Embed Size (px)
DESCRIPTION
C itoskelets 6. tēma. Citoskelets. Vairāk kā 300 gadus šūnas ir bijuši vairāk vai mazāk nekustīgi veidojumi. - PowerPoint PPT Presentation
Citation preview

Citoskelets6. tēma

Citoskelets
• Vairāk kā 300 gadus šūnas ir bijuši vairāk vai mazāk nekustīgi veidojumi.
• Tikai sākot ar 20 gadsimtu atklāja pirmās struktūras, kas veic kustības. Tās bija centriolas, kuras piedalījās hromosomu atvilkšanā. Nedaudz vēlāk muskuļu šūnās atklāja mikrofilamentus un to lomu kontrakcijās un viciņas ar to lomu šūnu pārvietošanā.
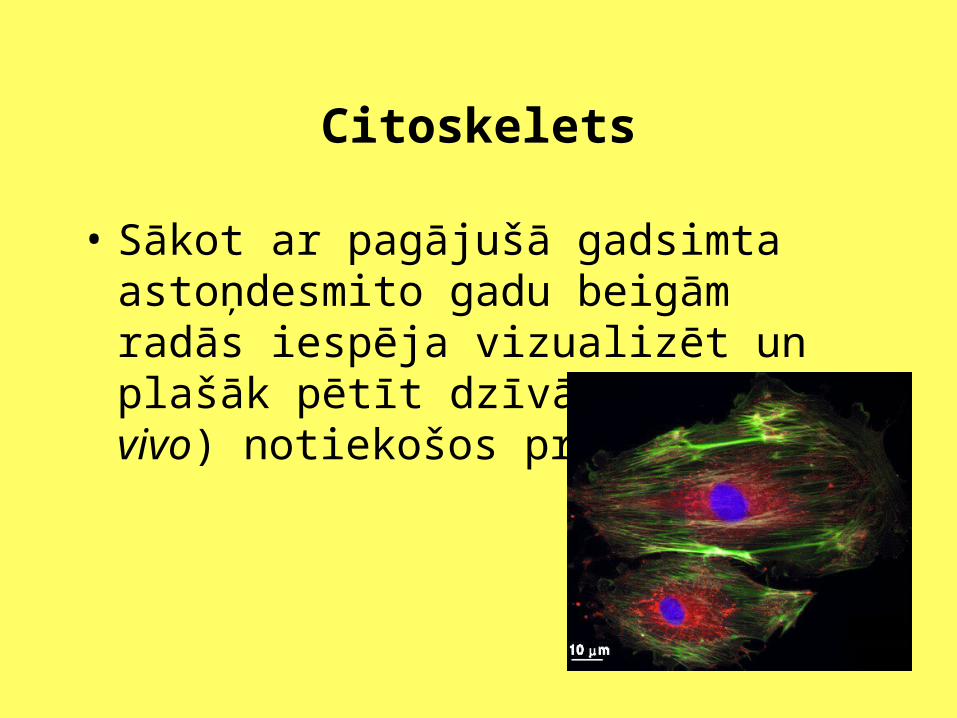
Citoskelets
• Sākot ar pagājušā gadsimta astoņdesmito gadu beigām radās iespēja vizualizēt un plašāk pētīt dzīvās šūnās (in vivo) notiekošos procesus.

Citoskelets
• Hloroplastu kustība elodejas lapā• http://priede.bf.lu.lv/ramis.shtml

Citoskeletam līdzīgie elementi prokariotu šūnās
Prokariotu šūnās nav novērojamas pat viskonservatīvākās eikariotu citoskeleta olbaltumvielas. Tomēr līdzīga situācija ir arī ar DNS iepakojumu prokariotu un eikariotu šūnās. Prokariotiem nav histonu olbaltumvielas, bet ir histoniem līdzīgas olbaltumvielas. Tās veic līdzīgas funkcijas, bet atšķiras pēc savas uzbūves. Pat visvienkāršākajās prokariotu šūnās - mikoplazmās - ir atrastas tubulīnam līdzīgas olbaltumvielas, kurām ir citoskeleta funkcijas. Šie citoskeleta elementi pašlaik tiek aktīvi pētīti. Tomēr to uzbūve un funkcijas pagaidām ir neskaidras. Citā prokariotu šūnu grupā Archaebacteria ir konstatētas filamentveidīgas struktūras. Taču to uzbūve atšķiras no eikariotu šūnu mikrofilamentiem un starpfilamentiem. Uzskata, ka tā varētu būt pārejas forma starp eikariotu un prokariotu šūnām.
Skat. filmu:
http://www-micro.msb.le.ac.uk/Video/Rhodospirillum.mov
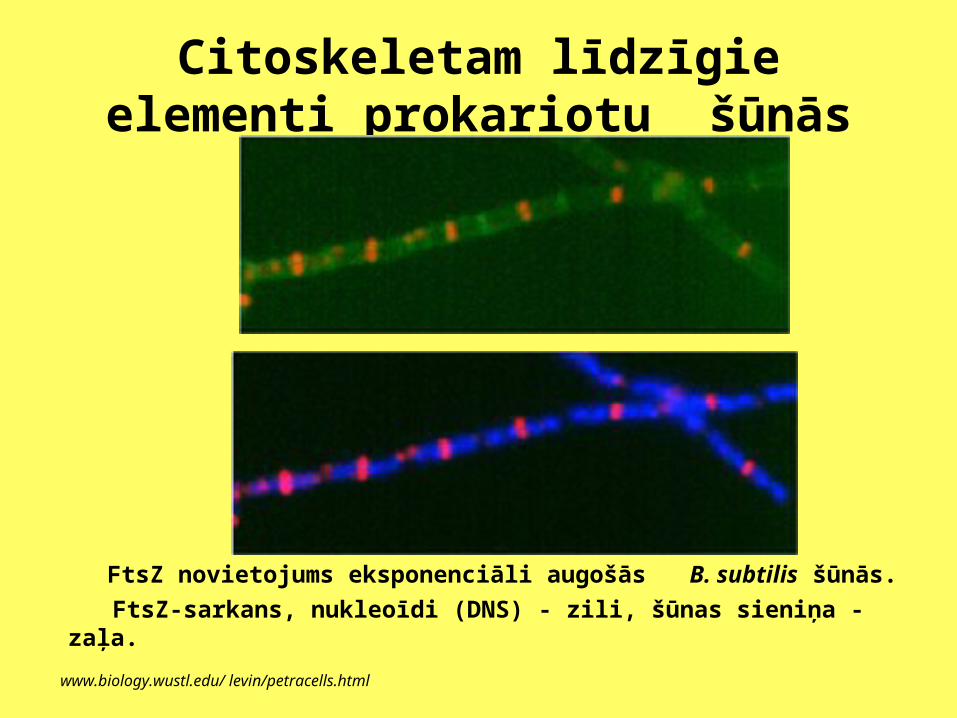
Citoskeletam līdzīgie elementi prokariotu šūnās
FtsZ novietojums eksponenciāli augošās B. subtilis šūnās.
FtsZ-sarkans, nukleoīdi (DNS) - zili, šūnas sieniņa - zaļa.
www.biology.wustl.edu/ levin/petracells.html

Prokariotu viciņas
Baktērijas ir kustīgi organismi. Tās spēj attālināties no vietām ar nepiemērotiem apstākļiem, un tās spēj pārvietoties uz īpaši labvēlīgām vietām. Tās reaģē uz dažāda veida ķīmiskajiem kairinājumiem. Baktēriju kustības, kuras izraisa ķīmisko vielu klātbūtne, sauc par hemotaksijām.
Baktēriju kustības nodrošina viciņu rotācijas kustība. Viciņu diametrs ir tikai 20 nm, tas ir, mazāk nekā vienas mikrocaurulītes diametrs. Eikariotu šūnās viciņu iekšienē atradīsies mikrocaurulīšu kūlītis.

Prokariotu viciņas
• Baktēriju viciņas garums ir no 10 - 100 m. Baktēriju kustība ir haotiska, ja tās atrodas labvēlīgā vai neitrālā vidē. Tas ir tāpēc, ka viciņas brīžiem rotē pulksteņrādītāja virzienā, bet brīžiem pretēji. Ja vidē parādās barības vielas vai inde, tad baktēriju viciņu pavedienveida olbaltumvielas izveido kūlīti, kur tās kustas pretēji pulksteņa rādītāja virzienam.
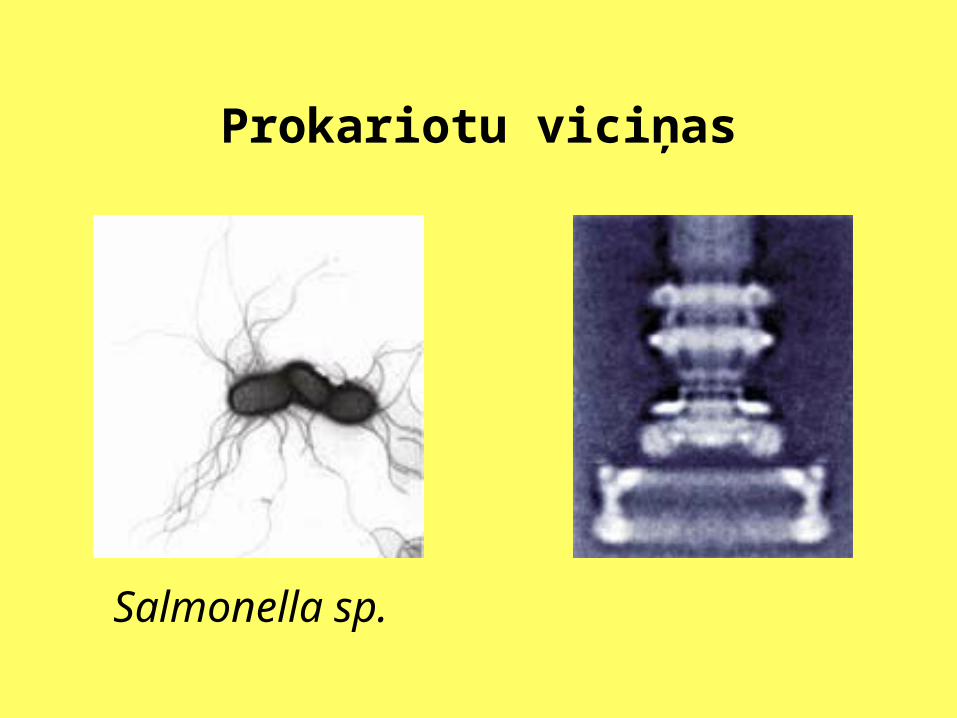
Prokariotu viciņas
Salmonella sp.

Prokariotu viciņas Flagelīna viciņas pamatnē ir elastīga un kustīga āķa
daļa, kas iet cauri šūnas sieniņai. Āķa pamatne savukārt ir piestiprināta pie diskveidīgu olbaltumvielu kompleksa, kas atrodas plazmatiskajā membrānā. To sauc par bazālo ķermenīti. Bazālais ķermenītis un āķa pamatne ir ietverta šūnas sieniņā un plazmatiskajā membrānā. Bazālais ķermenītis ir kompleksa struktūra un to veido apmēram 20 olbaltumvielas. Tā uzbūve ir atšķirīga grampozitīvajās un gramnegatīvajās baktērijās.
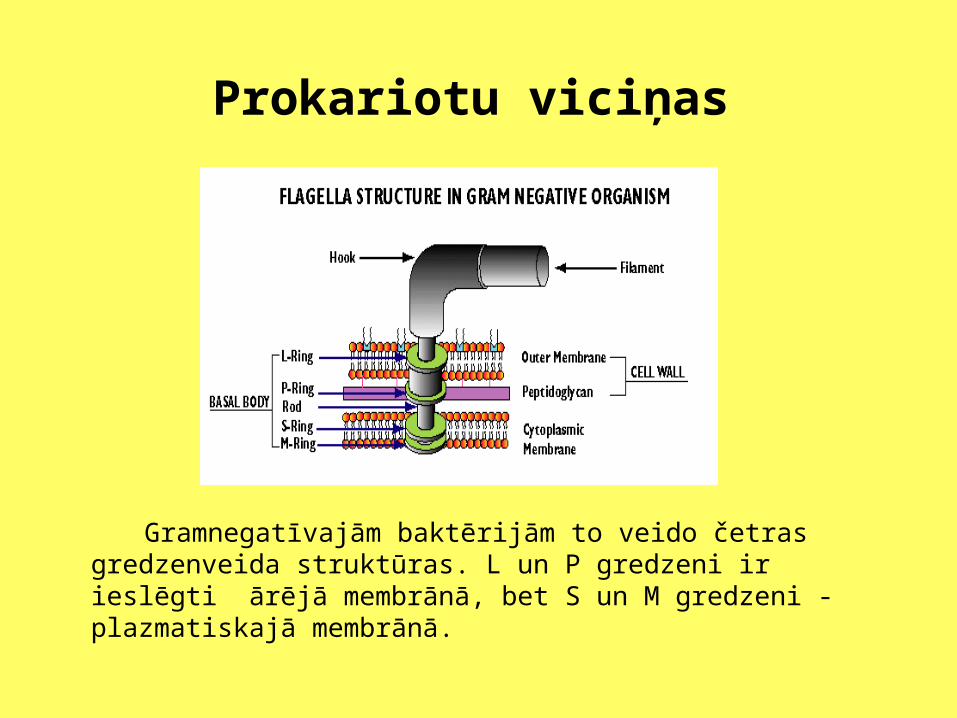
Prokariotu viciņas
Gramnegatīvajām baktērijām to veido četras gredzenveida struktūras. L un P gredzeni ir ieslēgti ārējā membrānā, bet S un M gredzeni - plazmatiskajā membrānā.

Prokariotu viciņas
Šie olbaltumvielu diski darbojas līdzīgi elektromotoram.
Diska ārējo daļu veido nelielu olbaltumvielu gredzens - stators, bet centrā atrodas rotors.
Gredzena ārējā daļa ir stabili nostiprināta baktēriju šūnas sieniņā. Rotācijas kustību šeit nodrošina nevis elektromagnētiskais lauks, bet H+ gradients, kas pastāv starp citoplazmu un periplazmatisko telpu.

Prokariotu viciņas
Rotācijas kustība var notikt samērā lielā temperatūras diapazonā. Turklāt tai ir ārkārtīgi augsts lietderības koeficients - gandrīz 100%.
Tā kā viciņas āķa daļa ir piestiprināta pie rotora, tad tā rotē kopā ar to. Rotācijas ātrums var būt līdz 150 apgriezieniem sekundē. Kustības virziena regulācija notiek statora daļā. Tur var piesaistīties signālmolekulas un izmainīt protonu plūsmu caur membrānu.

