Embed Size (px)
Citation preview

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 1/12
110
Journal of Vertebrate Paleontology 22(1):110–121, March 2002᭧ 2002 by the Society of Vertebrate Paleontology
LOCOMOTOR EVOLUTION IN CAMELS REVISITED: A QUANTITATIVE ANALYSIS OF PEDALANATOMY AND THE ACQUISITION OF THE PACING GAIT
CHRISTINE M. JANIS, JESSICA M. THEODOR*, and BETHANY BOISVERT†
Department of Ecology and Evolutionary Biology, Brown University, Providence, Rhode Island 02912,Christine [email protected]
ABSTRACT—Extant camelids (llamas and camels) are unique among wild mammals in their regular employment of a pacing gait. They also have a unique foot morphology, assumed to be an adaptation for this mode of locomotion:their feet are secondarily digitigrade, with the loss of hooves and the addition of a broad foot pad. We examined 22measurements of the metapodials and phalanges of camelids and ruminants with bivariate and multivariate analyses,including 18 genera of extinct camelids from the Tertiary of North America. Extant camelids and ruminants wereclearly distinguishable from each other. Most extinct camelids showed some morphological features typical of extantforms, five out of eighteen clustered with the extant camelids. Pacing may have evolved independently within thesubfamilies Camelinae and Protolabinae. Additionally, evolutionary change towards a condition resembling that of extant camelids also occurred within the subfamilies Stenomylinae and Miolabinae. These parallel changes in camelidlocomotor anatomy occurred in the late Oligocene or early Miocene, preceding the formation of widespread opengrassland habitats in the late Miocene.
INTRODUCTION
Camelids, represented today by camels and llamas, are fas-cinating and little-studied animals. Camelids are artiodactyls,and they probably represent the extant sister-group to ruminants(Webb and Taylor, 1980), although some molecular studieswould place them in a more basal position within the Artio-dactyla (Gatesy et al., 1999; but see Norris, 1999). Camelidsare unique among artiodactyls in their secondarily digitigradefoot posture, derived from an ancestral unguligrade condition,with a splay-toed foot and the typical ungulate hooves replacedby a broad foot pad. This foot morphology is characterized by
metapodials that are splayed at their distal end (Fig. 1A), andalso by the derived condition of the loss of the interdigital lig-aments, allowing for the divergence of the third and fourth dig-its (Webb, 1972).
Extant camelids are also distinguished from all other wildmammals by their mode of locomotion, employing a true pac-ing gait (i.e., a ‘‘running pace,’’ Hildebrand, 1976). The paceresembles the trot, in which pairs of fore and hind legs aremoved together, with a period of suspension between the move-ment of each fore-/hindlimb pair during which all four feet areoff the ground. However, in the trot, contralateral pairs of foreand hind legs are moved simultaneously (e.g., left fore withright hind), whereas the pace involves ipsilateral pairs of legs(e.g., left fore with left hind). The pace prevents the fore andhind leg on the same side from interfering with each other dur-
ing fast locomotion, allowing a longer stride length, and hencea faster and more efficient mode of locomotion than the trot(Howell, 1944; Webb, 1972; Gauthier-Pilters and Dagg, 1981).Three distinct types of pace gait can be distinguished in extantCamelus: a slow pace, where the animal may be supported forfour legs during some part of the locomotory cycle; a mediumpace, where the animal is only ever supported by two legs; anda fast pace, where there are times during the cycle when nolegs are on the ground (Gauthier-Pilters and Dagg, 1981). The
* Present address: Ilinois State Museum, Research and CollectionsCenter, 1011 East Ash Street, Springfield, Illinois 62703.
† Present address: 265 Blackberry Hill Road, Berwick, Maine 03901.
fast pace is the one that would be termed a ‘‘running pace’’ byHildebrand (1976).
With the exception of some long-legged domestic dogs, andsome breeds of domestic horses (e.g., standardbreds, used inharness races), camelids are the only mammals that use thistype of pacing gait in place of a trot. Some long-legged mam-mals employ a slower, walking pace, also seen in camelids, inwhich there is no period of suspension. The walking pace, usedby some long-legged ungulates (e.g., giraffe and gerenuk) andthe cheetah, is actually a modified fast version of the regularlateral walk of mammals and is not the same as the runningpace of camelids (Hildebrand, 1976). Many long-legged mam-
mals, including camelids, giraffe, and cheetah, also employ alateral ‘‘rotary’’ gallop (involving ipsilateral pairs of limbs be-ing moved in sequence), instead of the transverse gallop (in-volving contralateral pairs of limbs) seen in most mammals.Thus, while camelids are not unique in their use of lateral gaits,they differ from other wild mammals in their use of the runningpace.
Webb (1972) provided an extensive discussion of the advan-tages and disadvantages of the pacing gait in camelids. A primedisadvantage is reduced lateral stability. Webb considered thedevelopment of the broad foot pad and splay-footed digitigradestance to be adaptations to mitigate this instability, and listedother morphological features that would aid in increasing lateralstability. Camelids have a narrow chest with broad, flat ribs.They also have enlarged the areas of attachment for the prox-
imal limb abductors, whose action would prevent the body fromcollapsing towards the unsupported side. These areas include arelatively large scapular spine and acromion process in theshoulder girdle for attachment of the trapezius and deltoid mus-cles; a large deltopectoral crest on the humerus for the insertionof the deltoids; and a broadened greater trochanter of the femurfor the insertion of the gluteals and vastus lateralis. To this listwe add a laterally expanded ridge on the dorsal ilium for theorigin of the tensor fascia latae, and transverse processes of thelumbar vertebrae that are more robust and more horizontal inposition than in ruminants, for the attachment of the longissi-mus dorsi.
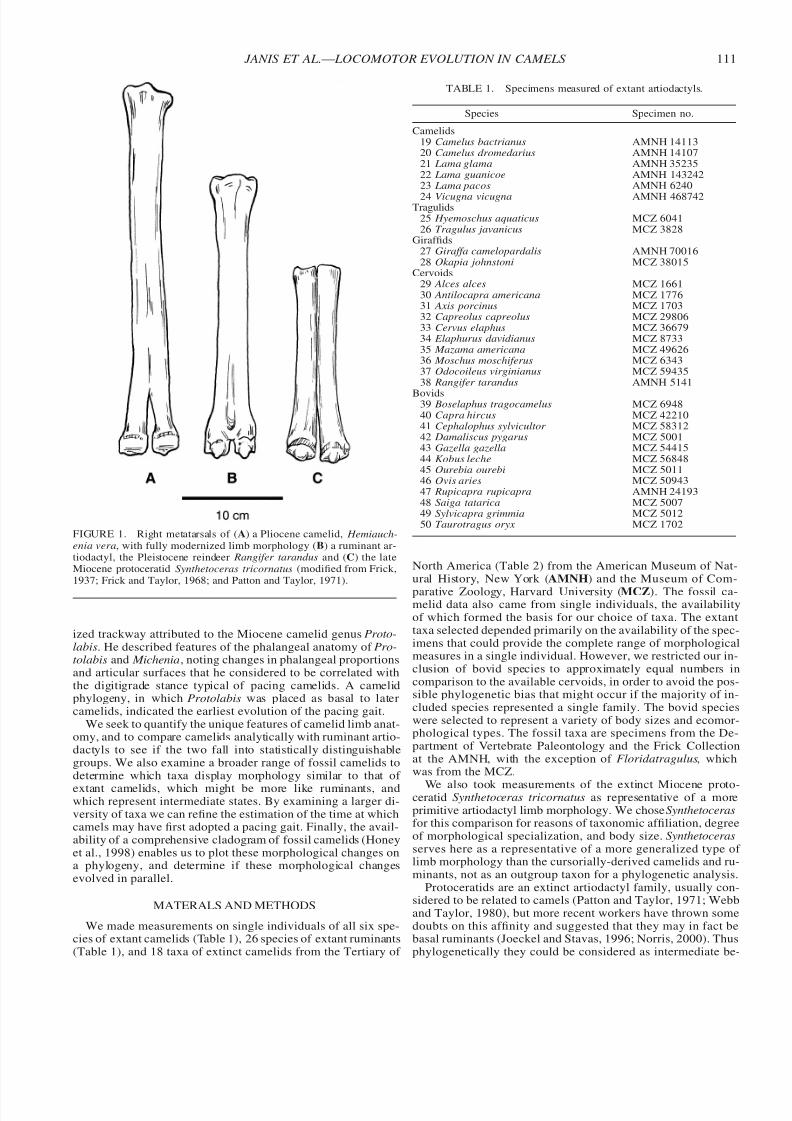
To trace the evolution of the pacing gait in camelids, Webb(1972) used evidence from both limb morphology and a fossil-
7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 2/12
111 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
FIGURE 1. Right metatarsals of (A) a Pliocene camelid, Hemiauch-enia vera, with fully modernized limb morphology (B) a ruminant ar-tiodactyl, the Pleistocene reindeer Rangifer tarandus and (C) the lateMiocene protoceratid Synthetoceras tricornatus (modified from Frick,1937; Frick and Taylor, 1968; and Patton and Taylor, 1971).
