Embed Size (px)
Citation preview

Vietnam J. Agri. Sci. 2018, Vol. 16, No. 1: 36-45 Tạp chí Khoa học Nông nghiệp Việt Nam 2018, 16(1): 36-45 www.vnua.edu.vn
36
SỰ LƯU HÀNH CỦA VIRUS GÂY BỆNH THIẾU MÁU TRUYỀN NHIỄM Ở GÀ (CIAV)
TẠI HÀ NỘI VÀ VÙNG PHỤ CẬN
Đào Đoan Trang1*
, Cao Thị Bích Phượng2, Vũ Thị Ngọc
2,
Nguyễn Văn Giáp2, Huỳnh Thị Mỹ Lệ
2
1Trung tâm Thực nghiệm và Bảo tồn vật nuôi
2Khoa Thú y, Học viện Nông nghiệp Việt Nam
Email*: [email protected]
Ngày gửi bài: 08.11.2017 Ngày chấp nhận: 20.03.2018
TÓM TẮT
Trên thế giới đã có nhiều nghiên cứu về bệnh thiếu máu truyền nhiễm và virus gây bệnh. Bài báo này trình bày
kết quả nghiên cứu sự hiện diện của virus gây bệnh thiếu máu truyền nhiễm ở gà (chicken infectious anemia virus -
CIAV) nuôi tại Hà Nội và vùng phụ cận. Bằng phương pháp PCR, đã xác định được virus lưu hành phổ biến với
92,86% số trang trại và 50,81% số mẫu gà ốm dương tính CIAV. Kết quả phân tích trình tự gen mã hóa capsid
protein cho biết: (i) có hai nhóm CIAV (nhóm 2 và nhóm 3) lưu hành ở gà nuôi tại các địa phương lấy mẫu và (ii) các
chủng virus thực địa này khác với chủng virus vacxin (chủng Cux-1, P4 và 3711).
Từ khóa: Virus gây bệnh thiếu máu truyền nhiễm, lưu hành, Hà Nội, vùng phụ cận.
Prevalence of Chicken Infectious Anemia Virus (CIAV) Circulating in Hanoi and Surrounding Provinces
ABSTRACT
Chicken infectious anemia virus (CIAV) and its related disease are well known in all major poultry- producing
countries.. This paper reported the results of investigation on the prevalence of CIAV in Hanoi and surrounding
provinces. By PCR method, the virus was detected in 92.86% of sampling farms and 50.81% of tested samples. The
analyses of capsid protein coding gene revealed two genetic groups (group 2 and 3) of CIAV circulating in
investigated areas. It was also demonstrated that all field strains of CIAV differed from the vaccine strains (Cux-1, P4
and 3711).
Keywords: Chicken infectious anemia virus, prevalence, Hanoi, neighboring provinces.
1. ĐẶT VẤN ĐỀ
Ức chế miễn dðch (immunosuppression) là
träng thái täm thời hoặc låu dài, trong đò khâ
nëng đáp ứng miễn dðch cþa cơ thể bð ânh hưởng
do tùn thương hệ miễn dðch (Dohms & Saif,
1984). Tình träng ức chế miễn dðch gây ra nhiều
hêu quâ như: tëng tỷ lệ chết, giâm tëng trõng
(McNulty et al., 1991), giâm hiệu lực cþa vacxin
(Sun et al., 2009) và mở đường cho nhiều bệnh
kế phát dễ xây ra (Subler et al., 2006). Ở gà, có
hai nhóm nguyên nhân dén tới hiện tượng ức
chế miễn dðch, đò là: (i) nguyên nhån khöng
truyền nhiễm như nuöi dưỡng kém, đûc tø nçm
møc trong thức ën, ... và (ii) nguyên nhån
truyền nhiễm, g÷m mût sø loäi virus như:
Chicken Infectious Anemia Virus (CIAV),
Infectious Bursal Disease Virus (IBDV),
Marek’s Disease Virus (MDV), Avian Leucosis
Virus (ALV), Reticuloendotheliosis Virus
(REV),... (Balamurugan & Kataria, 2006, Hoerr,
2010, Schonewille et al., 2008, Islam et al.,

Đào Đoan Trang, Cao Thị Bích Phượng, Vũ Thị Ngọc, Nguyễn Văn Giáp, Huỳnh Thị Mỹ Lệ
37
2002). CIAV (giö ng Gyrovirus, ho Circoviridae) là
mût virus thường gặp trong nhóm virus gây ức
chế miễn dðch (McNulty, 1991). CIAV có bû gen
là sợi ADN đơn, åm, däng vòng (MacLachlan &
Dubovi, 2017). Bû gen virus chứa 3 cçu trúc
phiên mã mở (ORF), trong đò cò 1 gen cçu trúc
(ORF1) mã hóa capsid protein VP1 (Noteborn et
al., 1991). Gen cçu trýc này cò tính đa däng di
truyền cao nhçt và thường được dùng trong
nghiên cứu đặc điểm dðch tễ hõc phân tử cþa
CIAV (Ducatez et al., 2006, Rimondi et al.,
2014, Olszewska-Tomczyk et al., 2016). Về đặc
điểm dðch tễ hõc, bệnh thiếu máu truyền nhiễm
do CIAV gây ra xuçt hiện ở hæu hết các nước
chën nuöi gà trên thế giới (Bougiouklis et al.,
2007; Toro, 2006; Oluwayelu, 2010). Virus
thường gây bệnh thể lâm sàng ở gà dưới 3 tuæn
tuùi (McIlroy et al., 1992) và thể cên lâm sàng ở
gà trên 3 tuæn tuùi (Adair, 2000). Ở Việt Nam,
mặc dü đã cò bìng chứng dương tính huyết
thanh hõc với CIAV ở gà nuôi täi Hà Nûi và Hà
Nam vào nëm 2013 (Trinh et al., 2015), nhưng
hướng nghiên cứu về bệnh thiếu máu truyền
nhiễm vén còn rçt mới. Do đò, việc nghiên cứu
về bệnh do CIAV gây ra ở Việt Nam là cæn thiết.
Với các cën cứ nêu trên, nghiên cứu này đã được
thực hiện nhìm tìm hiểu sự lưu hành cþa CIAV
ở đàn gà nuöi täi Hà Nûi và vùng phụ cên cÿng
như mût sø đặc điểm dðch tễ hõc phân tử cþa
virus phát hiện được.
