngoại trừ một số loại quá mẩn cảm như lợn (Singleton và Kratzer, 1973).
Thí nghiệm xác định ảnh hưởng của gossypol trên rainbow cho thấy cá chậm sinh trưởng khi hàm lượng 290ppm gossypol tự do trong khẩu phần (Herman, 1970). Tuy nhiên, kết quả TN của Roehm et al (1967) không thấy ảnh hưởng đến tăng trọng của rainbow khi ăn 250ppm gossypol acetate trong 18 tháng liền.
Thí nghiệm của Dorsa et al (1982) trên cá da trơn cho ăn khô dầu bông và gossypol acetate thì sinh trưởng giảm 17% ở khẩu phần khô dầu bông. Trong khi, Tilapia aurea đề kháng với gossypol và với mức dưới 0,18% không ảnh hưởng tăng trọng.
Ảnh hưởng của gossypol với các loài cá khác chưa được biết rõ. Sức kháng gossypol rất khác nhau ở các loài cá như cá hồi, cá da trơn, cá rophi, và mức gossypol vượt quá ngưởng đề kháng làm giảm sinh trưởng, tích lũy mô và thay đổi mô bệnh.
Alkaloid Alcaloid là nhóm độc tố thực vật lớn và đa dạng. Alkaloid là những hợp chất hữu
cơ có chứa nitơ và có tính kiềm nhẹ, đa số có nguồn gốc từ thảo mộc, chỉ với một liều rất nhỏ cũng tạo ra tác dụng sinh học rất mạnh trên cơ thể. Chất alkaloide đơn giản được tổng hợp sớm nhất vào năm 1886 là chất coniine, tên hóa học là 2 - propyl - piperidine (C5H10NC3H7). Đây là chất mà ngày xưa người xấu sử dụng để đầu độc nhà triết học vĩ đại Socrates. Hiện nay, người ta tìm thấy có gần 6000 chất alkaloid khác nhau và có khoảng 30 trong số này đã được sử dụng rộng rãi trong y học và được nghiên cứu kỹ. Ví dụ: chất nicotine trong cây thuốc lá được sử dụng để làm thuốc trừ sâu, cafein trong hạt cà phê, cocaine trong cây coca để làm hưng phấn thần kinh, morphine trong nhựa cây á phiện được sử dụng để làm thuốc giảm đau, reserpine trong cây dừa cạn làm thuốc giảm đau, atropin được tìm thấy trong cây cà độc dược (belladonna) để làm thuốc giãn đồng tử (Encarta, McRosoft, 1997). Theo tài liệu của Petterson (1991) thì alkaloid được chia ra làm 3 nhóm: nhóm alkaloid thực (True - alkaloids), nhóm alkaloid giả (Pseudo - alkaloids) và nhóm tiền alkaloid (Proto - alkaloids). Những alkaloid được biết rõ nhất là trong cây khoai tây và trong cỏ lupin. Ngày nay, người ta còn biết được trong một số cây nhiệt đới cũng có nhiều alkaloid. Ví dụ: cây vông nem (Erythrina sp) có chứa trên 60 loại alkaloid. Alkaloid có chứa nhiều trong hạt Erythrina americana, E. breviflora (Sotelo, 1930).
Phần củ khoai tây trồi lên trên mặt đất có vỏ xanh và phần củ khoai tây có chứa chất solanin. Khi thủy phân nó giải phóng ra đường, vì vậy người ta gọi nó đúng nghĩa hoá học là các glucoside cũng được.
Trên người: hiện tượng ngộ độc solanin thường xảy ra do ăn củ khoai tây mọc mầm. Tùy theo giống khoai tây mà hàm lượng solanin có khác nhau. Giống khoai tây Rosevall ở Angieri có chứa đến 0,49g solanin/1kg ruột củ và 1,22g/1kg vỏ củ. Các giống khác có hàm lượng solanin thấp hơn, trung bình 0,04 - 0,07 g/kg ruột củ và 0,30 - 0,55 g/kg vỏ củ. Khi khoai tây mọc mầm thì solanin có thể lên đến 1,34g/kg củ. Liều gây chết người của salonin từ 0,2 - 0,4 g/1kg thể trọng người.
Triệu chứng ngộ độc trên người ở thể nhẹ là đau bụng, tiêu chảy rồi sau đó táo bón. Thể nặng thì giãn đồng tử và liệt nhẹ hai chân. Khi khu thần kinh bị tê liệt dẫn đến ngừng hô hấp, ngừng tim và tử vong.
Trên động vật: Trong thực tiễn chăn nuôi trên đồng cỏ chăn thả có một số loài thực vật họ đậu như cỏ ngôi sao hoa trắng (Lupinus albus) hoặc hoa vàng (Lupinus luteus) có

158
một loại chất độc gây bệnh cho gia súc trên đồng cỏ xứ ôn đới, trước đây người ta gọi tên bệnh do loại cỏ này gây ra là lupinozis. Sau này, xác định trong cây cỏ Lupin có chứa nhiều loại alkaloid mà trong đó có chất kinolizidin là rất độc gây hại cho gan, làm thoái hoá và mỡ hoá gan. Alkaloid trong loại cỏ này không bị phá hủy bởi qúa trình phơi và sấy, do đó sự ngộ độc trên gia súc thường xảy ra khi cho bò ăn cỏ Lupin khô. Theo tài liệu của Humphreys (1988) thì sự ngộ độc do loại cỏ này xảy ra ở bò sữa mang thai kỳ cuối hoặc mới đẻ còn gây ra bệnh ketosis cho bò.
Trong một số cỏ họ đậu thuộc giống Medicago sativa sau một thời gian dài trồng để lấy hạt cỏ già tích lũy chất độc được biết là chất latirin còn là loại alkaloid gây ngộ độc cho gia súc ăn nhiều. Triệu chứng bệnh xuất hiện trên hệ thần kinh dẫn đến bại liệt người ta gọi là bệnh latirizmus.
Biện pháp phòng ngừa: ngâm nước và rửa nhiều lần có thể làm giảm chút ít alkaloid. Tuy vậy, điều quan trọng là về mặt di truyền, người ta cố gắng tuyển chọn giống có hàm lượng alkaloid thấp.
Nhóm chất tannin Tannin là một hợp chất ester giữa đường glucose và một nhóm chất khác, thường là
một phức hợp của axit phenolic hoặc axit oxyphenolic. Nếu đem thủy phân ra ta thu được đường glucose và một thành phần khác không phải đường, đó là axit gallic và m - digallic, như thế ta gọi là "gallotannins". Ngoài ra, người ta còn biết có một loại tannin khác gọi là "ellagitannins" nếu cắt liên kết ra ta thu được axit ellagic. Theo Kumar và D’Mello (1995) thì tannin là những hợp chất có chứa phenolic hòa tan, có phân tử lượng >500, có khả năng kết tủa với gelatin và các protein trong môi trường nước. Trong thực vật có 2 loại tannin: một loại tannin có khả năng thủy phân gọi là hydrolysable tannin (HTs) và một loại không có khả năng thủy phân gọi là condensed tannin (CTs).
Tannin phân bố rất rộng trong các loại thực vật, tuy nhiên có loại thực vật chứa nhiều, có loại ít. Thực vật càng già, đã hóa gỗ thì tannin càng nhiều. Chất gallotannin có nhiều trong lá cây non cây Shin - oak (Quereus havardi) gây tổn thất lớn cho động vật chăn thả trong mùa nó phát triển gia súc phải sử dụng loại cây này (Pigeon et al.,1962) ở Bắc Mỹ. Một dẫn chứng khác về loại độc tố này đã gây thiệt hại kinh tế khá lớn trong chăn nuôi, đó là một loại cây ở châu Úc: Terminalia oblongata (Evereist, 1974; Payner, 1975). Độc tố của nó còn có ở cây Acacia salicina.
Từ xa, người ta biết sử dụng tannin để thuộc da, bảo vệ chất đạm chống lại sự lên men phân giải của vi khuẩn. Một hướng khác, người ta còng sử dụng tannin để làm se niêm mạc ruột trị các bệnh tiêu chảy.
Cơ chế tác động của tannin trong dinh dưỡng động vật: phản ứng với protein gây kết tủa và biến tính protein làm cho nó trở nên khó tiêu hóa. Trong các loại cây cao lương, những giống nguyên thủy hàm lượng tannin rất cao. Những giống cao lương cải tiến có hàm lượng protein khá cao (11 - 13%) nhưng vì có chứa tannin nên khả năng tiêu hóa kém, protein bao bọc xung quanh hạt tinh bột, dưới tác động của tannin làm cho nó kết tủa, tiêu hóa kém, từ đó tỉ lệ tiêu hóa tinh bột còng kém theo. Nếu cao lương được hấp hơi hoặc ép dẹp làm khô cho gia súc ăn sẽ tiêu hóa tốt hơn rất nhiều.
Tannin còn có ảnh hưởng như một chất kháng dinh dưỡng. Sự có mặt của tannin trong một vài cây cỏ làm thức ăn gia súc quan trọng không những làm giảm khả năng tiêu hóa, mà còn làm giảm tính ngon miệng của gia súc, làm thay đổi trao đổi chất trong dạ cỏ và gây hại cho động vật.

159
Các axit béo cyclopropene Các axit béo cyclopropene (CPFA), axit sterculic, malvalic là các axit trong thực
vật họ Malvale. Khô dầu bông chứa các CPFA nói trên. Hàm lượng CPFA trong dầu bông khoảng 0,6-1,2% (Bailey et al. 1966). CPFA còn có trong khô dầu loại bỏ dầu nhưng hàm lượng phụ thuộc dầu còn lại, khoảng 0,01% khi chế biến bằng phương pháp hiện đại (Levi et al., 1967) và tỷ lệ giữa axit sterculic và malvic là 2:1. Nếu khẩu phần chiếm 20-30% khô dầu bông - thường dùng cho cá hồi - thì CPFA có 20-30ppm. Như vậy, chỉ có 7-10% axit sterculic hữu hiệu có trong khẩu phần cá hồi. Mức này không đủ gây ảnh hưởng sinh trưởng ở cá, đặc biệt cá lớn.
Một trong các ảnh hưởng của CPFA đến động vật có vú và lớp chim là ức chế hệ thống enzyme desaturase axit béo (Raju và Reiser, 1967; Johnson et al., 1969) và cá rainbow (Roehm et al., 1970). Một số ảnh hưởng khác lên cá rainbow là làm biến đổi trao đổi lipid (Truthers et al., 1975), biến đổi thành phần và cấu trúc lipid (Scarpelli et al., 1974), biến đổi hoạt tính của oxidase chức năng trong gan và các enzyme khác (Eisele et al., 1978, 1983), tăng tỷ lệ khối lượng giữa gan và cơ thể (Hendricks et al., 1980)... Dầu bông chứa 0,35% CPFA được sử dụng trong khẩu phần 7,5% làm cho 30% cá có khối u trong gan lúc 12 tháng. Khẩu phần này chứa khoảng 90ppm axit sterculic.
14.1.2. Độc tố có nguồn gốc động vật Trong thủy sản, thức ăn có nguồn gốc động vật như bột cá, bột máu, bột lông vũ, bột
thịt, bột đầu tôm.. được sử dụng với một lượng nhỏ trong khẩu phần. Nhìn chung, các loại này không có độc tố. Tuy nhiên, gan và buồng trứng cá có chứa 2 độc tố tự nhiên có thể ảnh hưởng khi sử dụng làm thức ăn, đó là tetrodotoxin và dinogunellin.
Tetrodotoxin Khoảng 80 loại thuộc họ
Tetraodontiformes (cá puffer hay globefish) sản sinh và chứa tetrodotoxin – rất độc đối với tât cả động vật xương sống trừ loài sản sinh độc tố trên (Fuhrman, 1974). Chưa thử nghiệm trên cá, nhưng đối với người và động vật thí nghiệm, chất này gây tê cóng môi, lưỡi và ngón tay, mất cảm giác cơ, mất cảm giác hô hấp và chết. Hầu hết cá chứa độc tố trên từ Ấn Độ dương và Tây Thái Bình dương. Ảnh hưởng của độc tố lên thức ăn cá
là chưa được hiểu rõ. Tuy nhiên, với tỷ lệ bột cá rất thấp trong khẩu phần cho cá thì độ độc hiếm khi xảy ra.
Dinogunellin Dinogunellin là lipoprotein độc, được sản sinh trong tinh dịch của hai loài cá (Stichaeus
grigorgjewi và Scorpaenichthys marmoratus). Chất này không độc như tetrodotoxin nhưng gây ỉa chảy, nôn mữa, hoại tử gan và lách trên người (Fuhrman, 1974, 1985). Khả năng gây độc cho thủy sản là rất thấp vì tỷ lệ sử dụng thấp.

