Embed Size (px)
DESCRIPTION
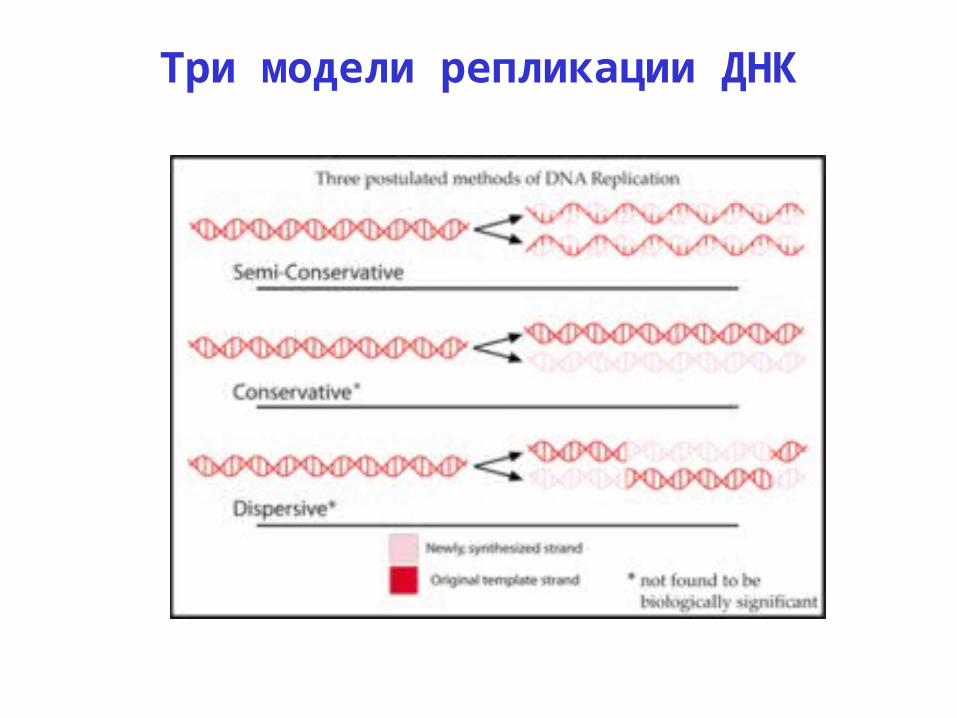
Три модели репликации ДНК. Правильная - полуконсервативная (Matthew Meselson и Franklin Stahl , 1958). Коетки растут на среде с 15 N ( тяжелый изотоп ) потом их переносят на среду с 14 N ( легкий изотоп ). Q: Что было бы для двух других моделей ?. Реплицирующаяся ДНК. - PowerPoint PPT Presentation
Citation preview
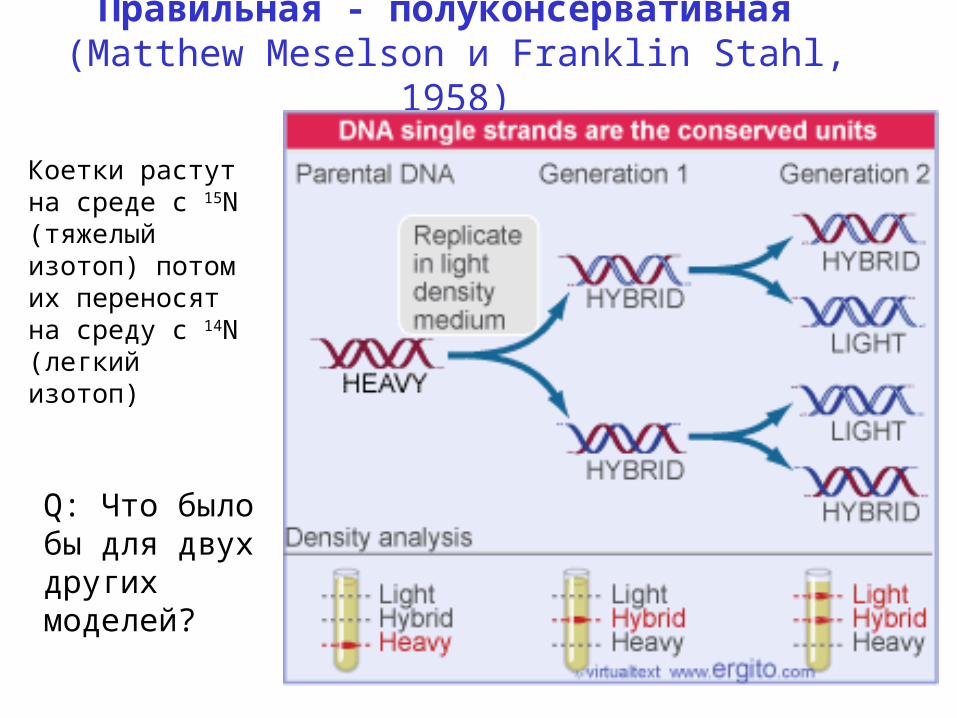
Правильная - полуконсервативная (Matthew Meselson и Franklin Stahl, 1958)
Q: Что было бы для двух других моделей?
Коетки растут на среде с 15N (тяжелый изотоп) потом их переносят на среду с 14N (легкий изотоп)