2. PHƯƠNG PHÁP NGHIÊN CỨU
2.1. Nguyên liệu
- Méu gûp phþ täng cþa gà øm g÷m:
tuyến Harder, tuyến ức, túi bursal Fabricius,
tþy xương, häch lympho manh tràng; tim, gan,
lách, thên. Phäm vi thu méu là mût sø đàn gà
chưa được tiêm vacxin phòng bệnh thiếu máu
truyền nhiễm nuôi täi Hà Nûi và vùng phụ cên.
- Hóa chçt dùng tách chiết ADN tùng sø
g÷m: (i) dung dðch ly giâi méu có chứa 27%
sucrose, 15 mM trisodium citrate, 0,15 M NaCl,
1 mM ethylene diaminetetraacetic acid, 1%
sodium dodecyl sulphate, 200 µg/ml proteinase
K; (ii) phenol-chloroform-isoamyl alcohol (PCI,
25:24:1); (iii) isopropyl; (iv) c÷n 70%; (v) đệm TE
(pH = 8).
- Vêt liệu cho phân ứng PCR: (i) cặp m÷i
đặc hiệu dùng phát hiện CIAV (CAVVP3F:
TTAAGATGGACGCTCTCCAAGAAGATACT,
CAV2: GGCTGAAGGATCCCTCATTC) được
tham khâo theo nghiên cứu trước đåy (van
Santen et al., 2001); (ii) kít PCR (Maxime PCR
PreMix i-Taq, iNtRON, Hàn Quøc).
- Hóa chçt dùng phân tích sân phèm PCR
g÷m: (i) agarose; (ii) redsafe nucleic acid staining
solution (20,000x); (iii) 100bp DNA ladder.
- Kit tinh säch sân phèm PCR: GeneJET
gel extraction kit (Thermo Fisher Scientific)
2.2. Phương pháp nghiên cứu
2.2.1. Lấy và xử lý mẫu
Do gà nhiễm CIAV thường ở thể cên lâm
sàng (Gholami-Ahangaran, 2011, Haridy et al.,
2012, Rimondi et al., 2014) nên có thể không
biểu hiện triệu chứng điển hình. Cÿng do đåy là
nghiên cứu tương đøi mới về CIAV ở nước ta nên
méu bệnh phèm đã được lçy từ gà øm chưa rô
nguyên nhân, thuûc mõi lứa tuùi. Trong quá
trình thực hiện, đã cò 124 méu được thu thêp ở
14 träi chën nuöi. Méu được chia thành 3 nhóm
(tuæn tuùi): < 3, từ 3 ÷ 5 và ≥ 6. Sau khi đ÷ng
nhçt hoàn nguyên thành huyễn dðch 10% trong
dung dðch PBS 1x, bâo quân ở nhiệt đû -20oC
cho đến khi xét nghiệm.
2.2.2. Tách chiết ADN
ADN tùng sø được tách và tinh säch theo
các bước như sau:
(i) Ly giâi méu: 250 µl huyễn dðch méu
bệnh phèm được trûn đều trong 500 µl dung
dðch sucrose/proteinase K. Ủ ở 56oC/90 phút
hoặc 37oC/12 giờ;
(ii) Tách pha ADN: bù sung 200 µl dung
dðch PCI vào øng méu sau khi ly giâi, Vortex
hún hợp, ly tâm 12.000 vòng/phút/15 phút ở 4oC;
(iii) Tþa ADN: thu 450 μl dðch nùi phía trên
ở bước (ii), trûn đều với 450 μl isopropyl. Tþa
ADN ở -20oC/15 phút, ly tâm 12.000
vòng/phút/15 phút ở 4oC;

Sự lưu hành của virus gây bệnh thiếu máu truyền nhiễm ở gà (CIAV) tại Hà Nội và vùng phụ cận
38
(iv) Rửa tþa ADN: rửa bìng 1 ml c÷n 70%
(pha trong nước cçt đã xử lý DEPC). Ly tâm
12.000 vòng/phút/15 phút ở 4oC, loäi bó hết c÷n,
hong khô ở nhiệt đû phòng trong 15 phút;
(v) Hòa tan tþa ADN: tþa được hòa tan
trong 30 µl đệm TE (pH = 8,0).
2.2.3. PCR phát hiện CIAV
Phân ứng PCR phát hiện CIAV được thực
hiện bìng cặp m÷i CAVVP3F/ CAV2 (van
Santen et al., 2001) với hiệu chînh điều kiện
phân ứng: (i) nhiệt đû bít m÷i 58,5oC; (ii) n÷ng
đû cuøi cùng 0,5 µM cho m÷i xuöi/ ngược. Phân
tích sân phèm PCR bìng điện di trong thäch
agarose 2% có bù sung thuøc nhuûm ADN
(RedSafe) 1x.
2.2.4. Giải trình tự gen
Sân phèm PCR tinh säch được giâi trình tự
theo hai chiều (xuöi và ngược) bìng phương
pháp Sanger (thực hiện täi công ty 1st BASE,
Singapore). Trình tự nucleotide sau đò được
phân tích bìng chương trình tin sinh hõc
BioEdit v7.1.3.0 (Hall, 1999) trên cơ sở đøi chiếu
so sánh (i) giữa trình tự nucleotide được giâi
trình tự theo chiều xuôi và chiều ngược và (ii)
với trình tự gen ORF1 tham chiếu công bø trên
ngân hàng gen.
2.2.5. Phân tích trình tự gen
Nhìm làm rõ møi liên hệ di truyền, 7 chþng
CIAV đäi diện trong nghiên cứu này được so
sánh với (i) 6 chþng CIAV cþa Việt Nam thu
thêp nëm 2013 (BN1, HN1, VP7- VP10, tương
ứng với mã sø truy cêp từ KP780287-
KP780292) và (ii) 3 chþng virus vacxin: Cux-1
(M55918), P4 (AJ890284) và 3711 (EF683159).
Các trình tự gen được síp xếp theo cût
(alignment) bìng phæn mềm ClustalW tích hợp
trong chương trình BioEdit v7.1.3.0 (Hall,
1999). Công cụ Highlighter (https://www.hiv.
lanl.gov/content/sequence/highlight) düng để
hiển thð sự sai khác trình tự nucleotide giữa các
chþng CIAV thực đða với chþng virus vacxin.
Xây dựng cây phát sinh chþng loäi
(phylogenetic tree) bìng phương pháp
Neighbor-joining (với sø bootstrap là 1.000 læn),
dựa trên mô hình Kimura-2 parameter mô
phóng sự biến đùi cþa nucleotide. Phân tích kể
trên được thực hiện bởi phæn mềm MEGA6
(Tamura et al., 2013). Các chþng CIAV được
phân chia thành 3 nhóm di truyền (nhóm 1, 2
và 3a, 3b) dựa theo nghiên cứu đã cöng bø trước
đåy (Olszewska-Tomczyk et al., 2016).
