Embed Size (px)
Citation preview

BỘ GIÁO DỤC VIỆN HÀN LÂM KHOA HỌC
VÀ ĐÀO TẠO VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
-----------------------------
Phạm Thị Huế
NGHIÊN CỨU TINH CHẾ VÀ XÁC ĐỊNH HOẠT TÍNH KHÁNG
KHUẨN CỦA PEPTIDE MASTOPARAN TỪ NỌC ONG Vespa
velutina THU Ở VIỆT NAM
LUẬN VĂN THẠC SĨ SINH HỌC
Hà Nội- 2021

BỘ GIÁO DỤC VIỆN HÀN LÂM KHOA HỌC
VÀ ĐÀO TẠO VÀ CÔNG NGHỆ VIỆT NAM
HỌC VIỆN KHOA HỌC VÀ CÔNG NGHỆ
-----------------------------
Phạm Thị Huế
NGHIÊN CỨU TINH CHẾ VÀ XÁC ĐỊNH HOẠT TÍNH KHÁNG
KHUẨN CỦA PEPTIDE MASTOPARAN TỪ NỌC ONG Vespa
velutina THU Ở VIỆT NAM
Chuyên ngành: Sinh học thực nghiệm
Mã số: 8420114
LUẬN VĂN THẠC SĨ SINH HỌC
NGƯỜI HƯỚNG DẪN KHOA HỌC:
Hướng dẫn
TS. Lê Thị Bích Thảo
Hà Nội-2021

Lời cam đoan
Tôi xin cam đoan đề tài nghiên cứu trong luận văn này là công trình nghiên
cứu của tôi dựa trên những tài liệu, số liệu do chính tôi tự tìm hiểu và nghiên
cứu. Chính vì vậy, các kết quả nghiên cứu đảm bảo trung thực và khách quan
nhất. Đồng thời, kết quả này chưa từng xuất hiện trong bất cứ một nghiên cứu
nào. Các số liệu, kết quả nêu trong luận văn là trung thực nếu sai tôi hoàn chịu
trách nhiệm.
Tác giả luận văn
Phạm Thị Huế

Lời cảm ơn
Lời đầu tiên, tôi xin bày tỏ lòng biết ơn chân thành và sâu sắc tới TS. Lê
Thị Bích Thảo, người cô đã tận tình chỉ bảo, hướng dẫn và giúp đỡ tôi trong
suốt quá trình học tập, nghiên cứu và hoàn thiện luận văn.
Tôi xin gửi lời cảm ơn chân thành tới tới GS.TS. Phan Văn Chi, TS. Bùi
Thị Huyền, TS. Phạm Đình Minh cùng toàn thể các anh chị làm việc tại phòng
Hóa sinh Protein, Phòng thí nghiệm Trọng điểm Công nghệ Gen, Viện Công
nghệ Sinh học;
Tôi xin gửi lời cảm ơn tới PGS.TS. Nguyễn Thị Phương Liên, Viện Sinh
thái và Tài nguyên sinh vật, Viện Hàn lâm Khoa học và Công nghệ Việt Nam;
TS. Phạm Thị Hằng, Viện Sốt rét – Ký sinh trùng- Côn trùng Trung ương;
Trung tâm Bệnh nhiệt đới, Bệnh viện Bạch Mai; và ban Lãnh đạo, phòng Đào
tạo, các phòng chức năng của Học viện Khoa học và Công nghệ đã hết lòng
giúp đỡ tôi thực hiện thành công luận văn này.
Cuối cùng, tôi vô cùng biết ơn gia đình, bạn bè, những người thân đã
luôn khích lệ, động viên, giúp đỡ và là chỗ dựa vững chắc cho tôi trong quãng
thời gian qua.
Xin chân thành cảm ơn!
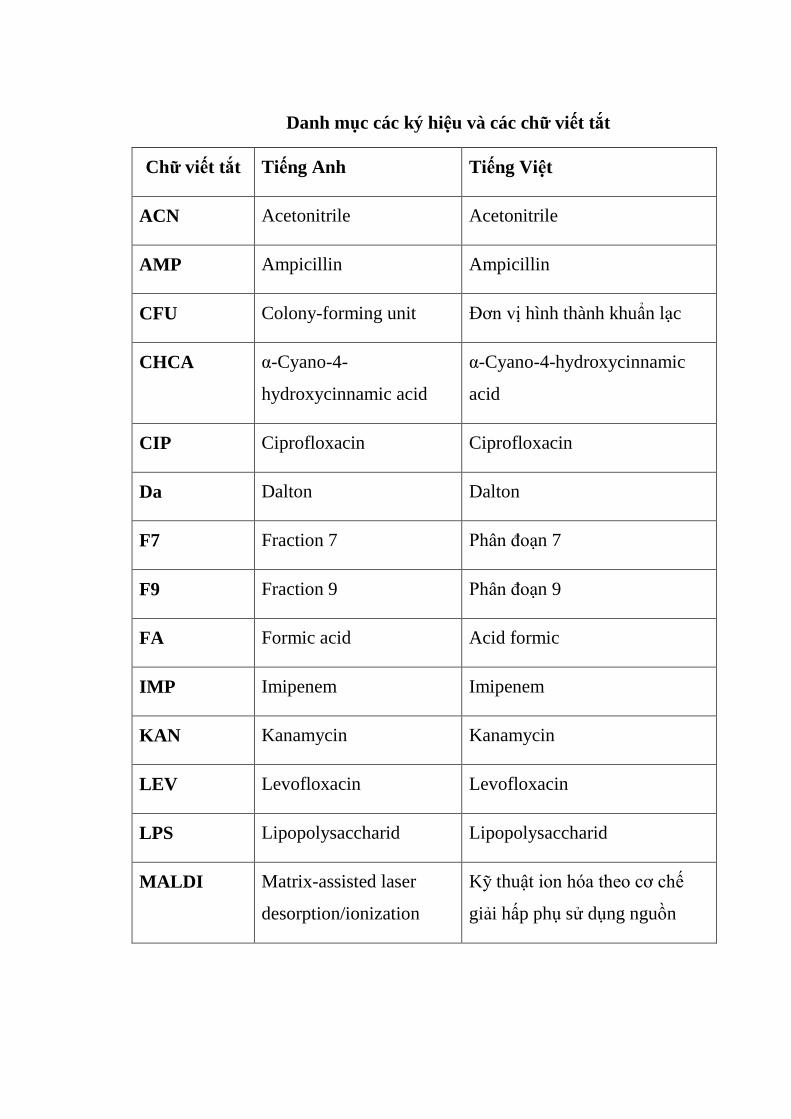
Danh mục các ký hiệu và các chữ viết tắt
Chữ viết tắt Tiếng Anh Tiếng Việt
ACN Acetonitrile Acetonitrile
AMP Ampicillin Ampicillin
CFU Colony-forming unit Đơn vị hình thành khuẩn lạc
CHCA α-Cyano-4-
hydroxycinnamic acid
α-Cyano-4-hydroxycinnamic
acid
CIP Ciprofloxacin Ciprofloxacin
Da Dalton Dalton
F7 Fraction 7 Phân đoạn 7
F9 Fraction 9 Phân đoạn 9
FA Formic acid Acid formic
IMP Imipenem Imipenem
KAN Kanamycin Kanamycin
LEV Levofloxacin Levofloxacin
LPS Lipopolysaccharid Lipopolysaccharid
MALDI Matrix-assisted laser
desorption/ionization
Kỹ thuật ion hóa theo cơ chế
giải hấp phụ sử dụng nguồn
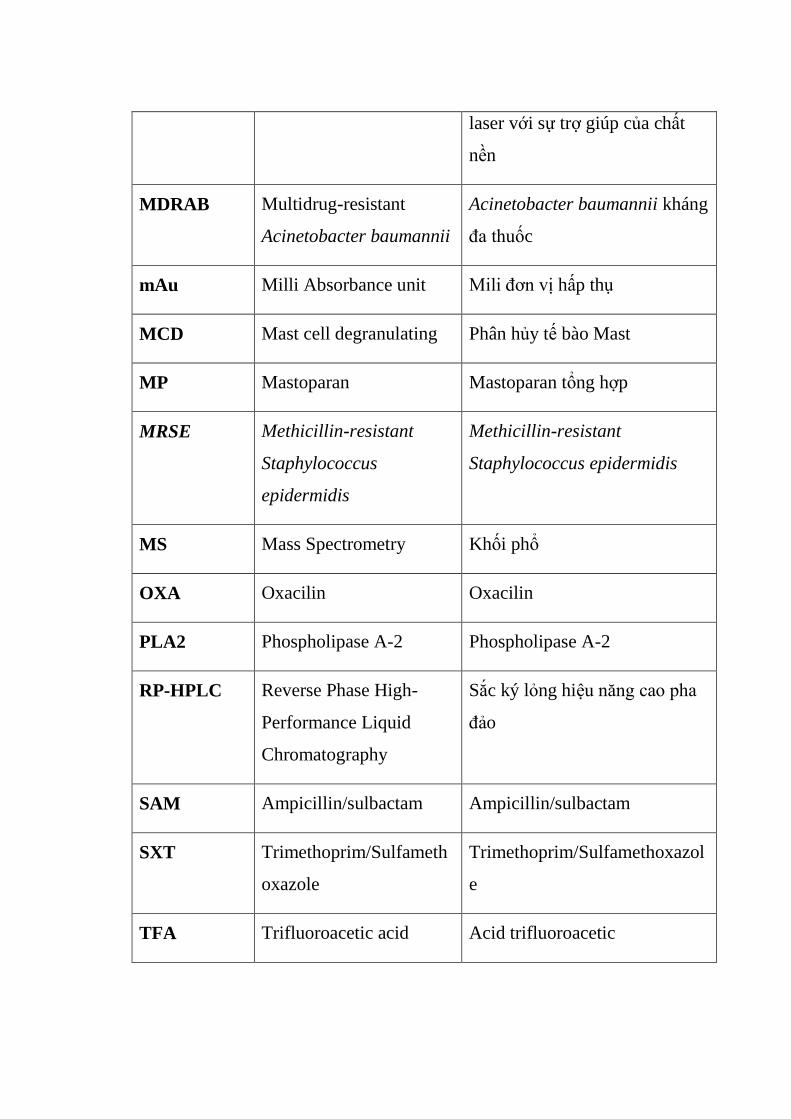
laser với sự trợ giúp của chất
nền
MDRAB Multidrug-resistant
Acinetobacter baumannii
Acinetobacter baumannii kháng
đa thuốc
mAu Milli Absorbance unit Mili đơn vị hấp thụ
MCD Mast cell degranulating Phân hủy tế bào Mast
MP Mastoparan Mastoparan tổng hợp
MRSE Methicillin-resistant
Staphylococcus
epidermidis
Methicillin-resistant
Staphylococcus epidermidis
MS Mass Spectrometry Khối phổ
OXA Oxacilin Oxacilin
PLA2 Phospholipase A-2 Phospholipase A-2
RP-HPLC Reverse Phase High-
Performance Liquid
Chromatography
Sắc ký lỏng hiệu năng cao pha
đảo
SAM Ampicillin/sulbactam Ampicillin/sulbactam
SXT Trimethoprim/Sulfameth
oxazole
Trimethoprim/Sulfamethoxazol
e
TFA Trifluoroacetic acid Acid trifluoroacetic

TFE Tetrafluoroethylene Tetrafluoroethylene
TOF Time of Flight Thời gian bay

Danh mục bảng
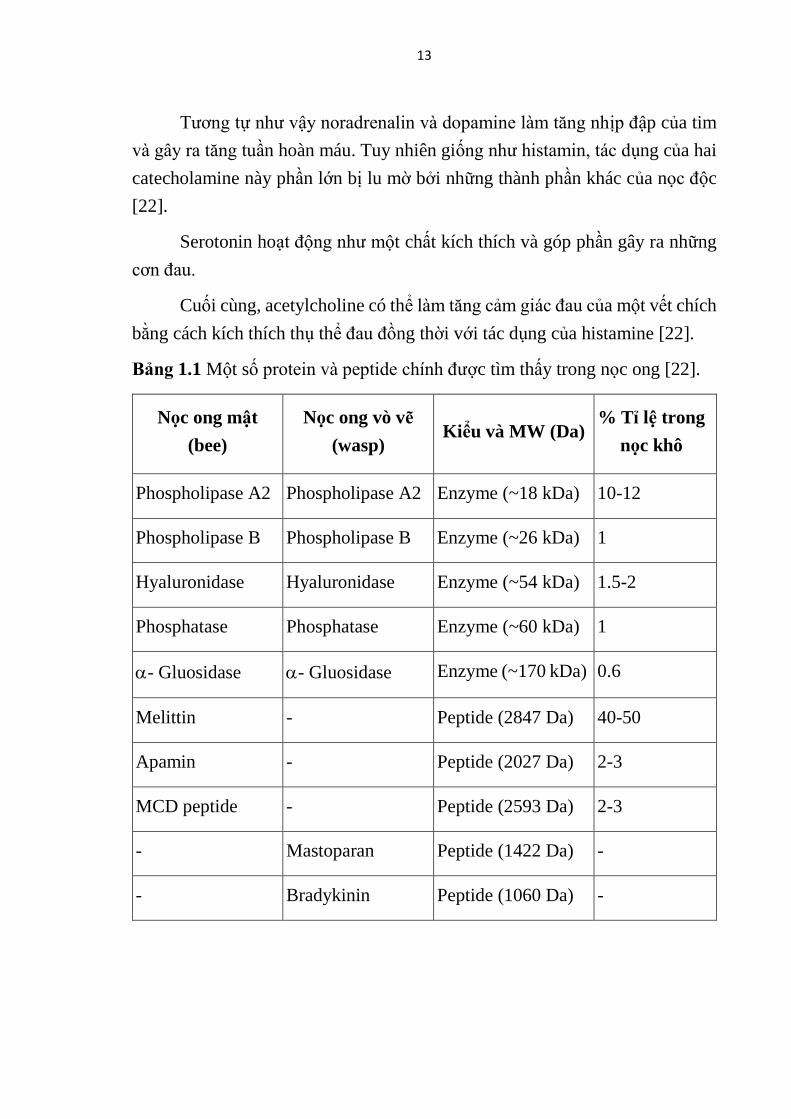
Bảng 1.1 Một số protein và peptide chính được tìm thấy trong nọc ong . ..... 13
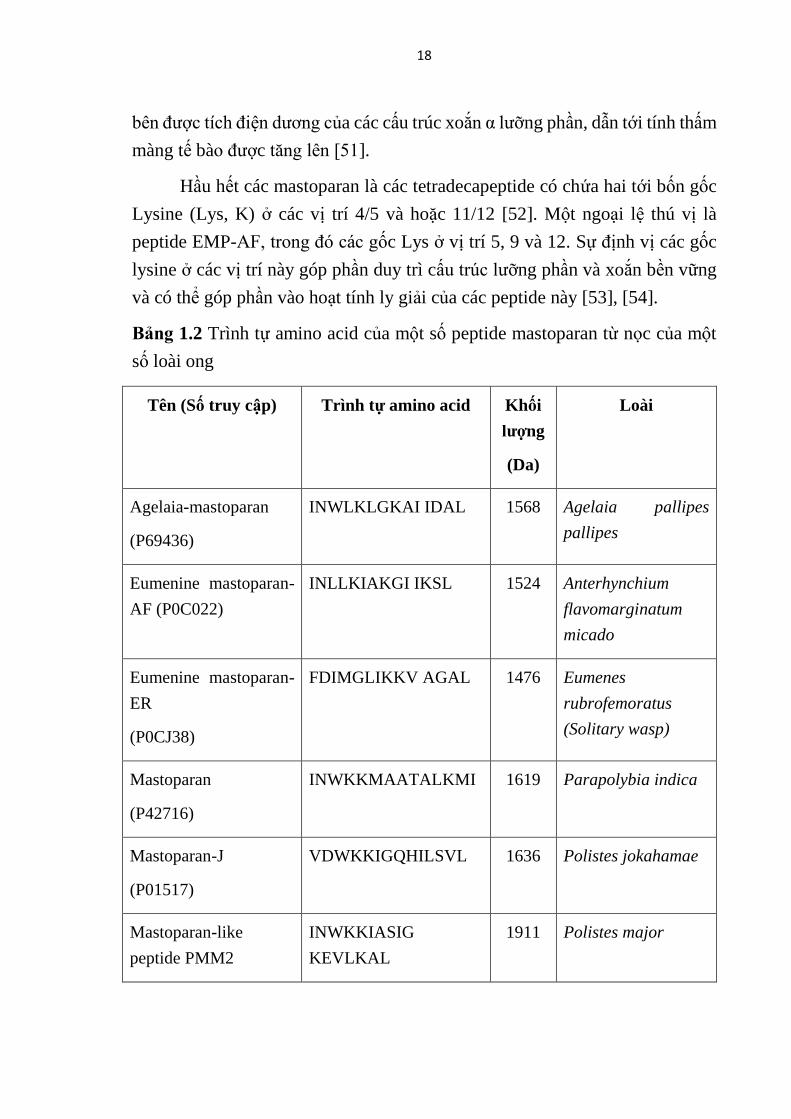
Bảng 1.2 Trình tự amino acid của một số peptide mastoparan từ nọc của một
số loài ong ....................................................................................................... 18
Bảng 2.1 Các hóa chất chính được sử dụng trong nghiên cứu ....................... 29
Bảng 2.2 Các thiết bị được sử dụng chính trong nghiên cứu ......................... 30
Bảng 3.1 Kết quả nhận diện phân đoạn 7 bằng phần mềm PEAK. ............... 40
Bảng 3.2 Kết quả nhận diện phân đoạn 9 bằng phần mềm PEAK. ................ 41
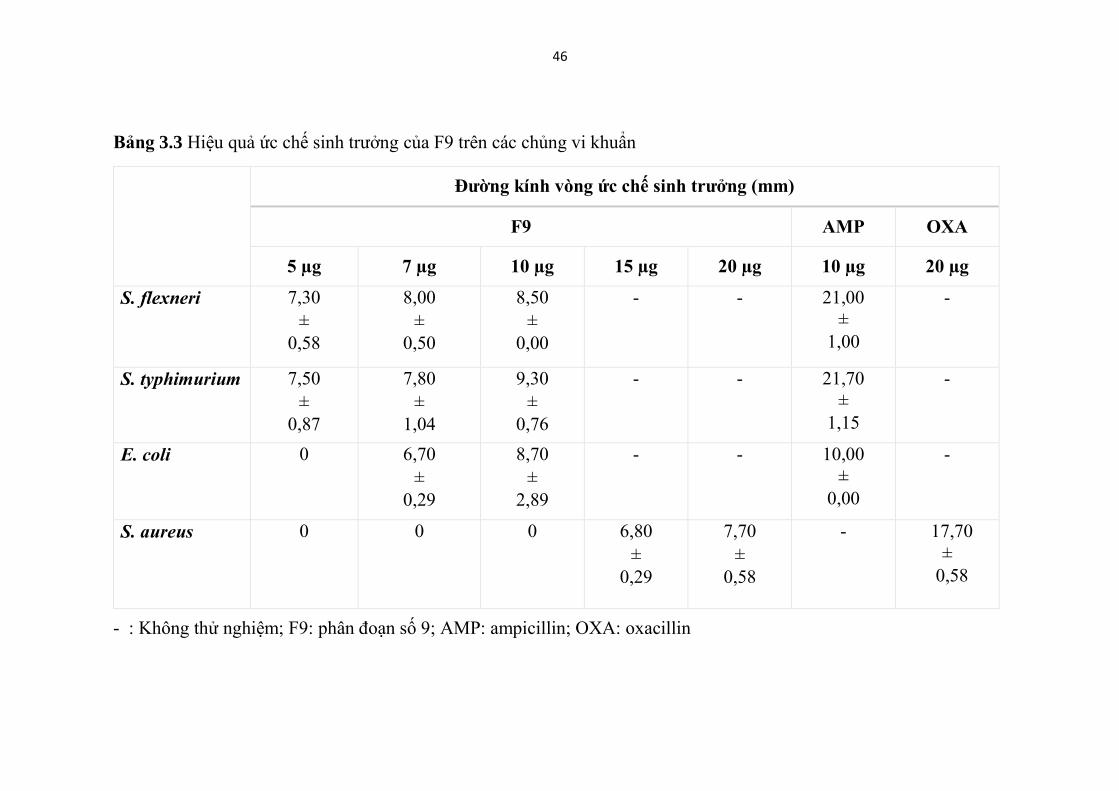
Bảng 3.3 Hiệu quả ức chế sinh trưởng của F9 trên các chủng vi khuẩn ........ 46
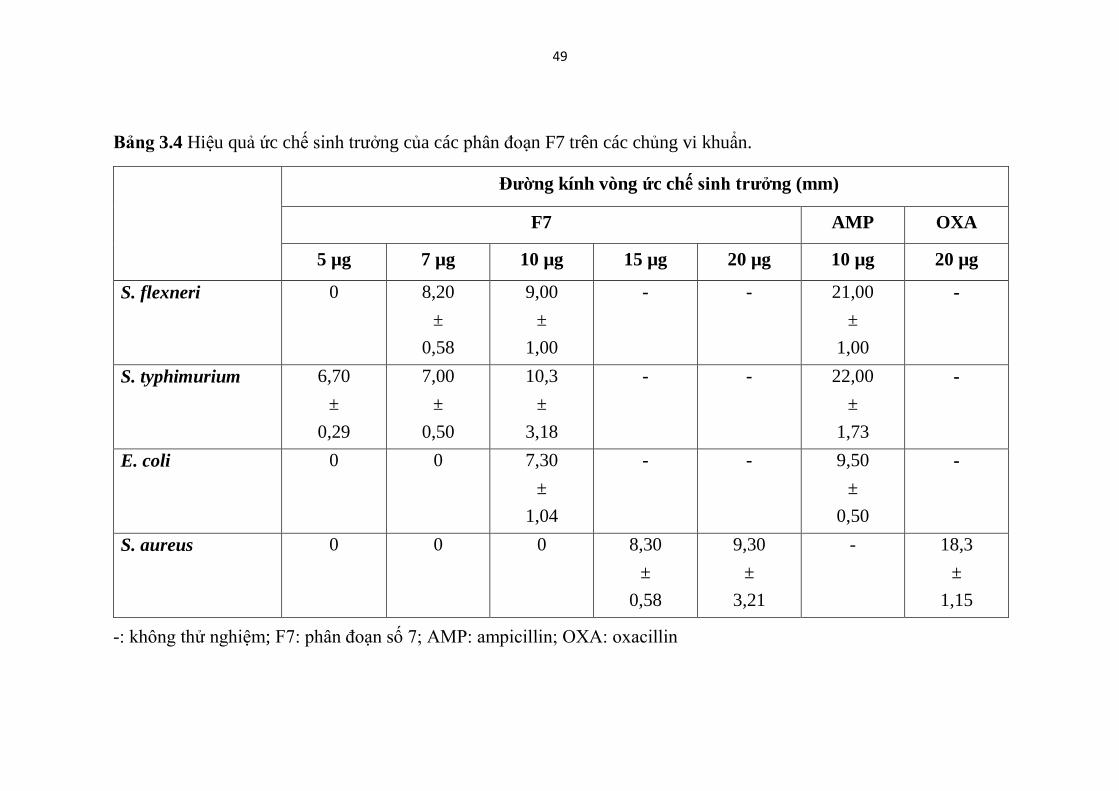
Bảng 3.4 Hiệu quả ức chế sinh trưởng của các phân đoạn F7 trên các chủng vi
khuẩn. .............................................................................................................. 49
Bảng 3.5 Kết quả thử nghiệm xác định nồng độ ức chế tối thiểu của MP trên
các chủng vi khuẩn kháng kháng sinh ............................................................ 52
Bảng 3.6 Mức độ nhạy cảm của các vi khuẩn đối với các MP và các loại thuốc
kháng sinh ....................................................................................................... 56

