Embed Size (px)
Citation preview

− 53 −
宮崎医会誌 2014 ; 38 : 53-7.
総 説
は じ め に
樹状細胞(dendritic cells ; DCs)は樹状突起を有する系統マーカー陰性,主要組織適合遺伝 子 複 合 体(major h i s t o compa t ib i l i t y comp lex ; MHC)クラスII陽性の抗原提示細胞であり,造血幹細胞よりミエロイド系やリンパ 球 系 分 化 経 路 を 経 て 未 熟DCs(immature DCs ; iDCs)へ分化し,さらに様々な刺激により成熟DCs(mature DCs ; mDCs)に至る1−2)。生体においてDCsは分化系列,成熟段階の異なる多様なサブセットとして末梢非リンパ組織やリンパ組織に広く存在しており,機能的には通常型DCs(conventional DCs ; cDCs)と 形 質 細 胞 様DCs(pla smacy to id DCs ; pDCs)に大別される1−2)。 病原性微生物の外来抗原の末梢組織への侵襲により惹起された炎症状態では,DCsは自然免疫と獲得免疫を繋ぐ最も強力な抗原提示細胞として免疫系を賦活化する1−2)。一方,非炎症状
態(定常状態)では末梢においてDCsは免疫寛容を誘導し,免疫学的恒常性の維持に重要であると推測されている1−2)。 pDCsはウイルス感染で多量のI型インターフェロン(inter f e ron ; IFN)を産生する免疫細胞として同定された3−4)。pDCsはcDCsと異なり,エンドソーム内核酸受容体であるTol l様受容体(toll - l ike receptor ; TLR)7とTLR9のみを高発現し,核酸認識後にMyD88依存的なnuclear factor κB (NF-κB)の活性化を介して炎症性サイトカインを産生し,inhibitor o f NF-κB (IκB) kinase (IKK)-αとIFN regulatory factor- 7 (IRF-7)の活性化を介してI型IFNを産生する5)。 pDCsはI型IFN高産生能に基づく抗ウイルス感染防御への主要なメディエーターとして考えられている3−4)。また,pDCsはT細胞免疫応答において,活性化あるいは抑制的に多面的な役割を担っていると推測されている6−8)。しかしながら,これらの推察は試験管内実験や免疫細胞移入実験の知見に基づいており,pDCsの生体内での詳細な免疫学的役割は不明である。生
形質細胞様樹状細胞によるT細胞の反応の制御
佐藤 克明
要約:樹状細胞(dendritic cells ; DCs)は樹状突起を有する系統マーカー陰性,主要組織適合遺伝子複合体(major histocompatibility complex ; MHC)クラスII陽性の抗原提示細胞であり,通常型樹状細胞(conventional DCs ; cDCs)と形質細胞様樹状細胞(plasmacytoid DCs ; pDCs)に大別されるサブセットから構成される。pDCsはToll様受容体(toll-like receptor ; TLR)7とTLR9を高発現し,ウイルス感染で多量のI型インターフェロン(interferon; IFN)を産生することから,抗ウイルス感染防御への主要なメディエーターとして考えられている。しかしながら,生体内での免疫応答におけるpDCsの役割は不明である。著者らはpDCsの免疫学的役割を解明するために,pDCs特異的発現分子として同定したSiglec-Hの欠損マウスとpDCs自身が消失したpDCs特異的消失マウスを作製した。本稿では,著者らが得た知見に基づいてpDCsの炎症反応とT細胞免疫応答における役割を概説する。 〔平成26年3月27日入稿,平成26年6月9日受理〕
宮崎大学医学部医学科感染症学講座免疫学分野

− 54 −
宮崎医会誌 第38巻 第2号 2014年9月
体 内 で の 免 疫 応 答 に お け るpDCsの 解 析 で はpDCsで 発 現 が 認 め ら れ るGr - 1 やbone marrow stromal antigen 2 (BST2)に対する抗体を用いた除去法が用いられてきた4)。しかしながら,この手法の問題点として,これらの分子は他の免疫細胞でも発現が示され,抗Gr- 1抗体や抗BST2抗体の投与ではpDCsを含む 多 種 の 免 疫 細 胞 が 除 去 さ れ る こ と か ら,pDCsの生体内での免疫学的役割の解釈に齟齬が生じている4)。従って,生体内でのpDCsの機能解析はその特異的消失法の欠如により妨げられている。 著者らはpDCsの機能制御機構を明らかにするためにその特異的発現分子であるSiglec -Hを欠損したSig l ec -H欠損マウスを作製した9)。さらに,pDCsの生体内での免疫学的役割を解明するために,Siglec -H遺伝子の一部にジフテリア毒素受容体遺伝子を導入後,ジフテリア毒素を 投 与 す る こ と に よ りpDCs自 身 が 消 失 し たpDCs特異的消失マウスを作製した 9)。 本稿ではこれまで不明であった生体内でのpDCsの免疫応答における役割とその機能制御機構について紹介する9)。
