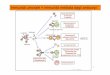
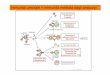
Reacii umorale si celulare cu antigenul
Reacii umorale si celulare cu antigenulRolul complementului
Rspunsul imun este rezultatul interaciunii dintre antigen si
factorii imunologiei efectori ai organismului , acetia din urm
putnd fi grupai in dou mari categorii : factori umorali (anticorpi)
i factori celulari.
Reaciile factorilor imunologici efectori cu antigenul au loc n
mod spontan in vivo, n organismul imunizat ; ele au fost ns obinute
i in vitro , condiii care au permis lrgirea cunotinelor asupra
mecanismelor imunologice i au condus la concretizarea unui mare
numr de tehnici diagnostice , prognostice , de evolutivitate etc.,
cu vast aplicativitate in clinic.Reacii umorale cu antigenul (
reacii antigen-anticorp) Prezena anticorpilor intr-un organism
poate fi constatat prin punerea in eviden a anumitor reacii
caracteristice pe care serul acestui organism le dezvolt in prezena
antigenului imunizat. n condiii similare, observarea unor reacii
imulogice caracteristice intre serul unui organism i un anumit
antigen permite precizarea unor relaii cauzale intre antigenul
respectiv ( bacterie, virus, agent sensibilizant etc. ) i starea
patologic a organismului de la care s-a prelevat serul. Reaciile
imunologice caracteristice sunt expresia combinrii anticorpilor cu
antigenul corespunztor, complexul antigen-anticorp care ia astfel
natere prezentnd proprieti diferite de acelea ale componenilor si
(anticorpii i antigenul) , ceea ce permite punerea lui n eviden.
Astfel, cnd un antigen solubil este amestecat cu serul organismului
imunizat, complexul antigen-anticorp format nu mai are acelai grad
de solubilitate ca cel al componenilor si; i, atunci cnd proporiile
de antigen si anticorp puse in contact sunt adecvate , complexul
format va precipita ; iar aspectul tulbure al amestecului care
rezult va permite constatarea c reacia dintre antigen si anticorp a
avut loc. Posibilitile de punere in eviden a complexului
antigen-anticorp format sunt foarte variate si ele sunt n mare msur
dependente de calitile componenilor ce intr n reacie i care, n
ultim instan , definesc noile proprieti ale complexului ce ia
natere. Astfel,spre exemplu, dac antigenul intrat in reacie nu este
solubil , ci corpuscular , atunci agregatele formate de complexul
antigen-anticorp vor fi deosebit de mari , lund forma unor
aglutinate. Dar, dac anticorpul este monovalent sau dac in reacie
intervine numai haptena antigenului atunci complexul format nu va
mai fi precipitalul ; n schimb , n asemenea situaii , se pot
constata modificri ale tensiunii superficiale i vscozitii
amestecului sau schimbrii ale coeficientului de sedimentare, ale
mobilitii electroforetice etc. , care traduc formarea complexului
antigen-anticorp. Mai mult , n asemenea situaii , dac antigenul
avea propriti biologice concrete (citotoxicitate, activitate
enzimatic sau biostimulatoare etc.) , dup punerea lui n contact cu
antigenul, chiar n absena formrii unui precipitat, se poate
constata c i-a pierdut proprietile biologice ( a fost neutralizat
). Exist o alt serie de reacii ntre antigen i anticorp care, pentru
a fi evideniate , necesit prezena n sistemul de reacie al unor
anumite celule, deoarece n aceste cazuri anticorpii au proprietatea
de a se fixa pe celule. Acesta este cazul anticorpilor opsonizani,
a celor anafilactizani, eliberatori de substane H i a celor
citofili pentru macrofage. Trebuie subliniat c, n aceste cazuri,
dei la reacie particip i o component celular, nu este vorba de
reacii propriu-zise ntre antigen si celul, deoarece factorul
imunologic efector este tot imunoglobulina-anticorp. n sfrit , o
categorie important de reacii antigen-anticorp, care pentru iniiere
i desfurare au nevoie de prezena complementului , constituie
substratul unui mare numr de procese imunologice , printre care
bacterioliza , bactericidia, imunoaderena i o parte important
dintre fenomenele de imobilizare i conglutinare. ntruct reaciile
dintre antigen si anticorp nu pot fi ntotdeauna direct vizualizate
, ci necesit adesea procedee indirecte de evideniere, s-a
considerat util s se descrie dou stadii in desfurarea acestor
procese : a) formarea complexului antigen-anticorp ca rezultat al