Danh mục hình
Hinh 1.1 Ong Vespa velutina (Lepeletier) ....................................................... 5
Hinh 1.2 Ong Vespa velutina nigrithorax cái mặt lưng (phía trên) và bụng (phía
dưới) .................................................................................................................. 7
Hinh 1.3 Ong Vespa velutina nigrithorax cái (bên trái) và đực (bên phải) ..... 8
Hinh 1.4 Nọc ong săn mồi như một nguồn của các hợp chất có hoạt tính sinh
học và các ứng dụng của chúng. ....................................................................... 9
Hinh 1.5 Một số hợp chất trong thành phần trong nọc ong ........................... 10
Hinh 1.6 Cấu trúc 3D của Mastoparan X được xác định bằng NMR. ........... 16
Hinh 1.7 Sự so sánh trình tự của các gốc amino acid của các polypeptide tiền
thân của mastoparan. ....................................................................................... 17
Hinh 1.8 Các mô hình để giải thích các cơ chế của sự thấm qua màng ......... 24
Hinh 2.1 Sơ đồ quy trình các bước tiến hành ................................................. 31
Hinh 3.1 Ong Vespa velutina cái .................................................................... 35
Hinh 3.2 Cơ quan nọc độc của ong Vespa velutina sau khi được giải phẫu. 36
Hinh 3.3 Sắc ký đồ của các phân đoạn của dịch chiết nọc ong Vespa velutina
......................................................................................................................... 37
Hinh 3.4 Phổ MALDI-TOF của phân đoạn 7 (A) và phân đoạn 9 (B) ......... 39
Hinh 3.5 Giải trình tự peptide các phân đoạn F7 (A) và F9 (B) có chứa
mastoparan sử dụng De novo .......................................................................... 42
Hinh 3.6 Sự so sánh trình tự của F7 và F9 với mastoparan từ các loài ong trong
chi Vespa ......................................................................................................... 43
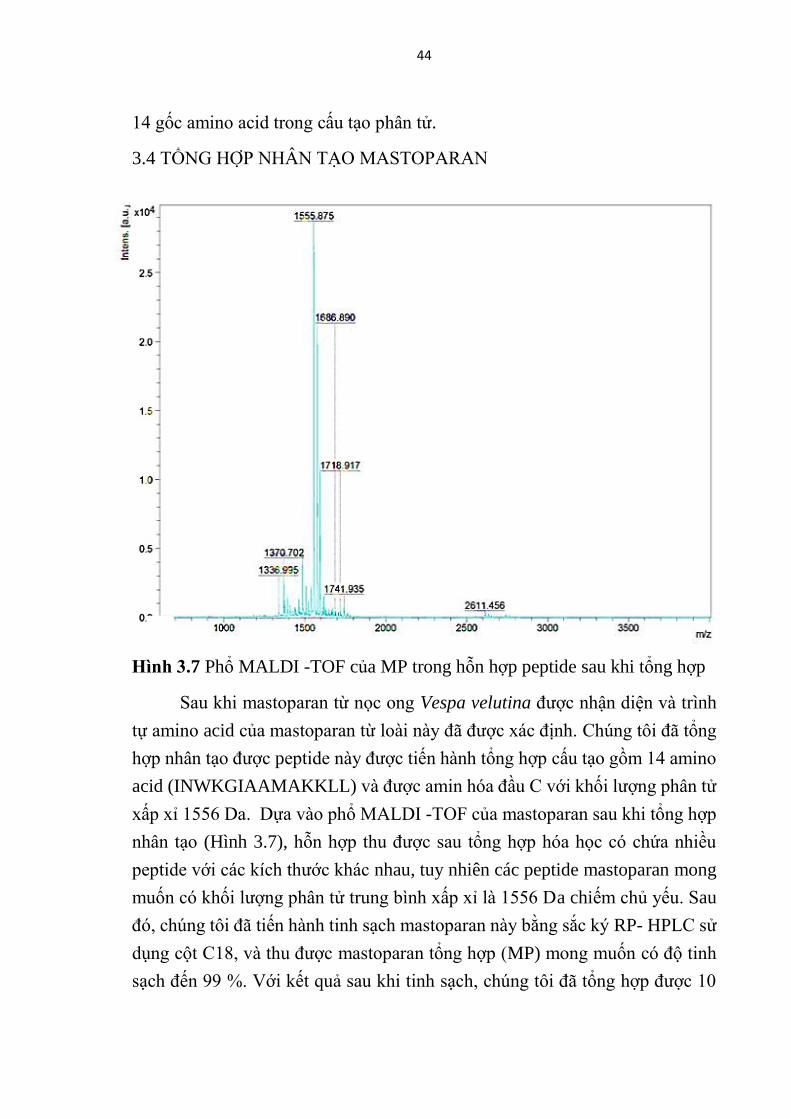
Hinh 3.7 Phổ MALDI -TOF của MP trong hỗn hợp peptide sau khi tổng hợp
......................................................................................................................... 44
Hinh 3.8 Hiệu quả ức chế sự sinh trưởng của mastoparan phân đoạn số 9 trên
các chủng vi khuẩn .......................................................................................... 45

Hinh 3.9 Hiệu quả ức chế sự sinh trưởng của mastoparan phân đoạn số 7 trên
các chủng vi khuẩn .......................................................................................... 48
Hinh 3.10 Kết quả Thử hoạt tính kháng khuẩn của MP trên các chủng vi khuẩn
kháng kháng sinh theo các nồng độ ................................................................ 53
Hinh 3.11 Kết quả xác định mức độ nhạy cảm của MP và các kháng sinh đối
với các chủng vi khuẩn kháng kháng sinh ...................................................... 55

1
MỤC LỤC
MỞ ĐẦU ------------------------------------------------------------------------------- 3
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU ----------------------------------------- 5
1.1 GIỚI THIỆU VỀ ONG Vespa velutina -------------------------------------- 5
1.1.1 Phân loại khoa học -------------------------------------------------------- 6
1.1.2 Phân bố của ong Vespa velutina ---------------------------------------- 6
1.1.3 Hình thái của ong Vespa velutina -------------------------------------- 6
1.2 GIỚI THIỆU VỀ NỌC ONG ------------------------------------------------- 8
1.2.1 Các thành phần của nọc ong -------------------------------------------- 9
1.2.1.1 Các enzyme ----------------------------------------------------------- 11
1.2.1.2 Các peptide ----------------------------------------------------------- 11
1.2.1.3 Các phân tử nhỏ khác ----------------------------------------------- 12
1.2.2 Ứng dụng của nọc ong ------------------------------------------------- 14
1.3 MASTOPARAN -------------------------------------------------------------- 15
1.3.1 Cấu tạo -------------------------------------------------------------------- 15
1.3.2. Hoạt tính của mastoparan -------------------------------------------- 20
1.3.2.1 Hoạt tính chung của mastoparan ---------------------------------- 20
1.3.2.2 Hoạt tính kháng khuẩn của mastoparan -------------------------- 21
1.3.2.3 Cơ chế hoạt động của các mastoparan chung trên màng của vi
sinh vật ------------------------------------------------------------------------- 23
1.4 TÌNH HÌNH NGHIÊN CỨU TẠI VIỆT NAM --------------------------- 25
CHƯƠNG 2. NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU -- 28
2.1 NGUYÊN VẬT LIỆU ------------------------------------------------------- 28
2.1.1 Vật liệu sinh học --------------------------------------------------------- 28
2.1.2 Các hóa chất ------------------------------------------------------------- 29
2.1.3 Trang thiết bị ------------------------------------------------------------ 30
2.2 SƠ ĐỒ NGHIÊN CỨU CHUNG ------------------------------------------- 31

2
2.3 PHƯƠNG PHÁP NGHIÊN CỨU ------------------------------------------ 31
2.3.1. Thu thập và tách chiết nọc ong -------------------------------------- 31
2.3.2. Phân đoạn nọc ong và tinh sạch mastoparan --------------------- 32
2.3.3. Phân tích MALDI-TOF ----------------------------------------------- 32
2.3.4 Tổng hợp hóa học mastoparan bằng phương pháp tổng hợp
peptide pha rắn ---------------------------------------------------------------- 33
2.3.5 Phương pháp khoanh giấy khuếch tán trên đĩa thạch ---------- 34
2.3.6 Xử lý số liệu -------------------------------------------------------------- 34
CHƯƠNG 3. KẾT QUẢ VÀ THẢO LUẬN ---------------------------------- 35
3.1 THU THẬP VÀ TÁCH CHIẾT NỌC ONG Vespa velutina ----------- 35
3.1.1 Thu thập ong Vespa velutina ------------------------------------------ 35
3.1.2 Tách chiết nọc ong Vespa velutina ----------------------------------- 36
3.2 PHÂN ĐOẠN NỌC ONG VÀ TINH SẠCH MASTOPARAN -------- 36
3.3 NHẬN DIỆN VÀ XÁC ĐỊNH TRÌNH TỰ AMINO ACID CỦA
MASTOPARAN ------------------------------------------------------------------ 38
3.4 TỔNG HỢP NHÂN TẠO MASTOPARAN ------------------------------ 44
3.5 THỬ NGHIỆM HOẠT TÍNH KHÁNG KHUẨN CỦA MASTOPARAN
--------------------------------------------------------------------------------------- 45
3.5.1 Hoạt tính kháng khuẩn của mastoparan tự nhiên --------------- 45
3.5.2 Thử nghiệm hoạt tính kháng khuẩn của mastoparan được tổng
hợp nhân tạo (MP).------------------------------------------------------------ 51
CHƯƠNG 4. KẾT LUẬN VÀ KIẾN NGHỊ ---------------------------------- 58
4.1 KẾT LUẬN -------------------------------------------------------------------- 58
4.2 KIẾN NGHỊ ------------------------------------------------------------------- 58
TÀI LIỆU THAM KHẢO -------------------------------------------------------- 59

3
MỞ ĐẦU
Thuốc kháng sinh là một trong những phát minh quan trọng nhất trong
nghiên cứu y sinh. Với sự phát hiện ra penicillin của Alexander Fleming vào
năm 1928, “kỷ nguyên kháng sinh” và nghiên cứu về thuốc kháng sinh đã được
bắt đầu. Tuy nhiên, việc sử dụng rộng rãi các loại thuốc chống nhiễm trùng
trong bệnh viện và công nghiệp đã làm phát sinh và gia tăng số lượng các chủng
vi khuẩn đa kháng thuốc. Các thuốc kháng sinh đang mất dần hiệu lực, vì vậy
hướng nghiên cứu tìm kiếm các chất có tính kháng khuẩn, đặc biệt là kháng lại
một số dòng vi khuẩn kháng kháng sinh là một nhu cầu cấp thiết của xã hội.
Các peptide kháng khuẩn đã được nghiên cứu và cho thấy tiềm năng trong việc
điều trị các bệnh nhiễm trùng do vi khuẩn, nấm. Hiện nay, các peptide trong
nọc độc của các loài động vật/côn trùng đang được quan tâm nghiên cứu ngày
càng nhiều cho những ứng dụng phát triển chất kháng sinh mới. Trong số đó,
mastoparan là ứng cử viên được quan tâm nhiều nhất.
Vespa velutina là một trong những loài ong vò vẽ nguy hiểm có xuất xứ
từ các nước Đông Nam Á. Đây là loài côn trùng có nọc độc cực mạnh, có thể
tái tạo nọc độc nhiều lần. Nọc ong là một hỗn hợp phức tạp trong thành phần
có chứa các enzyme có bản chất là protein (hyaluronidase, phospholipase A và
B, phosphatase acid, các protease, esterase và glucidase); các peptide nhỏ
(apamin là chất độc thần kinh, melittin là chất độc gây tan máu, MCD peptide
- Mast cell degranulating peptide, kinin, bradykinin, kháng nguyên 5,
mastoparan); và cuối cùng là các phân tử nhỏ khác như các amin gây viêm
(histamin, noradrenalin, dopamin, serotonin, acetylcholine). Chúng đã được
phát hiện có tiềm năng trong việc điều trị nhiều loại bệnh như một số bệnh rối
loạn thần kinh, bảo vệ phóng xạ, chống đột biến, chống viêm, chống kích hoạt
đau, và còn có khả năng sử dụng cho mục đích khác như kháng ung thư, kháng
khuẩn,..vv.
Từ những cơ sở dữ liệu đầu tiên về thành phần các protein/peptide có
trong nọc ong Vespa velutina thu thập ở Việt Nam, mastoparan đã được nhận
diện trong tổng số hơn 80 protein và peptide từ cơ sở dữ liệu này. Trên cơ sở
đó, chúng tôi thực hiện đề tài “Nghiên cứu tinh chế và xác định hoạt tính

4
kháng khuẩn của peptide mastoparan từ nọc ong Vespa velutina thu ở Việt
Nam” với mong muốn phát hiện một loại mastoparan mới từ loài ong này và
đánh giá hoạt tính kháng khuẩn của peptide này trên một số chủng vi khuẩn
nhằm tìm kiếm khả năng ứng dụng của trong y dược của chúng, phục vụ đời
sống con người như một chất kháng sinh mới.

5
CHƯƠNG 1. TỔNG QUAN TÀI LIỆU
1.1 GIỚI THIỆU VỀ ONG Vespa velutina
Ong bắt mồi chân vàng, Vespa velutina (Hình 1.1) là một loài ong có
nguồn gốc từ các nước khu vực Châu Á [1] và đã xâm nhập vào một số khu
vực của Châu Âu, lần đầu tiên xuất hiện ở Pháp vào năm 2004 [1]. Như một kẻ
săn mồi nói chung, chúng là một loài gây hại cho ong mật và là mối quan tâm
chính của nhiều người nuôi ong.
Hinh 1.1 Ong Vespa velutina (Lepeletier) [1]

6
1.1.1 Phân loại khoa học
Giới (regnum) : Animalia
Ngành (phylum) : Arthropoda
Lớp (class) : Insecta
Bộ (ordo) : Hymenoptera
Họ (familia) : Vespidae
Phân họ (subfamilia) : Vespinae
Chi (genus) : Vespa
Loài (species) : V. velutina
1.1.2 Phân bố của ong Vespa velutina
Vespa velutina có nguồn gốc từ Châu Á, là loài bản địa ở Đông Nam Á,
xuất hiện ở Hàn Quốc [1], Trung Quốc [2] và Nhật Bản [3] và mốt số quốc gia
Châu Á khác. Nó được du nhập vào Pháp vào năm 2004, rất có thể thông qua
việc nhập khẩu các chậu cây cảnh [2]. Loài này có khả năng xâm lấn rất cao và
dễ dàng lây lan sang các khu vực châu Âu khác như Ý [4], Anh [5], Quần đảo
Balearic [6] và miền bắc nước Đức [7]
Phân bố của loài ong Vespa velutina ở Việt Nam
Loài ong Vespa velutina có thể được tìm thấy ở các tỉnh của Việt Nam như: Hà
Giang, Cao Bằng, Thái Nguyên, Phú Thọ, Vĩnh Phúc, Hà Tây, Nghệ An, Hà
Tĩnh, Quảng Trị, Thừa Thiên Huế, Quảng Nam, Lâm Đồng, v.v. [8]
1.1.3 Hình thái của ong Vespa velutina
V. velutina rất đa dạng về màu sắc và 10 phân loài đã được xác định [8],
[9], [10].
C. Villemant (Bảo tàng Lịch sử Tự nhiên Quốc gia, Paris, thông tin cá nhân của
Pháp, 2013) mô tả Vespa velutina nigrithorax (Hình 1.2 và Hình 1.3) như sau:

7
Ong cái: Đầu và ngực nhọn, có lông tơ mịn mọc rải rác, bụng có rãnh.
Cơ thể màu nâu sẫm, mặt và miệng có màu cam (trừ răng sẫm màu); râu màu
nâu ở mặt lưng, màu cam ở bụng. Mặt lưng của các đoạn bụng màu nâu với các
rìa đỉnh rõ ràng hơn: một dải mỏng màu vàng ở đoạn thứ nhất và dải mỏng màu
cam ở đoạn thứ hai và thứ ba; đoạn bụng thứ tư gần như hoàn toàn màu cam
với một vạch đen hình tam giác ở giữa, thường không nhìn thấy khi ong bắp
cày còn sống; đoạn bụng thứ năm và thứ sáu có màu nâu cam nhiều hơn hoặc
ít hơn. Phần bụng màu nâu, đỉnh hình tam giác thường nhạt hơn; phân đoạn thứ
hai và thứ ba màu vàng toàn thân với một vạch đen ở giữa. Chân màu nâu trừ
tarsi vàng; cánh màu nâu xám. Không có sự khác biệt rõ rệt về hình thái giữa
con cái (ong thợ) hữu tính và bất dục. Một số ong thợ nhỏ hơn (đặc biệt là vào
mùa xuân) nhưng vào mùa thu, nhiều ong thợ lớn như những ong chúa tương
lai. Bên trong đàn, có thể nhận ra ong chúa đẻ trứng bằng phần bụng căng phồng
và vào cuối mùa, đôi cánh bị hư hại.
Hinh 1.2 Ong Vespa velutina nigrithorax cái mặt lưng (phía trên) và bụng
(phía dưới) [11]

8
Con đực: rất giống con cái về kích thước và màu sắc, râu dài hơn. Mặt
bụng của bụng màu nâu, đỉnh cụt có một đôi đốm vàng.
Ong đực và ong cái có thể phân biệt với nhau bằng các râu của chúng,
với các râu ong cái có vẻ mỏng hơn và chiều dài ngắn hơn so với râu của ong
đực. Như với tất cả các Bộ cánh màng, con cái có ngòi độc còn con đực thì
không.
Hinh 1.3 Ong Vespa velutina nigrithorax cái (bên trái) và đực (bên phải) [12]
1.2 GIỚI THIỆU VỀ NỌC ONG
Các sinh vật sử dụng nọc độc như một thứ vũ khí hoá học hiệu quả để tự
vệ và săn bắt con mồi, vì vậy nọc của chúng được tiến hoá theo hướng đặc
trưng cho loài và độc tính cao. Bên cạnh đó, nọc độc cũng là nguồn dự trữ các
hợp chất có hoạt tính sinh học quý, mang lại nhiều hứa hẹn trong việc tìm tòi
và phát triển các loại thuốc chữa bệnh (Hình 1.4).

9
Hinh 1.4 Nọc ong săn mồi như một nguồn của các hợp chất có hoạt tính sinh
học và các ứng dụng của chúng [13].
1.2.1 Các thành phần của nọc ong
Nọc ong là một hỗn hợp phức tạp chứa 3 nhóm chất chính: gồm các
enzyme có bản chất là protein (hyaluronidase, phospholipase A và B,
phosphatase acid, các protease, esterase và glucidase); các peptide nhỏ (apamin
là chất độc thần kinh, melittin là chất độc gây tan máu, MCD peptide - Mast
cell degranulating peptide, kinin, bradykinin, kháng nguyên 5, mastoparan); và
cuối cùng gồm các phân tử nhỏ khác như các amin gây viêm (histamin,
noradrenalin, dopamin, serotonin, acetylcholine) (Bảng 1.1) [14].
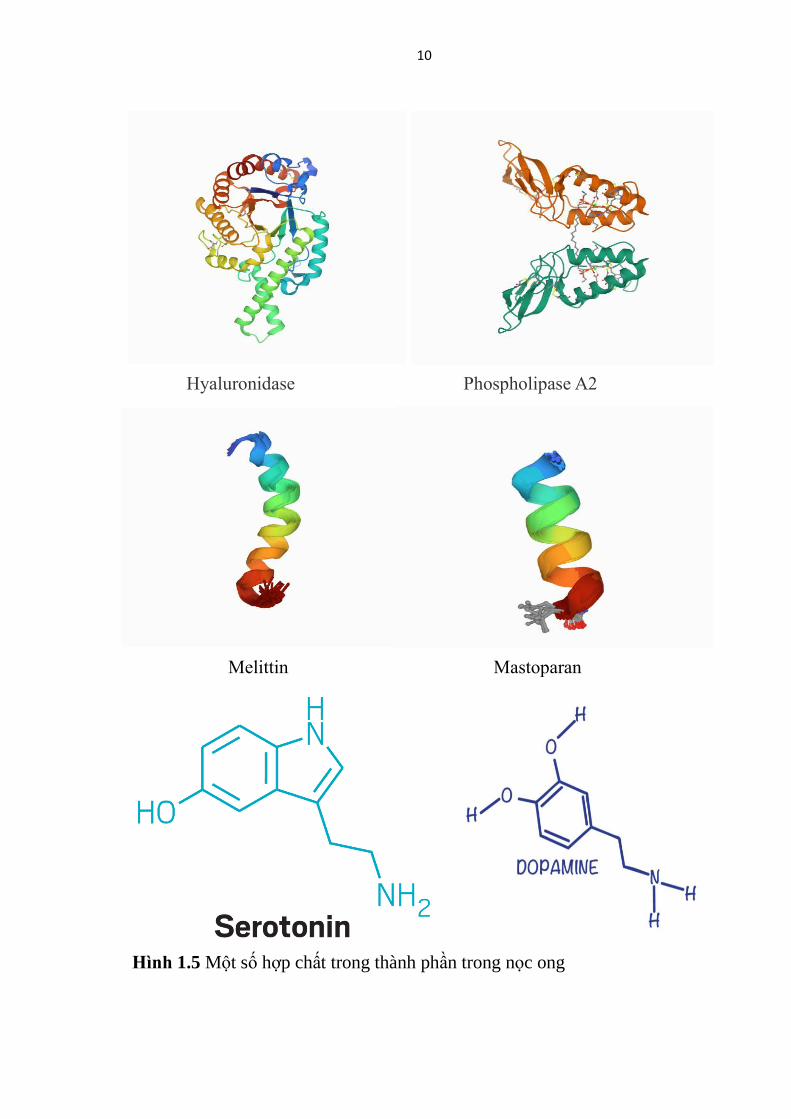
10
Hinh 1.5 Một số hợp chất trong thành phần trong nọc ong
Hyaluronidase Phospholipase A2
Melittin Mastoparan