pDCsの機能制御におけるSiglec-Hの役割
は じ め にSig l ec -H欠 損pDCsの 性 状 解 析 を行った。野生型pDCsと比較してSig l ec -H欠損pDCsではCD11cとBST2の発現亢進,B220とMHCクラスIIの発現低下が認められた。このことから,Siglec -Hは転写因子の発現調節によりpDCsの分化を制御することが示唆された。ま た,TLR9リ ガ ン ド で あ るCpG-A刺 激 後,Sig lec -H欠損pDCsは野生型pDCsと比較して,I型IFNと 炎 症 性 サ イ ト カ イ ン で あ るinter leukin (IL)-12の著しい産生亢進を示した。さらに,Siglec -H欠損pDCsではCpG-A刺激によるIKK-αとIRF -7の活性化やNF-κBの活性化が野生型pDCsと比較して増強していた。pDCsにおいてSig l ec -Hはアダプター分子であるDAP12と会合していることから,Sig l ec -Hと未知リガンドとの結合はDAP12の細胞内ド
メイン中の活性化アミノ酸配列モチーフである immunoreceptor tyrosine-based activation mot i f(ITAM)をリン酸化し,さらにチロシンキナーゼであるSykの活性化を誘導することが推察された9)。これらの結果から,Sig l ec -HはDAP12-Syk経路を介してTLR9-MyD88依存性シグナルを阻害することにより,TLR9リガンド誘導性サイトカイン産生を抑制すると考えられた。 さらに,Siglec-HによるpDCsのT細胞活性化能 の 制 御 に つ い て 検 討 し た。 そ の 結 果,Siglec -H欠損pDCsは野生型pDCsと比較して抗原特異的CD4+T細胞活性化能が減弱し,これはMHCクラスII分子の低下によることが考えられた。また,Siglec -H欠損pDCsは野生型pDCsと比較して抗原特異的CD8+T細胞活性化能も減弱していた。野生型pDCsとSiglec -H欠損pDCsは同程度のMHCクラスIの発現を示していることから,これは抗原クロスプレゼンテーション経路10)の障害によることが考えられた。
pDCsによる炎症反応の制御
pDCsのS ig l e c -Hを介したTLR9リガンド誘発性炎症反応に対する制御について検討した。野生型マウスではCpG-AとD-ga l a c t o sam ineの投与により血清中でのI型IFNや炎症性サイトカインの産生が認められ,投与後24時間以内に炎症性ショックにより全例死亡した。一方,S ig l e c -H欠損マウスではこれら血清中サイ ト カ イ ン 産 生 の 著 し い 亢 進 が 認 め ら れ た。しかしながら,pDCs特異的消失マウスではI型IFNの ほ ぼ 完 全 な 産 生 抑 制, 炎 症 生 サ イ トカインについては部分的な産生抑制が認められ,死亡率の低下が示された。これらの結果から,生体内においてpDCsはTLR9リガンド刺 激 に 対 す るI型IFNの 主 要 な 産 生 細 胞 で あり,炎症性サイトカインの産生には部分的に寄与していることが示され,S ig l e c -Hが直接的にこれらサイトカイン産生を制御していることが明らかとなった。また,TLR9リガンド誘 導 性 のcDCsの 成 熟 に つ い て 検 討 し た と こ

− 55 −
佐藤 克明:形質細胞様樹状細胞による感染防御
ろ, 野 生 型 マ ウ ス のcDCsに お け るMHCク ラスI Iと 共 刺 激 分 子 の 発 現 増 加 と 比 較 し てS ig l e c -H欠損マウスではこれらのさらなる発現増強が示されたが,pDCs特異的消失マウスではこれらの分子の発現が低下していた。以上の結果から,cDCsのTLR9リガンド誘導性成熟機構にはpDCsによるサイトカイン産生が関与することが示された。
pDCsによる抗原特異的CD4+ T細胞応答の制御
