Embed Size (px)
Citation preview

ĐẠI CƢƠNG VỀ CẤU TRÚC CỦA PROTEIN
Nguyên lý cơ bản của vấn đề này nằm trong câu nói “Chức năng bắt nguồn từ cấu trúc lập thể, và
cấu trúc lập thể do trình tự amino acid quyết định”. Thực tế, chuỗi protein gấp nếp thành các cấu
trúc lập thể đặc thù. Cấu trúc này được giữ ổn định bởi nhiều loại tương tác không cộng hóa trị
giữa các amino acid.
Chúng ta sẽ nghiên cứu lần lượt từng bậc tổ chức của protein.
4.1 Cấu trúc bậc 1: Amino acid mạch thẳng
Trong protein, các amino acid gắn kết với nhau thông qua liên kết amide cộng hóa trị, liên kết
này được gọi là liên kết peptide, chúng tạo thành chuỗi mạch thẳng, không phân nhánh. Đôi khi
cũng có các liên kết disulfide nối cộng hóa trị nối các mạch phụ (nhóm R) lại với nhau. Một đầu
của protein có nhóm amin tự do (không tạo liên kết) được gọi là đầu N, và đầu kia có nhóm
carboxyl tự do gọi là đầu C.
Cấu trúc bậc 1 của protein là sự xếp mạch thẳng một cách đơn giản, kích cỡ được tính bằng đơn
vị Dalton, trọng lượng phân tử trung bình của mỗi amino acid là 113, do vậy ta có thể dùng giá
trị này để ước lượng được số phần tử amino acid của một protein và ngược lại.

Hình 4.1:Mô tả cấu trúc thẳng (2D và 3D) của chuỗi polypeptide(Theo Color Atlas of
Biochemistry 2ed – Koolman)
4.2 Cấu trúc bậc 2: Thành phần tạo thành kiến trúc trung tâm, quan trọng
nhất của protein
Đây là cách sắp xếp không gian bền của các vùng trong chuỗi polypeptide (tạo thành cấu trúc
bậc 1). Những đoạn này gắn kết với nhau thông qua liên kết Hydrogen. Tùy theo trình tự, mỗi
chuỗi polypeptide có thể chứa nhiều kiểu cấu trúc bậc 2 trên nhiều vùng. Các cấu trúc bậc 2 cơ
bản là:
Xoắn α
Hình 4.2:Mô tả Xoắn α(Theo Color Atlas of Biochemistry 2ed – Koolman)
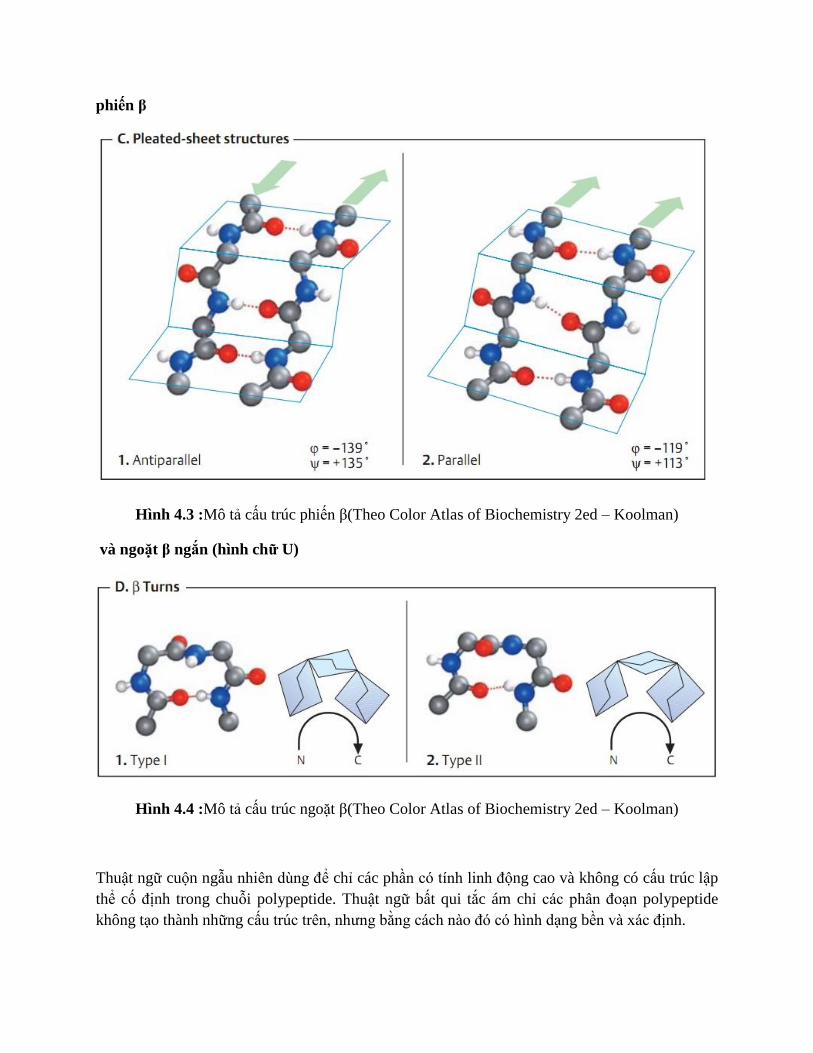
phiến β
Hình 4.3 :Mô tả cấu trúc phiến β(Theo Color Atlas of Biochemistry 2ed – Koolman)
và ngoặt β ngắn (hình chữ U)
Hình 4.4 :Mô tả cấu trúc ngoặt β(Theo Color Atlas of Biochemistry 2ed – Koolman)
Thuật ngữ cuộn ngẫu nhiên dùng để chỉ các phần có tính linh động cao và không có cấu trúc lập
thể cố định trong chuỗi polypeptide. Thuật ngữ bất qui tắc ám chỉ các phân đoạn polypeptide
không tạo thành những cấu trúc trên, nhưng bằng cách nào đó có hình dạng bền và xác định.

Có khoảng 60% chuỗi polypeptide của protein có dạng xoắn α và phiến β, phần còn lại mang cấu
trúc cuộn và ngoặt. Do đó, xoắn α và phiến β là các thành phần chống đỡ bên trong quan trọng
của protein.
Xoắn α là một phân đoạn polypeptide có kiểu gấp nếp, mạch khung xoắn lại. Ở cấu trúc này, mọi
O của nhóm –CO– của amino acid này sẽ tạo liên kết H với H thuộc nhóm –NH– của amino
acid kế cận, trừ đầu tận cùng không tạo liên kết với nhau. Kiểu sắp xếp có chu kì này tạo thành
cấu trúc xoắn từ đầu amin đến đầu carboxyl vì toàn bộ chất cho liên kết hydro có cùng chiều từ
trên xuống dưới, và cũng do đó, mỗi vòng xoắn chứa 3,6 amino acid.
Cơ cấu này do Linus Pauling đề xuất (Nobel hóa học), cách tạo cầu nối Hydrogen này làm cho
vòng xoắn α bền vững và đồng thời xác định tính chất kị nước của xoắn. Trong dung dịch, xoắn
ưa nước thường nằm tại bề mặt, nơi chúng có thể tương tác với môi trường nước, trong khi các
xoắn kị nước thường vùi vào trong lõi của protein gấp nếp. Hiện tượng này giải thích tại sao một
tổ hợp vòng xoắn α như thụ thể liên kết với protein G (GPCR) có thể thay đổi hình dạng sau khi
liên kết với ligand.
