Embed Size (px)
Citation preview

Capsular Polysaccharide of Erysipelothrix rhusiopathiae, the CausativeAgent of Swine Erysipelas, and Its Modification withPhosphorylcholine
Fang Shi,a Tomoyuki Harada,a,b Yohsuke Ogawa,a Hiroshi Ono,c Mayumi Ohnishi-Kameyama,c Toru Miyamoto,a Masahiro Eguchi,a
and Yoshihiro Shimojia,d
National Institute of Animal Health, Tsukuba, Ibaraki, Japana; Graduate School of Agriculture and Life Sciences, University of Tokyo, Bunnkyoku, Tokyo, Japanb; NationalFood Research Institute, Tsukuba, Ibaraki, Japanc; and Research Institute for Biological Sciences, Tokyo University of Sciences, Noda, Chiba, Japand
The capsule has been implicated in the virulence of the swine pathogen Erysipelothrix rhusiopathiae, a rod-shaped, intracellularGram-positive bacterium that has a unique phylogenetic position in the phylum Firmicutes and is a close relative of Mollicutes(mycoplasma species). In this study, we analyzed the genetic locus and composition of the capsular polysaccharide (CPS) of theFujisawa strain of E. rhusiopathiae. Genome analysis of the Fujisawa strain revealed that the genetic locus for capsular polysac-charide synthesis (cps) is located next to an lic operon, which is involved in the incorporation and expression of phosphorylcho-line (PCho). Reverse transcription-PCR analysis showed that cps and lic are transcribed as a single mRNA, indicating that theloci form an operon. Using the cell surface antigen-specific monoclonal antibody (MAb) ER21 as a probe, the capsular materialswere isolated from the Fujisawa strain by hot water extraction and treatment with DNase, RNase, pronase, and N-acetylmurami-dase SG, followed by anion-exchange and gel filtration chromatography. The materials were then analyzed by high-performanceliquid chromatography, mass spectrometry, and nuclear magnetic resonance (NMR) spectroscopy. The CPS of E. rhusiopathiaeis heterogeneous and consists of the major monosaccharides galacturonic acid, galactose, mannose, glucose, arabinose, xylose,and N-acetylglucosamine and some minor monosaccharides containing ribose, rhamnose, and N-acetylgalactosamine. In addi-tion, the capsule is modified by PCho, which comigrates with the capsular materials, as determined by Western immunoblotting,and colocalizes on the cell surface, as determined by immunogold electron microscopy. Virulence testing of PCho-defective mu-tants in mice demonstrated that PCho is critical for the virulence of this organism.
Capsules are important virulence determinants on the cell sur-faces of many bacterial pathogens, and they play a critical role
in protection against innate host defenses during infection. Somepathogens have evolved several mechanisms to evade host im-mune responses. In mucosal pathogens, including Haemophilusinfluenzae (33), Pseudomonas aeruginosa (32), Neisseria species(5), and some serotypes of Streptococcus pneumoniae (12, 31),phosphorylcholine (PCho) is a surface component of lipopolysac-charide (LPS) or capsular polysaccharides (CPSs), and this surfacemodification plays an important role in the pathogenesis of infec-tion.
Erysipelothrix rhusiopathiae is a Gram-positive bacterium thatrepresents a new class, Erysipelotrichia, in the phylum Firmicutesand is a facultative intracellular pathogen that causes a variety ofdiseases in many species of birds and mammals, including hu-mans. In pigs, E. rhusiopathiae can cause swine erysipelas, whichmay occur as acute septicemia or chronic endocarditis and poly-arthritis (34). E. rhusiopathiae expresses a capsule (29), whichplays an important role in the resistance of the organism to phago-cytosis by polymorphonuclear leukocytes and intracellular killingby macrophages (24, 28). Whereas the importance of the capsulein virulence has long been recognized, the biochemical featuresand mechanisms by which the capsule prevents the recognition ofE. rhusiopathiae by phagocytes or the immune system remainpoorly understood.
Recently, we sequenced the genome of E. rhusiopathiae andreported that E. rhusiopathiae is phylogenetically related to Molli-cutes (Mycoplasma species) (17). Based on genomic phylogenetictrees, we proposed that E. rhusiopathiae and other Erysipelotrichia
strains may be separate from Firmicutes and classified as a distinctphylum (17). Furthermore, our genome analyses revealed that E.rhusiopathiae lacks many orthologous genes for the biosynthesisof wall teichoic acids (WTA) and lipoteichoic acids (LTA). In ad-dition, E. rhusiopathiae lacks the dltABCD operon, which is re-sponsible for D-alanine incorporation into WTA and LTA, sug-gesting that the organism may have an atypical cell wall (17). Thus,the phylogenetic position of E. rhusiopathiae is unique amongGram-positive pathogens, highlighting the need for detailed stud-ies of E. rhusiopathiae cell surface antigens and cell wall structure.In the present study, we genetically and immunochemically ana-lyzed the capsule of E. rhusiopathiae.
MATERIALS AND METHODSStrain and growth conditions. The wild-type E. rhusiopathiae strain Fu-jisawa (serotype 1a), which was originally isolated from a septicemic pig(29), a capsule-defective nonreverting mutant, YS-1, which was previ-ously defined as an acapsular mutant of Fujisawa (26), and two PCho-defective insertional mutants of Fujisawa, clones 100 and 112, were cul-
Received 13 June 2012 Returned for modification 16 July 2012Accepted 28 August 2012
Published ahead of print 4 September 2012
Editor: J. F. Urban, Jr.
Address correspondence to Yoshihiro Shimoji, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.00635-12
November 2012 Volume 80 Number 11 Infection and Immunity p. 3993–4003 iai.asm.org 3993
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

tured in brain heart infusion (BHI; Difco Laboratories, Detroit, MI)medium supplemented with 0.1% Tween 80 (pH 8.0) (BHI-T80) at 37°C.
MAbs. The monoclonal antibodies (MAbs) ER21 and TEPC-15 (Sig-ma-Aldrich) were used to detect capsular material(s) and PCho, respec-tively. MAb ER21, which was prepared from a hybridoma generated frommice that had been immunized with irradiated E. rhusiopathiae bacteria,specifically agglutinates intact E. rhusiopathiae strains and recognizes thesurface antigen(s) (25).
RT-PCR. For reverse transcription-PCR (RT-PCR), total RNA fromthe Fujisawa strain was prepared with the RNeasy minikit (Qiagen, Va-lencia, CA) based on the protocol supplied. The RNA was further treatedwith RNase-free cloned DNase I (TaKaRa, Shiga, Japan) to remove DNAcontaminants and quantified with a spectrophotometer (DU series 500;Beckman, Fullerton, CA). One microgram of total RNA in 20 �l reactionsolution was converted to cDNA with Verso reverse transcriptase(Thermo Scientific Verso cDNA synthesis kit; ABgene, Surrey, UnitedKingdom) at 42°C for 55 min followed by 95°C for 2 min. cDNA wasamplified with the primers shown in Table 1, and the amplicons wereseparated on a 1% agarose gel. The corresponding controls lacking reversetranscriptase or template RNA were included for each reaction to confirmthe absence of genomic DNA.