Реплицирующаяся ДНК
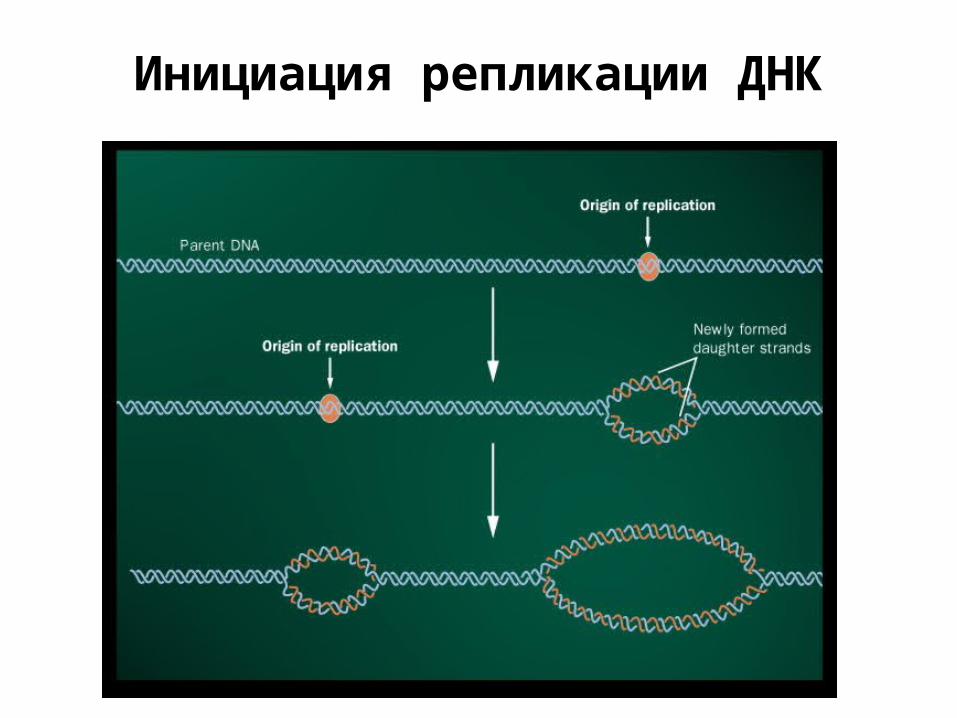
Инициация репликации ДНК
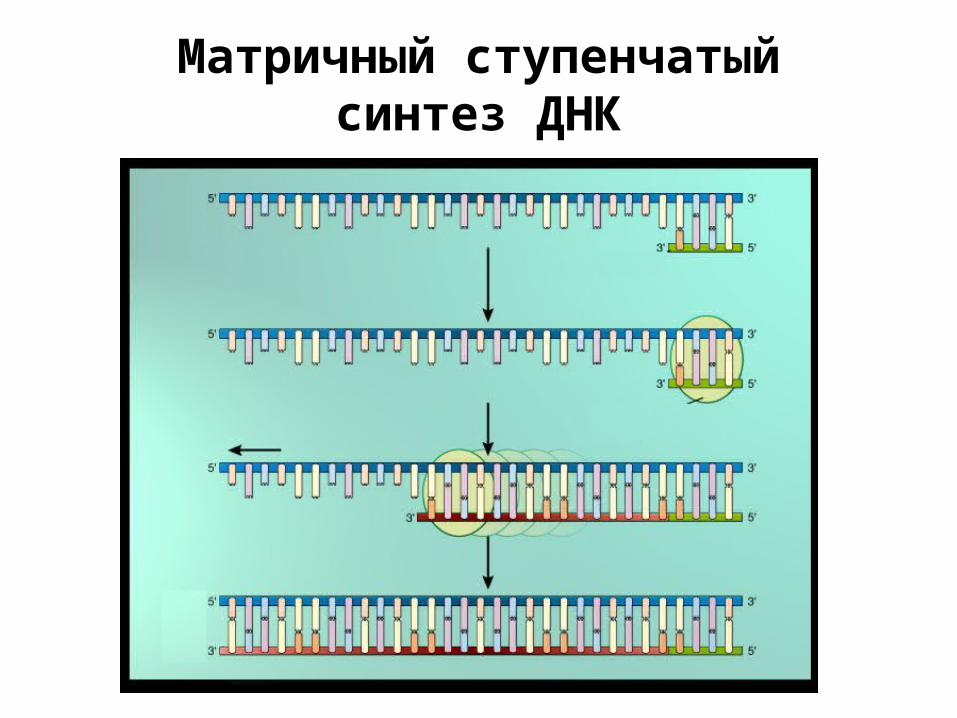
Матричный ступенчатый синтез ДНК
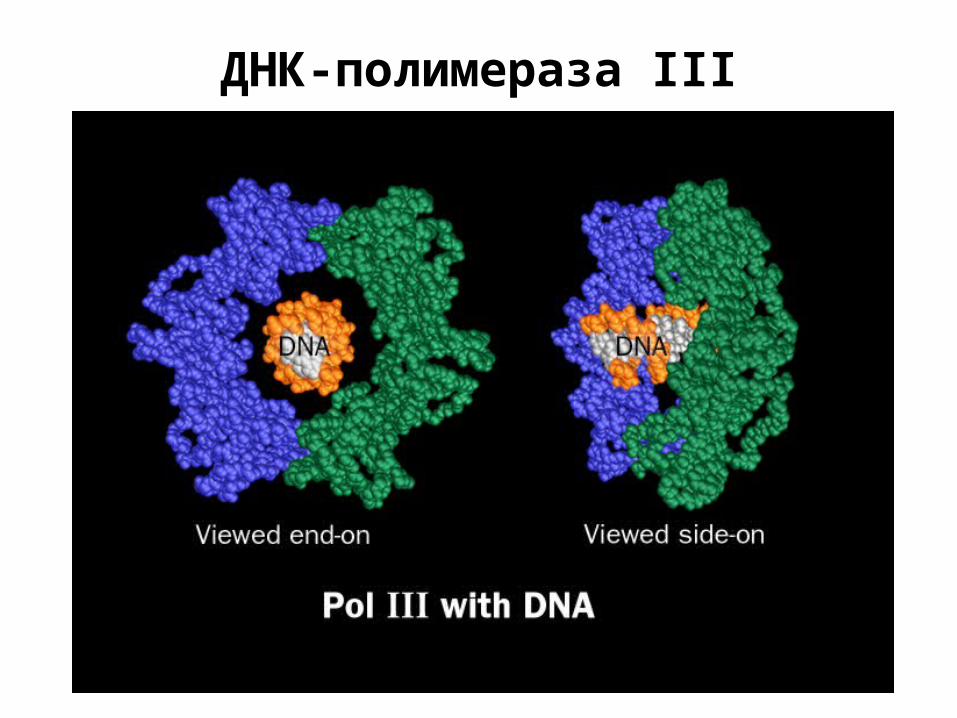
ДНК-полимераза III
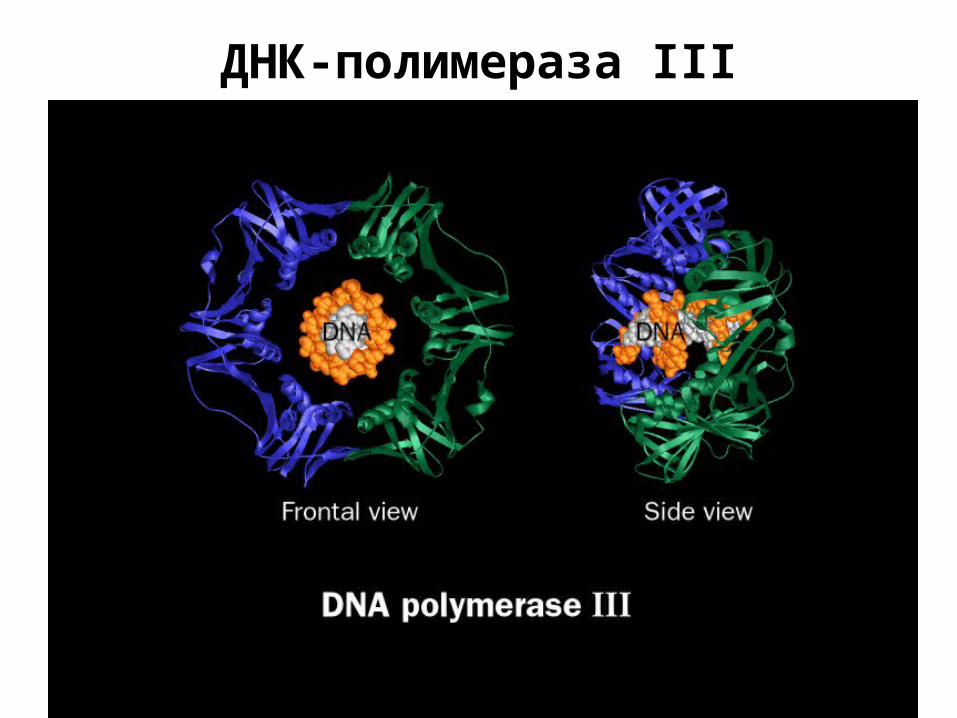
ДНК-полимераза III
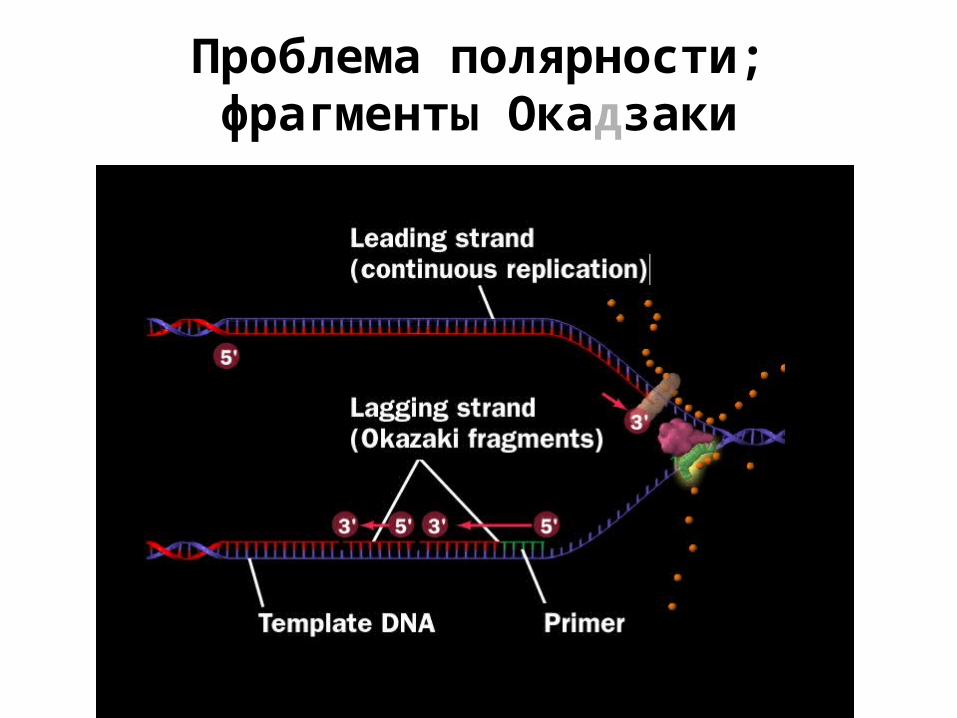
Проблема полярности; фрагменты Окадзаки
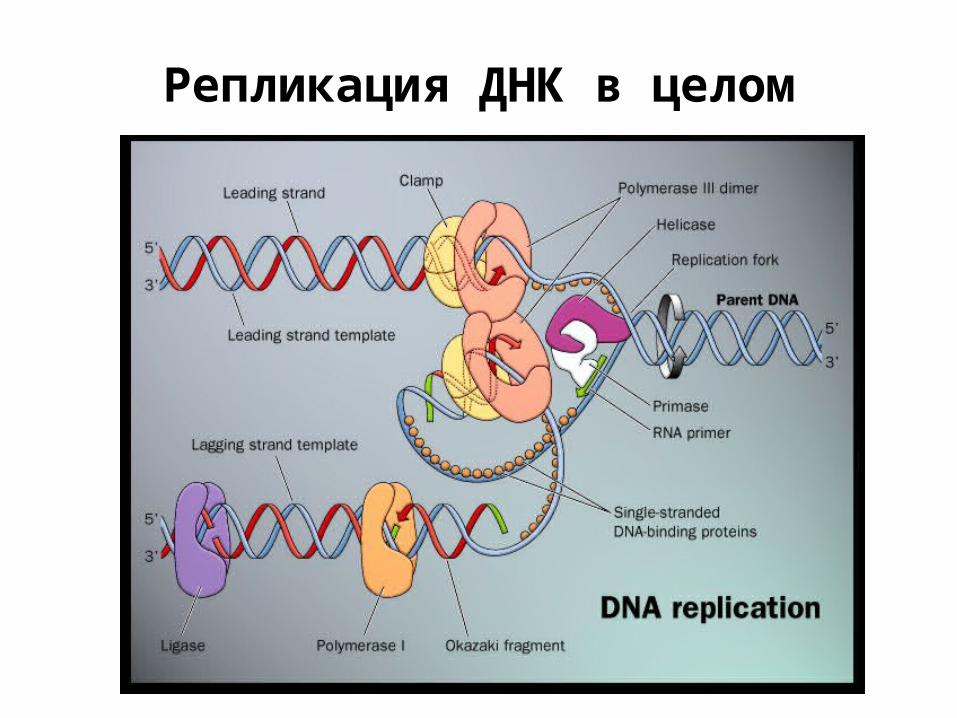
Репликация ДНК в целом
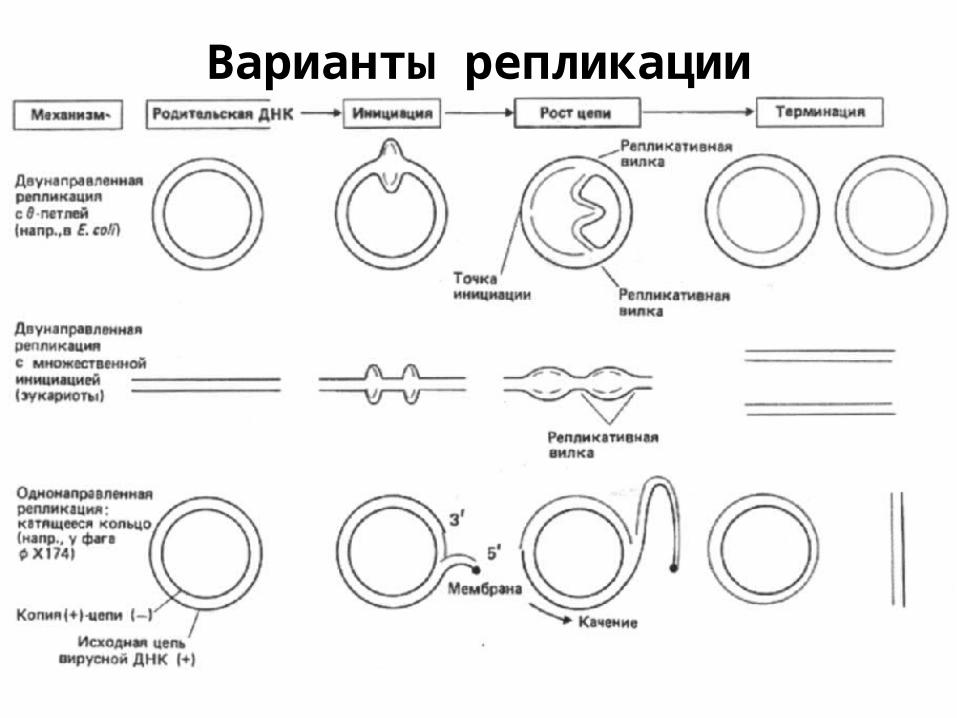
Варианты репликации
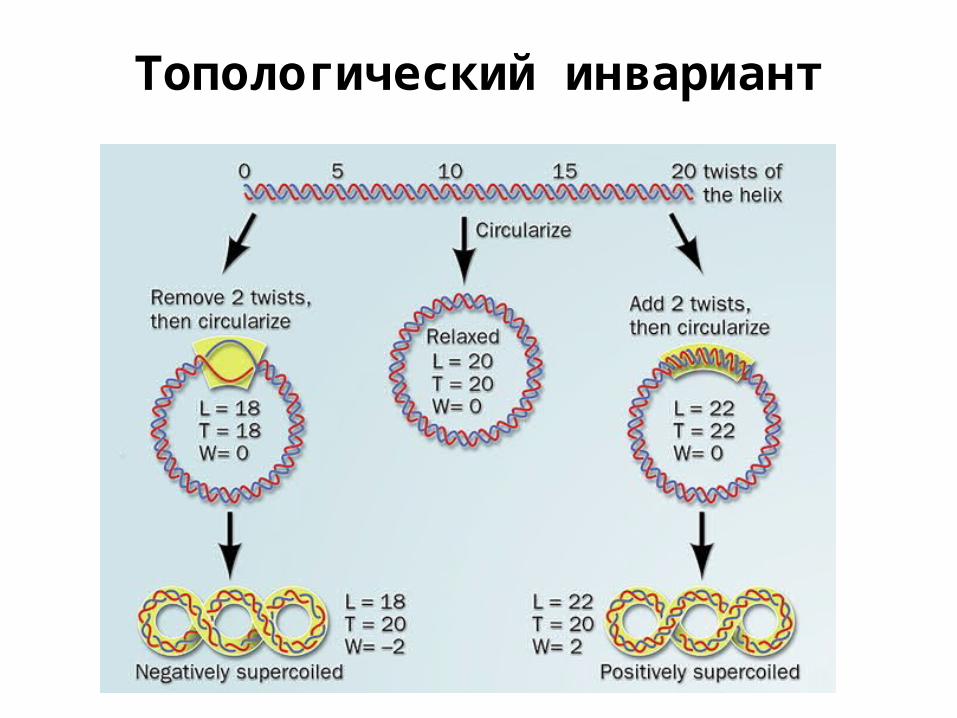
Топологический инвариант
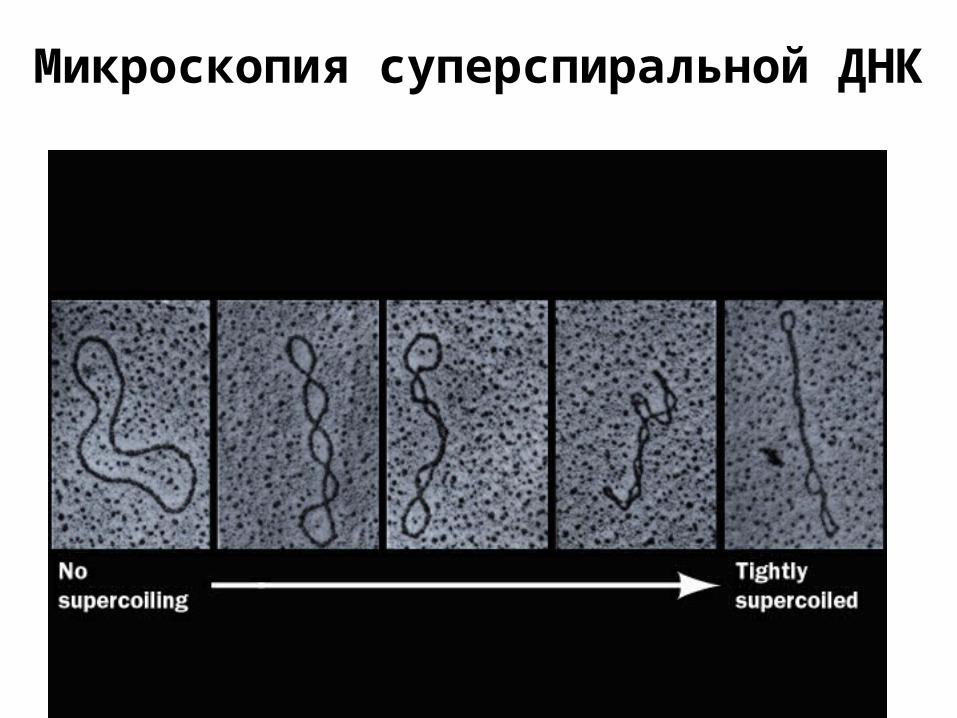
Микроскопия суперспиральной ДНК