Prokariotu viciņas
Kustības regulē receptoru sistēma, kas atrodas baktēriju plazmatiskajā membrānā. Ārējā membrānā ir poras, kas ļauj periplazmatiskajā telpā iekļūt vielām no apkārtējās vides. Šīs vielas saistās ar pārnesējolbaltumvielām, kas atrodas periplazmatiskajā telpā. Pārnesējmolekulas saistās ar kādu no receptoriem, kas atrodas plazmatiskajā membrānā. Vienā gadījumā receptora signāls šūnā izpaudīsies kā signālmolekulas fosforilēšana, otrā - kā defosforilēšana.
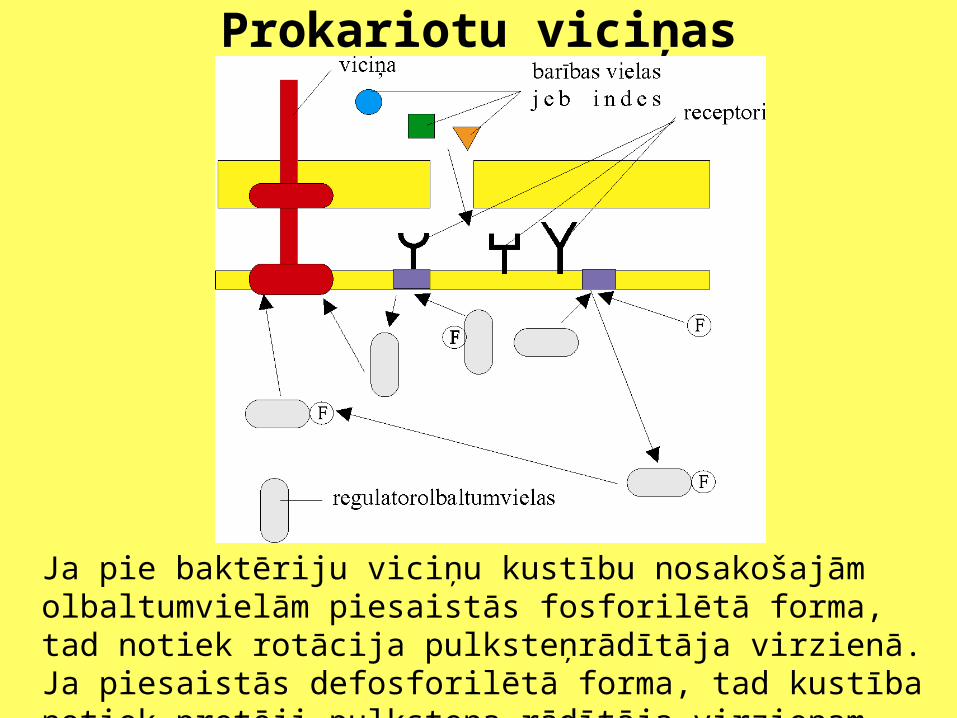
Prokariotu viciņas
Ja pie baktēriju viciņu kustību nosakošajām olbaltumvielām piesaistās fosforilētā forma, tad notiek rotācija pulksteņrādītāja virzienā. Ja piesaistās defosforilētā forma, tad kustība notiek pretēji pulksteņa rādītāja virzienam.

Citoskelets
Visās eikariotu šūnās ir konstatētas dažādas pavedienveida struktūras. Visvairāk tās ir izpētītas dzīvnieku šūnās. Dažos gadījumos pavedieni ir apvienojušies augstāk organizētās struktūrās, kuras var redzēt pat gaismas mikroskopā. Tādi veidojumi ir centriolas, bazālā plātnīte u.c.
Visiem pavedieniem ir dots kopīgs nosaukums - citoskelets.
Citoskelets
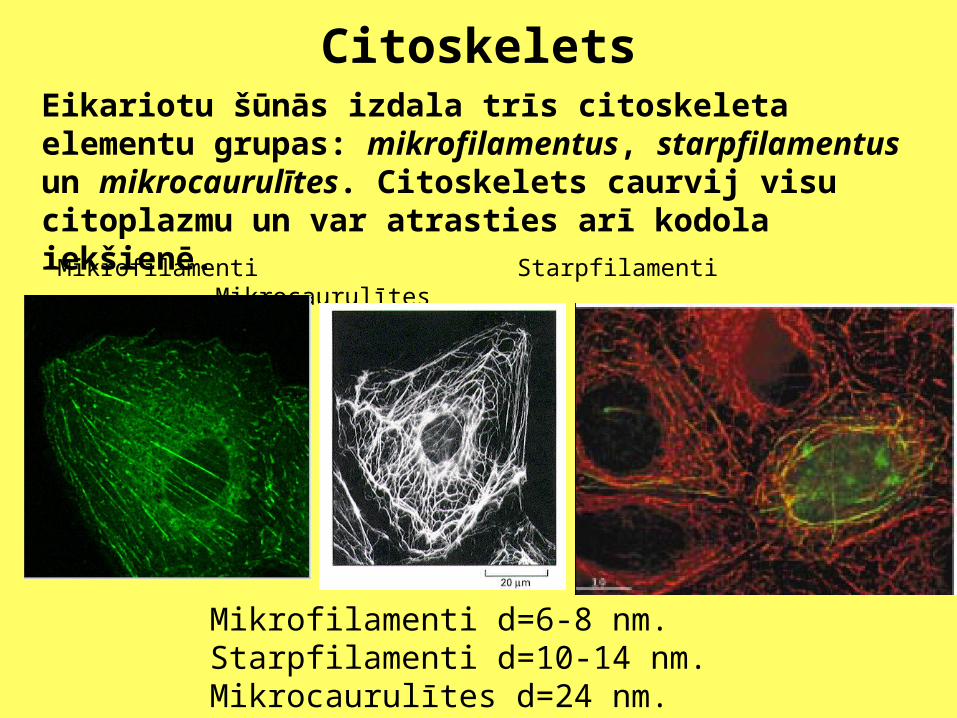
Mikrofilamenti Starpfilamenti Mikrocaurulītes
Mikrofilamenti d=6-8 nm.Starpfilamenti d=10-14 nm.Mikrocaurulītes d=24 nm.
Eikariotu šūnās izdala trīs citoskeleta elementu grupas: mikrofilamentus, starpfilamentus un mikrocaurulītes. Citoskelets caurvij visu citoplazmu un var atrasties arī kodola iekšienē.

Mikrocaurulītes
Mikrocaurulītes ir sastopamas visās eikariotiskajās šūnās. Tās kā blīvs tīkls ietver kodolu un atiet uz šūnas perifēriju. Mikrocaurulītes sastāv no tubulīna. Tubulīns ir globulāra olbaltumviela, kurai piemīt spēja polimerizēties.
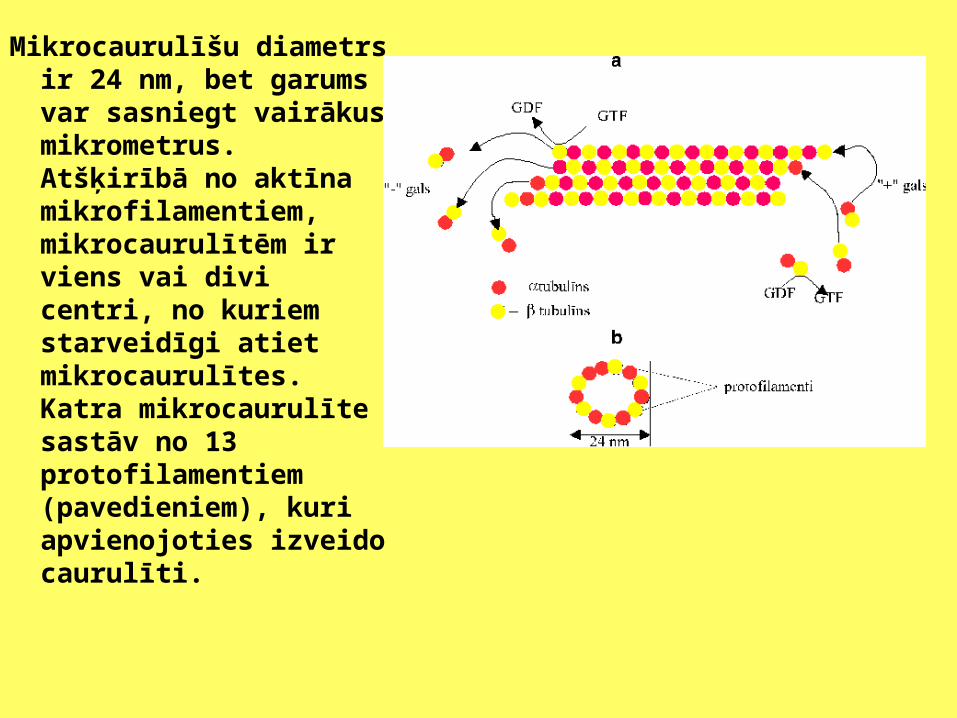
Mikrocaurulīšu diametrs ir 24 nm, bet garums var sasniegt vairākus mikrometrus. Atšķirībā no aktīna mikrofilamentiem, mikrocaurulītēm ir viens vai divi centri, no kuriem starveidīgi atiet mikrocaurulītes. Katra mikrocaurulīte sastāv no 13 protofilamentiem (pavedieniem), kuri apvienojoties izveido caurulīti.
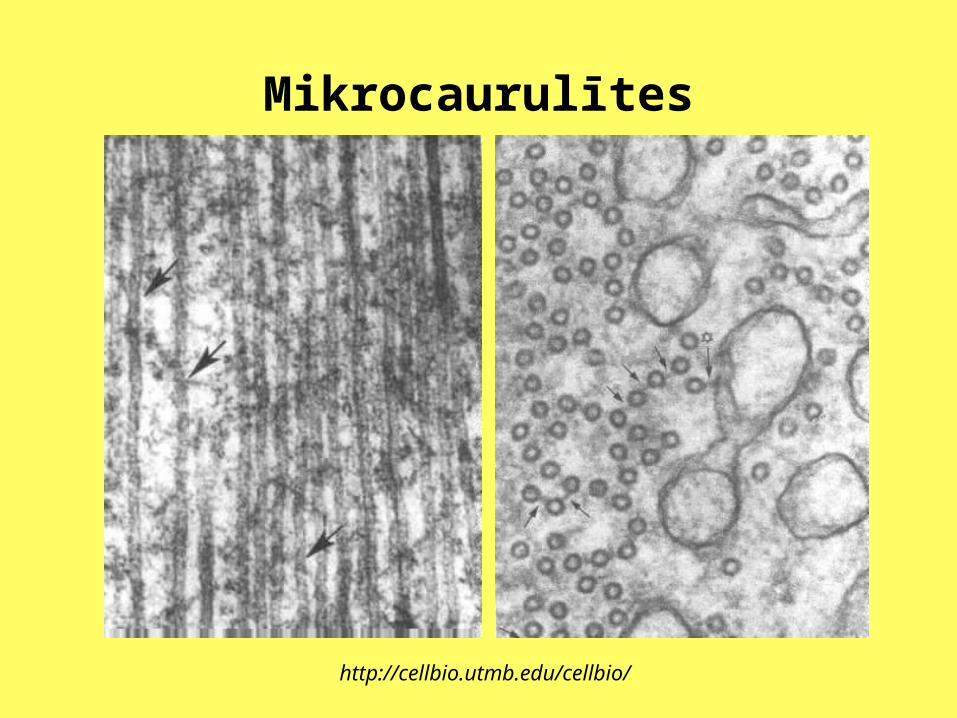
Mikrocaurulītes
http://cellbio.utmb.edu/cellbio/

Mikrocaurulītes
• Mikrocaurulīšu polimerizāciju traucē kolhicīns un citi savienojumi, kuri neļauj tubulīna dimēriem pievienoties pie mikrocaurulītes.
http://cellbio.utmb.edu/cellbio/

Vielu un organellu transports, izmantojot mikrocaurulītes
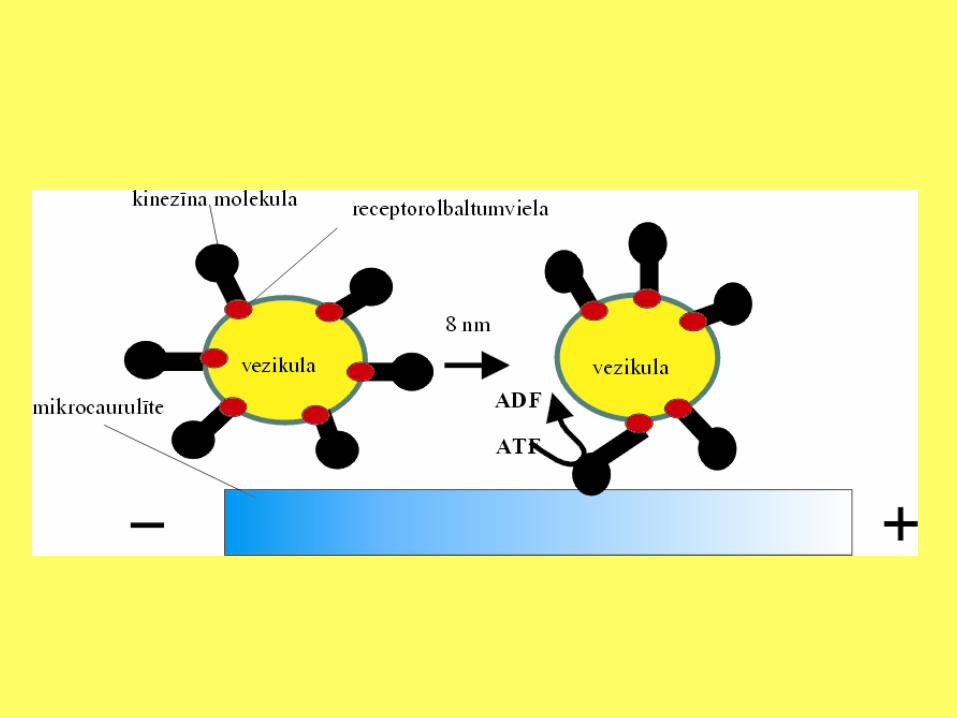
Gandrīz visās eikariotu šūnās makromolekulu, vezikulu un organellu transportu nodrošina mikrocaurulītes. Dažos gadījumos to pagarināšanās un saīsināšanās nodrošina pārvietošanu. Tomēr parasti to veic motorās olbaltumvielas, kas vienlaicīgi var pievienoties mikrocaurulītei un transportējamam objektam. Motorās olbaltumvielas pieder pie divām grupām. Vienas sauc par dineīniem, otras par kinezīniem. Katrā grupā ietilpst dažāda lieluma un nedaudz atšķirīgas aminoskābju secības polipeptīdu ķēdes. Dineīns nodrošina vielu transportu no šūnu perifērijas uz šūnu centru, bet kinezīns no šūnu centra uz perifēriju.
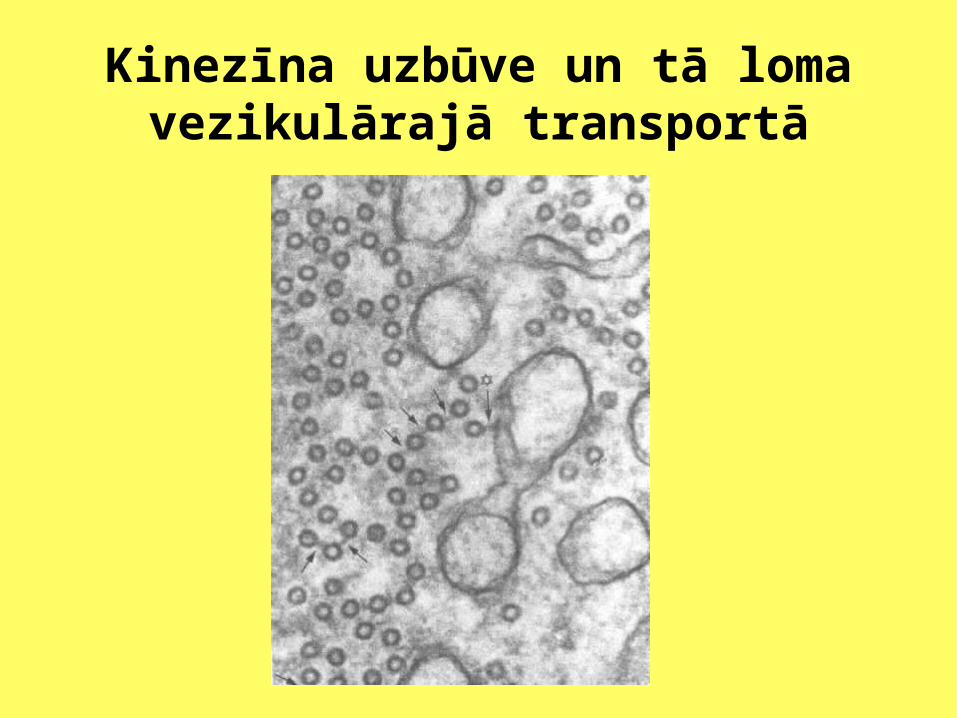
Kinezīna uzbūve un tā loma vezikulārajā transportā