Extraction and purification of CPS. Bacteria were grown at 37°C inBHI-T80 and harvested at the end of logarithmic growth by centrifuga-tion. The capsular polysaccharide (CPS) was then extracted with 65°C hotwater for 20 min with constant stirring. The extracts were subjected totreatment with DNase (116 U/ml; Invitrogen, Carlsbad, CA), RNase (500�g/ml; Invitrogen), pronase (1 mg/ml; Roche, Mannheim, Germany),and N-acetylmuramidase SG (66 U/ml; Seikagaku Biobusiness, Tokyo,Japan), followed by trichloroacetic acid (TCA) precipitation. The super-natant was neutralized with Trizma base, concentrated by ultrafiltration,and further subjected to methanol and chloroform extraction (1:1:0.5methanol-chloroform-H2O) (2) to remove possible lipophilic compo-nents of the cytoplasmic membrane. The crude CPS was exhaustivelydialyzed against H2O (cellulose tube; molecular weight cutoff [MWCO],14,000). The CPS was further dialyzed against an equilibration buffer of10 mM Tris-HCl (pH 7.5) containing 50 mM NaCl. The concentratedcrude CPS was applied to a 2.5- by 12-cm DEAE-cellulose anion-exchangechromatography column (DE-52; Whatman), and fractions were elutedwith a linear gradient of 50 mM to 500 mM NaCl in Tris-HCl (pH 7.5).Five-milliliter fractions were collected, and the CPS-containing fractionswere pooled and further fractionated by gel filtration chromatography(GFC) on a 60- by 16-cm Sephacryl S-200 column (GE Healthcare Bio-Sciences AB, Uppsala, Sweden). The column was equilibrated and elutedwith 150 mM NaCl in 10 mM Tris-HCl (pH 7.5). Each fraction was as-
sayed with phenol sulfuric acid to determine the total amount of carbo-hydrates (10) and by dot immunoblotting to detect CPS with reactivityagainst MAb ER21. The approximate molecular weight of the CPS wasestimated by comparing the elution volume of the CPS in GFC with thevoid volume, which was determined with dextran blue 2000 (GE Health-care, Uppsala, Sweden).
Peroxidate oxidation of CPS. CPS was oxidized with metaperiodateas previously described (4). Briefly, the CPS (400 ng) from each fractioneluted from GFC was dried in vacuo and resuspended in 10 �l 50 mMsodium acetate (pH 4.5) containing 20 mM metaperiodate. The CPS wasthen incubated in the dark with gentle shaking at room temperature for 1h. The CPS was dissolved in 10 �l 50 mM sodium acetate (pH 4.5) withoutmetaperiodate as a control.
Phospholipase C treatment of CPS. The CPS was treated with phos-pholipase C, as described previously (16), with the following modifica-tions. CPS (168 ng) dried in vacuo was dissolved in 0.5% deoxycholic acid(pH 8.0), followed by the addition of phospholipase C (1 U; Sigma-Al-drich, St. Louis, MO) dissolved in 10 mM 3,3-dimethylglutaric acid-NaOH with 0.1% bovine serum albumin (BSA). The solution was incu-bated at 37°C for 30 min, and the reaction was stopped with the additionof 1 M Tris-HCl containing 0.2% SDS. The corresponding control reac-tions were performed in the absence of CPS or phospholipase C. Thereaction mixture was separated by SDS-PAGE and subjected to Westernimmunoblotting.
Dot and Western immunoblotting. For dot immunoblotting, theCPS-containing solution or bacterial culture was spotted onto a nylonmembrane (MagnaGraph; Funakoshi, Tokyo, Japan). For Western im-munoblotting, the CPS-containing solution was separated by 15% SDS-PAGE and transferred to a polyvinylidene difluoride (PVDF) membrane(Immobilon-P; Millipore, Bedford, MA). The air-dried nylon membraneor PVDF membrane was blocked with 1% skim milk in phosphate-buff-ered saline (PBS) containing 0.05% Tween 20 and incubated with MAbER21 or TEPC-15. The membranes were further treated with horseradishperoxidase conjugated with goat anti-mouse immunoglobulin antibody(IgG, IgM, and IgA; H�L) (Zymed Laboratories, San Francisco, CA). Theblots were developed with 3,3-diaminobenzidine tetrahydrochloride– hy-drogen peroxide (Wako, Tokyo, Japan).
Analytical procedures. (i) HPLC. The monosaccharide componentsof the CPS from Fujisawa were released and labeled with p-aminobenzoicacid ethyl ester (ABEE) according to the manufacturer’s instructions forthe ABEE labeling kit Plus S (J-Oil Mills Co., Ltd., Tokyo, Japan). Briefly,the dried CPS (5 �g) was successively subjected to neuraminidase treat-ment, acid hydrolysis with 4 M trifluoroacetic acid (TFA), and N-acetyla-tion. After the solvent was evaporated in vacuo, 10 �l distilled H2O and 40�l ABEE solution were added into the tube, and the mixture was incu-bated at 80°C for 1 h for labeling. The solution was cooled to room tem-perature, and 200 �l distilled H2O and 200 �l chloroform were added andmixed vigorously. The aqueous phase was collected and used for high-performance liquid chromatography (HPLC) analysis. The ABEE-labeledmonosaccharides were analyzed on a Honenpak C18 column (75 by 4.6mm) (Seikagaku Biobusiness) on a Prominence HPLC instrument (Shi-madzu Corp., Kyoto, Japan) according to the method previously de-scribed (35). A 20-�l aliquot of ABEE-labeled monosaccharide was in-jected into the column and separated with 7% acetonitrile in 0.2 Mpotassium borate buffer (pH 8.9) over 50 min; the column was thenwashed with 50% acetonitrile in 0.02% TFA for 5 min at a flow rate of 1.0ml/min at 30°C. The elution profile was monitored by fluorescence (ex-citation of 305 nm and emission of 360 nm) with an RF-10A detector(Shimadzu Corp.). The monosaccharide constituents of CPS were iden-tified based on the retention time of each monosaccharide by comparisonwith the 11 standard monosaccharides supplied in the ABEE labeling kit,as well as D-galacturonic acid monohydrate (Fluka) and D-glucuronic acid(Sigma-Aldrich).
(ii) MALDI-TOF MS. A 0.5-�l aliquot of native (45 �g/ml) or par-tially hydrolyzed CPS (2% acetic acid, 100°C, 2.5 h) was mixed with 0.5 �l
TABLE 1 Primers used in RT–PCR
Primer Sequence Product size (bp)
1F 5=-CAA CCT TGC GAA CTG AAA CA-3= 2,5941R 5=-GGA GTT GGC TTC GTG CTT AG-3=2F 5=-AGC ATG ATG ACC GAA TCA CA-3= 3,2302R 5=-TTG CTT GAT ACC ATG CCT CA-3=3F 5=-AGG GAT GCA TCG TCA CAG TA-3= 3,4133R 5=-GGC GTA TCA GGA TCA AAT GG-3=4F 5=-CTT GCG TAT GAC CGC AAT AA-3= 3,4894R 5=-CTC TGC ACT GTC CGT TCG TA-3=5F 5=-TGA TCT CGC AAA CGG ACT AA-3= 1,9355R 5=-AGC CAG CAG TTC TCC CAG TA-3=6F 5=-CGG AGG GTT TTA TGT TTT CG-3= 3,4886R 5=-CGT GGA TTT TCG GAA GAA AG-3=7F 5=-GG CAT CCA CGA TGT TCT TA-3= 2,7027R 5=-GTG CAA GTG CTT TGT TTC CA-3=8F 5=-ACG CTA CAC ACG CTC TAG CA-3= 2,8078R 5=-AAC GGA AGG ATC AAC TGT GG-3=
Shi et al.
3994 iai.asm.org Infection and Immunity
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

�-cyano-4-hydroxycinnamic acid (CHCA) (Bruker Daltonics, Billerica,MA) saturated in a mixture of acetonitrile and 0.1% TFA (1:1). The solu-tion was spotted onto a stainless steel matrix-assisted laser desorptionionization (MALDI) mass spectrometry (MS) target and air dried. Massspectra were recorded on a 4800 plus MALDI-tandem time of flight (TOF/TOF) analyzer (AB SCIEX, Foster City, CA) in both the positive- andnegative-ion modes. The mass spectrometer was tuned and calibratedwith standard peptides or proteins (Bruker Daltonics) prior to measure-ments.
(iii) NMR spectroscopy. Nuclear magnetic resonance (NMR) spec-troscopy was performed as described previously (23). The lyophilizedpolysaccharide (200 �g) was dissolved in 0.25 ml deuterium oxide(99.96% D) and transferred into a 5-mm NMR tube (Shigemi, Tokyo,Japan). One-dimensional 1H and distortionless enhancement by polariza-tion transfer at 135° (DEPT-135) 13C, as well as two-dimensional doublequantum filtered-correlated spectroscopy (DQF-COSY), triple quantumfiltered (TQF)-COSY, total correlation spectroscopy (TOCSY), nuclearOverhauser effect spectroscopy (NOESY), 1H-13C heteronuclear singlequantum coherence (HSQC), 1H-13C heteronuclear multiple bond cor-relation (HMBC), 1H-15N HMBC, and 1H-13C HSQC-TOCSY were mea-sured on a Bruker Avance 800 spectrometer (Bruker Biospin,Rheinstetten, Germany) equipped with a TCI cryoprobe (1H-13C/15N).For two-dimensional 1H-31P HMBC, a Bruker DRX 600 spectrometer(Bruker Biospin) with a QXI probe (1H-13C/15N/31P) was used. Bothspectrometers were run with XWIN-NMR software. All NMR experi-ments were performed at a probe temperature of 25°C. The chemicalshifts are expressed as �-values relative to the internal standard sodium2,2-dimethyl-2-silapentane-5-sulfonate (DSS).
