Embed Size (px)
DESCRIPTION
第8章 核 酸. 本章主要内容. 8.1 核酸是遗传物质 8.2 核苷酸 8.3 脱氧核糖核酸( DNA) 8.4 核糖核酸( RNA) 8.5 核酸的某些物化性质和最常用的研究方法. 1928 Griffith 1944 Avery O.T. RII DNA SIII. 8.1 核酸是遗传物质. 核酸分脱氧核糖核酸( deoxyribonucleic acid, DNA ) 和核糖核酸( ribonucleic acid, RNA ) 两大类。. - PowerPoint PPT Presentation
Citation preview
第 8 章 核 酸

8.1 核酸是遗传物质
8.2 核苷酸
8.3 脱氧核糖核酸( DNA )
8.4 核糖核酸( RNA )
8.5 核酸的某些物化性质和最常用的研究方法
本章主要内容
核酸分脱氧核糖核酸( deoxyribonucleic acid, DNA )和核糖核酸 (ribonucleic acid, RNA) 两大类。
8.1 核酸是遗传物质
• DNA is a main genetic material
• RNA is genetic material also Tobacco Mosaic Virus (TMV)
Watson & Crich DNA double helix
RII DNA SIII
1928 Griffith 1944 Avery O.T
肺炎双球菌转化试验证明使肺炎双球菌的遗传性发生改变的转化因子是 DNA ,不是蛋白质
20 世纪自然科学领域的重大突破之一,揭开了分子生物学研究的序幕,奠定了分子遗传学研究的基础。
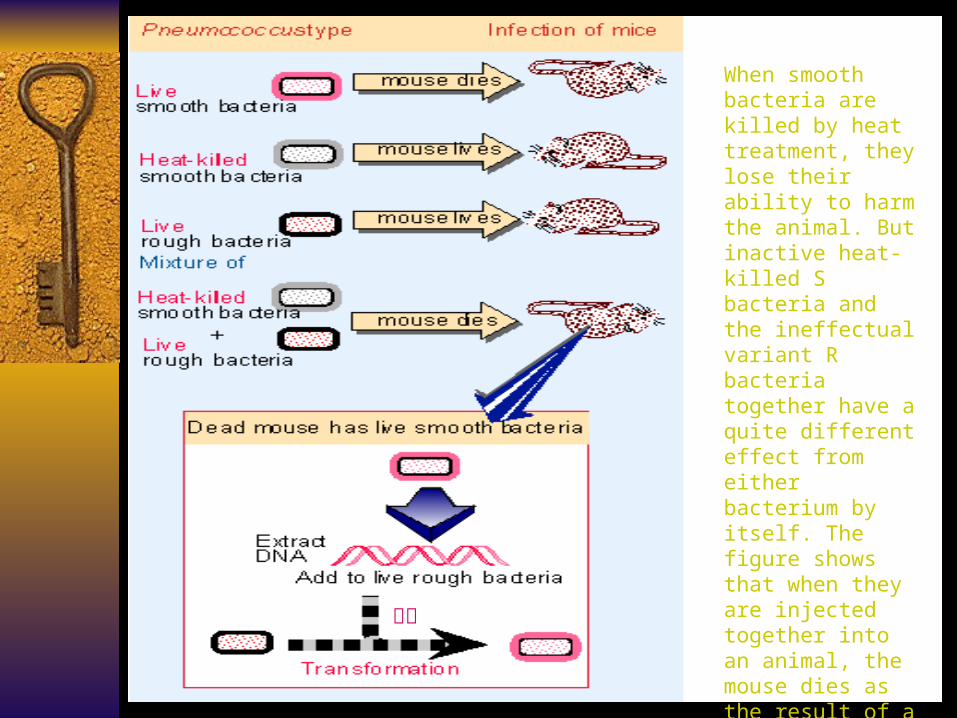
When smooth bacteria are killed by heat treatment, they lose their ability to harm the animal. But inactive heat-killed S bacteria and the ineffectual variant R bacteria together have a quite different effect from either bacterium by itself. The figure shows that when they are injected together into an animal, the mouse dies as the result of a Pneumococcal infection. Virulent S bacteria can be recovered from the
mouse postmortem. 转化

Francis Crick (35y)
1951
分子生物学的重要里程碑
James Watson (23y)
丹麦 哥本哈根
剑桥大学 Cavendish Lab.
1953 , DNA Double Helix model
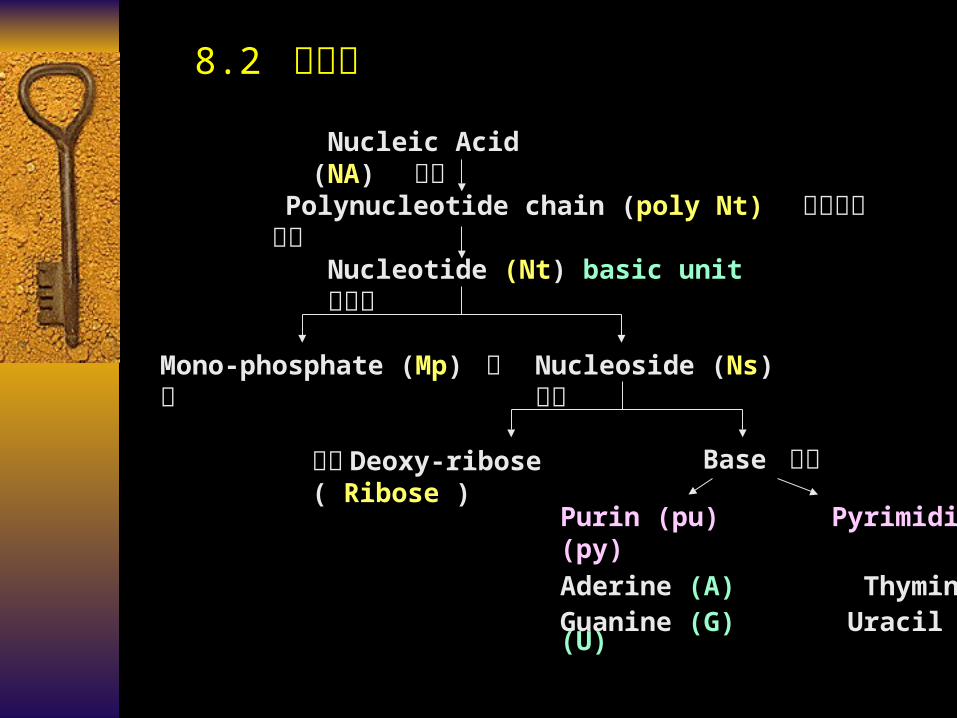
Nucleic Acid (NA) 核酸 Polynucleotide chain (poly Nt) 多聚核
苷酸链Nucleotide (Nt) basic unit 核苷酸
Mono-phosphate (Mp) 磷酸
Nucleoside (Ns) 核苷
戊糖 Deoxy-ribose ( Ribose )
Base 碱基
Purin (pu) Pyrimidine (py)Aderine (A) Thymine (T) Guanine (G) Uracil (U) Cytosine (C)
8.2 核苷酸

两类核酸的基本化学组成的区别
• 核糖 Ribose
DNA :脱氧核糖 Deoxy-ribose
RNA :核糖 Ribose
• 碱基 base
DNA : A 、 G 、 C 、 T
RNA : A 、 G 、 C 、 U
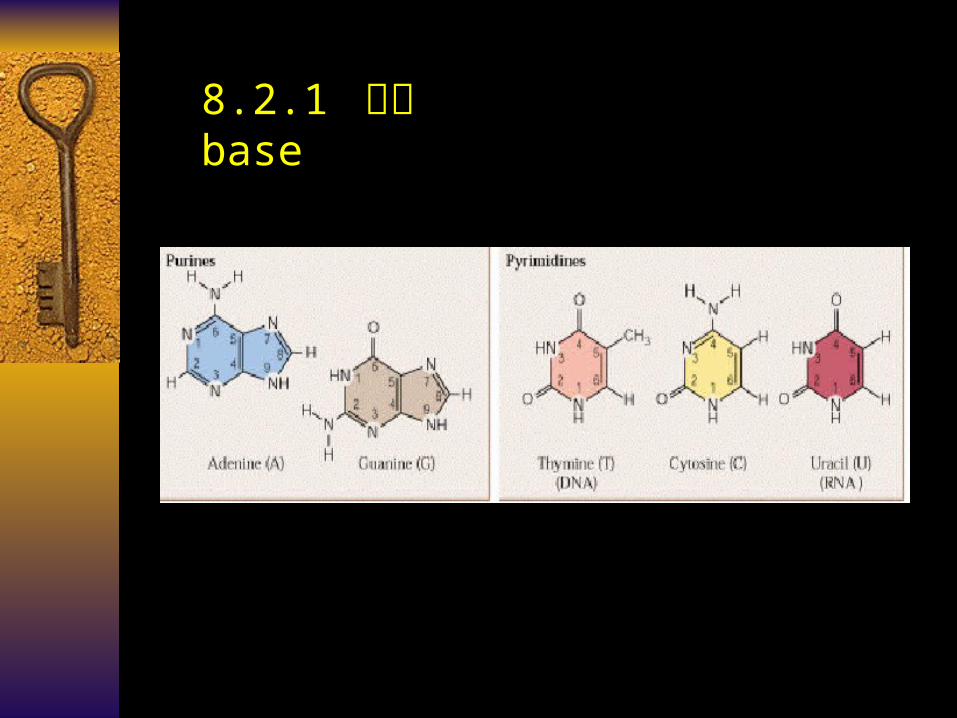
8.2.1 碱基 base
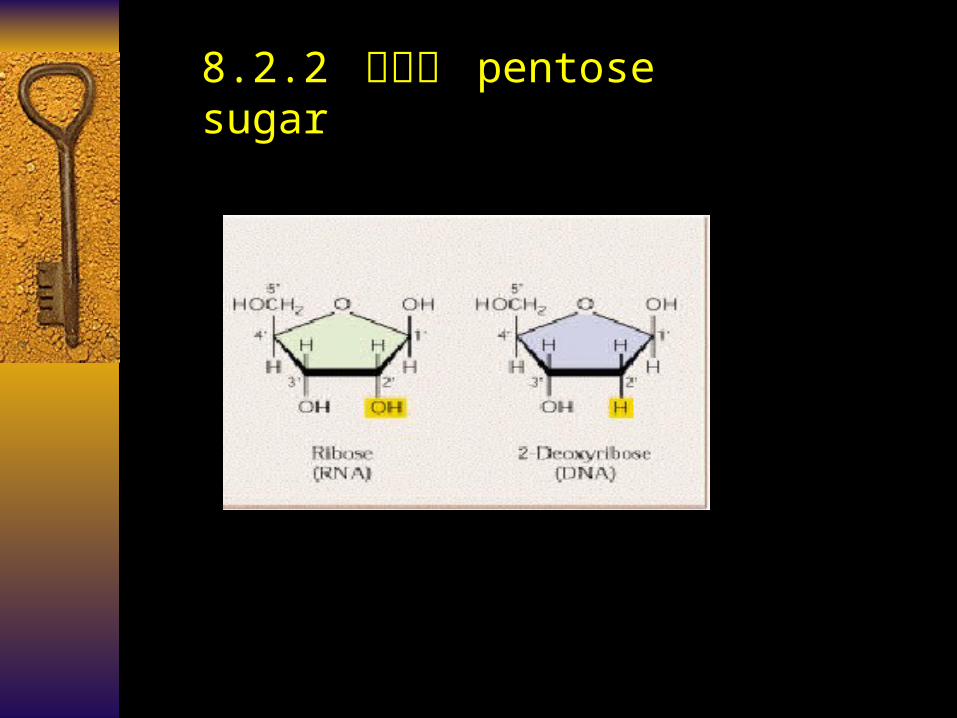
8.2.2 戊糖基 pentose sugar
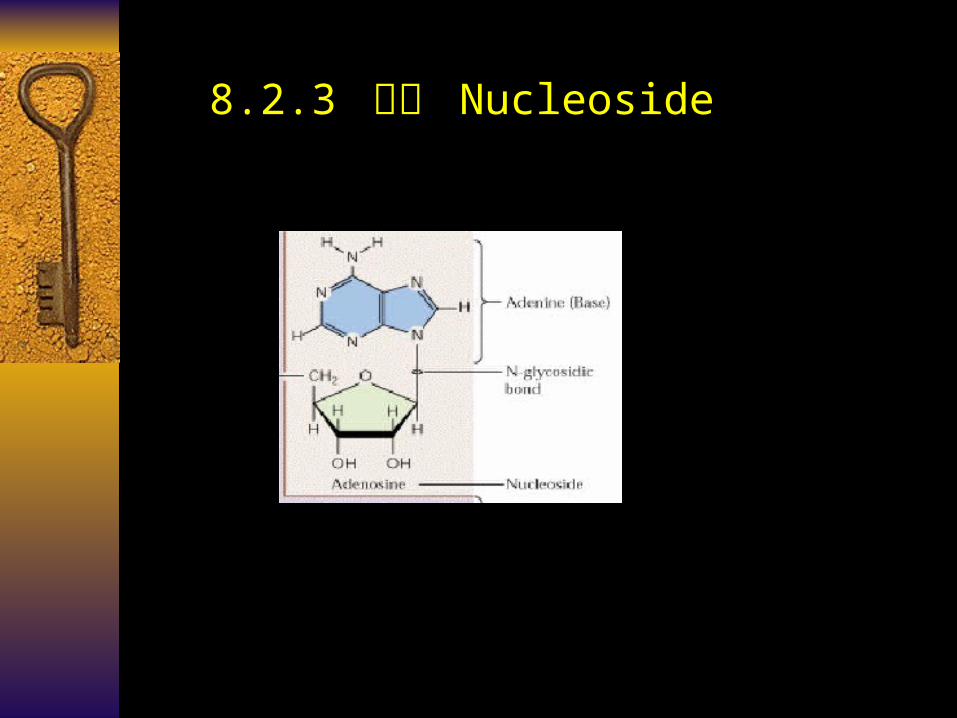
8.2.3 核苷 Nucleoside
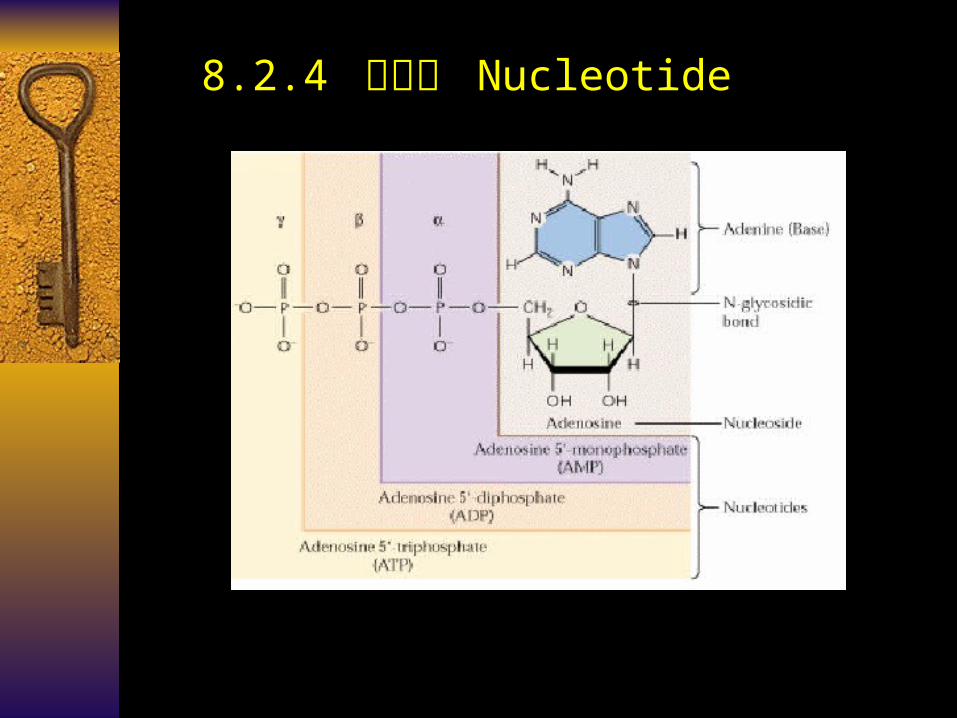
8.2.4 核苷酸 Nucleotide
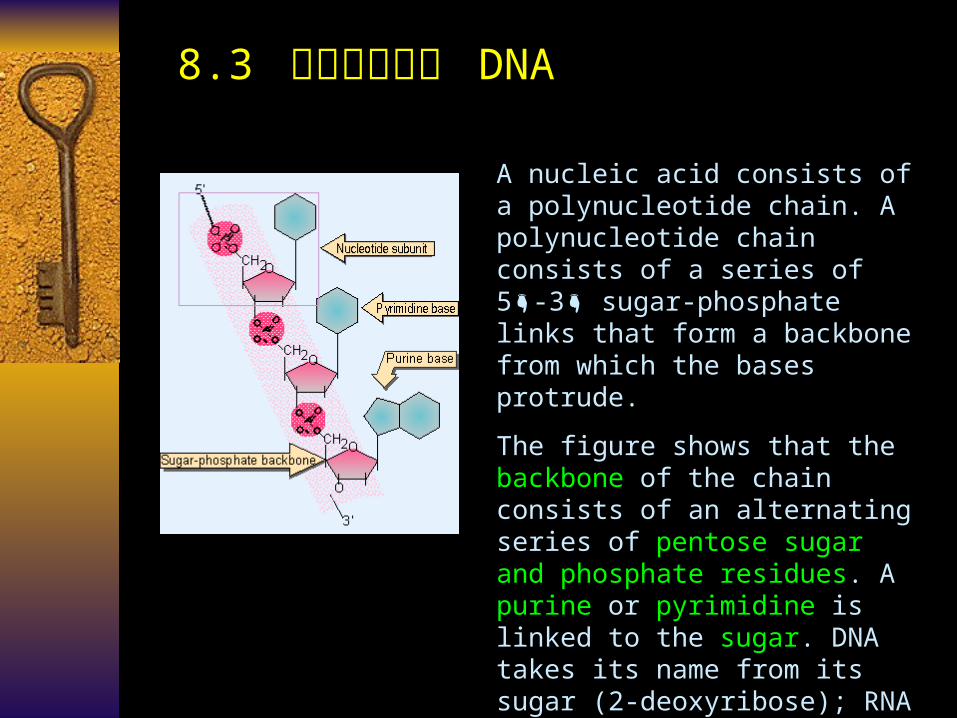
8.3 脱氧核糖核酸 DNA
A nucleic acid consists of a polynucleotide chain. A polynucleotide chain consists of a series of 5 -3 sugar-phosphate links that form a backbone from which the bases protrude.
The figure shows that the backbone of the chain consists of an alternating series of pentose sugar and phosphate residues. A purine or pyrimidine is linked to the sugar. DNA takes its name from its sugar (2-deoxyribose); RNA is named for its sugar (ribose). The difference is that the sugar in RNA has an OH group at the 2 position of the pentose ring.