11
1.2.1.1 Các enzyme
Hyaluronidase và Phospholipase là hai enzyme chính có trong nọc độc
của các loài ong. Hai enzyme này có thể kích hoạt phản ứng miễn dịch, bằng
cách cảm ứng IgE gây ra các phản ứng quá mẫn dẫn đến phá huỷ lớp lipid kép
trong màng tế bào, gây độc tế bào hoặc kích thích miễn dịch đối với các loại tế
bào khác [15], [16].
Hyaluronidase thuỷ phân và cắt đặc hiệu acid hyaluronic, có thể được sử
dụng như các tác nhân gây mê, giảm đau, chống ung thư và cũng hỗ trợ sự phân
tán thuốc trong mô [17].
Phospholipase A-2 (PLA2) là thành phần chủ yếu thứ 2 của nọc ong,
chiếm 10-12% trọng lượng khô của nọc. PLA2 là một enzyme phụ thuộc vào
canxi mà thủy phân sn-2 este của glycerophospholipids để tạo ra một acid béo
và một lysophospholipid. Nó có thể phá hủy phospholipids, phá vỡ tính toàn
vẹn của lớp lipid kép, do đó làm cho các tế bào dễ bị suy thoái hơn. Trên thực
tế, các sản phẩm phản ứng PLA2, chẳng hạn như lysophosphatidylcholine, acid
lysophosphatidic và sphingosine 1- phosphate, có thể có tác dụng gây độc tế
bào hoặc kích thích miễn dịch trên các loại tế bào khác nhau, gây viêm và phản
ứng miễn dịch [16]
1.2.1.2 Các peptide
MCD peptide (Mast cell degranulating peptide) là một peptide tích điện
dương gồm 22 amino acid chứa 2 cầu disulfide giữa Cys3 và Cys15, Cys5 và
Cys19, có cấu trúc giống như apamin, [18]. Peptide này có tiềm năng kháng
viêm, tuy nhiên khi ở nồng độ thấp MCD peptide lại là chất trung gian mạnh
của quá trình phân rã tế bào mast và giải phóng chủ động histamine từ các tế
bào mast, mà được hiện diện trong việc truyền máu và trong tất cả các cô được
truyền dịch bằng máu [19].
Bradykinin là peptide thuộc nhóm protein kinin, tác động vào hai thụ thể
được ký hiệu là B1 và B2 gây đau và làm giảm huyết áp [19]. Đối với thụ thể
B1 chỉ được biểu hiện do tổn thương mô và nó được cho là có vai trò gây đau
mãn tính. Ngược lại, thụ thể B2 được biểu hiện chủ yếu, tham gia vào quá trình

12
giãn mạch thông qua việc giải phóng prostacyclin, oxit nitric, và yếu tố tăng
phân cực có nguồn gốc nội mô, do đó góp phần làm giảm huyết áp [19], [20].
Adolapin là peptide đầu tiên được tinh sạch từ nọc ong trong thập kỷ 80,
ức chế hoạt động của protstaglandin gây ra hiệu quả giảm đau và kháng viêm
mạnh khi thử nghiệm trên chuột [21], [22].
Melittin là peptide độc hoà tan trong nước chiếm 50% trọng lượng khô
của nọc. Đây là peptide xoắn lưỡng phần ( một phân tử có cả tính chất ưa nước
và kỵ nước ) bao gồm 26 gốc amino acid và trong đó cùng đầu N chủ yếu kỵ
nước và vùng đầu C là ưa nước. Melittin là thành phần hoạt động chính của nọc
ong (apitoxin) và chịu trách nhiệm phá vỡ và giết các tế vào. Khi một số peptide
melittin tích tụ trong màng tế bào, quá trình đóng gói phospholipid bị phá vỡ
nghiêm trọng, do đó dẫn đến ly giải tế bào [23]. Melittin và phospholipase A-2
ức chế sự tăng sinh tế bào khối u và do đó có giá trị tiềm năng cho chữa trị ung
thư và giảm khả năng nhiễm HIV-1 [24].
Apamin là độc tố thần kinh trong nọc ong gồm có 18 gốc amino acid với
hai cầu nối disulfide mà liên kết vị trí 1 với 11 và vị trí 3 với 15 [25]. Apamin
được biết từ rất lâu như là chất ức chế chọn lọc cao của các kênh K+ đã kích
hoạt Ca2+ (kênh SK) được biểu hiện trong hệ thống thần kinh trung ương. Loại
kênh này đóng một vai trò quan trọng trong các hoạt động lặp đi lặp lại trong
các tế bào thần kinh, ngăn chặn nhiều hiệu ứng ức chế siêu phân cực [26].
Mastoparan là một peptide lưỡng phần hoạt động trên màng với 14
amino acid, cảm ứng sự vận chuyển tính thấm của màng nên ảnh hưởng đến
khả năng sống sót của tế bào [27].
1.2.1.3 Các phân tử nhỏ khác
Ngoài 2 thành phần cơ bản ở trên, trong nọc ong còn chứa một số phân
tử nhỏ khác như các chất khoáng, các amino acid và các chất có hoạt tính giống
như amin.
Trong nhóm này kể đến histamine là một chất chính điển hình, tham gia
vào quá trình gây viêm thông qua việc làm tăng tính thấm của thành mao mạch
[22].
13
Tương tự như vậy noradrenalin và dopamine làm tăng nhịp đập của tim
và gây ra tăng tuần hoàn máu. Tuy nhiên giống như histamin, tác dụng của hai
catecholamine này phần lớn bị lu mờ bởi những thành phần khác của nọc độc
[22].
Serotonin hoạt động như một chất kích thích và góp phần gây ra những
cơn đau.
Cuối cùng, acetylcholine có thể làm tăng cảm giác đau của một vết chích
bằng cách kích thích thụ thể đau đồng thời với tác dụng của histamine [22].
Bảng 1.1 Một số protein và peptide chính được tìm thấy trong nọc ong [22].
Nọc ong mật
(bee)
Nọc ong vò vẽ
(wasp) Kiểu và MW (Da)
% Tỉ lệ trong
nọc khô
Phospholipase A2 Phospholipase A2 Enzyme (~18 kDa) 10-12
Phospholipase B Phospholipase B Enzyme (~26 kDa) 1
Hyaluronidase Hyaluronidase Enzyme (~54 kDa) 1.5-2
Phosphatase Phosphatase Enzyme (~60 kDa) 1
- Gluosidase - Gluosidase Enzyme (~170 kDa) 0.6
Melittin - Peptide (2847 Da) 40-50
Apamin - Peptide (2027 Da) 2-3
MCD peptide - Peptide (2593 Da) 2-3
- Mastoparan Peptide (1422 Da) -
- Bradykinin Peptide (1060 Da) -

14
1.2.2 Ứng dụng của nọc ong
Nọc ong là một chất độc với con người, vì vậy hiểu biết cặn kẽ đặc tính
về thành phần phân tử, tác dụng phụ và độc học của chúng đóng vai trò đặc biệt
quan trọng trong việc bảo vệ con người cũng như tìm kiếm các thuốc mới một
cách an toàn, hiệu quả. Nọc ong mật đã được nghiên cứu rất nhiều trong thế kỷ
trước [18], [28]. Trình tự của hệ gen ong mật đã được công bố vào năm 2006
[29] và dữ liệu tổng thể nọc ong mật đã có được từ năm 2005 trên Peptide Atlas.
Tổng số có 2288 protein ong mật đã được xác định với ít nhất 2 peptide với
FDR (false discovery rate) là 1,2 % [30].
Các thành phần peptide trong nọc độc của ong rất khác nhau về cấu trúc
hóa học và có nhiều tác dụng dược lý trên các đích đặc hiệu. Chúng được tạo
ra từ quá trình tiến hóa hàng triệu năm để đối phó với sự cạnh tranh khắc nghiệt
của môi trường sống. Các hợp chất này đặc biệt hữu dụng và triển vọng trong
các nghiên cứu sinh y học nhằm tìm cơ chế tác động ở trạng thái bệnh lý và
bình thường trên cơ thể người cũng như trong việc thiết kế tạo các dược phẩm
mới.
Trong lịch sử y dược thế giới, nọc ong đã được sử dụng trong y học
phương Đông (Trung Quốc và Hàn quốc) và trong cả thời kỳ Ai cập và Hy Lạp
cổ đại để kiểm soát một số bệnh rối loạn thần kinh [31], [32]. Trong những năm
trước, các nghiên cứu chú trọng đến lợi ích của nọc ong như bảo vệ phóng xạ
[33], chống đột biến [34], chống viêm [35], chống kích hoạt đau [36].
Gần đây, nọc ong không chỉ được phát hiện có tiềm năng điều trị bệnh
trên mà nó còn có khả năng sử dụng cho một số mục đích khác như kháng ung
thư [36], kháng khuẩn [22] làm suy giảm sự nhiễm HIV-1 [24], v.v.
Dược phẩm chứa melittin có tác dụng kháng viêm giảm đau mạnh gấp
nhiều lần hydrocortison, dùng trong bệnh thấp khớp, dùng trong phẫu thuật
chỉnh hình và thẩm mỹ. Một vài loại thuốc sử dụng nọc ong mật thô đã được
đăng ký và xuât hiện trên thị trường Châu Âu và toàn cầu như Forapin (Đức),
Virapin (Slovakia), Apiven (Pháp), Melivenon (Bulgaria), và Apifor (Nga).
Trong những năm gần đây, nọc ong và các thành phần của nó đã cho thấy

15
hoạt tính kháng khuẩn tiềm năng. Jalaei và cộng sự (2014) đã thực hiện sàng
lọc tác dụng kháng khuẩn của nọc độc thô Vespa Orientalis trên hai vi khuẩn
gram dương (Staphylococcus aureus và Bacillus subtillis) và hai gram âm
(Escherichia coli và Klesiella pneumonia). Kết quả cho thấy nọc độc của Vespa
Orientalis có tác dụng đáng kể đối với các chủng vi khuẩn được thử nghiệm (ở
nồng độ nọc độc 30 µg/ khoanh giấy, vùng ức chế là 12,6; 22,7; 22,4 và 10,2
mm đối với S.aureus, B.subtilis, E. coli, và K.pneumonia tương ứng) [37].
Các peptide được nghiên cứu đặc tính và được tách chiết gần đây cho
thấy hoạt tính kháng khuẩn, đóng vai trò quan trọng trong việc ngăn ngừa sự
lây nhiễm tiềm ẩn bởi vi sinh vật trong quá trình tiêu thụ con mồi của ấu trùng
côn trùng. Melittin đã thể hiện đặc tính kháng khuẩn mạnh và nó cũng có hoạt
tính tan máu và đặc tính gây dị ứng rõ rệt [38]. Các peptide kháng khuẩn này
không chỉ được áp dụng để điều trị ở người mà còn để chống lại các mầm bệnh
cây trồng quan trọng về mặt kinh tế mà hiện nay đang là một trong những yếu
tố hạn chế chính trong việc sản xuất cây trồng trên toàn thế giới [39]. Các
peptide ly giải như mastoparan và anoplin cũng thể hiện khả năng ức chế tăng
trưởng mạnh đối với vi khuẩn gram dương và gram âm, vì chúng có thể chèn
vào lớp lipid kép của vi khuẩn và hình thành lỗ.
Như vậy có thể nói rằng nọc ong là nguồn cung cấp dồi dào nhiều loại
thuốc khác nhau phục vụ sức khỏe con người nếu ta nghiên cứu phát hiện được
ra chúng. Ong có thể giết hại hàng nghìn người mỗi năm nhưng nọc độc chết
người của nó tiềm tàng khả năng cứu chữa cho hàng nghìn người khác.
1.3 MASTOPARAN
1.3.1 Cấu tạo
Mastoparan là một peptide lưỡng phần, cation, ngắn, hoạt động màng,
gồm 10 tới 14 gốc amino acid, giàu các gốc kỵ nước như leucine, isoleucine,
valine or alanine, 2-4 gốc lysine, trong môi trường TFE tạo thành dạng cấu trúc
xoắn lưỡng phần, đặc biệt cấu trúc amin hoá đầu C đã tạo nên những hoạt tính
quý của mastoparan.
Mastoparan đại diện cho lớp thành phần phong phú nhất của các peptide

16
trong nọc độc của các các loài ong săn mồi riêng lẻ và bày đàn [40], [41]. Chúng
được tìm thấy trong các chi như Vespa, Parapolybia, Protonectarina, Polistes,
và Protopolybia [22].
Hinh 1.6 Cấu trúc 3D của Mastoparan X được xác định bằng NMR [42].
Mastoparan đầu tiên được nhận dạng, tách chiết và nghiên cứu đặc tính
hóa học bởi Harai và cộng sự [43] từ ong săn mồi bầy đàn Vespula lewisii. Hirai
và cộng sự [44] cũng là những người đầu tiên tinh chế và nghiên cứu đặc tính
của mastoparan từ Vespa xanthoptera. Từ đó, các peptide mastoparan tương tự
đã được nhận diện và nghiên cứu đặc tính từ nọc của nhiều loài ong đất và ong
săn mồi bầy đàn.
Kể từ năm 1979, các nghiên cứu về cấu trúc đã đề xuất một cấu trúc xoắn
ốc, dưới dạng một bánh xe lưỡng phần (vừa có tính ưa nước vừa có tính kỵ
nước). Todokoro và cộng sự (2006) đã sử dụng kỹ thuật cộng hưởng từ hạt nhân
(NMR) để xác nhận cấu trúc này của mastoparan được tách chiết từ nọc độc
của ong Vespa xanthoptera (Hình 1.6) [45]. Trong cấu trúc xoắn của nó,
mastoparan thường tạo thành một bánh xe lưỡng phần, trong đó tất cả các gốc
lysine nằm ở một phía của vòng xoắn và tất cả các gốc acid amin kỵ nước được
đặt ở phía đối diện [45].
Lin và cộng sự (2011) đã so sánh các đặc tính sinh học và cấu trúc của
các mastoparan trong nọc độc của sáu loài Vespa: V. affinis, V. analis, V.
basalis, V. ducalis, V. mandarinia, và V. velutina flavitarsus ở Đài Loan. Tiền
thân của các mastoparan này được bao gồm một trình tự tín hiệu đầu N, một
prosequence, mastoparan trưởng thành và một glycine bổ sung ở đầu C. Trình
tự tín hiệu bao gồm 23 gốc amino acid, là giống nhau ở tất cả sáu mastoparan

17
tiền thân [46]. Các mastoparan trưởng thành, sau khi cải biến sau dịch mã, có
các đặc tính đặc trưng của các peptide điện tích dương mạch thẳng (giàu các
amino acid cơ bản và kỵ nước, không có liên kết disulfua). Chúng được mong
đợi như một cấu trúc bậc hai xoắn α lưỡng phần.
Quá trình tổng hợp mastoparan bắt đầu với các polypeptide tiền thân bao
gồm một trình tự tín hiệu vùng đầu N, một prosequence anion, một peptide
mastoparan trưởng thành và một glycine thêm vào ở đầu C. Từ dạng này, quá
trình cải biến sau dịch mã bao gồm việc phân tách của trình tự tín hiệu, loại bỏ
prosequence và amin hóa đầu C, mastoparan trưởng thành được tạo ra [47].
Signal sequence Prosequence Mature peptide
MP-AF MKNTILILFTAFIALLGFFGMSA EADPIADPIADPISGPNAEADPEA INLKAIAALAKKLFG 62
MP-A MKNTILILFTAFIALLGFFGMSA E--ALADPIADPLAGPNAEADPEA IKWKAILDAVKKVIG 60
MP-B MKSTILILFTAFIALLGFFGMSA E--ALADPLAEPLADPNAEADPEA LKLKSIVSWAKKVLG 60
MP-D MKNTILILFTAFIALLGFFGMSA E--ALADPIADPVAGPNPEADPEA INLKAIAAFAKKLLG 60
MP-M MKNTILILFTAFIALLGFFGMSA E--ALADPKADPLAGPNPDADPEA INLKAIAALAKKLLG 60
MP-V MKNTILILFTAFIALLGFFGMSA E--ALADPVADPLAGPNAEADPEA INWKGIAAMAKKLLG 60
**.******************** * :*** *:*::.** :***** :: *.* .**::*
Hinh 1.7 Sự so sánh trình tự của các gốc amino acid của các polypeptide tiền
thân của mastoparan.
Sự amin hóa đầu C của các mastoparan là một đặc tính chung của các
loài thuộc chi Vespa [43-44], [48], [49]. Nó làm bền cấu trúc xoắn α bằng cách
cung cấp thêm một liên kết hydro [46]. Hơn thế nữa, hoạt tính kháng khuẩn của
các mastoparan được amin hóa được tăng cường so với các peptide được khử
amin trong các loài ong săn mồi Polistinae [50].
Mastoparan có một điện tích dương toàn phần (cation) và một cấu trúc
xoắn α lưỡng phần (một phân tử có cả tính chất ưa nước và kỵ nước), trong đó
tất cả các chuỗi bên của các amino acid kỵ nước là nằm một phía của trục và
bazo của các amin acid này hoặc các gốc amino acid ưa nước là nằm ở vị trí
phía đối diện. Các peptide này là các phân tử polycation có hoạt tính sinh học
mà có thể đính với các màng sinh học và hình thành các lỗ thông qua các chuỗi
18
bên được tích điện dương của các cấu trúc xoắn α lưỡng phần, dẫn tới tính thấm
màng tế bào được tăng lên [51].
Hầu hết các mastoparan là các tetradecapeptide có chứa hai tới bốn gốc
Lysine (Lys, K) ở các vị trí 4/5 và hoặc 11/12 [52]. Một ngoại lệ thú vị là
peptide EMP-AF, trong đó các gốc Lys ở vị trí 5, 9 và 12. Sự định vị các gốc
lysine ở các vị trí này góp phần duy trì cấu trúc lưỡng phần và xoắn bền vững
và có thể góp phần vào hoạt tính ly giải của các peptide này [53], [54].
Bảng 1.2 Trình tự amino acid của một số peptide mastoparan từ nọc của một
số loài ong
Tên (Số truy cập) Trình tự amino acid Khối
lượng
(Da)
Loài
Agelaia-mastoparan
(P69436)
INWLKLGKAI IDAL 1568 Agelaia pallipes
pallipes
Eumenine mastoparan-
AF (P0C022)
INLLKIAKGI IKSL 1524 Anterhynchium
flavomarginatum
micado
Eumenine mastoparan-
ER
(P0CJ38)
FDIMGLIKKV AGAL 1476 Eumenes
rubrofemoratus
(Solitary wasp)
Mastoparan
(P42716)
INWKKMAATALKMI 1619 Parapolybia indica
Mastoparan-J
(P01517)
VDWKKIGQHILSVL 1636 Polistes jokahamae
Mastoparan-like
peptide PMM2
INWKKIASIG
KEVLKAL
1911 Polistes major
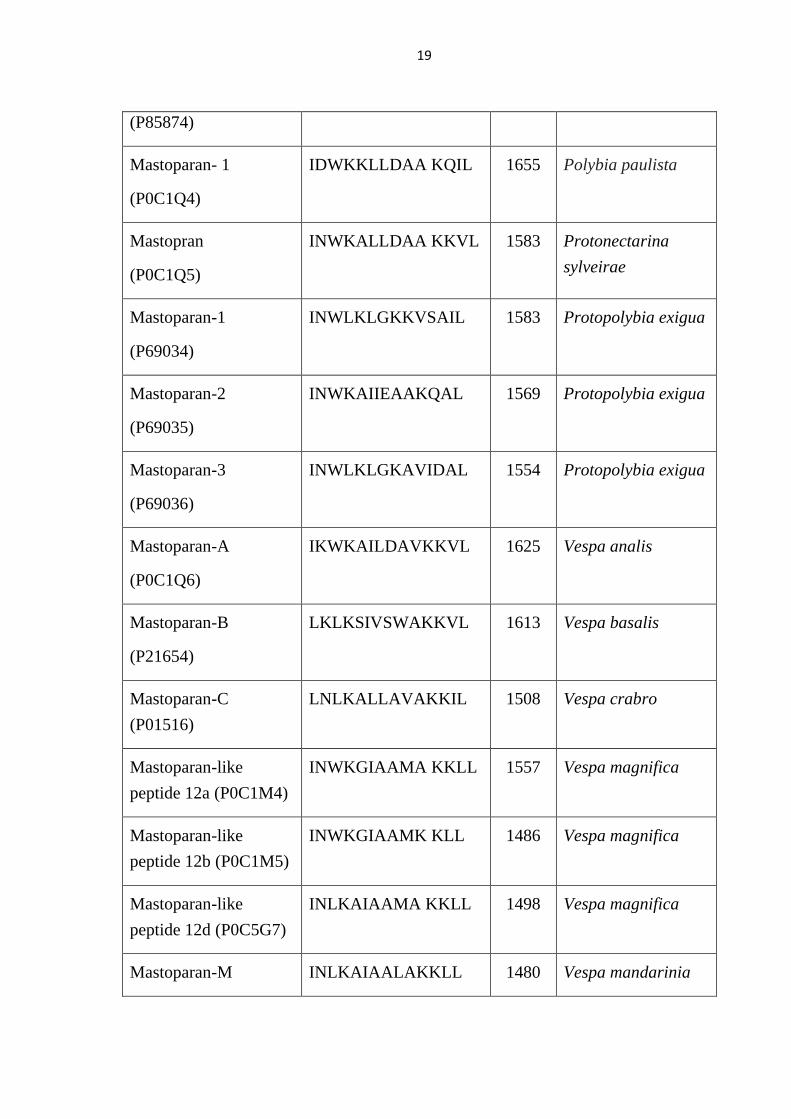
19
(P85874)
Mastoparan- 1
(P0C1Q4)
IDWKKLLDAA KQIL 1655 Polybia paulista
Mastopran
(P0C1Q5)
INWKALLDAA KKVL 1583 Protonectarina
sylveirae
Mastoparan-1
(P69034)
INWLKLGKKVSAIL 1583 Protopolybia exigua
Mastoparan-2
(P69035)
INWKAIIEAAKQAL 1569 Protopolybia exigua
Mastoparan-3
(P69036)
INWLKLGKAVIDAL 1554 Protopolybia exigua
Mastoparan-A
(P0C1Q6)
IKWKAILDAVKKVL 1625 Vespa analis
Mastoparan-B
(P21654)
LKLKSIVSWAKKVL 1613 Vespa basalis
Mastoparan-C
(P01516)
LNLKALLAVAKKIL 1508 Vespa crabro
Mastoparan-like
peptide 12a (P0C1M4)
INWKGIAAMA KKLL 1557 Vespa magnifica
Mastoparan-like
peptide 12b (P0C1M5)
INWKGIAAMK KLL 1486 Vespa magnifica
Mastoparan-like
peptide 12d (P0C5G7)
INLKAIAAMA KKLL 1498 Vespa magnifica
Mastoparan-M INLKAIAALAKKLL 1480 Vespa mandarinia
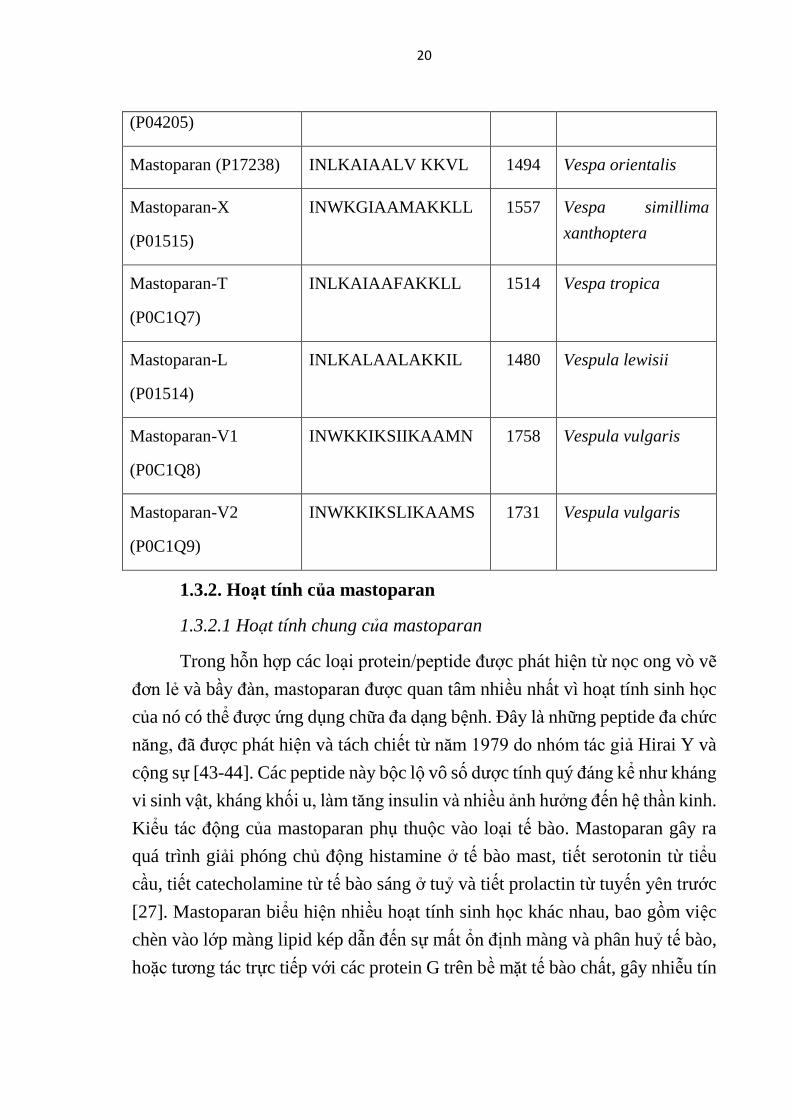
20
(P04205)
Mastoparan (P17238) INLKAIAALV KKVL 1494 Vespa orientalis
Mastoparan-X
(P01515)
INWKGIAAMAKKLL 1557 Vespa simillima
xanthoptera
Mastoparan-T
(P0C1Q7)
INLKAIAAFAKKLL 1514 Vespa tropica
Mastoparan-L
(P01514)
INLKALAALAKKIL 1480 Vespula lewisii
Mastoparan-V1
(P0C1Q8)
INWKKIKSIIKAAMN 1758 Vespula vulgaris
Mastoparan-V2
(P0C1Q9)
INWKKIKSLIKAAMS 1731 Vespula vulgaris
1.3.2. Hoạt tính của mastoparan
1.3.2.1 Hoạt tính chung của mastoparan
Trong hỗn hợp các loại protein/peptide được phát hiện từ nọc ong vò vẽ
đơn lẻ và bầy đàn, mastoparan được quan tâm nhiều nhất vì hoạt tính sinh học
của nó có thể được ứng dụng chữa đa dạng bệnh. Đây là những peptide đa chức
năng, đã được phát hiện và tách chiết từ năm 1979 do nhóm tác giả Hirai Y và
cộng sự [43-44]. Các peptide này bộc lộ vô số dược tính quý đáng kể như kháng
vi sinh vật, kháng khối u, làm tăng insulin và nhiều ảnh hưởng đến hệ thần kinh.
Kiểu tác động của mastoparan phụ thuộc vào loại tế bào. Mastoparan gây ra
quá trình giải phóng chủ động histamine ở tế bào mast, tiết serotonin từ tiểu
cầu, tiết catecholamine từ tế bào sáng ở tuỷ và tiết prolactin từ tuyến yên trước
[27]. Mastoparan biểu hiện nhiều hoạt tính sinh học khác nhau, bao gồm việc
chèn vào lớp màng lipid kép dẫn đến sự mất ổn định màng và phân huỷ tế bào,
hoặc tương tác trực tiếp với các protein G trên bề mặt tế bào chất, gây nhiễu tín