TABLE 1. Specimens measured of extant artiodactyls.
Species Specimen no.
Camelids19 Camelus bactrianus20 Camelus dromedarius21 Lama glama22 Lama guanicoe23 Lama pacos
24 Vicugna vicugna
AMNH 14113AMNH 14107AMNH 35235AMNH 143242AMNH 6240
AMNH 468742Tragulids
25 Hyemoschus aquaticus26 Tragulus javanicus
MCZ 6041MCZ 3828
Giraffids27 Giraffa camelopardalis28 Okapia johnstoni
AMNH 70016MCZ 38015
Cervoids29 Alces alces30 Antilocapra americana31 Axis porcinus32 Capreolus capreolus33 Cervus elaphus
MCZ 1661MCZ 1776MCZ 1703MCZ 29806MCZ 36679
34 Elaphurus davidianus35 Mazama americana36 Moschus moschiferus37 Odocoileus virginianus38 Rangifer tarandus
MCZ 8733MCZ 49626MCZ 6343MCZ 59435AMNH 5141
Bovids39 Boselaphus tragocamelus40 Capra hircus41 Cephalophus sylvicultor 42 Damaliscus pygarus43 Gazella gazella44 Kobus leche
MCZ 6948MCZ 42210MCZ 58312MCZ 5001MCZ 54415MCZ 56848
45 Ourebia ourebi46 Ovis aries47 Rupicapra rupicapra48 Saiga tatarica49 Sylvicapra grimmia50 Taurotragus oryx
MCZ 5011MCZ 50943AMNH 24193MCZ 5007MCZ 5012MCZ 1702
ized trackway attributed to the Miocene camelid genus Proto-labis. He described features of the phalangeal anatomy of Pro-tolabis and Michenia, noting changes in phalangeal proportionsand articular surfaces that he considered to be correlated withthe digitigrade stance typical of pacing camelids. A camelidphylogeny, in which Protolabis was placed as basal to latercamelids, indicated the earliest evolution of the pacing gait.
We seek to quantify the unique features of camelid limb anat-omy, and to compare camelids analytically with ruminant artio-dactyls to see if the two fall into statistically distinguishablegroups. We also examine a broader range of fossil camelids todetermine which taxa display morphology similar to that of
extant camelids, which might be more like ruminants, andwhich represent intermediate states. By examining a larger di-versity of taxa we can refine the estimation of the time at whichcamels may have first adopted a pacing gait. Finally, the avail-ability of a comprehensive cladogram of fossil camelids (Honeyet al., 1998) enables us to plot these morphological changes ona phylogeny, and determine if these morphological changesevolved in parallel.
MATERALS AND METHODS
We made measurements on single individuals of all six spe-cies of extant camelids (Table 1), 26 species of extant ruminants(Table 1), and 18 taxa of extinct camelids from the Tertiary of
North America (Table 2) from the American Museum of Nat-ural History, New York (AMNH) and the Museum of Com-
parative Zoology, Harvard University (MCZ). The fossil ca-melid data also came from single individuals, the availabilityof which formed the basis for our choice of taxa. The extanttaxa selected depended primarily on the availability of the spec-imens that could provide the complete range of morphologicalmeasures in a single individual. However, we restricted our in-clusion of bovid species to approximately equal numbers incomparison to the available cervoids, in order to avoid the pos-sible phylogenetic bias that might occur if the majority of in-cluded species represented a single family. The bovid specieswere selected to represent a variety of body sizes and ecomor-phological types. The fossil taxa are specimens from the De-partment of Vertebrate Paleontology and the Frick Collectionat the AMNH, with the exception of Floridatragulus, whichwas from the MCZ.
We also took measurements of the extinct Miocene proto-ceratid Synthetoceras tricornatus as representative of a moreprimitive artiodactyl limb morphology. We chose Synthetocerasfor this comparison for reasons of taxonomic affiliation, degreeof morphological specialization, and body size. Synthetocerasserves here as a representative of a more generalized type of limb morphology than the cursorially-derived camelids and ru-minants, not as an outgroup taxon for a phylogenetic analysis.
Protoceratids are an extinct artiodactyl family, usually con-sidered to be related to camels (Patton and Taylor, 1971; Webband Taylor, 1980), but more recent workers have thrown somedoubts on this affinity and suggested that they may in fact bebasal ruminants (Joeckel and Stavas, 1996; Norris, 2000). Thusphylogenetically they could be considered as intermediate be-

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 3/12
112 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 1, 2002
TABLE 2. Morphological characteristics of extinct camelids. Key: Mets: ϭ metatarsals. C ϭ cameline-like; L ϭ lamine-like; R ϭ ruminant-like; R/L; ϭ morphology in between that of ruminants and lamines; L/C ϭ morphology in between that of lamines and camelines. Note thatHoney et al. (1998) considered ‘‘ Aepycamelus’’ priscus (taxon 14) to belong to a separate genus from other species of Aepycamelus. Met. lengthϭ length of metatarsals in comparison with extant artiodactyls. All specimen numbers refer to AMNH specimens unless otherwise noted. A #beside the specimen no. indicates a field number.
Taxon Specimen no. Mets. fused? Mets. splayed? Met. length PCA Ternary plot
1 Poebrotherium 6520 no no long R RStenomylinae
2 Pseudolabis3 Miotylopus4 Stenomylus
419423644614226
partialfullymostly
slightslightslight/mod
mediumlonglong
R/LR/LR/L
RRR/L
(Not assigned to subfamily)5 Floridatragulus6 Gentilicamelus
MCZ 77847510
nopartial
slightno
shortmedium
RR/L
R/LR/L
Miolabinae7 Paramiolabis8 Miolabis9 Nothotylopus
652-39#2419968554
partialnopartial
slightslightno
mediummediummedium
R/LR/LL
LRL/C
Protolabinae10 Tanymykter 11 Michenia12 Protolabis
3658439628400-2988#
mostlymostlyfully
no/slightmod/fullyfully
mediumlongvery long
LLL
R/LLL
(Not assigned to subfamily)13 Oxydactylus 17620 partial no long L R
Camelinae14 ‘‘ A.’’ priscus 14188 fully mod/fully long C R/L
Camelinae (Lamini)15 Aepycamelus16 Hemiauchenia
910938179
fullyfully
fullyfully
very longvery long
CC
CL
Camelinae (Camelini)17 Procamelus18 Titanotylopus
7500310703
fullyfully
fullyfully
mediumlong
CC
LL/C
tween camelids and ruminants. Protoceratids are derived cra-nially, and later forms (synthetoceratines) had characteristic‘‘sling-shot’’ nasal horns. However, postcranially they retainprimitive features such as short, unfused metapodials with in-complete distal metapodial keels.
It might seem that a more basal taxon would be preferableto determine primitive artiodactyl limb anatomy, such as theEocene Diacodexis, or an oreodont. However, the problem with
both of these taxa is that they are small animals, and largeranimals such as the camelids and ruminants under study mightbe expected to have differences in their limb anatomy simplybecause of their larger size, and further, they retains full-sizedlateral digits, a primitive condition not found in any of the taxain our analysis. Synthetoceras is of comparable size to the ca-melid and ruminant taxa compared in Figures 1 and 2 (bodymass estimated at around 150 to 200 kg). Some suids are fairlylarge and retain a relatively primitive postcranial anatomy, andthe same is also true for some extinct oreodonts. But theseartiodactyls are more distantly related to camels and ruminantsthan protoceratids, and might be expected to have their ownanatomical specializations, making them less appropriate forcomparison.