reaciei fizico-chimice dintre cei doi componeni; b) reaciile
secundare care urmeaz formrii complexului i care permit punerea lui
n eviden. Aceste dou etape nu pot fi ns net disociate n timp , iar
mecanismele care particip sunt strict dependente ntre ele . Totui,
deoarece formarea complexului antigen-anticorp este un proces
comun, coordonat de legi generale , iar manifestrile secundare sunt
variate , condiionate de caliti particulare ale componentilor (care
definesc tipul de reacie : agregare, neutralizare, anafilaxie,
opsonizare, adeziune etc.) , este preferabil s fie descrise in
prealabil mecanismele formrii complexului antigen-anticorp i
ulterior tipurile evideniabile ale reaciilor dintre anticorpi i
antigene.1. Formarea complexului antigen-anticorp
ntre moleculele de anticorp i cele de antigen corespunztor exist
o serie de regiuni complementare din punct de vedere fizico-chimic
: aa numitele situsuri combinative de pe molecula anticorpului i
determinani antigenici de pe molecula antigenului. Aceste dou zone
complementare determin reacia dintre antigen i anticorp, care n
esen este de natur stereochimic.
Modul de reacie ntre situsurile combinative i determinanii
antigenici constituie substratul specificitii acestor reacii i n
ultim analiz , substratul caracterului dominant al proceselor
imunologice efectoare. Acest mecanism de reacie este determinat de
o serie de factori, printre care natura i talia determinanilor ce
intr n reacie , energia caloric a forelor de legatur ,
reversibilitatea legturilor i implicit influena temperaturii,
pH-ului, forei ionice etc. asupra vitezei de reacie. Studiul
contribuiei fiecruia dintre aceti factori la formarea complexului
antigen-anticorp a permis formularea unei imagini de ansamblu
asupra tipurilor de legtur ce se realizeaz ntre molecula de antigen
i cea de anticorp.Aceste cercetri au avut ca punct de pornire
specificitatea foarte marcat a reaciei dintre antigen si anticorpul
su. Dou antigene care au determinani comuni pot reaciona fiecare cu
anticorpul produs de cellalt, dar reacia este mai puin intens dect
cea pe care antigenul o d in contact cu anticorpul produs de el
nsui. Cu ct dou antigene conin un numr mai mare de determinani
comuni, cu att reaciile ncruciate pe care le dau sunt mai intense;
de unde rezult c specificitatea combinrii antigenului cu anticorpul
este direct corelat cu numrul de situsuri combinative i determinani
antigenici complementari care pot intra n reacie.
Natura i talia determinanilor antigenici care intr in reacie cu
molecula de anticorp au fost studiate n special cu ajutorul
antigenelor artificiale. Asemenea studii au artat c formarea
complexului antigen-anticorp este modificat prin introducerea n
molecula de antigen a unor grupri cu talie molecular redus, ca
gruprile azofenil, naftalen, antracen, arsanil etc. O importan mare
deine tipul de legtur dintre grupare i restul moleculei de antigen;
n acest sens nca din 1936 , Mutsaars si Gregoir au evideniat o
specificitate serologic diferit ntre proteina n care au acelai
determinant este legat la nucleul lizinei prin legtur ureidic i cel
conjugat prin azo-legtur. Cercetri mai recente au artat , cu
ajutorul haptenelor DNP-polilizinice, c ndeprtarea gruprii reactive
de masa moleculei de antigen (ceea ce se obine prin creterea
numrului de molecule de lizin n gruparea haptenic) duce la alt tip
de legtur ntre antigen i anticorp i ,prin aceasta, la o modificare
a specificitii antigenului.Aceste cercetri au evideniat deci c
determinanii antigenici sunt de fapt grupri chimice cu talie
relativ mic , al caror mod de legare n molecula antigenic i a cror
poziie spaial este adesea hotartoare.Natura chimic a acestor
determinani i a legturilor din reacia lor cu anticorpii a fost n
plus atestat de influena pe care o au condiiile de pH, de
salinitate i temperatur asupra formrii complexului
antigen-anticorp. Astfel, spre exemplu, pentru formarea unui anumit
tip de complec antigen-anticorp este optim o anumit valoare a pH
ului mediului , ceea ce nseamn c , combinaia depinde de gradul de
ionizare al anumitor grupri. n acelai sens s-a constatat c exist o
concentraie optim de sruri, de obicei cea fiziologic , care asigur
o intesitate maxim a formrii complexului antigen-anticorp. O zon
optima similar ntre 15 i 40 C.