Mikrocaurulītes organizējošie centri
Mikrocaurulītes visās eikariotu šūnās dažās zonās ir apvienotas un veido vairāk vai mazāk kompaktas struktūras, no kurām starveidīgi atiet mikrocaurulītes:
• dzīvnieku šūnā tāda struktūra ir centrosoma,
• vienšūņiem - bazālais ķermenītis,
• daudzām sēnēm - vārpstu veidojošais ķermenis,
• augiem - kodola apvalka ārējā membrāna.
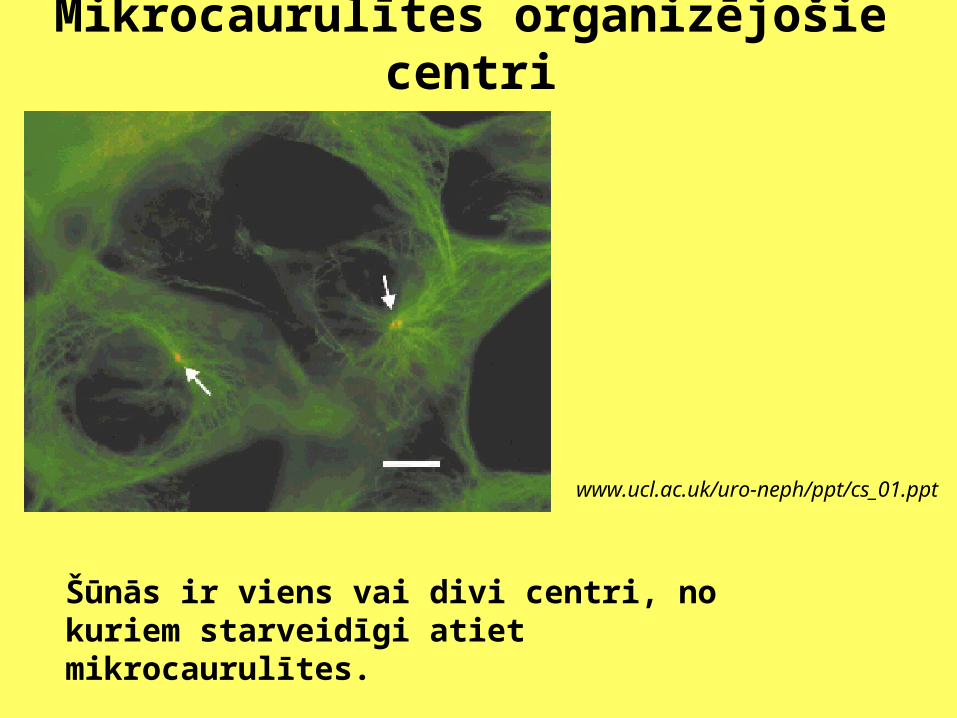
www.ucl.ac.uk/uro-neph/ppt/cs_01.ppt
Šūnās ir viens vai divi centri, no kuriem starveidīgi atiet mikrocaurulītes.
Mikrocaurulītes organizējošie centri

Centriolas
Nobriedušas centriolas distālās daļas mikrocaurulītes no ārpuses ir klātas ar olbaltumvielām - satelītiem un apendiksiem. Abu centriolu proksimālās daļas savieno nelieli pavedieni. To molekulārā uzbūve ir neskaidra. Nenobriedusī centriola arī ir saistīta ar jaunu mikrocaurulīšu nukleāciju. Šajā centrosomas daļā mitozes laikā ir augsta -tubulīna koncentrācija. Mikrocaurulītēm, kas starveidīgi iet ārā no šīs centrosomas zonas, "-" gals atrodas centrosomā, bet - "+" gals citoplazmā.
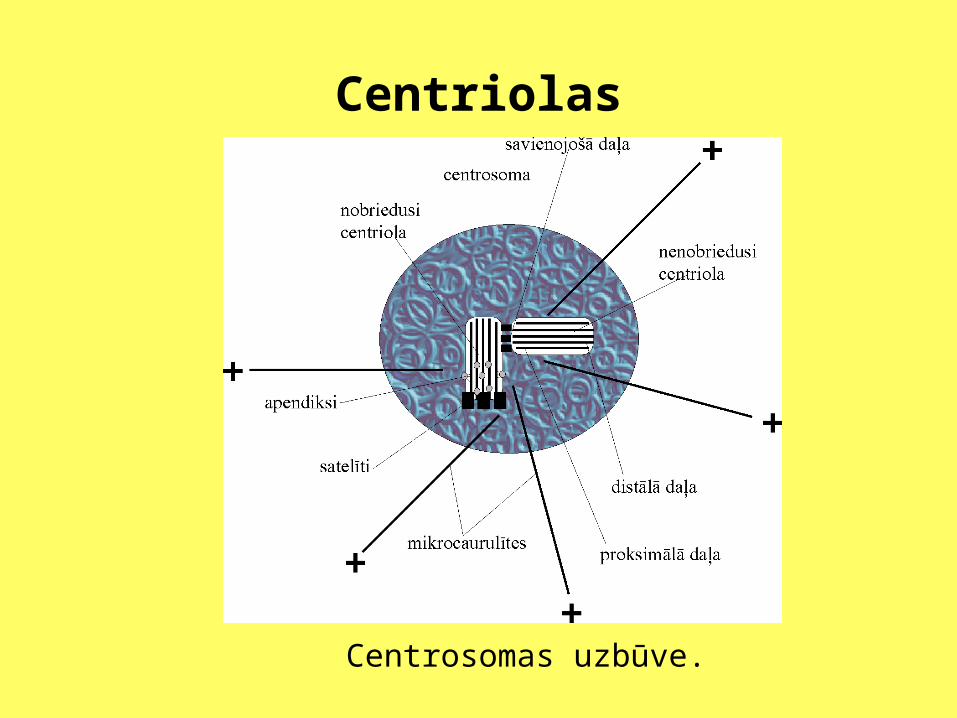
Centriolas
Centrosomas uzbūve.
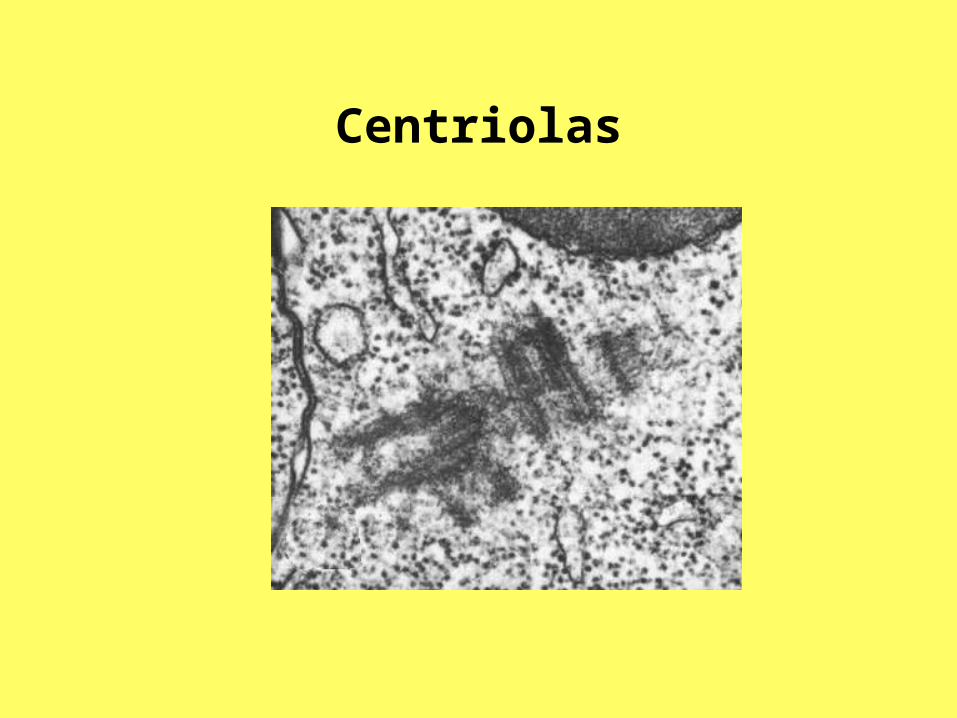
Centriolas

Centriolas Centriolas veido mikrocaurulīšu
kūlīšu komplekss. Kompleksu veido deviņi mikrocaurulīšu tripleti. Tripletā izšķir A, B un C mikrocaurulītes. Apskatot centriolu iekšējo uzbūvi, var redzēt, ka tā atšķiras gan nobriedušām un nenobriedušām centriolām, gan arī centriolas proksimālajam un distālajam galam.

Centriolas• Tripleti ir izkārtoti gredzenveidīgi un ir savstarpēji
savienoti. Perifērajos mikrocaurulīšu tripletos mikrocaurulītes nav vienādas. Centram tuvāko apzīmē ar A, vidējo ar B, bet ārējo ar C. Iekšējo mikrocaurulīti veido 13 protofilamenti, līdzīgi kā citoplazmas mikrocaurulītēs. Ārējo un vidējo mikrocaurulīti veido tikai 10 protofilamenti. Tas ir tādēļ, ka, apvienojoties saskarsmes plaknē, tām ir viena kopīga sieniņa.
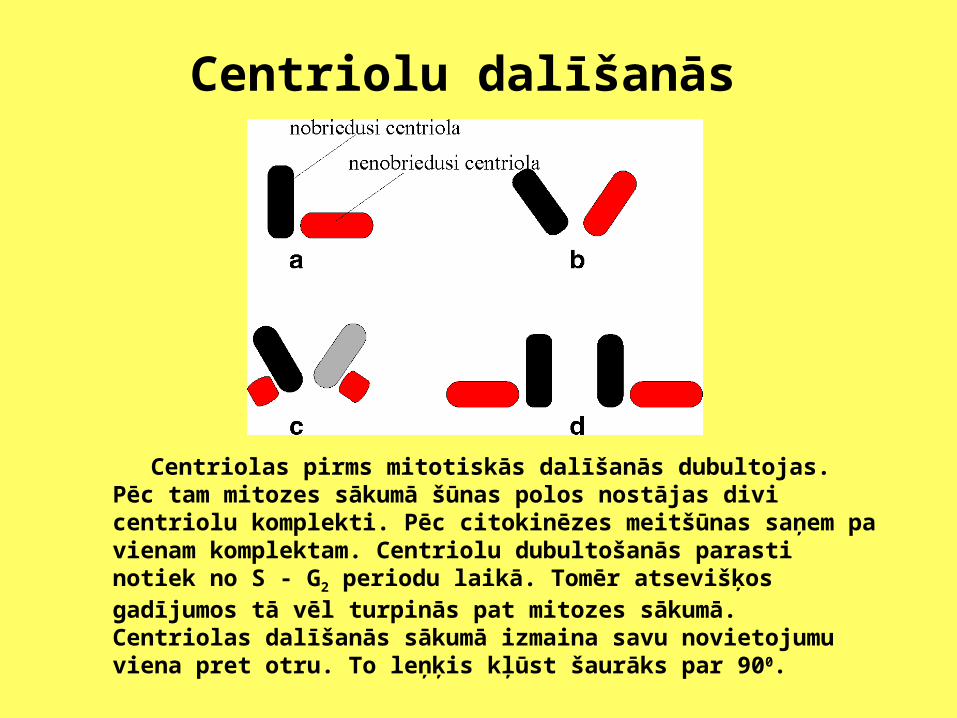
Centriolu dalīšanās
Centriolas pirms mitotiskās dalīšanās dubultojas. Pēc tam mitozes sākumā šūnas polos nostājas divi centriolu komplekti. Pēc citokinēzes meitšūnas saņem pa vienam komplektam. Centriolu dubultošanās parasti notiek no S - G2 periodu laikā. Tomēr atsevišķos gadījumos tā vēl turpinās pat mitozes sākumā. Centriolas dalīšanās sākumā izmaina savu novietojumu viena pret otru. To leņķis kļūst šaurāks par 900.
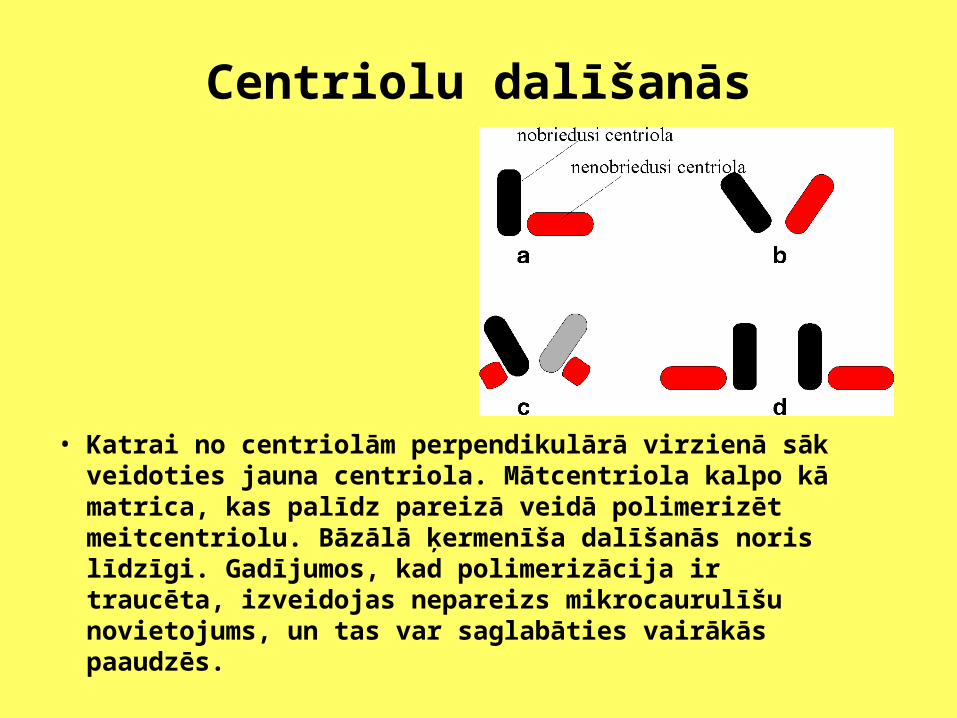
Centriolu dalīšanās
• Katrai no centriolām perpendikulārā virzienā sāk veidoties jauna centriola. Mātcentriola kalpo kā matrica, kas palīdz pareizā veidā polimerizēt meitcentriolu. Bāzālā ķermenīša dalīšanās noris līdzīgi. Gadījumos, kad polimerizācija ir traucēta, izveidojas nepareizs mikrocaurulīšu novietojums, un tas var saglabāties vairākās paaudzēs.