21
hiệu màng tế bào [55], [56] hoặc kích thích phospholipase, tăng nồng độ Ca 2+
từ ty thể và mạng lưới nội bào, gây hoại tử tế bào hoặc quá trình apoptosis [57],
[58]. Hiện nay, mastoparan đang được nghiên cứu khả năng kháng khuẩn, phân
huỷ tế bào máu và tiêu diệt các tế bào ung thư [46], [59-62].
1.3.2.2 Hoạt tính kháng khuẩn của mastoparan
Trong 30 năm qua, người ta đã quan sát thấy sự gia tăng đáng kể của các
siêu vi khuẩn phân lập trong môi trường lâm sàng, đặc biệt là các vi khuẩn từ
nhóm được gọi là ESKAPE (Enterococcusfoecium, Staphylococcus aureus,
Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa,
Enterobacter spp.) cho thấy khả năng đề kháng cao đối với tất cả các chất kháng
khuẩn hiện có. Sự xuất hiện của các vi khuẩn kháng kháng sinh là một mối đe
dọa tới loài người trên toàn thế giới và do đó đã có một sự quan tâm đáng kể
trong sự phát triển của một lớp chất kháng khuẩn mới như một phương pháp trị
liệu để điều trị các bệnh truyền nhiễm.
Việc tìm kiếm các chất kháng sinh mới đã tìm thấy một nguồn gần như
vô tận của các chất điều trị tiềm năng trong số các peptide kháng khuẩn, với
các nguồn bao gồm cả nọc côn trùng. Các peptide kháng khuẩn là một nhóm
peptide đa dạng và độc đáo có chức năng thiết yếu trong đáp ứng miễn dịch
bẩm sinh của vật chủ khi bị xâm nhập với các sinh vật gây bệnh. Tuy nhiên, sự
hứa hẹn tiềm ẩn đã trở thành những trở ngại đầy thách thức, chẳng hạn như các
khó khăn và chi phí tổng hợp các chuỗi peptide dài hơn, phạm vi hoạt động hẹp
hơn, độ nhạy của chuỗi peptide ngắn, độc tính thấp đối với vật chủ và tính phù
hợp của dược động học thuộc tính. Trong số nhiều peptide kháng khuẩn đã biết,
mastoparan và các peptide tương tự mastoparan là những ứng cử viên có đặc
điểm tốt nhất. Các mastoparan được tìm thấy trong nọc ong săn mồi, chúng đại
diện cho các ví dụ điển hình về peptide kháng khuẩn đã được nghiên cứu [63-
68]. Mastoparan là các peptide kháng khuẩn với 14 gốc amino acid được amin
hóa đầu C, có tác dụng rộng rãi chống lại vi khuẩn gram dương và gram âm và
các loài nấm [59].
Mastoparan từ nọc của các loài ong đất khác các loài Vespa cũng như V.
basalis [69], [70] và V. magnifica đã được báo cáo có hoạt tính kháng khuẩn.

22
Chen và cộng sự (2008) đã tinh sạch và nghiên cứu đặc tính của
mastoparan từ nọc của ong Vespa bicolor [71]. Peptide này cho thấy hoạt tính
kháng khuẩn chống lại các chủng nấm, vi khuẩn gram âm, vi khuẩn gram dương
chuẩn được thử nghiệm. Hơn thế nữa, nó có hoạt tính kháng khuẩn mạnh chống
lại các chủng vi khuẩn kháng thuốc được phân lập lâm sàng, nhưng hầu như
không thể hiện hoạt tính tan huyết chống lại các tế bào hồng cầu.
Yang và cộng sự (2013) đã tinh sạch hai mastoparan (VT1 và VT2) và
đã nhận diện năm peptide giống mastoparan bổ sung thuộc các peptide kháng
khuẩn tự nhiên từ tuyến độc của ong đất Vespa tropica [72]. Một điều thú vị,
trình tự amino acid của mastoparan VT1 (NLKAIAALAKKLL-NH2) là giống
hoàn toàn với mastoparan M từ Vespa mandarinia [49]. Mastoparan VT2 có
một isoleucine đầu N ngắn hơn so với mastoparan VT1. Hoạt tính kháng khuẩn
của các peptide này được thử nghiểm chống lại các chủng vi khuẩn, nấm chuẩn
và các chủng vi khuẩn kháng thuốc được phân lập từ lâm sàng. Chúng đã chứng
minh hoạt tính kháng khuẩn phổ rộng chống lại tất các các chủng thử nghiệm
bao gồm nấm, cả vi khuẩn gram âm và gram dương và có thể cung cấp các mẫu
cho phát triển các kháng sinh peptide mới [72].
Nghiên cứu đã được thảo luận trước đây của mastoparan trong nọc độc
của sáu loài ong Vespa của Lin và cộng sự (2011) [46] chứng minh rằng tất cả
sáu mastoparan cho thấy hoạt tính kháng khuẩn chống lại cả vi khuẩn gram âm
và gram dương được thử nghiệm, và sự thẩm thấu màng của các tế bào E.coli.
Các nghiên cứu sâu hơn về hoạt tính kháng khuẩn của mastoparan từ V. affinis,
mastoparan AF cũng đã được thực hiện bởi nhóm tác giả này [59], [73]. Họ đã
chứng minh rằng mastoparan AF có hoạt tính kháng khuẩn chống lại chủng
E.coli đa kháng thuốc được phân lập từ các động vật [59] và các chủng
Acinetobacter baumannii lâm sàng đa kháng thuốc được phân lập từ máu của
các bệnh nhân [73]. Ngoài ra, các kết quả đã cho thấy rằng mastoparan AF thể
hiện sự tan huyết không đáng kể hồng cầu người ở nồng độ kháng khuẩn hiệu
quả của nó [46], và kết hợp với các thuốc kháng sinh nhất định chống lại một
số vi sinh vật kháng đa kháng sinh, điều này gợi ý rằng mastoparan AF đơn lẻ
hoặc kết hợp với các thuốc kháng sinh khác có thể được phát triển như một loại

23
thuốc hứa hẹn chống lại các vi khuẩn cho các ứng dụng lâm sàng [59], [73].
Tầm quan trọng của các gốc Lys tới hoạt tính kháng khuẩn của
mastoparan cũng đã được điều tra. Yoon và cộng sự (2015) cho thấy
mastoparan của V. analis với bốn gốc Lys cho thấy hoạt tính kháng khuẩn và
kháng khối u tốt hơn so với mastoparan từ V. crabro với ba gốc Lys [74]. Họ
đã làm sáng tỏ điều này có thể là do gốc Lys bổ sung trong peptide mastoparan
trưởng thành của V. analis. Li và cộng sự (2000) cho thấy rằng hoạt tính kháng
khuẩn của D-mastoparan là tốt hơn so với mastoparan tự nhiên. Họ sử dụng D-
mastoparan được tổng hợp hóa học, chứa D-amino acid thay vì các dạng L tự
nhiên, cái mà có thể giết vi khuẩn bằng việc phá vỡ các tế bào [75]. Tiềm năng
các mastoparan tổng hợp tương tự như các chất kháng khuẩn được điều tra sâu
hơn bởi Chen và cộng sự (2018) [76]. Họ đã thiết kế hai mô hình của
mastoparan C (V. crabro) với mục đích cải thiện hoạt tính sinh học của chúng
và khả năng thẩm thấu màng. Nghiên cứu tiết lộ rằng mastoparan C là một ứng
cử viên tốt cho việc phát triển các chất kháng khuẩn.
1.3.2.3 Cơ chế hoạt động của các mastoparan chung trên màng của vi
sinh vật
Các mastoparan có chung một đặc tính sinh lý về hoạt tính kháng vi sinh
vật của chúng, peptide này có tổng điện tích dương, có lực hút tĩnh điện đối với
các bề mặt vi sinh vật tích điện âm, và tạo thành một chuỗi xoắn α lưỡng phần,
kết quả của sự phơi bày các gốc kỵ nước trên bề mặt cho việc cài vào phần kỵ
nước của bề mặt màng vi sinh vật [77], [78]. Trong quá trình tương tác của
chúng với màng, các mastoparan đã được biết là gây ra sự phá vỡ màng thông
qua một cơ chế như mô hình thảm (carpet model), mô hình có hình xuyến
(toroidal model) hoặc mô hình thùng (barrel-stave model), tuy nhiên, không có
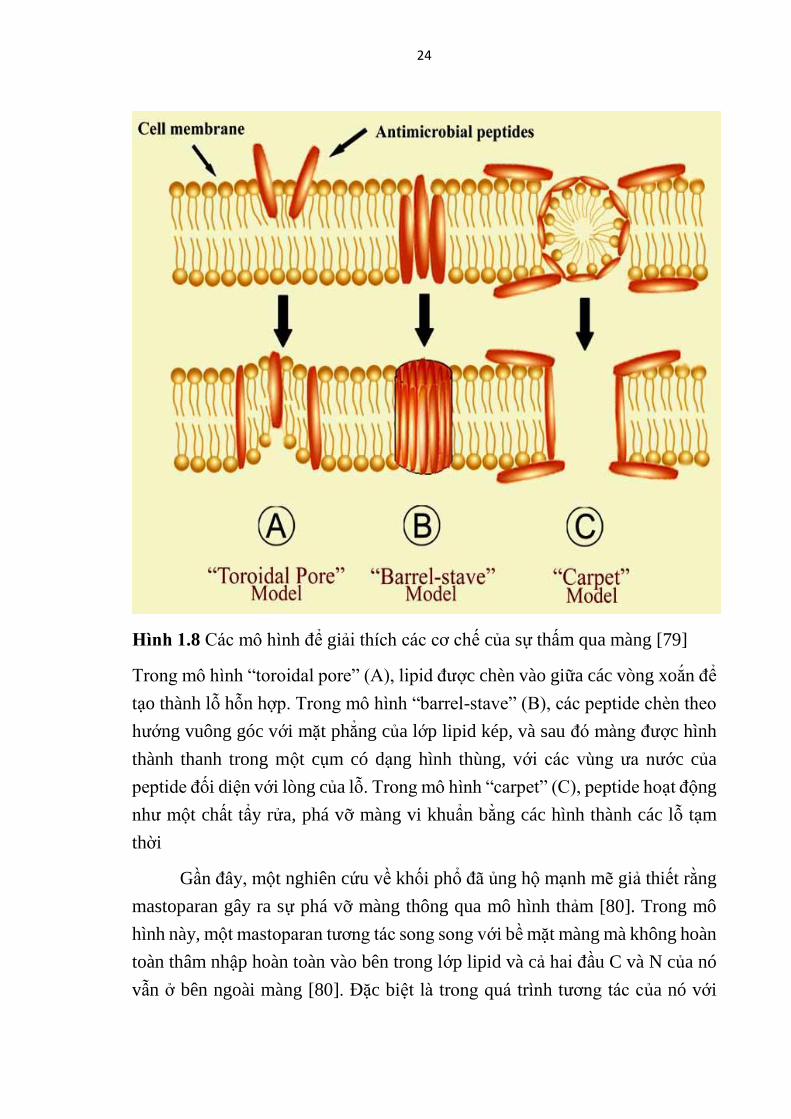
sự mô tả rõ ràng nào thu được cho đến nay.
24
Hinh 1.8 Các mô hình để giải thích các cơ chế của sự thấm qua màng [79]
Trong mô hình “toroidal pore” (A), lipid được chèn vào giữa các vòng xoắn để
tạo thành lỗ hỗn hợp. Trong mô hình “barrel-stave” (B), các peptide chèn theo
hướng vuông góc với mặt phẳng của lớp lipid kép, và sau đó màng được hình
thành thanh trong một cụm có dạng hình thùng, với các vùng ưa nước của
peptide đối diện với lòng của lỗ. Trong mô hình “carpet” (C), peptide hoạt động
như một chất tẩy rửa, phá vỡ màng vi khuẩn bằng các hình thành các lỗ tạm
thời
Gần đây, một nghiên cứu về khối phổ đã ủng hộ mạnh mẽ giả thiết rằng
mastoparan gây ra sự phá vỡ màng thông qua mô hình thảm [80]. Trong mô
hình này, một mastoparan tương tác song song với bề mặt màng mà không hoàn
toàn thâm nhập hoàn toàn vào bên trong lớp lipid và cả hai đầu C và N của nó
vẫn ở bên ngoài màng [80]. Đặc biệt là trong quá trình tương tác của nó với

25
màng, mastoparan tạo thành chuỗi xoắn α để tối ưu hóa tính chất lưỡng phần,
thay đổi cấu trúc ở giữa peptide để làm cho cấu trúc của nó thuận lợi hơn về
mặt năng lượng và dẫn đến sự mất ổn định của màng kép, trong khi dưới các
điều kiện nước, nó hình thành chuỗi cuộn ngẫu nhiên [80]. Đáng chú ý, đầu C
của mastoparan có gắn nhóm amide kích hoạt sự ổn định của các cấu trúc xoắn
ốc của chúng, cho phép cài vào màng tế bào của vi khuẩn và động vật [80]. Do
đó, tổng điện tích dương, tính kỵ nước và sự ổn định của cấu trúc xoắn nên
được xem xét để cải thiện hoạt tính kháng khuẩn của các mastoparan.
1.4 TÌNH HÌNH NGHIÊN CỨU TẠI VIỆT NAM
Theo đông y, có thể sử dụng nọc ong trong điều trị một số chứng bệnh
khá hiệu quả, như u bã đậu (mà không cần phẫu thuật), cai nghiện ma túy, viêm
khớp, v.v.. Đã có nhiều nghiên cứu cho thấy tác dụng chữa bệnh của nọc ong
như dùng làm các dạng cao xoa giảm đau, chống viêm, cho ong đốt trực tiếp
vào các huyệt tương ứng trên cơ thể. Việc ứng dụng cách chữa bệnh bằng nọc
ong này từng được tiến hành ở một số quốc gia trên thế giới, trong đó có cả
Việt Nam. Song ở Việt Nam, phương pháp này hiện vẫn chưa được phổ biến
rộng rãi.
Những nghiên cứu về khảo sát, đặc tính hóa sinh của các protein nọc ong
ở Việt Nam còn khá hạn chế. Từ thế kỷ trước, trong khoảng thời gian từ năm
1966-1970, GS. Nguyễn Năng An cùng nhóm cộng sự của ông đã thực hiện đề
tài "Nghiên cứu nọc ong trên thực nghiệm". Trong nghiên cứu này, các ong thợ
thuộc giống Apisarthron của Ðức được chọn lọc có độ tuổi từ 15-40 ngày tuổi,
khỏe mạnh cho việc lấy nọc độc. Sau đó, nọc ong cùng chế phẩm nọc ong để
tiêm tĩnh mạch chó, thỏ, chuột lang, chuột nhắt, chuột cống trắng, ếch. Tiếp
theo đó GS. Nguyễn Năng An đã hợp tác cùng GS. Nguyễn Tài Thu tại Trạm
E, Sở Y tế Hà Nội thực hiện đề tài đề tài "Thủy châm chữa một số bệnh bằng
sản phẩm ong" [81].
Năm 2016, PGS.TS. Cấn Văn Mão và cộng sự đã thực hiện xây dựng
quy trình chiết xuất nọc ong (Apis mellifera) và đánh giá tác dụng dược lý theo
hướng sử dụng hỗ trợ điều trị viêm khớp” [82]. Kết quả cho thấy các thành
phần chính của nọc ong như apamin, phospholipase A2 và melittin được xác

26
định bằng phương pháp RP-HPLC/Q-TOF [83]. Độc tính cấp tính, bán trường
diễn của nọc ong trên động vật thực nghiệm cũng đã được xác định [82]. Cụ
thể LD50 được xác định là 20,5 mg/kg, nọc ong ở liều 2 mg/kg/ngày và liều
6mg/kg/ngày không gây độc tính bán trường diễn trên chuột. Các thí nghiệm
nghiên cứu trên động vật còn cho thấy nọc ong óc khả năng giảm đau, kháng
viêm. Nhóm tác giả đã định hướng nghiên cứu ứng dụng nọc ong trong điều trị
viêm khớp ở người.
Trong giai đoạn 2017-2020, nhóm nghiên cứu của PGS. TS. Nguyễn Bá
Quang và cộng sự đã thực hiện đề tài “Nghiên cứu tác dụng chống viêm giảm
đau của thủy châm chế phẩm chứa nọc ong trên bệnh nhân thoái hóa cột sống,
thoái hóa khớp gối, viêm quanh khớp vai”. Kết quả cho thấy, chế phẩm nọc ong
có tác dụng chống viêm, giảm đau, cải thiện vận động trên động vật thực
nghiệm được thuỷ châm. Chế phẩm nọc ong có thể được ứng dụng trong điều
trị các bệnh lý xương khớp mạn tính [84].
Tại Viện Công nghệ Sinh học có nhóm nghiên cứu của PGS.TS. Nguyễn
Thị Ngọc Dao đã nghiên cứu tạo phân tử hai chức năng Carrier-melittin và đã
tạo được phân tử protein tái tổ hợp có khả năng gắn kết và làm tan các tế bào
lympho tách từ máu ngoại vi của bệnh nhân mắc bệnh Hodgkin [85]. Trong
giai đoạn 2015-2016, TS. Lê Thị Bích Thảo và cộng sự đã thực hiện đề tài
“Nghiên cứu khảo sát các protein/peptide có hoạt tính kháng tế bào ung thư
trong nọc ong mặt quỷ Vespa velutina của Việt Nam”, kết quả đã có được cơ
sở dữ liệu đầu tiên về thành phần các protein/peptide có trong nọc ong Vespa
velutina thu thập ở Việt Nam [86]. Mastoparan đã được nhận diện trong tổng
số hơn 80 protein và peptide trong thành phần nọc ong Vespa velutina [86].
Mastoparan ngoài khả năng kháng ung thư cũng cho thấy tiềm năng
kháng khuẩn một mình hay khi điều trị phối hợp với các kháng sinh khác. Tuy
nhiên, Việt Nam có rất ít nghiên cứu về mastoparan. Đi theo hướng nghiên cứu
này, chúng tôi thực hiện đề tài “Nghiên cứu tinh sạch và xác định hoạt tính
kháng khuẩn của peptide mastoparan từ nọc ong Vespa velutina thu thập ở Việt
Nam” với mong muốn phát hiện một loại mastoparan mới từ loài ong này và

27
tìm kiếm khả năng ứng dụng trong y dược của chúng phục vụ đời sống con
người như một chất kháng sinh mới.