Viteza de reacie ntre antigen i anticorp s-a observat a fi
variabil n timp: astfel, dei combinarea iniial a antigenului cu
serul imun anti-protein de cal sau anti-polizaharid pneumococic are
loc practic n cateva secunde de la amestecarea preparatelor , totui
reacia nu este complet dect dup un timp mai ndelungat.Viteza de
reacie dintre antigen i anticorp este dependent de raportul dintre
antigen i anticorp, de temperatura mediului de racie , de pH-ul
mediului i de concentraia n sruri.ns ,la condiii identice, viteza
de racie variaz mult n funcie de tipul antigenului: polizaharidul
pneumococic se cupleaz 90% cu anticorpii n numai 3 secunde , n timp
ce reacia pentru antigenul Rh necesit 60 de minute. n determinarea
vitezei de reacie pot s intervin, desigur, prezena unor anticorpi
deficitari, valena antigenului etc. ; dar principala cauz o
contituie tipul de legare a antigenului cu
anticorpul.Reversibilitatea complexelor antigen-anticorp este un
alt aspect al tipului de legtur realizat ntre antigen i anticorp.
Posibilitatea de disociere a complexelor antigen-anticorp a fost
semnalat de numeroi cercettori. S-au folosit n acest scop metode
diferite , prin care adugarea unui exces de antigen constituie o
cale curent pentru solubilizarea complexului antigen-anticorp att
in vitro ct i in vivo. Acest fenomen a fost interpretat ca o
inversare a unor reacii dintre determinanii antigenici i situsurile
combinative , proces care este coordonat de legea maselor i poate
fi prelucrat matematic. n conformitate cu legea maselor, aplicat la
amestecul de antigen i anticorp, rezult ca trebuie s existe o
relaie linear ntre logaritmul raportului anticorp|antigen n
precipitat i logaritmul concentraiei componentei n exces (antigenul
sau anticorpul) aflat n supernatant.Verificarea experimental a
acestei ipoteze a artat existena relaiilor lineare prevzute prin
calculul matematic.Reversibilitatea reaciilor serologice a fost
demonstrat prin tehnici variate. Iniial, Eisen i Karush au folosit
dializa consecutiv a haptenei i anticorpului contra hapten,
constatnd un anumit echilibru, bine definit, ntre compuii
solubilizai din complexul antigen-anticorp. Ulterior, un echilibru
dinamic ntre hapter , anticorp i compuii ce conin una , dou sau
trei molecule de anticorp a fost evideniat cu ajutorul tehnicilor
combinate de dializ de echilibru, electrodializ , electroforez i
imunoelectroforez , ultracentrifugare, trasori radioactivi etc.
Aplicarea legii aciunii maselor la procesul de formare a
complexului antigen-anticorp a permis, prin determinarea
experimental a constantei de echilibru ntre complexul format i
componenii solubili , s se determine de la caz la caz gradul de
stabilitate al complexului antigen-anticorp. Aceste studii au facut
de asemenea posibil calcularea energiei libere i variaiile
entalpiei i entropiei la temperaturi diferite.