160
Mỡ bị oxy hóa Mỡ của cá biển chứa nhiều các axit béo không no (PUFA) và thường bị oxy hóa bởi oxy
trong không khí (Watanabe, 1982). Sử dụng mỡ bị oxy hóa hoặc mỡ trong khẩu phần để lâu ngày bị oxy hóa gây ra độc. Các chất độc này ảnh hưởng trực tiếp hay gián tiếp đên cá, ví dụ thông qua sự thiếu hụt các vitamin, đặc biệt vitamin E (Hardy et al., 1983). Ảnh hưởng của mỡ bị oxy hóa đã được nghiên cứu trên các nhóm cá nuôi như cá hồi, da trơn và họ cá chép. Ảnh hưởng bao gồm ngưng sinh trưởng, tăng tỷ lệ chết, thiếu máu và thoái hóa mỡ gan ở cá hồi (Smith, 1979), loạn dưỡng cơ ở cá chép (Hashimoto et al., 1966). Tuy nhiên, các triệu chứng nói trên có thể khắc phục khi bổ sung vitamin E vào khẩu phần có mỡ bị oxy hóa.
14.1.3. Độc tố có nguồn gốc vi sinh vật Độc tố có nguồn gốc vi sinh vật là chất độc sinh ra từ nấm mốc, nó không phải là
hợp chất có trong nguyên liệu thức ăn do sự tổng hợp của thực vật hay động vật. Nó xuất hiện trong nguyên liệu sau quá trình thu hoạch, bảo quản và chế biến do các loại nấm mốc tạo ra. Nó gây tác hại rất lớn cho vật nuôi thủy sản và cả cho sức khỏe con người. Ví dụ, nấm Aspergillus flavus có trong khô dầu bông gây bệnh ung thư gan cho cá hồi (rainbow trout). Trong thực tế, nấm Aspergillus có thể lây nhiễm qua khô dầu bông, lạc, các sản phẩm ngô, lúa mì, gạo và đại mạch. Như đã biết, loại nấm này sản sinh các aflatoxin, trong đó aflatoxin B1 (AFB1) là chủ yếu. Nhiều nghiên cứu ảnh hưởng của AFB1 trên cá hồi đã được công bố, tuy nhiên, ít có nghiên cứu trên các loại cá khác.
Aflatoxin có trong tự nhiên Aflatoxin là nhóm độc tố thường gặp trong thức ăn gia súc. Aflatoxin do nấm
Aspergillus flavus, Aspergilus parasiticus sinh ra, là hợp chất hữu cơ có nhân đa mạch vòng có 16 dẫn xuất hóa học khác nhau, trong đó có 4 loại có tính độc lớn nhất: B1, B2, G1 và G2. Cơ chế gây bệnh của aflatoxin là do khả năng liên kết của chất này với DNA trong nhân tế bào, sự liên kết này gây ức chế enzyme polymerase của RNA làm hạn chế sự tổng hợp RNA và ức chế polymerase t - RNA. Đây là nguyên nhân làm giảm sút sự tổng hợp protein trong tế bào. Người ta còn chứng minh rằng α - , β - lactone không bão hoà có trong phân tử aflatoxin làm cho hợp chất này có hoạt tính gây ung thư, đồng thời cũng lactone này gây ức chế tổng hợp DNA nhân tế bào, do đó, làm rối loạn sự tăng trưởng bình thường của tế
bào. Các loại nông sản dễ nhiễm aflatoxin bao gồm: hạt lạc, khô dầu lạc, các hạt cốc (bắp), hạt họ đậu, khô cùi dừa, hạt hướng dương (bảng 14.1). Aflatoxin gây thương tổn gan và cú

161
thể gây ung thư. Nó còng làm giảm khả năng đẻ trứng, tiết sữa, sức đề kháng cho gia súc và gia cầm.
Giữa bốn loại trên thì loại aflatoxin B1 và G1 chiếm số lượng nhiều nhất và còn gây tác hại nhiều nhất trên cá hồi (Halver, 1965, 1967; Bauer et al., 1969); giá trị LD50 đã được xác lập cho AFB1 là 0,81 mg/kg và AFG1 là 1,90 mg/kg (Bauer et al., 1969). Trong khi đó, cá Zebra thì nhạy cảm hơn, với giá trị liều LD50 đối với AFL1: 0,44-0,58 mg/kg và AFG1: 0,75-0,83 mg/kg; AFB2: 1,0 mg/kg; AFG2: 4,2 mg/kg (Abedi, McKinley, 1968). Đối với cá hồi coho, liều LD50 là 5-10 mg/kg AFB1 (Halver et al., 1969). Thử nghiệm ảnh hưởng AFB1 trên cá da trơn cho thấy, cá này đề kháng tốt và chưa tìm thấy liều LD50. Các số liệu trên còn cho thấy, ảnh hưởng khác nhau của AF trên các đối tượng là không giống nhau.
Bảng 14.1. Hàm lượng aflatoxin trong một số thức ăn dùng cho chăn nuôi ở Việt Nam
Tên thức ăn N Hàm lượng trung bình aflatoxin (ppb)
Hàm lượng tối đa aflatoxin (ppb)
Ngô hạt Gạo và tấm Đậu nành hạt Cám gạo Khô dầu mỡ Khô dầu dừa Khô dầu đậu nành Khô dầu lạc Bột sắn lát Thức ăn hỗn hợp
25 2 1 3 3 7 4 29 1 28
205 22 50 29 8 17 12 1200 40 105
600 25 50 55 10 50 50 5000 40 500
Nguồn: Trần Văn An, 1991. Các loại mycotoxin khác Ochratoxin là nhóm độc nhất, do nấm Aspergillus ochraceus sản sinh ra khi ngô và
lúa mì bị nhiễm (Shotwell et al., 1969; Scott et al., 1970). Nhóm này gồm ochratoxin A và B và có hai hoạt chất a và b (Doster et al., 1972). Các loại thực vật dễ nhiễm là gạo, lúa mạch, lúa mì, ngô, cao lương, ớt, hạt tiêu, đậu nành, cafe. Độc tố này gây hại đến gan, thận động vật, với nồng độ lớn hơn 1 ppm làm giảm sản lượng trứng ở gà đẻ, liều trên 5 ppm có thể gây tổn hại đến gan và ruột, giảm sức đề kháng và gây ung thư ở người. Ochratoxin A gây chết người, với liều LD50 4,67 mg/kg. Nó gây thoái hóa gan, hoại tử ống lượn gần và tiểu cầu thận, trong khi đó các loại khác không gây độc. Ngoài ochratoxin, patulin được sản sinh từ nhiều loại nấm Penicillium và Aspergillus, và sinh sản rất nhanh trên các loại thức ăn hạt cốc (Scott, 1977). Ít có nghiên cứu độc tố này trên cá. Rubratoxin A và B được sản sinh do Penicellium rubrum, rất độc với động vật nhưng chưa thử nghiệm trên cá (Scott, 1977). Citrinin được sản sinh do Penicillium và Aspergillus và phát triển nhanh trên lúa mì, đại mạch và yến mạch (Scott, 1977).
Các giai đoạn và nguồn gây nhiễm độc tố nấm Trong thực tế sản xuất, có 3 nguồn nhiễm quan trọng: (i) Nhiễm ngoài đồng lúc thu
hoạch: Điều này được nhận biết rõ nhất là ngô. Muốn khắc phục tình trạng này thì khi thu hoạch không nên để lâu ngoài đồng mà đem về sấy, phơi khô liền. (ii) Nhiễm trong kho trong khi chế biến, bảo quản, dự trữ thức ăn. Nguyên nhân chủ yếu là do ẩm độ trong thức ăn còn cao (>14%), sau đó đem dự trữ hoặc do ẩm độ không khí trong kho cao hấp thu vào nguyên liệu. Ngoài ra, do chênh lệch nhiệt độ ngày đêm làm cho nước ngưng tụ bề mặt lớp thức ăn gây ra hiện tượng ẩm cục bộ, tạo điều kiện tốt cho nấm phát triển. (iii) Nhiễm khi

162
cho ăn: trường hợp này ít xảy ra với nuôi trồng thủy sản vì thời gian thức ăn tiếp xúc với môi trường nhiễm bẩn ngắn. Nhưng thường xãy ra ở trong môi trường chăn nuôi khi thức ăn tiếp xúc với máng ăn, nền chuồng nhiễm bẩn..
Những giải pháp phòng ngừa mycotoxin Có 3 nội dung quan trọng và đơn giản cần thực hiện khi tiến hành dự trữ thức ăn,
như sau: Kiểm tra đánh giá tình trạng nguyên liệu trước khi dự trữ. Giảm thiểu tối đa tình
trạng hô hấp của hạt và mầm vi sinh vật có trong nguyên liệu dự trữ. Nơi dự trữ phải có cấu trúc hợp lý để duy trì môi trường ổn định và phải ngăn chặn
không cho côn trùng, sâu mọt, các loài gặm nhấm xâm nhập vào kho. Hạt dự trữ trong kho qua một thời gian, nếu cần thiết phải xử lý thêm nhiệt để duy
trì tình trạng tốt với sự cung thờm nhiệt và hút ẩm. Có sự liên quan giữa nấm và sâu mọt. Nếu để sâu mọt phát triển vừa làm tổn thất
chất dinh dưỡng, vừa làm tăng độ ẩm nguyên liệu và vừa mang mầm vi sinh như bào tử nấm gây lan truyền nấm mốc rất mau lẹ, từ đó sản sinh ra độc tố gây hại cho người và động vật.
1. Kiểm tra, khống chế độ ẩm và nhiệt độ thớch hợp. Phải sấy khô nguyên liệu trước khi đưa vào kho dự trữ. Độ ẩm và nhiệt độ có liên quan nhau. Muốn giữ nguyên liệu tốt ta cần có qui định tình trạng hạt trong điều kiện dự trữ cụ thể. Có sự cân đối giữa ẩm độ không khí và ẩm độ nguyên liệu, và phụ thuộc vào nhiệt độ môi trường (bảng 14.2).
Bảng 14.2. Sự cân đối độ ẩm nguyên liệu và ẩm độ không khí (70%) ở nhiệt độ 27oC
Nguyên liệu Sự cân đối ẩm độ nguyên liệu ở độ ẩm không khí 70% Ngô 13,5 Lúa 15,0 Gạo 13,0 Đậu các loại 15,0 Lạc nhân 7,0
Lượng nước trong nguyên liệu có 2 dạng: dạng kết hợp, liên kết chặt với chất hữu cơ, khóang trong nguyên liệu và dạng nước tự do dễ dàng bay ra hoặc hấp thu vào nguyên liệu, ta gọi là nước hoạt động (free water hay active water). Chính lượng nước này có thể gây ra tình trạng ẩm cục bộ cho nguyên liệu, tạo điều kiện thuận lợi cho nấm mốc phát triển.
2. Kiểm soát và trừ khử côn trùng, sâu mọt trong kho bải. Người ta nhận thấy có mối liên hệ giữa sự phá hoại của sâu mọt, côn trùng trong nguyên liệu và sự phát triển nấm mốc. Điều này có thể giải thích bởi 2 lý do sau đây:
Hoạt động trao đổi chất của côn trùng sử dụng chất hữu cơ trong nguyên liệu, hô hấp sinh ra nước làm cho môi trường trữ thức ăn ngày càng ẩm thêm, tạo điều kiện thuận lợi cho nấm mốc phát triển.
Côn trùng, sâu mọt di chuyển trong nguyên liệu mang trên mình nó những bào tử nấm phát tán nhanh trong nguyên liệu. Theo tài liệu của FAO (1979) thì côn trùng, sâu mọt có thể làm tăng sự phát triển của nấm mốc lên từ 10 - 30%.
Chính vì vậy, ta cần phải kiểm tra độ nhiễm côn trùng để có biện pháp kiểm tra độ nhiễm côn trùng.
3. Sử dụng hóa chất. Người ta sử dụng nhiều loại hóa chất như Aureofugin, Thiramtan, các axit: propionic, lactic, sorbic, benzoic và axetic để trộn vào thức ăn khống

163
chế nhiễm độc nấm. Tùy từng loại nấm và nguồn lây nhiễm mà sử dụng hóa chất cho hợp lý. Ví dụ, nấm Aspergillus flavus nhiễm trên ngô hạt có thể bị khống chế bởi ammonia 2% hay axit propionic 1%.
4. Biện pháp vật lý. Nhiều phương pháp vật lý để loại trừ nấm mycotoxin trong thức ăn như loại bỏ hạt bị nhiễm để sử dụng hạt không nhiễm; loại bỏ aflatoxin trong dầu. Ngày nay, người ta đẫ chế ra hệ thống lọc hấp phụ để tách aflatoxin ra khỏi dầu 95 - 100%.
5. Làm mất hiệu lực aflatoxin. Làm mất hiệu lực aflatoxin bởi nhiệt độ: Nguyên lý của phương pháp là biến đổi thành phần hóa học của aflatoxin hoặc thay đổi nhóm hoạt động trong phân tử aflatoxin.
Ở Ấn Độ, người ta thấy khi đem gạo nhiễm aflatoxin nấu dưới áp suất hơi nước có gần 70% aflatoxin bị phá hủy. Phương pháp nhiệt như rang, nướng nhờ nhiệt độ cao có thể làm giảm aflatoxin.
Làm mất hiệu lực aflatoxin bởi ánh sáng: Ánh sáng còn được dùng để phá hủy aflatoxin. Như vậy, ánh sáng mặt trời có tác dụng tốt để phá hủy aflatoxin.
Làm mất hiệu lực aflatoxin bởi NH3 dưới áp suất cao. Mặc dù còn một số điểm liên quan đến khả năng gây biến tính của sản phẩm chưa được làm rõ, song nó được coi là một phương pháp thành công và được sử dụng ở mức công nghiệp với những đảm bảo cần thiết.
Làm mất hiệu lực aflatoxin bởi chất hấp phụ bề mặt: Người ta chọn các chất hấp phụ aflatoxin trong đường tiêu hóa, làm cho nó không hấp thu được vào cơ thể mà theo phân thải ra ngoài, từ đó không gây tác hại cho cơ thể.
Mặt trái của chất hấp phụ bề mặt là nếu không chọn lọc kỹ thì chất hấp phụ có thể gây ra sự hấp phụ vitamin và một số hoạt chất sinh học khác thải ra ngoài.
14.2. CÁC HỢP CHẤT KHÔNG TỰ NHIÊN VÀ PHỤ GIA TRONG KHẨU PHẦN
14.2.1. Hóa chất hữu cơ Các chất hữu cơ xâm nhiễm vào thức ăn dưới 2 con đường: dài hạn, xâm nhiễm
mức thấp thông qua môi trường tích lũy dần vào thức ăn; ngắn hạn, nhiễm mức cao từ rũi ro công nghiệp hay chất thải. Cả hai phương thức ô nhiễm này đều ảnh hưởng đến cá nuôi hay cá tự nhiên (Nadeau, Davis, 1976; Murchelano, Wolke, 1985).
Toxaphene Toxaphene là hỗn hợp chứa 67-69% chlorine, được sử dụng rộng rãi để phun cho
cây trồng khi DDT bị nghiêm cấm. Toxaphene đã được thấy trong cá với hàm lượng cao do sự tích lũy sinh học. Toxaphane độc hơn DDT đối với các loại cá chép nhưng ít độc hơn ở cá hồi (Macek, McAllister, 1970). Thức ăn, như đậu tương, ngô, bong và lạc trồng ở vùng có sử dụng toxaphene thì có tiềm năng nhiễm rất cao.
Polychlorinated Biphenyls (PCB) PCB là hợp chất hóa học bền, có mặt khắp nơi và gây nhiều ảnh hưởng đến người,
gia súc và cả cá. Ảnh hưởng cấp tính của PCB đã được phát hiện trên nhiều loại cá thong qua đường nước (Stalling, Mayer, 1972). Một trong số nguồn nhiễm PCB trong khẩu phần cá là dầu cá làm nguồn bổ sung lipid. Hàm lượng PCB trong dầu cá biển thấp nhưng nếu kéo dài thời gian cho ăn dầu này thì hàm lượng trong cá nuôi sẽ cao. Nguồn PCB lây nhiễm chủ yếu từ dầu họ cá trích hoặc từ mỡ thịt cá và tôm. Nói chung, PCB là nguồn lây nhiễm đối với cá nhưng không nguy hiểm.