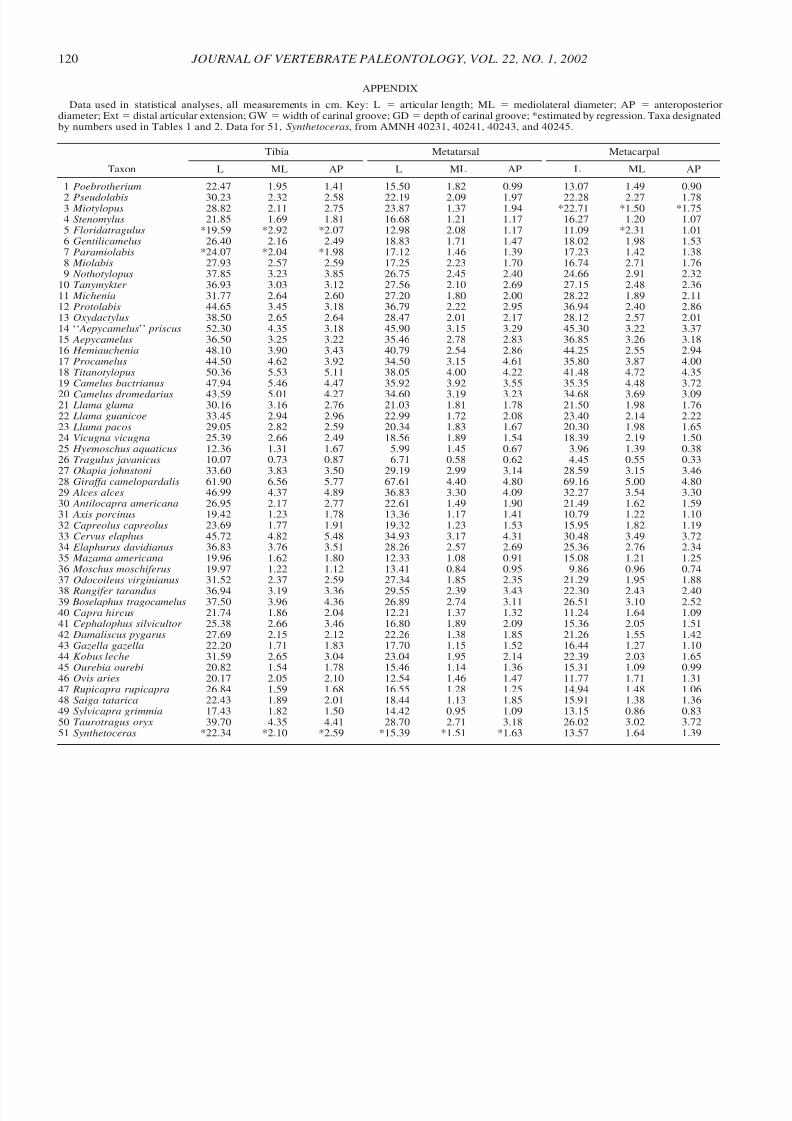
The following measurements were taken on all specimens,
as allowed by preservation. Measurements were taken of thelengths (maximum articular length) and the diameter (at themidshaft, of both anteroposterior and mediolateral dimensions)of the tibia, metatarsal, and metacarpal, and of the length anddiameters of the three hindfoot phalanges. Additional measure-ments were taken on articular surfaces of the phalanges. Theseincluded the extension onto the volar surface of the distal ar-ticular surface of the proximal and medial phalanges (Fig. 2.1and 2.3) and the width and the depth of the carinal groove onthe proximal articular surface of the proximal phalanx (Fig.2.2). All measurements are listed in the appendix. In the fossiltaxa we also noted whether or not the metapodials were fused,and if they had splayed distal ends, as in extant camelids (Table2).
A principal components analysis was conducted using the 22morphological variables described above. Two principal com-ponents were extracted using a Varimax rotation as imple-mented in StatView 5.0. Missing data points for Synthetoceras,
Miotylopus, Paramiolabis and Floridatragulus were estimatedusing regression equations (Appendix), while Tragulus was ex-cluded because of two missing data points. To visualize differ-ences in foot posture and proportions we used a ternary diagram
to examine the proportions of the phalanges, following themethods outlined by Gatesy and Middleton (1997:310). Wemade no attempt to collect data for all extant ruminant taxa,nor have we corrected our data for phylogenetic biases.
RESULTS
General Features of Extant Camelid Foot Morphology
Figures 1 and 2 show some general differences in foot mor-phology between camelids, ruminants, and protoceratids.
Metapodial Morphology The primitive condition is forrelatively short metapodials. The primitive tragulid ruminant
Hyemoschus has metatarsals that are approximately six timeslonger than their average mid-shaft width. Metapodials are lon-ger in all extant ruminants and camelids, although certain mon-
tane bovids have secondarily shortened metapodials (Scott,1985). Extant Rangifer and Lama (the llama) both have meta-tarsals that are about eleven times as long as their width. Notethat metapodial length tends to scale with negative isometry(Bertram and Biewener, 1990), except in Giraffa (the giraffe)which has exceedingly long metapodials, with metatarsalsaround twenty times as long as their width.
Other primitive character states, seen in both Hyemoschusand Synthetoceras, are as follows: small ‘‘side toes’’ (short,complete or partial digits 2 and 5) are present; metapodials 3and 4 are unfused and not splayed at their distal ends; and themetapodials lack complete distal keels. Extant camelids and ru-minants are both derived with respect to these conditions, butin different ways. Both have fused metapodials; all extant ca-

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 4/12
113 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
FIGURE 2. Phalangeal morphology of (A) a camelid, (B) a ruminantand (C) a protoceratid. All phalanges are right pedal phalanges, all scalebars are 1 cm. Taxa are the same as Figure 1. (1) Anterior (volar) viewof proximal phalanx. (2) Posterior (plantar) view of proximal phalanx.(3) Anterior view of medial phalanx. (4) Anterior view of ungual pha-lanx. (5) Lateral view of ungual phalanx.
melids and almost all fossil taxa retain only digits 3 and 4(Honey et al., 1998), whereas small side toes may be retainedin the Cervidae and Moschidae, but are lost in members of other extant ruminants. Ruminants are derived in possessingcomplete distal metapodial keels that act to lock the foot in theunguligrade position (Webb and Taylor, 1980; Janis and Scott,1987). Camelids lack complete keels, but have the derived fea-ture of distally splayed metapodials (Fig. 1; Webb, 1972).
Proximal Phalanx Morphology Camelids have a relative-ly long proximal phalanx, with a length of about five times thatof the diameter. The relative length in ruminants is shorter, witha length around four times that of the diameter (Fig. 2.1). The
condition in Synthetoceras resembles the ruminant one: thus,the camelid condition is likely derived.
In camelids, the distal articular surface of the proximal pha-lanx is extended up onto the volar surface (Fig. 2.1). This ex-tension is not found in ruminants or in Synthetoceras, so thecamelid condition is likely the derived one. The extent of thedistal articular surface (not figured) is also greater on the plantarside in camelids than in Synthetoceras, but in ruminants it ac-tually appears to be less extended than in Synthetoceras. Thus,in this morphology both camelids and ruminants appear to bemodified from the probable primitive condition. The camelidmorphology would allow for a greater degree of extension of the medial phalanx on the proximal one in association with adigitigrade foot posture. In contrast, the restriction of the artic-
ular surface in ruminants would serve to limit mobility betweenthe proximal and medial phalanges, perhaps in association witha more derived unguligrade foot posture.
The carinal groove on the posterior border of the proximalarticulation of the proximal phalanx in camelids is relativelywider and shallower than in ruminants and Synthetoceras (Fig.2.2). Additionally, the proximal articular surface of camelids isrelatively smooth, with little surface relief, and is broader in
the mediolateral direction than in the anteroposterior one. Syn-thetoceras has a relatively deep carinal groove, and a square-shaped articular surface with higher relief. If Synthetoceras rep-resents the more primitive condition, the derived camelid con-dition could be interpreted as reflecting a less stabilized meta-podial-phalangeal joint in conjunction with a digitigrade footposture. The ruminant condition differs from the camelid onein the opposite direction from that of Synthetoceras. The artic-ular surface is longer anteroposteriorly, the carinal groove isdeepened, presumably in association with the complete meta-podial keels, and the articular surface is deeper, allowing formore interlocking of the joint.
Medial Phalanx Morphology The medial phalanx of cam-elids is relatively shorter in comparison to the width (about twoand a half as long as it is wide) than in ruminants (about threetimes as long as wide; Fig. 2.3). However, the medial phalanxof Synthetoceras is also short, so this may represent the prim-itive condition. The distal articular surface has only a slightextension onto the volar surface in both camelids and Synthe-toceras, whereas in ruminants this surface is extended (Fig.2.3). The derived ruminant condition is presumably related toa greater extension of the ungual phalanx on the medial onewith an unguligrade foot posture that is more highly stabilizedthan in Synthetoceras.