Skropstiņas, viciņas un bazālais ķermenītis
Skropstiņas ir smalki pavedieni, kuru diametrs ir apmēram 0,25 m. To garums svārstās dažādos audos un sugās, bet parasti ir apmēram 3 - 5 m. To centrālo daļu veido mikrocaurulīšu kūlītis, bet ārējo daļu - irdens matrikss, kas ir ietverts plazmatiskajā membrānā. Skropstiņas ir novērojamas vairākos dzīvnieku audu tipos, vienšūņos un zemākajos augos. Līdzīga iekšējā uzbūve ir arī eikariotu šūnu viciņām, tikai tās ir daudz garākas un sasniedz pat 200 m.
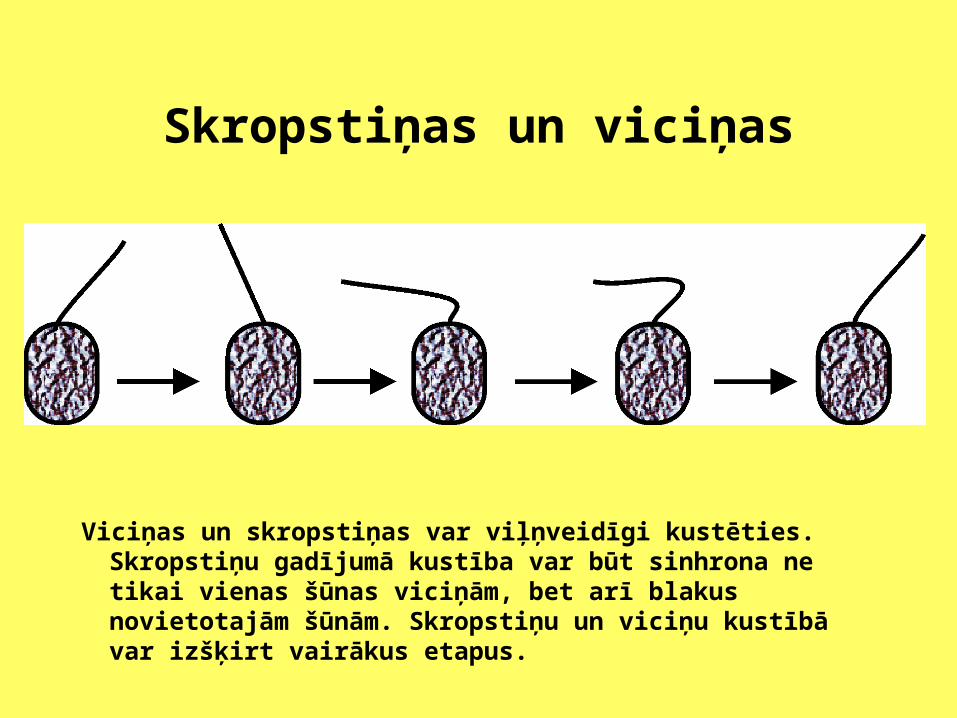
Skropstiņas un viciņas
Viciņas un skropstiņas var viļņveidīgi kustēties. Skropstiņu gadījumā kustība var būt sinhrona ne tikai vienas šūnas viciņām, bet arī blakus novietotajām šūnām. Skropstiņu un viciņu kustībā var izšķirt vairākus etapus.
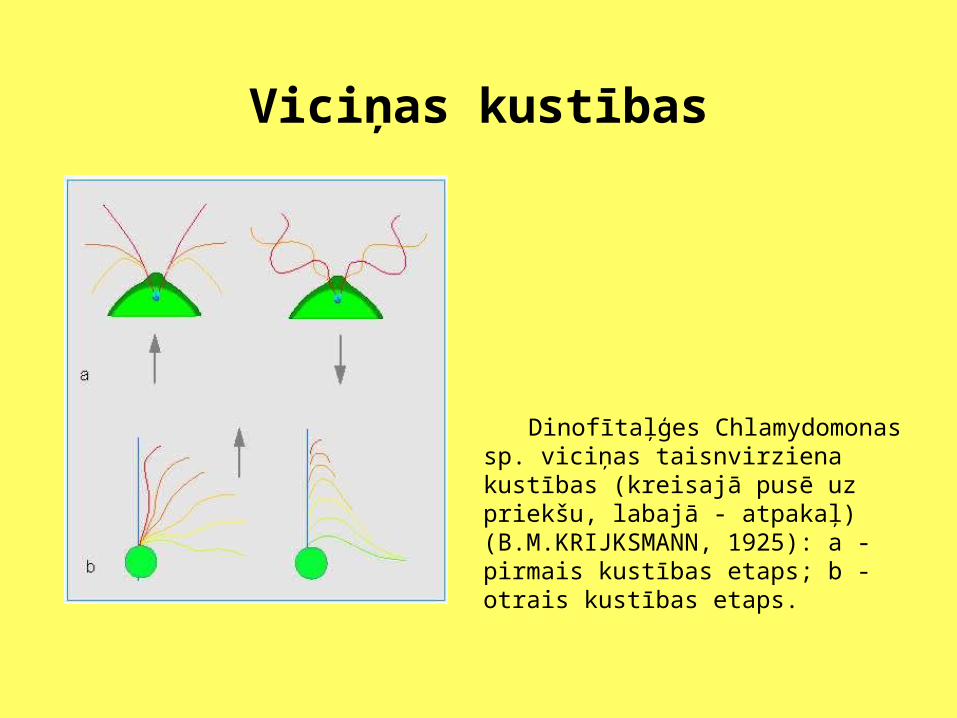
Viciņas kustības
Dinofītaļģes Chlamydomonas sp. viciņas taisnvirziena kustības (kreisajā pusē uz priekšu, labajā - atpakaļ) (B.M.KRIJKSMANN, 1925): a - pirmais kustības etaps; b - otrais kustības etaps.

Viciņas kustības
Eiglena video.Zvaniņinfuzorija video.

Skropstiņu kustības
Skropstaiņu kustība video.

Viciņas un skropstiņas
Kustības nodrošina mikrocaurulīšu kūlīša izmaiņas. Kūlīti veido deviņi mikrocaurulīšu dupleti. Tie ir izkārtoti gredzenveidīgi. Gredzena centrā atrodas vēl viens mikrocaurulīšu pāris, kas starveidīgi ir savienots ar perifērajām mikrocaurulītēm.

Viciņas un skropstiņas
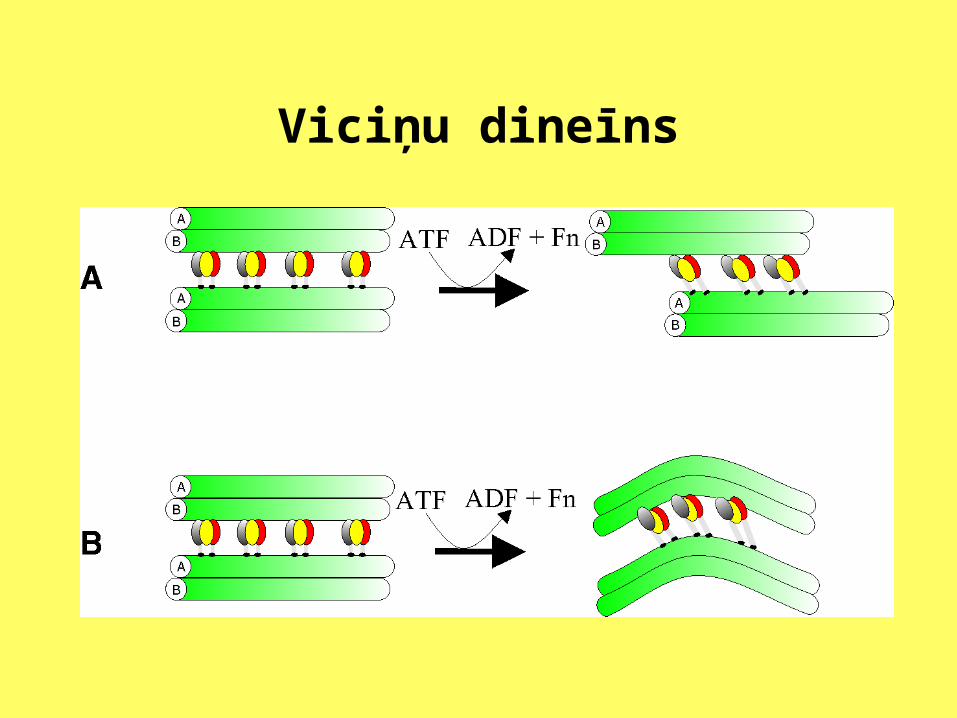
Viciņu dineīns

Mikrocaurulītēm piesaistītās olbaltumvielas
Pie tubulīna var pievienoties trīs olbaltumvielu grupas. Pirmo grupu veido mikrocaurulītēm pievienotās olbaltumvielas (MPO), tās reizēm sauc arī par strukturālajām MPO, jo tās stabilizē tubulīna molekulas, un var būt saistītas ar tām vairākos polimerizācijas un depolimerizācijas ciklos.
Otru grupu veido motorās olbaltumvielas. Tās sauc par motorajām olbaltumvielām tādēļ, ka, izmantojot ATF hidrolīzē iegūto enerģiju, tās slīd pa mikrocaurulītēm. Tās ļoti bieži ir cieši piesaistītas pie mikrocaurulītēm. Tomēr tās var vieglāk atdalīt kā pirmās grupas olbaltumvielas.

Mikrocaurulītēm piesaistītās olbaltumvielas
Trešo grupu veido hetorogēna olbaltumvielu grupa. Tās ir ciešāk piesaistītas mikrocaurulītēm nekā motorās olbaltumvielas. Tajās ietilpst glikolītiskie fermenti (aldolāze u.c.), kināzes (proteīnkināze A u.c.), olbaltumvielu biosintēzē izmantojamās olbaltumvielas (elongācijas faktors EF un pat ribosomas), olbaltumvielas, kas pievienojas membrānu receptoriem (dinamīns u.c.), ribonukleoproteīni. Šo molekulu mijiedarbības mehānismi ir nepilnīgi noskaidroti, tomēr tie atspoguļo citoskeleta lomu daudzu citoplazmas olbaltumvielu telpiskajā novietošanā.
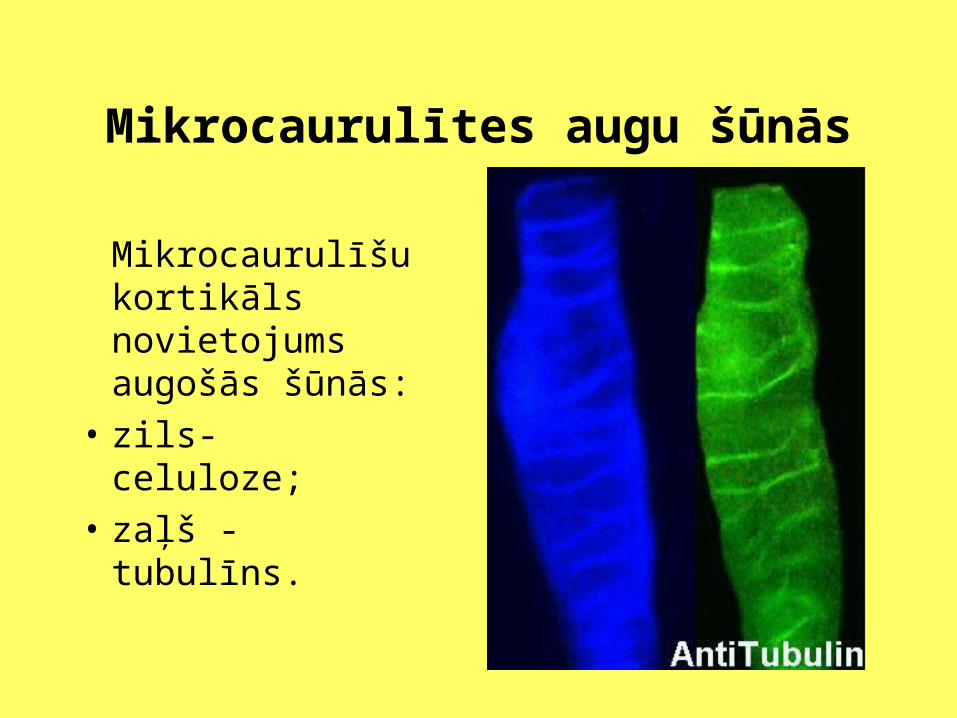
Mikrocaurulītes augu šūnās
Mikrocaurulīšu kortikāls novietojums augošās šūnās:
• zils- celuloze;
• zaļš - tubulīns.
http://www-lbcmcp.ups-tlse.fr/documents/videomitose.mov
Mikrocaurulīšu loma mitozē

Mikrofilamentus veido vissmalkākie olbaltumvielu pavedieni. To diametrs ir apmēram 8 nm. Vislabāk izpētīti ir aktīna mikrofilamenti. Aktīns ir plaši izplatīta olbaltumviela un sastopams visās eikariotiskajās šūnās. Īpaši lielos daudzumos (20% no kopējā šūnas olbaltumvielu daudzuma) aktīns ir atrodams muskuļu šūnās, kur tas rada muskuļu kontrakcijas.
Mikrofilamenti