28
CHƯƠNG 2. NGUYÊN LIỆU VÀ PHƯƠNG PHÁP NGHIÊN CỨU
2.1 NGUYÊN VẬT LIỆU
2.1.1 Vật liệu sinh học
Ong Vespa velutina thu thập ở Việt Nam
Các chủng vi sinh vật
Các chủng vi sinh vật không kháng kháng sinh được cung cấp từ Viện
Sốt rét- Ký sinh trùng-Côn trùng Trung ương bao gồm:
+ Các chủng vi khuẩn gram âm:
- Escherichia coli (E. coli ATCC 25922),
- Shigella flexneri (S. flexneri ATCC 12022),
- Salmonella typhimurium (S. typhimurium ATCC 14028),
+ Chủng vi khuẩn gram dương:
- Staphylococus aureus (S. aureus ATCC 26923).
Các chủng vi sinh vật kháng kháng sinh bao gồm
+ Các chủng vi khuẩn gram âm được phân lập từ Việt Nam được cung
cấp từ Trung tâm bệnh nhiệt đới, Bệnh Viện Bạch Mai:
- Salmonella spp kháng ampicillin
- Klebsiella pneumonia (K. pneumonia) đa kháng kháng sinh
- Acinetobacter baumannii (A. baumannii) đa kháng kháng sinh
+ Chủng vi khuẩn gram dương được cung cấp từ Viện Công nghệ sinh
học, Viện Hàn lâm Khoa học và Công nghệ Việt Nam:
- Methicillin-resistant Staphylococcus epidermidis (MRSE ATCC
35984) kháng methicillin
29
2.1.2 Các hóa chất
Bảng 2.1 Các hóa chất chính được sử dụng trong nghiên cứu
Tên hóa chất Nhà sản xuất
Acetonitrile Merck (Đức)
Acid trifluoroacetic Sigma-aldrich
Acid formic Prolabo (VWR) (Mỹ)
Acid α-Cyano-4-hydroxycinnamic Thermo fisher scientific (Mỹ)
N-α F-moc amino acid Intavis Bioanalytical
Instruments AG (Đức)
Môi trường Mueller- Hinton lỏng, agar
(Beef extract, casein hydrolysate, starch,
agar)
Merck (Đức)
Môi trường BHI ( beef heart, calf brain,
disodium hydrogen phosphate, glucose,
peptone, sodium chloride
BD ( Mỹ)
Các thuốc kháng sinh: Imipenem (IMP –
10 µg), Levofloxacin (LEV – 5 µg),
Trimethoprim/Sulfamethoxazole (SXT-25
µg ), Ciprofloxacin (CIP- 5 µg), Ampicillin
(AMP- 10 µg, AMP-20 µg),
Ampicillin/sulbactam (SAM-20 µg),
Oxacilin (OXA)-20 µg, Kanamycin
(KAN)-30 µg, Gentamicin (GEN)- 20 µg
Oxoid (Anh)
30
2.1.3 Trang thiết bị
Bảng 2.2 Các thiết bị được sử dụng chính trong nghiên cứu
Thiết bị Nhà sản xuất
Máy vortex OSI (Mỹ)
Máy khấy từ IKA (Đức)
Máy ly tâm Thermo fisher Scientific (Mỹ)
Hệ thống SpeedVac Thermo fisher scientific (Mỹ)
Tủ lạnh -20oC, -80 oC Sanyo (Nhật Bản)
Cột sắc ký Luna C18 Phenomenex (Mỹ)
Hệ thống HPLC Shimadzu, (Nhật Bản)
Hệ thống máy khối phổ MALDI-TOF Bruker (Mỹ)
Hệ thống máy tổng hợp peptide Intavis Bioanalytical
Instruments AG (Đức)
Tủ cấy vi sinh (tủ an toàn sinh học) Sanyo (Nhật Bản)
Tủ ấm vi sinh Amerex Instruments (Mỹ)
Lò vi sóng Sharp (Nhật Bản)
Nồi hấp tiệt trùng Hirayama (Nhật Bản)

31
2.2 SƠ ĐỒ NGHIÊN CỨU CHUNG
Sơ đồ các bước thí nghiệm trong luận văn được tiến hành như trong Hình 2.1
2.3 PHƯƠNG PHÁP NGHIÊN CỨU
2.3.1. Thu thập và tách chiết nọc ong
Ong Vespa velutina cái được thu thập và phân loại dựa vào các đặc điểm
hình thái học bởi PGS. TS. Nguyễn Thị Phương Liên (Viện Sinh thái và Tài
nguyên sinh vật, Viện Hàn Lâm Khoa học và Công nghệ Việt Nam). Sau khi
thu thập, ong được làm đông lạnh ngay lập tức và bảo quản ở -20 oC để tránh
protein trong nọc ong bị phân huỷ. Vì nọc ong có rất ít nên để lấy được cơ quan
nọc độc của ong phải lựa chọn các con ong cái to và tiến hành giải phẫu để thu
nhận ngòi độc và túi chứa nọc độc của ong.
Thu thập, tách chiết nọc ong
Vespa velutina
Phân đoạn nọc ong và tinh sạch mastoparan
Nhận diện và xác định trình tự amino acid
của mastoparan
Tổng hợp hóa học mastoparan
Xác định hoạt tính kháng khuẩn của mastoparan
Hinh 2.1 Sơ đồ quy trình các bước tiến hành

32
Cơ quan nọc ong bao gồm phần ngòi (sting) và túi chứa nọc (reservoir)
được tách ra sau khi giải phẫu phần bụng cuối của ong. Tiến hành cắt nhỏ cơ
quan nọc ong và chiết xuất protein/peptide bằng dung dịch đệm ACN 50%
trong 3 h ở 4oC. Sau khi ly tâm 13000 vòng/phút trong 30 phút, thu dịch nổi và
làm khô không nhiệt bằng Speedvac để bảo đảm các protein không bị biến tính.
Sau khi làm khô, nọc ong được bảo quản -20oC cho các thí nghiệm tiếp theo.
2.3.2. Phân đoạn nọc ong và tinh sạch mastoparan
Hỗn hợp protein nọc ong sau khi được tách chiết và làm khô được hòa
trong dung dịch ACN 5% (v/v) và cho qua hệ sắc ký đảo pha HPLC với cột
Luna C18 (250 x 4,6 mm; 5µm) cho việc tinh sạch mastoparan [42]
Chuẩn bị dung môi
- Dung dịch A: Nước cất deion vô trùng
- Dung dịch B: Dung dịch ACN chứa 0,1% TFA.
Tất cả các dung dịch đều được lọc qua màng lọc 0,22 µm, khử khí bằng
siêu âm trước khi sử dụng. Việc này nhằm loại bỏ các hạt mà có thể ngăn cản
các dòng dung môi hoặc cột và cũng như các bộ khử khí trong dung môi.
Phân tích mẫu
100 μL mẫu được tiêm qua hệ thống tiêm mẫu tự động. Mẫu được đưa
lên cột Luna C18 column (250 x 4,6mm; 5µm) pha đảo và được rửa giải theo
một gradient nồng độ 5-60% acetonitrile có chứa 0,1% trifluoracetic acid ở tốc
độ dòng 2 mL/phút, trong 45 phút ở nhiệt độ phòng. Protein/peptide được đo ở
bước sóng 215 nm bằng PDA detector. Các phân đoạn được thu bằng tay trong
các ống Eppendorf 2 mL sạch, sau đó các phân đoạn được mang đi nhận diện
và phân tích MALDI-TOF.
2.3.3. Phân tích MALDI-TOF
Các phân đoạn thu được sau khi thực hiện sắc ký RP-HPLC sẽ được nhận
diện và giải trình tự bằng phân tích MALDI-TOF. Mẫu được hòa trong
0,1%FA/50%ACN. Chuẩn bị một lượng dung dịch matrix (chất nền) có chứa
25 mg CHCA trong 70% ACN/0,1% FA. 1 μl của dung dịch matrix được thêm

33
vào và trộn kỹ với 1 μl mẫu trước khi được đưa vào tấm thép không gỉ của máy
khối phổ. Phân tích MALDI-TOF được thực hiện. Các mẫu được phân tích trên
máy reflectron TOF mass spectrometer. Máy được vận hành ở chế độ ion dương
và tuyến tính ở một điện áp gia tốc 20 kV. Công suất laser được điều chỉnh đến
một giá trị cao hơn ngưỡng hấp phụ / ngưỡng ion hóa, và phổ khối lượng được
thu lại bằng cách lấy trung bình 100 phát bắn xung laser. Phổ được ghi lại sau
đó được xử lý bằng phần mềm FlexAnalysis (Bruker Daltonics). Phổ MS/MS
thu được được phân tích bằng phần mềm PEAK (Bioinformatics Solution Inc.,
Canada) cho việc nhận diện và xác định trình từ peptide.
2.3.4 Tổng hợp hóa học mastoparan bằng phương pháp tổng hợp
peptide pha rắn
Mastoparan được tổng hợp hóa học bằng phương pháp tổng hợp peptide
pha rắn trên máy tổng hợp tự động (Intavis Bioanalytical Instruments AG, Đức)
từ nguyên liệu là N-α F-moc amino acid theo hướng dẫn của nhà sản xuất. Các
α amino acid đã được khóa nhóm amin bằng nhóm F-moc (9-
fluorenylmethoxycarbonyl) còn khóa chuỗi bên bởi các dẫn xuất este, ether và
urethane của tert.-butanol. Amino acid cuối cùng của chuỗi peptide được gắn
vào một giá thể rắn bởi đầu cacboxyl liên kết hóa học với nhóm chức trên giá
thể đó, sau đó được tháo khóa nhóm amin bằng piperidine (loại bỏ nhóm khóa
Fmoc). Các amino acid liền kề tiếp theo đã được hoạt hóa phần cacboxyl và
khóa nhóm amin được bổ sung tiếp vào amino acid đã gắn trên giá thể và liên
kết peptide đã được hình thành. Sau mỗi bước liên kết với một amino acid của
chuỗi peptide, oligopeptide được lọc vả rửa để có được phân tử sạch trước khi
quá trình tổng hợp được tiếp tục. Các amino acid được liên kết với nhau theo
một chương trình đã được cài đặt trên máy tổng hợp tự động cho đến khi
mastoparan đã được tổng hợp. Cuối cùng, các peptide tổng hợp được loại bỏ
giá thể bằng acid flohydric. Hỗn hợp peptide thu được được phân tích bằng
khối phổ để đánh giá kết quả tổng hợp nhân tạo. Các mastoparan sau đó được
tinh sạch bằng sắc kí RP- HPLC sử dụng Cột C18 (250 x 4,6 mm; 5µm) nhằm
loại bỏ những peptide không mong muốn cũng như các amino acid còn dư trong
phản ứng.

34
2.3.5 Phương pháp khoanh giấy khuếch tán trên đĩa thạch
Chúng tôi đã tiến hành thử nghiệm hoạt tính kháng khuẩn của
mastoparan được tách chiết từ nọc ong Vespa velutina trên bốn chủng vi khuẩn
bao gồm: Escherichia coli (E. coli ATCC 25922), Shigella flexneri (S. flexneri
ATCC 12022), Salmonella typhimurium (S. typhimurium ATCC 14028) và
Staphylococus aureus (S. aureus ATCC 26923).
Thử nghiệm hoạt tính kháng khuẩn của mastoparan tổng hợp nhân tạo
trên bốn chủng vi khuẩn kháng kháng sinh bao gồm: Salmonella spp,
Klebsiella pneumonia (K. pneumonia), Acinetobacter baumannii (A.
baumannii), Methicillin-resistant Staphylococcus epidermidis (MRSE ATCC
35984)
Xác định hoạt tính kháng khuẩn của các peptide mastoparan được thực
hiện bằng phương pháp khoanh giấy khuếch tán trên đĩa thạch sử dụng môi
trường Muller- Hinton agar. Các chủng vi khuẩn được hoạt hóa qua đêm ở 37
ºC. Sau đó, chúng được cấy chuyển sang môi trường Muller-Hinton lỏng, nuôi
lắc ở 37 ºC cho đến khi mật độ tế bào đạt xấp xỉ 106 CFU/mL. Sử dụng các
mảnh giấy Whatman vô trùng (đường kính 6 mm) cho việc bổ sung mastoparan
với các nồng độ khác nhau hoặc các chất kháng sinh dùng làm đối chứng. Quan
sát sự phát triển của vi khuẩn sau thời gian 24 giờ nuôi ủ ở 36 ºC ± 1 ºC, trong
điều kiện hiếu khí. Xác định đường kính vùng ức chế (mm) của mẫu nghiên
cứu và đối chứng, từ đó đánh giá hoạt tính kháng khuẩn của các petide
mastoparan. Các thí nghiệm được thực hiện lặp lại 3 lần.
2.3.6 Xử lý số liệu
Các số liệu được xử lí bằng Excel và được trình bày dưới dạng giá trị
trung bình ± sai số chuẩn

35
CHƯƠNG 3. KẾT QUẢ VÀ THẢO LUẬN
3.1 THU THẬP VÀ TÁCH CHIẾT NỌC ONG Vespa velutina
3.1.1 Thu thập ong Vespa velutina
Các cá thể ong Vespa velutina đã được thu thập và phân loại dựa vào đặc
điểm hình thái học. Ong sau khi được thu thập được đông lạnh và bảo quản ở
tủ - 20 oC cho các thí nghiệm tiếp theo. Các ong Vespa velutina cái có phần
ngòi độc được lộ rõ ở phần cuối bụng (Hình 3.1) và chỉ có những con ong cái
mới có ngòi độc bởi vì ngòi độc có liên quan đến cơ quan sinh sản của ong cái.
Do đó chỉ những con ong Vespa velutina cái được thu nhận để tách chiết nọc
ong.
Hinh 3.1 Ong Vespa velutina cái

36
3.1.2 Tách chiết nọc ong Vespa velutina
Các ong Vespa velutina cái được giải phẫu để thu nhận cơ quan nọc ong bao
gồm: ngòi độc và bầu độc (túi độc) (Hình 3.2)
Các cơ quan nọc ong sau khi được giải phẫu, được cắt nhỏ và chiết xuất
protein/peptide bằng dung dịch đệm ACN 50% trong 3 giờ ở 4 ºC. Sau đó ly
tâm 13000/phút trong 30 phút, thu dịch nổi và làm khô.
3.2 PHÂN ĐOẠN NỌC ONG VÀ TINH SẠCH MASTOPARAN
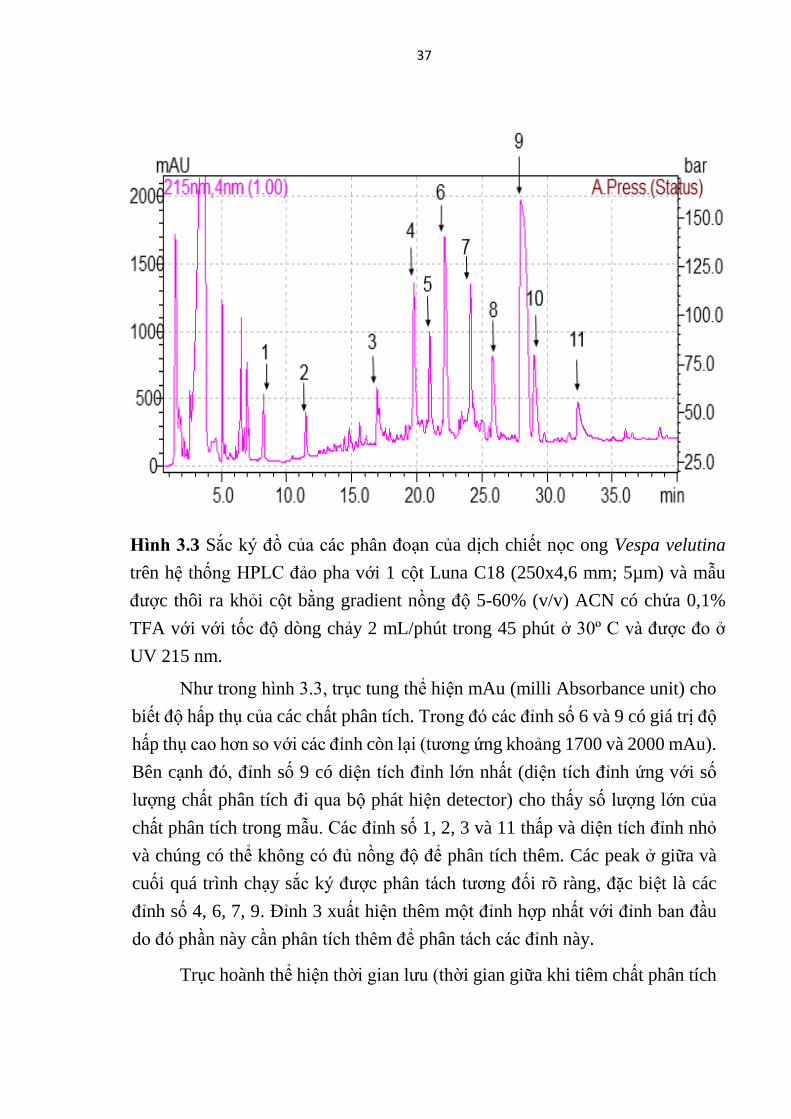
Mastoparan được tinh sạch bằng sắc ký HPLC đảo pha và mẫu được thôi
bằng gradient nồng độ tuyến tính từ 5-60% (v/v) ACN có chứa 0.1 % (v/v) TFA
với tốc độ dòng 2 mL/phút trong 45 phút và thu các phân đoạn. Cường độ mẫu
được đo ở UV 215 nm. Thông tin rửa giải được hiển thị trên sắc ký đồ (Hình
3.3).
Từ hình ảnh sắc ký đồ (Hình 3.3) cho thấy các tính hiệu tạo thành các
peak (đỉnh) khác nhau, mỗi peak đại diện cho một protein/peptide trong mẫu đi
qua cột hấp phụ và hấp phụ tia UV 215 nm. Chúng tôi đã tiến hành thu 11 phân
đoạn rửa giải tương ứng với các peak được đánh số lần lượt theo thứ tự từ 1 tới
11 như trong Hình 3.3.
Hinh 3.2 Cơ quan nọc độc của ong Vespa velutina
sau khi được giải phẫu.
37
Như trong hình 3.3, trục tung thể hiện mAu (milli Absorbance unit) cho
biết độ hấp thụ của các chất phân tích. Trong đó các đỉnh số 6 và 9 có giá trị độ
hấp thụ cao hơn so với các đỉnh còn lại (tương ứng khoảng 1700 và 2000 mAu).
Bên cạnh đó, đỉnh số 9 có diện tích đỉnh lớn nhất (diện tích đỉnh ứng với số
lượng chất phân tích đi qua bộ phát hiện detector) cho thấy số lượng lớn của
chất phân tích trong mẫu. Các đỉnh số 1, 2, 3 và 11 thấp và diện tích đỉnh nhỏ
và chúng có thể không có đủ nồng độ để phân tích thêm. Các peak ở giữa và
cuối quá trình chạy sắc ký được phân tách tương đối rõ ràng, đặc biệt là các
đỉnh số 4, 6, 7, 9. Đỉnh 3 xuất hiện thêm một đỉnh hợp nhất với đỉnh ban đầu
do đó phần này cần phân tích thêm để phân tách các đỉnh này.
Trục hoành thể hiện thời gian lưu (thời gian giữa khi tiêm chất phân tích
Hinh 3.3 Sắc ký đồ của các phân đoạn của dịch chiết nọc ong Vespa velutina
trên hệ thống HPLC đảo pha với 1 cột Luna C18 (250x4,6 mm; 5µm) và mẫu
được thôi ra khỏi cột bằng gradient nồng độ 5-60% (v/v) ACN có chứa 0,1%
TFA với với tốc độ dòng chảy 2 mL/phút trong 45 phút ở 30º C và được đo ở
UV 215 nm.