8.3.1 DNA 的碱基组成
• 参与 DNA 组成的碱基主要有四种:A 、 T 、 G 、 C
• 所有 DNA 中, A=T , G=C 。因此,嘧啶总含量与嘌呤总含量相等,即 A+G=C+T 。此发现提示了 A 与 T , G 与 C 之间碱基互补的可能性,为 DNA 双螺旋结构的建立提供了重要依据。 DNA 的碱基组成具有种的特异性,但没有组织和器官的特异性
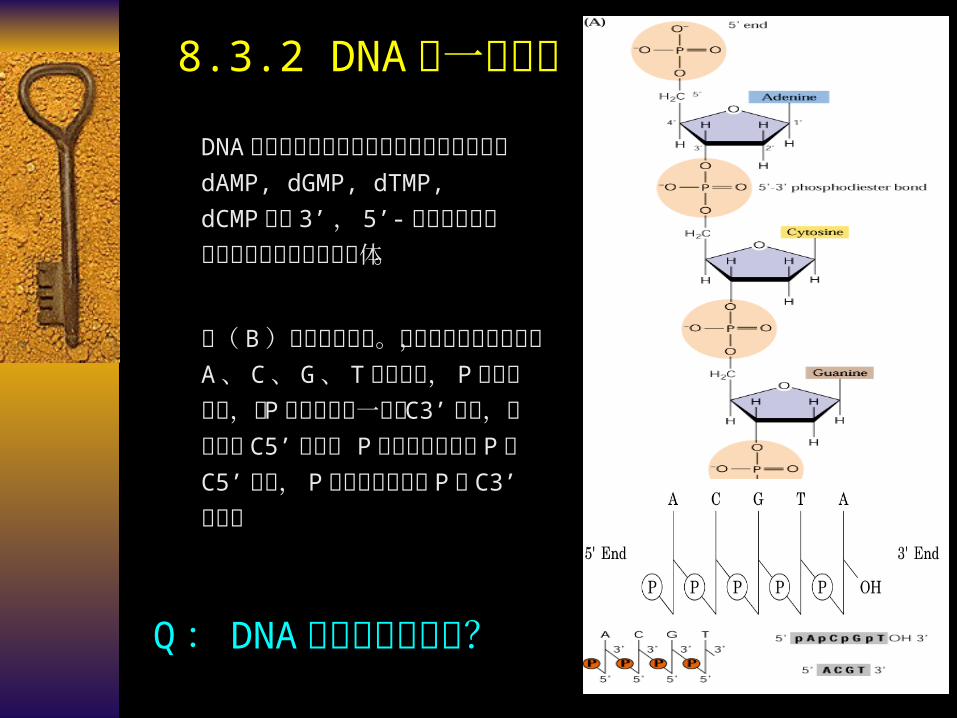
8.3.2 DNA 的一级结构
DNA 是由数量极其庞大的四种脱氧核糖核苷酸 dAMP,
dGMP, dTMP, dCMP 通过3’ , 5’- 磷酸二酯键连接成的直线形或环形多聚体。
Q : DNA 为什么没有侧链?
图( B )为线条式缩写。竖线表示核糖的碳链, A 、 C 、 G 、 T 表示碱基,P 代表磷酸基,由 P 引出的斜线一端与 C3’ 相连,另一端与C5’ 相连。 P 在碱基左侧表示 P与 C5’ 相连, P 在碱基右侧表示 P 与 C3’ 相连。
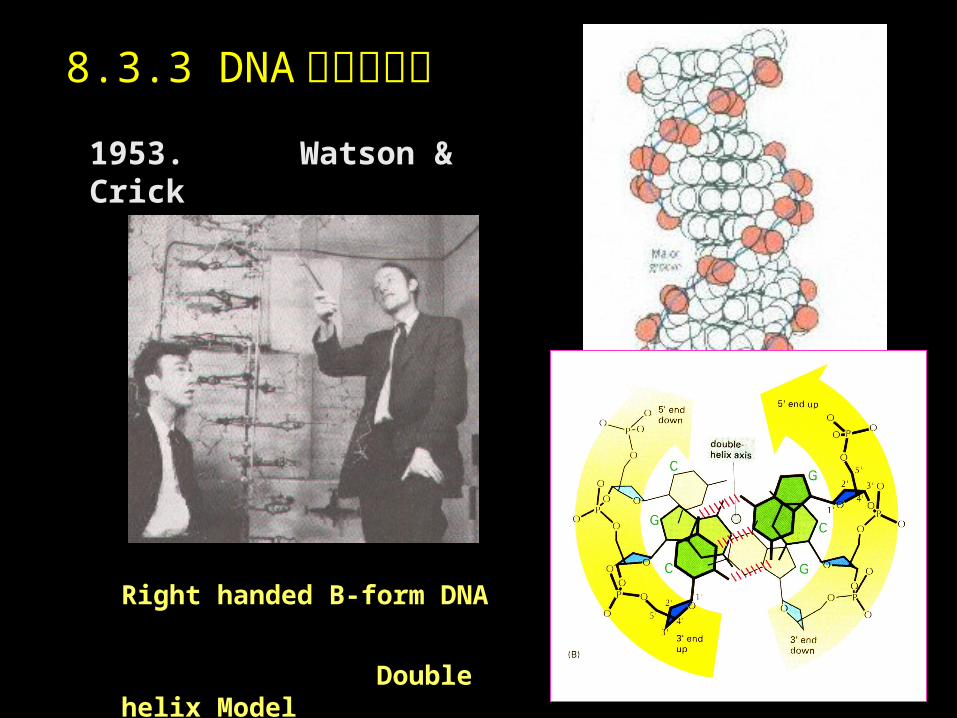
1953. Watson & Crick
Right handed B-form DNA
Double helix Model
8.3.3 DNA 的空间结构
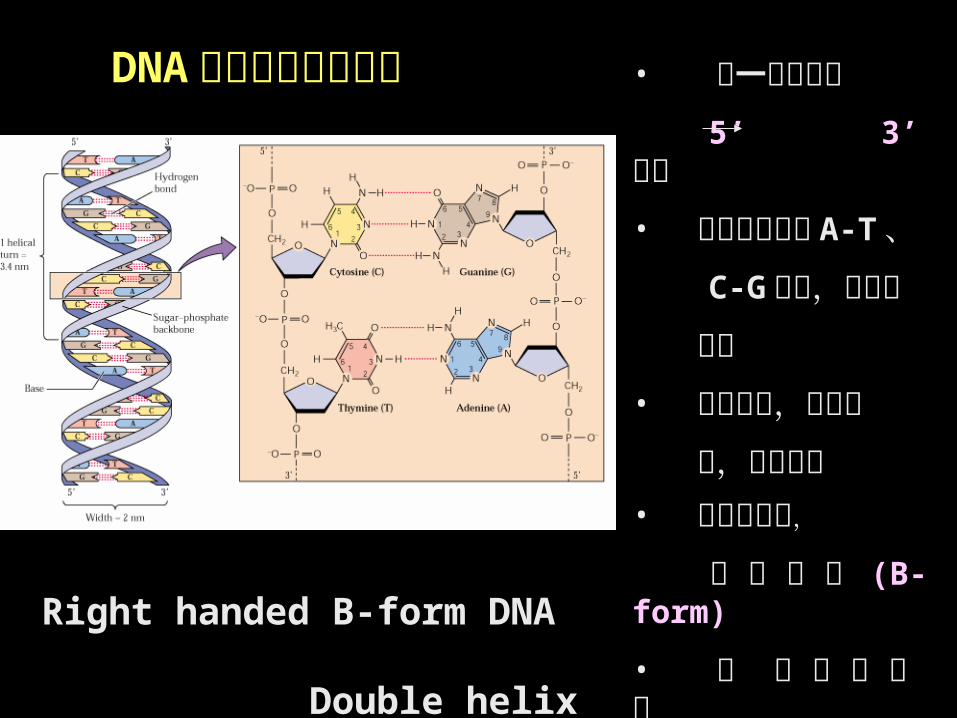
Right handed B-form DNA Double helix Model
• 每一单链具有
5’ 3’ 极性• 两条单链间按 A-T 、
C-G 配对,以氢键
连接• 两条单链,极性相
反,反向平行• 以中心为轴,
向 右 盘 旋 (B-form)
• 双 螺 旋 中 存 在
大沟、小 沟
DNA 双螺旋的结构特点
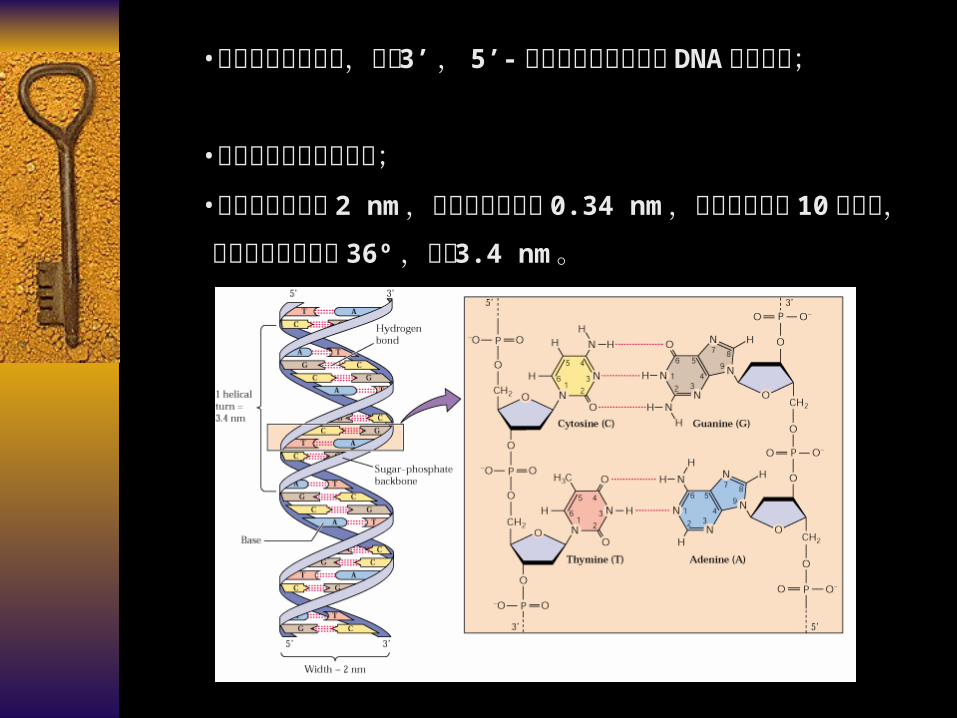
•碱基位于双螺旋的内侧;•双螺旋平均直径 2 nm ,相邻碱基对高度 0.34 nm ,每个螺旋包含 10 核苷酸,两核苷酸之间夹角 36º ,螺距 3.4 nm 。
•磷酸与核糖在外侧,通过 3’ , 5’- 磷酸二酯键相连形成DNA 分子骨架;
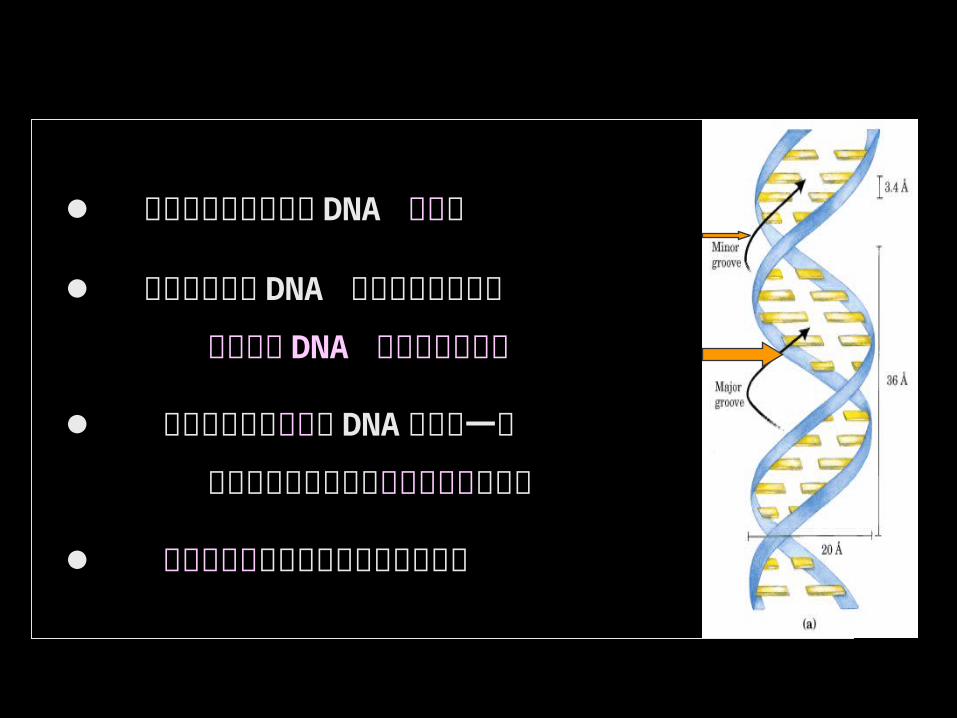
碱基顶部基团裸露在 DNA 大沟内
蛋白质因子与 DNA 的特异结合依赖于
氨基酸与 DNA 间的氢键的形成
蛋白质因子沿大沟与 DNA 形成专一性
结合的机率与多样性高于沿小沟的结合 大沟的空间更有利于与蛋白质的结合
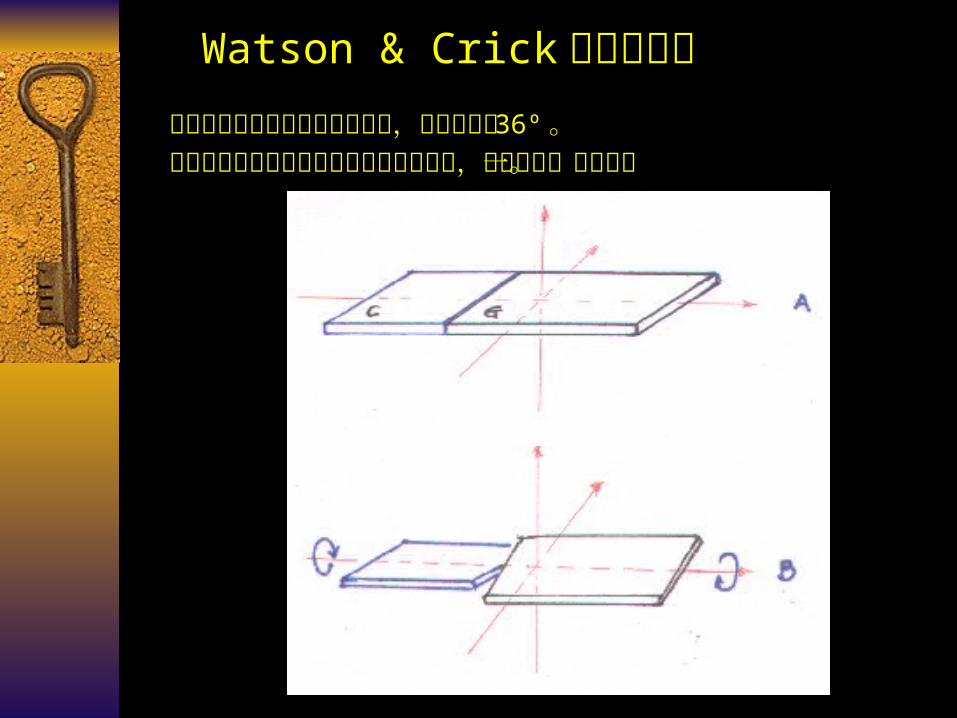
Watson & Crick 模型的修正两个核苷酸之间的夹角是可变的,并非固定的 36º 。
组成碱基对的碱基之间发生螺旋桨状扭曲,而非在同一平面上。