Aktīns šūnās ir atrodams divos veidos: kā atsevišķas globulāras aktīna molekulas(G-aktīns) un fibrillu veidā kā polimerizētas aktīna molekulas (F-aktīns). Noteiktā zonā, pēc atslēgas slēdzenes principa, G-aktīna moleklai ir pievienota ATF molekula.
Mikrofilamenti
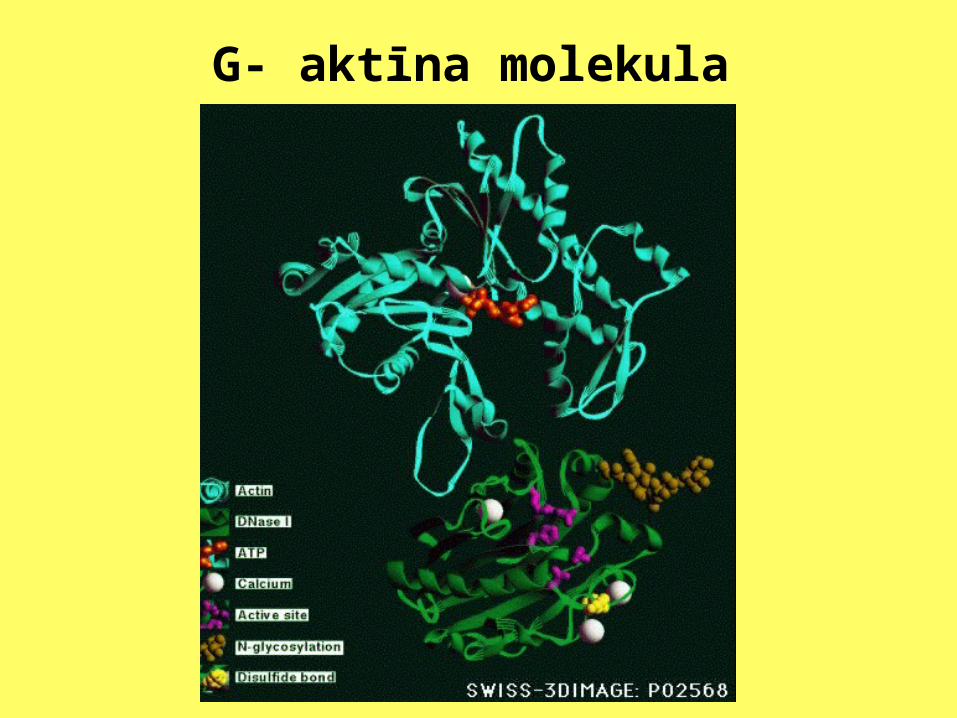
G- aktīna molekula
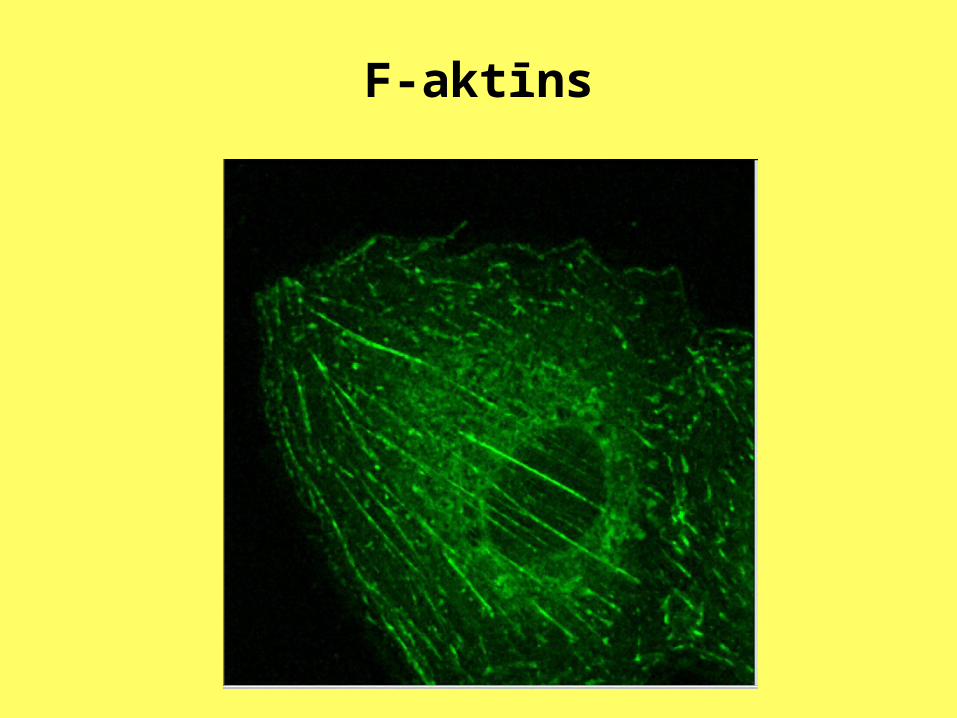
F-aktīns

Aktīna polimerizācija
• Jaunu aktīna fibrillu (F-aktīna) veidošanās procesu Jaunu aktīna fibrillu (F-aktīna) veidošanās procesu sauc par sauc par nukleācijunukleāciju. .
• Esoša aktīna filamenta pagarināšanos sauc par Esoša aktīna filamenta pagarināšanos sauc par elongācijuelongāciju. .
• F-aktīna molekulu kopējais garums daudzkārt F-aktīna molekulu kopējais garums daudzkārt pārsniedz šūnas garumu. pārsniedz šūnas garumu.

Aktīna polimerizācija 2
In vitroIn vitro apstākļos aktīna apstākļos aktīna polimerizācija ir atkarīga no polimerizācija ir atkarīga no globulārā aktīna globulārā aktīna koncentrācijas. Ja tā ir koncentrācijas. Ja tā ir lielāka par kritiskolielāka par kritisko - 0,1- 0,1M, M, tad var notikt polimerizācija. tad var notikt polimerizācija. Šajos apstākļos molekulu Šajos apstākļos molekulu polimerizācijai nav polimerizācijai nav nepieciešama ATF hidrolīze. nepieciešama ATF hidrolīze. ATF vietā var izmantot ATF vietā var izmantot sintētiskus analogus, kuri sintētiskus analogus, kuri nevar atdalīt fosfāta grupas.nevar atdalīt fosfāta grupas.

Aktīna polimerizācija
Filma
• Axis292c30/The cell
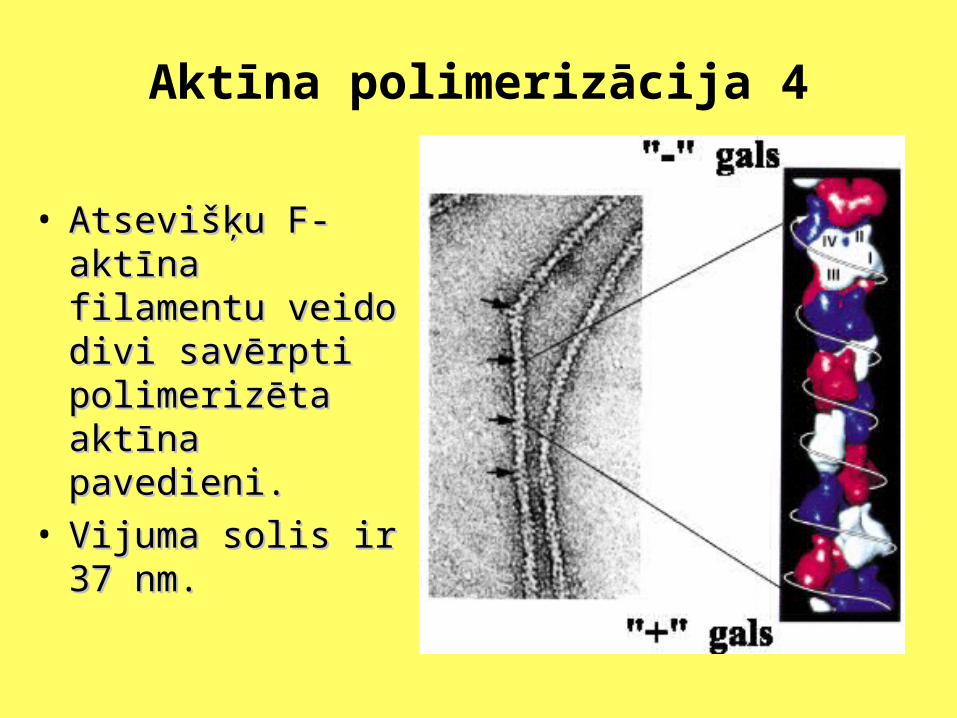
Aktīna polimerizācija 4
• Atsevišķu F-aktīna Atsevišķu F-aktīna filamentu veido divi filamentu veido divi savērpti polimerizēta savērpti polimerizēta aktīna pavedieni. aktīna pavedieni.
• Vijuma solis ir 37 nm.Vijuma solis ir 37 nm.

Aktīna polimerizācija 5
• F-aktīna filamentam izšķir plus galu un mīnus galu. Pie F-aktīna filamentam izšķir plus galu un mīnus galu. Pie plus gala aktīvāk pievienojas jaunas G-aktīna molekulas, plus gala aktīvāk pievienojas jaunas G-aktīna molekulas, bet no mīnus gala tās aktīvāk atdalās. Pie plus gala bet no mīnus gala tās aktīvāk atdalās. Pie plus gala pievienojas aktīna molekulas, kuras satur ATF. Mīnus galā pievienojas aktīna molekulas, kuras satur ATF. Mīnus galā ATF hidrolizējas un izveido ADF. Tas vājina saites starp ATF hidrolizējas un izveido ADF. Tas vājina saites starp galējo aktīna molekulu un filamentu un tā rezultātā galējo aktīna molekulu un filamentu un tā rezultātā molekula atdalās.molekula atdalās.
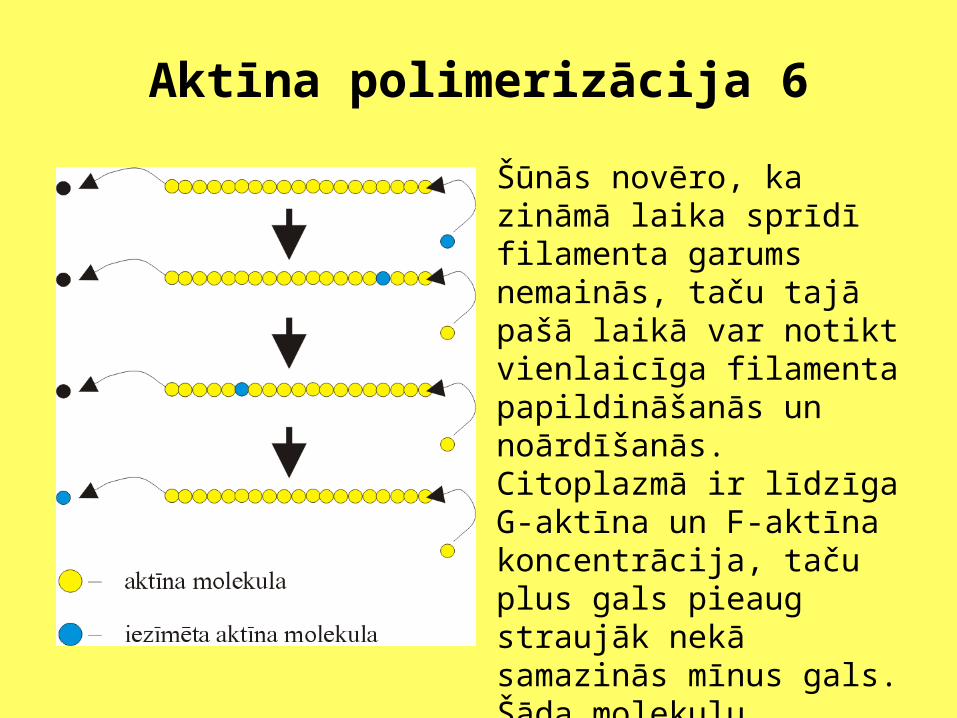
Aktīna polimerizācija 6
Šūnās novēro, ka zināmā laika sprīdī filamenta garums nemainās, taču tajā pašā laikā var notikt vienlaicīga filamenta papildināšanās un noārdīšanās. Citoplazmā ir līdzīga G-aktīna un F-aktīna koncentrācija, taču plus gals pieaug straujāk nekā samazinās mīnus gals. Šāda molekulu cirkulācija, filamenta mīnus galā, hidrolizējot ATF, patērē enerģiju.

Eksperimentos izmantojamās vielas
• CITOHALAZĪNS D (sēņu alkaloids) -neļauj mikrofilamenta “+” galā pievienot monomērus
• LATRUNKULĪNS - pievienojas G-aktīnam un neļauj šiem monomēriem pievienoties pie filamenta
• FALOIDĪNS - pievienojas G-aktīna monomēriem to savienojuma vietā un stabilizē mikrofilamentus. Izmanto arī kā aktīna specifisku krāsvielu.

Aktīnu pārraujošās un aktīnu pārklājošās olbaltumvielas 2
• Ar aktīna mikrofilamentiem saistās aktīnu pārraujošās un aktīnu pārklājošās olbaltumvielas. To darbību var pamanīt, vērojot citoplazmas konsistenci dažādās šūnas zonās. Šūnas centrā citoplazma ir šķidrāka (t.i. sola stāvoklī), bet šūnas perifērijā tā ir viskozāka (t.i. gēla stāvoklī). Turklāt šie stāvokļi dažādās šūnas zonās var strauji mainīties. Lielā mērā to nodrošina mikrofilamentu polimerizācija un depolimerizācija. Mikrofilamentu noārdīšanos nodrošina aktīnu pārraujošās olbaltumvielas.

Aktīnu pārraujošās olbaltumvielas
• Pie aktīnu pārraujošajām olbaltumvielām pieder gelozīns, severīns un villīns. Šo olbaltumvielu molekulmasa ir lielāka nekā tām olbaltumvielām, kas saistās ar globulāro aktīnu.
• Visas šīs olbaltumvielas piestiprinās kādā aktīna mikrofilamenta zonā un pārrauj saiti starp divām blakus stāvošām aktīna molekulām.

Aktīnu pārraujošās olbaltumvielasFilma
• Axis292c30/The cell

Aktīnu pārklājošās olbaltumvielas
• Savukārt no pastiprinātas noārdīšanas mikrofilamentus pasargā aktīnu pārklājošās olbaltumvielas. Tās biežāk ir sastopamas šūnās, kuru skeleta struktūra ir samērā stabila. Šķērssvītroto muskuļu šūnās pie aktīna mikrofilamentu + gala var pievienoties CapZ olbaltumvielas. Tās pārtrauc aktīna molekulu atdalīšanos. Tās arī ir ārkārtīgi stabilas un atdalās no mikrofilamenta apmēram pēc trīsdesmit dienām. Pie Aktīna mikrofilamentu stabilitāte palielinās, ja tiem ir pievienotas tropomiozīna molekulas.Tad to “-” galā var vēl piestiprināties tropomodulīna molekulas.