2.2.6. Xử lý số liệu
Sø liệu được tính toán bìng phæn mềm
Microsoft Excel 2007. Phép thử t (t- test) tích
hợp trong phæn mềm Minitab 14 dùng kiểm
đðnh sự sai khác về tỷ lệ dương tính.
3. KẾT QUẢ VÀ THẢO LUẬN
3.1. Sự lưu hành CIAV ở gà nuôi tại Hà Nội
và vùng phụ cận
3.1.1. Kết quả PCR phát hiện CIAV
Bâng 1 trình bày kết quâ PCR phát hiện
CIAV trong 124 méu bệnh phèm thu thêp ở 14
trang träi.
Trong 124 méu gà øm được kiểm tra, đã
phát hiện 63 méu dương tính (chiếm tỷ lệ
50,81%). Tùng hợp kết quâ theo trang träi cho
biết có 13/14 träi cò lưu hành CIAV, chiếm tỷ lệ
92,86%. Câu hói tiếp theo được đặt ra cho nhóm
nghiên cứu là phâi xác đðnh virus lưu hành là
chþng gây bệnh tự nhiên hay chþng virus
vacxin. Trên thực tế, vacxin nhược đûc phòng
bệnh thiếu máu truyền nhiễm có giçy phép
nhêp khèu vào Việt Nam từ nëm 2007. Theo
Bâng 1. Kết quâ PCR phát hiện CIAV trong mẫu bệnh phẩm
Phân nhóm Số mẫu kiểm tra
Âm tính Dương tính
Số mẫu Tỷ lệ (%) Số mẫu Tỷ lệ (%)
Theo cá thể 124 61 49,19 63 50,81
Theo trang trại 14 1 7,14 13 92,86

Đào Đoan Trang, Cao Thị Bích Phượng, Vũ Thị Ngọc, Nguyễn Văn Giáp, Huỳnh Thị Mỹ Lệ
39
thöng tư 10/2016/TT-BNNPTNT, hiện có 2 loäi
vacxin phòng bệnh thiếu máu truyền nhiễm
được phép lưu hành là Nobilis CAV P4
(Intervet) và AviPro Thymovac (Lohmann
Animal Health GmbH). Theo tìm hiểu cþa
chýng töi, cho đến nay, người chën nuöi gà ít
biết tới bệnh thiếu máu truyền nhiễm nên đäi
đa sø các cơ sở chën nuöi (trong đò cò 14 trang
träi kể trên) không sử dụng vacxin phòng bệnh
do CIAV gây ra. Vì vêy, mặc dù kỹ thuêt PCR
dùng trong nghiên cứu này không phân biệt
được chþng virus vacxin và chþng virus gây
bệnh nhưng kết quâ dương tính CIAV ở gà chưa
từng sử dụng vacxin phòng bệnh thiếu máu
truyền nhiễm cho thçy 50,81% méu gà øm
dương tính CIAV là do nhiễm tự nhiên.
Trên thế giới, đã cò nhiều công bø khîng đðnh
CIAV lưu hành với tỷ lệ cao ở gà. Ví dụ, bìng
phân ứng PCR đã xác đðnh được 55,40% méu
dương tính với CIAV täi vùng Ontario (Canada)
(Eregae, 2014). Täi Thù Nhï Kỳ, có tới 80% sø
trang träi kiểm tra dương tính với CIAV với tỷ lệ
nhiễm virus trung bình là 55,80% (Hadimli et al.,
2008). Trong khi đò, täi Iran, CIAV cñn được phát
hiện ở gà không có biểu hiện triệu chứng lâm sàng
cþa bệnh thiếu máu truyền nhiễm với tỷ lệ dao
đûng từ 24,58 - 58,40% (Gholami-Ahangaran,
2011, Gholami-Ahangaran, 2012). Cùng với kết
quâ phát hiện 73,1% méu dương tính huyết thanh
hõc với CIAV ở gà thu thêp täi Hà Nûi và Hà Nam
nëm 2013 (Trinh et al., 2015), nghiên cứu này góp
phæn khîng đðnh CIAV lưu hành phù biến không
chî ở đàn gà nuöi täi Hà Nûi và vùng phụ cên mà
còn ở nhiều nước chën nuöi gà trên thế giới, trong
đò cò Việt Nam.
3.1.2. Sự lưu hành của CIAV theo lứa tuổi
Bìng phân ứng huyết thanh hõc, mût sø
nghiên cứu đã phát hiện được CIAV ở gà từ 1 -
43 ngày tuùi (Roussan, 2006) và từ 2 - 6 tuæn
tuùi (Karimi, 2010). Tuy nhiên, CIAV thường
gây bệnh thể låm sàng cho gà dưới 3 tuæn tuùi
(McIlroy et al., 1992) và gây bệnh thể cên lâm
sàng ở gà trên 3 tuæn tuùi (Adair, 2000). Với cën
cứ trên, nghiên cứu này đã phån tích sự lưu
hành CIAV theo 3 nhóm tuùi (Bâng 2).
Bâng 2 cho biết tỷ lệ méu dương tính CIAV
cao nhçt ở nhóm gà 3 - 5 tuæn tuùi (56,94%) và
cao hơn so với nhóm gà ≥ 6 tuæn tuùi (50,00%).
Tuy nhiên, sự sai khác này khöng cò ý nghïa
thøng kê (P > 0,05). Theo khuyến cáo sử dụng
vacxin phòng bệnh thiếu máu truyền nhiễm
(Nobilis CAV P4), trong mõi trường hợp, không
được dùng vacxin cho gà nhó hơn 6 tuæn tuùi.
Do đò, việc phát hiện CIAV ở các đàn gà chưa
được tiêm vacxin phòng bệnh thiếu máu truyền
nhiễm (Bâng 1) và tỷ lệ dương tính cao ở nhóm
gà 3 - 5 tuæn tuùi mût læn nữa khîng đðnh CIAV
ở đàn gà nuöi täi Hà Nûi và vùng phụ cên không
phâi là các chþng virus vacxin.
Mặc dù CIAV có thể nhiễm cho gà ở mõi lứa
tuùi (McNulty, 1991) nhưng nghiên cứu này
không phát hiện được CIAV ở nhóm gà < 3 tuæn
tuùi. Cÿng bìng kỹ thuêt PCR, mût nghiên cứu
ở Ấn Đû đã xác đðnh CIAV nhiễm cao nhçt ở gà
< 3 tuæn tuùi (80,30%), tiếp đến là ở gà 3 - 7
tuæn tuùi (66,60%) và thçp nhçt ở gà 7 - 12 tuæn
tuùi (25%) (Wani et al., 2013). Khi so sánh tỷ lệ
nhiễm CIAV ở nhóm gà < 3 tuæn tuùi, kết quâ
cþa nghiên cứu này (0%) và nghiên cứu kể trên
(80,30%) là trái ngược. Sự tương phân kể trên,
theo chýng töi là do dung lượng méu nhó cþa
nhóm gà < 3 tuæn tuùi (8/124 méu xét nghiệm).