B-DNA的重要特性
1. 天然 DNA分子的形态十分细长。2. DNA分子中的碱基存在互变异构现象。碱基偶尔发生互变异构可引起 DNA突变,推动生物进化,但通常情况下,氢原子在碱基上有较固定的位置,保持遗传信息的稳定。3. DNA分子的双螺旋结构在生理状态下十分稳定。4. DNA分子在溶液中具有较大的可塑性。
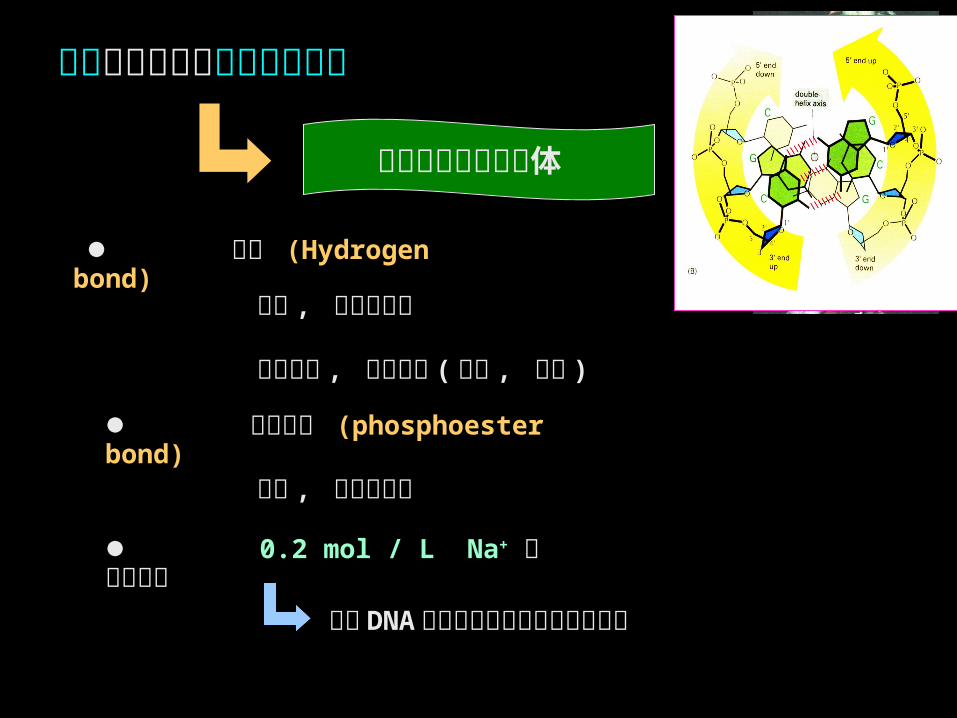
影响双螺旋结构稳定性的因素
碱基堆积的棒状实体
氢键 (Hydrogen bond)
消除 DNA 单链上磷酸基团间的静电斥力
弱键 , 可加热解链
氢键堆积 , 有序排列 ( 线性 , 方向 )
磷酸酯键 (phosphoester bond)
0.2 mol / L Na+ 生理盐条件
强键 , 需酶促解链

碱基堆积力 ( 非特异性结合力 )
3.4A°
☆ Van de waals force (1.7A°/ 嘌呤环与嘧啶环作用半径 )
(0.34 nm/ 碱基对间距 )
☆ 疏水相互作用 (Hydrophobic interaction) 不溶于水的非极性分子在水中相互联合 , 成串结合的趋势力 即为熵 Entrophy
成为碱基间的部分堆积力
DNA 分子中非极性的嘌呤、嘧啶环,使有序的以氢键连接的水分子层绕,熵值降低。
DNA 分子中非极性碱基的聚集,使有序的水分子层绕减低到最低的限度,减少熵值的降低。
DNA 分子中的亲水的磷酸骨架,氨基,酮基与水分子形成氢键。熵值增加
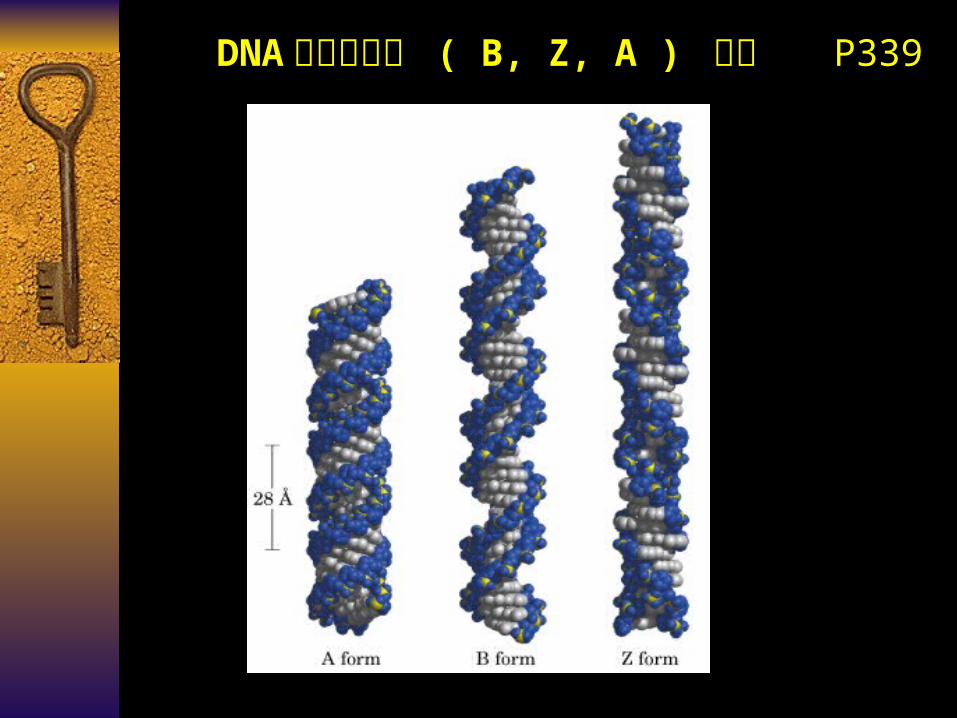
DNA 的分子构型 ( B, Z, A ) 比较 P339
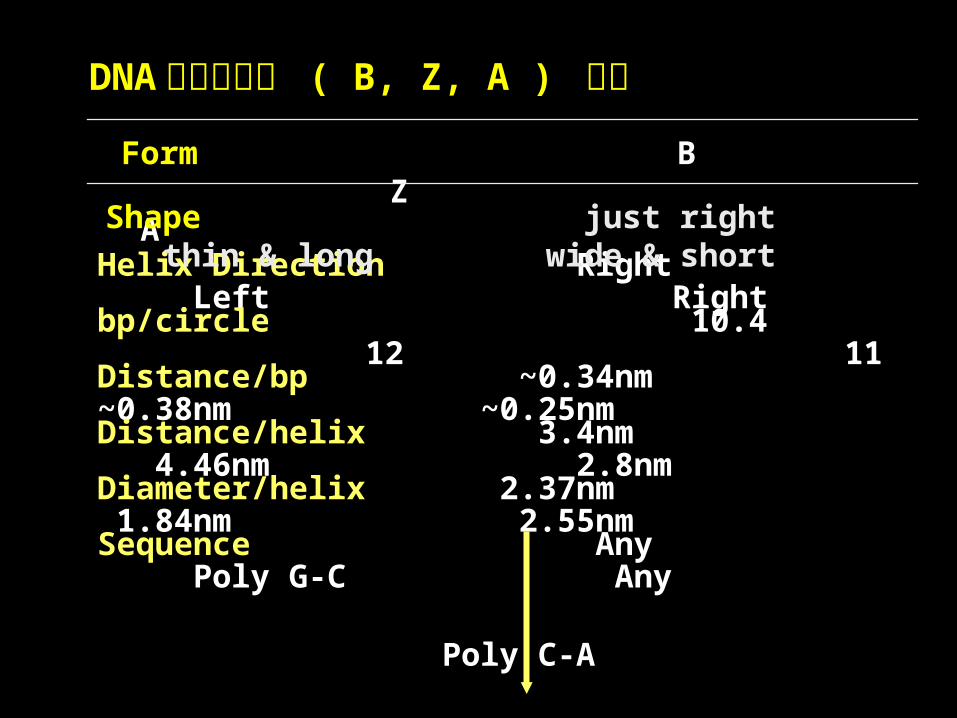
DNA 的分子构型 ( B, Z, A ) 比较
Form B Z A
Helix Direction Right Left Right
bp/circle 10.4 12 11
Distance/bp ~0.34nm ~0.38nm ~0.25nm
Distance/helix 3.4nm 4.46nm 2.8nm
Diameter/helix 2.37nm 1.84nm 2.55nm
Sequence Any Poly G-C Any
Poly C-A
Poly T-G
Poly T-A
Shape just right thin & long wide & short

Condition Normal 4M Na+ DNA-Na Crytic
D.S RNA DNA/RNA
Major Groove wide & deep flat narrow & deepMinor Groove narrow & deep narrow & deep wide & shallow
Conformation A,G-anti A, G-syn A, G-anti
( dGMP, dAMP ) C, T-anti T, C-anti T, C-anti
Form B Z A
8.3.4 环形 DNA P340
• 连环数( L , linking number ):在双螺旋DNA 中,一条链以右手螺旋绕另一条链缠绕的次数。
• 缠绕数( T , twisting number ): DNA 分子中的 Watson-Crick 螺旋数目。
• 超螺旋周数或扭曲数( W , writhing
number )。
• 比连环差( λ , specific linking
difference ) : DNA 的超螺旋程度, λ= ( L-
L0 ) /L0
L=T+W
• 拓扑异构酶的作用改变 DNA 的拓扑结构。
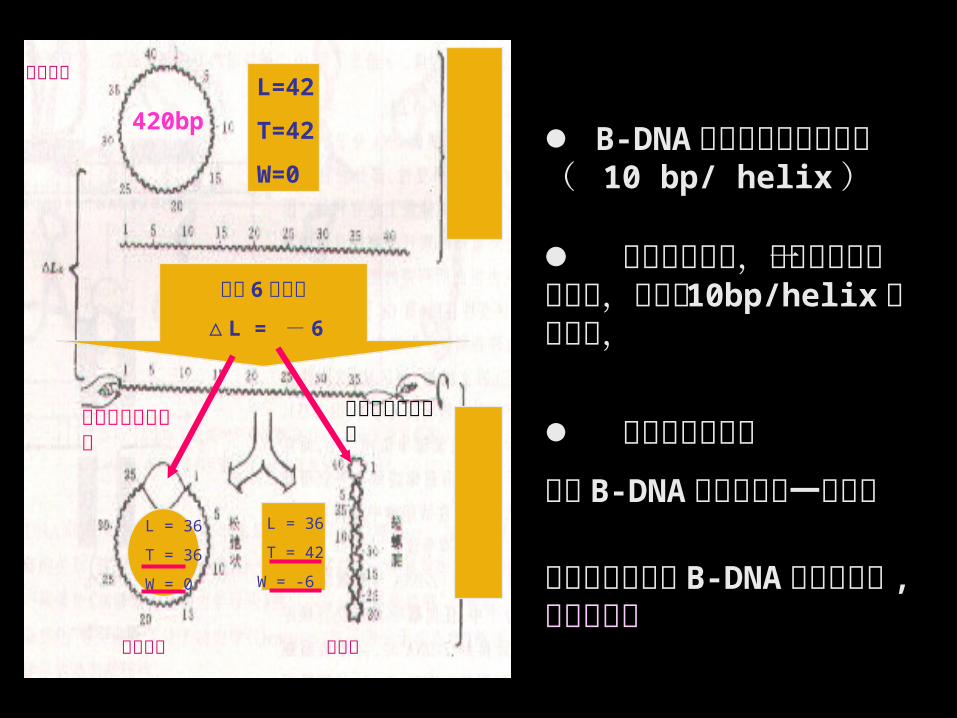
B-DNA 是力学上稳定的结构( 10 bp/ helix ) 虽交叉数减少,但需转换为一种应力,以维持10bp/helix 的螺旋数,
应力的重新分配
或在 B-DNA 状态中保留一单链区或螺旋力将维持 B-DNA 的右旋结构 , 形成超螺旋
420bp
L=42
T=42
W=0
无应力松弛状
态
应力的分配
L = 36
T = 36
W = 0
L = 36
T = 42
W = -6
链松弛后再结成环
链未松弛再结成环
松开 6 圈螺旋
△ L = - 6
松弛环形
解链环形 超螺旋