репликация - кино
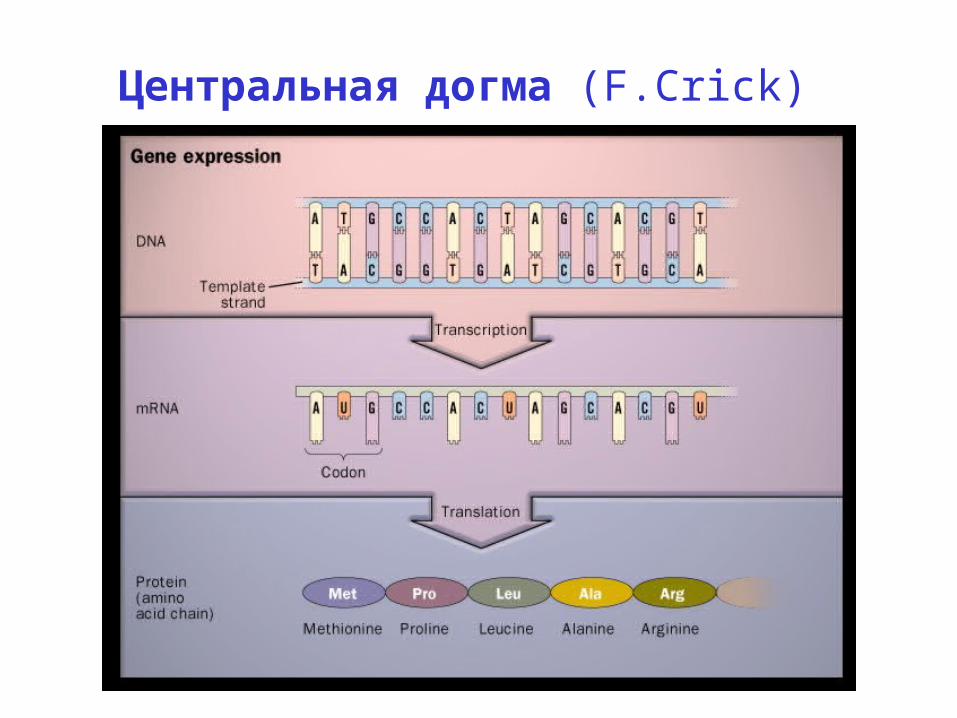
Центральная догма (F.Crick)
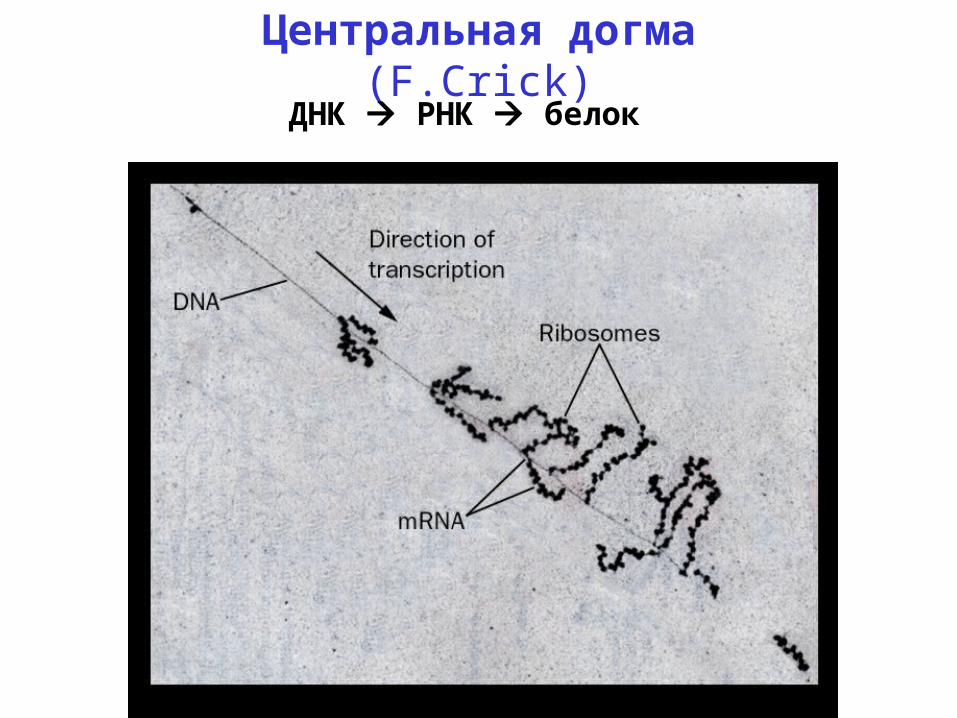
Центральная догма (F.Crick)ДНК РНК белок

РНК
• мРНК - перенос информации от ДНК к белку
• хранение генетической информации (вирусы и фаги)
• рибозимы - катализ• рРНК - структура (+ катализ)• тРНК - трансляция• регуляция
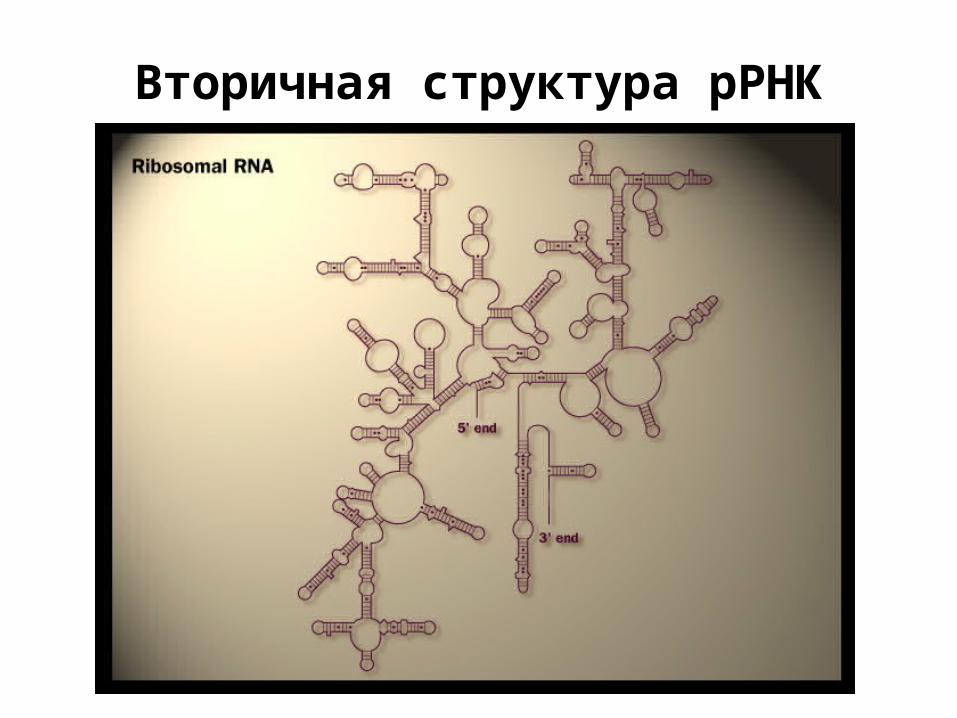
Вторичная структура рРНК
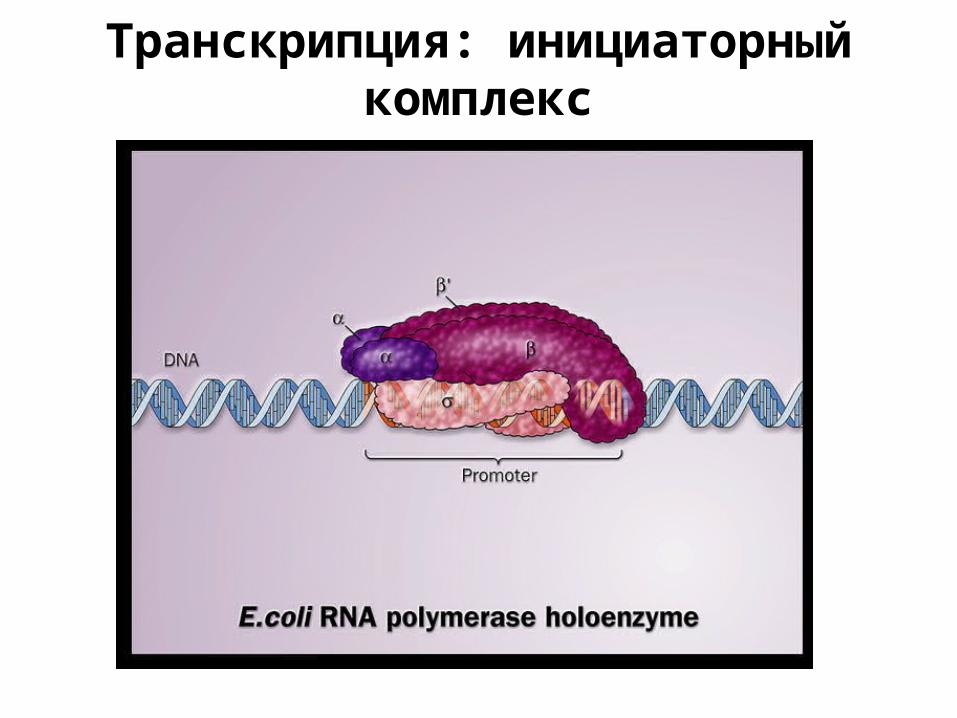
Транскрипция: инициаторный комплекс

ПромоторыrecA ttgatactgtat---gagcatacagtataatrpsL ttgacaccttttc--ggcatcgccctaaaataroF ttgaaaacttta---ctttatgtgttatcgtlpdA tttaaaaattgtt--aacaattttgtaaaatcyoA tttacagtaatgt--aaccttcccgtaaaatuvrD ttggcatctctg---acctcgctgatataatatpI ttgaaatcacgg---gggcgcaccgtataattonB ttgaatatgattgc-tatttgcatttaaaatrpsU tttacaaagcagca-gcaattgcagtaaaatcirA ttgataattgtta--tcgtttgcattatcgtfur ttttcatttaggc--gtggcaattctataatampC ttgtcacgctga---ttggtgtcgttacaatuvrB gtgatgaactgttt-ttttatccagtataatargC ttgacacacct----ctggtcatgatagtatrpmH ttgactccggagt--gtacaattattacaatrplT ttaacgttttta---actttttaattagaatgyrA tttacctcaaactgcgcggctgtgttataat
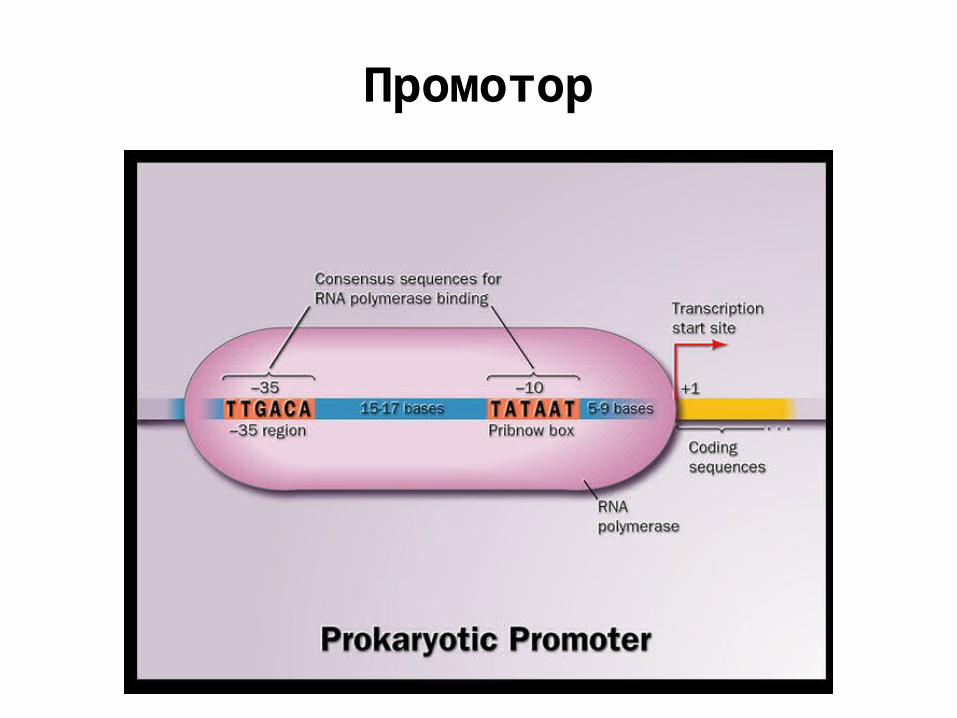
Промотор

Слабые промоторыcarB ttatcgagttaattgagcagtaccgtaaaacnagB tttactggctaaa--ccagaaaacttattttilvG ttgccagcccacgg-tcggtcgacttactgtcysK atgtcattattt---cccttctgtatatagasmp ttgcttctggca---acattaagtctcaaatamn gtgacatacta----tcggatgtgcggtaatileR atgccagcagag---ttttttatgcgttaatfabB ttgttcggcgtac--aagtgtacgctattgtdnaN atatcaaagaag---atttttcaaatttaatargC tcgataaatggcg--gtaatttgtttttcataraB ctgacgcttttt---atcgcaactctctactsulA ttgtcactggatgt-actgtacatccatacatrpR gttactgatccgc--acgtttatgatatgctdnaN ttgcaggaaaaact-ggtcaccatcgacaatcspA ttgcatcacccgc--caatgcgtggcttaatrpsU tcgcccctggaga--aagcctcgtgtatactrpoH ttcacaagcttgcattgaacttgtggataaa

Lac - промотор
gcgcccaatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagctggca
cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct
cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaat
tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct
cggtacccggggatcctctagagtcgacctgcaggcatgcaagcttggcactggccgtcg
ttttacaacgtcgtgactgggaaaaccctggcgttacccaacttaatcgccttgcagcac
atccccctttcgccagctggcgtaatagcgaagaggcccgcaccgatcgcccttcccaac
agttgcgcagcctgaatggcgaatggcgcctgatgcggtattttctccttacgcatctgt

Lac - промотор
cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct
cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggaat
tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct

Lac - промотор
cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct
cactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtggAat
tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct

Lac - промотор
cgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagttagct
cactcattaggcaccccaggcTTtACActttatgcttccggctcgTATgtTgtgtggAat
tgtgagcggataacaatttcacacaggaaacagctatgaccatgattacgaattcgagct