38
tới khi phát hiện chất phân tích). Người ta cũng biết rằng các đỉnh có thời gian
lưu lâu hơn có tính kỵ nước cao hơn các đỉnh có thời gian lưu ngắn hơn bởi vì
khi "đầu” kỵ nước của polypeptite hấp phụ vào bề mặt kỵ nước của vật liệu pha
đảo, nơi nó vẫn duy trì cho đến khi nồng độ dung môi hữu cơ tăng đến nồng độ
tới hạn và khử polypeptide. Mastoparan là các peptide giàu các gốc kỵ nước, vì
vậy nó có thể mất thời gian lâu hơn để khử hấp phụ. Do đó, các phân đoạn có
thời gian lưu lâu hơn có thể được kỳ vọng là peptide được quan tâm.
3.3 NHẬN DIỆN VÀ XÁC ĐỊNH TRÌNH TỰ AMINO ACID CỦA
MASTOPARAN
Mười một phân đoạn peptide đã được tinh sạch bằng sắc ký RP-HPLC
đã được phân tích, nhận diện bằng MALDI-TOF/MS. Các kết quả phổ MALDI-
TOF/MS được phân tích bằng phần mềm PEAK (Bioinformatics Solutions Inc.,
Canada) (Bảng 3.1 và Bảng 3.2).
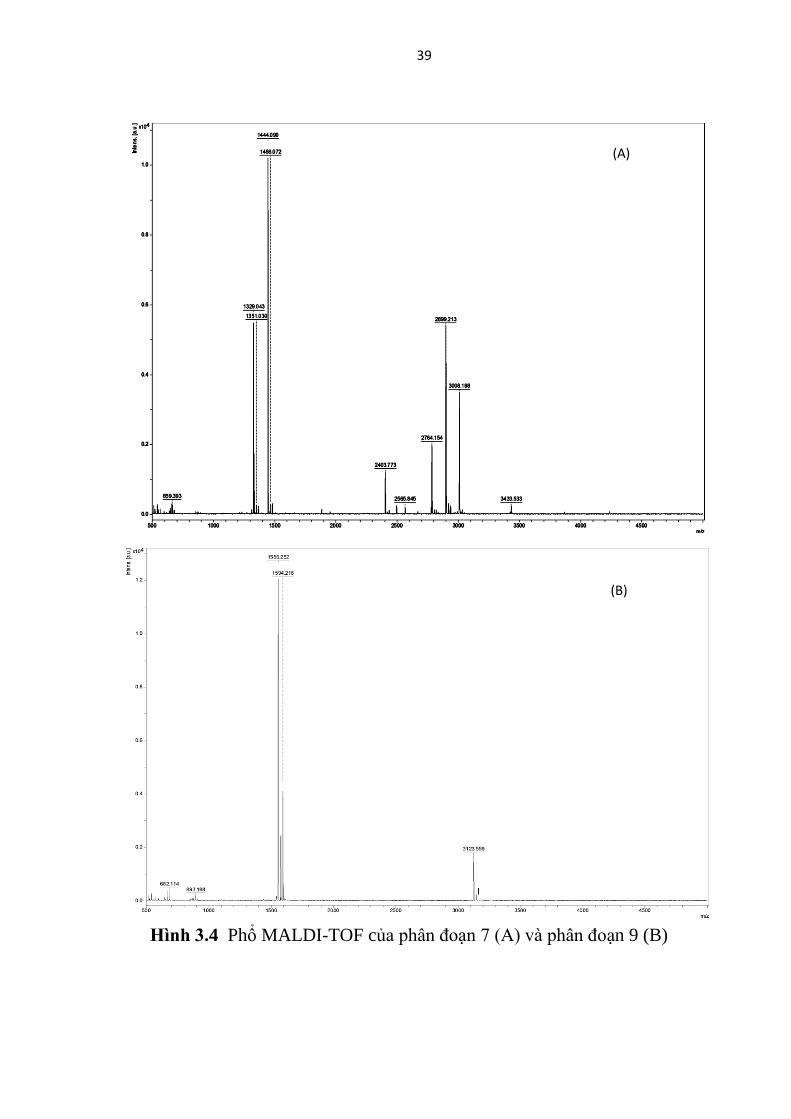
Hai phân đoạn số 7 và phân đoạn số 9 với khối lượng phân tử xấp xỉ
1444.93 và 1557.03 Da (Hình 3.4) được nhận dạng là Mastoparan-X
(mastoparan được tách chiết từ loài ong Vespa simillima xanthoptera) hoặc
Mastoparan-like peptide 12a (mastoparan được tách chiết từ loài ong Vespa
magnifica).
39
Hinh 3.4 Phổ MALDI-TOF của phân đoạn 7 (A) và phân đoạn 9 (B)
(A)
(B)
40
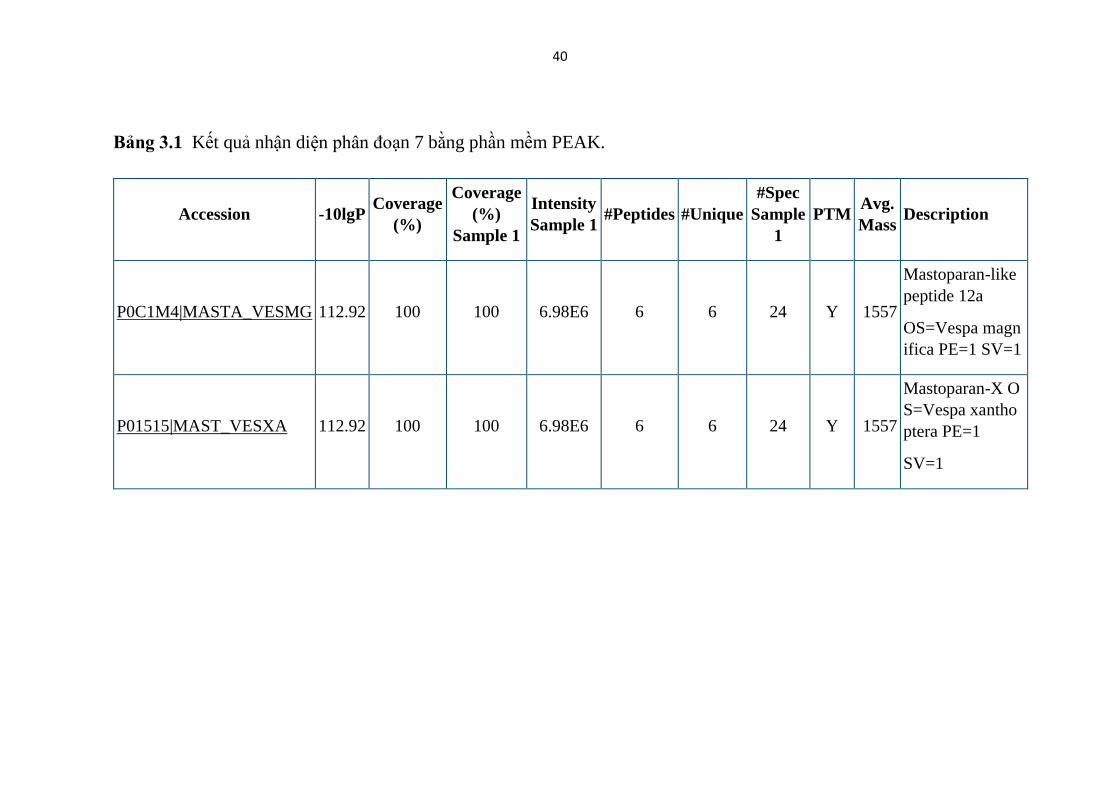
Bảng 3.1 Kết quả nhận diện phân đoạn 7 bằng phần mềm PEAK.
Accession -10lgP Coverage
(%)
Coverage
(%)
Sample 1
Intensity
Sample 1 #Peptides #Unique
#Spec
Sample
1
PTM Avg.
Mass Description
P0C1M4|MASTA_VESMG 112.92 100 100 6.98E6 6 6 24 Y 1557
Mastoparan-like
peptide 12a
OS=Vespa magn
ifica PE=1 SV=1
P01515|MAST_VESXA 112.92 100 100 6.98E6 6 6 24 Y 1557
Mastoparan-X O
S=Vespa xantho
ptera PE=1
SV=1
41
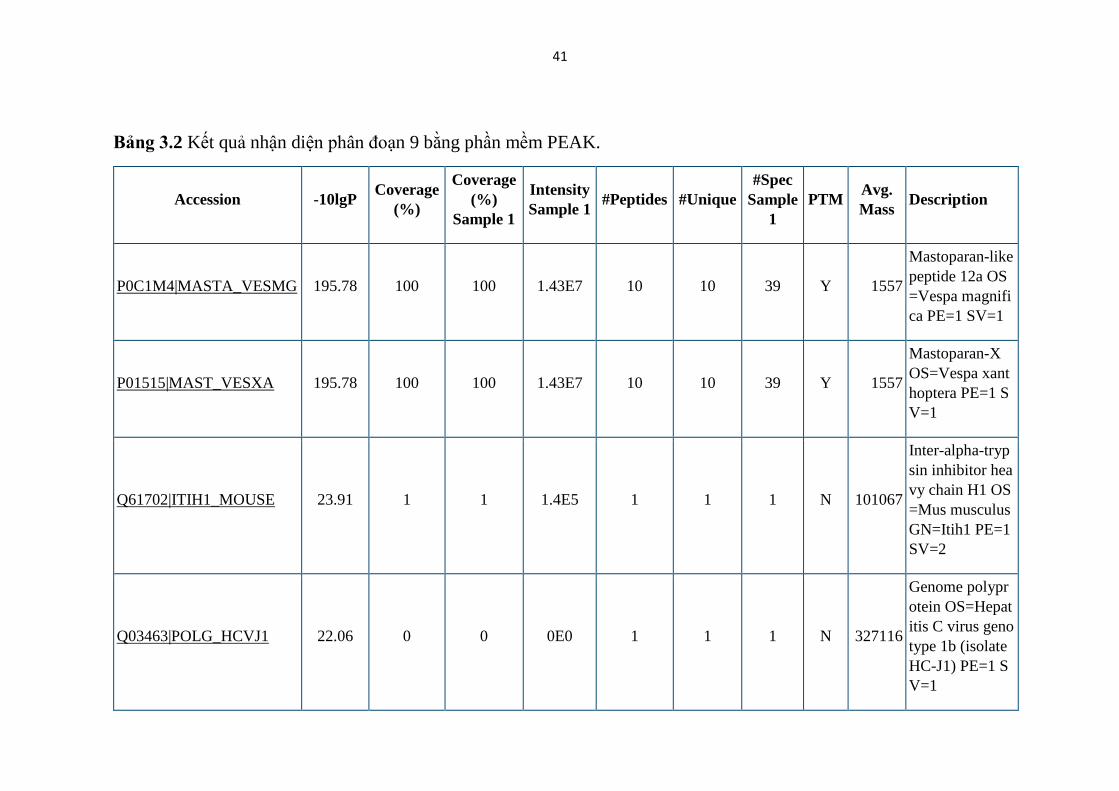
Bảng 3.2 Kết quả nhận diện phân đoạn 9 bằng phần mềm PEAK.
Accession -10lgP Coverage
(%)
Coverage
(%)
Sample 1
Intensity
Sample 1 #Peptides #Unique
#Spec
Sample
1
PTM Avg.
Mass Description
P0C1M4|MASTA_VESMG 195.78 100 100 1.43E7 10 10 39 Y 1557
Mastoparan-like
peptide 12a OS
=Vespa magnifi
ca PE=1 SV=1
P01515|MAST_VESXA 195.78 100 100 1.43E7 10 10 39 Y 1557
Mastoparan-X
OS=Vespa xant
hoptera PE=1 S
V=1
Q61702|ITIH1_MOUSE 23.91 1 1 1.4E5 1 1 1 N 101067
Inter-alpha-tryp
sin inhibitor hea
vy chain H1 OS
=Mus musculus
GN=Itih1 PE=1
SV=2
Q03463|POLG_HCVJ1 22.06 0 0 0E0 1 1 1 N 327116
Genome polypr
otein OS=Hepat
itis C virus geno
type 1b (isolate
HC-J1) PE=1 S
V=1
42
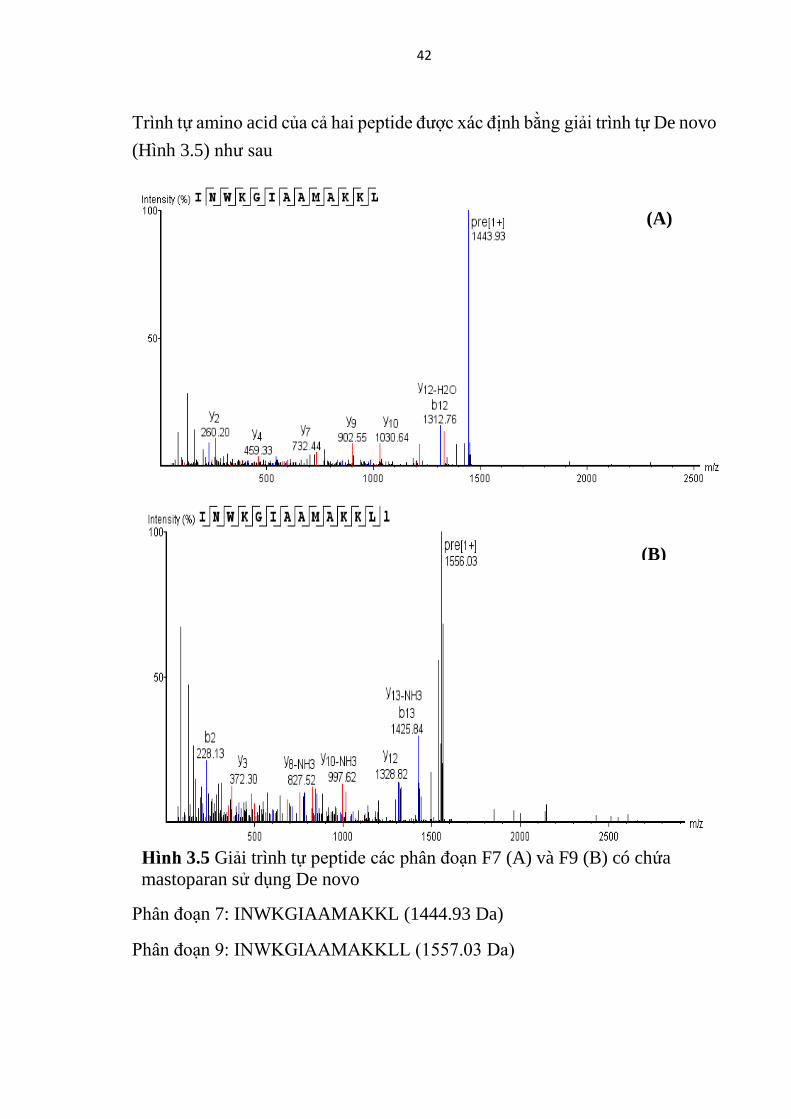
Trình tự amino acid của cả hai peptide được xác định bằng giải trình tự De novo
(Hình 3.5) như sau
Hinh 3.5 Giải trình tự peptide các phân đoạn F7 (A) và F9 (B) có chứa
mastoparan sử dụng De novo
Phân đoạn 7: INWKGIAAMAKKL (1444.93 Da)
Phân đoạn 9: INWKGIAAMAKKLL (1557.03 Da)
(A)
(B)

43
Một điều có thể dễ dàng nhận thấy rằng, phân đoạn số 7 có cấu tạo gồm
13 amino acid, phân đoạn số 9 có cấu tạo gồm 14 amino acid. Đặc biệt, phân
đoạn số 7 và phân đoạn số 9 có trình tự amino acid giống hệt nhau chỉ khác
nhau ở điểm phân đoạn số 9 có thêm một gốc L ở vị trí số 14. Có thể nói hai
phân đoạn số 7 và số 9 là các đồng phân của mastoparan (isoforms).
Dựa vào sự so sánh trình tự của F7 và F9 với các trình tự của mastoparan khác
từ các loài ong thuộc chi Vespa (Hình 3.6) cho thấy sự có mặt của các gốc
Lysine ở vị trí thứ 4, 11 và 12 ở cả hai trình tự của các peptide (F7 và F9) cũng
là một cấu trúc thường gặp đối với hầu hết tất cả các mastoparan được tách
chiết từ các loài ong thuộc chi Vespa đã được biết như Mastoparan-A,
Mastoparan-B, Mastoparan-C, Mastoparan-L, Mastoparan-M, Mastoparan-T,
Mastoparan-X, Mastoparan-like peptide 12a, Mastoparan-like peptide 12b,
Mastoparan-like peptide 12d, Mastoparan. Các gốc Lys này đã được chứng
minh là có liên quan tới quyết định hoạt tính kháng khuẩn của mastoparan.
Ngoài ra, các amino acid bao gồm Ile1, Asn2, Ile6, Leu13, Leu14 của mastoparan
từ F7 và F9 cũng có vị trí tương đồng với nhiều mastoparan thành viên từ các
loài ong thuộc chi Vespa.
Cả hai peptide từ phân đoạn số 7 và 9 đều giàu các amino acid kỵ nước
như Leucine, Isoleucine, Alanine, Methionine. Phân đoạn số 7 chứa 9 gốc trên
13 gốc amino acid trong cấu tạo phân tử còn phân đoạn số 9 chứa 10 gốc trên
Hinh 3.6 Sự so sánh trình tự của F7 và F9 với mastoparan từ các loài ong
trong chi Vespa
44
14 gốc amino acid trong cấu tạo phân tử.
3.4 TỔNG HỢP NHÂN TẠO MASTOPARAN
Sau khi mastoparan từ nọc ong Vespa velutina được nhận diện và trình
tự amino acid của mastoparan từ loài này đã được xác định. Chúng tôi đã tổng
hợp nhân tạo được peptide này được tiến hành tổng hợp cấu tạo gồm 14 amino
acid (INWKGIAAMAKKLL) và được amin hóa đầu C với khối lượng phân tử
xấp xỉ 1556 Da. Dựa vào phổ MALDI -TOF của mastoparan sau khi tổng hợp
nhân tạo (Hình 3.7), hỗn hợp thu được sau tổng hợp hóa học có chứa nhiều
peptide với các kích thước khác nhau, tuy nhiên các peptide mastoparan mong
muốn có khối lượng phân tử trung bình xấp xỉ là 1556 Da chiếm chủ yếu. Sau
đó, chúng tôi đã tiến hành tinh sạch mastoparan này bằng sắc ký RP- HPLC sử
dụng cột C18, và thu được mastoparan tổng hợp (MP) mong muốn có độ tinh
sạch đến 99 %. Với kết quả sau khi tinh sạch, chúng tôi đã tổng hợp được 10
Hinh 3.7 Phổ MALDI -TOF của MP trong hỗn hợp peptide sau khi tổng hợp

45
mg peptide đạt chuẩn cho các thử nghiệm tiếp theo.
3.5 THỬ NGHIỆM HOẠT TÍNH KHÁNG KHUẨN CỦA MASTOPARAN
3.5.1 Hoạt tính kháng khuẩn của mastoparan tự nhiên
Sau khi tinh sạch, chúng tôi đã thu được mastoparan tự nhiên (F7, F9)
với hàm lượng 2 mg có độ tinh khiết 99 %.
Kết quả thử nghiệm hoạt tính kháng khuẩn của mastoparan tự nhiên từ dịch
chiết nọc ong phân đoạn số 9 được thể hiện trong Hình 3.8 và Bảng 3.3
Hinh 3.8 Hiệu quả ức chế sự sinh trưởng của mastoparan phân đoạn số 9
trên các chủng vi khuẩn S. typhimurium (A), S. flexneri (B), E. coli (C), và
S.aureus (D). Hình (A), (B), (C), 1: Kháng sinh AMP (ampicillin 10 µg); 2;
3; 4 lần lượt là mastoparan 5 µg, 7 µg; 10 µg. Hình (D) 1: Kháng sinh OXA
(oxacillin 20 µg); 2; 3; 4 lần lượt là mastoparan 10 µg, 15 µg; 20µg
1
1 1
1
4
2
4
3
2
3 4
2
4 3
2
3
(A)
(D) (C)
(B)
46
Bảng 3.3 Hiệu quả ức chế sinh trưởng của F9 trên các chủng vi khuẩn
Đường kính vòng ức chế sinh trưởng (mm)
F9 AMP OXA
5 µg 7 µg 10 µg 15 µg 20 µg 10 µg 20 µg
S. flexneri 7,30
±
0,58
8,00
±
0,50
8,50
±
0,00
- - 21,00
±
1,00
-
S. typhimurium 7,50
±
0,87
7,80
±
1,04
9,30
±
0,76
- - 21,70
±
1,15
-
E. coli 0 6,70
±
0,29
8,70
±
2,89
- - 10,00
±
0,00
-
S. aureus 0 0 0 6,80
±
0,29
7,70
±
0,58
- 17,70
±
0,58
- : Không thử nghiệm; F9: phân đoạn số 9; AMP: ampicillin; OXA: oxacillin

47
Theo kết quả Hình 3.8 và bảng 3.3, cho thấy mastoparan từ F9 có hoạt
tính chống lại cả bốn chủng vi sinh vật thử nghiệm bao gồm S. typhimurium, S.
flexneri, E. coli và S. aureus.
F9 cho thấy khả năng kháng khuẩn phổ rộng khi chống lại cả vi khuẩn
gram âm (S. typhimurium, S. flexneri, E. coli ) và vi khuẩn gram dương (S.
aureus )
Và nó thể hiện hoạt lực kháng khuẩn mạnh nhất trên chủng S.
typhimurium với đường kính kháng khuẩn 9,3 mm ở nồng độ thử nghiệm 10
µg tiếp sau đó là S. flexneri, E. coli và hoạt lực yếu nhất trên chủng S. aureus.
Nồng độ ức chế thấp nhất đối với các chủng S. typhimurium, S. flexneri, E. coli
và S. aureus của mastoparan từ F9 được xác định lần lượt là 5; 5; 7; 15µg với
vòng ức chế sinh trưởng có đường kính tương ứng/; 7,50; 7,30; 6,70 và 6,80
mm.
So sánh hoạt tính kháng khuẩn của F9 với kháng sinh đối chứng
(ampicillin và oxacillin) cho thấy F9 tạo ta vòng kháng khuẩn nhỏ hơn.
48
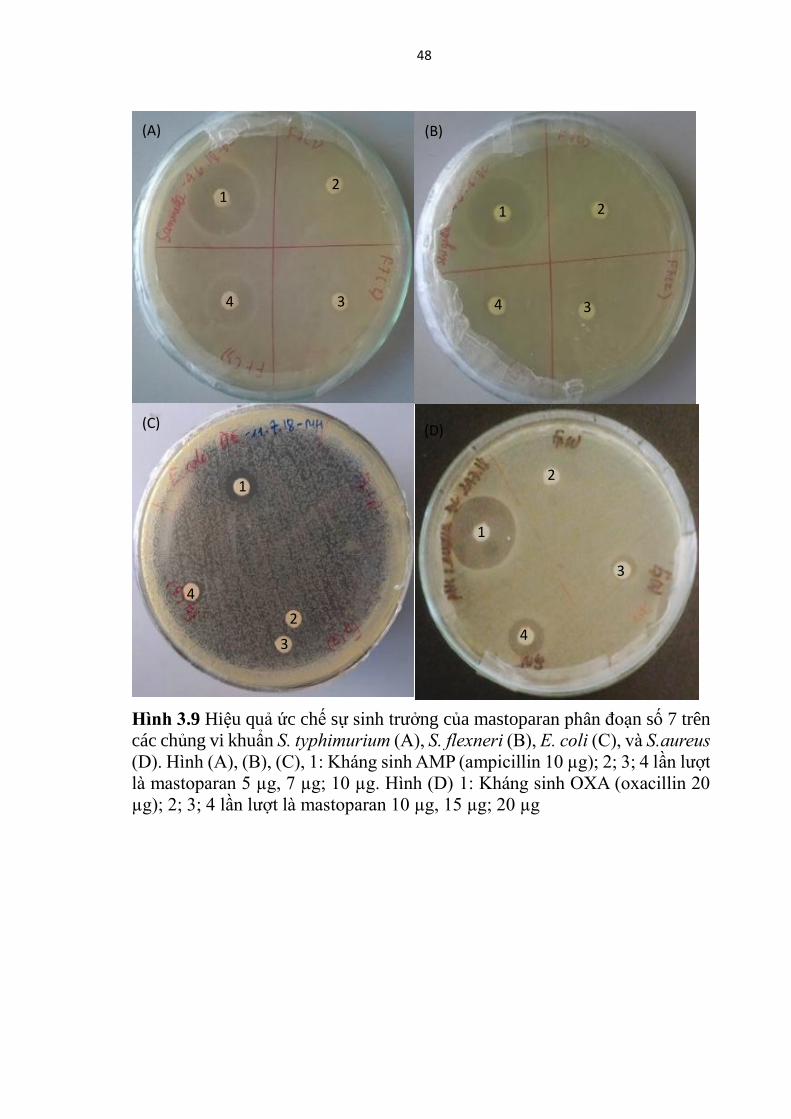
Hinh 3.9 Hiệu quả ức chế sự sinh trưởng của mastoparan phân đoạn số 7 trên
các chủng vi khuẩn S. typhimurium (A), S. flexneri (B), E. coli (C), và S.aureus
(D). Hình (A), (B), (C), 1: Kháng sinh AMP (ampicillin 10 µg); 2; 3; 4 lần lượt
là mastoparan 5 µg, 7 µg; 10 µg. Hình (D) 1: Kháng sinh OXA (oxacillin 20
µg); 2; 3; 4 lần lượt là mastoparan 10 µg, 15 µg; 20 µg
1
1
1
1
2
2
2
2
3
3
3
3 4 4
4
4
(A) (B)
(C) (D)
49
Bảng 3.4 Hiệu quả ức chế sinh trưởng của các phân đoạn F7 trên các chủng vi khuẩn.
Đường kính vòng ức chế sinh trưởng (mm)
F7 AMP OXA
5 µg 7 µg 10 µg 15 µg 20 µg 10 µg 20 µg
S. flexneri 0 8,20
±
0,58
9,00
±
1,00
- - 21,00
±
1,00
-
S. typhimurium 6,70
±
0,29
7,00
±
0,50
10,3
±
3,18
- - 22,00
±
1,73
-
E. coli 0 0 7,30
±
1,04
- - 9,50
±
0,50
-
S. aureus 0 0 0 8,30
±
0,58
9,30
±
3,21
- 18,3
±
1,15
-: không thử nghiệm; F7: phân đoạn số 7; AMP: ampicillin; OXA: oxacillin