8.3.5 DNA 的生物学功能 P342
DNA 是遗传物质
• DNA 是染色体的主要成分,染色体与遗传直接相关• 细胞核内 DNA 含量与染色体数目的多少有平行关系
• DNA 在代谢上稳定
• 作用于 DNA 的物化因素均可引起遗传特性的改变• 肺炎双球菌转化实验直接证明 DNA 是遗传物质
8.4 核糖核酸 RNA P343
The central dogma states that information in nucleic acid can be perpetuated or transferred, but the transfer of information into protein is irreversible.
Natural RNA is mainly synthesized from DNA, but some RNA-virus can synthesize RNA from RNA.
the central dogma
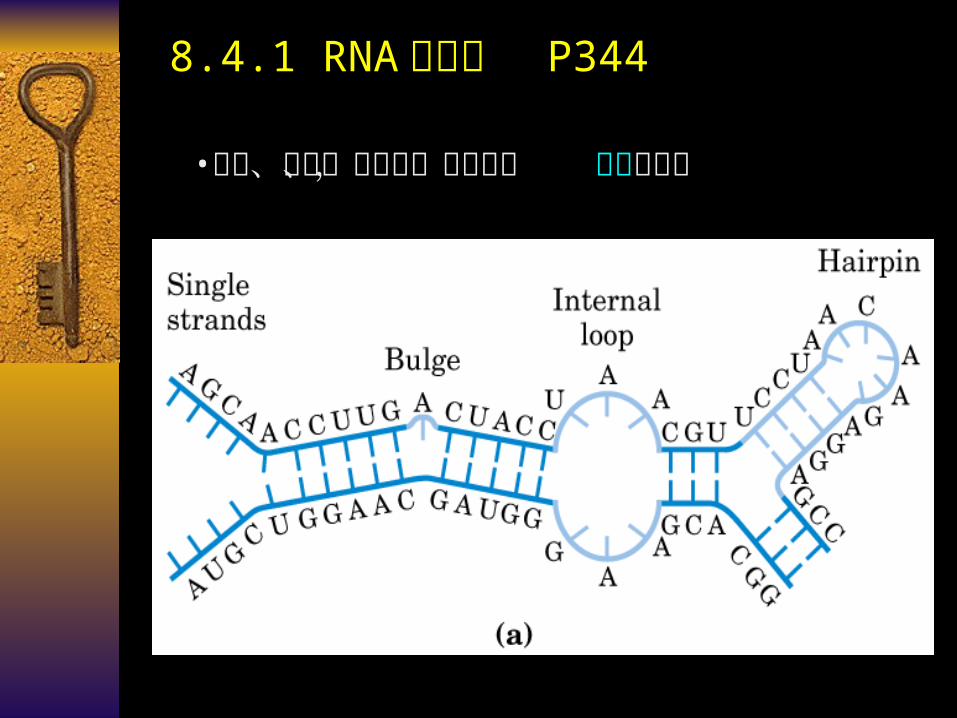
•单链、线形、无分支,局部形成 链内双螺旋
8.4.1 RNA 的结构 P344
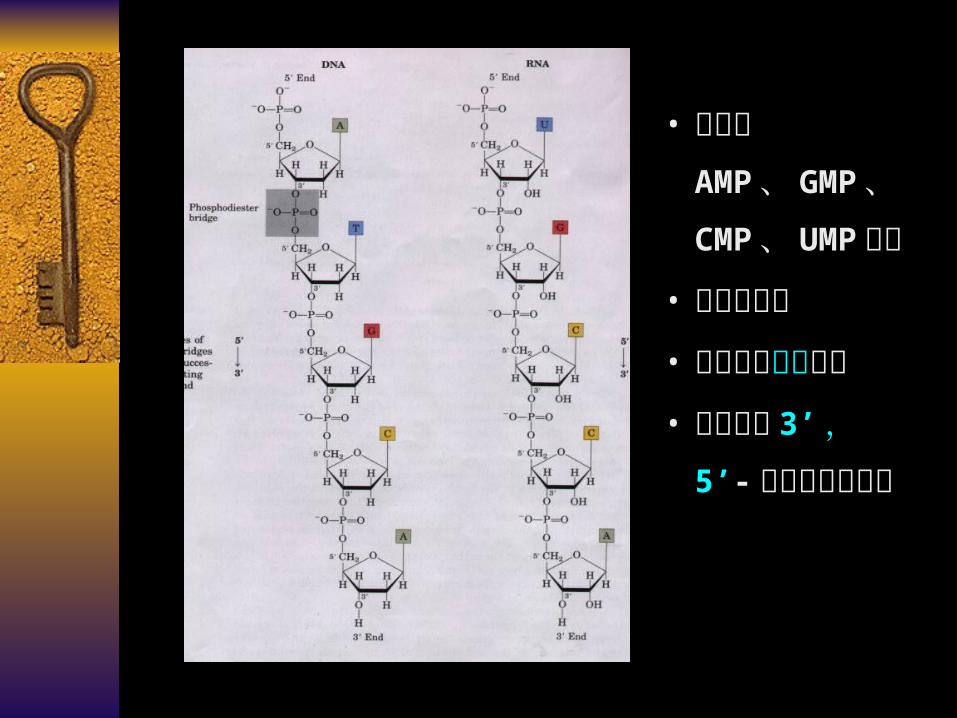
• 主要由AMP 、 GMP
、 CMP 、 U
MP 组成
• 戊糖为核糖
• 含有某些稀有碱基
• 核苷酸以 3’ ,5’- 磷酸二酯键连接

8.4.2 RNA 的类型
• messenger RNA
(mRNA)
• transfer RNA (tRNA)
• ribosomal RNA (rRNA)
生物细胞内含有三种主要的 RNA :
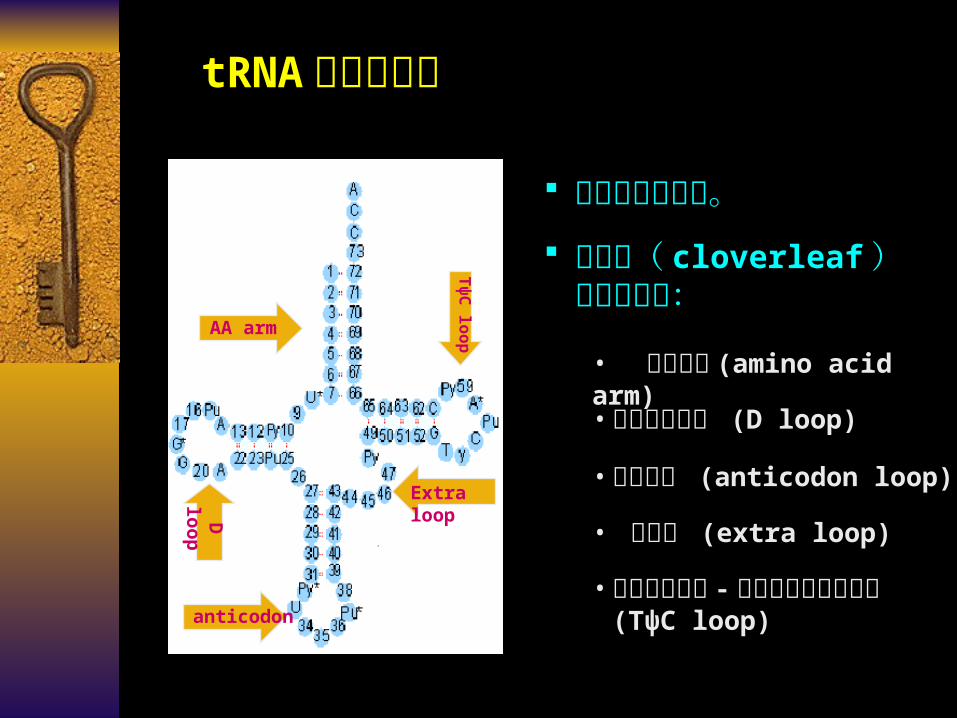
AA arm
Tψ
C loop
D loop
anticodon
Extra loop
有较多稀有碱基。
tRNA 的二级结构
• 氨基酸臂 (amino acid arm)• 二氢尿嘧啶环 (D loop)
• 反密码环 (anticodon loop)
• 额外环 (extra loop)
• 假尿嘧啶核苷 - 胸腺嘧啶核糖核苷环 (TψC loop)
三叶草( cloverleaf )形二级结构:
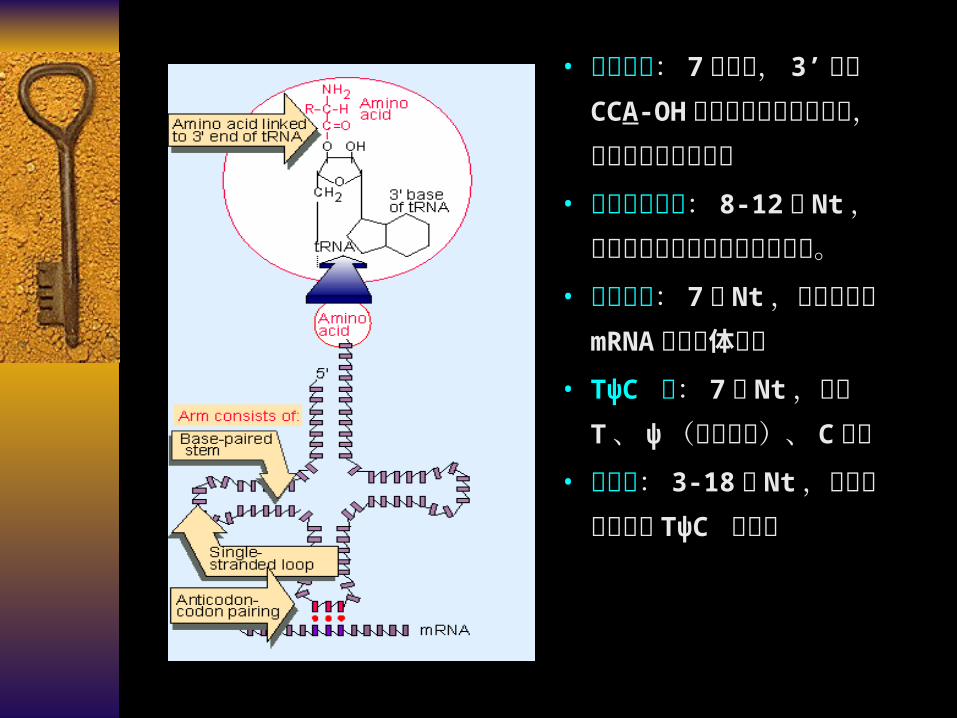
• 氨基酸臂: 7 对碱基, 3’
末端 CCA-OH 与氨基酸通过酯键相连,起转运氨基酸的作用
• 二氢尿嘧啶环: 8-12 个
Nt ,因含稀有碱基二氢尿嘧啶而得名。
• 反密码环: 7 个 Nt ,读取和识别 mRNA 的三联体密码
• TψC 环: 7 个 Nt ,因含T 、 ψ (假尿嘧啶)、 C
得名• 额外环: 3-18 个 Nt ,位
于反密码环和 TψC 环之间
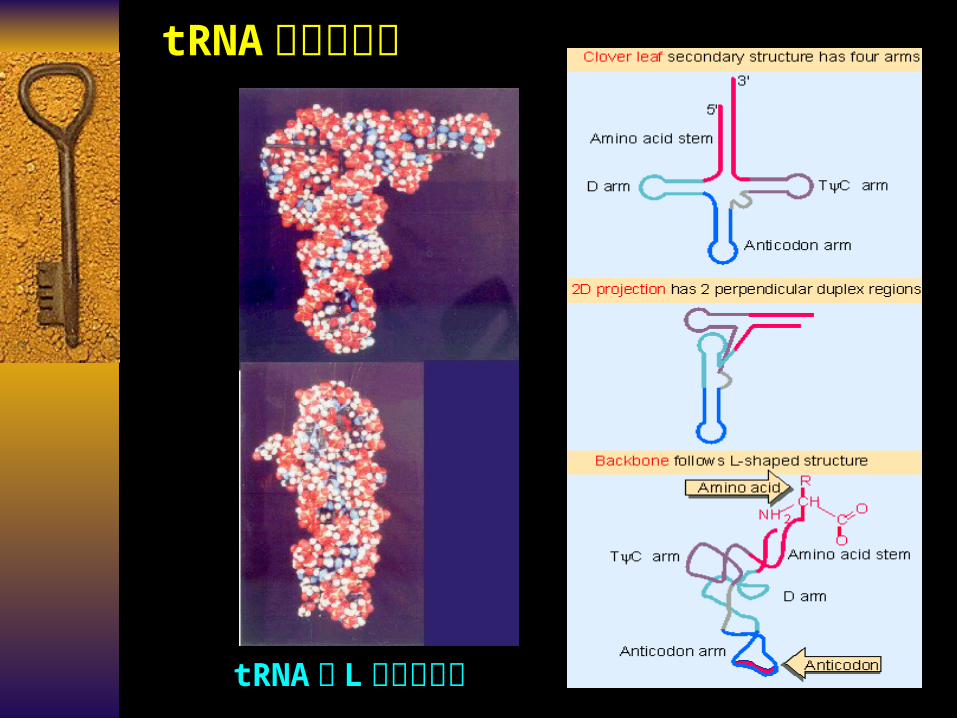
tRNA 的三级结构
tRNA 的 L 形三级结构
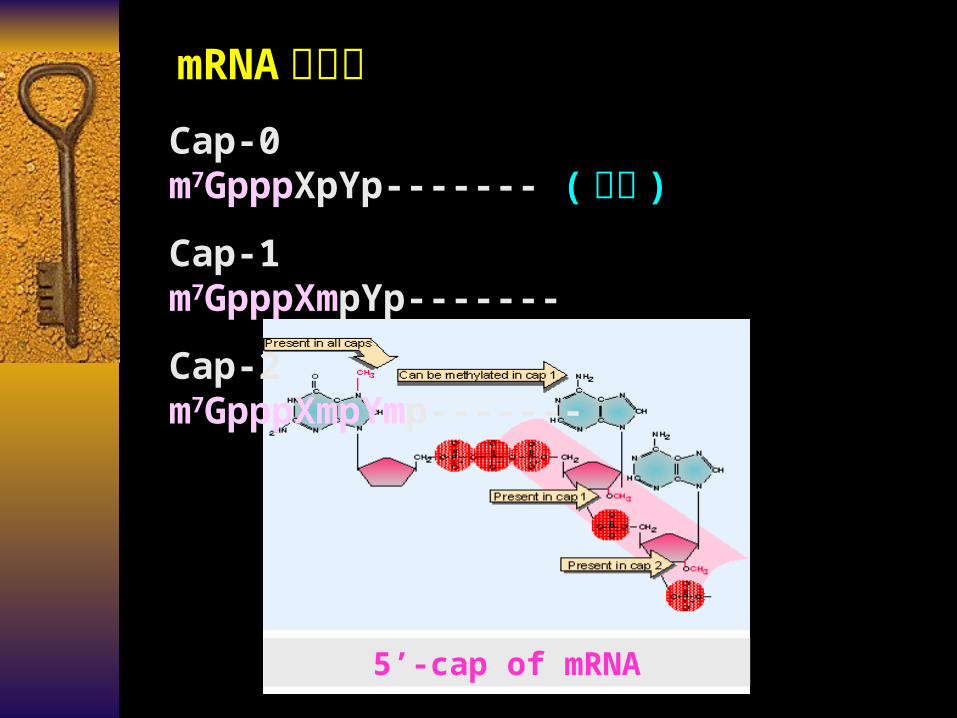
mRNA 的结构Cap-0 m7GpppXpYp------- ( 共有 )
Cap-1 m7GpppXmpYp-------
Cap-2 m7GpppXmpYmp-------
5’-cap of mRNA

mRNA 的结构
Man α- globin mRNA 5’-------CUUUGAAUAAA------------poly(A) 3’ -20Man β- globin mRNA 5’-------UGCCUAAUAAA------------poly(A) 3’ -20
3’-tail of mRNA
AAUAAA 为加尾信号序列
A poly(A) tail (50-200±) be added at -20 Nt ± tailing
signal (AAUAAA) from 3’-end of Pre-RNA

rRNA 的特点
• 含量最大的一类 RNA ,构成核糖体的骨架
• 原核生物核糖体中三类 rRNA : 5S
rRNA , 16S rRNA , 23S rRNA
• 真核生物核糖体中四类 rRNA : 5S
rRNA , 5.8S rRNA , 18S rRNA , 28S
rRNA