Mikrofilamentu novietojums šūnā
Eikariotu šūnās var novērot vairāku veidu aktīna mikrofilamentu kompleksus. Mikrofilamenti var būt sakārtoti kūlīšos vai veidot tīklu. Kūlīšos mikrofilamentiem ir paralēls vai antiparalēls novietojums. Savukārt tīklā mikrofilamenti ir daudz retāki un viens pret otru cenšas novietoties apmēram 900 leņķī. Izdala paralēlos kūlīšus, kontraktilos kūlīšus un gēlam līdzīgo tīklojumu.
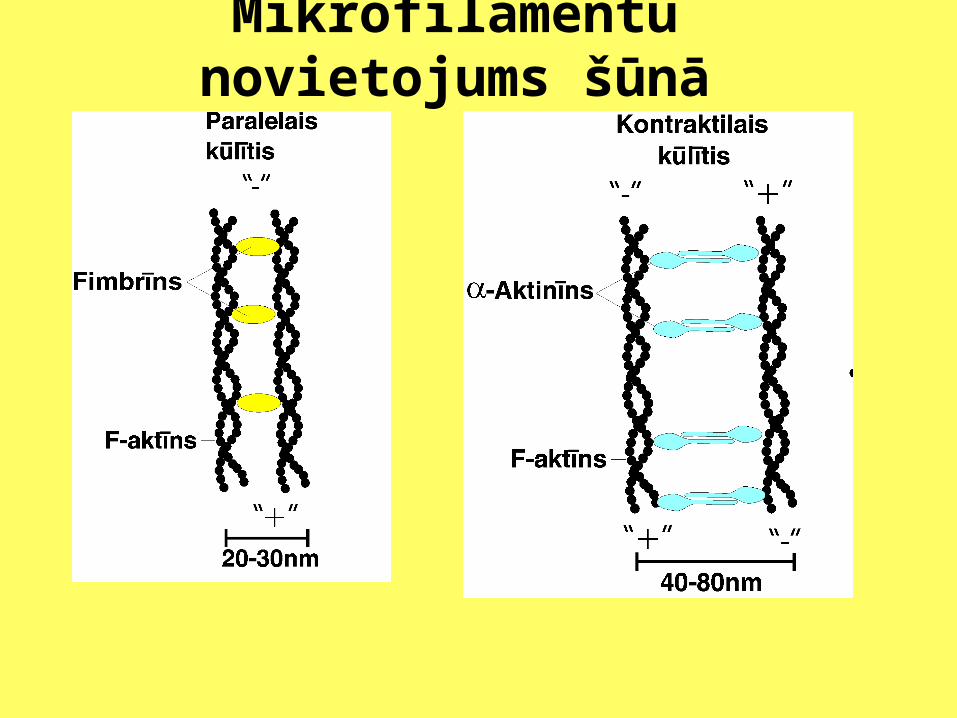
Mikrofilamentu novietojums šūnā
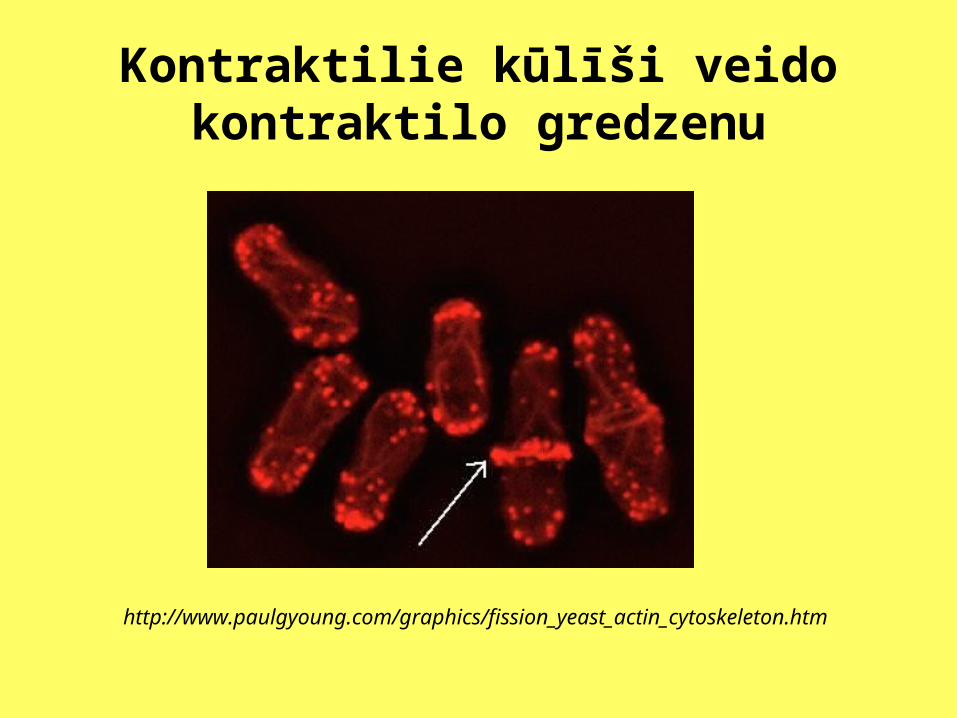
Kontraktilie kūlīši veido kontraktilo gredzenu
http://www.paulgyoung.com/graphics/fission_yeast_actin_cytoskeleton.htm
Kontraktilie kūlīši veido kontraktilo gredzenu
Filma
• Axis292c30/The cell
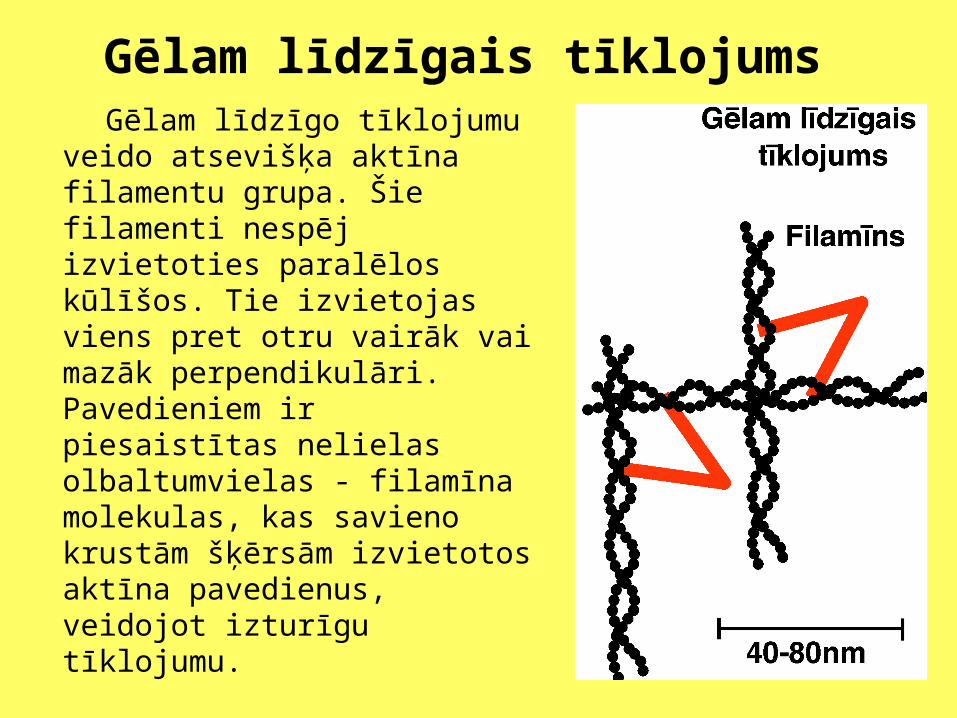
Gēlam līdzīgais tīklojums Gēlam līdzīgo tīklojumu veido
atsevišķa aktīna filamentu grupa. Šie filamenti nespēj izvietoties paralēlos kūlīšos. Tie izvietojas viens pret otru vairāk vai mazāk perpendikulāri. Pavedieniem ir piesaistītas nelielas olbaltumvielas - filamīna molekulas, kas savieno krustām šķērsām izvietotos aktīna pavedienus, veidojot izturīgu tīklojumu.
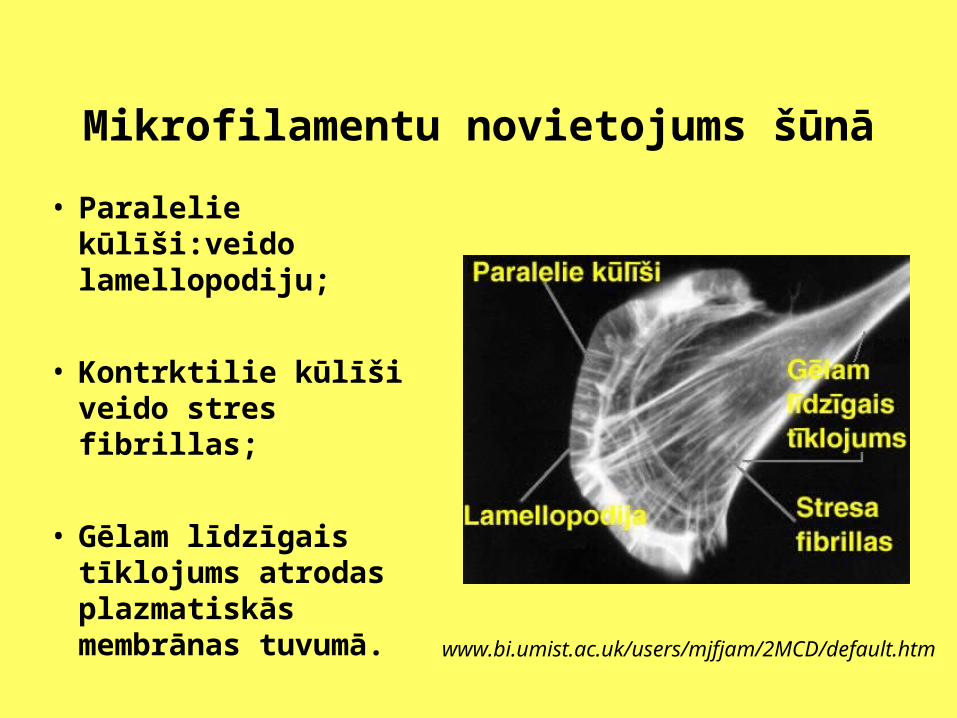
Mikrofilamentu novietojums šūnā
• Paralelie kūlīši:veido lamellopodiju;
• Kontrktilie kūlīši veido stres fibrillas;
• Gēlam līdzīgais tīklojums atrodas plazmatiskās membrānas tuvumā. www.bi.umist.ac.uk/users/mjfjam/2MCD/default.htm
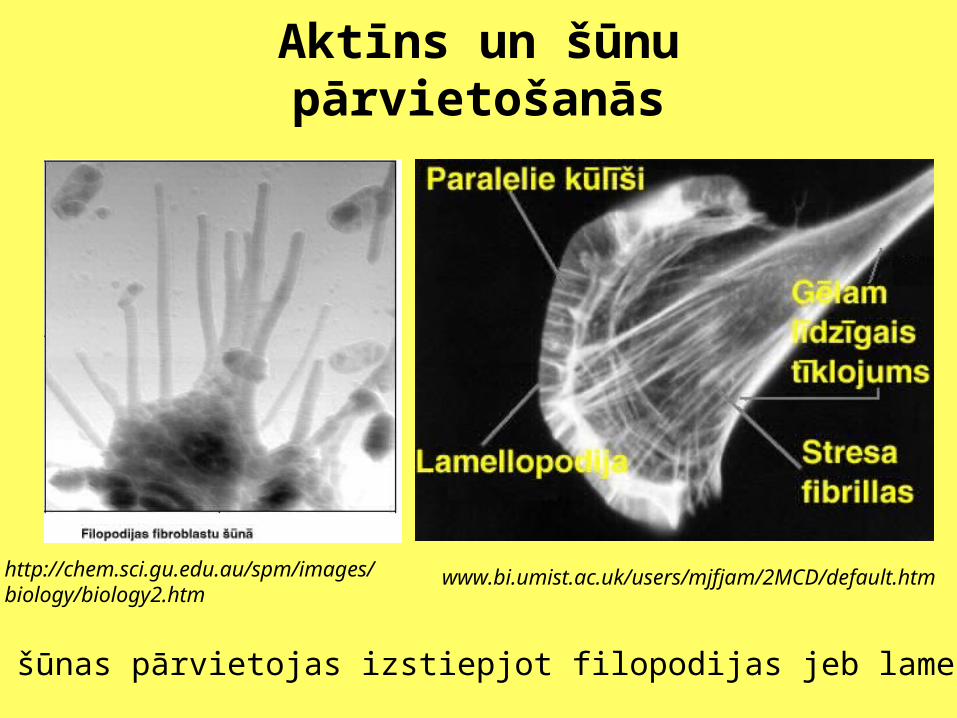
Aktīns un šūnu pārvietošanās
http://chem.sci.gu.edu.au/spm/images/biology/biology2.htm
www.bi.umist.ac.uk/users/mjfjam/2MCD/default.htm
Eikariotu šūnas pārvietojas izstiepjot filopodijas jeb lamelopodijas.
Pārvietošanās ar lamelopodiju palīdzību
http://www.bio.davidson.edu/misc/movies/fishlamellipodia.mov
Makrofāgu pārvietošanās
http://www.bio.davidson.edu/misc/movies/macwalk.mov
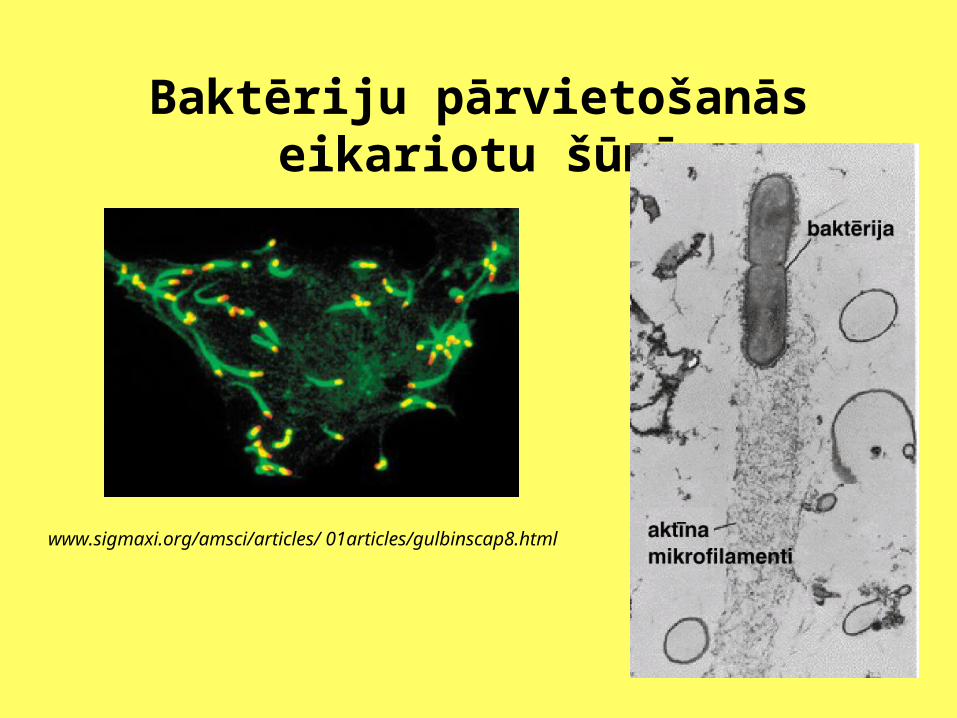
Baktēriju pārvietošanās eikariotu šūnā
www.sigmaxi.org/amsci/articles/ 01articles/gulbinscap8.html
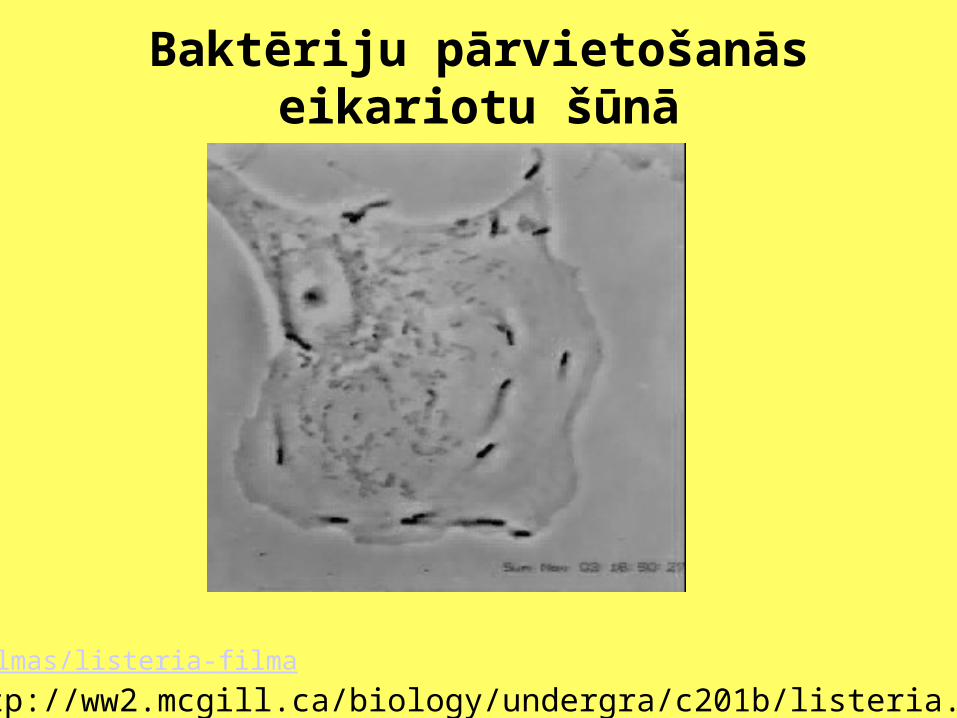
Baktēriju pārvietošanās eikariotu šūnā
/filmas/listeria-filma
http://ww2.mcgill.ca/biology/undergra/c201b/listeria.mov