The proximal articular surface of the medial phalanx (notfigured) is longer in the mediolateral direction than in the an-teroposterior direction in camelids, with little surface grooving.The articular surface is more square in shape and more deeplydepressed in both Synthetoceras and ruminants, with extensiverelief in the ruminant condition. The derived condition in ca-melids probably represents greater mobility between proximal
and medial phalanges.Ungual Phalanx Morphology The ungual phalanx of ca-
melids is relatively short and flat, with a very shallow, relativelyflat articular surface. In ruminants, the phalanx is long, distallypointed, and high in relief, with extensive depressions on thearticular surface (Fig. 2.4, 2.5). The morphology of the ungualphalanx in Synthetoceras is similar to the ruminant condition,but with less extensive relief on the articular surface. The dif-ferences between camelids and ruminants would again reflectfoot posture. The longer and deeper ungual phalanx of rumi-nants is encased within a hoof, and the deep relief on the artic-ular surface reflects greater restriction of interphalangeal mo-bility with an unguligrade foot posture.
Relative Metapodial Lengths
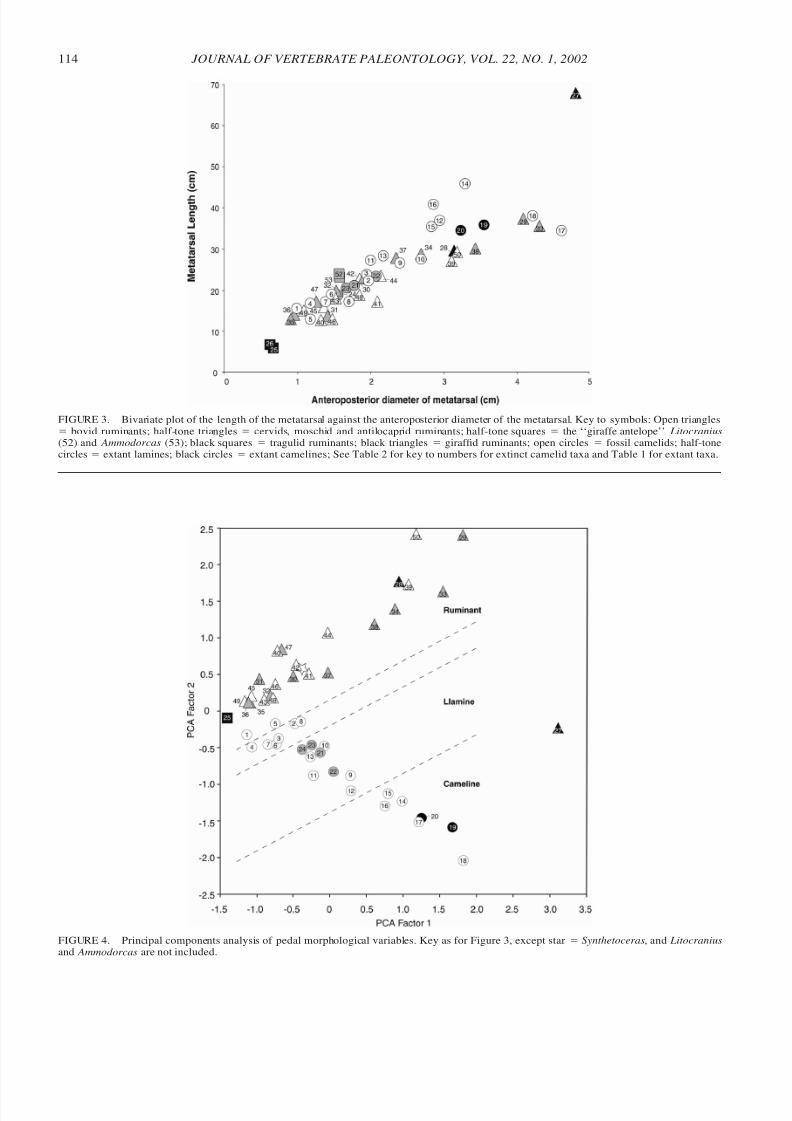
Figure 3 shows a plot of metatarsal length against the an-teroposterior diameter of the metatarsal, including measure-ments for the long-legged, so-called ‘‘giraffe antelope,’’ the ge-renuk, Litocranius walleri, and the dibatag, Ammodorcas clar-kei, (taken from Scott, 1985). Metatarsal diameter was chosenas a proxy of body size, because the diameters of distal limbbones provide better estimates of body mass in artiodactyls thanlength measurements (Scott, 1990). Additionally, the mass of extant camelids can be estimated reliably from the correlationsof limb diameters with body mass derived from ruminants(Scott, 1990). However, metatarsal diameter does not here pro-vide a perfect proxy of size: for example, as shown in Figure3, the metatarsal of Giraffa has a greater diameter than that of
7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 5/12
114 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 1, 2002
FIGURE 3. Bivariate plot of the length of the metatarsal against the anteroposterior diameter of the metatarsal. Key to symbols: Open triangles
ϭ bovid ruminants; half-tone triangles ϭ cervids, moschid and antilocaprid ruminants; half-tone squares ϭ the ‘‘giraffe antelope’’ Litocranius(52) and Ammodorcas (53); black squares ϭ tragulid ruminants; black triangles ϭ giraffid ruminants; open circles ϭ fossil camelids; half-tonecircles ϭ extant lamines; black circles ϭ extant camelines; See Table 2 for key to numbers for extinct camelid taxa and Table 1 for extant taxa.
FIGURE 4. Principal components analysis of pedal morphological variables. Key as for Figure 3, except star ϭ Synthetoceras, and Litocraniusand Ammodorcas are not included.

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 6/12
115 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
TABLE 3. Factor loadings for variables used in the principal com-ponents analysis.
Variable
Factor loading
Factor 1 Factor 2
Tibia lengthTibia diameter, mediolateralTibia diameter, anteroposterior
Metatarsal lengthMetatarsal diameter, mediolateralMetatarsal diameter, anteroposterior
0.9310.9670.950
0.9000.9500.954
Ϫ0.196Ϫ0.146
0.044Ϫ
0.202Ϫ0.142Ϫ0.019
Metacarpal lengthMetacarpal diameter, mediolateralMetacarpal diameter, anteroposteriorProximal phalanx lengthProximal phalanx diameter, mediolateralProximal phalanx diameter, anteroposterior
0.8720.9400.9570.9050.9570.973
Ϫ0.300Ϫ0.181Ϫ0.128Ϫ0.344
0.1230.093
Proximal phalanx distal articular extensionProximal phalanx carinal groove depthProximal phalanx carinal groove widthMedial phalanx lengthMedial phalanx diameter, mediolateral
0.7460.7320.8820.9060.949
Ϫ0.4180.309Ϫ0.334Ϫ0.016Ϫ0.027
Medial phalanx diameter, anteroposteriorMedial phalanx distal articular extensionUngual phalanx lengthUngual phalanx diameter, mediolateral
Ungual phalanx diameter, anteroposterior
0.9040.8020.5470.879
0.765
0.3620.4790.7370.155
0.581
the extinct camelid Titanotylopus, but other skeletal elementsindicate that the latter is the larger taxon.
In comparing extant forms, it can be observed that Giraffais highly aberrant in its extremely long metatarsals, but the re-lated Okapia (the okapi) clusters with the other ruminants. Ad-ditionally, note that camelids are relatively long-legged in com-parison with most ruminants, especially at larger body sizes,although they do not fall completely outside of the range of
extant ruminants. The two species of Camelus are long-leggedin comparison with the majority of ruminants of a similar size(i.e., body mass of around 500 kg), but are of comparable pro-portions to the moose, Alces.
Many of the extinct camelids are longer-legged than extantcamels, with extremely long metapodials in comparison withany living ruminant except Giraffa. This is especially true inthe size range corresponding to body masses of around 200–500 kg. These extinct camelids include not only those taxa tra-ditionally considered as ‘‘giraffe camels,’’ such as Aepycame-lus, but also the taxa Hemiauchenia and Protolabis. The smallercamels, mainly Eocene or Oligocene in age, cluster with theextant ruminants and lamines. The larger ones (extinct membersof the tribe Camelini) have metatarsals of similar length to Ca-melus and Alces.
Our data on Protolabis shows it to have relatively long me-tapodials, although Honey et al. (1998) claimed that the meta-podials are not very long in this genus, only equal (or less than)in length to the basal length of the skull. However, Honey etal. (1998) also stated that the metapodials of Protolabis arerelatively shorter in more derived species. Our specimen is afairly early one, from the early Hemingfordian Running WaterFormation, so perhaps this specimen is primitive in its longmetatarsals.