50
Tính nhạy cảm của bốn chủng vi khuẩn: S. aureus (ATCC 25923), E.
coli (ATCC 25922), S. typhimurium (ATCC 14028) và S. flexneri (ATCC
12022) đối với mastoparan của phân đoạn 7 được thể hiện trong Hình 3.9 và
Bảng 3.4. Kết quả cho thấy rằng tất cả các vi khuẩn được thử nghiệm đều nhạy
cảm với F7
Theo kết quả Hình 3.9, và Bảng 3.4, mastoparan của F7 có khả năng ức
chế sinh trưởng tốt nhất đối với chủng S. typhimurium, sau đó là chủng
S.flexneri, tiếp đến là E. coli và cuối cùng là S. aureus. Nồng độ ức chế tối
thiểu của F7 khi thử nghiệm trên các chủng S. typhimurium, S. flexneri, E. coli
và S. aureus được xác định lần lượt là 5; 7; 10 và 15 µg /khoanh giấy với vòng
ức chế sinh trưởng có đường kính tương ứng là 6,7; 8,2; 7,3; và 8,3 mm.
Một điều có thể dễ dàng nhận thấy rằng, cả hai phân đoạn F7 và F9 đều
cho thấy hiệu quả ức chế sinh trưởng tốt nhất đối với chủng S. typhimurium và
kém hiệu quả nhất trên chủng S. aureus.
Kết quả thử nghiệm trên bốn chủng vi khuẩn chuẩn cho thấy mastoparan
từ nọc ong Vespa velutina có hoạt tính kháng khuẩn phổ rộng, chống lại cả vi
khuẩn gram âm (E. coli, S. flexneri, S. typhimurium) và vi khuẩn gram dương
(S. aureus) điều này cũng tương đồng với các nghiên cứu trước đây của Lin và
cộng sự (2011) [46]. F7 và F9 ở nồng độ 10 µg/khoanh giấy có thể gây ức chế
sinh trưởng của tất cả các vi khuẩn gram âm chuẩn được thử nghiệm bao gồm
E. coli, S. typhimurium và S. flexneri, tuy nhiên, sự phát triển của chủng vi
khuẩn gram dương S. aureus chỉ bị ức chế khi sử dụng cả hai phân đoạn ở nồng
độ thử nghiệm cao hơn - 15 µg/khoanh giấy.
Tính nhạy cảm với mastoparan của các chủng vi sinh vật thử nghiệm
khác nhau do sự khác biệt về cấu trúc màng ngoài và tính chất giữa các loài vi
khuẩn. Các mastoparan có thể thay đổi tính thấm của màng vi khuẩn theo cách
phụ thuộc vào liều lượng. Các kết quả thử nghiệm trước đây cho thấy rằng các
mastoparan có thể gây ra tổn thương nghiêm trọng cho màng tế bào vi khuẩn ở
nồng độ cao và sau đó dẫn đến chết tế bào. Có nhiều mô hình được đề xuất để
giải thích sự thấm qua màng, chẳng hạn như mô hình thùng [87], thảm [88], và
mô hình hình xuyến [87]. Tuy nhiên, cơ chế của các mastoparan tương tác với

51
màng và gây ra tính thấm qua màng vẫn chưa thực sự rõ ràng và cần được
nghiên cứu thêm. Theo kết quả của phép đo các vùng ức chế, chúng ta có thể
kết luận rằng đường kính của các vùng ức chế sinh trưởng được tạo ra tỷ lệ
thuận với sự gia tăng nồng độ thử nghiệm của cả hai phân đoạn.
Bên cạnh việc xác định tính nhạy cảm của các chủng vi khuẩn đối với
các mastoparan, chúng tôi cũng so sánh hiệu quả kháng khuẩn của mastoparan
với các thuốc kháng sinh đối chứng. Dựa vào kết quả đường kính vòng kháng
khuẩn tạo ra bởi các mastoparan và các kháng sinh đối chứng (Bảng 3.3 và
Bảng 3.4) cho thấy hiệu quả ức chế sinh trưởng của mastoparan trên các chủng
vi sinh vật thử nghiệm kém hơn so với các thuốc kháng sinh đối chứng được
sử dụng.
3.5.2 Thử nghiệm hoạt tính kháng khuẩn của mastoparan được
tổng hợp nhân tạo (MP).
Khi thử nghiệm hoạt tính kháng khuẩn của mastoparan tự nhiên trên bốn
chủng vi khuẩn không kháng kháng sinh chỉ ra rằng mastoparan có hoạt tính
kháng khuẩn phổ rộng, chống lại cả vi khuẩn gram âm và gram dương. Vì vậy
trong nghiên cứu này, để đánh giá khả năng kháng khuẩn của mastoparan từ
nọc ong Vespa velutina trên các chủng vi sinh vật kháng kháng sinh chúng tôi
đã sử dụng mastoparan tổng hợp nhân tạo cho việc thử nghiệm hoạt tính trên
các chủng vi khuẩn kháng kháng sinh bao gồm K. pneumonia, A. baumannii,
Salmonella spp., MRSE.
Kết quả cho thấy MP có khả năng chống lại các chủng vi khuẩn kháng
kháng sinh, đồng thời cũng đã thể hiện hoạt tính kháng khuẩn phổ rộng, chống
lại cả vi khuẩn gram âm (K. pneumonia, A. baumannii) và vi khuẩn gram dương
(MRSE) (Bảng 3.5, Hình 3.10) tương đồng kết quả các nghiên cứu trước đây
của Lin và cộng sự (2011) [42]. Tuy nhiên MP cho thấy không có hoạt lực
kháng vi khuẩn mạnh, thể hiện ở vòng kháng khuẩn nhỏ (7,07-12,00 mm) với
nồng độ ở mức 15-90 µg/khoanh giấy.
Nồng độ ức chế tối thiểu của MP đối với các chủng K. pneumonia, A.
baumannii, MRSE đã được xác định lần lượt là 90; 90; 15 µg /khoanh giấy với
các vòng kháng khuẩn có kích thước tương ứng là 10,00; 10,00 và 7,07 mm.

52
Với nồng độ thử nghiệm ≤ 90 µg /khoanh giấy, MP chưa thể hiện hoạt tính
chống lại chủng Salmonella spp. Điều đó cho thấy, chủng MRSE là vi khuẩn
chỉ kháng methicillin thể nhạy cảm với MP hơn các chủng vi khuẩn kháng
kháng sinh được phân lập từ lâm sàng.
Đối với chủng A. baumanii, một chủng siêu vi khuẩn kháng đa thuốc
đáng lo ngại nhất hiện nay, theo nghiên cứu của Lin và cộng sự (2017) [73]
cũng cho thấy kết quả mastoparan từ chi Vespa (Mastoparan-AF từ nọc ong
Vespa affinis) thể hiện hoạt tính kháng khuẩn mạnh ở mức 2-16 μg / mL đối
với chủng tham chiếu A. baumannii ATCC 15151 và bảy chủng MDRAB phân
lập lâm sàng, đặc biệt là MDRAB kháng colistin, E0158. Và trong nghiên cứu
này của chúng tôi, MP từ Vespa velutina cũng cho thấy hiệu quả ức chế sinh
trưởng đối với chủng A. baumannii. Điều đó càng khẳng định mastoparan có
hoạt tính kháng khuẩn đối với chủng A. baumanii kháng đa thuốc.
Bảng 3.5 Kết quả thử nghiệm xác định nồng độ ức chế tối thiểu của MP
trên các chủng vi khuẩn kháng kháng sinh
Chủng vi khuẩn Đường kính vòng kháng khuẩn theo nồng độ (mm)
22,5
µg/khoanh giấy
45
µg/khoanh giấy
90
µg/khoanh giấy
K. pneumonia 0 0 10,00±0.00
A. baumannii 0 0 10,00±0.00
Salmonella spp. 0 0 0
15
µg/khoanh giấy
30
µg/khoanh giấy
60
µg/khoanh giấy
MRSE 7,07±1,68 9,10±0,79 10,90±0.75
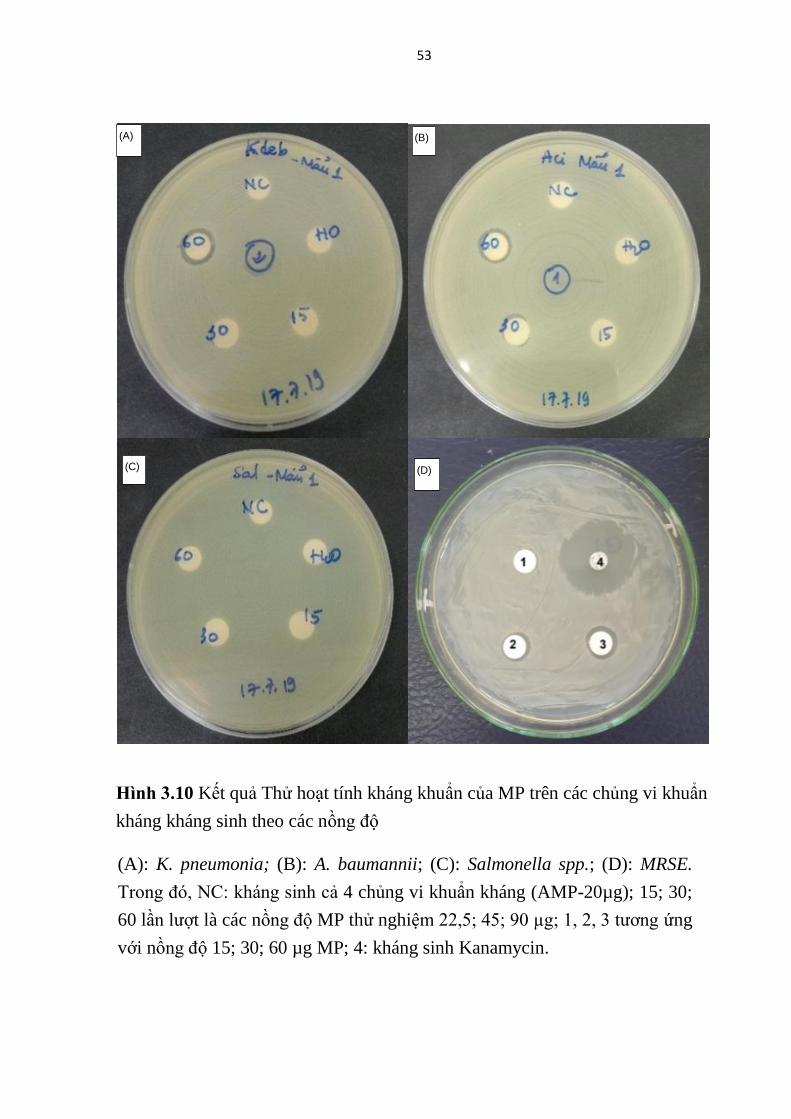
53
(A) (B)
(C) (D)
(A): K. pneumonia; (B): A. baumannii; (C): Salmonella spp.; (D): MRSE.
Trong đó, NC: kháng sinh cả 4 chủng vi khuẩn kháng (AMP-20µg); 15; 30;
60 lần lượt là các nồng độ MP thử nghiệm 22,5; 45; 90 µg; 1, 2, 3 tương ứng
với nồng độ 15; 30; 60 µg MP; 4: kháng sinh Kanamycin.
Hinh 3.10 Kết quả Thử hoạt tính kháng khuẩn của MP trên các chủng vi khuẩn
kháng kháng sinh theo các nồng độ

54
Như một số nghiên cứu của Katsu và cộng sự (1990) [55] và Nakahata
và cộng sự (1990) [56] đã đề cập, mastoparan có thể chèn vào lớp màng lipid
kép dẫn đến sự mất ổn định màng và phân huỷ tế bào, hoặc tương tác trực tiếp
với các protein G trên bề mặt tế bào chất, gây nhiễu tín hiệu màng tế bào [55-
56]. Đối với khả năng kháng khuẩn của mastoparan, kết quả thử nghiệm MP
của chúng tôi trên các chủng vi khuẩn kháng kháng sinh là khác nhau với mỗi
chủng có thể liên quan đến sự khác biệt về cấu trúc màng ngoài và tính chất
giữa các loài vi khuẩn.
Ngoài việc xác định nồng độ ức chế tối thiểu của MP trên các chủng vi
khuẩn, chúng tôi cũng đã tiến hành xác định mức độ nhạy cảm của MP và các
kháng sinh đối với các chủng vi khuẩn kháng kháng sinh (Hình 3.11, Bảng 3.6)
Chủng A. baumanii là chủng vi khuẩn kháng các loại kháng sinh như
Ampicillin, Pipercillin/tazobactam, Cefazoline, Gentamycin, Ciprofloxacin,
Levofloxacin. K. pneumoniae kháng với kháng sinh: Trimethoprim/
Sulfamethoxazone, Ampicillin, Ceftazidime. Còn Salmonella spp. chỉ kháng
với Ampicillin và MRSE kháng Methicillin. Do đó các kháng sinh được lựa
chọn làm đối chứng dương là kháng sinh mà vi khuẩn vẫn còn nhạy cảm và vẫn
đang được sử dụng chính trong điều trị, các kháng sinh làm đối chứng âm là
kháng sinh mà các vi khuẩn đó đã kháng.
Khi tiến hành thử nghiệm mức độ nhạy cảm của vi khuẩn đối với MP và
các kháng sinh, nồng độ MP 180 µg/khoanh giấy được sử dụng đối với các
chủng K. pneumonia, A. baumannii, Salmonella spp., cho thấy các vòng kháng
khuẩn với đường kính lần lượt là 12,00; 12,00 và 10,00 mm; tuy nhiên, đối với
chủng MRSE, MP với nồng độ là 60 µg/khoanh giấy đã cho thấy vòng kháng
khuẩn có đường kính 10,09 mm.
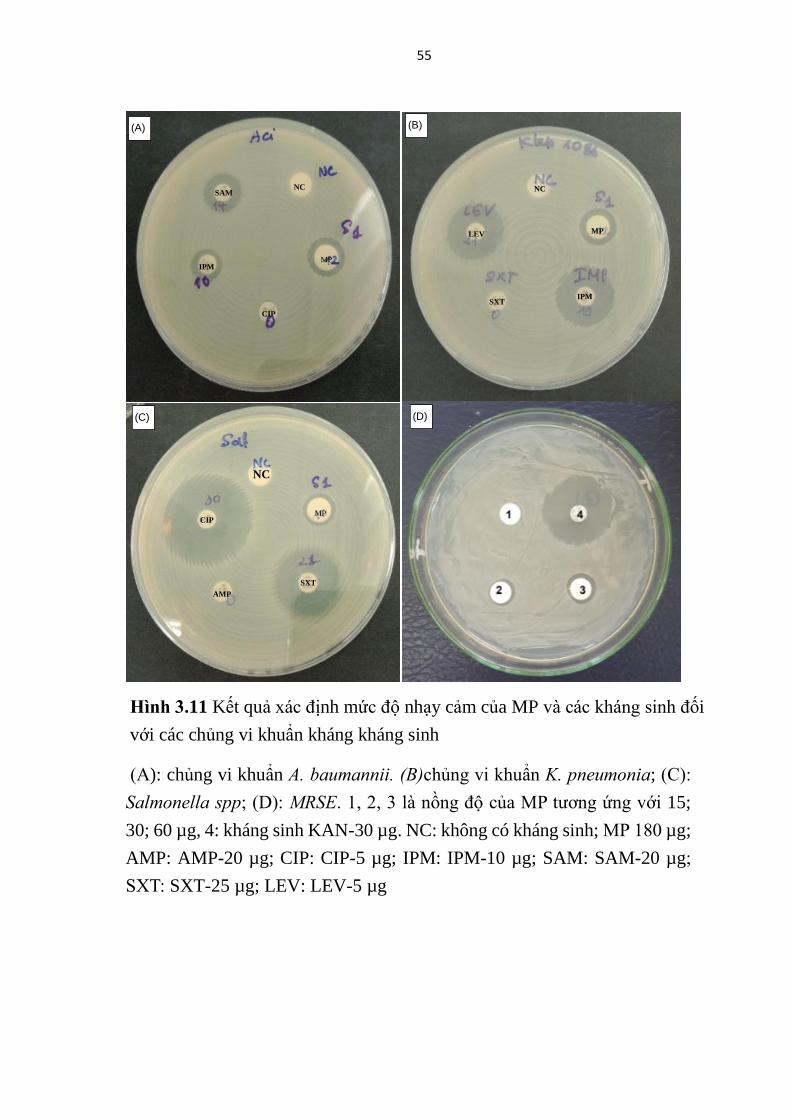
55
Hinh 3.11 Kết quả xác định mức độ nhạy cảm của MP và các kháng sinh đối
với các chủng vi khuẩn kháng kháng sinh
(A): chủng vi khuẩn A. baumannii. (B)chủng vi khuẩn K. pneumonia; (C):
Salmonella spp; (D): MRSE. 1, 2, 3 là nồng độ của MP tương ứng với 15;
30; 60 µg, 4: kháng sinh KAN-30 µg. NC: không có kháng sinh; MP 180 µg;
AMP: AMP-20 µg; CIP: CIP-5 µg; IPM: IPM-10 µg; SAM: SAM-20 µg;
SXT: SXT-25 µg; LEV: LEV-5 µg
(A) (B)
(C) (D)
NC 1 NC
NC
IPM SXT
LEV MP
MP
MP
CIP
IPM
SAM
CIP
AMP
SXT
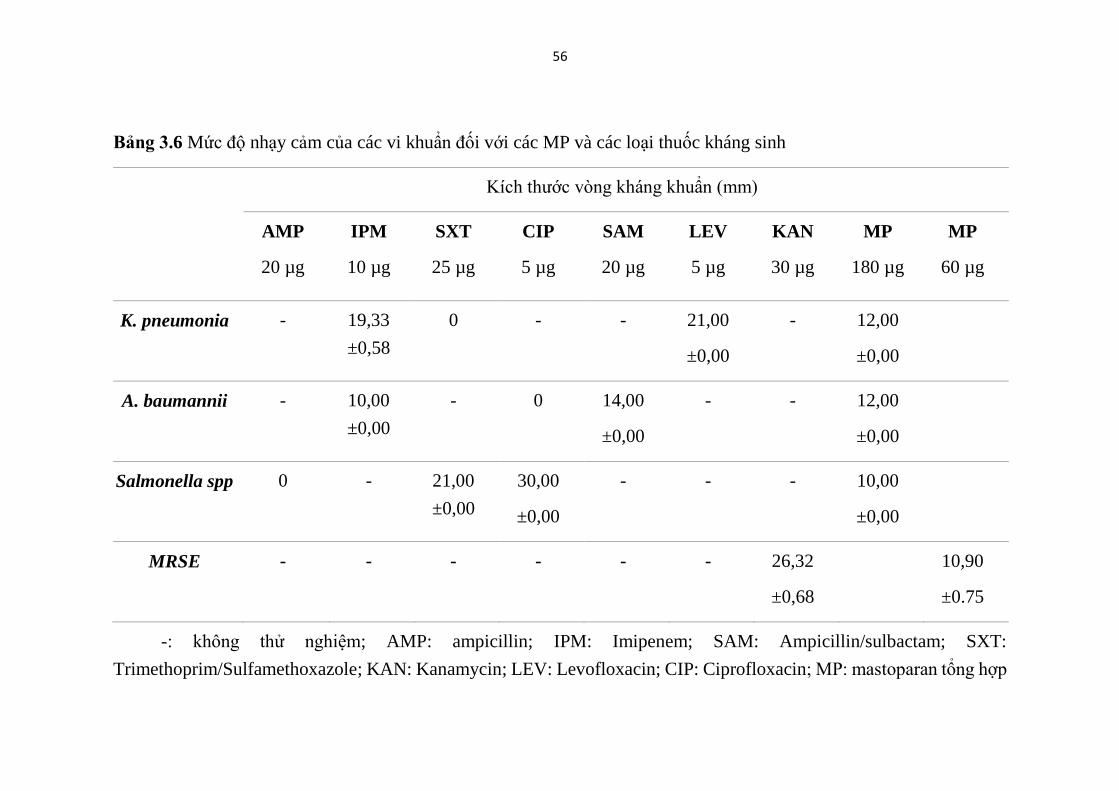
56
Bảng 3.6 Mức độ nhạy cảm của các vi khuẩn đối với các MP và các loại thuốc kháng sinh
Kích thước vòng kháng khuẩn (mm)
AMP
20 µg
IPM
10 µg
SXT
25 µg
CIP
5 µg
SAM
20 µg
LEV
5 µg
KAN
30 µg
MP
180 µg
MP
60 µg
K. pneumonia - 19,33
±0,58
0 - - 21,00
±0,00
- 12,00
±0,00
A. baumannii - 10,00
±0,00
- 0 14,00
±0,00
- - 12,00
±0,00
Salmonella spp 0 - 21,00
±0,00
30,00
±0,00
- - - 10,00
±0,00
MRSE - - - - - - 26,32
±0,68
10,90
±0.75
-: không thử nghiệm; AMP: ampicillin; IPM: Imipenem; SAM: Ampicillin/sulbactam; SXT:
Trimethoprim/Sulfamethoxazole; KAN: Kanamycin; LEV: Levofloxacin; CIP: Ciprofloxacin; MP: mastoparan tổng hợp

57
So sánh giữa MP và các kháng sinh thử nghiệm thì MP mặc dù có độ
nhạy thấp hơn các kháng sinh đã thử, tuy nhiên MP có một khối lượng phân tử
lớn (xấp xỉ 1556 Da), nên có thể cần tối ưu hàm lượng MP dùng cho thử
nghiệm.

58
CHƯƠNG 4. KẾT LUẬN VÀ KIẾN NGHỊ
4.1 KẾT LUẬN
- Tách chiết được mastoparan từ nọc ong Vespa velutina có 2 đồng phân với
các trình tự amino acid cơ bản lần lượt là: INWKGIAAMAKKL (1444,93 Da)
và INWKGIAAMAKKLL (1557,03 Da).
- Mastoparan tự nhiên được tách chiết từ nọc ong Vespa velutina có hàm lượng
2 mg với độ tinh khiết 99 %. Chúng đã cho thấy hiệu quả kháng khuẩn trên các
chủng vi khuẩn bao gồm E. coli, S. flexneri, S. typhimurium, S. aureus.
- Mastoparan được tổng hợp nhân tạo với trình tự INWKGIAAMAKKLL có
hàm lượng 10 mg với độ tinh khiết 99 %. MP cho thấy hoạt tính kháng vi khuẩn
trên các chủng vi khuẩn kháng kháng sinh bao gồm: Salmonella spp, K.
pneumonia, A. baumannii, MRSE.
4.2 KIẾN NGHỊ
- Tiến hành thử nghiệm hoạt tính kháng vi sinh vật của mastopraran từ nọc ong
Vespa velutina thêm trên các chủng vi sinh vật khác như nấm
- Nghiên cứu hoạt tính kháng khuẩn của mastoparan khi kết hợp với kháng sinh
khác.
- Thử nghiệm các hoạt tính khác của mastoparan từ nọc ong Vespa velutina
như hoạt tính chống ung thư.