pDCsのS i g l e c -Hを介した抗原特異的CD4+ T細胞応答に対する制御について検討した。抗原と完全フロイントアジュバントの投与(免疫)後,野生型マウスと比較してS ig l e c -H欠損マウスではより強い抗原特異的CD4+ T細胞増殖が認められ,pDCs特異的消失マウスではさらなるその増強が示された。一方,抗原特異 的 IFN -γ 産 生 CD4+ T 細 胞(type 1 T h e l p e r T細胞 ; T H1細胞)の誘導では野生型マウスと比較してS ig l e c -H欠損マウスでは減弱し,pDCs特異的消失マウスでは増強が認められた。これらの結果から,生体内においてpDCsはS i g l e c -Hが関与するMHCクラスI I拘束性抗原提示とサイトカイン産生の制御に基づいて抗原特異的CD4+ T細胞の活性化を抑制することが考えられた。
pDCsによる抗原特異的CD8+ T細胞応答の制御
pDCsのSiglec-Hを介した抗原特異的CD8+ T細胞応答への制御について検討した。野生型マウスでは抗原とCpG-Aの免疫により抗原特異的細胞障害性T細胞(cytotoxic T lymphocytes ; CTLs)が生成した。一方,Siglec-H欠損マウスでは抗原特異的CTLsの生成が低下し,pDCs特異的消失マウスではさらなる低下が認められた。こ れ ら の 結 果 か ら, 生 体 内 に お い てpDCsは
Sig lec -Hが関与する抗原クロスプレゼンテーシ ョ ン を 介 し てCD8+ T細 胞 の 惹 起 を 行 い,CTLsの生成に寄与することが明らかとなった。
pDCsのウイルス感染免疫応答における役割
pDCsのSig l e c -Hを介したウイルス感染免疫応答の制御について単純ヘルペスⅠ型ウイルス
(herpes simplex virus-1;HSV-1)感染モデル を 用 い て 検 討 し た。HSV-1感 染 に よ りSiglec -H欠損マウスでは野生型マウスと比較して,血清中でのI型IFNと IL -12の高産生を示し,pDCs特異的消失マウスではI型IFNの著しい産生抑制,IL -12の弱い産生抑制が認められた。また,野生型マウスでのHSV-1感染によるHSV-1特異的CTLs生成と比較して,Sig lec -H欠 損 マ ウ ス とpDCs特 異 的 消 失 マ ウ ス で はI型IFN産生に対する異なった効果が認められたにも関わらず,ともにHSV-1特異的CTLs生成が減弱していた。さらに,野生型マウスでは感染後,脾臓でHSV-1が検出されないが,pDCs特異的消失マウスはSiglec -H欠損マウスよりも高いHSV-1感染量を示した。これらの結果から,pDCsはウイルス感染においてI型IFNの産生とウイルス抗原のクロスプレゼンテーションによる抗ウイルスCTLsの生成を介してウイルス感染防御免疫応答に寄与することが考えられた。
お わ り に
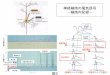
こ の 研 究 か ら,Sig l ec -HはpDCsの 分 化,TLR7/9依存性性サイトカイン産生能,T細胞活性化能を制御することが明らかとなった(図1)。自然免疫応答ではpDCsは初めにウイルスを感知して,I型IFNや炎症性サイトカインを産生して炎症反応を惹起し,cDCsなどの他の免疫細胞を活性化して炎症反応を亢進することを初めてつきとめた(図2)。また,ウイルス

− 56 −
宮崎医会誌 第38巻 第2号 2014年9月
感染に対する適応免疫応答ではpDCsはエフェク タ ー CD4+ T細 胞 の 誘 導 を 抑 制 す る が,CTLsを積極的に生成することによりウイルスやその感染細胞を生体から効率的に排除することを解明した(図2)。従って,この研究の成
果を応用することで,感染症に対する新しい治療法の開発につながる可能性が期待された。今後は自己免疫疾患の発症や増悪におけるpDCsの役割とその制御機構を解明したい。
図2.pDCsによる炎症反応とT細胞免疫応答の制御.左図: 感染誘発性炎症反応では,pDCsは初めにウイルスを感知して,I型IFNや炎症性サイトカインの産生により炎症
反応を惹起し,さらにcDCsを含む他の免疫細胞を活性化して炎症反応を亢進する.右図: 環境抗原やウイルス抗原に対するT細胞免疫応答では,pDCsはcDCsによるエフェクターCD4+ T細胞の誘導を抑
制するが,抗原クロスプレゼンテーションによるCTLsの生成ではpDCsはcDCsと協調的に作用する.