Transposon mutagenesis. PCho-defective transposon Fujisawa mu-tants were generated by a mariner-based transposition system, as previ-ously described (3). Briefly, pMC plasmids were transferred into strainFujisawa by electroporation according to the previously describedmethod (27). Transformants were selected at 30°C on BHI-T80 platessupplemented with 1 �g/ml erythromycin. Individual colonies weregrown in BHI-T80 with erythromycin at 30°C. The cultures were dilutedand plated on BHI-T80 containing erythromycin at 30°C and then shiftedto 40°C to eliminate the thermosensitive pMC plasmids. Erythromycin-resistant (Emr) clones were picked and passaged on BHI-T80 plates sup-
plemented with erythromycin. The transposon insertion site was deter-mined by sequencing the transposon-flanking DNA regions oftransformants that contained a single transposon insertion. The mutantsof interest were analyzed for CPS and PCho production, and their viru-lence was further tested.
Effects of gene inactivation at the cps-lic locus. Effects of gene inac-tivation at the cps-lic chromosomal region on CPS and PCho expressionwere examined. For electron microscope studies, strain Fujisawa, YS-1,clone 100, and clone 112 were cultured in BHI-T80 medium at 37°Covernight, harvested by centrifugation, and resuspended in a PBS solutioncontaining 5% fetal calf serum (FCS). For metaperiodate treatment, Fu-jisawa cells were washed once with 50 mM sodium acetate (pH 4.5) andincubated with 50 mM sodium acetate (pH 4.5) containing 20 mM meta-periodate in the dark with gentle shaking at room temperature for 1 h. Thebacteria were placed on Formvar-coated nickel grids (400 mesh; Nisshin,Tokyo, Japan) for 2 min. After being washed with a PBS solution contain-ing 10% FCS, the grids were incubated with MAbs ER21 and TEPC-15,followed by incubation with 10-nm-diameter colloidal gold-conjugatedgoat anti-mouse immunoglobulin (IgG, IgM, and IgA) (BB International,Cardiff, United Kingdom). The grids were briefly stained with 1% ammo-nium molybdate and air dried after successive washing with PBS contain-ing 10% FCS and 1% ammonium acetate solution. The dried grids wereobserved by a transmission electron microscopy (H-7500; Hitachi, Ja-pan). Samples treated with secondary antibody alone were included ascontrols. For morphological analysis with light microscopy, Fujisawa,YS-1, and clones 100 and 112 were cultured at 37°C in BHI-T80 overnight,and aliquots of each culture were smeared on a glass slide. After air dryingand heat fixation, the slides were Gram stained using the Favor G Gramstain kit (Nissui Pharmacological, Tokyo, Japan) and viewed under a100� objective lens with a Leica DM1000 microscope. Images were cap-tured with LAS EZ software.
The effect of gene inactivation at the cps-lic region on CPS and PChoexpression was further examined by dot immunoblotting and Westernblotting. The Fujisawa and YS-1 strains and clones 100 and 112 werecultured in 500 ml of BHI-T80 at 37°C overnight. For dot immunoblot-ting, 3 �l of the overnight cultures was spotted onto a nylon membrane.Subsequently, 500-ml cultures of the strains were pelleted by centrifuga-tion, and the pellets were washed with ice-cold PBS and suspended in 15
FIG 1 (A) Schematic representation of the cps-lic region of E. rhusiopathiae. The arrows indicate the orientation and corresponding locations of the primer pairsused (Table 1) in the RT-PCR assay. The black arrowhead denotes the transposon insertion site. (B) RT-PCR analysis of the E. rhusiopathiae cps-lic cluster. Thenumbers above the lanes correspond to the Arabic numerals of the primer names. All lanes labeled with a represent the analysis of PCRs performed with cDNA.All lanes labeled with b represent the corresponding control reactions with RNA extracts not subjected to reverse transcription. All lanes labeled with c showcontrol reactions with mixtures not containing template. Lane 1a represents a control reaction with primers 1F and 1R, which were both designed to amplify anintragenic region of the 23S rRNA gene. A molecular size marker (1-kb ladder; Gibco BRL) is shown to the left (M).
Capsule of Erysipelothrix rhusiopathiae
November 2012 Volume 80 Number 11 iai.asm.org 3995
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
ml of water. Water-soluble antigens were extracted by treatment with65°C water, as described above, and the crude capsular antigens wereanalyzed by Western blotting.
Virulence testing in mice. Five female 6- to 8-week-old ddY mice(purchased from Japan SLC, Inc., Hamamatsu, Japan) were subcutane-ously (s.c.) inoculated with 2.0 � 108 CFU (approximately 107 times the50% lethal dose [LD50] of the Fujisawa strain) of either the PCho-defec-tive mutant clone 100 or clone 112. Five mice were inoculated s.c. with2.0 � 108 CFU of strain Fujisawa as a control. Mice were observed for 14days for clinical symptoms and death.
RESULTSOrganization of a cps-lic gene cluster. We previously constructeda nonreverting mutant Fujisawa strain of E. rhusiopathiae, YS-1, inwhich Tn916 was inserted into a gene (ERH_0855) and excisedfrom the chromosome, resulting in a permanent inactivation ofthe gene (26). Genome analysis of the Fujisawa strain revealed that
this gene is located in a cluster of genes encoding seven proteins(ERH_0855 to ERH_0861) that appear to be involved in capsularpolysaccharide synthesis (cps genes) (17). The putative lic operoninvolved in the production and incorporation of PCho includesfour genes (ERH_0862 to ERH_0865: licC-licA of ERH_0862,licD, licB, and licC-licA of ERH_0865) and lies immediately down-stream of the cps region. In E. rhusiopathiae, the choline kinase(licA) and phosphorylcholine cytidylyltransferase (licC) genes arefused, suggesting that they form bifunctional enzymes (Fig. 1A).RT-PCR analysis was performed with seven primer sets that weredesigned to create overlapping PCR products encompassing thecps and lic regions and the downstream gene (ERH_0866; ethanol-amine utilization protein gene [eutH]). A single band of the ex-pected length was amplified (Fig. 1B), indicating that these genesare transcribed as a polycistronic mRNA, forming an operon. Eth-anolamine can replace choline in the cell wall and substitute for a
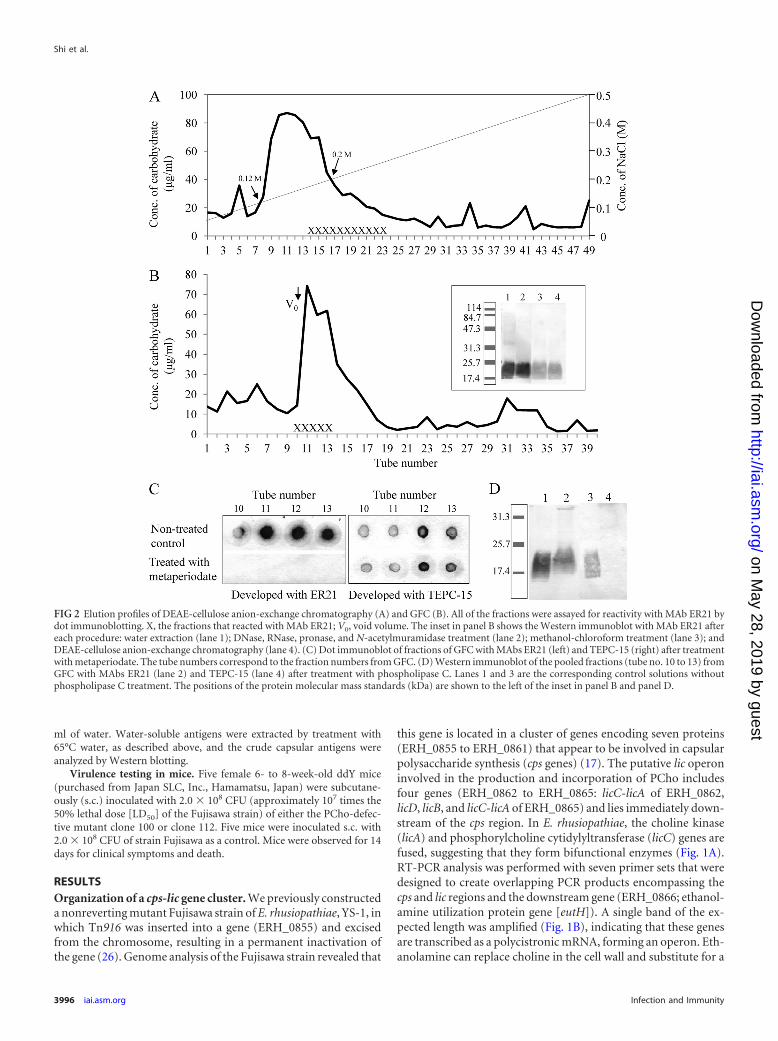
FIG 2 Elution profiles of DEAE-cellulose anion-exchange chromatography (A) and GFC (B). All of the fractions were assayed for reactivity with MAb ER21 bydot immunoblotting. X, the fractions that reacted with MAb ER21; V0, void volume. The inset in panel B shows the Western immunoblot with MAb ER21 aftereach procedure: water extraction (lane 1); DNase, RNase, pronase, and N-acetylmuramidase treatment (lane 2); methanol-chloroform treatment (lane 3); andDEAE-cellulose anion-exchange chromatography (lane 4). (C) Dot immunoblot of fractions of GFC with MAbs ER21 (left) and TEPC-15 (right) after treatmentwith metaperiodate. The tube numbers correspond to the fraction numbers from GFC. (D) Western immunoblot of the pooled fractions (tube no. 10 to 13) fromGFC with MAbs ER21 (lane 2) and TEPC-15 (lane 4) after treatment with phospholipase C. Lanes 1 and 3 are the corresponding control solutions withoutphospholipase C treatment. The positions of the protein molecular mass standards (kDa) are shown to the left of the inset in panel B and panel D.