Energia de legtur ntre antigen i anticorp a fost iniial
determinat experimental de ctre Boyd, cu ajutorul unui calorimetru
sensibil. Folosind ca antigen hemocianina i anticorpi specifici de
cal, Boyd gsete c formarea complexului antigen-anticorp dezvolt
circa 3 calorii per gram de azot al antigenului; ceea ce, pentru
greutatea molecular de 6 800 000 a hemocianinei reprezint circa 3
300 kcal/mol antigen. Prin extrapolare, s-a calculat c aceast
energie corespunde la circa 40 kcal/mol de anticorp.
ntruct metoda iniial a lui Boyd utiliza un antigen cu talie
foarte mare, ceea ce contribuie la constituirea unui complex
antigen-anticorp cu un raport Atc/Atg deosebit de mare, s-au refcut
msurtorile asupra energiei libere dezvoltat in reaciile
antigen-anticorp cu ajutorul unor antigene obinuite i folosind
metode de estimare termodinamic variate: a) msurarea direct a
cldurii de reacie ; b) dialize de echilibru ; c)analiza
precipitatelor i supernatantelor ; d) observaii electroforetice i
prin ultacentrifugare; e) difuziunea luminii etc. Rezultatele
obinute au permis aprecieri multiple:a) Variaia energiei libere
care reprezint energia de legare ntre situsurile combinative ale
anticorpului i determinanii antigenici ; pentru diferite tipuri de
complexe antigen-anticorp , s-au gsit valori ale variaiei energiei
libere ntre -4,8 i 14 kcal/mol; b) Variaia entalpiei care
corespunde schimbului total de cldur din cursul reaciei;valorile
variaiei entalpiei au fost gsite ca variind ntre 0 i 9;
c) Variaia entropiei care reprezint diferena dintre entalpie i
energia liberat n reacii , n condiii termodinamice date i
corespunde nivelului de particule libere din sistem ; faptul c
pentru variaia entropiei s-au gsit valori pozitive arat c , n
sistem , dup formarea complexului antigen-anticorp , se gsesc mai
multe particule libere (ceea ce pare s fie rezultatul moleculelor
de ap provenite din unirea antigenului cu anticorpul i modificrilor
consecutive de structur teriar ).
Tipuri de legtur ntre antigen i anticorp. Rezultatele privind
natura i talia gruprilor reactive, energia de legtur, condiiile de
reversibilitate ale complexului antigen-anticorp au permis adncirea
cunotinelor asupra modului de reacie dintre moleculele de antigen i
anticorpii respectivi.
Teoretic, exist un numr mai mare de lagturi chimice care ar
putea interveni la formarea complexului antigen-anticorp. Dintre
acestea, unele sunt legturi puternice, ca cele ionice si cele
covalente; n aceste cazuri, energia de legtur este mare , de
ordinul a 50 -100 kcal/mol. Alte fore de legtur necesit energii
mult mai mici ; astfel , atracia ntre gruprile polare aprotice, de
tipul punilor de hidrogen, implic energii de a 5 kcal/mol ; atracia
de tip coulombian , ntre grupri cu sarcin opus , implic energii de
ordinul 4,5 kcal/mol; iar forele de atracie de tip van der Waals
sunt de ordinul a 0,25-1,9 kcal/mol. ntruct energia de lagtur in
complexele antigen-anticorp este de ordinul a 4-15 kcal/mol , exist
toate argumentele pentru a admite implecarea, n aceste cazuri, a
legturilor slabe. Dealtfel, n acest sens demonstreaz valorile
cldurii de reacie rezultat din combinarea anticorpilor aglutinani
anti-B cu un antigen corespunztor, care s-au dovedit egale cu 4
legturi de hidrogen sau 20 legturi van der Waals.