Phiến β là một phân đoạn ngắn và hầu như trải ra hết cỡ. Trong cấu trúc này, liên kết Hydrogen
hình thành trong phiến giữa các nguyên tử nằm trên khung những mạch β riêng biệt. Các mạch β
riêng biệt này có thể cùng nằm trong một chuỗi polypeptide và kết nối thông qua những vòng dài
hoặc ngắn, hoặc nằm trên các chuỗi polypeptide khác nhau. Từ các đặc điểm trên, hai hoặc nhiều
mạch β sắp xếp kề nhau, tạo thành tấm β hai chiều gầu như xếp ly (hoặc gọi là tấm xếp có nếp).
Liên kết Hydrogen trong mặt phẳng các phiến giữ mạch với nhau và nhóm R gắn phía trên hoặc
phía dưới mặt phẳng này. Giống như xoắn α, phiến β có tính định hướng do chiều liên kết
peptide. Do đó, trong phiến xếp, các mạch β sát nhau nằm cùng chiều (song song) hoặc ngược
chiều (đối song song). Trong một số protein, phiến β tạo thành phần nền của hốc gắn hoặc lõi kị
nước; ở các protein xuyên màng, phiến β cuộn lại tạo thành lỗ trung tâm ưa nước, cho phép ion
và tiểu phân tử đi qua.
Loại cấu trúc phiến β hiếm gặp trên thụ thể cũng như các kênh ion nhưng lại rất phổ biến trong
enzyme. Điều đó được giải thích là do lực nối Hydrogen liên phân tử trong cấu trúc này tương
đối yếu nên tạo ra được sự linh động lớn trong cấu trúc lập thể cần thiết cho hoạt động của
enzyme.
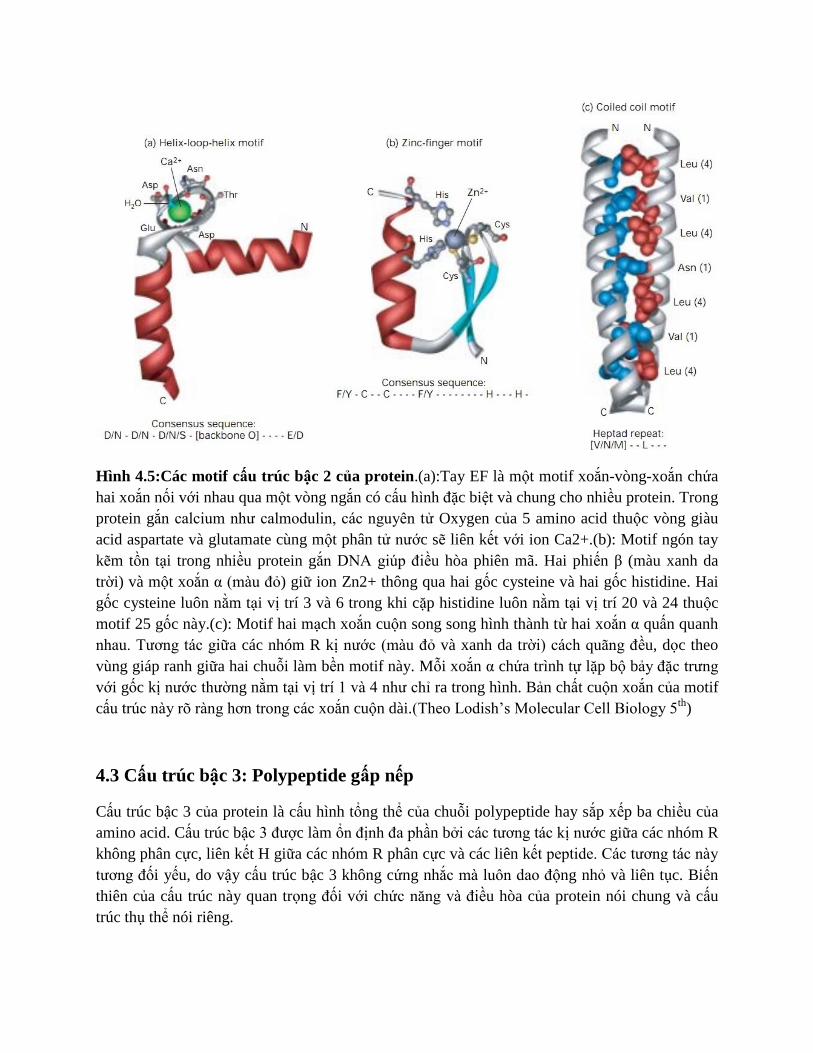
Hình 4.5:Các motif cấu trúc bậc 2 của protein.(a):Tay EF là một motif xoắn-vòng-xoắn chứa
hai xoắn nối với nhau qua một vòng ngắn có cấu hình đặc biệt và chung cho nhiều protein. Trong
protein gắn calcium như calmodulin, các nguyên tử Oxygen của 5 amino acid thuộc vòng giàu
acid aspartate và glutamate cùng một phân tử nước sẽ liên kết với ion Ca2+.(b): Motif ngón tay
kẽm tồn tại trong nhiều protein gắn DNA giúp điều hòa phiên mã. Hai phiến β (màu xanh da
trời) và một xoắn α (màu đỏ) giữ ion Zn2+ thông qua hai gốc cysteine và hai gốc histidine. Hai
gốc cysteine luôn nằm tại vị trí 3 và 6 trong khi cặp histidine luôn nằm tại vị trí 20 và 24 thuộc
motif 25 gốc này.(c): Motif hai mạch xoắn cuộn song song hình thành từ hai xoắn α quấn quanh
nhau. Tương tác giữa các nhóm R kị nước (màu đỏ và xanh da trời) cách quãng đều, dọc theo
vùng giáp ranh giữa hai chuỗi làm bền motif này. Mỗi xoắn α chứa trình tự lặp bộ bảy đặc trưng
với gốc kị nước thường nằm tại vị trí 1 và 4 như chỉ ra trong hình. Bản chất cuộn xoắn của motif
cấu trúc này rõ ràng hơn trong các xoắn cuộn dài.(Theo Lodish’s Molecular Cell Biology 5th
)
4.3 Cấu trúc bậc 3: Polypeptide gấp nếp
Cấu trúc bậc 3 của protein là cấu hình tổng thể của chuỗi polypeptide hay sắp xếp ba chiều của
amino acid. Cấu trúc bậc 3 được làm ổn định đa phần bởi các tương tác kị nước giữa các nhóm R
không phân cực, liên kết H giữa các nhóm R phân cực và các liên kết peptide. Các tương tác này
tương đối yếu, do vậy cấu trúc bậc 3 không cứng nhắc mà luôn dao động nhỏ và liên tục. Biến
thiên của cấu trúc này quan trọng đối với chức năng và điều hòa của protein nói chung và cấu
trúc thụ thể nói riêng.