Bâng 2. Sự lưu hành CIAV theo lứa tuổi
Tuần tuổi
Theo cá thể Theo trang trại
Số mẫu kiểm tra
Số mẫu dương tính
Tỷ lệ (%) Số trại
kiểm tra Số trại
dương tính Tỷ lệ (%)
< 3 8 0 0 1 0 0
3 ÷ 5 72 41 56,94 6 6 100
≥ 6 44 22 50,00 7 7 100
Tổng hợp 124 63 50,81 14 13 92,86

Sự lưu hành của virus gây bệnh thiếu máu truyền nhiễm ở gà (CIAV) tại Hà Nội và vùng phụ cận
40
Chính vì hän chế này, kết quâ nghiên cứu hiện
thời chưa phân ánh đæy đþ tình hình nhiễm
CIAV theo nhóm tuùi. Để làm rô đặc điểm lưu
hành cþa CIAV theo lứa tuùi ở gà nuôi täi Hà
Nûi và vùng phụ cên, cæn thực hiện các nghiên
cứu tiếp theo với dung lượng lớn hơn.
3.2. Đặc điểm sinh học phân tử của gen ORF1
3.2.1. Trình tự nucleotide của gen ORF1
Trong sø 63 méu dương tính CIAV cþa 13
trang träi, 2 méu ngéu nhiên cþa mût träi được
chõn để giâi trình tự. Sau khi lược bó méu có
trình tự giøng nhau, có 7 trình tự gen đäi diện
được giữ läi (Hình 1). Kết quâ so sánh trình tự
phån đoän gen ORF1 giữa 7 chþng CIAV thực
đða (C1, C5, C8, C10, C12, C13 và C16) cho thçy
giữa chúng có mức tương đ÷ng rçt cao về trình
tự nucleotide (96,7 - 99,8%); có 561/582 (93,39%)
vð trí khöng đût biến nucleotide và 21/582
(3,61%) vð trí đût biến (đa hình nucleotide,
polymorphic).
Khi so sánh phân đoän gen ORF1
(nucleotide 1 - 580) giữa các chþng CIAV cþa
Việt Nam với 3 chþng CIAV dùng sân xuçt
vacxin phòng bệnh thiếu máu truyền nhiễm, kết
quâ được trình bày ở hình 2.
Hình 1. Trình tự gen ORF1 của CIAV (nucleotide 1-580)
Ghi chú: Dçu “.” biểu thị các nucleotide giống với trình tự cûa chûng virus tham chiếu Vx_P4_AJ890284
10 20 30 40 50 60 70 80 90 100 110 120 130 140 150
....|....|....|....|.. ..|....|....|....|....|... .|....|....|....|....|....| ....|....|....|....|....|. ...|....|....|....|....|.. ..|....|....|....|....|
Vx_P4_AJ890284 ATGGCAAGACGAGCTCGCAGACCGAGAGGCCGGTTTTACGCCTTCAGAAGAGGACGGTGGCACCACCTCAAGCGACTTCGACGAAGATATAAATTTCGACATCGGAGGAGACAGCGGTATCGTAGACGAGCTTTTAGGAAGGCCTTTCAC
Vx_3711_EF683159 ................................A.....................................................................................................................
Cux1_M55918 ................................A......T..............................................................................................................
Vn_VP10_KP780292 ................................A................................G....................................................................................
Vn_VP9_KP780291 ................................A.....................................................................................................................
Vn_VP8_KP780290 ................................A.....................................................................................................................
Vn_VP7_KP780289 ................................A................................G....................................................................................
Vn_HN1_KP780288 ................................A.....................................................................................................................
Vn_BN1_KP780287 ................................A.....................................................................................................................
C1_2016 ................................A..............................A......................................................................................
C5_2016 ................................A.....................................................................................................................
C8_2016 ................................A..............................A......................................................................................
C10_2016 ................................A..............................A......................................................................................
C13_2017 ................................A.....................................................................................................................
C12_2017 ................................A.....................................................................................................................
C16_2017 ................................A.....................................................................................................................
160 170 180 190 200 210 220 230 240 250 260 270 280 290 300
....|....|....|....|.. ..|....|....|....|....|... .|....|....|....|....|....| ....|....|....|....|....|. ...|....|....|....|....|.. ..|....|....|....|....|
Vx_P4_AJ890284 AACCCCCGCCCCGGTACGTATAGTGTGAGGCTGCCGAACCCCCAATCTACTATGACTATCCGATTCCAAGGAGTCATCTTTCTCACGGAAGGACTCATTCTGCCTAAAAACAGCACAGCGGGGGACTATGCAGACCACATGTACGGGGCG
Vx_3711_EF683159 ...................................A........G..C..G...........C.......................C..............A.................T....G......G..................
Cux1_M55918 ..............................................................C........G....................................................G.........................
Vn_VP10_KP780292 ................................A...........G..A..G...........C..T......A....T.....T..C.....................................G.............C...........
Vn_VP9_KP780291 ..............................................................C....................................T........................G.........................
Vn_VP8_KP780290 ..............................................................C....................................T........................G.........................
Vn_VP7_KP780289 ................................A...........G..A..G...........C..T......A....T.....T..C.....................................G.............C...........
Vn_HN1_KP780288 ..............................................................C....................................T........................G.........................
Vn_BN1_KP780287 ..............................................................C....................................T........................G.........................
C1_2016 ..............................................................C........CA...................................................G.............T...........
C5_2016 ..............................................................C....................................T........................G.........................
C8_2016 ..............................................................C........CA...........................................C.......G.............T...........
C10_2016 ..............................................................C........CA...........................................C.......G.............T...........
C13_2017 .............................A................................C.............................................................G.........................
C12_2017 .............................A................................C....................................T........................G.........................
C16_2017 ..............................................................C....................................T........................G.........................
310 320 330 340 350 360 370 380 390 400 410 420 430 440 450
....|....|....|....|.. ..|....|....|....|....|... .|....|....|....|....|....| ....|....|....|....|....|. ...|....|....|....|....|.. ..|....|....|....|....|
Vx_P4_AJ890284 AGAGTCGCCAAGATCTCTGTAAACCTGAAAGAGTTCCTGCTAGCGTCAATGAACCTGACATACGTGAGCAAAATCGGAGGACCCATCGCCGGTGAGTTGATTGCGGACGGGTCTAAATCACAAGCCGCGGAGAATTGGCCTAATTGCTGG
Vx_3711_EF683159 .................A..G.................C.....A..........................G..A.....C......................................G..............C.....A.........