164
14.2.2. Kim loại nặng Trong lĩnh vực độc tố học thủy sản, ảnh hưởng của kim loại nặng trong nước đến
cá đã được đề cập rất rõ ràng. Cá thường ngộ độc một số kim loại nặng như thủy ngân (Hg), cadmium (Cd) và chì (Pb). Tuy nhiên, sự ngộ độc này ít khi xảy ra.
Thủy ngân Thí nghiệm nuôi cá hồi với các mức Hg 4, 8, 16 và 24 ppm dưới dạng methyl HgCl
trên 105 ngày (Wobeser, 1975) cho thấy, hai mức cao nhất làm tăng hematocrit và tăng sinh biểu mô mang nhưng không chết. Hg đã tích tụ trong cơ dưới 30 ppm.
Cadmium Cd gây hoại tử gan, trong khi PCB làm giảm tác động tổn thương lên cá. Tỷ lệ chết
cao ở cả hai trường hợp (Rhodes et al., 1985).
Bảng 14.3. Tóm tắt một số chất kháng dinh dưỡng và độc tố trong thức ăn (De Silva, Anderson, 1995; NRC, 1993)
Hoạt chất Mô tả ảnh hưởng Nguồn thức ăn Xử lý Kháng trypsin Các chất ức chế trao đổi protein.
Nhiều loại nhưng chủ yếu ức chế trypsin và chrysotrypsin
Hầu hết hạt cốc, khô dầu (lạc, đỗ tương, bông..) và hạt bộ đậu
Xử lý nhiệt
Gossypol Giảm chất lượng protein, đặc biệt lysine. Giảm lượng ăn vào và hạn chế sinh trưởng
Hạt bông Không thể xử lý nhiệt hoặc ngâm trong nước
Mimosine Ảnh hưởng sản sinh thyroxin và sinh trưởng của cá
Keo dậu Ngâm nước
Glucosinonate Khi thủy phân sinh ra ion thiocynate, goitrin, nitrate làm giảm hoạt động tuyến giáp
Cải dầu Xử lý nhiệt, ngâm nước
Phytic acid 70% phospho trong đỗ tương.. ở dạng phytate. Phytate giảm availability của protein, Zn, Mn, Cu và Fe
Đỗ tương và nhiều cây trồng khác
Không có phương pháp xử lý hữu hiệu. Sử dụng enzyme phytase hay bổ sung khoáng
Mycotoxin Mọc trên các loại thức ăn dự trữ ẩm mốc.
Khô lạc Nên dự trữ thức ăn nơi khô, mát. Không dùng TĂ đã bị nhiễm
Erucic acid Axit béo không no gây tích lũy mỡ và hoại tử tim, da, mang và thận
Cải dầu Không xử lý được
Kim loại nặng Kim loại nặng vừa là chất dinh dưỡng nhưng vừa chất độc, ví du, Hg, Cd, As, Cu, Pb, Al, Zn..
Thỉnh thoảng gây ảnh hưởng trên cá
Tạo ra chất mang (chelator) kim loại để hạn chê độc khi bổ sung vào TĂ

165
Câu hỏi: 1. Hãy so sánh các nhóm độc tố trong thức ăn thủy sản? 2. Đặc điểm và ảnh hưởng của nhóm độc tố mycotoxin? 3. Những biện pháp loại bỏ ảnh hưởng của nhóm mycotoxin? 4. Đặc điểm của nhóm độc tố từ các chất phụ gia trong khẩu phần và biện pháp
giảm thiểu ảnh hưởng?

166
PHỤ LỤC I HỖN HỢP THỨC ĂN, PREMIX KHOÁNG, VITAMIN CHO TÔM, CÁ (Feed and Feeding of Fish and Shrimp, Michael B. New; FAO, Rome, 1987)
Bảng 1.1. Hỗn hợp khoáng số 1 (cá hồi, chép, rôphi, da trơn) (Chow, 1982)
Chất khoáng mg/g premix Fe Cu Co Mn Zn I
Se
50 3
0,01 20 30 0,1 0,1
Premix dung với mức 0,1% khẩu phần
Bảng 1.2. Hỗn hợp khoáng số 2 (Cá da trơn - Catfish) (Halver, 1982)
Chất khoáng Số lượng ( g ) CaCO3 MnSO4.H2O ZnSO4.7H2O CuSO4.5H2O FeSO4.7H2O KI (hoặc KIO4) NaH2PO4 MgSO4 NaCl
150,0 15,0 35,0 2,0 25,0 0,1
1000,0 50,0 223,0
Premix dùng với mức 1,2% trong khẩu phần ẩm
Bảng 1.3. Hỗn hợp khoáng số 3 (Cá chép Ấn Độ - Indian Carps) (Chow, 1982)
Chất khoáng ppm trong khẩu phần cuối cùng
CuSO4.5H2O FeSO4.7H2O MnSO4.H2O ZnO CoCl.6H2O KI
Cu = 10 Fe = 100 Mn = 50 Zn = 50
Co = 0,05 I = 0,1
Dùng ở mức 0,1% trong khẩu phần

167
Bảng 1.4. Hỗn hợp khoáng số 4 (Tilapia) (Jauncey và Ross, 1982) Chất khoáng Trong khẩu phần cá nước
ngọt (g/kg premix)1 Trong khẩu phần cá biển (g/kg premix)2
CaHPO4.H2O MgSO4.7H2O
NaCl KCl
FeSO4.7H2O ZnSO4.7H2O MnSO4.4H2O CuSO4.7H2O CoSO4.7H2O Ca(IO3).6H2O CrCl3.6H2O
727,7775 127,5000 60,0000 50,0000 25,0000 5,5000 2,5375 0,7850 0,4775 0,2950 0,1275
- 510.00 200,00 151,11 100,00 22,00 10,15 3,14 1,91 1,18 0,51
1Dùng ở mức 4% trong khẩu phần; 2 Dùng ở mức 1% trong khẩu phần
Bảng 1.5. Hỗn hợp vitamin (Cá hồi, chép, rôphi, da trơn) (Chow, 1982)
Vitamin mg/kg khẩu phần khô
Vitamin A - D3 - E - K - C - B1 - B2
Axit panthothenic Niacin
Vitamin B6 Biotin
Axit folic Cholin chloride
Vitamin B12 Ethoxyquin
1000 IU 200 IU 10 IU
2 40 4 4 10 20 4
0,02 1 90
0,004 16

168
Bảng 1.6. Hỗn hợp vitamin (Tôm biển, tôm nước ngọt ) Vitamin mg/g premix
(Chow, 1982) mg/kg thức ăn (Kanazawa, 1984)
Vitamin A - D3 - B1 - B2 Pyridoxine Vitamin B12 Axit Nicotinic Calcium Panthothenate Axit Folic Vitamin K Vitamin C
500 IU 100 IU 0,1 0,3 0,2 0,001 2,0 0,6 0,05 0,2 5,0
5000 IU 1000 IU 120 40 120 0,02 150 100 5 40 5000
Bảng 1.7. Công thức thức ăn hỗn hợp cá da trơn (catfish) (Lee, 1981); Tacon và Beveridge, 1981) Channel Catfish (USA- Viên khô)
Catfish (Clarias&Ophicephalus) (Thái Lan- Viên khô)
Bột cá (60% CP) 12,0 Bột máu (80%CP) 5,0 Bột lông vũ 5,0 Khô đỗ tương chiết ly 22,5 Khô dầu bông bỏ vỏ 12,5 Distillers solubles khô 8,0 Cám gạo 20,0 Tấm lúa mì 10,0 Bột cỏ alfalfa (17%CP) 3,5 Muối iot 1,0 Premix vitamin 0,5 TC: 100,0
Bột cá 56,0 Cám gạo 12,0 Khô lạc 12,0 Tinh bột (alpha) 14,0 Dầu cá 4,0 Premix K+V 1,6 Basfin (Chất kết dính) 0,4 TC: 100,0
Bảng 1.8. Hỗn hợp cho cá chép, da trơn và rôphi (Chow, 1982) Nguyên liệu Thức ăn cho cá sinh trưởng
(Mehico) Đỗ tương ép dầu Khô dầu hướng dương Khô mầm ngô Cao lương Cám lúa mì Mật rỉ Calcium orthophosphat Bột đá vôi Premix vitamin Premix khoáng
54,00% 11,07 7,34
15,00 3,00 5,00 2,35 2,54 0,60 0,10