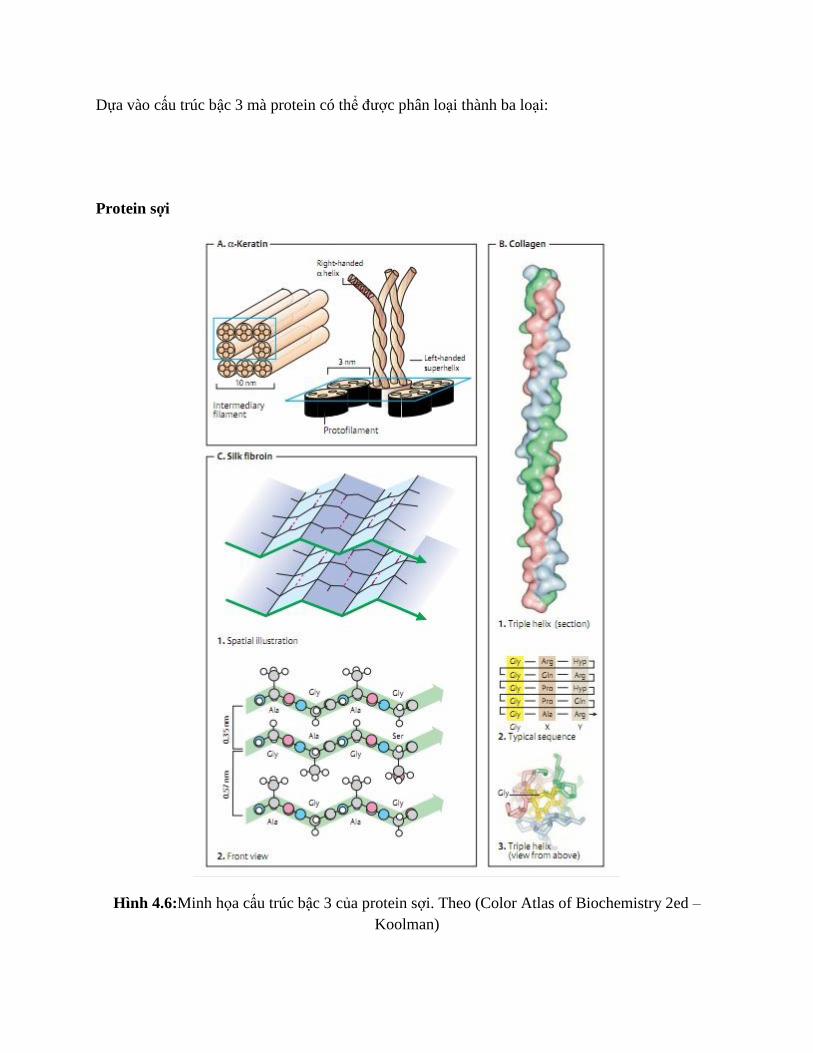
Dựa vào cấu trúc bậc 3 mà protein có thể được phân loại thành ba loại:
Protein sợi
Hình 4.6:Minh họa cấu trúc bậc 3 của protein sợi. Theo (Color Atlas of Biochemistry 2ed –
Koolman)
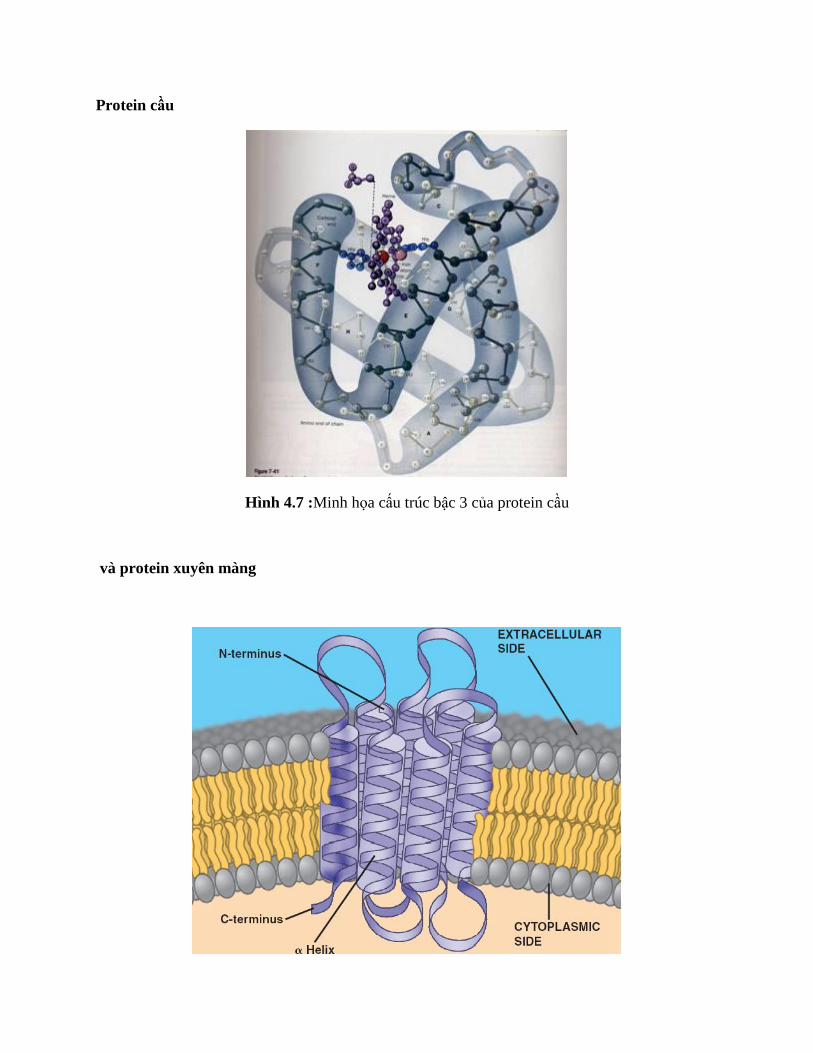
Protein cầu
Hình 4.7 :Minh họa cấu trúc bậc 3 của protein cầu
và protein xuyên màng

Hình 4.8:Minh họa cấu trúc bậc 3 của protein xuyên màng
Protein sợi là phân tử lớn, dài, cứng và thường cấu tạo từ nhiều trình tự ngắn lặp lại liên tiếp, tạo
thành cấu trúc bậc 3 lặp đơn (xem cấu trúc của collagen). Protein sợi thường tụ thành các sợi lớn
gồm nhiều protein và chúng không tan trong nước, có vai trò lớn trong vận động tế bào.
Protein cầu thường chứa tập hợp các cấu trúc bậc 2, hòa tan trong nước, gấp nếp chặt và không
có hình cầu hoàn hảo.
Protein xuyên màng nhúng trong lớp phospholipid kép của màng.
3 loại protein trên không phải lúc nào cũng được phân định rõ ràng. Một số protein cấu thành từ
tổ hợp 2 hay cả 3 loại này.