Cux1_M55918 ....................G.......................C...................................C..................................................C..................
Vn_VP10_KP780292 .................A..G.................C...................................A.....C.................................C..........A...C................T...
Vn_VP9_KP780291 ....................G........G..........................C...............C.......C.....................................................C........C......
Vn_VP8_KP780290 ....................G........G..........................C...............C.......C.....................................................C........C......
Vn_VP7_KP780289 .................A..G.................C...................................A.....C.................................C..........A...C................T...
Vn_HN1_KP780288 ....................G........G..........................C...............C.......C.....................................................C........C......
Vn_BN1_KP780287 ....................G........G..........................C...............C.......C.....................................................C........C......
C1_2016 .................C..G...........................................................C.................................C....T.........C....C...............
C5_2016 ....................G........G..........................C...............C.......C.....................................................C........C......
C8_2016 .................C..G.............................................................................................C....T.........C....C...............
C10_2016 .................C..G...........................................................C.................................C....T.........C....C...............
C13_2017 ....................G........G..........................A...............C.......C.....................................................C...............
C12_2017 ....................G........G..........................A...............C.......C.....................................................C...............
C16_2017 ....................G........G..........................C...............C.......C.....................................................C........C......
460 470 480 490 500 510 520 530 540 550 560 570 580
....|....|....|....|.. ..|....|....|....|....|... .|....|....|....|....|....| ....|....|....|....|....|. ...|....|....|....|....|.. ..|..
Vx_P4_AJ890284 CTGCCGCTAGATAATAACATGCCCTCCGCGACACCATCGGCATGGTGGAGATGGGCCTTAATGATGATGCAGCCCACGGACTCTTGCCGGTTCTTTAATCACCCTAAGCAGATGACCCTGCAAGACATGGGT
Vx_3711_EF683159 ..................G..........T........T.................T.................A........C........T..............A..A.....................
Cux1_M55918 ..................G..........T..........................................................................A...........................
Vn_VP10_KP780292 ..................G..........T..........................T..................................................A..A.....................
Vn_VP9_KP780291 ..A...............G.A.......................................................................T.................A.....................
Vn_VP8_KP780290 ..A...............G.A.......................................................................T.................A.....................
Vn_VP7_KP780289 ..................G..........T..........................T..................................................A..A.....................
Vn_HN1_KP780288 ..A...............G...........................................................................................A.....................
Vn_BN1_KP780287 ..A...............G...........................................................................................A.....................
C1_2016 ..................G...................A.............................................................................................
C5_2016 ..A...............G...................A.................G.....................................................A.....................
C8_2016 ..................G.................................................................................................................
C10_2016 ..................G.................................................................................................................
C13_2017 ..................G...................A..................................................A....................A.....................
C12_2017 ..................G...................A..................................................A....................A.....................
C16_2017 ..A...............G...........................................................................................A.....................
Đào Đoan Trang, Cao Thị Bích Phượng, Vũ Thị Ngọc, Nguyễn Văn Giáp, Huỳnh Thị Mỹ Lệ
41
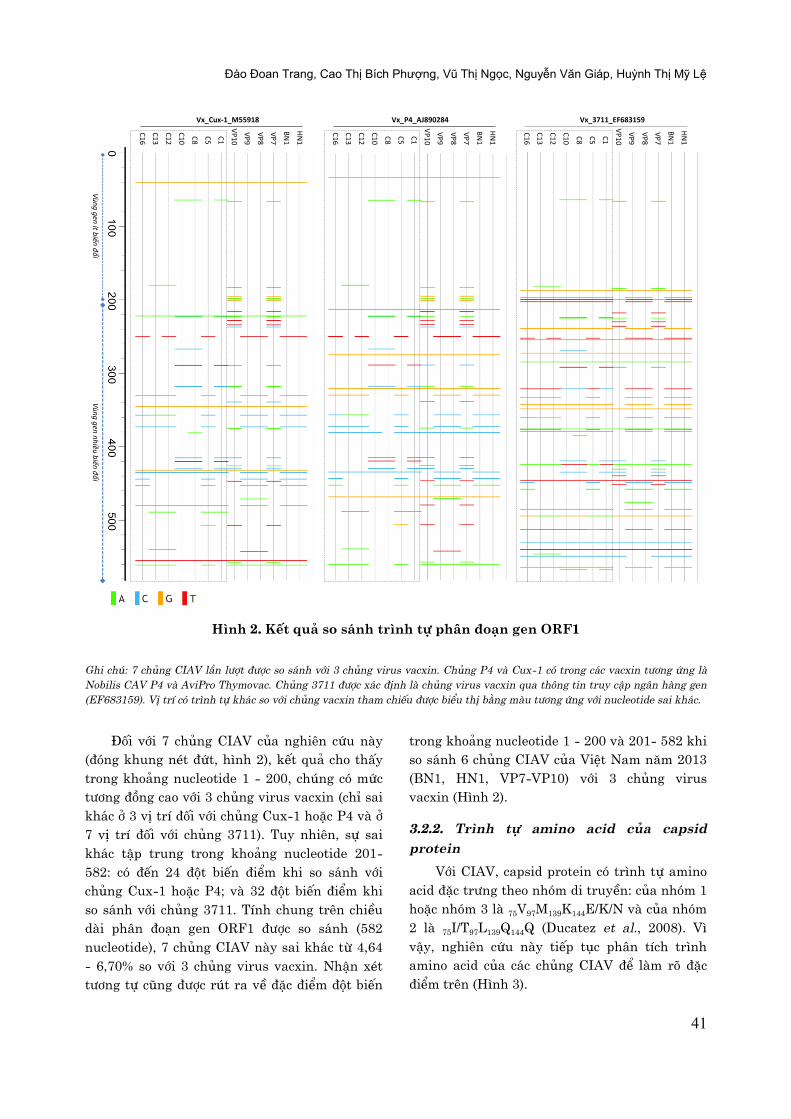
Hình 2. Kết quâ so sánh trình tự phân đoạn gen ORF1
Ghi chú: 7 chûng CIAV læn lượt được so sánh với 3 chûng virus vacxin. Chûng P4 và Cux-1 có trong các vacxin tương ứng là
Nobilis CAV P4 và AviPro Thymovac. Chûng 3711 được xác định là chûng virus vacxin qua thông tin truy cập ngån hàng gen
(EF683159). Vị trí có trình tự khác so với chûng vacxin tham chiếu được biểu thị bằng màu tương ứng với nucleotide sai khác.