169
Bảng 1.9. Hỗn hợp cho rô phi nước ngọt (Jauncey và Ross, 1982)
Nguyên liệu Hương 0,5 g ( % )
0,5 - 35 g ( % )
35g – cá bột ( % )
Bột cá Bột lông vũ Bột thịt Khô đỗ tương Khô lạc Khô dầu bông Cám gạo Bổng bia rượu Premix vitamin Premix khoáng Lipit bổ sung Tổng cộng
30 15 5 5 10 5 10 10 2 4 4 100
10 5 5 12 12 20 20 10 2 4 - 100
5 3 5 4 10 20 37 10 2 4 - 100
Bảng 1.10. Một số công thức thức ăn cho cá măng – Chanos chanos (Lim, 1991)
Loại thức ăn 40% protein cho cá giống 27% protein Bột cá 30 10 Khô đỗ tương 20 34,5 Bột tôm, bột mực 15 - Bột mì 25,45 - Dầu cá biển 8 - Premix vitamin 1 0,5 Premix khoáng 0,5 0,5 Chất chống oxy hóa 0,05 - Hạt cốc hay phụ phẩm - 48 Hemicellulose viên - 3 Dầu - 2 Dicalcium phosphate - 1,5
Bảng 1.11. Một số công thức thức ăn cho cá hồi Atlantic (Storebakken, 2000)
Cá bột Cá giống
Cá con Trưởng thành
Vỗ béo Brood stock
Loại thức ăn (%) Bột cá 35-60 35-60 35-60 25-50 25-50 25-50 Sản phẩm đỗ tương 0 0-5 0-5 0-15 0-15 0-15 Gluten 0-15 0-15 0-15 5-20 5-20 5-20 Hạt ngũ cốc 8-15 8-15 8-15 10-18 10-18 10-18 Dầu 10-15 10-20 10-20 20-30 10-30 10-30 Khác 3-5 3-5 3-5 3-5 3-5 3-5 Thành phần dinh dưỡng (%) Protein 50-60 50-60 50-55 35-55 35-55 50-60 Mỡ 18-25 18-30 18-30 30-40 20-40 20-35 Tinh bột 6-12 6-12 6-12 7-15 7-15 7-15
170
Bảng 1.12. Một số công thức thức ăn tự sản xuất cho ấu trùng cá da trơn và rôphi ở Thái Lan (Supis Thongrod et al., 2002 trong Feed and Feeding for Inland Aquaculture, FAO)
Nguyên liệu thức ăn Ấu trùng cá da trơn 1
Ấu trùng cá da trơn 1
Ấu trùng cá da
trơn/rôphi
Ấu trùng rôphi
Ấu trùng rôphi đơn
tính Tinh bột sắn 0 15 15 15 0 Cám gạo 0 3 10 30 30 Gluten mì 25 0 0 0 0 Bột cá 50 72 65 47 68 Dầu 7 7 7 5 0 Dicalcium phosphate 1 1 1 1 0 Premix 2 2 2 2 2 Chất mang (binder) 15 0 0 0 0 Protein thô (%) 45 40 37,5 30 41 Năng lượng (kcal/kg) 3000 3000 3000 2800 2700
Bảng 1.13. Một số công thức thức ăn tự phối chế cho cá roophi và các loại cá ăn cỏ ở Thái Lan (Supis Thongrod et al., 2002, trong Feed and Feeding for Inland Aquaculture, FAO)
Nguyên liệu phối trộn Cá giống đến 2-4 tháng
tuổi
Đến cá thương phẩm (nuôi
lồng)
Đến cá thương phẩm 1
Đến cá thương phẩm
2 Sắn khô 23 23 35 22 Khô dừa 0 0 0 30 Cám gạo 15 20 15 0 Khô đỗ tương 35 25 25 25 Bột cá 25 25 20 20 Dầu 4 4 2 0 Dicalcium phosphate 1 1 1 1 Premix 2 2 2 2 Protein thô (%) 31 27,5 24,5 26 Năng lượng (kcal/kg) 2700 2700 2500 2500 Bảng 1.14. Hỗn hợp cho tôm biển (Kanazawa, 1984); NRC, 1983)
Viên khô cho P.monodon & P. merrguiensis (Malaysia)
Khẩu phần thực hành ở ao ( USA )
Nguyên liệu Bột cá Bột thịt xương Khô đỗ tương Khô vừng ép Khô lạc ép Ngô Khô dừa
( % ) 27,0 10,0 15,0 5,0 5,0 4,0 10,0
Nguyên liệu Bột cá (61%CP) Khô đỗ tương chiết ly Bột đầu tôm Bột mì giầu gluten Cám gạo Mỡ
( % )
15,000 36,000 10,000 20,000 12,000 2,000
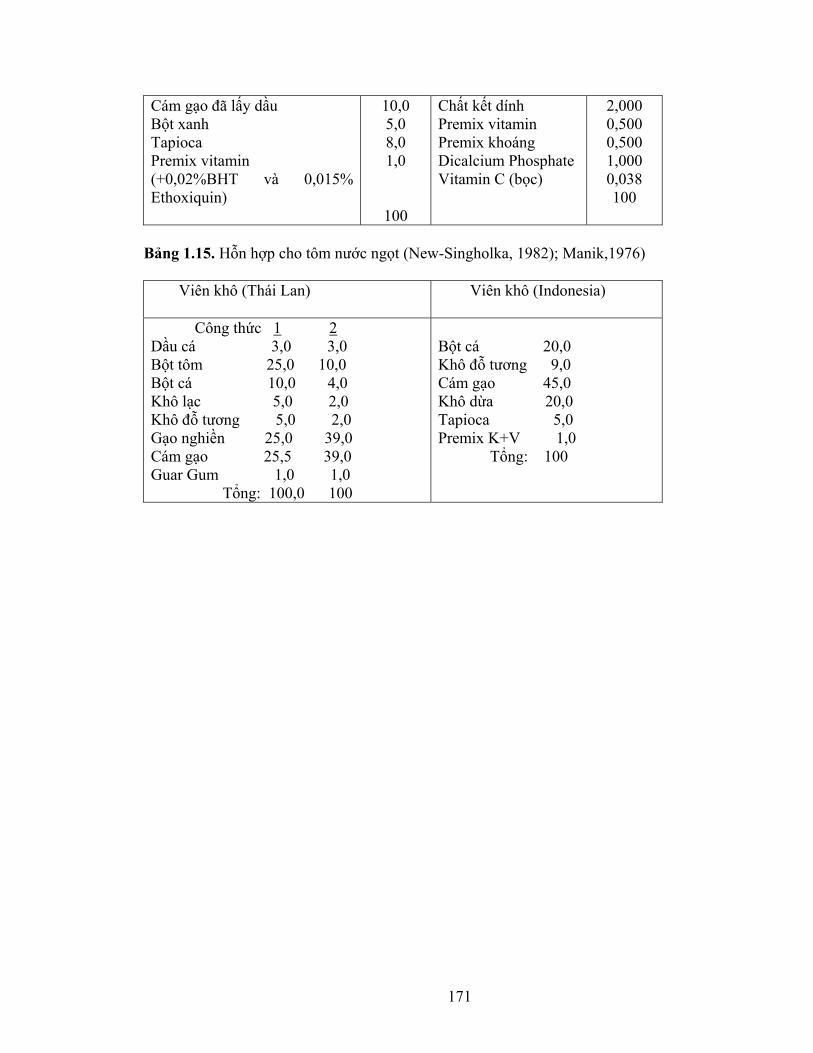
171
Cám gạo đã lấy dầu Bột xanh Tapioca Premix vitamin (+0,02%BHT và 0,015% Ethoxiquin)
10,0 5,0 8,0 1,0
100
Chất kết dính Premix vitamin Premix khoáng Dicalcium Phosphate Vitamin C (bọc)
2,000 0,500 0,500 1,000 0,038 100
Bảng 1.15. Hỗn hợp cho tôm nước ngọt (New-Singholka, 1982); Manik,1976)
Viên khô (Thái Lan)
Viên khô (Indonesia)
Công thức 1 2 Dầu cá 3,0 3,0 Bột tôm 25,0 10,0 Bột cá 10,0 4,0 Khô lạc 5,0 2,0 Khô đỗ tương 5,0 2,0 Gạo nghiền 25,0 39,0 Cám gạo 25,5 39,0 Guar Gum 1,0 1,0 Tổng: 100,0 100
Bột cá 20,0 Khô đỗ tương 9,0 Cám gạo 45,0 Khô dừa 20,0 Tapioca 5,0 Premix K+V 1,0 Tổng: 100

172
PHỤ LỤC II BẢNG NHU CẦU DINH DƯỠNG CỦA MỘT SỐ LOÀI CÁ Những giá trị về nhu cầu dinh dưỡng trình bày trong bảng dưới đây là nhu cầu tối
thiểu của cá trong điều kiện thí nghiệm. Trong thực tế sản xuất những nhu cầu này phải được thêm một số dư an toàn (margin of safety) để bù đắp cho những mất mát khi chế biến và dự trữ, sự chênh lệch về khả năng lợi dụng sinh học của các chất dinh dưỡng và chênh lệch về nhu cầu do tác động của môi trường. Nếu nồng độ năng lượng hay protein khẩu phần cao hơn hay thấp hơn các giá trị ghi trong bảng thì nhu cầu các chất dinh dưỡng khác cũng phải thay đổi theo .
Nhu cầu axit amin, axit béo, vitamin và khoáng được xác định với khẩu phần tinh chế có độ lợi dụng sinh học gần 100%, điều này cần được xem xét khi khẩu phần được phối hợp từ thức ăn tự nhiên có độ lợi dụng sinh học kém hơn.
Bảng 2.1. Nhu cầu dinh dưỡng của một số loài cá (NRC, 1993)
Cá da trơn Cá hồi
(Rainbow Trout)
Cá hồi Pacific
Cá chép Rô phi
DE (kcal/kg) Protein thô (%) Protein tiêu hoá (%) Axit amin (%): - Arginine - Histidine - Isoleucine - Leucine - Lysine - Met + Cys - Phe + Tyr - Threonine - Tryptophan - Valin n-3 fatty acids (%) n-6 fatty acids (%) Chất khoáng: Ca (%) Cl (%) Mg (%) P (%) K (%) Na (%) Cu (mg/kg) I (mg/kg) Fe (mg/kg) Mn (mg/kg)
3000 32 28
1,2 0,42 0,73 0,98 1,43 0,64 1,40 0,56 0,14 0,84
0,5 - 1,0 -
R R
0.04 0,45
R R 5
1,1 E 30 2,4
3600 38 34
1,5 0,7 0,9 1,4 1,8 1,0 1,8 0,8 0,2 1,2 1 1
1 E 0,9 E 0,05 0,6 0,7
0,6 E 3
1,1 60 13
3600 38 34
2,04 0,61 0,75 1,33 1,70 1,36 1,73 0,75 0,17 1,09 1 - 2
-
NT NT NT 0,6 0,8 NT NT
0,6 - 1,1 NT R
3200 35 28
1,31 0,64 0,76 1,00 1,74 0,94 1,98 1,19 0,24 1,1 1 1
NT NT 0,05 0,6 NT NT 3
NT 150 13
3000 32 28
1,18 0,48 0,87 0,95 1,43 0,90 1,55 1,05 0,28 0,78
- 0,5 - 1
R
NT 0,06 0,5 NT NT R
NT NT R

173
Zn (mg/kg) Se (mg/kg) Vitamin: A, UI/kg D, UI/kg E, UI/kg K, mg/kg Riboflavin, mg/kg Pantothenic acid, mg/kg Niacin, mg/kg Vitamin B12, mg/kg Choline, mg/kg Biotin, mg/kg Folate, mg/kg Thiamin, mg/kg Vitamin B6, mg/kg Myoinositol, mg/kg Vitamin C, mg/kg
20 0,25
1000-2000
500 50 R 9
15 14 R
400 R
1,5 1 3
NR 25-50
30 0,3
2500 2400 50 R 4
20 10
0,01 E 1000 0,15 1,0 1 3
300 50
R R
2500 NT 50 R 7
20 R R
800 R 2 R 6
300 50
30 NT
4000 NT 100 NT 7
30 28 NR 500 1
NR 0,5
6 NT 440 R
20 NT
NT NT 50 NT 6 10
NT NR NT NT NT NT NT NT 50
Ghi chú: R: có nhu cầu nhưng không xác định được số lượng NR: không thấy có nhu cầu khi xác định trong điều kiện thí nghiệm. NT: không thử E: ước tính
Bảng 2.2. Nhu cầu protein cho cá chép, rôphi và cá da trơn châu Á (Jantrarotai, 1996; Takeuchi et al., 2002; Murthy, 2002; Shiau, 2002; Paripatananont, 2002) Loài Nhu cầu (%) Kích cở Cá chép 30-38 Cá giống Trắm cỏ 28-35 Cá giống Cá mè 37-42 Cá bột và giống
45 Cá bột 30-36 Cá giống Rôphi 28-35 Juvenile
Cá da trơn: Clarias 40 C. macrocephalus 35
27-29 Cá giống Pangasius hypothalmus >18 Juvenile 43 Cá bột Cá lóc 30 Cá giống

174
Bảng 2.3. Nhu cầu axit amin thiết yếu (g/16 g N) của cá chép, rô phi và da trơn (NRC, 1993; Jantrarotai, 1996; Murthy, 2002; Shiau, 2002) Axit amin Cá chép Catla catla Labeo rohita Rô phi Ictalurus Agr 4,2 4,8 5,8 4,2 4,4 His 2,1 2,5 2,3 1,7 1,5 Iso 2,3 2,4 3,0 3,1 2,6 Leu 3,4 3,7 4,6 3,4 3,5 Lys 5,7 6,2 5,6 5,1 5,1 Met 3,1 3,6 2,9 2,7 2,3 Phe 6,5 3,7 4,0 3,8 5,0 Thr 3,9 5,0 4,3 3,8 2,0 Try 0,8 1,0 1,1 1,0 0,5 Val 3,6 3,6 3,8 2,8 3,0
Bảng 2.4. Nhu cầu dinh dưỡng cho các loại cá ăn thịt (Tacon, 1990)
Cá bột Giống Juvenile Sinh trưởng
Vỗ béo
Mỡ thô, % min(*) 16 14 14 12 10 Protein thô, % min 52 49 47 45 47 Lys, % min 3,08 2,9 2,78 2,66 2,78 Met 1 0,94 0,9 0,87 0,9 Cys 0,36 0,34 0,33 0,31 0,33 Carbohydrate, % 15 20 25 25 25 Ca, % max 2,5 2,5 2 2 2 P, % min 1 0,8 0,8 0,7 0,8 Mg, % min 0,08 0,07 0,07 0,06 0,07 * Tỷ lệ mỡ cá: thực vật là 7:1
Bảng 2.5. Nhu cầu dinh dưỡng cho các loại cá ăn tạp (Tacon, 1990)
Cá bột Giống Juvenile Sinh trưởng
Vỗ béo
Mỡ thô, % min(*) 8 7 7 6 5 Protein thô, % min 42 39 37 35 37 Lys, % min 2,48 2,31 2,19 2,07 2,19 Met 0,81 0,75 0,71 0,67 0,71 Cys 0,29 0,27 0,26 0,24 0,26 Carbohydrate, % 30 35 40 40 40 Ca, % max 2,5 2,5 2 2 2 P, % min 1 0,8 0,8 0,7 0,8 Mg, % min 0,08 0,07 0,07 0,06 0,07 * Tỷ lệ mỡ cá: thực vật là 7:1

175
PHỤ LỤC III THÀNH PHẦN HOÁ HỌC CỦA MỘT SỐ THỨC ĂN CHỦ YẾU
CHO CÁ Ở VIỆT NAM (%)
Loại thức ăn C
hất k
hô
Prot
ein
thô
Lipt
thô
Xơ
thô
Dẫn
suất
kh
ông đạ
m
Kho
áng
tổng
số
Ca
P
Cỏ tự nhiên hỗn hợp
24.10 2.60 0.70 6.90 11.60 2.30 0.11 0.05
Bèo cái 8.60 1.20 0.30 1.20 4.30 1.60 0.09 0.04 Bèo dâu 7.00 1.30 0.30 0.70 3.20 1.50 0.05 0.02 Bèo tấm 8.50 1.60 0.20 0.50 5.00 1.20 0.07 0.05 Bèo tây 7.60 0.80 0.30 1.50 3.60 1.40 0.16 0.04 Rau muống 10.60 2.10 0.70 1.60 4.70 1.50 0.12 0.05 Cỏ Ghi-nê 23.30 2.47 0.51 7.30 10.62 2.40 0.13 0.03 Cây Trichantera
13.68 2.08 0.60 1.72 6.07 3.21 0.86 0.04
Cỏ Ruzi 22.43 2.91 0.32 7.11 10.73 1.37 0.11 0.06 Thân lá Stylô 24.27 4.13 0.54 6.81 10.80 1.99 0.61 0.05 Củ sắn bỏ vỏ 31.5 0.90 0.60 0.70 28.6 0.70 0.08 0.05 Củ sắn bỏ vỏ khô
88.36 2.06 1.39 2.01 81.33 1.58 0.14 0.40
Củ sắn cả vỏ 27.70 0.90 0.40 1.00 24.70 0.70 0.05 0.04 Củ sắn cả vỏ khô
89.10 2.91 2.38 4.07 77.56 2.18 0.17 0.19
Cây bắp cải- lá già
11.00 2.20 0.70 1.60 4.50 2.00 0.26 0.05
Hạt đậu tương
88.48 37.02 16.30 6.39 23.87 4.91 0.29 0.56
Khô dầu bông
91.49 33.54 7.43 17.54 27.19 5.79 0.53 0.98
Khô dầu đậu tương
86.46 42.57 7.40 5.86 24.65 5.97 0.26 0.76
Khô dầu lạc nhân
90.19 45.54 6.96 5.25 26.70 5.74 0.18 0.53
Cám bổi 89.01 7.96 7.61 31.39 31.51 10.54 - - Cám gạo nếp 87.40 11.20 12.80 7.10 47.00 9.30 0.11 1.22 Cám gạo tẻ giả chày
89.00 13.70 17.90 6.80 37.00 13.60 - 1.50
Cám gạo tẻ loại I
87.57 13.00 12.03 7.77 46.40 8.37 0.17 1.56