Figure 3 also illustrates possible differences in scaling ex-ponents of the metatarsals between camelids and ruminants.Scott (1985) noted a ‘‘breakpoint’’ in the scaling of metatarsallengths in extant bovids: metatarsal length scales allometricallybelow a body size of around 200 kg, but at greater mass thereis little increase in absolute metatarsal length. This breakpointin limb length scaling has been attributed to the physical de-
mands of support at larger body size (Bertram and Biewener,1990). Note that in Figure 3 the ‘‘breakpoint’’ occurs at a meta-tarsal width of about 2.8 cm; ruminants with metapodials of this size are Elaphurus davidianus and Cervus elaphus, whichboth have body masses of around 200 kg.
The members of the tribe Camelini (Camelus, Procamelus,and Titanotylopus) appear to obey the same scaling laws thatapply to the majority of ruminants. Their metatarsals are aroundthe same absolute length as smaller taxa such as Protolabis.However, metatarsal length in other large extinct taxa, princi-pally the members of the tribe Lamini, Aepycamelus and Hem-iauchenia, have metatarsal lengths that appear to fall along acontinuation of the scaling exponent that applies to the smallerartiodactyls, as does Giraffa.
Why the giraffe and some larger camelids should be exempt
from the scaling rules that appear to affect ruminants and mem-bers of the tribe Camelini is not clear. Additionally, preliminarydata suggest that, unlike any ruminant or camelid, Giraffa isunique in having a metatarsal that is longer than the tibia.
Principal Components Analysis
Figure 4 shows the results of the principal component anal-ysis using the dimensions described in the Materials and Meth-ods section. The first principal component (PC1) accounts for78.5% of the variance. The loadings on PC1 are all positive,and almost all are close to unity (Table 3). Thus this axis clearlydiscriminates taxa based on body size. Some shape influence isinvolved, as seen in the slightly lower loading for the length of
the ungual phalanx (Table 3), which is the variable that sepa-rates Giraffa from the other taxa on this axis. This results inGiraffa scoring more positively on this axis than the much larg-er extinct camelid Titanotylopus. When Giraffa is removedfrom the analysis, the loading of the ungual phalanx lengthvariable on PC1 is similar to the other variables, and the othertaxa are more evenly spread out along PC1 in accordance withtheir body masses.
The second principal component explains 9.3% of the vari-ance. This axis clearly separates the camelids (negative load-ings) from the ruminants (positive loadings). Giraffa has nearzero loadings on PC2, similar to the tragulids, while Okapia
clusters with the other ruminants. PC2 also contains a size com-ponent, in that larger ruminants (except Giraffa ) have higherpositive loadings, and larger camelids have higher negativeloadings. This may reflect greater locomotor specializations inlarger taxa, or somehow reflect a size component in PC2.
Variables with high positive values on PC2 (i.e., ruminantfeatures) are as follows: the length and anteroposterior diameterof the ungual phalanx, the anteroposterior diameter of the me-dial phalanx, the length of the volar extension of the distalarticular surface on the medial phalanx, and the depth of thecarinal groove on the proximal phalanx. As previously dis-cussed, these articular features reflect the limitation of inter-phalangeal mobility, and the large ungual phalanx reflects itsenclosure within a hoof.
Variables with high negative values (i.e., camelid features)
are as follows: the length of the volar extension of the distalarticular surface on the proximal phalanx, the length of theproximal phalanx, the width of the carinal groove on the prox-imal phalanx, and the length of the metacarpal. These articularfeatures reflect increased interphalangeal mobility. The lengthof the proximal phalanx may be related to a general elongationof the limb proximal to the foot pad.
Thus PC2 appears to be primarily an axis of digitigrady (neg-ative loadings) versus unguligrady (positive loadings). Thelength of the metacarpal (high negative values on PC2) reflectsthe fact that all living and extinct camelids have fore- and hindlimbs of approximately equal length. In ruminants, equally-pro-portioned forelimbs and hind limbs are seen only in animalsliving in more open habitats, that make habitual use of the trot

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 7/12
116 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 1, 2002
FIGURE 5. (A) Ternary plot of phalangeal lengths. (B) Close up of ternary plot. Key as for Figure 4.
gait, instead of using the bound or gallop for speeds faster thana walk (Scott, 1985). In contrast, animals living in closed hab-itats have forelimbs that are shorter than the hind limbs (Scott,1985).
Among the fossil camelids, Poebrotherium and Floridatra-gulus fall fairly close to the ruminants, although with slightlyless positive scores. Stenomylus, Gentilicamelus , Miolabis,
Miotylopus, Paramiolabis, and Pseudolabis fall in the mor-
phospace between ruminants and lamines. Michenia, Oxydac-tylus, Nothotylopus, Protolabis, and Tanymykter cluster withextant lamines. Aepycamelus, ‘‘ Aepycamelus’’ priscus, Hem-iauchenia, Procamelus, and Titanotylopus cluster with extantcamelines (Table 2).
The third principal component (not shown) explains 3.4% of the variance. This component appears to be a ‘‘giraffe’’ axis,separating Giraffa from the other taxa. The variables with highloadings on PC3 are long metapodials, a short ungual phalanx,a short extension to the distal articular surface on the proximalphalanx, and a wide and deep carinal groove on the proximalphalanx. The separation of the giraffe from other taxa appearsto be a result of its similarity to extant camelids in limb length,but with an unguligrade foot posture and the anomalous con-dition (for unguligrady) of a short ungual phalanx.
Ternary Diagram
The ternary diagram of phalangeal lengths clearly separatesout all extant camelids from all ruminants (with the exceptionof Giraffa, which here ‘‘out-camels’’ Camelus!; Fig. 5). Ca-melids score higher on length of the proximal phalanx and low-er on the other two axes (lengths of medial and ungual phalan-ges, respectively) than ruminants. As with the PCA, Okapiaclusters with other ruminants, towards the high end of the ru-minant range, while Hyemoschus falls within the low end of the ruminant range, as do Moschus and Saiga in this analysis.Synthetoceras clusters within the ruminants. Again, as with thePCA, the extant camelines fall further from the ruminants thando the extant lamines.
In this analysis, the following fossil taxa cluster with the
ruminants: Miolabis, Miotylopus, Oxydactylus, Poebrotherium,and Pseudolabis (although Miotylopus and Oxydactylus are sit-uated at the edge of the ruminant morphospace). The followingtaxa fall in the morphospace between ruminants and lamines:‘‘ Aepycamelus’’ priscus, Floridatragulus, Gentilicamelus , Sten-omylus, and Tanymykter . Finally, Hemiauchenia, Michenia,
Nothotylopus, Paramiolabis, Procamelus, and Protolabis clus-ter with lamines, and Aepycamelus clusters with camelines (Ta-ble 2). As in the PCA (Fig. 4), Titanotylopus again is morederived along the camelid trend in morphospace than any extantform, here falling close to the giraffe.
DISCUSSION
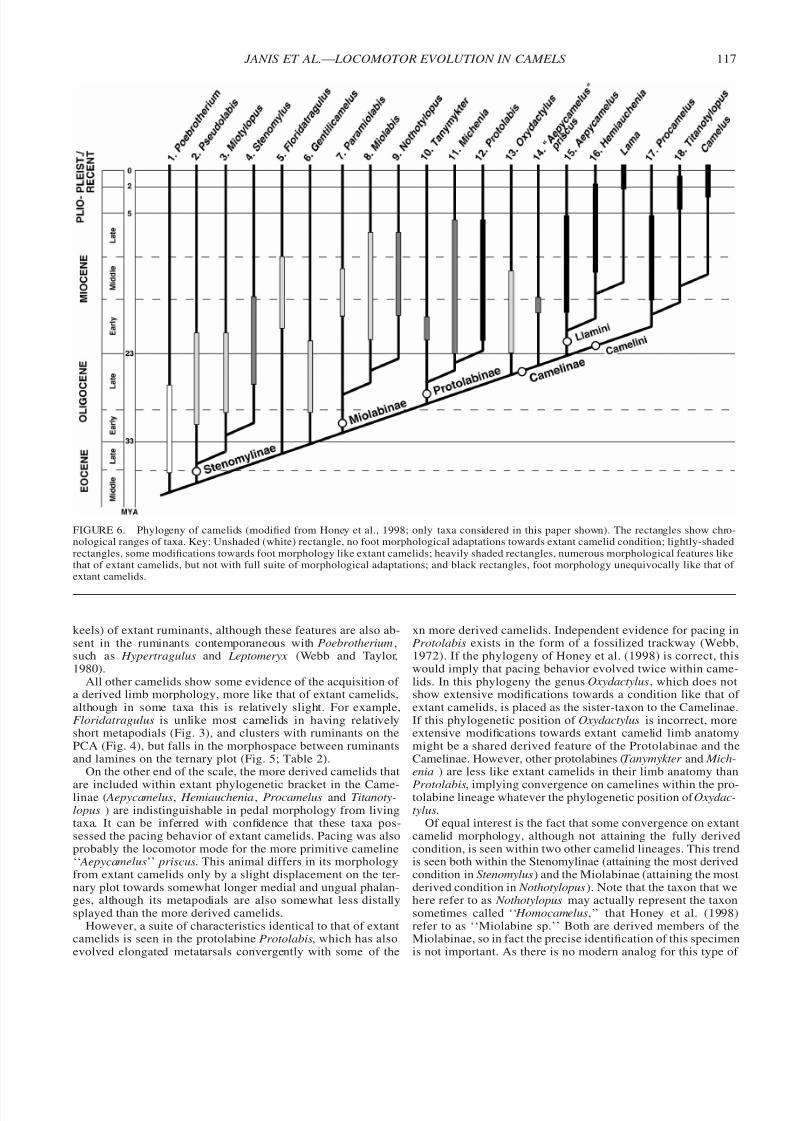
Figure 6 illustrates the only comprehensive phylogeny of ca-melids, and additionally shows the chronological range of these
taxa (both from Honey et al., 1998). The fossil taxa have beenidentified in terms of varying degrees of foot posture that re-sembles the condition in extant camelids (see also Table 2).