Shi et al.
3996 iai.asm.org Infection and Immunity
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

growth requirement of S. pneumoniae (8), and it is intriguing thateutH is located in this region.
Transposon mutagenesis and the construction of PCho-de-fective mutants. To obtain PCho-defective mutants, we con-structed approximately 300 distinct transformants that containedsingle transposon insertions and determined the insertion sites bysequencing. Two mutants, clone 100 and clone 112, which con-tained transposons inserted into licC-licA (ERH_0862) and licD(ERH_0863), respectively (Fig. 1A), were successfully obtainedand used for further analysis.
Preparation of CPS. To determine the chemical composition,the crude CPS of Fujisawa was extracted with hot water, treatedwith nucleases, pronase, and N-acetylmuramidase, delipidated,and purified by DEAE-cellulose anion-exchange chromatogra-phy. As shown in Fig. 2A, a peak that contained a large amount ofcarbohydrate was eluted at salt concentrations of 0.12 M to 0.2 MNaCl; the fractions from the latter part of the peak showed immu-noreactivity with MAb ER21. The fractions containing MAbER21-reacting antigen(s) were pooled and further purified bySephacryl S-200 GFC. As shown in Fig. 2B, the fractions contain-ing MAb ER21-reacting antigen(s) were eluted near the void vol-ume. After Western immunoblotting with MAb ER21, a broadlysmeared band that reacted with MAb ER21 was observed in the17.4- to 25.7-kDa region. The patterns of reactivity were not al-tered when checked after each step, including water extraction,DNase, RNase, pronase, and N-acetylmuramidase treatment,methanol and chloroform treatment, and DEAE-cellulose anion-exchange chromatography (Fig. 2B, inset). To investigate the na-ture of the epitopes recognized by MAb ER21 and the presence ofphosphorylcholine in the fractions, the antigenic fractions fromGFC were treated with sodium metaperiodate or phospholipase Cand analyzed by dot and Western immunoblotting. As shown inFig. 2C, treatment with sodium metaperiodate, which cleaves thecarbon-carbon bond between vicinal diols, abolished the bindingof MAb ER21, suggesting that the epitopes recognized by MAbER21 are carbohydrates. The CPS-containing fractions (no. 10 to13) were pooled and further treated with phospholipase C. Treat-ment with phospholipase C resulted in a loss of reactivity with thePCho-specific MAb TEPC-15 (Fig. 2D). Taken together, these re-sults indicate that the CPS of E. rhusiopathiae is modified by PCho,as has been shown for other mucosal pathogens, including N.meningitidis, S. pneumoniae, and H. influenzae (5, 31, 33). Thepooled CPS-containing fractions were further analyzed.
Analysis of CPS components. To identify the monosaccharidecomponents of the CPS of Fujisawa, the ABEE-labeled monosac-charides were analyzed by HPLC. The CPS comprises seven majormonosaccharides, galacturonic acid (GalA), galactose (Gal), man-nose (Man), glucose (Glc), arabinose (Ara), xylose (Xyl), and N-acetylglucosamine (GlcNAc), and some minor monosaccharides,including ribose (Rib), rhamnose (Rha), and N-acetylgalac-tosamine (GalNAc). The identities of other minor monosaccha-rides were not determined due to a lack of standards (Fig. 3).
The purified native CPS and partially hydrolyzed CPS wereanalyzed by MALDI-TOF mass spectrometry to confirm themonosaccharide components of the CPS that were identified byHPLC and the presence of PCho in the CPS and to detect othernoncarbohydrate moieties. MALDI-TOF mass spectra of the pu-rified native CPS included a broad peak in the range of m/z 5,000to 10,000, and the partially hydrolyzed CPS exhibited no distinctpeaks in the high-mass range (data not shown). However, several
peaks were observed in the low-mass range of both samples (datanot shown). Figure 4 shows the representative tandem MS (MS/MS) spectra generated from four precursor ions in the positive- ornegative-ion modes. The precursor ion at m/z 1,547.9 gave severalions by losing m/z 146, 203, and 243, indicating that the precursorion contains Rha, N-acetylhexosamine, and an unidentified com-ponent (Fig. 4A). Likewise, the precursor ion at m/z 1,289.4 gen-erated a series of ions by losing m/z 132, 162, and 275, indicatingthat it contains a pentose (Ara, Xyl, or Rib), five hexoses, andprobably N-acetylmuramic acid (MurNAc), the latter of whichmay have been derived from peptidoglycan while preparing theCPS (Fig. 4B). The precursor ion at m/z 604 (Fig. 4C) gave productions with mass differences of m/z 203 and 43, indicating the ioncontains N-acetylhexosamine. The precursor ion at m/z 620 (Fig.4D) gave product ions with mass differences of m/z 97, 44, 59, and183; the latter two ions were also detected in the precursor ion atm/z 604 (Fig. 4C), which corresponds to neutral loss of H2PO4,(CH2)2O, trimethyl amine, and PCho, respectively, indicatingthat the ions contain PCho. Thus, mass spectrometric analysisconfirmed the modification of the CPS by PCho and the mono-saccharide composition of the CPS determined by HPLC.
Structural analysis of the CPS. To reveal the chemical struc-ture of the CPS, lyophilized CPS was dissolved in deuterium oxideand analyzed by NMR. Figure 5 shows the one-dimensional spec-tra of 1H and 13C DEPT-135 (1D-1H and 1D-13C). 1D-1H analysisrevealed several characteristic spectral clusters for the anomericregion at 4.4 to 5.5 ppm, the nonanomeric region at 3.3 to 4.4ppm, and three kinds of methyl residues at 3.21 ppm, 1.90 to 2.10ppm, and 1.10 to 1.17 ppm (Fig. 5A). The 1D-13C spectra were inaccordance with those of 1D-1H, except that the spectra of themethylenes of C-6 and PCho are shown downward in the DEPT-
FIG 3 HPLC chromatograms of ABEE-labeled 13 standard monosaccharides(A) and component monosaccharides released from the CPS of E. rhusio-pathiae (B). The CPS was hydrolyzed, N-acetylated, and converted to ABEE-monosaccharide. The HPLC analysis was carried out as described in Materialsand Methods. Asterisks represent peaks that were not determined due to a lackof standards. Peaks are numbered as follows: 1, GlcA; 2, GalA; 3, Gal; 4, Man;5, Glc; 6, Ara; 7, Rib; 8, ManNAc; 9, Xyl; 10, GlcNAc; 11, Fuc; 12, Rha; and 13,GalNAc.