транскрипция - кино

Регуляция транскрипции у прокариот
• Фактор транскрипции: взаимодействие с РНК-полимеразой– репрессоры– активаторы
• Лиганд (кофактор):– способствует/препятствует
связыванию с ДНК
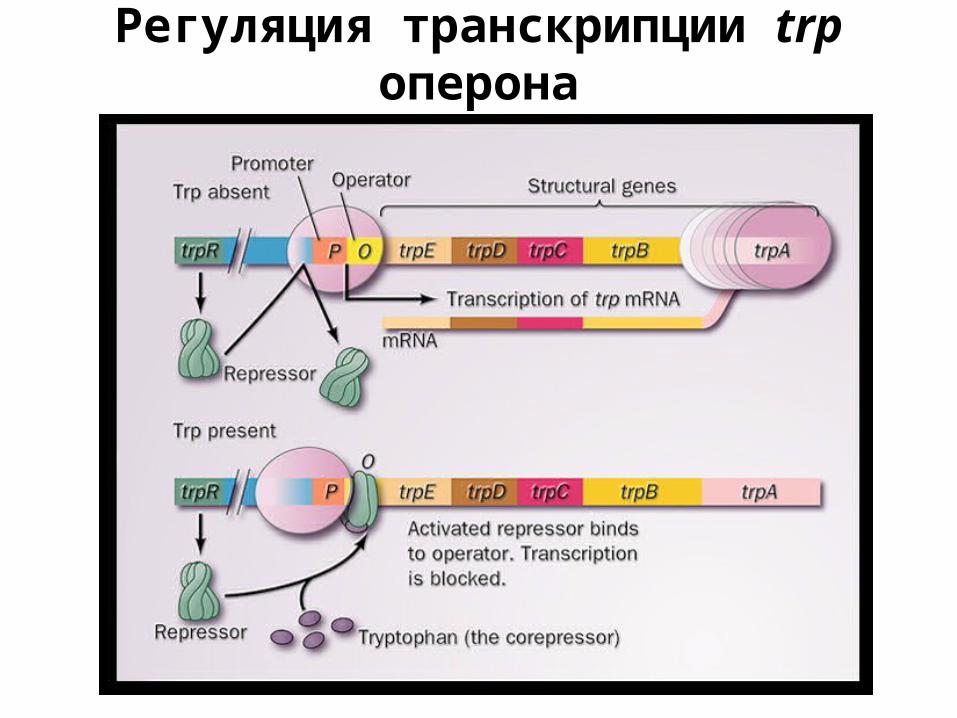
Регуляция транскрипции trp оперона
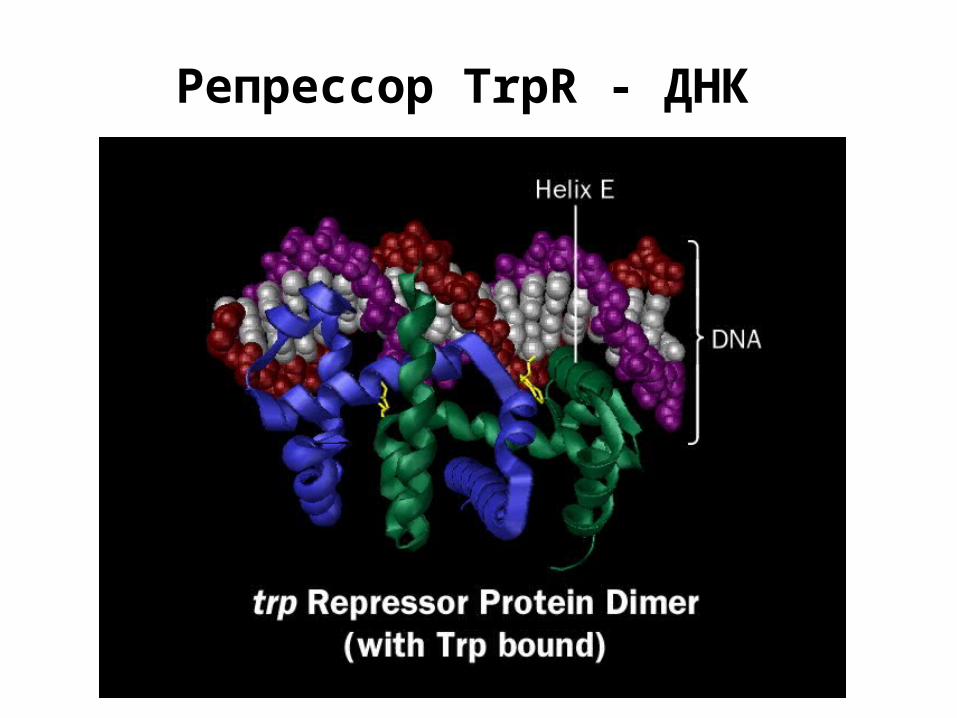
Репрессор TrpR - ДНК
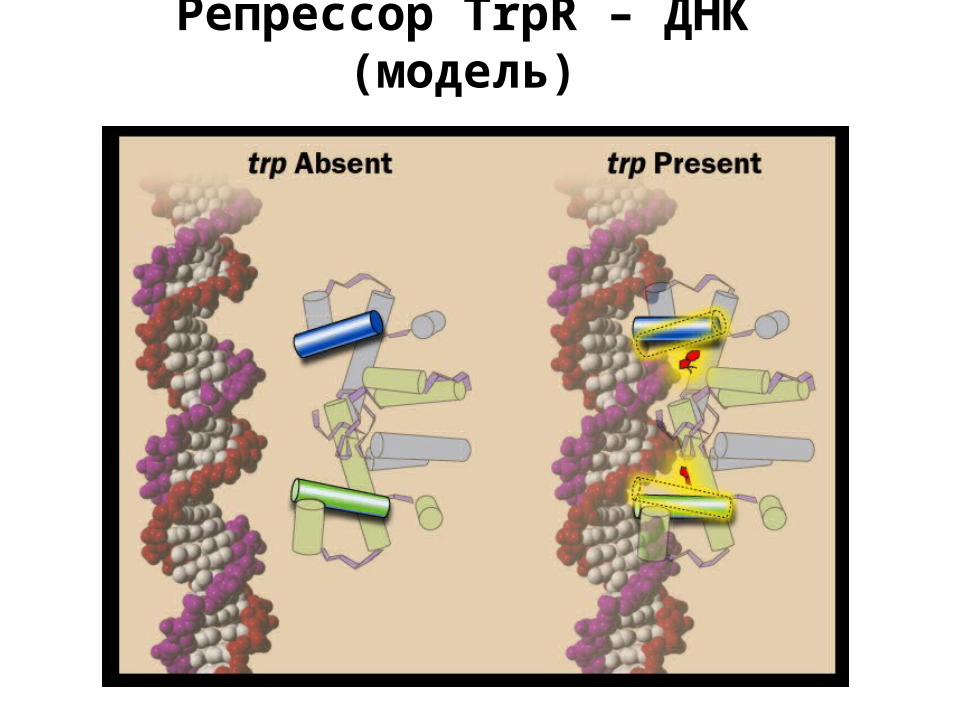
Репрессор TrpR – ДНК (модель)
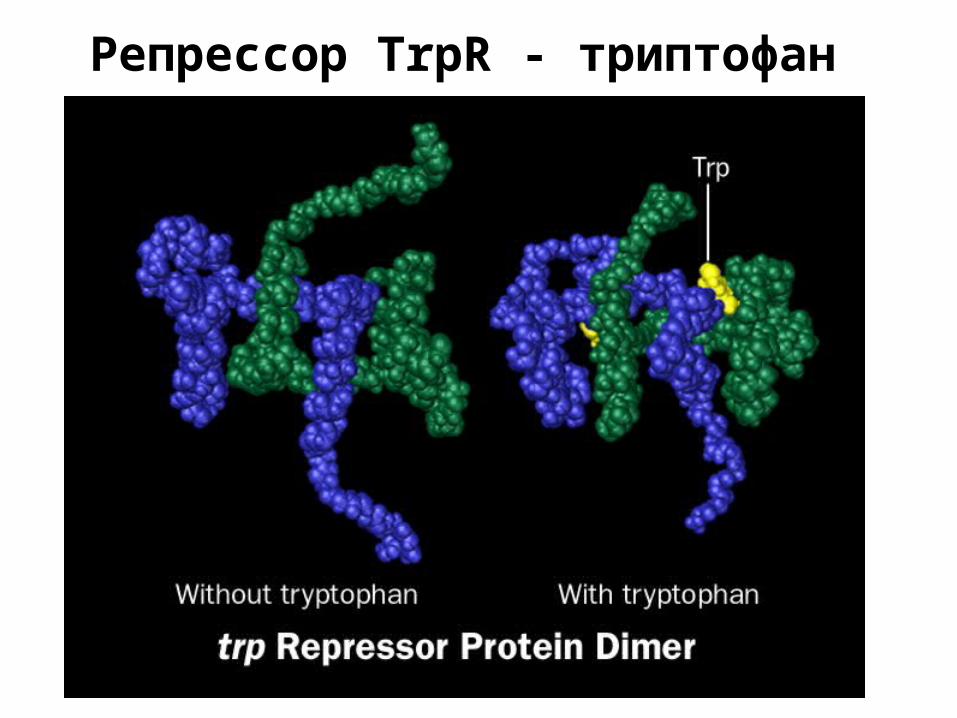
Репрессор TrpR - триптофан