176
Bột cá lợ 50% protein
89.00 52.80 6.10 1.80 0.10 28.20 5.35 2.79
Bột cá nhạt 40% protein
87.50 38.30 4.80 1.50 42.90 7.34 1.67
Bột cá nhạt 45% protein
89.71 64.19 10.91 1.06 0.52 31.03 5.52 2.53
Bột cá nhạt 50% protein
88.98 51.58 10.52 2.97 1.37 22.54 4.34 3.01
Bột cá nhạt 55% protein
90.26 35.55 10.25 0.89 1.17 24.40 5.09 2.88
Bột tôm 87.50 57.60 10.50 13.10 0.00 4.50 2.00 0.60 Sữa bột 96.50 26.50 26.00 38.50 5.50 0.90 0.80

177
PHỤ LỤC IV TIÊU CHUẨN NGÀNH MỘT SỐ LOẠI THỨC ĂN THỦY SẢN
4.1. Thức ăn hỗn hợp dạng viên cho tôm càng xanh - Compound pellet feed for Giant freshwater prawn - Tiêu chuẩn ngành 28 TCN 190 : 2004 4.1.1 Đối tượng và phạm vi ứng dụng
1.1 Tiêu chuẩn này quy định các yêu cầu về chất lượng và an toàn vệ sinh thú y của thức ăn hỗn hợp dạng viên (gọi tắt là thức ăn viên); được phối chế từ nhiều loại nguyên liệu đảm bảo có đủ các chất dinh dưỡng; sử dụng để ương giống và nuôi tôm càng xanh thương phẩm.
1.2 Tiêu chuẩn được áp dụng trong phạm vi cả nước đối với các cơ sở sản xuất và kinh doanh thức ăn viên cho tôm càng xanh. 4.1.2. Phân loại
Thức ăn viên cho tôm càng xanh gồm 6 loại sử dụng cho các giai đoạn phát triển của tôm với các số hiệu như sau:
Số 1: Loại dạng mảnh sử dụng cho cỡ tôm có khối lượng: 0,01 - 0,20 g/con Số 2: Loại dạng mảnh sử dụng cho cỡ tôm có khối lượng: 0,20 -2,00 g/con Số 3: Loại dạng mảnh (hoặc viên) sử dụng cho cỡ tôm có khối lượng: 2 - 5 g/con Số 4: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: 5,00 -10,00 g/con Số 5: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: 10,00 - 20,00 g/con Số 6: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: > 20,00 g/con
4.1.3. Yêu cầu kỹ thuật 3.1 Các chỉ tiêu cảm quan của thức ăn viên phải theo đúng quy định trong bảng 4.1.
Bảng 4.1. Chỉ tiêu cảm quan của thức ăn viên
TT Chỉ tiêu Yêu cầu 11 Hình dạng bên ngoài Viên hình trụ (hoặc mảnh) đều nhau, bề mặt mịn, kích
cỡ theo đúng số hiệu của từng loại thức ăn quy định trong bảng 4.2.
22 Màu sắc Nâu vàng đến nâu, đặc trưng của nguyên liệu phối chế. 33 Mùi vị Ðặc trưng của nguyên liệu phối chế, không có mùi
men mốc và mùi lạ khác. 3.2 Các chỉ tiêu lý, hóa của thức ăn viên phải theo đúng quy định trong bảng 4.2.
Bảng 4.2. Chỉ tiêu lý, hóa của thức ăn viên
Loại thức ăn TT Chỉ tiêu Số 1 Số 2 Số 3 Số 4 Số 5 Số 6 0,4 - 0,7
0,7 - 0,9
1,5
2,2
2,5
3,0
1
Kích cỡ: - Ðường kính viên (hoặc mảnh) tính bằng mm, không lớn hơn - Chiều dài so với đường kính viên (lần) nằm trong khoảng
1,5 - 2,0
2 Tỷ lệ vụn nát, tính bằng tỷ lệ % khối lượng,
2

178
không lớn hơn 3 Ðộ bền, tính theo số
giờ quan sát, không nhỏ hơn
2
4 Năng lượng thô, tính bằng kcal cho 1 kg thức ăn, không nhỏ hơn
3100 3000 2860 2800 2750 2700
5 Ðộ ẩm, tính bằng tỷ lệ % khối lượng, không lớn hơn
11
6 Hàm lượng protein thô, tính bằng tỷ lệ % khối lượng, không nhỏ hơn
35 32 30 27 25 23
7 Hàm lượng lipid thô, tính bằng tỷ lệ % khối lượng, không nhỏ hơn
5 5 5 4 4 4
8 Hàm lượng xơ thô, tính bằng tỷ lệ % khối lượng, không lớn hơn
3 3 4 4 5 5
9 Hàm lượng tro, tính bằng tỷ lệ % khối lượng, không lớn hơn
16
10 Cát sạn (tro không hòa tan trong HCl 10%), tính bằng tỷ lệ % khối lượng, không lớn hơn
2
11 Hàm lượng canxi, tính bằng tỷ lệ % khối lượng, không lớn hơn
2,3
12 Tỷ lệ canxi/ phospho nằm trong khoảng
1,0 - 1,5
13 Hàm lượng natri clorua (tỷ lệ % khối lượng), không lớn hơn
2,5
14 Hàm lượng lysine, tính bằng tỷ lệ % khối lượng, không nhỏ hơn
1,8 1,8 1,7 1,7 1,6 1,5
5 Hàm lượng
methionin, tính bằng tỷ lệ % khối lượng, không nhỏ hơn
0,89
0,89
0,84
0,76
0,72
0,72
3.3. Các chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên phải theo đúng yêu
cầu được quy định trong bảng 4.3. 3.4.

179
Bảng 4.3. Chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Côn trùng sống Không cho phép 2 Vi khuẩn gây bệnh (Salmonella) Không cho phép 3 Nấm mốc độc (Aspergillus flavus) Không cho phép 4 Chất độc hại (Aflatoxin) Không cho phép 5 Các loại kháng sinh và hóa chất đã bị cấm sử dụng theo Quyết
định số 01/2002/QÐ-BTS ngày 22/01/2002 của Bộ trưởng Bộ Thuỷ sản.
Không cho phép
4.1.4. Phương pháp thử
4.1 Lấy mẫu theo Tiêu chuẩn Việt Nam (TCVN) 4325-86, chuẩn bị mẫu thử theo Tiêu chuẩn Việt Nam TCVN 6952 : 2001.
4.2 Thử chỉ tiêu cảm quan theo Tiêu chuẩn Việt Nam TCVN 1532-1993 4.3 Thử các chỉ tiêu lý, hóa 4.3.1 Ðo kích cỡ (đường kính và chiều dài) viên thức ăn bằng thước kẹp 4.3.2 Thử tỷ lệ vụn nát của thức ăn viên bằng phương pháp sàng 4.3.3 Thử độ bền trong nước của thức ăn viên: 4.3.3.1 Dụng cụ thử :
- Cốc thủy tinh dung tích 50 ml - Ðũa thủy tinh
4.3.3.2 Cách thử: - Lấy khoảng 5,0 g thức ăn cho vào cốc thủy tinh có chứa nước trong để yên trong
vài phút. Sau đó, cứ khoảng 15 phút dùng đũa thuỷ tinh khuấy nhẹ một vòng rồi quan sát. Nếu hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng và có thể cầm nhẹ lên mà không bị vỡ nát là thức ăn chưa bị rã.
- Ðộ bền của viên thức ăn được tính bằng số giờ quan sát. Kể từ khi thả thức ăn vào cốc thuỷ tinh chứa nước cho đến khi hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng phải đúng theo quy định trong bảng 4.2.
4.3.4 Xác định năng lượng thô theo phương pháp hiện hành của Bộ Nông nghiệp và Phát triển nông thôn.
4.3.5 Xác định độ ẩm theo TCVN 4326 : 2001. 4.3.6 Xác định hàm lượng protein thô theo TCVN 4328 : 2001. 4.3.7 Xác định hàm lượng lipid thô theo TCVN 4331 : 2001. 4.3.8 Xác định hàm lượng xơ thô theo TCVN 4329 - 1993. 4.3.9 Xác định hàm lượng tro và cát sạn theo TCVN 4327 - 1993. 4.3.10 Xác định hàm lượng canxi theo TCVN 1526 - 86. 4.3.11 Xác định hàm lượng phospho theo TCVN 1525 : 2001. 4.3.12 Xác định hàm lượng natri clorua theo TCVN 4330 - 86. 4.3.13 Xác định hàm lượng lyzin theo TCVN 5281- 90. 4.3.14 Xác định hàm lượng methionin theo TCVN 5282 - 90. 4.4 Thử các chỉ tiêu vi sinh và an toàn vệ sinh thú y 4.4.1 Xác định độ nhiễm côn trùng sống theo TCVN 1540-86. 4.4.2 Xác định vi khuẩn gây bệnh theo TCVN 4829 : 2001. 4.4.3 Xác định nấm mốc độc theo TCVN 5750 -1993. 4.4.4 Xác định chất độc hại theo TCVN 4804 - 89.

180
4.4.5 Xác định các loại kháng sinh và hoá chất bị cấm sử dụng theo các quy định hiện hành.
4.1.5. Bao gói, ghi nhãn, bảo quản và vận chuyển 5.1 Bao gói 5.1.1 Tuỳ theo điều kiện sản xuất, thức ăn viên phải được đóng gói trong các loại bao
PE, hoặc bao PP, hoặc bao giấy 3 lớp. 5.1.2 Bao đựng thức ăn phải bền, kín, không rách, đã được tẩy trùng. 5.2 Ghi nhãn 5.2.1 Việc ghi nhãn trên bao đựng thức ăn viên phải theo đúng các quy định tại
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thủy sản (hướng dẫn thực hiện Quyết định số 178/1999/QÐ-TTg ngày 30/8/1999 của Thủ tướng Chính phủ ban hành Quy chế ghi nhãn hàng hóa lưu thông trong nước và hàng hóa xuất khẩu, nhập khẩu đối với hàng hóa thủy sản).
5.2.1.1 Các nội dung bắt buộc phải ghi trên nhãn: a. Tên hàng hoá b. Tên và địa chỉ của thương nhân chịu trách nhiệm về hàng hóa c. Ðịnh lượng của hàng hóa (khối lượng tịnh) d. Thành phần cấu tạo (nguyên liệu chính được sử dụng) đ. Chỉ tiêu chất lượng chủ yếu (hàm lượng protein thô, chất béo thô, độ ẩm, chất xơ
thô, hàm lượng khoáng ...) e. Ngày sản xuất, thời hạn sử dụng, thời hạn bảo quản g. Hướng dẫn bảo quản, hướng dẫn sử dụng (lượng cho ăn, số lần cho ăn, và cách
theo dõi lượng thức ăn hàng ngày) h. Xuất xứ của hàng hoá (với thức ăn được nhập khẩu) 5.2.1.2 Ngoài các nội dung bắt buộc, trên nhãn phải ghi thêm các nội dung sau: a. Cam kết: Thức ăn không chứa các chất bị cấm sử dụng theo Quyết định của Bộ
trưởng Bộ Thủy sản số 01/2002/QÐ-BTS ngày 22/01/2002. b. Số hiệu tiêu chuẩn đăng ký chất lượng của thức ăn (cấp cơ sở hoặc cấp ngành). c. Các nội dung không bắt buộc khác (nếu thấy cần thiết) ghi theo quy định trong
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thuỷ sản. 5.3 Bảo quản 5.3.1 Thức ăn viên phải đựơc bảo quản trong kho khô, sạch; để trên bục kê cao ráo,
thoáng mát và được tẩy trùng. Kho phải có biện pháp chống chuột và côn trùng phá hoại. 5.3.2 Thời gian bảo quản sản phẩm kể từ ngày sản xuất cho đến khi sử dụng không
quá 90 ngày. 5.4 Vận chuyển 5.4.1 Phương tiện vận chuyển thức ăn viên phải khô, sạch, được che mưa nắng,
không có chất độc hại, đảm bảo yêu cầu về an toàn vệ sinh thú y. 5.4.2 Khi bốc dỡ thức ăn viên phải nhẹ nhàng, tránh va đập mạnh.
4.2. Thức ăn hỗn hợp dạng viên cho tôm sú - Compound pellet feed for tiger shrimp (Penaeus monodon)- Tiêu chuẩn ngành 28 TCN 102 : 2004 4.2.1. Ðối tượng và phạm vi áp dụng
1.1 Tiêu chuẩn này quy định các yêu cầu về chất lượng và an toàn vệ sinh thú y của thức ăn hỗn hợp dạng viên (gọi tắt là thức ăn viên); được phối chế từ nhiều loại nguyên liệu đảm bảo có đủ các chất dinh dưỡng; sử dụng để ương giống và nuôi tôm Sú thương phẩm.