Capsule of Erysipelothrix rhusiopathiae
November 2012 Volume 80 Number 11 iai.asm.org 3997
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

135 measurement (Fig. 5B). Analysis of the anomeric region fromHSQC revealed that the CPS contains at least nine monosaccha-rides; the H-2 sites of these monosaccharides could be traced un-equivocally from the anomeric H-1 sites in DQF-COSY (data notshown). These monosaccharides were labeled A, B, C, D, E, F, G,
H, and I. The integrals of the H-1 proton peaks of the nine mon-osaccharides were different (Fig. 6). These results confirmed theheterogeneity of the CPS with deoxyhexose, N-acetylhexosamineand other monosaccharides, and PCho. The coupling constants(3JH1,H2) of vicinal protons in monosaccharides A to F are no
FIG 4 MALDI-TOF MS/MS spectra of precursor ions at m/z 1,547.9 (A) and m/z 1,289.4 (B) in the positive-ion mode and precursor ions at m/z 604.2 (C) andm/z 620.2 (D) in the negative-ion mode. Spectra were obtained from native CPS (A and B) or CPS partially hydrolyzed with 2% acetic acid at 100°C for 2.5 h (Cand D).
Shi et al.
3998 iai.asm.org Infection and Immunity
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
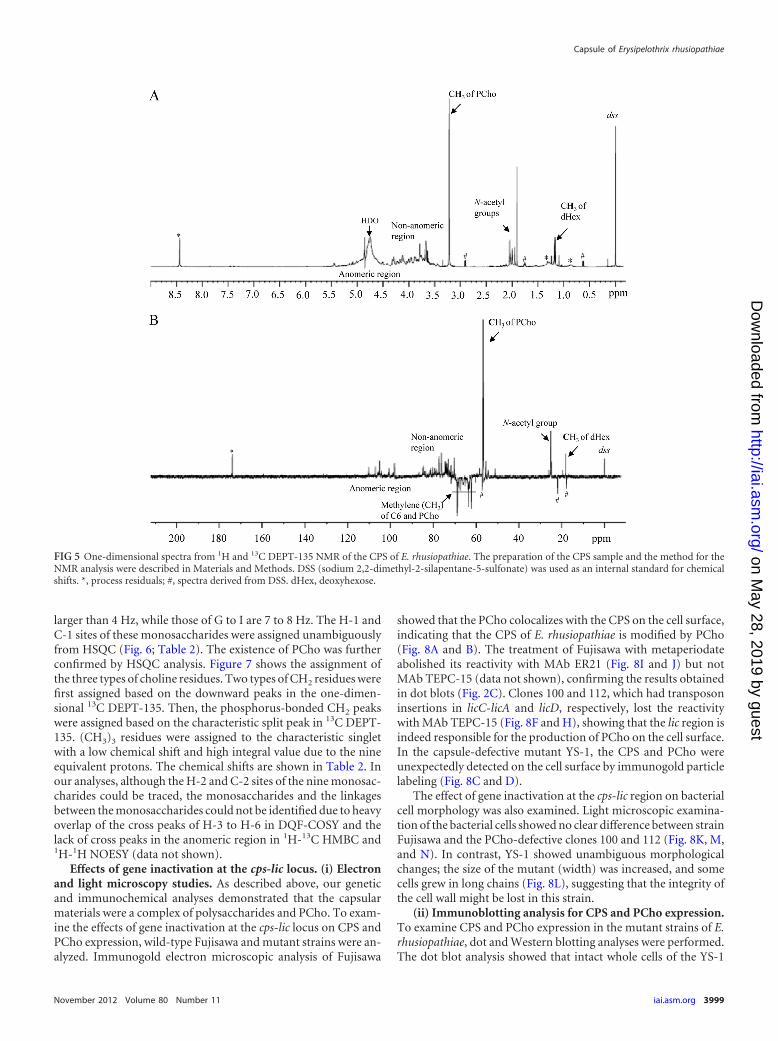
larger than 4 Hz, while those of G to I are 7 to 8 Hz. The H-1 andC-1 sites of these monosaccharides were assigned unambiguouslyfrom HSQC (Fig. 6; Table 2). The existence of PCho was furtherconfirmed by HSQC analysis. Figure 7 shows the assignment ofthe three types of choline residues. Two types of CH2 residues werefirst assigned based on the downward peaks in the one-dimen-sional 13C DEPT-135. Then, the phosphorus-bonded CH2 peakswere assigned based on the characteristic split peak in 13C DEPT-135. (CH3)3 residues were assigned to the characteristic singletwith a low chemical shift and high integral value due to the nineequivalent protons. The chemical shifts are shown in Table 2. Inour analyses, although the H-2 and C-2 sites of the nine monosac-charides could be traced, the monosaccharides and the linkagesbetween the monosaccharides could not be identified due to heavyoverlap of the cross peaks of H-3 to H-6 in DQF-COSY and thelack of cross peaks in the anomeric region in 1H-13C HMBC and1H-1H NOESY (data not shown).
Effects of gene inactivation at the cps-lic locus. (i) Electronand light microscopy studies. As described above, our geneticand immunochemical analyses demonstrated that the capsularmaterials were a complex of polysaccharides and PCho. To exam-ine the effects of gene inactivation at the cps-lic locus on CPS andPCho expression, wild-type Fujisawa and mutant strains were an-alyzed. Immunogold electron microscopic analysis of Fujisawa
showed that the PCho colocalizes with the CPS on the cell surface,indicating that the CPS of E. rhusiopathiae is modified by PCho(Fig. 8A and B). The treatment of Fujisawa with metaperiodateabolished its reactivity with MAb ER21 (Fig. 8I and J) but notMAb TEPC-15 (data not shown), confirming the results obtainedin dot blots (Fig. 2C). Clones 100 and 112, which had transposoninsertions in licC-licA and licD, respectively, lost the reactivitywith MAb TEPC-15 (Fig. 8F and H), showing that the lic region isindeed responsible for the production of PCho on the cell surface.In the capsule-defective mutant YS-1, the CPS and PCho wereunexpectedly detected on the cell surface by immunogold particlelabeling (Fig. 8C and D).
The effect of gene inactivation at the cps-lic region on bacterialcell morphology was also examined. Light microscopic examina-tion of the bacterial cells showed no clear difference between strainFujisawa and the PCho-defective clones 100 and 112 (Fig. 8K, M,and N). In contrast, YS-1 showed unambiguous morphologicalchanges; the size of the mutant (width) was increased, and somecells grew in long chains (Fig. 8L), suggesting that the integrity ofthe cell wall might be lost in this strain.