Đøi với 7 chþng CIAV cþa nghiên cứu này
(đòng khung nét đứt, hình 2), kết quâ cho thçy
trong khoâng nucleotide 1 - 200, chúng có mức
tương đ÷ng cao với 3 chþng virus vacxin (chî sai
khác ở 3 vð trí đøi với chþng Cux-1 hoặc P4 và ở
7 vð trí đøi với chþng 3711). Tuy nhiên, sự sai
khác têp trung trong khoâng nucleotide 201-
582: cò đến 24 đût biến điểm khi so sánh với
chþng Cux-1 hoặc P4; và 32 đût biến điểm khi
so sánh với chþng 3711. Tính chung trên chiều
dài phån đoän gen ORF1 được so sánh (582
nucleotide), 7 chþng CIAV này sai khác từ 4,64
- 6,70% so với 3 chþng virus vacxin. Nhên xét
tương tự cÿng được rút ra về đặc điểm đût biến
trong khoâng nucleotide 1 - 200 và 201- 582 khi
so sánh 6 chþng CIAV cþa Việt Nam nëm 2013
(BN1, HN1, VP7-VP10) với 3 chþng virus
vacxin (Hình 2).
3.2.2. Trình tự amino acid của capsid
protein
Với CIAV, capsid protein có trình tự amino
acid đặc trưng theo nhòm di truyền: cþa nhóm 1
hoặc nhóm 3 là 75V97M139K144E/K/N và cþa nhóm
2 là 75I/T97L139Q144Q (Ducatez et al., 2008). Vì
vêy, nghiên cứu này tiếp tục phân tích trình
amino acid cþa các chþng CIAV để làm rô đặc
điểm trên (Hình 3).
C16
C13
C12
C10
C8
C5
C1
Vx_Cux-1_M55918
HN1
BN1
VP8
VP7
VP9
VP10
C16
C13
C12
C10
C8
C5
C1
Vx_P4_AJ890284
HN1
BN1
VP8
VP7
VP9
VP10
C16
C13
C12
C10
C8
C5
C1
Vx_3711_EF683159
HN1
BN1
VP8
VP7
VP9
VP10
Vù
ng
gen
ít biến
đổ
iV
ùn
g g
en n
hiều
biến
đổ
i

Sự lưu hành của virus gây bệnh thiếu máu truyền nhiễm ở gà (CIAV) tại Hà Nội và vùng phụ cận
42
Hình 3. Trình tự amino acid suy diễn của một phần protein VP1
Ghi chú: Các vị trí amino acid đặc trưng được đánh dçu màu xám. Đóng khung là vùng siêu biến đổi (amino acid 139-151) cûa
protein VP1. Dçu “.” biểu thị các amino acid giống với trình tự tham chiếu Vx_P4_AJ890284
Phân tích trình tự amino acid suy diễn (vð
trí 1 - 194 cþa protein VP1) cþa 7 chþng CIAV
đäi diện cho biết sự tương đ÷ng rçt cao (giøng
nhau ở 188/194 vð trí). Đøi với trình tự amino
acid đặc trưng, cò 4 chþng (C5, C12, C13 và
C16) mang trình tự amino acid 75V97M139K144E là
đặc trưng cþa nhóm 1 hoặc nhóm 3; 3 chþng còn
läi (C1, C8 và C10) mang trình tự amino acid
75I97L139Q144Q là đặc trưng cþa nhòm 2. Như vêy,
dự đoán cò ít nhçt hai nhóm di truyền cþa CIAV
lưu hành ở gà täi các đða điểm lçy méu. Ở 4 vð
trí amino acid đặc trưng về nhóm di truyền, 2 vð
trí nìm ở vùng siêu biến đùi. Mût sø nghiên cứu
trước đåy cho biết vùng siêu biến đùi có ânh
hưởng tới đûc lực cþa CIAV (Renshaw et al.,
1996, Yamaguchi et al., 2001). Vì vêy, cæn tiếp
tục nghiên cứu phân lêp virus và đánh giá đûc
lực cþa các chþng CIAV lưu hành ở Việt Nam.
3.2.3. Cây phát sinh chủng loại của CIAV
dựa vào gen ORF1
Bìng phân tích cây phát sinh chþng loäi và
sự sai khác trình tự amino acid cþa capsid
protein VP1, sự khác biệt giữa 7 chþng CIAV
cþa nghiên cứu này với 3 chþng virus vacxin và
6 chþng CIAV cþa Việt Nam (công bø nëm 2013)
được làm rõ ở hình 4.
Giøng như mût sø nghiên cứu đã cöng bø
(Olszewska-Tomczyk et al., 2016, Snoeck et al.,
2012), cây phát sinh chþng loäi dựa vào mût
phæn trình tự gen ORF1 (Hình 4) cho biết CIAV
có thể chia thành 3 nhòm. Trong đò, 100%
chþng virus lưu hành ở Việt Nam thuûc về
nhòm 2 và 3. Đáng chý ý, 7/7 chþng CIAV cþa
nghiên cứu này không nìm chung nhánh với các
chþng virus vacxin (chþng P4, Cux-1 và 3711).
Thêm vào đò, kết quâ so sánh trình tự amino
acid với chþng virus vacxin (P4) cho biết: (i) 4
chþng CIAV thực đða trong nghiên cứu này
(chþng C5, C12, C13 và C16) có 3 vð trí sai khác
và (ii) 3 chþng (C1, C8 và C10) có 7 vð trí sai
khác (đòng khung, Hình 2). Như vêy, các kết
quâ trình bày ở trên cho thçy: (i) có hai nhóm di
truyền cþa CIAV lưu hành ở gà nuôi täi Hà Nûi
và vùng phụ cên, (ii) các chþng virus này khác
với chþng virus vacxin phòng bệnh thiếu máu
10 20 30 40 50 60 70 80 90 100
....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|
Vx_P4_AJ890284 MARRARRPRGRFYAFRRGRWHHLKRLRRRYKFRHRRRQRYRRRAFRKAFHNPRPGTYSVRLPNPQSTMTIRFQGVIFLTEGLILPKNSTAGDYADHMYGA
Vx_3711_EF683159 ...........................................................................................G........
Cux1_M55918 .............S.............................................................................G........
Vn_VP10_KP780292 .....................Q....................................................I................G....L...
Vn_VP9_KP780291 ...........................................................................................G........
Vn_VP8_KP780290 ...........................................................................................G........