Các tổ hợp cấu trúc bậc 2 và bậc 3 nhất định được gọi là motif cấu trúc hay kiểu gấp nếp. Motif
cấu trúc góp phần hình thành cấu trúc tổng thể của toàn bộ protein và mỗi loại motif cấu trúc
thường thực hiện một chức năng chung trong các protein khác nhau (ví dụ với tiểu phân tử hay
ion, đặc biệt quan trọng đối với thụ thể). Các trình tự bậc 1 mã hóa cho một loại motif nhất định
có thể rất giống nhau. Hay một motif trình tự chung có thể tạo ra một motif cấu trúc lập thể
chung. Tuy nhiên, cũng có những trình tự không giống nhau lại có thể gấp nếp lại thành những
motif cấu trúc chung. Đôi khi các motif trình tự ngắn chứa một loại amino acid lớn bất thường
(proline, aspartate, glutamate, …) sẽ được gọi là miền.
Nhiều motif cấu trúc sử dụng xoắn α. Một motif gắn calcium phổ biến được gọi là tay EF (EF
hand) sử dụng hai xoắn ngắn kết nối với nhau qua vùng vòng. Motif này tồn tại trong hơn 100
protein cảm biến dò nồng độ calcium trong tế bào. Calcium gắn với nguyên tử Oxygen tại các
gốc bảo tồn ở vùng vòng phụ thuộc vào nồng độ Calcium và thường gây biến đổi hoạt tính của
protein. Do đó, nồng độ calcium có thể điều kiển trực tiếp cấu trúc và chứng năng của protein.
Protein thường sử dụng motif cấu trúc dạng xoắn – ngoặt – xoắn (helix – turn – helix) và xoắn –
vòng – xoắn cơ sở (basic helix – loop – helix, bHLH) để gắn DNA và qua đó điều hòa hoạt tính
gene. Ví dụ như cấu trúc dạng ngón tay kẽm (zinc finger) nằm trong các protein gắn DNA và
RNA có cấu trúc 1 xoắn α và 2 xoắn β nằm đối song gắn với nhau bởi ion Zn giống như ngón
tay.
Ngoài ra còn có các cấu tạo khác như “miền cấu trúc”, “miền chức năng”, … tuy nhiên do không
nằm trong phạm vi nghiên cứu của đề tài nên không được đề cập ở đây.
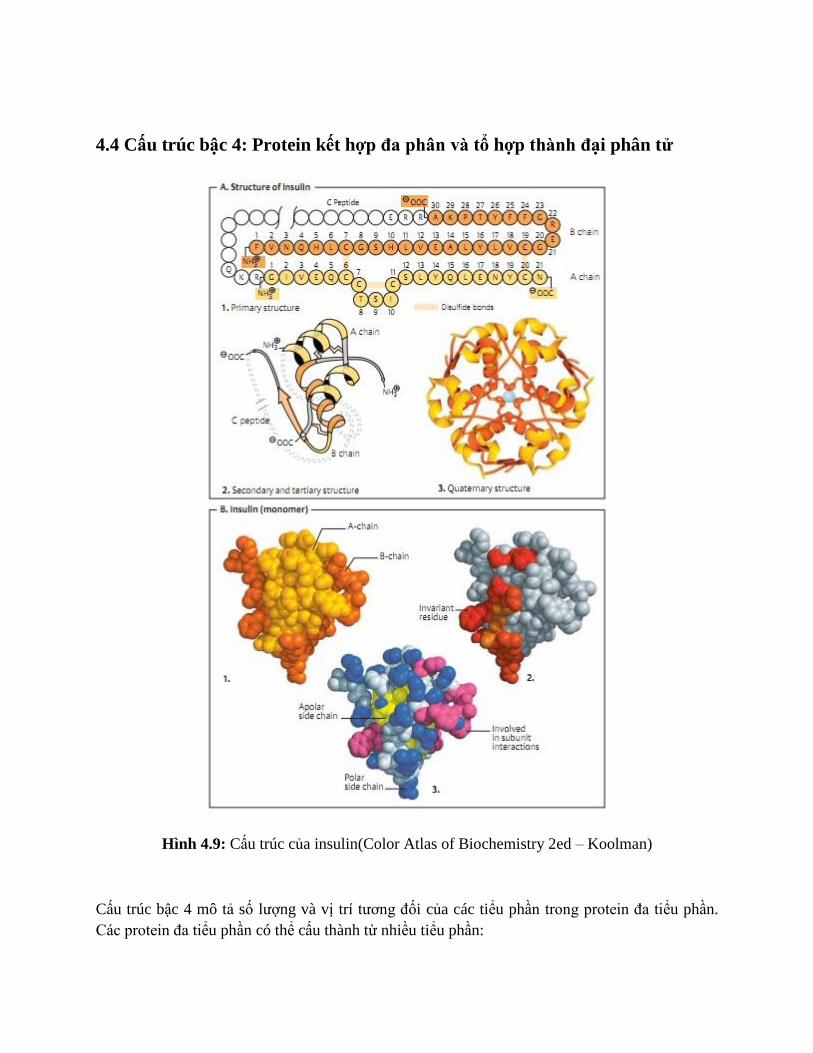
4.4 Cấu trúc bậc 4: Protein kết hợp đa phân và tổ hợp thành đại phân tử
Hình 4.9: Cấu trúc của insulin(Color Atlas of Biochemistry 2ed – Koolman)
Cấu trúc bậc 4 mô tả số lượng và vị trí tương đối của các tiểu phần trong protein đa tiểu phần.