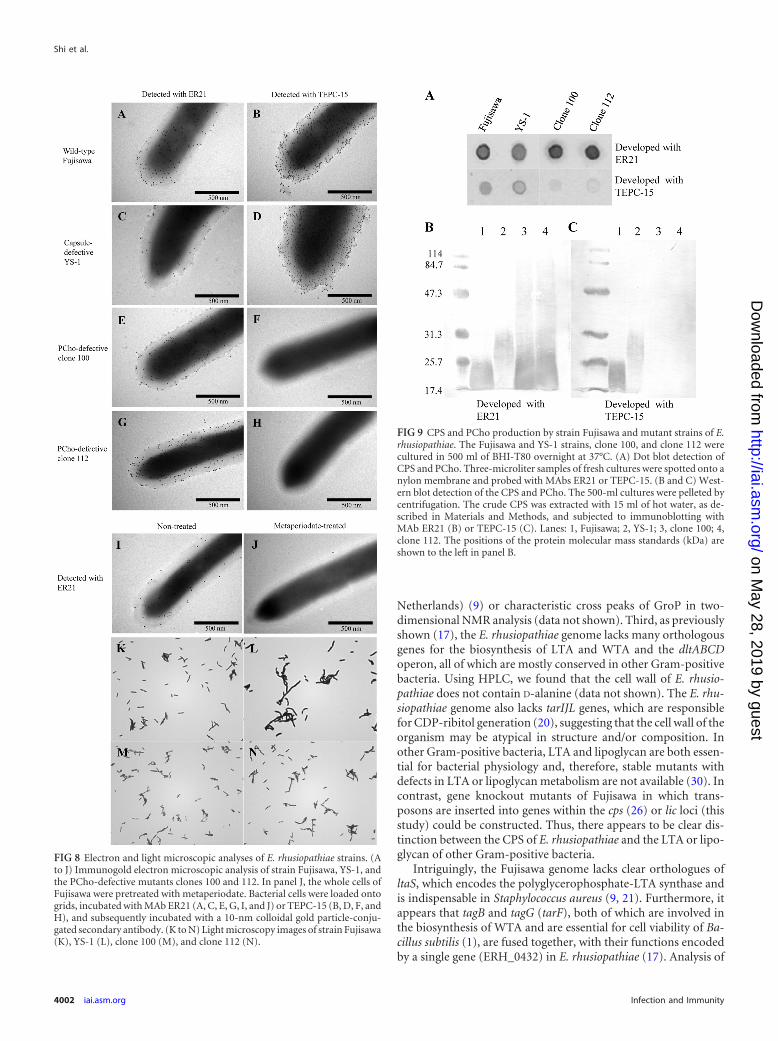
(ii) Immunoblotting analysis for CPS and PCho expression.To examine CPS and PCho expression in the mutant strains of E.rhusiopathiae, dot and Western blotting analyses were performed.The dot blot analysis showed that intact whole cells of the YS-1
FIG 5 One-dimensional spectra from 1H and 13C DEPT-135 NMR of the CPS of E. rhusiopathiae. The preparation of the CPS sample and the method for theNMR analysis were described in Materials and Methods. DSS (sodium 2,2-dimethyl-2-silapentane-5-sulfonate) was used as an internal standard for chemicalshifts. *, process residuals; #, spectra derived from DSS. dHex, deoxyhexose.
Capsule of Erysipelothrix rhusiopathiae
November 2012 Volume 80 Number 11 iai.asm.org 3999
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

strain reacted with the MAbs ER21 and TEPC-15 (Fig. 9A), whichwas also observed in the electron microscopy analysis. However,in the Western blot analysis, the band that reacted with MAb ER21was faint in the YS-1 strain, and retarded electrophoretic mobilitywas observed (Fig. 9B, lane 2). Taken together, these resultsstrongly suggest that the YS-1 strain had changes in the capsulestructure that affected its solubility and charge density. Likewise,the ER21-reactive bands of clones 100 and 112 also migrated withretarded mobility (Fig. 9B, lanes 3 and 4), suggesting that both the
CPS and PCho are required to maintain the molecular integrity ofthe capsule.
Virulence testing in mice. To determine whether PCho playsan important role in the virulence of E. rhusiopathiae, we inocu-lated mice s.c. with 2.0 � 108 CFU (approximately 107 times theLD50) of the Fujisawa strain or the PCho-defective mutants. Miceinoculated with the Fujisawa strain died within 2 days of inocula-tion, whereas all of the mice inoculated with the PCho-defectivemutants survived for 14 days without clinical symptoms. Thisresult indicates that PCho plays a critical role in the virulence ofthis organism.
DISCUSSION
Capsular polysaccharides are important virulence factors in awide range of bacterial species. The presence of a capsule in E.rhusiopathiae and the role of the capsule in virulence have beendetermined (26, 29). However, the chemical and biological prop-erties of the capsule have not been characterized.
Lachmann and Deicher (14) analyzed the surface componentsof E. rhusiopathiae by sulfate-polyacrylamide gel electrophoresisand immunoblotting. They found that the major antigen with amolecular weight of 14,000 to 22,000 had a positive anthrone re-action and that its antigenicity was lost after treatment withsodium metaperiodate but not with alkali or organic acids; there-fore, they concluded that the antigen is the capsular polysaccha-ride antigen. Schubert and Fiedler (22) investigated the cell sur-face of E. rhusiopathiae and found that the carbohydrate moiety ofthe cell wall is very complicated. The authors concluded that thecell surface polysaccharide is mainly composed of N-acetylfuco-
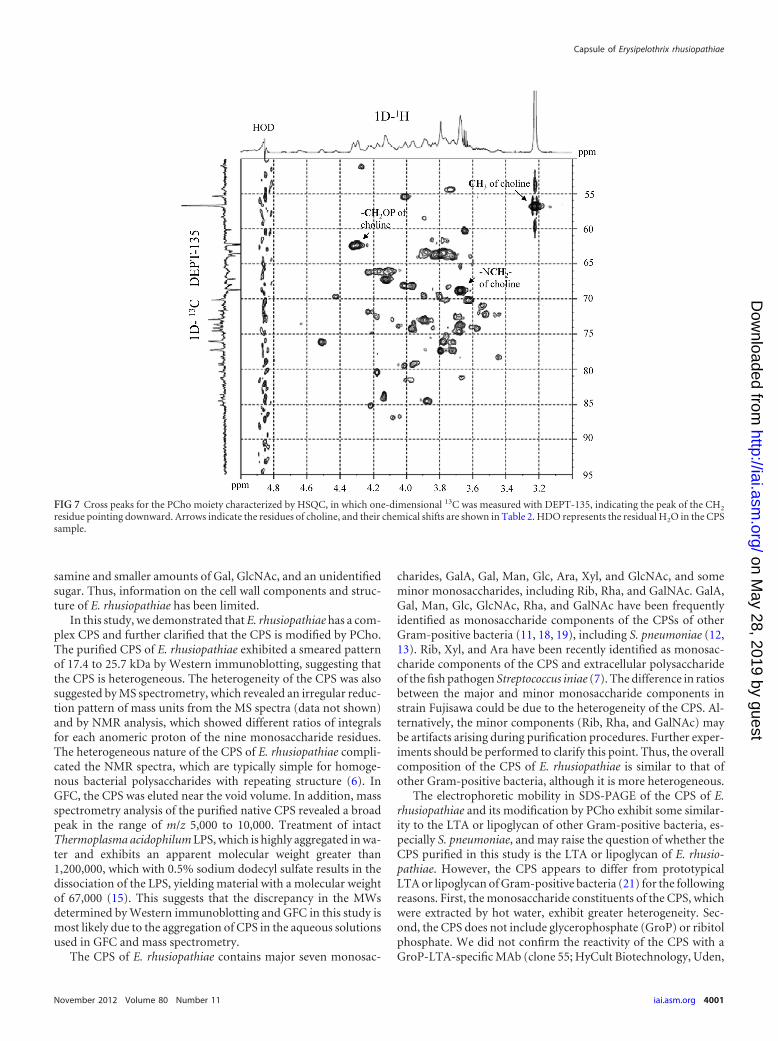
FIG 6 HSQC of the anomeric region of the CPS of E. rhusiopathiae. The cross peaks of nine monosaccharides for which H-2 sites could be unequivocally tracedfrom anomeric H-1 sites in DQF-COSY are labeled from A to I, and their chemical shifts are shown in Table 2. The integrals of the H-1 protons of thesemonosaccharides were calculated from the area of each peak. HDO represents the residual H2O in the CPS sample.
TABLE 2 Chemical shifts of each anomeric resonance of the ninemonosaccharides and phosphorylcholine of strain Fujisawa CPS
Residue
Chemical shift (ppm)
H–1 C–1
MonosaccharidesA 5.44 98.2B 5.14 98.8C 5.10 99.1D 5.08 110.3E 5.01 100.6F 4.96 100.5G 4.73 105.1H 4.64 107.2I 4.61 105.6
PhosphorylcholineCH3N– 3.21 56.8–NCH2– 3.66 68.8–CH2P 4.29 62.2
Shi et al.