Vn_VP7_KP780289 .....................Q....................................................I................G....L...
Vn_HN1_KP780288 ...........................................................................................G........
Vn_BN1_KP780287 ...........................................................................................G........
C1_2016 .....................N....................................................I................G....L...
C5_2016 ...........................................................................................G........
C8_2016 .....................N....................................................I................G....L...
C10_2016 .....................N....................................................I................G....L...
C13_2017 ...........................................................................................G........
C12_2017 ...........................................................................................G........
C16_2017 ...........................................................................................G........
110 120 130 140 150 160 170 180 190
....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....|....
Vx_P4_AJ890284 RVAKISVNLKEFLLASMNLTYVSKIGGPIAGELIADGSKSQAAENWPNCWLPLDNNMPSATPSAWWRWALMMMQPTDSCRFFNHPKQMTLQDMG
Vx_3711_EF683159 ........................................................V.....................................
Cux1_M55918 ...........................................D............V.....................................
Vn_VP10_KP780292 ......................................Q....Q............V.....................................
Vn_VP9_KP780291 ........................L...............................V.....................................
Vn_VP8_KP780290 ........................L...............................V.....................................
Vn_VP7_KP780289 ......................................Q....Q............V.....................................
Vn_HN1_KP780288 ........................L...............................V.....................................
Vn_BN1_KP780287 ........................L...............................V.....................................
C1_2016 ......................................Q....Q............V.....................................
C5_2016 ........................L...............................V.....................................
C8_2016 ......................................Q....Q............V.....................................
C10_2016 ......................................Q....Q............V.....................................
C13_2017 ........................L...............................V.....................................
C12_2017 ........................L...............................V.....................................
C16_2017 ........................L...............................V.....................................

Đào Đoan Trang, Cao Thị Bích Phượng, Vũ Thị Ngọc, Nguyễn Văn Giáp, Huỳnh Thị Mỹ Lệ
43
Hình 4. Cây phát sinh chủng loại (phylogenetic tree) của CIAV dựa vào gen ORF1
Ghi chú: Dựa vào các chûng tham chiếu đã biết nhóm di truyền, CIAV được chia thành nhóm 1, nhóm 2 và nhóm 3 (phån
nhóm 3a, 3b). Giá trị bootstrap täi các nút (node) cûa cåy phát sinh chûng loäi được hiển thị cho phån nhánh chính. Bâng đính
kèm liệt kê các vị trí có thay đổi amino acid được mã hóa, trong đó các vị trí amino acid đặc trưng nhóm di truyền được đánh
dçu màu xám. Những vị trí amino acid có sai khác giữa 7 chûng CIAV trong nghiên cứu này với chûng virus vacxin P4 được
đóng khung.
truyền nhiễm. Các kết quâ thu được cþa nghiên
cứu này góp phæn làm rõ thêm sự lưu hành rçt
phù biến và đa nhòm di truyền cþa CIAV ở Việt
Nam và ở các nước trên thế giới (Kim et al.,
2010, Snoeck et al., 2012, Wani et al., 2013, van
Santen et al., 2001).
4. KẾT LUẬN
CIAV lưu hành phù biến ở đàn gà nuöi täi
Hà Nûi và vùng phụ cên với 92,86% sø trang
träi và 50,81% sø gà øm kiểm tra dương tính
CIAV.
7 chþng CIAV nëm 2016 - 2017 thuûc về 2
nhóm di truyền là nhóm 2 và nhóm 3. Các
chþng này khác với chþng virus vacxin phòng
bệnh thiếu máu truyền nhiễm.
LỜI CẢM ƠN
Têp thể tác giâ xin chân thành câm ơn dự
án Việt- Bî nëm 2017 (mã sø 04/DAVB) đã tài
trợ kinh phí cho nghiên cứu này.
TÀI LIỆU THAM KHẢO
Adair, B. M. (2000). Immunopathogenesis of chicken
anemia virus infection. Dev Comp Immunol., 24,
247-255.
Balamurugan, V. and J. M. Kataria (2006).
Economically Important Non-oncogenic
Immunosuppressive Viral Diseases of Chicken-
Current Status. Veterinary Research
Communications, 30: 541-566.
Bougiouklis, P. A., M. Sofia, G. Brellou, I. Georgopoulou,
C. Billinis and I. Vlemmas (2007). A clinical case of
chicken infectious anemia disease and virus DNA
Vị trí amino acid
3a
2
1
14 22 75 92 97 125 139 144 152
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V G M L K E V
A H V D M I K E M
S H V G M I K D V
A N I G L I Q Q V
A N I G L I Q Q V
A N I G L I Q Q V
A Q I G L I Q Q V
A Q I G L I Q Q V
A H V G M I K E V
3b94,2%

Sự lưu hành của virus gây bệnh thiếu máu truyền nhiễm ở gà (CIAV) tại Hà Nội và vùng phụ cận
44
detection in naturally infected broilers in Greece.
Avian Dis., 51: 639-642.
Dohms, J. E. and Y. M. Saif, 1984). Criteria for
evaluating immunosuppression. Avian Dis., 28:
305-310.
Ducatez, M. F., H. Chen, Y. Guan and C. P. Muller
(2008). Molecular epidemiology of chicken anemia
virus (CAV) in southeastern Chinese live birds
markets. Avian Dis., 52: 68-73.
Ducatez, M. F., A. A. Owoade, J. O. Abiola and C. P.
Muller (2006). Molecular epidemiology of chicken
anemia virus in Nigeria. Arch Virol., 151.
Eregae, M. E. (2014). The Epidemiology of chicken anaemia virus, fowl adenovirus, and infectious bursal disease virus. Ontario broiler flocks, p. 350. Guelph, Ontario, Canada.
Gholami-Ahangaran, M., Momtaz, H., Zia-Jahromi, N. and Momeni, M. (2011). Genomic detection of the chicken anaemia virus from apparently healthy commercial broiler chickens in Iran. Revue de Médecine Vétérinaire, 162: 604-606.
Gholami-Ahangaran, M. a. Z.-J., N. (2012). Serological and molecular identification of subclinical chicken anaemia virus infection in broiler chickens in Iran. African Journal of Microbiology Research, 6: 4471-4474.
Hadimli, H. H., O. Erganis, L. Guler and U. S. Ucan (2008). Investigation of chicken infectious anemia virus infection by PCR and ELISA in chicken flocks. Turk. J. Vet. Anim. Sci., 32: 79-84.
Hall, T. A. (1999). BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser., 41: 95-98.
Haridy, M., J. Sasaki, M. Ikezawa, K. Okada and M. Goryo (2012). Pathological and immunohistochemical studies of subclinical infection of chicken anemia virus in 4-week-old chickens. J Vet Med Sci., 74.