The most primitive camelid, Poebrotherium, shows no evi-dence of limb features like those of extant camelids, clusteringwith the ruminants in every analysis. Note that while it tendsto fall close to less derived ruminants such as the tragulids onthe PCA and the ternary plot, its metatarsals are relatively long,even in comparison with similar-sized ruminants (Fig. 3). Thus,if Poebrotherium can be taken as the primitive camelid condi-tion, we can see that the original camelid morphological designwas for moderately long legs with an unguligrade limb posture.But primitive camelids lack the additional cursorial limb spe-cializations (fused metapodials, complete distal metapodial
7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 8/12
117 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
FIGURE 6. Phylogeny of camelids (modified from Honey et al., 1998; only taxa considered in this paper shown). The rectangles show chro-nological ranges of taxa. Key: Unshaded (white) rectangle, no foot morphological adaptations towards extant camelid condition; lightly-shadedrectangles, some modifications towards foot morphology like extant camelids; heavily shaded rectangles, numerous morphological features likethat of extant camelids, but not with full suite of morphological adaptations; and black rectangles, foot morphology unequivocally like that of extant camelids.
keels) of extant ruminants, although these features are also ab-sent in the ruminants contemporaneous with Poebrotherium,such as Hypertragulus and Leptomeryx (Webb and Taylor,1980).
All other camelids show some evidence of the acquisition of a derived limb morphology, more like that of extant camelids,although in some taxa this is relatively slight. For example,Floridatragulus is unlike most camelids in having relativelyshort metapodials (Fig. 3), and clusters with ruminants on thePCA (Fig. 4), but falls in the morphospace between ruminantsand lamines on the ternary plot (Fig. 5; Table 2).
On the other end of the scale, the more derived camelids that
are included within extant phylogenetic bracket in the Came-linae ( Aepycamelus, Hemiauchenia, Procamelus and Titanoty-lopus ) are indistinguishable in pedal morphology from livingtaxa. It can be inferred with confidence that these taxa pos-sessed the pacing behavior of extant camelids. Pacing was alsoprobably the locomotor mode for the more primitive cameline‘‘ Aepycamelus’’ priscus. This animal differs in its morphologyfrom extant camelids only by a slight displacement on the ter-nary plot towards somewhat longer medial and ungual phalan-ges, although its metapodials are also somewhat less distallysplayed than the more derived camelids.
However, a suite of characteristics identical to that of extantcamelids is seen in the protolabine Protolabis, which has alsoevolved elongated metatarsals convergently with some of the
xn more derived camelids. Independent evidence for pacing inProtolabis exists in the form of a fossilized trackway (Webb,1972). If the phylogeny of Honey et al. (1998) is correct, thiswould imply that pacing behavior evolved twice within came-lids. In this phylogeny the genus Oxydactylus, which does notshow extensive modifications towards a condition like that of extant camelids, is placed as the sister-taxon to the Camelinae.If this phylogenetic position of Oxydactylus is incorrect, moreextensive modifications towards extant camelid limb anatomymight be a shared derived feature of the Protolabinae and theCamelinae. However, other protolabines (Tanymykter and Mich-enia ) are less like extant camelids in their limb anatomy than
Protolabis, implying convergence on camelines within the pro-tolabine lineage whatever the phylogenetic position of Oxydac-tylus.
Of equal interest is the fact that some convergence on extantcamelid morphology, although not attaining the fully derivedcondition, is seen within two other camelid lineages. This trendis seen both within the Stenomylinae (attaining the most derivedcondition in Stenomylus) and the Miolabinae (attaining the mostderived condition in Nothotylopus). Note that the taxon that wehere refer to as Nothotylopus may actually represent the taxonsometimes called ‘‘ Homocamelus,’’ that Honey et al. (1998)refer to as ‘‘Miolabine sp.’’ Both are derived members of theMiolabinae, so in fact the precise identification of this specimenis not important. As there is no modern analog for this type of

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 9/12
118 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 1, 2002
intermediate morphology, without associated trackways it is im-possible to determine what the actual gait of these animalsmight have been.
Stenomylus and Nothotylopus both lack the splaying of thedistal ends of the metapodials seen in more derived camelids,and thus it seems unlikely that they had a digitigrade foot pos-ture. Yet, on both the PCA and ternary plots, Stenomylus fallsin the morphospace between the ruminants and lamines and
Nothotylopus clusters with the lamines, indicative of changesin relative proportions of the foot bones in both taxa. Notho-tylopus is more derived in its pedal anatomy than Stenomylus,but Stenomylus is more derived than the other stenomylines,Pseudolabis and Miotylopus.
We also note that Stenomylus possesses other skeletal fea-tures like those of extant camelids that may be indicative of lateral stabilization, such as broad, flat ribs and an expandediliac crest. We were unable to determine the condition of thesefeatures in Nothotylopus. In contrast, the primitive camelid Poe-brotherium, which possesses a ruminant-like foot morphology,shows little evidence of features promoting lateral stability.Poebrotherium lacks an expanded iliac crest, and has only aslight broadening of the anteriormost ribs (nos. 1–7, as seen onthe mounted specimen in the American Museum of NaturalHistory; Scott, 1940:630–635). Perhaps these camelids of ‘‘in-termediate’’ morphology used some types of lateral gaits (forexample, the slow pace or lateral walk and the rotary gallop,as employed by present-day long-legged ruminants), but did nothave the running pace gait of extant camelids.
When Webb (1972) considered the evolution of pacing lo-comotion in camelids, he noted that the pacing trackways of Protolabis were middle Miocene (Barstovian) in age, and cor-related the locomotor shift with the spread of open grasslandhabitats in North America. He pointed out that, in extant cam-elids, the pacing gait is adapted for long distance travel in openterrain, and interpreted the fossil record as supportive of thehypothesis that pacing first evolved along with the open habitatgrasslands.
Figure 6 shows the chronological ranges of camelid taxaalong with their phylogeny, and it is apparent that the trend
towards a more derived morphology resembling that of extantcamelids occurred at an earlier date than proposed by Webb.Consider those taxa where partial modifications are attained.Stenomylus (Stenomylinae) first appeared in the late Oligocene,and Nothotylopus (Miolabinae) and Tanymykter and Michenia(Protolabinae) in the early Miocene. Furthermore, some taxapossessing modifications fully like those of extant camelids,Protolabis (Protolabinae) and Aepycamelus (Camelinae) alsofirst appeared in the early Miocene. Thus, both taxa with ad-aptations indicating a fully-derived pacing gait, and those withsome sort of modifications leading towards this gait, made theirappearance before the spread of open savanna or grasslands(Jacobs et al., 1999). The significance of this observation isunclear. One possible explanation is that more open habitatswere present earlier in North America than previously consid-
ered, as also suggested by other workers (Webb, 1977; Janis,1982; Leopold et al., 1992), although such habitats were notdominated by grass until later in the Miocene.
Another interesting feature of this study is the observationof how different Giraffa is from other extant ruminants. Wedocument here the convergences with camelids in certain as-pects of foot morphology; but the giraffe also has other peculiaraspects of postcranial morphology, some convergent with theextant camelid condition and some unique. The description of giraffe morphology below is based on general osteological ob-servations rather than on quantitative analysis, but neverthelessserves to highlight features that are deserving of further study.