Miozīns un aktīna mikrofilamentu loma vielu un organellu transportā
Mikrofilamenti un to motorās olbaltumvielas nodrošina aktīvus iekššūnas transporta procesus. Motorajām olbaltumvielām aminoterminālā daļa var piesaistīties pie aktīna mikrofilamentiem, bet blakus domēni piesaista un hidrolizē ATF. Molekulu karboksilterminālā daļa, atkarībā no tās uzbūves, pievienojas pie noteiktiem lipīdiem vai olbaltumvielām.
Pie mikrofilamentu motorajām olbaltumvielām pieder miozīna molekulas.
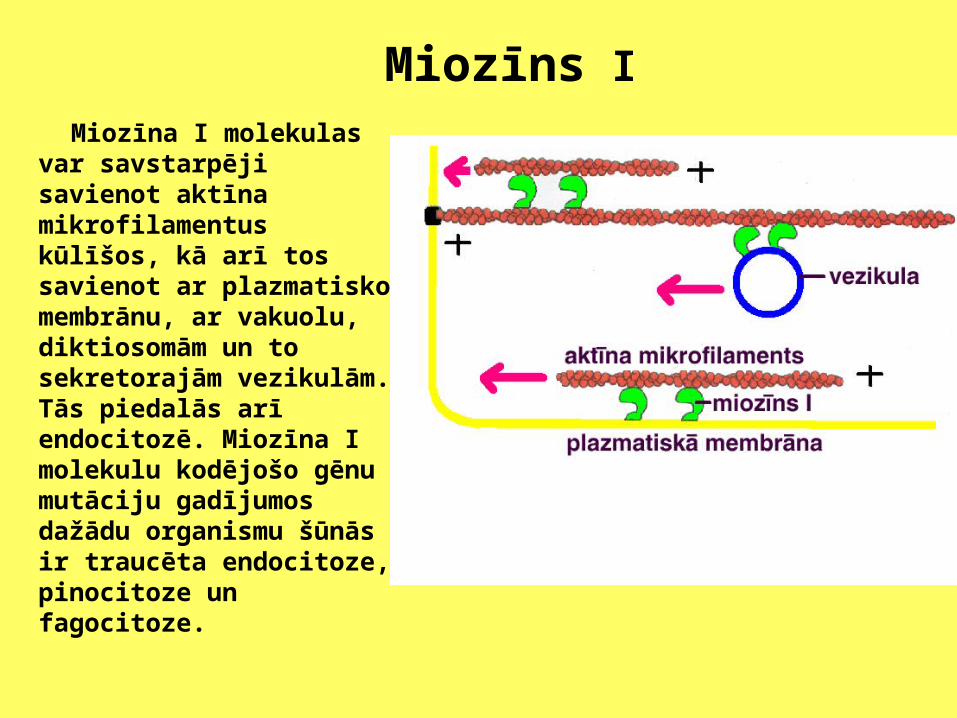
Miozīns I
Miozīna I molekulas var savstarpēji savienot aktīna mikrofilamentus kūlīšos, kā arī tos savienot ar plazmatisko membrānu, ar vakuolu, diktiosomām un to sekretorajām vezikulām. Tās piedalās arī endocitozē. Miozīna I molekulu kodējošo gēnu mutāciju gadījumos dažādu organismu šūnās ir traucēta endocitoze, pinocitoze un fagocitoze.

Miozīns II
Miozīns II nodrošina mugurkaulnieku muskuļu šūnu kontrakcijas, bet citos šūnu tipos - kontraktilā gredzena veidošanos un šūnu atdalīšanu citokinēzē.
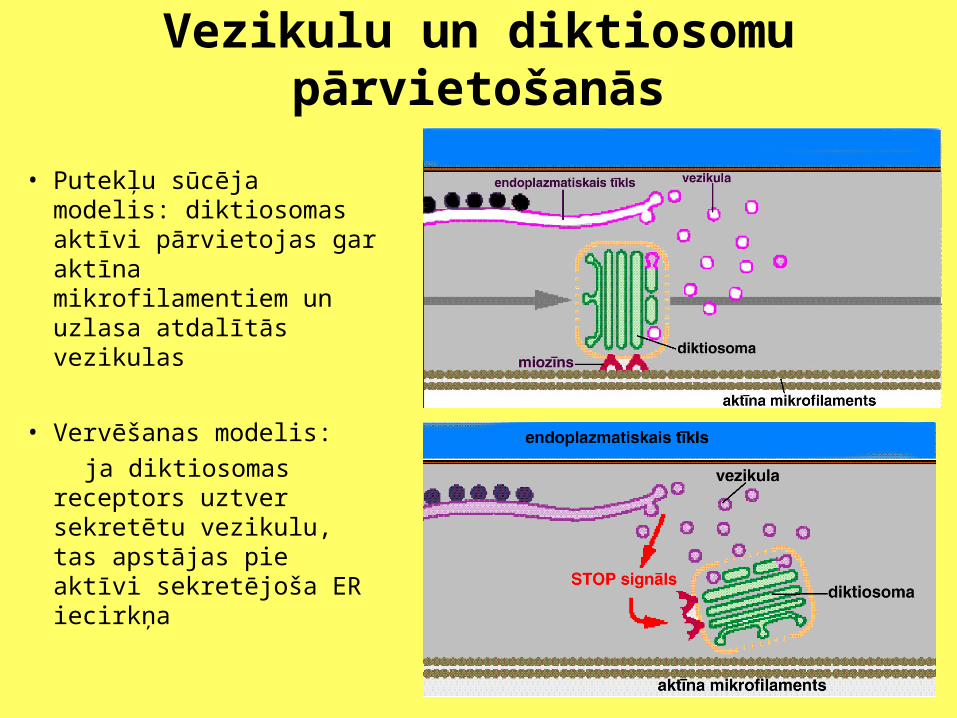
Vezikulu un diktiosomu pārvietošanās
• Putekļu sūcēja modelis: diktiosomas aktīvi pārvietojas gar aktīna mikrofilamentiem un uzlasa atdalītās vezikulas
• Vervēšanas modelis:
ja diktiosomas receptors uztver sekretētu vezikulu, tas apstājas pie aktīvi sekretējoša ER iecirkņa
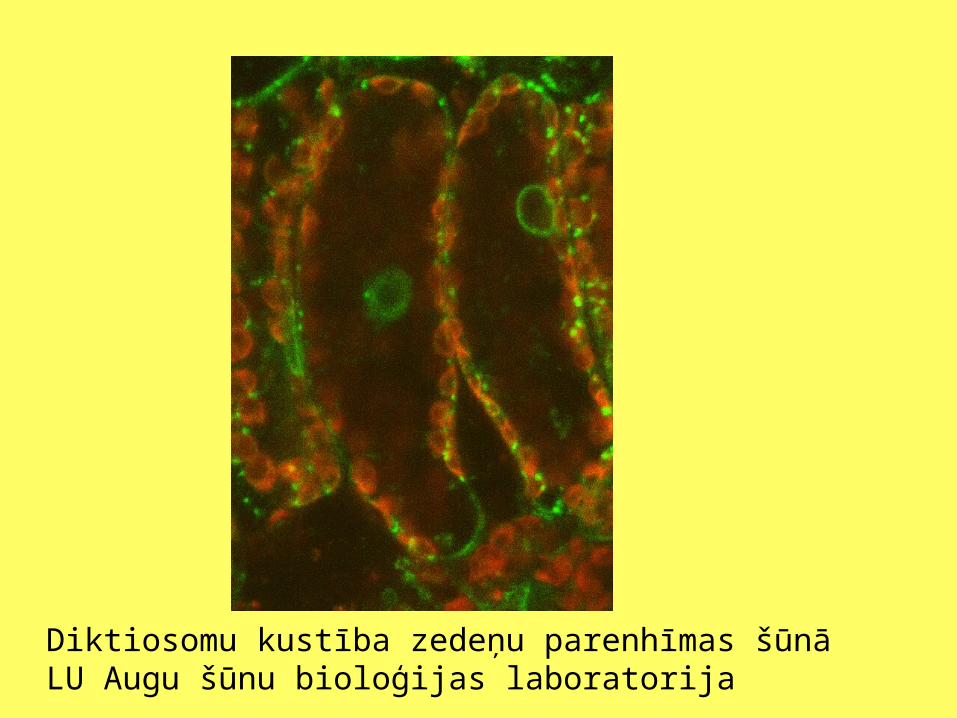
Diktiosomu kustība zedeņu parenhīmas šūnāLU Augu šūnu bioloģijas laboratorija

Citoskelets muskuļu šūnās
SR-sarkoplazmatiskais retikuls,M - mitohondriji,K-kodols
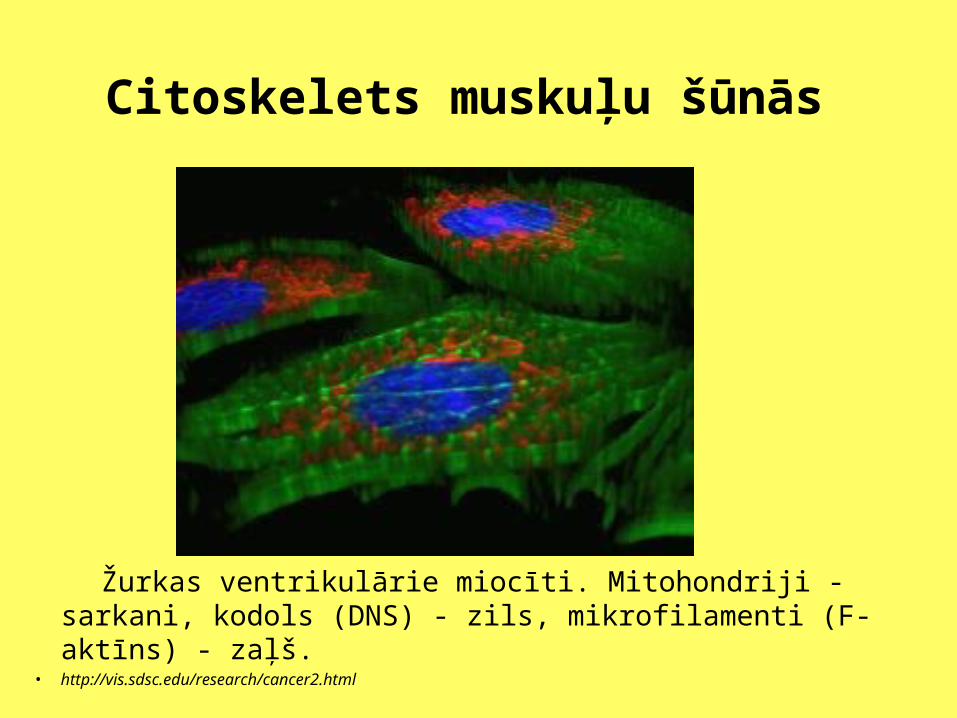
Citoskelets muskuļu šūnās
Žurkas ventrikulārie miocīti. Mitohondriji - sarkani, kodols (DNS) - zils, mikrofilamenti (F-aktīns) - zaļš.
• http://vis.sdsc.edu/research/cancer2.html
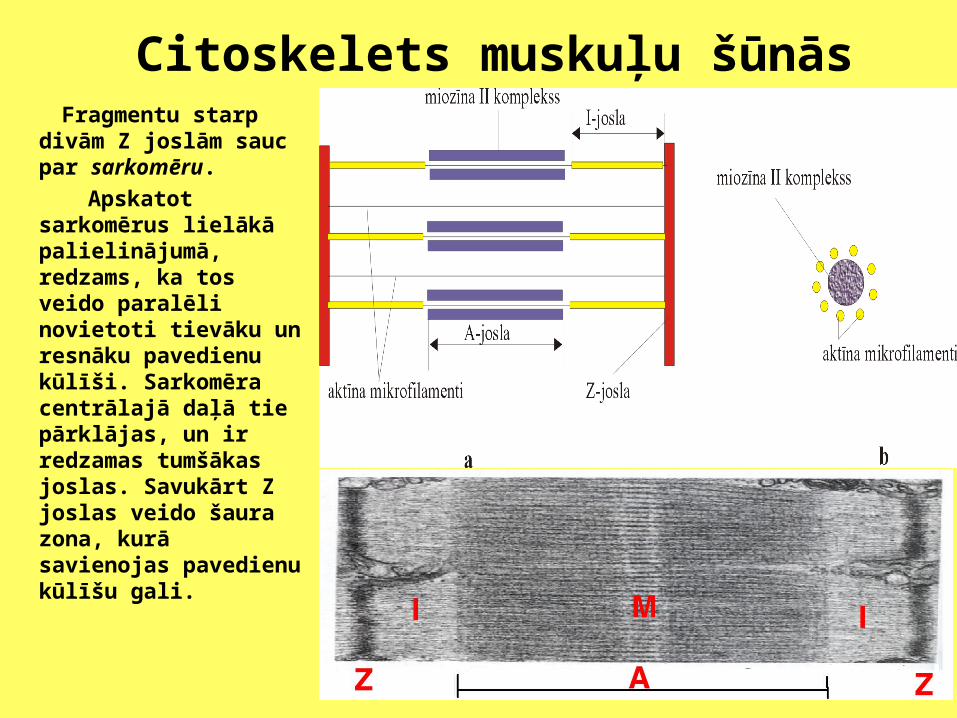
Citoskelets muskuļu šūnās Fragmentu starp divām
Z joslām sauc par sarkomēru.
Apskatot sarkomērus lielākā palielinājumā, redzams, ka tos veido paralēli novietoti tievāku un resnāku pavedienu kūlīši. Sarkomēra centrālajā daļā tie pārklājas, un ir redzamas tumšākas joslas. Savukārt Z joslas veido šaura zona, kurā savienojas pavedienu kūlīšu gali.