181
1.2 Tiêu chuẩn được áp dụng trong phạm vi cả nước đối với các cơ sở sản xuất và kinh doanh thức ăn viên cho tôm sú. 4.2.2. Phân loại
Thức ăn viên cho tôm sú gồm 6 loại sử dụng cho các giai đoạn phát triển của tôm với các số hiệu như sau:
Số 1: Loại dạng mảnh sử dụng cho cỡ tôm PL15 có khối lượng: 0,01 - 0,20 g/con Số 2: Loại dạng mảnh sử dụng cho cỡ tôm có khối lượng: 0,20 -1,00 g/con Số 3: Loại dạng mảnh (hoặc viên) sử dụng cho cỡ tôm có khối lượng: 1 - 5 g/con Số 4: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: 5,00 -10,00 g/con Số 5: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: 10,00 - 20,00 g/con Số 6: Loại dạng viên sử dụng cho cỡ tôm có khối lượng: > 20,00 g/con
4.2.3. Yêu cầu kỹ thuật 3.1 Các chỉ tiêu cảm quan của thức ăn viên phải theo đúng yêu cầu quy định trong
bảng 4.3.
Bảng 4.3. Chỉ tiêu cảm quan của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Hình dạng bên ngoài Viên hình trụ hoặc mảnh đều nhau, bề mặt mịn, kích cỡ
theo đúng số hiệu của từng loại thức ăn quy định trong bảng 4.4.
2 Màu sắc Nâu vàng đến nâu, đặc trưng của nguyên liệu phối chế. 3 Mùi vị Ðặc trưng của nguyên liệu phối chế, không có mùi men
mốc và mùi lạ khác. 3.2 Các chỉ tiêu lý, hóa của thức ăn viên phải theo đúng mức được quy định trong bảng 4.4.
Bảng 4.4. Chỉ tiêu lý, hóa của thức ăn viên
Loại thức ăn TT Chỉ tiêu Số 1 Số 2 Số 3 Số 4 Số 5 Số 6 0,6
0,8
1,2
1,8
2,2
2,5
1
Kích cỡ: - Ðường kính viên (hoặc mảnh) tính bằng mm, không lớn hơn - Chiều dài so với đường kính viên (lần) nằm trong khoảng
1,5 - 2,5
2 Tỷ lệ vụn nát (% khối lượng), không lớn hơn
2
3 Ðộ bền, tính theo số giờ quan sát, không nhỏ hơn
1
4 Năng lượng thô, tính bằng kcal cho 1 kg thức ăn, không nhỏ hơn
3400 3400 3200 3200 3000 3000
5 Ðộ ẩm, tính bằng tỷ lệ % khối lượng, không lớn hơn
11
6 Hàm lượng protein thô, % 42 40 39 38 37 35

182
khối lượng, không nhỏ hơn 7 Hàm lượng lipid thô, % khối
lượng, trong khoảng 6 - 8 6 - 8 5 - 7 5 - 7 4 - 6 4 - 6
8 Hàm lượng xơ thô, % khối lượng, không lớn hơn
3 3 4 4 5 5
9 Hàm lượng tro, % khối lượng, không lớn hơn
14 14 15 15 16 16
10 Cát sạn (tro không hòa tan trong HCl 10%), tỷ lệ % khối lượng, không lớn hơn
1 1 2 2 2 2
11 Hàm lượng canxi, % khối lượng, không lớn hơn
2,3
12 Tỷ lệ canxi/phospho nằm trong khoảng
1,0 - 1,5
13 Hàm lượng natri clorua, % khối lượng, không >
2,5
14 Hàm lượng lyzin, % khối lượng, không nhỏ hơn
2,10 2,10 1,80 1,80 1,70 1,70
15 Hàm lượng methionin, % khối lượng, không nhỏ hơn
0,90 0,90 0,80 0,80 0,70 0,70
3.3. Các chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên phải theo đúng yêu cầu được quy định trong bảng 4.5.
Bảng 4.5. Chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Côn trùng sống Không cho phép 2 Vi khuẩn gây bệnh (Salmonella) Không cho phép 3 Nấm mốc độc (Aspergillus flavus) Không cho phép 4 Chất độc hại (Aflatoxin) Không cho phép 5 Các loại kháng sinh và hóa chất đã bị cấm sử dụng theo Quyết
định số 01/2002/QÐ-BTS ngày 22/01/2002 của Bộ trưởng Bộ Thuỷ sản.
Không cho phép
4.2.4. Phương pháp thử 4.1 Lấy mẫu theo Tiêu chuẩn Việt Nam TCVN 4325-86, chuẩn bị mẫu thử theo
Tiêu chuẩn Việt Nam TCVN 6952 : 2001. 4.2 Thử chỉ tiêu cảm quan theo Tiêu chuẩn Việt Nam TCVN 1532-1993. 4.3 Thử các chỉ tiêu lý, hóa 4.3.1 Ðo kích cỡ (đường kính và chiều dài) viên thức ăn bằng thước kẹp. 4.3.2 Thử tỷ lệ vụn nát của thức ăn viên bằng phương pháp sàng. 4.3.3 Thử độ bền trong nước của thức ăn viên: 4.3.3.1 Dụng cụ thử :
- Cốc thủy tinh dung tích 50 ml - Ðũa thủy tinh.
4.3.3.2 Cách thử:

183
- Lấy khoảng 5,0 g thức ăn cho vào cốc thủy tinh có chứa nước trong để yên trong vài phút. Sau đó, cứ khoảng 15 phút dùng đũa thuỷ tinh khuấy nhẹ một vòng rồi quan sát. Nếu hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng và có thể cầm nhẹ lên mà không bị vỡ nát là thức ăn chưa bị rã.
- Ðộ bền của viên thức ăn được tính bằng số giờ quan sát. Kể từ khi thả thức ăn vào cốc thuỷ tinh chứa nước cho đến khi hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng phải đúng theo quy định trong bảng 4.4.
4.3.4 Xác định năng lượng thô theo phương pháp hiện hành của Bộ Nông nghiệp và Phát triển nông thôn.
4.3.5 Xác định độ ẩm theo Tiêu chuẩn Việt Nam TCVN 4326 : 2001. 4.3.6 Xác định hàm lượng protein thô theo TCVN 4328 : 2001. 4.3.7 Xác định hàm lượng lipid thô theo TCVN 4331 : 2001. 4.3.8 Xác định hàm lượng xơ thô theo TCVN 4329 - 1993. 4.3.9 Xác định hàm lượng tro và cát sạn theo TCVN 4327 - 1993. 4.3.10 Xác định hàm lượng canxi theo TCVN 1526 - 86. 4.3.11 Xác định hàm lượng phospho theo TCVN 1525 : 2001. 4.3.12 Xác định hàm lượng natri clorua theo TCVN 4330 - 86. 4.3.13 Xác định hàm lượng lysine theo TCVN 5281- 90. 4.3.14 Xác định hàm lượng methionine theo TCVN 5282 - 90. 4.4 Thử các chỉ tiêu vi sinh và an toàn vệ sinh thú y 4.4.1 Xác định độ nhiễm côn trùng sống theo TCVN 1540-86. 4.4.2 Xác định vi khuẩn gây bệnh theo TCVN 4829 : 2001. 4.4.3 Xác định nấm mốc độc theo TCVN 5750 -1993. 4.4.4 Xác định chất độc hại theo TCVN 4804 - 89. 4.4.5 Xác định các loại kháng sinh và hoá chất bị cấm sử dụng theo các quy định
hiện hành. 4.2.5. Bao gói, ghi nhãn, bảo quản và vận chuyển 5.1 Bao gói 5.1.1 Tuỳ theo điều kiện sản xuất, thức ăn viên phải được đóng gói trong các loại bao
PE, hoặc bao PP, hoặc bao giấy 3 lớp. 5.1.2 Bao đựng thức ăn phải bền, kín, không rách, đã được tẩy trùng. 5.2 Ghi nhãn 5.2.1 Việc ghi nhãn trên bao đựng thức ăn viên phải theo đúng các quy định tại
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thủy sản (hướng dẫn thực hiện Quyết định số 178/1999/QÐ-TTg ngày 30/8/1999 của Thủ tướng Chính phủ ban hành Quy chế ghi nhãn hàng hóa lưu thông trong nước và hàng hóa xuất khẩu, nhập khẩu đối với hàng hóa thủy sản).
5.2.1.1 Các nội dung bắt buộc phải ghi trên nhãn: a. Tên hàng hoá b. Tên và địa chỉ của thương nhân chịu trách nhiệm về hàng hóa c. Ðịnh lượng của hàng hóa (khối lượng tịnh) d. Thành phần cấu tạo (nguyên liệu chính được sử dụng) đ. Chỉ tiêu chất lượng chủ yếu (hàm lượng protein thô, chất béo thô, độ ẩm, chất xơ
thô, hàm lượng khoáng ...) e. Ngày sản xuất, thời hạn sử dụng, thời hạn bảo quản

184
g. Hướng dẫn bảo quản, hướng dẫn sử dụng (lượng cho ăn, số lần cho ăn, và cách theo dõi lượng thức ăn hàng ngày)
h. Xuất xứ của hàng hoá (với thức ăn được nhập khẩu) 5.2.1.2 Ngoài các nội dung bắt buộc, trên nhãn phải ghi thêm các nội dung sau: a. Cam kết: Thức ăn không chứa các chất bị cấm sử dụng theo Quyết định của Bộ
trưởng Bộ Thủy sản số 01/2002/QÐ-BTS ngày 22/01/2002. b. Số hiệu tiêu chuẩn đăng ký chất lượng của thức ăn (cấp cơ sở hoặc cấp ngành). c. Các nội dung không bắt buộc khác (nếu thấy cần thiết) ghi theo quy định trong
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thuỷ sản. 5.3 Bảo quản 5.3.1 Thức ăn viên phải đựơc bảo quản trong kho khô, sạch; để trên bục kê cao ráo,
thoáng mát và được tẩy trùng. Kho phải có biện pháp chống chuột và côn trùng phá hoại. 5.3.2 Thời gian bảo quản sản phẩm kể từ ngày sản xuất cho đến khi sử dụng không
quá 90 ngày. 5.4 Vận chuyển 5.4.1 Phương tiện vận chuyển thức ăn viên phải khô, sạch, được che mưa nắng,
không có chất độc hại, đảm bảo yêu cầu về an toàn vệ sinh thú y. 5.4.2 Khi bốc dỡ thức ăn viên phải nhẹ nhàng, tránh va đập mạnh.
4.3. Thức ăn hỗn hợp dạng viên cho cá tra và cá ba sa (Compound pellet feed for Pangasianodon hypophthalmus and Pangasius bocourti) - Tiêu chuẩn ngành 28 TCN 188 : 2004 4.3.1. Ðối tượng và phạm vi áp dụng
1.1 Tiêu chuẩn này quy định các yêu cầu về chất lượng và an toàn vệ sinh thú y của thức ăn hỗn hợp dạng viên (gọi tắt là thức ăn viên); được phối chế từ nhiều loại nguyên liệu đảm bảo có đủ các chất dinh dưỡng; sử dụng để ương giống và nuôi cá Tra và cá Ba sa thương phẩm.
1.2 Tiêu chuẩn được áp dụng trong phạm vi cả nước đối với các cơ sở sản xuất và kinh doanh thức ăn viên cho cá Tra và cá Ba sa.
4.3.2. Phân loại Thức ăn viên cho cá Tra và cá Ba sa gồm 6 loại sử dụng cho các giai đoạn phát
triển của cá với các số hiệu như sau: Số 1: Loại dạng mảnh (hoặc viên) sử dụng cho cỡ cá có khối lượng: < 1 g/con Số 2: Loại dạng mảnh (hoặc viên) sử dụng cho cỡ cá có khối lượng: 1 - 5 g/con Số 3: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 5 - 20 g/con Số 4: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 20 - 200 g/con Số 5: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 200 - 500 g/con Số 6: Loại dạng viên sử dụng cho cỡ cá có khối lượng: >500 g/con
4.3.3. Yêu cầu kỹ thuật 3.1 Các chỉ tiêu cảm quan của thức ăn viên cho cá Tra và cá Ba sa phải theo đúng
yêu cầu quy định trong bảng 4.6.
Bảng 4.6. Chỉ tiêu cảm quan của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Hình dạng bên ngoài Viên hình trụ (hoặc mảnh) đều nhau, bề mặt mịn, kích cỡ
theo đúng số hiệu của từng loại thức ăn quy định trong Bảng 2.