Forele slabe de lagtur menionate sunt eficiente numai la distane
foarte mici; ele sunt invers proporionale cu distana dintre grupele
reactive , i anume ;cu ptratul aceste distane n cazul forelor
coulombiene, cu puterea a asea n cazul punilor de hidrogen i cu
puterea a aptea n cazul forelor van der Waals. Aceasta presupune ca
distana dintre determinanii antigenici i situsurile anticorpice
care intr n reacie trebuie s fie foarte mic , ceea ce a condus la
concepia c reacia antigen-anticorp trebuie s aib un mecanism
similar cu cel al reaciilor dintre enzime i substanele lor :
anticorpul trebuie s prezinte o concavitate de grupri reactive n
care s se muleze o convexitare de grupri complementare reactive
aparinnd antigenului. Calculele fcute n aceste sens au dus la
concluzia c cele dou feluri de grupe reactive trebuie s se afle la
distane de 5-6 angstromi una de alta, iar un situs combinativ
trebuie s aib cam 34 x 12 x 7 A , ceea ce nseamna cam 2 500 -3 000
A2 ca arie total. Aceast suprafat nu depaete 1% din cea total a
unei molecule de IgG. Cum pe o molecul de anticorp exist un numr
mare de situsuri conbinative , s-ar putea considera c polivalena
acestor anticorpi este egal cu cea a locurilor de combinare. Din
punct de vedere practic ns , datorit taliei moleculei de antigen,
nu este posibil s reacioneze simultan toate situsurile combinative
ale anticorpului. ntre numrul de molecule de antigen i cele de
anticorp care se combin exist posibilitatea unor proporii variate ,
ntre o limit superioar i una inferioar, determinate de valenele
celor doi componei ce intr n reacie. Proporia relativ a fiecrui
compenent poate varia n complexul antigen-anticorp n funcie de
valena lor.2) Reacii de agregare i mecanismele de formare
Reaciile de precipitare dintre anticorp i un antigen solubil ca
i cele de aglutinare realizate de anticorp cu un antigen
particulat, sunt rezultatul unor procese de agregare. Diferena ntre
precipitarea specific i aglutinarea const n diferena de talie a
particulelor de antigen; ambele reacii constau n unirea specific a
anticorpului cu antigenul. n acest sens, Jones i Pol au observat c
particulele de colodiu, mbrcate cu albu de ou, erau aglutinate de
diluii foarte mari de ser anti-ovalbumin, chiar dac acelai ser imun
ddea reacii de precipitare cu ovalbumina n diluii mult mai mici.
Prezena determinanilor antigenici la suprafaa celulelor bacteriene,
protozoarelor , eritrocitelor etc.a fost evideniat prin contrast de
faz, microscopie electronic, anticorpi fuloresceni etc.Exist
variate procese de combinare a antigenului cu anticorpul care dau
nastere la agregate. Printre reaciile de precipitare tipice, n afar
de precipitarea i flocularea clasic , trebuie amintite : a) reacia
inelului (de precipitare) care apare la interfaza de separare a
antiserului pus n contact cu soluia de antigen ; b) imunodifuzia de
tip Ouchterlony , care const n formarea unor benzi de precipitat ca
urmare a difuziei concomitente a antigenului i anticorpilor ntr-un
gel de agar ; c) imunoelectroforeza , care este rezultatul separrii
electroforetice a serurilor urmat de dubl difuzie n gel; d)
evidenierea complexului antigen-anticorp format prin utilizarea
marcrii cu radioizotopi, metale grele etc. a antigenului sau prin
separarea lui prin ultracentrifugare, cromatografie etc.n ceea ce
privete imunoaglutinarea , tehnicile care folosesc acest procedeu
au cptat o extindere larg deoarece permit identificarea i dozarea
unor cantiti mici de anticorp (sau antigen ). La baza baza acestor
tehnici st constatarea c polizaharidele antigenice pot fi adsorbite
pe suprafaa membranei hematiilor, proteinele antigenice pot fi
absorbite de hematiile tanate , iar lipidele antigenice pot fi
absorbite pe particule inerte, cum este kaolinul, polistirenul,
colodiul etc. Hematiile sau particulele coloidale astfel pregtite
constituie antigene cu talie mult marit , care pot fi uor
vizualizate dup agregare n prezena unor mici cantiti de
anticorpi.
Tehnicile cantitative de imunoagregare au fost utilizate ca
metode analitice pentru estimarea anticorpilor intrai n reacie i
pentru analiza mecanismelor imune de agregare. Iniiate de ctre
Heidelberger, aceste tehnici analizeaz N-proteic n complexul
antigen-anticorp, dup tratarea adecvat a preparatului. Cercetri
similare, ntreprinse cu polizaharidul pneumococic tip I i serurile
specifice, au demonstrat c aglutininile i precipitinele respective
sunt identice.