Примеры
• триптофановый репрессор – биосинтез триптофана – связан с ДНК при наличии триптофана
• лактозный репрессор– катаболизм (использование) лактозы– связан с ДНК при отсутствии лактозы
• арабинозный активатор– катаболизм (использование) арабинозы– связан с ДНК при наличии арабинозы
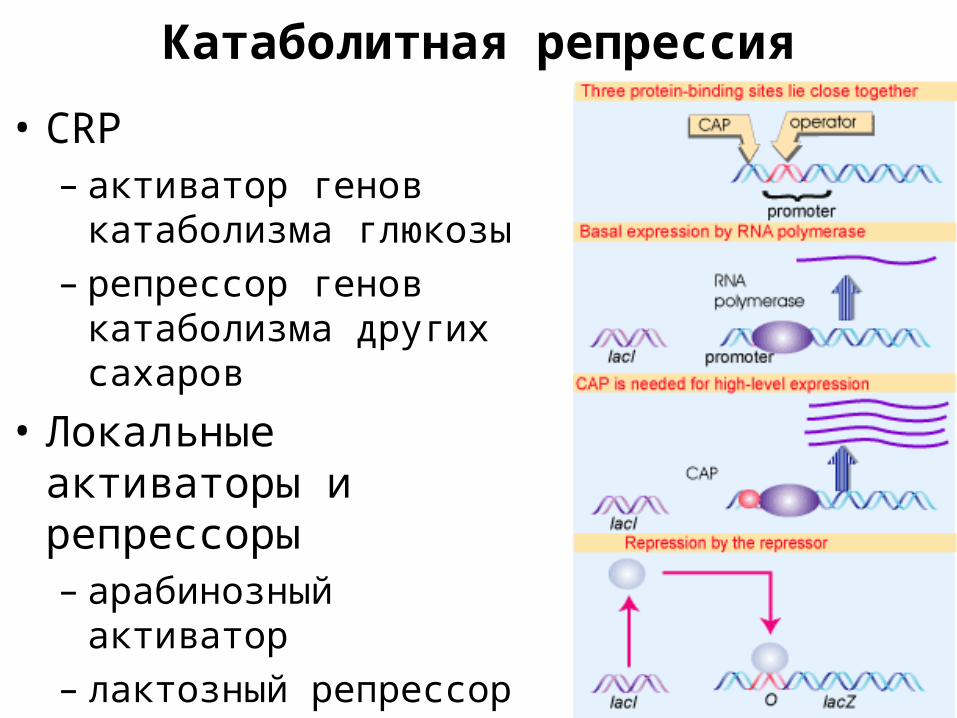
Катаболитная репрессия• CRP
– активатор генов катаболизма глюкозы
– репрессор генов катаболизма других сахаров
• Локальные активаторы и репрессоры– арабинозный активатор– лактозный репрессор
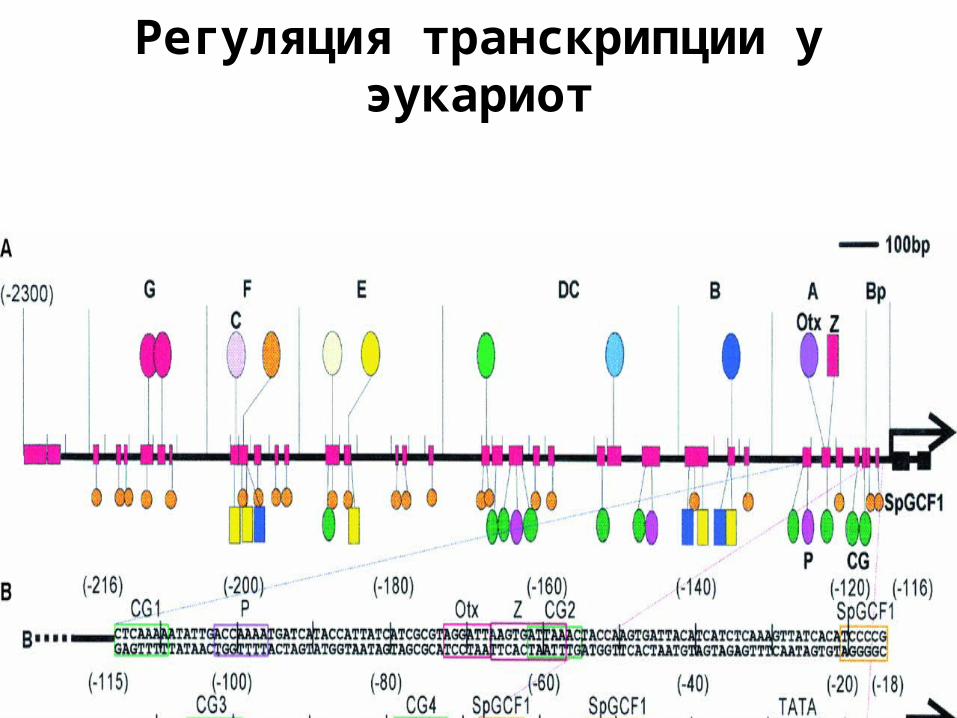
Регуляция транскрипции у эукариот
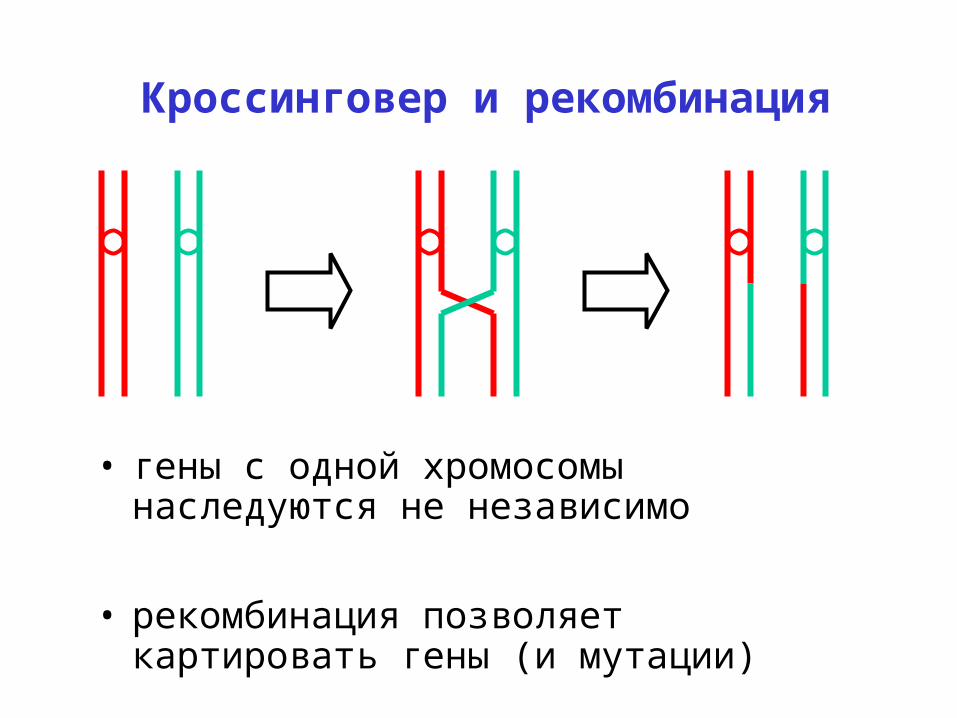
Кроссинговер и рекомбинация
• гены с одной хромосомы наследуются не независимо
• рекомбинация позволяет картировать гены (и мутации)
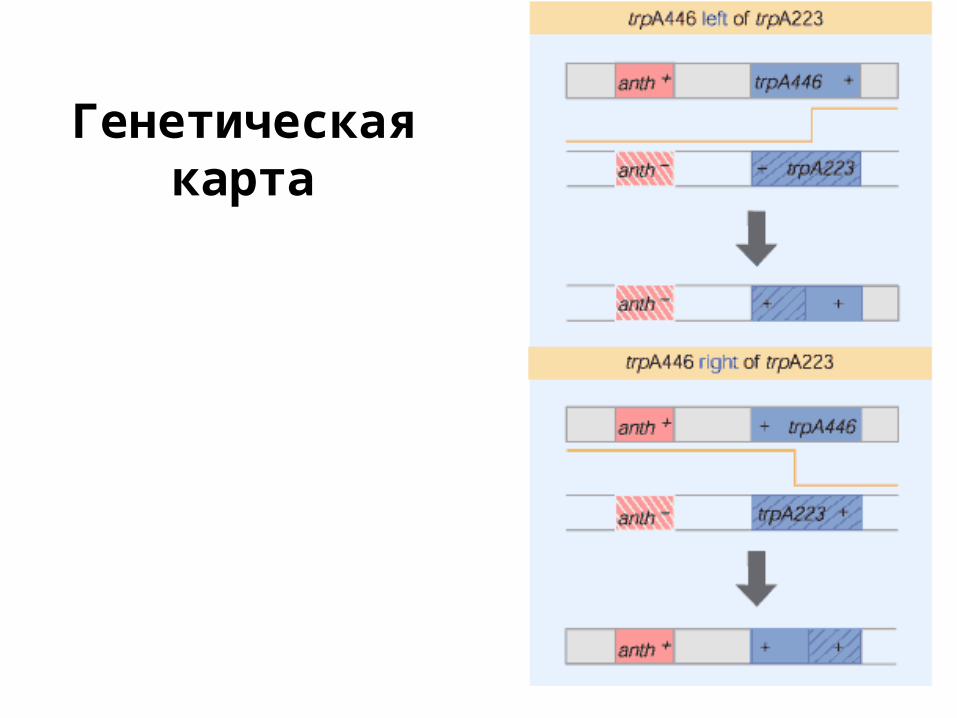
Генетическаякарта
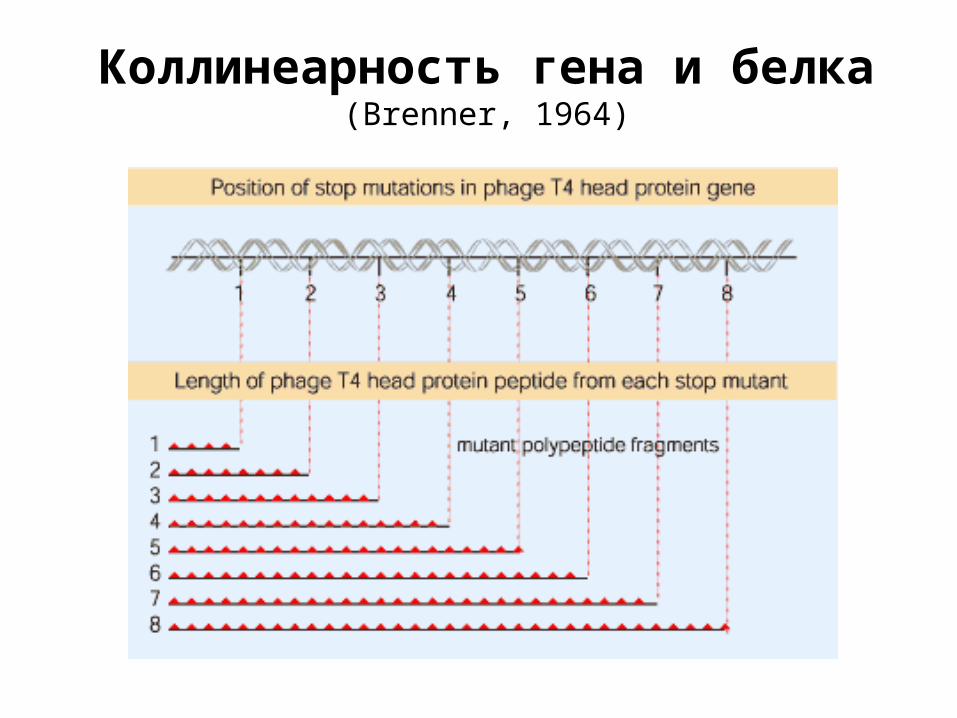
Коллинеарность гена и белка (Brenner, 1964)
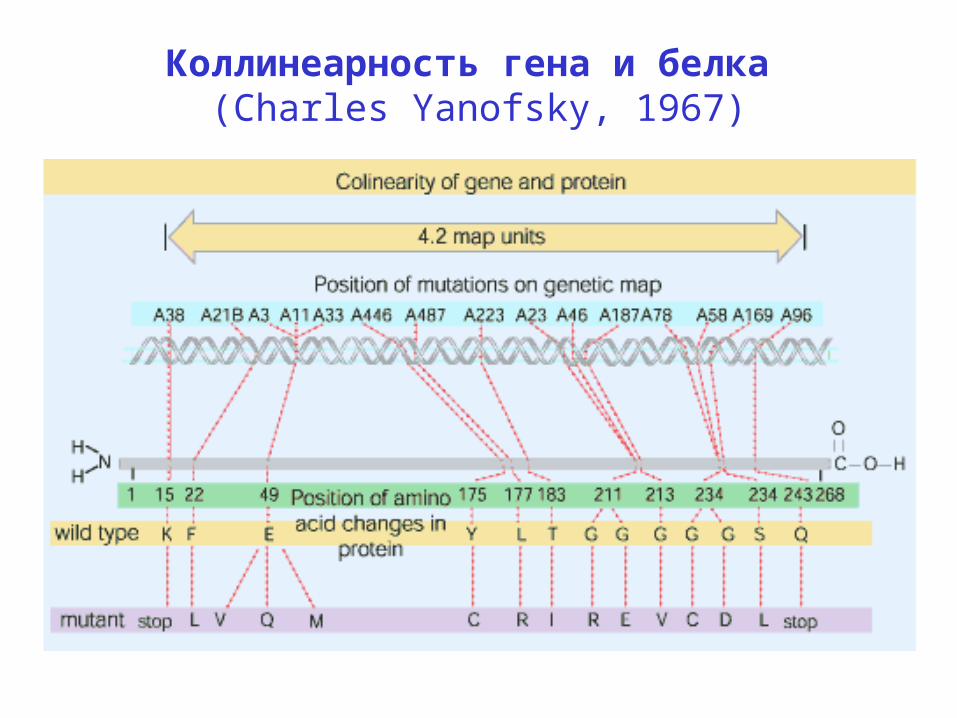
Коллинеарность гена и белка (Charles Yanofsky, 1967)

Генетический код• Генетический код – соответствие между ДНК и белком
(George Gamow, 1954 (Георгий Гамов)
• Крик и соавторы (1961):– не перекоывающийся (одна мутация – одна
аминокислота)– вырожденный (несоклько кодонов для одной
аминокислоты)– без запятых (меркеров между кодонами)– периодический

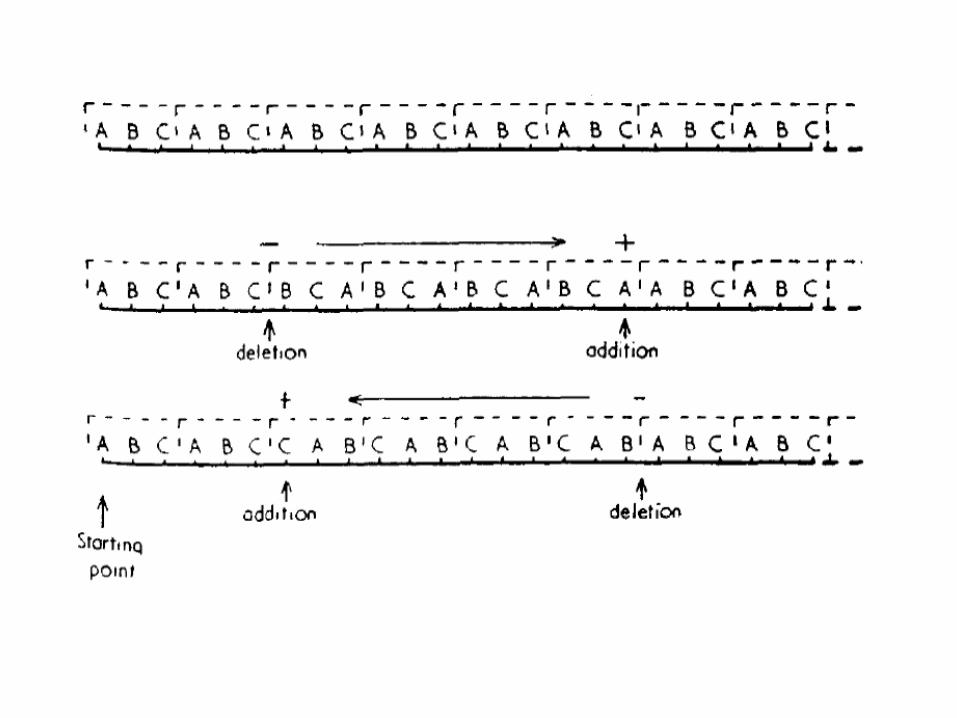
кодон - триплет• мутации, вызванные актидиновыми красителями
– не протекающие (функция не ослабляется, а исчезает)– механизм – вставки/делеции оснований
(дальше все путается код без запятых)
CUACUACUACUACUACUACUACUACUACUACUACUACUALeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeuLeu
вставкаCUACUACUACGUACUACUACUACUACUACUACUACUACULeuLeuLeuArgThrThrThrThrThrThrThrThrThr
делецияCUACUACUACUACUACUACUACUACUACACUACUACUACLeuLeuLeuLeuLeuLeuLeuLeuLeuHisTyrTyrTyr
U
G