185
2 Màu sắc Nâu vàng đến nâu, đặc trưng của nguyên liệu phối chế. 3 Mùi vị Ðặc trưng của nguyên liệu phối chế, không có mùi men
mốc và mùi lạ khác.
3.2 Các chỉ tiêu lý, hóa của thức ăn viên cho cá Tra và cá Ba sa phải theo đúng mức được quy định trong bảng 4.7.
Bảng 4.7. Chỉ tiêu lý, hóa của thức ăn viên
Loại thức ăn TT Chỉ tiêu Số 1 Số 2 Số 3 Số 4 Số 5 Số 6 1,0
1,5
2,5
5,0
10,0
12,0
1
Kích cỡ: - Ðường kính viên tính bằng mm, không lớn hơn - Chiều dài so với đường kính viên (lần) nằm trong khoảng
1,0 - 1,5
2 Tỷ lệ vụn nát, tính bằng % khối lượng, không lớn hơn
2
3 Ðộ bền, tính theo số phút quan sát, không nhỏ hơn
30
4 Năng lượng thô, kcal/1 kg thức ăn, không nhỏ hơn
3300 2800 2400 2100 1800 1500
5 Ðộ ẩm, tính bằng tỷ lệ % khối lượng, không lớn hơn
11
6 Hàm lượng protein thô, % khối lượng, không nhỏ hơn
40 35 30 26 22 18
7 Hàm lượng lipid thô, % khối lượng, không nhỏ hơn
8 6 5 5 4 3
8 Hàm lượng xơ thô, tỷ lệ % khối lượng, không lớn hơn
6 6 7 7 8 8
9 Hàm lượng tro, tỷ lệ % Khối lượng, không lớn hơn
16 14 12 10 10 10
10 Cát sạn (tro không hòa tan trong HCl 10%), tỷ lệ % khối lượng, không lớn hơn
2
11 Hàm lượng phospho, tính bằng tỷ lệ % khối lượng, không nhỏ hơn
1
12 Hàm lượng natri clorua, % khối lượng, không lớn hơn
2,5
13 Hàm lượng lysine, % khối lượng, không nhỏ hơn
2,0 1,8 1,5 1,3 1,1 0,9
14 Hàm lượng methionine, % khối lượng, không nhỏ hơn
0,9 0,8 0,7 0,6 0,5 0,4

186
3.3. Các chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên cho cá Tra và cá Ba sa phải theo đúng yêu cầu được quy định trong Bảng 4.8.
Bảng 4.8. Chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Côn trùng sống Không cho phép 2 Vi khuẩn gây bệnh (Salmonella) Không cho phép 3 Nấm mốc độc (Aspergillus flavus) Không cho phép 4 Chất độc hại (Aflatoxin) Không cho phép 5 Các loại kháng sinh và hóa chất đã bị cấm sử dụng theo Quyết
định số 01/2002/QÐ-BTS ngày 22/01/2002 của Bộ trưởng Bộ Thuỷ sản.
Không cho phép
4.3.4. Phương pháp thử 4.1 Lấy mẫu theo Tiêu chuẩn Việt Nam TCVN 4325-86, chuẩn bị mẫu thử theo
Tiêu chuẩn Việt Nam TCVN 6952 : 2001. 4.2 Thử chỉ tiêu cảm quan theo Tiêu chuẩn Việt Nam TCVN 1532-1993. 4.3 Thử các chỉ tiêu lý, hóa 4.3.1 Ðo kích cỡ (đường kính và chiều dài) viên thức ăn bằng thước kẹp. 4.3.2 Thử tỷ lệ vụn nát của thức ăn viên bằng phương pháp sàng. 4.3.3 Thử độ bền trong nước của thức ăn viên: 4.3.3.1 Dụng cụ thử : - Cốc thủy tinh dung tích 50 ml - Ðũa thủy tinh 4.3.3.2 Cách thử: - Lấy khoảng 5,0 g thức ăn cho vào cốc thủy tinh có chứa nước trong để yên trong
vài phút. Sau đó, cứ khoảng 15 phút dùng đũa thuỷ tinh khuấy nhẹ một vòng rồi quan sát. Nếu hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng và có thể cầm nhẹ lên mà không bị vỡ nát là thức ăn chưa bị rã.
- Ðộ bền của viên thức ăn được tính bằng số phút quan sát. Kể từ khi thả thức ăn vào cốc thuỷ tinh chứa nước cho đến khi hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng phải đúng theo quy định trong bảng 4.7.
4.3.4 Xác định năng lượng thô theo phương pháp hiện hành của Bộ Nông nghiệp và Phát triển nông thôn.
4.3.5 Xác định độ ẩm theo TCVN 4326 : 2001. 4.3.6 Xác định hàm lượng protein thô theo TCVN 4328 : 2001. 4.3.7 Xác định hàm lượng lipid thô theo TCVN 4331 : 2001. 4.3.8 Xác định hàm lượng xơ thô theo TCVN 4329 - 1993. 4.3.9 Xác định hàm lượng tro và cát sạn theo TCVN 4327 - 1993. 4.3.10 Xác định hàm lượng phospho theo TCVN 1525 : 2001. 4.3.11 Xác định hàm lượng natri clorua theo TCVN 4330 - 86. 4.3.12 Xác định hàm lượng lyzin theo TCVN 5281- 90. 4.3.13 Xác định hàm lượng methionin theo TCVN 5282 - 90. 4.4 Thử các chỉ tiêu vi sinh và an toàn vệ sinh thú y 4.4.1 Xác định độ nhiễm côn trùng sống theo TCVN 1540-86. 4.4.2 Xác định vi khuẩn gây bệnh theo TCVN 4829 : 2001. 4.4.3 Xác định nấm mốc độc theo TCVN 5750 -1993.

187
4.4.4 Xác định chất độc hại theo TCVN 4804 - 89. 4.4.5 Xác định các loại kháng sinh và hoá chất bị cấm sử dụng trong thức ăn theo
các quy định hiện hành 4.3.5. Bao gói, ghi nhãn, bảo quản và vận chuyển
5.1 Bao gói 5.1.1 Tuỳ theo điều kiện sản xuất, thức ăn viên phải được đóng gói trong các loại bao
PE, hoặc bao PP, hoặc bao giấy 3 lớp. 5.1.2 Bao đựng thức ăn phải bền, kín, không rách, đã được tẩy trùng. 5.2 Ghi nhãn 5.2.1 Việc ghi nhãn trên bao đựng thức ăn viên phải theo đúng các quy định tại
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thủy sản (hướng dẫn thực hiện Quyết định số 178/1999/QÐ-TTg ngày 30/8/1999 của Thủ tướng Chính phủ ban hành Quy chế ghi nhãn hàng hóa lưu thông trong nước và hàng hóa xuất khẩu, nhập khẩu đối với hàng hóa thủy sản).
5.2.1.1 Các nội dung bắt buộc phải ghi trên nhãn: a. Tên hàng hoá b. Tên và địa chỉ của thương nhân chịu trách nhiệm về hàng hóa c. Ðịnh lượng của hàng hóa (khối lượng tịnh) d. Thành phần cấu tạo (nguyên liệu chính được sử dụng) đ. Chỉ tiêu chất lượng chủ yếu (hàm lượng protein thô, chất béo thô, độ ẩm, chất xơ
thô, hàm lượng khoáng ...) e. Ngày sản xuất, thời hạn sử dụng, thời hạn bảo quản g. Hướng dẫn bảo quản, hướng dẫn sử dụng (lượng cho ăn, số lần cho ăn, và cách
theo dõi lượng thức ăn hàng ngày) h. Xuất xứ của hàng hoá (với thức ăn được nhập khẩu) 5.2.1.2 Ngoài các nội dung ắt buộc, trên nhãn phải ghi thêm các nội dung sau: a. Cam kết: Thức ăn không chứa các chất bị cấm sử dụng theo Quyết định của Bộ
trưởng Bộ Thủy sản số 01/2002/QÐ-BTS ngày 22/01/2002. b. Số hiệu tiêu chuẩn đăng ký chất lượng của thức ăn (cấp cơ sở hoặc cấp ngành). c. Các nội dung không bắt buộc khác (nếu thấy cần thiết) ghi theo quy định trong
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thuỷ sản. 5.3 Bảo quản 5.3.1 Thức ăn viên phải đựơc bảo quản trong kho khô, sạch; để trên bục kê cao ráo,
thoáng mát và được tẩy trùng. Kho phải có biện pháp chống chuột và côn trùng phá hoại. 5.3.2 Thời gian bảo quản sản phẩm kể từ ngày sản xuất cho đến khi sử dụng không
quá 90 ngày. 5.4 Vận chuyển 5.4.1 Phương tiện vận chuyển thức ăn viên phải khô, sạch, được che mưa nắng,
không có chất độc hại, đảm bảo yêu cầu về an toàn vệ sinh thú y. 5.4.2 Khi bốc dỡ thức ăn viên phải nhẹ nhàng, tránh va đập mạnh.
4.4. Thức ăn hỗn hợp dạng viên cho cá Rô phi (Compound pellet feed for Oreochromiss sp) - Tiêu chuẩn ngành 28 TCN 189 : 2004 4.4.1. Ðối tượng và phạm vi áp dụng
1.1 Tiêu chuẩn này quy định các yêu cầu về chất lượng và an toàn vệ sinh thú y của thức ăn hỗn hợp dạng viên (gọi tắt là thức ăn viên); được phối chế từ nhiều loại nguyên

188
liệu đảm bảo có đủ các chất dinh dưỡng; sử dụng để ương giống và nuôi cá Rô phi thương phẩm.
1.2 Tiêu chuẩn được áp dụng trong phạm vi cả nước đối với các cơ sở sản xuất và kinh doanh thức ăn viên cho cá Rô phi. 4.4.2. Phân loại
Thức ăn viên cho cá Rô phi gồm 6 loại sử dụng cho các giai đoạn phát triển của cá với các số hiệu như sau:
Số 1: Loại dạng mảnh (hoặc viên) sử dụng cho cỡ cá có khối lượng: nhỏ hơn 5,0 g/con Số 2: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 5,0 -10,0 g/con Số 3: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 10,0 - 20,0 g/con Số 4: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 20,0 - 200,0 g/con Số 5: Loại dạng viên sử dụng cho cỡ cá có khối lượng: 200,0 - 500,0 g/con Số 6: Loại dạng viên sử dụng cho cỡ cá có khối lượng: lớn hơn 500,0 g/con
4.4.3. Yêu cầu kỹ thuật 3.1 Các chỉ tiêu cảm quan của thức ăn viên cho cá Rô phi phải theo đúng yêu cầu
quy định trong bảng 4.9.
Bảng 4.9. Chỉ tiêu cảm quan của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Hình dạng bên ngoài Viên hình trụ/hình tròn (hoặc mảnh), đều nhau, bề mặt
mịn, kích cỡ theo đúng số hiệu của từng loại thức ăn quy định trong Bảng 2.
2 Màu sắc Nâu vàng đến nâu, đặc trưng của nguyên liệu phối chế. 3 Mùi vị Ðặc trưng của nguyên liệu phối chế, không có mùi men
mốc và mùi lạ khác. 3.2 Các chỉ tiêu lý, hóa của thức ăn viên cho cá Rô phi phải theo đúng mức được
quy định trong bảng 4.10.
Bảng 4.10. Chỉ tiêu lý, hóa của thức ăn viên
Loại thức ăn TT Chỉ tiêu Số 1 Số 2 Số 3 Số 4 Số 5 Số 6 1,0 1,5 2,0 4,0 4,0 6,0
1
Kích cỡ: - Ðường kính viên tính bằng mm, không lớn hơn - Chiều dài so với đường kính viên (lần) nằm trong khoảng
1,0 - 1,5
2 Tỷ lệ vụn nát, % khối lượng, không lớn hơn
2
3 Ðộ bền, tính theo số phút quan sát, không nhỏ hơn
30
4 Năng lượng thô, kcal/1 kg thức ăn, không nhỏ hơn
3200 3000 2860 2800 2750 2700
5 Ðộ ẩm, tính bằng tỷ lệ % khối lượng, không lớn hơn
11
6 Hàm lượng protein thô, % 40 35 30 27 25 20

189
khối lượng, không nhỏ hơn 7 Hàm lượng lipid thô, % khối
lượng, không nhỏ hơn 6 6 5 5 4 4
8 Hàm lượng xơ thô, % khối lượng, không lớn hơn
5 5 6 6 7 7
9 Hàm lượng tro, % khối lượng, không lớn hơn
16
10 Cát sạn (tro không hòa tan trong HCl 10%), % khối lượng, không lớn hơn
2
11 Hàm lượng canxi, % khối lượng, không lớn hơn
2,5
12 Tỷ lệ canxi/phospho nằm trong khoảng
1,0 - 1,5
13 Hàm lượng natri clorua, % khối lượng, không lớn hơn
2,5
14 Hàm lượng lyzin, % khối lượng, không nhỏ hơn
1,7 1,6 1,4 1,3 1,1 0,9
15 Hàm lượng methionin, % khối lượng, không <
1,0 0,9 0,8 0,7 0,6 0,5
3.3. Các chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên cho cá Rô phi phải theo đúng yêu cầu được quy định trong bảng 4.11.
Bảng 4.11. Chỉ tiêu vi sinh và an toàn vệ sinh thú y của thức ăn viên
TT Chỉ tiêu Yêu cầu 1 Côn trùng sống Không cho phép 2 Vi khuẩn gây bệnh (Salmonella) Không cho phép 3 Nấm mốc độc (Aspergillus flavus) Không cho phép 4 Chất độc hại (Aflatoxin) Không cho phép 5 Các loại kháng sinh và hóa chất đã bị cấm sử dụng theo Quyết
định số 01/2002/QÐ-BTS ngày 22/01/2002 của Bộ trưởng Bộ Thuỷ sản.
Không cho phép
4.4.4. Phương pháp thử 4.1 Lấy mẫu theo Tiêu chuẩn Việt Nam TCVN 4325-86, chuẩn bị mẫu thử theo
Tiêu chuẩn Việt Nam TCVN 6952 : 2001. 4.2 Thử chỉ tiêu cảm quan theo Tiêu chuẩn Việt Nam TCVN 1532-1993. 4.3 Thử các chỉ tiêu lý, hóa 4.3.1 Ðo kích cỡ (đường kính và chiều dài) viên thức ăn bằng thước kẹp. 4.3.2 Thử tỷ lệ vụn nát của thức ăn viên bằng phương pháp sàng. 4.3.3 Thử độ bền trong nước của thức ăn viên: 4.3.3.1 Dụng cụ thử : - Cốc thủy tinh dung tích 50 ml - Ðũa thủy tinh 4.3.3.2 Cách thử: - Lấy khoảng 5,0 g thức ăn cho vào cốc thủy tinh có chứa nước trong để yên trong
vài phút. Sau đó, cứ khoảng 15 phút dùng đũa thuỷ tinh khuấy nhẹ một vòng rồi quan sát.