Cantitile de antigen necesare pentru agregare sunt n general
deosebit de mici: cu 0,2-1 g/ml se pot obine precipitate vizibile
cu ovalbumina sau polizaharidul pneumococic, puse n prezena unor
cantiti variabile de ser imun. La diluii mari de ser imun,
precipitarea nu mai are loc. Acest proces nu este datorit lipsei
totale de anticorpi deoarece, dac se reduce cantitatea de antigen,
precipitatul se formeaz i cu diluii mari de ser imun; este vorba de
o inhibiie prin exces de antigen sau despre o solubilizare a
complexului antigen-anticorp n prezena unui exces de antigen. Un
fenomen similar, dar care se produce mai rar, este cel al inhibiiei
prin exces de anticorpi. Solubilizarea complexului antigen-anticorp
prin exces de anticorpi a fost semnalat n unele boli autoimune, n
cazul serurilor imune de cal antiprotein.Procesele de inhibiie prin
exces de antigen sau anticorpi nu sunt provocate prin blocarea
formrii complexului antigen-anticorp, ci datorit excesului unuia
dintre componeni-complecii formai conin un numr mare din
componentul ce satureaz i, astfel, agregabilitatea sistemului este
mult diminuat.
Mecanismul imunoagregrii. O molecul de imunoglobulin-anticorp
posed dou lanuri polipeptidice uoare (L) i doup grele (H), legate
ntre ele printr-o serie de legturi disulfurice. Hidroliza menajat
cu tripsin a acestor molecule duce la eliberarea a dou fragmente
Fab (t) i un fragment Fc (t), dintre care fiecare fragment Fab are
capacitatea de a se lega monovalent cu antigenul, reprezentnd deci
o grupare combinativ monovalent. Devine astfel evident c, dac o
structur imunoglobulinic incomplet de tipul fragmentului menovalent
Fab este pus n contact cu antigenul ea se va comporta bivalent,
fiecare fragment Fab reacionnd cu o molecula de antigen. n cazul
cnd o aceeai molecul de antigen este fixat-monovalent-de mai multe
molecule de anticorpi i acestea reacioneaz prin cealalt valen cu
alt molecul de antigen, se va forma o reea tridimensional de
multiple complexe antigen-anticorp. Talia acestei formaiuni de
supercompleci imuni poate fi enorm, atingnd n reaciile de floculare
volume n jur de 1 mm, vizibile cu ochiul liber. Formarea unor
asemenea supercompleci face uor de neles capacitatea lor de
agregare, spre deosebire de cazurile cnd n reacie intr anticorpi
sau antigene incomplete, care nu permit formarea de reele
tridimensionale.Teoria reelei a fost formulat iniial de Marrack i
dezvoltat de Heidelberger, Pauling i alii. Conform acestei
concepii, astzi larg verificat experimental, imunoagregarea nu
comport dect reacii specifice ntre anticorpii i antigenele
corespunztoare; influena altor factori are un substrat nespecific,
putnd accelera procesul de agregare, fr s participe direct la
formarea complecilor imuni.
Reelele de antigen i anticorpi au putut fi dealtfel vizualizate
prin microscopie electronic. Atunci cnd se folosesc antigene cu
talie mare, n imaginile electronice obinute cu aceste preparate
moleculele de anticorpi se observ ca nite puni ce leag ntre ele
structurile antigenice.