8.5 核酸的某些物化性质和最常用的研究方法
5.5.1 核酸的紫外吸收
5.5.2 核酸的沉降特性
5.5.3 凝胶电泳
5.5.4 核酸的变性、复性及杂交
5.5.5 DNA 的固相合成
5.5.6 DNA 的限制酶图谱
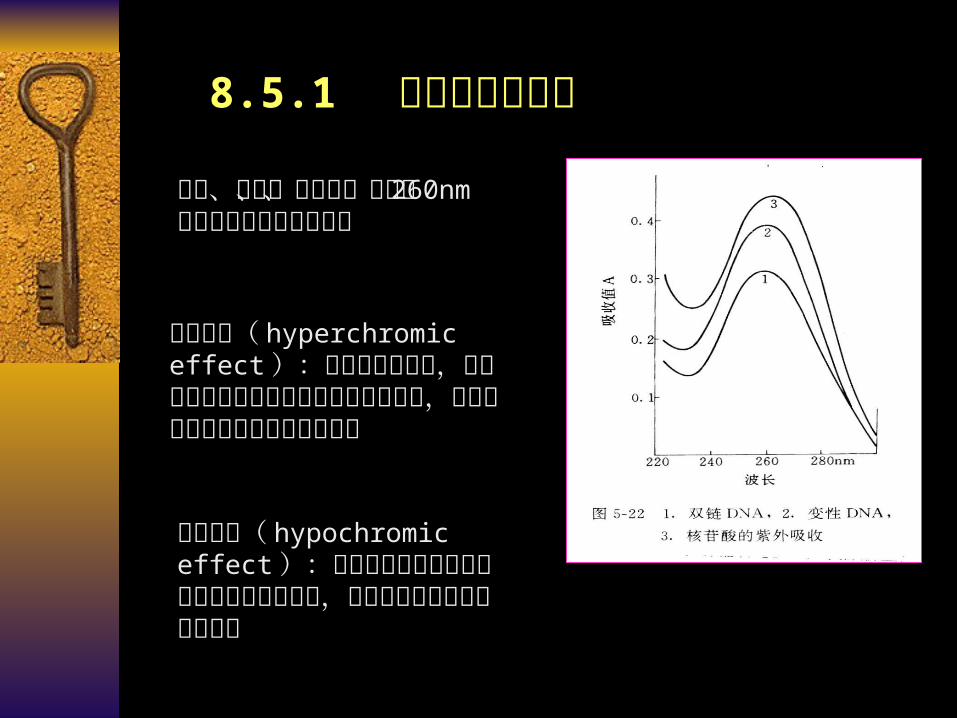
8.5.1 核酸的紫外吸收
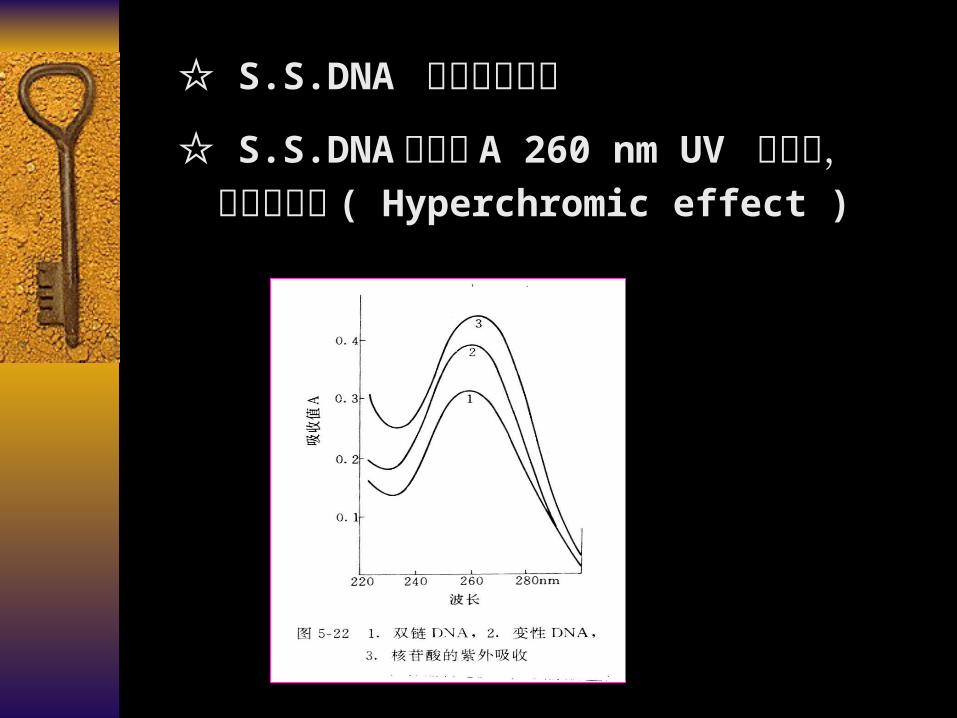
碱基、核苷、核苷酸、核酸在 260nm 紫外波长处有最大光吸收
增色效应( hyperchromic effect ):核酸发生变性时,超螺旋结构多核苷酸转变成单链多核苷酸,对紫外光的吸收值随之增加的现象
减色效应( hypochromic effect ):单链多核苷酸复性成超螺旋结构多核苷酸时,对紫外光的吸收值降低的现象
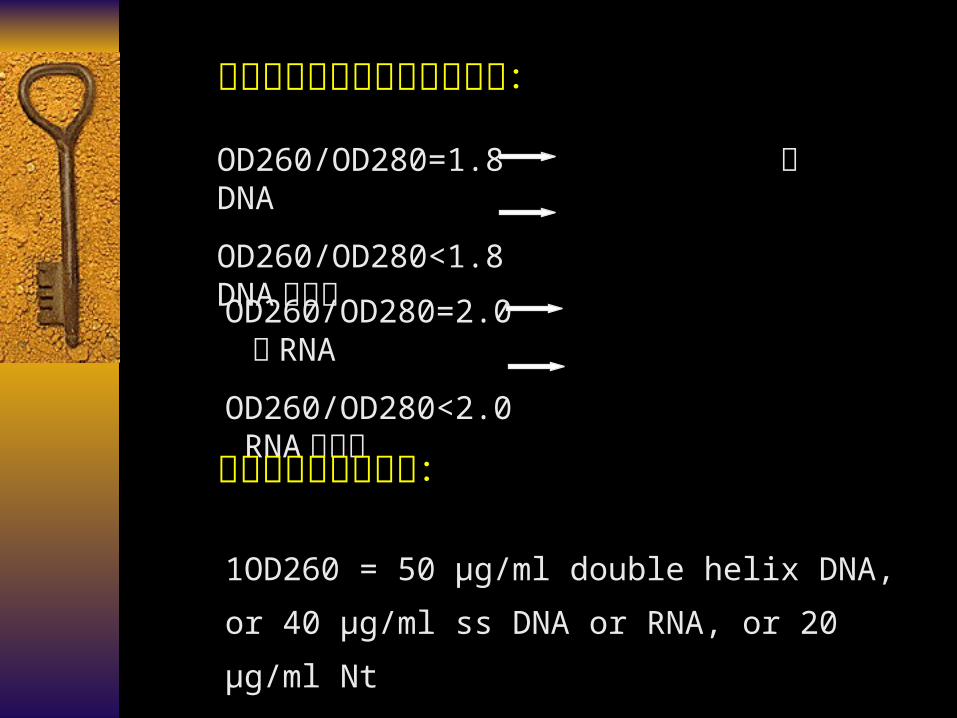
判断核酸样品是否纯品的方法:
OD260/OD280=1.8 纯 DNA
OD260/OD280<1.8 DNA 含杂质
OD260/OD280=2.0 纯 RNA
OD260/OD280<2.0 RNA 含杂质
核酸纯品的含量测定:
1OD260 = 50 µg/ml double helix DNA, or 40 µg/ml
ss DNA or RNA, or 20 µg/ml Nt

8.5.2 核酸的沉降特性不同构象的核酸、蛋白质及其它杂质在引力场中的沉降速率有很大差异
•蔗糖梯度离心法分离 RNA
•氯化铯梯度离心分离 DNA
•染料 -氯化铯密度梯度超离心
蛋白质
开环形及线形 DNA
闭环超螺旋 DNA
RNA
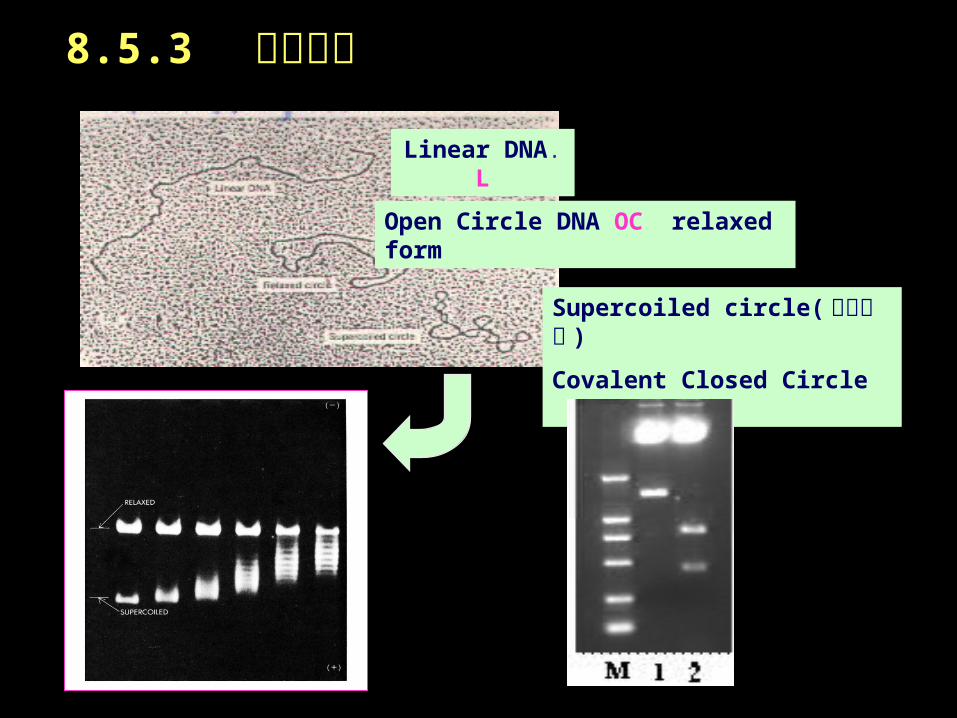
Linear DNA. L
Open Circle DNA OC relaxed form
Supercoiled circle( 高级结构 )
Covalent Closed Circle CCC
polymerpolymer
OCOC
LLCCCCCC
8.5.3 凝胶电泳
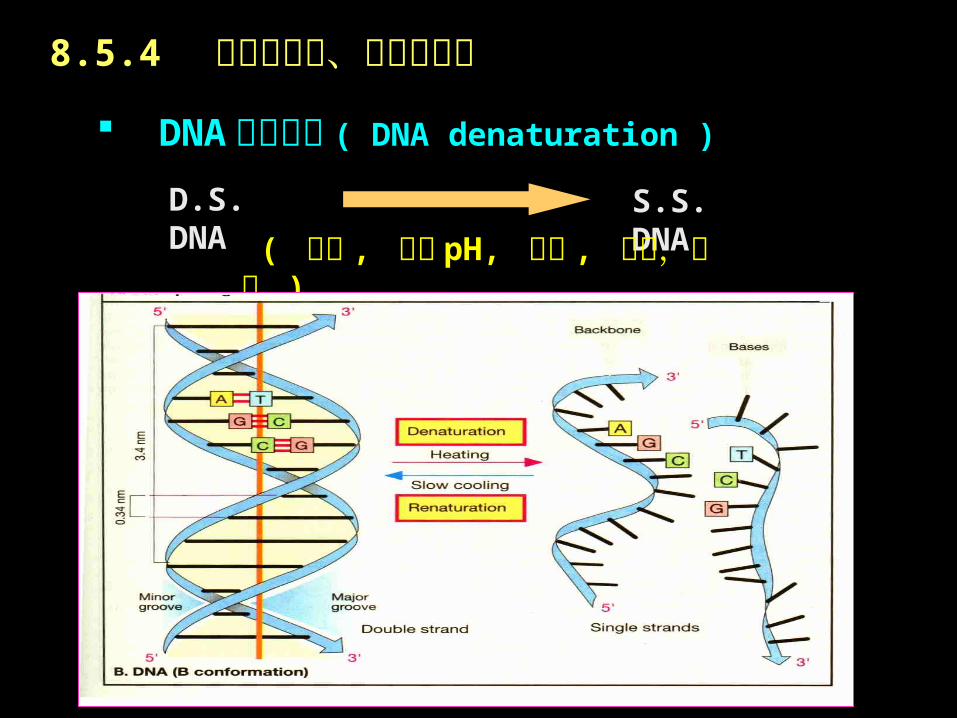
DNA 分子变性 ( DNA denaturation )
D.S. DNA S.S. DNA
( 加温 , 极端 pH, 尿素 , 甲醛,酰胺 )
8.5.4 核酸的变性、复性及杂交
变性过程的表现 ☆ S.S. DNA 粘度降低
溶液粘度取决于分子流动过程中的内摩擦和阻力
高分子溶液 > 普通溶液
线状分子 > 不规则线团 > 球形分子
D.S. DNA 钢性较强,结构较为舒展的 Double helix
S.S. DNA 没有氢键的支撑
由螺旋结构向折叠和线团结构转变
D.S DNA S.S DNA 粘度降低
☆ S.S.DNA 沉降速度加快
☆ S.S.DNA 分子的 A 260 nm UV 值上升,即增色效应 ( Hyperchromic effect )
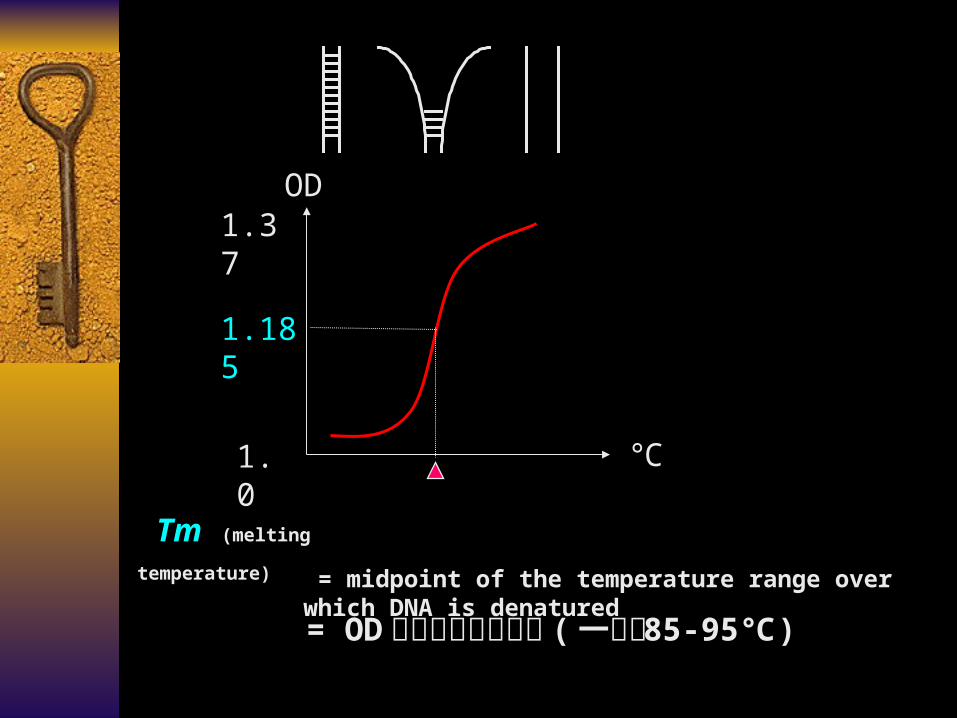
1.185
1.0
1.37OD
℃
= OD 增加值的中点温度 ( 一般为 85-95)℃
Tm (melting temperature) = midpoint of the temperature range over which DNA is denatured
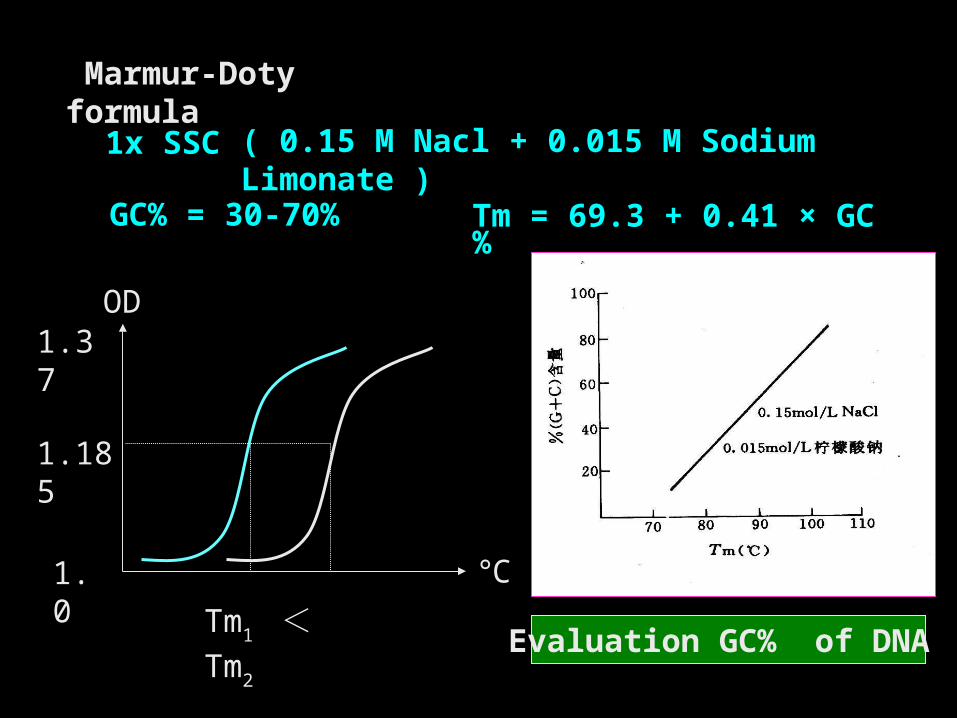
Marmur-Doty formula
Evaluation GC% of DNA
Tm = 69.3 + 0.41 × GC%
1x SSC
GC% = 30-70%
( 0.15 M Nacl + 0.015 M Sodium Limonate )
Tm1 < Tm2
℃1.0
1.185
1.37OD
what is meaning ?
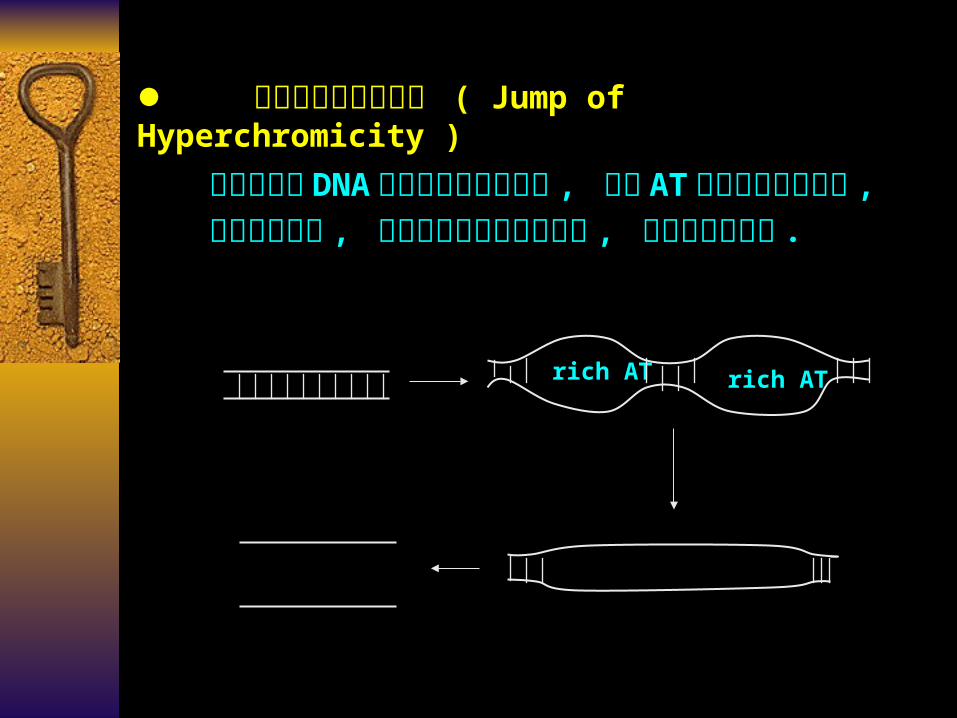
增色效应的跳跃现象 ( Jump of Hyperchromicity )
高分子量的 DNA分子在热变性过程中 , 富含 AT区域首先发生变性 , 然后逐步扩展 , 表现增色效应的跳跃现象 , 使变性过程加快 .
rich AT rich AT