190
Nếu hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng và có thể cầm nhẹ lên mà không bị vỡ nát là thức ăn chưa bị rã.
- Ðộ bền của viên thức ăn được tính bằng số phút quan sát. Kể từ khi thả thức ăn vào cốc thuỷ tinh chứa nước cho đến khi hầu hết các viên thức ăn vẫn còn giữ nguyên hình dạng phải đúng theo quy định trong Bảng 4.9.
4.3.4 Xác định năng lượng thô theo phương pháp hiện hành của Bộ Nông nghiệp và Phát triển nông thôn.
4.3.5 Xác định độ ẩm theo Tiêu chuẩn Việt Nam TCVN 4326 : 2001. 4.3.6 Xác định hàm lượng protein thô theo TCVN 4328 : 2001. 4.3.7 Xác định hàm lượng lipid thô theo TCVN 4331 : 2001. 4.3.8 Xác định hàm lượng xơ thô theo TCVN 4329 - 1993. 4.3.9 Xác định hàm lượng tro và cát sạn theo TCVN 4327 - 1993. 4.3.10 Xác định hàm lượng canxi theo TCVN 1526 - 86. 4.3.11 Xác định hàm lượng phospho theo TCVN 1525 : 2001. 4.3.12 Xác định hàm lượng natri clorua theo TCVN 4330 - 86. 4.3.13 Xác định hàm lượng lysine theo TCVN 5281- 90. 4.3.14 Xác định hàm lượng methionine theo TCVN 5282 - 90. 4.4 Thử các chỉ tiêu vi sinh và an toàn vệ sinh thú y 4.4.1 Xác định độ nhiễm côn trùng sống theo TCVN 1540-86. 4.4.2 Xác định vi khuẩn gây bệnh theo TCVN 4829 : 2001. 4.4.3 Xác định nấm mốc độc theo TCVN 5750 -1993. 4.4.4 Xác định chất độc hại theo TCVN 4804 - 89. 4.4.5 Xác định các loại kháng sinh và hoá chất bị cấm sử dụng theo các quy định
hiện hành. 4.4.5. Bao gói, ghi nhãn, bảo quản và vận chuyển
5.1 Bao gói 5.1.1 Tuỳ theo điều kiện sản xuất, thức ăn viên phải được đóng gói trong các loại bao
PE, hoặc bao PP, hoặc bao giấy 3 lớp. 5.1.2 Bao đựng thức ăn phải bền, kín, không rách, đã được tẩy trùng. 5.2 Ghi nhãn 5.2.1 Việc ghi nhãn trên bao đựng thức ăn viên phải theo đúng các quy định tại
Thông tư số 03/2000/TT-BTS ngày 22/9/2000 của Bộ Thủy sản (hướng dẫn thực hiện Quyết định số 178/1999/QÐ-TTg ngày 30/8/1999 của Thủ tướng Chính phủ ban hành Quy chế ghi nhãn hàng hóa lưu thông trong nước và hàng hóa xuất khẩu, nhập khẩu đối với hàng hóa thủy sản).
5.2.1.1 Các nội dung bắt buộc phải ghi trên nhãn: a. Tên hàng hoá b. Tên và địa chỉ của thương nhân chịu trách nhiệm về hàng hóa c. Ðịnh lượng của hàng hóa (khối lượng tịnh) d. Thành phần cấu tạo (nguyên liệu chính được sử dụng) đ. Chỉ tiêu chất lượng chủ yếu (hàm lượng protein thô, chất béo thô, độ ẩm, chất xơ
thô, hàm lượng khoáng ...) e. Ngày sản xuất, thời hạn sử dụng, thời hạn bảo quản g. Hướng dẫn bảo quản, hướng dẫn sử dụng (khuyến cáo về mật độ nuôi, lượng cho
ăn, số lần cho ăn, và cách theo dõi lượng thức ăn hàng ngày) h. Xuất xứ của hàng hoá (với thức ăn được nhập khẩu)

191
5.2.1.2 Ngoài các nội dung bắt buộc, trên nhãn phải ghi thêm các nội dung sau: a. Cam kết: Thức ăn không chứa các chất bị cấm sử dụng theo Quyết định của Bộ
trưởng Bộ Thủy sản số 01/2002/QÐ-BTS ngày 22/01/2002. b. Số hiệu tiêu chuẩn đăng ký chất lượng của thức ăn (cấp cơ sở hoặc cấp ngành). c. Các nội dung không bắt buộc khác (nếu thấy cần thiết) ghi theo quy định trong
Thông tư số 03/2000/TT-BTS ngày 22/9/2000. 5.3 Bảo quản 5.3.1 Thức ăn viên phải đựơc bảo quản trong kho khô, sạch; để trên bục kê cao ráo,
thoáng mát và được tẩy trùng. Kho phải có biện pháp chống chuột và côn trùng phá hoại. 5.3.2 Thời gian bảo quản sản phẩm kể từ ngày sản xuất cho đến khi sử dụng không
quá 90 ngày. 5.4 Vận chuyển 5.4.1 Phương tiện vận chuyển thức ăn viên phải khô, sạch, được che mưa nắng,
không có chất độc hại, đảm bảo yêu cầu về an toàn vệ sinh thú y. 5.4.2 Khi bốc dỡ thức ăn viên phải nhẹ nhàng, tránh va đập mạnh
192
PHỤ LỤC V TÊN KHOA HỌC CỦA MỘT SỐ LOẠI CÁ
Tiếng Anh Tên latin Tiếng Việt Anabantids
Climbing perch Anabas testudibneus Gour Osphronemus goramy
Carp, common Cyprinus carpio Cá chép Chinese Parabramis pekinensis Cá chép Trung Quốc Grass Ctenopharyngodon idella Cá trắm cỏ Mud Cirrhina molitorella Cá trôi Silver Hypophthalamichthys molitrix Cá mè
Catfish Họ cá da trơn African Clarias gariepinus Brackish water Chrysichthys nigrodigitalus Channel Ictalurus punctatus Malaysian Pangasius pangasius Walking Clarias batrachus
Eel Họ cá chình Japanese Anguilla japonica Cá chình Nhật Bản European Angiulla anguilla Cá chình châu Âu
Rabbit fish Siganus spp Nhóm cá dìa Salmonids Họ cá hồi
Atlantic salmon Salmo salar Pink salmon Oncorhynchus gorbuscha Lake trout Salvelinus namaycush Rainbow trout Oncorhynchus mykiss
Bass Họ cá vược Black sea Centropristis striata European sea Dicentrarchus labrax Giant sea perch Lates calcarifer
Snakehead Ophicephalus spp Cá lóc Tilapia Họ cá rôphi
Blue Oreochromis discolor Java O. aureus Nile O. niloticus
Whitefish Coregonus clupeaformis Yellow perch Perca flavescens Yellowtail Seriola quiqueradiata Cá cam Milkfish Chanos chanos Cá măng Snapper Lujanus spp. Cá hanh, cá chỉ vàng

193
TÀI LIỆU THAM KHẢO CHÍNH Tiếng Việt Vũ Duy Giảng, Nguyễn Thị Lương Hồng, Tôn Thất Sơn, 1997. Dinh dưỡng và thức ăn gia
súc. Nhà XBNN, Hà Nội. Vũ Duy Giảng, 2001. Giáo trình dinh dưỡng và thức ăn gia súc (Dành cho sinh viên cao
học). Nhà XBNN, Hà Nội. Lại Văn Hùng, 2004. Dinh dưỡng và thức ăn trong nuôi trồng thủy sản. Nhà XBNN, Hồ
Chí Minh. Lê Đức Ngoan, 2002. Giáo trình dinh dưỡng gia súc. Nhà XBNN, Hà Nội. Lê Đức Ngoan, Nguyễn Thị Hoa Lý, Dư Thanh Hằng, 2005. Giáo trình thức ăn gia súc.
Nhà XBNN, Hà Nội. Viện chăn nuôi, 2002. Bảng thành phần hóa học và giá trị dinh dưỡng thức ăn gia súc. Nhà
XBNN, Hà nội. Tiếng Anh Akiyama, T., Oohara, I. and Yamamoto, T., 1997. Comparision of essential amino acid
requirements with A/E ratio among fish species (review paper). Fisheries Science 63, 963-970.
AOAC, 1990. Official Methods of Analysis. 15th Ed. Washington, DC.Arai, S., 1991. Eel, Anguilla spp. In: Wilson, R.P. (ed) Handbook of Nutrient requirements of Finfish, CRC Press, Boca Raton, Florida, pp. 69-75.
Arai, S., Nose, T. and Hashimoto, Y., 1972. Amino acids essential for the growth of eels, Anguilla anguilla and A.japonica. Bulletin of the Japanese Society of Scientific Fisheries 38, 753 - 759.
Boonyaratpalin, M., 1981. Vitamin Requirement in Snakehead Diets. Progress report of the Regional Project RAS/76/003. Network of Aquaculture Centers in Asia, Bangkok, Thailand, 18pp.
Carter, C. G., Houlihan, D. F., and Owen, S. F. (1998). J. Fish Biol. 53, 272 - 284. Cowey, C. B., and Walton, M.J. (1989). In “Fish nutrition,” 2nd ed. (J. E. Halver, ed), pp.
259 - 329. Acedemic Press, New York. De Silva, K.H.G.M, 1989. Cheap source of alternate feed for the farming of snakehead fish
(Teleostei: Channidae) in Sri Lanka. In: Huisman, E.A., Zonneveld, N. and Bouwmans, A.H.M. (eds) Aquacultural Research in Asia: Management Techniques and Nutrition. Proceedings of the Asia Seminar on Aquaculture, Malang, Indonesia, pp, 241 - 267.
Degani, G., Gallagher, M.L, and Levanon, D., 1987. Effect of dietary protein and energy levels on growth of European eels (Anguilla anguilla). Boletim de fisiologia animal da Universidale de Sao Paulo 11, 95 - 100.
Ensminger, M.E., 1991. Animal Science. 9th Edition. Interstate Publishers, INC., Illinois FAO. 1995. Farm-made Aquafeed. FAO Fisheries Technical Paper 343. Rome. Gerald M. Ludwig, 1999. Zooplankton Succession and Larval Fish Culture in Freshwater
Ponds - SRAC Publication No. 700. Granvil D. Treece and D. Allen Davis, 2000. Culture of Small Zooplankton for the
Feeding of Larval Fish – SRAC Publication No. 701

194
Granvil D. Treece, 2000. Arrtemia Procduction for Marine Larval Fish Culture - SRAC Publication No. 702.
Garzon, A., preagon, J., Hidalogo, M., Cardenete, G., Lupianez, J. A., and de la Higuera, M.(1994). Aquaculture 124, 64 (abstr).
Gurr, M. I., and Harwood, J.l. (1991). In “Lipid Biochemistry,” vol. 93, p.375. Chapman and Hall, London.
Edwards, P. and Allan, G. L. (eds), 2004. Feeds and Feeding for Inland Aquaculture in Mekong Region Countries. Australian Centre for International Agricultural Research, Canberra 2004
Halver, J. E., Hardy, R.W., 2002. Fish Nutrition. 3rd Eds. Academic Press, Imprint of Elsevier Science, 823pp.
Laurent Verschuere, Geert Rombaut, Patrick Sorgeloos, Willy Verstraete, 2000. Probiotic Bacteria as Biological Control Agents in Aquaculture
Ling, S.W., 1977. Aquaculture in Southeast Asia: A Historical Overview. University of Washington Press, Seattle, Washington, 108 pp.
McDonald, Edward, Greenhalgh và Morgan, 2002. Animal Nutrition. 6th Ed, Longman Scientific & Technical.
Michael B. New, 1987. Feed and Feeding of Fish and Shrimp (A manual on the preparation and presentatition of compound feed for shrimp and fish in aquaculture), UNDP, FAO, Rome.
Nose, T. and Arai, S., 1979. Recent advances in studies on mineral nutrition of fish in Japan. In: Pillay, T.V.R and Dill, W.A. (eds) Advances in Aquaculture, Fishing News, Farnham, England, pp, 584 - 590.
Nose, T., 1979. Summary report the requirements of essential amino acids for carp. In: Tiews, K. and Halver, J.E (eds) Finfish Nutrition and fishfeed technology. Heenemann, Berlin, pp, 145-146
NRC (National Research Council), 1993. Nutrient Requirements of Fish. National Academy Press, Washington, DC, USA.
Primary Industries and Resources SA (www.pir.sa.gov.au/factsheets). Water quality in freswater aquaculture ponds - FS No. 60/01
Robert R. Stickney, 2000. Encyclopedia of Aquaculture. Rottmann, R.W., J. Scott Graves, Craig Watson and Roy P.E. Yanong, 2003. Culture
Techniques of Moina: The Ideal daphnia for Feeding freshwater Fish Fry – University Florida, IFAS Extension.
Schmitz, O., Greuel, E. and Peffer, E., 1984. Digestibility of crude protein and organic matter of potential source of dietary protein for eels (A. anguilla, L.). Aquaculture 41,21-30.
Steffens, W., 1989. Principles of Fish Nutrition. Ellis Horwood Limited, England Stumn, W. and Morgan, J.J., 1996. Aquatic Chemistry: Chemical equilibria and rates in
natural. 3rd Ed, A Wiley-Interscience Publication. Swift, D.R., 1985. Aquaculture Training Manual. Fishing New Books Ltd., England Takeuchi, T., Arai, S., Watanabe, T. and Shimma, Y., 1980. Requirement of eel Anguilla
japonica for essential fatty acids. Bullentin of the Japanese Society of Scientific Fisheries 49, 345-353.
Webster, C.D. and Lim, C. (eds), 2002. Nutrient Requirement and Feeding of Finfish for Aquaculture. CAB international.

195
Wee, K.L., 1982. Snakeheads - Their Biology and Culture. In: Muir, J.F and Roberts, R.J (eds) Recent Advances in Aquaculture. Croom Helm, London, pp. 179 - 213.
Yamakawa, T., Arai, S., Watanabe, T. and Shimma, Y., 1975. Vitamin E requirement for Japanaese eel, Vitamin 49, 62.





