O alt posibilitatea de evideniere a reelelor de antigene i
anticorpi a fost obinut prin utilizarea trasorilor radioactivi i a
metodei autoradiografice. Astfel, Punescu si colaboratorii pun n
contact antigenele marcate cu I i anticorpii citofilipentru
macrofage i complexele obinute le pun n eviden pe cale
autoradiografic; n frotiurile finale, antigenele apar fixate pe
celule, prin anticorpii citofili. Mai multe mononucleare sunt
reinute mpreun, formnd compleci alternani de tipul
celul-anticorp-antigen-anticorp-celul. Existena acestor compleci a
fost dealtfel sugerat nc de experinele lui Pauling i colab. Care au
preparat dou seruri imune: anti-A i anti-B; fiecare ser n parte nu
precipita cu haptena corespunztoare, dar, dac cele dou haptene A i
B se cuplau i se puneau n contact simultan cu cele dou seruri, avea
loc formarea de precipitat. Acest precipitat trebuie s fie
rezultatul constituirii unor compleci de tip alternant: ser
anti-B-BA-ser anti-A-AB-ser Anti-B
3) Reacii de neutralizareAntigenele dotate cu activitate
biologic (toxine, enzime, hormoni, etc.) induc formarea de
anticorpi neutralizani, care au capacitatea de a le bloca
activitatea biologic. Reacia dintre aceste antigene i anticorpii
corespunztori poate duce uneori la formarea de precipitate; cum nsa
dozele minime biologic active din asemenea antigene sunt adesea att
de mici nct nu pot asigura, prin reacia cu anticorpul, formarea
unor precipitate vizibile, pentru punerea n evidena a reactiilor de
neutralizare se prefer teste specifice care pot demonstra blocarea
activitii biologice a antigenului cnd este pus n contact cu serul
imun.
Pentru toxine, testele de neutralizare necesit de obicei probe
in vivo, direct pe animalul de experien sau pe organe izolate.
Aceste teste se practic n dou etape: 1) realizarea in vitro de
amestecuri toxin-antitoxin (ser imun) n care toxina este pstrat n
concentraie constant (egal de obicei cu o doz limit mortal: DLM),
iar antitoxina este introdus n concentraii crescnde; 2)
administrarea amestecurilor astefl obinute la animale sensibile i
observarea mortalitii obinute n funcie de doza de antiser folosit n
amestec (cantitatea cea mai mic de ser imun care poate neutraliza
efectul unei DLM din toxina respectiv este denumit unitate
antitoxic: U.A)n prezena anticorpilor specifici, blocarea activitii
enzimatice poate fi total sau numai parial, nsoit sau nu de
formarea unor precipitate. Activitatea enzimatic a precipitatelor
imune formate este cu att mai redus cu ct valoarea raportului
anticorp/enzim este mai mare att n amestecul esenial, ct i n
complexul format. ntre neutralizarea activitii enzimatice i gradul
de precipitare al amestecului nu exist o relaie liniar: n primul
rnd, neutralizarea se poate constata nainte ca precipitatul s se
formeze; n al doilea rnd, neutralizarea este maxim n prezena unui
exces de anticorpi, spre deosebire de precipitare care este maxim n
zona de echivalen, neutralizarea se poate dealtfel obine i cu
fragmente Fab(t) monovalente ale Ig-anticorpi respectivi, care nu
produc i precipitare.Spre deosebire de imunoreaciile de
neutralizare a toxinelor, n cazul neutralizrii enzimelor reacia
poate fi inhibat de prezena substratului, care intr n competiie cu
anticorpul pentru determinanii antigenici ai centrului activ
catalitic. Acest inhibiie prin competiie are un mecanism
stereochimic, aa cum dealtfel demonstreaz faptul c n timp ce
competiia dintre anticopr i un substrat cu talie mic poate bloca
numai neutralizarea, lsnd intact capacitatea precipitant a serului
imun; dac substratul competitiv are talie mare atunci se inhib nu
numai fixarea anticorpilor neutralizani, ci i a situsurilor
combinative de precipitare.
Dac n cazul enzimelor substratul poate anihila determinantul
antigenic al centrului activ biologic, un fenomen similar poate fi
indus la toxine prin formalizare, procedeu care modific exotoxinele
bacteriene n anatoxine.
Att n cazul enzimelor, ct i al toxinelor blocarea funciei active
biologic nu poate suprima total funcia antigenic a moleculei.
Serurile preparate cu anatoxine au capacitatea antitoxic integral;
desigur, aceast afirmaie este valabil numai pentru exotoxine,
singurele care pot fi transformate n anatoxine prin formolizare.
CUPRINS1. Rolul complementului ..1
2. Reacii umorale cu antigenul 1
3. Formarea complexului antigen-anticorp ..3
4. Reacii de agregare i mecanismele de formare ..9
5. Reacii de neutralizare 12PAGE 1