The giraffe resembles extant camelids in having an elongatedand laterally projecting iliac crest, which presumably also
serves for the origin of an enlarged tensor fascia latae to aid inlateral stability, although the giraffe lacks the camelid featurein the scapula of a relatively large scapular spine and acromionprocess. This morphology of the ilium is not simply related tolarge size, because it is absent in other large ruminants such as
Alces and Bison. The giraffe also resembles extant camelids inhaving a relatively narrow chest, but lacks the camelid-likebroad, flattened ribs.
The giraffe is unique among all ruminants and camelids, in-cluding the extinct ‘‘giraffe camels’’ with highly elongatedlimbs, in having metapodials that are longer than the tibia. Ad-ditionally, there appears to be a peculiar morphology of thepedal sesamoid bones observed on mounted specimens in theHarvard Museum of Zoology. In ruminants (e.g., Bison), thedistal sesamoids have their major articulation with the base of the medial phalanx. In contrast, Giraffa has relatively largerdistal sesamoids that appear to have a greater articulation withthe ungual phalanx, possibly acting as a posterior extension forthe effective plantar surface of the ungual phalanx.
CONCLUSIONS
The peculiar foot morphology of present-day camelids, in-
dicative of a secondarily digitigrade foot posture in associationwith a pacing mode of locomotion, may have evolved morethan once within the lineage. The fully modern version of thismorphology is seen not only in the extinct taxa closely relatedto the living camels and llamas, but also within the Protolabi-nae, the sister taxon to the Camelinae, where it may representindependent evolution. Fossilized trackways also support thehypothesis that the derived protolabine Protolabis had a run-ning pace gait like that of extant camelids (Webb, 1972). Partialconvergence on the foot morphology of extant camelids wasalso seen within the more primitive families Stenomylinae (pri-marily in the genus Stenomylus) and the Miolabinae (primarilyin the genus Nothotylopus). However, some deviation from themore primitive condition (as exemplified by the extinct proto-ceratid Synthetoceras, or in extant tragulid ruminants) could be
observed in all camelids studied with the exception of the mostprimitive taxon, Poebrotherium.
We consider that most extinct camelids may have engagedin some types of lateral gaits, such as the lateral walk and therotary gallop, observed in extant long-legged antelope and thegiraffe. However, the running pace gait, unique to camelidsamong extant wild mammals, was probably used only by thosetaxa with a foot fully like that of extant camelids. The parallelemergence of more derived types of foot morphology amongfossil camelids occurred in the late Oligocene to early Miocenetimes, preceding the development of more open savanna habi-tats in the North American late Miocene.
ACKNOWLEDGMENTS
We thank the following people for access to specimens intheir collections. Maria Rutzmoser (Mammalogy) and Chuck Schaff (Vertebrate Paleontology) at the Museum of Compara-tive Zoology (Harvard University), and Dick Tedford (Verte-brate Paleontology) and Chris Norris (Mammalogy) at theAmerican Museum of Natural History (New York). Thanks alsoto Dave Webb for discussions, Kay Earls for checking speci-mens on display, Brian Regal for the osteological drawings, IanTattersall for accommodations, and Mark Norell for being un-derstanding of various problems. This project was funded inpart by a Salomon Faculty Award (Brown University) to CMJ,and by Brown University undergraduate honors thesis funds toBB.

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 10/12
119 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
LITERATURE CITED
Bertram, J. E. A., and A. A. Biewener. 1990. Differential scaling of thelong bones in the terrestrial Carnivora and other mammals. Journalof Morphology 20:157–169.
Frick, C. 1937. Horned ruminants of North America. Bulletin of theAmerican Museum of Natural History 69:1–669.
———, and B. E. Taylor. 1968. A generic review of stenomyline cam-els. American Museum Novitates 2353:1–51.
Gatesy, J., M. Milinkovitch, V. Waddell, and M. Stanhope. 1999. Sta-bility of cladistic relationships between Cetacea and higher-levelartiodactyl taxa. Systematic Biology 48:6–20.
Gatesy, S. M., and K. M. Middleton. 1997. Bipedalism, flight, and theevolution of theropod locomotor diversity. Journal of VertebratePaleontology 17:308–329.
Gauthier-Pilters, H., and A. I. Dagg. 1981. The Camel: its Evolution,Ecology, Behavior, and Relationships to Man. University of Chi-cago Press, Chicago, 208 pp.
Hildebrand, M. 1976. Analysis of tetrapod gaits: general considerationsand symmetrical gaits; pp. 203–236 in R. M. Herman, G. Grillner,P. S. G. Stein, and P. G. Stuart (eds.), Neural Control of Locomo-tion. Plenum Press, New York.
Honey, J. G., J. A. Harrison, D. R. Prothero, and M. S. Stevens. 1998.Camelidae; pp. 439 –462 in C. M. Janis, K. M. Scott, and L. L.Jacobs (eds.), Tertiary Mammals of North America. CambridgeUniversity Press, Cambridge and New York.
Howell, A. B. 1944. Speed in Animals. Hafner Publishing Co., NewYork, 270 pp.
Jacobs, B. F., J. D. Kingston, and L. L. Jacobs. 1999. The origin of grass-dominated ecosystems. Annals of the Missouri Botanic Gar-den 86:590–643.
Janis, C. M. 1982. Evolution of horns in ungulates: ecology and paleo-
ecology. Biological Reviews 57:261–318.
———, and K. M. Scott. 1987. The interrelationships of higher rumi-nant families with special emphasis on the members of the Cer-
voidea. American Museum Novitates 2893:1–85.
Joeckel, R. M., and J. M. Stavis. 1996. Basicranial anatomy of Syndy-
oceras cooki (Artiodactyla: Protoceratidae) and the need for a re-appraisal of tylopod relationships. Journal of Vertebrate Paleontol-ogy 16:320–327.
Leopold, L., G. Liu, and S. Clay-Poole. 1992. Low biomass vegetationin the Oligocene; pp. 399–420 in D. R. Prothero and W. A. Berg-gren (eds.), Eocene–Oligocene Climatic and Biotic Evolution.Princeton University Press, Princeton.
Norris, C. A. 1999. The cranium of Bunomeryx (Artiodactyla: Homa-codontidae) from the upper Eocene Uinta deposits of Utah and its
implications for tylopod systematics. Journal of Vertebrate Pale-ontology 19:742–751.
——— 2000. The cranium of Leptotragulus, a hornless protoceratid(Artiodactyla, Protoceratidae) from the middle Eocene of NorthAmerica. Journal of Vertebrate Paleontology 20:341–348.
Patton, T. H., and B. E. Taylor. 1971. The Synthetoceratinae (Mam-malia, Tylopoda, Protoceratidae). Bulletin of the American Muse-um of Natural History 145:119–218.
Scott, K. M. 1985. Allometric trends and locomotor adaptations in theBovidae. Bulletin of the American Museum of Natural History 179:197–288.
——— 1990. Postcranial dimensions of ungulates as predictors of bodymass; pp. 301–335 in J. Damuth and B. J. MacFadden (eds.), BodySize in Mammalian Paleobiology: Estimation and Biological Im-plications. Cambridge University Press, Cambridge and New York.
Scott, W. B. 1940. The mammalian fauna of the White River Oligocene,Part IV. Artiodactyla. Transactions of the American Philosophical
Society 28:363–746.Webb, S. D. 1972. Locomotor evolution in camels. Forma et Functio
5:99–112.——— 1977. A history of savanna vertebrates in the New World, Part
I. North America. Annual Review of Ecology and Systematics 8:355–380.
———, and B. E. Taylor. 1980. The phylogeny of hornless ruminantsand a description of the cranium of Archaeomeryx. Bulletin of theAmerican Museum of Natural History 167:117–158.
Received 5 June 2000; accepted 15 May 2001.
7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 11/12
120 JOURNAL OF VERTEBRATE PALEONTOLOGY, VOL. 22, NO. 1, 2002
APPENDIX
Data used in statistical analyses, all measurements in cm. Key: L ϭ articular length; ML ϭ mediolateral diameter; AP ϭ anteroposteriordiameter; Ext ϭ distal articular extension; GW ϭwidth of carinal groove; GDϭ depth of carinal groove; *estimated by regression. Taxa designatedby numbers used in Tables 1 and 2. Data for 51, Synthetoceras, from AMNH 40231, 40241, 40243, and 40245.