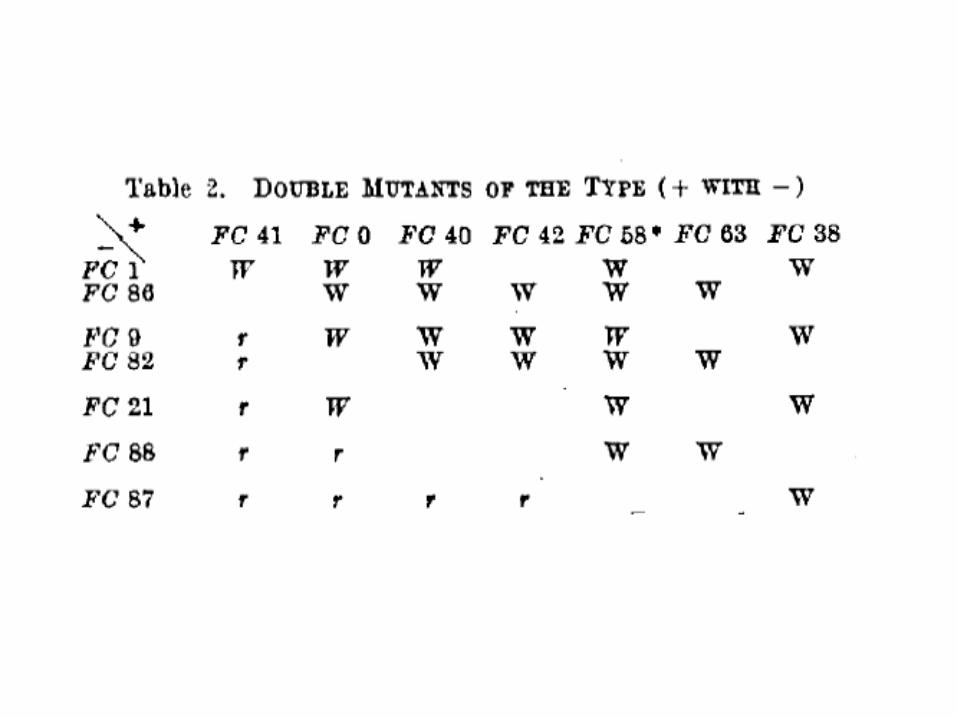
Двойные мутанты и ревертанты• два класса мутаций: (+) и (–) • двойной мутант (+)¤(+) и (–)¤(–) – все еще мутантные
фенотипы• двойные мутанты (+)¤(–) и (–)¤(+) – протекающие
фенотипы
CUACUACUACGUACUACUACUACUACUACUACUACUACULeuLeuLeuArgThrThrThrThrThrThrThrThrThr
¤CUACUACUACUACUACUACUACUACUACACUACUACUACLeuLeuLeuLeuLeuLeuLeuLeuLeuHisTyrTyrTyr
CUACUACUACGUACUACUACUACUACUACACUACUACUALeuLeuLeuArgThrThrThrThrThrThrLeuLeuLeu
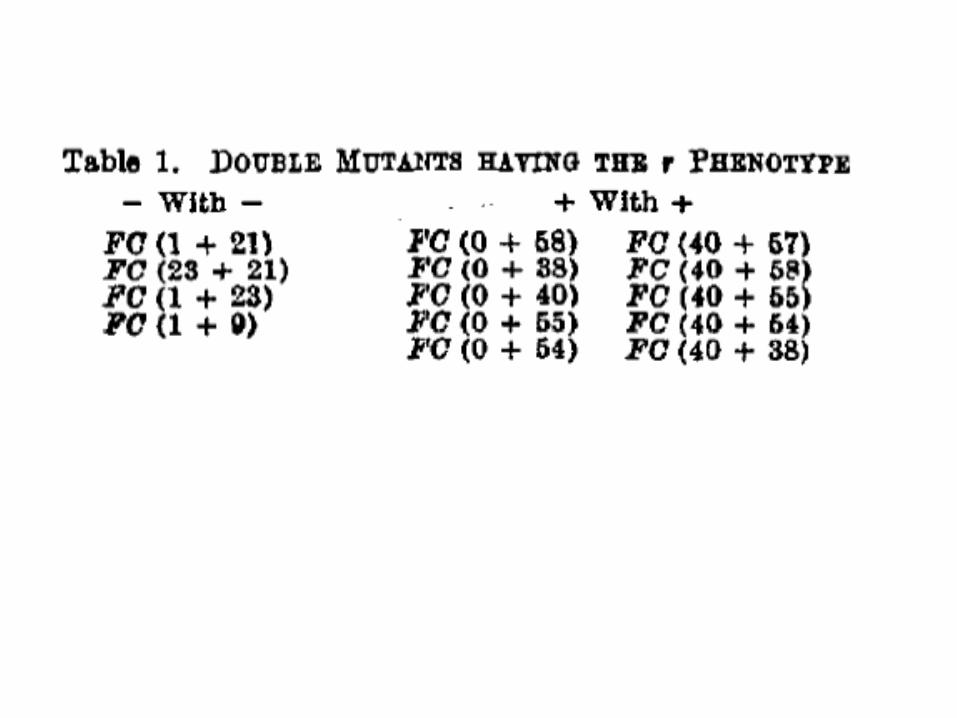

Тройные мутанты – реверсия!• тройные мутанты из одного класса, (+)¤(+)¤(+) и (–)¤(–)¤(–),
дают протекающие фенотипы
CUACUACUACGUACUACUACUACUACUACUACUACUACUACULeuLeuLeuArgThrThrThrThrThrThrThrThrThrThr¤CUACUACUACUACUACUACGUACUACUACUACUACUACUACULeuLeuLeuLeuLeuLeuArgThrThrThrThrThrThrThr
двойной мутант – нет функции
CUACAUCUACGUACUACUACGUACUACUACUACUACUACUACLeuLeuLeuArgThrThrThrTyrTyrTyrTyrTyrTyrTyr¤CUACUACUACUACUACUACUACUACUACGUACUACUACUACULeuLeuLeuLeuLeuLeuLeuLeuLeuArgThrThrThrThr
тройной мутант – слабая функцияCUACUACUACGUACUACUACGUACUACUACGUACUACUACUALeuLeuLeuArgThrThrThrTyrTyrTyrValLeuLeuLeu

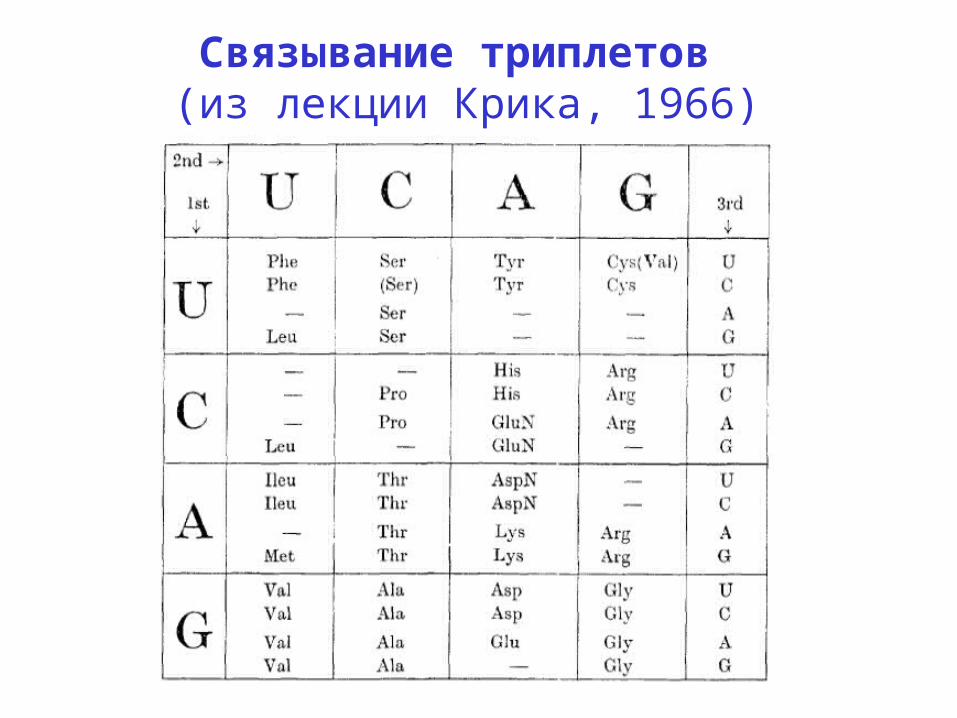
Связывание триплетов (из лекции Крика, 1966)
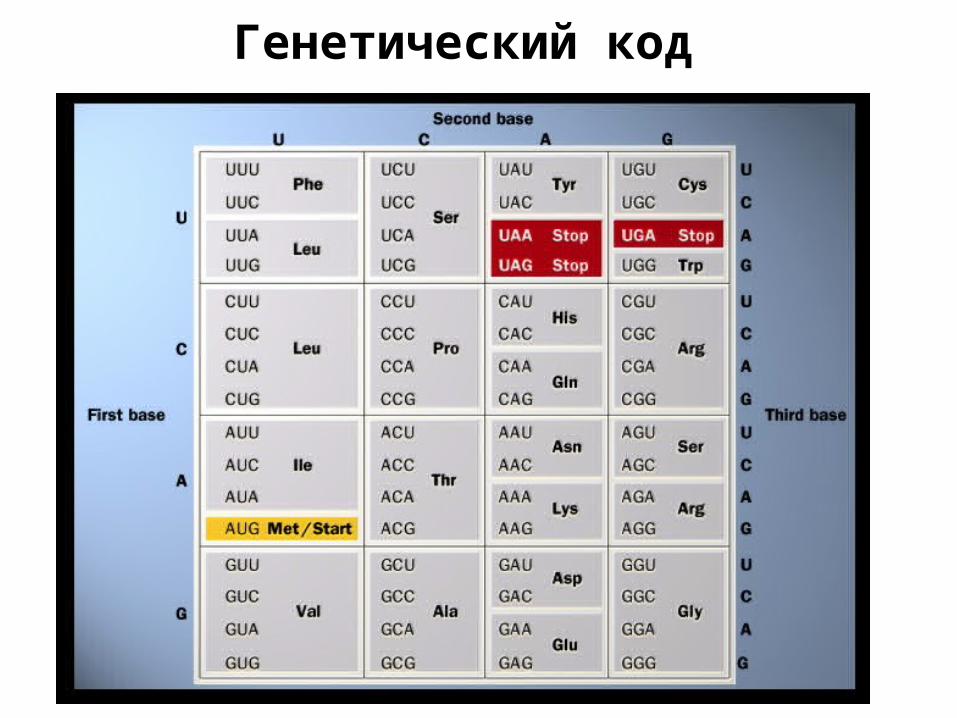
Генетический код
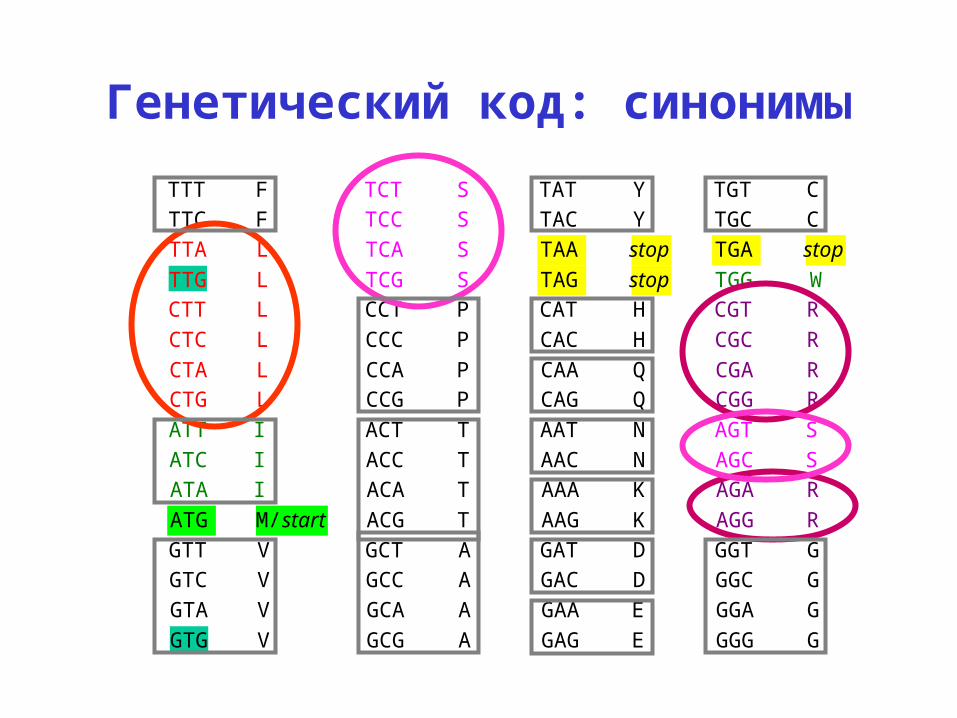
Генетический код: синонимыTTT F TCT S TAT Y TGT CTTC F TCC S TAC Y TGC CTTA L TCA S TAA stop TGA stopTTG L TCG S TAG stop TGG WCTT L CCT P CAT H CGT RCTC L CCC P CAC H CGC RCTA L CCA P CAA Q CGA RCTG L CCG P CAG Q CGG RATT I ACT T AAT N AGT SATC I ACC T AAC N AGC SATA I ACA T AAA K AGA RATG M/ start ACG T AAG K AGG RGTT V GCT A GАT D GGT GGTC V GCC A GАC D GGC GGTA V GCA A GАA E GGA GGTG V GCG A GАG E GGG G
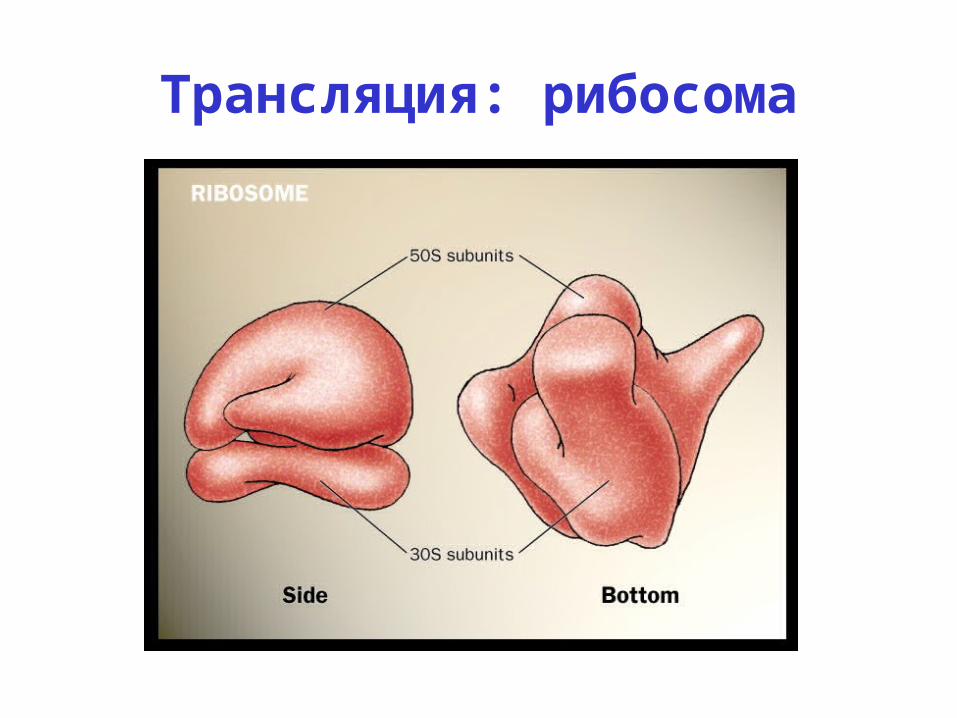
Трансляция: рибосома
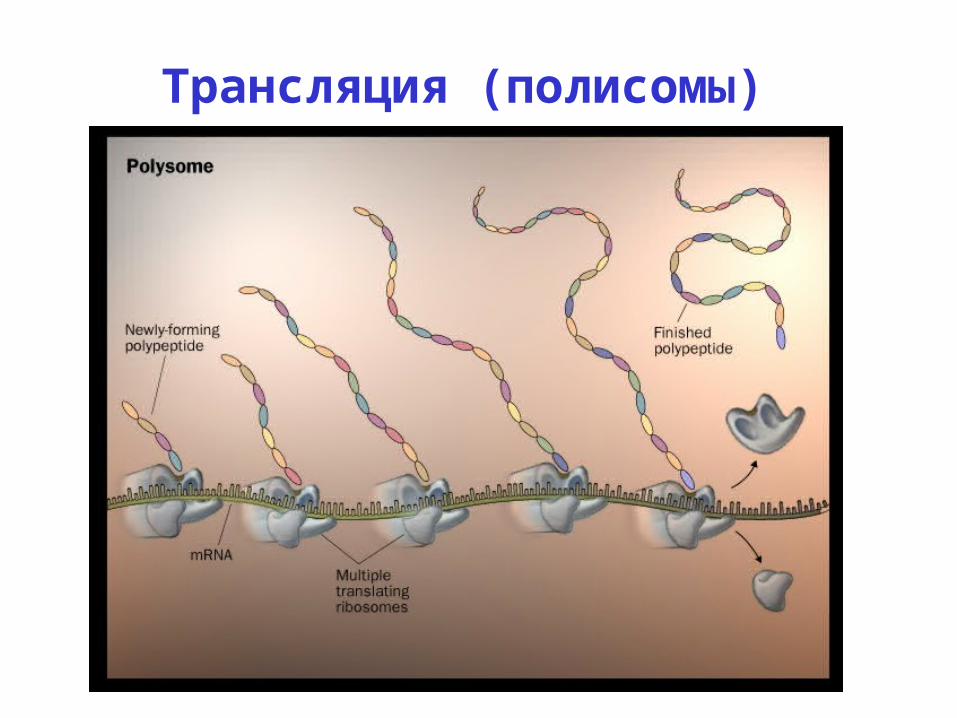
Трансляция (полисомы)