Các protein đa tiểu phần có thể cấu thành từ nhiều tiểu phần:

Đồng nhất (homomeric)
Hình 4.10: Minh họa protein có tiểu phần đồng nhất(Color Atlas of Biochemistry 2ed –
Koolman)
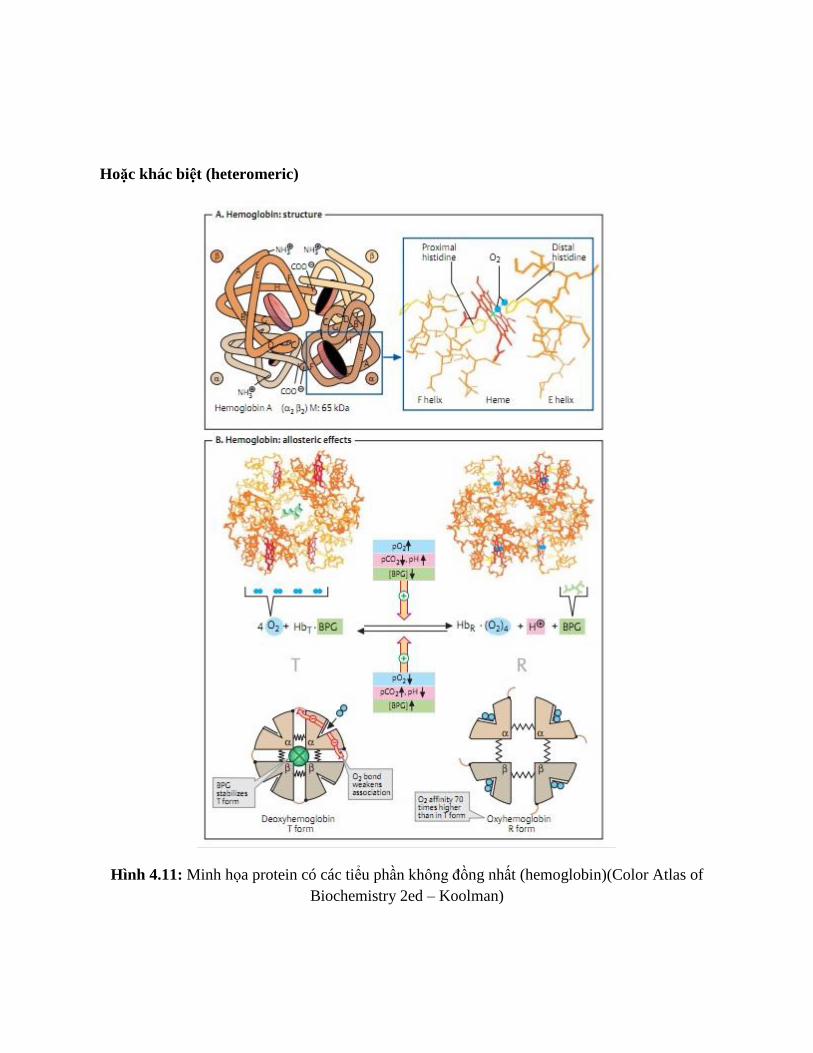
Hoặc khác biệt (heteromeric)
Hình 4.11: Minh họa protein có các tiểu phần không đồng nhất (hemoglobin)(Color Atlas of
Biochemistry 2ed – Koolman)

Thông thường, các tiểu phần riêng biệt không có chức năng trừ khi chúng lắp ráp thành protein
đa tiểu phần. Trong một số trường hợp, protein đa tiểu phần bố trí tiểu phần kề nhau theo chuỗi
phản ứng cần cho một con đường tế bào và chính điều này làm tăng hiệu quả vận hành của
chúng. Cấu trúc bậc cao nhất này của protein là sự kết hợp các protein thành tổ hợp đại phân tử
có kích thước và khối lượng lớn ( trên 1 Mda và 30-300 nm). Tổ hợp đại phân tử với chức năng
cấu trúc bao gồm capsid (bao bọc bộ gen của virus) và các bó sợi khung tế bào (hình thành màng
tế bào chất), hoạt động như bộ máy truyền tin, thực hiện hầu hết các quá trình phức tạp trong quá
trình truyền tin nội và ngoại bào, … Ví dụ bộ máy phiên mã có vai trò tổng hợp RNA thông tin
(mRNA) từ khuôn DNA. Bộ máy này chứa RNA polymerase (protein đa tiểu phần) và ít nhất 50
thành phần khác, bao gồm các yếu tố phiên mã, protein gắn promoter, helicase, …
4.5 HIỆU QUẢ GẤP NẾP CỦA PROTEIN: Chaperone & Chaperonin
Các chất biến tính (pH, nhiệt độ, β-mercaptoethanol …) có thể phá hủy các tương tác không
cộng hóa trị của protein và làm biến tính protein. Dưới những điều kiện biến tính như vậy,
entropy tăng khi quần thể đồng nhất chứa các phân tử gấp nếp bị mất ổn dịnh và chuyển hóa
thành các tập hợp chứa nhiều phân tử không gấp nếp (hay biến tính). Tập hợp protein biến tính
này sẽ tạo thành rất nhiều protein không có hoạt tính sinh học và thực tế chúng cũng không tồn
tại trong trạng thái tự nhiên (mặc dù theo lý thuyết có tới 8n-1
cấu hình).
Lời giải chính xác cho thực tế trên nằm trong tập hợp protein gọi là chaperone. Chúng giúp cho
protein gấp nếp. Chaperone có vai trò rất quan trọng và có thể thấy điều này bởi vì chúng được
bảo tổn qua tiến hóa. Có 2 họ chaperone thường thấy:
- Chaperone phân tử gắn và ổn định protein chưa gấp nếp hoặc đã phần nào gấp nếp do đó
ngăn chặn các protein này kết tụ và phân hủy.
- Chaperonin tạo thành hốc gấp nếp nhỏ. Trong hốc này protein chưa gấp nếp bị cô lập,
mang lại thời gian và môi trường thích hợp để nó gấp nếp chính xác.
Lý do làm nên tầm quan trọng của chaperone là chúng giúp cho protein chưa gấp nếp không bị
kết tụ.
Các protein chưa gấp nếp hoặc mới gấp nếp một phần có khuynh hướng kết tụ thành khối lớn và
thường không hòa tan trong nước, do vậy protein trong những khối này rất khó tách ra để gấp
nếp thành cấu hình chính xác. Sự kết tụ này một phần là do các mạch nhánh kị nước lộ ra khi
chưa kịp vùi vào lõi của protein, những mạch nhánh kị nước lộ ra trên các phân tử khác nhau sẽ
bám vào nhau do hiệu ứng kị nước, do đó thúc đẩy sự kết tụ. Protein mới tổng hợp có nguy cơ bị
kết tụ trước khi hình thành quá trình gấp nếp. Chaperone phân tử gắn với polypeptide đích hoặc
tách nó khỏi các protein chưa gấp nếp hoặc mới gấp nếp một phần, nhờ đó ngăn chặn sự kết tụ
và mang lại thời gian để protein mới sinh gấp nếp chính xác.

Chaperone phân tử. Chaperone phân tử gồm protein sốc nhiệt (Hsp: heat-shock protein). Hsp70
và các thể tương đồng của nó (Hsp70 trong tương bào và chất nền ti thể, BiP trong lưới nội chất,
DnaK của vi khuẩn). Chúng được xác định lần đầu tiên bởi sự xuất hiện nhanh chóng sau khi tế
bào chịu sốc nhiệt. Hsp70 và các thể tương đồng của nó là những chaperone quan trọng trong
mọi sinh vật. Khi gắn với ATP, các protein dạng Hsp70 có cấu hình mở, với các hốc kị nước lộ
ra và gắn tạm thời với vùng kị nước của protein đích chưa gấp nếp. ATP bị thủy phân làm
chaperone phân tử đóng lại và thúc đẩy protein đích gấp nếp trong đó một phần do ngăn các
protein không gấp nếp kết tụ. ATP chuyển hóa thành ADP làm biến đổi cấu hình chaperone và
giải phóng protein đích. Chu trình này được đẩy nhanh hơn bởi protein gọi là đồng chaperone
Hsp40 ở sinh vật nhân chuẩn. Những đồng chaperone này tăng hiệu quả gấp nếp do Hsp70 trung
gian của nhiều protein bằng cách kích thích thủy phân ATP nhờ Hsp70/DnaK. Nhiều chaperone
phân tử được coi là gắn với mọi chuỗi polypeptide mới sinh khi chúng đang tổng hợp từ
ribosome.
Chaperonin. Để gấp nếp chính xác, rất nhiều protein mới tổng hợp cũng cần chaperonin phụ trợ.