4000 iai.asm.org Infection and Immunity
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
samine and smaller amounts of Gal, GlcNAc, and an unidentifiedsugar. Thus, information on the cell wall components and struc-ture of E. rhusiopathiae has been limited.
In this study, we demonstrated that E. rhusiopathiae has a com-plex CPS and further clarified that the CPS is modified by PCho.The purified CPS of E. rhusiopathiae exhibited a smeared patternof 17.4 to 25.7 kDa by Western immunoblotting, suggesting thatthe CPS is heterogeneous. The heterogeneity of the CPS was alsosuggested by MS spectrometry, which revealed an irregular reduc-tion pattern of mass units from the MS spectra (data not shown)and by NMR analysis, which showed different ratios of integralsfor each anomeric proton of the nine monosaccharide residues.The heterogeneous nature of the CPS of E. rhusiopathiae compli-cated the NMR spectra, which are typically simple for homoge-nous bacterial polysaccharides with repeating structure (6). InGFC, the CPS was eluted near the void volume. In addition, massspectrometry analysis of the purified native CPS revealed a broadpeak in the range of m/z 5,000 to 10,000. Treatment of intactThermoplasma acidophilum LPS, which is highly aggregated in wa-ter and exhibits an apparent molecular weight greater than1,200,000, which with 0.5% sodium dodecyl sulfate results in thedissociation of the LPS, yielding material with a molecular weightof 67,000 (15). This suggests that the discrepancy in the MWsdetermined by Western immunoblotting and GFC in this study ismost likely due to the aggregation of CPS in the aqueous solutionsused in GFC and mass spectrometry.
The CPS of E. rhusiopathiae contains major seven monosac-
charides, GalA, Gal, Man, Glc, Ara, Xyl, and GlcNAc, and someminor monosaccharides, including Rib, Rha, and GalNAc. GalA,Gal, Man, Glc, GlcNAc, Rha, and GalNAc have been frequentlyidentified as monosaccharide components of the CPSs of otherGram-positive bacteria (11, 18, 19), including S. pneumoniae (12,13). Rib, Xyl, and Ara have been recently identified as monosac-charide components of the CPS and extracellular polysaccharideof the fish pathogen Streptococcus iniae (7). The difference in ratiosbetween the major and minor monosaccharide components instrain Fujisawa could be due to the heterogeneity of the CPS. Al-ternatively, the minor components (Rib, Rha, and GalNAc) maybe artifacts arising during purification procedures. Further exper-iments should be performed to clarify this point. Thus, the overallcomposition of the CPS of E. rhusiopathiae is similar to that ofother Gram-positive bacteria, although it is more heterogeneous.
The electrophoretic mobility in SDS-PAGE of the CPS of E.rhusiopathiae and its modification by PCho exhibit some similar-ity to the LTA or lipoglycan of other Gram-positive bacteria, es-pecially S. pneumoniae, and may raise the question of whether theCPS purified in this study is the LTA or lipoglycan of E. rhusio-pathiae. However, the CPS appears to differ from prototypicalLTA or lipoglycan of Gram-positive bacteria (21) for the followingreasons. First, the monosaccharide constituents of the CPS, whichwere extracted by hot water, exhibit greater heterogeneity. Sec-ond, the CPS does not include glycerophosphate (GroP) or ribitolphosphate. We did not confirm the reactivity of the CPS with aGroP-LTA-specific MAb (clone 55; HyCult Biotechnology, Uden,
FIG 7 Cross peaks for the PCho moiety characterized by HSQC, in which one-dimensional 13C was measured with DEPT-135, indicating the peak of the CH2
residue pointing downward. Arrows indicate the residues of choline, and their chemical shifts are shown in Table 2. HDO represents the residual H2O in the CPSsample.
Capsule of Erysipelothrix rhusiopathiae
November 2012 Volume 80 Number 11 iai.asm.org 4001
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from
Netherlands) (9) or characteristic cross peaks of GroP in two-dimensional NMR analysis (data not shown). Third, as previouslyshown (17), the E. rhusiopathiae genome lacks many orthologousgenes for the biosynthesis of LTA and WTA and the dltABCDoperon, all of which are mostly conserved in other Gram-positivebacteria. Using HPLC, we found that the cell wall of E. rhusio-pathiae does not contain D-alanine (data not shown). The E. rhu-siopathiae genome also lacks tarIJL genes, which are responsiblefor CDP-ribitol generation (20), suggesting that the cell wall of theorganism may be atypical in structure and/or composition. Inother Gram-positive bacteria, LTA and lipoglycan are both essen-tial for bacterial physiology and, therefore, stable mutants withdefects in LTA or lipoglycan metabolism are not available (30). Incontrast, gene knockout mutants of Fujisawa in which trans-posons are inserted into genes within the cps (26) or lic loci (thisstudy) could be constructed. Thus, there appears to be clear dis-tinction between the CPS of E. rhusiopathiae and the LTA or lipo-glycan of other Gram-positive bacteria.
Intriguingly, the Fujisawa genome lacks clear orthologues ofltaS, which encodes the polyglycerophosphate-LTA synthase andis indispensable in Staphylococcus aureus (9, 21). Furthermore, itappears that tagB and tagG (tarF), both of which are involved inthe biosynthesis of WTA and are essential for cell viability of Ba-cillus subtilis (1), are fused together, with their functions encodedby a single gene (ERH_0432) in E. rhusiopathiae (17). Analysis of
FIG 8 Electron and light microscopic analyses of E. rhusiopathiae strains. (Ato J) Immunogold electron microscopic analysis of strain Fujisawa, YS-1, andthe PCho-defective mutants clones 100 and 112. In panel J, the whole cells ofFujisawa were pretreated with metaperiodate. Bacterial cells were loaded ontogrids, incubated with MAb ER21 (A, C, E, G, I, and J) or TEPC-15 (B, D, F, andH), and subsequently incubated with a 10-nm colloidal gold particle-conju-gated secondary antibody. (K to N) Light microscopy images of strain Fujisawa(K), YS-1 (L), clone 100 (M), and clone 112 (N).
FIG 9 CPS and PCho production by strain Fujisawa and mutant strains of E.rhusiopathiae. The Fujisawa and YS-1 strains, clone 100, and clone 112 werecultured in 500 ml of BHI-T80 overnight at 37°C. (A) Dot blot detection ofCPS and PCho. Three-microliter samples of fresh cultures were spotted onto anylon membrane and probed with MAbs ER21 or TEPC-15. (B and C) West-ern blot detection of the CPS and PCho. The 500-ml cultures were pelleted bycentrifugation. The crude CPS was extracted with 15 ml of hot water, as de-scribed in Materials and Methods, and subjected to immunoblotting withMAb ER21 (B) or TEPC-15 (C). Lanes: 1, Fujisawa; 2, YS-1; 3, clone 100; 4,clone 112. The positions of the protein molecular mass standards (kDa) areshown to the left in panel B.
Shi et al.
4002 iai.asm.org Infection and Immunity
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from

the E. rhusiopathiae genome also suggests that tagO, which is in-dispensable for the growth of Gram-positive bacteria (1), is mostlikely fused with mraY (17), a gene essential for peptidoglycanbiosynthesis. Given the unique phylogenetic position of E. rhusio-pathiae, there may be some overlap between the pathways for cpsand other cell wall components, including WTA and/or LTA, dueto reductive genome evolution. Such an overlap could explain theproduction of an atypical CPS by E. rhusiopathiae and the mor-phological changes observed in YS-1, in which the integrity of thecell wall might be lost. The gene ERH_0855, which was inactivatedin the YS-1 strain, encodes a protein containing a domain that ishighly conserved in a large number of different bacterial sugartransferases (pfam02397) in a broad spectrum of Gram-negativeand Gram-positive bacteria (17). It is therefore possible that inac-tivation of this gene might have pleiotropic effects on the structureof the CPS and cell wall biosynthesis, which are important formaintaining cell wall integrity.
In summary, E. rhusiopathiae has a complex CPS, which ismodified by PCho. The complexity of the E. rhusiopathiae CPSmay be related to the unique phylogenetic position of the organ-ism. To fully understand the pathogenicity and physiology of E.rhusiopathiae, further clarification of the cell wall structure andcomposition is needed.