Taxon
Tibia
L ML AP
Metatarsal
L ML AP
Metacarpal
L ML AP
1 Poebrotherium
2 Pseudolabis3 Miotylopus4 Stenomylus5 Floridatragulus
22.47
30.2328.8221.85
*19.59
1.95
2.322.111.69
*2.92
1.41
2.582.751.81
*2.07
15.50
22.1923.8716.6812.98
1.82
2.091.371.212.08
0.99
1.971.941.171.17
13.07
22.28*22.71
16.2711.09
1.49
2.27*1.50
1.20*2.31
0.90
1.78*1.75
1.071.01
6 Gentilicamelus7 Paramiolabis8 Miolabis9 Nothotylopus
10 Tanymykter
26.40*24.07
27.9337.8536.93
2.16*2.04
2.573.233.03
2.49*1.98
2.593.853.12
18.8317.1217.2526.7527.56
1.711.462.232.452.10
1.471.391.702.402.69
18.0217.2316.7424.6627.15
1.981.422.712.912.48
1.531.381.762.322.36
11 Michenia12 Protolabis13 Oxydactylus14 ‘‘ Aepycamelus’’ priscus15 Aepycamelus
31.7744.6538.5052.3036.50
2.643.452.654.353.25
2.603.182.643.183.22
27.2036.7928.4745.9035.46
1.802.222.013.152.78
2.002.952.173.292.83
28.2236.9428.1245.3036.85
1.892.402.573.223.26
2.112.862.013.373.18
16 Hemiauchenia17 Procamelus18 Titanotylopus19 Camelus bactrianus
20 Camelus dromedarius
48.1044.5050.3647.94
43.59
3.904.625.535.46
5.01
3.433.925.114.47
4.27
40.7934.5038.0535.92
34.60
2.543.154.003.92
3.19
2.864.614.223.55
3.23
44.2535.8041.4835.35
34.68
2.553.874.724.48
3.69
2.944.004.353.72
3.0921 Llama glama22 Llama guanicoe23 Llama pacos24 Vicugna vicugna25 Hyemoschus aquaticus
30.1633.4529.0525.3912.36
3.162.942.822.661.31
2.762.962.592.491.67
21.0322.9920.3418.56
5.99
1.811.721.831.891.45
1.782.081.671.540.67
21.5023.4020.3018.39
3.96
1.982.141.982.191.39
1.762.221.651.500.38
26 Tragulus javanicus27 Okapia johnstoni28 Giraffa camelopardalis29 Alces alces30 Antilocapra americana
10.0733.6061.9046.9926.95
0.733.836.564.372.17
0.873.505.774.892.77
6.7129.1967.6136.8322.61
0.582.994.403.301.49
0.623.144.804.091.90
4.4528.5969.1632.2721.49
0.553.155.003.541.62
0.333.464.803.301.59
31 Axis porcinus32 Capreolus capreolus33 Cervus elaphus34 Elaphurus davidianus35 Mazama americana
19.4223.6945.7236.8319.96
1.231.774.823.761.62
1.781.915.483.511.80
13.3619.3234.9328.2612.33
1.171.233.172.571.08
1.411.534.312.690.91
10.7915.9530.4825.3615.08
1.221.823.492.761.21
1.101.193.722.341.25
36 Moschus moschiferus37 Odocoileus virginianus
38 Rangifer tarandus39 Boselaphus tragocamelus40 Capra hircus
19.9731.52
36.9437.5021.74
1.222.37
3.193.961.86
1.122.59
3.364.362.04
13.4127.34
29.5526.8912.21
0.841.85
2.392.741.37
0.952.35
3.433.111.32
9.8621.29
22.3026.5111.24
0.961.95
2.433.101.64
0.741.88
2.402.521.09
41 Cephalophus silvicultor 42 Damaliscus pygarus43 Gazella gazella44 Kobus leche45 Ourebia ourebi
25.3827.6922.2031.5920.82
2.662.151.712.651.54
3.462.121.833.041.78
16.8022.2617.7023.0415.46
1.891.381.151.951.14
2.091.851.522.141.36
15.3621.2616.4422.3915.31
2.051.551.272.031.09
1.511.421.101.650.99
46 Ovis aries47 Rupicapra rupicapra48 Saiga tatarica49 Sylvicapra grimmia50 Taurotragus oryx51 Synthetoceras
20.1726.8422.4317.4339.70
*22.34
2.051.591.891.824.35
*2.10
2.101.682.011.504.41
*2.59
12.5416.5518.4414.4228.70
*15.39
1.461.281.130.952.71
*1.51
1.471.251.851.093.18
*1.63
11.7714.9415.9113.1526.0213.57
1.711.481.380.863.021.64
1.311.061.360.833.721.39

7/29/2019 Janis Theodor Boisvert 2002
http://slidepdf.com/reader/full/janis-theodor-boisvert-2002 12/12
121 JANIS ET AL.— LOCOMOTOR EVOLUTION IN CAMELS
APPENDIX (Continued )
Taxon
Proximal phalanx
L ML AP Ext GW GD
Medial phalanx
L ML AP Ext
Ungual phalanx
L ML AP
1234
56789
10
2.844.353.583.72
4.314.384.574.607.105.70
0.710.970.960.68
0.931.030.771.241.541.32
0.861.280.950.90
1.071.071.011.281.731.37
0.350.480.350.50
0.500.400.550.501.170.51
0.220.320.380.29
0.370.340.370.490.630.47
0.250.300.340.25
0.370.260.420.300.480.33
1.572.211.802.19
2.212.122.102.373.803.21
0.661.010.850.67
0.980.870.851.061.591.12
0.691.050.710.68
0.980.870.771.101.161.07
0.510.860.590.56
0.590.620.500.840.840.92
1.692.611.841.48
1.881.781.402.762.122.53
0.480.700.670.64
0.710.720.580.931.180.91
0.621.120.910.55
0.940.790.751.081.101.22
11121314151617181920
6.997.735.358.129.69
10.229.99
12.6710.5610.09
0.921.311.051.701.771.711.952.382.402.14
1.511.611.141.872.322.042.523.212.772.44
0.870.740.481.311.541.221.912.172.211.94
0.590.650.450.870.830.740.850.971.020.82
0.360.420.330.400.460.440.370.310.450.39
3.233.712.374.664.373.964.776.476.175.98
1.051.181.131.781.821.832.012.522.642.81
1.051.060.861.581.551.551.702.191.861.58
0.760.860.671.231.080.990.911.171.221.17
1.982.502.593.022.572.692.782.652.962.57
0.810.980.871.441.451.241.651.682.241.90
1.081.131.151.791.541.401.771.741.661.50
2122
2324252627282930
6.967.95
6.526.062.041.446.62
12.968.305.28
1.231.45
1.101.060.850.412.323.572.510.95
1.491.60
1.251.290.730.412.503.732.901.53
1.041.25
0.961.030.200.050.541.001.460.36
0.670.67
0.580.480.34
—0.561.550.690.19
0.390.31
0.310.270.24
—0.430.850.550.21
3.463.84
3.012.921.480.963.896.446.182.99
1.371.56
1.501.140.750.312.083.782.171.01
1.071.18
1.091.300.550.382.193.683.191.26
0.931.00
0.840.940.400.111.843.622.881.14
2.282.17
2.442.001.221.006.132.998.113.37
1.071.24
1.191.050.590.261.811.902.240.66
1.241.23
1.291.020.500.323.022.503.511.51
31323334353637383940
3.453.927.446.723.034.155.236.637.113.83
0.750.782.292.060.810.631.331.602.311.10
1.231.072.802.421.050.901.601.862.901.08
0.710.340.950.980.350.360.730.780.870.66
0.130.310.650.680.170.440.490.650.590.30
0.200.270.490.650.150.380.460.430.480.45
2.622.675.445.732.292.984.314.614.802.61
0.910.842.121.810.810.601.321.592.251.00
1.081.052.592.300.940.781.611.742.570.99
0.930.832.421.620.660.601.261.851.991.06
2.632.726.555.482.062.013.565.356.023.29
0.770.662.062.050.660.551.051.971.680.50
1.171.092.792.270.980.881.532.312.741.54
4142434445464748495051
4.475.464.525.743.213.744.594.893.076.515.37
1.760.980.891.370.831.231.090.920.632.171.67
1.641.271.051.651.001.171.151.180.852.681.43
0.740.490.580.550.210.570.530.450.470.830.49
0.200.310.180.270.210.340.320.250.220.510.31
0.330.380.250.430.180.240.480.310.290.730.40
2.993.012.353.742.122.433.312.452.064.120.32
1.271.030.721.410.641.050.840.820.672.141.97
1.481.220.731.450.801.070.970.930.792.541.55
1.061.130.781.360.760.941.160.900.671.860.69
3.043.672.915.032.542.713.732.452.077.770.39
0.810.780.641.110.550.700.720.590.641.681.98
1.231.581.121.650.881.301.351.120.822.992.33





