Tievie un resnie filamenti
Tievos pavedienus veido aktīna mikrofilamenti. Resnos pavedienus veido galvenokārt miozīna II molekulu kompleksi.
Mugurkaulniekiem miozīna II molekulu kompleksos apvienojas 300 - 400 miozīna molekulas.
Mugurkaulnieku gadījumā katrā resnā filamenta zonā ir deviņas paralēli novietotas miozīna molekulas.
Katrs resnais filaments pievieno vairākus tievos filamentus. To daudzums ir atkarīgs no organisma un muskuļu šūnu tipa.
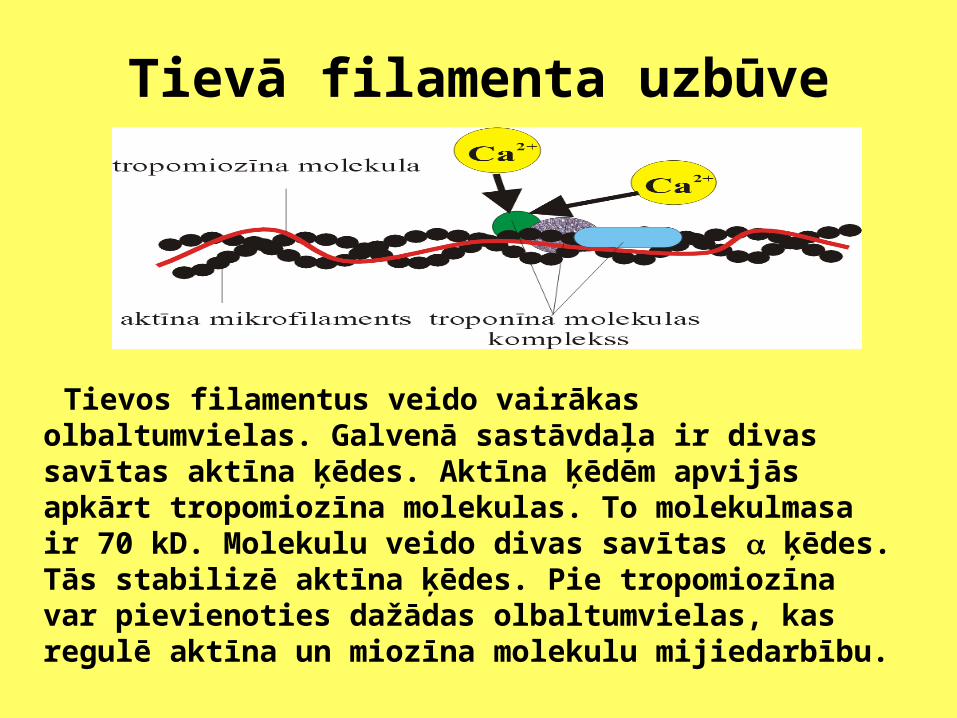
Tievā filamenta uzbūve
Tievos filamentus veido vairākas olbaltumvielas. Galvenā sastāvdaļa ir divas savītas aktīna ķēdes. Aktīna ķēdēm apvijās apkārt tropomiozīna molekulas. To molekulmasa ir 70 kD. Molekulu veido divas savītas ķēdes. Tās stabilizē aktīna ķēdes. Pie tropomiozīna var pievienoties dažādas olbaltumvielas, kas regulē aktīna un miozīna molekulu mijiedarbību.

Tievā filamenta uzbūve 2
Šķērssvītrotajos muskuļos pie tropomiozīna pievienojas troponīna molekulu komplekss. Gludajos muskuļos pievienojas kaldesmons. Savukārt citos šūnu tipos var pievienoties arī vinkulīns un citas olbaltumvielas, kas savieno tievos filamentus vai arī pievieno tos pie plazmatiskās membrānas vai starpfilamentiem.

Z-joslu uzbūve
Z-joslu veido aktīns, filamīns, sinemīns un Z-proteīns.
Z-josla pēc savas uzbūves ir diskveidīgs veidojums. Tajā pārklājas:tievo mikrofilamentu gali, Z-proteīni
aktīns nodrošina šo filamentu savienošanu.
Z-joslu stabilitāti palielina desmīns un vimentīns. Tie palīdz saistīt miofibrillas ar starpfilamentiem.
Starpfilamenti nodrošina miofibrillu paralēlu sakārtojumu šūnā un saista tos ar plazmatisko membrānu.

Muskuļu kontrakciju molekulārais mehānisms
• Muskuļu kontrakcijas nodrošina miozīna II molekulu pārslīdēšana pa aktīna mikrofilamentiem "+" gala virzienā.
• Katrs sarkomērs kontrakcijas rezultātā saīsinās par vairākiem nanometriem.
• Relaksācijas laikā miozīnu molekulu galvas daļa ir cieši pievienota aktīna molekulām.
• Kad pie miozīna molekulas pievienojas ATF, tad notiek neliela molekulas konformācijas maiņa. Tās rezultātā pavājinās saites starp miozīnu un aktīnu. Tāpēc miozīna molekulas galvas daļa pārslīd gar aktīna mikrofilamentu par apmēram 10 nm "+" gala virzienā.

Muskuļu kontrakciju molekulārais mehānisms
http://www.bio.davidson.edu/misc/movies/tropotropo.mov
Kontrakcija.
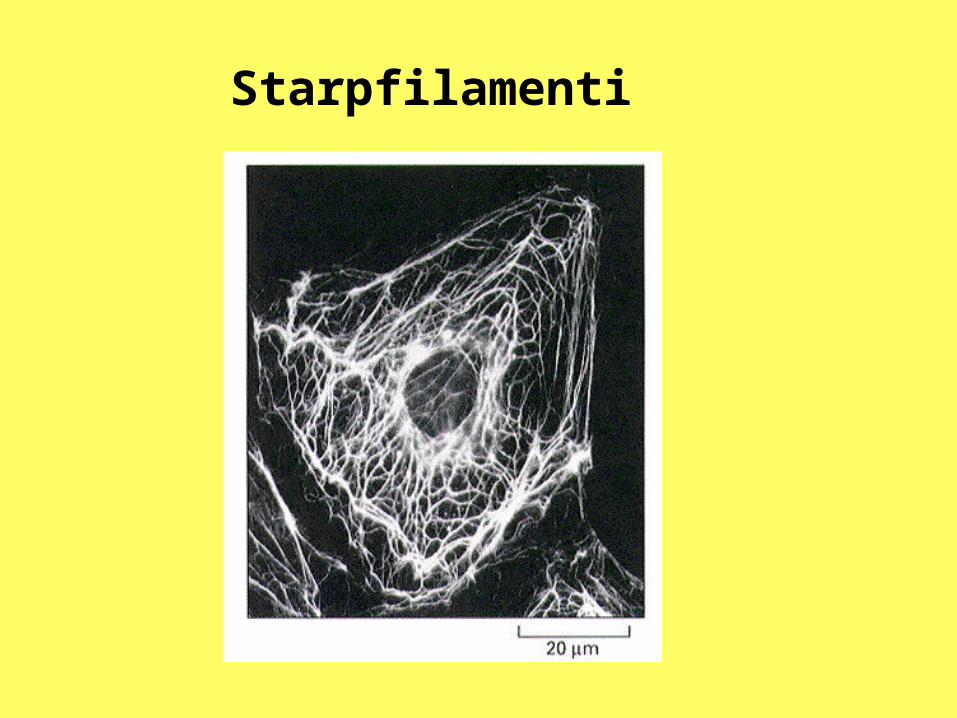
Starpfilamenti

Starpfilamenti
Starpfilamenti ir atrodami visos eikariotisko šūnu tipos. Tie ir ļoti daudzveidīgi pēc garuma un ķīmiskā sastāva. Pašreizējais nosaukums radies tāpēc, ka to diametrs ir mazāks kā mikrocaurulītēm, bet lielāks kā mikrofilamentiem, un tas ir apmēram 10 nm. No mikrofilamentiem un mikrocaurulītēm tie atšķiras arī ar to, ka šo elementu polimerizācijai nav nepieciešama GTF vai ATF hidrolīze.

Tie, atšķirībā no mikrofilamentiem un mikrocaurulītēm, ir arī stabilāki. Tieši tie šūnās pilda skeleta funkciju, saista savā starpā dažādus citoskeleta elementus, plazmatisko membrānu un lielāko daļu no šūnu organoīdiem.
Mugurkaulnieku šūnās var izšķirt piecas starpfilamentu grupas:
1) laminas,
2) vimentīns un tam līdzīgās olbaltumvielas,
3) bāziskie keratīni,
4) skābie keratīni un
5) neirofilamenti.

Keratīna starpfilamentiepitēlija šūnās
Epitēlija šūnās keratīna starpfilamenti pievienojas pie desmosomām un tādējādi savieno blakus novietotās šūnas.

Keratīna novietojums
Citokeratīni ietver kodolu un kodola tuvumā novietotos mitohondrijus. Tie saista arī ietvertās organellas ar šūnas perifēriju un plazmatisko membrānu.
http://froglab.biology.utah.edu/Cytoskeleton/Keratins/body_keratins.html

Kodola laminas
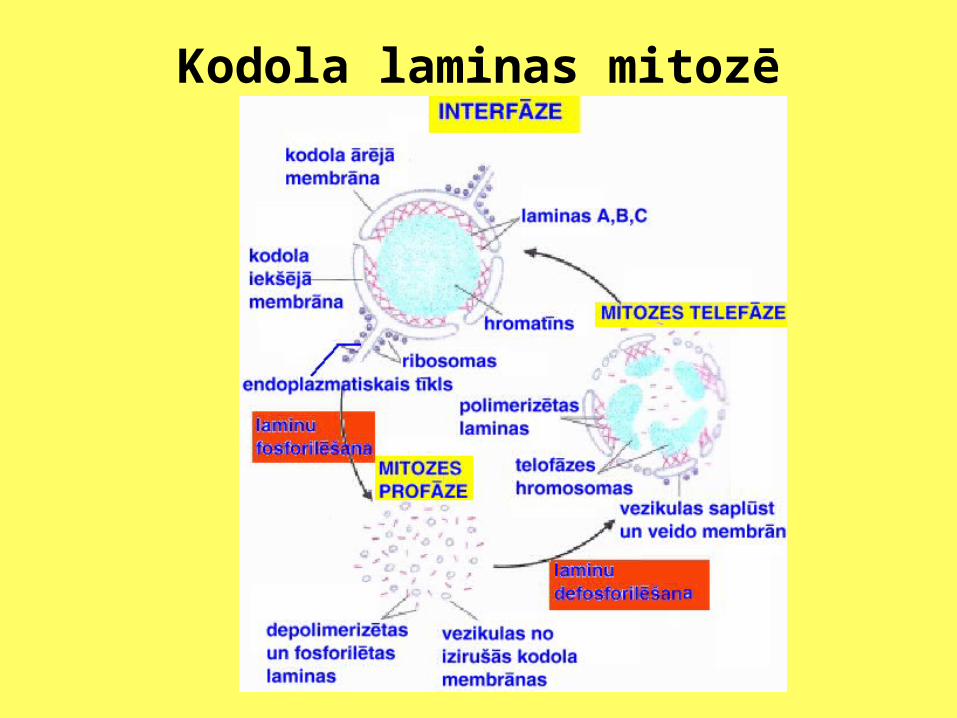
Kodola laminas mitozē
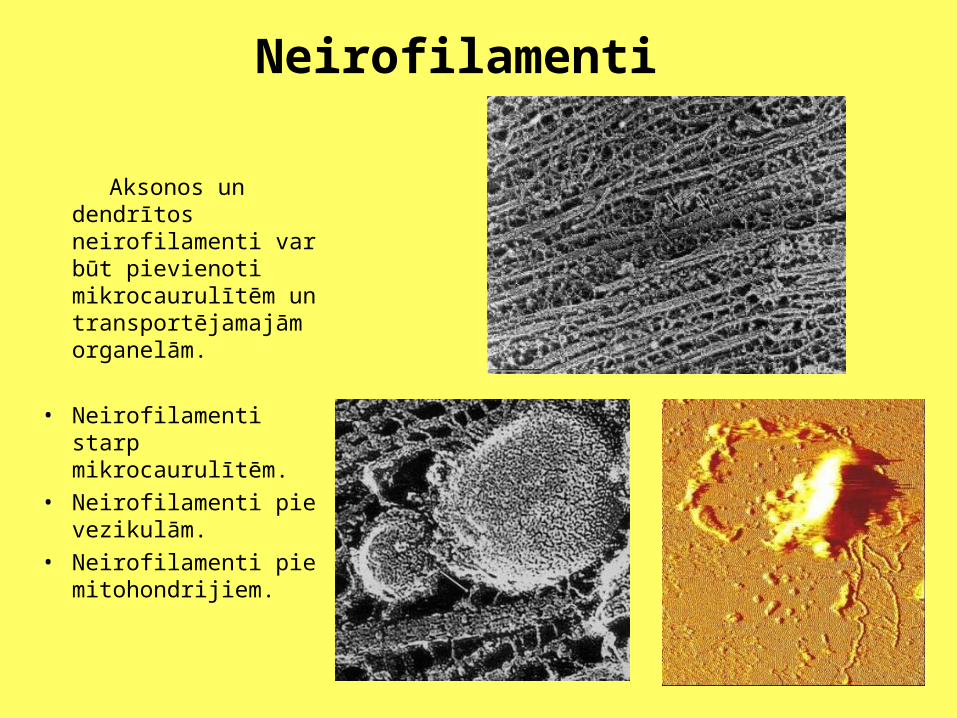
Neirofilamenti
Aksonos un dendrītos neirofilamenti var būt pievienoti mikrocaurulītēm un transportējamajām organelām.
• Neirofilamenti starp mikrocaurulītēm.
• Neirofilamenti pie vezikulām.
• Neirofilamenti pie mitohondrijiem.

Slimības
Centriolu skaita izmaiņas noved pie daudzpolu mitozes un nepareiza hromosomu sadalījuma. Aktīna mikrofilamentiem ievienoto olbaltumvielu mutāciju gadījumā tiek traucēts vezikulārais transports (eksocitoze un endocitozē) var veidoties muskuļu distrofija.Mikrocaurulīšu polimerizācijas traucējumu gadījumā ir traucējumi vielu transportā šūnā un dalīšanās vārpstas veidošanā.Mikrocaurulītēm pievienotās olbaltumvielas, piemēram tau bojājuma gadījumā parādās Alcheimera slimība.Starpfilamenti:keratīna starpfilamentu veidošanās traucējumi izraisa ādas epitēlija šūnu funkciju traucējumus.Neirofilamentu bojājumi izraisa traucējumus aksonu veidošanās procesā.Plektīna bojājumi izraisa muskuļu distrofiju.





