Polysomes

Адапторная гипотеза (F.Crick и S.Brenner)
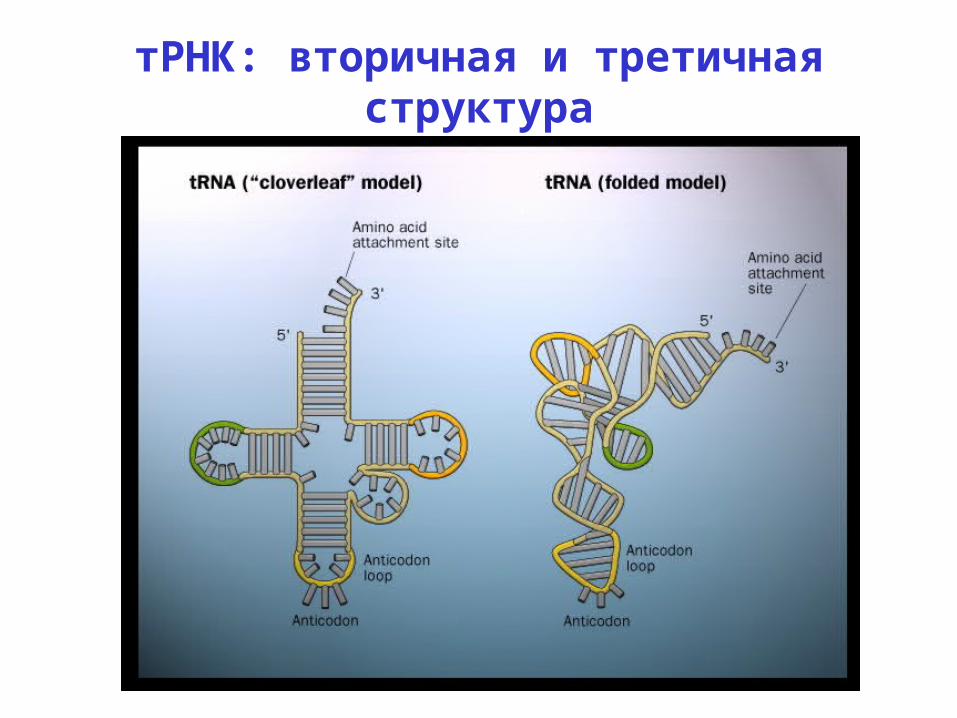
тРНК: вторичная и третичная структура
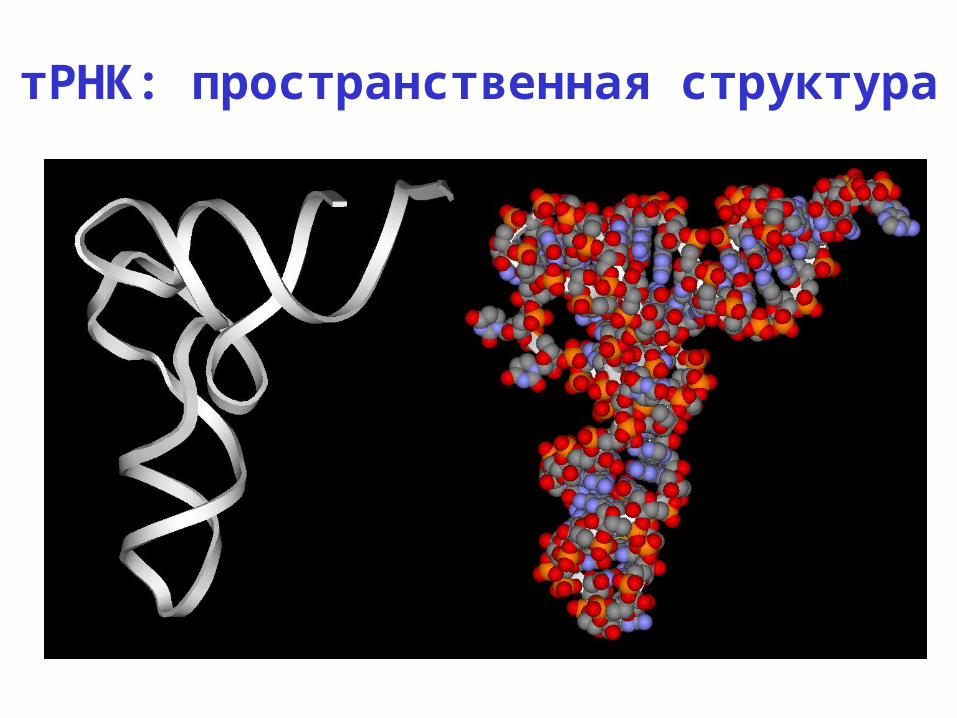
тРНК: пространственная структура
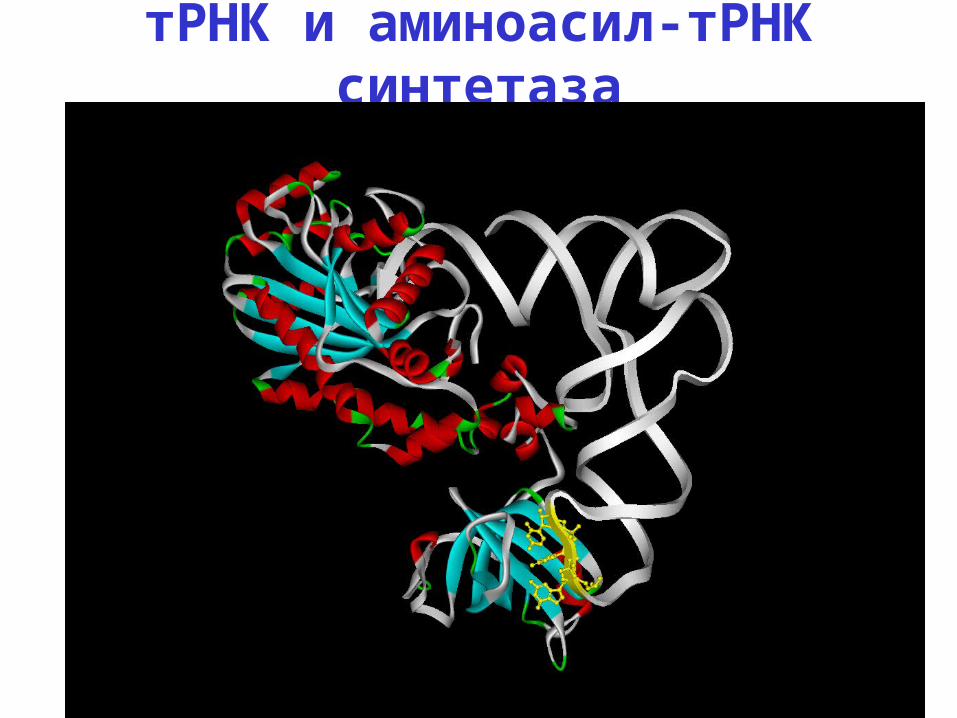
тРНК и аминоасил-тРНК синтетаза
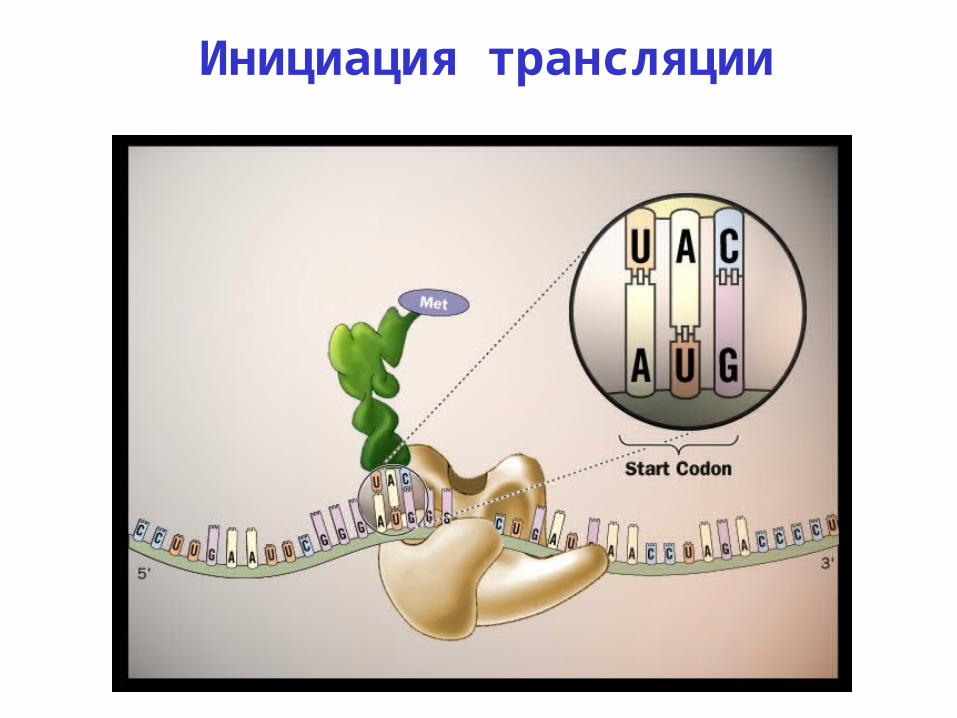
Инициация трансляции

Старт трансляцииdnaN ACATTATCCGTTAGGAGGATAAAAATGgyrA GTGATACTTCAGGGAGGTTTTTTAATGserS TCAATAAAAAAAGGAGTGTTTCGCATGbofA CAAGCGAAGGAGATGAGAAGATTCATGcsfB GCTAACTGTACGGAGGTGGAGAAGATGxpaC ATAGACACAGGAGTCGATTATCTCATGmetS ACATTCTGATTAGGAGGTTTCAAGATGgcaD AAAAGGGATATTGGAGGCCAATAAATGspoVC TATGTGACTAAGGGAGGATTCGCCATGftsH GCTTACTGTGGGAGGAGGTAAGGAATGpabB AAAGAAAATAGAGGAATGATACAAATGrplJ CAAGAATCTACAGGAGGTGTAACCATGtufA AAAGCTCTTAAGGAGGATTTTAGAATGrpsJ TGTAGGCGAAAAGGAGGGAAAATAATGrpoA CGTTTTGAAGGAGGGTTTTAAGTAATGrplM AGATCATTTAGGAGGGGAAATTCAATG