Hoerr, F. J. (2010). Clinical aspects of immunosuppression in poultry. Avian Dis., 54: 2-15.
Islam, A. F., C. W. Wong, S. W. Walkden-Brown, I. G. Colditz, K. E. Arzey and P. J. Groves (2002). Immunosuppressive effects of Marek's disease virus (MDV) and herpesvirus of turkeys (HVT) in broiler chickens and the protective effect of HVT vaccination against MDV challenge. Avian Pathol., 31: 449-461.
Karimi, I., Mahzounieh, M., Bahadoran, S. and Azad,
F. (2010). Chicken anemia virus infection in
broiler chickens in Shahrekord, Iran: Serological,
hematological, and histopathological findings.
Comparative Clinical Pathology, 19: 63-67.
Kim, H. R., Y. K. Kwon, Y. C. Bae, J. K. Oem and O.
S. Lee (2010). Molecular characterization of
chicken infectious anemia viruses detected from
breeder and broiler chickens in South Korea. Poult
Sci., 89: 2426-2431.
MacLachlan, N. J. and E. J. Dubovi (2017). Chapter 13
- Circoviridae and Anelloviridae. In: Dubovi E. J.
(Ed.), Fenner's Veterinary Virology (Fifth Edition),
pp. 259-268. Academic Press, Boston.
McIlroy, S. G., M. S. McNulty, D. W. Bruce, J. A.
Smyth, E. A. Goodall and M. J. Alcorn (1992).
Economic effects of clinical chicken anemia agent
infection on profitable broiler production. Avian
Dis., 36: 566-574.
McNulty, M. S., 1991). Chicken anaemia agent: a
review. Avian Pathol., 20: 187-203.
McNulty, M. S., S. G. McIlroy, D. W. Bruce and D.
Todd (1991). Economic effects of subclinical
chicken anemia agent infection in broiler chickens.
Avian Dis., 35: 263-268.
Noteborn, M. H., G. F. de Boer, D. J. van Roozelaar, C.
Karreman, O. Kranenburg, J. G. Vos, S. H.
Jeurissen, R. C. Hoeben, A. Zantema, G. Koch et
al. (1991). Characterization of cloned chicken
anemia virus DNA that contains all elements for
the infectious replication cycle. J Virol., 65: 3131-
3139.
Olszewska-Tomczyk, M., E. Swieton, Z. Minta and K.
Smietanka (2016). Occurrence and phylogenetic
studies of chicken anemia virus from Polish broiler
flocks. Avian Dis., 60: 70-74.
Oluwayelu, D. O. (2010). Diagnosis and epidemiology
of chicken infectious anemia in Africa. African
Journal of Biotechnology, 9: 2043-2049.
Renshaw, R. W., C. Soine, T. Weinkle, P. H.
O'Connell, K. Ohashi, S. Watson, B. Lucio, S.
Harrington and K. A. Schat (1996). A
hypervariable region in VP1 of chicken infectious
anemia virus mediates rate of spread and cell
tropism in tissue culture. J Virol, 70: 8872-8878.
Rimondi, A., S. Pinto, V. Olivera, M. Dibarbora, M.
Perez-Filgueira, M. I. Craig and A. Pereda (2014).
Comparative histopathological and immunological
study of two field strains of chicken anemia virus.
Vet Res., 45: 102.
Roussan, D. A. (2006). Serological survey on the
prevalence of chicken infectious anemia virus in
commercial broiler chicken flocks in Northern
Jordan. International Journal of Poultry Science, 5:
544-546.
Schonewille, E., A. Singh, T. W. Gobel, W. Gerner, A.
Saalmuller and M. Hess (2008). Fowl adenovirus
(FAdV) serotype 4 causes depletion of B and T
cells in lymphoid organs in specific pathogen-free
chickens following experimental infection. Vet
Immunol Immunopathol., 121: 130-139.
Snoeck, C. J., G. F. Komoyo, B. P. Mbee, E. Nakoune,
A. Le Faou, M. P. Okwen and C. P. Muller (2012).
Epidemiology of chicken anemia virus in Central
African Republic and Cameroon. Virol J., 9: 189.

Đào Đoan Trang, Cao Thị Bích Phượng, Vũ Thị Ngọc, Nguyễn Văn Giáp, Huỳnh Thị Mỹ Lệ
45
Subler, K. A., C. S. Mickael and D. J. Jackwood
(2006). Infectious bursal disease virus-induced
immunosuppression exacerbates Campylobacter
jejuni colonization and shedding in chickens.
Avian Dis., 50: 179-184.
Sun, S., Z. Cui, J. Wang and Z. Wang (2009). Protective
efficacy of vaccination against highly pathogenic
avian influenza is dramatically suppressed by early
infection of chickens with reticuloendotheliosis virus.
Avian Pathol., 38: 31-34.
Tamura, K., G. Stecher, D. Peterson, A. Filipski and S.
Kumar (2013). MEGA6: Molecular Evolutionary
Genetics Analysis version 6.0. Mol Biol Evol., 30:
2725-2729.
Toro, H., S. Ewald and F. J. Hoerr (2006). Serological
evidence of chicken infectious anemia virus in the
United States at least since 1959. Avian Diseases,
50: 124-126.
Trinh, D. Q., H. Ogawa, V. N. Bui, T. T. Nguyen, D.
Gronsang, T. Baatartsogt, M. K. Kizito, M.
AboElkhair, S. Yamaguchi, V. K. Nguyen and
K. Imai (2015). Development of a blocking latex
agglutination test for the detection of antibodies
to chicken anemia virus. J Virol Methods, 221:
74-80.
van Santen, V. L., L. Li, F. J. Hoerr and L. H.
Lauerman (2001). Genetic characterization of
chicken anemia virus from commercial broiler
chickens in Alabama. Avian Dis., 45: 373-388.
Wani, M. Y., K. Dhama, R. Barathidasan, V.
Gowthaman, R. Tiwari, P. Bhatt, N. K. Mahajan, M.
M. Chawak, S. D. Singh and J. M. Kataria (2013).
Molecular detection and epidemiology of chicken
infectious anaemia virus in India. South Asian
Journal of Experimental Biology, 3: 145-151.
Yamaguchi, S., T. Imada, N. Kaji, M. Mase, K.
Tsukamoto, N. Tanimura and N. Yuasa (2001).
Identification of a genetic determinant of
pathogenicity in chicken anaemia virus. J Gen
Virol., 82: 1233-1238.





