Tổ hợp đại phân tử hình trụ lớn này cấu thành từ hai vòng oligomer. Hai vòng này có cấu hình
chặt (tight) gắn peptide và cấu hình lỏng lẻo để giải phóng peptide. Mỗi vòng chaperronin TriC
của sinh vật nhân chuẩn chứa 8 tiểu phần. Cơ chế gấp nhờ GroEL được hiểu kĩ hơn thông qua
TriC và trở thành mô hình chung. Polypeptide gấp nếp một phần hoặc mất nếp gấp gắn vào
khoang của GroEL dạng thùng, nơi nó gắn với thành trong và gấp thành cấu hình tự nhiên. Trong
một bước phụ thuộc ATP, GroEL biến đổi hình dạng và giải phóng protein đã gấp nếp. Một
protein gọi là đồng chaperonin (GroES) trợ giúp cho quá trình này. ATP và GroEL ở trạng thái
chặt làm hốc của nó mở rộng gấp hai lần, chuyển dịch cân bằng sang trạng thái lỏng và giải
phóng peptide. Chú ý rằng thiết kế thùng có nắp của GroEL/GroES rất giống với cấu trúc
proteasome 26S (tham gia phân hủy protein).

Hình 4.12:Protein gấp nếp nhờ Chaperonin. Sự gắp nếp chính xác của một số protein phụ
thuộc vào các chaperonin như GroEL của sinh vật nhân sơ. GroEL là một phức hệ dạng thùng
rỗng cấu thành từ 14 tiểu phần đồng nhất, trọng lượng phân tử khoảng 60000MW, tổ chức thành
hai vòng chồng lên nhau. Khi không gắn ATP hoặc gắn ADP, GroEL mang cấu hình đóng chặt
và gắn với protein không gấp nếp hoặc gấp nếp một phần. ATP gắn vào làm GroEL mở ra, giải
phòng protein đã gấp nếp.
4.6 ĐIỀU HÒA CHỨC NĂNG PROTEIN:
PHẦN 1: PHÂN HỦY PROTEIN
Hầu hết các quá trình trong tế bào xảy ra với tốc độ không đổi và có sự tương tác với nhau chặt
chẽ để tạo ra hiểu quả tổ chức tối đa cho sự sống.
Có 3 cơ chế điều hòa hoạt tính protein, đóng vai trò không thể thiếu trong sự sống và sự phối hợp
hoạt động của protein:
- Tế bào tăng hoạt giảm mức protein tại trạng thái ổn định bằng cách thay đổi tốc độ tổng
hợp, tốc độ phân hủy hoặc cả hai yếu tố đó.
- Tế bào biến đổi hoạt tính nội tại của protein (ví dụ thay đổi ái lực gắn kết cơ chất, thời
gian bất hoạt/hoạt hóa, …)
- Thay đổi vị trí hoặc nồng độ của protein trong tế bào hoặc thay đổi một số phân tử khác
cần cho hoạt tính của protein.

Điều hòa quá trình tổng hợp và phân hủy protein là tính chất cơ bản của tế bào:
- Kiểm soát tổng hợp protein: Tốc độ tổng hợp protein được quyết định bởi tốc độ chuyển
DNA mã hóa cho protein thành mRNA qua quá trình phiên mã cũng như lượng mRNA
hoạt động trong tế bào ở trạng thái ổn định và tốc độ chuyển mRNA thành protein thông
qua quá trình dịch mã.
- Kiểm soát phân hủy protein: Protein nội bào có thời gian tồn tại khác nhau, có thể tính
bằng phút hoặc suốt đời. Thời gian tồn tại của protein được quá trình điều hòa phân hủy
protein kiểm soát.
Có 2 nguyên lý quan trọng đặc biệt trong phân hủy protein:
- Thứ nhất là quá trình phân hủy các protein có độc tính, gấp nếp hoặc lắp ráp sai hay bị hư
hại (sản phẩm đột biến gene, …). Theo ước tính, có 30% protein mới tổng hợp đã nhanh
chóng bị phân hủy do gấp nếp sai hoặc tạo thành một tập hợp có sai phạm trong cấu trúc,
…
- Thứ hai là quá trình phân hủy protein có kiểm soát là cơ chế rất hữu hiệu để duy trì nồng
độ và hoạt tính phụ hợp của các protein, và cho phép thay đổi một cách nhanh chóng hàm
lượng của chúng khi tế bào đáp ứng với các điều kiện nội sinh hay ngoại sinh thay đổi.
Cơ chế tác động của Proteasome và hệ thống Ubiquitin:
Proteasome là một đại phân tử protein có khối lượng 2-2,4 x 106 Da. Chúng có dạng hình trụ với
lõi xúc tác dạng thùng rỗng (proteasome 20S). Một hoặc hai vùng mũ gắn với hai đầu của phần
lõi này giúp điều hòa hoạt tính của proteasome. Protein này rất quan trọng, do vậy có tới 40%
năng lượng tế bào dùng để tổng hợp ra nó.
Dạng proteasome 20S có cấu trúc: Cao khoảng 14,8 nm, đường kính 11,3 n và chứa vùng mũ
điều hòa có kích thước 19S ở hai đầu. Có một số loại phức hợp mũ điều hòa khác nhau. Mũ 19S
có 16-18 tiểu phần protein, 6 trong số đó để thủy phân ATP (chúng có bản chất là ATPase) để
cung cấp năng lượng cho quá trình làm protein cơ chất mất gấp nếp và chuyển có chọn lọc chúng
đến buồng trong của proteasome. Lõi xúc tác của proteasome gồm hai vòng trong và hai vòng
ngoài. Hai vòng trong có 6 vị trí xúc tác quay vào buồng trong có đường kính khoảng 1,7 nm).
Những trung tâm hoạt động này có vai trò phân hủy protein. Hai vòng ngoài đóng vai trò kiểm
soát mức truy cập của cơ chất. Proteasome có thể phân hủy hoàn toàn hầu hết mọi protein vì
chúng có các trung tâm hoạt động với khả năng phân cắt ngay sau các gốc kị nước (vốn tiêu tốn
nhiều năng lượng), các gốc acid và base. Cơ chất là polypeptide phải đi vào buồng theo khe hở
được điều hòa ở trung tâm các vòng ngoài. Trong proteasome 26S, mức mở khe do ATPase
trong vùng mũ 19S kiểm soát. Các sản phẩm peptide ngắn sau khi bị phân hủy dài khoảng 2-24

amino acid thoát khỏi buồng và nhanh chóng bị thủy phân tiếp bởi các peptidase trong bào
tương, cuối cùng chúng hình thành nên từng mono amino acid. (Người ta có thể ứng dụng chất
kiểm hãm chức năng của proteasome để chữa ung thư).
Hình 4.13: Phân hủy protein nhờ ubiquitin và proteasome.(a) Sau khi polyubiquitin hóa, các
protein bị phân hủy trong proteasome. Enzyme E1 hoạt hóa khi gắn với một phân tử Ubiquitin
(Ub) (bước 1). Sau đó E1 chuyển phân tử Ub này tới gốc Cystein của E2 (bước 2). Ubiquitin
ligase (E3) truyền phân tử Ub gắn E2 tới -NH2 trong nhóm R của lysine thuộc protein đích
(bước 3). Các bước 1 và 3 lặp lại dẫn đến nhiều phân tử Ub gắn với protein đích, tạo thành chuỗi

polyubiquitin (bước 4). Vùng mũ của proteasome nhận biết protein đích đã gắn với chuỗi
polyubiquitin và sử dụng năng lượng thủy phân ATP để loại bỏ các nhóm Ub, khử gấp nếp rồi
chuyển protein mất nếp gấp tới buồng phân hủy protein nằm trong lõi. Tại đây, protein bị cắt vụn
thành các đoạn peptide nhỏ và giải phóng ra ngoài (bước 5). (b) Hình ảnh kiến tạo bằng máy tính
cho thấy proteasome có hình trụ chứa mũ 19S (màu xanh da trời) tại mỗi đầu của vùng lõi 20S.