● 影响 Tm 值的因素
☆ 在 A, T, C, G 随机分布的情况下
☆ GC% 含量相同的情况下
GC%愈高 → Tm 值愈大 GC%愈低 → Tm 值愈小
AT 形成变性核心,变性加快, Tm 值小
例:常温下,活体内 D.S DNA 分子中富含 AT 的变性
核心区( promoter, terminator region )
常表现氢键的断裂与形成的 “ DNA呼吸现象”
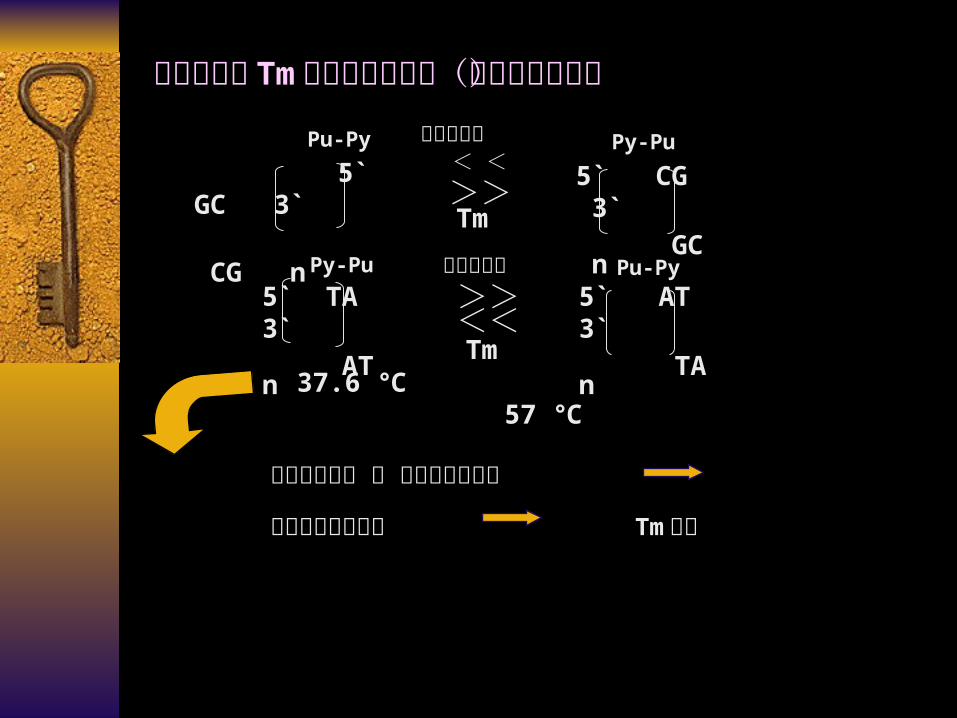
5` CG 3`
GC n
5` GC 3`
CG n
5` TA 3`
AT n
5` AT 3`
TA n
37.6 57 ℃ ℃
Tm>>
<<
< <
>>Tm
碱基堆积面大 和 碱基积压程度高
双链间的稳定性差 Tm变低
Pu-Py Py-Pu碱基堆积面
Pu-PyPy-Pu 碱基堆积面
碱基排列对 Tm 值具有明显影响(除变性核心外)
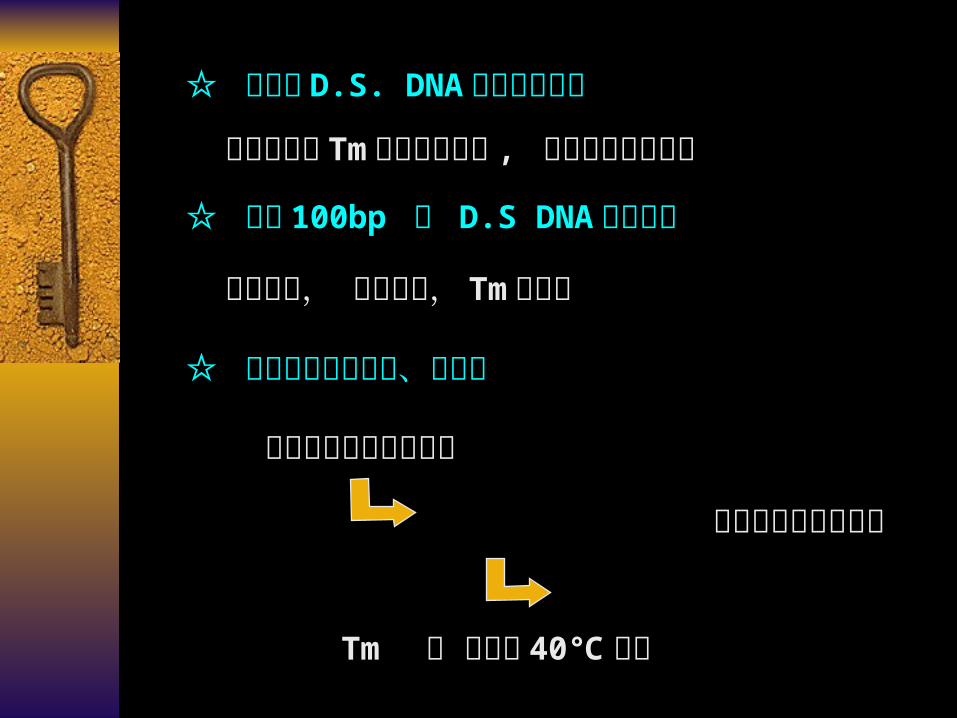
☆ 大片段 D.S. DNA 分子之间比较
片段长短对 Tm 值的影响较小 , 与组成和排列相关
☆ 小于 100bp 的 D.S DNA 分子比较片段愈短, 变性愈快, Tm 值愈小
☆ 变性液中含有尿素、酰胺等
尿素与碱基间形成氢键
改变碱基对间的氢键
Tm 值 可降至 40℃左右
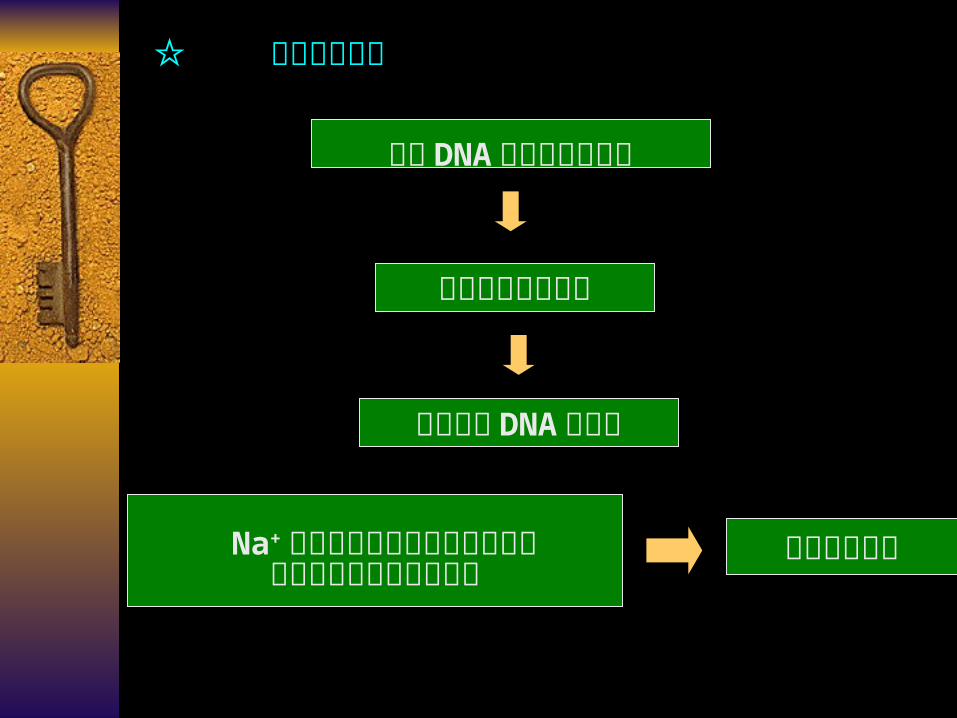
☆ 盐浓度的影响
单链 DNA 主链的磷酸基团
负电荷的静电斥力
两条单链 DNA 的分离
Na+ 在磷酸基团周围形成的电子云对静电斥力产生屏蔽作用
减弱静电斥力
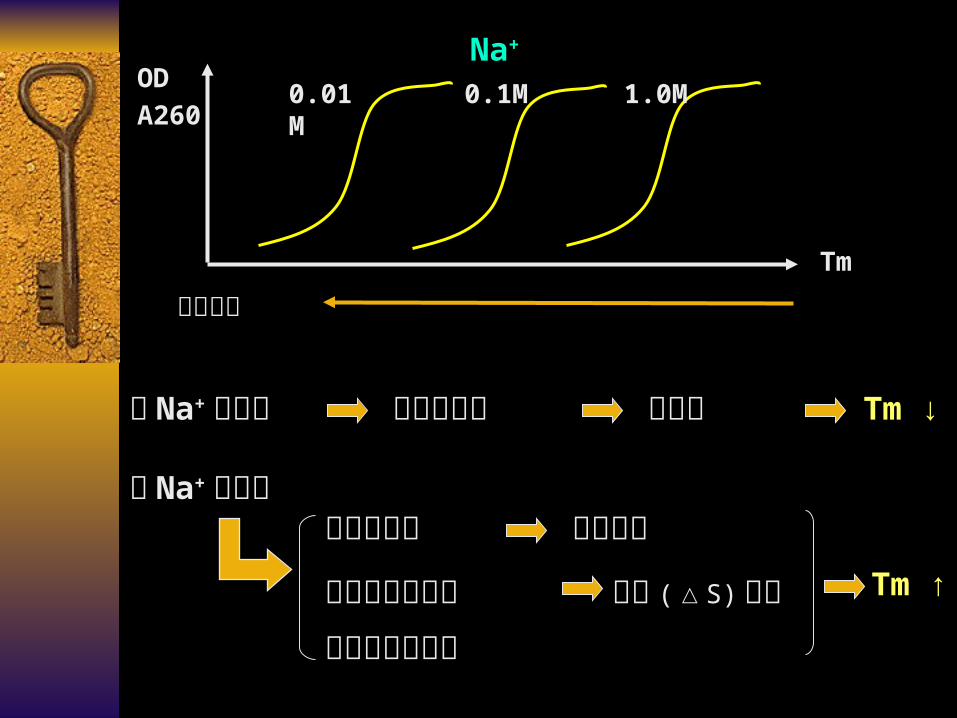
Tm ↑
当 Na+浓度低
屏蔽作用小 斥力强 Tm ↓
静电斥力 Tm
OD
A260 0.01M 0.1M 1.0M
Na+
当 Na+浓度高 屏蔽作用大 斥力减弱
熵值 (△S) 上升
碱基溶解性降低疏水作用力增加
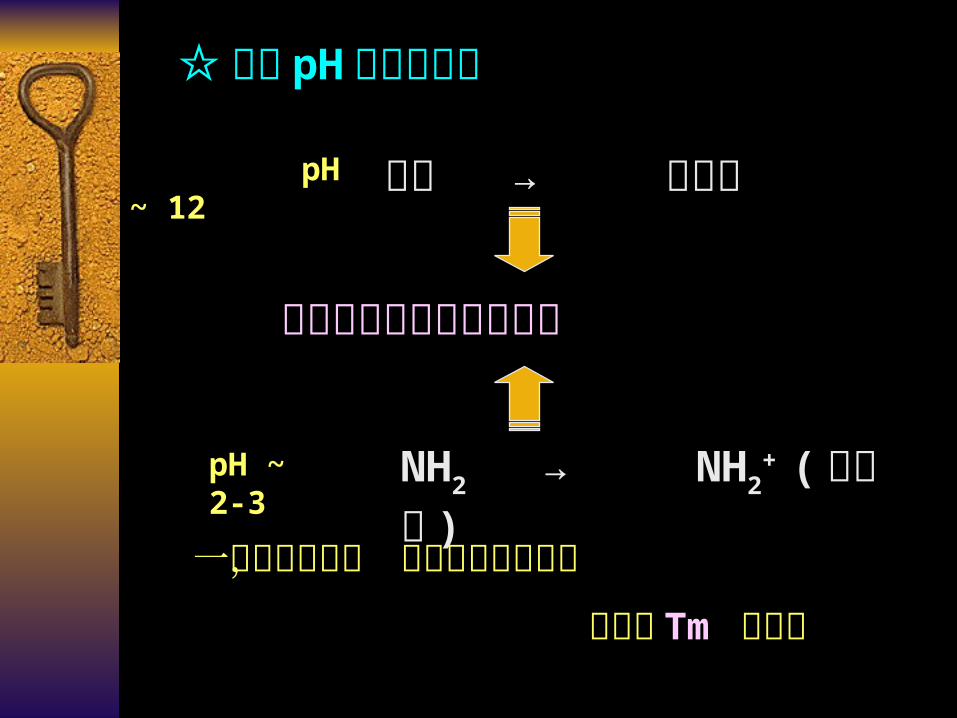
pH ~ 12 酮基 → 烯醇基
pH ~ 2-3 NH2 → NH2+ ( 质子化 )
改变氢键的形成与结合力
☆极端 pH 条件的影响
一切减弱氢键, 碱基堆积力的因素
均将使 Tm 值降低
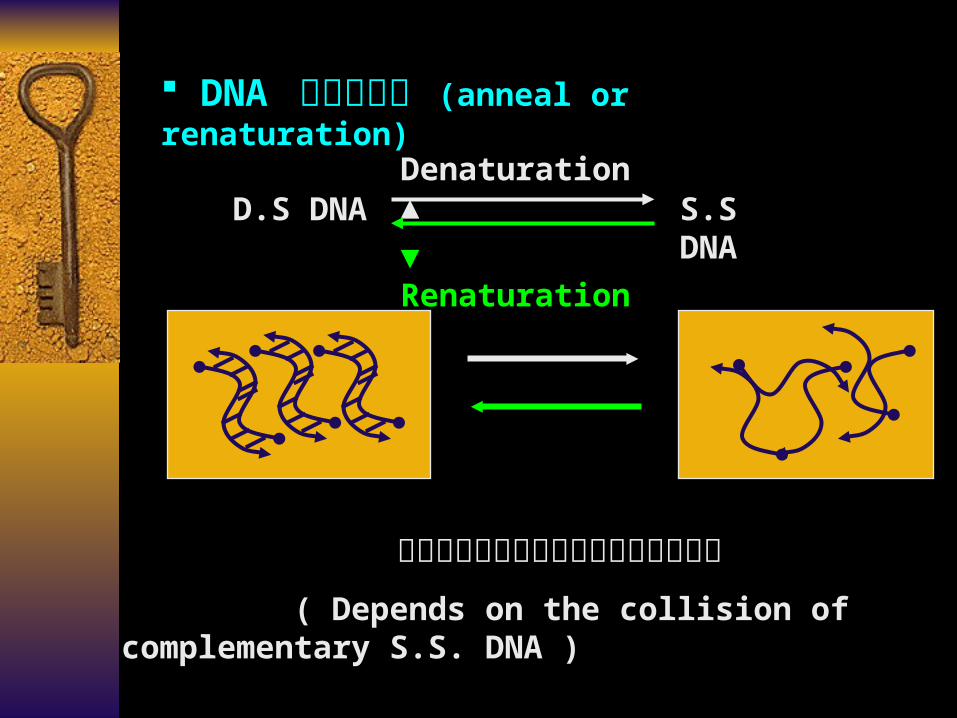
DNA 分子的复性 (anneal or renaturation)
D.S DNA S.S DNADenaturation ▲
▼ Renaturation
复性过程依赖于单链分子间的随机碰撞
( Depends on the collision of complementary S.S. DNA )