Выравнивание сайтов связывания рибосом
dnaN ACATTATCCGTTAGGAGGATAAAAATGgyrA GTGATACTTCAGGGAGGTTTTTTAATGserS TCAATAAAAAAAGGAGTGTTTCGCATGbofA CAAGCGAAGGAGATGAGAAGATTCATGcsfB GCTAACTGTACGGAGGTGGAGAAGATGxpaC ATAGACACAGGAGTCGATTATCTCATGmetS ACATTCTGATTAGGAGGTTTCAAGATGgcaD AAAAGGGATATTGGAGGCCAATAAATGspoVC TATGTGACTAAGGGAGGATTCGCCATGftsH GCTTACTGTGGGAGGAGGTAAGGAATGpabB AAAGAAAATAGAGGAATGATACAAATGrplJ CAAGAATCTACAGGAGGTGTAACCATGtufA AAAGCTCTTAAGGAGGATTTTAGAATGrpsJ TGTAGGCGAAAAGGAGGGAAAATAATGrpoA CGTTTTGAAGGAGGGTTTTAAGTAATGrplM AGATCATTTAGGAGGGGAAATTCAATG
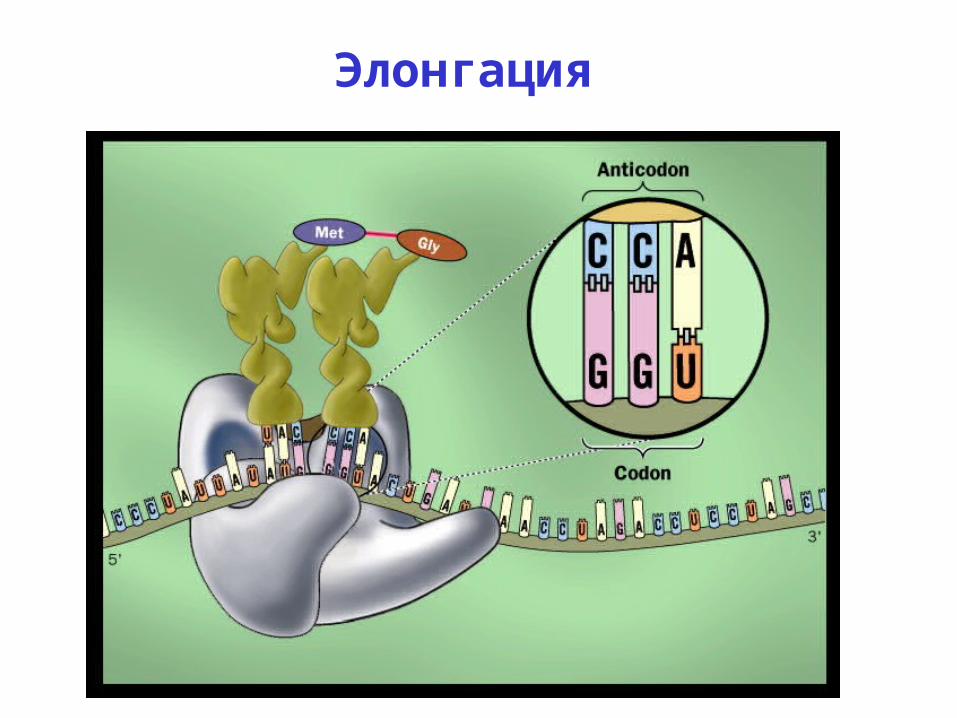
Элонгация
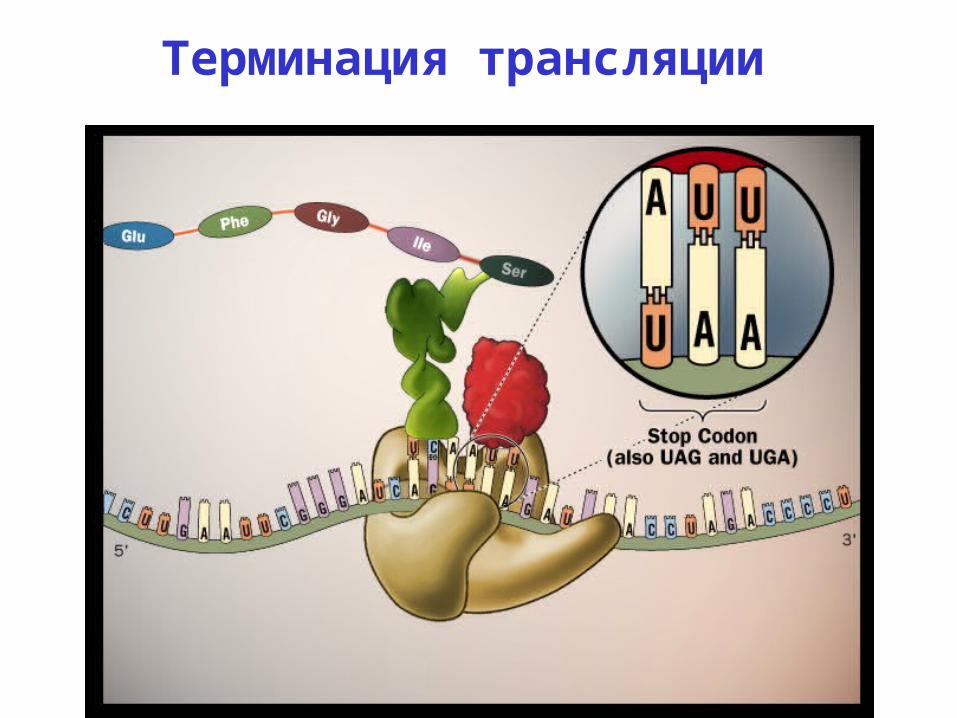
Терминация трансляции

трансляция - кино

Диалекты кода
• Генетический код не универсален, но– различия невелики– встречаются обычно в небольших геномах и
геномах органелл (и у инфузорий)– затрагивают ограниченный набор семейств.– Часто увеличивается симметрия.– многие изменения затрагивают стоп-кодоны.
• Дополнительные аминокислоты– формилметионин– селеноцистеин– пирролизин
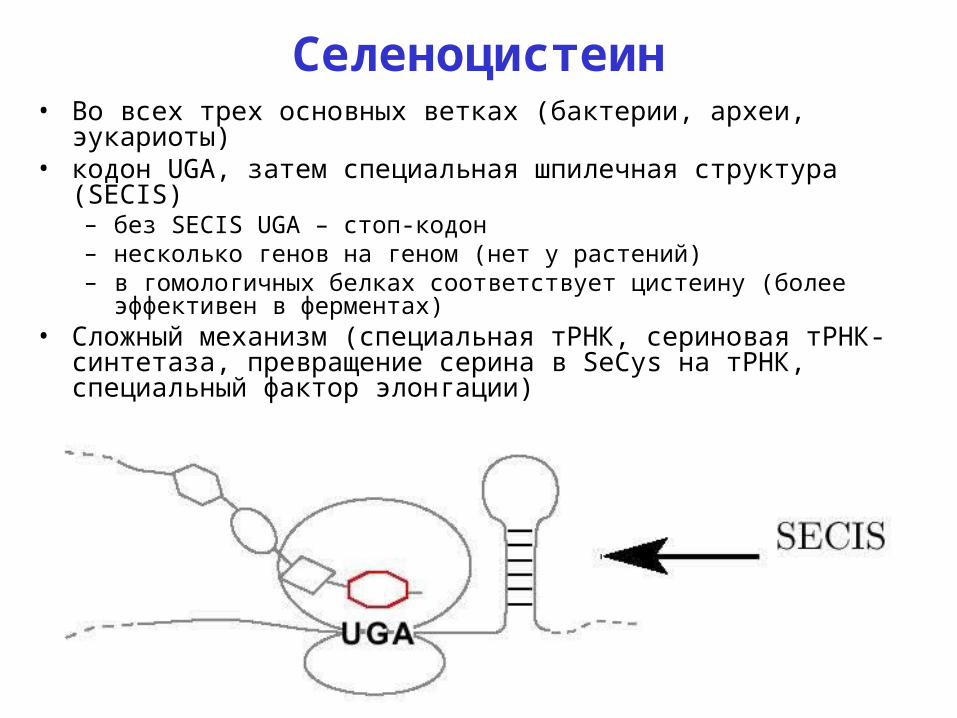
Селеноцистеин• Во всех трех основных ветках (бактерии, археи, эукариоты)• кодон UGA, затем специальная шпилечная структура (SECIS)
– без SECIS UGA – стоп-кодон– несколько генов на геном (нет у растений)– в гомологичных белках соответствует цистеину (более эффективен в
ферментах)• Сложный механизм (специальная тРНК, сериновая тРНК-
синтетаза, превращение серина в SeCys на тРНК, специальный фактор элонгации)

Выравнивание SECIS
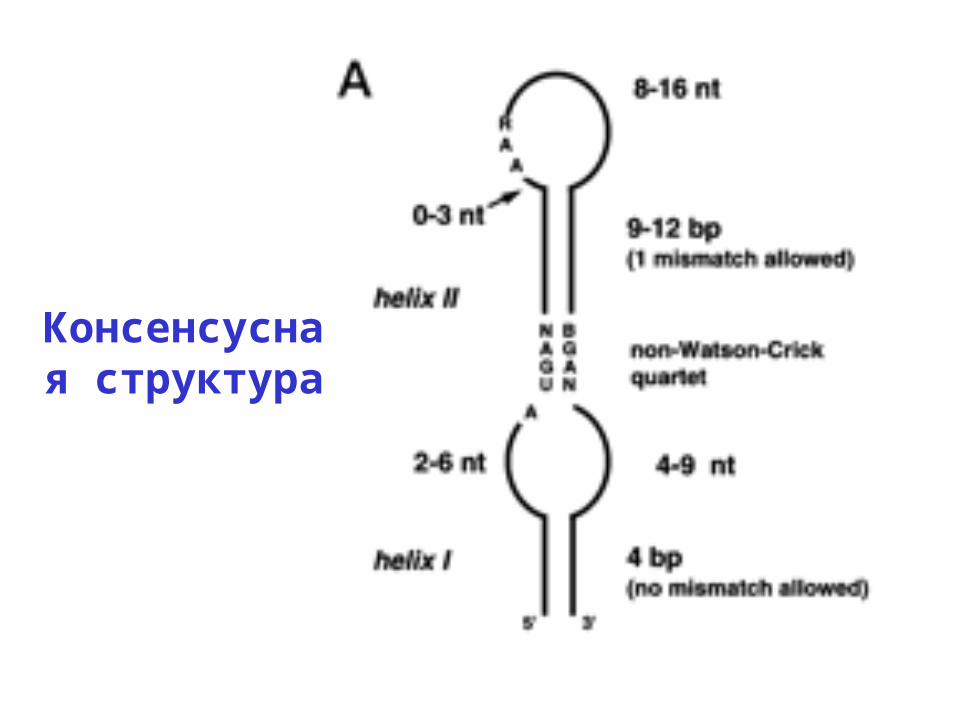
Консенсусная структура
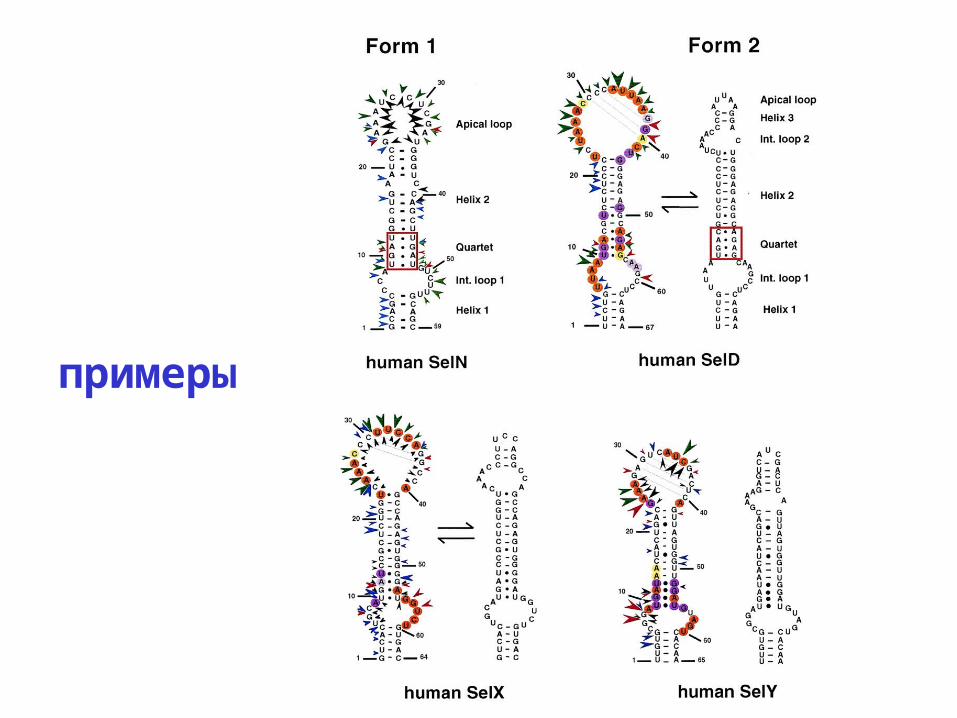
примеры