Ubiquitin có chức năng đánh dấu protein trong bào tương cho quá trình phân hủy trong
proteasome. Do đó, proteasome có thể “phân biệt” được giữa protein cần phân hủy và protein
không.
Ubiquitin là một polypeptide dài 76 amino acid, nó là một hệ cảm biến phức tạp quyết định
protein nào sẽ bị phân hủy và sau đó sẽ xảy ra quá trình gắn nhiều phân tử ubiquitin vào phân tử
protein đích. Phần mũ điều hòa 19S của proteasome 26S sau đó nhận biết protein đã gắn
ubiquitin và khử gấp nếp cũng như đưa nó vào phân hủy trong proteasome.
Cơ chế ubiquitin hóa gồm 3 bước như sau:
1. Kích hoạt enzyme hoạt hóa ubiquitin (E1) khi phân tử gắn ubiquitin. Giai đoạn này cần
tiêu tốn ATP.
2. Truyền phân tử ubiquitin này tới gốc cysteine của enzyme liên hợp ubiquitin (E2)
3. Hình thành liên kết peptide giữa đầu C của ubiquitine gắn trên E2 và amino acid trong
nhóm R của lysine ở protein đích. Phản ứng này được xúc tác bởi enzyme ubiquitin-
protein ligase (E3). Các phản ứng sau đó gắn cộng hóa trị thêm nhiều ubiquitin vào mạch
nhánh của lysine trong ubiquitin đã gắn vào trước đó để tạo ra một chuỗi polimer
ubiquitin mạch thẳng.
Quá trình phân hủy này có tính đặc hiệu bởi vì cơ chất của E3 ligase là yếu tố quyết định protein
nào sẽ gắn với ubiquitin. Có tới hàng trăm loại E3 ligase của tế bào đảm nhận việc polyubiquitin
hóa các protein khác nhau. Ngoài ra, phản ứng còn có thể bị đảo ngược bởi một số enzyme khử
ubiquitine.
Ngoài chức năng phân hủy protein đích, việc đánh dấu ubiquitin còn có các chức năng khác như:
- Gắn cộng hóa trị một phân tử ubiquitin vào lysine trên protein đích.
- Gắn một ubiquitin vào nhiều nơi.
- Ubiquitin liên kết với đầu N của protein đích
- Polyubiquitin hóa theo liên kết của ubiquitin với gốc lysine khác thay vì Lysine 48.
Các tác động trên ảnh hưởng đến quá trình vận chuyển protein trong tế bào (ẩm bào), kiểm soát
sửa chữa DNA và điều hòa quá trình phiên mã, …
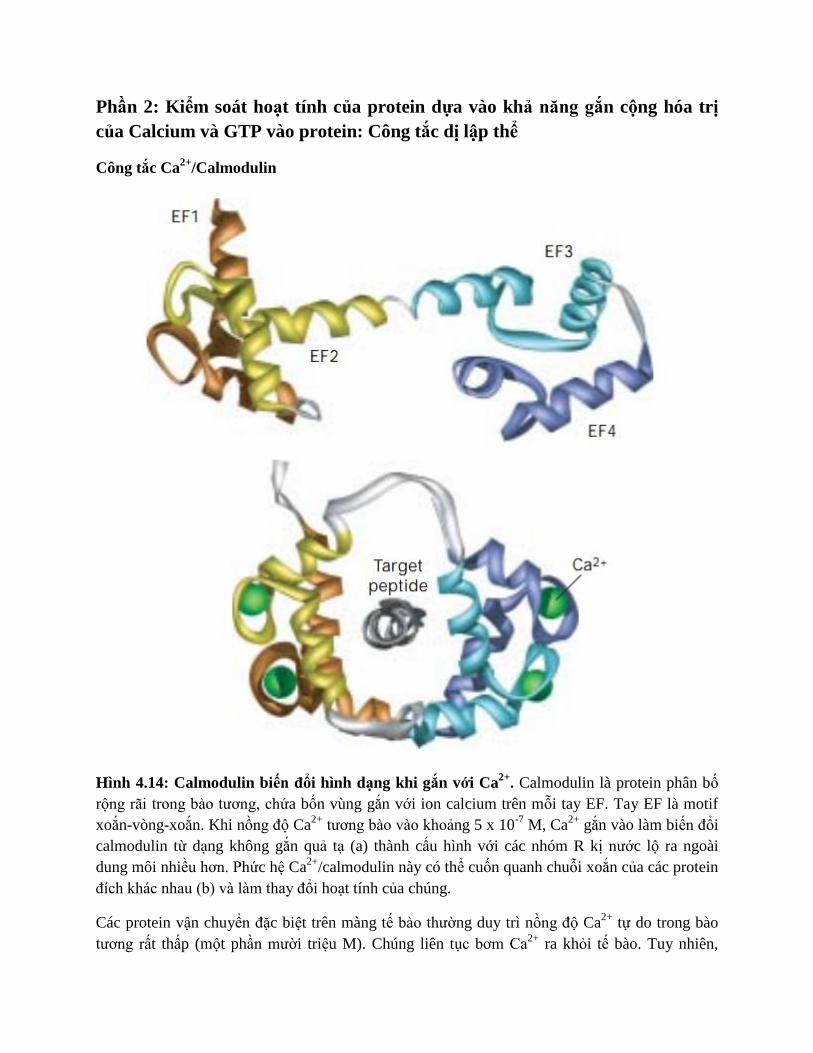
Phần 2: Kiểm soát hoạt tính của protein dựa vào khả năng gắn cộng hóa trị
của Calcium và GTP vào protein: Công tắc dị lập thể
Công tắc Ca2+
/Calmodulin
Hình 4.14: Calmodulin biến đổi hình dạng khi gắn với Ca2+
. Calmodulin là protein phân bố
rộng rãi trong bào tương, chứa bốn vùng gắn với ion calcium trên mỗi tay EF. Tay EF là motif
xoắn-vòng-xoắn. Khi nồng độ Ca2+
tương bào vào khoảng 5 x 10-7
M, Ca2+
gắn vào làm biến đổi
calmodulin từ dạng không gắn quả tạ (a) thành cấu hình với các nhóm R kị nước lộ ra ngoài
dung môi nhiều hơn. Phức hệ Ca2+
/calmodulin này có thể cuốn quanh chuỗi xoắn của các protein
đích khác nhau (b) và làm thay đổi hoạt tính của chúng.