影响 DNA复性过程的因素 :
• 阳离子浓度 0.18 ~ 0.2M Na+ 可消除 polydNt 间的静电斥力
• 复性反应的温度 Tm - 25 (50-65 ) ℃ ℃
以消除 S.S. DNA 分子内的部分二级结构
• S.S. DNA 分子的长度S.S. DNA 愈长S.S. DNA愈短
→ 分子扩散愈慢 → 复性愈慢→ 分子扩散愈快 → 复性愈快
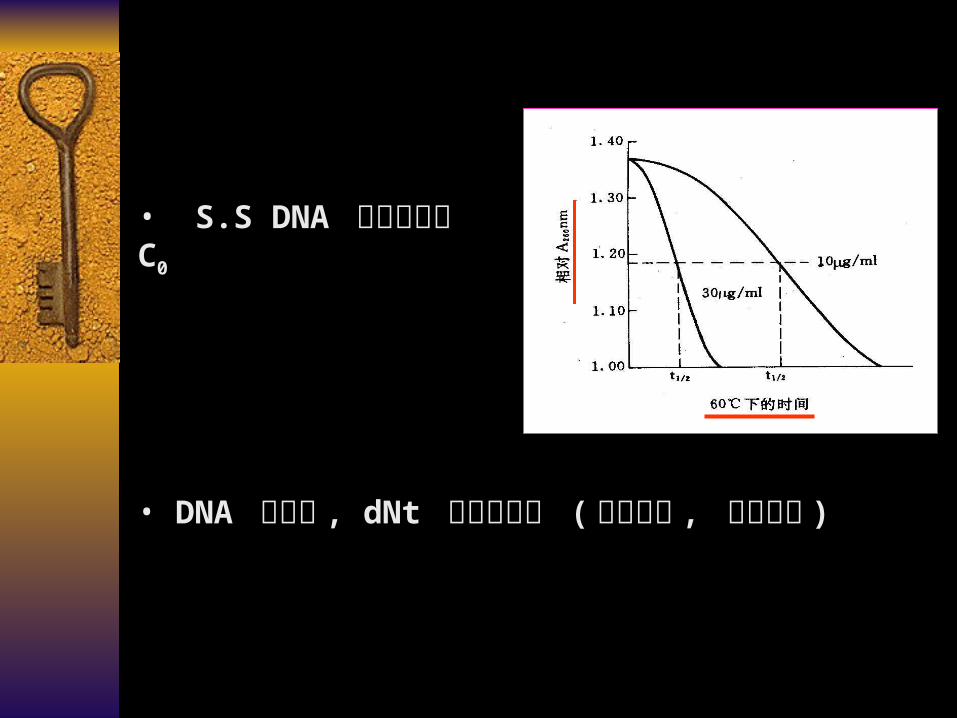
• DNA 分子中 , dNt 的排列状况 (随机排列 , 重复排列 )
• S.S DNA 的初始浓度 C0
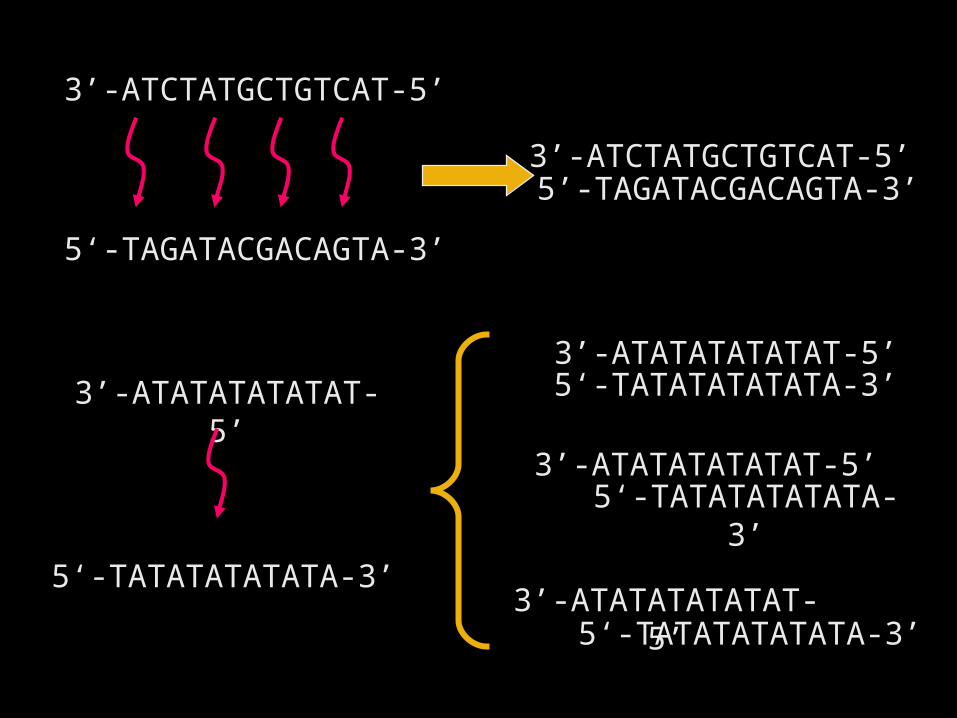
3’-ATCTATGCTGTCAT-5’
5‘-TAGATACGACAGTA-3’
5’-TAGATACGACAGTA-3’3’-ATCTATGCTGTCAT-5’
3’-ATATATATATAT-5’
5‘-TATATATATATA-3’
3’-ATATATATATAT-5’5‘-TATATATATATA-3’
5‘-TATATATATATA-3’3’-ATATATATATAT-5’
3’-ATATATATATAT-5’5‘-TATATATATATA-3’

复性发生的过程的讨论
遵循 second – order kinetics formula (二级反应动力学)
dCt / dt = -KCt2
反应初始 t = 0
• 两条部分同源 ( 小于 20dNt) 的 S.S. DNA, 在复性过程中
形成的部分双链区是不稳定的• 单链 DNA 的随机碰撞 过程( randomly collision )
单链 DNA浓度 = C0
反应达 t 时
单链 DNA浓度 = Ct
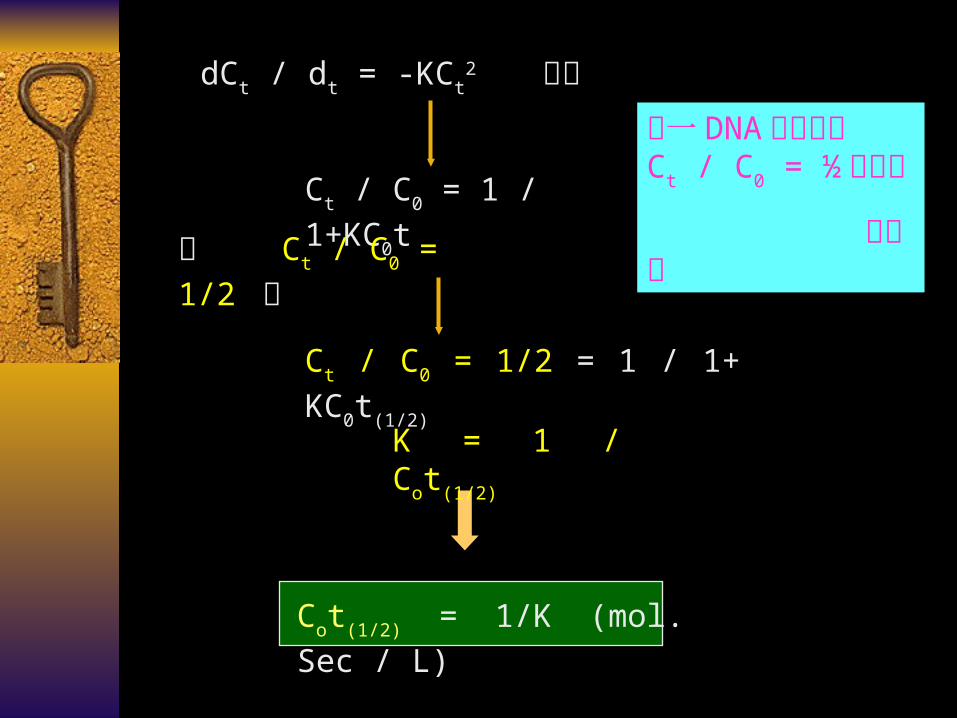
dCt / dt = -KCt2 积分
Ct / C0 = 1 / 1+KC0t
当 Ct / C0 = 1/2 时
Ct / C0 = 1/2 = 1 / 1+ KC0t(1/2)
K = 1 / Cot(1/2)
Cot(1/2) = 1/K (mol. Sec / L)
任一 DNA 分子达到 Ct / C0 = ½ 的速率
是定值