図1.Siglec-HによるpDCsの機能制御.自己傷害細胞からの自己DNA・RNA,ウイルスなどの病原性微生物由来のCpG DNA・RNAを認識したpDCsにおいて,Siglec-Hと未知リガンドとの結合はDAP12-Syk経路の活性化によりTLR7/9-MyD88依存性のIKK-αとIRF-7の活性化やNF-κBの活性化を阻害し,I型IFNと炎症性サイトカインの産生を抑制する.また,Siglec-H介在性シグナルは転写因子(E2-2・IRF-8・Spi-B・PU.1・Id2)の発現調節を介してpDCsの分化を制御すると考えられる.さらに,この分化制御に基づくMHCクラスII分子発現や抗原クロスプレゼンテーション経路の調節によりMHCクラスII拘束性抗原提示とMHCクラスI依存性クロスプレゼンテーションを介するCD4+ T細胞 とCD8+ T細胞のプライミングを制御すると考えられる.

− 57 −
佐藤 克明:形質細胞様樹状細胞による感染防御
参考文献
1) Sato K, Fujita S. Dendritic cells-Nature and classification. Allergol Int 2007 ; 56 : 183-91.
2) Shor tman K, Na ik SH. S t eady-s t a t e and inflammatory dendritic-cell development. Nat Rev Immunol 2007 ; 7 : 19-30.
3) Gilliet M, Cao W, Liu YJ. Plasmacytoid dendritic cells : sensing nucleic acids in viral infection and autoimmune diseases. Nat Rev Immunol 2008 ; 8 : 594-606.
4) Swiecki M, Colonna M. Unraveling the functions of plasmacytoid dendritic cells during viral infections, autoimmunity, and tolerance.Immunol Rev 2010 ; 234 : 142-62.
5) Hoshino K, Sugiyama T, Matsumoto M, et al. IκB kinase-α is critical for interferon-α production induced by Toll-like receptors 7 and 9. Nature 2006 ; 440 : 949-53.
6) Salio M, Palmowski MJ, Atzberger A, et al. CpG-matured murine plasmacytoid dendritic cells are capable of in vivo priming of functional CD8 T cell responses to endogenous but not exogenous antigens. J Exp Med 2004 ; 199 : 567-79.
7) Ochando JC, Homma C, Yang Y, et al. Alloantigen-presenting plasmacytoid dendritic cells mediate tolerance to vascularized grafts. Nat Immunol 2006 ; 6 : 652-62.
8) Irla M, Küpfer N, Suter T, et al. MHC class II-restricted antigen presentation by plasmacytoid dendr i t i c c e l l s i nh i b i t s T ce l l -med i a t ed autoimmunity. J Exp Med 2010 ; 207 : 1891-905.
9) Takagi H, Fukaya T, Eizumi K, et al. Plasmacytoid dendritic cells are crucial for the initiation of inflammation and T cell immunity in vivo. Immunity 2011 ; 35 : 958-71.
10) Villadangos JA, Young L. Antigen-presentation properties of plasmacytoid dendrit ic cel ls. Immunity 2008 ; 29 : 352-61.