ACKNOWLEDGMENTS
We thank Kazuma Shiraiwa for technical assistance.This work was supported in part by a Research and Development
Project for Application in Promoting New Policy of Agriculture, Forestryand Fisheries grant from the Ministry of Agriculture, Forestry and Fish-eries of Japan (to Y.S.).
REFERENCES1. Bhavsar AP, Brown ED. 2006. Cell wall assembly in Bacillus subtilis: how
spirals and spaces challenge paradigms. Mol. Microbiol. 60:1077–1090.2. Bligh GE, Dyer JW. 1959. A rapid method of total lipid extraction and
purification. Can. J. Biochem. Physiol. 37:911–917.3. Cao M, Bitar AP, Marquis H. 2007. A mariner-based transposition
system for Listeria monocytogenes. Appl. Environ. Microbiol. 73:2758 –2761.
4. Casal J, Jado I, Fenoll A, Perez A, Toraño A. 1998. Periodate oxidationof R36A pneumococci greatly enhances production of hybridomas secret-ing anti-protein antibodies. Microb. Pathog. 24:111–116.
5. Casey R, Newcombe J, McFadden J, Bodman-Smith KB. 2008. Theacute-phase reactant C-reactive protein binds to phosphorylcholine-expressing Neisseria meningitidis and increases uptake by human phago-cytes. Infect. Immun. 76:1298 –1304.
6. Christopher J. 2005. NMR assays for carbohydrate-based vaccines. J.Pharm. Biomed. Anal. 38:840 – 850.
7. Eyngor M, et al. 2008. Emergence of novel Streptococcus iniae exopoly-saccharide-producing strains following vaccination with nonproducingstrains. Appl. Environ. Microbiol. 74:6892– 6897.
8. Fischer W. 2000. Phosphocholine of pneumococcal teichoic acids: role inbacterial physiology and pneumococcal infection. Res. Microbiol. 151:421– 427.
9. Gründling A, Schneewind O. 2007. Synthesis of glycerol phosphate lipo-teichoic acid in Staphylococcus aureus. Proc. Natl. Acad. Sci. U. S. A. 104:8478 – 8483.
10. Hodge JE, Hofreiter BT. 1962. Determination of reducing sugar andcarbohydrates. Methods Carbohydr. Chem. 1:380 –394.
11. Kalelkar S, Glushka J, Halbeek H, Morris LC, Cherniak R. 1997.Structure of the capsular polysaccharide of Clostridium perfringens Hobbs5 as determined by NMR spectroscopy. Carbohydrate Res. 299:119 –128.
12. Karlsson C, Jansson P, Søreson UB. 1998. The chemical structures of thecapsular polysaccharides from Streptococcus pneumoniae types 32F and32A. Eur. J. Biochem. 255:296 –302.
13. Karlsson C, Jansson P, Widmalm G, Søreson UB. 1997. Structural
elucidation of the capsular polysaccharide from streptococcus pneumoniaetype 18B. Carbohydrate Res. 304:165–172.
14. Lachmann PG, Deicher H. 1986. Solubilization and characterization ofsurface antigenic components of Erysipelothrix rhusiopathiae T28. Infect.Immun. 52:818 – 822.
15. Mayberry-Carson KJ, Roth IL, Smith PF. 1975. Ultrastructure of lipo-polysaccharide isolated from Thermoplasma acidophilum. J. Bacteriol.121:700 –703.
16. Nagahama M, Nakayama T, Michiue K, Sakurai J. 1997. Site-specificmutagenesis of Clostridium perfringens alpha-toxin: replacement of Asp-56, Asp-130, or Glu-152 causes loss of enzymatic and hemolytic activities.Infect. Immun. 65:3489 –3492.
17. Ogawa Y, et al. 2011. The genome of Erysipelothrix rhusiopathiae, thecausative agent of swine erysipelas, reveals new insights into the evolutionof Firmicutes and the organism’s intracellular adaptations. J. Bacterial.193:2959 –2971.
18. Ovodov YS. 2006. Bacterial capsular antigens. Structural patterns of cap-sular antigens. Biochemistry (Mosc.) 71:937–954.
19. Park S, et al. 2010. Characterization of the structure and biological func-tions of a capsular polysaccharide produced by Staphylococcus saprophyti-cus. J. Bacteriol. 192:4618 – 4626.
20. Pereira MP, Brown ED. 2004. Bifunctional catalysis by CDP-ribitol syn-thase: convergent recruitment of reductase and cytidylyltransferase activ-ities in Haemophilus influenzae and Staphylococcus aureus. Biochemistry43:11802–11812.
21. Rahman O, Dover LG, Sutcliffe IC. 2009. Lipoteichoic acid biosynthesis:two steps forwards, one step sideways? Trends Microbiol. 17:219 –225.
22. Schubert K, Fiedler F. 2001. Structural investigations on the cell surfaceof Erysipelothrix rhusiopathiae. Syst. Appl. Microbiol. 24:26 –30.
23. Sheng S, Cherniak R. 1998. Structure of the capsular polysaccharide ofClostridium perfringens Hobbs 10 determined by NMR spectroscopy. Car-bohydr. Res. 305:65–72.
24. Shimoji Y. 2000. Pathogenicity of Erysipelothrix rhusiopathiae: virulencefactors and protective immunity. Microbes Infect. 2:965–972.
25. Shimoji Y, Asato H, Sekizaki T, Mori Y, Yokomizo Y. 2002. Hyaluron-idase is not essential for the lethality of Erysipelothrix rhusiopathiae infec-tion in mice. J. Vet. Med. Sci. 64:173–176.
26. Shimoji Y, Mori M, Sekizaki T, Shibahara T, Yokomizo Y. 1998.Construction and vaccine potential of acapsular mutants of Erysipelothrixrhusiopathiae: use of excision of Tn916 to inactivate a target gene. Infect.Immun. 66:3250 –3254.
27. Shimoji Y, et al. 2002. Erysipelothrix rhusiopathiae YS-1 as a live vaccinevehicle for heterologous protein expression and intranasal immunizationof pigs. Infect. Immun. 70:226 –232.
28. Shimoji Y, Yokomizo Y, Mori Y. 1996. Intracellular survival and repli-cation of Erysipelothrix rhusiopathiae within murine macrophages: failureof induction of oxidative burst of macrophages. Infect. Immun. 64:1789 –1793.
29. Shimoji Y, Yokomizo Y, Sekizaki T, Mori Y, Kubo M. 1994. Presence ofa capsule in Erysipelothrix rhusiopathiae and its relationship to virulencefor mice. Infect. Immun. 62:2806 –2810.
30. Sutcliffe IC, Shaw N. 1991. Atypical lipoteichoic acids of gram-positivebacteria. J. Bacteriol. 173:7065–7069.
31. Thornton JA, Durick-Eder K, Tuomanen EI. 2010. Pneumococcalpathogenesis: “innate invasion” yet organ-specific damage. J. Mol. Med.88:103–107.
32. Weiser JN, Goldberg JB, Pan N, Wilson L, Virji M. 1998. The phos-phorylcholine epitope undergoes phase variation on a 43-kilodalton pro-tein in Pseudomonas aeruginosa and on pili of Neisseria meningitidis andNeissseria gonorrhoeae. Infect. Immun. 66:4263– 4267.
33. Weiser JN, et al. 1998. Phosphorylcholine on the lipopolysaccharide ofHaemophilus influenzae contributes to persistence in the respiratory tractand sensitivity to serum killing mediated by C-reactive protein. J. Exp.Med. 187:631– 640.
34. Wood RL. 1999. Erysipelas, p 419 – 430. In Straw BE, D’Allaire S, Menge-ling WL, Taylor DJ (ed), Diseases of swine, 8th ed. Iowa State UniversityPress, Ames, IA.
35. Yasuno S, Kokubo K, Kamei M. 1999. New method for determining thesugar composition of glycoproteins, glycolipids, and oligosaccharides byhigh-performance liquid chromatography. Biosci. Biotechnol. Biochem.63:1353–1359.
Capsule of Erysipelothrix rhusiopathiae
November 2012 Volume 80 Number 11 iai.asm.org 4003
on May 28, 2019 by guest
http://iai.asm.org/
Dow
nloaded from