在控制反应条件相同的前提下 , 两种 DNA 分子的 C0t(1/2) 值 ,
取决于 dNt 的排列复杂性
( 复性动力学的复杂性 , Kinetic complexity, K.C )
AAAAAAAA K. C. = 1 C0t(1/2) = 2×10-6
ATCGATCGATCG K.C. = 4
K.C. = 5 ×105 C0t(1/2) = 1
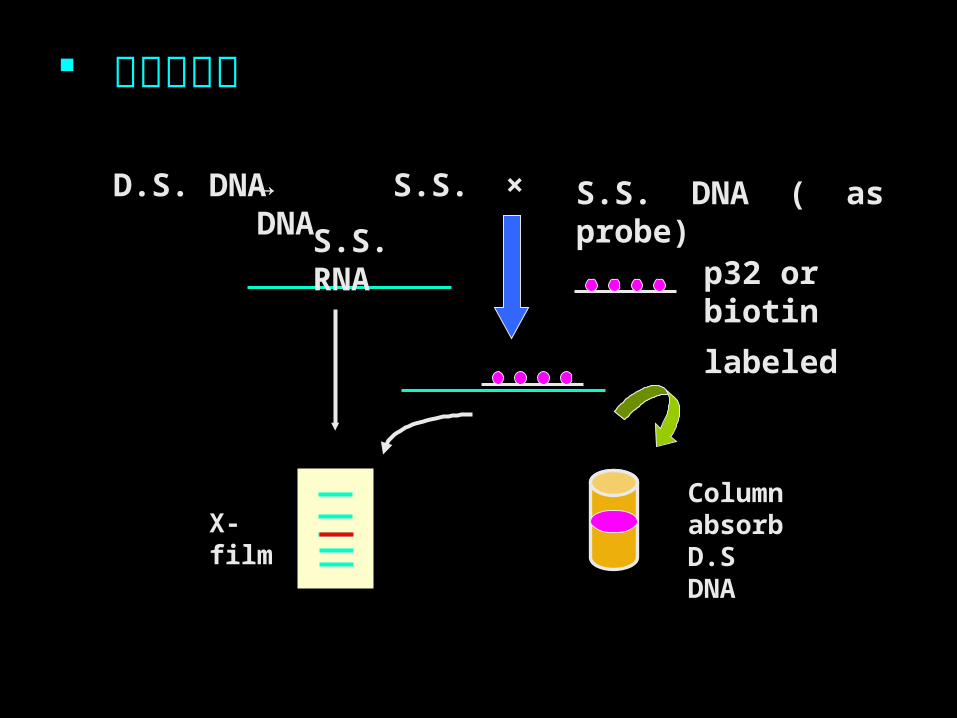
D.S. DNA
X-filmColumn absorb D.S DNA
p32 or biotin
labeled
S.S. DNA ( as probe)×
→ S.S. DNA
S.S. RNA
核酸的杂交
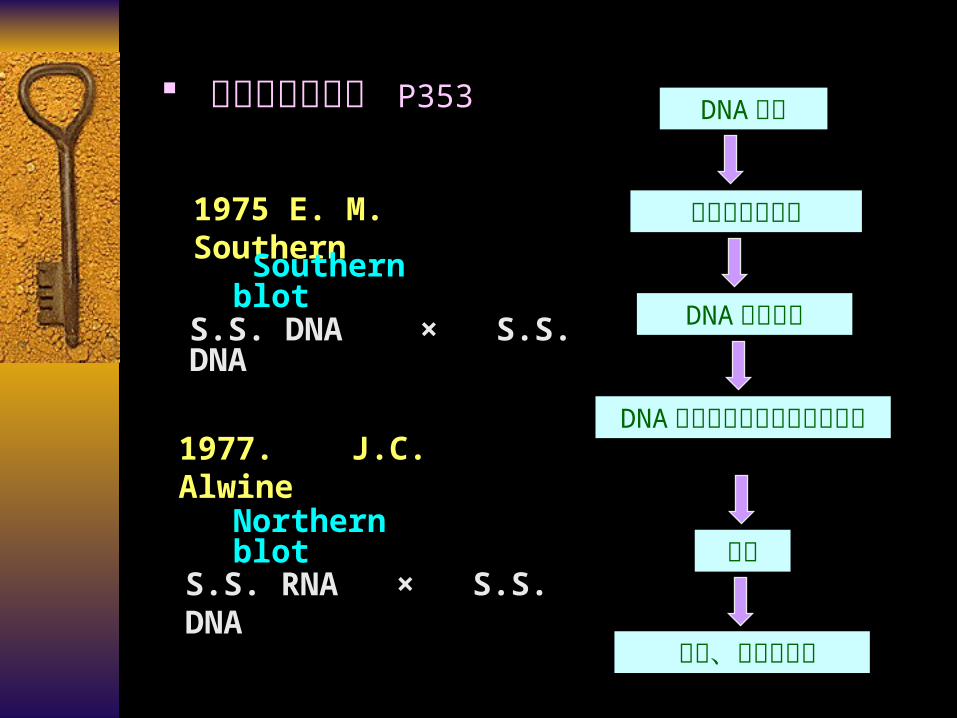
核酸的固相杂交 P353 DNA 酶解
琼脂糖凝胶电泳
DNA 片段转膜
DNA 膜与同位素标记的探针杂交
洗膜
压片、放射自显影
1975 E. M. Southern
Southern blot
S.S. DNA × S.S. DNA
1977. J.C. Alwine
Northern blot
S.S. RNA × S.S. DNA
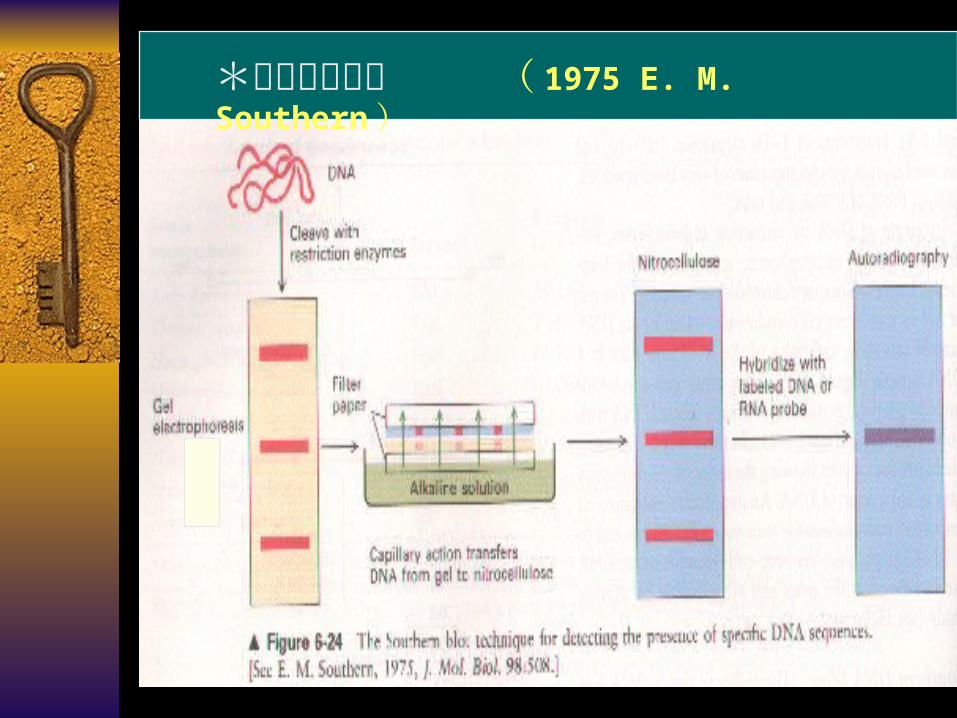
*固相分子杂交 ( 1975 E. M. Southern )
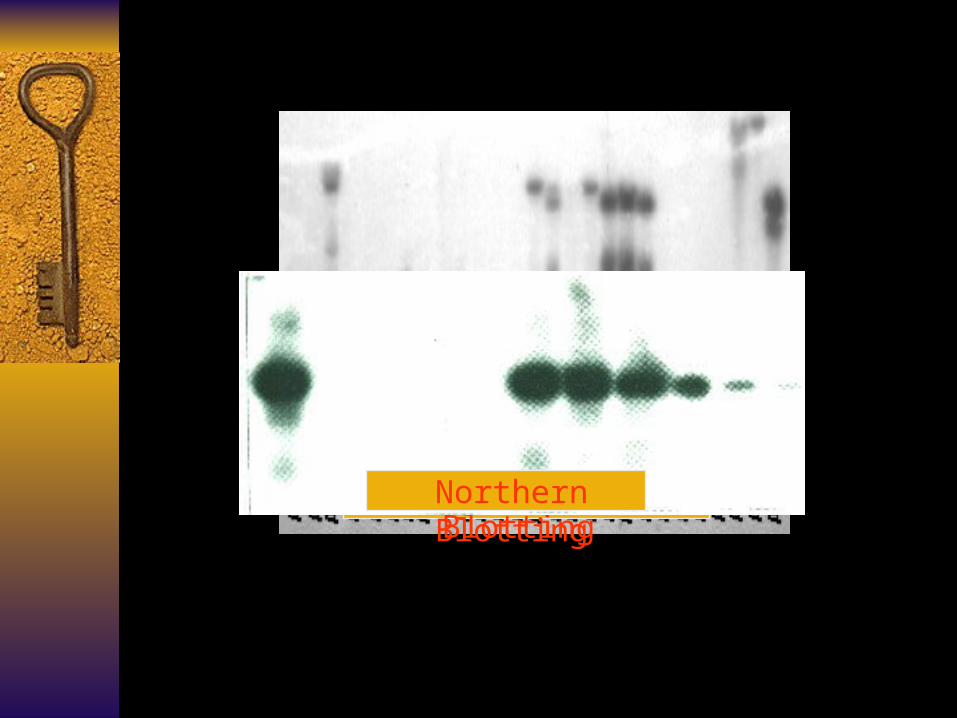
Southern BlottingNorthern Blotting

8.5.5 DNA 的固相合成原理
首先将待活化的核苷酸上的某些基团( 5’-羟基,碱基上的氨基)保护起来,然后将 3’-羟基活化。第一个核苷酸的 3’-羟基与树脂结合,5’-羟基与活化核苷酸的 3’-羟基间形成亚磷酸三酯。亚磷酸三酯氧化成磷酸三酯。 5’-羟基解保护,进入下一轮延伸反应。

8.5.6 DNA 的限制酶图谱
限制性内切酶 restriction
endonuclease (RE)
自身的 DNA经过甲基化修饰
降解外来 DNA但不降解自身 DNA
作用位点具有极高的专一性

Restriction Endonuclease 的种类
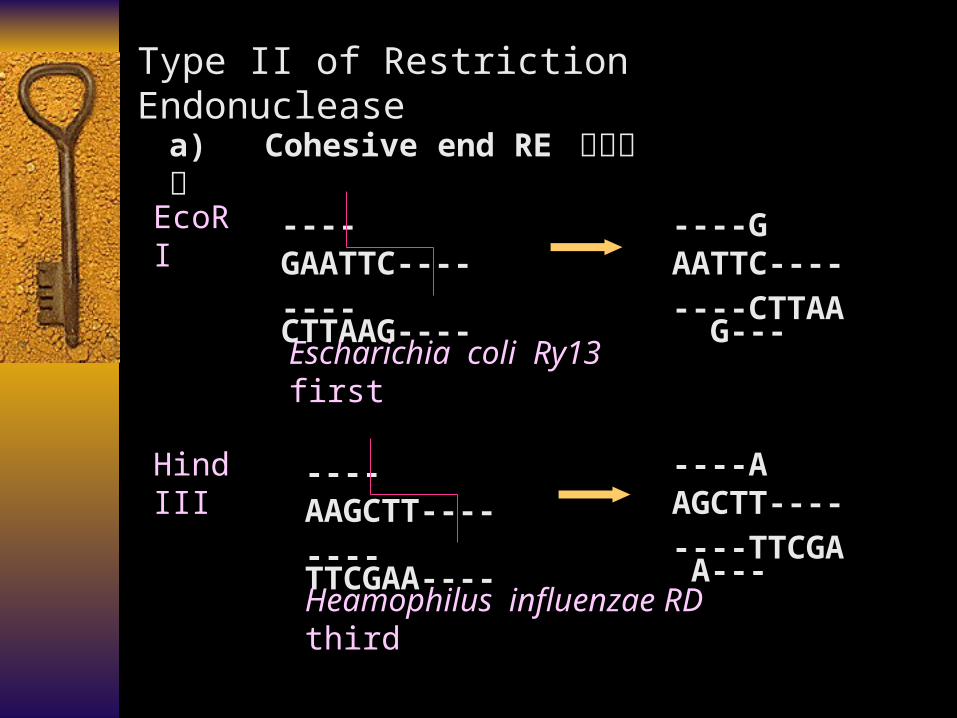
Type II of Restriction Endonuclease
a) Cohesive end RE 粘性末端
----GAATTC----
----CTTAAG----
----AAGCTT----
----TTCGAA----
Escharichia coli Ry13 first
----G AATTC----
----CTTAA G---
EcoR I
Hind III ----A AGCTT----
----TTCGA A---
Heamophilus influenzae RD third
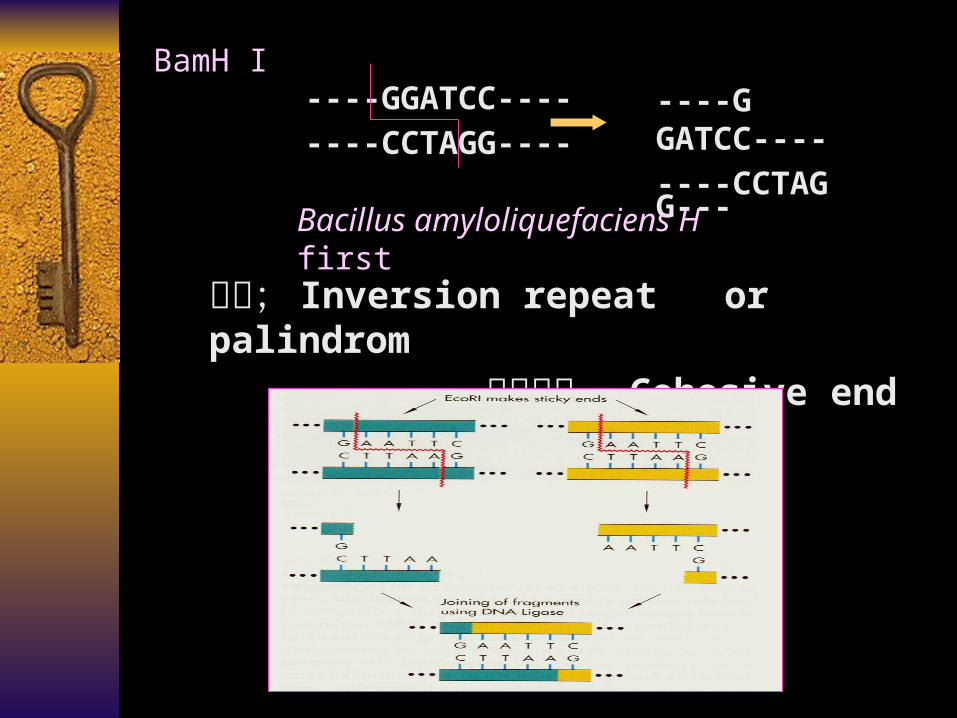
特点; Inversion repeat or palindrom
粘性末端 Cohesive end
----GGATCC----
----CCTAGG----
BamH I----G GATCC----
----CCTAG G---
Bacillus amyloliquefaciens H first
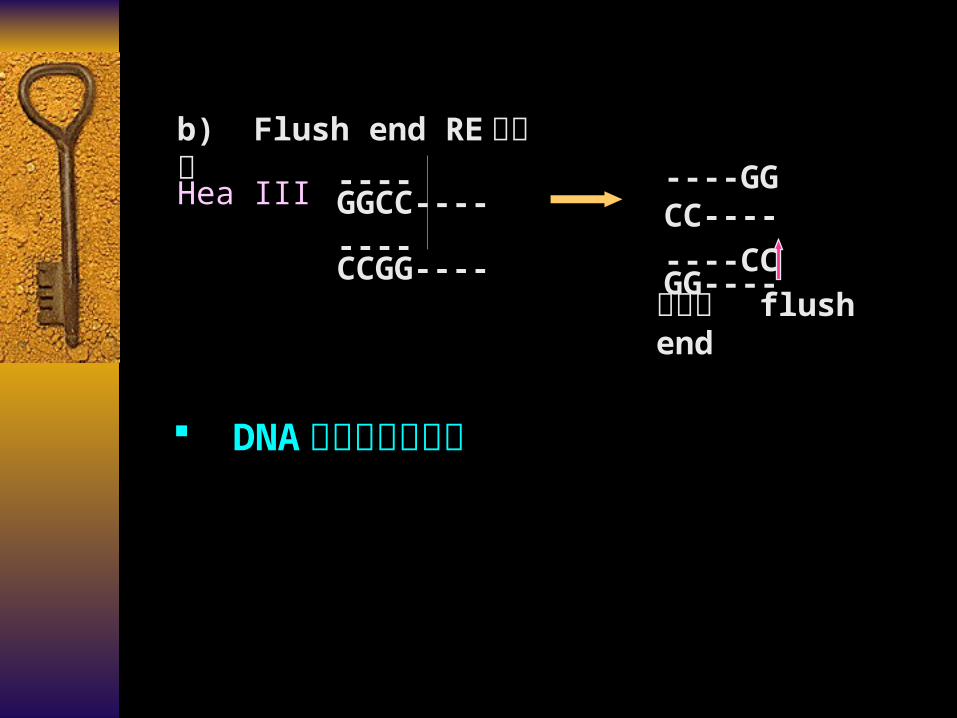
b) Flush end RE 平末端----GG CC----
----CC GG----Hea III ----GGCC----
----CCGG----
平末端 flush end
DNA 的限制性酶图谱





