Các protein vận chuyển đặc biệt trên màng tế bào thường duy trì nồng độ Ca2+
tự do trong bào
tương rất thấp (một phần mười triệu M). Chúng liên tục bơm Ca2+
ra khỏi tế bào. Tuy nhiên,

nồng độ Ca2+
trong bào tương sẽ có thể tăng từ 10 đến 100 lần khi các kênh dẫn ion Ca2+
trong
màng tế bào mở ra và cho phép Ca2+
chảy ngược từ ngoài vào trong tế bào. Các protein gắn Ca2+
đặc biệt cảm thụ sự tăng nồng độ Ca2+
ngoại bào đối với hoạt tính của tế bào bằng cách “bật”
hoặc “tắt” các protein khác. Năm 1883, S.Ringer đã chứng minh được sự quan trọng của Ca2+
ngoại bào khi ông phát hiện tim chuột sẽ đập hoàn hảo khi nhúng trong dung dịch NaCl pha
loãng bằng nước cứng giàu Ca2+
nhưng sẽ đập rất yếu và ngừng đập nếu dung dịch chỉ có NaCl
mà thôi. Có nhiều protein gắn Ca2+
chứa motif dạng xoắn – vòng – xoắn/tay EF. Protein tay EF
điển hình là Calmodulin – nằm trong mọi tế bào nhân chuẩn và có thể tồn tại ở dạng protein đa
tiểu phần. Khi Ca2+
gắn vào calmodulin (4 vùng gắn) thì tạo thành phức hệ Ca2+
/calmodulin gắn
vào trình tự bảo tồn của nhiều loại protein đích và do đó bật hoặc tắt hoạt tính của những proteiin
này. Ghi nhận những điều trên, ta thấy rằng calmodulin và các protein tay EF tương tự có chức
năng như những protein công tắc, phối hợp nhịp nhàng với biến thiên nồng độ Ca2+
để điều khiển
hoạt tính của các protein khác.
Guanine nucleotide
Hình 4.15: Công tắc GTPase. GTPase có hoạt tính khi gắn với GTP. GTP bị thủy phân làm bất
hoạt enzyme này. Quá trình được thúc đẩy bởi GAP (protein trợ giúp GTPase) và GRS (yếu tố
điều hòa quá trình truyền tín hiệu protein G); bị kiềm hãm bởi GDI (yếu tố kìm hãm quá trình
phân ly Guanine nucleotide). GEF (yếu tố trao đổi Guanine nucleotide) chuyển hóa GDP thành
GTP do đó tái hoạt hóa Enzyme.
Siêu họ GTPase cũng là một công tắc. Enzyme GTPase với khả năng thủy phân GTP thành GDP,
chúng chứa protein Ras đơn phân và tiểu phần Gα của protein G tam phân. Ras và Gα có thể gắn
với tế bào chất, có chức năng truyền tín hiệu và đóng vai trò quan trọng trong quá trình tăng sinh
cũng như biệt hóa tế bào. Những thành viên khác thuộc họ này thì lại có chức năng tổng hợp

protein, vận chuyển protein giữa nhân và bào tương, tạo các túi lysosome và dung hợp chúng với
màng tế bào tương tác đặc hiệu, tái sắp xếp tế bào actin. Chaperone Hsp70 là một ví dụ điển hình
về công tắc ATP/ADP, có cơ chế tương tự như công tắc GTP/GDP.
Tất cả các protein công tắc GTPase tồn tại 2 dạng:
- Hoạt hóa khi gắn với GTP với khả năng gắn và điều chỉnh hoạt tính của protein đích.
- Bất hoạt khi gắn GDP tạo ra từ sự thủy phân GTP.
Tốc độ hoạt tính của công tắc GTPase quyết định thời gian nó tồn tại ở dạng hoạt hóa. Do đó
hoạt tính của GTP đóng vai trò như đồng hồ hẹn giờ để điều khiển công tắc này. Tế bào chưa
nhiều loại protein với khả năng điều tiết tốc độ nội tại của hoạt tính GTPase trong một công tắc
GTPase nhất định.
Phản ứng Phosphoryl hóa và khử Phosphoryl hóa
Hình 4.16: Điều hòa hoạt tính của protein qua công tắc Kinase/phosphatase. Chu trình
phosphoryl hóa và khử phosphoryl hóa protein là cơ chế chung giúp điều hòa hoạt tính protein
của tế bào. Trong ví dụ này, protein đích (R) có hoạt tính (phía trên) khi bị khử phosphoryl hóa
và bất hoạt (phía dưới) khi bị khử phosphoryl hóa. Lưu ý rằng đa số protein đáp ứng ngược lại
với quá trình tương tác của sự phosphoryl hóa này.
Proten kinase xúc tác phản ứng phosphoryl hóa còn phosphatase có vài trò ngược lại. Do tác
động trái ngược nhau này của hai loại enzyme trên nên nó đã hình thành một công tắc “bật” và
“tắt” nhiều loại protein khác nhau. Nhiều loại protein kinase và phosphatase đặc hiệu cho các
protein đích khác nhau và do đó có thể điều hòa nhiều con đường khác nhau.

4.7 Phân cắt protein
Đây là quá trình mang tính chất không thuận nghịch. Ví dụ như nhiều loại protein (ví dụ như
insulin) được tổng hợp ở trạng thái tiền chất trước rồi khi ra khỏi tế bào, một số liên kết peptide
của chúng cần bị thủy phân để tạo thành một peptide gấp nếp chính xác. Trong một số trường
hợp, polypeptide dài là tiền thân của tiền hormone (preprohormone) có thể bị cắt thành các loại
hormone có hoạt tính. Để không bị phân hủy sai protein trước khi đến ruột non, protease serine
trong tụy được tổng hợp thành dạng zymogen (tiền enzyme) không có hoạt tính. Lên kết peptide
gần đầu N của trypsinogen bị phân cắt tạo ra một gốc đầu N mới là Ile-16. Nhóm amino acid của
Ile-16 này có thể tạo liên kết ion với nhóm carboxyl trên nhóm R của acid aspartic nằm trong
enzyme. Điều này thay đổi cấu hình của enzyme thành trạng thái hoạt hóa với vùng gắn cơ chất
mở ra. Trypsin đã kích hoạt sau đó có thể hoạt hóa trypsinogen, chymotrypsinogen và các tiền
enzyme khác. Tương tự nhưng phức tạp hơn nhiều là tầng protease (một protease kích hoạt tiền
chất đang bất hoạt của protease khác) có thể khuếch đại tín hiệu ban đầu và đóng vai trò quan
trọng đối với một số hệ thống như hệ thống đông máu chẳng hạn. Việc điều hòa cẩn thận các hệ
thống như vậy rõ ràng là rất quan trọng do máu đông không thích hợp có thể làm tắc hệ thống
tuần hoàn gây tử vong hoặc nếu không đủ có thể gây chảy máu không ngừng, …
Một loạt sự ly giải protein hiếm và đáng chú ý được gọi là tự cắt protein, xảy ra ở một số sinh vật
nhân chuẩn và đa số vi khuẩn. Cơ chế tương đối đơn giản và các tính chất như sau:
- Một phân đoạn ở giữa polypeptide bị loại bỏ và hai đầu chuỗi sẽ nối lại.
- Quá trình này là quá trình tự xúc tác, không cần enzyme.
- Peptide bị cắt tự tách khỏi protein theo cơ chế tương tự cơ chế ly giải của một số phân tử
RNA khác.
- Điều đáng lưu ý là sự tự phân cắt phân tử Hedgehog – một phân tử truyền tín hiệu gắn
trên màng rất quan trọng đối với một số quá trình phát triển (tuy nhiên không được đề cập
ở đây do vượt quá phạm vi nghiên cứu).





