59
TÀI LIỆU THAM KHẢO
1. Monceau K., Bonnard O. and Thiéry D., 2014, Vespa velutina: a new
invasive predator of honeybees in Europe, Journal of Pest Science, 87(1), pp.1-
16.
2. Robinet C., Suppo C. and Darrouzet E., 2017, Rapid spread of the invasive
yellow‐legged hornet in F rance: the role of human‐mediated dispersal and the
effects of control measures, Journal of Applied Ecology, 54(1), pp.205-215.
3. Kishi S. and Goka K., 2017, Review of the invasive yellow-legged hornet,
Vespa velutina nigrithorax (Hymenoptera: Vespidae), in Japan and its possible
chemical control, Applied Entomology and Zoology, 52(3), pp.361-368.
4. Bertolino S., Lioy S., Laurino D., Manino A. and Porporato M., 2016, Spread
of the invasive yellow-legged hornet Vespa velutina (Hymenoptera: Vespidae)
in Italy, Applied entomology and zoology, 51(4), pp.589-597.
5. Budge G.E., Hodgetts J., Jones E.P., Ostojá-Starzewski,J.C., Hall J.,
Tomkies V., Semmence N., Brown M., Wakefield M. and Stainton K., 2017,
The invasion, provenance and diversity of Vespa velutina Lepeletier
(Hymenoptera: Vespidae) in Great Britain, PLoS One, 12(9), p.e0185172.
6. Leza M., Miranda M.Á. and Colomar V., 2018, First detection of Vespa
velutina nigrithorax (Hymenoptera: Vespidae) in the Balearic Islands (Western
Mediterranean): a challenging study case, Biological Invasions, 20(7),
pp.1643-1649.
7. Husemann M., Sterr A., Maack S. and Abraham R., 2020, The northernmost
record of the Asian hornet Vespa velutina nigrithorax (Hymenoptera,
Vespidae), Evolutionary Systematics, 4, p.1.
8. Nguyen L.T., Saito F., Kojima J.I. and Carpenter J.M., 2006, Vespidae of
Viet Nam (Insecta: Hymenoptera) 2. Taxonomic Notes on
Vespinae, Zoological science, 23(1), pp.95-104.

60
9. Carpenter J.M. and Kojima J.I., 1997, Checklist of the species in the
subfamily Vespinae (Insecta: Hymenoptera: Vespidae), Natural history
bulletin of Ibaraki University, 1, pp.51-92.
10. van der Vecht J., 1957, The Vespinae of the Indo-Malayan and Papuan areas
(Hymenoptera, Vespinae), Zoologische Verhandelingen, 34, pp 1-83.
11 www.cabi.org
12 Ueno T., 2014, Establishment of the invasive hornet Vespa velutina
(Hymenoptera: Vespidae) in Japan, Int J Chem Environ Biol Sci, 2(4), pp.220-
222.
13. El-Wahed A., Yosri N., Sakr H.H., Du M., Algethami A.F., Zhao C.,
Abdelazeem A.H., Tahir H.E., Masry S.H., Abdel-Daim M.M. and Musharraf
S.G., 2021, Wasp Venom Biochemical Components and Their Potential in
Biological Applications and Nanotechnological Interventions, Toxins, 13(3),
p.206.
14. De Lima P.R. and Brochetto-Braga M.R., 2003, Hymenoptera venom
review focusing on Apis mellifera, Journal of Venomous Animals and Toxins
including Tropical Diseases, 9(2), pp.149-162.
15. Cichocka-Jarosz E., 2012, Hymenoptera venom allergy in humans, Folia
medica Cracoviensia, 52(3-4), pp. 43-60.
16. Gräler M.H. and Goetzl E.J., 2002, Lysophospholipids and their G protein-
coupled receptors in inflammation and immunity, Biochimica et Biophysica
Acta (BBA)-Molecular and Cell Biology of Lipids, 1582(1-3), pp.168-174.
17. Girish K.S. and Kemparaju K., 2007, The magic glue hyaluronan and its
eraser hyaluronidase: a biological overview, Life sciences, 80(21), pp.1921-
1943.
18. Dotimas E.M., Hamid K.R., Hider R.C. and Ragnarsson U., 1987, Isolation
and structure analysis of bee venom mast cell degranulating
peptide, Biochimica et Biophysica Acta (BBA)-Protein Structure and
Molecular Enzymology, 911(3), pp.285-293.

61
19. Ziai M.R., RUSSEK S., WANG H.C., BEER B. and BLUME A.J., 1990,
Mast cell degranulating peptide: a multi‐functional neurotoxin, Journal of
pharmacy and pharmacology, 42(7), pp.457-461.
20. Sharma J.N., 2014, Basic and clinical aspects of bradykinin receptor
antagonists, Recent Developments in the Regulation of Kinins, pp.1-14.
21. Shkenderov S. and Koburova K., 1982, Adolapin-a newly isolated analgetic
and anti-inflammatory polypeptide from bee venom, Toxicon, 20(1), pp.317-
321.
22. Moreno M. and Giralt E., 2015, Three valuable peptides from bee and wasp
venoms for therapeutic and biotechnological use: melittin, apamin and
mastoparan, Toxins, 7(4), pp.1126-1150.
23. Raghuraman H. and Chattopadhyay A., 2007, Melittin: a membrane-active
peptide with diverse functions, Bioscience reports, 27(4-5), pp.189-223.
24. Hood J.L., Jallouk A.P., Campbell N., Ratner L. and Wickline S.A., 2013,
Cytolytic nanoparticles attenuate HIV-1 infectivity, Antivir Ther, 18(1), pp.95-
103.
25. Pease J.H. and Wemmer D.E., 1988, Solution structure of apamin
determined by nuclear magnetic resonance and distance
geometry, Biochemistry, 27(22), pp.8491-8498.
26. Banks B.E.C., Brown C., Burgess G.M., Burnstock G., Claret M., Cocks
T.M. and Jenkinson D.H., 1979, Apamin blocks certain neurotransmitter-
induced increases in potassium permeability, Nature, 282(5737), pp.415-417.
27. Higashijima T., Uzu S., Nakajima T. and Ross E.M., 1988, Mastoparan, a
peptide toxin from wasp venom, mimics receptors by activating GTP-binding
regulatory proteins (G proteins), Journal of Biological Chemistry, 263(14),
pp.6491-6494.
28. Wood C.L. and Hoffman D.R., 1983, Two-dimensional polyacrylamide gel
electrophoresis of hymenoptera venom and venom sac extracts, Toxicon, 21(2),
pp.291-299.

62
29. Weinstock G.M., Robinson G.E., Gibbs R.A., Worley K.C., Evans J.D.,
Maleszka R., Robertson H.M., Weaver D.B., Beye M., Bork P. and Elsik C.G.,
2006, Insights into social insects from the genome of the honeybee Apis
mellifera, Nature, 443(7114), pp.931-949.
30. Chan Q.W., Chan M.Y., Logan M., Fang Y., Higo H. and Foster L.J., 2013,
Honey bee protein atlas at organ-level resolution, Genome research, 23(11),
pp.1951-1960.
31. Kwon Y.B., Lee J.D., Lee H.J., Han H.J., Mar W.C., Kang S.K., Beitz A.J.
and Lee J.H., 2001, Bee venom injection into an acupuncture point reduces
arthritis associated edema and nociceptive responses, Pain, 90(3), pp.271-280.
32. Park H.J., Lee S.H., Son D.J., Oh K.W., Kim K.H., Song H.S., Kim G.J.,
Oh G.T., Yoon D.Y. and Hong J.T., 2004, Antiarthritic effect of bee venom:
Inhibition of inflammation mediator generation by suppression of NF‐κB
through interaction with the p50 subunit, Arthritis & rheumatism, 50(11),
pp.3504-3515.
33. Varanda E.A. and Tavares D.C., 1998, Radioprotection: mechanisms and
radioprotective agents including honeybee venom, Journal of Venomous
Animals and Toxins, 4(1), pp.5-21.
34. Varanda E.A., Monti R. and Tavares D.C., 1999, Inhibitory effect of
propolis and bee venom on the mutagenicity of some direct‐and indirect‐acting
mutagens, Teratogenesis, carcinogenesis, and mutagenesis, 19(6), pp.403-413.
35. Nam K.W., Je K.H., Lee J.H., Han H.J., Lee H.J., Kang S.K. and Mar W.,
2003, Inhibition of COX-2 activity and proinflammatory cytokines (TNF-α and
IL-1β) production by water-soluble sub-fractionated parts from bee (Apis
mellifera) venom, Archives of pharmacal research, 26(5), pp.383-388.
36. Son D.J., Lee J.W., Lee Y.H., Song H.S., Lee C.K. and Hong J.T., 2007,
Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects
of bee venom and its constituent compounds, Pharmacology &
therapeutics, 115(2), pp.246-270.

63
37. Jalaei J., Fazeli M., Rajaian H. and Shekarforoush S.S., 2014, In vitro
antibacterial effect of wasp (Vespa orientalis) venom, Journal of Venomous
Animals and Toxins including Tropical Diseases, 20(1), pp.1-6.
38. Blondelle S.E. and Houghten R.A., 1991, Hemolytic and antimicrobial
activities of the twenty-four individual omission analogs of
melittin, Biochemistry, 30(19), pp.4671-4678.
39. Stockwell V.O. and Duffy B., 2012, Use of antibiotics in plant
agriculture, Revue Scientifique Et Technique-Office International Des
Epizooties, 31(1).
40. dos Santos-Pinto J.R.A., Perez-Riverol A., Lasa A.M. and Palma M.S.,
2018, Diversity of peptidic and proteinaceous toxins from social Hymenoptera
venoms, Toxicon, 148, pp.172-196.
41. Lee S.H., Baek J.H. and Yoon K.A., 2016, Differential properties of venom
peptides and proteins in solitary vs. social hunting wasps, Toxins, 8(2), p.32.
42. Todokoro Y., Yumen I., Fukushima K., Kang S.W., Park J.S., Kohno T.,
Wakamatsu K., Akutsu H. and Fujiwara T., 2006, Structure of tightly
membrane-bound mastoparan-X, a G-protein-activating peptide, determined
by solid-state NMR, Biophysical journal, 91(4), pp.1368-1379.
43. Hirai Y., Yasuhara, T., Yoshida H., Nakajima T., Fujino M. and Kitada C.,
1979a, A new mast cell degranulating peptide" mastoparan" in the venom of
Vespula lewisii, Chemical and Pharmaceutical Bulletin, 27(8), pp.1942-1944.
44. Hirai Y., Kuwada M., Yasuhara T., Yoshida H. and Nakajima T., 1979b, A
new mast cell degranulating peptide homologous to mastoparan in the venom
of Japanese hornet (Vespa xanthoptera), Chemical and Pharmaceutical
Bulletin, 27(8), pp.1945-1946.
45. Todokoro Y., Yumen I., Fukushima K., Kang S.W., Park J.S., Kohno T.,
Wakamatsu K., Akutsu H. and Fujiwara T., 2006, Structure of tightly
membrane-bound mastoparan-X, a G-protein-activating peptide, determined
by solid-state NMR, Biophysical journal, 91(4), pp.1368-1379.

64
46. Lin C.H., Tzen J.T., Shyu C.L., Yang M.J. and Tu W.C., 2011, Structural
and biological characterization of mastoparans in the venom of Vespa species
in Taiwan, Peptides, 32(10), pp.2027-2036.
47. Lee V.S.Y., Tu W.C., Jinn T.R., Peng C.C., Lin L.J. and Tzen J.T.C., 2007,
Molecular cloning of the precursor polypeptide of mastoparan B and its
putative processing enzyme, dipeptidyl peptidase IV, from the black‐bellied
hornet, Vespa basalis, Insect molecular biology, 16(2), pp.231-237.
48. Argiolas A. and Pisano J.J., 1984, Isolation and characterization of two new
peptides, mastoparan C and crabrolin, from the venom of the European hornet,
Vespa crabro, Journal of Biological Chemistry, 259(16), pp.10106-10111.
49. Hirai Y., Yasuhara T., Yoshida H. and Nakajima T., 1981, A new mast cell
degranulating peptide, mastoparan-M, in the venom of the hornet Vespa
mandarinia, Biomedical Research, 2(4), pp.447-449.
50. Čeřovský V., Slaninová J., Fučík V., Hulačová H., Borovičková L., Ježek
R. and Bednárová L., 2008, New potent antimicrobial peptides from the venom
of Polistinae wasps and their analogs, Peptides, 29(6), pp.992-1003.
51. Brogden K.A., 2005, Antimicrobial peptides: pore formers or metabolic
inhibitors in bacteria?, Nature reviews microbiology, 3(3), pp.238-250.
52. dos Santos Cabrera M.P., Arcisio-Miranda M., da Costa L.C., de Souza,
B.M., Costa S.T.B., Palma M.S., Neto J.R. and Procopio J., 2009, Interactions
of mast cell degranulating peptides with model membranes: a comparative
biophysical study, Archives of biochemistry and biophysics, 486(1), pp.1-11.
53. de Souza B.M., dos Santos Cabrera M.P., Neto J.R. and Palma M.S., 2011,
Investigating the effect of different positioning of lysine residues along the
peptide chain of mastoparans for their secondary structures and biological
activities, Amino Acids, 40(1), pp.77-90.
54. de Souza B.M. and Palma M.S., 2008, Monitoring the positioning of short
polycationic peptides in model lipid bilayers by combining
hydrogen/deuterium exchange and electrospray ionization mass

65
spectrometry, Biochimica et Biophysica Acta (BBA)-Biomembranes, 1778(12),
pp.2797-2805.
55. Katsu T., Kuroko M., Morikawa T., Sanchika K., Yamanaka H., Shinoda
S. and Fujita Y., 1990, Interaction of wasp venom mastoparan with
biomembranes, Biochimica et Biophysica Acta (BBA)-Biomembranes, 1027(2),
pp.185-190.
56. Nakahata N., Abe M.T., Matsuoka I. and Nakanishi H., 1990, Mastoparan
inhibits phosphoinositide hydrolysis via pertussis toxin-intensive G-protein in
human astrocytoma cells, FEBS letters, 260(1), pp.91-94.
57. Rocha T., de Souza B.M., Palma M.S. and da Cruz-Höfling M.A., 2007,
Myotoxic effects of mastoparan from Polybia paulista (Hymenoptera,
Epiponini) wasp venom in mice skeletal muscle, Toxicon, 50(5), pp.589-599.
58. Dongol Y., L Dhananjaya B., K Shrestha R. and Aryal G., 2016, Wasp
venom toxins as a potential therapeutic agent, Protein and peptide
letters, 23(8), pp.688-698.
59. Lin C.H., Hou R.F., Shyu C.L., Shia W.Y., Lin C.F. and Tu W.C., 2012, In
vitro activity of mastoparan-AF alone and in combination with clinically used
antibiotics against multiple-antibiotic-resistant Escherichia coli isolates from
animals, Peptides, 36(1), pp.114-120.
60. dos Santos Cabrera M.P., Alvares D.S., Leite N.B., Monson de Souza B.,
Palma M.S., Riske K.A. and Ruggiero Neto J., 2011, New insight into the
mechanism of action of wasp mastoparan peptides: lytic activity and clustering
observed with giant vesicles, Langmuir, 27(17), pp.10805-10813.
61. Leite N.B., Da Costa L.C., dos Santos Alvares D., dos Santos Cabrera M.P.,
de Souza B.M., Palma M.S. and Neto J.R., 2011, The effect of acidic residues
and amphipathicity on the lytic activities of mastoparan peptides studied by
fluorescence and CD spectroscopy, Amino Acids, 40(1), pp.91-100.
62. Yamamoto T., Ito M., Kageyama K., Kuwahara K., Yamashita K.,
Takiguchi Y., Kitamura S., Terada H. and Shinohara Y., 2014, Mastoparan
peptide causes mitochondrial permeability transition not by interacting with

66
specific membrane proteins but by interacting with the phospholipid phase, The
FEBS journal, 281(17), pp.3933-3944.
63. De Souza B.M., da Silva A.V.R., Resende V.M.F., Arcuri H.A., dos Santos
Cabrera M.P., Neto J.R. and Palma M.S., 2009, Characterization of two novel
polyfunctional mastoparan peptides from the venom of the social wasp Polybia
paulista, Peptides, 30(8), pp.1387-1395.
64. Dos Santos Cabrera M.P., Arcisio-Miranda M., Gorjao R., Leite N.B., de
Souza B.M., Curi R., Procopio J., Ruggiero Neto J. and Palma M.S., 2012,
Influence of the bilayer composition on the binding and membrane disrupting
effect of Polybia-MP1, an antimicrobial mastoparan peptide with leukemic T-
lymphocyte cell selectivity, Biochemistry, 51(24), pp.4898-4908.
65. Henriksen J.R. and Andresen T.L., 2011, Thermodynamic profiling of
peptide membrane interactions by isothermal titration calorimetry: a search for
pores and micelles, Biophysical journal, 101(1), pp.100-109.
66. Xu X., Li J., Lu Q., Yang H., Zhang Y. and Lai R., 2006, Two families of
antimicrobial peptides from wasp (Vespa magnifica) venom, Toxicon, 47(2),
pp.249-253.
67. Xu X., Yang H., Yu H., Li J. and Lai R., 2006, The mastoparanogen from
wasp, Peptides, 27(12), pp.3053-3057.
68. Yandek L.E., Pokorny A. and Almeida P.F., 2009, Wasp mastoparans
follow the same mechanism as the cell-penetrating peptide transportan 10,
Biochemistry, 48(30), pp.7342-7351.
69. Park N.G., Yamato Y., Lee S. and Sugihara G., 1995, Interaction of
mastoparan‐B from venom of a hornet in Taiwan with phospholipid bilayers
and its antimicrobial activity, Biopolymers: Original Research on
Biomolecules, 36(6), pp.793-801.
70. Yu H.M., Wu T.M., Chen S.T., Ho C.L., Her G.R. and Wang K.T., 1993,
Mastoparan B, synthesis and its physical and biological
properties, Biochemistry and molecular biology international, 29(2), pp.241-
246.

67
71. Chen W., Yang X., Yang X., Zhai L., Lu Z., Liu J. and Yu H., 2008,
Antimicrobial peptides from the venoms of Vespa bicolor
Fabricius, Peptides, 29(11), pp.1887-1892.
72. Yang X., Wang Y., Lee W.H. and Zhang Y., 2013, Antimicrobial peptides
from the venom gland of the social wasp Vespa tropica, Toxicon, 74, pp.151-
157.
73. Lin C.H., Lee M.C., Tzen J.T., Lee H.M., Chang S.M., Tu W.C. and Lin,
C.F., 2017, Efficacy of Mastoparan-AF alone and in combination with
clinically used antibiotics on nosocomial multidrug-resistant Acinetobacter
baumannii, Saudi journal of biological sciences, 24(5), pp.1023-1029.
74. Yoon K.A., Kim K., Nguyen, P., Seo J.B., Park Y.H., Kim K.G., Seo H.Y.,
Koh Y.H. and Lee S.H., 2015, Comparative bioactivities of mastoparans from
social hornets Vespa crabro and Vespa analis, Journal of Asia-Pacific
Entomology, 18(4), pp.825-829.
75. Li M.L., Liao R.W., Qiu J.W., Wang Z.J. and Wu T.M., 2000,
Antimicrobial activity of synthetic all-D mastoparan M, International journal
of antimicrobial agents, 13(3), pp.203-208.
76. Chen X., Zhang L., Wu Y., Wang L., Ma C., Xi X., Bininda-Emonds O.R.,
Shaw C., Chen T. and Zhou M., 2018, Evaluation of the bioactivity of a
mastoparan peptide from wasp venom and of its analogues designed through
targeted engineering, International journal of biological sciences, 14(6), p.599.
77. Etzerodt T., Henriksen J.R., Rasmussen P., Clausen M.H. and Andresen
T.L., 2011, Selective acylation enhances membrane charge sensitivity of the
antimicrobial peptide mastoparan-x, Biophysical journal, 100(2), pp.399-409.
78. Hancock R.E., 1997, The bacterial outer membrane as a drug barrier,
Trends in microbiology, 5(1), pp.37-42.
79. Meng S., Xu H. and Wang F., 2010, Research advances of antimicrobial
peptides and applications in food industry and agriculture, Current Protein and
Peptide Science, 11(4), pp.264-273.

68
80. Da Silva A.V., De Souza B.M., dos Santos Cabrera M.P., Dias N.B., Gomes
P.C., Neto J.R., Stabeli R.G. and Palma M.S., 2014, The effects of the C-
terminal amidation of mastoparans on their biological actions and interactions
with membrane-mimetic systems, Biochimica Et Biophysica Acta (BBA)-
Biomembranes, 1838(10), pp.2357-2368.
81. N. N. An, L.V. Khang, D.H. Lợi, H.N. Oanh, 1970, Một số kết quả
bước đầu nghiên cứu nọc ong trên thực nghiệm, Dược học, 3, pp15-18.
82. Cấn Văn Mão, Nguyễn Minh Núi, 2016, Nghiên cứu độc tính cấp tính bán
trường diễn của nọc ong trên động vật thực nghiệm, 2, pp 162-165.
83. Lê Hữu Thọ, Nguyễn Huy Du, Nguyễn Xuân Hải, Đỗ Văn Nhật Trường,
Nguyễn Trung Nhân, Nguyễn Thị Thanh Mai, 2015, Estabishment on
procedure for determination of apamin, phospholipaza a2 and melittin in bee
venom from Apis melifera by HPLC/UV, Tạp chí phân tích Hóa, Lý và Sinh
học, 20 (4), tr 13-19.
84.
85. https://www.ibt.ac.vn/index.php/12-cong-trinh-khoa-hoc
86. Nguyễn Tiến Dũng, Đỗ Thị Vân Anh, Nguyễn Thị Minh Phương, Bùi Thị
Huyền, Phạm Đình Minh, Đỗ Hữu Chí, Nguyễn Thị Phương Liên, Phan Văn
Chi, Lê Thị Bích Thảo, 2017, Phân tích hệ protein/peptide nọc độc ong Vespa
velutina phân lập ở Việt Nam bằng kỹ thuật proteomics, Tạp chí Công nghệ
Sinh học, 15(2), pp 1-7 .
87. Yang L., Harroun T.A., Weiss T.M., Ding L. and Huang H.W., 2001,
Barrel-stave model or toroidal model? A case study on melittin
pores, Biophysical journal, 81(3), pp.1475-1485.
88. Naito A., Nagao T., Norisada K., Mizuno T., Tuzi S. and Saitô H., 2000,
Conformation and dynamics of melittin bound to magnetically oriented lipid
bilayers by solid-state 31P and 13C NMR spectroscopy, Biophysical
Journal, 78(5), pp.2405-2417.